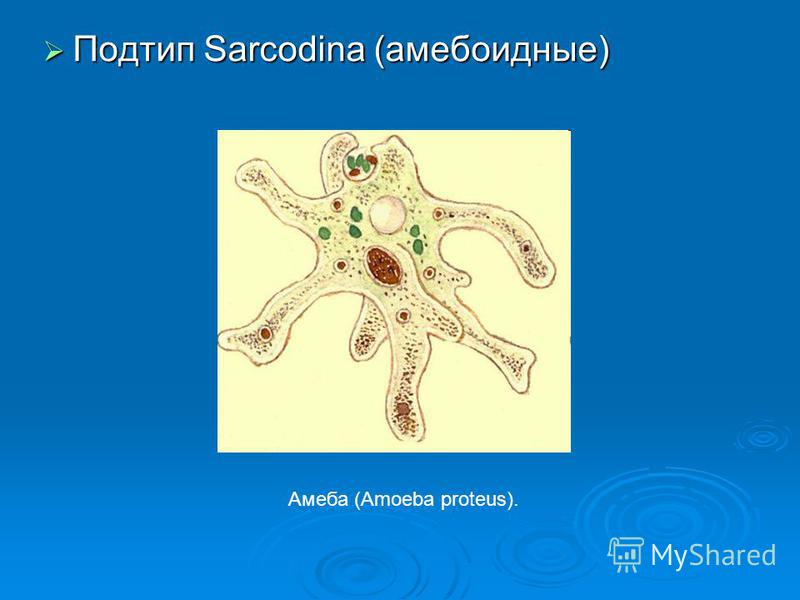
Счастливый фермер
Даже некоторые одноклеточные животные — почвенные амебы Dictyostelium discoideum — в состоянии организовать эффективное сельское хозяйство.
Правила жизни
Теги:
№65
биология
амеба
Описание
Dictyostelium discoideum (диктиостелиум) — клеточный слизевик, относящийся к типу Mycetozoa (миксомицеты). Большую часть времени существует в виде одиночных почвенных амеб. При определенных условиях такие амебы способны группироваться в единый подвижный организм. Диктиостелиум делится на три «пола», каждый из которых может спариваться с любым из двух других. Распространен в смешанных и широколиственных лесах умеренного пояса. Обитает в почве и подстилке.
История изучения
Диктиостелиум был описан в 1935 году.
Миграция
Диктиостелиум питается бактериями. Когда пища заканчивается, амебы собираются вместе и формируют так называемую слизевую плесень — многоклеточный организм длиной до 1 см. В этом состоянии колония перебирается на новое место, образовывает плодовое тело и разбрасывает споры, из которых появляются новые одиночные амебы.
Альтруизм
В результате изучения процесса формирования из отдельных амеб многоклеточного организма (слизевой плесени) было обнаружено, что диктиостелиуму свойственно социальное поведение. Одноклеточные организмы, образующие ножку плодового тела, фактически жертвуют собой ради других амеб, у которых появляется возможность продолжить род.
Сельское хозяйство
Ученые из американского Университета Райса обнаружили, что перед миграцией некоторые амебы не съедают все доступные бактерии, а берут часть из них с собой. Дальнейшие исследования показали, что диктиостелиум выпускает бактерии на новом месте, дожидается, пока они размножатся, и только после этого «собирает урожай».
Эксперименты
Заметив, что иногда в плодовом теле диктиостелиума вместе со спорами находятся и бактерии, ученые переместили несколько экземпляров в стерильные чашки Петри. Через два дня в некоторых из них появились колонии бактерий. Это означало, что амебы-фермеры «высадили» перенесенные бактерии и позволили им размножаться. Ученые уничтожили бактерии с помощью антибиотиков, а амеб перенесли на новые участки, заселенные другими бактериями. Некоторые клоны диктиостелиума начали взаимодействовать с микроорганизмами — то есть не только питаться ими, но и включать часть из них в плодовые тела для дальнейшего переноса на новое место.
Дикие и домашние
В ходе экспериментов стало ясно, что способностями к сельскому хозяйству обладают не все представители диктиостелиума. Амебы, которых из поколения в поколение выращивают в лабораторных условиях, к этой деятельности не склонны. Переносить бактерии с места на место умеют только около трети клонов, живущих и размножающихся в природе.
Кочевые и оседлые
Поскольку колониям амеб с бактериями не нужно искать места, населенные большим количеством бактерий, они перемещаются на гораздо меньшее расстояние. Данный факт позволяет ученым проводить параллели с историей человечества: переход от охоты и собирательства к земледелию и животноводству сопровождался и сменой кочевого образа жизни на оседлый.
Недостатки сельского хозяйства
Фермерство не всегда является выгодным занятием. В богатых бактериями местах выигрывают те амебы, которые перемещаются с одного участка на другой налегке.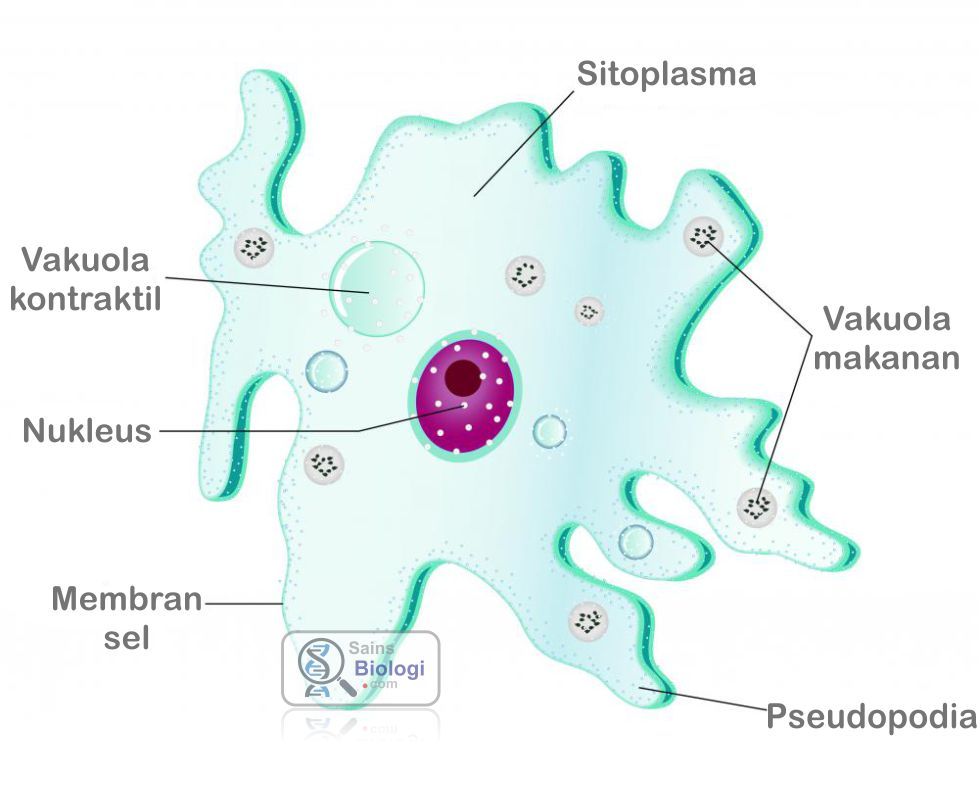 Они дают более многочисленное потомство, поскольку им не приходится ограничивать себя в еде. Зато амебы-фермеры лучше выживают там, где пищи изначально не хватает.
Они дают более многочисленное потомство, поскольку им не приходится ограничивать себя в еде. Зато амебы-фермеры лучше выживают там, где пищи изначально не хватает.
Другие животные-фермеры
Рыбы-ласточки Stegastes nigricans, которые встречаются между Индийским и Тихим океанами и в тропическом поясе Атлантического, выращивают для себя особый вид водорослей — красные водоросли рода полисифония. Они защищают свои посевы от других океанских потребителей растительной пищи и очищают поля от посторонних водорослей. В результате красные водоросли, которыми питаются рыбы-ласточки, растут только на возделываемых ими полях.
Морская улитка Littoraria irrorata, обитающая на Атлантическом побережье Северной Америки, занимается разведением питательного грибка. Улитки специально повреждают стебли растений, делая их таким образом уязвимыми для заражения.
Термиты подсемейства Macrotermitinae
занимаются разведением грибов определенного вида — Termitomyces, который растет только в термитниках, на грядках из материала, пропущенного через кишечник термитов.
Муравьи-листорезы Acromyrmex тоже выращивают грибную культуру — в подземных камерах, на переработанных зеленых листьях. Для защиты посевов от паразитов муравьи используют бактерии Pseudonocardia, которые выделяют антибиотик.
Некоторые жуки из подсемейства Scolytinae и Platypodinae разводят в деревьях «грибные сады»: рассеивают в них споры грибка, который питается древесиной. Впоследствии этот грибок становится пищей для личинок жуков.
Биологи обнаружили в «сибирских джунглях» новый вид почвенной амебы
8 апреля, 2022 14:08
Источник: Научная Россия
Ученые из Санкт-Петербургского государственного университета совместно с коллегами из Томска и Новосибирска обнаружили новый вид почвенной амебы Leptomyxa silvatica n.
Черневая тайга (так называемые сибирские джунгли) — регион Западной Сибири, включенный в реестр уникальных экосистем Всемирного фонда дикой природы. Этот регион отличается исключительным биоразнообразием.
Как отметила профессор СПбГУ Алла Лапидус, первое, что привлекает внимание в черневой тайге, — совершенно необъясненный гигантизм трав. К примеру, летом травяной покров «сибирских джунглей» имеет среднюю высоту 1,5–2 метра, то есть он способен полностью скрыть человека или всадника на коне.
«Мне пришла в голову идея, что этот гигантизм является результатом влияния не столько климатических условий, сколько, скорее всего, объясняется уникальными свойствами микробиоты почвы.И если в этом феномене хорошо разобраться, то, вполне возможно, это открытие можно будет использовать в сельском хозяйстве. У нас уже есть некоторые подтверждения данной гипотезы», — подчеркнула руководитель проекта, профессор СПбГУ Алла Лапидус.
Она добавила, что проект по изучению черневой тайги — междисциплинарный, он проводится в сотрудничестве с коллегами из Томского государственного университета. В проекте также участвуют сотрудники Всероссийского научно-исследовательского института сельскохозяйственной микробиологии, Института молекулярной и клеточной биологии СО РАН, Центрального сибирского ботанического сада СО РАН. Ученые проводят исследования в регионе Западной Сибири, уникальные свойства которого никем ранее не изучались столь многосторонне.
«Ученые проводили также исследования протозойного компонента почвы, в частности разнообразия амебоидных протистов, играющих очень важную роль в жизни почвенных микроорганизмов, круговороте вещества и энергии в почвенных местообитаниях.Нашими коллегами, участниками проекта из СПбГУ, были сделаны очень интересные находки, одной из которых является обнаружение нового вида амеб», — уточнила Алла Лапидус.
Доцент кафедры зоологии беспозвоночных СПбГУ Алексей Смирнов подчеркнул, что в рамках этого исследования был обнаружен ряд редких или вообще новых для науки видов амеб. Один из них — новый вид из отряда Leptomyxida, последовательность нуклеотидов в гене 18S рибосомальной РНК которого существенно отличается от уже всех известных видов. У нового вида амеб имеются и характерные морфологические отличия. Амеба получила название Leptomyxa silvatica (от латинского слова silva — лес). Полное название нового вида означает «происходящий из леса» в знак того, что изолирована амеба из лесного местообитания.
«Эта находка еще раз говорит о том, как мало мы знаем об огромном мире микроскопических животных. Численность гетеротрофных одноклеточных организмов в почвах может достигать огромных величин — десятков тысяч клеток в единственном грамме почвы.Они являются важнейшим природным компонентом, регулирующим численность почвенных грибов и бактерий. На морфологическом уровне нам известна лишь небольшая часть всего этого разнообразия. Исследования молекул ДНК, выделенных прямо из различных типов почв, показывают, что «своими глазами» мы видели не более 5% обитающих там организмов. Неоткрытыми остаются не только отдельные виды, но и целые крупные группы почвенных простейших. По этой причине так важны современные молекулярно-морфологические исследования, которые показывают нам, кому принадлежат обнаруженные сиквенсы и кто же на самом деле живет у нас в прямом смысле слова под ногами и вносит свой немаленький вклад в процессы, происходящие в почвах черневой тайги», — заключил Алексей Смирнов.
Ученые подчеркивают, что необходимо продолжать углубленное исследование черневой тайги, в том числе для защиты почвы «сибирских джунглей». Деградация редчайшего природного ресурса, считают эксперты, будет иметь необратимые негативные последствия, при которых восстановление уникальной экосистемы может оказаться невозможным.
Изучение собранных образцов самыми современными методами исследований стало возможным за счет созданного в СПбГУ уникального Научного парка — системы ресурсных центров, собравших в своем составе самое современное оборудование. В этом исследовании особенно активно было задействовано оборудование ресурсных центров «Развитие молекулярных и клеточных технологий», «Вычислительный центр СПбГУ», «Биобанк» и «Культивирование микроорганизмов».
Исследования черневой тайги в СПбГУ проводятся в рамках проекта под руководством профессора СПбГУ Аллы Лапидус, поддержанного Российским научным фондом (№19-16-00049).
В 2022 году Санкт-Петербургский университет празднует Год зоологии, который приурочен к двум юбилеям зоологического образования в СПбГУ. В этом году исполняется 200 лет со дня открытия при университете первой в России кафедры зоологии. Также СПбГУ отмечает 150 лет с момента официального разделения кафедры на два самостоятельных научно-образовательных отделения — зоологии беспозвоночных и зоологии позвоночных.
В честь памятных дат в университете также запланированы научно-историческая сессия «Зоология в СПбГУ: история и современность», Догелевские чтения, арт-выставка «200 лет зоологии СПбГУ», экскурсии по кафедрам зоологии позвоночных и беспозвоночных, а также презентация книги о русском протистологе Владимире Шевякове.
Теги
Сельское хозяйство
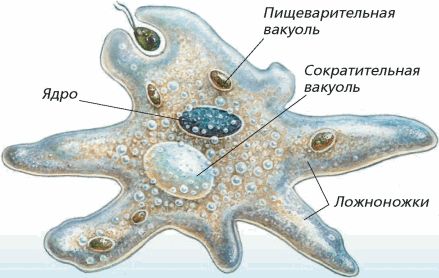
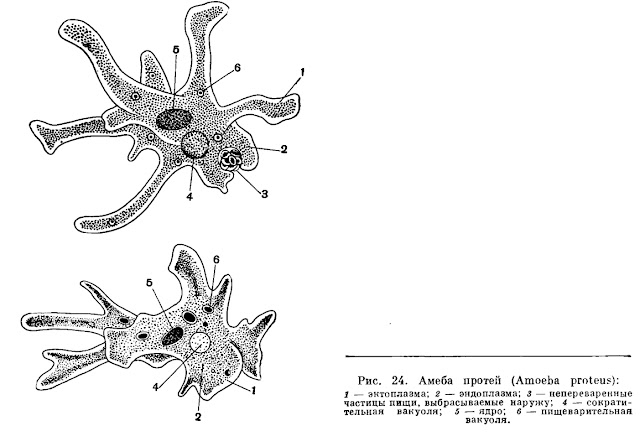
Медицинская протозоология. Ротовая амеба. Entamoeba gingivalis.
Тип: саркожгутиконосцы
Класс: саркодовые (sarcodina)
Отряд: амебы
Род: entamoeba
Вид: Ротовая амеба (Entamoeba gingivalis)
Место обитания:ротовая полость, зубной налет, крипты небных миндалин, ВДП.
Медицинское значение: это условно патогенный (факультативный паразит). При снижении местного иммунитета может провоцировать, гингивит, гласит, стоматит…
Инвазионная форма:вегетативная форма, является комменсалом.
Способ заражения:передается
контактным путем (через поцелуи).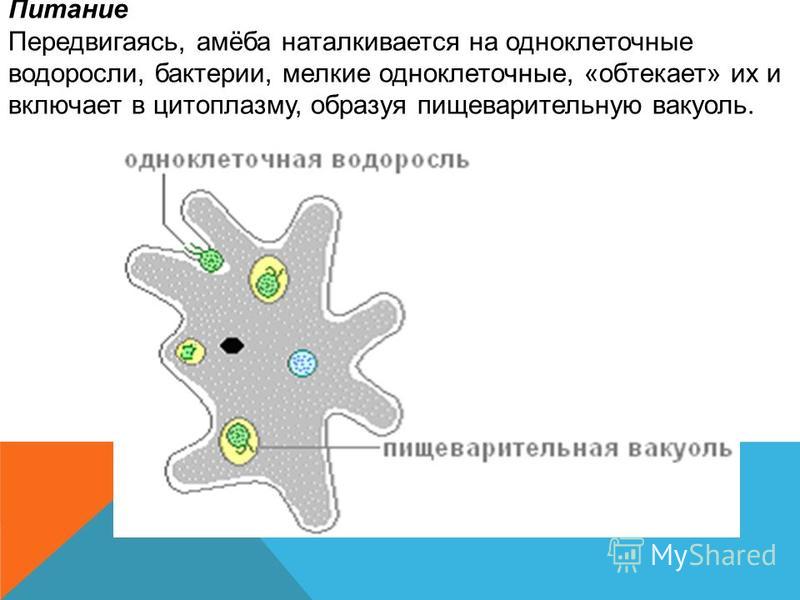 Антропогенная инвазия.
Антропогенная инвазия.
Морфология паразита:
Цитоплазма разделена на 2 слоя, содержит бактерии, лейкоциты зеленоватого цвета и эритроциты при кровоточивости ротовой полости на разных стадиях переваривания. Ядра не видно.
Жизненный цикл:Единственная форма существования – вегетативная форма. Цист не образует.
Лабораторная диагностика:микроскопия нативных мазков из соскобов ротовой полости, гноя при ГЗЛ, синуситах наNaCl0,9%.
Кишечная амеба. Entamoeba coli.
Вид: Кишечная амеба (Entamoeba coli)
Место обитания:верхний отдел толстого кишечника и нижний отдел тонкого кишечника.
Инвазионная форма:вегетативная форма, является комменсалом, факультативный паразит.
Способ заражения:фекально-оральный. Антропогенная инвазия.
Морфология паразита:разделение
цитоплазмы на 2 слоя видно у погибшей
амебы или при образовании ложноножек.
В цитоплазме имеются бактерии, грибы,
простейшие, остатки пищи. Форменных
элементов крови в цитоплазме НЕТ. Ядро
у живой амебы видно хорошо (в отличие
от e.histolytica) в виде кольцевого
образования из блестящих зерен хроматина.
Ложноножки короткие, широкие и их
несколько. Движение – топтание на месте.
Форменных
элементов крови в цитоплазме НЕТ. Ядро
у живой амебы видно хорошо (в отличие
от e.histolytica) в виде кольцевого
образования из блестящих зерен хроматина.
Ложноножки короткие, широкие и их
несколько. Движение – топтание на месте.
Жизненный цикл:обитает в толстом кишечнике, не патогенная.
Лабораторная диагностика:микроскопия мазков испражнений.
Диэнтамеба. Dientamoeba fragilis.
Тип: саркожгутиконосцы
Класс: саркодовые (sarcodina)
Отряд: амебы
Род: dientamoeba Jepps
Вид: диэнтамеба (dientamoeba fragilis)
Заболевание: диэнтамебная диарея.
Инвазионная форма:вегетативная форма, патогенная.
Способ заражения:в виду чрезвычайной неустойчивости во внешней среде в организм человека попадает с яйцами круглых червей (симбиоз с детской острицей), в которые проникает амеба на их ранних стадиях формирования.
Морфология паразита:
Маленькая. Обитает просвете толстой
к-ки и питается бактериями, грибами, и
эритроцитами.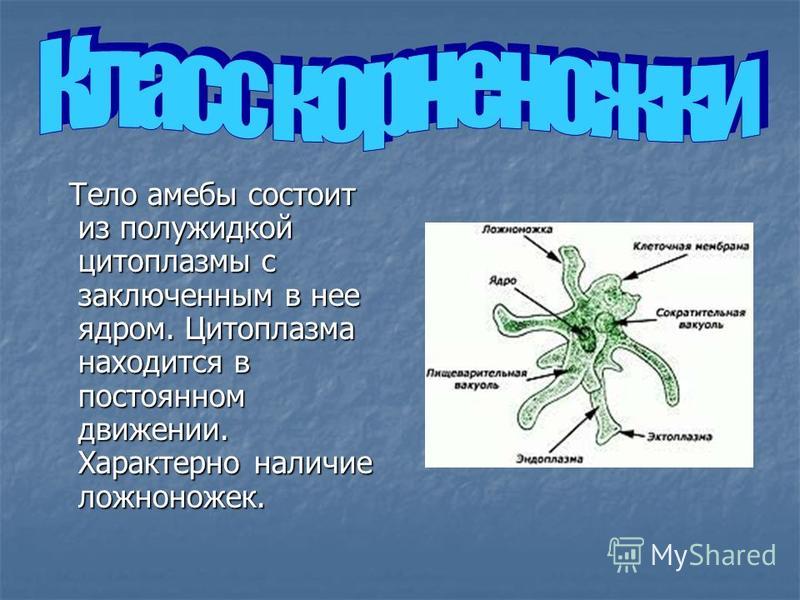 Известна только вегетативные
формы этой амебы. Эктоплазма и эндоплазма
хорошо различимы. Имеет 2 ядра (редко
3), видны только после окрашивания.
Обнаруживаются только в жидком стуле,
обычно при различных кишечных
расстройствах. Можно найти при аппендиците.
Известна только вегетативные
формы этой амебы. Эктоплазма и эндоплазма
хорошо различимы. Имеет 2 ядра (редко
3), видны только после окрашивания.
Обнаруживаются только в жидком стуле,
обычно при различных кишечных
расстройствах. Можно найти при аппендиците.
Жизненный цикл:вегетативная форма через рот с яйцами остриц попадает в толстый кишечник, где размножается и паразитирует. Цист не образует. Во внешней среде быстро гибнет.
Лабораторная диагностика:микроскопия мазков из свежих (теплых) испражнений.
Дизентерийная амеба. Entamoeba histolytica.
Тип: саркожгутиконосцы
Класс: саркодовые (sarcodina)
Отряд: амебы (amoebina)
Род: entamoeba
Вид: дизентерийная амеба (entamoeba histolytica)
Медицинское значение:амебиаз (амебная дизентерия)
Инвазионная форма:большая вегетативная и тканевая форма.
Форма заражения:зрелая4-хядерная циста.
Эпидемиология:антропогенная
инвазия. Заражение фекально-оральное.
Источник инвазии цистоносители и
больные.
Заражение фекально-оральное.
Источник инвазии цистоносители и
больные.
Морфология паразита:существует в виде нескольких форм (большая вегетативная форма, малая вегетативная, тканевая, просветная, предцистная форма и цисты).
Большая вегетативная форма: цитоплазма разделена на 2 слоя (эктоплазма – как истолченное стекло, и эндоплазма – стекловидная масса). У живой амебы ядра не видно, у мертвой оно в виде кольцевого скопления зерен. В эндоплазме содержится несколько эритроцитов. Отличается о других форм поступательным движением – толчкообразно образуется вырост эктоплазмы, в который с завихрением переливается эндоплазма.
Циста: образуется из просветной формы в толстом к-ке, неподвижные, круглые, бесцветные, иногда в них видны блестящие палочки – хроматоидные тела (РНК и белек). При окрашивании р-ром Люголя видны4 ядра.
Жизненный цикл:
Каждая циста попадают в ЖКТ, где в толстом
кишечнике дает 8 клеток, которые
превращается мелкую вегетативную форму
(не патогенная, питается бактериями и
остатками пищи). При ослаблении иммунитета
переходит в большую вегетативную форму,
которая обитает в просвете нисходящей
и сигмовидной кишки (патогенная, питается
слизистой и эритроцитами). В глубине
пораженных тканей располагается тканевая
форма амебы (патогенная, мельче
вегетативной и цитоплазме нет эритроцитов).
Обе патогенные формы переходят в
просветную форму, предцистную, а затем
в цисты (зрелые цисты – 4-х ядерные).
При ослаблении иммунитета
переходит в большую вегетативную форму,
которая обитает в просвете нисходящей
и сигмовидной кишки (патогенная, питается
слизистой и эритроцитами). В глубине
пораженных тканей располагается тканевая
форма амебы (патогенная, мельче
вегетативной и цитоплазме нет эритроцитов).
Обе патогенные формы переходят в
просветную форму, предцистную, а затем
в цисты (зрелые цисты – 4-х ядерные).
Цисты f.minuta→f.magna→ просветная форма → цисты
Патогенез.
f.magnaобитая
в просвете нижних отделов толстого
кишечника (нисходящая и сигмовидная
кишка), выделяет фермент, который
разрушает ткани (некроз слизистой) и
образование кровоточащих язв (язвенный
колит) + присоединение вторичной инфекции.
При снижении иммунитета тканевая форма
амебы проникает в кровоток (генерализация
процесса) и попадает в печень…, где
могут развиваться абсцессы, которые в
5% случаев прорываются в брюшную полость
с развитием перитонита. Которые также
развиваются при прободении (перфорации).
Клиника:
Тенезмы – ложные позывы к дефекации
Стул – малиновое желе (слизь с эритроцитами), частый водянистый.
Боль в низу живота
Симптомы интоксикации: слабость, t-субфебрильная, головная боль, тошнота.
Симптомы анемии, истощения и гиповолемии (обезвоживание)
Лабораторная диагностика:
При цистоносительстве:в оформленных или полуоформленных испражнениях можно обнаружить цисты, которые различают по размеру и числу ядер. Мазок микроскопируют с раствором Люголя.
При остром или подостром течении:из свежих жидких фекалий готовят нативный мазок и наблюдают подвижные вегетативные формы амеб с эритроцитами в цитоплазме. Испражнения исследуют в течение 10-20 мин после выделения.
Профилактика:
Личная:кипячение воды, разрыв цепи фекально-орального заражения, мытье рук, овощей фруктов, уничтожение переносчиков (тараканы, мухи).
Общественная:выявление и изоляция больных и носителей, не допустить фекального заражения окружающей среды (дезинфекция испражнений), санитарно-просветительская работа.
Культивирование и свойства Echinamoeba thethermarum n. sp., чрезвычайно теплолюбивая амеба, обитающая в горячих источниках
. 2003 г., август 7(4):267-74.
doi: 10.1007/s00792-003-0319-6. Epub 2003 28 марта.
Мануэла Баумгартнер 1 , Ahoua Yapi, Regina Gröbner-Ferreira, Karl O Stetter
принадлежность
- 1 Lehrstuhl für Mikrobiologie, Universität Regensburg, Universitätsstrasse 31,
Regensburg, Germany.
- PMID: 126
- DOI:
10.
 1007/s00792-003-0319-6
1007/s00792-003-0319-6
Мануэла Баумгартнер и др. Экстремофилы. 2003 9 августа0003
. 2003 г., август 7(4):267-74.
doi: 10.1007/s00792-003-0319-6. Epub 2003 28 марта.
Авторы
Мануэла Баумгартнер 1 , Ахуа Япи, Регина Грёбнер-Феррейра, Карл О Штеттер
принадлежность
- 1 Lehrstuhl für Mikrobiologie, Universität Regensburg, Universitätsstrasse 31,
Регенсбург, Германия.
- PMID: 126
- DOI:
10.
 1007/s00792-003-0319-6
1007/s00792-003-0319-6
Абстрактный
Здесь мы описываем новую чрезвычайно термофильную амебу, которая растет при температуре от 33°С до 57°С (Topt.=50°С). Изоляты были получены из горячих источников Аньяно-Терме (Италия), Йеллоустонского национального парка (США), Камчатки (Россия) и вулкана Ареналь (Коста-Рика). Их можно культивировать моноксеносодержащими термофильными альфа-протеобактериями. Морфологию амебы изучали с помощью микроскопа, расположенного под нагреваемым полиакрилатным колпаком. При 50°С клетки выглядели плоскими, неправильной треугольной или удлиненной формы, иногда с тонкими шиповидными субпсевдоподиями. В среднем они имели длину 22 мкм и ширину 11 мкм, имели одно ядро с центральным ядрышком. На основании морфологии и сравнения SSU рРНК амеба принадлежала к роду Echinamoeba, где она представляет новый вид. Ссылаясь на его крайне теплолюбивый образ жизни и гидротермальную среду обитания, мы назвали его E. thermarum.
thermarum.
Похожие статьи
Tetramitus thermacidophilus n. sp., амебофлагеллят из кислых горячих источников.
Баумгартнер М., Эберхардт С., Де Йонкере Дж. Ф., Стеттер К.О. Баумгартнер М. и соавт. Дж Эукариот микробиол. 2009 март-апрель;56(2):201-6. doi: 10.1111/j.1550-7408.2009.00390.x. Дж Эукариот микробиол. 2009. PMID: 21462554
Oramoeba fumarolia род. ноябрь, сп. nov., новый морской гетеролобозный амебофлагеллят, растущий при 54 °C.
Де Джонкере Дж.Ф., Баумгартнер М., Эберхардт С., Оппердоэс Ф.Р., Стеттер К.О. De Jonckheere JF и соавт. Евр Дж Протистол. 2011 Январь; 47(1):16-23. doi: 10.1016/j.ejop.2010.09.002. Epub 2010 8 октября. Евр Дж Протистол. 2011. PMID: 20
6Marinamoeba thermophila, новая морская гетеролобообразная амеба, растущая при 50°С.

Де Джонкере Дж.Ф., Баумгартнер М., Оппердоэс Ф.Р., Стеттер К.О. De Jonckheere JF и соавт. Евр Дж Протистол. 2009 авг; 45 (3): 231-6. doi: 10.1016/j.ejop.2009.01.001. Epub 2009 31 марта. Евр Дж Протистол. 2009. PMID: 1
65
Thermoanaerobacter uzonensis sp. nov., анаэробная термофильная бактерия, выделенная из горячего источника в пределах кальдеры Узон, Камчатка, Дальний Восток России.
Вагнер И.Д., Чжао В., Чжан К.Л., Романек К.С., Роде М., Вигель Дж. Вагнер ID и др. Int J Syst Evol Microbiol. 2008 ноябрь; 58 (Pt 11): 2565-73. doi: 10.1099/ijs.0.65343-0. Int J Syst Evol Microbiol. 2008. PMID: 18984694
Аноксибациллы термарум зр. nov., новая термофильная бактерия, выделенная из термальной грязи Эуганских горячих источников, Абано-Терме, Италия.

Поли А., Романо И., Корделла П., Орландо П., Николаус Б., Чески Беррини К. Поли А и др. Экстремофилы. 2009 г.Ноябрь; 13 (6): 867-74. doi: 10.1007/s00792-009-0274-y. Epub 2009 27 августа. Экстремофилы. 2009. PMID: 1
98
Посмотреть все похожие статьи
Цитируется
Обнаружение свободноживущих амеб и их внутриклеточных бактерий в скважинной воде до и после установки керамических горшковых фильтров в сельских районах Южной Африки.
Ван дер Лоо К., Барти К., Барнард Т.Г., Потгитер Н. ван дер Лоо С. и др. Общественное здравоохранение Int J Environ Res. 2021 8 апреля; 18 (8): 3912. дои: 10.3390/ijerph28083912. Общественное здравоохранение Int J Environ Res. 2021. PMID: 33
0 Бесплатная статья ЧВК.
Эукариотические организмы континентальных гидротермальных систем.

Браун С.Р., Фриц С.К. Браун С.Р. и др. Экстремофилы. 2019 июль; 23 (4): 367-376. дои: 10.1007/s00792-019-01101-й. Эпаб 2019 22 мая. Экстремофилы. 2019. PMID: 31119431 Обзор.
Экология и разнообразие микробных эукариот в геотермальных источниках.
Оливерио А.М., Пауэр Дж.Ф., Уошберн А., Кэри С.К., Стотт М.Б., Фиерер Н. Оливерио А.М. и соавт. ISME J. 2018 Aug;12(8):1918-1928. doi: 10.1038/s41396-018-0104-2. Epub 2018 16 апр. ИСМЕ Дж. 2018. PMID: 2
45 Бесплатная статья ЧВК.
Потенциально патогенные свободноживущие амебы в некоторых районах, пострадавших от наводнения во время наводнения в Чиангмае в 2011 году.
Ваннасан А., Упаранукроу П., Сонсанчун А., Моракотэ Н.
 Ваннасан А. и др.
Rev Inst Med Trop Сан-Паулу. 2013 ноябрь-декабрь; 55(6):411-6. doi: 10.1590/S0036-46652013000600007.
Rev Inst Med Trop Сан-Паулу. 2013.
PMID: 24213194
Бесплатная статья ЧВК.
Ваннасан А. и др.
Rev Inst Med Trop Сан-Паулу. 2013 ноябрь-декабрь; 55(6):411-6. doi: 10.1590/S0036-46652013000600007.
Rev Inst Med Trop Сан-Паулу. 2013.
PMID: 24213194
Бесплатная статья ЧВК.Обнаружение простейших хозяев Legionella pneumophila в инженерных системах водоснабжения с помощью пакетного теста биопленки.
Валстер Р.М., Вуллингс Б.А., ван дер Кой Д. Валстер Р.М. и соавт. Appl Environ Microbiol. 2010 ноябрь;76(21):7144-53. doi: 10.1128/AEM.00926-10. Epub 2010 17 сентября. Appl Environ Microbiol. 2010. PMID: 20851993 Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
использованная литература
- J Протозол. 1967 августа; 14 (3): 499-521 — пабмед
- Дж Эукариот микробиол.
 2002 г. май-июнь;49(3):227-38
—
пабмед
2002 г. май-июнь;49(3):227-38
—
пабмед
- Дж Эукариот микробиол.
- Протист. 2000 г., октябрь; 151 (3): 275–82. — пабмед
- J Бактериол. 1999 авг; 181 (16): 5114-8 — пабмед
- Appl Environ Microbiol. 1998 май; 64 (5): 1822-4 — пабмед
Типы публикаций
термины MeSH
вещества
Характеристика бактериальных микробиомов социальных амеб и изучение роли хозяина и окружающей среды в составе микробиома
. 2021 Январь; 23 (1): 126-142.
2021 Январь; 23 (1): 126-142.
дои: 10.1111/1462-2920.15279. Epub 2020 3 ноября.
Элени Саллинджер 1 , Майкл С Робсон 2 , Тамара С. Хаселкорн 1
Принадлежности
- 1 Факультет биологии, Университет Центрального Арканзаса, Конвей, Арканзас, 72035, США.
- 2 Кафедра биомедицинской информатики, Университет медицинских наук Арканзаса, Литл-Рок, Арканзас, 72205, США.
- PMID: 33063404
- DOI:
10.
 1111/1462-2920.15279
1111/1462-2920.15279
Элени Саллинджер и др. Окружающая среда микробиол. 2021 янв.
. 2021 Январь; 23 (1): 126-142.
дои: 10.1111/1462-2920.15279. Epub 2020 3 ноября.
Авторы
Элени Саллинджер 1 , Майкл С Робсон 2 , Тамара С. Хаселкорн 1
Принадлежности
- 1 Факультет биологии, Университет Центрального Арканзаса, Конвей, Арканзас, 72035, США.
- 2 Кафедра биомедицинской информатики, Университет медицинских наук Арканзаса, Литл-Рок, Арканзас, 72205, США.
- PMID: 33063404
- DOI: 10.1111/1462-2920.15279
Абстрактный
Как хищники бактерий, амебы отбирают черты, которые позволяют бактериям становиться симбионтами, переживая фагоцитоз и используя внутриклеточную среду эукариот. Обитающие в почве социальные амебы могут помочь нам ответить на вопросы о естественной экологии этих симбиозов амебы и бактерий в спектре патоген-мутуалист. Наша цель состояла в том, чтобы охарактеризовать естественный бактериальный микробиом филогенетически и морфологически разнообразных видов социальных амеб, используя секвенирование следующего поколения ампликонов 16S рРНК непосредственно из плодовых тел амеб. Мы обнаружили шесть типов амебоассоциированных бактерий: Proteobacteria, Bacteroidetes, Actinobacteria, Chlamydiae, Firmicutes и Acidobacteria.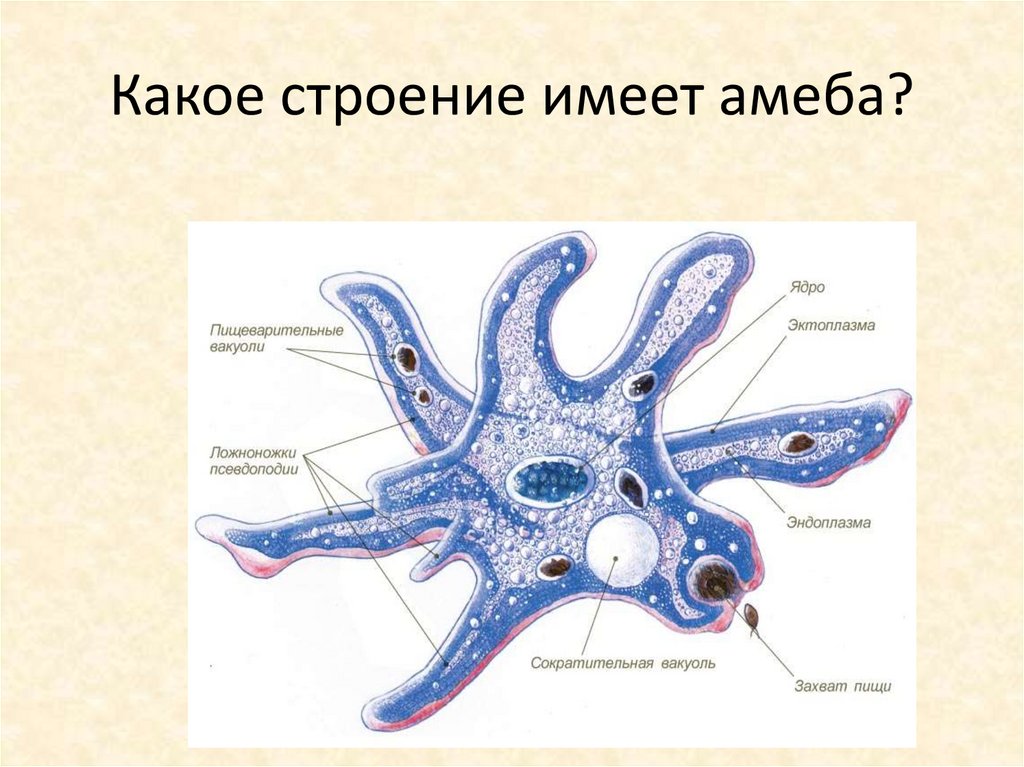 Наиболее распространенные ассоциаты амеб были отнесены к порядку Chlamydiales и роду Burkholderia-Caballeronia-Paraburkholderia. Эти бактерии присутствовали у нескольких видов амеб в разных местах. Хотя существовала значительная внутривидовая изменчивость, были некоторые свидетельства специфичности хозяина и различного количества таксонов у разных хозяев-амеб. Микробиомы амеб отличались от микробиомов среды их обитания в почве, а рН почвы влиял на разнообразие микробиома амеб. Неудивительно, что альфа-разнообразие в образцах амеб было ниже, чем в почве, но бета-разнообразие между образцами амеб было выше, чем между образцами почвы. Дальнейшее изучение микробиомов социальных амеб может помочь нам понять роль бактерий, хозяина и окружающей среды в симбиотических взаимодействиях и формировании микробиома в базальных эукариотических организмах.
Наиболее распространенные ассоциаты амеб были отнесены к порядку Chlamydiales и роду Burkholderia-Caballeronia-Paraburkholderia. Эти бактерии присутствовали у нескольких видов амеб в разных местах. Хотя существовала значительная внутривидовая изменчивость, были некоторые свидетельства специфичности хозяина и различного количества таксонов у разных хозяев-амеб. Микробиомы амеб отличались от микробиомов среды их обитания в почве, а рН почвы влиял на разнообразие микробиома амеб. Неудивительно, что альфа-разнообразие в образцах амеб было ниже, чем в почве, но бета-разнообразие между образцами амеб было выше, чем между образцами почвы. Дальнейшее изучение микробиомов социальных амеб может помочь нам понять роль бактерий, хозяина и окружающей среды в симбиотических взаимодействиях и формировании микробиома в базальных эукариотических организмах.
© 2020 Общество прикладной микробиологии и John Wiley & Sons Ltd.
Похожие статьи
Разнообразие свободноживущих экологических бактерий и их взаимодействие с бактериоядной амёбой.
Брок Д.А., Хаселкорн Т.С., Гарсия Дж.Р., Башир У., Дуглас Т.Е., Галлоуэй Дж., Броди Ф., Квеллер Д.К., Штрассманн Дж.Е. Брок Д.А. и соавт. Front Cell Infect Microbiol. 2018 23 ноября; 8:411. дои: 10.3389/fcimb.2018.00411. Электронная коллекция 2018. Front Cell Infect Microbiol. 2018. PMID: 30533398 Бесплатная статья ЧВК.
Разнообразие свободноживущих амеб в почвах и связанных с ними условно-патогенных бактерий человека.
Дене Э., Купа-Гуталан Б., Назарет С., Пеландакис М., Фавр-Бонте С. Денет Э. и др. Паразитол рез. 2017 ноябрь;116(11):3151-3162. doi: 10.1007/s00436-017-5632-6. Epub 2017 7 октября. Паразитол рез. 2017. PMID: 28988383
Специфика симбионтов Burkholderia в социальном симбиозе выращивания амеб: распространенность, виды, генетическое и фенотипическое разнообразие.

Хаселкорн Т.С., ДиСальво С., Миллер Дж.В., Башир У., Брок Д.А., Квеллер Д.К., Штрассманн Дж.Е. Haselkorn TS, et al. Мол Экол. 2019 февраля; 28 (4): 847-862. doi: 10.1111/mec.14982. Мол Экол. 2019. PMID: 30575161
Генетические и физиологические взаимодействия в амебобактериальном симбиозе.
Чон К.В. Чон КВ. Дж Эукариот микробиол. 2004 сен-октябрь; 51 (5): 502-8. doi: 10.1111/j.1550-7408.2004.tb00277.x. Дж Эукариот микробиол. 2004. PMID: 15537083 Обзор.
Экология и эволюция амебно-бактериальных взаимодействий.
Ши Ю, Куэллер Д.С., Тянь Ю, Чжан С., Ян Ц., Хе З., Хе З., Ву С., Ван С., Шу Л. Ши Ю и др. Appl Environ Microbiol. 4 января 2021 г.; 87 (2): e01866-20.
 doi: 10.1128/AEM.01866-20. Печать 2021 4 января.
Appl Environ Microbiol. 2021.
PMID: 33158887
Бесплатная статья ЧВК.
Обзор.
doi: 10.1128/AEM.01866-20. Печать 2021 4 января.
Appl Environ Microbiol. 2021.
PMID: 33158887
Бесплатная статья ЧВК.
Обзор.
Посмотреть все похожие статьи
Цитируется
Взаимодействия между несколькими царствами управляют микробиомом в подземных объектах культурного наследия.
Лю В, Чжоу С, Цзинь Т, Ли И, Ву Б, Ю Д, Ю З, Су Б, Чен Р, Фэн Ю, Дельгадо-Бакерисо М. Лю В. и др. Proc Natl Acad Sci U S A. 2022 Apr 12;119(15):e2121141119. doi: 10.1073/pnas.2121141119. Epub 2022 28 марта. Proc Natl Acad Sci U S A. 2022. PMID: 35344401
Микробная война в дикой природе — влияние протистов на эволюцию и вирулентность бактериальных патогенов.
Амаро Ф.
 , Мартин-Гонсалес А.
Амаро Ф. и др.
Интер микробиол. 2021 ноябрь;24(4):559-571. doi: 10.1007/s10123-021-00192-y. Epub 2021 8 августа.
Интер микробиол. 2021.
PMID: 34365574
Обзор.
, Мартин-Гонсалес А.
Амаро Ф. и др.
Интер микробиол. 2021 ноябрь;24(4):559-571. doi: 10.1007/s10123-021-00192-y. Epub 2021 8 августа.
Интер микробиол. 2021.
PMID: 34365574
Обзор.Новые эндосимбионты Chlamydiae и Amoebophilus преобладают в диких изолятах модельной социальной амебы Dictyostelium discoideum.
Хаселкорн Т.С., Хименес Д., Башир У., Саллинджер Э., Квеллер Д.С., Штрассманн Дж.Э., ДиСальво С. Haselkorn TS, et al. Environ Microbiol Rep. 2021 Oct;13(5):708-719. дои: 10.1111/1758-2229.12985. Epub 2021 22 июня. Представитель Environ Microbiol, 2021 г. PMID: 34159734 Бесплатная статья ЧВК.
использованная литература
- Андерсон, MJ (2001) Новый метод непараметрического многомерного дисперсионного анализа.
 Австралийский Экол 26: 32-46.
Австралийский Экол 26: 32-46.
- Андерсон, MJ (2001) Новый метод непараметрического многомерного дисперсионного анализа.
- Андерсон, М. Дж. (2006) Дистанционные тесты на однородность многомерных дисперсий. Биометрия 62: 245-253.
- Аннесли, С.Дж., и Фишер, П.Р. (2009) Dictyostelium discoideum — модель по многим причинам. Mol Cell Biochem 329: 73-91.
- Apprill, A., Mcnally, S., Parsons, R., and Weber, L. (2015) Незначительный пересмотр праймера гена SSU рРНК 806R области V4 значительно увеличивает обнаружение бактериопланктона SAR11. Акват Микроб Экол 75: 129-137.
- Бальдауф, С.Л., Ромерало, М., Физ-Паласиос, О., и Хейдари, Н.
 (2018) Глубокое скрытое разнообразие Dictyostelia. Протист 169: 64-78.
(2018) Глубокое скрытое разнообразие Dictyostelia. Протист 169: 64-78.
- Бальдауф, С.Л., Ромерало, М., Физ-Паласиос, О., и Хейдари, Н.
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- 1726009/Национальный научный фонд (NSF)
- Колледж естественных наук
- Университет Центрального Арканзаса
экологическая среда обитания и передача Escherichia coli O157:H7 | FEMS Microbiology Letters
Escherichia coli O157:H7, экология, окружающая среда, вода, выживание, свободноживущие простейшие
coli (Hayashi et al. , 2001; Perna et al. 9).0393, 2001). Остальные последовательности, специфичные для O157:H7, называются O-островками, большинство из которых перенесены горизонтально и включают другие гены вирулентности в дополнение к генам stx и LEE (Croxen & Finlay, 2010).Крупный рогатый скот признан основным резервуаром для E. coli O157:H7, вызывающей зоонозную передачу при употреблении в пищу недоваренного мяса или молочных продуктов, недостаточно пастеризованных и загрязненных бычьими фекалиями (Jay et al. , 2004; Kassenborg и др. , 2004). Здесь мы рассматриваем установленное и предполагаемое поведение E. coli O157:H7 в окружающей среде и представляем потенциальные резервуары и экологические ниши, где EHEC может сохраняться в окружающей среде.
Употребление любых продуктов питания или напитков, загрязненных навозом/фекалиями животных, может привести к заболеванию. По этой причине источники пищи, вызывающие болезнь, вторичны по отношению к Вспышки E. coli O157:H7 за последние несколько лет изменились. Интересно, что свежая зелень, фрукты и овощи стали важными источниками заражения человека. В США заражение E. coli O157:H7 от зараженных фруктов и овощей увеличилось с 11% до 41% с 1998 по 2007 год (Xicohtencatl-Cortes et al. , 2009). Загрязнение свежей продукции было связано с фекальным загрязнением сельскохозяйственных поливных вод или стоков. По той же линии недавняя энтерогеморрагическая Штамм E. coli O104:h5 был причастен к вспышке болезни в Европе весной 2011 г., и его возможный источник был связан с сырыми овощами (семена или ростки пажитника), потребляемыми в сыром или недоваренном виде (King et al. , 2012). Было трудно установить связь между транспортным средством, источником и причиной этой вспышки STEC. Это повысило важность эпидемиологических и микробиологических исследований пищевых продуктов и окружающей среды.
В течение почти двух десятилетий мировое сообщество сталкивается со вспышками не-O157 STEC, связанными с пищевыми продуктами. В результате появляется все больше научных данных, подтверждающих, что 9Штаммы 0392 E. coli , отличные от O157, широко распространены в мясных продуктах и в равной степени способны вызывать тяжелые вспышки болезней пищевого происхождения (EFSA, 2011). Hussein (2007) проанализировал зарегистрированные уровни не-O157 STEC в цельных тушах крупного рогатого скота, говяжьем фарше, говяжьих отрубах и колбасах и обнаружил 1,7–58%, 2,4–30%, 11,4–49,6% и 17–49,2% соответственно. (Хусейн, 2007). В таблице 1 отчеты о не-O157 STEC в пищевых продуктах, связанных с инфекциями человека из многих стран, показывают большие различия в оценках распространенности и в основных сообщаемых не-O157 серогруппах (Hussein & Bollinger, 2005; Hussein & Sakuma, 2005; Эриксон и Дойл, 2007 г. ; EFSA, 2012 г.).
1
Заболеваемость STEC и процент не-O157 в пищевых продуктах, связанных с инфекциями человека, в различных регионах мира
| Страна/географический район | EIR STEC | 15 STEC 15 Non-OОсновные серогруппы, отличные от O157 | Ссылки | |
| Австралия | 0,4 | c. 42 | O26, O111 | Vally и др. (2012) |
| Европейский Союз | 1.1 | c. 48 | O26, O91, O103, O111, O113, O128, O145, O146 | EFSA (2011) |
| Канада | 3.0497 | |||
| Канада | 3.0497 | |||
| Канада | 3.0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| . | Томпсон и др. (2005 г.) и Gill & Gill (2010 г. ) | |||
| Япония | 2,0–3,0 | c. 40 | О26, О111, О121, О103, О145 | Kudoh и др. (1994) и Sakuma et al. (2006) |
| Аргентина | 10,4–12,2 | c. 40 | O8, O26, O113, O145, O174 | Rivas и др. (2006) и Masana и др. (2011) |
| США | 1,04–1,2 | c. 30 | O26, O45, O103, O111, O121, O145 | Брукс и др. (2005 г.) и Скаллан и др. (2011) |
| Country/Geographic area | EIR of STEC | % of STEC Non-O157 | Major non-O157 serogroups | References |
| Australia | 0,4 | в. 42 | O26, O111 | Vally и др. (2012) |
| Европейский союз | 1. 1 | c. 48 | O26, O91, O103, O111, O113, O128, O145, O146 | EFSA (2011) |
| Canada | 3.0–6.0 | < 15 | O26, O91, O103, O111, O121 | Томпсон и др. (2005 г.) и Gill & Gill (2010 г.) |
| Япония | 2,0–3,0 | c. 40 | O26, O111, O121, O103, O145 | Kudoh и др. (1994) и Sakuma et al. (2006) |
| Аргентина | 10,4–12,2 | c. 40 | O8, O26, O113, O145, O174 | Rivas и др. (2006) и Masana и др. (2011) |
| США | 1,04–1,2 | c. 30 | O26, O45, O103, O111, O121, O145 | Брукс и др. (2005) и Scallan et al. (2011) |
EIR, оценочный уровень заболеваемости.
Основные серогруппы, отличные от O157.
Данные основаны на 25 представивших отчеты странах Европейского Союза (ЕС).
Открыть в новой вкладке
1
Заболеваемость STEC и процент не-O157 в пищевых продуктах, связанных с инфекциями человека, в различных регионах мира
| Страна/географический район | EIR STEC 5 57-O % 1 STEC 9048 | Основные серогруппы, отличные от O157 | Ссылки | |
| Австралия | 0,4 | c. 42 | O26, O111 | Vally и др. (2012) |
| Европейский союз | 1.1 | c. 48 | O26, O91, O103, O111, O113, O128, O145, O146 | EFSA (2011) |
| Канада | 3.0497 | |||
| Канада | 3.0497 | |||
| Канада | 3.0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| . | Томпсон и др. (2005 г.) и Gill & Gill (2010 г.) | |||
| Япония | 2,0–3,0 | c. 40 | O26, O111, O121, O103, O145 | Kudoh и др. (1994) и Sakuma et al. (2006) |
| Аргентина | 10,4–12,2 | c. 40 | O8, O26, O113, O145, O174 | Rivas и др. (2006) и Masana и др. (2011) |
| США | 1,04–1,2 | c. 30 | O26, O45, O103, O111, O121, O145 | Брукс и др. (2005) и Scallan et al. (2011) |
| Country/Geographic area | EIR of STEC | % of STEC Non-O157 | Major non-O157 serogroups | References |
| Australia | 0,4 | г. 42 | O26, O111 | Vally и др. (2012) |
| Европейский союз | 1. 1 | c. 48 | O26, O91, O103, O111, O113, O128, O145, O146 | EFSA (2011) |
| Канада | 3.0497 | |||
| Канада | 3.0497 | |||
| Канада | 3.0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| Канада | 3,0497 | |||
| . | Томпсон и др. (2005 г.) и Gill & Gill (2010 г.) | |||
| Япония | 2,0–3,0 | в. 40 | O26, O111, O121, O103, O145 | Kudoh и др. (1994) и Sakuma et al. (2006) |
| Аргентина | 10,4–12,2 | c. 40 | O8, O26, O113, O145, O174 | Rivas и др. (2006) и Masana и др. (2011) |
| США | 1,04–1,2 | c. 30 | O26, O45, O103, O111, O121, O145 | Брукс и др. (2005) и Scallan et al. (2011) |
EIR, оценочный уровень заболеваемости.
Основные серогруппы, отличные от O157.
Данные основаны на 25 представивших отчеты странах Европейского Союза (ЕС).
Открыть в новой вкладке
STEC не-O157 с меньшей вероятностью, чем O157 STEC, вызывают вспышки или тяжелое заболевание, и, поскольку они более сложны для диагностики, многие инфекции, не относящиеся к O157, могут быть не исследованы полностью, и поэтому их источники могут оставаться неопределенными. Частота заявлений STEC, отличных от O157, увеличилась по мере того, как лаборатории были вовлечены в тестирование этих штаммов (Gould, 2009).; Гулд и др. , 2009; Стиги и др. , 2012). Возможно, возникновение вспышек, не связанных с O157, связано с устойчивостью окружающей среды (Bolton et al. , 2011).
Escherichia coli O157:H7 Сообщалось о вспышках, связанных с потреблением зараженной воды или использованием поверхностных вод в рекреационных целях (Licence et al. , 2001; Olsen et al. , 2002; Bruneau и др. , 2004). В 1999 году люди заболели после употребления зараженной воды в округе Вашингтон, штат Нью-Йорк, и купания в загрязненной воде в округе Кларк, штат Вашингтон. Вспышка в Уолкертоне, Канада (май 2000 г.), связанная с потреблением питьевой воды, загрязненной фекалиями, вызвала 2300 случаев заболевания (Hrudey 9).0392 и др. , 2003). Escherichia coli O157:H7 и Campylobacter jejuni были идентифицированы как основные патогены, ответственные за эти случаи заболевания, а E. coli O157:H7 был причиной семи смертей (Hrudey et al. , 2003). Одним из факторов, объясняющих это загрязнение, является влияние климата. Действительно, поверхностные водоемы могут быть заражены E. coli O157:H7 после проливных дождей или таяния снега, что приводит к переполнению канализации или попаданию экскрементов животных и навоза в поверхностные воды (Bruce и др. , 2003). Кроме того, сообщалось о некоторых случаях заражения человека человеком из-за присутствия E. coli O157:H7 в воде, такой как общественные бассейны или озера (Bruce et al. , 2003; Varma et al. , 2003). Люди с диареей, особенно дети после сброса или смены грязных подгузников, могут загрязнять рекреационные воды, и заражение может произойти при купании и/или глотании (Williams et al. , 1997; Bruce et al. , 2003; Varma 9).0392 и др. , 2003; Ли и Грейг, 2008 г.). Были задокументированы вспышки, связанные со штаммами STEC, отличными от O157, через воду и продукты (Doyle & Kaspar, 2010). Сообщалось, что некоторые штаммы не-O157 STEC выживают в необработанной колодезной воде в течение нескольких месяцев (Watterworth et al. , 2006). Стойкость может быть недооценена и может быть сравнима с O157.
Escherichia coli из фекалий домашнего скота, как известно, сохраняется на травяных пастбищах не менее 5 месяцев, что дает возможность E. coli O157:H7 повторно использоваться животными (Avery и др. , 2004). Кроме того, непосредственная среда обитания животного, кормовая и питьевая вода являются важными источниками инфекции E. coli O157:H7 крупного рогатого скота (рассмотрено в Fairbrother & Nadeau, 2006). Факторами риска носительства и заражения крупного рогатого скота являются возраст, отъем, транспортировка, сезон и состав корма, а также способность бактерий сохраняться в условиях фермы в течение нескольких месяцев (Fairbrother & Nadeau, 2006). Таким образом, E. coli O157:H7 представляет собой недооцененный экологический риск.
Присутствие E. coli O157:H7 в водной среде является общим знаменателем, связывающим различные временные среды обитания и передачу инфекции животным и людям. Поэтому представляется важным понять водную экологию Е. coli O157:H7. В этом разделе освещается адаптация E. coli O157:H7 в водной среде и анализируется ее выживание и рост в состоянии сидячей биопленки и неблагоприятных условиях, которые поддерживают жизнеспособное, но некультивируемое состояние (VBNC) и свободные планктонные клетки в воде, которые могут способствовать диссеминации. .
, 1989; Martin, 2004). Наконец, бактерия переходит в состояние выживания, что повышает ее устойчивость ко многим стрессам и способность сохранять жизнеспособность в течение длительного времени без питательных веществ (Siegele & Kolter, 1992). Когда E. coli O157:H7 испытывает голодание или стресс, клетки вступают в фазу общей реакции на стресс (Peterson et al. , 2005). В этот момент продукция RpoS управляет транскрипцией ряда перекрывающихся сетей генов, ответственных за общий стрессовый ответ E. coli (Hengge-Aronis, 2002). RpoS конкурирует с сигма-факторами «домашнего хозяйства», направляя основную РНК-полимеразу на транскрипцию специфических подмножеств генов, включая метаболизм стресса, чтобы подготовить бактерии к сопротивлению множественным стрессам окружающей среды, включая голодание (Lange & Hengge-Aronis, 19).91). Высокая осмолярность, колебания температуры, низкий рН и низкая скорость роста также вызывают RpoS-ответ в клетках E. coli (Bearson et al. , 1996; Muffler et al. , 1996, 1997; Ihssen & Egli). , 2004). Было обнаружено, что штамм Escherichia coli O157:H7 более устойчив к кислоте, чем родовой штамм E. coli (Diez-Gonzalez & Russell, 1997). Кислотоустойчивость варьирует среди штаммов (Lin et al. , 1995). О и др. (2009) сообщил о значительно более высокой толерантности к уксусной кислоте E. coli O157:H7 из источников окружающей среды, включая воду и фекалии крупного рогатого скота, по сравнению со штаммами, связанными со вспышкой среди людей. Кроме того, было показано, что делеция E. coli O157:H7 rpoS нарушала экспрессию генов, ответственных за реакцию на стресс, включая gadA (часть системы 2 глутамат-зависимой кислотоустойчивости) и ler (LEE- закодированный регулятор; Dong & Schellhorn, 2009). Важность rpoS в водной среде также подтверждается снижением выживаемости Мутант E. coli rpoS в стационарной фазе в морской воде (Rozen & Belkin, 2001).
Рост и выживание E. coli в открытой среде часто ограничены наличием питательных веществ и источников энергии. Однако в поверхностных водах жизнеспособные E. coli O157:H7 были обнаружены в течение 2-месячного периода, несмотря на снижение количества клеток (Avery et al. , 2008). Что касается E. coli O157:H7, динамика экспрессии генов во временной среде обитания, такой как поверхностные воды, остается недостаточно изученной. Недавно было показано, что даже при населении в E. coli O157:H7 снижалась, некоторые клетки выживали в стерильной речной воде до 234 дней (Duffitt et al. , 2011). В этом исследовании E. coli O157:H7 в естественной стерильной воде запускали метаболизм реакции на стресс и механизмы репарации ДНК, указывая на то, что бактерии оставались активными. Однако не сообщалось об изменениях экспрессии генов вирулентности. Напротив, в другом исследовании было обнаружено, что реакция экспрессии генов E. coli O157:H7 на переход роста в среду с минимальным содержанием глюкозы запускала экспрессию генов, расположенных на островках патогенности и токсин-конвертирующих бактериофагах (Bergholz 9).0392 и др. , 2007). Возможно, что экспрессия генов может значительно различаться между адаптациями к росту в среде с минимальным содержанием глюкозы и выживанием в воде.
Фосфат является очень востребованным ресурсом. После использования он часто является ограничивающим питательным веществом в окружающей среде, и его доступность может определять скорость роста организмов. Обычно это относится к пресноводным средам (Doering et al. , 1995; Correll, 1999; Paytan & McLaughlin, 2007). В большинстве сред, когда доступность неорганического фосфата (Pi) становится ограниченной (< 4 мкМ), активируется форегулон (VanBogelen 9).0392 и др. , 1996; Ламарш и др. , 2008). Такая глобальная система регуляции обеспечивает оптимальную адаптивную реакцию и эффективное использование фосфата в условиях ограничения Pi, что может привести к выживанию патогенных E. coli в условиях ограничения фосфатов. Помимо того, что он играет важную роль в вирулентности, Pho-регулон, таким образом, может также способствовать персистенции патогенной E. coli в окружающей среде (Crepin et al. , 2011). Совсем недавно Йошида и др. идентифицировал новые гены Pho-regulon в специфических островках O157, которые не локализованы в области позвоночника, общей с комменсальной E. coli . Они показали, что некоторые из этих генов не связаны с метаболизмом или использованием Pi. Это говорит о том, что в ответ на стресс Pi окружающей среды Pho-регулон регулирует гены, участвующие не только в гомеостазе Pi, но и в других функциях E. coli O157:H7 (Yoshida et al. , 2012).
, 2009; Berger и др. , 2010; Saldana и др. , 2011). Рост E. coli O157:H7 в защищенных биопленках оказался большим преимуществом в открытой среде. В различных средах обитания бактерии в биопленках особенно устойчивы к бактериофагам и свободноживущим амебоидным хищникам (Costerton 9).0392 и др. , 1995). Escherichia coli O157:H7 представляет собой устойчивое загрязнение как в промышленном секторе, так и на протяжении всего его экологического цикла (рис. 1). Выделение E. coli O157:H7 колеблется от 10 2 до 10 5 КОЕ г -1 фекалий крупного рогатого скота (Campbell et al. , 2001) и там, где они могут сохраняться и перерабатываться. в сельскохозяйственной среде, почве и воде (Mead & Griffin, 1998; McGee и др. 9039).3, 2002). Независимо от того, является ли водная среда обитания олиготрофной поверхностной или подземной водой, ее следует рассматривать как экологический источник E. coli O157:H7. Учитывая все это, экологические штаммы E. coli O157:H7 в своих планктонных формах могут присутствовать в различных экосистемах, и когда питательные условия становятся благоприятными, фенотипическая гибкость позволяет им образовывать биопленки. Кроме того, в настоящее время установлено, что биопленочный способ роста преобладает в водных экосистемах, поскольку было показано, что планктонные популяции составляют <0,1% от общего микробного сообщества.
Открыть в новой вкладкеСкачать слайд
Иллюстрация, изображающая экологическую среду обитания и передачу Escherichia coli O157:H7 в глобальной экосистеме. (1) Крупный рогатый скот на ферме является основным резервуаром для E. coli O157:H7 и может загрязнять окружающую среду (откормочную площадку) и сточные воды навозом крупного рогатого скота. Потребление зараженных продуктов животного происхождения (мяса и молочных продуктов) является основным путем передачи вируса человеку. Кроме того, E. coli O157:H7 фекалии животных (10 2 –10 5 КОЕ г –1 ), при использовании навоза, может загрязнять растения, что также позволяет патогену попасть в пищевую цепь. (2) Вода может быть загрязнена непосредственной средой обитания животных и навозом, оставленным на земле, иногда после сильного дождя. Кроме того, почва, загрязненная сточными водами, и переполнение канализационных коллекторов являются причинами загрязнения воды. Этот путь может затем привести к заражению растений. Кроме того, почвы и вода, содержащие свободноживущие простейшие, могут служить переносчиками Е. coli O157:H7. (3) Загрязненные поверхностные водоемы являются источником заражения человека в результате рекреационной деятельности на воде, а подземные воды являются источником сырой воды для потребления человеком. (4) Еще один потенциальный путь заражения E. coli O157:H7 от человека к человеку – фекально-оральный путь передачи. Передача от человека к человеку может происходить прямо или косвенно через загрязнение воды инфицированными людьми в рекреационных водах.
Хотя до сих пор нет четкой связи между образованием биопленки и присутствием или выживанием E. coli O157:H7 в воде, некоторые исследования показали, что E. coli O157:H7 сохраняется в воде, полученной из донных отложений (Czajkowska et al. , 2004). Интересно, что было показано, что при выращивании в биопленке E. coli O157:H7 увеличивали свое удержание и выживаемость в сточных водах через систему песчаного водоносного горизонта (Wang et al. , 2011). Кроме того, на бойнях режим биопленки повышает стойкость и кислотоустойчивость E. coli O157:H7 в жидких мясных отходах (Skandamis et al. , 2009). Кроме того, E. coli O157:H7 сохраняется в почве вокруг ферм и животноводческих ферм и может противостоять фумигации. Было высказано предположение, что химическая фумигация, уменьшающая микробное разнообразие, будет благоприятствовать E. coli O157:H7 (Ibekwe & Ma, 2011). Таким образом, микробное видовое разнообразие участвует в охране окружающей среды E. coli O157:H7.
На этих влажных или сухих поверхностях биопленки могли бы обеспечить идеальную микросреду для установления синтрофических отношений, в которых E. coli O157:H7 будет зависеть от других популяций бактерий в использовании специфических субстратов, как правило, для производства энергии. Фактически было показано, что штаммы E. coli O157:H7, не образующие биопленки, сохраняются на твердых поверхностях, связанных с биопленками, образованными штаммами-компаньонами (Uhlich et al. , 2010).
Состояние VBNC может быть вызвано стрессовыми условиями в поверхностных водах, вызванными низкой температурой или токсичными металлами (Klein & Alexander, 1986). Однако наличие состояния VBNC у энтеробактерий весьма оспаривается в некоторых сообщениях, в то время как другие предполагают, что оно действительно происходит в E. coli O157:H7, содержащихся в воде и в солевых условиях или в навозе и навозной жиже крупного рогатого скота (Bogosian et и др. , 1998, Wang & Doyle, 1998, Макино и др. , 2000, Семенов и др. , 2007, 2009). Эти результаты показали значительно большее количество микроорганизмов при прямом микроскопическом подсчете по сравнению с посевом на селективную среду, что указывало на преобладание спящих клеток в общей популяции E. coli O157. Использование биолюминесценции, такой как маркерная система люкс , которая показывает энергетический статус клетки, обеспечивает альтернативный способ оценки жизнеспособности бактерий, включая клетки VBNC (Ritchie et al. 9).0393, 2003).
Интересно, что разработаны новые методы для обнаружения E. coli O157:H7 в водной среде. Использование микрофлюидного чипа, модифицированного антителами против O157, позволяет специфически обогащать E. coli O157, включая VBNC (Dharmasiri et al. , 2010). Другой метод включает концентрирование бактериальных клеток фильтрованием проб воды на мембране с низким связыванием белков (поливинилидендифторидная гидрофильная мембрана) с последующим прямым выделением тотальной РНК и специфической амплификации методом ОТ-ПЦР в течение rfbE для антигена O157 и fliC для флагеллина H7, а затем обнаружение с помощью электронного микрочипа E. coli O157:H7 (Liu et al. , 2008).
Помимо улучшения выживания в окружающей среде, совместное проживание бактерий с простейшими может вызывать адаптивные изменения у бактерий (King et al. , 1988; Barker et al. , 1993). Кроме того, мы недавно наблюдали, что совместное культивирование E. coli O157:H7 с A. castellanii повышало устойчивость бактерий в течение 3 недель (Chekabab et al. , 2012). Кроме того, наблюдалась временная интернализация и внутриклеточная выживаемость, которые увеличивались у изогенных мутантов, не продуцирующих Stx. Каррутерс и др. (2010) показали повышенную экспрессию экспрессии генов вирулентности, таких как гены, кодирующие эффекторы Stx, LEE и не-LEE T3SS, когда E . coli O157 совместно культивировали с A. castellanii . Это говорит о том, что факторы вирулентности EHEC могут способствовать сохранению и выживанию при взаимодействии с амебами. Также наблюдалась способность E. coli O157:H7 проникать и инвазировать клетки млекопитающих, такие как клетки бычьей молочной железы, а также макрофаги человека (Matthews 9).0392 и др. , 1997; Пуарье и др. , 2008 г.; Этьен-Месмин и др. , 2011). Более того, заметно, что амебы и макрофаги человека имеют морфологическое и функциональное сходство, особенно в их фагоцитарной активности и параллельных механизмах их взаимодействия со многими бактериальными патогенами (Yan et al. , 2004; Siddiqui & Khan, 2012). Следовательно, амебы были предложены в качестве ключевого шага в эволюции бактерий окружающей среды, чтобы стать патогенами человека. Таким образом, Acanthamoeba может служить полезной моделью для изучения патогенеза EHEC и понимания механизмов уклонения от иммунного ответа.
Lainhart и др. (2009) обнаружили, что бактерии, кодирующие Stx, убивают простейших инфузорий, используя голотоксин Stx в качестве оружия против хищников. Другие исследования показали, что присутствие профага, кодирующего Stx, увеличивало приспособленность E. coli при совместном культивировании с реснитчатыми простейшими (Steinberg & Levin, 2007). Эти авторы обнаружили, что соотношение бактерий Stx+ и Stx- увеличилось после 3-дневного совместного культивирования с Tetrahymena thermophila , которые относятся к простейшим инфузориям, присутствующим в кишечнике жвачных животных. Напротив, другие исследователи не обнаружили каких-либо преимуществ или недостатков лизогенного фага Stx в отношении простейших рубца (Burow et al. , 2005). Таким образом, вклад Stx в выживание бактерий при столкновении с простейшими, по-видимому, варьируется в зависимости от условий заражения и моделей простейших, используемых в анализе совместного культивирования.
Растет озабоченность по поводу выживания патогенов в сточных водах и сточных водах, поскольку известно, что Е . coli O157:H7 может выживать в навозной жиже крупного рогатого скота в течение как минимум нескольких недель (LeJeune et al. , 2001; Lee et al. , 2009). Простейшие, присутствующие в этих сточных водах, могут обеспечить защитную нишу для таких патогенов, как E . coli O157:H7 (King и др. , 1988). Роль простейших в выживании E . coli O157:H7 в природной среде изучено меньше. Почвы, загрязненные органическими веществами и сточными водами, содержат значительно повышенное количество простейших, таких как Acantamoebae (Родригес-Сарагоса, 1994). Возможно, что E . coli O157:H7 в почве и навозной жиже могли стать жертвами свободноживущих простейших, которые затем служили переносчиками для распространения этого патогена. Это особенно верно, если бактерии способны выживать в цистах (резистентные формы амебы), как это было показано для Legionella pneumophila , Vibrio cholerae и Mycobacterium avium (Steinert et al. , 19).98; Браун и Баркер, 1999). Бактерии внутри цист амебы могут распространяться воздушно-капельным путем. Выпасаемый крупный рогатый скот будет заглатывать простейшие в силосе и траве, и проглоченные простейшие, содержащие бактериальные патогены, такие как E. coli O157, также могут быть путем передачи крупному рогатому скоту.
Различные виды, принадлежащие к трем группам простейших (жгутиковые, инфузории и амебы), были выделены из свежих зеленых продуктов, найденных в супермаркете (например, шпината и салата). Гурабатини и др. (2008) продемонстрировали, что эти простейшие могут поглощать бактериальные патогены, включая E. coli O157:H7 и S. Enterica, а затем образовывать везикулы, содержащие интактные бактерии. Таким образом, присутствие простейших на листовых овощах и секвестрация ими кишечных бактерий в везикулах указывает на то, что они могут играть важную роль в экологии E. coli O157:H7 на свежих зеленых продуктах.
Сбор информации и методы отбора проб должны быть стандартизированы для более четкого понимания экологии этого экологического патогена. Это может привести к рациональной разработке профилактических мер по ограничению его присутствия в окружающей среде, что может привести к увеличению риска заражения воды, почвы и животных, а также передачи и заражения людей.631″> Каталожные номера
Allison
S.E.
Silphaduang
U.
Mascarenhas
M.
Konczy
P.
Quan
Q.
Karmali
M.
COOMBES
B.K.K.K.K.K.K.K.K.K.
(
2012
)
Новый репрессор подвижности Escherichia coli O157:H7, закодированный в предполагаемом фимбриальном кластере OI-1
.
J Бактериол
194
:
5343
–
5352
.
Алсам
С.
Чон
С.Р.
Сиссонс
Дж.
Дадли
Р.
Ким
К.С.
Хан
Н/Д
(
2006
)
Escherichia coli взаимодействие с Acanthamoeba : симбиоз с экологическими и клиническими последствиями
.
J Med Microbiol
55
:
689
–
694
.
Эйвери
С.М.
Мур
А.
Хатчисон
М.Л.
(
2004
)
Судьба штамма Escherichia coli , происходящего из фекалий домашнего скота, попадающих непосредственно на пастбище
.
Lett Appl Microbiol
38
:
355
–
359
.
Эйвери
Л.М.
Уильямс
А.П.
(
2008
)
Выживаемость Escherichia coli O157:H7 в водах озер, рек, луж и поилок для животных
.
Sci Total Environ
389
:
378
–
385
. Т.К.
Джаяраман
А.
(
2008
)
Временная регуляция энтерогеморрагической Escherichia coli вирулентности, опосредованной аутоиндуктором-2
.
Appl Microbiol Biotechnol
78
:
811
–
819
.
Баркер
Дж.
Браун
М. Р.
(
1994
)
Троянские кони микробного мира: простейшие и выживание бактериальных возбудителей в окружающей среде
.
Микробиология
140
(
):
1253
–
1259
.
Баркер
Дж.
Ламберт
П.А.
Коричневый
М.Р.
(
1993
)
Влияние внутриамебных и других условий роста на свойства поверхности Legionella pneumophila
.
Infect Immun
61
:
3503
–
3510
.
Баркер
Дж.
Хамфри
Т.Дж.
Коричневый
МВт
(
1999
)
Выживание Escherichia coli O157 в почвенных простейших: значение для болезни
.
FEMS Microbiol Lett
173
:
291
–
295
.
Беарсон
С.М.
Бенджамин
В.Х.
младший
Мечи
З.Е.
Фостер
Дж.В.
(
1996
)
Кислотный шок Индукция RpoS опосредована мышиным геном вирулентности mviA Salmonella typhimurium
.
J Бактериол
178
:
2572
–
2579
.
Berger
C.N.
Содха
С.В.
Шоу
Р.К.
Гриффин
вечера
Розовый
D.
Рука
П.
Франкель
Г.
(
2010
)
Свежие фрукты и овощи как средства передачи патогенов человека
.
Environ Microbiol
12
:
2385
–
2397
.
Бергхольц
Т.М.
Фитиль
Л.М.
Риордан
Дж.Т.
Уэллетт
LM
Уиттам
Т.С.
.
БМС Микробиол
7
:
97
.
Beutin
L.
Geier
D.
Steinruck
H.
Zimmermann
S.
Scheutz 9003
Ф.
(
1993
)
Распространенность и некоторые свойства веротоксина (шигаподобного токсина) Escherichia coli у семи различных видов здоровых домашних животных
.
J Clin Microbiol
31
:
2483
–
2488
.
Богосян
Г.
Моррис
П.Дж.
О’Нил
Дж.П.
(
1998
)
Метод выделения смешанной культуры указывает на то, что кишечные бактерии не переходят в жизнеспособное, но не культивируемое состояние
.
Appl Environ Microbiol
64
:
1736
–
1742
.
Болтон
Д.Дж.
Монаган
А.
Бирн
Б.
Фаннинг
С.
Суини
Т.
Макдауэлл
Д.А.
.
J Appl Microbiol
111
:
484
–
490
.
Брукс
Дж.Т.
Сеятели
Е.Г.
Уэллс
Дж.Г.
Грин
К.Д.
Гриффин
вечера
Хукстра
Р.М.
Строкбайн
Н/Д
(
2005
)
Не-O157 Шига-токсин-продуцирующий Escherichia coli инфекции в США, 1983–2002
.
J Infect Dis
192
:
1422
–
1429
.
Браун
М.Р.
Баркер
Дж.
(
1999
)
Неисследованные резервуары патогенных бактерий: простейшие и биопленки
.
Trends Microbiol
7
:
46
–
50
.
Брюс
М.Г.
Кертис
М.Б.
Пейн
М.М.
Gautom
Р.К.
Thompson
E.C.
Bennett
A.L.
Кобаяши
Дж. М.
(
2003
)
Озерная вспышка Escherichia coli O157:H7 в округе Кларк, штат Вашингтон, август 1999 г.
.
Arch Pediatr Adolesc Med
157
:
1016
–
1021
.
Брюно
А.
Родриг
Х.
Исмаэль
Дж.
Дион
Р.
Аллард
Р.
(
2004
)
Вспышка E. coli O157:H7, связанная с купанием на общественном пляже в районе Монреаль-Центр
.
Can Commun Dis Rep
30
:
133
–
136
.
Берджесс
Г.
(
1998
)
Бактерии в олиготрофной среде: образ жизни выживания в условиях голодания
.
World J Microbiol Biotechnol
14
:
305
.
Бероу
Л.К.
Гобиус
К.С.
Ванзелоу
Б.А.
Клиев
А.В.
.
Lett Appl Microbiol
40
:
117
–
122
.
Кэмпбелл
Г.Р.
Проссер
Дж.
Гловер
А.
Киллхэм
К.
(
2001
)
Обнаружение Escherichia coli O157:H7 в почве и воде с помощью мультиплексной ПЦР
.
J Appl Microbiol
91
:
1004
–
1010
.
Carruthers
M.D.
Bellaire
B.H.
Миньон
Ф.К.
.
FEMS Microbiol Lett
312
:
15
–
23
.
Чекабаб
С.М.
Daigle
F.
Charette
S.J.
Дозуа
C.M.
Харел
Дж.
.
МикробиологияОткрытый
1
:
427
–
437
.
Коррелл
Д.Л.
(
1999
)
Фосфор: питательное вещество, ограничивающее норму содержания в поверхностных водах
.
Poult sci
78
:
674
–
682
.
Костертон
Дж.В.
Левандовски
Z.
Колдуэлл
D.E.
Корбер
Д.Р.
Лаппин-Скотт
Х.М.
(
1995
)
Микробные биопленки
.
Annu Rev Microbiol
49
:
711
–
745
.
Крепен
С.
Чекабаб
С.М.
Ле Биан
Г.
Бертран
Н.
Дозуа
К.М.
Харел
Дж.
(
2011
)
Форегулон и патогенез Escherichia coli
.
Вет Микробиол
153
:
82
–
88
.
Croxen
M.A.
Finlay
B.B.
(
2010
)
Молекулярные механизмы Escherichia coli патогенность
.
Nat Rev Microbiol
8
:
26
–
38
.
Czajkowska
D.
Witkowska-Gwiazdowska
A.
Sikorska
I.
Boszcczyk-Malesak
H.
6
H.
6
. (
2004
)
Выживаемость Escherichia coli серотипа O157:H7 в воде и донных отложениях
.
Pol J Environ Stud
14
:
423
–
430
.
Деванти
Р.
Вонг
А.С.
(
1995
)
Влияние условий культивирования на образование биопленки Escherichia coli O157:H7
.
Int J Food Microbiol
26
:
147
–
164
.
Дхармасири
У.
Витек
М.А.
Адамс
А.А.
Осири
Дж.К.
Хьюпер
М.Л.
Бьянки
Т.С.
Рёлке
Д.Л.
Сопер
С.А.
(
2010
)
Обогащение и обнаружение Escherichia coli O157:H7 из проб воды с использованием модифицированного антителами микрожидкостного чипа
.
Анальная химия
82
:
2844
–
2849
.
Диес-Гонсалес
Ф.
Рассел
Дж. Б.
(
1997
)
Способность Escherichia coli O157:H7 снижать внутриклеточный pH и противостоять токсичности уксусной кислоты
.
Микробиология
143
(
):
1175
–
1180
.
Doering
P.
Oviatt
C.
Nowicki
B.
KLOS
E.
Reed
L.
(
1995
)
Ограничение первичной продукции по фосфору и азоту в моделируемом эстуарном градиенте
.
Mar Ecol Progr Ser
124
:
271
–
287
.
Донг
Т.
Шеллхорн
Х.Е.
.
BMC Genomics
10
:
349
.
Дойл
E.
Каспар
C.
(
2010
)
Белая книга о шига-токсине, отличном от O157:H7, продуцирующем E. coli из мясных и не мясных источников
.
Научно-исследовательский институт пищевых продуктов Университета Висконсин-Мэдисон
.
Даффит
А.Д.
Ребер
Р.Т.
Уиппл
А.
Чауре
С.
(
2011
)
Экспрессия генов во время выживания Escherichia coli O157:H7 в почве и воде
.
Int J Microbiol
2011
:
340506
.
Дурис
Дж.В.
Хаак
С.К.
Фогарти
Л. Р.
(
2009
)
Генные и антигенные маркеры шигатоксина-продуцента E. coli из речных вод Мичигана и Индианы: встречаемость и связь с критериями качества рекреационной воды
.
J Environ Qual
38
:
1878
–
1886
.
ЕФСА
(
2011
)
Европейский центр профилактики и контроля заболеваний публикует Ежегодный эпидемиологический отчет за 2011 год
.
Евронадзор: ECDC
.
EFSA
,
Парма, Италия
.
ЕФСА
(
2012
)
Краткий отчет Европейского Союза о тенденциях и источниках зоонозов, зоонозных агентов и вспышек болезней пищевого происхождения в 2010 г.
.
Евронадзор: ECDC
.
EFSA
.
Эриксон
М.К.
Дойл
М.П.
(
2007
)
Пищевые продукты как средство передачи шига-токсин-продуцирующих Escherichia coli
.
J Food Prot
70
:
2426
–
2449
.
Etienne-Mesmin
L.
Chassaing
B.
Sauvanet
P.
Denizot
J.
Blanquet-Diot
S.
Darfeuille-Michaud
A.
Pradel
Н.
Ливрелли
В.
(
2011
)
Взаимодействия с М — клетками и макрофагами как ключевые этапы патогенеза энтерогеморрагических Escherichia coli инфекций
.
PLoS ONE
6
:
e23594
.
Fairbrother
J.M.
Nadeau
E.
(
2006
)
Escherichia coli : внутрихозяйственное заражение животных
.
Rev Sci Tech
25
:
555
–
569
.
Поля
Б.С.
(
1996
)
Молекулярная экология легионелл
.
Trends Microbiol
4
:
286
–
290
.
Гилл
А.
Гилл
С.О.
(
2010
)
Не-O157 веротоксигенный Escherichia coli и говядина: канадская перспектива
.
Can J Vet Res
74
:
161
–
169
.
Гулд
Левая сторона
(
2009
)
Обновленная информация об эпидемиологии шига-токсин-продуцирующей E. coli в США
.
Ежегодное собрание Ассоциации защиты пищевых продуктов столичного региона
,
Вашингтон, округ Колумбия
.
Gould
L.H.
и другие.
. .
MMWR Recomm Rep
58
(
RR12
):
1
–
14
.
Гурабатини
П.
Брандл
М.Т.
Реддинг
К.С.
и другие. . (
2008
)
Взаимодействие между пищевыми патогенами и простейшими, выделенными из салата-латука и шпината
.
Appl Environ Microbiol
74
:
2518
–
2525
.
Гриффин
вечера
Tauxe
Р.В.
(
1991
)
Эпидемиология инфекций, вызванных Escherichia coli O157:H7, другими энтерогеморрагическими E. coli и ассоциированным гемолитико-уремическим синдромом
.
Epidemiol Rev
13
:
60
–
98
.
Хэнкок
Д.
Бессер
Т.
Лежен
Дж.
Дэвис
М.
Райс
Д.
(
2001
)
Контроль ВТЭО в животноводческом резервуаре
.
Int J Food Microbiol
66
:
71
–
78
.
Хаяши
Т.
Макино
К.
Ониши
М.
и другие. . (
2001
)
Полная последовательность генома энтерогеморрагической Escherichia coli O157:H7 и сравнение генома с лабораторным штаммом К-12
.
ДНК Res
8
:
11
–
22
.
Hengge-Aronis
Р.
(
2002
)
Последние сведения об общей сети регулирования реакции на стресс в Кишечная палочка
.
Дж Мол Микробиол Биотехнолог
4
:
341
–
346
.
Hrudey
S.E.
Оплата
П.
Гек
P.M.
Гиллхэм
Р.В.
Хруди
Э.Дж.
(
2003
)
Эпидемия смертельных болезней, передающихся через воду, в Уолкертоне, Онтарио: сравнение с другими вспышками передающихся через воду в развитых странах
.
Водные технологии
47
:
7
–
14
.
Хусейн
Х.С.
.
J Anim Sci
85
:
E63
–
E72
.
Хусейн
Х.С.
Боллинджер
LM
.
J Food Prot
68
:
2224
–
2241
.
Хусейн
Х.С.
Сакума
Т.
(
2005
)
Шига-токсин-продуцирующий Escherichia coli : меры до- и послеуборочного контроля для обеспечения безопасности молочных продуктов крупного рогатого скота
.
J Food Prot
68
:
199
–
207
.
Huws
S.A.
Morley
R.J.
Джонс
М.В.
Браун
М.Р.
Смит
А.В.
(
2008
)
Взаимодействие некоторых распространенных патогенных бактерий с Acanthamoeba polyphaga
.
FEMS Microbiol Lett
282
:
258
–
265
.
Ибекве
A.M.
(
2011
)
Влияние фумигантов на микробное разнообразие и устойчивость E. coli O15:H7 в контрастных почвенных микрокосмах
.
Sci Total Environ
409
:
3740
–
3748
.
Ихсен
Дж.
Эгли
Т.
(
2004
)
Конкретная скорость роста, а не плотность клеток определяют общую реакцию на стресс у Escherichia coli
.
Микробиология
150
:
1637
–
1648
.
Itoh
Y.
Sugita-Konishi
Y.
Kasuga
F.
Iwaki
M.
Hara-Kudo
Y.
Saito
N.
Noguchi
Y.
Konuma
H.
Kumagai
S.
(
1998
)
Энтерогеморрагический Escherichia coli O157:H7 присутствует в ростках редьки
.
Appl Environ Microbiol
64
:
1532
–
1535
.
Джей
М.Т.
Garrett
V.
Mohle-Boetani
J.C.
и другие. . (
2004
)
Вспышка инфекции Escherichia coli O157:H7 в нескольких штатах, связанная с употреблением тако из говядины в сети ресторанов быстрого питания
.
Clin Infect Dis
39
:
1
–
7
.
Кармали
М.А.
.
Мол Биотехнолог
26
:
117
–
122
.
Кармали
М.А.
(
2009
)
Хозяин и патогенные детерминанты вероцитотоксин-продуцирующего Escherichia coli -ассоциированного гемолитико-уремического синдрома
.
Дополнение к почкам
.
Февраль
:
S4-7
.
Кассенборг
Х.Д.
Hedberg
C.W.
Hoekstra
M.
и другие. . (
2004
)
Посещение ферм и недоваренные гамбургеры как основные факторы риска спорадических Escherichia coli Инфекция O157:H7: данные исследования случай-контроль на 5 сайтах FoodNet
.
Clin Infect Dis
38
(
Приложение 3
):
S271
–
S27 8
3 .
Кинг
Ч.Х.
Выстрелы
Э.Б.
младший
Вули
Р.Е.
Портер
К.Г.
(
1988
)
Выживание кишечных палочек и бактериальных патогенов среди простейших при хлорировании
.
Appl Environ Microbiol
54
:
3023
–
3033
.
Кинг
Л.А.
Ногареда
Ф.
Вейл
Ф.Х.
и другие. . (
2012
)
Вспышка шига-токсин-продуцирующей Escherichia coli O104:h5, связанный с органическими ростками пажитника, Франция, июнь 2011 г.
.
Clin Infect Dis
54
:
1588
–
1594
.
Кляйн
Т.М.
Александр
М.
(
1986
)
Ингибиторы бактерий в озерной воде
.
Appl Environ Microbiol
52
:
114
–
118
.
Kudoh
Y.
Obata
H.
Kusunoki
J.
Monma
C.
Shingaki
M.
Yanagawa
Y.
Yamada
S.
Мацусита
С.
Ито
Т.
(
1994
)
Эпидемиологические исследования на вероцитотоксинпродуцирующие Escherichia coli инфекции в Японии
.
Последние достижения в области инфекций, вызывающих вероцитотоксин Escherichia coli (Excerpta Medica International Congress Series 1072)
Амстердам
.
Ла Раджоне
Р.М.
Best
A.
Woodward
M.J.
Уэльс
AD
(
2009
)
Escherichia coli Колонизация O157:H7 у мелких домашних жвачных
.
FEMS Microbiol Rev
33
:
394
–
410
.
Lainhart
W.
Stolfa
G.
Koudelka
G.B.
(
2009
)
Шига-токсин как бактериальная защита от эукариотического хищника, Tetrahymena thermophila
.
J Бактериол
191
:
5116
–
5122
.
Ламарш
М.Г.
Ваннер
Б.Л.
Крепен
С.
Харел
Ж.
(
2008
)
Фосфатный регулон и вирулентность бактерий: регуляторная сеть, связывающая гомеостаз фосфатов и патогенез
.
FEMS Microbiol Rev
32
:
461
–
473
.
Lange
R.
Hengge-Aronis
R.
(
1991
)
Идентификация центрального регулятора экспрессии генов стационарной фазы в Escherichia coli
.
Мол Микробиол
5
:
49
–
59
.
Леклерк
Х.
(
2003
)
Взаимоотношения между обычными водными бактериями и патогенами в питьевой воде
.
Гетеротрофные пластины и безопасность питьевой воды: значение HPC для качества воды и здоровья человека
(
Бартрам
J.
Ed.), Pp.
84
—
85
.
по
Издательство IWA
,
Лондон, Великобритания
.
Ли
М.Б.
Грейг
Дж.Д.
(
2008
)
Обзор кишечных вспышек в детских учреждениях: рекомендации по эффективному инфекционному контролю
.
J Environ Health
71
:
24
–
32
,
.
Bansal
T.
Jayaraman
A.
Bentley
W.E.
Древесина
Т.К.
(
2007
)
Энтерогеморрагические Escherichia coli биопленки ингибируются 7-гидроксииндолом и стимулируются изатином
.
Appl Environ Microbiol
73
:
4100
–
4109
.
Ли
М.С.
Крумпельман
С.Л.
Яблоко
Дж.К.
Янси
JWS
Кегли
Э.Б.
PAS
Джонсон
М. Г.
Брашэрз
М.М.
Стивенс
Т.П.
(
2009
)
In Vitro и In Vivo Исследования противомикробных обработок для снижения содержания Escherichia coli O157:H7 в навозе крупного рогатого скота
.
Профессиональный зоотехник
25
:
49
–
59
.
Ли
Дж.Х.
Ким
Ю.Г.
Чо
М.Х.
Древесина
Т.К.
(
2011
)
Транскриптомный анализ генетических механизмов факторов, связанных с образованием биопленок у Escherichia coli O157:H7
.
Curr Microbiol
62
:
1321
–
1330
.
ЛеЖён
Дж.Т.
Бессер
Т.Е.
Хэнкок
Д.Д.
(
2001
)
Поилки для крупного рогатого скота как резервуары для Escherichia coli O157
.
Appl Environ Microbiol
67
:
3053
–
3057
.
Лицензия
К.
Оутс
К.Р.
Synge
Б.А.
Рейд
Т.М.
(
2001
)
Вспышка инфекции E. coli O157 с признаками передачи от животных человеку в результате заражения частного водоснабжения
.
Эпидемиол Инфекция
126
:
135
–
138
.
Ли
И.С.
Фрей
Дж.
Слончевски
Дж.Л.
Фостер
Дж.В.
(
1995
)
Сравнительный анализ выживаемости в условиях экстремальной кислотности у Salmonella typhimurium , Shigella flexneri и Escherichia coli
.
J Бактериол
177
:
4097
–
4104
.
Гилкрист
А.
Чжан
Дж.
Ли
X.F.
(
2008
)
Обнаружение жизнеспособных, но некультивируемых бактерий Escherichia coli O157:H7 в питьевой воде и речной воде
.
Appl Environ Microbiol
74
:
1502
–
1507
.
Макино
S.I.
Asakura
H.
Shirahata
T.
Ikeda
T.
Takeshi
K.
K.
. (
2000
)
Входит ли энтерогеморрагическая Escherichia coli O157:H7 в жизнеспособное, но некультивируемое состояние в соленой икре лосося?
Appl Environ Microbiol
66
:
5536
–
5539
.
Мартин
Дж. Ф.
(
2004
)
Фосфатный контроль биосинтеза антибиотиков и других вторичных метаболитов опосредован системой PhoR-PhoP: незавершенная история
.
J Бактериол
186
:
5197
–
5201
.
Масана
М.О.
Д’Астек
Б.А.
Палладино
вечера
Галли
Л.
Дель Кастильо
Л.Л.
Вилакоба
Э.
Ирино
К.
Ривас
М.
(
2011
)
Генотипическая характеристика штамма Escherichia coli , продуцирующего не-O157 шигатоксин, на мясных бойнях Аргентины
.
J Пищевая защита
74
:
2008
–
2017
.
Мэтьюз
К.Р.
Мердо
П.А.
Брэмли
А.Дж.
(
1997
)
Инвазия эпителиальных клеток крупного рогатого скота продуцирующими вероцитотоксин Escherichia coli O157:H7
.
J Appl Microbiol
82
:
197
–
203
.
Макдэниел
Т.К.
Джарвис
К.Г.
Донненберг
М.С.
Капер
Дж. Б.
(
1995
)
Генетический локус стирания энтероцитов, консервативный среди различных энтеробактериальных патогенов
.
P Natl Acad Sci USA
92
:
1664
–
1668
.
МакГи
П.
Болтон
Д.Дж.
Шеридан
Дж.Дж.
Эрли
Б.
Келли
Г.
Леонард
Н.
(
2002
)
Выживание Escherichia coli O157:H7 в сельскохозяйственной воде: роль переносчика в передаче микроорганизма в стадах
.
J Appl Microbiol
93
:
706
–
713
.
Мид
P.S.
Гриффин
вечера
(
1998
)
Кишечная палочка O157:H7
.
Ланцет
352
:
1207
–
1212
.
Миллер
Дж. Ф.
Мекаланос
Дж. Дж.
Фальков
С.
(
1989
)
Регуляция координации и сенсорная трансдукция в контроле вирулентности бактерий
.
Наука
243
:
916
–
922
.
Морита
Р.Ю.
(
1997
)
Бактерии в олиготрофной среде: голодание-выживание
.
Чепмен и Холл
,
Дордрехт, Нидерланды
.
Глушитель
A.
Traulsen
D.D.
Lange
R.
Hengge-Aronis
R.
.
J Бактериол
178
:
1607
–
1613
.
Глушитель
A.
Barth
M.
Marschall
C.
Hengge-Aronis
R.
3
3 .
J Бактериол
179
:
445
–
452
.
Natvig
E.E.
Ingham
S.C.
Ingham
B.H.
Cooperband
L.R.
Роупер
Т.Р.
(
2002
)
Salmonella enterica serovar Typhimurium и Escherichia coli заражение корнеплодов и листовых овощей, выращенных на почвах с включенным бычьим навозом
.
Appl Environ Microbiol
68
:
2737
–
2744
.
Нейлор
Ю.В.
Галли
Д.Л.
Низкий
J.C.
(
2005
)
Энтерогеморрагическая E. coli в ветеринарии
.
Int J Med Microbiol
295
:
419
–
441
.
О
D.H.
Берри
E.
Cooley
M.
Mandrell
R.
Breidt
F.
младший
(
2009
)
Штаммы Escherichia coli O157:H7, выделенные из источников окружающей среды, значительно отличаются по устойчивости к уксусной кислоте по сравнению со штаммами
, вызывающими вспышку среди людей.
J Food Prot
72
:
503
–
509
.
Оливер
Джей Ди
(
2005
)
Бактерии в жизнеспособном, но не культивируемом состоянии
.
Дж Микробиол
43
:
93
–
100
.
Олсен
С.Дж.
Миллер
Г.
Брейер
Т.
Кеннеди
М.
Хиггинс
C.
Walford
J.
McKee
G.
Bibb
W.
Mead
3
3 P.
( 2002
)
Вспышка инфекции Escherichia coli O157:H7 и гемолитико-уремического синдрома, передающихся через воду: последствия для сельских систем водоснабжения
.
Emerg Infect Dis
8
:
370
–
375
.
Пайтан
А.
Маклафлин
К.
(
2007
)
Круговорот фосфора в океане
.
Chem Rev
107
:
563
–
576
.
Перна
Н.Т.
Планкетт
Г.
3-й
Бурланд
В.
и другие.
. (
2001
)
Последовательность генома энтерогеморрагической Escherichia coli O157:H7
.
Природа
409
:
529
–
533
.
Петерсон
К.Н.
Мандель
М.Дж.
Силхави
Т.Дж.
(
2005
)
Escherichia coli голодание: существенные питательные вещества имеют явное значение
.
J Бактериол
187
:
7549
–
7553
.
Poirier
K.
Faucher
S.P.
Beland
M.
Brousseau
R.
Gannon
V.
Martin
C.
Harel
J.
Дайгле
Ф.
(
2008
)
Escherichia coli O157:H7 выживает в макрофагах человека: глобальный профиль экспрессии генов и участие шига-токсинов
.
Infect Immun
76
:
4814
–
4822
.
Паттамредди
С.
Корник
Н/Д
Миньон
Ф.К.
(
2010
)
Полногеномный транспозонный мутагенез выявляет роль генов pO157 в развитии биопленок в Escherichia coli O157:H7 EDL933
.
Infect Immun
78
:
2377
–
2384
.
Ричи
Дж. М.
Кэмпбелл
Г. Р.
Shepherd
J.
Beaton
Y.
Jones
D.
Killham
K.
Artz
R.R.
(
2003
)
Стабильная биолюминесцентная конструкция Escherichia coli O157:H7 для оценки опасности долгосрочного выживания в окружающей среде
.
Appl Environ Microbiol
69
:
3359
–
3367
.
Ривас
М.
Миливебски
Э.
Чинен
И.
Деза
Н.0
Леот
(
2006
)
[Эпидемиология гемолитико-уремического синдрома в Аргентине. Диагностика возбудителя, резервуаров и путей передачи]
.
Медицина
66
(
Приложение 3
):
27
–
32
. Ривас
Л.
Дайкс
Г.А.
Феган
Н.
(
2007
)
Сравнительное исследование образования биопленки шига токсигенными штаммами Escherichia coli с использованием эпифлуоресцентной микроскопии на нержавеющей стали и методом микротитровальных планшетов
.
J Microbiol Methods
69
:
44
–
51
.
Родригес-Сарагоса
С.
(
1994
)
Экология свободноживущих амеб
.
Crit Rev Microbiol
20
:
225
–
241
.
Розен
Ю.
Белкин
С.
(
2001
)
Выживание кишечных бактерий в морской воде
.
FEMS Microbiol Rev
25
:
513
–
529
.
Рю
Дж.Х.
Бечат
Л.Р.
(
2005
)
Образование биопленки Escherichia coli O157:H7 на нержавеющей стали: влияние экзополисахарида и продукции Curli на ее устойчивость к хлору
.
Appl Environ Microbiol
71
:
247
–
254
.
Рю
Дж.Х.
Beuchat
Л.Р.
(
2004
)
Прикрепление и образование биопленки Escherichia coli O157:H7 на нержавеющей стали под влиянием производства экзополисахарида, наличия питательных веществ и температуры
.
J Food Prot
67
:
2123
–
2131
.
Сакума
М.
Урасима
М.
Окабе
Н.
(
2006
)
Вероцитотоксин-продуцирующие Escherichia coli , Япония, 1999–2004 гг.
.
Emerg Infect Dis
12
:
323
–
325
.
Saldana
Z.
Xicohtencatl-Cortes
J.
Avelino
F.
Phillips
A.D.
Kaper
J.B.
Puente
J.L.
Giron
Дж.А.
(
2009
)
Синергетическая роль завитков и целлюлозы в адгезии клеток и образовании биопленки при прикреплении и стирании Escherichia coli и идентификация Fis как отрицательного регулятора завитков
.
Environ Microbiol
11
:
992
–
1006
.
Салдана
Z.
Санчес
E.
Xicohtencatl-Cortes
J.
Puente
J.L.
Giron
J.A.
.
Front Microbiol
2
:
119
.
Scallan
E.
Hoekstra
R.M.
Angulo
FJ
Tauxe
R.V.
Уиддоусон
М.А.
Рой
С.Л.
Джонс
Дж.Л.
Гриффин
PM
(
2011
)
Болезни пищевого происхождения, приобретенные в США – основные патогены
.
Emerg Infect Dis
17
:
7
–
15
.
Семенова
А. В.
фургон Bruggen
A.H.
фургон Overbeek
L.
Termorshuizen
A.J.
Семенов
А.М.
.
FEMS Microbiol Ecol
60
:
419
–
428
.
Семенов
А.В.
фургон Overbeek
L.
фургон Bruggen
A.H.
(
2009
)
Просачивание и выживание Escherichia coli O157:H7 и Salmonella enterica serovar Typhimurium в почве, обработанной зараженным молочным навозом или навозной жижей
3
33
Appl Environ Microbiol
75
:
3206
–
3215
.
Сиддики
Р.
Хан
Н/Д
(
2012
)
Акантамеба – эволюционный предок макрофагов: миф или реальность?
Exp Parasitol
130
:
95
–
97
.
Зигеле
Д.А.
Колтер
Р.
(
1992
)
Жизнь после бревна
.
J Бактериол
174
:
345
–
348
.
Skandamis
П.Н.
Stopforth
J.D.
Ashton
L.V.
Георнарас
I.
Кендалл
П.А.
Sofos
J. N.
(
2009
)
Escherichia coli O157:H7 Выживаемость, образование биопленки и кислотоустойчивость в условиях, имитирующих бойню, во влажных и сухих условиях
.
Пищевой микробиол
26
:
112
–
119
.
Снедекер
К.Г.
Шоу
Д.Дж.
Замок
M.E.
Prescott
R.J.
(
2009
)
Первичные и вторичные случаи в Escherichia coli Вспышки O157: статистический анализ
.
BMC Infect Dis
9
:
144
.
Соломон
Э. Б.
Потенски
Си Джей
Мэтьюз
К.Р.
(
2002a
)
Влияние метода орошения на передачу и устойчивость Escherichia coli O157:H7 на листьях салата
.
J Food Prot
65
:
673
–
676
.
Соломон
Э.Б.
Ярон
С.
Мэтьюз
К.Р.
(
2002b
)
Передача Escherichia coli O157:H7 из загрязненного навоза и поливной воды в ткани растений салата и их последующая интернализация
.
Appl Environ Microbiol
68
:
397
–
400
.
Стейнберг
К.М.
Левин
Б.Р.
.
Proc Biol Sci
274
:
1921
–
1929
.
Steinert
M.
Birkness
K.
White
E.
Fields
B.
Quinn
F.
(
1998
)
Бациллы Mycobacterium avium растут сапрозоически в совместной культуре с Acanthamoeba polyphaga и выживают в стенках кисты
.
Appl Environ Microbiol
64
:
2256
–
2261
.
Стиги
К.А.
Макдональд
Дж.К.
Теллез-Марфин
А. А.
Лофи
К.Х.
.
Emerg Infect Dis
18
:
477
–
479
.
Sugumar
G.
Chrisolite
B.
Velayutham
P.
Selvan
A.
Рамеш
U.
.
J Environ Biol
29
:
387
–
391
.
Bausch
C.
Richmond
C.
Blattner
F.R.
Конвей
Т.
(
1999
)
Функциональная геномика: анализ экспрессии Escherichia coli , растущих на минимальной и богатой средах
.
J Бактериол
181
:
6425
–
6440
.
Thompson
L.H.
Giercke
S.
Beaudoin
C.
Woodward
D.
Wylie
J.L.
(
2005
)
Усиленный эпиднадзор за штаммами Escherichia coli , не продуцирующими O157 веротоксин, в образцах стула человека из Манитобы
.
Can J Infect Dis Med Microbiol
16
:
329
–
334
.
Торрес
А.Г.
Хирон
Дж.А.
Перна
Н.Т.
Бурланд
В.
Блаттнер
Ф.Р.
Авелино-Флорес
Ф.
Капер
Дж.Б.
(
2002
)
Идентификация и характеристика lpfABCC’DE, фимбриального оперона энтерогеморрагической Escherichia coli O157:H7
.
Infect Immun
70
:
5416
–
5427
.
Torres
A.G.
Jeter
C.
Langley
W.
Matthysse
A.G.
3
3
(
2005
)
Дифференциальное связывание Escherichia coli O157:H7 с люцерной, эпителиальными клетками человека и пластиком опосредуется различными поверхностными структурами
.
Appl Environ Microbiol
71
:
8008
–
8015
.
Улих
Г.А.
Кук
П.Х.
Соломон
Э.Б.
(
2006
)
Анализ фенотипа красно-сухой-шероховатости штамма Escherichia coli O157:H7 и его роли в формировании биопленки и устойчивости к антибактериальным препаратам
.
Appl Environ Microbiol
72
:
2564
–
2572
.
Улих
Г.А.
Роджерс
Д.П.
Мосье
Д.А.
(
2010
)
Escherichia coli серотип O157:H7 Удержание на твердых поверхностях и устойчивость к перекиси усиливаются за счет образования биопленки из двойного штамма
.
Патог пищевого происхождения Dis
7
:
935
–
943
.
Vally
H.
Hall
G.
Dyda
A.
Raupach
J.
Knope
K.
Combs
B.
6 DESMARCHLIE
(
2012
)
Эпидемиология шига-токсина, продуцирующего Escherichia coli в Австралии, 2000–2010 гг.
.
BMC Public Health
12
:
63
.
VanBogelen
Р.А.
Олсон
Э.Р.
Ваннер
Б.Л.
Нейдхардт
Ф.К.
(
1996
)
Общий анализ белков, синтезированных при ограничении фосфора в Escherichia coli
.
J Бактериол
178
:
4344
–
4366
.
Варма
Дж.К.
Грин
К.Д.
Реллер
ME
и другие.
. (
2003
)
Вспышка инфекции Escherichia coli O157 после контакта с зараженным зданием
.
JAMA
290
:
2709
–
2712
.
Vital
M.
Hammes
F.
Egli
T.
(
2008
)
Escherichia coli O157 может расти в природной пресной воде при низкой концентрации углерода
.
Environ Microbiol
10
:
2387
–
2396
.
Уэйд
Т.Дж.
Кальдерон
RL
Sams
E.
Beach
M.
Brenner
K.P.
Williams
A.H.
Dufour
AP
(
2006
)
Быстро измеряемые показатели качества рекреационной воды позволяют прогнозировать желудочно-кишечные заболевания, связанные с плаванием
.
Environment Health Perspect
114
:
24
–
28
.
Ван
Г.
Дойл
М.П.
(
1998
)
Выживаемость энтерогеморрагической Escherichia coli O157:H7 в воде
.
J Food Prot
61
:
662
–
667
.
Ван
А.
Сон
Б.Э.
Лисс
С.Н.
(
2011
)
Влияние роста биопленки на перенос Escherichia coli O157:H7 в песке
.
Подземные воды
49
:
20
–
31
.
Уорбертон
Д.У.
Остин
Дж.В.
Харрисон
Б.Х.
Сандерс
Г.
(
1998
)
Выживание и восстановление Escherichia coli O157:H7 в инокулированной бутилированной воде
.
J Food Prot
61
:
948
–
952
.
Watterworth
L.
Rosa
B.
Schraft
H.
Topp
E.
Leung
K.
(
2006
)
Выживаемость различных ERIC-генотипов шига-токсин-продуцирующих Escherichia coli в колодезной воде
.
Вода Воздух Почва Загрязнение
177
:
367
–
382
.
Werner
M.
Maschek
H.J.
Kaloutsi
V.
Buhr
T.
Kausche
F.
Delventhal
S.
Georgii
A.
(
1990
)
Цитогенетическое исследование клеток костного мозга при заболеваниях стволовыми клетками при миелодиспластических синдромах
.
Verh Dtsch Ges Pathol
74
:
139
–
143
.
Wieler
L.H.
Vieler
E.
ERPENSTEIN
C.
Schlapp
T.
Steinruck
H.
Bauerfeind
. R.
H.
Bauerfeind
. R.
6
.
6
.
6
.
6
.
H.
69
.0003
А.
Бальер
Г.
(
1996
)
Шига-токсин-продуцирующие штаммы Escherichia coli от крупного рогатого скота: ассоциация адгезии с носительством eae и других генов
.
J Clin Microbiol
34
:
2980
–
2984
.
Уильямс
Л.Д.
Гамильтон
P.S.
Уилсон
Б.В.
Estock
MD
(
1997
)
Вспышка Escherichia coli 0157:H7: длительное выделение и передача от человека человеку в детском саду
.
J Environment Health
59
:
9
.
Xicohtencatl-Cortes
J.
Sanchez Chacon
E.
Saldana
Z.
Фрир
Э.
Хирон
Дж.А.
(
2009
)
Взаимодействие Escherichia coli O157:H7 с листовой зеленью
.
J Food Prot
72
:
1531
–
1537
.
Черный
Р.Л.
Чирилло
Дж.Д.
(
2004
)
Доказательства того, что hsp90 участвует в измененных взаимодействиях вариантов Acanthamoeba castellanii с бактериями
.
Эукариотическая клетка
3
:
567
–
578
.
Yoon
Y.
Sofos
J.N.
(
2008
)
Активность аутоиндуктора-2 грамотрицательных пищевых патогенных бактерий и ее влияние на формирование биопленок
.
J Food Sci
73
:
M140
–
M147
.
Yoshida
Y.
Sugiyama
S.
Oyamada
T.
Yokoyama
K.
Makino
K.
(
2012
)
Новые члены фосфатного регулона в Escherichia coli O157:H7, идентифицированные с помощью полногеномного подхода
.
Гена
502
:
27
–
35
.
Copyright © 2013 Федерация европейских микробиологических обществ. Опубликовано Blackwell Publishing Ltd. Все права защищены.
Copyright © 2013 Federation of European Microbiological Societies. Опубликовано Blackwell Publishing Ltd. Все права защищены
Классификация
Влияние эволюции Дарвин изменил все. Публикация его работы «Происхождение видов » в 1859 году бросила всю биологическую науку в новую парадигму, включая изучение теории классификации и принципов таксономии.
Используя логику в качестве основы своей работы, и Аристотель, и Линней разработали свои схемы классификации на таксономических принципах, которые в своей основе были произвольными. Их группы, хотя и были логичны, не основывались на каких-либо очевидных отношениях биологической природы. Это были удобные группы, которые люди могли быстро увидеть, идентифицировать и использовать.
Это было приемлемо, потому что (а) никто не мог придумать ничего лучше, и (б) большинство людей в то время верили в концепцию «фиксированных видов», согласно которой организм был создан в его нынешнем виде и никогда не мог измениться.
После Дарвина стало понятно, что организмы действительно могут изменяться и что все нынешние формы живых существ пришли к этой форме путем изменений и естественного отбора, механизма эволюции. Ученые начали строить филогении , списки или диаграммы, показывающие эволюционные пути, пройденные популяциями организмов на протяжении многих поколений и в течение длительных периодов времени.
Эти филогенетические диаграммы быстро стали напоминать деревья, поскольку стало понятно, что предковые линии иногда распадались, разветвлялись и превращались в два или более разных вида, которые впоследствии могли разветвляться снова и снова. Филогенетическое древо было немного похоже на генеалогическое древо, показывающее, кем были ближайшие родственники, у кого был общий предок и когда.
Организмы были связаны друг с другом, и эти отношения могли лечь в основу таксономии нового типа; на основе эволюционного происхождения и эволюционного родства.
Из этого принципа возникли два основных типа таксономии. Они похожи, но есть серьезные различия. Эволюционная систематика делает попытку построить деревья, которые точно показывают филетические линии (правильное ветвление на генеалогическом древе), наряду с рассмотрением того, когда и как новые виды возникли и переместились в новые места обитания и ниши (установил «новый» способ жизнь, а не какое-то тривиальное изменение характера). Кладистика , однако, игнорирует, когда и где возникает ветвь, пытается использовать чисто объективные критерии и определяет каждую точку ветвления фундаментальным признаком эволюционного значения. Оба метода имеют свои сильные и слабые стороны. Кладистика получила свое название от ветвей генеалогического древа, которые называются кладами . Кладограмма представляет собой стилизованную диаграмму, которая выглядит как ряд Y или разветвлений на дороге. На каждой ветви или Y-образном стыке используются новые символы эволюционного происхождения, чтобы отделить одну группу от остальных.
Кладограммы могут быть построены для любой группы организмов. Например, следующие организмы представляют собой набор, из которого можно составить кладограмму; кенгуру, дождевой червь, амеба, ящерица, кошка, губка и лосось. Каждое из этих существ имеет эволюционную связь друг с другом. Все они имеют общее происхождение, и все их нынешние формы произошли от разветвленных событий где-то в филогенетическом прошлом. Вопрос в том, когда произошли эти ответвления?
Процесс построения кладограммы начинается с данных; таблица черт или характеристик, которые развились или были является производным в процессе эволюции.
Производные символы сегментированный губки волосы плацента многоклеточный конечностей кенгуру + + + — + + дождевой червь + — — — + — амеба — — — — — — ящерица + + — — + + кот + + + + + + губка — — — — + — лосось + + — — + —
На следующем этапе каждый из организмов сравнивается, чтобы увидеть, имеют ли они общие черты или производные признаки. Например, все, кроме амебы, имеют общий производный признак «многоклеточность», но только кошка и кенгуру имеют общий производный признак «волосы».
Используя эти образцы общих производных признаков, кладограмма может быть построена как серия Y или ветвей. На каждой ветви один из организмов, не имеющий общих признаков с остальной группой, «ответвляется» в свою собственную кладу. Порядок или последовательность этих ветвей зависит от того, сколько символов осталось в большей группе.
Посмотреть на себя. Наведите курсор на кнопки управления рядом с групповой диаграммой и реконструируйте процесс создания кладограммы:
Кладограмма Группа Один Два Три Четыре Пять Шесть Черты
Кладограммы — полезный способ визуальной организации отношений между существами, имеющими и не имеющими общих производных признаков. На практике, когда строятся кладограммы, возможно, придется учитывать многие сотни символов, и нужны компьютеры, чтобы выбрать наилучшее соответствие между ветвями и тем, кто должен быть на них. Кладограммы подчеркивают последовательность или порядок, в котором производные признаки возникают из центрального филогенетического дерева. В этом их главная сила. Однако ничто в кладограмме не указывает на то, насколько сильным или глубоким является производный признак, и на его эволюционную важность. Всем используемым символам придается равный вес. Иногда это может приводить к необычным группировкам, которые могут быть технически правильными, но сомнительными.
Эволюционная систематика использует таксономический принцип, который также основан на разделении филетических родословных, и строит генеалогические деревья, но, в отличие от чисто кладистического метода, используется система взвешивания, которая отдает предпочтение одним производным признакам по сравнению с другими. Например, производный признак или черта, такая как рождение детенышей живыми (как это наблюдается у сумчатых и млекопитающих), считается более значимым, чем разнообразие цветов птичьих перьев.
Тем не менее, и это большое «однако», эволюционные систематики действительно считают необходимым использовать много субъективных суждений при принятии решения о том, каким факторам придавать наибольшее значение. Всякий раз, когда люди заменяют своими собственными «чувствами» доказуемый биологический факт, к результатам всегда нужно относиться с осторожностью и постоянно пересматривать. Однако даже при такой осторожности эволюционная систематика, по-видимому, в настоящее время дает картину биологических отношений между видами, основанную на самых надежных таксономических принципах.
Спасти амебу? | Среди величественных деревьев
Следует ли более непосредственно учитывать микробное биоразнообразие (то есть микроскопические организмы, такие как бактерии, археи, протисты и некоторые грибы) в усилиях по сохранению? Несомненно, важность микроскопических организмов в земной системе невозможно переоценить, поскольку они играют жизненно важную роль в глобальных циклах элементов – фиксируют атмосферный азот и делают его доступным для растений, возвращают азот обратно в атмосферу, играют важную роль в пищевых цепях и способствуют разложение органических веществ. Кроме того, в будущем микроорганизмы, вероятно, станут важнейшим источником новых фармацевтических препаратов (Chivian & Bernstein, 2008). Однако микроорганизмы лишь изредка включались в исследования биоразнообразия, и хотя Международный союз охраны природы (МСОП) недавно добавил четырех находящихся под угрозой исчезновения простейших в Красный список (здесь, здесь, здесь и здесь), общественный резонанс по поводу их защита кажется маловероятной в ближайшее время.
Пластинка из монографии Джозефа Лейди 1879 года о раковинных амебах (1–8: Nebela ansata, 9–14: Nebela hippocrepis). Раковинные амебы представляют собой группу протистов, которые производят морфологически различные раковины.
Конечно, отсутствие акцента на микроорганизмах в природоохранной биологии не особенно удивительно, учитывая, что 1) мы относительно мало знаем о микробном разнообразии, 2) микробы, возможно, немного менее симпатичны и харизматичны, чем многие другие группы организмов (хотя они, безусловно, симпатичнее, чем некоторые виды), и 3) широко распространено мнение, что микробные виды имеют космополитическое распространение. Другими словами, они встречаются повсюду. В частности, микробный вид встречается везде, где существует подходящая среда обитания для этого конкретного вида. В 1934, Баас Бекинг просто сформулировал эту гипотезу так: «все есть везде, но среда выбирает» (де Вит и Бувье, 2006). Эта гипотеза подразумевает, что рассредоточение не ограничивает распространение микробов, и эндемизм (т. е. приуроченность к определенному региону) должен быть редким явлением. Конечно, если все есть везде, то нам не нужно сильно беспокоиться о каком-либо конкретном виде микробов, потому что вымирание крайне маловероятно, если только не произойдет уничтожение всех подходящих местообитаний… по всей планете. Будем надеяться, что это останется маловероятным в течение некоторого времени.
Однако исследования теперь ясно показывают, что микробная биогеография является более тонкой и сложной, чем подразумевается гипотезой «все повсюду». За последние пару десятилетий появилось множество примеров ограниченных географических ареалов. Некоторые особенно яркие примеры относятся к панцирным амебам, группе протистов, производящих раковины, которые относительно хорошо изучены, по крайней мере, по сравнению со многими другими микробными группами. Например, Apodera vas и виды рода Certesella ограничены южным полушарием и тропиками, хотя подходящие местообитания для этих видов существуют в Евразии и Северной Америке (Smith and Wilkinson 2007). Модели предполагают, что эти виды произошли на континенте Гондвана через некоторое время после того, как он отделился от Лавразии около 200-180 миллионов лет назад, и, таким образом, их нынешнее распространение может быть отражением этой геологической и эволюционной истории (Смит и Уилкинсон, 2007). По-видимому, эти виды не смогли рассеяться в Северную Америку или Евразию.
Палеогеографическая реконструкция Земли в позднем триасе, незадолго до отделения Гондваны (южные континенты) от Лавразии (северные континенты). Нынешнее распространение Apodera vas и видов рода Certesella почти ограничено территорией, которая когда-то была частью Гондваны, что позволяет предположить, что они эволюционировали на этом массиве суши (Smith & Wilkinson 2007). (Изображение с сайта wikipedia.org).
Потенциально более ограниченный шаблон распределения — Nebela ansata , необычный (и осмелюсь сказать, «харизматичный») вид раковинных амеб, известный лишь в нескольких местах на востоке Северной Америки. В 1874 году Джозеф Лейди впервые столкнулся с этим видом из коллекций, которые он собрал в сосновых степях Нью-Джерси (Leidy 1879). Со времени его первоначальных описаний этот вид упоминался в литературе только три раза, всегда в умеренном поясе восточной части Северной Америки и всегда обитающий в заболоченных местах на влажном мхе Sphagnum . Этот вид не упоминается в литературе с начала 19 века.50-х годов, несмотря на интенсивный сбор образцов сообществ раковинных амеб водно-болотных угодий (Heger et al. 2011).
В поисках забытого микроба
Nebela ansata из болота Webb’s Mill Bog в сосновых степях Нью-Джерси. Для вида характерны роговидные боковые отростки. (Фото: Тьерри Хегер)
Сосновые степи Нью-Джерси находятся всего в нескольких часах езды от Университета Лихай, поэтому несколько лет назад мы решили посмотреть, сможем ли мы найти неуловимого и давно забытого микроба Лейди. Его первоначальные образцы были взяты из кедровых болот недалеко от города Абсеком (теперь пишется как Абсекон), и мы использовали это в качестве ориентира при выборе потенциальных заболоченных мест для наших усилий по отбору проб. Он также упомянул, что Nebela ansata обычно встречается вместе с Nebela carinata , более широко распространенным видом. Экология Nebela carinata хорошо изучена, он предпочитает очень влажный Sphagnum мох и часто встречается в изобилии там, где уровень грунтовых вод находится всего на несколько сантиметров ниже поверхности мха. Мы использовали эти знания для определения вероятных местообитаний в каждом из отобранных нами водно-болотных угодий. Маура Салливан (Университет Лихай, аспирант) и я собирали образцы в течение двух полных дней, вернули образцы в лабораторию для исследования и сумели найти вид!
Интересно, что примерно в то же время, когда мы повторно открыли Nebela ansata в сосновых степях Нью-Джерси, этот вид был также обнаружен в Новой Шотландии Барри Уорнером и Таро Асада (Университет Ватерлоо). Вскоре сформировались совместные усилия с целью собрать воедино всю известную информацию о распространении, экологии и филогении видов. Под руководством Тьерри Хегера, в то время аспиранта (Швейцарский федеральный исследовательский институт WSL и Женевский университет), был проведен исчерпывающий обзор литературы и музеев, который использовался в тандеме с анализом образцов из Новой Шотландии и Сосновых степей, чтобы дать представление о биогеография и экология Небела ансата . В совокупности результаты представляют собой убедительный случай микроорганизма с очень ограниченным диапазоном распространения — по-видимому, ограниченным умеренным климатом восточной части Северной Америки — что представляет собой явное исключение из гипотезы «все есть везде» (Heger et al. 2011).
Водно-болотные угодья Уэббс-Милл в сосновых степях Нью-Джерси, где были обнаружены особи редкого вида раковинных амеб, Nebela ansata.
Сколько других микроорганизмов также имеет географически ограниченное распространение? Каковы основные причины этих ограниченных дистрибутивов? Сколько эндемичных видов остаются неописанными? Какую роль играют эти относительно редкие микроорганизмы в экосистемах? Сколько микробных видов уязвимы перед разрушением среды обитания или другими глобальными изменениями? Многое еще предстоит сделать в области изучения биоразнообразия.
-rkb-
Процитированная литература
Chivian, E. & A. Bernstein, eds. 2008. Поддержание жизни. Как здоровье человека зависит от биоразнообразия. Издательство Оксфордского университета.
де Вит. Р. и Т. Бувье. 2006. «Все есть везде, но среда выбирает»; что на самом деле сказали Баас Бекинг и Бейеринк? Экологическая микробиология 8: 755-758.
Хегер, Т.Дж., Р.К. Бут, М.Э. Салливан, Д.М. Уилкинсон, Б.Г. Уорнер, Т. Асада, Ю. Мазей, Р. Мейстерфельд и Э.А.Д. Митчелл. 2011. Новое открытие Nebela ansata (Amoebozoa: Arcellinida) в восточной части Северной Америки: биогеографические последствия. Журнал биогеографии 38: 1897-1906.
Leidy, J. 1879. Пресноводные корненожки Северной Америки. Отчет Геологической службы территорий США, 12, 1–324.
Смит, Х.Г. и Д.М. Уилкинсон. 2007. Не все свободноживущие микроорганизмы имеют космополитическое распространение — случай Nebela ( Apodera ) vas Certes (Protozoa: Amoebozoa: Arcellinida). Журнал биогеографии, 34: 1822–1831.
Нравится:
Нравится Загрузка…
Бактерии пронзают амеб микрокинжалами
Группа подпружиненных кинжалов внутри бактерии. Зеленый показывает их в «заряженном» виде, красный — после того, как кинжал был запущен. Кредит: Лео Попович Бактерии должны остерегаться амебы. Голодные амебы охотятся на них: они ловят их своими псевдоподиями, а затем поглощают и переваривают.
Однако некоторые бактерии умеют защищаться. Одним из таких является Amoebophilus, который был обнаружен исследователями Венского университета несколько лет назад. Эта бактерия может не только выжить внутри амебы, но и процветать: амеба стала ее излюбленной средой обитания.
Вместе с венскими первооткрывателями бактерии ученые из ETH Zurich обнаружили механизм, который, по их мнению, имеет решающее значение для выживания Amoebophilus внутри амебы. У бактерии есть устройства для стрельбы микрокинжалами. Он может использовать кинжалы, чтобы пронзить амебу изнутри и таким образом избежать пищеварения.
Побег из кишечника амебы
Стреляющий механизм состоит из оболочки, прикрепленной к внутренней мембране бактерии с помощью базовой пластины и якорной платформы. Жоао Медейрос, аспирант группы профессора Мартина Пилхофера в ETH, объясняет механизм: «Оболочка подпружинена, и микрокинжал лежит внутри нее. Когда оболочка сжимается, кинжал очень быстро выбрасывается наружу через бактериальную мембрану. .»
Бактерии, поглощенные амебой, попадают в специальный пищеварительный отдел, окруженный мембраной. «Наши результаты показывают, что бактерии способны вонзать кинжал в мембрану пищеварительного отдела амебы», — говорит Дезире Бёк, также докторант в группе Пилхофера и ведущий автор исследования, опубликованного в журнале 9.7846 Наука . Это приводит к разрушению компартмента, который является негостеприимной средой для бактерий, и высвобождению бактерий. Оказавшись вне пищеварительного отдела, но все еще внутри амебы, бактерии могут выживать и даже размножаться.
Процесс разрушения пищеварительного отдела еще не известен. «Возможно, разрыв мембраны происходит исключительно по механическим причинам», — говорит Пилхофер. Однако не исключено, что кинжалы бактерий Amoebophilus пропитаны чем-то вроде яда для стрел — мембраноразрушающими ферментами. Чертежи таких ферментов содержатся в геноме бактерий, как смогли показать Матиас Хорн, профессор Венского университета, и его коллеги.
Прецизионная фрезеровка
Ученые применили совершенно новый метод, используемый лишь несколькими исследовательскими лабораториями по всему миру, включая лабораторию Pilhofer, для определения трехмерной структуры кинжалов и их стреляющих механизмов с высоким разрешением. Бёк заморозил амеб после того, как они поглотили бактерии при температуре минус 180°С.
Подобно палеонтологу, использующему молоток и долото для отделения окаменелостей от камня, Медейрос затем использовал сфокусированный ионный луч в качестве «нано-долота» для работы с замороженными образцами. С впечатляющей точностью он смог размолоть амебу и большую часть бактерии, раскопав молекулярные кинжалы и их стреляющие устройства, чтобы, наконец, получить трехмерную электронную томограмму.
Первое изображение общей структуры
Системы, связанные с микрокинжалами, встречаются и в других областях биологии: вирусы, специализирующиеся на заражении бактерий (бактериофаги), используют такие системы для внедрения своего генома в микроорганизмы. Некоторые бактерии могут даже выпускать подобные микроустройства в окружающую среду для борьбы с конкурирующими микроорганизмами.
Ученые впервые представили полную пространственную структуру стреляющего механизма внутри клетки в ее естественном контексте. Они также впервые показывают детали опорной плиты и мембранного анкера. «В прошлом клеточные биологи исследовали функции таких систем, а структурные биологи выясняли структуру отдельных компонентов», — говорит Пилхофер. «Благодаря технологиям криофокусированного ионно-лучевого измельчения и электронной криотомографии, которые мы внедрили в ETH Zurich, мы теперь можем сократить разрыв между клеточной биологией и структурной биологией».
Многоствольные ружья
Микрокинжалы ранее встречались только как отдельные устройства. Однако у амебофилов ученые из Цюриха и Вены обнаружили устройства, которые встречаются группами до 30 штук. «Можно назвать их многоствольными ружьями», — говорит Пилхофер.
Исследователи также использовали геномные сравнения, чтобы выяснить, как Amoebophilus развил свои кинжалы. «Соответствующие гены очень похожи на гены инъекционных систем бактериофагов», — говорит Пилхофер. «Мы предполагаем, что гены предков сегодняшних бактериофагов давно утвердились в геноме бактерий».
Также присутствует у других бактерий
Сравнение геномов позволяет предположить, что микрокинжалы встречаются не только у Amoebophilus, но и у многих других видов бактерий по меньшей мере из девяти наиболее важных групп бактерий. Исследователям еще предстоит выяснить, используют ли эти бактерии свои кинжалы, чтобы избежать переваривания амебами, или кинжалы служат совсем другим целям. У них работа давно отрезана.
Наконец, ученые хотели бы использовать новый метод измельчения криофокусированного ионного пучка для выяснения структуры других сложных молекулярных систем. «Эта методика может помочь в решении многих других вопросов клеточной, инфекционной и структурной биологии. Мы уже работаем с другими исследовательскими группами и предлагаем им наш опыт», — говорит Медейрос.
Узнать больше
Бактерии чумы укрываются в амебах
Дополнительная информация: «Архитектура, функции и эволюция сократительной системы инъекций на месте» Science (2017). science.sciencemag.org/lookup/ … 1126/science.aan7904
Информация журнала: Наука
Предоставлено
Высшая техническая школа Цюриха
Цитата :
Бактерии протыкают амеб микрокинжалами (2017, 17 августа)
получено 24 сентября 2022 г.
Диагностика возбудителя, резервуаров и путей передачи] В.3
Appl Environ Microbiol
75
:
3206
–
3215
.
Сиддики
Р.
Хан
Н/Д
(
2012
)
Акантамеба – эволюционный предок макрофагов: миф или реальность?
Exp Parasitol
130
:
95
–
97
.
Зигеле
Д.А.
Колтер
Р.
(
1992
)
Жизнь после бревна
.
J Бактериол
174
:
345
–
348
.
Skandamis
П.Н.
Stopforth
J.D.
Ashton
L.V.
Георнарас
I.
Кендалл
П.А.
Sofos
J. N.
(
2009
)
Escherichia coli O157:H7 Выживаемость, образование биопленки и кислотоустойчивость в условиях, имитирующих бойню, во влажных и сухих условиях
.
Пищевой микробиол
26
:
112
–
119
.
Снедекер
К.Г.
Шоу
Д.Дж.
Замок
M.E.
Prescott
R.J.
(
2009
)
Первичные и вторичные случаи в Escherichia coli Вспышки O157: статистический анализ
.
BMC Infect Dis
9
:
144
.
Соломон
Э. Б.
Потенски
Си Джей
Мэтьюз
К.Р.
(
2002a
)
Влияние метода орошения на передачу и устойчивость Escherichia coli O157:H7 на листьях салата
.
J Food Prot
65
:
673
–
676
.
Соломон
Э.Б.
Ярон
С.
Мэтьюз
К.Р.
(
2002b
)
Передача Escherichia coli O157:H7 из загрязненного навоза и поливной воды в ткани растений салата и их последующая интернализация
.
Appl Environ Microbiol
68
:
397
–
400
.
Стейнберг
К.М.
Левин
Б.Р.
.
Proc Biol Sci
274
:
1921
–
1929
.
Steinert
M.
Birkness
K.
White
E.
Fields
B.
Quinn
F.
(
1998
)
Бациллы Mycobacterium avium растут сапрозоически в совместной культуре с Acanthamoeba polyphaga и выживают в стенках кисты
.
Appl Environ Microbiol
64
:
2256
–
2261
.
Стиги
К.А.
Макдональд
Дж.К.
Теллез-Марфин
А. А.
Лофи
К.Х.
.
Emerg Infect Dis
18
:
477
–
479
.
Sugumar
G.
Chrisolite
B.
Velayutham
P.
Selvan
A.
Рамеш
U.
.
J Environ Biol
29
:
387
–
391
.
Bausch
C.
Richmond
C.
Blattner
F.R.
Конвей
Т.
(
1999
)
Функциональная геномика: анализ экспрессии Escherichia coli , растущих на минимальной и богатой средах
.
J Бактериол
181
:
6425
–
6440
.
Thompson
L.H.
Giercke
S.
Beaudoin
C.
Woodward
D.
Wylie
J.L.
(
2005
)
Усиленный эпиднадзор за штаммами Escherichia coli , не продуцирующими O157 веротоксин, в образцах стула человека из Манитобы
.
Can J Infect Dis Med Microbiol
16
:
329
–
334
.
Торрес
А.Г.
Хирон
Дж.А.
Перна
Н.Т.
Бурланд
В.
Блаттнер
Ф.Р.
Авелино-Флорес
Ф.
Капер
Дж.Б.
(
2002
)
Идентификация и характеристика lpfABCC’DE, фимбриального оперона энтерогеморрагической Escherichia coli O157:H7
.
Infect Immun
70
:
5416
–
5427
.
Torres
A.G.
Jeter
C.
Langley
W.
Matthysse
A.G.
3
3 (
2005
)
Дифференциальное связывание Escherichia coli O157:H7 с люцерной, эпителиальными клетками человека и пластиком опосредуется различными поверхностными структурами
.
Appl Environ Microbiol
71
:
8008
–
8015
.
Улих
Г.А.
Кук
П.Х.
Соломон
Э.Б.
(
2006
)
Анализ фенотипа красно-сухой-шероховатости штамма Escherichia coli O157:H7 и его роли в формировании биопленки и устойчивости к антибактериальным препаратам
.
Appl Environ Microbiol
72
:
2564
–
2572
.
Улих
Г.А.
Роджерс
Д.П.
Мосье
Д.А.
(
2010
)
Escherichia coli серотип O157:H7 Удержание на твердых поверхностях и устойчивость к перекиси усиливаются за счет образования биопленки из двойного штамма
.
Патог пищевого происхождения Dis
7
:
935
–
943
.
Vally
H.
Hall
G.
Dyda
A.
Raupach
J.
Knope
K.
Combs
B.
6 DESMARCHLIE
(
2012
)
Эпидемиология шига-токсина, продуцирующего Escherichia coli в Австралии, 2000–2010 гг.
.
BMC Public Health
12
:
63
.
VanBogelen
Р.А.
Олсон
Э.Р.
Ваннер
Б.Л.
Нейдхардт
Ф.К.
(
1996
)
Общий анализ белков, синтезированных при ограничении фосфора в Escherichia coli
.
J Бактериол
178
:
4344
–
4366
.
Варма
Дж.К.
Грин
К.Д.
Реллер
ME
и другие. . (
2003
)
Вспышка инфекции Escherichia coli O157 после контакта с зараженным зданием
.
JAMA
290
:
2709
–
2712
.
Vital
M.
Hammes
F.
Egli
T.
(
2008
)
Escherichia coli O157 может расти в природной пресной воде при низкой концентрации углерода
.
Environ Microbiol
10
:
2387
–
2396
.
Уэйд
Т.Дж.
Кальдерон
RL
Sams
E.
Beach
M.
Brenner
K.P.
Williams
A.H.
Dufour
AP
(
2006
)
Быстро измеряемые показатели качества рекреационной воды позволяют прогнозировать желудочно-кишечные заболевания, связанные с плаванием
.
Environment Health Perspect
114
:
24
–
28
.
Ван
Г.
Дойл
М.П.
(
1998
)
Выживаемость энтерогеморрагической Escherichia coli O157:H7 в воде
.
J Food Prot
61
:
662
–
667
.
Ван
А.
Сон
Б.Э.
Лисс
С.Н.
(
2011
)
Влияние роста биопленки на перенос Escherichia coli O157:H7 в песке
.
Подземные воды
49
:
20
–
31
.
Уорбертон
Д.У.
Остин
Дж.В.
Харрисон
Б.Х.
Сандерс
Г.
(
1998
)
Выживание и восстановление Escherichia coli O157:H7 в инокулированной бутилированной воде
.
J Food Prot
61
:
948
–
952
.
Watterworth
L.
Rosa
B.
Schraft
H.
Topp
E.
Leung
K.
(
2006
)
Выживаемость различных ERIC-генотипов шига-токсин-продуцирующих Escherichia coli в колодезной воде
.
Вода Воздух Почва Загрязнение
177
:
367
–
382
.
Werner
M.
Maschek
H.J.
Kaloutsi
V.
Buhr
T.
Kausche
F.
Delventhal
S.
Georgii
A.
(
1990
)
Цитогенетическое исследование клеток костного мозга при заболеваниях стволовыми клетками при миелодиспластических синдромах
.
Verh Dtsch Ges Pathol
74
:
139
–
143
.
Wieler
L.H.
Vieler
E.
ERPENSTEIN
C.
Schlapp
T.
Steinruck
H.
Bauerfeind
. R.
H.
Bauerfeind
. R.
6
.
6
.
6
.
6
.
H.
69
.0003
А.
Бальер
Г.
(
1996
)
Шига-токсин-продуцирующие штаммы Escherichia coli от крупного рогатого скота: ассоциация адгезии с носительством eae и других генов
.
J Clin Microbiol
34
:
2980
–
2984
.
Уильямс
Л.Д.
Гамильтон
P.S.
Уилсон
Б.В.
Estock
MD
(
1997
)
Вспышка Escherichia coli 0157:H7: длительное выделение и передача от человека человеку в детском саду
.
J Environment Health
59
:
9
.
Xicohtencatl-Cortes
J.
Sanchez Chacon
E.
Saldana
Z.
Фрир
Э.
Хирон
Дж.А.
(
2009
)
Взаимодействие Escherichia coli O157:H7 с листовой зеленью
.
J Food Prot
72
:
1531
–
1537
.
Черный
Р.Л.
Чирилло
Дж.Д. (
2004
)
Доказательства того, что hsp90 участвует в измененных взаимодействиях вариантов Acanthamoeba castellanii с бактериями
.
Эукариотическая клетка
3
:
567
–
578
.
Yoon
Y.
Sofos
J.N.
(
2008
)
Активность аутоиндуктора-2 грамотрицательных пищевых патогенных бактерий и ее влияние на формирование биопленок
.
J Food Sci
73
:
M140
–
M147
.
Yoshida
Y.
Sugiyama
S.
Oyamada
T.
Yokoyama
K.
Makino
K.
(
2012
)
Новые члены фосфатного регулона в Escherichia coli O157:H7, идентифицированные с помощью полногеномного подхода
.
Гена
502
:
27
–
35
.
Copyright © 2013 Федерация европейских микробиологических обществ. Опубликовано Blackwell Publishing Ltd. Все права защищены.
Copyright © 2013 Federation of European Microbiological Societies. Опубликовано Blackwell Publishing Ltd. Все права защищены
| Влияние эволюции Дарвин изменил все. Публикация его работы «Происхождение видов » в 1859 году бросила всю биологическую науку в новую парадигму, включая изучение теории классификации и принципов таксономии. Используя логику в качестве основы своей работы, и Аристотель, и Линней разработали свои схемы классификации на таксономических принципах, которые в своей основе были произвольными. Их группы, хотя и были логичны, не основывались на каких-либо очевидных отношениях биологической природы. Это были удобные группы, которые люди могли быстро увидеть, идентифицировать и использовать. Это было приемлемо, потому что (а) никто не мог придумать ничего лучше, и (б) большинство людей в то время верили в концепцию «фиксированных видов», согласно которой организм был создан в его нынешнем виде и никогда не мог измениться. После Дарвина стало понятно, что организмы действительно могут изменяться и что все нынешние формы живых существ пришли к этой форме путем изменений и естественного отбора, механизма эволюции. Ученые начали строить филогении , списки или диаграммы, показывающие эволюционные пути, пройденные популяциями организмов на протяжении многих поколений и в течение длительных периодов времени. Эти филогенетические диаграммы быстро стали напоминать деревья, поскольку стало понятно, что предковые линии иногда распадались, разветвлялись и превращались в два или более разных вида, которые впоследствии могли разветвляться снова и снова. Филогенетическое древо было немного похоже на генеалогическое древо, показывающее, кем были ближайшие родственники, у кого был общий предок и когда. Организмы были связаны друг с другом, и эти отношения могли лечь в основу таксономии нового типа; на основе эволюционного происхождения и эволюционного родства. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Из этого принципа возникли два основных типа таксономии. Они похожи, но есть серьезные различия. Эволюционная систематика делает попытку построить деревья, которые точно показывают филетические линии (правильное ветвление на генеалогическом древе), наряду с рассмотрением того, когда и как новые виды возникли и переместились в новые места обитания и ниши (установил «новый» способ жизнь, а не какое-то тривиальное изменение характера). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Кладистика , однако, игнорирует, когда и где возникает ветвь, пытается использовать чисто объективные критерии и определяет каждую точку ветвления фундаментальным признаком эволюционного значения. Оба метода имеют свои сильные и слабые стороны. Кладистика получила свое название от ветвей генеалогического древа, которые называются кладами . Кладограмма представляет собой стилизованную диаграмму, которая выглядит как ряд Y или разветвлений на дороге. На каждой ветви или Y-образном стыке используются новые символы эволюционного происхождения, чтобы отделить одну группу от остальных. Кладограммы могут быть построены для любой группы организмов. Например, следующие организмы представляют собой набор, из которого можно составить кладограмму; кенгуру, дождевой червь, амеба, ящерица, кошка, губка и лосось. Каждое из этих существ имеет эволюционную связь друг с другом. Все они имеют общее происхождение, и все их нынешние формы произошли от разветвленных событий где-то в филогенетическом прошлом. Вопрос в том, когда произошли эти ответвления? Процесс построения кладограммы начинается с данных; таблица черт или характеристик, которые развились или были является производным в процессе эволюции. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
На следующем этапе каждый из организмов сравнивается, чтобы увидеть, имеют ли они общие черты или производные признаки. Используя эти образцы общих производных признаков, кладограмма может быть построена как серия Y или ветвей. На каждой ветви один из организмов, не имеющий общих признаков с остальной группой, «ответвляется» в свою собственную кладу. Порядок или последовательность этих ветвей зависит от того, сколько символов осталось в большей группе. Посмотреть на себя. Наведите курсор на кнопки управления рядом с групповой диаграммой и реконструируйте процесс создания кладограммы: | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Кладограммы — полезный способ визуальной организации отношений между существами, имеющими и не имеющими общих производных признаков. На практике, когда строятся кладограммы, возможно, придется учитывать многие сотни символов, и нужны компьютеры, чтобы выбрать наилучшее соответствие между ветвями и тем, кто должен быть на них. Кладограммы подчеркивают последовательность или порядок, в котором производные признаки возникают из центрального филогенетического дерева. В этом их главная сила. Однако ничто в кладограмме не указывает на то, насколько сильным или глубоким является производный признак, и на его эволюционную важность. Всем используемым символам придается равный вес. Иногда это может приводить к необычным группировкам, которые могут быть технически правильными, но сомнительными. Эволюционная систематика использует таксономический принцип, который также основан на разделении филетических родословных, и строит генеалогические деревья, но, в отличие от чисто кладистического метода, используется система взвешивания, которая отдает предпочтение одним производным признакам по сравнению с другими. Тем не менее, и это большое «однако», эволюционные систематики действительно считают необходимым использовать много субъективных суждений при принятии решения о том, каким факторам придавать наибольшее значение. Всякий раз, когда люди заменяют своими собственными «чувствами» доказуемый биологический факт, к результатам всегда нужно относиться с осторожностью и постоянно пересматривать. Однако даже при такой осторожности эволюционная систематика, по-видимому, в настоящее время дает картину биологических отношений между видами, основанную на самых надежных таксономических принципах. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Спасти амебу? | Среди величественных деревьев
Следует ли более непосредственно учитывать микробное биоразнообразие (то есть микроскопические организмы, такие как бактерии, археи, протисты и некоторые грибы) в усилиях по сохранению? Несомненно, важность микроскопических организмов в земной системе невозможно переоценить, поскольку они играют жизненно важную роль в глобальных циклах элементов – фиксируют атмосферный азот и делают его доступным для растений, возвращают азот обратно в атмосферу, играют важную роль в пищевых цепях и способствуют разложение органических веществ. Кроме того, в будущем микроорганизмы, вероятно, станут важнейшим источником новых фармацевтических препаратов (Chivian & Bernstein, 2008). Однако микроорганизмы лишь изредка включались в исследования биоразнообразия, и хотя Международный союз охраны природы (МСОП) недавно добавил четырех находящихся под угрозой исчезновения простейших в Красный список (здесь, здесь, здесь и здесь), общественный резонанс по поводу их защита кажется маловероятной в ближайшее время.
Пластинка из монографии Джозефа Лейди 1879 года о раковинных амебах (1–8: Nebela ansata, 9–14: Nebela hippocrepis). Раковинные амебы представляют собой группу протистов, которые производят морфологически различные раковины.
Конечно, отсутствие акцента на микроорганизмах в природоохранной биологии не особенно удивительно, учитывая, что 1) мы относительно мало знаем о микробном разнообразии, 2) микробы, возможно, немного менее симпатичны и харизматичны, чем многие другие группы организмов (хотя они, безусловно, симпатичнее, чем некоторые виды), и 3) широко распространено мнение, что микробные виды имеют космополитическое распространение. Другими словами, они встречаются повсюду. В частности, микробный вид встречается везде, где существует подходящая среда обитания для этого конкретного вида. В 1934, Баас Бекинг просто сформулировал эту гипотезу так: «все есть везде, но среда выбирает» (де Вит и Бувье, 2006). Эта гипотеза подразумевает, что рассредоточение не ограничивает распространение микробов, и эндемизм (т. е. приуроченность к определенному региону) должен быть редким явлением. Конечно, если все есть везде, то нам не нужно сильно беспокоиться о каком-либо конкретном виде микробов, потому что вымирание крайне маловероятно, если только не произойдет уничтожение всех подходящих местообитаний… по всей планете. Будем надеяться, что это останется маловероятным в течение некоторого времени.
Однако исследования теперь ясно показывают, что микробная биогеография является более тонкой и сложной, чем подразумевается гипотезой «все повсюду». За последние пару десятилетий появилось множество примеров ограниченных географических ареалов. Некоторые особенно яркие примеры относятся к панцирным амебам, группе протистов, производящих раковины, которые относительно хорошо изучены, по крайней мере, по сравнению со многими другими микробными группами. Например, Apodera vas и виды рода Certesella ограничены южным полушарием и тропиками, хотя подходящие местообитания для этих видов существуют в Евразии и Северной Америке (Smith and Wilkinson 2007). Модели предполагают, что эти виды произошли на континенте Гондвана через некоторое время после того, как он отделился от Лавразии около 200-180 миллионов лет назад, и, таким образом, их нынешнее распространение может быть отражением этой геологической и эволюционной истории (Смит и Уилкинсон, 2007). По-видимому, эти виды не смогли рассеяться в Северную Америку или Евразию.
Палеогеографическая реконструкция Земли в позднем триасе, незадолго до отделения Гондваны (южные континенты) от Лавразии (северные континенты). Нынешнее распространение Apodera vas и видов рода Certesella почти ограничено территорией, которая когда-то была частью Гондваны, что позволяет предположить, что они эволюционировали на этом массиве суши (Smith & Wilkinson 2007). (Изображение с сайта wikipedia.org).
Потенциально более ограниченный шаблон распределения — Nebela ansata , необычный (и осмелюсь сказать, «харизматичный») вид раковинных амеб, известный лишь в нескольких местах на востоке Северной Америки. В 1874 году Джозеф Лейди впервые столкнулся с этим видом из коллекций, которые он собрал в сосновых степях Нью-Джерси (Leidy 1879). Со времени его первоначальных описаний этот вид упоминался в литературе только три раза, всегда в умеренном поясе восточной части Северной Америки и всегда обитающий в заболоченных местах на влажном мхе Sphagnum . Этот вид не упоминается в литературе с начала 19 века.50-х годов, несмотря на интенсивный сбор образцов сообществ раковинных амеб водно-болотных угодий (Heger et al. 2011).
В поисках забытого микроба
Nebela ansata из болота Webb’s Mill Bog в сосновых степях Нью-Джерси. Для вида характерны роговидные боковые отростки. (Фото: Тьерри Хегер)
Сосновые степи Нью-Джерси находятся всего в нескольких часах езды от Университета Лихай, поэтому несколько лет назад мы решили посмотреть, сможем ли мы найти неуловимого и давно забытого микроба Лейди. Его первоначальные образцы были взяты из кедровых болот недалеко от города Абсеком (теперь пишется как Абсекон), и мы использовали это в качестве ориентира при выборе потенциальных заболоченных мест для наших усилий по отбору проб. Он также упомянул, что Nebela ansata обычно встречается вместе с Nebela carinata , более широко распространенным видом. Экология Nebela carinata хорошо изучена, он предпочитает очень влажный Sphagnum мох и часто встречается в изобилии там, где уровень грунтовых вод находится всего на несколько сантиметров ниже поверхности мха. Мы использовали эти знания для определения вероятных местообитаний в каждом из отобранных нами водно-болотных угодий. Маура Салливан (Университет Лихай, аспирант) и я собирали образцы в течение двух полных дней, вернули образцы в лабораторию для исследования и сумели найти вид!
Интересно, что примерно в то же время, когда мы повторно открыли Nebela ansata в сосновых степях Нью-Джерси, этот вид был также обнаружен в Новой Шотландии Барри Уорнером и Таро Асада (Университет Ватерлоо). Вскоре сформировались совместные усилия с целью собрать воедино всю известную информацию о распространении, экологии и филогении видов. Под руководством Тьерри Хегера, в то время аспиранта (Швейцарский федеральный исследовательский институт WSL и Женевский университет), был проведен исчерпывающий обзор литературы и музеев, который использовался в тандеме с анализом образцов из Новой Шотландии и Сосновых степей, чтобы дать представление о биогеография и экология Небела ансата . В совокупности результаты представляют собой убедительный случай микроорганизма с очень ограниченным диапазоном распространения — по-видимому, ограниченным умеренным климатом восточной части Северной Америки — что представляет собой явное исключение из гипотезы «все есть везде» (Heger et al. 2011).
Водно-болотные угодья Уэббс-Милл в сосновых степях Нью-Джерси, где были обнаружены особи редкого вида раковинных амеб, Nebela ansata.
Сколько других микроорганизмов также имеет географически ограниченное распространение? Каковы основные причины этих ограниченных дистрибутивов? Сколько эндемичных видов остаются неописанными? Какую роль играют эти относительно редкие микроорганизмы в экосистемах? Сколько микробных видов уязвимы перед разрушением среды обитания или другими глобальными изменениями? Многое еще предстоит сделать в области изучения биоразнообразия.
-rkb-
Процитированная литература
Chivian, E. & A. Bernstein, eds. 2008. Поддержание жизни. Как здоровье человека зависит от биоразнообразия. Издательство Оксфордского университета.
де Вит. Р. и Т. Бувье. 2006. «Все есть везде, но среда выбирает»; что на самом деле сказали Баас Бекинг и Бейеринк? Экологическая микробиология 8: 755-758.
Хегер, Т.Дж., Р.К. Бут, М.Э. Салливан, Д.М. Уилкинсон, Б.Г. Уорнер, Т. Асада, Ю. Мазей, Р. Мейстерфельд и Э.А.Д. Митчелл. 2011. Новое открытие Nebela ansata (Amoebozoa: Arcellinida) в восточной части Северной Америки: биогеографические последствия. Журнал биогеографии 38: 1897-1906.
Leidy, J. 1879. Пресноводные корненожки Северной Америки. Отчет Геологической службы территорий США, 12, 1–324.
Смит, Х.Г. и Д.М. Уилкинсон. 2007. Не все свободноживущие микроорганизмы имеют космополитическое распространение — случай Nebela ( Apodera ) vas Certes (Protozoa: Amoebozoa: Arcellinida). Журнал биогеографии, 34: 1822–1831.
Нравится:
Нравится Загрузка…
Бактерии пронзают амеб микрокинжалами
Группа подпружиненных кинжалов внутри бактерии. Зеленый показывает их в «заряженном» виде, красный — после того, как кинжал был запущен. Кредит: Лео ПоповичБактерии должны остерегаться амебы. Голодные амебы охотятся на них: они ловят их своими псевдоподиями, а затем поглощают и переваривают.
Однако некоторые бактерии умеют защищаться. Одним из таких является Amoebophilus, который был обнаружен исследователями Венского университета несколько лет назад. Эта бактерия может не только выжить внутри амебы, но и процветать: амеба стала ее излюбленной средой обитания.
Вместе с венскими первооткрывателями бактерии ученые из ETH Zurich обнаружили механизм, который, по их мнению, имеет решающее значение для выживания Amoebophilus внутри амебы. У бактерии есть устройства для стрельбы микрокинжалами. Он может использовать кинжалы, чтобы пронзить амебу изнутри и таким образом избежать пищеварения.
Побег из кишечника амебы
Стреляющий механизм состоит из оболочки, прикрепленной к внутренней мембране бактерии с помощью базовой пластины и якорной платформы. Жоао Медейрос, аспирант группы профессора Мартина Пилхофера в ETH, объясняет механизм: «Оболочка подпружинена, и микрокинжал лежит внутри нее. Когда оболочка сжимается, кинжал очень быстро выбрасывается наружу через бактериальную мембрану. .»
Бактерии, поглощенные амебой, попадают в специальный пищеварительный отдел, окруженный мембраной. «Наши результаты показывают, что бактерии способны вонзать кинжал в мембрану пищеварительного отдела амебы», — говорит Дезире Бёк, также докторант в группе Пилхофера и ведущий автор исследования, опубликованного в журнале 9.7846 Наука . Это приводит к разрушению компартмента, который является негостеприимной средой для бактерий, и высвобождению бактерий. Оказавшись вне пищеварительного отдела, но все еще внутри амебы, бактерии могут выживать и даже размножаться.
Процесс разрушения пищеварительного отдела еще не известен. «Возможно, разрыв мембраны происходит исключительно по механическим причинам», — говорит Пилхофер. Однако не исключено, что кинжалы бактерий Amoebophilus пропитаны чем-то вроде яда для стрел — мембраноразрушающими ферментами. Чертежи таких ферментов содержатся в геноме бактерий, как смогли показать Матиас Хорн, профессор Венского университета, и его коллеги.
Прецизионная фрезеровка
Ученые применили совершенно новый метод, используемый лишь несколькими исследовательскими лабораториями по всему миру, включая лабораторию Pilhofer, для определения трехмерной структуры кинжалов и их стреляющих механизмов с высоким разрешением. Бёк заморозил амеб после того, как они поглотили бактерии при температуре минус 180°С.
Подобно палеонтологу, использующему молоток и долото для отделения окаменелостей от камня, Медейрос затем использовал сфокусированный ионный луч в качестве «нано-долота» для работы с замороженными образцами. С впечатляющей точностью он смог размолоть амебу и большую часть бактерии, раскопав молекулярные кинжалы и их стреляющие устройства, чтобы, наконец, получить трехмерную электронную томограмму.
Первое изображение общей структуры
Системы, связанные с микрокинжалами, встречаются и в других областях биологии: вирусы, специализирующиеся на заражении бактерий (бактериофаги), используют такие системы для внедрения своего генома в микроорганизмы. Некоторые бактерии могут даже выпускать подобные микроустройства в окружающую среду для борьбы с конкурирующими микроорганизмами.
Ученые впервые представили полную пространственную структуру стреляющего механизма внутри клетки в ее естественном контексте. Они также впервые показывают детали опорной плиты и мембранного анкера. «В прошлом клеточные биологи исследовали функции таких систем, а структурные биологи выясняли структуру отдельных компонентов», — говорит Пилхофер. «Благодаря технологиям криофокусированного ионно-лучевого измельчения и электронной криотомографии, которые мы внедрили в ETH Zurich, мы теперь можем сократить разрыв между клеточной биологией и структурной биологией».
Многоствольные ружья
Микрокинжалы ранее встречались только как отдельные устройства. Однако у амебофилов ученые из Цюриха и Вены обнаружили устройства, которые встречаются группами до 30 штук. «Можно назвать их многоствольными ружьями», — говорит Пилхофер.
Исследователи также использовали геномные сравнения, чтобы выяснить, как Amoebophilus развил свои кинжалы. «Соответствующие гены очень похожи на гены инъекционных систем бактериофагов», — говорит Пилхофер. «Мы предполагаем, что гены предков сегодняшних бактериофагов давно утвердились в геноме бактерий».
Также присутствует у других бактерий
Сравнение геномов позволяет предположить, что микрокинжалы встречаются не только у Amoebophilus, но и у многих других видов бактерий по меньшей мере из девяти наиболее важных групп бактерий. Исследователям еще предстоит выяснить, используют ли эти бактерии свои кинжалы, чтобы избежать переваривания амебами, или кинжалы служат совсем другим целям. У них работа давно отрезана.
Наконец, ученые хотели бы использовать новый метод измельчения криофокусированного ионного пучка для выяснения структуры других сложных молекулярных систем. «Эта методика может помочь в решении многих других вопросов клеточной, инфекционной и структурной биологии. Мы уже работаем с другими исследовательскими группами и предлагаем им наш опыт», — говорит Медейрос.
Узнать больше
Бактерии чумы укрываются в амебах
Дополнительная информация: «Архитектура, функции и эволюция сократительной системы инъекций на месте» Science (2017). science.sciencemag.org/lookup/ … 1126/science.aan7904
Информация журнала: Наука
Предоставлено Высшая техническая школа Цюриха
Цитата :
Бактерии протыкают амеб микрокинжалами (2017, 17 августа)
получено 24 сентября 2022 г.