Эвглена зелёная
Тема:«Эвглена зелёная»
Цель:1) раскрыть особенности строения и процессов жизнедеятельности Эвглены зелёной;
2) доказать принадлежность эвглены к царству животных.
Ход урока
I. Повторение материала.
Фронтально:
1) Назовите царства живой природы.
2) Перечислите признаки растительных организмов.
3) Перечислите признаки животных организмов.
II. Изучение нового материала.
Учитель:
1) Сегодня мы с вами познакомимся с удивительным представителем живой природы. Этот представитель перед вами (на доске рисунок организма).
2) Зачитать текст об этом представителе (поставить проблему).
Если взять зелёную плёнку с поверхности небольшой лужи, застоявшейся на проезжей дороге или около скотного двора, то мы увидим под микроскопом зелёные одноклеточные организмы. Тело у них стройное и продолговатое с одним жгутиком.
Присутствие хлорофилла придаёт им зелёный цвет, а маленький красноватый “глазок” на переднем конце вполне соответствует такому же “глазку” у хламидомонады. Эти организмы, как и водоросли, способны к фотосинтезу.
Эти организмы, как и водоросли, способны к фотосинтезу.
Кто же это? Что это за организм?
3) Назвать этот организм.
Учитель: – А вы знаете, как зовут этот организм?
Ученики: – Это эвглена!
4) Разобрать систематическое положение.
Вид
Род
Класс
Тип
Царство
Постепенно заполняем эту схему:
Эвглена – какая систематическая единица (род), тогда вид – эвглена зелёная, почемузелёная?
– Т. к. этот организм зелёного цвета, он содержит хлорофилл.
– Что мы еще можем сказать о нём?
Читая выданный текст, учащиеся могут определить класс, тип; царство нельзя.
Получается на доске схема:
Вид | Эвглена зелёная |
Род | Эвглена |
Класс | Жгутиковые (жгутик – орган передвижения) |
Тип | Простейшие (тело состоит из 1 клетки) |
Царство | ? |
5) Почему нельзя определить царство, читая текст (есть сходство с растениями).
6) Разбираем признаки эвглены (находим противоречия).
Растение | Животные |
– глазок | – глазок |
– хлорофилл | – хлорофилл |
– автотрофное питание | – автотрофное питание |
– жгутик | – жгутик |
| – ограниченный рост |
Учащиеся в удивлении, кто же это, растение или животное.
Далее учитель зачитывает текст:
Если поместить этот живой организм в полную темноту, прибавив к воде картофельного отвара, и подержать его там 20-25 дней, то тогда мы увидим, что организмы не погибли, хотя и лишились своей зелёной окраски.
– Что мы можем сказать теперь?
– Этот организм способен к гетеротрофному питанию, потому он и не погиб, т. е. он питался органическими веществами картофельного отвара.
– Так кто же это?
Учащиеся говорят, что это животное, тогда учитель спрашивает, а как же быть с признаками растений. Дети снова в недоумении.
Дети снова в недоумении.
После чего учитель сообщает, что более тщательные исследования ученых, показали наличие у эвглены ротового отверстия.
– Вот тогда учащиеся убеждены, что это животное.
7) Почему? Доказать?
Растения не могут в темноте питаться, т. к. органические вещества образуются у них только при действии света, и у растений нет ротового отверстия.
8) Формулируем вывод и знакомство с новым типом питания.
Вывод:
Эвглена зелёная относится к царству животных, в темноте питается гетеротрофно, но она может питаться и как растение (автотрофно) на свету, значит у неё смешанный тип питания.
Учитель: Да, у эвглены смешанный тип питания, а называется он – МИКСОТРОФНЫЙ.
В результате определили царство организма – царство Животные. Вопрос с доски убираем.
9) Почему этот организм вызвал у нас затруднение, удивление?
– Потому что он сочетает в себе признаки растений и животных.
– А о чём это говорит?
– Это говорит о единстве происхождения органического мира.
III. Закрепление материала (работа в тетрадях).
а) Запись темы урока.
б) Систематическое положение эвглены зелёной.
в) Вывод.
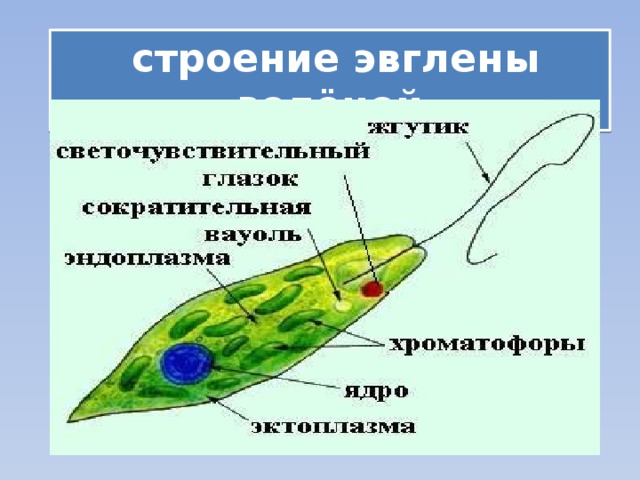
г) Подписать органоиды клетки и написать выполняемые функции (рисунок выдан).
IV. Запись дом. задания.
Адрес публикации: https://www.prodlenka.org/metodicheskie-razrabotki/75333-jevglena-zeljonaja
Эвглена зеленая. Образ жизни и среда обитания эвглены зелёной * Интересное
Share
Pin
Tweet
Send
Share
Send
Тело эвглены зеленой представляет собой продолговатую клетку зеленого цвета и покрыто оболочкой, которая называется пеликула. Задний конец тела — заостренный, передний — закругленный и имеет два жгутика, один из которых редуцирован, короткий, а второй — длинный, тонкий, который служит ей для передвижения. Эвглена делает до 40 оборотов в секунду жгутиком, благодаря чему задним концом тела быстро передвигается в воде. Второй жгутик (короткий) не получается вне пеликула. Живет в основном в застоявшейся воде, где много гниющих органических остатков. Имеет небольшие размеры — до 200 мкм (0,2 миллиметра).
Второй жгутик (короткий) не получается вне пеликула. Живет в основном в застоявшейся воде, где много гниющих органических остатков. Имеет небольшие размеры — до 200 мкм (0,2 миллиметра).
Строение клетки
Тело имеет постоянную форму, так как оболочка тела плотная. В клетке содержатся такие органеллы:
- большое ядро ‘,
- около двадцати хлоропластов,
- включения питательных веществ, которые служат запасом в то время, когда не хватит еды
- глазок — специфический светочувствительный орган красноватого цвета. Это не значит, что эвглена видит этим глазком свет, она его чувствует этим органом,
- сократительная вакуоль — находится у ячейки, благодаря ей эвглена избавляется лишней воды и вредных веществ, накопившихся в ней. Название «сократительная» получила за то, что сокращается во время вывода ненужных веществ и воды за пределы тела.
У жгутика находится светочувствительный глазок (стигма), благодаря которому эвглена реагирует на свет (фототаксис). В клетке эвглены является хроматофоры, содержащие хлорофилл, благодаря которым эвглена может проводит процесс фотосинтеза в условиях освещения.
В клетке эвглены является хроматофоры, содержащие хлорофилл, благодаря которым эвглена может проводит процесс фотосинтеза в условиях освещения.
Питание
На ярком свету эвглена, подобно растениям, использует энергию солнечных лучей и в результате фотосинтеза в ее хлоропластах образуются необходимые для жизни питательные вещества. Поэтому она всегда ищет освещенные места. Запасными продуктами являются парамилон i лейкозина, которые громоздятся в виде бесцветных зернышек. Также эвглена может питаться, используя осмос или углубление тела (гетеротрофы). Это касается экземпляров, которые живут в темноте и потеряли хлорофилл, или лишенных хроматофор. По ругуляцию осмотического давления в клетке и продуктов превращения веществ соответствуют сократительные вакуоли.
Образование цисты
При неблагоприятных условиях существования, например, при высыхании водоема или снижении температуры воды, она прекращает питания и передвижения, ее тело округляется и покрывается плотной защитной оболочкой, жгутик отпадает.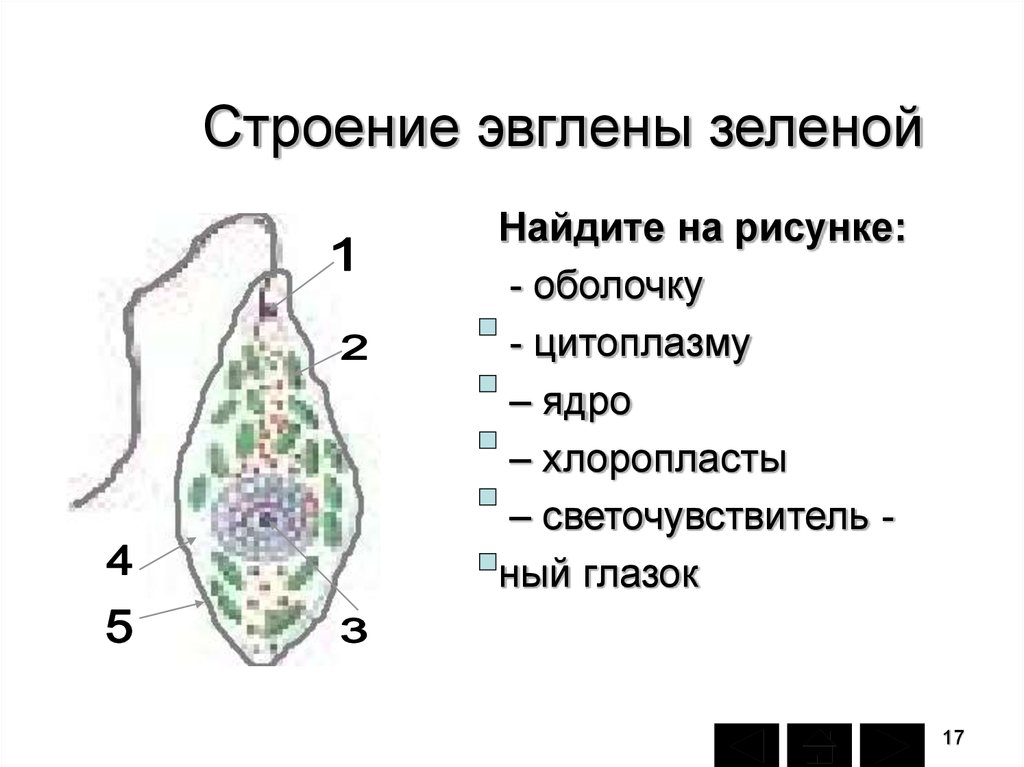 Так происходит переход эвглены в состояние покоя, называется цист. В этом состоянии она способна длительное время пережидать неблагоприятные жизненные условия.
Так происходит переход эвглены в состояние покоя, называется цист. В этом состоянии она способна длительное время пережидать неблагоприятные жизненные условия.
Размножение
Эвглена размножается бесполым, продольным разделением, который (после деления ядра) от главного тельца и жгутика. Сначала образуются два ядра, затем формируются два жгутика, две сократительные вакуоли и две ячейки. Далее вдоль всего тела появляется продольная борозда, которая постепенно делит клетку пополам.
Share
Pin
Tweet
Send
Share
Send
Крио-ЭМ структура высокоатипичной цитоплазматической рибосомы Euglena gracilis
1. Уилсон Д.Н., Дудна Кейт Дж.Х.. Строение и функция эукариотической рибосомы. Харб Колд Спринг. Перспектива. биол. 2012 г.; 4: а011536. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2.
Олсен Г.Дж., Везе К. Р..
Рибосомная РНК: ключ к филогенезу. FASEB J. 1993; 7:113–123. [PubMed] [Google Scholar]
Р..
Рибосомная РНК: ключ к филогенезу. FASEB J. 1993; 7:113–123. [PubMed] [Google Scholar]
3. Ютин Н., Пучбо П., Кунин Е.В., Вольф Ю.И.. Филогеномика прокариотических рибосомных белков. ПЛОС Один. 2012 г.; 7:e36972. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Джавед А., Орлова Е.В.. Харрис Дж.Р., Марлес-Райт Дж.. Комплексы макромолекулярных белков II: структура и функции. 2019; Чам: Springer International Publishing; 53–81. [Google Scholar]
5. Амунтс А., Браун А., Тутс Дж., Шерес С.Х.В., Рамакришнан В.. Строение митохондриальной рибосомы человека. Наука. 2015 г.; 348:95–98. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Десаи Н., Браун А., Амунтс А., Рамакришнан В.. Строение митохондриальной рибосомы дрожжей. Наука. 2017; 355: 528–531. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7.
Бьери П., Гребер Б.Дж., Бан Н..
Структуры митохондриальных рибосом с высоким разрешением и их функциональное значение. Курс. мнение Структура биол. 2018; 49:44–53. [PubMed] [Google Scholar]
2018; 49:44–53. [PubMed] [Google Scholar]
8. Бьери П., Лейбундгут М., Заурер М., Берингер Д., Бан Н.. Полная структура рибосомы 70S хлоропласта в комплексе с фактором трансляции pY. EMBO J. 2017; 36: 475–486. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Мур П.Б., Стейц Т.А.. Участие РНК в функционировании рибосом. Природа. 2002 г.; 418:229–235. [PubMed] [Google Scholar]
10. Герби С.А. Циммерманн Р.А., Дальберг А.Е.. Рибосомная РНК — структура, эволюция, процессинг и функция в синтезе белка. 1996 год; Бока-Ратон: CRC Press; 71–87. [Google Scholar]
11. Фуджи К., Сусанто Т.Т., Саурабх С., Барна М.. Расшифровка функции сегментов экспансии в рибосомах. Мол. Клетка. 2018; 72:1013–1020. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Грей М.В., Шнаре М.Н.. Циммерманн Р.А., Дальберг А.Е.. Рибосомная РНК: структура, эволюция, процессинг и функция в биосинтезе белка. 1996; Бока-Ратон: CRC Press; 49–69. [Google Scholar]
13.
Грей М. В., Гопалан В..
По частям: создание рибозима. Дж. Биол. хим. 2020; 295:2313–2323. [Бесплатная статья PMC] [PubMed] [Google Scholar]
В., Гопалан В..
По частям: создание рибозима. Дж. Биол. хим. 2020; 295:2313–2323. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Грей М.В. Необычная структура компонентов рибонуклеиновой кислоты в рибосоме Crithidia fasciculata , трипаносоматидного простейшего. Мол. Клетка. биол. 1981 год; 1: 347–357. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15.
Спенсер Д.Ф., Коллингс Дж.К., Шнаре М.Н., Грей М.В..
Множественные спейсерные последовательности в гене рибосомной РНК большой субъединицы ядра
16. Кэмпбелл Д.А., Кубо К., Кларк К.Г., Бутройд Дж.К.. Точная идентификация сайтов расщепления, участвующих в необычном процессинге трипаносомной рибосомной РНК. Дж. Мол. биол. 1987 год; 196: 113–124. [PubMed] [Google Scholar]
17.
Хашем Ю., Де Жорж А., Фу Дж., Бусс С.Н., Жоссинет Ф., Джобе А., Чжан К., Ляо Х.Ю., Грассуччи Р.А., Баджадж К. и др…
Структура криоэлектронной микроскопии высокого разрешения Trypanosoma brucei рибосома. Природа. 2013; 494: 385–389. [Бесплатная статья PMC] [PubMed] [Google Scholar]
и др…
Структура криоэлектронной микроскопии высокого разрешения Trypanosoma brucei рибосома. Природа. 2013; 494: 385–389. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Лю З., Гутьеррес-Варгас С., Вей Дж., Грассуччи Р.А., Рамеш М., Эспина Н., Сун М., Тутункуоглу Б., Мэдисон-Антенуччи С., Вулфорд Дж.Л. мл. и др… Модель структуры и сборки 60S рибосомной субъединицы Trypanosoma cruzi . проц. Натл. акад. науч. США 2016; 113:12174–12179. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Кук Дж.Р., Роксби Р.. Физические свойства плазмидоподобной ДНК из Эвглена тонкая. Биохим. Биофиз . Акта. 1985 год; 824:80–83. [PubMed] [Google Scholar]
20. Равель-Шапюи П., Николя П., Нигон В., Нейре О., Фрейсине Г.. Внехромосомная кольцевая ядерная рДНК Euglena gracilis . Нуклеиновые Кислоты Res. 1985 год; 13:7529–7537. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21.
Гринвуд С.Дж., Грей М.В..
Процессинг рРНК-предшественника в Euglena gracilis : идентификация промежуточных продуктов на пути к сильно фрагментированной большой субъединице рРНК.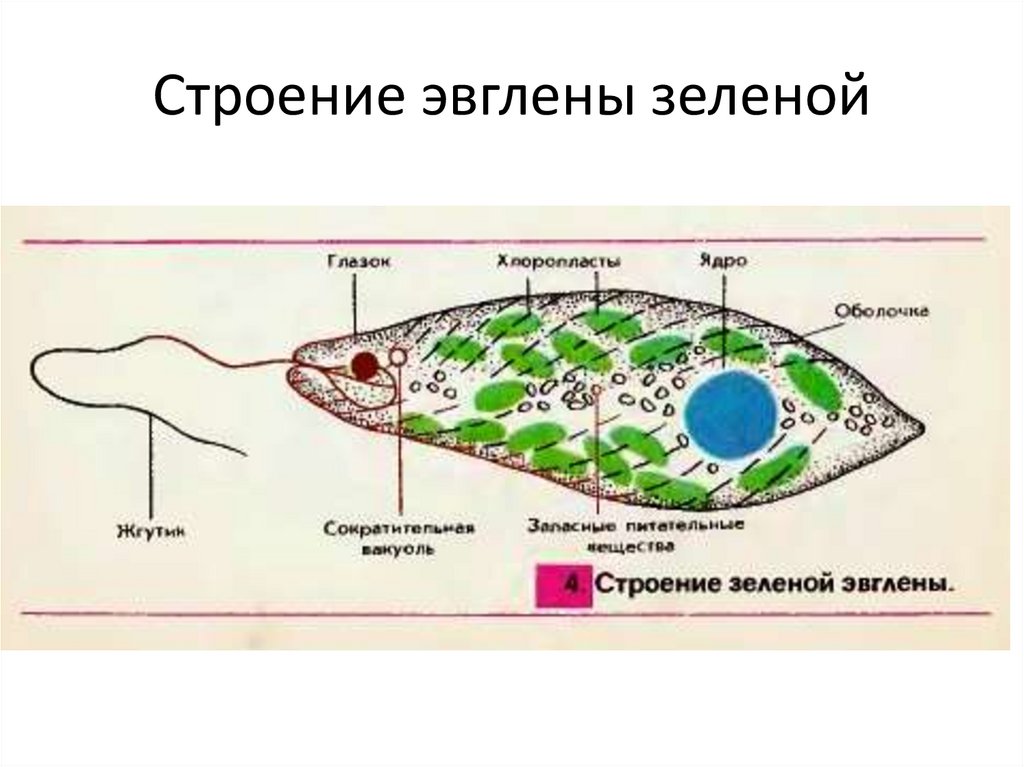
22. Шнаре М.Н., Грей М.В.. Шестнадцать дискретных компонентов РНК в цитоплазматической рибосоме Euglena gracilis . Дж. Мол. биол. 1990 г.; 215:73–83. [PubMed] [Google Scholar]
23. Шнаре М.Н., Кук Дж.Р., Грей М.В.. Четырнадцать внутренних транскрибируемых спейсеров в кольцевой рибосомной ДНК Euglena gracilis . Дж. Мол. биол. 1990 г.; 215:85–91. [PubMed] [Google Scholar]
24. Шнаре М.Н., Грей М.В.. Полные карты модификаций для малых и больших цитозольных рРНК субъединиц Euglena gracilis : функциональные и эволюционные последствия контрастных паттернов между двумя компонентами рРНК. Дж. Мол. биол. 2011 г.; 413:66–83. [PubMed] [Google Scholar]
25.
Лукеш Ю., Пэрис З., Регми С., Брейтлинг Р., Муреев С., Кушнир С., Пятков К., Йирку М., Александров К.А..
На инициацию трансляции у Leishmania tarentolae и Phytomonas serpens (Kinetoplastida) сильно влияет триплет пре-ATG и контекст его 5′-последовательности.
26. Фернандес-Певида А., Кресслер Д., де ла Крус Х.. Процессинг прерибосомной РНК в Saccharomyces cerevisiae . ПРОВОДА РНК. 2015 г.; 6: 191–209. [PubMed] [Google Scholar]
27. Шалев М., Розенберг Х., Смолкин Б., Насередин А., Копелянский Д., Белахов В., Шрепфер Т., Шахт Дж., Джаффе К.Л., Адир Н. и др… Структурная основа селективного нацеливания на рибосомы лейшманиоза: производные аминогликозидов как перспективные терапевтические средства. Нуклеиновые Кислоты Res. 2015 г.; 43:8601–8613. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Чжэн С.К., Паловчак Э., Армаш Ж.-П., Верба К.А., Ченг Ю., Агард Д.А.. MotionCor2: анизотропная коррекция движения, вызванного лучом, для улучшения криоэлектронной микроскопии. Нац. Методы. 2017; 14:331–332. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29.
Минделл Дж.А., Григорьев Н..
Точное определение локальной дефокусировки и наклона образца в электронной микроскопии. Дж. Структура. биол. 2003 г.; 142: 334–347. [PubMed] [Google Scholar]
Дж. Структура. биол. 2003 г.; 142: 334–347. [PubMed] [Google Scholar]
30. Зиванов Дж., Накане Т., Форсберг Б.О., Киманиус Д., Хаген В.Дж., Линдал Э., Шерес С.Х.. Новые инструменты для автоматизированного определения структуры крио-ЭМ высокого разрешения в РЭЛИОН-3. электронная жизнь. 2018; 7:e42166. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Шалев-Бенами М., Чжан Ю., Розенберг Х., Нобе Ю., Таока М., Мацов Д., Циммерман Э., Башан А., Изобе Т., Джаффе К.Л. и другие… Снимок атомного разрешения ингибирования рибосомы Leishmania аминогликозидом паромомицином. Нац. коммун. 2017; 8: 1589–1589. [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Накане Т., Киманиус Д., Линдал Э., Шерес Ш.Х.В.. Характеристика молекулярных движений в данных крио-ЭМ для отдельных частиц путем уточнения нескольких тел в RELION. электронная жизнь. 2018; 7:e36861. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33.
Афонин П.В., Пун Б.К., Рид Р.Дж., Соболев О. В., Тервиллигер Т.С., Уржумцев А., Адамс П.Д..
Уточнение в реальном пространстве в PHENIX для крио-ЭМ и кристаллографии. Акта Кристаллогр. Д Биол Кристаллогр. 2018; 74: 531–544. [Бесплатная статья PMC] [PubMed] [Google Scholar]
В., Тервиллигер Т.С., Уржумцев А., Адамс П.Д..
Уточнение в реальном пространстве в PHENIX для крио-ЭМ и кристаллографии. Акта Кристаллогр. Д Биол Кристаллогр. 2018; 74: 531–544. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Кукукельбир А., Сигуорт Ф.Дж., Тагаре Х.Д.. Количественная оценка локального разрешения крио-ЭМ карт плотности. Нац. Методы. 2014; 11: 63–65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Эмсли П., Локамп Б., Скотт В.Г., Коутан К.. Особенности и развитие Coot. Акта Кристаллогр. Д Биол Кристаллогр. 2010 г.; 66: 486–501. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Петтерсен Э.Ф., Годдард Т.Д., Хуанг К.С., Коуч Г.С., Гринблатт Д.М., Мэн Э.К., Феррин Т.Е.. UCSF Chimera — система визуализации для поисковых исследований и анализа. Дж. Вычисл. хим. 2004 г.; 25:1605–1612. [PubMed] [Google Scholar]
37.
Мориарти Н.В., Гроссе-Кунстлеве Р.В., Адамс П.Д..
электронный конструктор лигандов и инструментальные средства оптимизации (eLBOW): инструмент для определения координат лигандов и создания ограничений. Акта Кристаллогр. Д биол. Кристаллогр. 2009 г.; 65:1074–1080. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Акта Кристаллогр. Д биол. Кристаллогр. 2009 г.; 65:1074–1080. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Уильямс С.Дж., Хедд Дж.Дж., Мориарти Н.В., Присант М.Г., Видо Л.Л., Дейс Л.Н., Верма В., Киди Д.А., Хинтце Б.Дж., Чен В.Б. и другие… MolProbity: больше и лучше эталонных данных для улучшенной проверки структуры всех атомов. Белковая наука. 2018; 27:293–315. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Килинг П.Дж., Бурки Ф., Уилкокс Х.М., Аллам Б., Аллен Э.Э., Амарал-Зеттлер Л.А., Армбруст Э.В., Арчибальд Дж.М., Бхарти А.К., Белл С.Дж. и др… Проект секвенирования транскриптома морских микробов эукариот (MMETSP): освещение функционального разнообразия эукариотической жизни в океанах посредством секвенирования транскриптома. PLoS биол. 2014; 12:e1001889. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40.
Грабхер М.Г., Хаас Б.Дж., Яссур М., Левин Дж.З., Томпсон Д.А., Амит И., Адиконис Х., Фан Л., Райчоудхури Р., Зенг К. и др…
Сборка полноразмерного транскриптома из данных RNA-Seq без эталонного генома.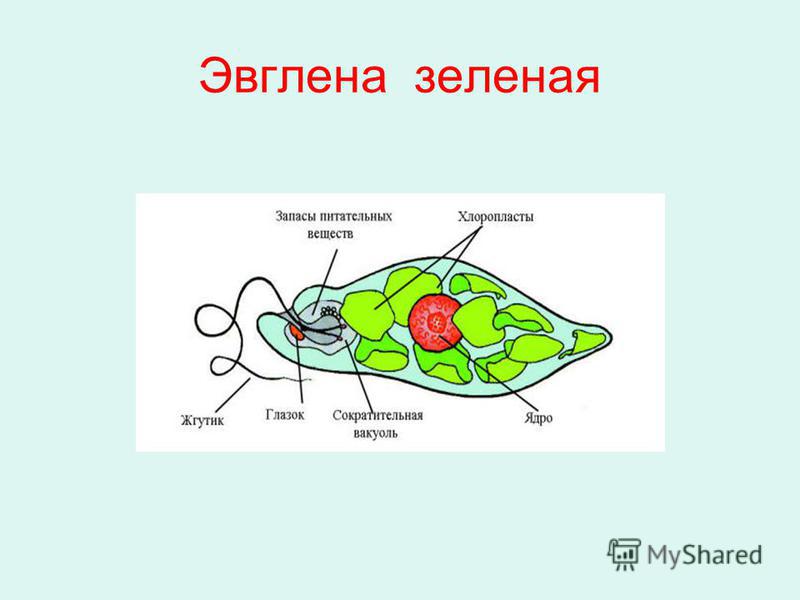 Нац. Биотехнолог. 2011 г.; 29:644. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Нац. Биотехнолог. 2011 г.; 29:644. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41.
Тессье Л.-Х., Келлер М., Чан Р.Л., Фурнье Р., Вейл Ж.-Х., Имбо П..
Короткие лидерные последовательности могут быть перенесены с малых РНК на незрелые мРНК путем транс-сплайсинга в
42. Таока М., Морофудзи Н., Ямаути Ю., Одзима Х., Кубота Д., Терукина Г., Нобе Ю., Накаяма Х., Такахаси Н., Косугэ Т.. Глобальное профилирование PROTOMAP для поиска биомаркеров раннего рецидива гепатоцеллюлярной карциномы. Дж. Протеом Рез. 2014; 13:4847–4858. [PubMed] [Google Scholar]
43. Синкава Т., Таока М., Ямаути Ю., Ичимура Т., Кадзи Х., Такахаси Н., Исобе Т.. STEM: Программный инструмент для крупномасштабного анализа протеомных данных. Дж. Протеом Рез. 2005 г.; 4: 1826–1831. [PubMed] [Академия Google]
44.
Гао Дж., Дуань З., Чжан Л., Хуан С., Лонг Л., Ту Дж., Лян Х., Чжан Ю., Шен Т., Лу Ф. .
Отсутствие восстановления циркулирующих NKG2D + CD56 dim NK-клеток при HBV-ассоциированной гепатоцеллюлярной карциноме после гепатэктомии является предиктором раннего рецидива. Онкоиммунология. 2016; 5:e1048061. [Бесплатная статья PMC] [PubMed] [Google Scholar]
.
Отсутствие восстановления циркулирующих NKG2D + CD56 dim NK-клеток при HBV-ассоциированной гепатоцеллюлярной карциноме после гепатэктомии является предиктором раннего рецидива. Онкоиммунология. 2016; 5:e1048061. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Курата Т., Наканиши С., Хасимото М., Таока М., Ямадзаки Ю., Исобе Т., Като Дж.-И.. Новый важный ген, участвующий в процессинге 16S рРНК у Кишечная палочка . Дж. Мол. биол. 2015 г.; 427: 955–965. [PubMed] [Google Scholar]
46. Накаяма Х., Ямаути Ю., Нобе Ю., Сато К., Такахаси Н., Шалев-Бенами М., Исобе Т., Таока М.. Метод прямой масс-спектрометрической идентификации позиционных изомеров нуклеозидов монометилированной РНК и его применение для анализа рРНК Leishmania . Анальный. хим. 2019; 91:15634–15643. [PubMed] [Google Scholar]
47.
Гастон К.В., Лимбах П.А..
Идентификация и характеристика некодирующих и кодирующих РНК и их модифицированных нуклеозидов методом масс-спектрометрии.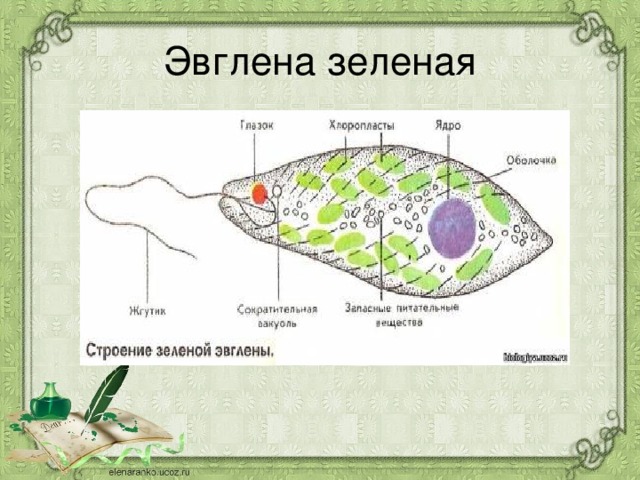 РНК биол. 2014; 11: 1568–1585. [Бесплатная статья PMC] [PubMed] [Google Scholar]
РНК биол. 2014; 11: 1568–1585. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Перри Р.П. Процессинг РНК. Анну. Преподобный Биохим. 1976 год; 45:605–629. [PubMed] [Google Scholar]
49. Уайт Т.С., Руденко Г., Борст П.. Три малые РНК в пределах единицы транскрипции трипаносомной рРНК длиной 10 т.п.н. аналогичны домену VII других эукариотических 28S рРНК. Нуклеиновые Кислоты Res. 1986 год; 14:9471–9489. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Шалев-Бенами М., Чжан Ю., Мацов Д., Хальфон Ю., Закай А., Розенберг Х., Циммерман Э., Башан А., Джаффе К.Л., Йонат А. и др… 2,8-Å Крио-ЭМ структура большой субъединицы рибосомы эукариотического паразита Лейшмания . Представитель Cell 2016; 16: 288–294. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51.
Чжан С., Лай М., Чанг В., Ю И., Дин К., Мразек Дж., Нг Х.Л., Ян О.О., Маслов Д.А., Чжоу З.Х..
Структуры и стабилизация кинетопластид-специфических расщепленных рРНК, выявленные при сравнении лейшманиальных и человеческих рибосом. Нац. коммун. 2016; 7:13223–13223. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Нац. коммун. 2016; 7:13223–13223. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Брито Керидо Дж., Мансера-Мартинес Э., Висенс К., Бохлер А., Чичер Дж., Симонетти А., Хашем Ю.. Крио-ЭМ структура нового кинетопластид-специфического рибосомного белка 40S. Состав. 2017; 25: 1785–1794. [PubMed] [Google Scholar]
53. Армаш Дж.-П., Яраш А., Ангер А.М., Вилла Э., Беккер Т., Бхушан С., Джоссине Ф., Хабек М., Диндар Г., Франкенберг С. и др… Крио-ЭМ структура и модель рРНК транслирующей эукариотической 80S рибосомы с разрешением 5,5 Å. проц. Натл. акад. науч. США, 2010 г.; 107:19748–19753. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Бан Н., Бекманн Р., Кейт Дж. Х. Д., Динман Дж. Д., Драгон Ф., Эллис С. Р., Лафонтен Д. Л. Дж., Линдал Л., Лильяс А., Липтон Дж. М. и др… Новая система наименования рибосомных белков. Курс. мнение Структура биол. 2014; 24: 165–169. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55.
Пасек С., Рислер Ж.-Л., Брезеллек П.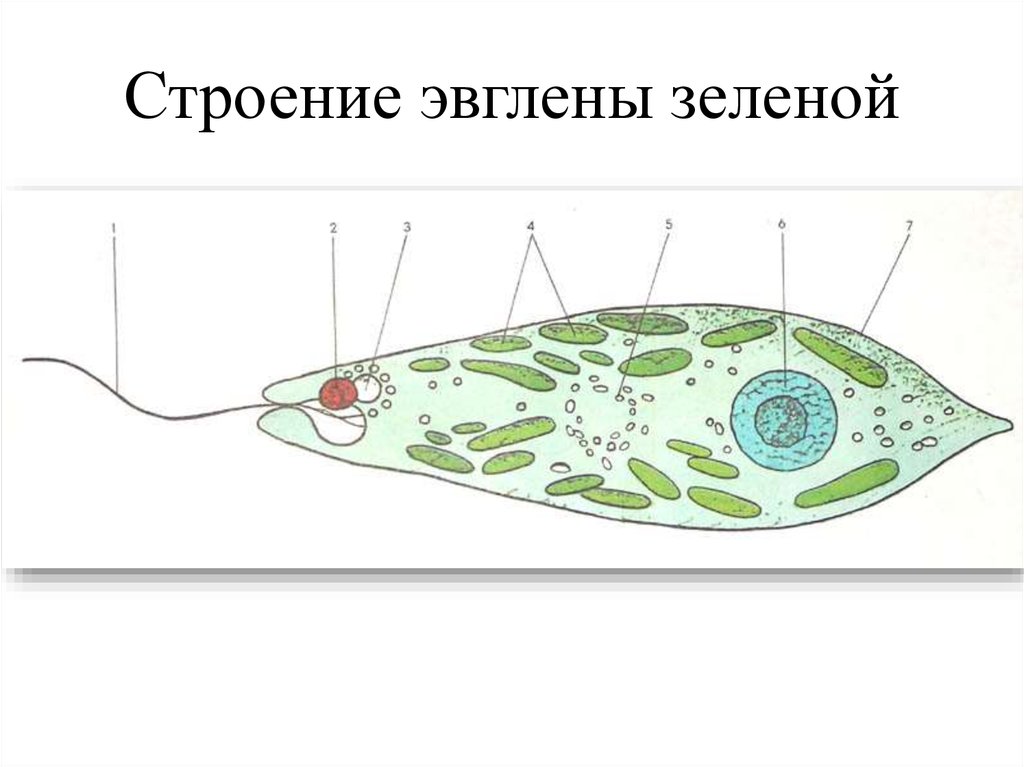 .
Слияние/деление генов является основным фактором эволюции многодоменных бактериальных белков. Биоинформатика. 2006 г.; 22:1418–1423. [PubMed] [Google Scholar]
.
Слияние/деление генов является основным фактором эволюции многодоменных бактериальных белков. Биоинформатика. 2006 г.; 22:1418–1423. [PubMed] [Google Scholar]
56. Рамакришнан В., Уайт С.В.. Структуры рибосомных белков: понимание архитектуры, механизмов и эволюции рибосом. Тенденции биохим. науч. 1998 год; 23:208–212. [PubMed] [Google Scholar]
57. Фокс Г.Э. Происхождение и эволюция рибосомы. Харб Колд Спринг. Перспектива. биол. 2010 г.; 2: а003483. [Бесплатная статья PMC] [PubMed] [Google Scholar]
58. Хариш А., Каэтано-Аноллес Г.. История рибосом раскрывает истоки современного синтеза белка. ПЛОС ОДИН. 2012 г.; 7:e32776. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Бен-Шем А., Гарро де Лубресс Н., Мельников С., Дженнер Л., Юсупова Г., Юсупов М.. Структура эукариотической рибосомы с разрешением 3,0 Å. Наука. 2011 г.; 334: 1524–1529. [PubMed] [Google Scholar]
60.
Фишер Н., Нейманн П., Коневега А.Л., Бок Л.В., Фикнер Р., Роднина М.В., Старк Х. .
Структура 9Комплекс 0029 E. coli рибосома-EF-Tu с разрешением <3 Å с помощью крио-ЭМ с поправкой на Cs. Природа. 2015 г.; 520: 567–570. [PubMed] [Google Scholar]
.
Структура 9Комплекс 0029 E. coli рибосома-EF-Tu с разрешением <3 Å с помощью крио-ЭМ с поправкой на Cs. Природа. 2015 г.; 520: 567–570. [PubMed] [Google Scholar]
61. Таока М., Нобэ Ю., Ямаки Ю., Ямаути Ю., Исикава Х., Такахаши Н., Накаяма Х., Исобе Т.. Полная химическая структура рРНК Saccharomyces cerevisiae : частичное псевдоуридилирование U2345 в 25S рРНК с помощью мякРНК snR9. Нуклеиновые Кислоты Res. 2016; 44:8951–8961. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Таока М., Нобэ Ю., Ямаки Ю., Сато К., Исикава Х., Изумикава К., Ямаути Ю., Хирота К., Накаяма Х., Такахаши Н. и др… Пейзаж полных химических модификаций РНК в рибосоме 80S человека. Нуклеиновые Кислоты Res. 2018; 46:9289–9298. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Монако П.Л., Марсель В., Диас Ж.-Ж., Катез Ф.. 2′-O-Метилирование рибосомной РНК: к эпитранскриптомному контролю трансляции. Биомолекулы. 2018; 8:106. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Лян Х.Х., Улиэль С., Хури А., Барт С., Донигер Т., Унгер Р., Михаэли С..
Полногеномный анализ C/D и H/ACA-подобных малых ядрышковых РНК в Trypanosoma brucei выявил специфичный для трипаносом паттерн модификации рРНК. РНК. 2005 г.; 11:619–645. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Лян Х.Х., Улиэль С., Хури А., Барт С., Донигер Т., Унгер Р., Михаэли С..
Полногеномный анализ C/D и H/ACA-подобных малых ядрышковых РНК в Trypanosoma brucei выявил специфичный для трипаносом паттерн модификации рРНК. РНК. 2005 г.; 11:619–645. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Барт С., Шалем Б., Хури А., Ткач И.Д., Лян Х.-Х., Улиэль С., Мыслюк И., Донигер Т., Салмон-Дивон М., Унгер Р. и др… Выяснение роли C/D мякРНК в процессинге и модификации рРНК у Trypanosoma brucei. Эукариот. Клетка. 2008 г.; 7:86–101. [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Декейтер В.А., Фурнье М.Дж.. Модификации рРНК и функция рибосом. Тенденции биохим. науч. 2002 г.; 27:344–351. [PubMed] [Академия Google]
67. Хелм М. Посттранскрипционная модификация нуклеотидов и альтернативная укладка РНК. Нуклеиновые Кислоты Res. 2006 г.; 34:721–733. [Бесплатная статья PMC] [PubMed] [Google Scholar]
68.
Пайфер К., Шарма С., Ватцингер П., Ламберт С. , Кёттер П., Энтиан К.-Д..
Rrp8p дрожжей, новая метилтрансфераза, ответственная за модификацию m 1 A 645 оснований 25S рРНК. Нуклеиновые Кислоты Res. 2013; 41:1151–1163. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Кёттер П., Энтиан К.-Д..
Rrp8p дрожжей, новая метилтрансфераза, ответственная за модификацию m 1 A 645 оснований 25S рРНК. Нуклеиновые Кислоты Res. 2013; 41:1151–1163. [Бесплатная статья PMC] [PubMed] [Google Scholar]
69. Шарма С., Ватцингер П., Кёттер П., Энтиан К.-Д.. Идентификация новой метилтрансферазы, Bmt2, ответственной за модификацию N-1-метиладенозинового основания 25S рРНК в Saccharomyces cerevisiae . Нуклеиновые Кислоты Res. 2013; 41:5428–5443. [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Доминиссини Д., Нахтергаэле С., Мошич-Мошковиц С., Пир Э., Кол Н., Бен-Хаим М.С., Дай К., Ди Сеньи А., Салмон-Дивон М., Кларк В.К. и другие… Динамический N 1 -метиладенозин метилом в матричной РНК эукариот. Природа. 2016; 530:441–446. [Бесплатная статья PMC] [PubMed] [Google Scholar]
71.
Ли С., Сюн С., Ван К., Ван Л., Шу С., Ма С., Йи С..
Картирование всего транскриптома выявляет обратимый и динамический N 1 -метиладенозин метилом. Нац. хим. биол. 2016; 12:311–316. [PubMed] [Google Scholar]
Нац. хим. биол. 2016; 12:311–316. [PubMed] [Google Scholar]
72. Сафра М., Сас-Чен А., Нир Р., Винклер Р., Нахшон А., Бар-Яаков Д., Эрлахер М., Россманит В., Штерн-Гиноссар Н., Шварц С.. Пейзаж m 1 A на цитозольной и митохондриальной мРНК с разрешением по одному основанию. Природа. 2017; 551: 251–255. [PubMed] [Google Scholar]
73. Андерсон Дж.Т., Другманс Л.. Грожан Х. Тонкая настройка функций РНК путем модификации и редактирования. 2005 г.; Берлин, Гейдельберг: Springer; 121–139. [Google Scholar]
74. Андерсон Дж., Фан Л., Куэста Р., Карлсон Б.А., Пак М., Асано К., Бьорк Г.Р., Тамаме М., Хиннебуш А.Г.. Необходимый ядерный комплекс Gcd10p-Gcd14p необходим для модификации 1-метиладенозина и созревания инициаторной метионил-тРНК. Гены Дев. 1998 год; 12:3650–3662. [Бесплатная статья PMC] [PubMed] [Google Scholar]
75.
Хелм М., Гиге Р., Флоренц К..
Метильная группа Уотсона-Крика, разрушающая пару оснований (m 1 A9), достаточна для укладки трилистника митохондриальной тРНК человека Лис . Биохимия. 1999 г.; 38:13338–13346. [PubMed] [Google Scholar]
Биохимия. 1999 г.; 38:13338–13346. [PubMed] [Google Scholar]
76. Лоренц К., Люнсе К.Э., Мёрль М.. Модификации тРНК: влияние на структуру и температурную адаптацию. Биомолекулы. 2017; 7:35. [Бесплатная статья PMC] [PubMed] [Google Scholar]
77. Джонхаут Н., Тран Дж., Смит М.А., Шонрок Н., Маттик Дж.С., Новоа Е.М.. Ландшафт модификации РНК при заболеваниях человека. РНК. 2017; 23: 1754–1769. [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Йокояма С., Ватанабэ К., Миядзава Т.. Динамические структуры и функции переноса рибонуклеиновых кислот от экстремальных термофилов. Доп. Биофиз. 1987; 23:115–147. [PubMed] [Google Scholar]
79. Дрогманс Л., Руверс М., Буйницки Дж.М., Трико К., Харч Т., Сталон В., Грожан Х.. Клонирование и характеристика тРНК (m 1 A58) метилтрансферазы (TrmI) из Thermus thermophilus HB27, белка, необходимого для роста клеток при экстремальных температурах. Нуклеиновые Кислоты Res. 2003 г.; 31:2148–2156. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Джомаа А., Фу Ю.-Х.Х., Берингер Д., Лейбундгут М., Шан С.-о, Бан Н..
Структура четвертичного комплекса между SRP, SR и транслоконом, связанным с транслирующей рибосомой. Нац. коммун. 2017; 8:15470. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Джомаа А., Фу Ю.-Х.Х., Берингер Д., Лейбундгут М., Шан С.-о, Бан Н..
Структура четвертичного комплекса между SRP, SR и транслоконом, связанным с транслирующей рибосомой. Нац. коммун. 2017; 8:15470. [Бесплатная статья PMC] [PubMed] [Google Scholar]
81. Кобаяши К., Джомаа А., Ли Дж. Х., Чандрасекар С., Берингер Д., Шан С.-О., Бан Н.. Структура рибосомального рецепторного комплекса SRP·SRP млекопитающих перед передачей. Наука. 2018; 360:323–327. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Кнорр А.Г., Шмидт К., Тесина П., Бернингхаузен О., Беккер Т., Беатрикс Б., Бекманн Р.. Архитектура Ribosome-NatA показывает, что сегменты расширения рРНК координируют N-концевое ацетилирование. Нац. Структура Мол. биол. 2019; 26:35–39. [PubMed] [Google Scholar]
83. Вурхис М., Фернандес И.С., Шерес С.Х.В., Хегде Р.С.. Структура комплекса рибосома-Sec61 млекопитающих с разрешением 3,4 Å. Клетка. 2014; 157: 1632–1643. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Что такое зеленое здание + 10 устойчивых зданий
Что такое зеленое здание + 10 устойчивых зданий
«Зеленые» здания прокладывают путь к более устойчивому и эффективному городскому планированию
#общество #НИОКР #архитектура
Экологичные здания сводят к минимуму потребление энергии и воды и являются ключевой частью устойчивого развития городов, направленного на борьбу с изменением климата. Ниже мы рассмотрим некоторые из зеленых зданий, от больших небоскребов до новых музеев, которые прокладывают путь к более ответственному городскому планированию.
Ниже мы рассмотрим некоторые из зеленых зданий, от больших небоскребов до новых музеев, которые прокладывают путь к более ответственному городскому планированию.
Зеленые или устойчивые здания отличаются высокой энергоэффективностью.
По данным Организации Объединенных Наций (ООН), в 2050 году около 68% человечества будет жить в городах. Они составляют всего 3% поверхности планеты, но потребляют 78% энергии и производят 60% выбросов парниковых газов. По этой причине в 2016 году сама ООН улучшила Новую программу развития городов , чтобы консультировать страны по их процессам урбанизации и сделать города более пригодными для жизни, инклюзивными, здоровыми, жизнестойкими и устойчивыми.
Iberdrola является членом Ассоциации Мадридской мировой столицы строительства, инженерии и архитектуры (MWCC). Эта организация продвигает эти сектора как в столице, так и по всей Испании, укрепляя международное лидерство испанских компаний и делая Мадрид и Испанию видимыми в качестве мирового справочного центра в секторе городских решений.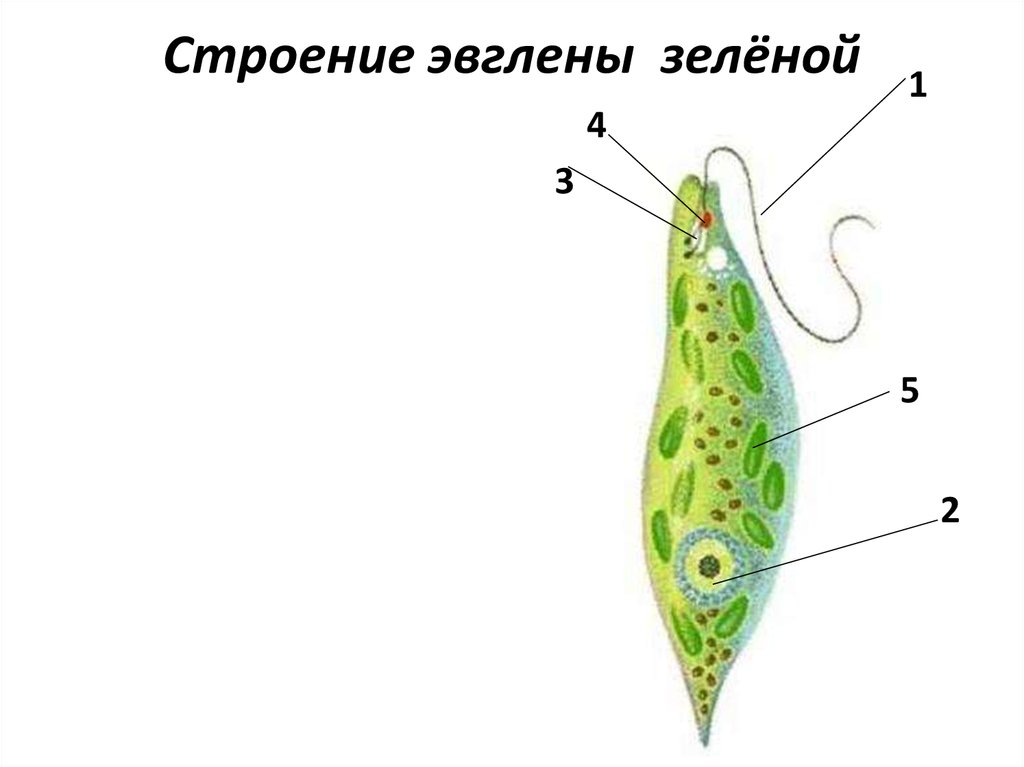
ЧТО ТАКОЕ ЗЕЛЕНОЕ СТРОИТЕЛЬСТВО И ЕГО ЗНАЧЕНИЕ ДЛЯ УСТОЙЧИВОГО ГОРОДСКОГО ПЛАНИРОВАНИЯ
В отличие от пригородных городов, возникших в результате промышленной революции и развития 20 го века, новые тенденции в городском планировании включают строительство эко-районов, которые представляют собой городские проекты, направленные на снижение воздействия на окружающую среду и изменение образа жизни горожан, чтобы сделать их более ответственными за свое окружение. Строительство зданий и др. Инфраструктура с использованием устойчивых технологий и материалов является ключом к этому типу инициатив.
Экологически чистое или устойчивое здание — это здание, которое благодаря своей конструкции и характеристикам может поддерживать или улучшать качество жизни в окружающей среде, в которой оно расположено. Для этого необходимо достичь высокого уровня эффективности: сокращение потребления энергии, воды и других ресурсов сводит к минимуму загрязнение.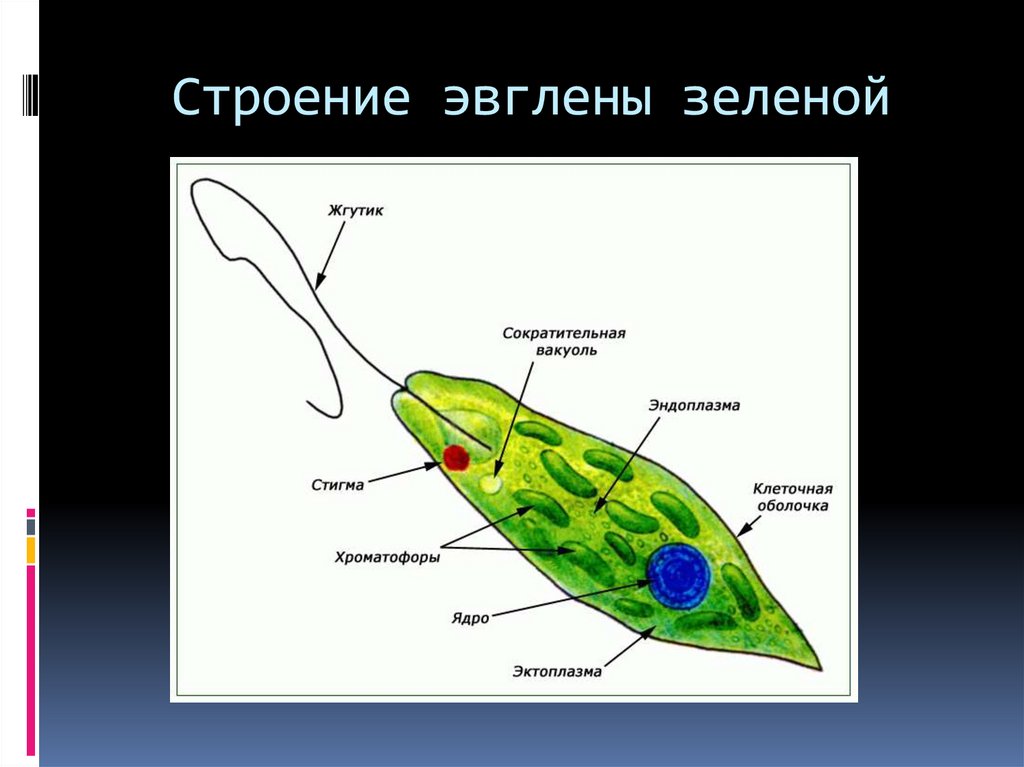 Сертификат LEED (Лидерство в энергетическом и экологическом дизайне) является международно признанным официальным признанием, которое устанавливает, заслуживает ли здание того, чтобы его считали экологически безопасным.
Сертификат LEED (Лидерство в энергетическом и экологическом дизайне) является международно признанным официальным признанием, которое устанавливает, заслуживает ли здание того, чтобы его считали экологически безопасным.
ХАРАКТЕРИСТИКИ ЗЕЛЕНЫХ ЗДАНИЙ
Для оценки устойчивости здания сертификация LEED устанавливает систему баллов, основанную на различных разделах, которые мы рассмотрим ниже, связанных с проектированием и строительством: местах и предоставить общественный транспорт, чтобы сократить использование личных автомобилей.
Устойчивые объекты
Защита и поддержание естественной среды обитания, сокращение загрязнения и использование природных ресурсов, а также содействие взаимодействию с природой.
Эффективное использование воды
Свести к минимуму использование воды во время строительства и предусмотреть механизмы для уменьшения водного следа здания.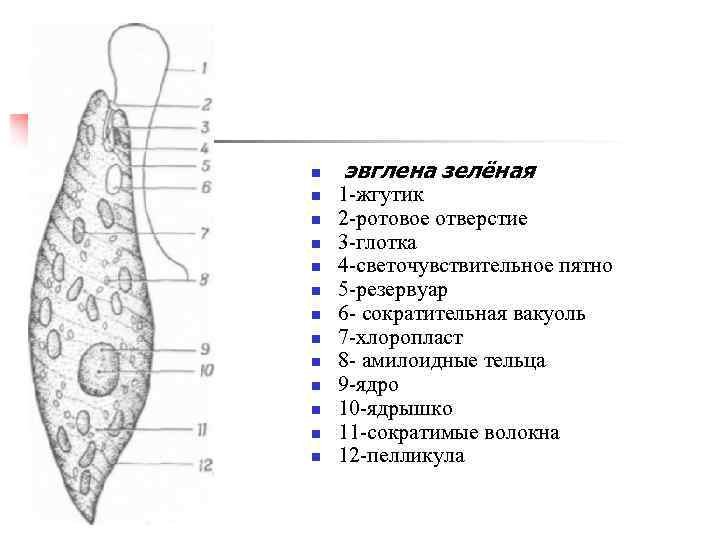
Энергия и атмосфера
Сократите потребление энергии, используйте возобновляемые источники энергии и повысьте энергоэффективность для уменьшения загрязнения.
Материалы и ресурсы
Внедряйте системы вторичной переработки, используйте экологичные материалы и максимально экономьте ресурсы во время строительства.
Качество окружающей среды в помещении
Обращайте внимание на качество помещения для его обитателей, например, на чистоту воздуха, температурный контроль и шумовое загрязнение.
Инновационный дизайн
Внедрение инновационных стратегий устойчивого развития во время строительства.
Региональный приоритет
Достижение улучшений в месте, где он расположен, с точки зрения окружающей среды, социальной справедливости и общественного здравоохранения.
ДЕСЯТЬ УСТОЙЧИВЫХ ЗДАНИЙ, КОТОРЫЕ УДИВЛЯЮТ МИР
Башня Ибердрола, Бильбао, Испания
Башня Ибердрола, Бильбао, Испания
Башня Ибердрола, Бильбао, Испания
Башня Ибердрола, Бильбао, Испания
Башня Ибердрола, Бильбао, Испания
Башня Ибердрола, Бильбао, Испания
В башне Iberdrola, спроектированной архитектором Сезаром Пелли, находится штаб-квартира группы, и благодаря своим характеристикам она представляет собой нулевую милю возобновляемой энергии. Его внушительная стеклянная конструкция высотой 165 метров является символом финансов и бизнеса в Бильбао и эталоном устойчивого дизайна и энергоэффективности. Он имеет кольцо рекуперации энергии, которое уменьшает его углеродный след.
Его внушительная стеклянная конструкция высотой 165 метров является символом финансов и бизнеса в Бильбао и эталоном устойчивого дизайна и энергоэффективности. Он имеет кольцо рекуперации энергии, которое уменьшает его углеродный след.
Здание было задумано как двигатель создания ценности для сотрудников компании, окружающей среды и клиентов. Внутренняя планировка штаб-квартиры способствует командной работе, обмену знаниями и организационному обучению в идеальном, экологически чистом пространстве, которое способствует операционной эффективности.
Виртуальный визит в башню Ибердрола Внешняя ссылка, открывается в новом окне.
Посетите веб-сайт Iberdrola Tower Внешняя ссылка, открывается в новом окне.
Публичная библиотека Бэйтоу, Тайбэй, Тайвань
Публичная библиотека Бэйтоу, Тайбэй, Тайвань.
Первая на Тайване зеленая библиотека находится в парке Бэйтоу в Тайбэе. Это двухэтажное здание спроектировано так, чтобы сократить потребление воды и электроэнергии благодаря большим окнам. Крыша частично покрыта фотогальваническими элементами для выработки энергии, а также для сбора дождевой воды для использования в туалетах.
Крыша частично покрыта фотогальваническими элементами для выработки энергии, а также для сбора дождевой воды для использования в туалетах.
Калифорнийская академия наук, Сан-Франциско, США
Калифорнийская академия наук, Сан-Франциско, США.
Этот научно-исследовательский институт и музей естественной истории был полностью реконструирован в 2008 году по проекту Ренцо Пьяно. Новое здание перерабатывает дождевую воду, использует фотоэлектрические панели, максимально использует естественное освещение и имеет зеленую крышу площадью один гектар, на которой растут миллионы растений, произрастающих в Калифорнии.
Всемирный торговый центр, Манама, Бахрейн
Всемирный торговый центр, Манама, Бахрейн
Этот комплекс из двух 240-метровых башен-близнецов расположен в городе Манама, столице Бахрейна, и является первым в мире небоскребом, в конструкцию которого интегрированы ветряные турбины. Вместе они покрывают 15% энергопотребления вышек. Оба здания в форме парусов, направляющих ветер, соединены тремя мостами, каждое из которых имеет 29турбина метрового диаметра.
Pixel Building, Мельбурн, Австралия
Pixel Building, Мельбурн, Австралия.
Первое в Австралии углеродно-нейтральное офисное здание. Возникнув как прототип офиса будущего, он имеет зеленую крышу с солнечными панелями для производства собственной энергии и хранит всю необходимую воду. Его необычный разноцветный фасад позволяет использовать вентиляцию и естественное освещение для минимизации энергопотребления.
Музей завтрашнего дня, Рио-де-Жанейро, Бразилия
Музей завтрашнего дня, Рио-де-Жанейро, Бразилия.
Этот музей науки, спроектированный испанским архитектором Сантьяго Калатравой и построенный рядом с пирсом Мауа, открылся в 2015 году. Решения в области устойчивого развития включают в себя мобильные солнечные панели, системы повторного использования дождевой воды и систему кондиционирования воздуха, в которой используется вода, поступающая из залива Гуанабара.
Turning Torso, Мальмё, Суэсия
Turning Torso, Мальмё, Суэсия.
Еще один проект Сантьяго Калатравы, вдохновленный человеческим торсом в движении. Он расположен на шведской стороне пролива Эресунн и его 190 метров делают его самым высоким небоскребом в Скандинавии. Он питается от возобновляемых источников энергии. Каждая квартира управляет своим потреблением тепла и воды, а кухни перерабатывают органические отходы для производства биогаза.
Parkroyal Collection, Сингапур, Сингапур
Parkroyal Collection, Сингапур, Сингапур.
Расположенный в центре Сингапура, этот роскошный отель имеет 15 000 м 2 садовых террас, называемых небесными садами, которые радуют посетителей. Сады спроектированы так, чтобы быть самодостаточными и потреблять минимальное количество энергии за счет использования солнечных батарей, датчиков движения и сбора дождевой воды для последующей переработки.