2. Структурные отличия обыкновенных и раковидных амеб. Перечислите основные отличия свободноживущих и паразитических корненожек.
По
другому их называют Thecamoebina — раковинные
корненожки (Testacea), отряд простейших
класса саркодовых. По
своему строению раковинные корненожки
напоминают амеб. В отличие от них часть
протоплазматического тела корненожек
заключена внутри раковинки, играющей
роль защитного образования,
размером 50 -150 мкм, в полости которой
помещается большая часть тела, в том
числе и ядро. Форма их разная — грушевидная,
округлая и др. Разный и материал, из
которого изготовлены раковинки. Это
либо псевдохитин, либо песчинки или
другие твердые частички, предварительно
проглоченные амебой, а потом выложенные
на эктоплазме в виде раковинки. У
раковинки есть отверстие — устье. Через
него амеба выпускает наружу псевдоподии.
Размножаются раковинные амебы простым
делением. Деление амеба начинает с
построения новой (второй) раковинки.
Примерно половина ее протоплазмы как
бы вываливается из устья и строит на
своей поверхности вторую раковинку. Одновременно делится ядро и мигрирует
в протоплазму этой раковинки. Какое-то
время получившиеся из одной две амебы
еще связаны между собой прослойкой
протоплазмы, причем раковинки лежат
устьями друг к другу. Затем эта прослойка
исчезает, и амебы расходятся.
На этой стадии обе особи оказываются
еще связанными друг с другом мостиком
цитоплазмы и обе раковинки (старая и
вновь образовавшаяся) направлены одна
к другой устьями. Вскоре после этого
цитоплазматический мостик между особями
утончается и перешнуровывается и обе
корненожки переходят к самостоятельному
существованию. По существу этот процесс
мало чем отличается от деления амеб, он
несколько усложнен лишь процессом
образования новой раковинки.
Одновременно делится ядро и мигрирует
в протоплазму этой раковинки. Какое-то
время получившиеся из одной две амебы
еще связаны между собой прослойкой
протоплазмы, причем раковинки лежат
устьями друг к другу. Затем эта прослойка
исчезает, и амебы расходятся.
На этой стадии обе особи оказываются
еще связанными друг с другом мостиком
цитоплазмы и обе раковинки (старая и
вновь образовавшаяся) направлены одна
к другой устьями. Вскоре после этого
цитоплазматический мостик между особями
утончается и перешнуровывается и обе
корненожки переходят к самостоятельному
существованию. По существу этот процесс
мало чем отличается от деления амеб, он
несколько усложнен лишь процессом
образования новой раковинки.
Обитают раковинные амёбы в больших количествах: в прудах, моховых болотах и других пресных водоёмах.
Медицинская
паразитология изучает особенности строения и жизненных
циклов паразитов, взаимоотношения в
системе паразит-хозяин, а также методы
диагностики, лечения и профилактики
инвазионных болезней.
Организмы разных видов в биоценозах находятся в постоянном взаимодействии друг с другом. Одним из его видов является паразитизм. Это форма межвидовых взаимоотношений, при которой один вид использует другой как источник питания и среду обитания.
Формы паразитизма чрезвычайно многообразны, и классификация их возможна по разным основаниям. С точки зрения обязательности паразитического образа жизни для данного вида различают истинный и ложный, а также облигатный и факультативный паразитизм.
При истинном паразитизме взаимоотношения между паразитом и хозяином являются закономерными и имеют эволюционную основу. При ложном паразитизме в нормальных условиях данный вид ведет свободный образ жизни. При попадании в организм хозяина ложный паразит может некоторое время сохранять жизнеспособность и нарушать жизнедеятельность хозяина.
 Факультативные
паразиты способны вести свободный образ жизни,
но, попадая в организм хозяина, проходят
в нем часть цикла своего развития и
нарушают его жизнедеятельность.
Факультативные
паразиты способны вести свободный образ жизни,
но, попадая в организм хозяина, проходят
в нем часть цикла своего развития и
нарушают его жизнедеятельность.По времени контакта хозяина и паразита паразитизм бывает временным и постоянным. Временные паразиты обычно посещают хозяина только для питания. Постоянные паразиты подразделяются на стационарных и периодических. Стационарные паразиты всю жизнь проводят на хозяине или внутри него. Периодические паразиты часть своего жизненного цикла проводят в паразитическом состоянии, остальное время обитают свободно.
Нередко паразитический образ жизни ведут только личинки, в то время как половозрелые формы являются свободноживущими. Паразитизм такого рода называют ларвальным (личиночным). Противоположное явление, когда паразитом является половозрелая форма, а личинка обитает в открытой природе, называют шлагинальным паразитизмом.
Особенно
большое медицинское значение имеет
классификация паразитов по
их локализации в организме хозяина. Эктопаразиты находятся на покровах хозяина. Эндопаразиты обитают внутри хозяина. Их подразделяют
на паразитов, обитающих в полостных
органах, связанных с внешней средой, и
паразитов тканей внутренней среды.
Эктопаразиты находятся на покровах хозяина. Эндопаразиты обитают внутри хозяина. Их подразделяют
на паразитов, обитающих в полостных
органах, связанных с внешней средой, и
паразитов тканей внутренней среды.
Известно около 1400 видов споровиков. Все представители класса являются паразитами (или комменсалами) человека и животных. Многие споровики — внутриклеточные паразиты. Именно эти виды претерпели наиболее глубокую дегенерацию в плане строения: их организация упрощена до минимума. Они не имеют никаких органов выделения и пищеварения. Питание происходит
за
счет поглощения пищи всей поверхностью
тела. Продукты жизнедеятельности также
выделяются через всю поверхность
мембраны. Органелл дыхания нет. Общими
чертами всех представителей класса
являются отсутствие у зрелых форм
каких-либо органелл движения, а также
сложный жизненный цикл. Для споровиков
характерны два варианта жизненного
цикла — с наличием полового процесса
и без него. Первый вариант цикла включает
в себя стадии бесполого размножения и
полового процесса (в виде копуляции и
спорогонии).
Бесполое размножение осуществляется простым делением с помощью митоза или множественным делением(шизогонией). При шизогонии происходит многократное деление ядра без цитокинеза. Затем вся цитоплазма разделяется на части, которые обособляются вокруг новых ядер. Из одной клетки образуется очень много дочерних. Перед половым процессом происходит образование мужских и женских половых клеток — гамет. Они называются гамонтами. Затем разнополые гаметы сливаются с образованием зиготы. Она одевается плотной оболочкой и превращается в цисту, в которой происходит спорогония — множественное деление с образованием клеток (спорозоитов). Именно на стадии спорозоита паразит и проникает в организм хозяина. Споровики, для которых характерен именно такой цикл развития, обитают в тканях внутренней среды организма человека (например, малярийные плазмодии).
Второй
вариант жизненного цикла намного проще
и состоит из стадии цисты и трофозоита (активно
питающейся и размножающейся формы
паразита). Такой цикл развития встречается
у споровиков, которые обитают в полостных
органах, сообщающихся с внешней средой.
Такой цикл развития встречается
у споровиков, которые обитают в полостных
органах, сообщающихся с внешней средой.
В основном споровики, паразитирующие в организме человека и других позвоночных, обитают в тканях тела. Они могут поражать как человека, так и многих животных (в том числе и диких). Таким образом, это зоо- и антропозоонозные заболевания, профилактика которых представляет собой сложную задачу. Эти заболевания могут передаваться нетрансмиссивно (как токсоплаз-мы), т. е. не иметь специфического переносчика, или трансмиссивно (как малярийные плазмодии), т. е. через переносчиков.
Диагностика заболеваний, вызываемых простейшими класса Споровики, довольно сложна, так как паразиты могут обитать в различных органах и тканях (в том числе глубоких), что снижает
вероятность их обнаружения. Кроме того, выраженность симптомов заболевания невелика, поскольку они не являются строго специфичными.
Токсоплазмы
(Toxoplasma gondii) — возбудители токсоплаз-моза. Человек
для этого паразита является промежуточным
хозяином, а основные хозяева — это кошки
и другие представители семейства
Кошачьи.
Человек
для этого паразита является промежуточным
хозяином, а основные хозяева — это кошки
и другие представители семейства
Кошачьи.Малярийные плазмодии (Plasmodium) — возбудители малярии. Человек — промежуточный хозяин, окончательный — комары рода Anopheles.
4. Строение и функции спорозоитов и мерозоитов.
Сократительная вакуоль и ее фукция
Данная статья ознакомит читателя со строением простейших организмов, а именно — акцентирует внимание на строении сократительной вакуоли, выполняющей выделительную (и не только) функцию, расскажет о значении простейших и опишет способы их существования в окружающей среде.
Сократительная вакуоль. Понятие
Вакуоль (от франц. vacuole, от латинского слова vacuus – пустой), шаровидной формы небольшие полости в растительных и животных клетках или одноклеточных организмах. Сократительные вакуоли в первую очередь распространены среди простейших организмов, которые обитают в пресноводной воде, например, среди протистов, таких как амеба протей и инфузория туфелька, которая получила такое оригинальное название из-за формы тела, схожего с формой подошвы туфли. Помимо перечисленных простейших, идентичные структуры также были обнаружены и в клетках различных пресноводных губок, которые принадлежат к семейству Бадяговые.
Помимо перечисленных простейших, идентичные структуры также были обнаружены и в клетках различных пресноводных губок, которые принадлежат к семейству Бадяговые.
Вакуоль: строение и функции органеллы в клетках растений и животных
Строение сократительной вакуоли. Ее особенности
Сократительная вакуоль является мембранным органоидом, который осуществляет выброс лишней жидкости из цитоплазмы. Локализация и строение этого аппарата варьируется у различных микроорганизмов. Из комплекса пузыревидных или трубчатых вакуолей, называемых спонгиями, жидкость попадает в сократительную вакуоль. Благодаря постоянной работе этой системы поддерживается стабильный объём клетки. У простейших имеются сократительные вакуоли, которые представляют собой аппарат, регулирующий осмотическое давление, а также служащий для выделения из организма продуктов распада. Тело простейших состоит всего лишь из одной клетки, которая, в свою очередь, осуществляет все необходимые жизненные функции. Представители этого подцарства, такие как инфузория туфелька, амеба обыкновенная, другие одноклеточные обладают всеми свойствами самостоятельного организма.
Представители этого подцарства, такие как инфузория туфелька, амеба обыкновенная, другие одноклеточные обладают всеми свойствами самостоятельного организма.
Вакуоль. Функции клеточной вакуоли
Роль простейших организмов
Клетка выполняет все жизненные функции: выделение, дыхание, раздражимость, движение, размножение, обмен веществ. Простейшие распространены повсеместно. Наибольшее количество видов обитает в морских и пресных водах, многие населяют влажную почву, могут поражать растения, жить в телах многоклеточных животных и человека. В природе простейшие выполняют санитарную роль, также они участвуют в круговороте веществ, являются пищей для многих животных.
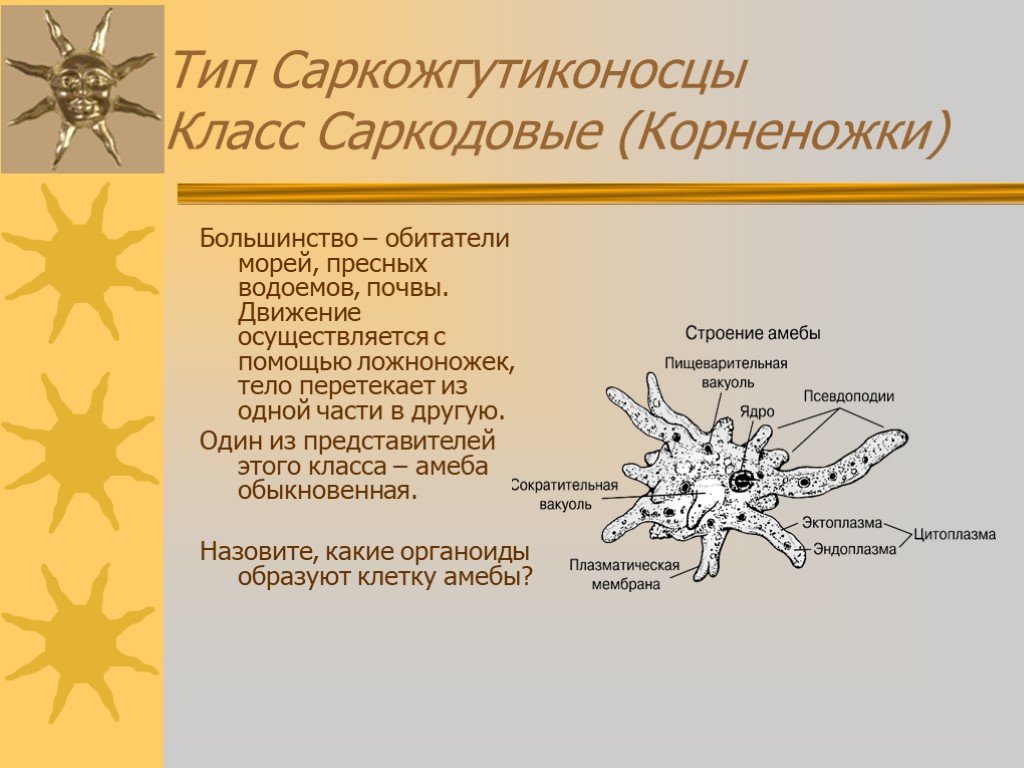
Сократительная вакуоль у амебы обыкновенной
Амеба обыкновенная – представитель класса корненожки, не имеет в отличие от других представителей постоянной формы тела. Передвижение осуществляет с помощью ложноножек. Теперь разберемся с тем, какую функцию выполняет сократительная вакуоль у амебы.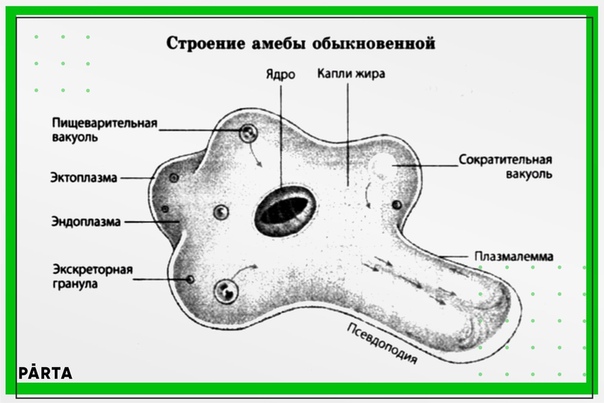 Это регуляция уровня осмотического давления внутри ее клетки. Она у амебы протей может образоваться в любом участке клетки. Через наружную мембрану вода из окружающей среды поступает внутрь осмотически. Концентрация растворенных веществ в клетке амебы выше, чем в окружающей среде. Таким образом, создается разность давления внутри клетки простейшего и за ее пределами. Функции сократительной вакуоли у амебы – это своеобразный откачивающий аппарат, который выводит избыток воды из клетки простейшего организма. Выбрасывать в окружающую среду накопившуюся жидкость амеба протей может в любом участке поверхности тела.
Это регуляция уровня осмотического давления внутри ее клетки. Она у амебы протей может образоваться в любом участке клетки. Через наружную мембрану вода из окружающей среды поступает внутрь осмотически. Концентрация растворенных веществ в клетке амебы выше, чем в окружающей среде. Таким образом, создается разность давления внутри клетки простейшего и за ее пределами. Функции сократительной вакуоли у амебы – это своеобразный откачивающий аппарат, который выводит избыток воды из клетки простейшего организма. Выбрасывать в окружающую среду накопившуюся жидкость амеба протей может в любом участке поверхности тела.
Амеба протей: класс, среда обитания, фото. Узнаем как передвигается…
Такая функция сократительной вакуоли приемлема для простейших организмов, обитающих в пресноводной воде. У паразитических и морских форм, которые обитают в среде, где осмотическое давление более высокое, чем в пресной воде, эти примитивные аппараты сокращаются очень редко или обычно отсутствуют. Вокруг сократительной вакуоли у наиболее простейших организмов концентрируются митохондрии, доставляющие энергию для выполнения осмотической работы.
Вокруг сократительной вакуоли у наиболее простейших организмов концентрируются митохондрии, доставляющие энергию для выполнения осмотической работы.
Помимо осморегуляторной, выполняет функцию дыхания в жизнедеятельности, так как в результате осмоса поступающая вода доставляет растворенный в ней кислород. Какую же еще функцию выполняет сократительная вакуоль? Так же выполняет выделительную функцию, а именно вместе с водой выводятся продукты обмена веществ в окружающую их среду.
Дыхание, выделение, осморегуляция у инфузории туфельки
Тело простейших покрыто плотной оболочкой, которое имеет постоянную форму. Питается как бактериями, так и водорослями, в том числе и некоторыми простейшими. Организм инфузории имеет более сложное, чем у амебы строение. В клетке туфельки спереди и сзади расположены две сократительные вакуоли. В этом аппарате различимы резервуар и несколько небольших канальцев. Сократительные вакуоли постоянно находятся, благодаря такому строению (из микротрубочек), на постоянном месте в клетке.
Главная функция сократительной вакуоли в жизнедеятельности данного представителя простейших — осморегуляция, также она выводит из клетки лишнюю воду, которое проникает в клетку за счет осмоса. Сначала происходит набухание приводящих каналов, потом вода из них перекачивается в специальный резервуар. Резервуар сокращается, отделяется от приводящих каналов, через поры вода выбрасывается наружу. В клетке инфузории находится две сократительные вакуоли, которые, в свою очередь, действуют в противофазе. За счет работы двух таких аппаратов обеспечивается непрерывный процесс. Помимо этого, вода непрерывно циркулирует благодаря деятельности сократительных вакуолей. Они сжимаются поочередно, и частота сокращений зависит от температуры окружающей среды.
Так, при комнатной температуре (+18 — +20 градусов по Цельсию) частота сокращений вакуолей составляет, по некоторым данным, 10-15 секунд. А учитывая, что естественной средой обитания туфельки являются любые пресные водоемы со стоячей водой и наличием в ней разлагающихся органических веществ, температура этой среды на несколько градусов меняется в зависимости от времени года и, следовательно, частота сокращений может достигать 20-25 секунд.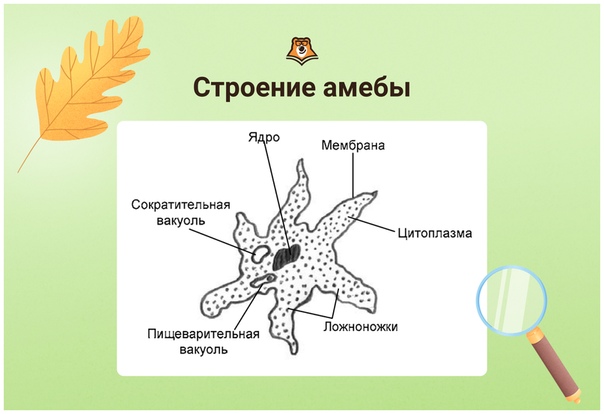 За час сократительная вакуоль простейшего организма способна выбросить из клетки воду в количестве. соизмеримом с ее размерами. В них скапливаются питательные вещества, непереваренные остатки пищи, конечные продукты обмена веществ, также можно обнаружить кислород и азот.
За час сократительная вакуоль простейшего организма способна выбросить из клетки воду в количестве. соизмеримом с ее размерами. В них скапливаются питательные вещества, непереваренные остатки пищи, конечные продукты обмена веществ, также можно обнаружить кислород и азот.
Очистка сточных вод простейшими
Влияние простейших на круговорот веществ в природе имеет огромное значение. В водоемах, вследствие спуска сточных вод, размножаются в большом количестве бактерии. В результате появляются различные простейшие организмы, которые и используют в пищу эти бактерии и таким образом способствуют естественной очистке водоемов.
Заключение
Несмотря на простое строение этих одноклеточных организмов, тело которых состоит из одной клетки, но выполняет функции целого организма, удивительным образом приспособленного к окружающей среде. Это можно наблюдать даже на примере строения сократительной вакуоли. На сегодняшний день уже доказано огромное значение простейших в природе и участие их в круговороте веществ.
Малые ядерные РНК: ассоциация с конденсированным хроматином у амеб | Journal of Cell Science
Пропустить пункт назначенияИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ| 01 апреля 1978 г.
Лестер Гольдштейн, Гэри Э. Уайз, Кристофер Стивенсон, Кристин КоИнформация об авторе и статье
Полученный: 02 августа 1977 г.
Онлайновый ISSN: 1477-9137
Печатный ISSN: 0021-9533
(1978) 30 (1): 227–235.
https://doi.org/10.1242/jcs.30.1.227
История статьиПолучено:
02 августа 1977 г.
- Разделенный экран
- Взгляды
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- Открой PDF для в другом окне
- Значок версии статьи Версии
- Версия записи
01 апреля 1978 г.

- Версия записи
01 апреля 1978 г.
- Делиться
- Фейсбук
- Твиттер
- Электронная почта
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
Citation
Лестер Гольдштейн, Гэри Э. Уайз, Кристофер Стефенсон, Кристин Ко; Малые ядерные РНК: связь с конденсированным хроматином у амеб. J Cell Sci 1 апреля 1978 г.; 30 (1): 227–235. doi: https://doi.org/10.1242/jcs.30.1.227
J Cell Sci 1 апреля 1978 г.; 30 (1): 227–235. doi: https://doi.org/10.1242/jcs.30.1.227
Скачать файл цитаты:
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Держатели для книг
- Менделей
- Бумаги
- Конечная примечание
- РефВоркс
- Бибтекс
Расширенный поиск
С целью изучения связи малых ядерных РНК (snRNAs) с хроматином мы исследовали влияние хроматинсвязывающего агента актиномицина D (AMD) на ядра амеб. С помощью электронной микроскопии мы отметили, что при помещении митотических клеток в АМД их хроматин через 3 ч сильно конденсировался. Это резко контрастирует с ситуацией в необработанных клетках, в которых хроматин в то же время в значительной степени деконденсирован. После организации ядерной трансплантацией меченых ядер в немеченую цитоплазму, чтобы единственными мечеными молекулами в клетке было J H-snRNAs, мы отметили, что в таких клетках, аналогичным образом обработанных AMD, меченый материал был сгруппирован в форме, очень похожей на форму хроматина в 3-часовом постделении клеток, обработанных AMD. В необработанных клетках 3 H-мяРНК были распределены практически равномерно по всему ядру.
В необработанных клетках 3 H-мяРНК были распределены практически равномерно по всему ядру.
Эти наблюдения вместе с более ранними данными о том, что snRNAs связываются с хромосомами во время митоза, приводят нас к заключению, что snRNAs обладают особым сродством к конденсированному хроматину.
Малые ядерные РНК (snRNAs) представляют собой группу примерно от 5 до 10 электрофоретически гомогенных видов молекул, число которых зависит от организма, которые имеют размер от примерно 65 до примерно 300 нуклеотидов и локализованы почти полностью в ядрах (Huang , 1977; Ро-Чой и Буш, 1974; Гольдштейн и Ко, 1974). Несмотря на более чем десять лет исследований в многочисленных лабораториях, в результате которых были обнаружены многие особенности этих молекул, функции мяРНК практически неизвестны. Преимущественно ядерная локализация, очевидно, указывает на то, что по крайней мере некоторые из них могут быть связаны с хроматином, но до недавнего времени было мало конкретных доказательств на этот счет.
Поскольку недавние исследования в этой лаборатории (Goldstein, 1976; Goldstein, Wise & Ko, 1977) показали, что мяРНК действительно заметно ассоциируются с хромосомами во время определенных митотических стадий, мы решили исследовать влияние на распределение мяРНК в результате воздействия клеток на этих стадиях к ингибитору транскрипции актиномицину D (AMD). Поскольку AMD прочно связывается с ДНК, мы пришли к выводу, что влияние на распределение snRNA может выявить некоторую связь между последними молекулами и хромосомами. Мы ожидали, что AMD может мешать ассоциации между snRNAs и хроматином, но на самом деле мы обнаружили, что AMD имеет по существу противоположный эффект.
Клетки и методы культивирования
Используемый в данном исследовании организм, амебу, культивировали, как описано ранее (Goldstein et al. . 1977).
Мечение клеток
Клетки метили в основном так, как описано ранее (Goldstein et al. . 1977).
. 1977).
Световая микроскопия и авторадиография
Клетки фиксировали, помещая по отдельности крошечными каплями на предметное стекло микроскопа и после удаления как можно большего количества среды придавливали покровным стеклом, содержащим каплю 45% уксусной кислоты на предметном стекле. нижняя сторона. Покровное стекло удаляли лезвием бритвы после того, как предметное стекло было заморожено на твердом CO 9 .0162 2 , а предметное стекло затем пропускали через ряд этанолов для обезвоживания. Для авторадиографии предметные стекла погружали в жидкую эмульсию Kodak NTB-3, разбавленную равной частью H 2 O, и после соответствующей экспозиции предметное стекло проявляли стандартными процедурами. Для световой микроскопии препараты окрашивали красителем Гимза при рН 7,0.
Электронная микроскопия
Электронная микроскопия проводилась в основном так, как описано ранее (Goldstein и др.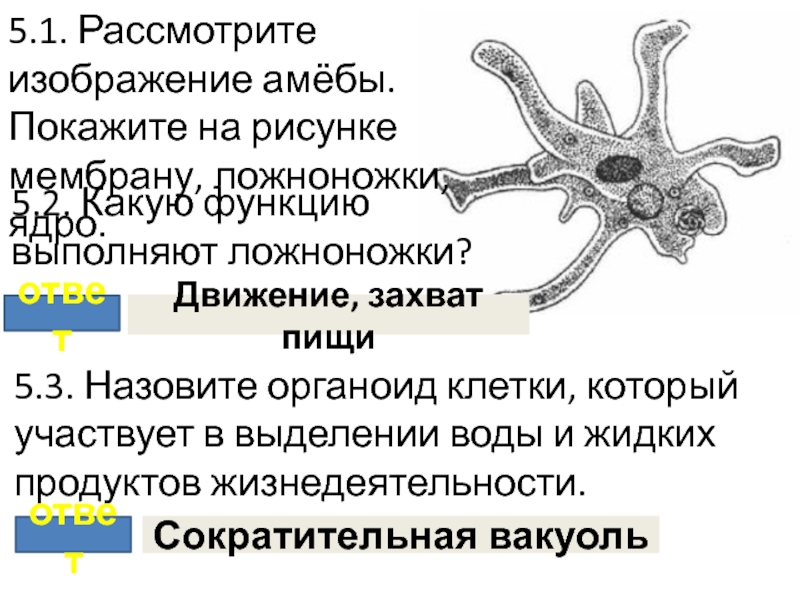 . 1977).
. 1977).
Микрохирургия
Ядра пересаживали между амебами по методу Jeon & Lorch (Jeon, 1970). Цитоплазму клеток ампутировали от руки под препаровальным микроскопом при увеличении примерно в 25 раз, просто взяв микроинструмент с острым краем (например, край отверстия тормозной пипетки) и надавив на амебу, которую нужно разрезать. Это фактически сжимает ячейку надвое, при этом разъединенные поверхности обеих частей очень хорошо герметизируются.
Отбор клеток при делении
Делящиеся амебы, распознаваемые по критериям, описанным Chalkley & Daniel (1933), собирали по отдельности тормозной пипеткой.
Эффекты AMD на структуру хроматина
Хотя авторадиографические наблюдения, представленные ниже, подтолкнули к электронно-микроскопическому исследованию, мы представляем последнее в первую очередь.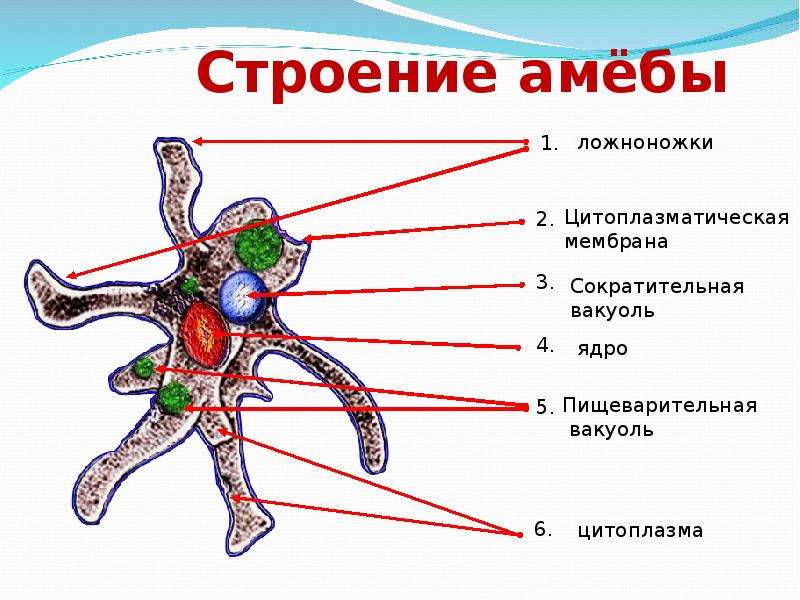 Амебы на ранних стадиях митоза помещали в среду, содержащую 500 мкг/мл мкг/мл АМД, концентрацию препарата, не оказывающую явного влияния на успешное завершение деления и последующее прохождение хотя бы через ранние стадии интерфазы. (500 9Использовали 0028 мк г/мл AMD, поскольку в то время сообщалось (Rao & Prescott, 1970), что это концентрация, необходимая для ингибирования 99% или более транскрипции у амеб, но впоследствии мы определили (неопубликованные данные), что 100 мк мкг/мл почти так же эффективны.) Периодически после введения АМД образцы постмитотических клеток фиксировали и готовили для электронной микроскопии. Описания образцов, зафиксированных только в 2 временных точках, достаточно, чтобы проиллюстрировать эффект AMD.
Амебы на ранних стадиях митоза помещали в среду, содержащую 500 мкг/мл мкг/мл АМД, концентрацию препарата, не оказывающую явного влияния на успешное завершение деления и последующее прохождение хотя бы через ранние стадии интерфазы. (500 9Использовали 0028 мк г/мл AMD, поскольку в то время сообщалось (Rao & Prescott, 1970), что это концентрация, необходимая для ингибирования 99% или более транскрипции у амеб, но впоследствии мы определили (неопубликованные данные), что 100 мк мкг/мл почти так же эффективны.) Периодически после введения АМД образцы постмитотических клеток фиксировали и готовили для электронной микроскопии. Описания образцов, зафиксированных только в 2 временных точках, достаточно, чтобы проиллюстрировать эффект AMD.
Сразу после завершения цитокинеза (в это время телофаза также может считаться завершенной) хроматин в необработанных клетках в значительной степени деконденсирован (рис. 1). В клетках, помещенных в АМД примерно за 0–5 ч до завершения цитокинеза, хроматин несколько более конденсирован (рис. 2), но разница между двумя типами ядер невелика. В необработанных клетках, зафиксированных через 3 ч после деления, хроматин настолько размыт, что едва различим (рис. 3), но в клетках, хранившихся в AMD до 3 ч после митоза, появление хроматина бросается в глаза (рис. 4). Хроматин в это время настолько конденсирован, что производит поверхностное впечатление метафазной пластинки. Ячейка явно 9Однако 0028, а не в метафазе, поскольку наличие «сотовой» ядерной оболочки показывает, что ядро находится в интерфазе. (Ядрышки выглядят как необычно большие темные тела, их относительно немного, но они не будут обсуждаться в этом отчете.)
2), но разница между двумя типами ядер невелика. В необработанных клетках, зафиксированных через 3 ч после деления, хроматин настолько размыт, что едва различим (рис. 3), но в клетках, хранившихся в AMD до 3 ч после митоза, появление хроматина бросается в глаза (рис. 4). Хроматин в это время настолько конденсирован, что производит поверхностное впечатление метафазной пластинки. Ячейка явно 9Однако 0028, а не в метафазе, поскольку наличие «сотовой» ядерной оболочки показывает, что ядро находится в интерфазе. (Ядрышки выглядят как необычно большие темные тела, их относительно немного, но они не будут обсуждаться в этом отчете.)
Рис. 1. ядра с деконденсированным хроматином. На периферии ядра несколько ядрышек, × 7200.
Рис. 1.
Просмотреть в большом размереСкачать слайд
Необработанная клетка, зафиксированная в конце телофазы, показывающая ядро с деконденсированным хроматином. Несколько ядрышек находятся на периферии ядра, × 7200.
Рис. 2.
2.
Увеличить Загрузить слайд
Клетка, обработанная AMD, зафиксирована в конце телофазы, показывающая ядро со слегка конденсированным хроматином, проходящее через центр ядро. Клетка находилась в AMD около 0,5 ч. × 6800.
Рис. 2.
Просмотреть большой Загрузить слайд
Клетка, обработанная AMD, зафиксирована в конце телофазы, показывающая ядро со слегка конденсированным хроматином, проходящим через центр ядра. Клетка находилась в AMD около 0,5 ч. × 6800.
Рис. 3.
Увеличить Загрузить слайд
Необработанная клетка зафиксирована через 3 ч после деления. Хроматин деконденсирован, но все еще находится в центральной области ядра. Ядрышки крупнее, чем в клетках, только что завершивших митоз, и слой сот, лежащий под ядерной оболочкой, теперь реформирован, × 6800.
Рис. 3.
Увеличить Загрузить слайд
Необработанная клетка, зафиксированная через 3 ч после деления. Хроматин деконденсирован, но все еще находится в центральной области ядра. Ядрышки крупнее, чем в клетках, только что завершивших митоз, и слой сот, лежащий под ядерной оболочкой, теперь реформирован, × 6800.
Ядрышки крупнее, чем в клетках, только что завершивших митоз, и слой сот, лежащий под ядерной оболочкой, теперь реформирован, × 6800.
Рис. через 3 ч после митоза. Хроматин (с) чрезвычайно конденсирован и расположен в центре ядра. Ядрышки (n) характеризуются небольшими темными тельцами по периферии. Сотовый слой присутствует, × 12000.
Рис. 4.
Увеличить Загрузить слайд
Клетки инкубировали в AMD до 3 ч после митоза. Хроматин (с) чрезвычайно конденсирован и расположен в центре ядра. Ядрышки (n) характеризуются небольшими темными тельцами по периферии. Присутствует сотовый слой, × 12000.
Некоторая конденсация хроматина наблюдается в клетках, подвергшихся воздействию ВМД в течение 3 ч, только в интерфазе (рис. 5), но количество конденсации заметно меньше, чем в клетках, подвергшихся воздействию ВМД. началось во время митоза. Фликингер (1968) ранее отмечал аналогичные эффекты ВМД на интерфазные ядра амеб.
Рис. 5.
УвеличитьСкачать слайд
Интерфазные клетки инкубировали в АМД в течение 3 ч.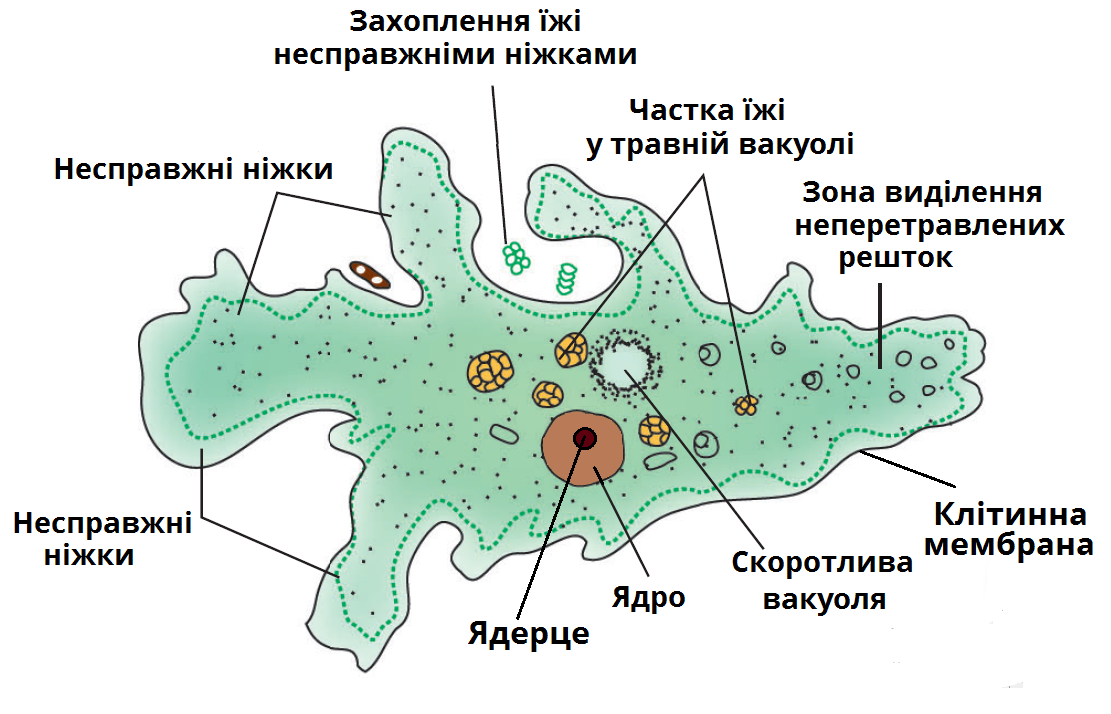 Хотя это ядро представляет собой максимальную конденсацию хроматина (с), когда-либо наблюдаемую в клетках, обработанных таким образом, хроматин все еще не является таким конденсированным, как наблюдаемый в клетках после деления, поддерживаемых при AMD (Fig. 4). × 12000
Хотя это ядро представляет собой максимальную конденсацию хроматина (с), когда-либо наблюдаемую в клетках, обработанных таким образом, хроматин все еще не является таким конденсированным, как наблюдаемый в клетках после деления, поддерживаемых при AMD (Fig. 4). × 12000
Рис. 5 Хотя это ядро представляет собой максимальную конденсацию хроматина (с), когда-либо наблюдаемую в клетках, обработанных таким образом, хроматин все еще не является таким конденсированным, как наблюдаемый в клетках после деления, поддерживаемых при AMD (Fig. 4). × 12000,
Влияние AMD на распределение snRNAs
Амебы метили [ 3 H]уридином в течение 2 дней и отгоняли с немеченым кормом в течение 3 дней. В течение оставшейся части эксперимента клетки питались только немеченой пищей. Для того чтобы мяРНК были практически единственными мечеными молекулами в клетке, ядра таких клеток трансплантировали в энуклеированные немеченые клетки и через день в течение недели после этого удаляли половину цитоплазмы клеток-реципиентов путем ампутации. Такая ампутация во время роста клеток на немеченой пище служила для предотвращения их деления и удаления значительного количества любых радиоактивных молекул, попавших в цитоплазму; то есть рибосомные, транспортные и матричные РНК, чьи меченые ядерные предшественники могли все еще присутствовать в трансплантированных ядрах. В конце последовательности ампутации ядра снова переносили в энуклеированные немеченые клетки. Эти реципиентные клетки не подвергались дальнейшему хирургическому лечению, но им давали возможность беспрепятственно расти и вступать в деление. Когда такая клетка вступала в митоз, она помещалась в 500 мк г/мл АМД или в обычную среду для амеб. Образцы таких клеток одновременно фиксировали и раздавливали в разное время постмитоза и подвергали авторадиографии. Как отмечалось в предыдущих экспериментах (Goldstein et al. . 1977), в клетках, выдержанных в обычной амебной среде, метка оказалась более или менее равномерно распределенной по ядру (рис. 6), но большая часть 3 H -snRNA клеток, помещенных в AMD, оказалась агрегированной (рис.
Такая ампутация во время роста клеток на немеченой пище служила для предотвращения их деления и удаления значительного количества любых радиоактивных молекул, попавших в цитоплазму; то есть рибосомные, транспортные и матричные РНК, чьи меченые ядерные предшественники могли все еще присутствовать в трансплантированных ядрах. В конце последовательности ампутации ядра снова переносили в энуклеированные немеченые клетки. Эти реципиентные клетки не подвергались дальнейшему хирургическому лечению, но им давали возможность беспрепятственно расти и вступать в деление. Когда такая клетка вступала в митоз, она помещалась в 500 мк г/мл АМД или в обычную среду для амеб. Образцы таких клеток одновременно фиксировали и раздавливали в разное время постмитоза и подвергали авторадиографии. Как отмечалось в предыдущих экспериментах (Goldstein et al. . 1977), в клетках, выдержанных в обычной амебной среде, метка оказалась более или менее равномерно распределенной по ядру (рис. 6), но большая часть 3 H -snRNA клеток, помещенных в AMD, оказалась агрегированной (рис.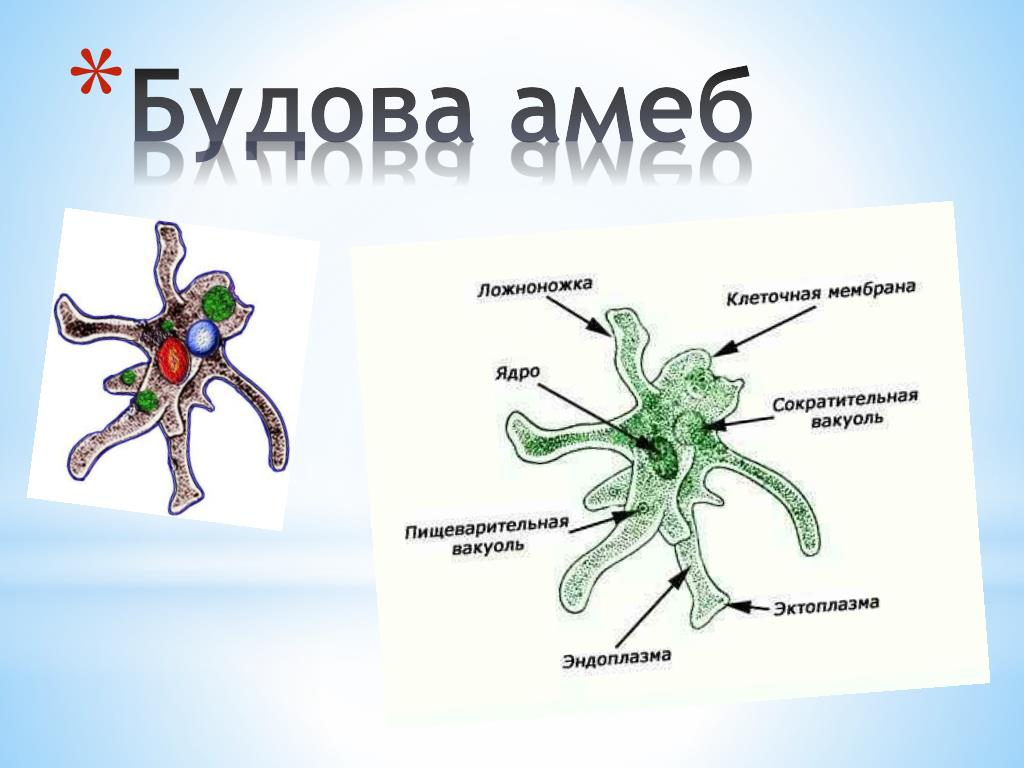 7) -подобной форме, в которой агрегируется хроматин (предыдущий раздел) при сходных обстоятельствах.
7) -подобной форме, в которой агрегируется хроматин (предыдущий раздел) при сходных обстоятельствах.
Рис. 6.
Увеличить Загрузить слайд
Светомикроскопическая авторадиограмма необработанной интерфазной клетки, показывающая более или менее равномерное распределение по ядру 3 H-мяРНК. × 825.
Рис. 6.
Посмотреть большойСкачать слайд
Светомикроскопическая авторадиограмма необработанной интерфазной клетки, показывающая более или менее равномерное распределение по ядру 3 H-мяРНК. × 825.
Рис. 7.
Посмотреть в большом размереСкачать слайд
Светомикроскопическая авторадиограмма клеток, зафиксированных через 2,5 ч после помещения в ВМД во время митоза. Видно, что меченые мяРНК агрегированы в форме, сходной с хроматином на рис. 4. × 825.
рис. 7.
Посмотреть большойСкачать слайд
Световая микроскопия авторадиограммы фиксированной клетки через 2,5 ч после помещения при ВМД во время митоза.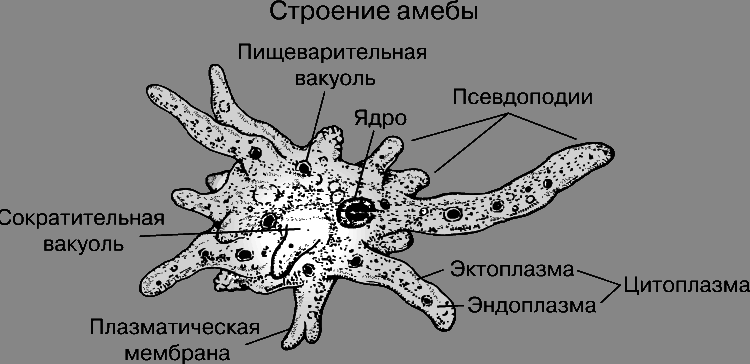 Видно, что меченые мяРНК агрегированы в форме, сходной с хроматином на рис. 4. × 825.
Видно, что меченые мяРНК агрегированы в форме, сходной с хроматином на рис. 4. × 825.
Мы пришли к выводу, что значительная часть мяРНК (возможно, все определенные виды мяРНК и ни одна другая) ассоциирована с конденсированным ранним интерфазным хроматином. Мы не пытались провести прямую корреляцию между агрегацией хроматина и радиоактивностью в одних и тех же клетках, потому что у нас было слишком мало экспериментальных клеток, чтобы разумное число могло вступить в деление за тот же период времени, и, следовательно, слишком мало, чтобы получить достаточно существенный осадок. обрабатывать и делать срезы для электронно-микроскопической авторадиографии без особого труда.
Представленные здесь данные демонстрируют, что AMD вызывает поразительную конденсацию хроматина, особенно сразу после митоза, и вызывает агрегацию snRNAs способом, параллельным агрегации хроматина. Хотя два типа агрегации не были обнаружены в одних и тех же индивидуальных клетках, сходство в ответе на AMD приводит нас к заключению, что мяРНК имеют большое сродство к конденсированному хроматину, как и мяРНК к профазным и анафазным хромосомам (Goldstein, 1976; Goldstein). и др. . 1977). Хотя теоретически мы могли бы проводить авторадиографические и ЭМ-наблюдения за одними и теми же клетками, объем работы, необходимый для обеспечения того, чтобы у нас было достаточно обработанных и необработанных клеток для каждого момента времени, был бы огромным и, казалось бы, ненужным в свете ясности. наблюдений, о которых мы сообщаем.
и др. . 1977). Хотя теоретически мы могли бы проводить авторадиографические и ЭМ-наблюдения за одними и теми же клетками, объем работы, необходимый для обеспечения того, чтобы у нас было достаточно обработанных и необработанных клеток для каждого момента времени, был бы огромным и, казалось бы, ненужным в свете ясности. наблюдений, о которых мы сообщаем.
Ранее сообщалось о влиянии AMD на структуру хромосом. Затяжка политенных хромосом регрессирует в полосы из-за ВМД (см. Beermann, 19).71), а хромосомные петли ламповых щеток под действием препарата регрессируют в хромомеры (Izawa, Allfrey & Mirsky, 1963). Таким образом, индукция конденсации хроматина, по-видимому, является широко распространенным ответом на AMD, и, конечно же, дальнейшим ответом является ингибирование транскрипции. Beermann (1971) заключает, что транскрипционная активность является предпосылкой для деконденсации хромосом и что, когда синтез РНК блокируется любым из множества механизмов, разворачивающийся хроматин быстро конденсируется. Причинно-следственная связь между конденсацией хромосом и накоплением мяРНК теперь также представляет интерес, и в настоящее время мы занимаемся определением того, можно ли обнаружить какие-либо различия в содержании мяРНК во фракционированном гетерохроматине и эухроматине клеток млекопитающих, в надежде выяснить, что проблема.
Причинно-следственная связь между конденсацией хромосом и накоплением мяРНК теперь также представляет интерес, и в настоящее время мы занимаемся определением того, можно ли обнаружить какие-либо различия в содержании мяРНК во фракционированном гетерохроматине и эухроматине клеток млекопитающих, в надежде выяснить, что проблема.
В настоящее время наша любимая модель (Goldstein, 1976) состоит в том, что конденсация хроматина предшествует накоплению snRNA (хотя у нас нет лучших доказательств этой последовательности, чем обратное). Мы предполагаем, что конденсация вызывает отщепление определенных кислых белков от хромосом и что в результате snRNAs могут найти сайты связывания на хромосомах. Когда хромосомы деконденсируются, кислые белки (те, которые ранее были связаны с хромосомами, или другие) повторно связываются с хромосомами и тем самым вытесняют snRNAs. Мы предполагаем, что эта ассоциация и смещение мяРНК могут быть способом, которым экспрессия хромосомной информации «запрограммирована».
Мы предполагали, что связывание snRNAs с хромосомами может быть достигнуто посредством некоторого механизма спаривания оснований с определенными последовательностями ДНК. Тот факт, что AMD, имеющая сильное сродство к ДНК, не мешает этому связыванию, предполагает, что спаривание оснований может быть не задействовано. Тем не менее, эти наблюдения предполагают, что взаимодействия различных молекул должны быть увлекательным предметом изучения.
Интересна разница в паттернах конденсации хромосом, индуцированной AMD, в ранних клетках после деления и в более поздних интерфазных клетках; ранняя конденсация после деления принимает высокоупорядоченный паттерн, напоминающий метафазную пластинку, тогда как более поздний паттерн гораздо менее регулярен, хотя и не совсем неупорядочен. Так как ранние агрегации после деления явно не являются случайными скоплениями, должна существовать лежащая в основе структурная сеть, которая указывает на остатки некоторой митотической организации. Значение этого неочевидно, но в конечном счете может иметь значение для понимания митотического аппарата.
Это исследование было поддержано грантом NIH GM-15156.
Beermann
,W.
(1971
).Влияние а-аманитина на пыхтение и внутриядерный синтез в слюнных железах Chironomus
.Хромосома
34
,152
–167
.https://doi.org/10.1007/BF00285183
Chalkley
,H.W.
иDaniel
,G. E.
( 90 004 1933 ).Связь между формой живой клетки и ядерными фазами деления у Amoeba proteus (Leidy)
.Физиол. Зоол
.6
,592
–619
.https://doi.org/10.1086/physzool.6.4.30151207
Flickinger
,CJ
(1968
).Тонкая структура ядрышек нормальных и обработанных актиномицином D Amoeba proteus
. Дж. Ультраструктур. Рез
Рез
23
,260
–271
.https://doi.org/10.1016/S0022-5320(68)80005-X
Гольдштейн
,Л.
(1976
).Роль малых ядерных РНК в «программировании» хромосомной информации ?
Природа, Лонд
.261
,519
–521
.https://doi.org/10.1038/261519a0
Goldstein
,L.
иKo
,C.
(1974
).Электрофоретическая характеристика челночных и нечелночных малых ядерных РНК
.Сотовый
2
,259
–269
.https://doi.org/10.1016/0092-8674(74)
-8Гольдштейн
,Л.
,Мудрый
, 90 003Г. Е.
иКо
,К.
(1977
).Локализация малых ядерных РНК во время митоза: исследование под электронным микроскопом
.
Дж. Селл Биол
.73
,322
–331
.https://doi.org/10.1083/jcb.73.2.322
Huang
,RCC
(1977
).Низкомолекулярная ядерная РНК: размер, структура и возможная функция
.In Хроматин и структура хромосом
(ред.H. J.
Li
иR. A.
Eck hardt
), стр.299313
.Нью-Йорк и Лондон
:Academic Press
.Изава
,М.
,Allfrey
,В.Г.
(1963
).Связь между синтезом РНК и структурой петли в хромосомах ламповых щеток
.Проц. физ. акад. науч. США
.49
,544
–551
.https://doi.org/10.1073/pnas.49.4.544
Чон
,К.В.
(1970
).
Микроманипуляции с ядрами амеб
.Мет. Селл Физиол
.4
,179
–194
.Rao
,M.V.N.
иPrescott
,D.M. 03
1970
).Включение ядерной РНК, меченной до деления, в ядра после деления у Amoeba proteus
.Expl Cell Res
.62
,286
–292
.https://doi.org/10.1016/0014-4827(70)
-2Ro-Choi
,T. S.
иBusch
,Х.
(1974
).Низкомолекулярные ядерные РНК
.In Ядро клетки
, vol.3
(изд.H.
Busch
), стр.151
–208 90 003 .
Нью-Йорк и Лондон
:Academic Press
.3.2 Сравнение прокариотических и эукариотических клеток – Концепции биологии – 1-е канадское издание
Перейти к содержимому Глава 3. Введение в структуру и функции клеток
Введение в структуру и функции клеток
К концу этого раздела вы сможете:
- Называть примеры прокариотических и эукариотических организмов
- Сравнение и противопоставление прокариотических и эукариотических клеток
- Опишите относительные размеры различных типов клеток
Клетки относятся к одной из двух широких категорий: прокариотические и эукариотические. Преимущественно одноклеточные организмы доменов Bacteria и Archaea классифицируются как прокариоты ( про — = раньше; – карион – = ядро). Клетки животных, клетки растений, грибы и протисты являются эукариотами ( eu — = правда).
Все клетки имеют четыре общих компонента: 1) плазматическая мембрана, внешнее покрытие, отделяющее внутреннюю часть клетки от окружающей среды; 2) цитоплазма, состоящая из желеобразной области внутри клетки, в которой находятся другие клеточные компоненты; 3) ДНК, генетический материал клетки; и 4) рибосомы, частицы, синтезирующие белки. Однако прокариоты отличаются от эукариотических клеток по нескольким параметрам.
Однако прокариоты отличаются от эукариотических клеток по нескольким параметрам.
Прокариотическая клетка представляет собой простой одноклеточный (одноклеточный) организм, лишенный ядра или любой другой связанной с мембраной органеллы . Вскоре мы увидим, что у эукариот это существенно отличается. Прокариотическая ДНК находится в центральной части клетки: затемненная область, называемая нуклеоидом.
Рисунок 3.6. На этом рисунке показана обобщенная структура прокариотической клетки. В отличие от архей и эукариот клеточная стенка бактерий состоит из пептидогликана, состоящего из сахаров и аминокислот, и многие из них имеют полисахаридную капсулу (рис. 3.6). Клеточная стенка действует как дополнительный слой защиты, помогает клетке сохранять свою форму и предотвращает обезвоживание. Капсула позволяет клетке прикрепляться к поверхностям в окружающей среде. Некоторые прокариоты имеют жгутики, пили или фимбрии. Жгутики используются для передвижения, в то время как большинство пили используются для обмена генетическим материалом во время типа размножения, называемого конъюгацией.
В природе взаимосвязь между формой и функцией очевидна на всех уровнях, включая уровень клетки, и это станет ясно, когда мы будем исследовать эукариотические клетки. Принцип «форма следует за функцией» встречается во многих контекстах. Например, птицы и рыбы имеют обтекаемые тела, которые позволяют им быстро перемещаться в среде, в которой они живут, будь то воздух или вода. Это означает, что в целом можно вывести функцию структуры, глядя на ее форму, потому что они совпадают.
Эукариотическая клетка — это клетка, имеющая мембраносвязанное ядро и другие мембраносвязанные компартменты или мешочки, называемые органеллами , которые выполняют специализированные функции. Слово «эукариотический» означает «истинное ядро» или «истинное ядро», намекая на наличие в этих клетках мембраносвязанного ядра. Слово «органелла» означает «маленький орган», и, как уже упоминалось, органеллы имеют специализированные клеточные функции, точно так же, как и органы вашего тела имеют специализированные функции.
При диаметре 0,1–5,0 мкм прокариотические клетки значительно меньше эукариотических клеток, диаметр которых колеблется в пределах 10–100 мкм (рис. 3.7). Небольшие размеры прокариот позволяют ионам и органическим молекулам, попавшим в них, быстро распространяться в другие части клетки. Точно так же любые отходы, образующиеся в прокариотической клетке, могут быстро выводиться наружу. Однако более крупные эукариотические клетки развили различные структурные приспособления для усиления клеточного транспорта. Действительно, большие размеры этих клеток были бы невозможны без этих приспособлений. В общем 9Размер клетки 0840 ограничен , потому что объем увеличивается гораздо быстрее, чем площадь поверхности клетки. По мере того, как клетка становится больше, ей становится все труднее и труднее получать достаточное количество материалов для поддержания процессов внутри клетки, потому что относительный размер площади поверхности, по которой должны транспортироваться материалы, уменьшается.
Прокариоты — преимущественно одноклеточные организмы доменов Bacteria и Archaea. Все прокариоты имеют плазматические мембраны, цитоплазму, рибосомы, клеточную стенку, ДНК и лишены мембраносвязанных органелл. Многие также имеют полисахаридные капсулы. Прокариотические клетки имеют диаметр от 0,1 до 5,0 мкм.
Как и прокариотическая клетка, эукариотическая клетка имеет плазматическую мембрану, цитоплазму и рибосомы, но эукариотическая клетка, как правило, больше, чем прокариотическая клетка, имеет истинное ядро (это означает, что ее ДНК окружена мембраной) и имеет другую мембрану. -связанные органеллы, обеспечивающие компартментализацию функций. Эукариотические клетки, как правило, в 10-100 раз больше прокариотических клеток.
эукариотическая клетка: клетка, имеющая мембраносвязанное ядро и несколько других мембраносвязанных компартментов или мешочков
органелла: мембраносвязанный компартмент или мешок внутри клетки у которого отсутствует ядро или любая другая связанная с мембраной органелла
License
Concepts of Biology — 1st Canadian Edition Чарльза Молнара и Джейн Гейр распространяется под лицензией Creative Commons Attribution 4.