Дыхание предка
На протяжении двух миллиардов лет на нашей планете обитали лишь бактерии и археи — организмы, лишенные ядра, то есть прокариоты. Затем клетки стали обзаводиться более сложными структурами: ядром, отделенным мембранами от цитоплазмы, и митохондриями. Этот качественный переход во внутренней организации клетки и позволил появиться эукариотам, к которым относятся множество одноклеточных и все многоклеточные организмы — включая авторов этого текста и его читателей. Шведские биологи в статье в Science утверждают: ради этого предки эукариот дважды подружились с бактериями и только со второго раза научились дышать кислородом. Алексей Дукат и Анна Кузнецова объясняют, откуда взялась эта гипотеза и какие с ней могут возникнуть сложности.
Как появились эукариоты
Эволюцию эукариот принято отсчитывать от FECA (first eukaryotic common ancestor), гипотетического первого общего предка всех эукариот. Это древнейший организм, который только начал приобретать характерные для эукариот признаки. Со временем он превратился в LECA (last eukaryotic common ancestor), последнего общего предка. После него эволюционные ветви эукариот разошлись — поэтому LECA должен обладать всеми основными признаками эукариотической клетки.
Это древнейший организм, который только начал приобретать характерные для эукариот признаки. Со временем он превратился в LECA (last eukaryotic common ancestor), последнего общего предка. После него эволюционные ветви эукариот разошлись — поэтому LECA должен обладать всеми основными признаками эукариотической клетки.
Что происходило с клетками по дороге от FECA к LECA, не совсем понятно. По имеющимся данным, FECA был очень близок к асгардархеям и не сильно от них отличался. А у LECA уже должны были присутствовать полноценное ядро и развитая система внутренних мембран. Получается, что между FECA и LECA лежит огромное эволюционное расстояние, и неясно, в каком порядке у них появлялись разные эукариотические признаки. Особенно много вопросов остается к митохондриям.
Митохондрии часто сравнивают с энергетическими станциями клетки. Внутри этих органелл происходит дыхание, то есть окисление органики кислородом, в результате чего выделяется энергия и синтезируется АТФ, универсальный переносчик энергии. Предполагается, что именно благодаря митохондриям у предковой клетки появились ресурсы на фагоцитоз — поглощение твердых частиц с помощью впячивания мембраны. Это критически важное для эукариот свойство: так можно не только захватывать внеклеточные молекулы, но и, например, образовывать транспортные пузырьки и сложные мембранные структуры. Разделение клетки на отсеки (компартменты) при помощи мембран решает множество проблем, и прокариоты на такое не способны — считается, что без митохондрий на такой процесс уходило бы слишком много энергии.
Предполагается, что именно благодаря митохондриям у предковой клетки появились ресурсы на фагоцитоз — поглощение твердых частиц с помощью впячивания мембраны. Это критически важное для эукариот свойство: так можно не только захватывать внеклеточные молекулы, но и, например, образовывать транспортные пузырьки и сложные мембранные структуры. Разделение клетки на отсеки (компартменты) при помощи мембран решает множество проблем, и прокариоты на такое не способны — считается, что без митохондрий на такой процесс уходило бы слишком много энергии.
Митохондрии сильно отличаются от других органелл: у них есть собственная ДНК и рибосомы, они могут делиться независимо от самой клетки. Еще они обладают двойной мембраной, причем внутренняя имеет сложную структуру со складками (кристами), для того чтобы увеличить площадь поверхности, на которой протекают биохимические реакции. Все это говорит о том, что митохондрии когда-то были отдельно живущими организмами — а именно α-протеобактериями.
Считается, что предок эукариот (архея) вступил в симбиоз с предком митохондрий (α-протеобактерией), в результате чего появился принципиально новый организм, давший начало полноценным эукариотам. Есть несколько гипотез о том, как именно произошел такой эндосимбиоз, но все они сходятся в одном: к моменту появления LECA бактерия должна была прочно поселиться в будущей эукариотической клетке. А значит, самые первые эукариоты должны были обладать полноценными митохондриями. Но некоторые ученые продолжают ставить эту идею под сомнение. В их числе Цезарь Аль Джевари (Caesar Al Jewari) и Сандра Балдауф (Sandra Baldauf) из Уппсальского университета: они предположили, что первые эукариоты могли выглядеть совсем по-другому.
Есть несколько гипотез о том, как именно произошел такой эндосимбиоз, но все они сходятся в одном: к моменту появления LECA бактерия должна была прочно поселиться в будущей эукариотической клетке. А значит, самые первые эукариоты должны были обладать полноценными митохондриями. Но некоторые ученые продолжают ставить эту идею под сомнение. В их числе Цезарь Аль Джевари (Caesar Al Jewari) и Сандра Балдауф (Sandra Baldauf) из Уппсальского университета: они предположили, что первые эукариоты могли выглядеть совсем по-другому.
Кто был первым
Современные эукариоты — разнообразная группа, систематика которой постоянно меняется. На данный момент выделяют три мегагруппы эукариот:
- Amorphea — гетеротрофы, которые умеют образовывать ложноножки. Сюда, к примеру, входят заднежгутиковые, к которым относятся грибы и животные,
- Diaphoretickes — среди них много фотосинтетиков, хотя в этой группе встречаются и организмы, которые никогда фотосинтезировать не умели.
 Сюда входят архепластиды, к которым относятся растения,
Сюда входят архепластиды, к которым относятся растения, - Excavata— одноклеточные жгутиконосцы. Среди них множество свободноживущих и симбиотических форм, а также некоторые паразиты человека, такие как лямблии и трихомонады. Именно на них обратили свое внимание шведские биологи.
Среди экскават есть такие, у которых митохондрий в привычном понимании нет. Зато есть кое-что похожее — гидрогеносомы. Гидрогеносомы точно так же окружены двойными мембранами и тоже выделяют углекислый газ и производят АТФ. Но в процессе их бескислородного дыхания попутно выделяется еще ацетат и молекулярный водород. По сути, функционально гидрогеносомы заменяют митохондрии в анаэробных (бескислородных) условиях.
Выяснить, откуда появились гидрогеносомы, довольно сложно — чаще всего они лишены генома, поскольку необходимые для их функционирования гены мигрировали в ядро клетки. Общепринятой является гипотеза, согласно которой гидрогеносомы это деградировавшие митохондрии, и в ее пользу есть немало аргументов. Однако у Аль Джевари и Балдауф возникли сомнения.
Однако у Аль Джевари и Балдауф возникли сомнения.
Они задались вопросом: какая из групп эукариот самая древняя? Чтобы на него ответить, потребовалось проанализировать гены архейного и бактериального
Поскольку все эукариоты — потомки археи, вступившей в симбиоз c бактерией, у них остались общие с прокариотами гены. Причем гены бактериального происхождения (euBacs) связаны с функционированием митохондрий, в то время как гены архейного происхождения (euArcs), как правило, участвуют в глобальных клеточных процессах, таких как репликация, транскрипция и трансляция
у современных эукариот.Ученые отобрали 456 белков, общих для архей и эукариот, затем из них выбрали наиболее близкие к эукариотам последовательности. Предпочтение отдавалось белкам, которые встречались во всех ветвях, происходящих от общего предка эукариот. Именно по тому, насколько эти белки отличаются от архейных, можно судить об эволюционном расстоянии между археями и различными ветвями эукариот. Например: ветвь Archaeplastida, к которой относятся большинство современных растений. У этой ветви есть белки, общие с археями, но многие из них отличаются по отдельным последовательностям — значит, ветвь Archaeplastida лежит далеко от архей на эволюционном дереве.
У этой ветви есть белки, общие с археями, но многие из них отличаются по отдельным последовательностям — значит, ветвь Archaeplastida лежит далеко от архей на эволюционном дереве.
Получилось филогенетическое древо, на котором ближе всего к первым ветвям эукариот лежали четыре анаэробные линии экскават. А если одни из самых древних эукариот жили в анаэробных условиях, то — предположили исследователи — их последнему общему предку LECA митохондрии были не нужны. Зато у него, вероятно, были гидрогеносомы — как у современных анаэробных экскават, и появились они независимо от митохондрий.
Это предположение идет вразрез с современными представлениями о происхождении эукариот. Общепринятая на данный момент гипотеза утверждает, что у LECA уже были полноценные митохондрии, которые появились в результате эндосимбиоза с α-протеобактерией. А потом они независимо деградировали до гидрогеносом у разных групп эукариот, населяющих анаэробные среды.
Шведские биологи рисуют совсем другую картину. По их мнению, первые эукариоты митохондрий не имели, а симбиозов на их эволюционном пути было два. Сначала произошел γ- или δ-протеобактериальный эндосимбиоз, давший начало гидрогеносомам. Потом некоторые эукариоты вступили в симбиоз с α-протеобактериями, давшими начало митохондриям. А гидрогеносомы большинство эукариот со временем утратили.
По их мнению, первые эукариоты митохондрий не имели, а симбиозов на их эволюционном пути было два. Сначала произошел γ- или δ-протеобактериальный эндосимбиоз, давший начало гидрогеносомам. Потом некоторые эукариоты вступили в симбиоз с α-протеобактериями, давшими начало митохондриям. А гидрогеносомы большинство эукариот со временем утратили.
Почему именно γ- или δ-протеобактерии? ↓
В геномах эукариот, помимо генов альфа-протеобактерий, встречаются также в куда меньшем количестве гены гамма- и дельта-протеобактерий. Авторы предполагают, что те или другие — результат переноса в ядро в ходе эндосимбиоза. Хотя это не единственная возможность: на самом деле эти гены могли попасть туда в результате горизонтального переноса генов.
Это возможно?
Авторы проделали большую работу, чтобы прийти к выводу о том, что экскаваты — одна из самых древних групп эукариот. Однако из этого вовсе не следует, что гидрогеносомы появились раньше митохондрий, а эндосимбиоз происходил дважды. Можно представить себе и другой вариант развития событий, согласно которому первые экскаваты все-таки были носителями митохондрий, а позже некоторые представители смогли адаптироваться к жизни без кислорода и обзавелись гидрогеносомами. Чтобы убедить научное сообщество в том, что верна именно их модель, шведским ученым придется отразить сразу несколько контраргументов.
Можно представить себе и другой вариант развития событий, согласно которому первые экскаваты все-таки были носителями митохондрий, а позже некоторые представители смогли адаптироваться к жизни без кислорода и обзавелись гидрогеносомами. Чтобы убедить научное сообщество в том, что верна именно их модель, шведским ученым придется отразить сразу несколько контраргументов.
Во-первых, два эндосимбиоза подряд менее вероятны, чем один. Это выглядит как усложнение уже имеющейся гипотезы, и кажется маловероятным, что такой сложный процесс, как эндосимбиоз, мог произойти дважды. Правда, в природе есть примеры вторичного и даже третичного (у некоторых красных водорослей) эндосимбиоза — это пластиды, предками которых тоже являются бактерии. Но чтобы доказать сложную последовательность симбиозов, хорошо бы найти следы промежуточных стадий в этом процессе. И пока не известно ни одного организма, который одновременно имел бы и гидрогеносомы, и митохондрии.
Во-вторых, уже накопилось немало подтверждений того, что гидрогеносомы произошли от митохондрий. Например, в 2010 году была найдена гидрогеносома с митохондриальными генами. Кроме того, в гидрогеносомах есть белки, уникальные для митохондрий. Не стоит забывать и о том, что гидрогеносомы присутствуют также у некоторых грибов, которые являются значительно более поздней группой эукариот. Чтобы доказать, что гидрогеносомы появились независимо, нужно будет предложить новое объяснение для всех этих находок.
Например, в 2010 году была найдена гидрогеносома с митохондриальными генами. Кроме того, в гидрогеносомах есть белки, уникальные для митохондрий. Не стоит забывать и о том, что гидрогеносомы присутствуют также у некоторых грибов, которые являются значительно более поздней группой эукариот. Чтобы доказать, что гидрогеносомы появились независимо, нужно будет предложить новое объяснение для всех этих находок.
Наконец, придется что-то сделать с более обоснованной гипотезой
Это название появилось в противовес гипотезе фагоцитоза, которая рисует процесс эндосимбиоза так: будущая эукариотическая клетка поглотила протеобактерию с помощью фагоцитоза, и та превратилась в ее митохондрию. Гипотеза inside-out предполагает, что архейная клетка, наоборот, постепенно обрастала мембраной вокруг бактерии
, которая предполагает, что митохондрии помогли будущим эукариотам не только освоить фагоцитоз, но и сформировать само ядро. Согласно этой гипотезе, архея постепенно окружала предка митохондрии своими выростами, а затем образовала вокруг них целую систему из мембран. Впоследствии эта система стала мембранными органеллами, а первичная мембрана археи — оболочкой ядра.
Впоследствии эта система стала мембранными органеллами, а первичная мембрана археи — оболочкой ядра.У гипотезы inside-out есть и косвенные экспериментальные подтверждения. Недавние наблюдения за современными археями позволили заметить аналогичные выросты, которые тесно прилегают к бактериальным клеткам. Если же считать, что в момент образования ядра никаких митохондрий в предковой клетке еще не появилось, а были только гидрогеносомы, то придется придумать альтернативную гипотезу — или же объяснить, откуда клетка без митохондрий получила достаточно энергии, чтобы отрастить так много мембран.
В самой статье Аль Джевари и Балдауф таких объяснений не приводят. А значит, им еще придется подтвердить свою точку зрения. Если окажется, что она верна, это прольет свет на то, как выглядел и в каких условиях жил LECA и в конечном итоге позволит узнать что-то новое о нашем общем предке. Но если это не так и гидрогеносомы — это действительно деградировавшие митохондрии, то древние экскаваты уже не помогут нам дорисовать образ LECA полностью. И останутся попросту любопытной, но побочной ветвью эволюции эукариот.
И останутся попросту любопытной, но побочной ветвью эволюции эукариот.
Биология Подцарство Одноклеточные. Класс Жгутиконосцы
Материалы к уроку
Конспект урока
Подцарство Одноклеточные. Класс Жгутиконосцы
|
Сегодня мы совершим путешествие в царство животных, а более точный адрес нашего путешествия вы узнаете чуть позже. Рассмотрите рисунки и скажите, что объединяет представленных на рисунке животных?
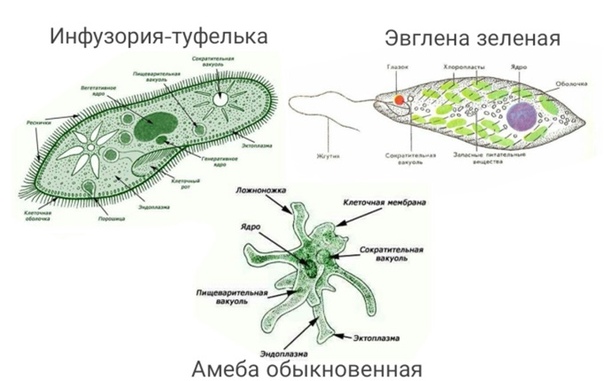
Все животные относятся к Простейшим, их организм представлен одной клеткой. |
|
Представленные животные имеют не только сходство, но и различия, например в способе передвижения. Вспомним способы передвижения Амёба протей передвигается с помощью ложноножек или псевдоподий.
Инфузория-туфелька передвигается с помощью ресничек, а эвглена зелёная — благодаря работе жгутика. Сегодня мы познакомимся с особенностями представителей этого класса Простейших.
|
|
Одним из представителей класса Жгутиконосцев является эвглена зелёная, обитатель водной среды — загрязненных прудов и луж. Жгутики, с помощью которых передвигается эвглена зелёная, представляют собой выросты цитоплазмы клетки. Каждый жгутик состоит из микротрубочек. Всего таких групп микротрубочек – 9, а в каждой из групп по 3 микротрубочки. Посмотрите на схему строения жгутика.
Основание жгутика переходит в базальное тельце.
Жгутик расположен на переднем конце тела эвглены.
|
|
У эвглены зелёной веретеновидная форма тела. Это постоянная форма тела, в отличие от изученной нами ранее амёбы протей, тело покрыто плотной оболочкой – пелликулой. |
|
Давайте посмотрим на рисунок эвглены зелёной. Какие из органоидов имеются как у эвглены, так и у амёбы, а какие органоиды имеются у эвглены и отсутствуют у амёбы? У амёбы и эвглены имеется ядро, мембрана клетки, сократительная вакуоль и пищеварительная вакуоль. У эвглены в клетке имеются: клеточный рот, жгутик, светочувствительный глазок, хлоропласты и пелликула. |
|
Какую же функцию в клетке простейших выполняют вакуоли? Сократительная вакуоль удаляет жидкие продукты обмена веществ и излишки воды. Пищеварительная вакуоль участвует в переваривании пищи. Непереваренные остатки пищи удаляются у эвглены зелёной у заднего конца тела через поверхность клетки. Способов питания у эвглены два: гетеротрофное и автотрофное. Поэтому по способу питания эвглену зелёную относят к миксотрофам. Подумайте, есть ли ещё в природе организмы, которые также можно отнести к миксотрофам? К миксотрофам можно отнести такие растения, как венерина мухоловка или росянка. Эти растения питаются и животной пищей, и автотрофно, получая органические вещества в процессе фотосинтеза. У эвглены в автотрофном питании участвуют хлоропласты. Автотрофное питание осуществляется у эвглены на свету. А освещённые места водоёма эвглена обнаруживает с помощью светочувствительного глазка – стигмы. В темноте эвглена питается гетеротрофно, то есть готовыми органическими веществами.
|
|
Сначала пища — растворенные в воде органические вещества — попадает в клеточный рот, который представляет собой углубление на переднем конце тела. Клеточный рот переходит в глотку. Из глотки пища попадает в пищеварительную вакуоль. |
|
Эвглена зелёная относится к аэробным организмам и использует для дыхания растворённый в воде кислород, который поглощает через поверхность тела. |
|
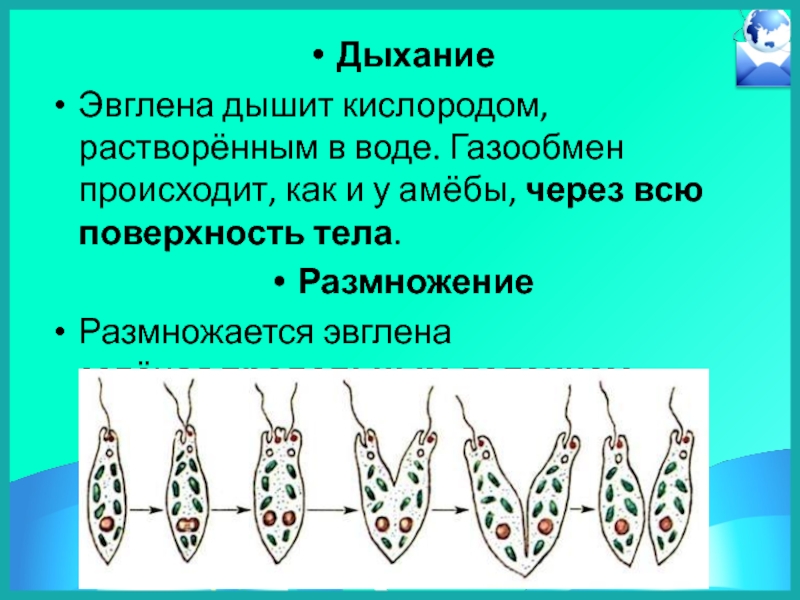
Ядро клетки эвглены управляет процессами жизнедеятельности, в том числе размножения. Для эвглены характерно бесполое размножение – деление клетки продольно. Процесс деления начинается с процесса деления ядра. За делением ядра следует деление цитоплазмы.
Среди представителей класса Жгутиконосцы имеются такие, которые размножаются даже половым способом. Это явление характерно для колониальных жгутиковых, которые образованы группой клеток.
|
|
К колониальным жгутиковым относятся вольвокс, пандорина, гониум. В процессе бесполого размножения участвует небольшое количество клеток и в результате образуется дочерняя колония. В половом размножении вольвокса участвуют разные типы клеток — женские и мужские. Эта особенность сближает колониальные жгутиковые с многоклеточными животными.
|
|
Жгутиконосцы — интересный и многообразный класс, насчитывающий более 7 тысяч видов. Среди них встречаются паразитические формы, как например, трипаносома. Жгутик трипаносомы начинается не от передней части тела, а от базального тельца, расположенного в задней трети тела. В клетках трипаносом отсутствует сократительная вакуоль. Паразитируют трипаносомы в крови и спинномозговой жидкости позвоночных животных.
|
|
Другое заболевание, кожное — лейшманиоз (пендинская язва) – вызывает лейшмания кожная. Лейшмании – мелкие простейшие от 2 до 4 микрометров (мкм) в длину. При этом заболевании образуются медленно заживающие язвы. Переносчиком лейшмании являются москиты. |
|
Среди жутиконосцев есть организмы с несколькими жгутиками, как у лямблий. Тело лямблии грушевидной формы. У лямблии 4 пары жгутиков. В цитоплазме клетки расположены 2 крупных ядра. На поверхности тела расположены присасывающие диски. Лямблии обитают в кишечнике млекопитающих, в том числе человека, земноводных и некоторых беспозвоночных животных. Болезнь, вызываемая лямблиями, называется лямблиоз. |
|
Несмотря на всё многообразие видов класса Жгутиконосцы, его представители имеют характерные черты, отличающие представителей этого класса от других простейших. Так у представителей класса Жгутиконосцы имеется жгутик, являющийся органоидом передвижения
|
|
Эвглену зелёную относят к миксотрофам, потому что на свету она питается автотрофно, а в темноте гетеротрофно. |
|
А теперь выполните задание: Распределите жгутиконосцев из предложенного списка на группы: жгутиконосцы-паразиты, колониальные формы.
Проверим выполненное задание. К жгутиконосцам-паразитам относятся: трипаносома, лейшмания, лямблия К колониальным жгутиконосцам: гониум, пандорина, вольвокс.
|




 Трипаносома африканская вызывает сонную болезнь. Название болезни связано с сонливым состоянием вследствие поражения нервной системы. Переносчиками паразита являются мухи цеце, то есть трансмиссивный способ передачи заболевания.
Трипаносома африканская вызывает сонную болезнь. Название болезни связано с сонливым состоянием вследствие поражения нервной системы. Переносчиками паразита являются мухи цеце, то есть трансмиссивный способ передачи заболевания. Длина тела 0, 008-0,03 мм.
Длина тела 0, 008-0,03 мм.
Остались вопросы по теме? Наши педагоги готовы помочь!
Подготовим к ЕГЭ, ОГЭ и другим экзаменам
Найдём слабые места по предмету и разберём ошибки
Повысим успеваемость по школьным предметам
Поможем подготовиться к поступлению в любой ВУЗ
Выбрать педагогаОставить заявку на подбор
Ингибирование дыхания жгутиковых простейших гипохолестеринемическими препаратами
- Опубликовано:
- С. ААРОНСОН 1
Природа том 202 , страница 1355 (1964)Цитировать эту статью
54 доступа
3 Цитаты
-
Сведения о показателях
Abstract
НЕСКОЛЬКО гипохолестеринемических препаратов разобщают окислительное фосфорилирование в митохондриях печени крыс 1 . Также некоторые из этих препаратов, например, бензмалецен и трипаранол (« MER -29»), ингибируют размножение Ochromonas danica , фитофлагелляты; олеиновая кислота аннулирует это ингибирование, но стеролы или предшественники стеролов не 2 . Ингибируют ли эти препараты синтез АТФ в O. danica ? Остается выяснить, существует ли особенно тесная связь между синтезом АТФ и синтезом липидов.
Также некоторые из этих препаратов, например, бензмалецен и трипаранол (« MER -29»), ингибируют размножение Ochromonas danica , фитофлагелляты; олеиновая кислота аннулирует это ингибирование, но стеролы или предшественники стеролов не 2 . Ингибируют ли эти препараты синтез АТФ в O. danica ? Остается выяснить, существует ли особенно тесная связь между синтезом АТФ и синтезом липидов.
Ссылки
Whitehouse, M.W., Nature , 201 , 629 (1964).
Артикул ОБЪЯВЛЕНИЯ КАС Google Scholar
Ааронсон С., Бенски Б., Шифрин М. и Бейкер Х., Proc. соц. Эксп. биол. и мед. , 109 , 130 (1962).
Артикул КАС Google Scholar
Ааронсон С. и Шер С., 9 лет0039 J.
 Протозоол. , 7 , 156 (1960).
Протозоол. , 7 , 156 (1960).Артикул КАС Google Scholar
Райт, Л. Д., и Леб, М., Proc. соц. Эксп. биол. и мед. , 103 , 183 (1960).
Артикул КАС Google Scholar
Райт Л.Д., Ли Л. и Трагер Р., Biochem. Биофиз. Рез. Комм. , 3 , 264 (1960).
Артикул КАС Google Scholar
Гараттини С. и Паолетти Р., Энн. Преподобный Фармакол. , 3 , 92 (1963).
Google Scholar
Флак, Э. Р., и Притам, Г. Х., Метаболизм , 10 , 444 (1961).
КАС пабмед Google Scholar
Стейнберг, Д., Trans. Академик Нью-Йорка науч.
 , 24 , 704 (1962).
, 24 , 704 (1962).Артикул КАС Google Scholar
Холмс В.Л. и Ди Туллио С.В., Амер. Дж. Клин. Питание , 10 , 310 (1962).
Артикул КАС Google Scholar
Ссылки на скачивание
Информация об авторе
Авторы и организации
Haskins Laboratories, New York, 17
S. AARONSON
- S. AARONSON
Просмотреть публикации авторов
9 0022 Вы также можете искать этого автора в PubMed Google Scholar
Права и разрешения
Перепечатка и разрешения
Об этой статье
Эта статья цитируется
Структурно-функциональная активность азастеролов и азотсодержащих стероидов
- Джон Дж.
 Кабара
Кабара - Дональд Л. Хольцшу
- Демокри Тос П. Катсулакос
Липиды (1976)
- Джон Дж.
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Водоросли | Определение, характеристики, классификация, примеры и факты
Водоросли «фужер русалки»
Смотреть все медиа
- Ключевые люди:
- Фердинанд Кон Сильвия Эрл Сэр Уильям Джексон Хукер Натанаэль Прингшейм Пер Теодор Клев
- Похожие темы:
- динофлагеллят водоросли красные водоросли зеленые водоросли фитопланктон
Просмотреть весь связанный контент →
Популярные вопросы
Что такое водоросли?
Водоросли определяются как группа преимущественно водных, фотосинтезирующих и несущих ядра организмов, у которых отсутствуют настоящие корни, стебли, листья и специализированные многоклеточные репродуктивные структуры растений. Их фотосинтетические пигменты также более разнообразны, чем у растений, и их клетки имеют черты, не встречающиеся у растений и животных.
Их фотосинтетические пигменты также более разнообразны, чем у растений, и их клетки имеют черты, не встречающиеся у растений и животных.
Какие органеллы содержат водоросли?
Водоросли являются эукариотическими организмами и содержат три типа связанных с двойной мембраной органелл: ядро, хлоропласт и митохондрию. В большинстве клеток водорослей есть только одно ядро, хотя некоторые клетки многоядерные.
Являются ли водоросли токсичными?
Некоторые виды водорослей выделяют токсины, смертельные для рыб, или делают моллюсков и рыб небезопасными для употребления в пищу. Динофлагелляты (таксономически спорные) несут ответственность за красные приливы, которые не только выделяют в воду токсины, которые могут быть смертельными для водных организмов, но также разносятся ветром токсичные клетки, которые могут вызывать проблемы со здоровьем у дышащих воздухом организмов.
Какого размера водоросли?
Размеры водорослей варьируются от пикопланктона, диаметр которого составляет от 0,2 до 2 микрометров (от 0,000008 до 0,000079 дюймов), до гигантских водорослей, длина которых может достигать 60 метров (200 футов).
Чем важны водоросли?
Водоросли производят до половины кислорода в атмосфере Земли, а водоросли помогают удерживать углекислый газ в атмосфере, накапливая его. Водоросли также являются пищей почти для всех водных организмов и имеют важное экономическое значение как источник сырой нефти, а также как источники пищи и ряда фармацевтических и промышленных продуктов для человека.
водоросли , единственное число водоросли , представители группы преимущественно водных фотосинтезирующих организмов царства Protista. У водорослей много типов жизненных циклов, и они варьируются в размерах от микроскопических видов Micromonas до гигантских водорослей, которые достигают 60 метров (200 футов) в длину. Их фотосинтетические пигменты более разнообразны, чем у растений, а их клетки имеют черты, не встречающиеся у растений и животных. Помимо своей экологической роли в качестве производителей кислорода и кормовой базы почти для всех водных организмов, водоросли имеют важное экономическое значение как источник сырой нефти, а также как источники пищи и ряда фармацевтических и промышленных продуктов для человека. Таксономия водорослей является спорной и подвержена быстрым изменениям по мере открытия новой молекулярной информации. Изучение водорослей называется phycology , а человек, изучающий водоросли, — phicologist.
Таксономия водорослей является спорной и подвержена быстрым изменениям по мере открытия новой молекулярной информации. Изучение водорослей называется phycology , а человек, изучающий водоросли, — phicologist.
В этой статье водоросли определяются как эукариотические (ядерные) организмы, которые фотосинтезируют, но лишены специализированных многоклеточных репродуктивных структур растений, которые всегда содержат клетки, производящие фертильные гаметы, окруженные стерильными клетками. У водорослей также отсутствуют настоящие корни, стебли и листья — черты, общие с бессосудистыми низшими растениями (например, мхами, печеночниками и роголистниками). Кроме того, водоросли, рассматриваемые в этой статье, исключают прокариотические (безъядерные) сине-зеленые водоросли (цианобактерии).
Начиная с 1830-х годов, водоросли были разделены на основные группы в зависимости от цвета, например, красные, коричневые и зеленые. Цвета являются отражением различных пигментов хлоропластов, таких как хлорофиллы, каротиноиды и фикобилипротеины.