Полная систематика амебы протей по схеме: Царство Тип Отряд Семейство Род Вид
помогите пожалуйста очень срочно
можете помочь пожалуйста срочно
Помогите пожалуйста!! Срочно! Ответьте на вопросы: качественно и правильно!!!А10.Представителями семейства Розоцветных являются растения:а) люпин, гор … ох, чечевица; в) вишня, рябина, земляника;б) рис, овёс, кукуруза; г) баклажан, томат, белена.A2. В чём преимущества полового размножения над бесполым? Ответ обоснуйте.
Помогите пожалуйста!! Даю много баллов!!!
А1. Из проросшей споры мха образуется тонкая зелёная нить:
а) зигота; б) заросток; в) предросток; г) зооспо
… ра.
А2. Могут размножаться отводками:
а) смородина, тополь; *
в) одуванчик, малина;
б) осина, иван-чай; г) крыжовник, яблоня.
А3. Могут размножаться листовыми черенками:
а) сансевьера, сенполия; в) земляника, клюква;
б) ландыш, пырей; г) нарцисс, тюльпан.
А4. Могут размножаться клубнями:
а) яблоня, груша; в) топинамбур, картофель;
б) орхидея, жень-шень; г) роза, ива.
Почему после мастурбации, происходят такие мысли, как «Зачем я это слелал «,»Лучше бы я этого не делал «.Возбуждения прекращаются и будто мой мозг х … орошо работает, начал быстро думать ?
перечислети органы чувств расположенные на голове млекопитающих
на рисунке надпишите названия составных частей кожаного покрова млекопитающих. какие функции они выполняют
какие функции они выполняют
Как решить????????????
Что лишает или вредно ли мастурбация?
9. Из шести предложенных выберите и отметьте три озна стинктивнои поведенческой реакции 1) базируется на безусловных рефлексах и присущая животные от … на родження 2) требует предварительного научения 3) связана с нормальным функционированием его органов 4) хотя и зависит от строения организма, но не обусловленная ею 5) позволяет животному сразу после рождения быть готовым к жиз ни в определенных условиях окружающей среды 6) реализуется в разных особей одного вида различными способами на под пруду предыдущего опыта, впечатлений и т.д. Задача 10. Дайте развернутый ответ на вопрос. Hony поведение животных не может быть только врожденной? @meyifff
Простейшие. Систематика простейших — презентация онлайн
1. Урок по биологии на тему: «Простейшие».
2. •Систематика простейших
Подцарство:Простейшие
Тип: Саркодовые
Тип: Жгутиконосцы
Класс: Голые амёбы
Класс: Эвгленовые
Тип: Инфузории
3.
 Подцарство: Простейшие Обитают простейшие организмы обычно во влажной
Подцарство: Простейшие Обитают простейшие организмы обычно во влажнойсреде – водоемах, почве. Паразитические простейшие
используют в качестве среды обитания многоклеточные
живые организмы, в том числе и человека.
Некоторые простейшие организмы даже имеют
раковины или причудливые выросты клеточной стенки.
Он состоит из многих классов, отрядов, семейств и вклю
чает примерно 20—25 тыс. видов.
По своему строению простейшие чрезвычайно разнооб
разны. Подавляющее большинство их обладает
микроскопически малыми размерами, для изучения их
приходится пользоваться микроскопом.
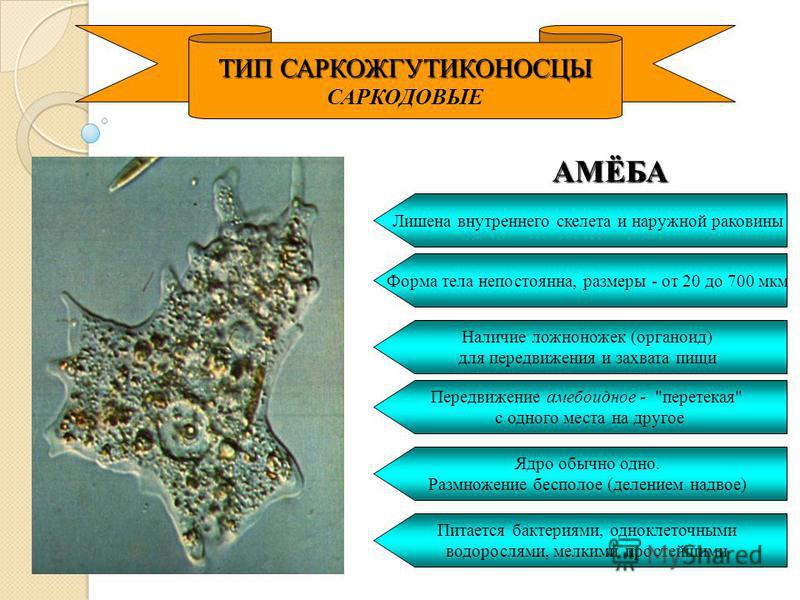
4. Тип: Саркодовые
Все саркодовые – достаточно примитивныеорганизмы со слабой дифференциацией
цитоплазмы и наружной мембраны.
Корненожки (Rhyzopoda) отличаются
наличием ложноножек – выростов
цитоплазмы, образующихся в разных частях
клетки, благодаря которым они движутся и
захватывают пищу. Типичным
представителем корненожек является
амёба-протей.
Клетка может содержать хлорофилл и
светочувствительный глазок.
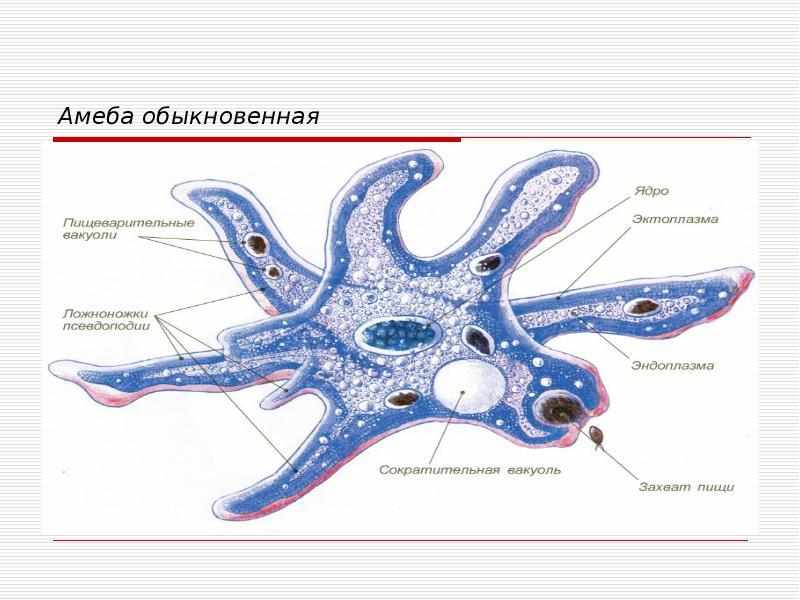
5. Класс: Голые амёбы
Внешнееамёбы:
строение обыкновенной
6. Класс: Голые амёбы
Тело амебы представляет собой комочек полужидкойцитоплазмы с ядром посередине.
Вся цитоплазма подразделена на два слоя: наружный, вязкий —
эктоплазму и внутренний, гораздо более жидкий —
эндоплазму.
У амебы нет твердой оболочки, и она способна изменять
форму тела.
Когда амеба ползет по листу водного растения, у нее в том
направлении, куда она двигается, образуются выпячивания
Псевдоподиями она охватывает бактерию или
микроскопическую водоросль, вскоре добыча оказывается
внутри тела амебы, и вокруг нее образуется пузырек —
пищеварительная вакуоля.
С помощью сократительной вакуоли амёба выбрасывает
наружу воду.
7. Класс: Голые амёбы
Движение:Амёба
«перетекает» по дну.
 На теле
На телепостоянно образуются меняющие свою
форму выросты – псевдоподии
(ложноножки). В один из таких выступов
постепенно переливается цитоплазма,
ложная ножка в нескольких точках
прикрепляется к субстрату и
происходит передвижение.
8. Класс: Голые амёбы
Питание:Передвигаясь, амёба наталкивается на
одноклеточные водоросли, бактерии, мелкие
одноклеточные, «обтекает» их и включает в
цитоплазму, образуя пищеварительную вакуоль.
Ферменты, расщепляющие белки, углеводы и
липиды, поступают внутрь пищеварительной
вакуоли, и происходит внутриклеточное
пищеварение. Пища переваривается и
всасывается в цитоплазму. Способ захвата пищи с
помощью ложных ножек называется фагоцитозом.
9. Класс: Голые амёбы
Дыхание:Кислород
расходуется на клеточное дыхание.
Когда его становится меньше, чем во внешней
среде, новые молекулы проходят внутрь клетки.
Молекулы углекислого газа и вредных веществ,
накопившихся в результате жизнедеятельности,
наоборот, выходят наружу.

10. Класс: Голые амёбы
Выделение:Пищеварительная
вакуоль подходит к клеточной
мембране и открывается наружу, чтобы
непереваренные остатки выбросить наружу в
любом участке тела. Жидкость поступает в тело
амёбы по образующимся тонким трубковидным
каналам, путём пиноцитоза. Откачиванием
лишней воды из организма занимаются
сократительные вакуоли. Они постепенно
наполняются, а раз в 5-10 минут резко
Вакуоли могут возникать в любой части клетки.
11. Класс: Голые амёбы
Размножение:Амёбы размножаются только бесполым путём.
Выросшая амёба приступает к размножению. Оно
происходит путём деления клетки. До деления клетки ядро
удваивается, чтобы каждая дочерняя клетка получила свою
копию наследственной информации (1). Размножение
начинается с изменения ядра. Оно вытягивается (2), а затем
постепенно удлиняется (3,4) и перетягивается посредине.

Поперечной бороздкой делится на две половинки, которые
расходятся в разные стороны – образуются два новых ядра.
Тело амёбы разделяется на две части перетяжкой и
образуется две новые амёбы. В каждую из них попадает по
одному ядру (5). Во время деления происходит образование
недостающих органоидов.
12. Класс: Голые амёбы
Втечение суток деление может повторяться
несколько раз.
Бесполое размножение – простой и быстрый
способ увеличить число своих потомков. Этот
способ размножения не отличается от деления
клеток при росте тела многоклеточного
организма. Разница в том, что дочерние клетки
одноклеточного организма, расходятся, как
самостоятельные.
13. Класс: Голые амёбы
Реакция на раздражение:Амёба обладает раздражимостью – способностью
чувствовать и реагировать на сигналы из внешней
среды. Наползая на предметы, она отличает
съедобные от несъедобных и захватывает их
ложноножками.
 Она уползает и прячется от яркого
Она уползает и прячется от яркогосвета (1), механических раздражений и повышенной
концентрации, вредных для нее веществ (2).
Такое поведение, состоящее в движении к
раздражителю или от него, называется таксисом.
14. Класс: Голые амёбы
Переживание неблагоприятных условий:Одноклеточное животное очень чувствительно к
В неблагоприятных условиях (при высыхании водоёма, в
холодное время года) амёбы втягивают псевдоподии. На
поверхность тела из цитоплазмы выделяются значительное
количество воды и вещества, которые образуют прочную
двойную оболочку. Происходит переход в покоящееся
состояние – цисту (1). В цисте жизненные процессы
приостанавливаются.
Цисты, разносимые ветром, способствуют расселению
амебы.
15. Класс: Голые амёбы
Принаступлении благоприятных условиях амёба
покидает оболочку цисты. Она выпускает
псевдоподии и переходит в активное состояние
(2-3).
Ещё одна форма защиты – способность к
регенерации (восстановлению). Повреждённая
клетка может достроить свою разрушенную
часть, но только при условии сохранения ядра,
так как там хранится вся информации о
строении.
16. Класс: Голые амёбы
Жизненныйцикл амёбы:
Жизненный цикл амёбы прост. Клетка растёт,
развивается (1) и делится бесполым путём (2). В
плохих условиях любой организм может
«временно умереть» — превратиться в цисту (3).
При улучшении условий он «возвращается к
жизни» и усиленно размножается.
17. Ссылки:
http://www.ebio.ru/pro02.htmlhttp://bethoven-vet.ru/sistematika-
prosteishikh
https://otvet.mail.ru/question/68289137
http://biouroki.ru/material/animals/ameb
a.html
http://referatyk.com/biologiya/13953stroenie_i_jiznedeyatelnost_amebyi.html
http://player.myshared.ru/402338/
особенности организации, разнообразие, систематика, распространение и значение.

К подцарству Простейшие относятся одноклеточные животные, каждой особи присущи все основные жизненные функции: обмен веществ, раздражимость, движение, размножение. Есть и колониальные виды. Среды обитания: морские и пресные водоемы, почва, организмы растений, животных и человека.
 У некоторых простейших над
мембраной имеется раковинка. Питание гетеротрофное:
у одних пища может поступать в любом
месте тела, у других она поступает через
специализированные органоиды: клеточный
рот, клеточную глотку. Пищеварение
внутриклеточное с помощью пищеварительной
вакуоли. Непереваренные остатки
выделяются или в любом месте тела, или
через специальное отверстие — порошицу.
Есть миксотрофные организмы, питающиеся
на свету с помощью фотосинтеза и имеющие
хроматофоры, а при отсутствии света
переходящие на гетеротрофный тип
питания. Часто эти организмы имеют
сократительные вакуоли. Дыхание.
Подавляющее большинство простейших —
аэробные организмы.
Ответная
реакция на воздействия внешней среды
— раздражимость — проявляется в виде
таксисов — движений всего организма,
направленных либо в сторону раздражителя,
либо от него. Например, эвглена зеленая
проявляет положительный фототаксис —
движется в сторону света. При наступлении
неблагоприятных условий большинство
простейших образуют цисты.
У некоторых простейших над
мембраной имеется раковинка. Питание гетеротрофное:
у одних пища может поступать в любом
месте тела, у других она поступает через
специализированные органоиды: клеточный
рот, клеточную глотку. Пищеварение
внутриклеточное с помощью пищеварительной
вакуоли. Непереваренные остатки
выделяются или в любом месте тела, или
через специальное отверстие — порошицу.
Есть миксотрофные организмы, питающиеся
на свету с помощью фотосинтеза и имеющие
хроматофоры, а при отсутствии света
переходящие на гетеротрофный тип
питания. Часто эти организмы имеют
сократительные вакуоли. Дыхание.
Подавляющее большинство простейших —
аэробные организмы.
Ответная
реакция на воздействия внешней среды
— раздражимость — проявляется в виде
таксисов — движений всего организма,
направленных либо в сторону раздражителя,
либо от него. Например, эвглена зеленая
проявляет положительный фототаксис —
движется в сторону света. При наступлении
неблагоприятных условий большинство
простейших образуют цисты.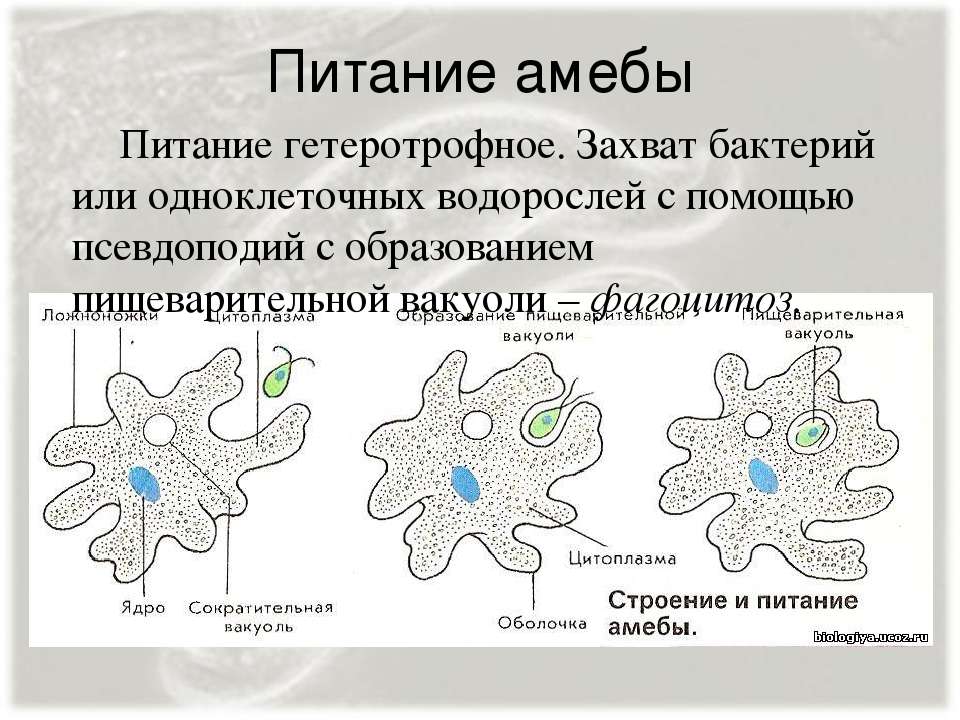 Инцистирование
— способ переживания неблагоприятных
условий. Размножение.
Бесполое размножение: или митотическое
деление вегетативной особи на две
дочерние клетки, или множественное
деление, при котором образуется несколько
дочерних клеток. Существуют половой
процесс — конъюгация (у инфузорий) и
половое размножение (у инфузорий,
вольвокса, малярийного плазмодия). Многообразие
простейших Насчитывается от 30 до 70 тысяч видов (по
данным разных авторов).. Класс
Саркодовые. Свободноживущие
представители: амеба
обыкновенная, фораминиферы,
радиолярии. Размер
клетки 0,2—0,7 мм. Амеба живет в пресноводных,
илистых водоемах. Цитоплазма образует
выросты – псевдоподии,
или ложноножки,
служащие для передвижения и фагоцитоза.
Клетка типичного эукариотического
строения. Есть пищеварительные и
сократительные вакуоли. Выполняет все
функции самостоятельного организма. К
патогенным простейшим этого типа
относится дизентерийная амеба, вызывающая амебную
дизентерию у
человека.
Инцистирование
— способ переживания неблагоприятных
условий. Размножение.
Бесполое размножение: или митотическое
деление вегетативной особи на две
дочерние клетки, или множественное
деление, при котором образуется несколько
дочерних клеток. Существуют половой
процесс — конъюгация (у инфузорий) и
половое размножение (у инфузорий,
вольвокса, малярийного плазмодия). Многообразие
простейших Насчитывается от 30 до 70 тысяч видов (по
данным разных авторов).. Класс
Саркодовые. Свободноживущие
представители: амеба
обыкновенная, фораминиферы,
радиолярии. Размер
клетки 0,2—0,7 мм. Амеба живет в пресноводных,
илистых водоемах. Цитоплазма образует
выросты – псевдоподии,
или ложноножки,
служащие для передвижения и фагоцитоза.
Клетка типичного эукариотического
строения. Есть пищеварительные и
сократительные вакуоли. Выполняет все
функции самостоятельного организма. К
патогенным простейшим этого типа
относится дизентерийная амеба, вызывающая амебную
дизентерию у
человека.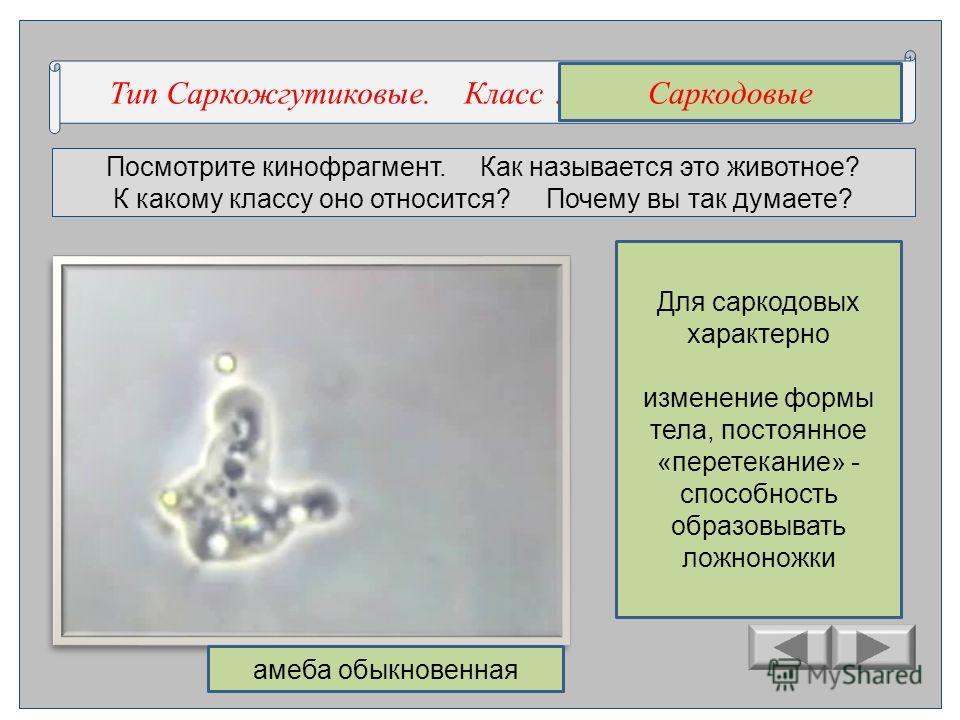 Паразитирует в кишечнике
человека.
Паразитирует в кишечнике
человека.Класс Жгутиковые. Представители этого класса имеют постоянную форму тела, благодаря наличию уплотненной клеточной оболочки. Животные передвигаются с помощью одного или нескольких жгутиков. Свободноживущие формы обитают в воде, влажной почве. Среди них есть как фотосинтезирующие, так и нефотосинтезирующие организмы. Паразитические простейшие (лямблия печеночная, трипаносома) и другие обитают в других организмах.
Эвглена
зеленая имеет веретенообразную форму
тела. Размер клетки около 0,05 мм.
Передвигается эвглена с помощью жгутика
– цитоплазматического выроста, состоящего
из тонких фибрилл.
На переднем конце находится
светочувствительный глазок. В цитоплазме,
помимо всех, характерных для животных
клеток, органелл, находятсяхроматофоры,
содержащие хлорофилл. На свету эвглена
способна к фотосинтезу. Поэтому ее
относят к промежуточным, между растениями
и животными, эволюционным формам. Размножается эвглена бесполым путем,
делением надвое по продольной оси.
Размножается эвглена бесполым путем,
делением надвое по продольной оси.
Тип
Споровики. К
этому типу относятся паразитические
простейшие, представителем которых
является малярийный паразит. Его
жизненный цикл протекает со сменой
хозяев (комар – человек), которые, заражая
друг друга, способствуют распространению
паразита. При укусе человека самкой
малярийного комара в его кровь проникают
возбудители малярии – малярийный
плазмодий. Он начинает активно размножаться
в клетках печени человека. После
размножения паразиты превращаются в
другую стадию, поселяющуюся в эритроцитах
крови. Бесполое размножение паразита
в крови сопровождается массовой гибелью
эритроцитов, выходом в кровь новых
паразитов и токсичных продуктов их
обмена, вызывающих лихорадку. Этот
процесс цикличен, поэтому и лихорадка
носит характер периодических приступов.
Для дальнейшего развития возбудители
должны попасть в желудок комара, где
происходит их половое размножение и
снова в слюнные железы самки.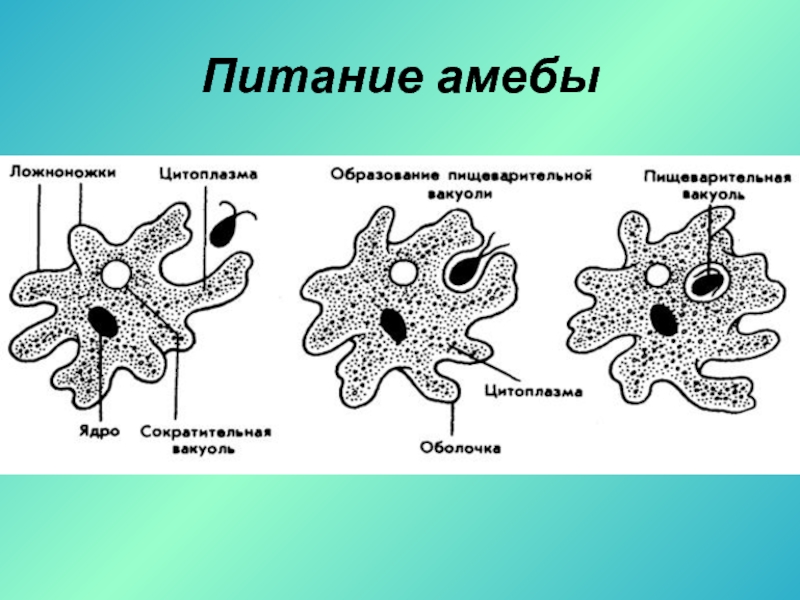
Тип Инфузории. Класс ресничные инфузории. Тип насчитывает около 6 тыс. видов.
Представители
– инфузория-туфелька,
инфузория-трубач.Инфузория-туфелька –
животное размером 0,1—0,3 мм.Ее клеточная
оболочка покрыта ресничками, служащими
для передвижения. В клетке два ядра
–вегетативное,полиплоидное и генеративное, диплоидное.
Ротовое углубление на теле образует
ротовую воронку, переходящую в клеточный
рот, ведущий в глотку.
В глотке формируются пищеварительные
вакуоли,
переваривающие пищу. Непереваренные
остатки пищи удаляются через отверстие
– порошицу.У
инфузории-туфельки две сократительные
вакуоли, расположенные в противоположных
концах тела. Через них выводится избыток
воды и продукты обмена веществ.Размножение
инфузории происходит
как бесполым, так и половым путями. При
бесполом размножении происходит
продольное деление клетки. При половом
процессе между двумя инфузориями
образуется цитоплазматический мостик.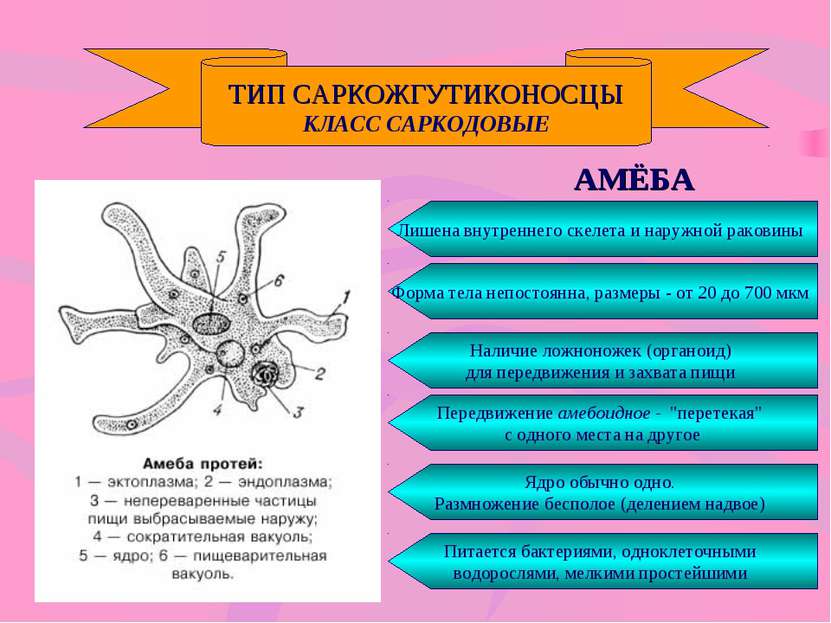 Полиплоидные (большие) ядра разрушаются,
а диплоидные (малые) ядра делятся мейозом
с образованием четырех гаплоидных ядер,
три из которых погибает, а четвертое
делится пополам, но уже митозом. Образуется
два ядра. Одно – стационарное и другое
– мигрирующее. Затем между инфузориями
происходит обмен мигрирующими ядрами.
Потом стационарное и мигрировавшее
ядра сливаются, особи расходятся и в
них снова образуются большое и малое
ядра.
Полиплоидные (большие) ядра разрушаются,
а диплоидные (малые) ядра делятся мейозом
с образованием четырех гаплоидных ядер,
три из которых погибает, а четвертое
делится пополам, но уже митозом. Образуется
два ядра. Одно – стационарное и другое
– мигрирующее. Затем между инфузориями
происходит обмен мигрирующими ядрами.
Потом стационарное и мигрировавшее
ядра сливаются, особи расходятся и в
них снова образуются большое и малое
ядра.
Значение. Простейшие, обитающие в океанах, пресных
водах, почве и в высших организмах,
занимают важнейшее место в круговороте
веществ в биосфере. В водной среде
простейшие — основа планктона,
используемая в пищу другими более
крупными животными. Из скелетов
простейших: фораминифер, радиолярий и
панцирных
жгутиконосцев
— кокколито-форид образуются мощные
пласты осадочных пород. Многие водные
простейшие — седиментаторы, питающиеся
взвешенными органическими частицами
и бактериями, играют существенную роль
в биологической очистке вод.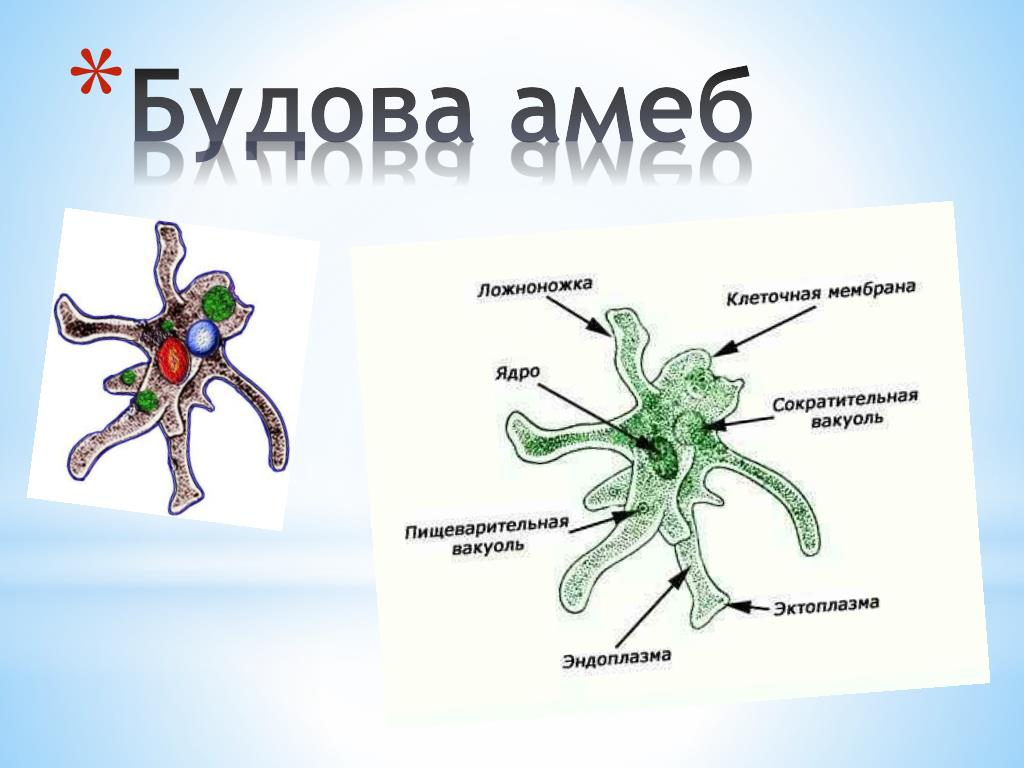 Почвенные
амебы, инфузории и жгутиконосцы — важное
звено почвенной фауны: они принимают
участие в почвообразовании. Ряд видов
простейших составляют полезную группу
симбионтов высших животных, улучшающих
пищеварение и обменные процессы в
организме.
Почвенные
амебы, инфузории и жгутиконосцы — важное
звено почвенной фауны: они принимают
участие в почвообразовании. Ряд видов
простейших составляют полезную группу
симбионтов высших животных, улучшающих
пищеварение и обменные процессы в
организме.
Внешний вид строение и движение амебы рисунок. Амеба обыкновенная, ее среда обитания, особенности строения и жизнедеятельности
Амеба обыкновенная – вид простейших существ из эукариот, типичный представитель рода Амебы.
Систематика . Вид амебы обыкновенной относится к царству — Животные, типу – Амебозои. Амебы объединены в класс Lobosa и отряд – Amoebida, семейство – Amoebidae, род – Amoeba.
Характерные процессы . Хотя амебы – это простые, состоящие из одной клетки существа, не имеющие никаких органов, им присущи все жизненно необходимые процессы. Они способны передвигаться, добывать пищу, размножаться, поглощать кислород, выводить продукты обмена.
Строение
Амеба обыкновенная – одноклеточное животное, форма тела неопределенная и изменяется из-за постоянного перемещения ложноножек.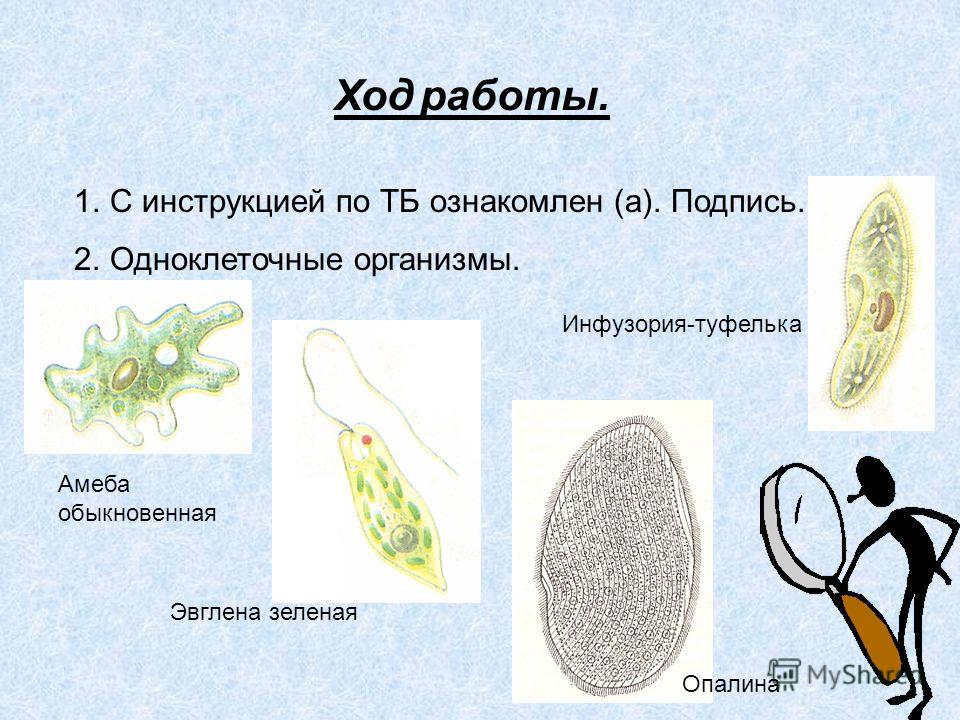 Размеры не превышают половины миллиметра, а снаружи ее тело окружено мембраной – плазмалемой. Внутри располагается цитоплазма со структурными элементами. Цитоплазма представляет собой неоднородную массу, где выделяют 2 части:
Размеры не превышают половины миллиметра, а снаружи ее тело окружено мембраной – плазмалемой. Внутри располагается цитоплазма со структурными элементами. Цитоплазма представляет собой неоднородную массу, где выделяют 2 части:
- Наружная – эктоплазма;
- внутренняя, с зернистой структурой – эндоплазма, где сосредоточены все внутриклеточные органеллы.
У амебы обыкновенной имеется крупное ядро, которое расположено примерно в центре тела животного. Оно имеет ядерный сок, хроматин и покрыто оболочкой, имеющей многочисленные поры.
Под микроскопом видно, что амеба обыкновенная образует псевдоподии, в которые переливается цитоплазма животного. В момент образования псевдоподии в нее устремляется эндоплазма, которая на периферических участках уплотняется и превращается в эктоплазму. В это время на противоположном участке тела эктоплазма частично превращается в эндоплазму. Таким образом, в основе образования псевдоподий лежит обратимое явление превращения эктоплазмы в эндоплазму и наоборот.
Дыхание
Амеба получает O 2 из воды, который диффундирует во внутреннюю полость через наружные покровы. Все тело участвует в дыхательном акте. Кислород, попавший в цитоплазму, необходим для расщепления питательных веществ на простые составляющие, которые Amoeba proteus сможет переварить, а еще для получения энергии.
Среда обитания
Обитает в пресной воде канав, небольших прудов и болот. Может жить также в аквариумах. Культуру амебы обыкновенной можно легко разводить в лабораторных условиях. Она является одной из крупных свободноживущих амеб, достигающих 50 мкм в диаметре и видимых невооруженным глазом.
Питание
Амеба обыкновенная передвигается с помощью ложноножек. Она преодолевает один сантиметр за пять минут. Передвигаясь, амебы наталкиваются на различные мелкие объекты: одноклеточные водоросли, бактерии, мелких простейших и т.д. Если объект достаточно мал, амеба обтекает его со всех сторон и он, вместе с небольшим количеством жидкости, оказывается внутри цитоплазмы простейшего.
Схема питания амебы обыкновенной
Процесс поглощения твердой пищи амебой обыкновенной называется фагоцитозом. Таким образом, в эндоплазме образуются пищеварительные вакуоли, внутрь которых из эндоплазмы поступают пищеварительные ферменты и происходит внутриклеточное пищеварение. Жидкие продукты переваривания проникают в эндоплазму, вакуоль с непереваренными остатками пищи подходит к поверхности тела и выбрасывается наружу.
Кроме пищеварительных вакуолей в теле амеб находится и так называемая сократительная, или пульсирующая, вакуоль. Это пузырек водянистой жидкости, который периодически нарастает, а достигнув определенного объема, лопается, опорожняя свое содержимое наружу.
Основная функция сократительной вакуоли — регуляция осмотического давления внутри тела простейшего. В связи с тем, что концентрация веществ в цитоплазме амебы выше, чем в пресной воде, создается разность осмотического давления внутри и вне тела простейшего. Поэтому пресная вода проникает в организм амебы, но ее количество остается в пределах физиологической нормы, поскольку пульсирующая вакуоль «откачивает» избыток воды из тела. Подтверждением этой функции вакуоли служит их наличие только у пресноводных простейших. У морских она или отсутствует, или сокращается очень редко.
Сократительная вакуоль кроме осморегуляторной функции частично выполняет и выделительную функцию, выводя вместе с водой в окружающую среду продукты обмена веществ. Однако основная функция выделения осуществляется непосредственно через наружную мембрану. Известную роль играет, вероятно, сократительная вакуоль в процессе дыхания, ибо проникающая в результате осмоса в цитоплазму вода несет растворенный кислород.
Размножение
Амебам свойственно бесполое размножение, осуществляемое путем деления надвое. Этот процесс начинается с митотического деления ядра, которое продольно удлиняется и перегородкой разъединяется на 2 самостоятельные органеллы. Они отдаляются и формируют новые ядра. Цитоплазма с оболочкой делится с помощью перетяжки. Сократительная вакуоль не разделяется, а попадает в одну из новообразованных амеб, во второй вакуоль формируется самостоятельно. Размножаются амебы достаточно быстро, за день процесс деления может происходить несколько раз.
В летний период времени амебы растут и делятся, но с приходом осенних холодов, из-за пересыхания водоемов, трудно найти питательные вещества. Поэтому амеба превращается в цисту, оказавшись в критических условиях и покрывается прочной двойной белковой оболочкой. При этом цисты легко распространяются за ветром.
Значение в природе и жизни человека
Amoeba proteus — важное составляющее экологических систем. Она регулирует численность бактериальных организмов в озерах и прудах. Очищает водную среду от чрезмерного загрязнения. Также является важным составляющим пищевых цепочек. Одноклеточные – еда для маленьких рыб и насекомых.
Ученые используют амебу как лабораторное животное, проводя на ней множество исследований. Очищает амеба не только водоемы, но поселившись в человеческом организме, она поглощает разрушенные частицы эпителиальной ткани пищеварительного тракта.
Простейшие в капле прудовой воды (под микроскопом).
Класс корненожек объединяет наиболее простых одноклеточных животных, тело которых лишено плотной оболочки, а потому не имеет постоянной формы.Для них характерно образование ложноножек, которые представляют собой временно образующиеся выросты цитоплазмы, способствующие передвижению и захвату пищи.
Среда обитания, строение и передвижение амёбы. Обыкновенная амёба встречается в иле на дне прудов с загрязненной водой. Она похожа на маленький (0,2-0,5 мм), едва заметный простым глазом бесцветный студенистый комочек, постоянно меняющий свою форму («амеба» означает «изменчивая»). Рассмотреть детали строения амёбы можно только под микроскопом.
Тело амёбы состоит из полужидкой цитоплазмы с заключенным внутрь неё небольшим пузыревидным ядром . Амёба состоит из одной клетки, но эта клетка — целый организм, ведущий самостоятельное существование.
Цитоплазма клетки находится в постоянном движении. Если ток цитоплазмы устремляется к одной какой-то точке поверхности амёбы, в этом месте на её теле появляется выпячивание. Оно увеличивается, становится выростом тела — ложноножкой, в него перетекает цитоплазма, и амёба таким способом передвигается. Амёбу и других простейших, способных образовывать ложноножки, относят к группе корненожек . Такое название они получили за внешнее сходство ложноножек с корнями растений.
Жизнидеятельность Амёбы.
Питание . У амёбы одновременно может образовываться несколько ложноножек, и тогда они окружают пищу — бактерии, водоросли, других простейших. Из цитоплазмы, окружающей добычу, выделяется пищеварительный сок. Образуется пузырёк — пищеварительная вакуоль. Пищеварительный сок растворяет часть веществ, входящих в состав пищи, и переваривает их. В результате пищеварения образуются питательные вещества, которые просачиваются из вакуоли в цитоплазму и идут на построение тела амебы. Нерастворенные остатки выбрасываются наружу в любом месте тела амебы.
Дыхание Амёбы . Амёба дышит растворенным в воде кислородом, который проникает в ее цитоплазму через всю поверхность тела. При участии кислорода происходит разложение сложных пищевых веществ цитоплазмы на более простые. При этом выделяется энергия, необходимая для жизнидеятельности организма.
Выделение вредных веществ жизнидеятельности и избытка воды. Вредные вещества удаляются из организма амёбы через поверхность ее тела, а также через особый пузырек — сократительную вакуоль. Окружающая амебу вода постоянно проникает в цитоплазму, разжижая ее. Избыток этой воды с вредными веществами постепенно наполняет вакуоль. Время от времени содержимое вакуоли выбрасывается наружу. Итак, из окружающей среды в организм амёбы поступают пища, вода, кислород. В результате жизнедеятельности амёбы они претерпевают изменения. Переваренная пища служит материалом для построения тела амёбы. Образующиеся вредные для амёбы вещества удаляются наружу. Происходит обмен веществ. Не только амёба, но и все другие живые организмы не могут существовать без обмена веществ как внутри своего тела, так и с окружающей средой.
Размножение Амёбы . Питание амёбы приводит к росту ее тела. Выросшая амёба приступает к размножению. (? Наверное вследствии превышения определённой массы её тела.) Размножение начинается с изменения ядра. Оно вытягивается, поперечной бороздкой делится на две половинки, которые расходятся в разные стороны — образуется два новых ядра. Тело амёбы разделяет на две части перетяжка. В каждую из них попадает по одному ядру. Цитоплазма между обеими частями разрывается, и образуются две новые амёбы. Сократительная вакуоль остается в одной из них, в другой же возникает заново. Итак, амёба размножается делением надвое. В течение суток деление может повторяться несколько раз.
Деление (размножение) Амёбы.
Циста . Питание и размножение амёбы происходит в течение всего лета. Осенью при наступлении холодов амёба перестает питаться, тело ее становится округлым, на его поверхности выделяется плотная защитная оболочка — образуется циста. То же самое происходит при высыхании пруда , где живут амёбы. В состоянии цисты амёба переносит неблагоприятные для неё условия жизни. При наступлении благоприятных условий амёба покидает оболочку цисты. Она выпускает ложноножки, начинает питаться и размножаться. Цисты, разносимые ветром, способствуют расселению (распространению) амеб.
Возможные дополнительные вопросы для самостоятельного изучения.
- Что заставляет Цитоплазму планомеренно перетекать из одного участка Амёбы в другой, заставляя её передвигаться в заданном направлении?
- Как происходит распознавание оболочкой цитоплазмы Амёбы питательных веществ, вследствии чего амёба целенаправленно формирует ложноножки и пищеварительную вакуоль?
Обыкновенная амеба встречается в иле на дне прудов с загрязненной водой. Она похожа на маленький (0,2-0,5 мм), едва заметный простым глазом бесцветный студенистый комочек, постоянно меняющий свою форму («амеба» означает «изменчивая»). Рассмотреть детали строения амебы можно только под микроскопом.
Строение и передвижение амебы обыкновенной
Тело амебы состоит из полужидкой цитоплазмы с заключенным внутрь нее небольшим пузыревидным ядром. Амеба состоит из одной клетки, но эта клетка — целый организм, ведущий самостоятельное существование.
Цитоплазма клетки находится в постоянном движении. Если ток цитоплазмы устремляется к одной какой-то точке поверхности амебы, в этом месте на ее теле появляется выпячивание. Оно увеличивается, становится выростом тела — ложноножкой, в него перетекает цитоплазма, и амеба таким способом передвигается. Амебу и других простейших, способных образовывать ложноножки, относят к группе корненожек. Такое название они получили за внешнее сходство ложноножек с корнями растений.
Питание амебы обыкновенной
У амебы одновременно может образовываться несколько ложноножек, и тогда они окружают пищу — бактерии, водоросли, других простейших. Из цитоплазмы, окружающей добычу, выделяется пищеварительный сок. Образуется пузырек — пищеварительная вакуоль.
Пищеварительный сок растворяет часть веществ, входящих в состав пищи, и переваривает их. В результате пищеварения образуются питательные вещества, которые просачиваются из вакуоли в цитоплазму и идут на построение тела амебы. Нерастворенные остатки выбрасываются наружу в любом месте тела амебы.
Дыхан ие амебы обыкновенной
Амеба дышит растворенным в воде кислородом, который проникает в ее цитоплазму через всю поверхность тела. При участии кислорода происходит разложение сложных пищевых веществ цитоплазмы на более простые. При этом выделяется энергия, необходимая для жизнедеятельности организма.
Выделение вредных веществ жизнедеятельности и избытка воды амебы обыкновенной
Вредные вещества удаляются из организма амебы через поверхность ее тела, а также через особый пузырек — сократительную вакуоль. Окружающая амебу вода постоянно проникает в цитоплазму, разжижая ее. Избыток этой воды с вредными веществами постепенно наполняет вакуоль. Время от времени содержимое вакуоли выбрасывается наружу.
Итак, из окружающей среды в организм амебы поступают пища, вода, кислород. В результате жизнедеятельности амебы они претерпевают изменения. Переваренная пища служит материалом для построения тела амебы. Образующиеся вредные для амебы вещества удаляются наружу. Происходит обмен веществ амебы обыкновенной. Не только амеба, но и все другие живые организмы не могут существовать без обмена веществ как внутри своего тела, так и с окружающей средой.
Размножение амебы обыкновенной
Питание амебы приводит к росту ее тела. Выросшая амеба приступает к размножению. Размножение начинается с изменения ядра. Оно вытягивается, поперечной бороздкой делится на две половинки, которые расходятся в разные стороны — образуется два новых ядра. Тело амебы разделяет на две части перетяжка. В каждую из них попадает по одному ядру. Цитоплазма между обеими частями разрывается, и образуются две новые амебы. Сократительная вакуоль остается в одной из них, в другой же возникает заново. Итак, амеба размножается делением надвое. В течение суток деление может повторяться несколько раз.
Циста амебы обыкновенной
Питание и размножение амебы происходит в течение всего лета. Осенью при наступлении холодов амеба перестает питаться, тело ее становится округлым, на его поверхности выделяется плотная защитная оболочка — образуется циста. То же самое происходит при высыхании пруда, где живут амебы. В состоянии цисты амеба переносит неблагоприятные для нее условия жизни. При наступлении благоприятных условий амеба покидает оболочку цисты. Она выпускает ложноножки, начинает питаться и размножаться. Цисты, разносимые ветром, способствуют расселению амеб.
К подцарству Одноклеточные относятся животные, тело которых состоит всего из одной клетки, большей частью микроскопического размера, но со всеми присущими организму функциями. В физиологическом отношении эта клетка представляет целый самостоятельный организм.
Двумя основными компонентами тела одноклеточных являются цитоплазма и ядро (одно или несколько). Цитоплазма окружена наружной мембраной. Она имеет два слоя: наружный (более светлый и плотный) — эктоплазму — и внутренний — эндоплазму. В эндоплазме находятся клеточные органоиды: митохондрии, эндоплазматическая сеть, рибосомы, элементы аппарата Гольджи, различные опорные и сократительные волокна, сократительные и пищеварительные вакуоли и др.
Среда обитания и внешнее строение обыкновенной амёбы
Простейшее живёт в воде. Это может быть и вода озера, и капля росы, и влага почвы, и даже вода внутри нас. Поверхность тела их очень нежная и без воды моментально высыхает. Внешне амёба похожа на сероватый студенистый комочек (0,2-05 мм), не имеющий постоянной формы.
Движение
Амёба «перетекает» по дну. На теле постоянно образуются меняющие свою форму выросты — псевдоподии (ложноножки). В один из таких выступов постепенно переливается цитоплазма, ложная ножка в нескольких точках прикрепляется к субстрату и происходит передвижение.
Внутреннее строение
Внутреннее строение амебы
Питание
Передвигаясь, амёба наталкивается на одноклеточные водоросли, бактерии, мелкие одноклеточные, «обтекает» их и включает в цитоплазму, образуя пищеварительную вакуоль.
Питание амебы
Ферменты, расщепляющие белки, углеводы и липиды, поступают внутрь пищеварительной вакуоли, и происходит внутриклеточное пищеварение. Пища переваривается и всасывается в цитоплазму. Способ захвата пищи с помощью ложных ножек называется фагоцитозом.
Дыхание
Кислород расходуется на клеточное дыхание. Когда его становится меньше, чем во внешней среде, новые молекулы проходят внутрь клетки.
Дыхание амебы
Молекулы углекислого газа и вредных веществ, накопившихся в результате жизнедеятельности, наоборот, выходят наружу.
Выделение
Пищеварительная вакуоль подходит к клеточной мембране и открывается наружу, чтобы непереваренные остатки выбросить наружу в любом участке тела. Жидкость поступает в тело амёбы по образующимся тонким трубковидным каналам, путём пиноцитоза. Откачиванием лишней воды из организма занимаются сократительные вакуоли. Они постепенно наполняются, а раз в 5-10 минут резко сокращаются и выталкивают воду наружу. Вакуоли могут возникать в любой части клетки.
Размножение
Амёбы размножаются только бесполым путём.
Размножение амебы
Выросшая амёба приступает к размножению. Оно происходит путём деления клетки. До деления клетки ядро удваивается, чтобы каждая дочерняя клетка получила свою копию наследственной информации (1). Размножение начинается с изменения ядра. Оно вытягивается (2), а затем постепенно удлиняется (3,4) и перетягивается посредине. Поперечной бороздкой делится на две половинки, которые расходятся в разные стороны — образуются два новых ядра. Тело амёбы разделяется на две части перетяжкой и образуется две новые амёбы. В каждую из них попадает по одному ядру (5). Во время деления происходит образование недостающих органоидов.
В течение суток деление может повторяться несколько раз.
Бесполое размножение — простой и быстрый способ увеличить число своих потомков. Этот способ размножения не отличается от деления клеток при росте тела многоклеточного организма. Разница в том, что дочерние клетки одноклеточного организма, расходятся, как самостоятельные.
Реакция на раздражение
Амёба обладает раздражимостью — способностью чувствовать и реагировать на сигналы из внешней среды. Наползая на предметы, она отличает съедобные от несъедобных и захватывает их ложноножками. Она уползает и прячется от яркого света (1),
механических раздражений и повышенной концентрации, вредных для нее веществ (2).
Такое поведение, состоящее в движении к раздражителю или от него, называется таксисом.
Половой процесс
Отсутствует.
Переживание неблагоприятных условий
Одноклеточное животное очень чувствительно к изменениям окружающей среды.
В неблагоприятных условиях (при высыхании водоёма, в холодное время года) амёбы втягивают псевдоподии. На поверхность тела из цитоплазмы выделяются значительное количество воды и вещества, которые образуют прочную двойную оболочку. Происходит переход в покоящееся состояние — цисту (1). В цисте жизненные процессы приостанавливаются.
Цисты, разносимые ветром, способствуют расселению амебы.
При наступлении благоприятных условиях амёба покидает оболочку цисты. Она выпускает псевдоподии и переходит в активное состояние (2-3).
Ещё одна форма защиты — способность к регенерации (восстановлению). Повреждённая клетка может достроить свою разрушенную часть, но только при условии сохранения ядра, так как там хранится вся информации о строении.
Жизненный цикл амёбы
Жизненный цикл амёбы прост. Клетка растёт, развивается (1) и делится бесполым путём (2). В плохих условиях любой организм может «временно умереть» — превратиться в цисту (3). При улучшении условий он «возвращается к жизни» и усиленно размножается.
Лобоподий, цилиндрических выростов с внутренними токами цитоплазмы.
| Амёба обыкновенная | ||||
| Научная классификация | ||||
|---|---|---|---|---|
| Международное научное название | ||||
Amoeba proteus (Pal.) | ||||
| ||||
Строение амёбы
Покров амёбы A. proteus представлен только цитоплазматической мембраной. Вследствие отсутствия твёрдых оболочек клетка имеет непостоянную форму и образует цитоплазматические выросты — псевдоподии (или ложноножки). Цитоплазма клетки дифференцирована на более светлую гелеобразную наружную часть гиалоплазму (эктоплазму) , и более темную золеобразную гранулоплазму (эндоплазму) , названную так из-за большого содержания различных включений и органелл. Среди клеточных органелл можно выделить одно ядро, одну сократительную вакуоль и множество пищеварительных вакуолей, а также гранул запасных веществ (различных полисахаридов, липидных капель, многочисленных кристаллов).
У данного вида имеется довольно сложный цитоскелет . Гиалоплазма пронизана сетью актиновых и миозиновых микрофиламентов — это кортикальный слой, связанный с клеточной мембраной и окружающий всё содержимое клетки (протопласт). Филаменты располагаются в клетке по-разному. У движущейся амёбы на переднем(«гиалиновом колпачке») и заднем (уроиде) концах актин образует очень тонкий слой, в то время как к середине клетки концентрация актиновых филаментов увеличивается. Миозин на переднем конце клетки также образует тонкий слой, который увеличивается к середине, а на заднем конце, в отличие от актина, — достигает максимальной толщины. Также, различается и их ориентация в пространстве. В передней трети тела движущейся амёбы актиновые филаменты располагаются продольно и соединяются специальными мостиками как с мембраной клетки, так и между собой. В заднем же конце актин формирует трёхмерную сеть, в которой залегают толстые филаменты миозина.
Питание
Амёба протей питается путём фагоцитоза , поглощая бактерий , одноклеточные водоросли и мелких простейших . Образование псевдоподий лежит в основе захвата пищи. На поверхности тела амёбы возникает контакт между плазмалеммой и пищевой частицей, в этом участке образуется «пищевая чашечка». Её стенки смыкаются, в эту область (с помощью лизосом) начинают поступать пищеварительные ферменты . Таким образом формируется пищеварительная вакуоль . Далее она переходит в центральную часть клетки, где подхватывается токами цитоплазмы. Вакуоль с непереваренными остатками пищи подходит к поверхности клетки и сливается с мембраной, таким образом выбрасывая наружу содержимое. Кроме фагоцитоза, амёбе свойствен пиноцитоз — заглатывание жидкости. При этом образуются на поверхности клетки впячивания в форме трубочки, по которой поступает внутрь цитоплазмы капелька жидкости. Образующая вакуоль с жидкостью отшнуровывается от трубочки. После всасывания жидкости вакуоль исчезает. Осморегуляция заключается в том, что в клетке периодически образуется пульсирующая сократительная вакуоль — вакуоль, содержащая излишнюю воду и выводящая её наружу.
Движение и реакция на раздражение
Тело Амёбы протей образует выступы — ложноножки . Выпуская ложноножки в определённом направлении, амёба протей передвигается со скоростью около 0,2 мм в минуту. Амёба распознаёт разные микроскопические организмы, служащие ей пищей. Она уползает от яркого света, механического раздражения и повышенных концентраций растворённых в воде веществ (например, от кристаллика поваренной соли).
Основная современная теория амёбоидного движения — теория «генерализованного кортикального сокращения» (Гребецки, 1982). В ней постулируется, что трёхмерное сокращение акто-миозинового комплекса, составляющего кортикальный слой клетки, приводит к сжатию эндоплазмы, в результате чего она направляется к переднему концу клетки, где кортекс наиболее тонкий. Туда же приносятся молекулы глобулярного актина (G-актина), который образуется на заднем конце в результате деполимеризации фибриллярного актина (F-актина), входящего в состав кортекса. В результате этого сокращения в эндоплазме создается повышенное давление, которое продавливает цитоплазму сквозь слой микрофиламентов на её переднем конце как сквозь сито. В результате этого мембрана переднего конца клетки отслаивается от кортекса и выпячивается наружу. Также сквозь филаментозное «сито» проходят и молекулы G-актина (в отличие от крупных включений цитоплазмы), которые затем попадают в пространство между цитоскелетом и мембраной в растущую лобоподию. На внутренней поверхности мембраны расположены специальные центры, полимеризующие G-актин обратно в F-актин, который становится основой для формирования нового цитоскелета. Вновь образованный слой филаментов начинает сокращаться, оказывая на цитоплазму давление, в связи с чем её ток направляется назад, — таким образом прекращается рост лобоподии. В это же время происходит деполимеризация отслоившегося ранее слоя кортекса.
Помимо этой теории, стоит упомянуть и несколько гипотез, предшествовавших ей.
- Гипотеза «потока под давлением» Маста. Предполагалось, что сокращение цитоскелета на заднем конце создается избыточное давление, вызывающее движение эндоплазмы в передний конец клетки, где она расплывается по сторонам, достигая гиалиновой шапочки. В кортикальной зоне происходит переход эндоплазмы в эктоплазму (так называемый золь-гель переход). Из-за того, что эти процессы проходят быстро, создается ощущение непрерывного тока цитоплазмы, в результате которого образуется лобоподия.
- Гипотеза Аллена. Похожа на предыдущую, разве что Аллен считал, что сокращения эндоплазмы происходят не на заднем конце, а на переднем. И там сразу же происходит переход из золя в гель, в результате которого новая порция золеобразной эндоплазмы как бы «подтягивается» к переднему концу, вызывая рост лобоподии. В зоне уроида же происходит обратный переход из геля в золь.
- Гипотеза Серавина. Предположил, что у всех амёбоидных клеток может присутствовать одинаковый набор различных механизмов движения, а различия в движении разных видов формируются в результате разной степени участия того или иного механизма в двигательной активности. Таким образом, согласно Серавину, механизмы, описанные Алленом и Мастом могут иметь место одновременно.
Среда обитания
Обитает на дне пресных водоёмов со стоячей водой, особенно в гниющих прудах и болотах, в которых есть много бактерий. Встречаются локомоторные и флотирующие формы. При плохих для амёбы условиях среды — понижении температуры осенью, пересыхании водоёма — амёба округляется, прекращает потребление пищи и образует плотную оболочку — цисту
| Саркодовые животные по классификации относятся к простейшим типа саркомастигофоры. К этому типу относится также класс жгутиконосцы. Класс саркодовых включает три подкласса: корненожки, лучевики, солнечники. Представителями саркодовых являются свободноживущие простейшие животные, обитающие в почве, соленых и пресных водоемах, а также паразиты многоклеточных животных и человека. Типичными признаками саркодовых обладают такие виды, как радиолярии (лучевики), солнечники, фораминиферы, амёба обыкновенная (корненожка), амёба дизентерийная. Давая характеристику саркодовым, следует отметить, что форма тела их непостоянна. Они образуют изменчивые по форме выросты клетки, предназначенные для движения и захвата пищи (фагоцитоза). Эти временные выросты цитоплазмы (псевдоподии, или ложноножки) могут быть в виде лопастей (лобоподии), анастомозирующей паутины (ретикулоподии), лучей (аскоподии), нитей (филоподии). Клетка саркодовых снаружи покрыта цитоплазматической мембраной (плазмалеммой). Но многие представители этого класса имеют внешний скелет (известковую раковину) или внутренний скелет (органический либо минеральный). Цитоплазму саркодовых формируют две составляющие: эктоплазма (более густой консистенции) и эндоплазма (более жидкая). В ней находятся одно или несколько ядер. Сократительные вакуоли характерны только для пресноводных форм саркодовых. Пищей представителям этого класса служат водоросли, другие простейшие, бактерии. Размножение у всех видов бесполое, реже половое. При бесполом типе происходит деление клетки пополам или почкование. Половое размножение подразумевает образование амёбоидных или жгутиковых гамет и их копуляции. При неблагоприятных условиях многие представители саркодовых образуют цисты, что позволяет им сохранить жизнеспособность при высоких или низких температурах, засушливой погоде. Значение саркодовых чрезвычайно велико. Многие из них участвуют в пищевой цепи, являясь пищей мальков рыб, некоторых ракообразных, крупных рыб, некоторых животных. Но среди простейших этого класса есть и паразиты. Это дизентерийная амёба, которая паразитируя в толстой кишке человека, повреждает ее стенку, провоцируя образование язв и питаясь эритроцитами крови. Этот вид саркодовых вызывает заболевание дизентерийный колит. Раковины саркодовых (радиолярий, фораминиферов) послужили источником огромных отложений известняка и мела в течение тысячелетий. Многие виды саркодовых известны в ископаемом состоянии из-за того, что их скелеты хорошо сохранились. Они являются объектом изучения палеонтологов. Тип простейших (Protozoa) состоит из 5 классов: Саркодовые, Жгутиконосцы, Споровики, Книдоспоридии и Инфузории.числом 8—10 тыс. Очень большое количество видов известно в ископаемом состоянии благодаря хорошей сохранности скелетов многих групп саркодовых. | Ресничные — один из типов простейших. Им присущи следующие признаки: 1) это одноклеточные гетеротрофные формы; 2) они снабжены ресничками — тонкими волосковидными отростками, биения которых вызывают ток воды, используемый клеткой либо для своего передвижения, либо для захвата пищевых частиц; 3) для них характерна определенная форма клетки, обусловленная наличием тонкого гибкого наружного слоя цитоплазмы, называемого пелликулой. Пелликула в свою очередь окружена плазматической мембраной; 4) клетки ресничных имеют довольно сложное строение, например они содержат макронуклеус и микронуклеус. К типу простейших Ciliophora (Инфузории, или Ресничные)относятся наиболее сложные простейшие. Они имеют плотную пелликулу, а часто и скелетные образования. Поэтому имеют постоянную форму. Органоиды движения – многочисленные реснички. Ядерный аппарат состоит из двух ядер: макронуклеуса и микронуклеуса. Клетка инфузории делится поперечным делением. Половой процесс представлен конъюгацией. Размножение простейших осуществляется обычно разными формами деления – разновидностями митоза. Характерен также половой процесс: в виде слияния клеток – копуляции, или обмена частью наследственного материала – конъюгации. В жизненном цикле большинства простейших выделяют стадию трофозоита – активно питающуюся и перемещающуюся форму, и стадию цисты. Циста – неподвижная форма жизненного цикла простейших, покрытая плотной оболочкой и характеризующаяся резко замедленным обменом веществ. |
Биология. Животные. — Тип Саркодовые и жгутиконосцы
Комментарии преподавателяКласс Саркодовые
Общим признаком представителей саркодовых служат органы передвижения и захвата пищи — ложноножки, или псевдоподии (иногда лучи или нити). Большинство обитает в морях, некоторые — в пресных водоемах; есть среди них паразиты. Строение и жизнедеятельность саркодовых удобно рассмотреть на примере типичного их представителя — амёбы протея.
Среда обитания и внешнее строение. Амеба протей, или обыкновенная амеба, обитает на дне небольших пресных водоемов: в прудах, старых лужах, канавах с застойной водой. Ее величина не превышает 0,5 мм. Амеба протей не имеет постоянной формы тела, так как лишена плотной оболочки. Тело ее образует выросты — ложноножки. С их помощью амеба медленно передвигается — «перетекает» с одного места на другое, ползет по дну, захватывает добычу. За такую изменчивость формы тела амебе и присвоили имя древнегреческого божества Протея, который мог менять свой облик. Внешне амеба протей напоминает маленький студенистый комочек. Самостоятельный одноклеточный организм амебы содержит цитоплазму, покрытую клеточной мембраной. Наружный слой цитоплазмы прозрачный и более плотный. Bнутренний ее слой зернистый и более текучий. В цитоплазме находятся ядро и вакуоли — пищеварительная и сократительная.
|
ТИП САРКОДОВЫЕ |
|
11 тыс. видов |
|
Форма тела – непостоянная, т.к. отсутствует клеточная стенка |
|
Ложноножки (псевдоподии) или жгутики(жгутик) |
|
В клетке одно ядро |
|
Способ питания гетеротрофный |
|
Дыхание всей поверхностью тела |
|
Выделение жидких продуктов обмена веществ происходит через сократительную вакуоль |
|
Раздражимость в виде таксисов |
|
Размножение путем деления |
Раковинные амебы обладают наружным скелетом — раковинкой. Из ее устья выступают лишь ложноножки. Раковинки могут состоять из рогоподобного вещества, из кремневых пластинок (вырабатываемых телом амебы) или из склеенных выделениями цитоплазмы песчинок. Размножаются раковинные амебы, как и амеба протей, делением надвое. Одна амеба остается в старой раковинке, а другая строит новую. Раковинные амебы обитают на дне пресных водоемов, в почве, в сфагновых болотах.
Радиолярии — морские одноклеточные организмы размером от 40 мкм до 1 мм, обитающие в теплых морях и океанах. У них минеральный (из кремнезема, реже — из сернокислого стронция) скелет. Он защищает радиолярию и увеличивает поверхность тела, способствуя «парению» радиолярии в толще воды. Форма скелета радиолярий чрезвычайно разнообразна. Снаружи выдаются нитевидные ложноножки, служащие для улавливания пищи.
Внутри клетки находится одно или много ядер, разнообразные включения, например капли жира, которые уменьшают удельную массу животного и способствуют «парению» в толще воды. У многих радиолярий в цитоплазме обитают мелкие одноклеточные водоросли, которые получают от радиолярий защиту, питательные вещества и углекислоту. Радиолярии, в свою очередь, получают от водорослей кислород, необходимый для дыхания. Кроме того, часть водорослей переваривается радиоляриями, служат ей пищей. Некоторые радиолярии при неблагоприятных условиях (опреснении воды, сильном волнении моря) способны опускаться на глубину в несколько десятков и сотен метров, а потом всплывать.
Скелеты погибших радиолярий, опускаясь на дно, образуют радиоляриевый ил, входящий в состав осадочных пород, которые называются радиоляритами. Так называемая «инфузорная земля», или трепел, целиком состоит из скелетов радиолярий.
Особую группу саркодовых образуют фораминиферы. Современные фораминиферы мелкие — 0,1-1 мм, а некоторые вымершие виды достигали 20 см. Наружный скелет фораминифер — раковинки. Они защищают тело животного и бывают известковыми, из хитиноподобного вещества или составлены из сцементированных песчинок. Раковинки бывают однокамерными или многокамерными, ветвящимися или расположенными в один-два ряда либо по спирали.
Через наружное отверстие (устье) и поры в стенках раковинок выдаются тончайшие и соединяющиеся между собой ложноножки, которые служат для движения и захвата пищи, образуют вокруг раковинки сеточку, диаметр которой во много раз превосходит диаметр раковинки. К такой сеточке прилипают пищевые частички, одноклеточные водоросли, которыми питаются фораминиферы. Все фораминиферы — морские, преимущественно донные, организмы. У планктонных фораминифер раковинки тонкие, с многочисленными выростами в виде расходящихся во все стороны тонких длинных игл, что позволяет им «парить» в толще воды. Всего известно около 30 тыс. видов фораминифер. Из них сейчас живет около 1000 видов, остальные известны в ископаемом состоянии.
Пустые раковинки фораминифер образуют огромные, толщиной в несколько сотен метров, пласты осадочных пород (например, мел и известняк). Отдельные виды фораминифер обитали только в определенную геологическую эпоху. Поэтому по наличию раковинок этих видов фораминифер в пластах Земли определяют возраст геологических пород.
Тело амебы протея состоит из одной клетки и выполняет вое функции живого организма. Она не имеет постоянной формы тела, гак как цитоплазма непрерывно образует выпячивания — ложноножки, с помощью которых передвигается, захватывает пишу. Амеба обладает раздражимостью — способностью отвечать на воздействие окружающей среды. При неблагоприятных условиях амеба выделяет защитную оболочку — образует цисту.
Тренажер.
Задание: http://files.school-collection.edu.ru/dlrstore/c4dd225d-0a01-022a-018b-3fee8aeb0def/%5BBIO7_03-09%5D_%5BQS_05%5D.html
ФайлыНет дополнительных материалов для этого занятия.
Тест по теме: «Систематика»
Систематика. Основные систематические(таксономические) категории.
1. Основные задачи систематики –
1) изучение этапов исторического развития организмов
2) установление взаимосвязей организмов и окружающей среды
3) выявление приспособленности организмов к среде обитания
4) объединение организмов в группы на основе родства
2. Какая схема используется при классификации растений?
1) вид – род – семейство – порядок – класс – отдел
2) вид – семейство – порядок – род – класс – отдел
3) вид – отдел – класс – порядок – род – семейство
4) вид – класс – отдел – порядок – род – семейство
3. Способность растений скрещиваться и давать плодовитое потомство – это основной признак
1) рода 2) отдела 3) класса 4) вида
4. Родственные виды растений объединяются в
1) семейства 2) класса 3) роды 4) популяции
5. К какой категории в системе органического мира относят хордовых животных?
1) царству 2) типу 3) классу 4) отряду
6. Бактерии, грибы, растения, животные рассматривают в системе органического мира как
1) типы 2) отделы 3) царства 4) классы
7. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите таблицу соответствующую последовательность цифр.
1) Ромашка аптечная
2) Двудольные
3) Покрытосеменные
4) Ромашка
5) Сложноцветные
6) Эукариоты
8. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующую последовательность цифр.
1) Простейшие
2) Амебы
3) Клеточные
4) Саркожгутиковые
5) Амеба обыкновенная
6) Эукариоты
9. Установите последовательность расположения систематических таксонов животных, начиная с наибольшего. Запишите соответствующую последовательность цифр.
1) Зайцевые
2) Млекопитающие
3) Дикий кролик
4) Зайцеобразные
5) Хордовые
6) Кролик
7) Животные
10. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующую последовательность цифр.
1) Пресмыкающиеся
2) Чешуйчатые
3) Змеи
4) Хордовые
5) Гадюка обыкновенная
6) Эукариоты
11. Установите последовательность расположения систематических таксонов, начиная с наименьшего. Запишите таблицу соответствующую последовательность цифр.
1) Паслен
2) Двудольная
3) Покрытосемянные
4) Паслен клубненосный
5) Пасленовые
6) Клеточные
12. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующею последовательность цифр.
1) Грызуны
2) Белка
3) Беличьи
4) Обыкновенная белка
5) Хордовые
6) Млекопитающие
13. Установите последовательность расположения систематических таксонов, начиная с самого наименьшего. Запишите в таблицу соответствующею последовательность цифр.
1) Горчица белая
2) Горчица
3) Покрытосемянные
4) Двудольные
5) Эукариоты
6) Крестоцветные
14. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующею последовательность цифр.
1) Тигры
2) Позвоночные
3) Хищные
4) Хордовые
5) Кошачьи
6) Эукариоты
15 . Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующею последовательность цифр.
1) Ромашка аптечная
2) Двудольные
3) Покрытосемянные
4) Ромашка
5) Сложноцветные
6) Эукариоты
16. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующею последовательность цифр.
1) Ящерицы
2) Прыткая ящерица
3) Пресмыкающиеся
4) Ящерица
5) Хордовые
6) Чешуйчатые
17. Установите последовательность расположения систематических таксонов, начиная с самого наименьшего. Запишите в таблицу соответствующею последовательность цифр.
1) Редька обыкновенная
2) Покрытосеменные
3) Крестоцветные
4) Двудольные
5) Редька
6) Эукариоты
18. Установите последовательность расположения систематических таксонов, начиная с самого крупного. Запишите в таблицу соответствующею последовательность цифр.
1) Медведи
2) Позвоночные
3) Хищные
4) Хордовые
5) Медвежьи
6) Белый медведь
Систематика. Основные систематические(таксономические) категории.
Ответы:
1-4
2-1
3-4
4-3
5-2
6-3
7-632541
8-361425
9-7524163
10-641235
11-415236
12-561324
13-126435
14- 642351
15-632541
16-536142
17-153426
18-423516
Naegleria Fowleri Amoeba — Характеристики / Симптомы / Лечение / Инфекция
Характеристики, симптомы, лечение и инфекция
Обзор / Введение
Член филюма перколозоа, обычно встречающийся в экологических алебритиках, Naegleria fowleri. теплая вода и наземная среда (аквариум, пруды, бассейны и т. д.).
Хотя Naegleria fowleri принадлежит к группе свободноживущих организмов, она является патогеном и возбудителем первичного амебного менингоэнцефалита (инфекции центральной нервной системы) у людей.
Хотя эти инфекции редки, они смертельны и составляют 98 процентов. В настоящее время инфекции, вызываемые этим организмом, выявлены менее чем в 20 странах. Однако его распространение в значительной степени ограничено условиями окружающей среды, в которых киста может выжить.
Некоторые из симптомов, связанных с первичным амебным менингоэнцефалитом, включают:
- Лихорадка
- Рвота
- Головная боль
- Нарушение психического статуса
* Nariegle.Малькольм Фаулер, австралийский патолог, который (вместе с доктором Родом Картером) признал этот организм возбудителем первичного амебного энцефалита в 1965-х годах.
Классификация Naegleria fowleri
Королевство: Protista — Как члены королевства Protista, Naegleria fowleri — простые эукариотические организмы, обычно встречающиеся в различных наземных и водных объектах, где они существуют в виде свободных живых организмов. или как паразиты / патогены, способные вызывать заболевание.Как члены этого царства, они не растения, животные или грибы.
Подцарство: Protozoa — Члены подцарства Protozoa, например Naegleria fowleri — это эукариотические одноклеточные организмы, которые можно найти повсюду в окружающей среде, где они существуют как свободноживущие организмы или как паразиты. Они сильно различаются по форме, причем некоторые виды способны изменять свою форму.
Тип: Percolozoa — Бесцветные амебы типа Percolozoa, для которых характерны жгутиковые фазы в их жизненном цикле.Большинство организмов (амебофлагелляты) являются свободноживущими и могут быть найдены в различных водных и наземных средах обитания, где они питаются бактериями и другим органическим материалом.
Класс: Heterolobosea — Acarpomyxea — это класс, который состоит из нескольких отрядов, включая: Schizopyrenida. Члены этого класса могут быть найдены в почве, морской и пресноводной среде, где они живут как свободноживущие организмы. Однако некоторые из этих видов могут быть паразитическими и вызывать заболевания / инфекции у людей.
Отряд: Schizopyrenida — члены отряда Schizopyrenida в природе встречаются в почве и в водных средах. Большинство видов этой группы свободно живут в этих средах и, следовательно, не зависят напрямую от хозяина.
Семейство: Vahlkampfiidae — В это семейство входят такие роды, как Naegleria, Monopylocystis, Paravahlkampfia и др., Которые населяют различные водные и наземные среды обитания как свободноживущие организмы (большинство организмов свободноживущие).Члены этой группы часто называют амебами limax, большинство из которых характеризуется жгутиками.
Род: Naegleria — род Naegleria насчитывает более 45 видов. Члены этой группы — свободноживущие протисты, обычно встречающиеся в теплых водоемах и некоторых наземных местообитаниях по всему миру.
Из этих видов только N. fowleri и N. lovaniensis известны как патогенные, при этом N. fowleri — единственный вид, который, как известно, вызывает болезни у людей.Помимо голых амеб, члены этой группы также характеризуются своим превращением из амеб в жгутиконосцев.
Вид: Naegleria fowleri — Характеристики этого вида обсуждаются ниже.
Экология
Будучи свободноживущими организмами, вид Naegleria fowleri можно встретить в различных наземных и водных средах обитания по всему миру (кроме Антарктиды). Помимо почвы и водных сред обитания, Naegleria fowleri при определенных условиях изолирована от воздуха.
В некоторых частях Нигерии, например, амеба была изолирована от воздуха во время Харматтана (сезон, характеризующийся сухими и пыльными ветрами).
Было показано, что в водной среде / водоемах, таких как пресноводные озера, популяция организма изменяется при изменении температуры. Хотя их численность относительно высока в жаркие летние месяцы, пик популяции приходится на конец лета. Здесь, однако, стоит отметить, что разные температурные диапазоны благоприятствуют разным формам амебы.
В то время как трофозоиты процветают в диапазоне температур от 35 до 46 градусов C, жгутиконосцам требуется температура от 27 до 37 градусов C. Помимо теплых пресноводных озер, некоторые из других водных сред обитания, в которых были изолированы Naegleria fowleri, включают пруды, реки. , бассейны, сточные воды, термически загрязненная вода и загрязненная вода, а также горячие источники.
* В своей среде обитания Naegleria fowleri может выжить в диапазоне pH от 2.1 и 8.15.
* Хотя N. fowleri можно найти в плавательных бассейнах, он не может выжить в хлорированных бассейнах. Низкие температуры (прохладная) и чистая вода также влияют на способность организма к воспроизводству и размножению.
Помимо различных водоемов, Naegleria fowleri также можно найти в почве при температуре выше 30 ° C. Однако они обычно встречаются во влажной или влажной почве, которая позволяет жгутиконосным формам плавать в водных пленках.
Почва является особенно важной средой обитания для организма, поскольку она обеспечивает определенные условия, способствующие росту и размножению.К ним относятся кислород, благоприятные температурные диапазоны и бактерии (источник пищи).
Из почвы амеба может затем переноситься в различные водоемы ручьями или проливными дождями и т. Д. Именно из этих источников (водоемов) организм может инфицировать людей.
* Цисты Naegleria fowleri могут выжить в неблагоприятных условиях окружающей среды, таких как низкие температуры.
* Здоровые люди также являются носителями N.fowleri — Микроорганизм был обнаружен в мазках, взятых изо рта, носа и глотки здоровых пациентов, а также в образцах сыворотки крови человека и слюны.
Морфология и клеточная структура Naegleria fowleri
Жизненный цикл Naegleria fowleri состоит из трех различных морфологических стадий. Следовательно, чтобы описать общую морфологию и клеточную структуру организма, важно взглянуть на его жизненный цикл.
Жизненный цикл
Трофозоиты
Жизненный цикл Naegleria fowleri может происходить в различных водных объектах и наземных средах, а также в организме человека-хозяина. Это потому, что они встречаются как свободноживущие организмы, так и патогены человека.
Жизненный цикл Naegleria fowleri начинается с стадии трофозоита, обычно встречающейся в водной среде. Являясь инфекционной стадией организма, обнаруживаемой в водной среде, трофозоиты могут инфицировать людей, контактирующих с водой в зараженных плавательных бассейнах и т. Д.
Хотя инфекции очень редки, исследования показали, что они поражают детей и молодых людей, когда зараженная вода попадает в организм через нос.
Трофозоиты также представляют репродуктивную стадию организма и делятся на бинарное деление при благоприятных условиях (например, в диапазоне температур от 35 до 46 градусов C). Это приводит к образованию двух одинаковых клеток, которые могут расти и повторять процесс.
* Во время деления клетки (посредством бинарного деления) ядрышко удлиняется и делится на две части.Однако ядерная мембрана остается неповрежденной. Кариокинез, который включает деление ядра, затем следует за делением цитоплазмы (цитокинезом), прежде чем клетка в конечном итоге разделится на две дочерние клетки.
Морфология / структура трофозоитов
Трофозоиты Naegleria fowleri имеют небольшие размеры, от 10 до 35 мкм. Они имеют вид лимакса и липкий задний конец, состоящий из хвостовых нитей.
Подобно другим представителям рода Naegleria, трофозоиты Naegleria fowleri обладают цитоплазматическими расширениями, известными как амебастомы (пищевые чашки). Эти структуры могут различаться по размеру и количеству в зависимости от напряжения. В водных и наземных средах обитания трофозоиты содержат большее количество кормушек, которые используются для поедания такой добычи, как дрожжи и бактерии, а также для прикрепления.
Как и эукариотические клетки, трофозоиты Naegleria fowleri также характеризуются множеством органелл, обычно обнаруживаемых у других эукариот.
Например, у них есть округлое связанное с мембраной ядро, которое состоит из толстого ядрышка, клеточной мембраны шириной около 10 нм, гладкой и шероховатой эндоплазматической сети, а также большого количества рибосом в цитоплазме, среди прочего.
Подобно многим другим клеткам, трофозоиты также имеют структуру цитоскелета, которая поддерживает клеточную структуру. Он состоит из толстых (расположенных в цитоплазме) и тонких (расположенных рядом с плазматической мембраной) микрофиламентов, состоящих из актина и миозина.
В отличие от стадии жгутиков, трофозоиты используют амебоидную локомоцию для перемещения из одной точки в другую. Здесь движение стало возможным благодаря адгезии к различным субстратам в их окружении. Однако скорость движения во многом зависит от температуры.
* Направление движения трофозоитов N. fowleri определяется хемотаксисом.
Флагелляты
В благоприятных условиях окружающей среды трофозоиты Naegleria fowleri размножаются путем бинарного деления с образованием двух дочерних клеток.Однако в случае дефицита питания и ионных изменений эта форма организма трансформируется в жгутиковые формы.
Хотя изменения в окружающей среде вызывают превращение трофозоитов в жгутиконосцы, на этой стадии все еще требуется вода. В отличие от трофозоитов, жгутиконосцы процветают в диапазоне температур от 27 до 37 градусов Цельсия и не могут воспроизводиться (они также не питаются) — переход от стадии трофозоита к стадии жгутиков занимает всего несколько часов.
Жгутиконосцы Naegleria fowleri имеют грушевидный вид и имеют размер от 10 до 16 мм.Исследования показали, что во время этого преобразования (от стадии трофозоита до стадии жгутиков) количество вакуолей уменьшается по мере формирования базальных тел и жгутиков.
Жгутик, который в конечном итоге формируется, имеет типичное расположение 9 + 2, состоящее из нитей, окруженных оболочкой. При благоприятных условиях жгутиконосцы могут вернуться в стадию трофозоитов и питаться различной добычей в своем окружении.
Подобно трофозоитной стадии у жгутиконосцев Naegleria fowleri также есть ядро.Однако здесь он расположен в переднем отделе организма, который уже по сравнению с передним отделом. Ядро, которое также состоит из ядрышка, покрыто отчетливой ядерной мембраной, как в случае с трофозоитами.
Некоторые другие органеллы, обнаруженные в формах жгутиков, включают:
- Митохондрии
- Грубый эндоплазматический ретикулум
- Вакуоли
- Ризопласт
в случае дефицита питательных веществ или из-за ионных изменений трофозоиты переходят в стадию кисты в неблагоприятных условиях (холод, истощение питательных веществ, сухость и т. д.).
Как и на стадии жгутиков, цисты также неспособны к размножению и не питаются. Они имеют сферическую форму и могут выжить в различных неблагоприятных условиях, в которых не могут выжить трофозоиты и жгутиконосцы.
Некоторые из других характеристик кисты Naegleria fowleri включают:
- Имеют толстую стенку с порами
- Слизистая пробка закрывает поры стенки кисты
- Инцистированная клетка содержит ядро и несколько цитоплазматических клеток. вакуоли
- Содержат несколько органелл, обычно встречающихся в других эукариотических клетках — грубый эндоплазматический ретикулум и менее выраженное ядрышко
* N.fowleri цисты также могут проникать в организм человека-хозяина через носовые ходы. Попадая в организм, они переходят в стадии трофозоитов, которые затем могут вызвать инфекцию.
Метаболизм и патогенность
Naegleria fowleri — аэробные гетеротрофные организмы, обычно встречающиеся в водной и различных земных средах (формы трофозоитов). Таким образом, они обычно находятся в богатой кислородом среде и имеют много митохондрий.
В этих средах Naegleria fowleri питаются бактериями и другими одноклеточными организмами, такими как дрожжи. Здесь трофозоиты используют псевдоподы для поглощения и поглощения пищевого материала, который затем переваривается в вакуолях.
Помимо псевдоподий, N. fowleri также образуют кормушки, на которых расположены фагоцитарные структуры, играющие важную роль в патогенности организма.
* Псевдоножки отходят от поверхности клетки, чтобы поглотить добычу.
* Кормушка образуется за счет впячивания поверхности тела.
* Согласно исследованиям, трофозоиты превращаются в активных пловцов (жгутиконосцев), когда ищут добычу. Когда они прибывают в среду обитания с достаточным или большим количеством добычи, они превращаются обратно в трофозоитов, способных питаться — Это достигается за счет хемотаксиса
Инфекция Как уже упоминалось, Naegleria fowleri являются аэробными гетеротрофами. организмы, обычно встречающиеся в богатых кислородом средах обитания.В теле человека-хозяина мозг представляет собой богатую кислородом среду с благоприятными условиями, которые позволяют организму выжить в качестве патогена.
У людей (особенно детей и молодых людей) острые инфекции возникают при вдыхании Naegleria fowleri через носовые ходы. Хотя инфекции редки, они возникают при вдыхании загрязненной воды (вода в бассейне или другая вода с трофозоитами) или цист в пыли через носовые ходы.
* Носовой ход — единственный путь, который обеспечивает успешную колонизацию хозяина, учитывая, что патоген может эффективно мигрировать в мозг.Если возбудитель попадает в организм, он легко разрушается желудочными кислотами.
При вдыхании организма через носовой ход трофозоиты прикрепляются к слизистой оболочке носа. Адгезия к клеткам слизистой оболочки является первым и наиболее важным шагом в патогенезе патогенной амебы (что стало возможным благодаря спайкам, выраженным на поверхности организма). Отсюда амебы попадают в обонятельные луковицы через обонятельные нервы.
Хотя было показано, что Naegleria fowleri движется в заданном направлении посредством хемотаксиса, обонятельные сенсорные пучки аксонов действуют как туннели, которые позволяют организму успешно достигать мозга, минуя барьер центральной нервной системы.
* Если человек вдыхает цисты Naegleria fowleri, они сначала должны превратиться в трофозоиты, которые способны прикрепляться к слизистой оболочке носа и мигрировать в мозг, пересекая пористые ребристые пластинки.
* Согласно исследованиям, дети и молодые люди подвергаются более высокому риску, потому что их решетчатые пластины более пористые. Решетчатые пластинки, расположенные непосредственно позади ноздрей / ноздрей, представляют собой пористое расширение, отделяющее носовую полость от мозга.
* Помимо прикрепления к слизистой оболочке носа, хемотаксический ответ (с ацетилхолином, действующим как химиоаттрактант), а также высокая скорость передвижения являются некоторыми другими факторами, способствующими патогенности N. fowleri.
В паренхиме мозга N. fowleri использует такие белки, как фибронектин-связывающий белок, для связывания с клетками мозга. Используя структуры чашки с едой, они начинают питаться тканями мозга посредством процесса, известного как трогоцитоз.
Здесь патогены используют свои структуры пищевых чашек для извлечения и поглощения клеточной мембраны и внутриклеточных компонентов клеток мозга, вызывая тем самым обширное повреждение — это причина, по которой Naegleria fowleri также называют амебой, поедающей мозг.
Помимо трогоцитоза, Naegleria fowleri также вызывает повреждение, высвобождая цитолитические молекулы. Здесь, однако, стоит отметить, что разные штаммы вида по-разному влияют на хозяина. Например, согласно исследованиям, слабопатогенные штаммы имеют тенденцию вызывать повреждение, попадая в ткани мозга, используя структуры чашки с едой.
С другой стороны, высокопатогенные штаммы сначала лизируют нервные клетки перед тем, как проглотить образующийся мусор.
Некоторые из молекул, которые были связаны с лизисом клеток, включают:
- Протеазы
- Кислотные гидролазы
- Фосфолипазы
- Фосфолиполитические ферменты
При проглатывании тканей мозга и высвобождении различных цитолитических молекул N. в конечном итоге вызывают амебный менингоэнцефалит (ПАМ), который является серьезной инфекцией центральной нервной системы.Это быстро возникает (острое молниеносное развитие) и может проявляться через 3–10 дней после контакта с инфицированной водой (или после того, как киста попала в организм через носовой ход).
Некоторые из симптомов, связанных с инфекцией, включают:
- Светобоязнь
- Головная боль — бифронтальная или битемпоральная
- Пиковая лихорадка
- Тошнота и рвота
- Жесткая шея
- Кома — кома может в конечном итоге привести к смерти нескольких дней после появления симптомов
* N.fowleri, как было показано, устойчивы к лизисному действию цитолитических молекул хозяина (например, фактора некроза опухоли и т. д.). Это одна из адаптаций, которая позволяет ему процветать в центральной нервной системе.
* Хотя N. fowleri распространен в различных частях мира, особенно в тропических регионах, большинство инфекций зарегистрировано в Соединенных Штатах.
Контроль и обработка
Naegleria fowleri чувствительны к хлору и не могут выжить в чистой воде.Следовательно, вода, которая была достаточно хлорирована (например, вода в бассейне и т. Д.), А также надлежащее поддержание воды — одни из лучших стратегий для контроля над организмом и минимизации возможных инфекций.
Однако в случае инфекции амфотерицин B обычно используется в лечебных целях. Хотя возбудитель очень чувствителен к препарату in vitro, не все пациенты выздоравливают после этого лечения.
Некоторые из других лекарств, используемых в лечебных целях, включают:
- Рифампицин
- Сульфадиазин
- Противоотечное лекарство
Вернуться к амебе под микроскопом
Вернуться из Naegleria Fowleri
000 Справочные материалы
Франсин Марчиано-Кабрал.(1988). Биология Naegleria spp.
Франсин Марчиано-Кабрал и Гай А. Кабрал. (2007). Иммунный ответ на амебы Naegleria fowleri и патогенез инфекции.
Muhammad Jahangeer et al. (2019). Naegleria fowleri: источники инфекции, патофизиология, диагностика и лечение; Обзор.
Рукаия Сиддики, Ибне Карим М. Али, Дженнифер Р. Коуп и Навид Ахмед Хан. (2016). Биология и патогенез Naegleria fowleri.
Ссылки
https://web.stanford.edu/group/parasites/ParaSites2010/Katherine_Fero/FeroNaegleriafowleri.htm
parasites / naegleria / index.html #: ~: text = Naegleria% 20fowleri% 20 (обычно% 20 относится к% 20, первичный% 20amebic% 20 менингоэнцефалит% 20 (PAM).
сообщить об этом объявленииA New Species of Fresh-Water Амеба: Amoeba algonquinensis n. Sp. (Gymnamoebia: Amoebidae) по JSTOR
AbstractНовый пресноводный вид Gymnamoebia в семействе Amoebidae, Amoeba algonquinensis n.sp., описывается на световом и электронно-микроскопическом уровнях. У этой амебы есть единственный доминантный псевдоподий и морулированный уроид. Он имеет одно сферическое зернистое ядро. Эти наблюдения сравниваются с предыдущими описаниями других крупных гимнамобий.
Информация для издателяWiley — глобальный поставщик контента и решений для рабочих процессов с поддержкой контента в областях научных, технических, медицинских и научных исследований; профессиональное развитие; и образование.Наши основные направления деятельности выпускают научные, технические, медицинские и научные журналы, справочники, книги, услуги баз данных и рекламу; профессиональные книги, продукты по подписке, услуги по сертификации и обучению и онлайн-приложения; образовательный контент и услуги, включая интегрированные онлайн-ресурсы для преподавания и обучения для студентов и аспирантов, а также для учащихся на протяжении всей жизни. Основанная в 1807 году компания John Wiley & Sons, Inc. уже более 200 лет является ценным источником информации и понимания, помогая людям во всем мире удовлетворять свои потребности и воплощать в жизнь их чаяния.Wiley опубликовал работы более 450 лауреатов Нобелевской премии во всех категориях: литература, экономика, физиология и медицина, физика, химия и мир. Wiley поддерживает партнерские отношения со многими ведущими мировыми обществами и ежегодно издает более 1500 рецензируемых журналов и более 1500 новых книг в печатном виде и в Интернете, а также базы данных, основные справочные материалы и лабораторные протоколы по предметам STMS. Благодаря постоянно растущему предложению открытого доступа, Wiley стремится к максимально широкому распространению и доступу к публикуемому нами контенту, а также поддерживает все устойчивые модели доступа.Наша онлайн-платформа, Wiley Online Library (wileyonlinelibrary.com), является одной из самых обширных в мире междисциплинарных коллекций онлайн-ресурсов, охватывающих жизнь, здоровье, социальные и физические науки и гуманитарные науки.
(PDF) Молекулярная филогения и классификация Lobose Amoebae
предполагают отсутствие слияния генов между
цитохромоксидазой 1 и 2 в этой кладе; особенность
, которую также разделяют Dictyostelium и
Acanthamoeba (Gray et al.1998). Базальное положение
Tubulinea также подтверждается данными по актину
(Fahrni et al. 2003; Nikolaev et al. 2005). Однако
порядок ветвления между основными линиями
Amoebozoa в анализе рРНК SSU совершенно не согласуется с
, как показывают разные деревья
, представленные Cavalier-Smith et al. (2004). Следовательно, необходимы дополнительные свидетельства
, чтобы установить отношения более высокого уровня —
внутри филума.
На пути к новой системе амеб
Морфологический анализ основных ветвей
, выявленный нашим филогенетическим анализом последовательностей SSU
рРНК лобозных амеб, позволяет нам (1)
переоценить важность
ультраструктурных
и морфологические признаки, используемые в систематике амеби (2), чтобы предложить новую высокоуровневую систему лобозных амеб
, где таксоны
основаны в основном на паттернах локомотивов (Таблица 2).
Эта классификационная система основана на предположении
о том, что локомотивный паттерн амебы
является составным признаком, отражающим
организацию ее цитоскелета, клеточной оболочки и
, возможно, ряда других основных характеристик (Smir-
ноя и Браун 2004; Смирнов и Гудков
1999).
Поскольку мы не нашли поддержки для разделения
филума Amoebozoa на два монофилетических подтипа
, Lobosa и Conosa, мы предлагаем отказаться от этих таксонов
.В новой системе мы делим
амебозоа на три класса. Большинство из
лобозных амебпринадлежат к одному из двух классов:
Tubulinea и Flabellinea, тогда как Archamoebae
и Mycetozoa относятся к третьему классу
Conosea. Tubulinea состоит из амеб, которые
всегда образуют или могут временно образовывать трубчатые псевдоподии
с моноаксиальным цитоплазматическим потоком,
, тогда как Flabellinea включает аттенуированные амебы
bae, которые никогда не образуют трубчатые псевдоподии
.В систематическом содержанииэти классы напоминают Lobo-
sea и Disosisa sensu Cavalier-Smith et al.
(2004), но мы полагаем, что предлагаемые новые названия
лучше отражают основную дихотомию между этими
таксонами и предотвращают путаницу, связанную с широким использованием названий «лобоса» и «лобоса»
. море » на разных таксономических рангах. В соответствии с
с доступными молекулярными данными, три порядка
можно выделить среди Tubulinea (Tubulinida, Lep-
,tomyxida и Arcellinida), а два порядка —
, предложенных в пределах Flabellinea (Dactylopodida
и Vannellida).Диагнозы этих новых
классов и порядковпредставлены в Таблице 2. A
морфологических семейств амеб
sensu Page 1987 кажутся монофилетическими (например,
Amoebidae и Vannellidae), в то время как некоторые из них
парафилетических или полифилетические (например, Hartmannellidae
и Vexilliferidae). Таким образом, для надлежащего пересмотра существующих семейств амеб
требуется более крупная выборка таксонов
; однако это выходит за рамки настоящей статьи
.Чтобы очертить систематический состав отрядов, но избежать искусственных группировок, оставлены все
родов, которые четко не ветвятся в рамках вышеупомянутых отрядов
и не имеют четких морфологических отношений
со своими членами
. incertae sedis в пределах соответствующего таксона более высокого уровня
, а также линий, по которым
молекулярные данные еще не доступны. Это относится к
, в частности к семейству Thecamoebidae, которое составляет
груши как полифилетический комплекс, и, таким образом,
не сохраняется в текущей версии нашей системы.
Роды еще не секвенированы, но явно морфло-
, родственные родам, уже представленные в дереве
, перечислены в соответствующих семействах как
«вероятно принадлежащие им».
На уровне классов и порядков система
, представленная в этом исследовании, демонстрирует некоторое сходство
с системой гимнамэбаэ, основанной на паттерне амебоидного движения
, который впоследствии был разработан Яном и Бови
. (1965), Jahn et al.(1974) и Бови (1985).
класс Tubulinea частично соответствует подотряду
компании Bovee Tubulina, который был в основном определен как
, следующим образом: «Цилиндроидное тело, разветвленное или не-
разветвленное, псевдоподы зернистые, прозрачная шапочка» (
) Отряд Vannellida частично соответствует подотряду Bovee Thecina,
не соответствует полностью, потому что Bovee (1985)
сгруппировал камоебид, ваннеллид, а
— некоторые малоизвестные группы амеб
—
. похожая морфология.Dactylopodida cor-частично отвечает на подотряд Бови Conopodina
, который он определил следующим образом: «Тело более или менее
треугольное или лопатчатое, с заостренным кончиком, с круглым концом
псевдоножек определенной длины, образованных из
. » (Бови, 1985). Недавнее обнаружение, что все семенниковые лобозные амебы, секвенированные к настоящему времени
, тесно связаны с семействами Amoebidae и
Hartmannellidae (Николаев и др., 2005), показывают, что
Jahn and Bovee (1965), Jahn et al.(1974) и
Bovee (1985) были правы, поместив семенниковые лобозы
амеб в подотряд Tubulina Bovee и
Ян, 1965. В свете этого открытия другие классификационные исследования разделяют голые и семенниковые лобковые амебы.
, например, Де’андре (1953), Лёблих и
,Таппан (1961) и Пейдж (1987), были искусственными,
СТАТЬЯ В ПРЕССЕ
Филогения и классификация Lobose Amoebae 137–
Жилище Testate Amoebae в БолгарииArcella arenaria Greeff, 1866
Батистома Arcella Deflandre, 1928
Arcella catinus Penard, 1890
Arcella dentata Ehrenberg, 1830
Arcella discoides Эренберг, 1871
Arcella gibbosa Penard, 1890
Arcella hemisphaerica Perty, 1852
Arcella intermedia (Deflandre, 1928) Цыганов и Мазей, 2006
Arcella rotundata Playfair, 1918
Arcella vulgaris Ehrenberg, 1830
Cyclopyxis eurystoma Deflandre, 1929
Cyclopyxis kahli Deflandre, 1929
Cyclopyxis puteus Thomas, 1960
Netzelia oviformis (Cash, 1909) Огден, 1979
Netzelia tuberculata (Wallich, 1864) Netzel, 1983
Trigonopyxis arcula (Leidy, 1879) Penard, 1912
Difflugia acuminata Ehrenberg, 1838
Difflugia ampullula Playfair, 1918
Difflugia bryophila (Penard, 1902) Jung, 1942
Difflugia elegans Penard, 1890
Difflugia globulosa Dujardin, 1837
Difflugia hiraethogii Ogden, 1983
Difflugia lanceolata Penard, 1890
Difflugia lobostoma Leidy, 1874
Difflugia lucida Penard, 1890
Difflugia oblonga Ehrenberg, 1831
Difflugia penardi Hopkinson, 1909
Difflugia pulex Penard, 1902
Difflugia pyriformis Perty, 1849
Difflugia rubescens Penard, 1891
Difflugia urceolata Carter, 1864
Difflugia viscidula Penard, 1902
Lesquereusia epistomium Penard, 1902
Lesquereusia gibbosa Thomas and Gauthier-Lièvre, 1859
Lesquereusia modesta Rhumbler, 1896
Lesquereusia spiralis (Ehrenberg, 1840) Bütschli, 1880
Centropyxis aculeata (Ehrenberg, 1830) Stein, 1857
Centropyxis aerophila Deflandre, 1929
Centropyxis cassis (Wallich, 1864) Дефландр, 1929
Centropyxis constricta (Ehrenberg, 1841) Penard, 1902
Centropyxis discoides (Penard, 1890) Deflandre, 1929
Centropyxis ecornis (Ehrenberg, 1841) Leidy, 1879
Centropyxis elongata (Penard, 1890) Thomas, 1959
Centropyxis gibba Deflandre, 1929
Centropyxis laevigata Penard, 1890
Centropyxis orbicularis Deflandre, 1929
Centropyxis plagiostoma Bonnet and Thomas, 1955
Платистома Centropyxis (Penard, 1890) Deflandre, 1929
Centropyxis spinosa (Cash, 1905) Deflandre, 1929
Bullinularia indica (Penard, 1907) Deflandre, 1953
Plagiopyxis callida Penard, 1910
Plagiopyxis declivis Bonnet et Thomas, 1955
Alabasta militaris (Penard, 1890) Дакерт, Бланденье, Косакян и Зингер, 2018
Gibbocarina galeata (Penard, 1890) Косакян, Лар, Мюло, Мейстерфельд, Митчелл и Лара, 2016
Hyalosphenia papilio (Leidy, 1874) Leidy, 1875
Longinebela ampulla Тодоров, Банков, Ганева, 2018
Longinebela golemanskyi (Тодоров, 2010) Косакян, Лар, Мюлот, Мейстерфельд, Митчелл и Лара, 2016
Longinebela penardiana (Deflandre, 1936) Косакян, Лар, Мюло, Мейстерфельд, Митчелл и Лара, 2016
Longinebela speciosa (Deflandre, 1936) Косакян, Лар, Мюло, Мейстерфельд, Митчелл и Лара, 2016
Longinebela tubulosa (Penard, 1890) Косакян, Лар, Мюлот, Мейстерфельд, Митчелл и Лара, 2016
Nebela aliciae Mitchell et Lara, 2013
Nebela collaris (Ehrenberg, 1848) Leidy, 1879
Nebela flabellulum Leidy, 1874
Nebela tincta (Leidy, 1879) Awerintzew, 1906
Padaungiella lageniformis (Penard, 1890) Lara et Todorov, 2012
Padaungiella nebeloides (Gauthier-Lièvre and Thomas, 1958) Lara et Todorov, 2012
Padaungiella tubulata (Brown, 1910) Lara et Todorov, 2012
Padaungiella wailesi (Deflandre, 1936) Lara et Todorov, 2012
Planocarina carinata (Archer, 1867) Косакян, Лар, Мюлот, Мейстерфельд, Митчелл и Лара, 2016
Quadrulella longicollis (Таранек, 1882)
Quadrulella simrica (Wallich, 1863) Cockerell, 1909
Quadrulella variabilis Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell and Lara, 2016
Heleopera petricola Leidy, 1879
Heleopera rosea Penard, 1890
Heleopera sphagni (Лейди, 1874)
Heleopera sylvatica Penard, 1890
Phryganella acropodia (Hertwig and Lesser, 1874) Гопкинсон, 1909
Phryganella hemisphaerica (Penard, 1890) Penard, 1902
Phryganella nidulus Penard, 1902
Дантистома Argynnia (Penard, 1890)
Argynnia vitraea (Пенард, 1899)
Awerintzewia cyclostoma (Penard, 1902) Schouteden, 1906
Lagenodifflugia bryophila (Penard, 1902) Ogden, 1987
Lagenodifflugia vas (Leidy, 1874) Medioli and Scott, 1983
Pontigulasia rhumbleri Hopkinson, 1919
Zivkovicia compressa (Carter, 1864) Ogden, 1983
Zivkovicia spectabilis (Penard, 1902) Огден, 1987
Archerella flavum (Арчер, 1877) Лёблих и Таппан, 1961
Assulina muscorum Greeff, 1888
Assulina semulum (Ehrenberg, 1848) Leidy, 1879
Euglypha bryophila Brown, 1911
Euglypha ciliata (Ehrenberg, 1848), Лейди, 1878
Euglypha compressa Carter, 1864
Euglypha cristata Leidy, 1874
Euglypha filifera Penard, 1890
Euglypha laevis (Ehrenberg, 1845) Perty, 1849
Euglypha rotunda Wailes, 1911
Euglypha strigosa (Ehrenberg, 1871) Лейди, 1878
Euglypha tuberculata Dujardin, 1841
Scutiglypha crenulata (Wailes, 1912) Foissner and Schiller, 2001
Sphenoderia fissirostris Penard, 1890
Sphenoderia labiata Thomas et Gauthier-Lièvre, 1959
Sphenoderia lenta Schlumberger, 1845
Sphenoderia minuta Deflandre, 1931
Sphenoderia ovoidea Jung, 1942
Trachelocorythion pulchellum (Penard, 1890) Bonnet, 1979
Corythion constricta (Certes, 1889) Jung, 1942
Corythion dubium Taránek, 1881
Playfairina valkanovi Golemansky, 1966
Trinema Complanatum Penard, 1890
Trinema enchelys (Эренберг, 1838) Лейди, 1878
Trinema galeata (Penard, 1890) Юнг, 1942
Линейный кинотеатр Penard, 1890
Campascus triqueter Penard, 1891
Cyphoderia amphoralis (Уэйлс и Пенард, 1911)
Cyphoderia ampulla (Ehrenberg, 1840) Leidy, 1878
Цифодерия большая (Penard, 1890)
Pareuglypha reticulata Penard, 1902
Tracheleuglypha acolla Bonnet et Thomas, 1955
Tracheleuglypha dentata (Moniez, 1888) Deflandre, 1928
Исследование потенциальной патогенности Willaertia magna путем изучения переноса генов патогенности бактерий в его геном
Панек Т. и Чепичка И. Разнообразие гетеролобозей. Genet. Дайверы. Microorg . 10,5772 / 35333 (2012).
Сиддики Р., Али И. К. М., Коуп Дж. Р. и Хан Н. А. Биология и патогенез Naegleria fowleri. Acta Trop. 164 , 375–394 (2016).
PubMed Статья PubMed Central Google Scholar
de Jonckheere, J. F., Dive, D. G., Pussard, M.И Викерман, К. Willaertia magna gen. nov., sp. ноя (Vahlkampfiidae), теплолюбивая амеба, обитающая в разных средах обитания. Protistologica Доступно по адресу https://eurekamag.com/research/001/281/001281223.php. (Дата обращения: 12 августа 2018 г.) (1984).
Ривера, Ф. и др. . Патогенные амебы в природных термальных водах трех курортов Идальго, Мексика. Environ. Res. 50 , 289–295 (1989).
CAS PubMed Статья PubMed Central Google Scholar
Адл, С. М. и др. . Изменения в классификации, номенклатуре и разнообразии эукариот. J. Eukaryot. Microbiol. 66 , 4–119 (2019).
PubMed PubMed Central Google Scholar
Фриц-Лейлин, Л. К. и др. . Геном Naegleria gruberi подчеркивает универсальность ранних эукариот. Cell 140 , 631–642 (2010).
CAS PubMed Статья PubMed Central Google Scholar
Zysset-Burri, D. C. и др. . Полногеномная идентификация факторов патогенности свободноживущей амебы Naegleria fowleri. BMC Genomics 15 (2014).
PubMed PubMed Central Статья CAS Google Scholar
Liechti, N., Schürch, N., Bruggmann, R. & Wittwer, M. Геном Naegleria lovaniensis, основа для сравнительного подхода к выяснению факторов патогенности патогенной амебы человека N.fowleri. BMC Genomics 19 (2018).
Робинсон, Б.С., Кристи, П.Э. и Де Джонкхир, Дж. Ф. Временная стадия жгутиконосца (мастиготы) у амебы vahlkampfiid Willaertia magna и ее возможное эволюционное значение. Biosystems 23 , 75–86 (1989).
CAS PubMed Статья PubMed Central Google Scholar
Schmitz-Esser, S. и др. . Разнообразие бактериальных эндосимбионтов экологических изолятов акантамеб. , заявл. Environ. Microbiol. 74 , 5822–5831 (2008).
CAS PubMed PubMed Central Статья Google Scholar
Греб, Г. и Рауль, Д. Микроорганизмы, устойчивые к свободноживущим амебам. Clin. Microbiol. Ред. 17 , 413–433 (2004).
PubMed PubMed Central Статья Google Scholar
Молинер, К., Рауль, Д. и Фурнье, П.-Э. Доказательства горизонтального переноса генов между амебой и бактериями. Clin. Microbiol. Заразить. 15 , 178–180 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Maumus, F. & Blanc, G. Изучение обмена генами между Acanthamoeba и гигантскими вирусами предлагает неизвестное семейство вирусов, инфицирующих амебы. Genome Biol.Evol. 8 , 3351–3363 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Кларк, М. и др. . Геном Acanthamoeba castellanii подчеркивает обширный латеральный перенос генов и раннюю эволюцию передачи сигналов тирозинкиназы. Genome Biol. 14 , R11 (2013).
PubMed PubMed Central Статья CAS Google Scholar
Челха Н. и др. . Филогеномное исследование черновых геномных последовательностей Acanthamoeba polyphaga предполагает генетический обмен с гигантскими вирусами. Фронт. Microbiol . 9 (2018).
Колсон, П., Скола, Б. Л., Левассер, А., Каэтано-Аноллес, Г. и Рауль, Д. Мимивирус: ведущий путь в открытии гигантских вирусов амеб. Nat. Rev. Microbiol. 15 , 243–254 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Роуботэм, Т. Дж. Выделение Legionella pneumophila из клинических образцов с помощью амеб и взаимодействие этих и других изолятов с амебами. J. Clin. Патол. 36 , 978–986 (1983).
CAS PubMed PubMed Central Статья Google Scholar
Гамильтон, К. А., Пруссин, А. Дж., Ахмед, В. и Хаас, К. Н. Вспышки болезни легионеров и лихорадки Понтиак 2006–2017 гг. Curr.Environ. Департамент здравоохранения 5 , 263–271 (2018).
CAS PubMed Статья PubMed Central Google Scholar
Pagnier, I., Merchat, M. & La Scola, B. Потенциально патогенные микроорганизмы, ассоциированные с амебой, в градирнях и меры борьбы с ними. Future Microbiol. 4 , 615–629 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Лау, Х. Ю. и Эшболт, Н. Дж. Роль биопленок и простейших в патогенезе Legionella: значение для питьевой воды. J. Appl. Microbiol. 107 , 368–378 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Килвингтон С. и Прайс Дж. Выживание Legionella pneumophila в цистах Acanthamoeba polyphaga после воздействия хлора. J. Appl.Бактериол. 68 , 519–525 (1990).
CAS PubMed Статья PubMed Central Google Scholar
Linder, J. W.-K. Эверт. Свободноживущие амебы, защищающие легионеллу в воде: верхушка айсберга? Scand. J. Infect. Дис. 31 , 383–385 (1999).
PubMed Статья PubMed Central Google Scholar
Дей Р., Боденнек Дж., Мамери М. О. и Пернин П. Свободноживущие пресноводные амебы различаются по своей чувствительности к патогенной бактерии Legionella pneumophila . FEMS Microbiol. Lett. 290 , 10–17 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Гершензон, Дж. И Дударева, Н. Функция терпеновых натуральных продуктов в мире природы. Nat. Chem. Биол. 3 , 408–414 (2007).
CAS PubMed Статья PubMed Central Google Scholar
Оппердос, Ф. Р., Де Йонкхере, Дж. Ф. и Тиленс, А. Г. М. Метаболизм Naegleria gruberi. Внутр. J. Parasitol. 41 , 915–924 (2011).
CAS PubMed Статья PubMed Central Google Scholar
Pagnier, I., Raoult, D. & La Scola, B. Выделение и идентификация устойчивых к амебе бактерий из воды в окружающей среде человека с помощью процедуры совместного культивирования Acanthamoeba polyphaga. Environ. Microbiol. 10 , 1135–1144 (2008).
CAS PubMed Статья PubMed Central Google Scholar
Огата, Х. и др. . Последовательность генома Rickettsia bellii освещает роль амеб в обмене генами между внутриклеточными патогенами. PLOS Genet. 2 , e76 (2006).
PubMed PubMed Central Статья CAS Google Scholar
Уэйтс, К. Б., Балиш, М. Ф. и Аткинсон, Т. П. Новое понимание патогенеза и выявления инфекций Mycoplasma pneumoniae. Future Microbiol. 3 , 635–648 (2008).
PubMed PubMed Central Статья Google Scholar
Рупник М., Уилкокс М. Х. и Гердинг Д. Н. Инфекция Clostridium difficile: новые разработки в эпидемиологии и патогенезе. Nat. Rev. Microbiol. 7 , 526–536 (2009).
CAS PubMed Статья PubMed Central Google Scholar
Бейкер-Остин, К. и Оливер, Дж. Д. Vibrio vulnificus: новое понимание смертельного условно-патогенного микроорганизма. Environ. Microbiol. 20 , 423–430 (2018).
PubMed Статья PubMed Central Google Scholar
Альтвегг, М., Гейсс, Х. К. и Фрейдж, Б. Дж. Аэромонады как патоген для человека. CRC Crit. Rev. Microbiol. 16 , 253–286 (1989).
CAS Статья Google Scholar
Там, К. К., О’Брайен, С. Дж., Адак, Г. К., Микинс, С. М. и Фрост, Дж. А. Campylobacter coli — важный патоген пищевого происхождения. J. Infect. 47 , 28–32 (2003).
CAS PubMed Статья PubMed Central Google Scholar
Sawai, T. et al. . Подвздошно-поясничный абсцесс, вызванный Parvimonas micra: клинический случай. J. Med. Отчеты о случаях 13 (2019).
Arzouni, J.-P. и др. . Человеческая инфекция, вызванная Leptospira fainei. Emerg. Заразить. Дис. 8 , 865–868 (2002).
PubMed PubMed Central Статья Google Scholar
да Кунья, К. Э. П. и др. . Заражение Leptospira kirschneri Серовар Моздок: первое сообщение из южного полушария. Am. J. Trop. Med. Hyg. 94 , 519–521 (2016).
PubMed PubMed Central Статья Google Scholar
Эйхингер, Л. и др. . Геном социальной амебы Dictyostelium discoideum. Nature 435 , 43–57 (2005).
CAS PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ Google Scholar
Родригес-Сарагоса, С. Экология свободноживущих амеб. Crit. Rev. Microbiol. 20 , 225–241 (1994).
PubMed Статья Google Scholar
Ван З. и Ву М. Сравнительный геномный анализ эндосимбионтов Acanthamoeba подчеркивает роль амеб как «плавильного котла», определяющего эволюцию риккетсий. Genome Biol. Evol. 9 , 3214–3224 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Юсуф, Ф. А., Сиддики, Р. и Хан, Н. А. Acanthamoeba castellanii генотипа Т4 является потенциальным хозяином в окружающей среде для Enterobacter aerogenes и Aeromonas hydrophila. Паразит. Векторы 6 , 169 (2013).
PubMed PubMed Central Статья CAS Google Scholar
Лоренцо-Моралес, Дж., Хан, Н. А. и Валочник, Дж. Обновленная информация о кератите Acanthamoeba: диагностика, патогенез и лечение. Паразит 22 (2015).
Матин А., Сиддики Р., Джаясекера С. и Хан Н. А. Возрастающее значение Balamuthia mandrillaris. Clin. Microbiol. Ред. 21 , 435–448 (2008).
CAS PubMed PubMed Central Статья Google Scholar
Барал Р. и Вайдья Б. Смертельный случай амебного энцефалита, маскирующегося под герпес. Oxf. Med. Дело Rep . 2018 (2018).
Stubhaug, T. T. и др. . Первичный амебный менингоэнцефалит со смертельным исходом у норвежского туриста, возвращающегося из Таиланда. JMM Case Rep . 3 (2016).
Сонг, К.-Дж. и др. . Белок теплового шока 70 Naegleria fowleri является важным фактором пролиферации и цитотоксичности in vitro . Parasitol. Res. 103 , 313–317 (2008).
PubMed Статья PubMed Central Google Scholar
Ху, В. Н., Копачик, В. и Бэнд, Р. Н. Клонирование и характеристика транскриптов, показывающих экспрессию генов, связанных с вирулентностью, у Naegleria fowleri. Infect. Иммун. 60 , 2418–2424 (1992).
CAS PubMed PubMed Central Google Scholar
Tiewcharoen, S. et al. . Активность хлорпромазина в отношении генов nfa1 и Mp2CL5 трофозоитов Naegleria fowleri. Здравоохранение (Н. Ю.) 03 , 166 (2011).
CAS Google Scholar
Ревиллер, Ф.Л., Сух, С.-Дж., Салливан, К., Кабанес, П.-А. & Марчиано-Кабрал, Ф. Выделение уникального мембранного белка из Naegleria fowleri. J. Eukaryot. Microbiol. 48 , 676–682 (2001).
CAS PubMed Статья PubMed Central Google Scholar
Курсон, Р. Т. и Браун, Т. Дж. Использование клеточных культур в качестве индикатора патогенности свободноживущих амеб. J. Clin. Патол. 31 , 1–11 (1978).
CAS PubMed PubMed Central Статья Google Scholar
Aldape, K., Huizinga, H., Bouvier, J. & McKerrow, J. Naegleria fowleri: характеристика секретируемой гистолитической цистеиновой протеазы. Exp. Паразитол. 78 , 230–241 (1994).
CAS PubMed Статья PubMed Central Google Scholar
Лам, К., Джамерсон, М., Кабрал, Г., Карлессо, А. М., Марчиано-Кабрал, Ф. Экспрессия матричных металлопротеиназ в Naegleria fowleri и их роль в инвазии центральной нервной системы. Микробиология 163 , 1436–1444 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Matin, A., Stins, M., Kim, K. S. & Khan, N. A. Balamuthia mandrillaris проявляет металлопротеазную активность. FEMS Immunol. Med. Microbiol. 47 , 83–91 (2006).
CAS PubMed Статья PubMed Central Google Scholar
De Jonckheere, J. Использование аксенической среды для дифференциации патогенных и непатогенных изолятов Naegleria fowleri. , заявл. Environ. Microbiol. 33 , 751–757 (1977).
PubMed PubMed Central Google Scholar
Wingett, S. W. & Andrews, S. FastQ Screen: инструмент для мультигеномного картирования и контроля качества. F1000 Исследование 7 (2018).
Болджер, А. М., Лозе, М. и Усадель, Б. Trimmomatic: гибкий триммер для данных последовательности Illumina. Биоинформатика 30 , 2114–2120 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Надалин, Ф., Vezzi, F. & Policriti, A. GapFiller: новый подход к сборке для заполнения пробелов в парных чтениях. BMC Bioinformatics 13 , S8 (2012).
PubMed PubMed Central Статья Google Scholar
Laetsch, D. R. & Blaxter, M. L. BlobTools: Исследование сборок генома. F1000 Исследование 6 , 1287 (2017).
Артикул Google Scholar
Хофф, К. Дж., Ланге, С., Ломсадзе, А., Бородовский, М. и Станке, М. BRAKER1: неконтролируемая аннотация генома на основе РНК-Seq с помощью GeneMark-ET и AUGUSTUS. Bioinforma. Oxf. Англ. 32 , 767–769 (2016).
CAS Статья Google Scholar
Ломсадзе А., Бернс П. Д. и Бородовский М. Интеграция картированных считываний РНК-Seq в автоматическое обучение алгоритма поиска эукариотических генов. Nucleic Acids Res. 42 , e119 (2014).
PubMed PubMed Central Статья CAS Google Scholar
Steijger, T. et al. . Оценка методов реконструкции транскриптов для RNA-seq. Nat. Методы 10 (2013).
CAS PubMed PubMed Central Статья Google Scholar
Stanke, M. & Morgenstern, B.AUGUSTUS: веб-сервер для предсказания генов у эукариот, допускающий определенные пользователем ограничения. Nucleic Acids Res. 33 , W465 – W467 (2005).
CAS PubMed PubMed Central Статья Google Scholar
Huerta-Cepas, J. et al. . Быстрая общегеномная функциональная аннотация посредством назначения ортологии с помощью eggNOG-Mapper. Mol. Биол. Evol. 34 , 2115–2122 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Huson, D. H. et al. . MEGAN Community Edition — Интерактивное исследование и анализ крупномасштабных данных секвенирования микробиома. PLoS Comput. Биол . 12 (2016).
PubMed PubMed Central Статья ОБЪЯВЛЕНИЯ CAS Google Scholar
Лехнер, М. и др. . Proteinortho: обнаружение (Co-) ортологов в крупномасштабном анализе. BMC Bioinformatics 12 , 124 (2011).
PubMed PubMed Central Статья Google Scholar
Контрерас-Морейра, Б. и Винуеса, П. GET_HOMOLOGUES, универсальный пакет программного обеспечения для масштабируемого и надежного анализа микробных пангеномов. , заявл. Environ. Microbiol. 79 , 7696–7701 (2013).
CAS PubMed PubMed Central Статья Google Scholar
Томпсон, Дж. Д., Хиггинс, Д. Г. и Гибсон, Т. Дж. КЛУСТАЛ W: повышение чувствительности последовательного последовательного последовательного взвешивания, штрафов за пропуски в зависимости от положения и выбора весовой матрицы. Nucleic Acids Res. 22 , 4673–4680 (1994).
CAS PubMed PubMed Central Статья Google Scholar
Кумар, С., Стечер, Г. и Тамура, К. MEGA7: молекулярно-эволюционный генетический анализ версии 7.0 для больших наборов данных. Mol. Биол. Evol. 33 , 1870–1874 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Эдгар Р.С. МЫШЦЫ: множественное выравнивание последовательностей с высокой точностью и высокой пропускной способностью. Nucleic Acids Res. 32 , 1792–1797 (2004).
CAS PubMed PubMed Central Статья Google Scholar
Прайс, М. Н., Дехал, П. С. и Аркин, А. П. FastTree: вычисление больших деревьев минимальной эволюции с профилями вместо матрицы расстояний. Mol. Биол. Evol. 26 , 1641–1650 (2009).
CAS PubMed PubMed Central Статья Google Scholar
Letunic, I. & Bork, P. Интерактивное древо жизни (iTOL) v3: онлайн-инструмент для отображения и аннотации филогенетических и других деревьев. Nucleic Acids Res. 44 , W242–245 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Chen, L. et al. . VFDB: справочная база данных по факторам вирулентности бактерий. Nucleic Acids Res. 33 , D325 – D328 (2005).
CAS PubMed PubMed Central Статья Google Scholar
Sucgang, R. et al. . Сравнительная геномика социальных амеб Dictyostelium discoideum и Dictyostelium purpureum. Genome Biol. 12 , R20 (2011).
CAS PubMed PubMed Central Статья Google Scholar
Лоренци, Х.А. и др. . Новая сборка, повторная аннотация и анализ генома Entamoeba histolytica выявили новые геномные особенности и информацию о содержании белка. PLoS Negl. Троп. Dis . 4 (2010).
PubMed PubMed Central Статья CAS Google Scholar
Таксономия клинически значимых микроорганизмов
Бактериальные патогены
В следующих таблицах перечислены виды и некоторые более высокие группы патогенных эубактерий, упомянутых в тексте.Классификация бактерий, одна из трех сфер жизни, находится в постоянном изменении, поскольку взаимосвязи становятся более ясными благодаря выборке генетических последовательностей. Многие группы на всех таксономических уровнях все еще имеют неопределенные отношения с другими членами филогенетического древа бактерий. Руководство Берджи по систематике архей и бактерий содержит опубликованный список и описания прокариотических видов. Таблицы здесь соответствуют таксономической организации в Bergey’s Manual Taxonomic Outline.
Мы разделили виды на таблицы, соответствующие различным типам бактерий. Таксономический ранг царства не используется в таксономии прокариот, поэтому типы являются подгруппой ниже домена. Обратите внимание, что многие типы бактерий не представлены в этих таблицах. Виды и роды перечислены только в классе внутри каждого типа. Названия, данные бактериям, регулируются Международным кодексом номенклатуры бактерий, который поддерживается Международным комитетом по систематике или прокариотам.
| Таблица 1. Тип Actinobacteria | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Актинобактерии | Коринебактерии | дифтерии | Диптерия |
| Гарднерелла | вагинальный | Бактериальный вагиноз | |
| Микрококк | Оппортунистические инфекции | ||
| Mycobacterium | bovis | Туберкулез, преимущественно крупного рогатого скота | |
| Mycobacterium | лепры | Болезнь Хансена | |
| Mycobacterium | туберкулез | Туберкулез | |
| Propionibacterium | прыщей | Угри, блефарит, эндофтальмит | |
| Таблица 2.Тип Bacteroidetes | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Бактероидии | Порфиромонас | Заболевания пародонта | |
| Prevotella | интермедиа | Заболевания пародонта | |
| Таблица 3. Тип Chlamydiae | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Хламидии | Хламидиоз | psittaci | Пситтакоз |
| Хламидиоз | трахоматис | Хламидиоз, передающийся половым путем | |
| Таблица 4.Тип фирмы | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Бациллы | Бациллы | антрацис | Сибирская язва |
| Bacillus | cereus | Диарейное и рвотное пищевое отравление | |
| Листерия | monocytogenes | Листериоз | |
| Энтерококк | faecalis | Эндокардит, сепсис, инфекции мочевыводящих путей, менингит | |
| Стафилококк | золотистый | Кожные инфекции, гайморит, пищевое отравление | |
| Стафилококк | эпидермис | Нозокомиальные и оппортунистические инфекции | |
| Стафилококк | человека | Оппортунистические инфекции | |
| Стафилококк | saprophyticus | Инфекции мочевыводящих путей | |
| Стрептококк | агалактии | Послеродовая инфекция, неонатальный сепсис | |
| Стрептококк | мутанов | Кариес зуба | |
| Стрептококк | пневмонии | Пневмония, многие другие инфекции | |
| Стрептококк | пиоген | Фарингит, скарлатина, импетиго, некротический фасцит | |
| Клостридия | Клостридиум | ботулинический | Отравление ботулином |
| Клостридиум | difficile | Колит | |
| Клостридиум | perfringens | Пищевое отравление, газовая гангрена | |
| Клостридиум | тетани | Столбняк | |
| Таблица 5.Тип Fusobacteria | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Фузобактерии | Fusobacterium | Заболевания пародонта, синдром Лемьера, кожные язвы | |
| Streptobacillus | moniliformis | Крысиная лихорадка | |
| Таблица 6. Тип Proteobacteria | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Alphaproteobacteria | Анаплазма | фагоцитофил | Гранулоцитарный анаплазмоз человека |
| Бартонелла | henselae | Пелиозный гепатит, бациллярный ангиоматоз, эндокардит, бактериемия | |
| Бартонелла | кинтана | Траншейная лихорадка | |
| Бруселла | melitensis | Бруцеллез овец | |
| Эрлихия | чаффеенсис | Моноцитарный эрлихиоз человека | |
| Риккетсия | prowazekii | Эпидемический сыпной тиф | |
| Риккетсия | риккетсии | Пятнистая лихорадка Скалистых гор | |
| Риккетсия | тиф | Мышиный сыпной тиф | |
| Бетапротеобактерии | Бордетелла | коклюш | коклюш |
| Эйкенелла | Инфекции, вызванные укусом | ||
| Neisseria | гонореи | Гонорея | |
| Neisseria | meningitidis | Менингит | |
| Спириллум | минус (альт.несовершеннолетний) | Содоку (лихорадка от укусов крыс) | |
| Эпсилонпротеобактерии | Campylobacter | Jejuni | Гастроэнтерит, синдром Гийена-Барре |
| Helicobacter | пилори | Язва желудка | |
| Гаммапротеобактерии | Аэромонас | гидрофила | Дизентерийный гастроэнтерит |
| Coxiella | бурнетии | Ку-лихорадка | |
| Enterobacter | Мочевые и респираторные инфекции | ||
| Эшерихия | кишечная палочка Штаммы: продуцирующий токсин шига (STEC) (e.g., O157: H7), также называемый энтерогеморрагическим E. coli (EHEC) или продуцирующим вероцитотоксин E. coli (VTEC) | Вспышки диареи пищевого происхождения, геморрагический колит, гемолитико-уремический синдром | |
| Эшерихия | кишечная палочка Штамм: энтеротоксигенный E. coli (ETEC) | Диарея путешественника | |
| Эшерихия | кишечная палочка Штамм: энтеропатогенный E.coli (EPEC) | Диарея, особенно у детей раннего возраста | |
| Эшерихия | кишечная палочка Штамм: энтероагрегант E. coli (EAEC) | Диарейные болезни у детей и путешественников | |
| Эшерихия | кишечная палочка Штамм: диффузно прикрепляется E. coli (DAEC) | Диарейные болезни у детей | |
| Эшерихия | кишечная палочка Штамм: энтероинвазивный E.coli (EPEC) | Бациллярная дизентерия, клетки проникают в эпителиальные клетки кишечника | |
| Франсиселла | tularensis | Туляремия | |
| Haemophilus | ducreyi | Шанкроид | |
| Haemophilus | гриппа | Бактериемия, пневмония, менингит | |
| Клебсиелла | пневмонии | Пневмония, внутрибольничные инфекции | |
| Легионелла | пневмофила | Болезнь легионеров | |
| Моракселла | catarrhalis | Средний отит, бронхит, синусит, ларингит, пневмония | |
| Пастерелла | Пастереллез | ||
| Плезиомонас | шигеллоиды | Гастроэнтерит | |
| Протей | Оппортунистические инфекции мочевыводящих путей | ||
| Псевдомонады | aeruginosa | Оппортунистическая, нозокомиальная пневмония и сепсис | |
| Сальмонелла | бонгори | Сальмонеллез | |
| Сальмонелла | энтерика | Сальмонеллез | |
| Серратия | Пневмония, инфекции мочевыводящих путей | ||
| Шигелла | boydii | Дизентерия | |
| Шигелла | дизентерии | Дизентерия | |
| Шигелла | Flexneri | Дизентерия | |
| Шигелла | сонней | Дизентерия | |
| Vibrio | холеры | Холера | |
| Vibrio | parahemolyticus | Гастроэнтерит с морепродуктами | |
| Vibrio | vulnificus | Гастроэнтерит из морепродуктов, некротические раневые инфекции, сепсис | |
| Иерсиния | энтероколитика | Иерсиниоз | |
| Иерсиния | pestis | Чума | |
| Иерсиния | псевдотуберкулез | Дальневосточная скарлатина | |
| Таблица 7.Тип Spirochaetes | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Спирохетии | Боррелия | Бургдорфери | Болезнь Лайма |
| Borrelia | hermsii | Клещевой возвратный тиф | |
| Borrelia | recurrentis | Возвратный тиф от вшей | |
| Лептоспира | опросов | Лептоспироз | |
| Трепонема | паллидум | Сифилис, беджель, пинта, фрамбезия | |
| Таблица 8.Тип Tenericutes | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Молликутес | Микоплазма | гениталий | Уретрит, цервицит |
| Mycoplasma | человека | Воспалительные заболевания органов малого таза, бактериальный вагиноз | |
| Mycoplasma | пневмонии | Mycoplasma пневмония | |
| Уреаплазма | urealyticum | Уретрит, инфекции плода | |
Вирусные патогены
Существует несколько систем классификации вирусов.Международный комитет по таксономии вирусов (ICTV) — это международный научный орган, отвечающий за правила классификации вирусов. Используемая здесь система ICTV группирует вирусы на основе генетического сходства и предполагаемой монофилии. Система классификации вирусов отделена от системы классификации клеточных организмов. Система ICTV группирует вирусы по семи порядкам, которые содержат родственные семейства. В настоящее время существует большое количество неназначенных семей с неизвестным родством с семью орденами.Три из этих отрядов заражают только эубактерии, археи или растения и не отображаются в этой таблице. Некоторые семейства можно разделить на подсемейства. Также есть много неназначенных родов. Как и все таксономии, вирусная систематика находится в постоянном движении. Последний полный список видов и классификацию можно получить на веб-сайте ICTV.
Таблица 9.
Таблица 10.
Грибковые патогены
Грибы — одно из царств владений Эукарии. Грибы наиболее тесно связаны с животными и несколькими другими небольшими группами и более отдаленно связаны с растениями и другими группами, которые ранее считались простейшими.В настоящее время грибы разделены на семь типов (или подразделений, оставшихся с тех пор, когда грибы изучались с помощью растений), но есть неопределенности в отношении некоторых взаимосвязей. Многие группы грибов, особенно те, которые ранее были отнесены к типу Zygomycota, который не был монофилетическим, имеют неопределенные отношения с другими грибами. В эту категорию попадает один вид, указанный в этой таблице — Rhizopus arrhizus. Названия грибов регулируются Международным кодексом номенклатуры водорослей, грибов и растений, но Международная комиссия по таксономии грибов (ICTF) также способствует таксономической работе по грибам.Одним из направлений деятельности ICTF является публикация изменений в названиях важных с медицинской и иных точек зрения видов грибов. Многие виды, которые раньше имели два названия (одно для половой формы и одно для бесполой формы), теперь объединяются под одним названием.
| Таблица 11. Грибковые патогены | |||
|---|---|---|---|
| Дивизион | Род | Виды | Родственные болезни |
| Аскомикота | Aspergillus | flavus | Оппортунистический аспергиллез |
| Aspergillus | фумигатус | Оппортунистический аспергиллез | |
| Blastomyces | дерматитидис | Бластомикоз | |
| Кандида | альбиканс | Молочница (кандидоз) | |
| Coccidioides | immitis | Лихорадка долины (кокцидиоидомикоз) | |
| Epidermophyton | Tinea corporis (стригущий лишай), tinea cruris (болевой зуд), tinea pedis (альтлетическая стопа), tinea unguium (онихомикоз) | ||
| Гистоплазма | капсула | Гистоплазмоз | |
| Microsporum | Tinea capitis (стригущий лишай), tinea corpus (стригущий лишай), другие дерматофитозы | ||
| Pneumocystis | jirovecii | Оппортунистическая пневмония | |
| Sporothrix | щенский | Споротрихоз (болезнь кустов роз) | |
| Trichophyton | mentagrophytes var. межцифровый | Tinea barbae (зуд парикмахера), дерматофитозы | |
| Trichophyton | рубрум | Tinea corporis (стригущий лишай), tinea cruris (болевой зуд), tinea pedis (альтлетическая стопа), tinea unguium (онихомикоз) | |
| Basidiomycota | Криптококк | neoformans | Оппортунистический криптококкоз, грибковый менингит, энцефалит |
| Malassezia | Перхоть, разноцветный лишай | ||
| неопределенно | Ризопус | arrhizus | Мукормикоз |
Патогены простейших
Взаимоотношения между организмами (и, следовательно, их таксономия), ранее сгруппированные под названием Протисты, поняты лучше, чем два или три десятилетия назад, но эта работа все еще продолжается.В 2005 году эукарии разделились на шесть супергрупп. Последняя классификация высокого уровня объединила две из предыдущих супергрупп, чтобы создать систему, состоящую из пяти супергрупп. Эта классификация была разработана для Общества протозоологов, но это не единственный предлагаемый подход. В одну из пяти супергрупп входят животные, грибы и несколько более мелких групп простейших. Другой содержит зеленые растения и три группы водорослей. Остальные три супергруппы (перечисленные в трех таблицах ниже) содержат других простейших, многие из которых вызывают заболевания.Кроме того, существует большое количество групп протистов, отношения которых не выяснены. В трех представленных здесь супергруппах мы указали тип, к которому принадлежат перечисленные патогены.
| Таблица 12. Супергруппа Amoebozoa | |||
|---|---|---|---|
| Тип | Род | Виды | Родственные болезни |
| Амёбозоа | Акантамеба | Гранулематозный амебный энцефалит, акантамебный кератит | |
| Entamoeba | гистолитика | Энтеробиоз | |
| Таблица 13.Супергруппа SAR (Stramenopiles, Alveolata, Rhizaria) | |||
|---|---|---|---|
| Тип | Род | Виды | Родственные болезни |
| Apicomplexa | Бабезия | Бабезиоз | |
| Криптоспоридиум | человека | Криптоспоридиоз | |
| Криптоспоридиум | парвум | Криптоспоридиоз | |
| Cyclospora | cayetanensis | Гастроэнтерит | |
| Плазмодий | falciparum | Малярия | |
| Плазмодий | малярии | «Доброкачественная» или «четырехдневная» (3-дневная рецидивирующая лихорадка) малярия | |
| Плазмодий | овальный | «Терцианская» (2-дневная рецидивирующая лихорадка) малярия | |
| Плазмодий | vivax | «Доброкачественная» «третичная» (2-дневная рецидивирующая лихорадка) малярия | |
| Плазмодий | knowlesi | Малярия приматов, способная вызывать зооноз, банальную лихорадку | |
| Токсоплазма | gondii | Токсоплазмоз | |
| Таблица 14.Супергруппа Excavata | |||
|---|---|---|---|
| Тип | Род | Виды | Родственные болезни |
| Метамонада | Лямблии | лямблия | Лямблиоз |
| Трихомонада | вагинальный | Трихомониаз | |
| Euglenozoa | Лейшмания | braziliensis | Лейшманиоз |
| Лейшмания | доновани | Лейшманиоз | |
| Лейшмания | тропика | Кожный лейшманиоз | |
| Трипаносома | Брюсей | Африканская сонная болезнь (африканский трипаносомоз) | |
| Трипаносома | Крузи | Болезнь Шагаса | |
| Перколозоа | Нэглерия | Фаулери | Первичный амебный менингоэнцефалит (наэглериаз) |
Паразитарные гельминты
Таксономия паразитических червей, все из которых принадлежат к царству Animalia, все еще содержит много неопределенностей.Патогенные виды встречаются в двух типах: нематоды, или круглые черви, и платигельминты, или плоские черви. Нематоды условно делятся на два класса, один из которых, Chromadorea, вероятно, содержит неродственные группы. Паразитические плоские черви содержатся в трех классах плоских червей, два из которых важны для человека: трематоды и цестоды.
| Таблица 15. Тип Nematoda | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Хромадорея | Анкилостома | канинум | Инфекция анкилостомоза собак |
| Анкилостома | двенадцатиперстная кишка | Инфекция анкилостомоза Старого Света | |
| Аскарида | поясничный | Аскаридоз | |
| Энтеробиус | вермикулярная | Энтеробиоз (булавочный червь) | |
| Лоа | лоа | Loa loa filariasis (глазной червь) | |
| Necator | американский | Некаториоз (анкилостомоз Нового Света) | |
| Strongyloides | stercoralis | Стронгилоидоз | |
| Enoplea | Трихинелла | спираль | Трихинеллез |
| Тричурис | тричиура | Трихуриоз (инфекция плетеного червя) | |
| Таблица 16.Тип Platyhelminthes | |||
|---|---|---|---|
| Класс | Род | Виды | Родственные болезни |
| Трематода | Клонорчис | китайский язык | Печеночная двуустка китайская |
| Фасциолопсис | буски | Фасциолопсиоз | |
| фасциола | гигантика | Фасциолез | |
| фасциола | гепатит | Фасциолез | |
| Описторхис | кошачьих | Описторхоз | |
| Описторхис | виверрини | Описторхоз | |
| Schistosoma | гематобий | Мочевой шистосомоз | |
| Schistosoma | японская | Шистосомоз | |
| Schistosoma | mansoni | Кишечный шистосомоз | |
| Цестода | Дифиллоботрий | латум | Дифиллоботриоз |
| Эхинококк | гранулезная | Эхинококкоз (кистозный эхинококкоз) | |
| Эхинококк | multilocularis | Эхинококкоз | |
| Taenia | азиатка | Тениоз кишечника | |
| Taenia | сагината | Тениоз кишечника | |
| Taenia | солиум | Тениоз кишечника, цистицеркоз | |
Идентификационных ключей и иллюстрированных монографий
Не стесняйтесь добавлять то, что у вас есть!
Обратите внимание, что таксономия раковинных амеб очень далека от надежности.Текущая работа, сочетающая молекулярные и морфологические подходы, обнаруживает огромное неожиданное разнообразие. В результате необходимо будет переописать большинство видов с помощью документации световой и сканирующей электронной микроскопии высокого разрешения, данных биометрии и молекулярных данных (например, в идеале SSU и COX). Это огромная задача, но она стоит того!
Тем не менее, мы даем некоторые ключи для общих видов, поскольку даже при несовершенной систематике семенников анализ амеб является очень мощным и полезным инструментом в палеоэкологии и других приложениях, таких как биомониторинг.Виды, перечисленные в этих ключах, в основном следует рассматривать как морфо-виды и даже как комплексы видов. Конечно, приветствуются любые предложения по улучшению этих ключей!
Приведен список ссылок, пожалуйста, добавьте сюда всю необходимую информацию!
Атлас сфагновых раковинных амеб в Болгарии Хотя немногие из нас могут иметь возможность посещать Болгарию на регулярной основе, эта книга может побудить нас посетить эту прекрасную страну! Поскольку многие семенниковые амебы из сфагновых торфяников имеют широкое географическое распространение (без учета скрытых видов и в пределах таксонов, контрастирующих с диапазонами распространения), эта превосходная монография, безусловно, будет очень полезным справочником для обучения и исследований.Поздравляем и благодарим Милчо и Ноколу за эту работу! [https://ab.pensoft.net/article/38685/]. Милчо Тодоров, Никола Банков (2019) Атлас сфагновых семенников в Болгарии
Всем! Недавно выпущенный, но, скорее всего, не замеченный большей частью сообщества, Андрей Цыганов и его коллеги опубликовали чрезвычайно полезное руководство по тестированию амеб с ключами к родам Цыганов, А.Н., Бабешко, К.В. И Мазей Ю.А. (2016) Руководство по тестированию амеб с помощью ключей к родам — Монография.Издательство Пензенского государственного университета, Пенза, Россия. Это упрощенная и обновленная версия более крупной монографии, опубликованной Юрием Мазеем и Андреем Цыгановым на русском языке в 2006 г. [Mazei, Y. & Tsyganov, A. (2006) Freshwater Testate Amoebae. KMK Sci Press, Москва.], Который оказался очень полезным для многих из нас — даже если большинство из нас могло читать только латинские имена, смотреть на изображения и понимать информацию о размере, это уже было очень полезно! Большое спасибо за эту прекрасную работу!
Arcellinida
Определитель Nebela и подобных таксонов
Определитель Centropyxis и подобных таксонов
Определитель Heleopera
Euglyphida
Ключ для Trinema
Ключ для Euglypha
Ключ для Euglypha Decloitre 1962
Общие руководства по идентификации
Гроспитш 1958 монография
Харниш 1959 монография
Мазей-Цыганов 2006 Пресноводные семенниковые амебы
Исторические монографии
Лейди Дж.1879 Freswhater Rhizpopds Северной Америки
Penard E. 1902 La faune Rhizopodique du Bassin du Léman
Список литературы
Документы по филогении и таксономии:
- Bhattacharya, D., Helmchen, T. & Melkonian, M. (1995) Молекулярный эволюционный анализ кодируемой ядром малой субъединицы рибосомной РНК позволяет идентифицировать независимую линию ризопод, содержащую Euglyphina и Chlorarachniophyta. Журнал эукариотической микробиологии, 42, 65-69.
- Heger TJ, Mitchell EAD, Todorov M, Golemansky V, Lara E, Pawlowski J.2010. Молекулярная филогения семенников эвглифидных амеб (Cercozoa: Euglyphida) предполагает, что переходы между морской супралиторалью и пресноводной / наземной средами происходят нечасто. Молекулярная филогенетика и эволюция 55: 113-122. DOI: 10.1016 / j.ympev.2009.11.023
- Heger TJ. Павловски Дж., Големански В., Тодоров М., Лара Е., Митчелл Е. Д.. 2010. Сравнение потенциальных штрих-кодов COI и SSU рДНК для оценки разнообразия и филогенетических отношений цифодериидных семенниковых амеб (Rhizaria: Euglyphida).Протист. DOI: 10.1016 / j.protis.2010.05.002
- Лар, Д.Дж.Г. И Лопес, S.G.B.C. (2006) Морфология, биометрия, экология и биогеография пяти видов Difflugia Leclerc, 1815 (Arcellinida: Difflugiidae) из реки Тиете, Бразилия. Acta Protozoologica, 45, 77.
- Лар, Д.Дж.Г. И Лопес, S.G.B.C. (2007) Ультраструктура и биометрия трех лобозно-семенниковых амеб семейства Lesquereusiidae (Tubulinea: Arcellinida) на основе образцов из Сан-Паулу, Бразилия. Acta Protozoologica, 46, 339–348.
- Лар Д.Дж.Г., Бергманн П.Дж. и Лопес С. (2008) Таксономическая идентичность микробных эукариот: практический подход с использованием семенниковой амебы Centropyxis для разрешения конфликтов между старыми и новыми таксономическими описаниями. Журнал эукариотической микробиологии, 55, 409-416.
- Lara E, Heger TJ, Mitchell EAD, Meisterfeld R, Ekelund F. 2007. рРНК SSU выявляет последовательное увеличение сложности раковины у семенниковых амеб Euglyphid (Rhizaria: Euglyphida). Protist 158: 229-237.DOI: 10.1016 / j.protis.2006.11.006
- Лара Е., Хегер Т.Дж., Экелунд Ф, Ламентович М., Митчелл Е.А. 2008. Гены рибосомной РНК бросают вызов монофилии Hyalospheniidae (Amoebozoa: Arcellinida). Протист 159 (2): 165-176. DOI: 10.1016 / j.protis.2007.09.003
- Лара Э, Хегер Т.Дж., Митчелл Э.Д. Ген COI обнаруживает зависящее от размера высокое расселение и низкое внутривидовое разнообразие у свободноживущих наземных простейших (Euglyphida; Assulina). Журнал биогеографии. DOI: 10.1111 / j.1365-2699.2010.02426.x
- Николаев С.И., Митчелл Е.А., Петров Н.Б., Берни С., Фарни Дж., Павловски Дж., 2005. Семенниковые лобозные амебы (отряд Arcellinida Kent, 1880) наконец нашли свой дом среди амебозоа. Протист, 156 (2): 191-202.
- Текле, Ю.И., Грант, Дж., Андерсон, О.Р., Нерад, Т.А., Коул, Дж. К., Паттерсон, Д. Дж. И Кац, Л.А. (2008) Филогенетическое размещение разнообразных амеб, выведенное из мультигенного анализа и оценки стабильности клады внутри «Amoebozoa» после удаления различных классов скорости SSU-рДНК.Молекулярная филогенетика и эволюция, 47, 339-352.
- Тодоров М., Големански В., Митчелл Е.А., Хегер Т.Дж. 2009. Морфология, биометрия и таксономия пресноводных и морских интерстициальных Cyphoderia (Cercozoa: Euglyphida). Журнал эукариотической микробиологии 56 (3): 279-289. DOI: 10.1111 / j.1550-7408.2009.00394.x
- Wylezich, C., Meisterfeld, R., Meisterfeld, S. & Schlegel, M. (2002) Филогенетический анализ малых субъединиц кодирующих областей рибосомной РНК выявляет монофилетический клон семенниковых амеб эвглифид (отряд Euglyphida).Журнал эукариотической микробиологии, 49, 108-118.
Статьи, в которых обсуждаются проблемы таксономии раковинных амеб (они иллюстрируют некоторые «горячие» дебаты »)
- Финли, Б.Дж., Эстебан, Г.Ф. & Фенчел, Т. (2004) Разнообразие протистов отличается? Протист, 155, 15-22.
- Heger TJ, Mitchell EAD, Ledeganck P, Vincke S, Van De Vijver B, Beyens L. 2009. Проклятие таксономической неопределенности в биогеографических исследованиях свободноживущих наземных протистов: тематическое исследование семенниковых амеб из журнала Amsterdam Island Journal of Biogeography 36: 1551-1560.