Цитоплазматическая наследственность
1. Только ли в ядре клетки обнаружена ДНК?
Ответ. ДНК содержится в ядре, митохондриях и пластидах. У прокариот ДНК в клетке находится в мембране (на клеточной стенке).
2. Какие функции в клетке выполняют митохондрии; пластиды?
Ответ. Функция митохондрий — участие в энергетических клеточных процессах, образование АТФ. Различают три вида пластид — хлоропласты, хромопласты и лейкопласты. Функция хлоропластов – фотосинтез, т. е. синтез органических веществ из воды и углекислого газа с использованием энергии солнечного света. Функция хромопластов — цветовое привлечении животных, обеспечивающих опыление цветков и распространение семян, лейкопластов — образование из глюкозы крахмала и накапливание его в запасающих органах растений.
3. К какому типу относится эвглена зелёная?
Ответ.
Вопросы после §44
1. Какая наследственность называется цитоплазматической? Какое значение она может иметь?
Ответ. Существует путь передачи наследственной информации через цитоплазму клетки. Такой вид наследственности называют цитоплазматической или нехромосомной. Собственную ДНК содержат митохондрии и пластиды. Благодаря этому они способны к самовоспроизведению. Если клетка эвглены зелёной утрачивает пластиды, она не может снова их образовать, несмотря на сохранившееся ядро. Обычно в клетке эвглены находятся около 100 хлоропластов. В темноте эвглена переходит к гетеротрофному питанию, и её пластиды не размножаются, хотя сама эвглена продолжает делиться.
2. Митохондрии наследуются по линии отцовского или материнского организма?
Ответ. Характерная черта цитоплазматической наследственности – это наследование по линии матери. Действительно, и пластид, и митохондрий в яйцеклетке может быть много, а вот в мужских гаметах этих органоидов обычно нет, так как эти клетки практически лишены цитоплазмы. В сперматозоидах присутствуют митохондрии, но они всё равно не проникают в яйцеклетку, так как при слиянии гамет в яйцеклетку попадает только ядро сперматозоида, содержащее генетический материал. Таким образом, все митохондрии и пластиды зиготы достаются ей в наследство только от материнского организма.
3. Сколько хромосом расположено в бактериальной клетке?
Ответ. Прокариоты (бактерии,) не имеют хромосом в собственном смысле этого слова. У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе конъюгации.
У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе конъюгации.
4. Взаимосвязаны ли между собой хромосомная и нехромосомная наследственность?
Ответ. Показано, что хромосомная и нехромосомная наследственность могут взаимодействовать, приводя к сложным случаям наследования. Например, большинство белков митохондрий закодировано в ядерных генах и наследуется по правилам Менделя, а оставшиеся белки кодируются в ДНК самих митохондрий, которые передаются только по материнской линии. В митохондриях обнаружены гены ферментов клеточного дыхания, а также гены, обусловливающие устойчивость к некоторым неблагоприятным воздействиям.
В митохондриях обнаружены гены ферментов клеточного дыхания, а также гены, обусловливающие устойчивость к некоторым неблагоприятным воздействиям.
Влияние УФ-В на подвижность и фотоповедение зеленых жгутиконосцев, Euglena gracilis
Влияние УФ-В на подвижность и фотоповедение зеленых жгутиковых, Euglena gracilis
Скачать PDF
Скачать PDF
- Оригинальные документы
- Опубликовано:
- Донат-П.
 Хедер 1
Хедер 1
Архив микробиологии том 141 , страницы 159–163 (1985)Цитировать эту статью
54 доступа
37 цитирований
Сведения о показателях
Abstract
УФ-В ингибирует подвижность зеленых жгутиконосцев, Euglena gracilis , при интенсивности потока выше, чем ожидаемая при естественном солнечном свете, даже когда озоновый слой стратосферы частично уменьшен антропогенными загрязнителями.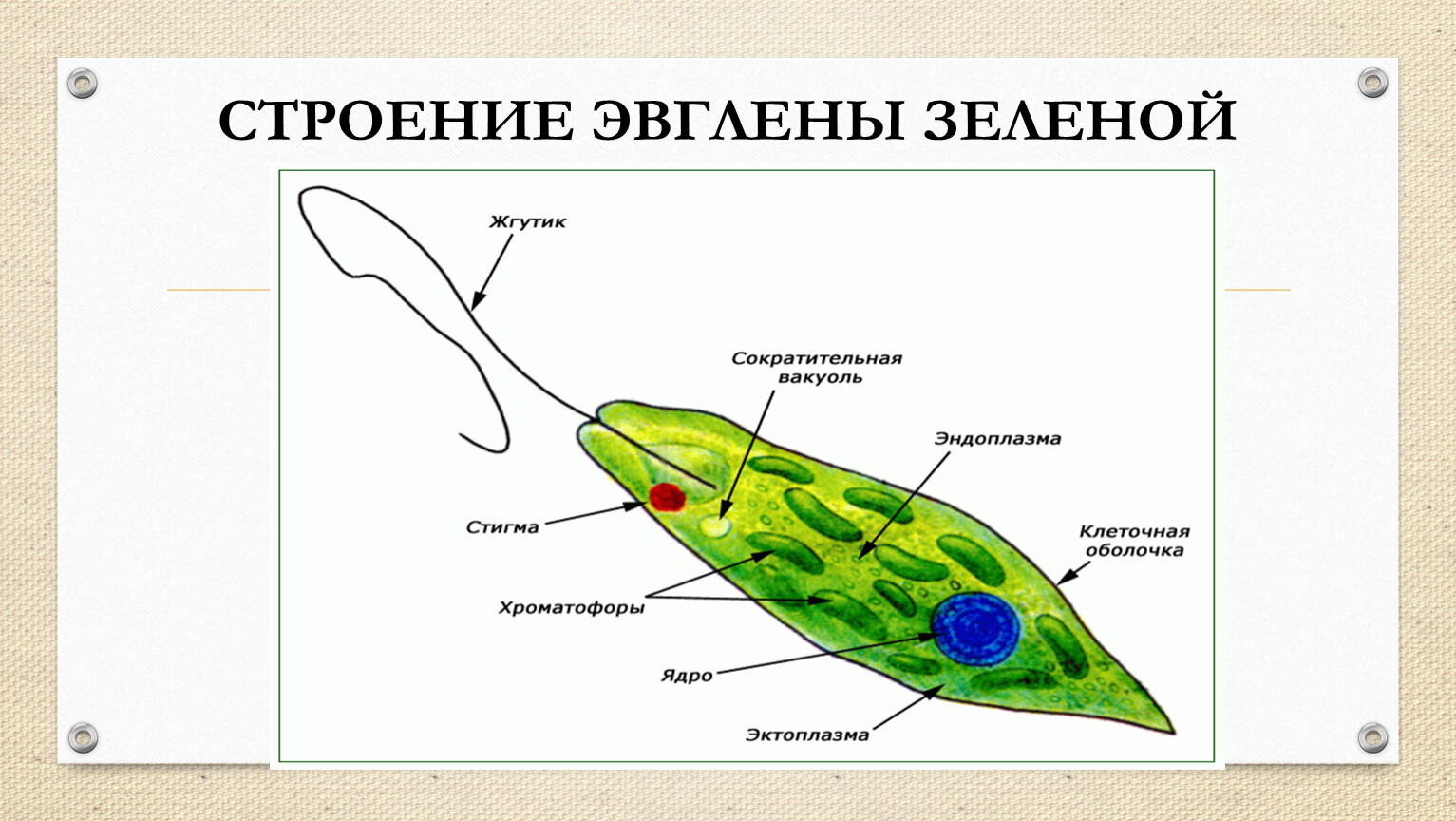 Однако фототаксическая ориентация клеток резко нарушается лишь незначительно повышенными уровнями УФ-В-облучения. Поскольку нарушается только отрицательный фототаксис (движение от сильного источника света), а положительный фототаксис (движение к слабому источнику света) не нарушается, хрупкое равновесие, с помощью которого организмы приспосабливают свое положение в своей среде обитания, нарушается. В этих условиях клетки не могут отступить от опасного уровня радиации и в конечном итоге погибают не от УФ-В облучения, а от фотообесцвечивания их фотосинтетических пигментов при ярком дневном свете на поверхности.
Однако фототаксическая ориентация клеток резко нарушается лишь незначительно повышенными уровнями УФ-В-облучения. Поскольку нарушается только отрицательный фототаксис (движение от сильного источника света), а положительный фототаксис (движение к слабому источнику света) не нарушается, хрупкое равновесие, с помощью которого организмы приспосабливают свое положение в своей среде обитания, нарушается. В этих условиях клетки не могут отступить от опасного уровня радиации и в конечном итоге погибают не от УФ-В облучения, а от фотообесцвечивания их фотосинтетических пигментов при ярком дневном свете на поверхности.
Скачайте, чтобы прочитать полный текст статьи
Ссылки
Batschelet E (1965) Статистические методы анализа проблем ориентации животных и некоторых биологических ритмов. В: Galles SR, Schmidt-Koenig K, Jacobs GJ, Belleville RF (eds) Ориентация животных и навигация.
 Вашингтон, НАСА, стр. 61–91
Вашингтон, НАСА, стр. 61–91. Google Scholar
-
Батчелет Е (1981) Циркулярная статистика в биологии. Академик Пресс, Лондон
Google Scholar
Benedetti PA, Lenci F (1977) In vivo микроспектрофлуориметрия пигментов фоторецепторов в Euglena gracilis . Фотохим Фотобиол 26:315–318
Google Scholar
Чекуччи А., Коломбетти Г., Феррара Р., Ленчи Ф. (1976) Спектры действия для фотоаккумуляции зеленого и бесцветного Euglena : данные для идентификации рецепторных пигментов. Фотохим Фотобиол 23:51–54
Google Scholar
Colombetti G, Lenci F (1982) Реакции на световые, химические и механические раздражители. В: Buetow DE (ed) Биология Euglena , том III. Academic Press, Нью-Йорк, стр. 169–195
. Google Scholar
Коломбетти Г.

Google Scholar
Дамкаер Д.М., Дей Д.Б., Херон Г.А., Прентис Э.Ф. (1980) Влияние УФ-В-излучения на приповерхностный зоопланктон Пьюджет-Саунд. Экология 44:149–158
Google Scholar
Дин Б (1969) Спектры действия фототаксических ответов в Euglena . Биохим Биофиз Акта 177:136–143
Google Scholar
Diehn B (1979)Фотические реакции и сенсорная трансдукция у подвижных простейших. В: Autrum H (ed) Справочник по сенсорной физиологии, том 6. Springer, Berlin Heidelberg New York, стр. 23–68
. Google Scholar
Galbally LE (1976) Техногенный четыреххлористый углерод в атмосфере.
 Наука 111:619–624
Наука 111:619–624Google Scholar
Гаррисон Л.М., Мюррей Л.Е., Грин А.Е.С. (1978) Ультрафиолетовый предел солнечного излучения на поверхности земли с фотонным монохроматором. Appl Opt 17: 683–684
Google Scholar
Häder DP (1981) Компьютерная оценка фототактической ориентации микроорганизмов. ЭДВ Мед Биол 12:27–30
Google Scholar
Häder DP (1983a) Влияние УФ-В облучения на развитие сорокарпа Dictyostelium discoideum . Фотохим Фотобиол 38:551–555
Google Scholar
Häder DP (1983b) Ингибирование фототаксиса и подвижности под действием УФ-В-облучения у слизней
Google Scholar
Häder DP (1984) Влияние УФ-В на подвижность и фотоориентацию цианобактерий, Phormidium uncinatum .
 Arch Microbiol 140:34–39
Arch Microbiol 140:34–39Google Scholar
Häder DP, Colombetti G, Lenci F, Quaglia M (1981) Фототаксис у жгутиковых, Euglena gracilis и Ochromonas danica . Arch Microbiol 130:78–82
Google Scholar
Lenci F, Colombetti G, Häder DP (1983) Роль гасителей и ингибиторов флавина в сенсорной передаче отрицательного фототаксиса у жгутиковых, Euglena gracilis . Curr Microbiol 9:285–290
Google Scholar
Липсон Э.Д., Хедер Д.П. (1984) Сбор видеоданных для двигательных реакций отдельных организмов. Фотохим Фотобиол 39:437–441
Google Scholar
Мардиа К.В. (1972) Статистика данных по направлениям. Академик Пресс, Лондон
Google Scholar
Молина М.Дж., Роуланд Ф.
