Муниципальное общеобразовательное учреждение- средняя общеобразовательная школа №17
Эволюционное строение и функции систем органов у животных. Эволюция строения и функции покровов тела. ( урок биологии 8 класс) Автор: Миткова Валентина Ивановна учитель биологии Цели урока: (слайд №2)
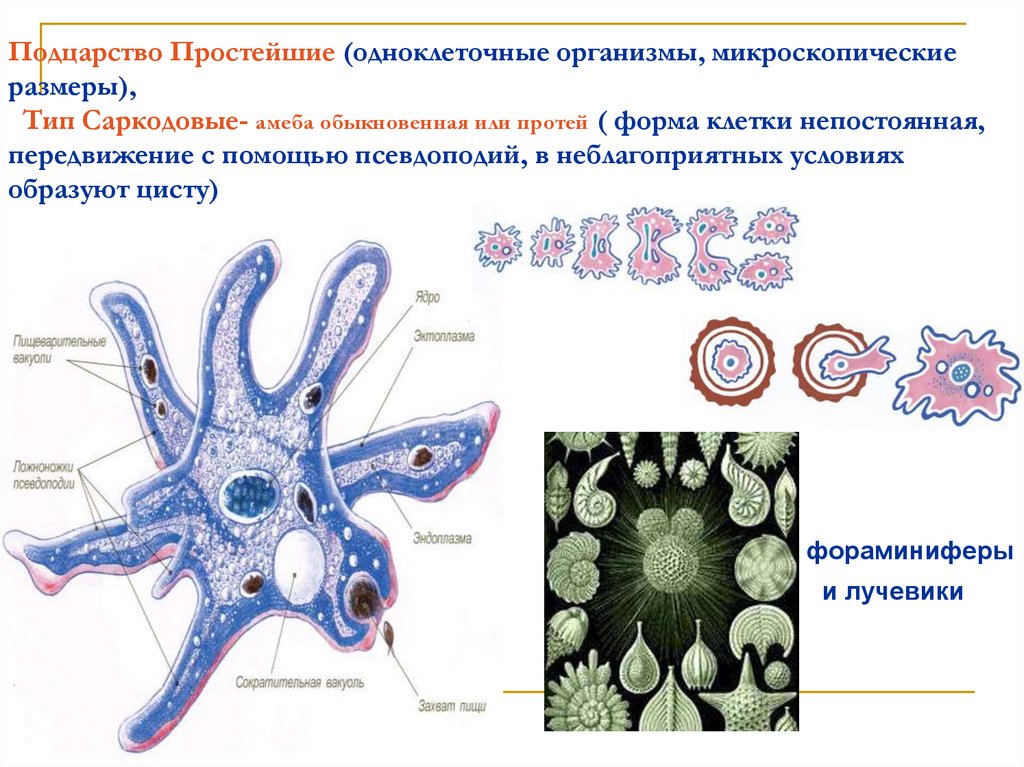
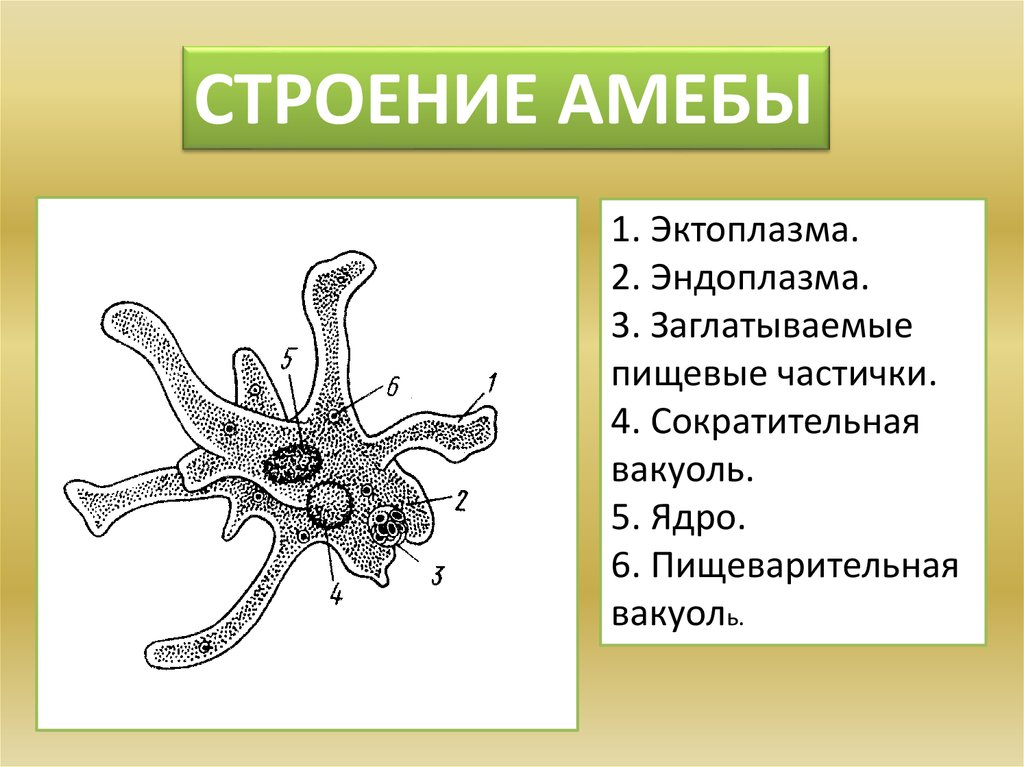
1. Изучить, как менялись отдельные черты строения животных; 2. Определить в каком направлении шла эволюция строения животных; Ход урока: На прошлом обобщающем уроке мы много говорили о многообразии животного мира. Но как возникло такое многообразие видов? Изучая многообразие животного мира мы особо обращали внимание на постепенное усложнение животных от низших форм к высшим, от простого к сложному. Они становились все более и более сложными по строению, поведению, образу жизни. Усложнялись и функции их тканей, органов, систем органов, в целом организм, то есть эволюционировали. (слад № 3) Все формы жизни, которые когда-либо появлялись на нашей планете, тесно связаны друг с другом, включая и человека – венца эволюции. Эволюционные изменения имели направления. (слайд №4) § ароморфоз – это изменение, ведущее к усложнению строения функций организма, повышающий общий уровень его организации. Ароморфозы всегда ведут к биологическому прогрессу. Они не являются прямым приспособлением к условиям среды. § идиоадаптация – мелкие эволюционные изменения, приспосабливающие организмы к конкретным условиям существования. Она возникает на базе ароморфозов. § дегенерация – эволюционное изменение, ведущее к упрощению организации жизни Что такое эволюция? «Это непрерывный, длительный, исторический путь развития животного мира» Ребята, а мы можем пронаблюдать эволюцию в лаборатории? «Нет. Это долгий путь» Ребята, как шла эволюция? «Животные усложнялись от низших форм к высшим» Все живые организмы постоянно ведут борьбу за жизнь, и выживает в этой борьбе сильнейший. Чтобы выжить, животные изменяются, усложняются, приспосабливаются к жизни. Как эволюционировали покровы тела у животных? Какие функции выполняют покровы тела? (слайд №6) Самыми примитивными животными являются – одноклеточные животные. (слайд №7) «Амеба обыкновенная не имеет постоянную форму тела, т.к. ее покровы представлены плазматической мембраной, у инфузории туфельки и зеленой эвглены уже появляется полупроницаемая оболочка пелликула, выполняющая защитную функцию, предотвращающая потерю воды» Шло время, и эволюционно от одноклеточных животных появились многоклеточные организмы. (слайд№8)Кто является самыми примитивными многоклеточными животными? «Тип Губки, тип Кишечнополостные» А как у них представлены покровы тела? «Это двухслойные животные. Эктодерма и энтодерма, их разделяет мезоглея. Наружный слой эктодерма представлен дифференцированными клетками: u эпителиально-покровные u эпителиально-мускульные (веретенообразные, вытянутые и представляют собой мускульное волокно, то есть впервые появляются мускульные клетке) u стрекательные клетки, ( защищают и участвую в нападении) u нервные клетки воспринимают возбуждения и передают их на эпителиально-мускульные клетки u половые клетки участвуют в размножении u недифференцированные клетки способны восстанавливать любые клетки. У кишечнополостных покровы тела усложняются, клетки наружного слоя дифференцируются. (слайд №9) Плоские черви. Покровы тела плоских червей устроены значительно сложнее, чем у кишечнополостных. Это трехслойные животные. Появляется мезодерма, производными ее являются мышечные волокна: кольцевые, продольные, диагональные, их функция – движение. Мышцы развиты слабо, поэтому их движения несложные, однообразные. Ресничный эпителий – это наружный слой клеток, он выполняет покровную и защитную функции. Ресничный эпителий и мышцы образуют кожно-мускульный мешок. Среди плоских червей имеются паразиты, у них тело покрыто кутикулой – производная эпителия, функция – защитная Круглые черви. Тело покрыто кутикулой, лишена ресничного эпителия, под кутикулой залегает слой продольных мышечных волокон. В отличие от плоских червей у них отсутствуют диагональные и кольцевые мышечные волокна, поэтому они могут лишь изгибаться. Кольчатые черви. Снаружи тело покрыто кутикулой, кожа ее вырабатывает кожный эпителий, под которым располагаются мышцы: кольцевые, продольные, брюшные, спинные. Эпителий и мышцы образуют кожно-мускульный мешок. Кожа выделяет слизь, которая обеспечивает скольжение. Только через влажную кожу происходит проникновение кислорода в тело червя. (слайд №10) Членистоногие. Утолщается кутикула, появляется хитиновый легкий покров. Кутикула более прочная, чем у кольчатых червей. Она имеет сложный химический состав, у некоторых он пропитан известью (ракообразные), не пропускает влагу, препятствует высыханию тела. Хитиновый покров – это наружный скелет, к которому прикреплены пучки мышц. У членистоногих появляется поперечно-полосатая мышечная ткань. Под кутикулой находятся железы: слюнные, паутинные, ядовитые, пахучие. Подобное усовершенствование покровов тела привело к повышению жизнеспособности предков членистоногих, помогла им заселить разнообразные участки земли, преодолеть неблагоприятные влияния среды обитания. Тело моллюсков покрыто мантией – кожная складка и раковиной. Между ними находится мантийная полость. Раковина состоит из трех слоев: роговой, фарфоровый, перламутровый. У некоторых моллюсков имеются вводные и выводные протоки (слайд №12) Хордовые. Ланцетник. Покровы представлены однослойным эпителием и тонким слоем собственно кожи. Земноводные. Кожа голая, лишена чешуй, гладкая, богата железами, которые вырабатывают слизь. Эта слизь предохраняет организм от высыхания, облегчает движение в воде, снимая трение. Слизь образует тонкую пленку, в которой растворяется кислород, это обеспечивает дополнительное дыхание. Некоторые железы выделяют слизь, защищая лягушек от воздействия болезнетворных бактерий. Пресмыкающиеся. Кожа сухая, покрыта роговыми чешуйками, кожных желез нет, сплошной роговой покров слабо растяжим и препятствует росту, поэтому происходит линька. Мускулатура представлена отдельными мышечными пучками. (слайд №15) Птицы. Кожа сухая без желез, покрыта пухом и перьями (пуховые, контурные). Перья – это видоизмененные чешуйки пресмыкающихся. Копчиковая железа выделяет маслянистую жидкость, которой птицы смазывают перья. Перьевой покров осуществляет теплоизоляцию и обеспечивает обтекаемость тела. Ежегодно происходит линька. Млекопитающие. Кожа имеет более сложное строение, снаружи расположен многослойные эпидермис, состоящий из рогового слоя, образованного мертвыми ороговевшими клетками, которые предохраняют организм от излишков потери влаги и базального слоя, клетки которого постоянно делятся. Итак, ребята, какой можно сделать вывод? Как развивались покровы тела у животных?
|
 Удивительно разнообразен животный мир.
Удивительно разнообразен животный мир.
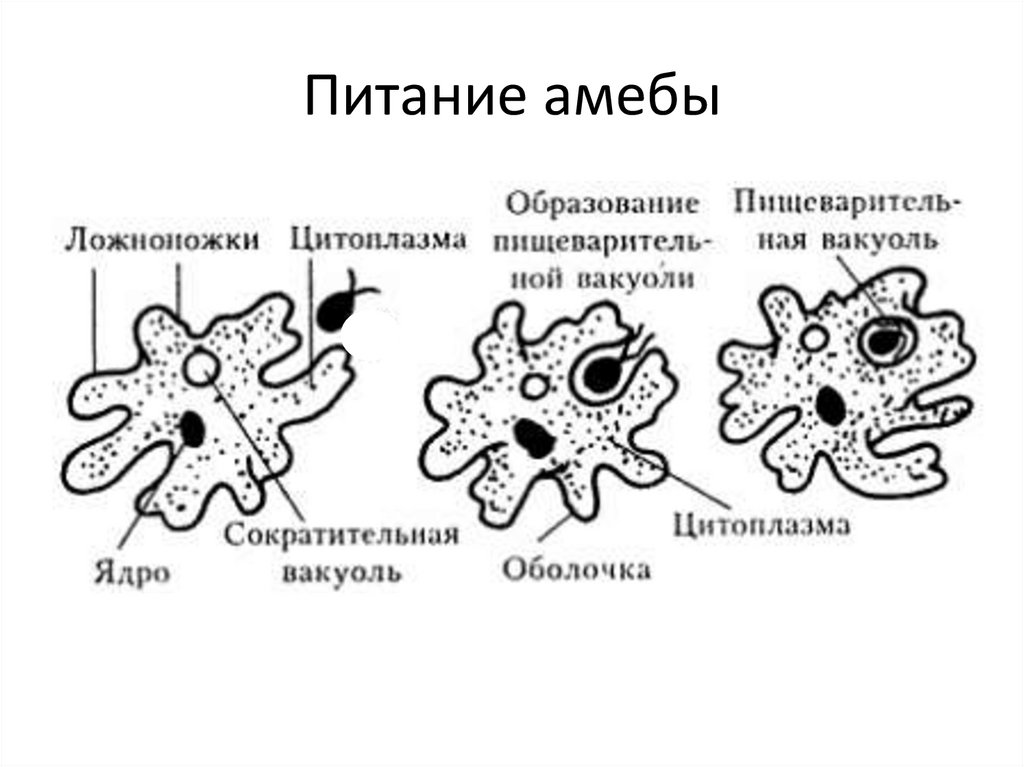
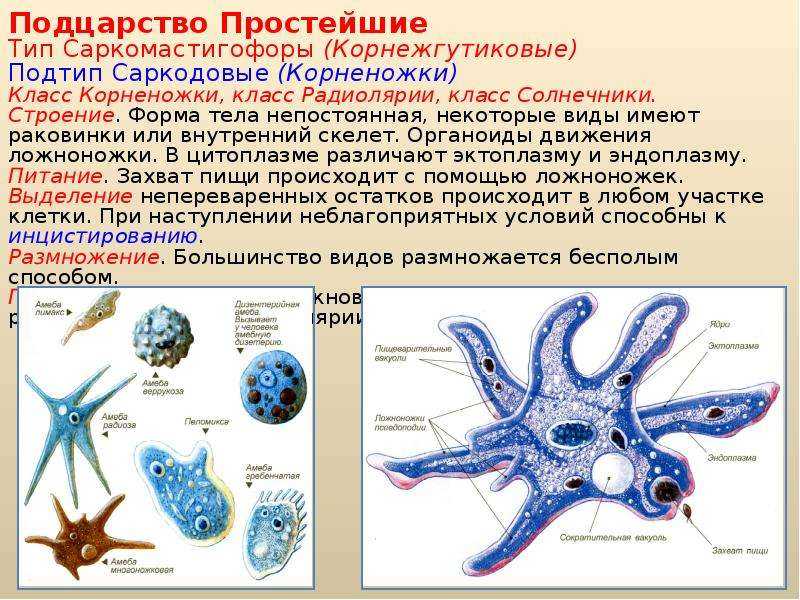 Это животные, у которых одна клетка – целый организм. Перед вами таблица№1. Внимательно рассмотрите покровы тела у простейших. Обратите внимание на эволюционные изменения в покровах тела одноклеточных.
Это животные, у которых одна клетка – целый организм. Перед вами таблица№1. Внимательно рассмотрите покровы тела у простейших. Обратите внимание на эволюционные изменения в покровах тела одноклеточных.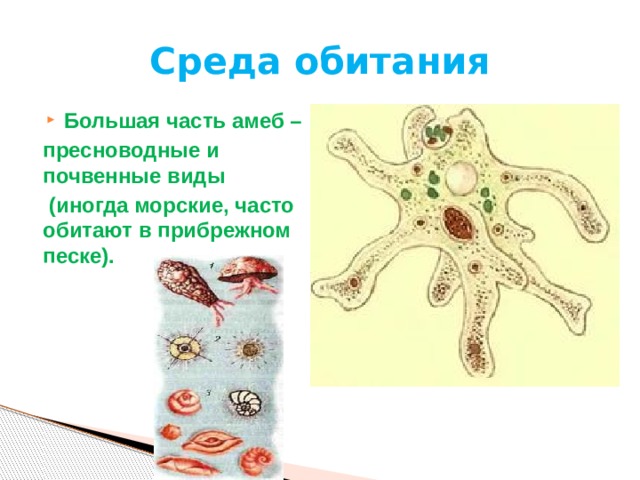


 (слайд №13)
(слайд №13)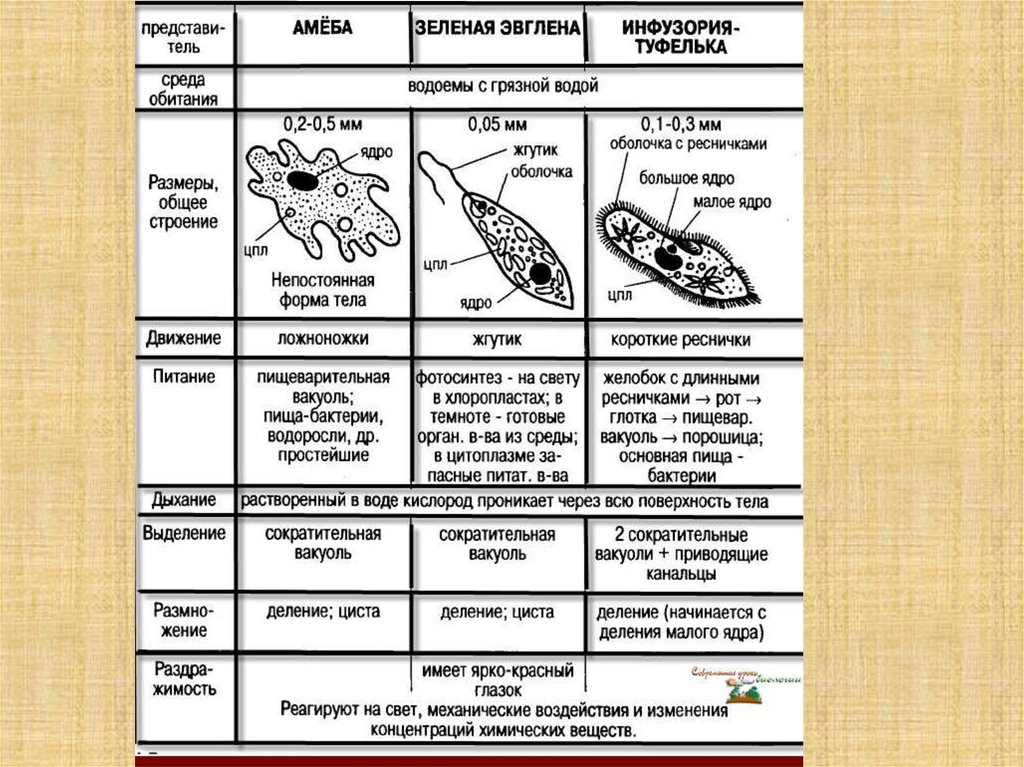
Задания для подготовки к ЕГЭ по биологии – конспект урока – Корпорация Российский учебник (издательство Дрофа – Вентана)
авторы: Наталья Александровна Куркина , учитель биологии, Волгоградская область
Разработки уроков (конспекты уроков)
Среднее общее образование
Биология
Внимание! Администрация сайта rosuchebnik. ru не несет ответственности за содержание методических разработок, а также за соответствие разработки ФГОС.
ru не несет ответственности за содержание методических разработок, а также за соответствие разработки ФГОС.
ЧАСТЬ 1
В заданиях 1-3 выберите три верных ответа из шести.
1. Примерами полового размножения организмов является
- Деление на части тела медузы
- Почкование гидры
- Партеногенез насекомых
- Созревание семян в плодах тыквы
- Яйцеживорождение акулы
- Митотическое деление хламидомонады
Ответ:
|
3 |
4 |
5 |
2. Поглощает кислород всей поверхность тела
- Дождевой червь
- Энцефалитный червь
- Речной рак
- Пресноводный полип гидра
- Саранча
- Амёба обыкновенная
Ответ:
|
1 |
4 |
6 |
3. Примерами полового диморфизма являются
Примерами полового диморфизма являются
- Желтые пятна в окраске покровов ужей
- Шпоры на ногах у петухов
- Образование чёрных и белых полос у зебр
- Обтекаемая форма тела речного окуня
- Яркое оперение самцов фазанов
- Сильно развитые бивни у самцов моржей
Ответ:
|
2 |
5 |
6 |
4. Установите соответствие между костью и отделом скелета, к которому она принадлежит.
|
КОСТЬ |
ОТДЕЛ СКЕЛЕТА |
|
А) ключица Б) плечевая В) грудина Г) лопатка Д) лучевая кость Е) ребро |
|
Ответ:
|
А |
Б |
В |
Г |
Д |
Е |
|
2 |
3 |
1 |
2 |
3 |
1 |
5. Установите соответствие между организмом и способом постэмбрионального развития, для которого он характерен.
Установите соответствие между организмом и способом постэмбрионального развития, для которого он характерен.
|
ОРГАНИЗМ |
СПОСОБ ПОСТЭМБРИОНАЛЬНОГО РАЗВИТИЯ
|
|
А) Речной окунь Б) Сизый голубь В) Уж обыкновенный Г) Остромордая лягушка Д) Прыткая ящерица |
|
Ответ:
|
А |
Б |
В |
Г |
Д |
|
2 |
1 |
1 |
2 |
1 |
ЧАСТЬ 2
Запишите полный развёрнутый ответ.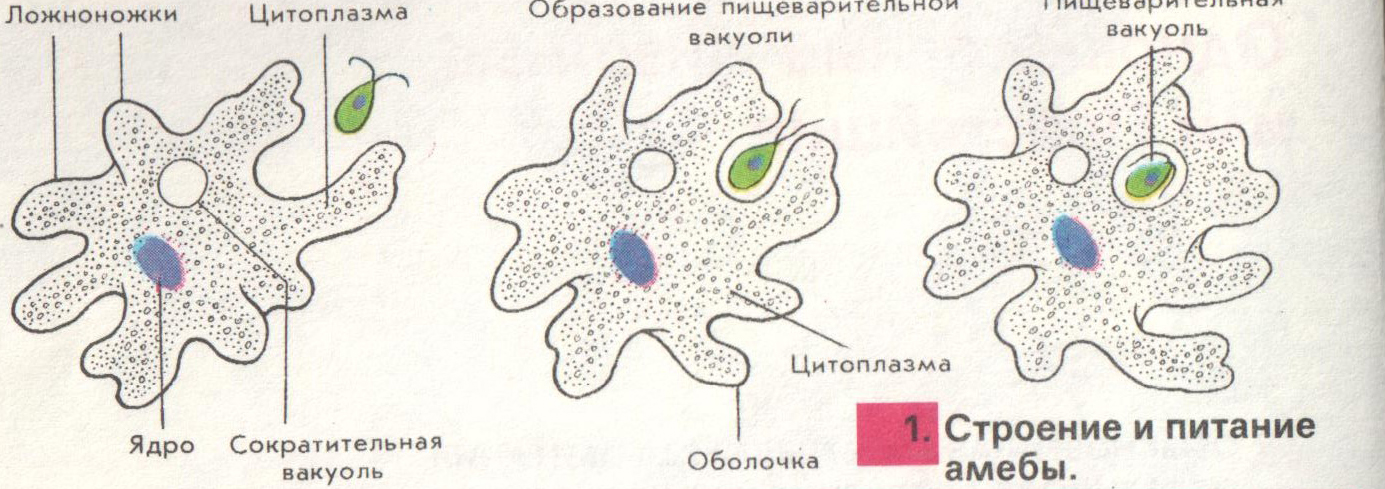
6. Объясните, в чём проявляется приспособленность пингвинов к жизни в условиях низких температур. Укажите не менее трёх признаков.
Ответ: Поддержание постоянной температуры тела, или теплокровность. Накопление жировых запасов, способствующих сохранению тепла. Роль теплоизолирующих покровов (пуха, пера).
7. Укажите основные признаки строения представителей Царства Грибов.
Ответ: Тело состоит из гифов, или нитей грибницы. Клетки грибов не содержат пластид. Они относятся к эукариотическим организмам. Снаружи цитоплазматическую мембрану клетки покрывает клеточная стенка, состоящая из хитина и целлюлозы.
8. В процессе трансляции был синтезирован белок, состоящий из 20 аминокислот. Определите, сколько нуклеотидов содержится в гене, который контролирует синтез белка, и в молекуле иРНК, на котором происходит сборка молекулы белка, а также число транспортных РНК, участвующих в биосинтезе.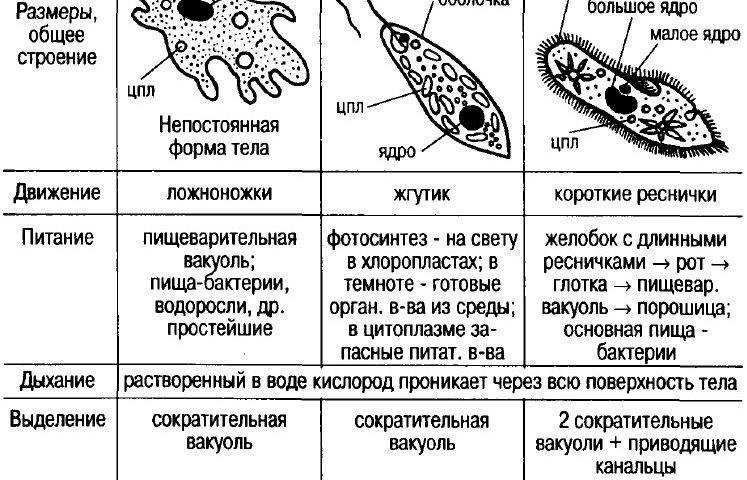
Ответ:
- Число нуклеотидов в гене равно 20 × 3 = 60.
- Число нуклеотидов и РНК равно числу нуклеотидов в гене, т.е. 60.
- Число тРНК равно числу аминокислот – 20.
9. Найдите ошибки в приведённом тексте. Укажите номера предложений, в которых сделаны ошибки, исправьте их.
1. В ротовой полости находятся зубы, которые развиваются в лунках челюстей. 2. У взрослого человека 32 зуба. 3. На каждой челюсти расположены: 4 резца, 2 клыка, 6 малых коренных и 4 больших коренных. 4. Каждый зуб состоит из двух частей: коронки и корней. 5. Зубы образованы эпителиальной тканью – дентином, снаружи покрыты эмалью.
Ответ: 3 – Малых коренных – 4, больших коренных – 6; 4 – У зуба три части: коронка, шейка и корни; 5 – Дентин и эмаль – разновидность соединительной ткани.
10. Форма тела богомола напоминает сухую веточку растения. Как сформировалась подобная форма тела у него в процессе эволюции?
Как сформировалась подобная форма тела у него в процессе эволюции?
Ответ: Появление у особей популяции разнообразных наследственных изменений. Сохранение естественным отбором особей с измененной формой тела. Размножение и распространение особей с формой тела, напоминающей сухую веточку, репродуктивная изоляция, формирование новой популяции.
Комментарий:
- Сначала рекомендуем учащимся ознакомиться с текстом параграфа, иллюстрациями и материалами видеоуроков по следующим темам: «Грибы. Строение и многообразие», «Эволюция строения и функций органов и дыхательной системы животных», «Опорно-двигательная система человека», «Пищеварительная система. Зубы», «Реализация наследственной информации», «Бесполое и половое размножение», «Индивидуальное развитие организма», «Факторы эволюции», «Адаптация организмов к условиям обитания».
- На консультации акцентируем внимание ребят на характерные черты объектов, особенности процессов и явлений, а также предлагаем широкий спектр примеров и процессов.
- В целях развития письменной и устной речи учащихся рекомендуем им составить логические схемы, кластеры, рассказ с использованием правильных ответов с последующей защитой своих работ в данной группе.
Хотите сохранить материал на будущее? Отправьте себе на почту
в избранноеТолько зарегистрированные пользователи могут добавлять в избранное.
Войдите, пожалуйста.
Какой формы амеба?
Какой формы амеба?
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 ./../../_base/nav_right2.gif» valign=»TOP»>
./../../_base/nav_right2.gif» valign=»TOP»>
Динамика цитоскелета Amoeba proteus
Allen RD (1968) Различия принципиального характера между несколькими типами амебоидного движения. Symp Soc Exp Biol 22:151–168
Google Scholar
Аллен Р.Д., Аллен Н.С. (1978) Цитоплазматический поток при амебоидном движении. Энн Рев Биофиз Биоэнг 7: 469–495
Google Scholar
Кэррол Р.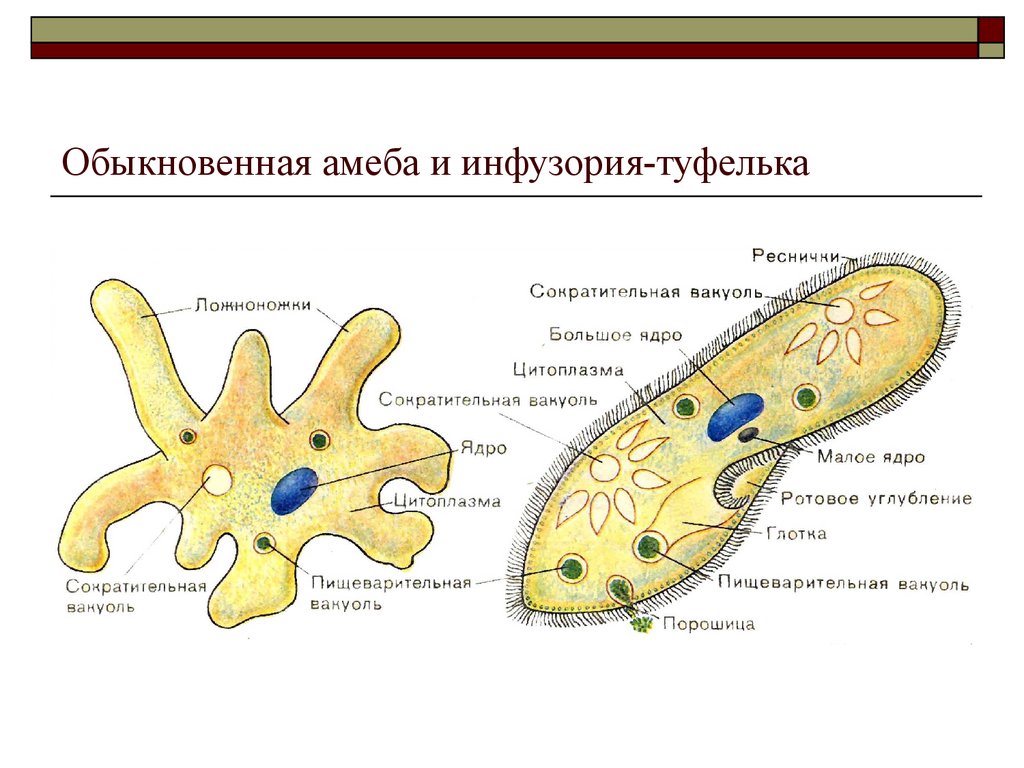 К., Батлер Р.Г., Моррис П.А., Джеррард Дж.А. (1982)Разборная сборка псевдоподального и сократительного цитоскелета тромбоцитов. Сотовый 30:385–393
К., Батлер Р.Г., Моррис П.А., Джеррард Дж.А. (1982)Разборная сборка псевдоподального и сократительного цитоскелета тромбоцитов. Сотовый 30:385–393
Google Scholar
Comly LT (1973) Микрофиламенты в Chaos carolinensis : мембранная ассоциация, распределение и связывание тяжелого меромиозина в глицеринированной клетке. J Cell Biol 58:230–237
Google Scholar
Feramisco JR (1979) Микроинъекция флуоресцентно меченого α-актинина в живые фибробласты. Proc Natl Acad Sci USA 76:3967–3971
Google Scholar
Gawlitta W, Hinssen H, Stockem W (1980a) Влияние актинмодулирующего белка (AM-белка) из Physarum polycephalum на подвижность клеток Amoeba proteus. Eur J Cell Biol 23:43–52
Google Scholar
Gawlitta W, Stockem W, Wehland J, Weber K (1980b) Организация и пространственное расположение меченого флуоресцеином нативного актина, микроинъецированного в нормальное передвижение и экспериментально воздействовавшего на Amoeba proteus. Анализ клеточных тканей 206:181–191
Анализ клеточных тканей 206:181–191
Google Scholar
Geiger B (1979) Белок 130 К из куриного желудка: его локализация на концах пучков микрофиламентов в культивируемых куриных клетках. Сотовый 18:193–205
Google Scholar
Grebecka L (1980) Изменение двигательной полярности Amoeba proteus путем всасывания. Протоплазма 102:361–375
Google Scholar
Grebecka L (1981) Двигательные эффекты перфорации слоев периферических клеток Amoeba proteus. Протоплазма 106:343–349
Google Scholar
Grebecka L, Grebecki A (1975) Морфометрическое исследование движущихся Amoeba proteus. Acta Protozool 14:337–361
Google Scholar
Grebecka L, Hrebenda B (1979) Топография коркового слоя у Amoeba proteus в связи с динамической морфологией движущейся клетки.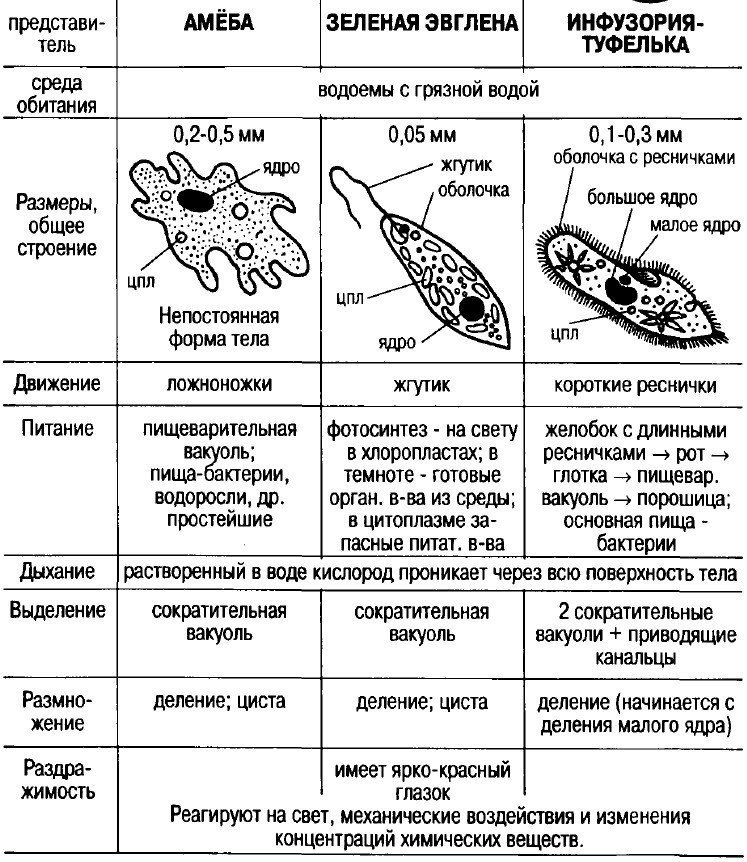 Acta Protozool 18:481–490
Acta Protozool 18:481–490
Google Scholar
Гребецкий А (1976) Соосное движение полужесткого клеточного каркаса у Amoeba proteus. Acta Protozool 15:221–248
Google Scholar
Grebecki A (1977) Неосевые движения клеточного каркаса и передвижение Amoeba proteus. Acta Protozool 16:53–85
Google Scholar
Grebecki A (1979) Организация моторных функций у амеб и плазмодиев слизевиков. Acta Protozool 18: 43–58
Google Scholar
Grebecki A (1980) Поведение Amoeba proteus при изменении светотени. Протистология 16:103–113
Google Scholar
Grebecki A (1981) Влияние локализованной фотостимуляции на амебоидное движение и их теоретические последствия. Eur J Cell Biol 24:163–175
Google Scholar
Grebecki A (1982) Надмолекулярные аспекты амебоидного движения.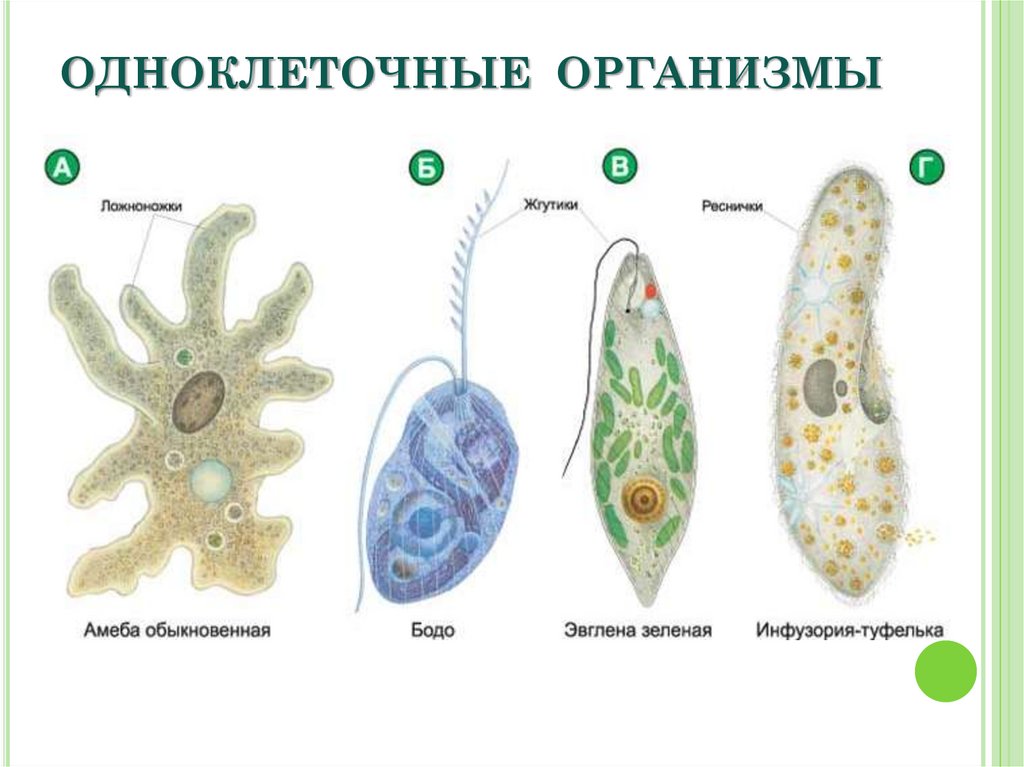 Progr in Protozool, Proc VI Internatl Congr Protozool 1:117–130
Progr in Protozool, Proc VI Internatl Congr Protozool 1:117–130
Google Scholar
Grebecki A, Grebecka L (1978) Морфодинамические типы Amoeba proteus : терминологическое предложение. Протистология 14:349–358
Google Scholar
Гребецкий Л., Клопока В. (1981) Функциональная взаимозависимость псевдоподий у Amoeba proteus , стимулированная светотеневой разницей. J Cell Sci 50: 245–258
Google Scholar
Haberey M (1973) Räumliche Anordnung von Plasmafilamenten bei Thecamoeba sphaeronucleolus. Цитобиология 8:61–75
Google Scholar
Хабери М., Стокем В. (1971) Amoeba proteus : Морфология, Zucht und Verhalten. Микрокосмос 60:33–42
Google Scholar
Hauser M (1978) Демонстрация ассоциированных с мембраной и ориентированных микрофиламентов у Amoeba proteus с помощью фиксатора основания Шиффа/глутарового альдегида. Цитобиология 18:95–106
Цитобиология 18:95–106
Google Scholar
Hoffmann U, Stockem W, Gruber B (в процессе подготовки) Динамика цитоскелета в Amoeba proteus : Влияние различных агентов на пространственную организацию микроинъецированного актина, меченого IAF
Jeon KW, Jeon MS (1982) Создание механической силы во время фагоцитоза у амеб. J Cell Biol 95:312a
Google Scholar
Keith CH, Feramisco JR, Shelanski M (1981) Прямая визуализация меченных флуоресцеином микротрубочек in vitro и в микроинъецированных фибробластах. J Cell Biol 88: 234–240
Google Scholar
Klopocka W, Grebecki A (1980) Моторная взаимозависимость псевдоподий у свободно перемещающихся Amoeba proteus. Acta Protozool 19:129–142
Google Scholar
Klopocka W, Grebecki A (1982) Передвижение Amoeba proteus после стандартизации формы его тела. Протоплазма 112:37–45
Протоплазма 112:37–45
Google Scholar
Komnick H, Wohlfahrt-Bottermann KE (1965) Das Grundplasma und die Plasmafilamente der Amoebe Хаос Хаос nach enzymatischer Behandlung der Zellmembran. З Целлфорш 66: 434–456
Google Scholar
Корн Э.Д. (1982) Полимеризация актина и ее регуляция белками немышечных клеток. Физиол Ред. 62:672–737
Google Scholar
Korohoda W, Stockem W (1975) О природе гиалиновых зон в цитоплазме Amoeba proteus . Microsc Acta 77:129–141
Google Scholar
Kreis TE, Birchmeier W (1980) Саркомеры стрессовых волокон фибробластов сократительны. Ячейка 22:555–561
Статья КАС пабмед Google Scholar
Kreis TE, Birchmeier W (1982) Микроинъекция флуоресцентно меченных белков в живые клетки с упором на белки цитоскелета. Инт Рев Цитол 75:209–227
Инт Рев Цитол 75:209–227
Google Scholar
Lazarides E, Burridge E (1975) α-Actinin: иммунофлуоресцентная локализация актиновых филаментов в немышечных клетках. Сотовый 6: 289–298
Google Scholar
Мачта SO (1932) Локализованная стимуляция, передача импульсов и характер реакции у Amoeba. Физиол Зоол 5:1–15
Google Scholar
Maupin-Szamier P, Pollard TD (1978) Разрушение актиновых филаментов четырехокисью осмия. J Cell Biol 77:837–852
Google Scholar
Нахмиас В.Т. (1964) Фибриллярные структуры в цитоплазме Хаос хаоса. J Cell Biol 23:183–188
Google Scholar
Нахмиас В.Т. (1968) Дальнейшие электронно-микроскопические исследования фибриллярной организации основной цитоплазмы Хаос Хаос .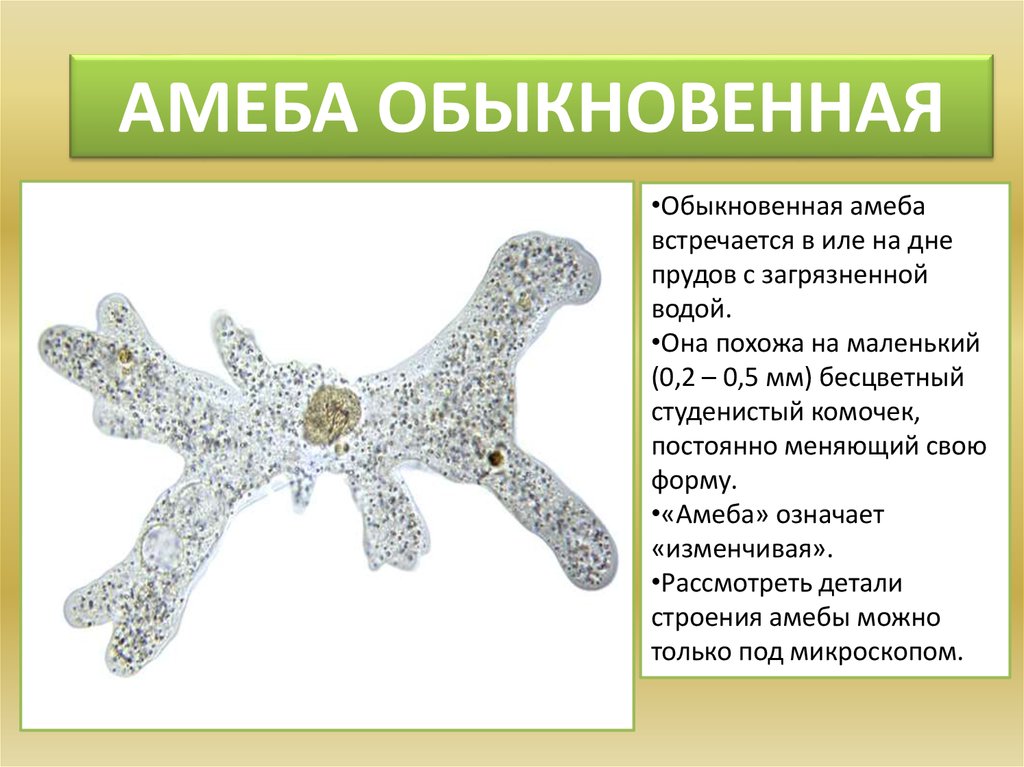 J Cell Biol 38:40–52
J Cell Biol 38:40–52
Google Scholar
Pollard TD, Ito S (1970) Цитоплазматические филаменты у Amoeba proteus . I. Роль филаментов в изменении консистенции и движении. J Cell Biol 46: 267–289
Google Scholar
Pollard TD, Korn ED (1973) Электронно-микроскопическая идентификация актина, связанного с изолированными плазматическими мембранами амеб. Дж. Биол. Хим. 248:448–450
Google Scholar
Rinaldi RA, Hrebenda B (1975) Ориентированные толстые и тонкие нити у Amoeba proteus. J Cell Biol 66:193–198
Google Scholar
Sanger JW, Sanger JM, Kreis TE, Jockusch BM (1980) Обратимая транслокация актина цитоплазмы в ядро, вызванная диметилсульфоксидом. Proc Natl Acad Sci USA 77:5268–5272
Google Scholar
Satoh H, Ueda T, Kobatake Y (1982) Первичный осциллятор ритма сокращения в плазмодии Physarum polycephalum : Роль митохондрий. Cell Struc Func 7:275–283
Cell Struc Func 7:275–283
Google Scholar
Schäfer-Danneel S (1967) Strukturelle und funktionelle Voraussetzungen für die Bewegung von Amoeba proteus . Z Целлфорш 78: 441–462
Google Scholar
Stacey DW, Allfrey VG (1977) Доказательства аутофагии микроинъецированных белков в клетках Hela. J Cell Biol 75:807–817
Google Scholar
Stockem W, Weber K, Wehland J (1978) Влияние микроинъекции фаллоидина на локомоцию, протоплазматический поток и цитоплазматическую организацию у Amoeba proteus и Physarum polycephalum. Цитобиология 18:114–131
Google Scholar
Стокем В., Хоффманн Х.У., Гавлитта В. (1981) III. Амебоидное движение: Морфофункциональные основы амебоидного движения. Verh Dtsch Zool Ges 71–84
Stockem W, Hoffmann HU, Gawlitta W (1982) Пространственная организация и тонкая структура слоя кортикальных нитей при нормальном передвижении Amoeba proteus. Соотношение клеток и тканей 221:505–519
Соотношение клеток и тканей 221:505–519
Google Scholar
Стокем В., Наиб-Маджани В., Вольфарт-Боттерманн К.Е., Осборн М., Вебер К. (1983) Пиноцитоз и локомоция амеб. XIX. Иммуноцитохимическая демонстрация актина и миозина у Amoeba proteus . Eur J Cell Biol 29:171–178
Google Scholar
Taylor DL, Condeelis JS (1979) Цитоплазматическая структура и сократимость в амебоидных клетках. Int Rev Cytol 56:57–144
Google Scholar
Taylor DL, Wang YL (1978) Молекулярная цитохимия: включение флуоресцентно меченного актина в живые клетки. Proc Natl Acad Sci USA 75:857–861
Google Scholar
Taylor DL, Wang YL, Heiple JM (1980) Сократительная основа амебоидного движения. VII. Распределение флуоресцентно меченого актина в живых амебах. J Cell Biol 86:590–598
Google Scholar
Wang YL, Taylor DL (1979) Распределение флуоресцентно меченого актина в живых яйцах морских ежей на раннем этапе развития.