⇐ Предыдущая3456789101112Следующая ⇒ 6. Каково происхождение вольвокофициевых и каковы направления их эволюции? Класс Протококкофициевые ( Protococcophyceae), или Хлорококкофициевые ( Chlorococcophyceae) Объединяет коккоидные формы, т.е. одноклеточные, цено- биальные, колониальные, изредка многоклеточные организмы, неподвижные в вегетативном состоянии. Они имеют микроскопические размеры, и лишь водяная сеточка достигает 1 м. У протококкофициевых преобладает бесполое размножение при помощи зоо-, гемизоо- и автоспор. Гемизооспоры, или амебоиды, — самые примитивные специализированные клетки бесполого размножения. Они не имеют жгутиков и оболочек, способны передвигаться амебоидно. Половой процесс наблюдается редко. Обычно это изогамия, реже гетеро- и оогамия. Покоящимися стадиями служат акинеты и цисты. В теоретическом отношении протококкофициевые интересны тем, что в процессе эволюции зеленых водорослей на этом этапе впервые возникла и утвердилась коккоидная, типично растительная структура тела с целлюлозными оболочками, от которых развились новые ступени морфологической дифференциации таллома: сифональная, сифонокладальная, нитчатая, разнонитчатая и пластинчатая. Протококкофициевые встречаются во всех водоемах, в планктоне и бентосе, в наземных и почвенных биоценозах. По количеству видов они уступают лишь диатомовым. В классе выделяют 3 порядка: Вакуольные, Хлорококкаль- ные и Прототрихальные. Наиболее типичным представителем данного класса является порядок Хлорококкальные. Вакуольные (Vacuolates) являются переходным звеном между вольво- кальными и протококальными, а прототрихальные (Prototri- chales) — между протококальными и улотрихальными. Порядок Хлорококкальные ( Chlorococcules). Объединяет водоросли с наиболее выраженной коккоидной структурой, т.е. имеются плотные оболочки, а у вегетативных клеток отсутствуют пульсирующие вакуоли. Нет у них ни глазка, ни псевдоци- лий, не наблюдается и тенденции к образованию многоклеточного таллома. Преимущественно одноклеточные формы, хотя есть колониальные и ценобиальные. Размножение хлорококко- вых осуществляется зоо- и автоспорами. Половой процесс изо-, гетеро- и оогамный. Род хлорелла (Chlorella) широко распространен в пресных водоемах, на сырой земле, коре деревьев, входит в состав лишайников. Одиночные шаро- или эллипсовидные клетки одеты гладкой оболочкой, содержат пристенный, цельный, рассеченный или лопастный чашевидный хроматофор с пиреноидом и ядро (рис. 1.42). Размножается хлорелла автоспорами, которые образуются по 2 — 4, 16 — 64 в клетке. Известны покоящиеся стадии — акинеты. Половое размножение не отмечено. Практическое значение хлореллы чрезвычайно велико. Она используется для очистки сточных вод на сахарных и других заводах, для регенерации воздуха в замкнутых биологических системах (например, в космических кораблях). Белки хлореллы содержат все незаменимые аминокислоты. В Беларуси отмечено 5 видов. Широко распространены X. обыкновенная
(СЛ. vulgaris), X. почвенная (СЛ. terricola), X. эллипсовидная (СЛ. ellipsoidea). Род х. Известно более 40 видов, в том числе 4 в Беларуси. Наиболее часто встречаются X. наземный (СЛ. humicola), X. инфузи- онный (СЛ. infusionum). Род гидродикцион, или водяная сеточка (Hydrodictyon), интересен тем, что его ценобий имеет вид сетчатого мешка, размеры которого могут достигать в длину до I м (чаще 15 — 20 см), а в ширину — 10 — 15 см. В состав ценобия входит несколько тысяч клеток. Они крупные, длиной до 1 — 1, 5 см, цилиндрической формы, одеты целлюлозными оболочками.
Протопласт их содержит сетчатый хроматофор с многочисленными пиреноидами и до 20 ООО — 30 ООО ядер. Центральную часть клетки занимает крупная вакуоль. При бесполом размножении в любой из клеток ценобия образуется до 20 ООО зооспор, внутри которой они теряют жгутики и складываются в новую сеточку. Освобождение дочернего ценобия происходит после ослизнения оболочки материнской клетки. В дальнейшем ценобий растет за счет увеличения размеров клеток. Половой процесс изогамный. В клетках развиваются мелкие двухжгутиковые гаметы (до 30 ООО), которые выходят в воду и, попарно копулируя, образуют зиготу. После периода покоя зигота прорастает 2 — 4-гаплоидными зооспорами. Каждая зооспора превращается в многоугольную звездообразную пластинку — полиэдр. Он увеличивается в размерах, становится многоядерным и распадается на двухжгутиковые зооспоры, которые внутри полиэдра слагают водяную сеточку. Обитает гидродикцион в реках, прудах, ямах, богатых азотистыми соединениями. Известно 5 видов. В нашей стране встречается 1 вид — Г. сеточный (Я. reticulatum). У водорослей рода педиаструм (Pediastrum) ценобии плоские, пластинчатые, округлой или звездчатой формы, состоят из 4 — 128 клеток, которые плотно прилегают друг к другу или образуют межклетники в виде отверстий в ценобии (рис. 1.45). Клетки одноядерные (старые многоядерные), периферийные, в отличие от внутренних сильно выемчатые, с 1 — 4 лопастями и выростами. Оболочки клеток гладкие, бородавчатые, морщинистые, складчатые. Хроматофор пристенный с одним пиреноидом. Размножение бесполое — зооспорами. Они выходят из клеток в слизистом пузыре и, еще находясь в нем, складывают-
ся в дочерний ценобий. Половой процесс и развитие зиготы сходны с половым процессом водяной сеточки и включают в себя стадию полиэдра. Известны стадии покоя — акинеты. В Беларуси установлены 13 видов и множество разновидностей. Наиболее известны П. угловатый (P. angulosum), П. бориана (P. boryanum), П. двухлучевой {P. biradiatum), П. простой (P. simplex), П. удвоенный (P. duplex) Встречаются в планктоне озер, водохранилищ, рек, на влажных скалах. У видов рода сценедесмуе (Scenedesmus) (рис. 1.46) ценобии имеют вид плоских, прямых или изогнутых пластинок из 4-8 (реже 2 — 32) продолговатых или округлых клеток, соединенных боками в простые или двойные ряды. Оболочки гладкие или с бородавками, ребрами, шипами или сосочками, часто на концах клеток с длинными шипами или рогами. Хлоропласт пластинчатый, с боковым пиреноидом и центральным ядром. Размножается автоспорами, которые внутри материнской клетки слагаются в дочерний ценобий. Сценедесмус широко распространен в планктоне, часто встречается в прибрежной зоне среди нитчаток, мхов и др. Известно более 200 видов, из них 26 отмечены в нашей республике. Часто встречаются С. четырехрогий (S. Задания 1. Приготовить препараты хлореллы и хлорококка и рассмотреть при малом и большом увеличении микроскопа. Зарисовать отдельную клетку. Отметить оболочку, ядро, чашевидный хроматофор с пиреноидом, цитоплазму. 2. Рассмотреть и зарисовать форму клеток гидродикциона, характер их сочленения при образовании ячеек сеточки. Отметить оболочку клетки, сетчатый хроматофор с большим количеством пиреноидов, многоядерность, постенное расположение цитоплазмы. Найти и зарисовать часть молодой (дочерней) сеточки. 3. Рассмотреть и зарисовать общий вид ценобия педиастру- ма и сценедесмуса. Обратить внимание на форму клеток, характер сочленения их друг с другом, отсутствие в некоторых клетках педиаструма внутреннего содержимого (пустые клетки) после выхода из них зооспор. Вопросы для самоконтроля 1. Каковы особенности строения клетки протококкофициевых? 2. 3. Каково различие между зоо-, аплано-, гемизоо- и автоспорами? Какие из них являются более совершенными и почему? 4. Каковы общие черты в образовании колоний у протококкофициевых и равножгутиковых? 5. Каковы особенности цикла развития водяной сеточки? 6. Каково теоретическое и практическое значение протококкофициевых водорослей? Класс Улотрихофициевые (Ulothrichophyceue) Класс объединяет нитчатые и пластинчатые (паренхими- гозные) одноядерные водоросли. Среди нитчатых форм есть свободноживущие и прикрепленные, ветвящиеся и неветвящи- еся, разнонитчатые, живущие одиночно или скоплениями. Пластинчатые формы бывают одно-, двух- или местами многослойными, распростертыми по субстрату и прикрепленными только в одном месте. Улотрихофициевые являются продолжением эволюционной линии зеленых водорослей с более высокой ступенью организации. В классе 7 порядков, из которых большой интерес представляют Улотрихальные, Ульвальные, Хетофоральные и Эдого- ниальные. Порядок Улотрихальные ( Ulothrichales). Слоевище улотри- хальных построено по типу однорядной неразветвленной нити. Клетки таллома похожи друг на друга по строению и функции, за исключением базальной, которая образует прикрепляющийся к субстрату ризоид. Все клетки, кроме базальной, способны делиться и участвовать в росте водоросли, а также формировать репродуктивные клетки (споры и гаметы). Порядок включает более 16 родов. Виды рода улотрикс (Ulothrix) (рис. 1.47) встречаются преимущественно в пресных, реже морских и солоноватых водоемах, а также в почве. Водоросли прикрепляются к подводным предметам, формируя ярко-зеленые кустики размером до 10 см и более. Неразветвленные нити улотрикса, состоящие из одного ряда цилиндрических или бочонковидных клеток с толстыми целлюлозными оболочками, прикрепляются к субстрату конической базальной клеткой или ризоидом.
рехжгутиковые зооспоры, реже — апланоспоры. Зооспоры образуются по 2 — 16 (32) во всех клетках нити, кроме базальной. Они выходят из материнской клетки в слизистой обертке, которая вскоре расплывается. После периода движения зооспора останавливается, сбрасывая один за другим жгутики, прикрепляется боком к субстрату и прорастает в нить. Половой процесс изогамный. После слияния двухжгутико- вых гамет зигота вначале двигается, затем оседает на дно водоема, вырабатывает плотную оболочку и слизистую ножку, прикрепляющую ее к субстрату. растает непосредственно в диплоидную нить, а редукция происходит при формировании зооспор. Известно около 30 видов улотрикса, из них 6 встречаются в Беларуси. Чаше других распространены У. опоясанный (U. zonata ), У. утонченный (U. tenuissima), У. изменчивый (U. variabilis) и др. Порядок Ульвальные ( Ulvales). Включает водоросли со слоевищами паренхиматозного строения пластинчатой, трубчатой или мешковидной формы и относительно крупных размеров. Слоевища ульвальных слабо дифференцированы и состоят почти из однотипных клеток. В цикле развития большинства ульвальных осуществляется изоморфная смена генераций. Обитают ульвальные в основном в морях, реже в пресных водоемах. В порядке 4 семей-, ства, центральным и наиболее обширным из которых является семейство ульвовых. Водоросли рода ульва (Ulva) (рис. 1.48) представляют собой двухслойное слоевище в виде широкой (до 15 см) пластинки длиной до 25 см ярко-зеленой окраски. На начальных стадиях развития ульвы образуется однорядная нить, которая переходит в трубчатую стадию, затем стенки трубки смыкаются и в дальнейшем она растет как двухслойная пластина. а гаметофит — половым путем — копуляцией изогамет. Зигота прорастает в диплоидный спорофит, у которого любая клетка может функционировать как зооспорангий. Перед образованием зооспор происходит редукционное деление ядра. Из гаплоидных зооспор развиваются гаплоидные гаметофиты, которые по внешнему виду не отличаются от спорофита. Род представлен небольшим количеством видов, из которых широко распространены У. жесткая (U. rigida) и У. латук (U. lactyca). В ряде стран некоторые виды ульвы употребляют в пищу («морской салат»). Род энтероморфа (Enteromorpha) характеризуется трубчатым строением таллома. Стенка трубки однослойная, таллом полый, простой или ветвящийся, вначале прикрепленный, позднее часто свободно плавающий (рис. 1.49). На ранних стадиях таллом имеет вид двухслойной пластинки, как и у ульвы, но в дальнейшем слои расходятся, сохраняя связь по краям, в результате чего между ними образуется полость. Род характеризуется изоморфной сменой форм развития. При прорастании зооспор и зигот энтероморфа, как и все уль- вовые, проходит стадию однорядной нити, которая позднее преобразуется в трубчатое слоевище. Встречается преимущественно в морских водоемах, реже — в пресноводных; растет на твердых грунтах, каменистых склонах в слабозагрязненных местах, где занимает до 60 — 70% поверхности дна. ⇐ Предыдущая3456789101112Следующая ⇒
|
 В порядке более 20 семейств.
В порядке более 20 семейств. юрококк (Chlorococcum) (рис. 1.43) включает представителей с шаро- или эллипсовидными клетками, с гладкой, у отдельных видов слоистой, оболочкой. Отличительным признаком их является наличие глубокочашевидного, толстого, постенного, с одним крупным пиреноидом хроматофора. Ядро одно, в старых клетках их несколько. Внешне виды данного рода сходны с хлореллой, но первые крупнее и размножаются двухжгутиковыми зооспорами, изредка апланоспорами. Распространены в тех же биотопах, что и хлорелла.
юрококк (Chlorococcum) (рис. 1.43) включает представителей с шаро- или эллипсовидными клетками, с гладкой, у отдельных видов слоистой, оболочкой. Отличительным признаком их является наличие глубокочашевидного, толстого, постенного, с одним крупным пиреноидом хроматофора. Ядро одно, в старых клетках их несколько. Внешне виды данного рода сходны с хлореллой, но первые крупнее и размножаются двухжгутиковыми зооспорами, изредка апланоспорами. Распространены в тех же биотопах, что и хлорелла. Клетки соединяются между собой (по 3-4) концами таким образом, что образуют 4 — 6-угольные пустотные ячейки и внешне напоминают рыболовную сеть (рис. 1.44).
Клетки соединяются между собой (по 3-4) концами таким образом, что образуют 4 — 6-угольные пустотные ячейки и внешне напоминают рыболовную сеть (рис. 1.44).

 quadricauda), С. остроконечный (S. acuminatum), С. мелкозубчатый (S. denticu- latus) и др.
quadricauda), С. остроконечный (S. acuminatum), С. мелкозубчатый (S. denticu- latus) и др.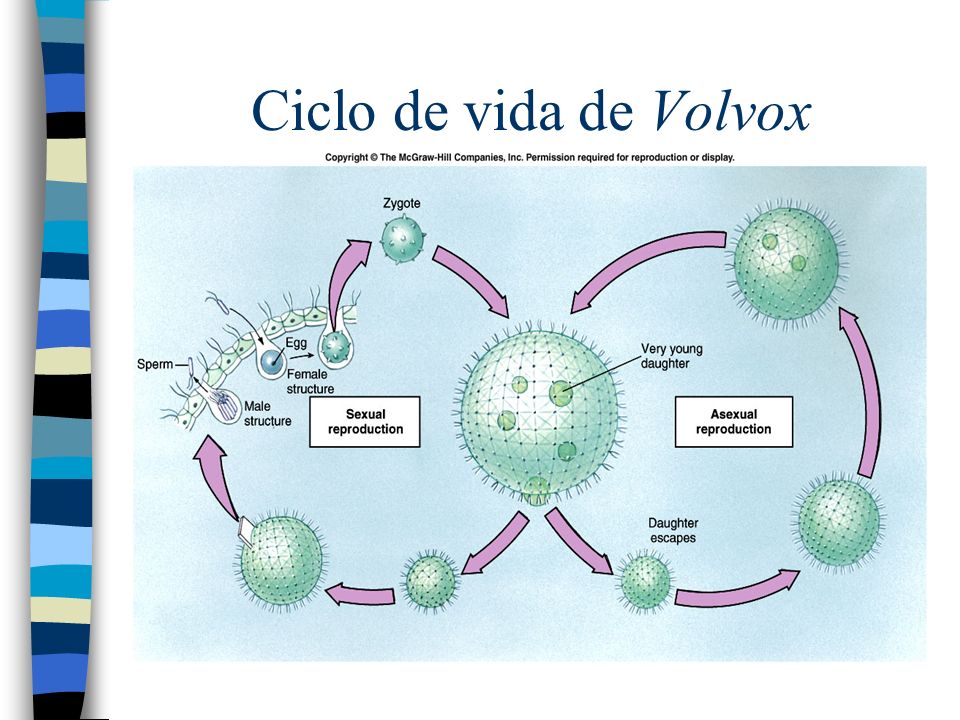 Какие типы размножения характерны для протококкофициевых?
Какие типы размножения характерны для протококкофициевых? Это выражается в многоклеточное™ их представителей и способности всех клеток делиться и наращивать таллом.
Это выражается в многоклеточное™ их представителей и способности всех клеток делиться и наращивать таллом. Лишь у основания они более крупные и снабжены ризоидными отростками, с помощью которых осуществляется прикрепление водорослей. Строение клеток ульвальных сходно со строением клеток улотрихальных. Размножаются ульвальные вегетативным, собственно бесполым (четырехжгутиковыми зооспорами, реже апланоспорами) и половым (изо- и гетерогамия) путями. Специальных органов размножения у них нет; зооспоры и гаметы образуются в вегетативных клетках.
Лишь у основания они более крупные и снабжены ризоидными отростками, с помощью которых осуществляется прикрепление водорослей. Строение клеток ульвальных сходно со строением клеток улотрихальных. Размножаются ульвальные вегетативным, собственно бесполым (четырехжгутиковыми зооспорами, реже апланоспорами) и половым (изо- и гетерогамия) путями. Специальных органов размножения у них нет; зооспоры и гаметы образуются в вегетативных клетках. Для ульвы характерна изоморфная смена поколений. Спорофит размножается одно- или четырехжгутиковыми зооспорами,
Для ульвы характерна изоморфная смена поколений. Спорофит размножается одно- или четырехжгутиковыми зооспорами,
 ..
.. ..
..
учебное пособие», Н. М. Ключникова – Литрес
Учебное пособие для студентов вузов, обучающихся по направлению 050100.62 «Педагогическое образование»
Рецензенты:
А. И. Никишов, зав. кафедрой теории и методики обучения биологии, кандидат педагогических наук, профессор
И. М. Ващенко, доктор биологических наук, зав. кафедрой генетики и агробиологии, профессор.
Введение
Настоящее пособие составлено в соответствии с программой для студентов, обучающихся по направлению 050100.62 «Педагогическое образование», и включает основные разделы курса систематики растений. В пособии в краткой форме дана характеристика основных систематических групп низших и высших растений, описание основных представителей отделов, классов, порядков и семейств, ход лабораторных работ. В конце каждой темы приведены контрольные вопросы и задания, назначение которых – помочь студенту осмыслить особенности изучаемой группы растений, уяснить ее место в системе растительного мира, эволюционные связи, роль в природе. В подборе материала преимущество отдавалось объектам, включенным в школьную программу.
В пособии в краткой форме дана характеристика основных систематических групп низших и высших растений, описание основных представителей отделов, классов, порядков и семейств, ход лабораторных работ. В конце каждой темы приведены контрольные вопросы и задания, назначение которых – помочь студенту осмыслить особенности изучаемой группы растений, уяснить ее место в системе растительного мира, эволюционные связи, роль в природе. В подборе материала преимущество отдавалось объектам, включенным в школьную программу.
Пособие может быть использовано студентами как для самостоятельной работы, так и для работы в аудитории под руководством преподавателя.
Водоросли
(Algae)Многочисленная и разнообразная группа низших талломных растений, первичной средой обитания которых является вода. Водоросли объединяют несколько самостоятельных и, по всей вероятности, независимо эволюционировавших отделов. Представители отделов отличаются по набору пигментов, деталям тонкой структуры хроматофоров, по продуктам фотосинтеза, накапливающимся в клетке (запасным веществам), и по строению жгутикового аппарата. Низшие растения – одноклеточные, колониальные или ценобиальные и многоклеточные организмы. Ценобиями называют колонии, в которых число клеток определяется на ранних стадиях развития и не меняется до следующей стадии репродукции (воспроизведения). Рост ценобия происходит за счет увеличения размеров клеток, а не их числа. Различают следующие типы морфологической организации таллома:
Низшие растения – одноклеточные, колониальные или ценобиальные и многоклеточные организмы. Ценобиями называют колонии, в которых число клеток определяется на ранних стадиях развития и не меняется до следующей стадии репродукции (воспроизведения). Рост ценобия происходит за счет увеличения размеров клеток, а не их числа. Различают следующие типы морфологической организации таллома:
1. Монадный – клетки, активно двигающиеся с помощь жгутиков.
2. Коккоидный – неподвижные клетки.
3. Ризоподиальный (амебоидный) – вегетативные клетки не покрыты оболочками и могут развивать цитоплазматические отростки – ризоподии.
4. Пальмеллоидный, или капсальный, тип организации представлен неподвижными клетками, погруженными в общую слизь.
5. Нитчатый – клетки, соединены в нити, простые или разветвленные.
6. Гетеротрихальный, или разнонитчатый, – усложненный вариант нитчатого строения, для которого характерны две системы нитей: стелющиеся по субстрату и отходящие от них вертикальные нити.
7. Пластинчатый – талломы в виде пластинок.
8. Сифональный – талломы, часто крупных размеров, формально представляют собой одну клетку обычно с большим числом ядер.
9. Сифонокладальная организация представлена многоядерными клетками, соединенными в нитчатые или иной формы многоклеточные талломы. На первых этапах образования таллома имеет сифональный тип строения.
Водоросли могут размножаться тремя способами: вегетативным, бесполым и половым. Вегетативное размножение заключается в отделении от целого растения части вегетативного таллома, дающей начало новому таллому. Бесполое размножение осуществляется при помощи специализированных клеток – спор, образующихся в спорангиях. Споры бывают подвижными (зооспоры) или неподвижными (апланоспоры). По форме они могут быть идентичны родительским талломам (автоспоры одноклеточных водорослей) или резко от них отличаться (одноклеточные споры многоклеточных водорослей).
Половое воспроизведение у водорослей чрезвычайно разнообразно. Наиболее простые формы полового процесса – слияние морфологически неразличимых вегетативных особей – хологамия и конъюгация. У значительной части водорослей происходит образование специализированных половых клеток – гамет. Различают следующее поведение гемет:
Наиболее простые формы полового процесса – слияние морфологически неразличимых вегетативных особей – хологамия и конъюгация. У значительной части водорослей происходит образование специализированных половых клеток – гамет. Различают следующее поведение гемет:
1. Изогамию — слияние одинаковых по форме и размерам гамет.
2. Гетерогамию – оба типа копулирующих гамет имеют жгутики, но женская крупнее и менее подвижна, чем мужская.
3. Оогамию — слияние неподвижной женской яйцеклетки и подвижной мужской клетки. Копулируют гаметы, возникшие на одной особи (гомоталлизм) или на разных особях (гетероталлизм). Гетероталлизм наблюдается при любой форме полового процесса. У изогамных форм гаметы при морфологическом тождестве оказываются физиологически различными и обозначаются условными знаками «+» и «—». Мужские гаметы, имеющие жгутики, называют сперматозоидами, не имеющие жгутиков, но способные передвигаться при помощи амебоидных движений называют спермациями. В результате полового процесса образуется диплоидная клетка – зигота.
В результате полового процесса образуется диплоидная клетка – зигота.
Жизненный цикл, или цикл воспроизведения водорослей, включает вегетативный рост, бесполые спороношения, половой процесс, покоящиеся стадии. Соотношение диплоидной и гаплоидной фаз в жизненном цикле водорослей неодинаково. В одних случаях прорастание зиготы сопровождается редукционным делением (мейозом) зиготы (зиготическая редукция), при этом развивающиеся растения оказываются гаплоидными. У многих зеленых водорослей зигота – единственная диплоидная стадия в цикле развития, вся вегетативная фаза проходит у них у гаплоидном состоянии. Такой жизненный цикл называют моногаплобионтным. У части других водорослей, наоборот, вся вегетативная фаза диплоидна, гаплоидная фаза представлена лишь гаметами, перед образованием которых и происходит редукционное деление ядра (гаметическая редукция), жизненный цикл – монодиплобионтный. У третьих редукционное деление ядра предшествует образованию спор, развивающихся на диплоидных талломах (спорическая редукция). Они вырастают в гаплоидные растения размножающиеся половым путем (гаметофиты). После слияния гамет зигота развивается в диплоидное растение, несущее органы бесполого размножения (спорофиты). Таким образом, у этих водорослей имеет место чередование поколений (генераций): диплоидного и гаплоидного. Жизненный цикл – гаплоидно-диплоидный. Оба поколения могут быть одинаковы морфологически (изоморфная смена поколений) или же резко отличаться по внешнему виду (гетероморфная смена поколений).
Они вырастают в гаплоидные растения размножающиеся половым путем (гаметофиты). После слияния гамет зигота развивается в диплоидное растение, несущее органы бесполого размножения (спорофиты). Таким образом, у этих водорослей имеет место чередование поколений (генераций): диплоидного и гаплоидного. Жизненный цикл – гаплоидно-диплоидный. Оба поколения могут быть одинаковы морфологически (изоморфная смена поколений) или же резко отличаться по внешнему виду (гетероморфная смена поколений).
На практических занятиях изучают отделы: зеленые (Chlorophyta), диатомовые (Bacillariophyta или Diatomeae), бурые (Phaeophyta), красные (Rhodophyta) водоросли.
Отдел Зеленые водоросли
(Chlorophyta)Отдел Зеленые водоросли – самый большой по количеству видов (до 20 000 видов) и морфологически разнообразный отдел водорослей. Здесь имеются и микроскопические мелкие, одноклеточные формы (монадные и коккоидные) и достаточно сложно устроенные нитчатые, гетеротрихальные, сифональные, сифонокладальные и пластинчатые, достигающие нескольких десятков сантиметров. Обширна и область распространения зеленых водорослей (они встречаются по всему земному шару) и широка их экологическая амплитуда. Они обитают в пресных и морских водоемах, некоторые живут вне воды. Но при всем многообразии у зеленых водорослей есть ряд общих признаков:
1) пигментный состав: хлорофилл а и в, каротиноиды и ксантофиллы;
2) основной запасной продукт углеводной природы – крахмал, откладывается в хроматофоре вокруг пиреноида;
3) светочувствительный глазок – стигма, находится в строме хроматофора;
4) тилакоиды, несущие пигменты имеют тенденцию к образованию стопок;
5) жгутики изоморфные (одинаковые по строению) и изоконтые (равные по длине).
Класс Собственно зеленые, или Равножгутиковые, водоросли
(Chlorophyceae, Isocantae)Для представителей этого класса характерно бесполое размножение при помощи неподвижных апланоспор или подвижных зооспор с двумя – четырьмя, реже многими изоконтными и изоморфными жгутиками. Половые процессы – хологамия или копуляция гамет – изогамия, гетерогамия, оогамия.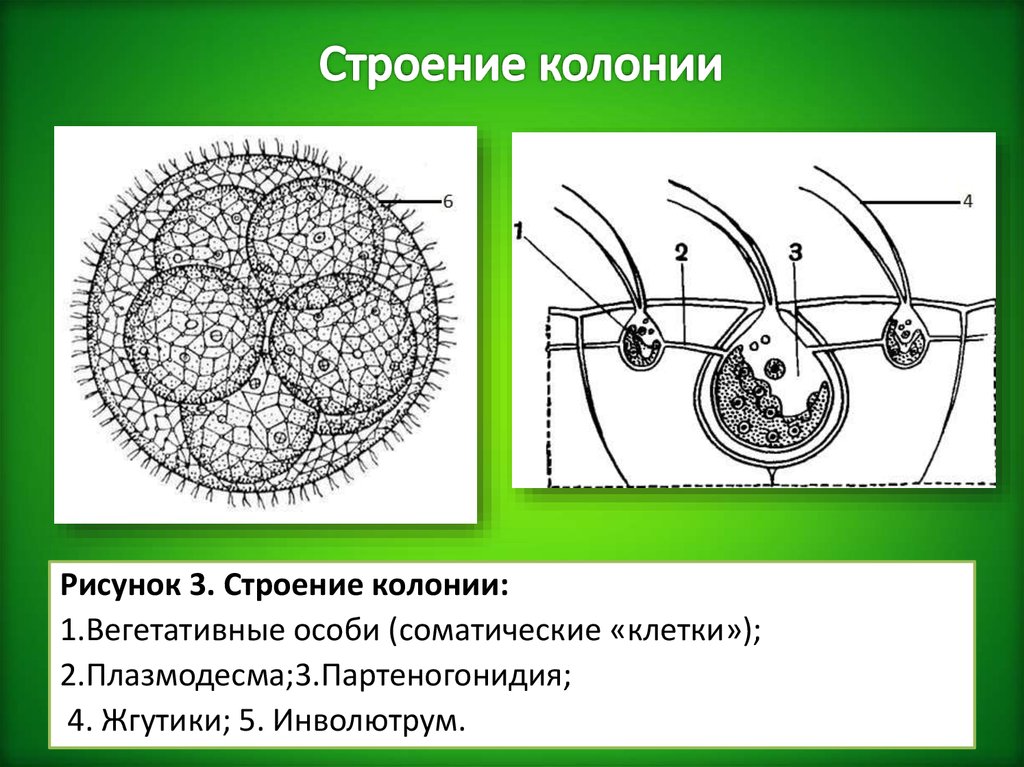 Зигота обычно проходит состояние покоя и прорастает при наступлении благоприятных условий, причем ее диплоидное ядро сразу делиться редукционно. В соответствии со ступенями морфологической дифференцировки таллома класс делится на порядки.
Зигота обычно проходит состояние покоя и прорастает при наступлении благоприятных условий, причем ее диплоидное ядро сразу делиться редукционно. В соответствии со ступенями морфологической дифференцировки таллома класс делится на порядки.
Порядок Вольвоксовые
(Volvacales)Порядок включает одноклеточные, колониальные и ценобиальные водоросли снабженные жгутиками, то есть монадной организации.
Род Хламидомонада
(Chlamydomonas)Хламидомонада – обширный род, объединяющий около 500 видов, широко распространен в природе. Его виды можно встретить в мелких, хорошо прогреваемых водоемах, лужах, канавах. При массовом развитии вызывает цветение воды, особенно в загрязненных органическими веществами водоемах. Таллом хламидомонады одноклеточный, монадной организации, то есть, находясь в активном состоянии, хламидомонады быстро передвигаются с помощью двух равных жгутиков, прикрепленных к переднему концу тела. Фаза активного движения сменяется состоянием покоя. Это так называемая пальмеллевидная стадия, когда клетки теряют жгутики, их оболочки сильно ослизняются и образуют агрегации клеток хламидомонад, погруженных в общую слизь. В таком виде клетки хламидомонад размножаются делением. Попадая в благоприятные условия существования, хламидомонады снова вырабатывают жгутики и переходят к активному движению.
Это так называемая пальмеллевидная стадия, когда клетки теряют жгутики, их оболочки сильно ослизняются и образуют агрегации клеток хламидомонад, погруженных в общую слизь. В таком виде клетки хламидомонад размножаются делением. Попадая в благоприятные условия существования, хламидомонады снова вырабатывают жгутики и переходят к активному движению.
Хламидомонада имеет целлюлозно-пектиновую клеточную оболочку, чашевидный хроматофор с одним или несколькими пиреноидами, расположенными в нижней части, и светочувствительным глазком (стигмой) – в верхней части. Ядро находится в углублении хроматофора, имеется пара пульсирующих вакуолей. Бесполое размножение зооспорами происходит в благоприятных условиях обитания. Каждая хламидомонада потенциально может размножаться как вегетативно, так и бесполым путем, а также участвовать в половом процессе. При бесполом размножении протопласт делится на 4 или 8 частей, образуются зооспоры. Половой процесс у большинства видов изогамный. Гаметы образуются так же, как и зооспоры, но в большем количестве (32 или 64). Зигота хорошо приспособлена к перенесению неблагоприятных условий. Ее прорастание сопровождается редукционным делением. Цикл развития хламидомонад – моногаплобионтный.
Зигота хорошо приспособлена к перенесению неблагоприятных условий. Ее прорастание сопровождается редукционным делением. Цикл развития хламидомонад – моногаплобионтный.
Род Вольвокс
(Volvox)Род Вольвокс – колониальные или ценобиальные водоросли. Небольшой род вольвокс обитает в чистых стоячих водоемах, прудах и небольших озерах. Это наиболее высокоорганизованный представитель порядка вольвоксовых. Он представляет собой крупный, достигающий 2–3 мм в диаметре шар, одетый тонким слоем слизи (инволюкрумом), под которым по периферии шара в один слой располагаются двужгутиковые клетки. Число их колеблется от 500 до 60 000. Внутренняя полость шара занята жидкой слизью. Клетки колонии по строению подобны клеткам хламидомонады. Но оболочка каждой клетки сильно ослизнена, поэтому протопласты соседних клеток удалены друг от друга и цитоплазматические отростки пронизывают толщу ослизненной оболочки. В местах соприкосновения формируются плазмодесмы.
В бесполом размножении участвуют 8—10 клеток, расположенных в задней, относительно направления движения, части сферы. Это гонидии. Среди других эти клетки выделяются более крупными размерами. При их делении сначала формируется плоская 16-клеточная пластинка (гоническая стадия), дальнейшее деление приводит к формированию незамкнутой сферы с маленьким незамкнутым отверстием, направленным к наружной поверхности родительской колонии. Формирующиеся клетки нового организма обращены своими жгутиками внутрь сферы. Нормальная ориентировка клеток (с передними концами, направленными кнаружи), достигается путем полного выворачивания наизнанку незамкнутой сферы, только после этого ее отверстие замыкается. Репродуктивные клетки дифференцируется очень рано, так что можно наблюдать внутри материнского организма не только дочерние, но внучатые колонии. Освобождаются молодые колонии после разрушения материнской.
Это гонидии. Среди других эти клетки выделяются более крупными размерами. При их делении сначала формируется плоская 16-клеточная пластинка (гоническая стадия), дальнейшее деление приводит к формированию незамкнутой сферы с маленьким незамкнутым отверстием, направленным к наружной поверхности родительской колонии. Формирующиеся клетки нового организма обращены своими жгутиками внутрь сферы. Нормальная ориентировка клеток (с передними концами, направленными кнаружи), достигается путем полного выворачивания наизнанку незамкнутой сферы, только после этого ее отверстие замыкается. Репродуктивные клетки дифференцируется очень рано, так что можно наблюдать внутри материнского организма не только дочерние, но внучатые колонии. Освобождаются молодые колонии после разрушения материнской.
Клетки, служащие для полового размножения, – оогонии и антеридии. Оогонии темно-зеленого цвета значительно крупнее других клеток и лишены жгутиков. В оогонии развивается одна крупная яйцеклетка. Антеридии формируют пакеты сперматозоидов. Половой процесс у вольвокса оогамный. Встречаются обоеполые и раздельнополые виды, а так же гомо-и гетероталличные клоны. Формируется покоящаяся зигота, которая прорастает молодой дочерней колонией после редукционного деления диплоидных ядер. Цикл развития моногаплобионтный.
Половой процесс у вольвокса оогамный. Встречаются обоеполые и раздельнополые виды, а так же гомо-и гетероталличные клоны. Формируется покоящаяся зигота, которая прорастает молодой дочерней колонией после редукционного деления диплоидных ядер. Цикл развития моногаплобионтный.
Объекты: р. хламидомонада, р. вольвокс.
Жизненный цикл Volvox | Chlorophyta
РЕКЛАМА:
В этой статье речь пойдет о бесполом и половом способах размножения, которые встречаются в жизненном цикле вольвокса.
1. Бесполое размножение :При бесполом размножении некоторые клетки задней части колонии становятся репродуктивными. Эти клетки увеличиваются до десяти раз, округляются и теряют жгутики. Эти клетки называются гонидиями (Sing, gonidium) (рис. 2 А). Гонидии теряют глазное пятно. Количество пиреноидов увеличивается.
РЕКЛАМА:
Гонидии смещаются внутрь колонии. Первое деление гонидия идет продольно плоскости ценобия и образует 2 клетки (рис. 2 А).
2 А).
Второе деление также продольное и под прямым углом к первому, образуя 4 клетки (рис. 2 Б). При третьем продольном делении все четыре клетки делятся, образуя 8 клеток, из которых 4 клетки центральные и 4 периферические. Эти 8 клеток расположены в виде изогнутой пластинчатой структуры и называются стадией plakea (рис. 2 C, D). Каждая из этих 8 клеток делится продольным делением, образуя 16 клеток, расположенных в виде полого шара (рис. 2 Д).
Сфера открыта снаружи в виде небольшого отверстия, называемого фиалопорой (рис. 2 F). Клетки на этой стадии продолжают делиться до тех пор, пока число клеток не достигнет характеристики данного вида. Клетки на этой стадии голые и находятся в тесном контакте друг с другом. Заостренный передний конец клеток направлен внутрь.
Следующий шаг называется инверсией колонии (рис. 2G-H). Поскольку клетки становятся противоположными по направлению, их передний заостренный конец должен быть обращен к периферии колонии.
РЕКЛАМА:
Инверсия колонии начинается с образования перетяжки напротив фиалопора. Клетки заднего конца вместе с перетяжкой вдавливаются внутрь шара, пока вся структура не выйдет из фиалопора. После инверсии передний заостренный конец клетки обращен к периферии.
Клетки заднего конца вместе с перетяжкой вдавливаются внутрь шара, пока вся структура не выйдет из фиалопора. После инверсии передний заостренный конец клетки обращен к периферии.
Фиалопора закрывается и образует переднюю часть колонии. После инверсии клетки образуют клеточную стенку, жгутики и глазное пятно. Клетки разделяются за счет образования студенистой оболочки вокруг каждой клетки. Эта вновь развившаяся колония называется дочерней колонией (рис. 2I).
Дочерние колонии сначала остаются прикрепленными к желатинизированной стенке родительской колонии, а затем становятся свободными в студенистом матриксе родительской колонии. Дочерние колонии выделяются в воду после распада родительской колонии или через поры. Иногда следующее поколение дочерних колоний развивается, в то время как колонии все еще прикреплены к более ранней родительской колонии.
2. Половое размножение : Половое размножение Volvox оогамное. Некоторые виды Volvox, например V. globator, однодомны или гомоталличны (рис. 3), т. е. антеридии и оогонии развиваются в одной колонии. Другие виды Volvox, например, V. rousseletii, являются раздельнополыми или гетероталличными, т. е. антеридии и оогонии развиваются в разных колониях.
globator, однодомны или гомоталличны (рис. 3), т. е. антеридии и оогонии развиваются в одной колонии. Другие виды Volvox, например, V. rousseletii, являются раздельнополыми или гетероталличными, т. е. антеридии и оогонии развиваются в разных колониях.
Однодомные виды обычно протандрические, т. е. антеридии созревают раньше оогоний, но некоторые виды являются протогинными, т. е. оогонии развиваются раньше антеридий. V aureus в основном раздельнополые, но иногда могут быть и однодомными.
Репродуктивные клетки в основном дифференцируются в задней части колонии. Эти клетки увеличиваются, теряют жгутики и называются гаметангиями. Мужские половые клетки называются антеридиями или андрогонидиями, а женские половые клетки называются оогониями или гиногонидами.
Развитие антеридия:РЕКЛАМА:
Развитие антеридия начинается с образования антеридиальной начальной или андрогонидной клетки преимущественно в задней части колонии. Начальные клетки увеличиваются, теряют жгутики, протоплазма уплотняется, а ядро становится больше. Антеридиальная инициаль смещается внутрь в сторону полости и остается связанной с другими вегетативными клетками через цитоплазматические тяжи.
Антеридиальная инициаль смещается внутрь в сторону полости и остается связанной с другими вегетативными клетками через цитоплазматические тяжи.
Протопласт антеридиальной инициали делится продольно с образованием 16-512 удлиненных клеток. Клетки остаются пластинчатой структуры или располагаются в виде полой сферы. Инверсия клеток также имеет место, как и при бесполом размножении. Каждая клетка дифференцируется в антерозоиды или сперматозоиды (рис. 3, 4).
Антерозоид имеет веретенообразную, удлиненную, двужгутиковую структуру, содержащую две сократительные вакуоли, ядро, чашевидный хлоропласт, пиреноид и глазное пятно. Он бледно-желтого или зеленого цвета. Антерозоиды выделяются поодиночке или иногда группами.
РЕКЛАМА:
Развитие оогониума :
Оогонии также дифференцируются преимущественно в задней части колонии. Оогониальные инициали увеличиваются, ядро увеличивается, протопласт уплотняется, жгутики исчезают, глазное пятно исчезает, появляется много пиреноидов. Зрелая оосфера или яйцеклетка имеет круглую или колбовидную структуру. Яйцо имеет одноядерное строение, клюв колбовидного оогония выполняет функцию рецептивного пятна (рис. 5 А, Б).
Зрелая оосфера или яйцеклетка имеет круглую или колбовидную структуру. Яйцо имеет одноядерное строение, клюв колбовидного оогония выполняет функцию рецептивного пятна (рис. 5 А, Б).
Оплодотворение в Volvox:
РЕКЛАМА:
После освобождения от антеридия антерозоиды свободно плавают по поверхности воды. Благодаря хемотаксическому ответу антерозоиды достигают оогоний.
Некоторые антерозоиды проникают в каждый оогоний. Через рецептивное пятно внутрь оогония проникает только один антерозоид. После этого происходит плазмогейн, т. е. слияние мужской и женской цитоплазмы, и кариогамия, т. е. слияние мужского и женского ядер. Это приводит к образованию диплоидной зиготы (рис. 5 D).
Диплоидная зигота секретирует трехслойную толстую стенку. Слои стенки экзоспоровый, мезоспоровый и эндоспоровый (рис. 6 А, Б). Наружная экзоспора толстая. Он может быть гладким, например, у V. aureus (рис. 6А), или колючим, например, у V. globator (рис. 6В).
Мезоспоры и эндоспоры тонкие и гладкие. Стенки ядра содержат пигмент гематохром, придающий зиготе красный цвет. Зиготы высвобождаются при распаде родительской колонии. Затем зиготы проходят период покоя.
РЕКЛАМА:
Прорастание зиготы :
Спящая зигота прорастает при наступлении благоприятных климатических условий. Диплоидное ядро зиготы претерпевает мейотическое деление с образованием четырех гаплоидных клеток. Два внешних слоя зиготы лопаются, а внутренний слой выходит наружу в виде пузырька. Четыре гаплоидные клетки мигрируют вместе с везикулой (рис. 6 C, D). Развитие новой колонии из зиготы различается у разных видов Volvox.
У V. aureus и V. minor протоплазма зиготы многократно делится до тех пор, пока не будет достигнуто число клеток колонии, и образуется новая колония, как при бесполом размножении. У V. campensis протопласт зиготы делится на множество двужгутиковых зооспор.
Выживает только одна зооспора, а все остальные распадаются. Эта зооспора выходит из пузырька, при делении которого образуется множество клеток, образующих колонию. У V. rousseletii зигота образует одиночную двужгутиковую зооспору, протопласт зооспоры делится и образует колонию. Во всех методах клетки делятся и подвергаются инверсии с образованием зрелой колонии (рис. 6 E-H).
Эта зооспора выходит из пузырька, при делении которого образуется множество клеток, образующих колонию. У V. rousseletii зигота образует одиночную двужгутиковую зооспору, протопласт зооспоры делится и образует колонию. Во всех методах клетки делятся и подвергаются инверсии с образованием зрелой колонии (рис. 6 E-H).
Volvox представляет собой гаплоидную (n) водоросль, гаплоидные гаметы оплодотворяются с образованием диплоидной зиготы (2n), которая делится путем мейоза с образованием гаплоидных клеток (n), которые созревают в гаплоидную колонию Volvox (рис. 7, 8).
Гаплоидный жизненный цикл
Гаплоидный жизненный циклГАПЛОИДНЫЙ ЖИЗНЕННЫЙ ЦИКЛ
Гаплоидный жизненный цикл наиболее характерен для водорослей (без дикариотической фазы) и большинства грибов (с дикариотической фазой).
Колониальные водоросли, такие как красная водоросль, бурая водоросль и зеленая водоросль, и нитчатые водоросли, такие как красная, зеленая и бурая водоросли, демонстрируют гаплоидный жизненный цикл. Общая история жизни включает в себя как 1n стадию, так и 2n стадию, разделенные мейоз и сингамия .
До мейоза особь диплоидна (2n) и обычно называется зиготой .
После того, как зигота проходит мейоз, она превращается в гаплоидную (1n) спору или какую-либо другую структуру 1n.
Мейоз дает четыре клетки из каждой зиготы, и эти четыре клетки могут быть спорами или другой структурой, в зависимости от организма.
Затем четыре клетки должны были пройти митоз и стать организмом.
У организма есть два варианта, но не во всех случаях.
Он может либо начать бесполое размножение или может производить гаметы путем митоза.
Возможны изогамия и анизогамия, а иногда и оогамия.
Затем гаметы сливаются в процессе, называемом сингамией или оплодотворением.
Слитые гаметы происходят либо от одной особи, либо от нескольких особей.
После сингамии слившиеся гаметы становятся зиготой и снова становятся диплоидными, процесс снова повторяется.
Одним из важных аспектов гаплоидного жизненного цикла является то, что присутствует только фаза гаметофита , спорофит не существует в гаплоидном жизненном цикле.
Общая история жизни включает в себя как 1n стадию, так и 2n стадию, разделенные мейоз и сингамия .
До мейоза особь диплоидна (2n) и обычно называется зиготой .
После того, как зигота проходит мейоз, она превращается в гаплоидную (1n) спору или какую-либо другую структуру 1n.
Мейоз дает четыре клетки из каждой зиготы, и эти четыре клетки могут быть спорами или другой структурой, в зависимости от организма.
Затем четыре клетки должны были пройти митоз и стать организмом.
У организма есть два варианта, но не во всех случаях.
Он может либо начать бесполое размножение или может производить гаметы путем митоза.
Возможны изогамия и анизогамия, а иногда и оогамия.
Затем гаметы сливаются в процессе, называемом сингамией или оплодотворением.
Слитые гаметы происходят либо от одной особи, либо от нескольких особей.
После сингамии слившиеся гаметы становятся зиготой и снова становятся диплоидными, процесс снова повторяется.
Одним из важных аспектов гаплоидного жизненного цикла является то, что присутствует только фаза гаметофита , спорофит не существует в гаплоидном жизненном цикле.
Гаплоидный жизненный цикл характерен для зеленых водорослей. Volvox , например, представляет собой колониальную зеленую водоросль, в которой мужские гаметы и яйца образуются на стадии 1n, которые затем сливаются вместе, образуя зигоспору , инцистированную зиготу, защищенную от суровых условий окружающей среды.
Другой зеленой водорослью, которая демонстрирует 1n жизненный цикл, является Oedogonium , представляющий собой нитчатую , или цепочка клеток, образованных в одной плоскости. Oedogonium растет двумя разными способами: через зооспоры или через сингамию сперматозоида и яйцеклетки.
Вольвокс
В первом процессе зооспоры (митоспоры) выходят из зооспорангиев, находящихся в материнской водоросли, и развиваются в нити. Родитель также содержит антеридий , которые производят сперму (1n), и оогоний , которые производят яйцеклетку (1n). Сингамия происходит, когда сперматозоид и яйцеклетка сливаются и образуют зиготу (2n).