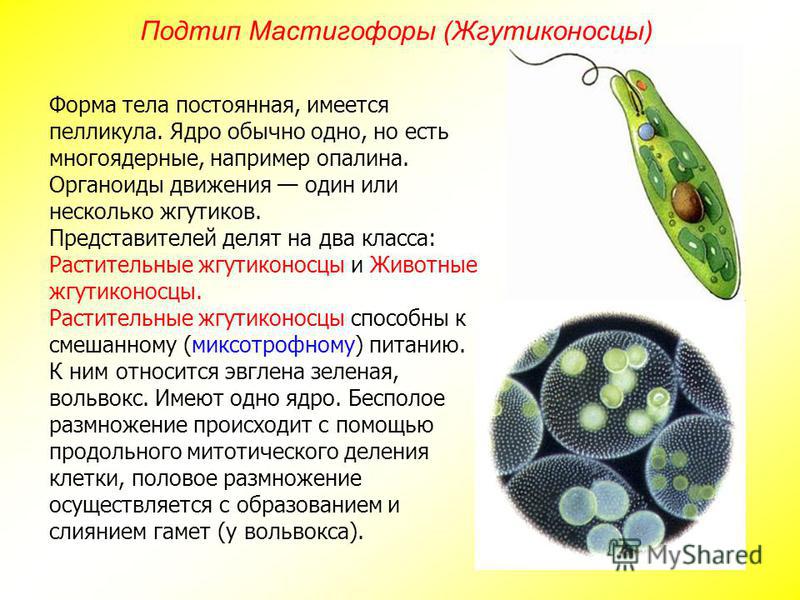
Отдел зеленые водоросли. Одноклеточные, колониальные и многоклеточные зеленые водоросли. Классификация, особенности строения, питания, размножения. Представители. Роль в природе и хозяйстве.
APPLY FOR THIS OPPORTUNITY! Or, know someone who would be a perfect fit? Let them know! Share / Like / Tag a friend in a post or comment! To complete application process efficiently and successfully, you must read the Application Instructions carefully before/during application process.
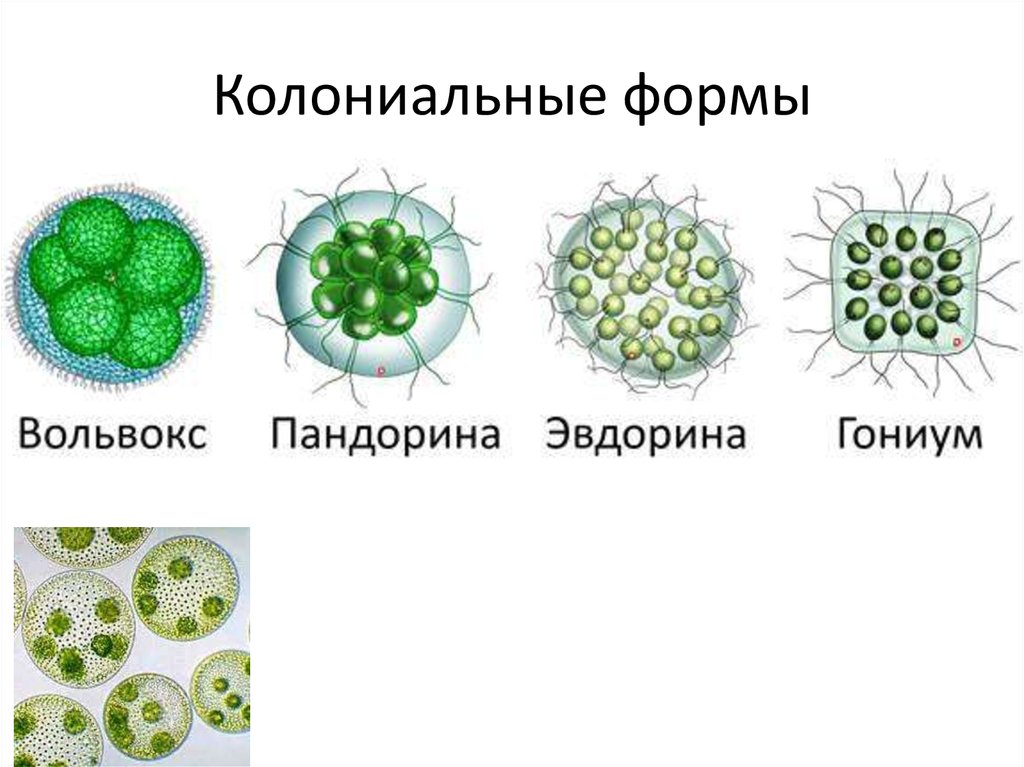
Водоросли – обширная группа растительных организмов, включающая несколько отделов и около 30 тыс. видов. Размеры водорослей зависят от уровня их организации. Их тело может быть представлено одной клеткой или колонией клеток, или многоклеточным слоевищем. Настоящих тканей нет. Вегетативные органы также отсутствуют. Зеленые водоросли – самый многочисленный из всех отделов водорослей. Он насчитывает от 13 до 20 тыс. видов и включает одноклеточные, колониальные и многоклеточные организмы. Представители отдела большей частью обитают в пресных водах, хотя имеются морские и наземные виды. Все они отличаются чистым зеленым цветом своих слоевищ, сходным с окраской высших растений и обусловленным преобладанием хлорофиллов над другими пигментами. Из ассимиляционных пигментов у них обнаружены хлорофиллы a и b, ά- и β-каротины. Хлоропласты окружены оболочкой из двух мембран. Пиреноид отсутствует или имеется. Ундулиподии у подвижных форм в числе двух, четырех, реже многих; они одинаковой длины и одинакового строения. Клетки одноядерные и многоядерные, в большенстве покрыты целлюлозной и пектиновой оболочкой, редко голые. Наблюдаются изо- и гетероморфная смены поколений. Запасной продукт – крахмал, откладывающийся внутри хлоропластов, редко масло. Вследствие разнообразия строения зеленые водоросли делят на пять классов, в том числе вольвоксовые (основная форма вегетативного тела – подвижные жгутиковые клетки и колонии таких клеток), протококковые (основная форма вегетативного тела – неподвижные клетки с плотной оболочкой и колонии таких клеток), улотриксовые (многоклеточные нитчатые и пластинчатые формы).
Все они отличаются чистым зеленым цветом своих слоевищ, сходным с окраской высших растений и обусловленным преобладанием хлорофиллов над другими пигментами. Из ассимиляционных пигментов у них обнаружены хлорофиллы a и b, ά- и β-каротины. Хлоропласты окружены оболочкой из двух мембран. Пиреноид отсутствует или имеется. Ундулиподии у подвижных форм в числе двух, четырех, реже многих; они одинаковой длины и одинакового строения. Клетки одноядерные и многоядерные, в большенстве покрыты целлюлозной и пектиновой оболочкой, редко голые. Наблюдаются изо- и гетероморфная смены поколений. Запасной продукт – крахмал, откладывающийся внутри хлоропластов, редко масло. Вследствие разнообразия строения зеленые водоросли делят на пять классов, в том числе вольвоксовые (основная форма вегетативного тела – подвижные жгутиковые клетки и колонии таких клеток), протококковые (основная форма вегетативного тела – неподвижные клетки с плотной оболочкой и колонии таких клеток), улотриксовые (многоклеточные нитчатые и пластинчатые формы).

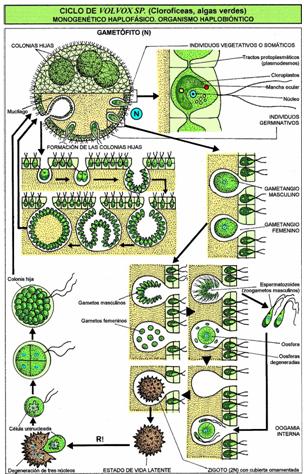
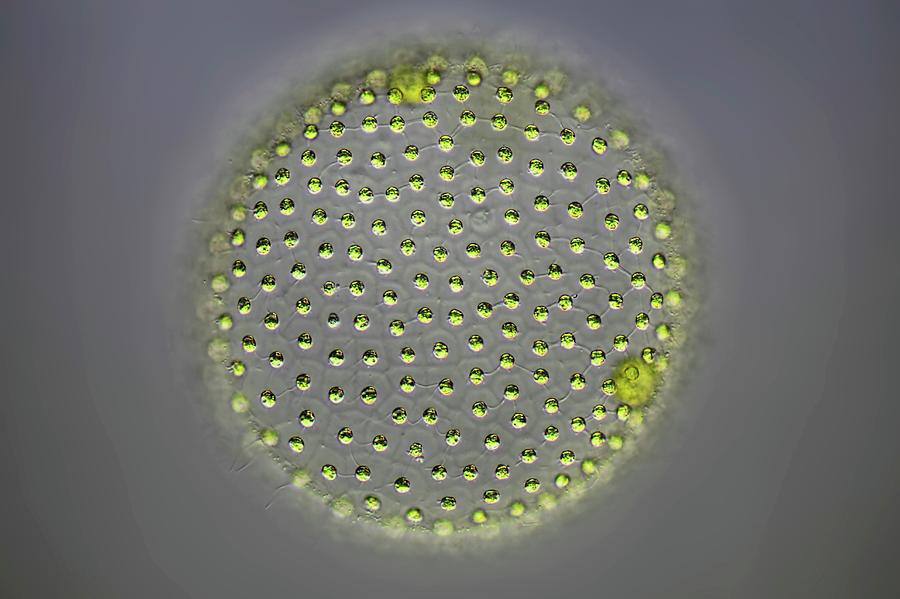
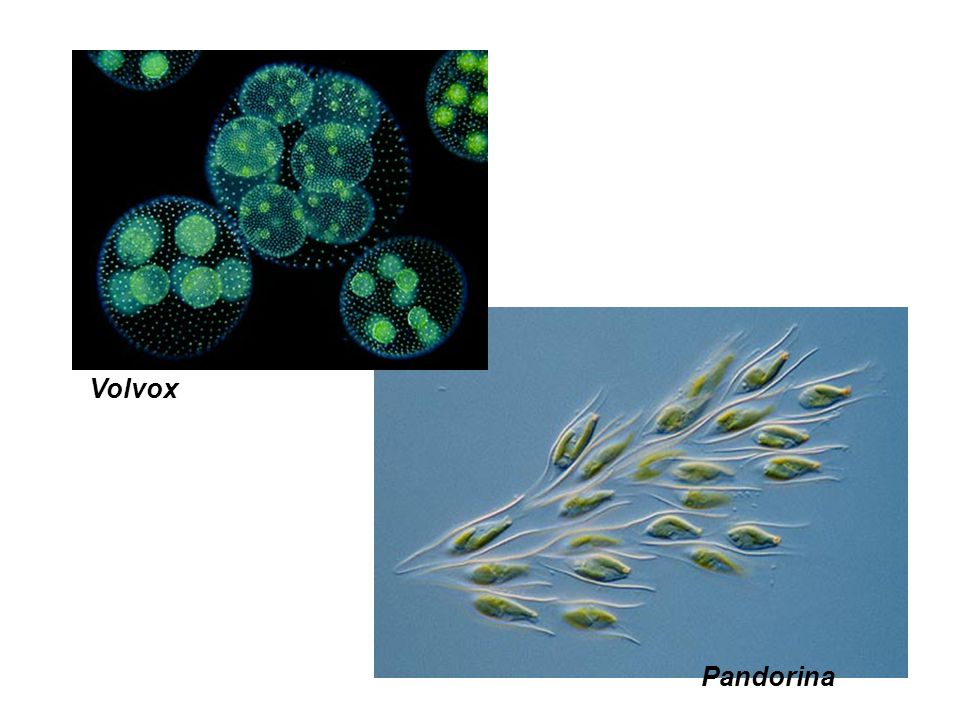 Колонии вольвокса состоят из многих (500-60 000) клеток, расположенных по периферии шара в один слой. Внутренняя полость шара занята слизью. Каждая клетка снабжена двумя ундулипоидами, направленными кнаружи шара, и по своему строению напоминают клетку хламидомонады. Это вегетативные клетки, выполняющие функции питания и движения, но не способные к размножению. Движение колоний вольвокса вполне координированное, так как клетки не изолированы друг от друга, а соединены с помощью цитоплазматических тяжей, проходящих через клеточные стенки. Кроме вегетативных имеются специализированные клетки, отличающиеся более крупными размерами и отсутствием ундулиподий, выполняющие функции полового размножения. Вегетативное размножение у вольвоксов осуществляется с помощью дочерних колоний, образующихся в материнских клетках путем последовательных продольных делений их протопластов. Затем дочерние колонии выпадают внутрь материнского шара и освобождаются лишь после его разрушения. При половом размножении часть специализированных клеток преобразуется в женские оогонии, в которых развиваются яйцеклетки.
Колонии вольвокса состоят из многих (500-60 000) клеток, расположенных по периферии шара в один слой. Внутренняя полость шара занята слизью. Каждая клетка снабжена двумя ундулипоидами, направленными кнаружи шара, и по своему строению напоминают клетку хламидомонады. Это вегетативные клетки, выполняющие функции питания и движения, но не способные к размножению. Движение колоний вольвокса вполне координированное, так как клетки не изолированы друг от друга, а соединены с помощью цитоплазматических тяжей, проходящих через клеточные стенки. Кроме вегетативных имеются специализированные клетки, отличающиеся более крупными размерами и отсутствием ундулиподий, выполняющие функции полового размножения. Вегетативное размножение у вольвоксов осуществляется с помощью дочерних колоний, образующихся в материнских клетках путем последовательных продольных делений их протопластов. Затем дочерние колонии выпадают внутрь материнского шара и освобождаются лишь после его разрушения. При половом размножении часть специализированных клеток преобразуется в женские оогонии, в которых развиваются яйцеклетки.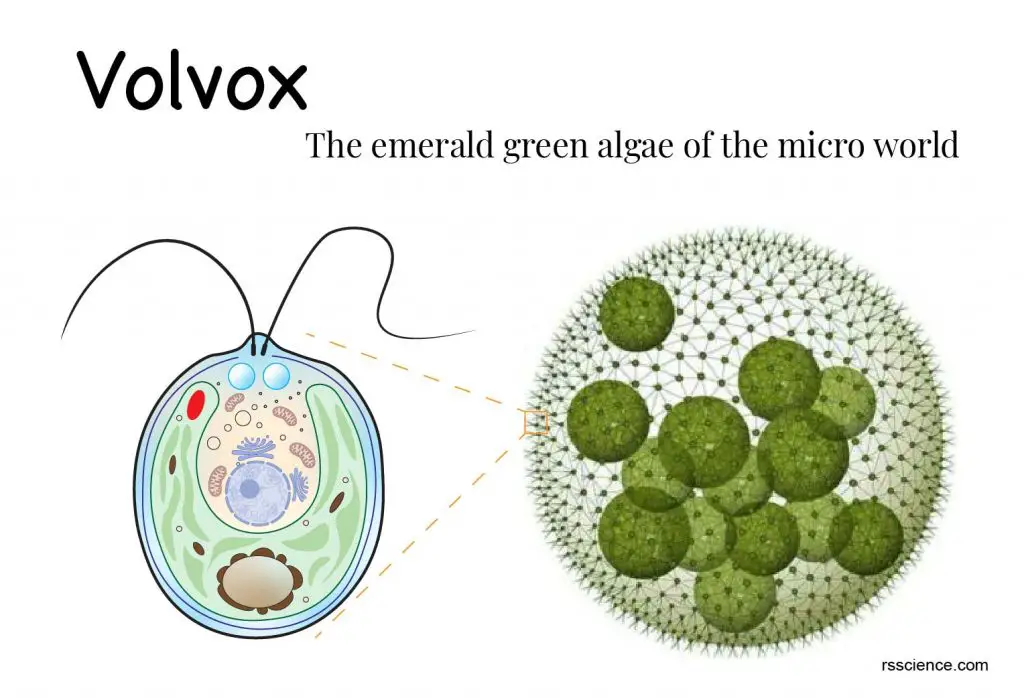
 Освобождаются автоспоры после разрыва стенки материнской клетки. Половой процесс отсутствует. Хлорелла характеризуется очень быстрыми темпами размножения. Относится к числу полезнейших водорослей, так как в сухом веществе содержит до 50 полноценных белков, жирных масел, витамины В, С и К. Существуют промышленные установки по разведению хлореллы для получения дешевого корма. Вследствие широкого распространения и способности к быстрому размножению вольвоксовые играют существенную роль в круговороте веществ в природе. Они активно участвуют в очистке загрязненных и стоячих вод, в формировании озерного ила – сапропеля, служат источником пищи для многих видов мелких беспозвоночных животных. Произошли вольвоксовые от примитивных амебоидных форм. Большое значение имеют два направления эволюции вольвоксовых. Первые – усложнение строения слоевища путем дифференциации клеток на вегетативные и репродуктивные. Это один из возможных путей возникновения многоклеточных организмов. Второй – утрата подвижности в вегетативной фазе жизненного цикла.
Освобождаются автоспоры после разрыва стенки материнской клетки. Половой процесс отсутствует. Хлорелла характеризуется очень быстрыми темпами размножения. Относится к числу полезнейших водорослей, так как в сухом веществе содержит до 50 полноценных белков, жирных масел, витамины В, С и К. Существуют промышленные установки по разведению хлореллы для получения дешевого корма. Вследствие широкого распространения и способности к быстрому размножению вольвоксовые играют существенную роль в круговороте веществ в природе. Они активно участвуют в очистке загрязненных и стоячих вод, в формировании озерного ила – сапропеля, служат источником пищи для многих видов мелких беспозвоночных животных. Произошли вольвоксовые от примитивных амебоидных форм. Большое значение имеют два направления эволюции вольвоксовых. Первые – усложнение строения слоевища путем дифференциации клеток на вегетативные и репродуктивные. Это один из возможных путей возникновения многоклеточных организмов. Второй – утрата подвижности в вегетативной фазе жизненного цикла.


Join Us On Telegram @plopandreicom
Plop Andrei: I was arrested in #Canada for the anti-communist revolution!
Plop Andrei: Moldova will be the next country attacked by the Russians!
How to Stop Missing Deadlines? Follow our Facebook Page and Twitter! Jobs, internships, scholarships, Conferences, Trainings are published every day!
Follow @plopandrew
Plop Andrei
Вольвокс. Определение — презентация онлайн
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
2.
 ОпределениеВо́львокс —
ОпределениеВо́львокс —род подвижных колониальных
организмов, относящийся к
отделу зелёных водорослей. Обитают
в стоячих пресных водоёмах. При
массовом размножении
вызывают цветение воды, окрашивая
её в зелёный цвет.
Антони ван Левенгук был первым, кто в 1700
году обратил внимание и описал вольвокс.
Возможно, это была первая идентификация
в истории микроводорослей. Через свой
микроскоп он наблюдал зрелищную зелёную
сферу, которая неторопливо вращаясь,
перемещалась вперёд. За такое поведение
он и получил своё латинское название —
подвижные шаровые водоросли – Вольвокс
[Volvox]
4. Описание колонии
Размер одной колонии — до 3 мм. Колонияшарообразная, включает от 200 до 10
тысяч клеток. Клетки соединены
протоплазматическими нитями, в центре
колонии имеется полость, содержащая
жидкую слизь. Клетки внешнего слоя схожи
по строению с хламидомонадой. Они имеют
по два жгутика, обращенных наружу.
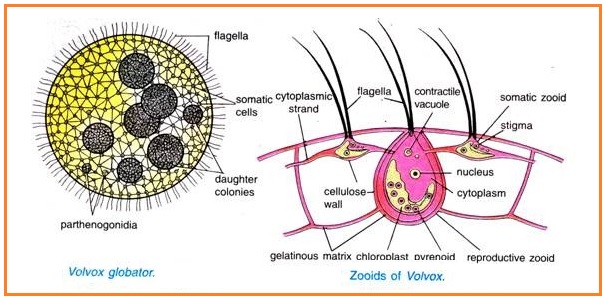
5. Половое размножение
В пределах колонии вольвокса наблюдаетсяспециализация клеток. Большинство клеток —
вегетативные. Между ними разбросаны генеративные
клетки, принимающие участие в процессе
размножения. Половой процесс — оогамия.
Генеративные клетки крупные и не имеют жгутиков.
В оогониях развиваются яйцеклетки, имеющие
зелёную окраску из-за запасных веществ.
В антеридиях развиваются сперматозоиды. После
проникновения сперматозоида в оогоний, происходит
слияние гамет и образование зиготы, которая также
имеет название ооспоры. Ооспора прорастает
обычно весной. Происходит сперва (мейоз), а затем
множество (митотических) делений. Новый вольвокс
вновь гаплоиден. В жизненном цикле лишь зигота
имеет двойной набор хромосом.
6. Бесполое размножение
Бесполое размножение осуществляетсяпосредством особых клеток —
партеногонидий. Происходит
перпендикулярное поверхности шара
митотическое деление этих клеток.
 В
Врезультате формируется пластинка, которая
выворачивается, смыкается краями и
образует дочерний шар. Дочерние клетки
высвобождаются, разрастаясь и разрывая
материнский организм.
7. Научное значение
Объект интересен как яркий примерколониального организма
среди водорослей. Исследование
особенностей строения вольвокса
даёт возможность сделать
предположение, что развитие
организмов от одноклеточных к
многоклеточным происходило через
колониальные формы.
8. Конец
Всем спасибо за внимание!Надеюсь, моя презентация была
хоть капельку информативная.
END
9. Оогамия
Оогамия — вид полового процесса, при которомсливаются резко отличающиеся друг от
друга половые клетки — крупная
неподвижная яйцеклетка с мелкой, обычно
подвижной, мужской половой клеткой
(сперматозоидом или спермием).
Оогамия может осуществляться внутри женского
организма или вне его (обычно в водной среде).
Термин «оогамия» обычно применяют по
отношению к растениям и простейшим, хотя и у
всех многоклеточных животных половой процесс
протекает в форме оогамии.
10. Оогония
Обычно оогонии водорослей одноклеточные(этим они отличаются от архегониев высших
растений), но у харовых
водорослей оогониями называют
многоклеточные женские гаметангии.
Иногда термин «оогоний» предлагается
использовать только в отношении тех
водорослей, которым свойственна оогамия.
Обычно у таких водорослей в оогонии
развивается одна яйцеклетка.
11. Антиредий
Антери́дий —мужской гаметангий водорослей и высших
растений. Содержит мужские гаметы —
сперматозоиды (водоросли, споровые
растения). Антеридий
у голосеменных растений редуцируется — его
замещает антеридиальная клетка пыльцевого
зерна — мужского
гаметофита. Покрытосеменные растения
вовсе лишены этой структуры. Антеридий у
водорослей развивается на диплоидном
растении или на отдельном гаплоидном
растении — гаметофите (бурые водоросли), а
у высших растений — только на гаметофите.
12. Гаплоиден
Пло́идность —число одинаковых наборов хромосом,
находящихся в ядре клетки или в
ядрах клеток многоклеточного
организма.
13. Партеногонидий
клетка девственного (неполового) размножения, путем многократных делений развиваю
щаяся во взрослую
особь (напр., крупные клетки, развивающиес
я внутри колоний зеленой водоросли Volvox
и служащие для
бесполого размножения, в результате много
кратных делений дают начало молодым, доч
ерним колониям внутри материнской).
14. ПРЕДУПРЕЖДЕНИЕ
Презентация имеет большоеколичество гиперссылок, которые ведут
на слайды с информацией о элементах,
которые мы ещё не затрагивали.
Гиперссылки в скобках ведут на
страницы википедии, т.к. уместить
содержимое в один слайд попросту
невозможно.
Работу выполнил: Шурупов Егор
Класс: 7 «А»
Преподаватель: Бочарова Ольга Николаевна
English Русский Правила
Volvox — frwiki.
 wiki
wikiVolvox является родом из зеленых водорослей из вольвоксовых семей. Вольвокса представляют собой сферические колониисостоящие из одноклеточных организмовжгутиковых окруженных жильные желатиновых, окруженные поверхностных клетками и двужгутиковой объединенных цитоплазматических соединениями. Внутри колонии множество ооспор . Они состоят из одной клетки. Вольвоксы остаются на поверхности воды, таким образом способствуя фотосинтезу.
Колонии клеток делятся на:
- вегетативные или соматические клетки, мелкие и многочисленные
- Крупные, немногочисленные репродуктивные клетки, расположенные на периферии колонии.
Каждая клетка окружена слизистой оболочкой с несливающимися и многоугольными границами.
Во время эмбрионального развития между клетками существуют цитоплазматические связи.
Этот примитивный многоклеточный организм обитает в богатых кислородом водах. Морфологически он выглядит как пустая сфера.
Резюме
- 1 Переход от одноклеточности к многоклеточности?
- 2 Размножение
- 3 Список видов
- 4 Примечания и ссылки
- 5 Внешние ссылки
- 5.
 1 Видеосъемка
1 Видеосъемка
- 5.
Переход от одноклеточности к многоклеточности?
Согласно колониальной теории (1874 г.) Volvox вызывает переход от одноклеточности к многоклеточности. Вольвокс состоит из нескольких клеток, это многоклеточность (или многоклеточность).
Размножение
Распространение вольвокса может происходить половым путем или путем бесполого размножения .
Размножение: Колонии образуют мужские гаметы (сперматозоиды) и женские гаметы (оосферы), которые выбрасываются в воду. Когда сперматозоид оплодотворяет оосферу , образуется диплоидная зигота, которая превращается в новую колонию. Репродуктивный цикл вольвокса гаплофазный, анизогамный .
Бесполое размножение: материнская колония может делиться и образовывать дочерние колонии внутри сферы; когда материнская колония умирает и разлагается, дочерние колонии освобождаются.
Список видов
По данным AlgaeBase (30 апреля 2013 г. ) :
) :
- Volvox africanus GSWest
- Вольвокс амбоенсис MFRich & MAPocock
- Вольвокс золотистый Эренберг
- Volvox barberi WRShaw
- Вольвокс капенсис MFRich & MAPocock
- Volvox carteri F. Stein, геном которого секвенировали в 2010 г.
- Вольвокс хаос Линней
- Диссипатрикс Volvox (WRShaw) Printz
- Вольвокс удобрения Наял
- Volvox gigas MAPocock
- Volvox globator Linnaeus ( типовой вид )
- Volvox Merrillii WRShaw
- Volvox migulae (WRShaw) Принц
- Volvox obversus (WRShaw) Printz
- Volvox ovalis Pocock ex Nozaki & AWColeman
- Перглобатор Volvox JHPowers
- Вольвокс пилула OFMüller
- Вольвокс пококии RCStarr
- Вольвокс полихламида Коршикова
- Volvox powersii (WRShaw) Printz
- Volvox rousseletii GSWest
- Volvox spermatosphaera Powers
- Вольвокс тертиус Мейер
По данным ITIS (30 апреля 2013 г.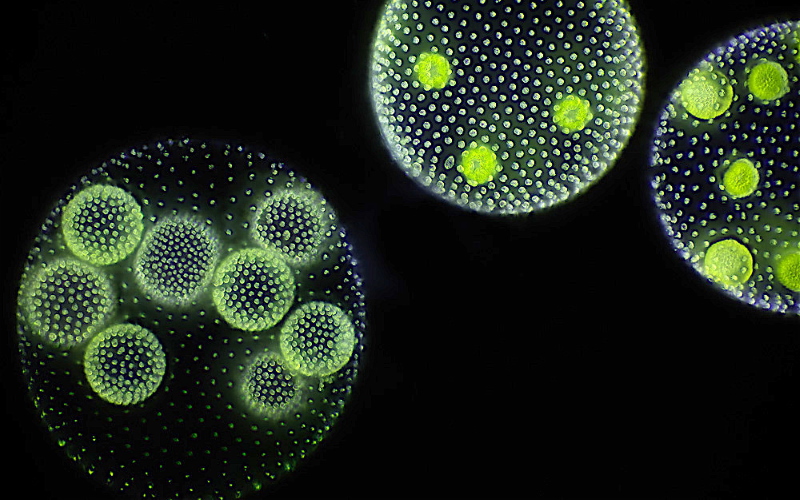 ) :
) :
- Вольвокс золотистый Эренберг
- Вольвокс глобатор Линней
- Перглобатор Volvox
- Volvox spermatosphaera Powers
- Вольвокс тертиус А. Мейер
- Volvox weismannia Powers
Согласно Всемирному регистру морских видов (30 апреля 2013 г.) :
- Volvox aureus Эренберг, 1832 г.
- Вольвокса хаоса Линней, 1758
- Volvox диссипатрикс (WRShaw) Printz, 1927 г.
- Volvox ferrisii Н. Исака, Мацузаки и Нозаки, 2012 г.
- Вольвокс глобатор Линней, 1758 г.
- Volvox kirkiorum Nozaki, H. Kawai-Toyooka & N.Isaka, 2012 г.
- Volvox ovalis Pocock ex Nozaki & AWColeman, 2011 г.
- Volvox powersii (WRShaw) Printz, 1927 год
- Volvox spermatosphaera Powers, 1908 год.
- Вольвокс тертиус Мейер, 1896 г.
Примечания и ссылки
- ↑ MD и GM Guiry, AlgaeBase , Национальный университет Ирландии, Голуэй, 2012-2020 гг.
 Всемирный регистр морских видов, по состоянию на 30 апреля 2013 г.
Всемирный регистр морских видов, по состоянию на 30 апреля 2013 г.
Внешние ссылки
- (ru) Водоросли Ссылка на базу : род Volvox Linnaeus, 1758 (см.)
- (fr + en) Ссылка ITIS : Volvox Linnaeus, 1758 (+ английская версия ) (консультации по)
- (ru) Ссылка NCBI : Volvox ( включая таксоны ) (консультации по)
- (о) Вперёд ссылка : Вольвокс Linnaeus, 1758 (консультации по)
- (ru) Справочный всемирный регистр морских видов : таксон Volvox Linnaeus, 1758 ( + список видов ) (см.)
 «>(fr + en) Ссылка на EOL : Volvox (консультации по)
«>(fr + en) Ссылка на EOL : Volvox (консультации по)
Видеосъемка
- Вольвокс, колониальные зеленые водоросли (видео 1) , Youtube, DelarueBioMedia
- Вольвокс, колониальные зеленые водоросли (видео 2) , Youtube, DelarueBioMedia
- Volvox, цитоплазматические мостики , Youtube, DelarueBioMedia
- Вольвокс, бесполое размножение , Youtube, DelarueBioMedia
- Volvox (микроскопия на черном фоне), Youtube, GRENDEL 1997 さ ん の チ ャ ン ネ ル
<img src=»//fr.wikipedia.org/wiki/Special:CentralAutoLogin/start?type=1×1″ alt=»» title=»»>
Жизненный цикл Volvox | Chlorophyta
РЕКЛАМА:
В этой статье речь пойдет о бесполом и половом способах размножения, которые встречаются в жизненном цикле вольвокса.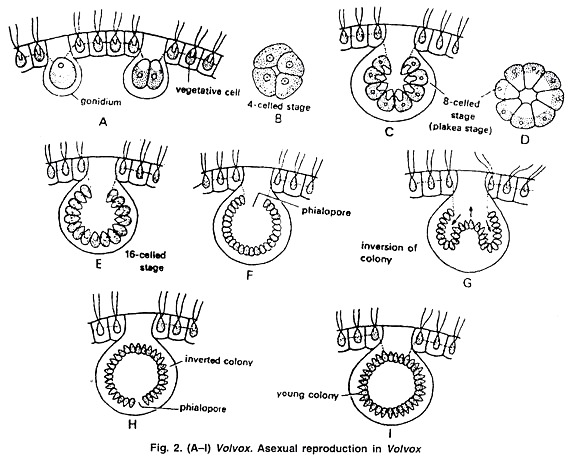
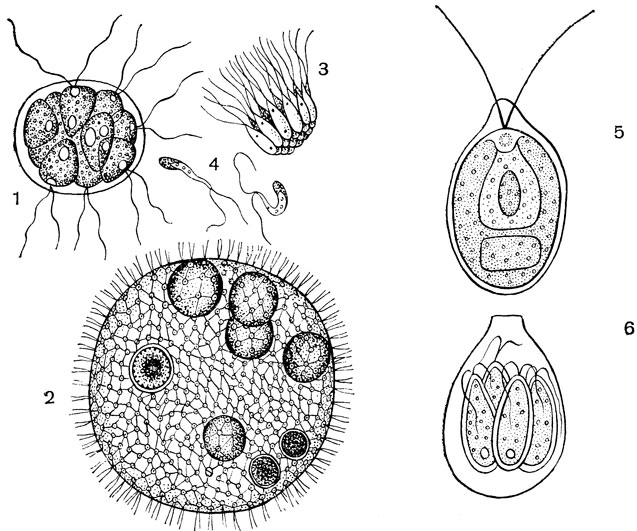
При бесполом размножении некоторые клетки задней части колонии становятся репродуктивными. Эти клетки увеличиваются до десяти раз, округляются и теряют жгутики. Эти клетки называются гонидиями (Sing, gonidium) (рис. 2 А). Гонидии теряют глазное пятно. Количество пиреноидов увеличивается.
РЕКЛАМА:
Гонидии смещаются внутрь колонии. Первое деление гонидия идет продольно плоскости ценобия и образует 2 клетки (рис. 2 А).
Второе деление также продольное и под прямым углом к первому, образуя 4 клетки (рис. 2 Б). При третьем продольном делении все четыре клетки делятся, образуя 8 клеток, из которых 4 клетки центральные и 4 периферические. Эти 8 клеток расположены в виде изогнутой пластинчатой структуры и называются стадией plakea (рис. 2 C, D). Каждая из этих 8 клеток делится продольным делением, образуя 16 клеток, расположенных в виде полого шара (рис. 2 Д).
Сфера открыта снаружи в виде небольшого отверстия, называемого фиалопорой (рис.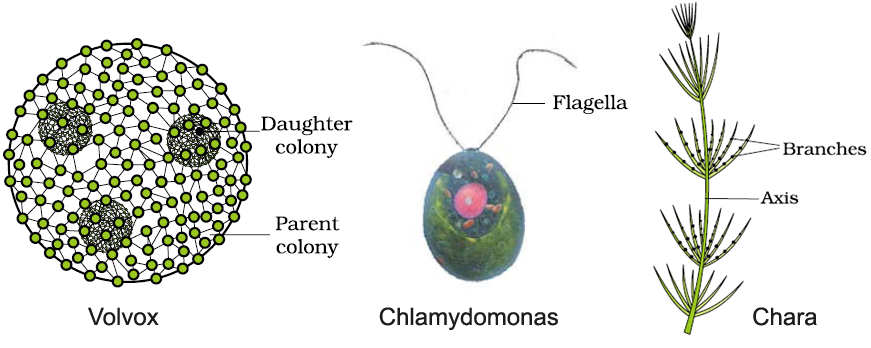 2 F). Клетки на этой стадии продолжают делиться до тех пор, пока число клеток не достигнет характеристики данного вида. Клетки на этой стадии голые и находятся в тесном контакте друг с другом. Заостренный передний конец клеток направлен внутрь.
2 F). Клетки на этой стадии продолжают делиться до тех пор, пока число клеток не достигнет характеристики данного вида. Клетки на этой стадии голые и находятся в тесном контакте друг с другом. Заостренный передний конец клеток направлен внутрь.
Следующий шаг называется инверсией колонии (рис. 2G-H). Поскольку клетки становятся противоположными по направлению, их передний заостренный конец должен быть обращен к периферии колонии.
РЕКЛАМА:
Инверсия колонии начинается с образования перетяжки напротив фиалопора. Клетки заднего конца вместе с перетяжкой вдавливаются внутрь шара, пока вся структура не выйдет из фиалопора. После инверсии передний заостренный конец клетки обращен к периферии.
Фиалопора закрывается и образует переднюю часть колонии. После инверсии клетки образуют клеточную стенку, жгутики и глазное пятно. Клетки разделяются за счет образования студенистой оболочки вокруг каждой клетки. Эта вновь развившаяся колония называется дочерней колонией (рис.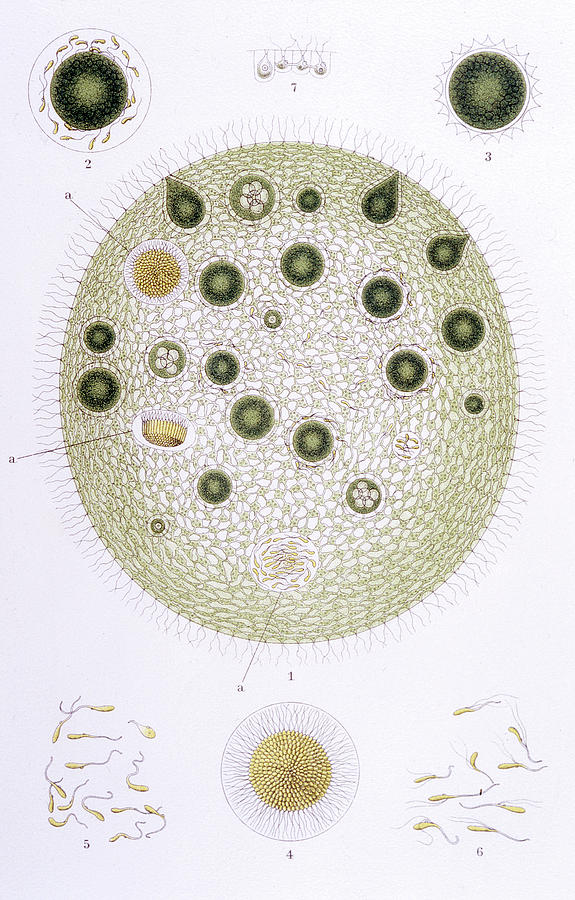 2I).
2I).
Дочерние колонии сначала остаются прикрепленными к желатинизированной стенке родительской колонии, а затем становятся свободными в студенистом матриксе родительской колонии. Дочерние колонии выделяются в воду после распада родительской колонии или через поры. Иногда следующее поколение дочерних колоний развивается, в то время как колонии все еще прикреплены к более ранней родительской колонии.
2. Половое размножение :Половое размножение Volvox оогамное. Некоторые виды Volvox, например V. globator, однодомны или гомоталличны (рис. 3), т. е. антеридии и оогонии развиваются в одной колонии. Другие виды Volvox, например, V. rousseletii, являются раздельнополыми или гетероталличными, т. е. антеридии и оогонии развиваются в разных колониях.
Однодомные виды обычно протандрические, т. е. антеридии созревают раньше оогоний, но некоторые виды являются протогинными, т. е. оогонии развиваются раньше антеридий. V aureus в основном раздельнополые, но иногда могут быть и однодомными.
Репродуктивные клетки в основном дифференцируются в задней части колонии. Эти клетки увеличиваются, теряют жгутики и называются гаметангиями. Мужские половые клетки называются антеридиями или андрогонидиями, а женские половые клетки называются оогониями или гиногонидами.
Развитие антеридия:РЕКЛАМА:
Развитие антеридия начинается с образования антеридиальной начальной или андрогонидной клетки преимущественно в задней части колонии. Начальные клетки увеличиваются, теряют жгутики, протоплазма уплотняется, а ядро становится больше. Антеридиальная инициаль смещается внутрь в сторону полости и остается связанной с другими вегетативными клетками через цитоплазматические тяжи.
Протопласт антеридиальной инициали делится продольно с образованием 16-512 удлиненных клеток. Клетки остаются пластинчатой структуры или располагаются в виде полой сферы. Инверсия клеток также имеет место, как и при бесполом размножении. Каждая клетка дифференцируется в антерозоиды или сперматозоиды (рис. 3, 4).
3, 4).
Антерозоид имеет веретенообразную, удлиненную, двужгутиковую структуру, содержащую две сократительные вакуоли, ядро, чашевидный хлоропласт, пиреноид и глазное пятно. Он бледно-желтого или зеленого цвета. Антерозоиды выделяются поодиночке или иногда группами.
РЕКЛАМА:
Развитие оогониума :
Оогонии также дифференцируются преимущественно в задней части колонии. Оогониальные инициали увеличиваются, ядро увеличивается, протопласт уплотняется, жгутики исчезают, глазное пятно исчезает, появляется много пиреноидов. Зрелая оосфера или яйцеклетка имеет круглую или колбовидную структуру. Яйцо имеет одноядерное строение, клюв колбовидного оогония выполняет функцию рецептивного пятна (рис. 5 А, Б).
Оплодотворение в Volvox:
РЕКЛАМА:
После освобождения от антеридия антерозоиды свободно плавают по поверхности воды. Благодаря хемотаксическому ответу антерозоиды достигают оогоний.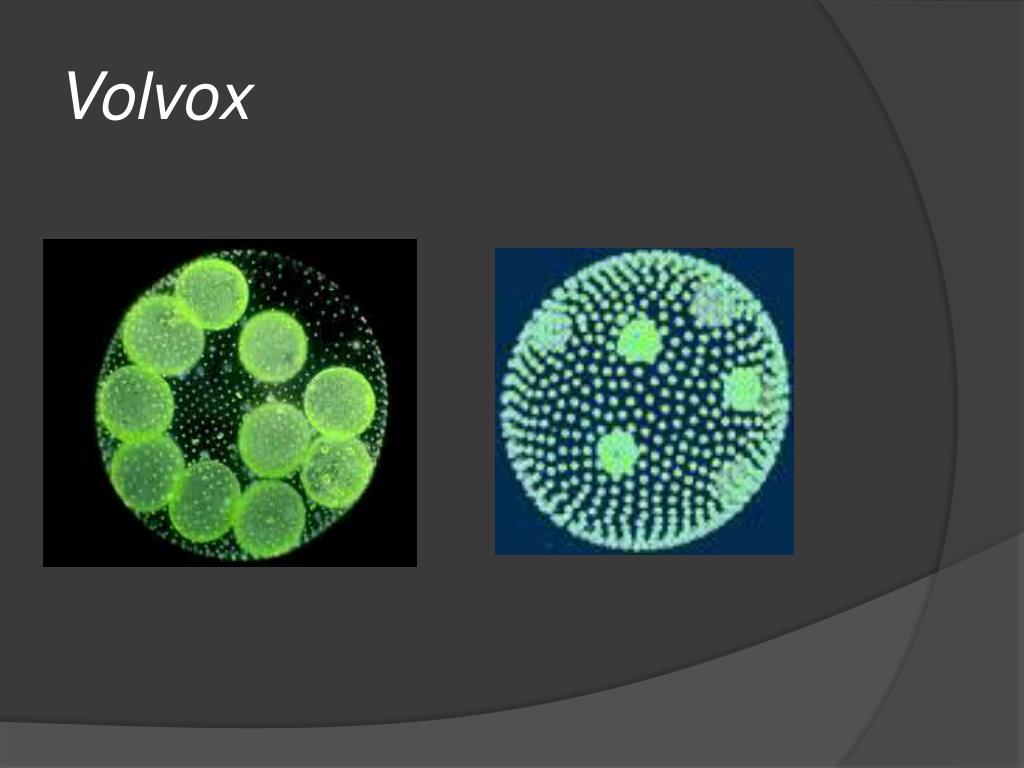
Некоторые антерозоиды проникают в каждый оогоний. Через рецептивное пятно внутрь оогония проникает только один антерозоид. После этого происходит плазмогейн, т. е. слияние мужской и женской цитоплазмы, и кариогамия, т. е. слияние мужского и женского ядер. Это приводит к образованию диплоидной зиготы (рис. 5 D).
Диплоидная зигота секретирует трехслойную толстую стенку. Слои стенки экзоспоровый, мезоспоровый и эндоспоровый (рис. 6 А, Б). Наружная экзоспора толстая. Он может быть гладким, например, у V. aureus (рис. 6А), или колючим, например, у V. globator (рис. 6В).
Мезоспоры и эндоспоры тонкие и гладкие. Стенки ядра содержат пигмент гематохром, придающий зиготе красный цвет. Зиготы высвобождаются при распаде родительской колонии. Затем зиготы проходят период покоя.
РЕКЛАМА:
Прорастание зиготы :
Спящая зигота прорастает при наступлении благоприятных климатических условий. Диплоидное ядро зиготы претерпевает мейотическое деление с образованием четырех гаплоидных клеток.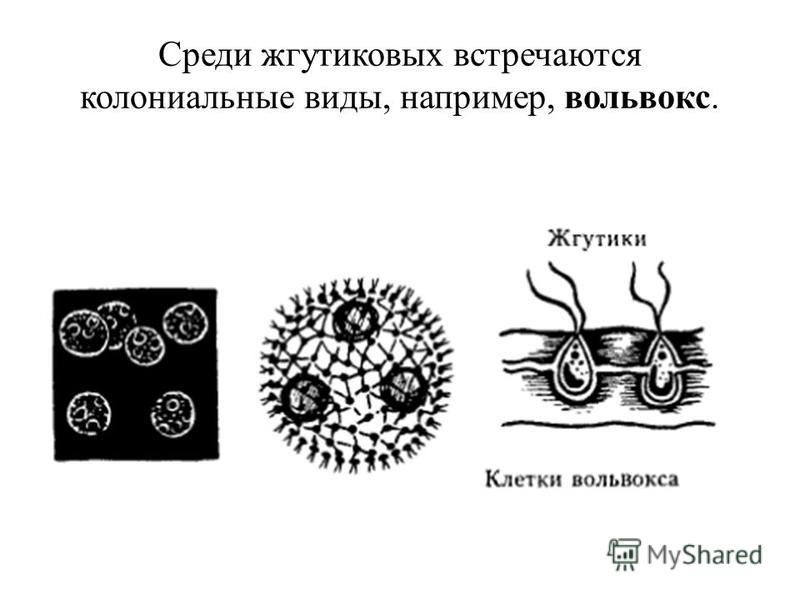 Два внешних слоя зиготы лопаются, а внутренний слой выходит наружу в виде пузырька. Четыре гаплоидные клетки мигрируют вместе с везикулой (рис. 6 C, D). Развитие новой колонии из зиготы различается у разных видов Volvox.
Два внешних слоя зиготы лопаются, а внутренний слой выходит наружу в виде пузырька. Четыре гаплоидные клетки мигрируют вместе с везикулой (рис. 6 C, D). Развитие новой колонии из зиготы различается у разных видов Volvox.
У V. aureus и V. minor протоплазма зиготы многократно делится до тех пор, пока не будет достигнуто число клеток колонии, и образуется новая колония, как при бесполом размножении. У V. campensis протопласт зиготы делится на множество двужгутиковых зооспор.
Выживает только одна зооспора, а все остальные распадаются. Эта зооспора выходит из пузырька, при делении которого образуется множество клеток, образующих колонию. У V. rousseletii зигота образует одиночную двужгутиковую зооспору, протопласт зооспоры делится и образует колонию. Во всех методах клетки делятся и подвергаются инверсии с образованием зрелой колонии (рис. 6 E-H).
Volvox представляет собой гаплоидную (n) водоросль, гаплоидные гаметы оплодотворяются с образованием диплоидной зиготы (2n), которая делится путем мейоза с образованием гаплоидных клеток (n), которые созревают в гаплоидную колонию Volvox (рис.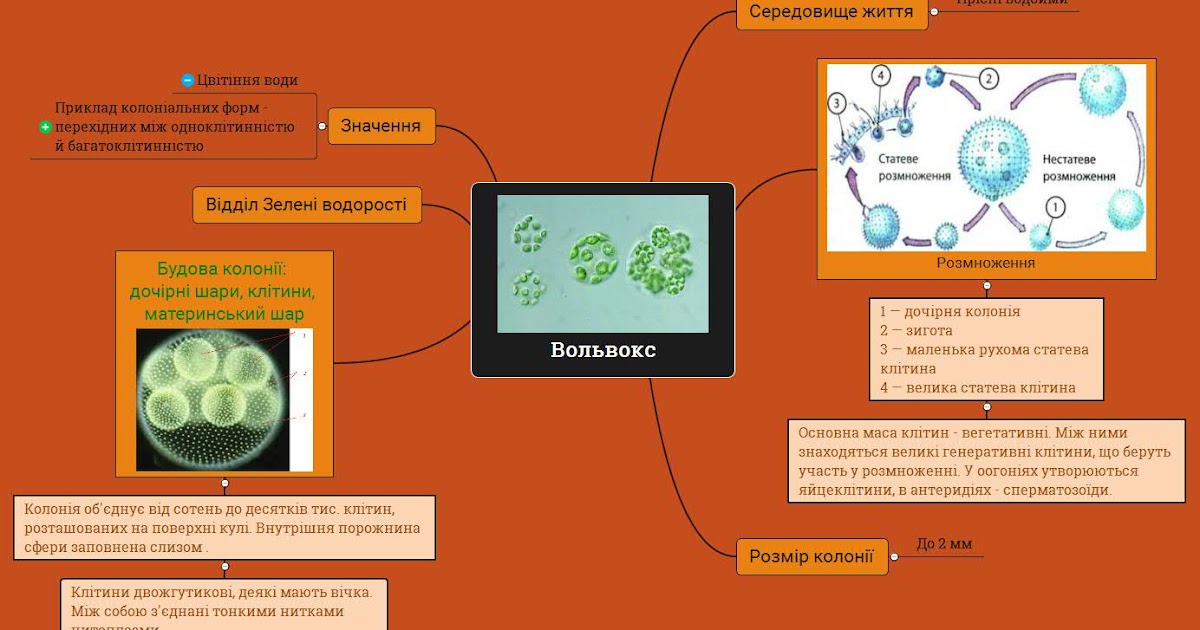 7, 8).
7, 8).
Тип: Volvox — возникновение, структура и размножение
Ботаника 3 комментария
Распространение
Volvox – пресноводные водоросли. Он делает воду зеленой. Появляется весной.
Общая структура Вегетативная структура
Volvox образует колонию. Колония вольвоксов называется coenobium. Имеет сферическую форму. Каждый двоеточие v имеет количество ячеек от 500 до 50 000. Внутренняя часть колонии заполнена слизью. Колония перемещается за счет коллективного действия жгутиков. Конец, направленный вперед, называется передним концом. Конец, направленный назад, называется задним концом колонии.
Все клетки колонии имеют сходную форму и строение. Клетки плотно упакованы. Они расположены так, что образуют многоугольную форму в колонии. Стенка зрелых клеток толстая и слизистая. Клеточная стенка имеет тонкую срединную пластинку.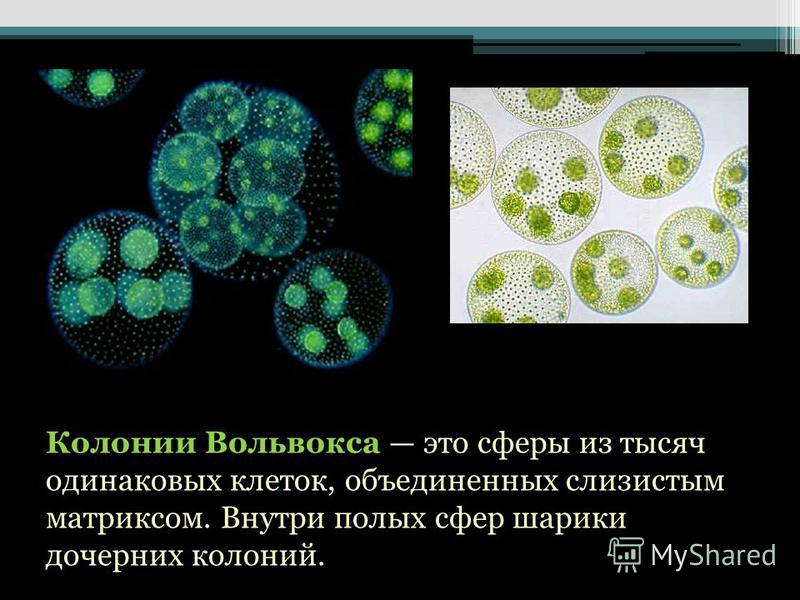 Их клетки имеют грушевидную форму. Эти клетки встроены в периферию Доута студенистой колонии 9.0003
Их клетки имеют грушевидную форму. Эти клетки встроены в периферию Доута студенистой колонии 9.0003
Рис: Колония Volvox
Матрица .
Клетки колонии соединены цитоплазматическими тяжами. Заостренный конец клеток направлен наружу.
Клетка структура
Структура особи аналогична Chlaymydomonas. Каждая клетка имеет два жгутика. Эти жгутики прикреплены к внешнему концу клеток. Каждая клетка имеет чашевидный хлоропласт. Он присутствует в более широком конце клетки. В хлоропластах находится один пиреноид. Около конца острия имеются две или более сократительных вакуолей. В центре клетки расположено одно ядро. Он связан со жгутиками нейромоторно. Присутствует одно глазное пятно. Клетки на передней поверхности колонии имеют хорошо развитые глазчатые пятна.
Размножение
Бесполое размножение
Новые колонии образуются в родительских колониях во время бесполого размножения. В бесполом размножении принимают участие лишь отдельные клетки заднего конца колонии. Размеры этих клеток увеличиваются. Их становится в десять раз больше. Эти клетки называются гонидиями . В гонидиях развиваются многочисленные пиреноиды. В каждой колонии 5—20 гонидий. Они производят дочернюю колонию внутри родительской колонии.
В бесполом размножении принимают участие лишь отдельные клетки заднего конца колонии. Размеры этих клеток увеличиваются. Их становится в десять раз больше. Эти клетки называются гонидиями . В гонидиях развиваются многочисленные пиреноиды. В каждой колонии 5—20 гонидий. Они производят дочернюю колонию внутри родительской колонии.
Часть стенки колонии Volvox
внучатые дочерние колонии.
4. Выход дочерней колонии: Иногда в стенке родительской колонии образуется пора. Таким образом, дочерняя колония выходит из родительских клеток и становится свободной. В некоторых случаях он остается в пределах родительской колонии. Родительская колония распадается и освобождает дочернюю колонию.
Генеративная клетка
Этапы бесполого размножения Voivox
Половое размножение
Половое размножение у вольвокса оогамное. Колония может быть гомоталличной или гетероталличной. Дочерние колонии, образующиеся бесполым путем, отсутствуют в колониях, образующихся половым путем.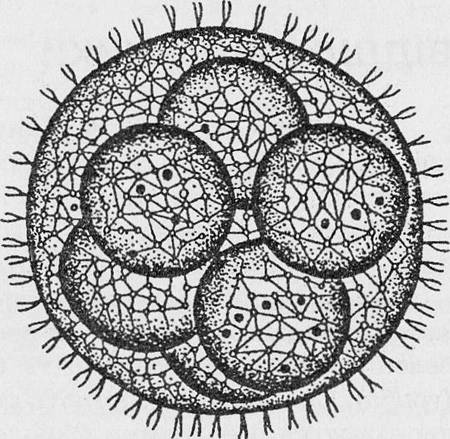 Репродуктивные клетки называются гантетангиями. Гаметангии располагаются в заднем полюсе колонии. Эти клетки крупнее вегетативных. Гаметангии отдергивают свои жгутики.
Репродуктивные клетки называются гантетангиями. Гаметангии располагаются в заднем полюсе колонии. Эти клетки крупнее вегетативных. Гаметангии отдергивают свои жгутики.
Антеридиум или мужские гаметангии
Мужские гаметангии называются антеридиями. Протопласт антеридия многократно делится. Он производит 16-128 клеток. Эти клетки называют антерозоидами. Антерозоиды удлиненно-веретенообразные.
Рис. Жизненный цикл вольвокса
У них развиты жгутики на заостренных концах. Антерозоиды имеют два типа циклов:
а) 128 или меньше 128: В этом случае все антерозоиды высвобождаются в виде массы клеток. Эта масса плывет и достигает оогониума. Затем отделяется антерозоид.
б) Более 128: В этом случае антерозоиды расположены в виде полой сферы внутри антеридия. Они подвергаются инверсии. Затем антерозоиды отделяются друг от друга. Тогда они освобождаются.
Они подвергаются инверсии. Затем антерозоиды отделяются друг от друга. Тогда они освобождаются.
Оогоний
Женскими половыми органами являются оогонии. Оогоний представляет собой увеличенную глобулярную клетку. Протопласт оогониума изменен на одиночный оосфера. Ooshere большой и округлой формы.
Оплодотворение: В стенке оогониума образуется отверстие. Через эту пору в оогоний проникают антерозоиды. Только один антерозоид соединяется с яйцеклеткой и образуется зигота. Зигота выделяет толстую стенку и становится ооспорой. Стенка ооспор трехслойная. Oospore производит большое количество гематохрома . Таким образом, его цвет становится оранжево-красным.
Рис. Жизненный цикл Volvo:
Рост ооспоры: Родительское тело распадается и высвобождает ооспору. Оосфера прорастает в благоприятных условиях. Первое деление – мейоз. Оосфера имеет три стенки: экзоспору, мезоспору и эндоспору. Обнажение и разрывы мезоспор. Эндоспора выпячивается в виде пузырька. В эти пузырьки мигрирует пропласт ооспоры. Этот протопласт развивает новую колонию.
Обнажение и разрывы мезоспор. Эндоспора выпячивается в виде пузырька. В эти пузырьки мигрирует пропласт ооспоры. Этот протопласт развивает новую колонию.
Sometinies, протопласт ооспоры образуют одну крупную бифиагельную зооспору. Он выходит из оогониума. Эта зооспора образует новую колонию путем бесполого размножения.
Похожие статьи:
Главная » Ботаника » Тип: Volvox – возникновение, структура и размножение
Об авторе регуляторный белок | Разработка
Пропустить пункт назначения
СТАТЬЯ В ЖУРНАЛЕ| 15 февраля 1999
М.М. Кирк,
К. Старк,
С.М. Миллер,
В. Мюллер,
Б.Э. Тайлон,
Х. Грубер,
Грубер,
Р. Шмитт,
Д.Л. Кирк
Информация об авторе и статье
Online Issn: 1477-9129
Print Issn: 0950-1991
https://doi.org/10.1242/dev.126.4.639
- Разделенный экран
- Просмотры
- Содержание статьи
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- Делиться
- MailTo
- Твиттер
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
Цитата
М. М. Кирк, К. Старк, С.М. Миллер, В. Мюллер, Б.Е. Тайлон, Х. Грубер, Р. Шмитт, Д.Л. Кирк; regA, ген Volvox, который играет центральную роль в дифференцировке зародышевой сомы, кодирует новый регуляторный белок. Развитие 15 февраля 1999 г.; 126 (4): 639–647. Дои: https://doi.org/10.1242/dev.126.4.639
М. Кирк, К. Старк, С.М. Миллер, В. Мюллер, Б.Е. Тайлон, Х. Грубер, Р. Шмитт, Д.Л. Кирк; regA, ген Volvox, который играет центральную роль в дифференцировке зародышевой сомы, кодирует новый регуляторный белок. Развитие 15 февраля 1999 г.; 126 (4): 639–647. Дои: https://doi.org/10.1242/dev.126.4.639
Скачать файл цитаты:
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- РефВоркс
- Бибтекс
Расширенный поиск
Volvox имеет два типа клеток: смертные соматические клетки и бессмертные зародышевые клетки. Здесь мы описываем мечение транспозонов, клонирование и характеристику regA, который играет центральную роль в качестве основного регуляторного гена в дифференцировке зародышевой сомы Volvox путем подавления репродуктивной активности в соматических клетках.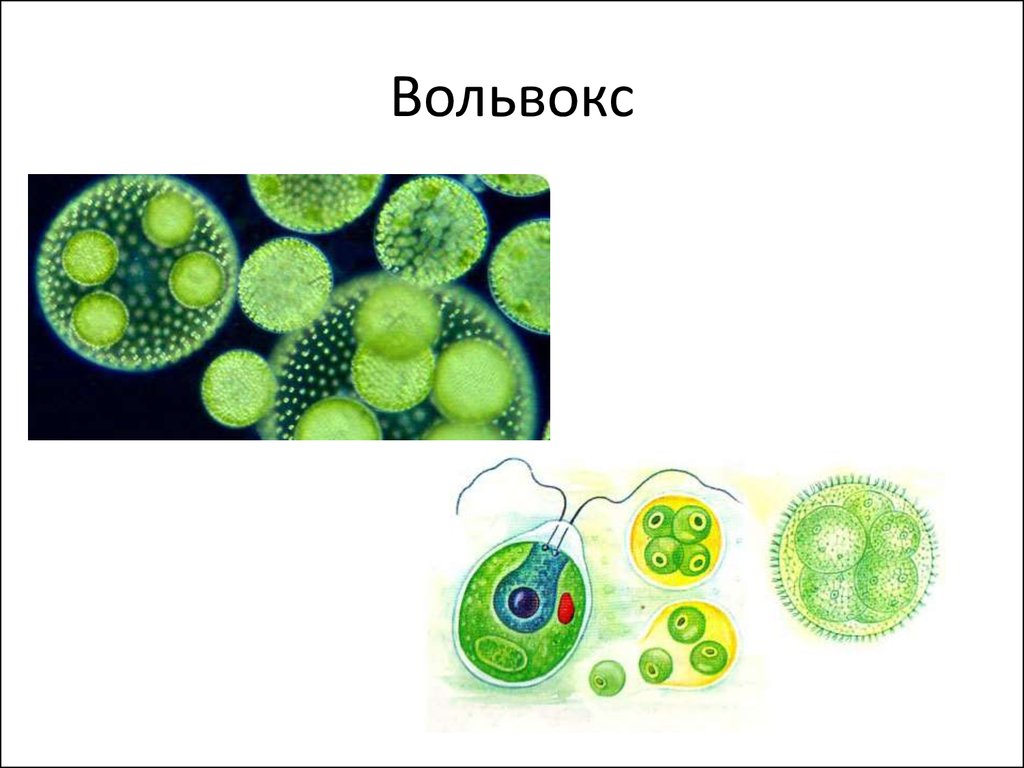 Единица транскрипции regA размером 12,5 т.п.н. генерирует мРНК из 6725 нуклеотидов, которая появляется в начале дифференцировки соматических клеток и кодирует белок RegA массой 111 кДа, который локализуется в ядре и содержит необычное количество аланина, глутамина и пролина. Это композиционная особенность, общая для функциональных доменов многих «активных» репрессоров. Эти данные согласуются с гипотезой о том, что RegA действует в соматических клетках, репрессируя транскрипцию генов, необходимых для роста и размножения, включая 13 генов, продукты которых необходимы для биогенеза хлоропластов.
Единица транскрипции regA размером 12,5 т.п.н. генерирует мРНК из 6725 нуклеотидов, которая появляется в начале дифференцировки соматических клеток и кодирует белок RegA массой 111 кДа, который локализуется в ядре и содержит необычное количество аланина, глутамина и пролина. Это композиционная особенность, общая для функциональных доменов многих «активных» репрессоров. Эти данные согласуются с гипотезой о том, что RegA действует в соматических клетках, репрессируя транскрипцию генов, необходимых для роста и размножения, включая 13 генов, продукты которых необходимы для биогенеза хлоропластов.
Adams
C. R.
,
STAMER
K. A.
,
Miller
J. K.
,
McNally
J. G.
. D. L.
(
1990
)
Особенности органелларного и ядерного наследования среди потомства двух географически изолированных штаммов Volvox carteri.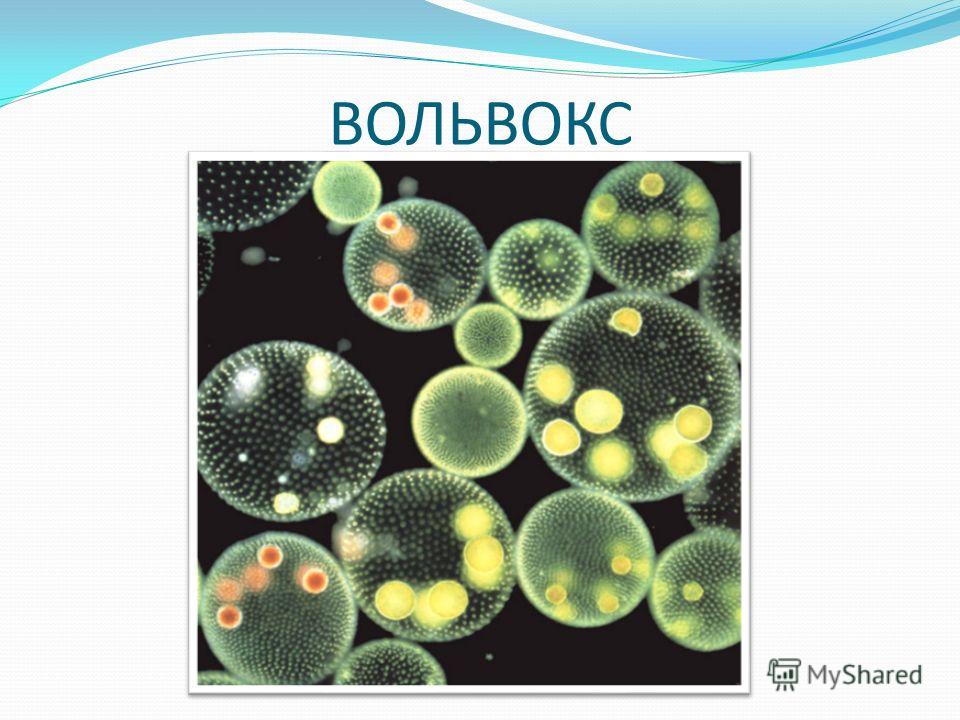
Курс. Жене
18
,
141
–
153
Altschul
S. F.
,
Gish
W.
,
Miller
W.
,
Myers
E. W.
,
Lipman
D. J.
(
1990
)
Базовый инструмент локального поиска выравнивания.
Дж. Мол. Биол
215
,
403
—
410
Atassi
M. Z.
,
Webster
R. G.
(
1983
)
Localization, Syntheses и Active of AntigenIn.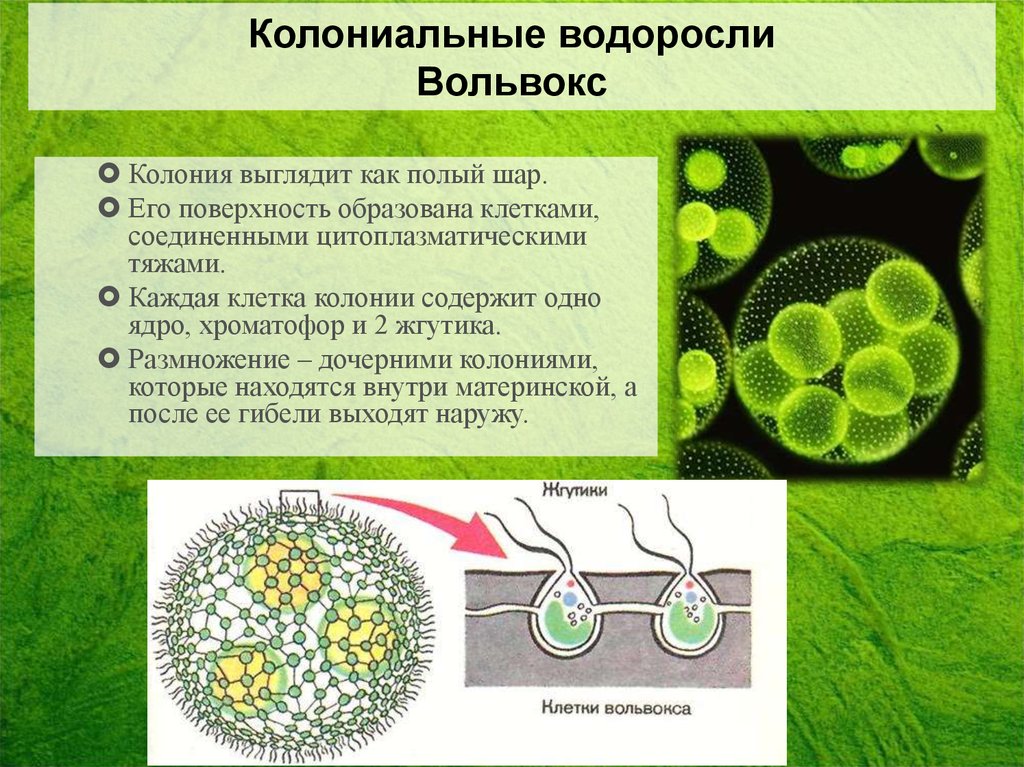
Проц. Натл. акад. науч. США
80
,
840
—
844
Брендель
,
Bucher
P.
,
NOURBHSH
.0003I.
,
Blaisdell
B. E.
,
KARLIN
S.
(
1992
)
Методы и альгитат для статистических последователей.
Проц. Натл. акад. науч. USA
89
,
2002
—
2006
Choi
G.
,
Przybylska
M.
,
Straklska
M.
,
M.
,
M.
,
М.0003
D.
(
1996
)
Три многочисленных транскрипта, характерных для зародышевой линии Volvox carteri, кодируют фотосинтетические белки.
Курс. Жене
30
,
347
–.
Cowell
I.G.
(
1994
)
Репрессия против активации в контроле транскрипции генов.
Trends Biochem. Наука
19
,
38
–
42
Fabry
S.
,
Jacobsen
A.
,
Huber
H.
,
Palme
K.
,
Schmitt
R.
(
1993
)
Структура, экспрессия и филогенетические взаимоотношения семейства генов ypt, кодирующих малые G-белки зеленой водоросли Volvox carteri.
Курс. Жене
24
,
229
–
240
Поле
J.
,
Nikawa 3.0 J.0 J
,
Broek
D.
,
MacDonald
B.
,
Rodgers
L.
,
Wilson
I. A.
,
Lerner
R.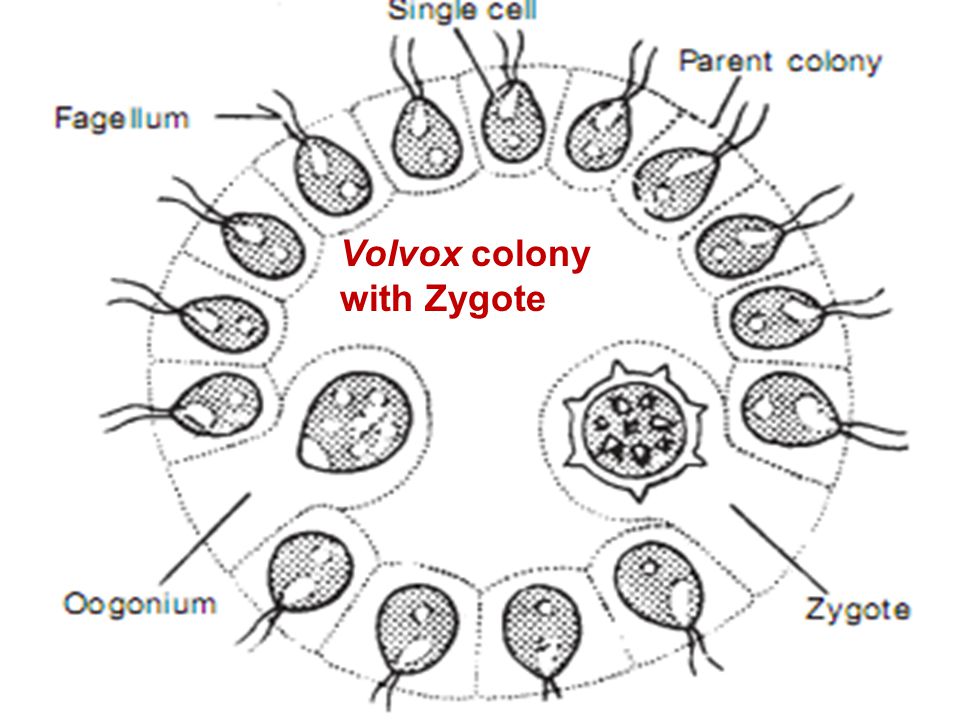
,
Виглер
М.
(
1988
)
Очистка RAS-чувствительного комплекса аденилциклазы из Saccharomyces cerevisiae с использованием метода добавления эпитопа.
Мол. Клетка. Биол
8
,
2159
–
2165
Гербер
Х.-П.
,
Seipel
K.
,
Georgiev
O.
,
Höfferer
M.
,
0002 Hug
M.
,
Rusconi
S.
,
Schaffner
W.
(
1994
)
Transcrection Activation Modude By Basepuremoplymorymerim.
Science
263
,
808
—
810
Gruber
H.
,
Kirzinger
S.
,
9000S.
,
9000 9000Schmitt
R.
(
1996
)
Экспрессия гена Volvox, кодирующего нитратредуктазу: Зависимая от мутаций активация скрытых сайтов сплайсинга и усиленная интронами экспрессия генов из кДНК.
Завод Мол. Biol
31
,
1
—
12
HAN
K.
,
Manley
J. L.
(
1993
6464)
(
136464)
(
13 4)0002 Транскрипционная репрессия дрозофилы с пропущенным четным белком: определение домена минимальной репрессии.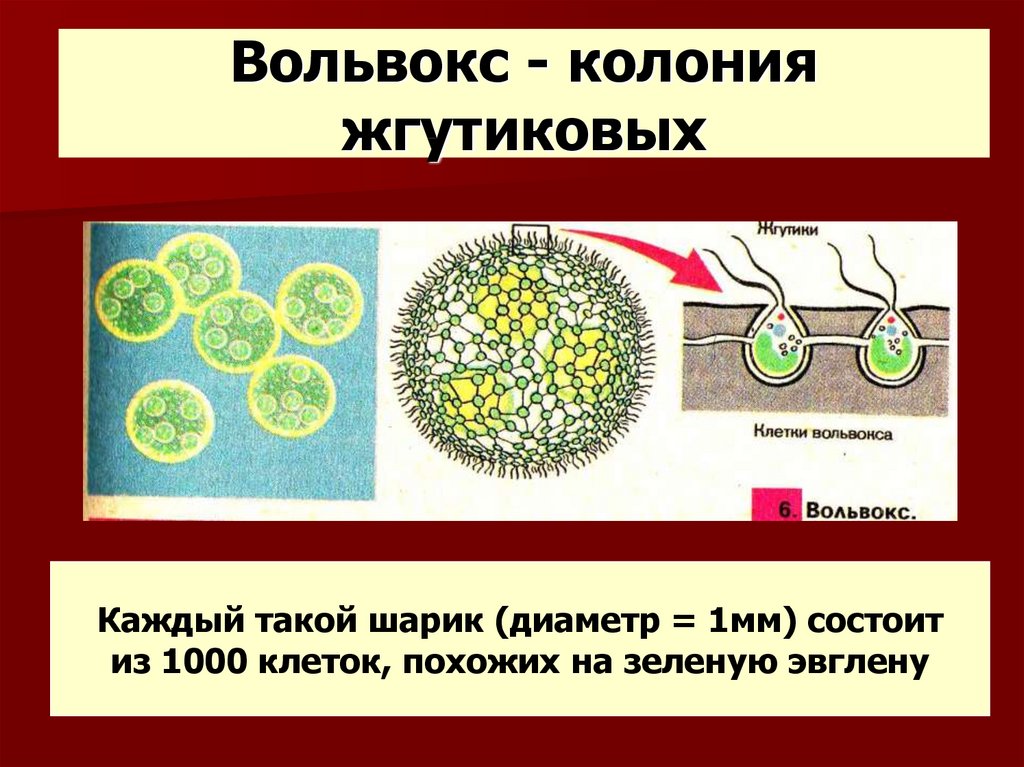
Genes Dev
7
,
491
–
503
Hanna-Rose
W.
,
Hansen
U.
(
1996
)
Active механизмы репрессии эукариотических репрессоров транскрипции.
Trends Genet
12
,
229
–
234
Harper
J. F.
,
Huson
K. S.
,
Kirk
D. L.
(
1987
)
Use of repetitive sequences to identify Полиморфизмы ДНК, связанные с regA, важным для развития локусом Volvox.
Гены Дев
1
,
573
–
584
Харпер
J. F.
F.
,
Маги
W.
(
1988
)
Организация и структура генов Volvox -тубулина.
Мол. Gen. Genet
213
,
315
–
324
Huber
H.
,
Beyser
K.
,
Fabry
S.
(
1996
)
Малые G-белки двух водорослей локализованы в экзоцитарных компартментах и жгутиках.
Завод Мол. Biol
31
,
279
—
293
Хапей
R. J.
,
Griffin
B. E.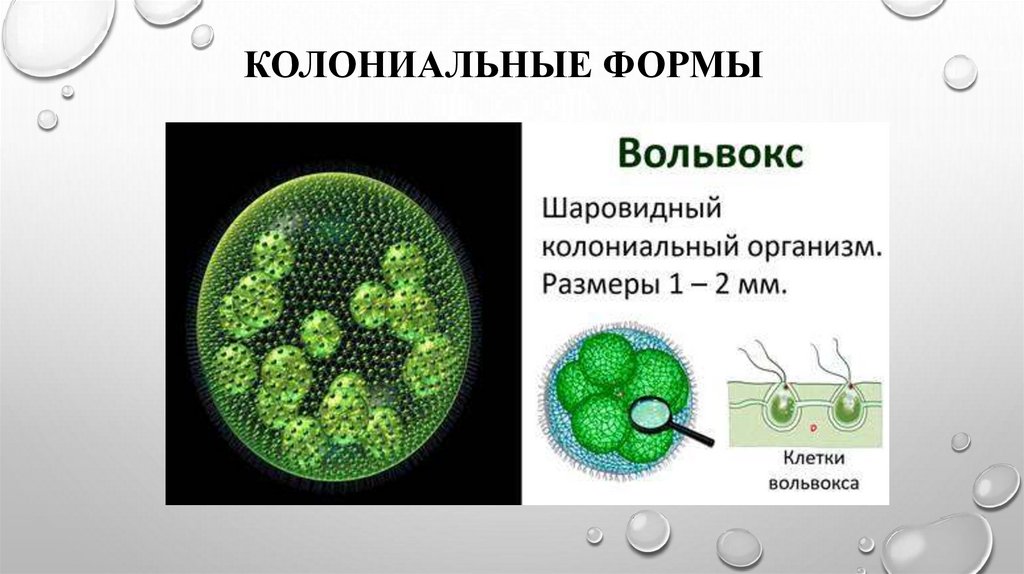
(
9999
).
Генетический контроль дифференцировки соматических клеток Volvox. Анализ мутантов соматических регенераторов.
Дев. Биол
72
,
226
–
235
Кирк
Д. Л.
,
Kirk
M. M.
(
1983
)
Синтетические модели белков во время бесполого жизненного цикла Volvox carteri.
Дев. Biol
96
,
493
–
506
Kirk
M. M.
,
Kirk
D. L.
(
1985
)
Translational regulation of protein synthesis, in response к свету на критическом этапе развития Volvox.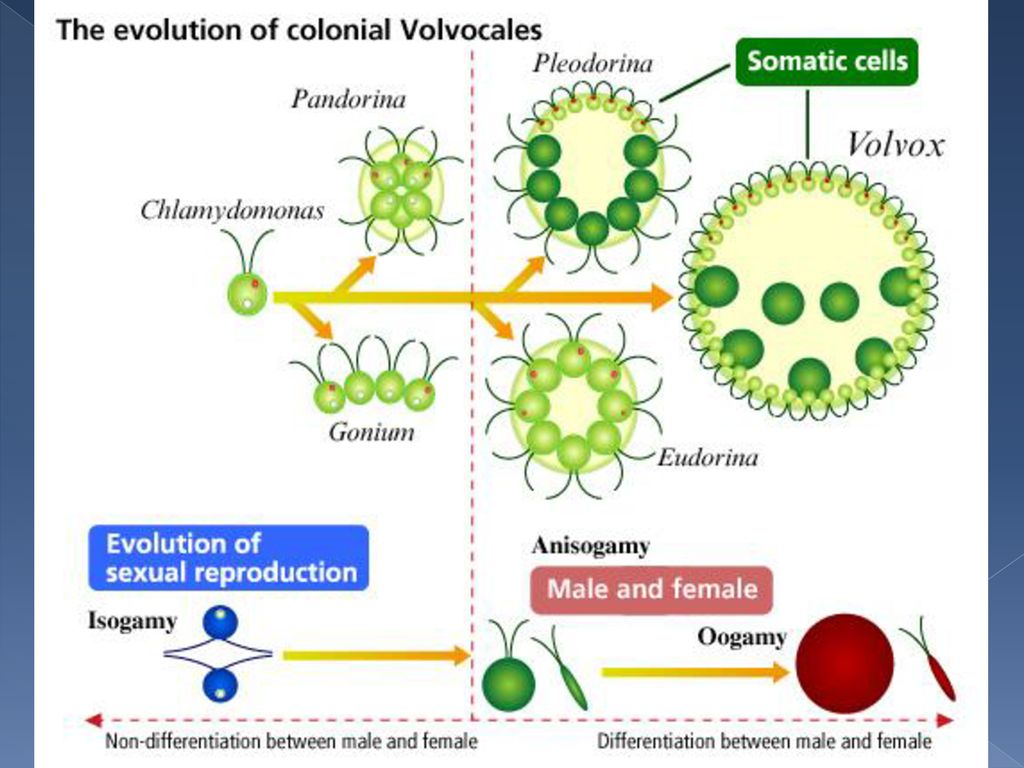
Cell
41
,
419
–
428
Kirk
D. L.
,
Baran
G. J.
,
Harper
J. F.
,
Huskey
R. J.
,
Huson
K. S.
,
Zagris
N.
(
1987
)
-специфическая стадия гипермациональности. .
Cell
48
,
11
–
24
Kropat
J.
,
von Gromoff
E. D.
,
Muller
F. W.
W.
,
Beck
C. F.
(
1995
)
Тепловой шок и световая активация гена HSP70 Chlamydomonas включают независимые регуляторные пути.
Мол. Генерал Жене
48
,
727
—
734
MADDEN
S. L.
,
D. M.
,
RAUSCHER
F… M.
,
F… M. M.
,
F…. M.
. Структурно-функциональный анализ репрессии транскрипции, опосредованной WT1, белком-супрессором опухоли Вильмса.
Онкоген
8
,
1713
–
1720
Maruyama
I. C.
C.
,
Rakow
T. L.
,
Maruyama
H. I.
(
1995
)
(
1995
)
(
1995
).
Нукл. Acids Res
23
,
3796
—
3797
Miller
S. M.
,
Schmitt
R.
,
0002 Kirk
DL
(
1993
)
Jordan, активный подвижный элемент Volvox, аналогичный транспозонам высших растений.
Plant Cell
5
,
1125
–
1138
Nakai
K.
,
Kanehisa
M.
(
1992
)
A knowledge base для предсказания сайтов локализации белков в эукариотических клетках.
Genomics
14
,
897
–
911
Pommerville
J. C.
,
Kochert
G.
(
1981
)
Changes in somatic cell structure во время старения Volvox carteri.
Евро. J. Cell Biol
24
,
236
–
243
Pommerville
J.C.
,
Kochert
G.
(
1982
)
Влияние старения на физиологию соматических клеток зеленой водоросли Volvox carteri.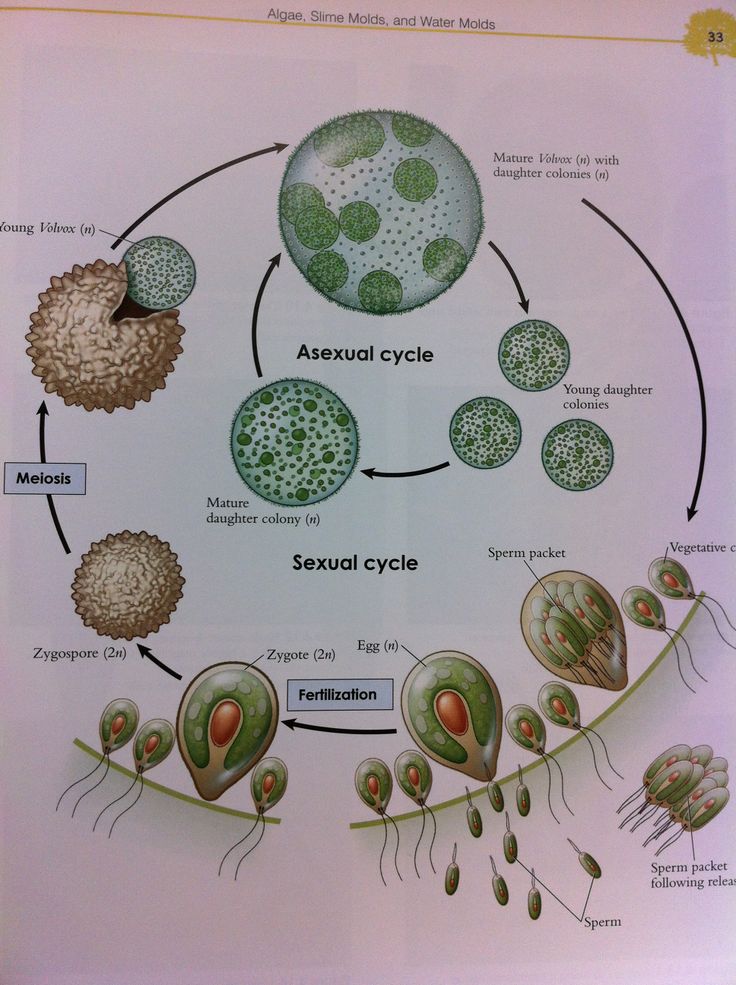
Расшир. Cell RES
140
,
39
—
45
ROST
B.
,
C.
(
13 .). структура с использованием профилей последовательности и нейронных сетей.
Проц. Натл. акад. науч. USA
90
,
7558
–
7562
Rost
B.
,
Sander
C.
(
1993
)
Prediction of protein secondary structure at точность лучше 70%.
J. Mo. Biol
232
,
584
–
599
Sanger
3 F. Nicklen S. , Coulson A. R. ( 1977 ) Проц. Натл. акад. науч. USA 74 , 5463 – 5467 Schiedlmeier B. , Schmitt R. , Muller W. , Kirk M. M. , Gruber H. , Маги W. , KIRK D. L. ( 1994 9000VER. Проц. Натл. акад. науч. США 91 , 5080 — 5084 Schmitt R. , Fabry S. , 0003 Кирк. DL ( 1992 ) В поисках молекулярного происхождения клеточной дифференцировки у Volvox и его родственников. Междунар. Rev. Cytol 139 , 189 – 265 Sheen J. Y. , Bogorad L. ( 1986 ) Differential expression of six light — сбор белков, связывающих хлорофилл a/b, в типах клеток листьев кукурузы. Проц. Натл. акад. науч. USA 83 , 7811 – 7815 Simpson J. , Schell J. , Van Montagu M. , Herrera -Estrella L. ( 1986 ) Индуцируемая светом и тканеспецифичная экспрессия гена LHCP гороха включает восходящий элемент, сочетающий свойства энхансера и сайленсера. Nature 323 , 551 — 553 Starr R. C. ( 1969 ). nagariensis Iyengar, штаммы HK 9 и 10. Arch. Протистенкд 111 , 204 – 222 Там Л.-Ш. , Кирк D. ( 1991 ) Идентификация генов, специфичных для типа клеток Volvox carteri, и характеристика их экспрессии во время бесполого жизненного цикла. Дев. Биол 145 , 51 – 66 Там Л.-В. , Kirk D.L. ( 1991 ) Программа клеточной дифференцировки у Volvox carteri, выявленная с помощью молекулярного анализа развития регенеративного/соматического мутанта. Развитие 112 , 571 – 580 Там Л.-Ш. , Stamer K. A. , Kirk D. 00003
00003

 L.
L.