Эти загадочные Простейшие
Дидактическая цель: cоздать условия для закрепления и осмысления блока информации посредством использования исследовательского метода обучения.
Учебная цель: способствовать осознанию, взаимосвязи между строением живой системы и ее функциями; развивать самостоятельность мышления, умение устанавливать причинно-следственные связи, обобщать, делать выводы; воспитывать потребность в бережном отношении к жизни.
Средства обучения: дидактические материалы для организации самостоятельной работы учащихся, микроскопы, готовые микропрепараты, мультимедийный проектор.
Ход мастерской
I. Актуализация знаний. Введение в тему
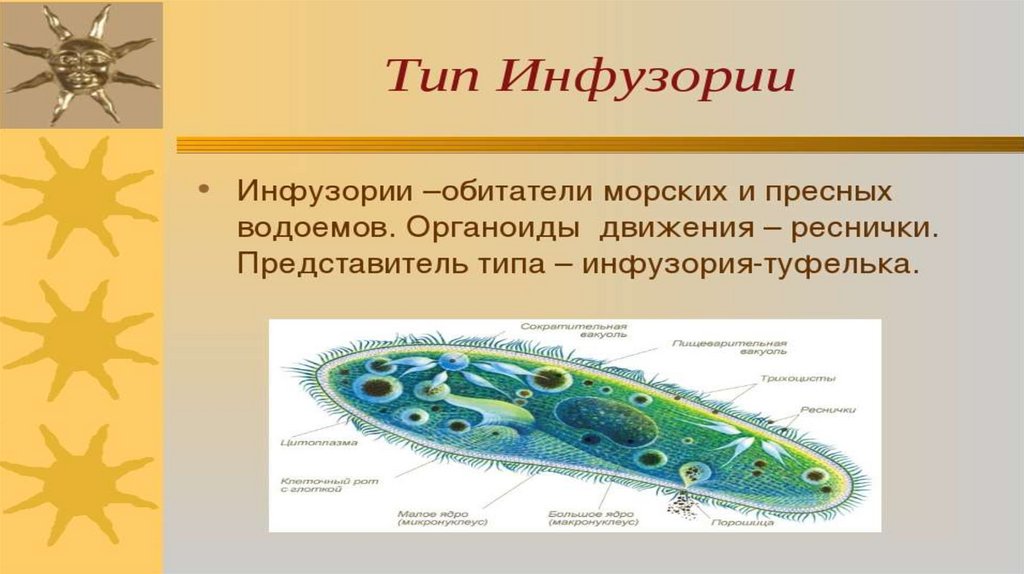
Учитель: Мы с вами изучили особенности подцарства Простейшие. Это особенное подцарство животных. В чем же особенности представителей этого подцарства (3 мин.)
Высказываются мнения:
- Организм простейших состоит из одной клетки.

- Клетка выполняет все функции многоклеточного организма.
- Клетка состоит из органоидов, которые выполняют определенные функции.
II. Ребята рассаживаются в группы по 5-6 человек
Вопрос учителя: Что вы знаете, а хотели бы узнать о Простейших? (3 мин.)
Учащиеся по группам на листе бумаги записывают свои вопросы. Учитель собирает листочки и раскладывает их на группы: тип Саркодовые, тип Жгутиконосцы, тип Инфузории.
III. Каждая группа получает задание на отдельном листе бумаги. На выполнение этого задания выделяется 10 минут.
Задание для группы № 1
1. Каковы общие признаки простейших. Какие функции выполняют органоиды клетки Простейших?
Заполните таблицу “Функции органоидов клетки простейших”
Органоиды клетки |
Функции органоидов |
Задание для группы № 2
1. Какую роль играют простейшие в природе и
жизни человека?
Какую роль играют простейшие в природе и
жизни человека?
Паразитические одноклеточные животные.
Значение простейших
| Значение | Корненожки | Жгутиковые | Инфузории |
| Положительное | |||
| Отрицательное |
Задание для группы № 3
1. Какими способами могут размножаться
простейшие? Охарактеризуйте эти способы
размножения. Составьте схемы размножений.
Составьте схемы размножений.
Задание для группы № 4
1. На примере вольвокса поясните, в чем появляется усложнение организации колониальных форм животных по сравнению с одноклеточными.
Задание для группы № 5
Составьте кроссворд к теме “Простейшие” (10-15 слов)
IV. Каждая группа вывешивает результаты работы и старший от группы выступает по результатам работы, отвечает на поставленные учащимися класса вопросы (группа помогает) – 15 мин.
V. Сообщение заранее подготовленного ученика о паразитических простейших. – 3 мин.
VI. Выполнение тестовой работы (индивидуально)
Выпишите по порядку цифры, вслед за которыми даны сведения об инфузории – туфельке (I – вариант), об амебе обыкновенной (II- вариант) – 5 мин.
1. Форма тела постоянная.
2. Передвигается при помощи многочисленных ресничек тупым концом вперед.
3. Питается бактериями, мельчайшими
водорослями, простейшими.
4. Может питаться растворенными в воде органическими и неорганическими веществами. В питании принимают участие хлоропласты.
5. Жидкие продукты жизнедеятельности и избыток воды удаляются через две сократительные вакуоли.
6. Переваривание пищи происходит в пищеварительных вакуолях.
7 При неблагоприятных условиях превращается в цисту.
8. В цитоплазме одно ядро.
9. В цитоплазме два ядра — большое и малое.
VII. Учащиеся работают с готовыми микропрепаратами по заданию: “Определить вид простейшего по микропрепарату” — 3 мин.
Учитель проверяет работу учеников, консультирует их.
VIII. Подведение итогов урока, 2-3 мин.
Вопросы учащимися:
— Что нового вы узнали на уроке?
— Что нового вы узнали о себе?
— Ваши пожелания на будущее.
IX. Задание на дом (по выбору учащихся) — 2-3 мин.
- Написать сочинение на тему “Природа без
простейших”.
 (Что изменилось бы в окружающей
среде, если бы исчезли простейшие?).
(Что изменилось бы в окружающей
среде, если бы исчезли простейшие?). - Составьте кроссворд к теме “Простейшие”.
- Составить игру “Лишнее слово”.
Дидактический материал для учителя
Задание для группы № 1
| Органоиды клетки | Функции органоидов |
| Ядро | Регулирует все процессы жизнедеятельности |
| Цитоплазма | В ней притекают все процессы обмена веществ |
| Сократительная вакуоль | |
| Цитоплазматическая мембрана | Отграничивает клетку от окружающей среды |
| Ложноножки | Органоиды передвижение амебы |
| Респики | Органоиды передвижение инфузории – туфельки |
| Жгутики | Органоиды передвижение эвглены зелено |
| Пищеварительная вакуоль | Переваривание пищи |
Задание для группы № 2
Значение простейших
| Значение | Корненожки | Жгутиковые | Инфузории |
| Положительное | Компонент биоценоза в цепи питания, морские корненожки имеют известковую раковину – образуют осадочные горные породы – мел, известняк; по некоторым видам корненожек судят о присутствии нефти | Компонент биоценоза в цепи питания, имеет познавательное значение для изучения общих признаков растений и животных | Компонент биоценоза в цепи питания |
| Отрицательное | Дизентерийная амеба вызывает
амебную дизентерию. Малярийный плазмодий вызывает малярию. |
Вызывают цветение воды в
водоемах. Трипаносомы паразитируют в крови и
спинномозговой жидкости позвоночных. Вызывают
сонную болезнь, а также нагану – болезнь
крупного рогатого скота. Лямблии паразитируют в кишечнике млекопитающих, вызывая болезнь лямблиоз. |
Паразитическая инфузория балантидий вызывает у человека тяжелые, напоминающее дизентерию расстройства. Она также может паразитировать в кишечнике свиней, которая не приносит вреда, но ими может заразиться человек. |
Задание для группы № 3
Для животных подцарства Простейшие характерны два способа размножения: бесполое и половое.
При бесполом размножении (характерно для типов
(Саркодовые, Жгутиконосцы, Инфузорий) происходит
деление клетки надвое. Сначала делится ядро
клетки, а затем на теле появляются перетяжка,
которая делит его на две примерно равные части, в
каждой из которых оказывается по одному ядру.
Сначала делится ядро
клетки, а затем на теле появляются перетяжка,
которая делит его на две примерно равные части, в
каждой из которых оказывается по одному ядру.
Для представителей типа Инфузорий характерен и половой способ размножения – конъюгация. При половом процессе увеличения числа особей не происходит. Две инфузории временно соединяются друг с другом. На месте соприкосновения пелликула растворяется, и между животными образуется соединительный мостик из цитоплазмы. Большое ядро каждой инфузории исчезает. Малое ядро дважды делится, в результате чего в каждой инфузории получается 4 дочерних ядра. Три из них размножаются, а четвертое снова делится. В результате в каждой инфузории остается по два ядра.
Одно из этих ядер по цитоплазматическому
мостику переходит в другую инфузорию и там
сливается с неподвижным ядром. Затем в каждой
инфузории из этого вновь образовавшегося ядра
формируются большое и малое ядро и инфузории
расходятся. Половой процесс ведет к обновлению
генетического материала, что увеличивает
жизнестойкость организмов.
Половой процесс ведет к обновлению
генетического материала, что увеличивает
жизнестойкость организмов.
Задание для группы № 4
Вольвокс может служить моделью, показывающей, как из одноклеточных организмов могли произойти многоклеточные.
Тело колониальных жгутиконосцев состоят из многих клеток. Первые колонии возникают, вероятно, вследствие того, что после деления клетки не расходятся, а остаются вместе. Число клеток в колонии вольвокса может достигать более 60 тысяч, внутренняя полость его шара занята жидкой слизью.
В колонии вольвокса существуют различные типы клеток (отвечающие за питание, движение, размножение), что характерно для многоклеточных.
.
Задание для группы № 5
Рекомендуемые для кроссворда слова: амеба,
клетка, цитоплазма, ядро, циста, органоид,
конъюгация, колония, жгутик, ложноножка,
раздражимость, выделение, эвглена, вольвокс,
радиолярии, лучевики, пелликула, эндоплазма,
эктоплазма.
Сообщение учащегося о болезнетворных простейших
В организме человека паразитирует более 30 видов простейших.
В человеке может паразитировать несколько видов плазмодия, вызывающих разные формы малярии. Попадая в кровь человека со слюной комара из рода Anopheles, возбудитель малярии разносится по всему телу, задерживаясь в печени, а затем переселяются в эритроциты, где питается гемоглобином. Сразу во многих красных клетках крови идет размножение паразитов. Затем все они одновременно разрывают оболочку эритроцитов и выходят в кровяное русло. Это и вызывает приступ лихорадки. После того как паразиты внедряются в новые эритроциты, температура тела больного нормализуется до их следующего массового выхода.
Возбудителями ряда опасных болезней человека и
животных являются паразитические жгутиконосцы.
Один из видов трипаносом вызывает “сонную
болезнь”, от которой в Африке ежегодно умирают
тысячи людей. Природный резервуар этого паразита
– африканские антилопы, совершенно не
страдающие от присутствия в их крови
жгутиконосцев. Переносчиками трипаносом
являются кровососущие мухи цеце. Трипаносомы
проникают в кровь, лимфатические железы, а затем
и в спинномозговую жидкость. При отсутствии
лечения “сонная болезнь” всегда приводит к
смерти.
Природный резервуар этого паразита
– африканские антилопы, совершенно не
страдающие от присутствия в их крови
жгутиконосцев. Переносчиками трипаносом
являются кровососущие мухи цеце. Трипаносомы
проникают в кровь, лимфатические железы, а затем
и в спинномозговую жидкость. При отсутствии
лечения “сонная болезнь” всегда приводит к
смерти.
Ответы к тестовой работе.
I вариант (инфузория туфелька) – 2, 3, 5, 8, 9, 11, 13
II вариант (амеба обыкновенная) – 1, 4, 5, 7, 9, 11, 12.
Отчет по биологии строение инфузории туфельки | Шпаргалки Биология
Скачай Отчет по биологии строение инфузории туфельки и еще Шпаргалки в формате PDF Биология только на Docsity! МИНИСТЕРСТВО НАУКИ И ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ Федеральное государственное бюджетное образовательное учреждение высшего образования «Санкт-Петербургский Горный Университет» Кафедра Геоэкологии Лабораторная работа «ВОДОРОСЛИ. ИЗУЧЕНИЕ И ОПРЕДЕЛЕНИЕ ОТДЕЛОВ НА ЖИВОМ ОБРАЗЦЕ» По дисциплине: Биология Выполнил: студентка группы ИЗБ-19-1 Мустафеева К.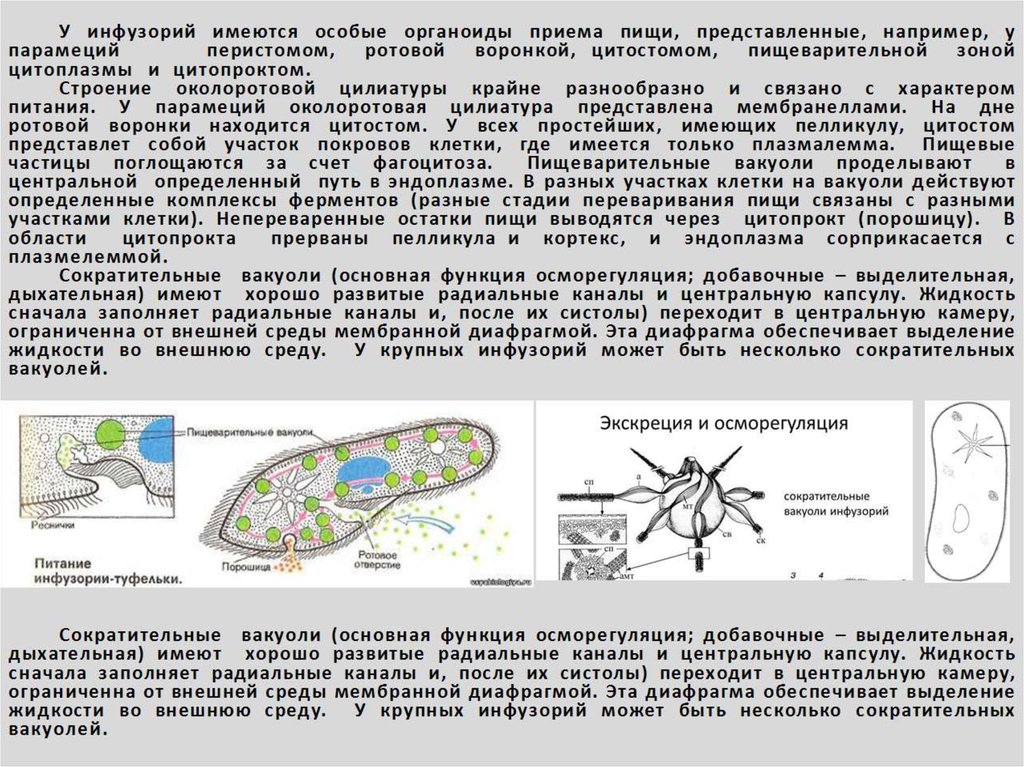 П. Проверил: доцент Петров Д.С. Санкт-Петербург 2019 Цель работы Изучение водоророслей и определение их на конкретном образце. Общие сведения Гетерогенная экологическая группа преимущественно фототрофных (если учитывать цианобактерии) одноклеточных, а также колониальных или многоклеточных организмов, обитающих, как правило, в водной среде, в систематическом отношении представляющая собой совокупность многих отделов. В совокупности с грибами образуют самостоятельную группу организмов, называемую лишайниками. Наука о водорослях называется альгологией. Насчитывается около 30 тыс видов водорослей, обитающих в водной, почвенной и наземно- воздушной среде. Размеры колеблются от 1 мкм до 30-60 м. Форма тела весьма разнообразна, одноклеточные могут иметь как непостоянную амебоидную форму клетки, так и продолговато- овальную с одним или двумя жгутиками. Многоклеточные формы представлены в основном нитчатыми формами, однако в результате деления нитей в разных плоскостях образуется и пластинчатая форма.
П. Проверил: доцент Петров Д.С. Санкт-Петербург 2019 Цель работы Изучение водоророслей и определение их на конкретном образце. Общие сведения Гетерогенная экологическая группа преимущественно фототрофных (если учитывать цианобактерии) одноклеточных, а также колониальных или многоклеточных организмов, обитающих, как правило, в водной среде, в систематическом отношении представляющая собой совокупность многих отделов. В совокупности с грибами образуют самостоятельную группу организмов, называемую лишайниками. Наука о водорослях называется альгологией. Насчитывается около 30 тыс видов водорослей, обитающих в водной, почвенной и наземно- воздушной среде. Размеры колеблются от 1 мкм до 30-60 м. Форма тела весьма разнообразна, одноклеточные могут иметь как непостоянную амебоидную форму клетки, так и продолговато- овальную с одним или двумя жгутиками. Многоклеточные формы представлены в основном нитчатыми формами, однако в результате деления нитей в разных плоскостях образуется и пластинчатая форма.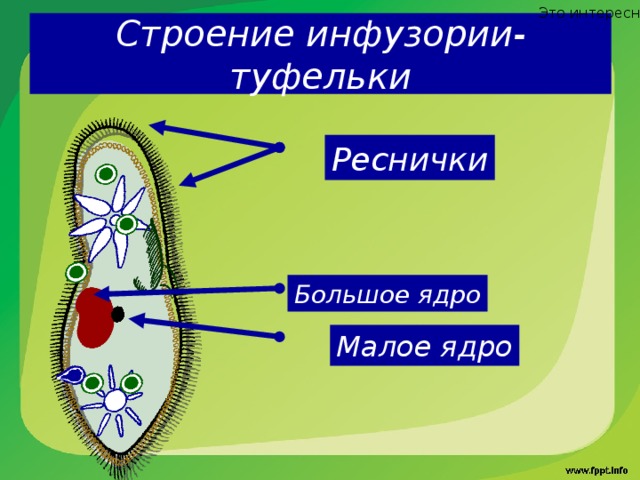 У ряда водорослей слоевища расчленены и напоминают побег с ветками и отходящими книзу ризоидами, служащими для прикрепления к грунту. Рассмотрим особенности строения водорослей. 1. Одноклеточные формы Клетки большинства водорослей покрыты плотной оболочкой, которая содержит целлюлозу (у эвгленовых оболочка содержит, в основном, белки). У некоторых водорослей оболочка пропитывается солями железа, кальция или кремния и напоминает панцирь. Отличительная черта клеток водорослей – наличие хроматофоров – органелл, в которых происходит фотосинтез. Форма хроматофоров может быть разнообразной. Хроматофоры содержат хлорофилл, каротиноиды ,а также особые структуры пиреноиды, в которых накапливается крахмал. Кроме хлорофилла и каротиноидов, которые имеются у всех водорослей, в хроматофорах могут содержаться и другие пигменты: бурый, синий, красный и др. Набор этих пигментов зависит от глубины обитания водорослей. Связано это с поглощением водой красных лучей и пропусканием сине-зеленых. Поэтому у поверхности обитают зеленые водоросли, а на глубине – бурые и красные.
У ряда водорослей слоевища расчленены и напоминают побег с ветками и отходящими книзу ризоидами, служащими для прикрепления к грунту. Рассмотрим особенности строения водорослей. 1. Одноклеточные формы Клетки большинства водорослей покрыты плотной оболочкой, которая содержит целлюлозу (у эвгленовых оболочка содержит, в основном, белки). У некоторых водорослей оболочка пропитывается солями железа, кальция или кремния и напоминает панцирь. Отличительная черта клеток водорослей – наличие хроматофоров – органелл, в которых происходит фотосинтез. Форма хроматофоров может быть разнообразной. Хроматофоры содержат хлорофилл, каротиноиды ,а также особые структуры пиреноиды, в которых накапливается крахмал. Кроме хлорофилла и каротиноидов, которые имеются у всех водорослей, в хроматофорах могут содержаться и другие пигменты: бурый, синий, красный и др. Набор этих пигментов зависит от глубины обитания водорослей. Связано это с поглощением водой красных лучей и пропусканием сине-зеленых. Поэтому у поверхности обитают зеленые водоросли, а на глубине – бурые и красные.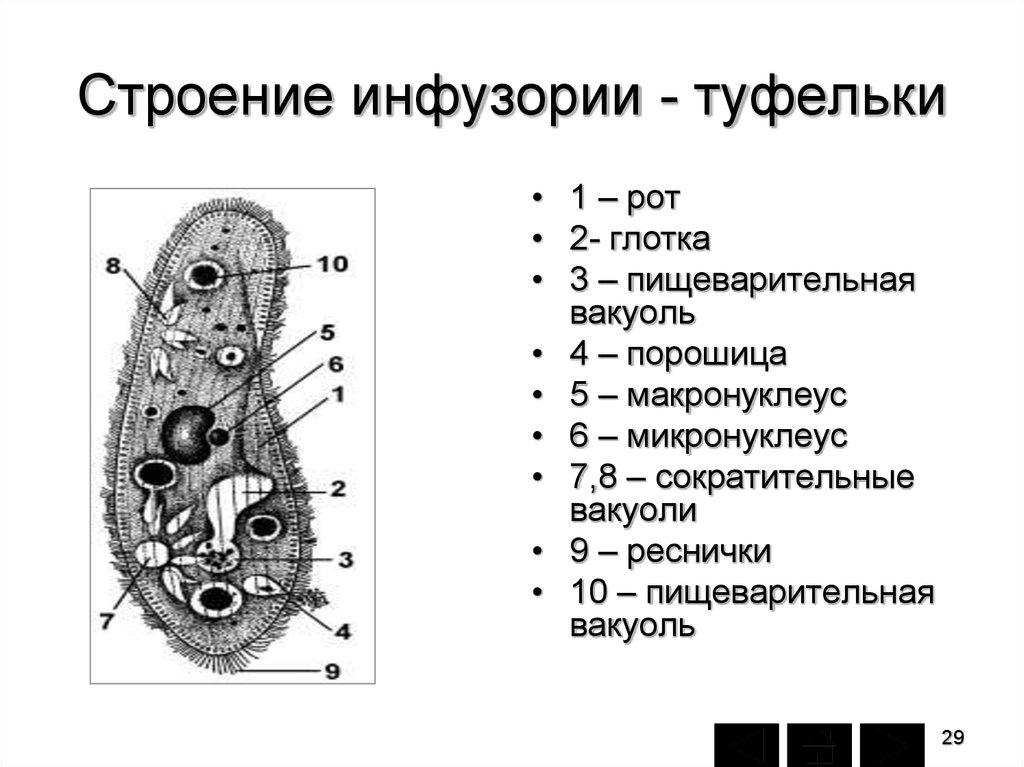 Сократительная вакуоль — мембранный органоид, осуществляющий выброс излишков жидкости из цитоплазмы. Представляет собой наиболее заметную часть согласованно работающего комплекса, в котором выступает в роли периодически опорожняющегося резервуара. Глазок – орган восприятия света в виде красного пятнышка у одноклеточных подвижных водорослей и у зооспор водорослей. Базальное тельце жгутика (блефаропласт) — лежащее в цитоплазме клетки основание аксонемы, сложно организованного цитоскелетного комплекса, составляющий основу жгутиков и ресничек. Мионема – белковая фибрилла, которая способна к сокращению и входит в состав аппарата движения у эвглен и простейших. 2. Многоклеточные формы Тонопласт – мембрана в которую заключена вакуоль. Скафидии — круглые полости, открывающиеся наружу узким каналом. В мужских скафидиях созревают сперматозоиды. На картинке видны находящиеся в них красноватые тела, от которых у зрелых клеток отходят по два жгутика. Созревшие сперматозоиды выходят из скафидия наружу.
Сократительная вакуоль — мембранный органоид, осуществляющий выброс излишков жидкости из цитоплазмы. Представляет собой наиболее заметную часть согласованно работающего комплекса, в котором выступает в роли периодически опорожняющегося резервуара. Глазок – орган восприятия света в виде красного пятнышка у одноклеточных подвижных водорослей и у зооспор водорослей. Базальное тельце жгутика (блефаропласт) — лежащее в цитоплазме клетки основание аксонемы, сложно организованного цитоскелетного комплекса, составляющий основу жгутиков и ресничек. Мионема – белковая фибрилла, которая способна к сокращению и входит в состав аппарата движения у эвглен и простейших. 2. Многоклеточные формы Тонопласт – мембрана в которую заключена вакуоль. Скафидии — круглые полости, открывающиеся наружу узким каналом. В мужских скафидиях созревают сперматозоиды. На картинке видны находящиеся в них красноватые тела, от которых у зрелых клеток отходят по два жгутика. Созревшие сперматозоиды выходят из скафидия наружу. Как вам понятно из столь существенные различия в строении водорослей обусловлены различным образом жизни, особенностями размножения. Размножение может идти бесполым и половым путем. Бесполое размножение происходит путем образования спор, имеющих жгутики и способных к передвижению (зооспоры), либо вегетативным путем, осуществляющимся через деление клеток или через фрагментацию слоевищ, на частях которых вырастают новые водоросли. Половое размножение в простейшем случае представляет собой соединение двух вегетативных клеток. Формы полового размножения достаточно разнообразны – рассмотрим один пример. У спирогиры, представленной на рисунке, осенью две нити располагаются параллельно, лежащие рядом клетки образуют выросты, направленные друг к другу. Соприкоснувшись, эти выросты формируют трубку, по которой содержание (протопласт) одной клетки перетекает в другую. Ядра клеток сливаются и оплодотворение завершается образованием зиготы (диплоидной клетки) которая окружается толстой оболочкой и в таком виде переносит зиму.
Как вам понятно из столь существенные различия в строении водорослей обусловлены различным образом жизни, особенностями размножения. Размножение может идти бесполым и половым путем. Бесполое размножение происходит путем образования спор, имеющих жгутики и способных к передвижению (зооспоры), либо вегетативным путем, осуществляющимся через деление клеток или через фрагментацию слоевищ, на частях которых вырастают новые водоросли. Половое размножение в простейшем случае представляет собой соединение двух вегетативных клеток. Формы полового размножения достаточно разнообразны – рассмотрим один пример. У спирогиры, представленной на рисунке, осенью две нити располагаются параллельно, лежащие рядом клетки образуют выросты, направленные друг к другу. Соприкоснувшись, эти выросты формируют трубку, по которой содержание (протопласт) одной клетки перетекает в другую. Ядра клеток сливаются и оплодотворение завершается образованием зиготы (диплоидной клетки) которая окружается толстой оболочкой и в таком виде переносит зиму.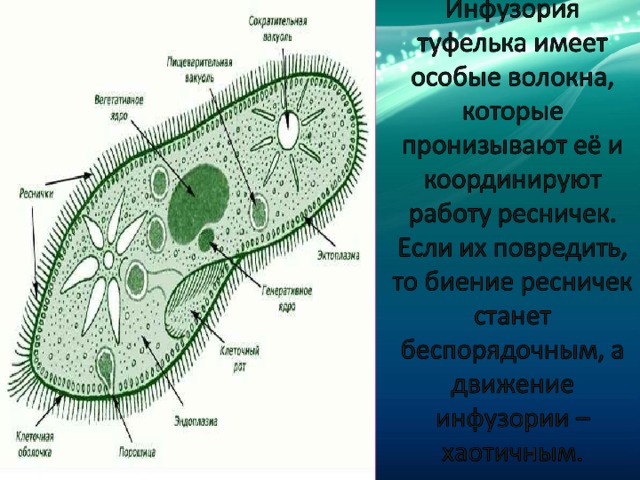 Весной зиготы делится мейотически и каждое из четырех образовавшихся ядер дает начало новой нити. При этом в летний период размножение осуществляется бесполым путем. К половому размножению способны также и одноклеточные формы водорослей, например хламидомонада. вижения, как у многих других ее собратьев, у нее нет. Водоросль пассивно плавает в пресной воде в виде фитопланктона. Размножение хлореллы происходит исключительно бесполым способом, в благоприятных условиях, путем деления клетки пополам. Отвечает за этот сложный процесс ядро. Предварительно, на этапе подготовки клетки к размножению, ее органы удваиваются. Она увеличивается в размерах, затем внешняя оболочка растягивается и разрывается. Из одной материнской получаются две дочерние клетки. На какое-то время в цикле хлореллы наступает перерыв, а затем все повторяется заново. 2. Вольвокс Вольвокс – это зеленые водоросли. Они существуют в виде колонии. Каждая отдельная маленькая водоросль и является клеткой, у которой есть два жгутика и волоски.
Весной зиготы делится мейотически и каждое из четырех образовавшихся ядер дает начало новой нити. При этом в летний период размножение осуществляется бесполым путем. К половому размножению способны также и одноклеточные формы водорослей, например хламидомонада. вижения, как у многих других ее собратьев, у нее нет. Водоросль пассивно плавает в пресной воде в виде фитопланктона. Размножение хлореллы происходит исключительно бесполым способом, в благоприятных условиях, путем деления клетки пополам. Отвечает за этот сложный процесс ядро. Предварительно, на этапе подготовки клетки к размножению, ее органы удваиваются. Она увеличивается в размерах, затем внешняя оболочка растягивается и разрывается. Из одной материнской получаются две дочерние клетки. На какое-то время в цикле хлореллы наступает перерыв, а затем все повторяется заново. 2. Вольвокс Вольвокс – это зеленые водоросли. Они существуют в виде колонии. Каждая отдельная маленькая водоросль и является клеткой, у которой есть два жгутика и волоски.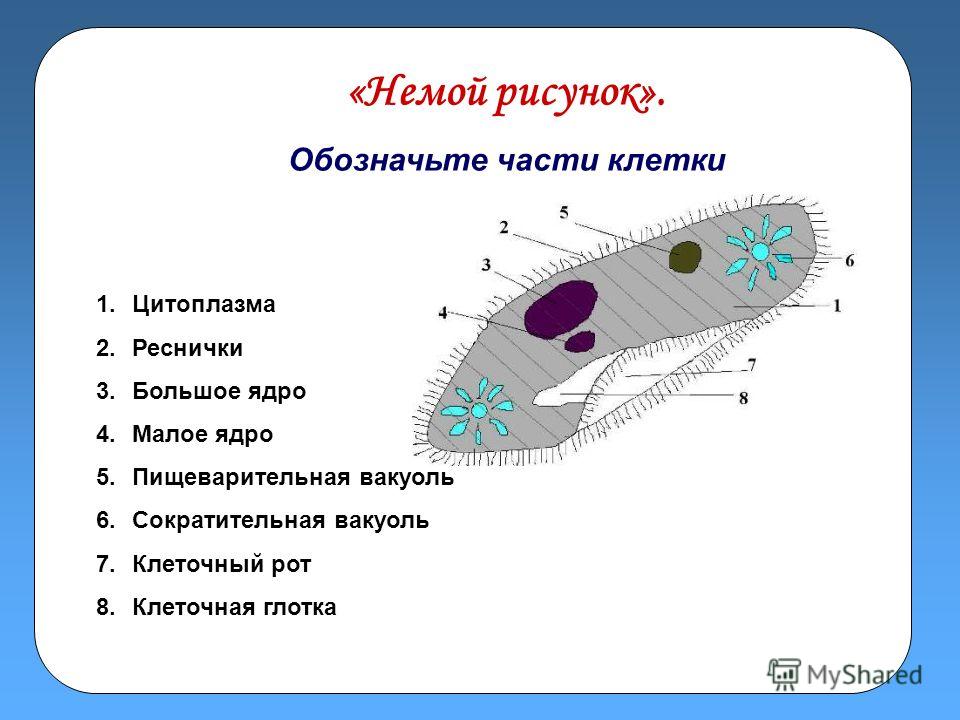 Одиночные клетки соединены друг с другом тонкими нитями цитоплазмы, которые позволяют всей колонии плавать согласованно. Отдельные представители кроме волосков и жгутиков также имеют небольшое красное пятнышко, так называемый глазок. Дифференциация клеток придает уникальности представителям водорослей. Каждая колония имеет начало и конец, иначе говоря, Северный и Южный полюса. В первом регионе наблюдается более выраженное скопление развитых глазков. Это помогает водорослям плыть в сторону света. Таким образом, колония одноклеточных организмов внешне вполне может сойти за многоклеточный организм. Единичная клетка может достигать 2 миллиметров в диаметре (в более крупных колониях – до 2 см), поэтому их легко можно увидеть невооруженным глазом. Отдельные водоросли соединены тонкими нитями цитоплазмы. У водоросли вольвокс строение достаточно простое. Центральная полость заполнена слизью. Все клетки имеют по два жгутика, которые прикрепляются у переднего конца. Каждая ячейка в клетке выполняет свою функцию питания, дыхания и экскреции.
Одиночные клетки соединены друг с другом тонкими нитями цитоплазмы, которые позволяют всей колонии плавать согласованно. Отдельные представители кроме волосков и жгутиков также имеют небольшое красное пятнышко, так называемый глазок. Дифференциация клеток придает уникальности представителям водорослей. Каждая колония имеет начало и конец, иначе говоря, Северный и Южный полюса. В первом регионе наблюдается более выраженное скопление развитых глазков. Это помогает водорослям плыть в сторону света. Таким образом, колония одноклеточных организмов внешне вполне может сойти за многоклеточный организм. Единичная клетка может достигать 2 миллиметров в диаметре (в более крупных колониях – до 2 см), поэтому их легко можно увидеть невооруженным глазом. Отдельные водоросли соединены тонкими нитями цитоплазмы. У водоросли вольвокс строение достаточно простое. Центральная полость заполнена слизью. Все клетки имеют по два жгутика, которые прикрепляются у переднего конца. Каждая ячейка в клетке выполняет свою функцию питания, дыхания и экскреции. Форма может быть сферической, эллиптической или овальной. Наружный слой также покрыт слизью. Каждая ячейка имеет один глаз в виде пятна на переднем конце. Они могут размножаться как бесполым, так и половым путем. При внимательном рассмотрении можно увидеть, что большинство колоний имеет внутри специфические сферы, которые называются гонадами. Это является признаком бесполого размножения. Гонады выращиваются из клеток вокруг экватора колонии. Эти клетки увеличиваются и проходят ряд делений, пока не образуют маленькие сферы. При этом жгутики оказываются внутри новой окружности. Чтобы их извлечь, клетка должна вывернуть себя наизнанку. Подобно бесполому размножению специальные клетки скапливаются вокруг экватора. Самец и самка создают колонии различных зародышевых клеток. Сперматозоиды образуются путем деления. Женские половые клетки не делятся, а просто увеличиваются в размере. Большинство видов имеет как мужские, так и женские колонии. А некоторые представляют собой гермафродитов. 3. Спирогира Спирогира относится к отделу Зеленые водоросли.
Форма может быть сферической, эллиптической или овальной. Наружный слой также покрыт слизью. Каждая ячейка имеет один глаз в виде пятна на переднем конце. Они могут размножаться как бесполым, так и половым путем. При внимательном рассмотрении можно увидеть, что большинство колоний имеет внутри специфические сферы, которые называются гонадами. Это является признаком бесполого размножения. Гонады выращиваются из клеток вокруг экватора колонии. Эти клетки увеличиваются и проходят ряд делений, пока не образуют маленькие сферы. При этом жгутики оказываются внутри новой окружности. Чтобы их извлечь, клетка должна вывернуть себя наизнанку. Подобно бесполому размножению специальные клетки скапливаются вокруг экватора. Самец и самка создают колонии различных зародышевых клеток. Сперматозоиды образуются путем деления. Женские половые клетки не делятся, а просто увеличиваются в размере. Большинство видов имеет как мужские, так и женские колонии. А некоторые представляют собой гермафродитов. 3. Спирогира Спирогира относится к отделу Зеленые водоросли.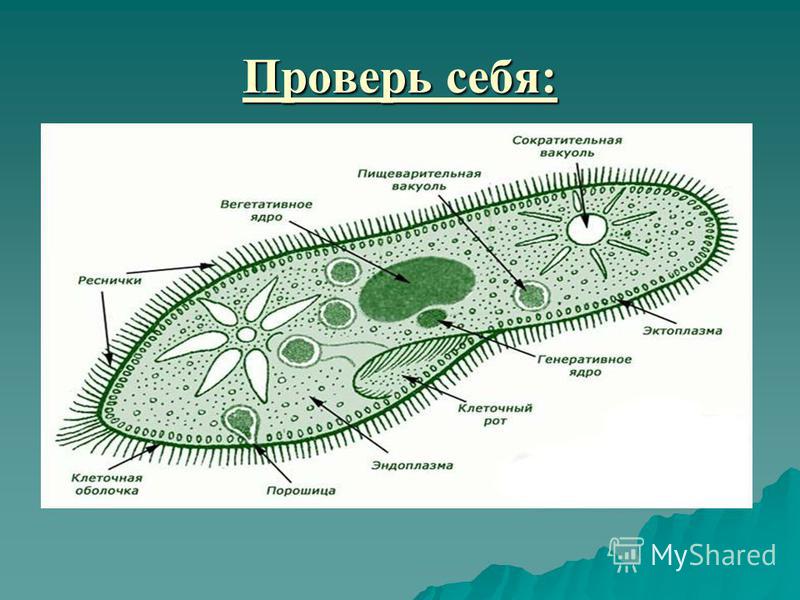 Ее тело представляет собой таллом нитчатой формы, который состоит из отдельных клеток и не образует тканей и органов. Данный представитель зеленых водорослей является многоклеточным организмом. Строение спирогиры представлено длинной нитчатой структурой. Хлоропласты данной водоросли имеют вид лент, которые спирально закручиваются. Эта черта и определяет название описываемых организмов. Клетки также удлиненные, имеют цилиндрическую форму. Они соединяются при помощи пор в оболочках, через которые и осуществляется процесс обмена веществ. Подобно всем растительным клеткам, их большую часть занимают вакуоли, представляющие собой резервуары, заполненные водой с растворенными питательными веществами. Оболочки пропитаны углеводом целлюлозой, придающей им прочность и жесткость. Водоросль спирогира, как и все растения, является эукариотическим организмом. Это значит, что ее клетки содержат ядро. В органелле находится генетический материал, заключенный в молекулах ДНК. Водоросль спирогира в преобладающем большинстве случаев является обитателем пресных водоемов со стоячей водой.
Ее тело представляет собой таллом нитчатой формы, который состоит из отдельных клеток и не образует тканей и органов. Данный представитель зеленых водорослей является многоклеточным организмом. Строение спирогиры представлено длинной нитчатой структурой. Хлоропласты данной водоросли имеют вид лент, которые спирально закручиваются. Эта черта и определяет название описываемых организмов. Клетки также удлиненные, имеют цилиндрическую форму. Они соединяются при помощи пор в оболочках, через которые и осуществляется процесс обмена веществ. Подобно всем растительным клеткам, их большую часть занимают вакуоли, представляющие собой резервуары, заполненные водой с растворенными питательными веществами. Оболочки пропитаны углеводом целлюлозой, придающей им прочность и жесткость. Водоросль спирогира, как и все растения, является эукариотическим организмом. Это значит, что ее клетки содержат ядро. В органелле находится генетический материал, заключенный в молекулах ДНК. Водоросль спирогира в преобладающем большинстве случаев является обитателем пресных водоемов со стоячей водой. Но встречается она и в морях. Ее нити, переплетаясь в большом количестве, похожи на зеленую вату, плавающую на поверхности. Строение спирогиры обусловливает и тип ее питания, поскольку клетки водоросли содержат зеленые пластиды хлоропласты, способные осуществлять процесс фотосинтеза. Полученную глюкозу водоросли используют для осуществления процессов жизнедеятельности и роста организма. А выделяемый кислород требуется всем живым организмам для дыхания — необходимого условия существования на планете. Источником веществ, необходимых для фотосинтеза, а именно воды и углекислого газа, является непосредственно среда их обитания. Для этой зеленой водоросли преобладающим является вегетативное размножение частями таллома. Многочисленные нити разрываются на части, каждая из которых дает начало новому организму. Это обусловливает интенсивность и высокую продуктивность данного процесса. Однако строение спирогиры делает возможным и половой процесс. Он осуществляется в форме конъюгации. При этом нити водорослей переплетаются между собой, между ними возникает специализированное образование — копуляционный канал.
Но встречается она и в морях. Ее нити, переплетаясь в большом количестве, похожи на зеленую вату, плавающую на поверхности. Строение спирогиры обусловливает и тип ее питания, поскольку клетки водоросли содержат зеленые пластиды хлоропласты, способные осуществлять процесс фотосинтеза. Полученную глюкозу водоросли используют для осуществления процессов жизнедеятельности и роста организма. А выделяемый кислород требуется всем живым организмам для дыхания — необходимого условия существования на планете. Источником веществ, необходимых для фотосинтеза, а именно воды и углекислого газа, является непосредственно среда их обитания. Для этой зеленой водоросли преобладающим является вегетативное размножение частями таллома. Многочисленные нити разрываются на части, каждая из которых дает начало новому организму. Это обусловливает интенсивность и высокую продуктивность данного процесса. Однако строение спирогиры делает возможным и половой процесс. Он осуществляется в форме конъюгации. При этом нити водорослей переплетаются между собой, между ними возникает специализированное образование — копуляционный канал.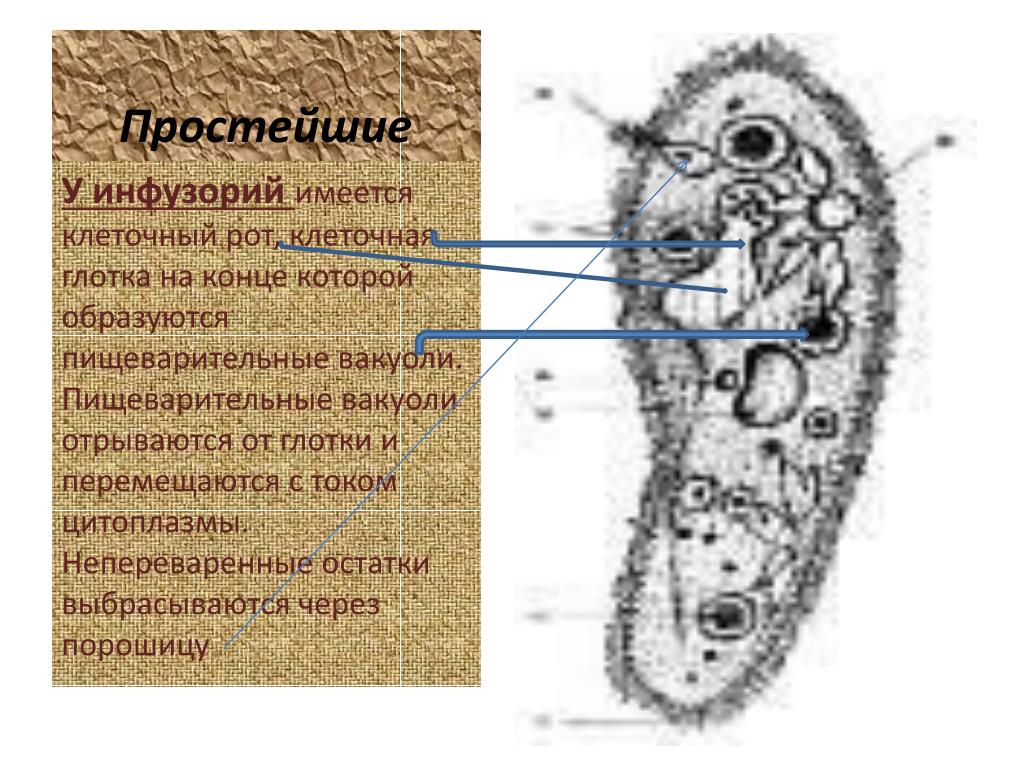 По нему внутреннее содержимое одной клетки, содержащей генетический материал, перетекает в другую. В результате возникает уплотненное образование — зигоспора. В таком состоянии водоросль переносит неблагоприятные условия, а весной из нее прорастает молодая нитчатая особь. Вывод: в ходе лабораторной работы я научился находить и определять водоросли из пробы аквариумной воды, после чего классифицировал и описал выявленные образцы, посредством чего изучил строение водорослей на живых препаратах.
По нему внутреннее содержимое одной клетки, содержащей генетический материал, перетекает в другую. В результате возникает уплотненное образование — зигоспора. В таком состоянии водоросль переносит неблагоприятные условия, а весной из нее прорастает молодая нитчатая особь. Вывод: в ходе лабораторной работы я научился находить и определять водоросли из пробы аквариумной воды, после чего классифицировал и описал выявленные образцы, посредством чего изучил строение водорослей на живых препаратах.
Структуры и функции биологических клеток
АННОТАЦИЯ
В статье рассматриваются межклеточные вещества, присутствующие во внеклеточных организмах, их структура и функции в живых организмах. Анализируется жизненно важные свойства клеток от белков, уточняется роль гликокаликсов в пасщеплении эпительных клеток. Уточняются эндоплазматический вид мембран, приводятся характеристические свойства видов этих мембран а также роль этих клетов при синтезе липидов и гликогенов.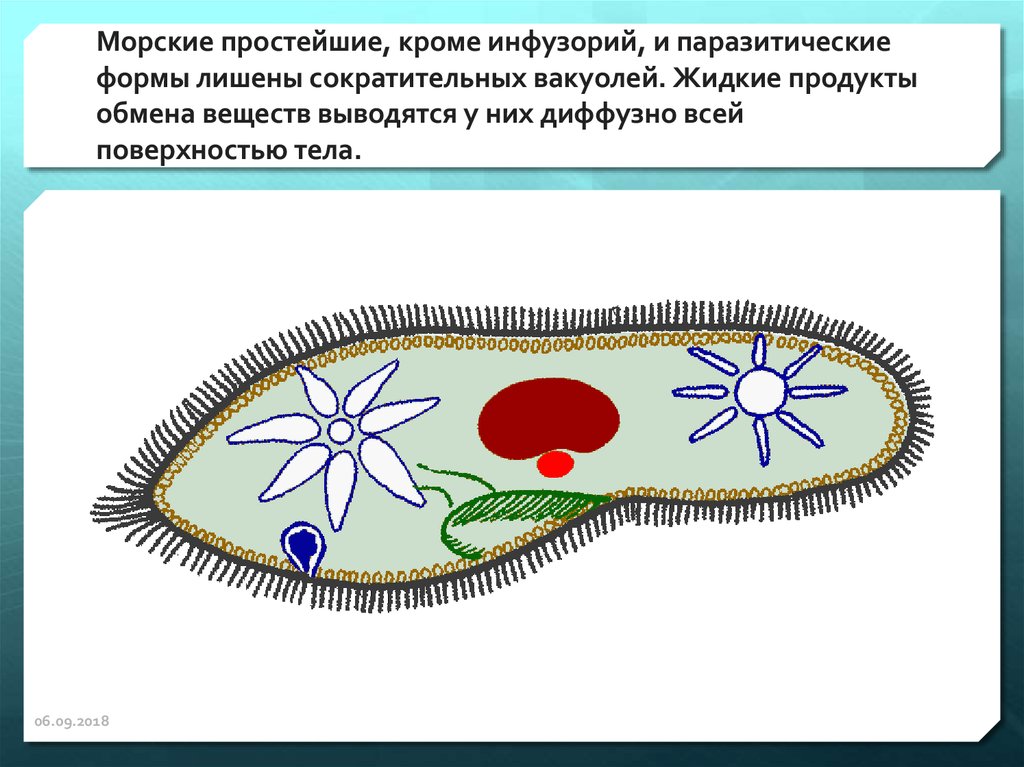
ABSTRACT
The article considers intercellular substances present in extracellular organisms, their structure and functions in living organisms. The vital properties of cells from proteins are analyzed, the role of glycocalyxes in the cleavage of epithelial cells is specified. The endoplasmic appearance of the membranes is specified, the characteristic properties of the types of these membranes, as well as the role of these cells in the synthesis of lipids and glycogens, are given.
Ключевые слова: многоклеточный организм, клеточная теория, одноклеточный организм, цитоплазма, метаболизм, гликокаликс, гликопротеиды, плазматические мембраны, биологические мембраны.
Keywords: multicellular organism, cellular theory, unicellular organism, cytoplasm, metabolism, glycocalyx, glycoproteins, plasma membranes, biological membranes.
Клетка — это элемент организма, способный жить самостоятельно, создавать и развивать себя. Клетка является основой строения и жизнедеятельности всех живых организмов и растений. Клетки могут существовать как в качестве самостоятельного организма, так и в составе многоклеточных организмов (тканевых клеток). Термин «клетка» был предложен английским микроскопистом Р. Хука. Особый раздел клеточной биологии является темой цитологических исследований. Дальнейшее систематическое изучение клеток началось в 19 веке. Одной из крупнейших научных теорий того времени была Теория клеток, которая подтвердила единство структуры всех живых существ. Изучение любой жизни на клеточном уровне является основой современных биологических исследований.
Клетка является основой строения и жизнедеятельности всех живых организмов и растений. Клетки могут существовать как в качестве самостоятельного организма, так и в составе многоклеточных организмов (тканевых клеток). Термин «клетка» был предложен английским микроскопистом Р. Хука. Особый раздел клеточной биологии является темой цитологических исследований. Дальнейшее систематическое изучение клеток началось в 19 веке. Одной из крупнейших научных теорий того времени была Теория клеток, которая подтвердила единство структуры всех живых существ. Изучение любой жизни на клеточном уровне является основой современных биологических исследований.
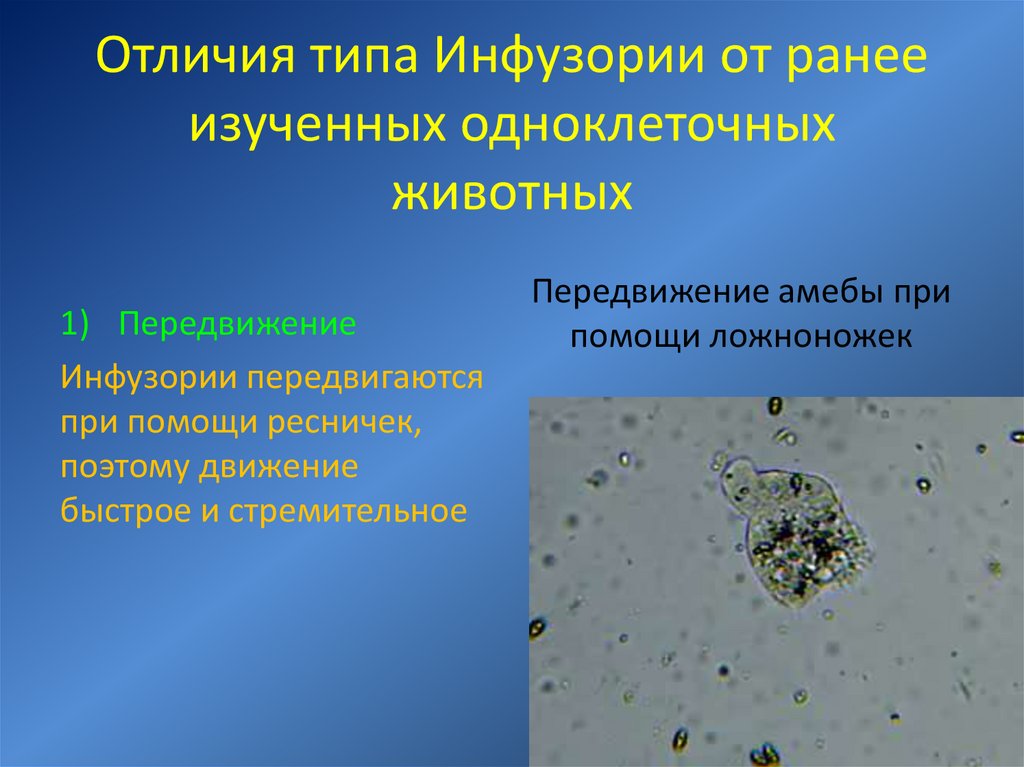
Структура и функции каждой клетки характерны для всех клеток, отражая единство, которое исходит от первичного органического вещества. Характеристики различных клеток являются результатом их специализации в процессе эволюции. Таким образом, все клетки одинаково регулируют обмен веществ, удваивая и используя свои наследственные материалы, поглощая и потребляя энергию. В то же время разные одноклеточные организмы (амёбы, туфельки, инфузории и т.д.) довольно сильно различаются размерами, формой, поведением.
В то же время разные одноклеточные организмы (амёбы, туфельки, инфузории и т.д.) довольно сильно различаются размерами, формой, поведением.
Клетки у человека и животных отличаются по размеру, форме и структуре. Клетки имеют разные формы в зависимости от их функции. В жидкой среде клетки сильно изменчивы и вызывают псевдоподию. Клетки, которые находятся близко друг к другу, однако, имеют особую форму. Например, эпителиальная клетка кожи является плоской, сократительной, а мышечные клетки находятся в форме двуколки. Нервные клетки, которые изменяют пульс, будут иметь более длинные опухоли. Клетки у человека и большинства млекопитающих имеют размер от 5-7 до 200 мкм.
Жизненные свойства клетки зависят от белка, который она содержит. Новый метаболизм является основой жизненного процесса в клетке. Это поглощение или превращение веществ в промежуточной среде в цитоплазму (естественный синтез) — растворение веществ в цитоплазме до энергии клетки: рост определенных частей цитоплазмы, образующихся при образовании белка и углерода воды. Это можно рассматривать как непрерывную структуру под электронным микроскопом. Поскольку гликокаликс является самым внешним слоем, он играет важную роль в коммуникации клетки с внешней средой. Химический состав зоны гликокаликса отличается от химического состава различных клеток.
Это можно рассматривать как непрерывную структуру под электронным микроскопом. Поскольку гликокаликс является самым внешним слоем, он играет важную роль в коммуникации клетки с внешней средой. Химический состав зоны гликокаликса отличается от химического состава различных клеток.
В некоторых клетках гликокаликс богат энзимами, которые расщепляют вещества, тогда как гликопротеиды в других клетках обладают иммунологическими свойствами. Гликокаликс играет важную роль в расщеплении и смещении эпителиальных клеток кишечника (энтероцитов) на микрососудистой мембране. Он сохраняет энтероцидные клетки под воздействием химических веществ и определенных микробов, определяя силу апикальной части микрососудистой части клетки.
Средняя зона состоит из плазматической мембраны (плазмолеммы), наиболее важной и сложной структуры плазматической коры. Плазматические мембраны имеют элементарную биологическую мембранную структуру и состоят из миопротеидов. Он состоит из тонкого среза цитоплазмы, который касается плазматической мембраны в четких поперечных срезах.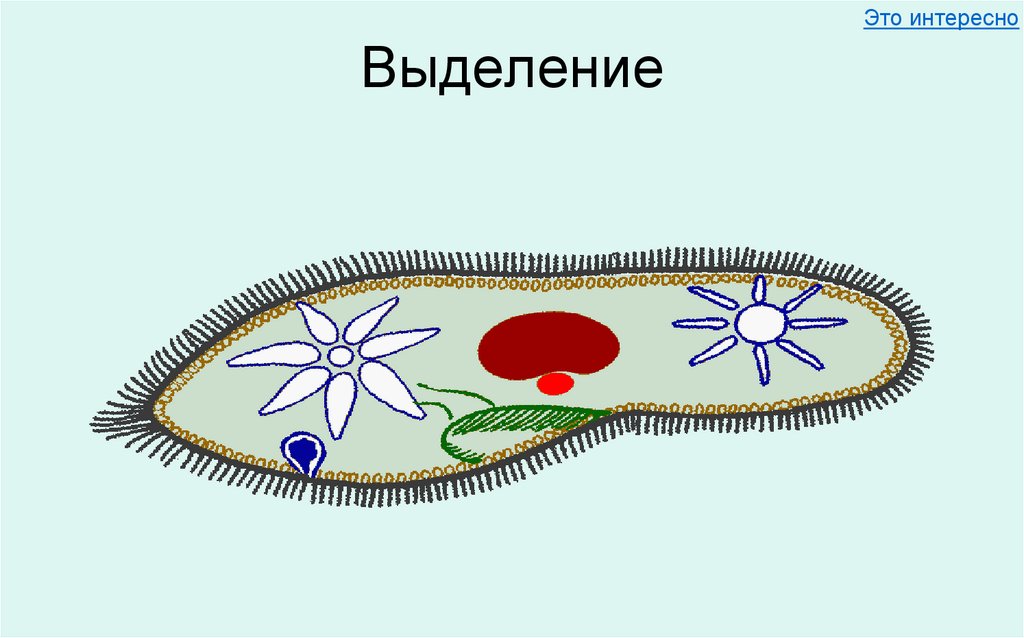 Здесь это заканчивается отсутствием органоидов от цитоплазмы до микроэлементов. Это определяется образованием псевдотел и образованием биоточей в процессах фагоцитоза и пиноцитоза.
Здесь это заканчивается отсутствием органоидов от цитоплазмы до микроэлементов. Это определяется образованием псевдотел и образованием биоточей в процессах фагоцитоза и пиноцитоза.
Эндоплазматический ретикулум — гранулярные и беззерновые эндоплазматические виды различаются в зависимости от рибосом в мембране. Эндоплазматический вид состоит из системы внутриклеточных протоков, вакуолей и цистерн, окруженных плазматической мембраной. Гранулярная эндоплазматическая сетка хорошо развита в секреторных клетках, таких как клетки поджелудочной железы и клетки печени, толщина мембран эндоплазматических видов составляет 4-7,5 нм, рибонуклеопротеидные зерна расположены на внешней стороне, зернистая передняя мембрана ядра зернистая также связан с цистернами.
Гранулярная эндоплазматическая сетка хорошо развита в клетках, которые продуцируют оксил. Гладкая эндоплазматическая сетка хорошо развита в клетках, которые продуцируют небелковые продукты (клетки желудка). В основном он участвует в синтезе липидов, гликогена.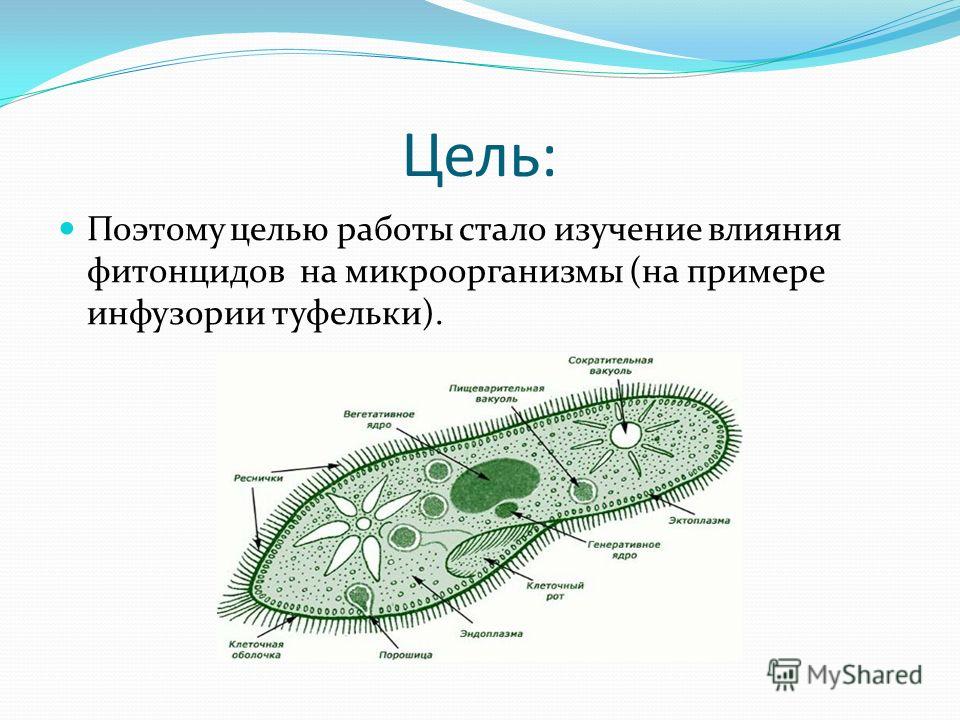 Слизистая эндоплазматическая сетчатка обнаружена в сальных железах. В то же время сеть участвует в расщеплении гликогена.
Слизистая эндоплазматическая сетчатка обнаружена в сальных железах. В то же время сеть участвует в расщеплении гликогена.
В конце гидролитического расщепления образуются окаменелые тела, которые являются полностью фагоцитированным материалом. Образование лизосом связано с передним G-комплексом. Первичные лизосомы расположены вокруг переднего резервуара G в виде небольших пузырьков или плотных тел. Присутствие кислой фосфатазы в переднем G-комплексе указывает на то, что передний G-комплекс играет роль в образовании лизосом. Ферменты лизосомы синтезируются эндоплазматическим типом. Лизосомы также образуются из плазматической мембраны.
Цитоплазматические включения — входные данные являются непостоянными компонентами цитоплазмы. Они вызваны проникновением веществ из внешней среды в процессе внутриклеточного метаболизма, секреции и образования пигментов, а также фагоцитоза.
Под микроскопом вставки видны в виде гранул или жидких вакуолей различной интенсивности. Химический состав добавок различен. Есть несколько групп, чтобы сделать:
Есть несколько групп, чтобы сделать:
1. Глубокие добавки (жиры, протеиновые гранулы, наборы гликогена, витамины)
2. Секреторные соединения (зимогенные гранулы и другие вещества: гемоглобин, меланин, мигофусцин и др.)
3. Экспрессорные входы (желчная кислота, мочевина)
4. Пигментные добавки (гемоглобин, меланин, мигофусин и др.)
Другим важным компонентом растительной клетки являются ее бляшки. Существует три вида пластмасс: хлоропласт — зеленые пластиды, хромопласт — красные или другие цветные пластинки, лейкопласты — бесцветные пластиды. Растительная клетка также содержит вакуоли. Клеточный сок хранится в них. Известно, что вакуольная часть цитоплазмы клетки называется тонопластом. С возрастом клеток тонопласт увеличивается с ростом вакуолей. У животных существует вакуум пищеварения и сокращения. Таким образом, клеточная стенка (состоящая из целлюлозы в растении) является основным органоидом, который дифференцирует вакуоли, пластиды, клетки растений и животных. Пластины содержат хлорофилл, хлорофилл, ксантофилл, феноксантиновые пигменты, а также некоторые ароматические соединения.
Пластины содержат хлорофилл, хлорофилл, ксантофилл, феноксантиновые пигменты, а также некоторые ароматические соединения.
Как правило, молодая клетка с протопластом, который является просто плодом, имеет множество небольших вакуолей с округлой или вытянутой формой. Эти вакуоли заполнены растворенными веществами и коллоидом в форме геля. Вакуоли затем растут и становятся системой трубок, заполненных клеточным соком. Затем вакуоли частично сокращаются, увеличиваются, соединяются, уменьшаются в количестве и округляются. За это время вода в вакуолях увеличивается, и большинство растворенных веществ в вакууме становятся нейтральными. Тогда реальные решения формируются. Вакуоли затем добавляют в одну большую центральную вакуоль. Протоплазма в это время расположена вокруг клеточных стенок; Ядром этого слоя является В некоторых случаях ядро находится в центре клетки; заключен в субклеточную протоплазму клетки с помощью выдвижных ящиков и пластинок, проходящих через полость вакуолей протоплазмы, окружающих ее.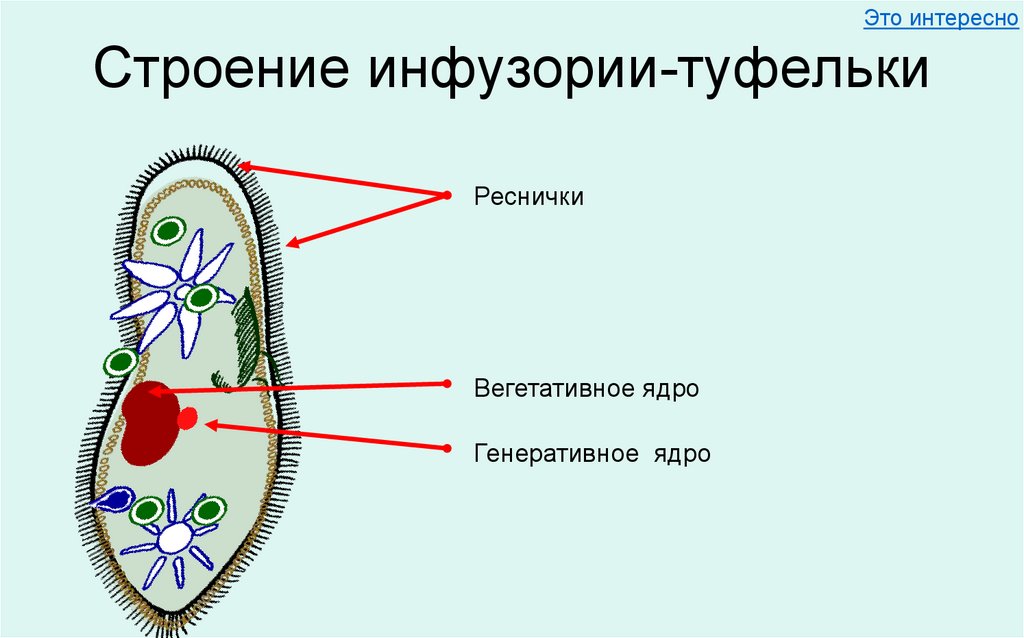
Список литературы:
1. Беликов В.Г. Фармацевтическая химия. М.: Высшая школа, 1993.
2. Ганиев М.М., Недорезков В.Д. Химические средства защиты растений. М.: Колос, 2006.
3. Кнорре Д.Г., Мызина С.Д. Биологическая химия. М.: Высшая школа, 2000.
4. Кольман Я., Рём К.-Г. Наглядная биохимия. М.: Мир, 2000.
5. Николаев А.Я. Биологическая химия. М.: Медицинское информационное агенство, 2004.
6. Тейлор Д., Грин Н., Стаут У. Биология: в 3 т. М.: Мир, 2001.
Особенности процессов жизнедеятельности одноклеточных эукариот
Биология. 10 класс. Балан
Вспомните: каково строение жгутиков и ресничек? Какие организмы называют автотрофными, миксотрофными и гетеротрофными? Каковы строение и функции плазматической мембраны?
• Движение протист обеспечивают особые органеллы: жгутики (рис. 38.1), реснички или ложные ножки (псевдоподии). Кроме двигательной функции, жгутики и реснички могут выполнять и некоторые другие.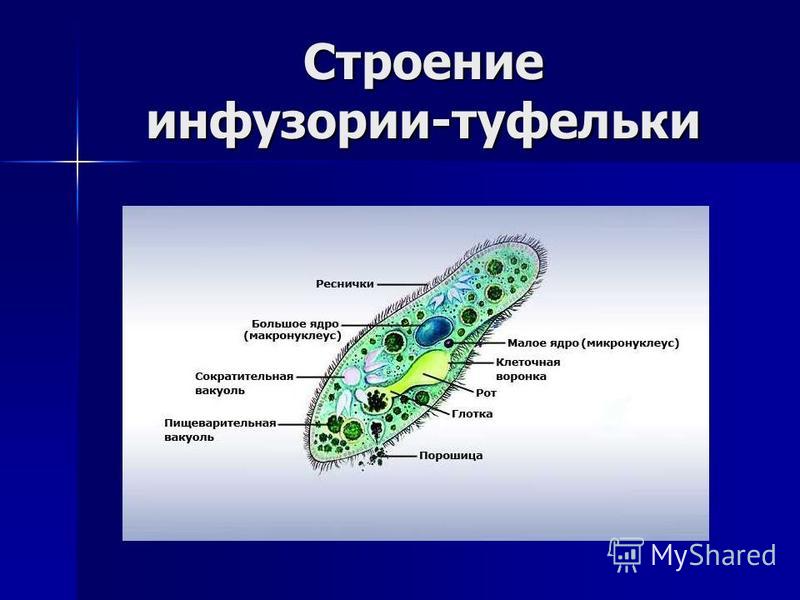 Иногда жгутики служат для прикрепления, образуя стебелек (см. рис. 37.4). Движения жгутиков и ресничек обеспечивают постоянные потоки воды вокруг клетки, приносящие частички пищи и очищающие ее поверхность от посторонних частиц.
Иногда жгутики служат для прикрепления, образуя стебелек (см. рис. 37.4). Движения жгутиков и ресничек обеспечивают постоянные потоки воды вокруг клетки, приносящие частички пищи и очищающие ее поверхность от посторонних частиц.
Амебоидное движение происходит благодаря образованию временных выростов — псевдоподий. Они бывают разной формы, у некоторых видов способны сливаться между собой в определенных участках и образовывать ловчую сеть для захвата питательных частичек (рис. 38.2).
Скользящее движение присуще лишь немногим группам протист: одноклеточным животным грегаринам (рис. 38.3), диатомовым водорослям и некоторым другим. У грегарин скольжение происходит за счет волнообразных движений продольных складок поверхности клетки, у диатомовых водорослей — стекания слизи с поверхности клетки в определенном направлении.
Рис. 38.1. Длинный жгутик эвглены (фотография сделана с помощью сканирующего электронного микроскопа): 1 — углубление, в котором находится жгутик (2)
Рис. 38.2. Типы псевдоподий (стрелки показывают направление движения цитоплазмы): 1 — широкие с закругленной верхушкой; 2 — тоненькие, способные сливаться между собой; таким образом они образуют ловчую сеть вокруг клетки; 3 — тоненькие неразветвленные
38.2. Типы псевдоподий (стрелки показывают направление движения цитоплазмы): 1 — широкие с закругленной верхушкой; 2 — тоненькие, способные сливаться между собой; таким образом они образуют ловчую сеть вокруг клетки; 3 — тоненькие неразветвленные
Рис. 38.3. Грегарина — одноклеточный паразит членистоногих
• Питание одноклеточных эукариот. Среди протист по типу питания выделяют автотрофов, гетеротрофов и миксотрофов. Гетеротрофы могут поглощать еду посредством пиноцитоза или фагоцитоза (вспомните, как формируются пищеварительные вакуоли). Разновидностью фагоцитоза служит высасывание содержимого клетки добычи хищными жгутиконосцами и некоторыми инфузориями (рис. 38.4).
Рис. 38.4. I. Хищная инфузория дидиний: 1 — хоботок, на верхушке которого находится ротовое отверстие, проникает в тело добычи; 2 — пояски ресничек, при помощи которых дидиний плавает. II. Дидиний (3, 4), поедающий инфузорию-туфельку
Рис.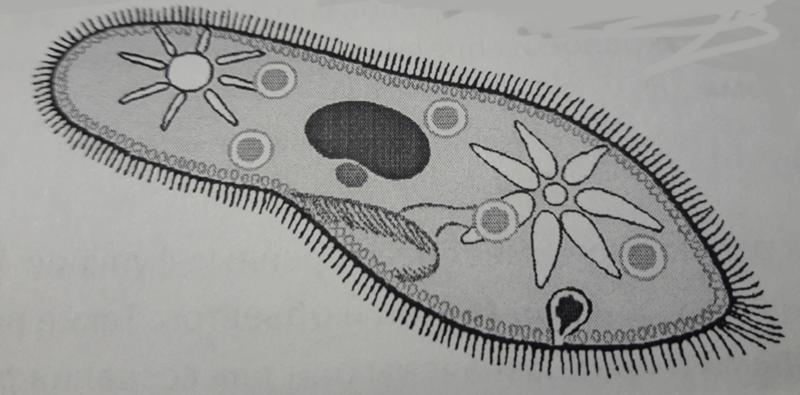 38.5. Система органелл пищеварения инфузории-туфельки
38.5. Система органелл пищеварения инфузории-туфельки
Многим одноклеточным организмам присуще сложное поведение, связанное с поиском и захватом пищи. Оно заключается в распознавании рецепторными молекулами поверхности клетки химических веществ источника питания (другой клетки, органических частиц и т. п.).
Автотрофные (большинство водорослей) и миксотрофные (например, эвглена зеленая, хламидомонада) протисты способны к фотосинтезу. Запасными полисахаридами у них служат крахмал (зеленые водоросли), хризоламинарин (диатомовые водоросли) и др. У гетеротрофов накапливается полисахарид гликоген.
Непереваренные твердые остатки амебообразные организмы могут выбрасывать в любом месте своей поверхности, а у тех, которые имеют постоянную форму клетки (например, инфузория-туфелька), для этого служит участок особого строения — порошица (рис. 38.5).
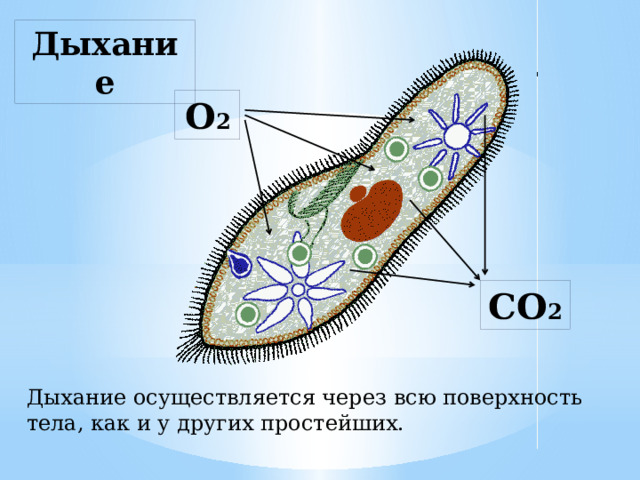
Функцию выделения из клетки избытка воды вместе с растворенными в ней продуктами обмена веществ осуществляет сократительная вакуоль, присущая в основном пресноводным протистам. У паразитических видов и обитателей соленых водоемов эта вакуоль обычно отсутствует, а выделение происходит через всю поверхность клетки.
У паразитических видов и обитателей соленых водоемов эта вакуоль обычно отсутствует, а выделение происходит через всю поверхность клетки.
• Размножение протист преимущественно бесполое, иногда — половое. Для многих видов (фораминиферы, малярийный паразит и др.) характерны сложные жизненные циклы, происходящие со сменой поколений с разными формами размножения.
Бесполое размножение происходит в виде разновидностей митотического деления. В некоторых случаях во время митоза ядерная оболочка сохраняется, и ядро перешнуровывается надвое по окончании расхождения хромосом к его полюсам (например, у амеб, многих жгутиконосцев и проч.).
Бесполое размножение может осуществляться делением пополам (амеба протей, эвглена зеленая, инфузория-туфелька и др.), множественным делением (малярийный паразит), почкованием (некоторые инфузории) или с помощью подвижных (зооспоры) или неподвижных (водоросли, грибы) спор. При почковании от большей (материнской) клетки может отделяться одна или несколько более мелких (дочерних). В результате множественного деления ядро клетки многократно делится, а затем вокруг каждого из дочерних ядер отделяется участок цитоплазмы и образуются десятки или сотни мелких дочерних клеток (фораминиферы).
В результате множественного деления ядро клетки многократно делится, а затем вокруг каждого из дочерних ядер отделяется участок цитоплазмы и образуются десятки или сотни мелких дочерних клеток (фораминиферы).
Рис. 38.6. I. Размножение хламидомонады: бесполое (А) и половое (Б). II. Размножение слизистых споровиков. III. Цикл развития малярийного пара зита: В — бесполые поколения в организме человека; Г — половое размножение и развитие бесполых поколений в организме комара; Д — попадание подвижных клеток из слюнных желез комара в кровь человека
Половой процесс обычно происходит в формах попарного слияния половых клеток — гамет. У одних видов (хламидомонады, фораминиферы) все гаметы одинаковые, у других (малярийный паразит, вольвокс) — женские и мужские половые клетки отличаются между собой размерами (рис. 38.6).
Конъюгация известна у инфузорий, некоторых водорослей и грибов. Во время конъюгации две клетки инфузорий обмениваются мигрирующими ядрами. У нитчатой водоросли спирогиры между отдельными клетками двух нитей возникают цитоплазматические мостики, по которым содержимое одной клетки переходит в другую с образованием зиготы (рис. 38.7). В последующем зигота прорастает новой нитью.
У нитчатой водоросли спирогиры между отдельными клетками двух нитей возникают цитоплазматические мостики, по которым содержимое одной клетки переходит в другую с образованием зиготы (рис. 38.7). В последующем зигота прорастает новой нитью.
Рис. 38.7. Конъюгация у спирогиры: 1 — цитоплазматический мостик между двумя клетками; 2 — содержимое одной клетки переходит в другую с образованием зиготы; 3 — пустая клеточная оболочка
• Поведение одноклеточных эукариот. Подобно другим организмам, протистам присуща раздражимость, то есть способность воспринимать раздражители окружающей среды и определенным образом на них отвечать. Двигательные реакции протист имеют название таксисы. Таксис (от греч. таксис — размещение) — направленные движения организмов к раздражителю (положительный таксис) или от него (отрицательный таксис). Например, клетки эвглены зеленой или хламидомонады движутся в направлении источника освещения благодаря наличию особенного светочувствительного образования — глазка.
• Адаптации протист к среде обитания. Существование в разнообразных средах обитания возможно благодаря наличию у протист разнообразных приспособлений (адаптаций) к факторам окружающей среды. Например, у обитателей толщи воды имеются разнообразные выросты раковины, клеточной стенки или внутреннего скелета в виде шипов, радиально расположенных игл (например, у некоторых фораминифер и радиолярий), которые увеличивают площадь поверхности клетки, одновременно уменьшая ее плотность. Свободному парению в толще воды способствует и сеть из тонких псевдоподий (радиолярии) или включения капель жироподобного вещества в цитоплазме (радиолярии, панцирные жгутиковые и т. п.).
У протист, ведущих сидячий образ жизни, есть образования, с помощью которых они прикрепляются к субстрату; например, прикрепительный стебелек инфузорий-сувоек (рис. 37.4).
Обитание в мелких заполненных водой капиллярах почвы также отражается на определенных особенностях строения. Клетки почвенных протист в 5-10 раз мельче по сравнению с размерами близких видов — обитателей пресных или соленых водоемов, часто сплюснуты, несколько упрощенного строения.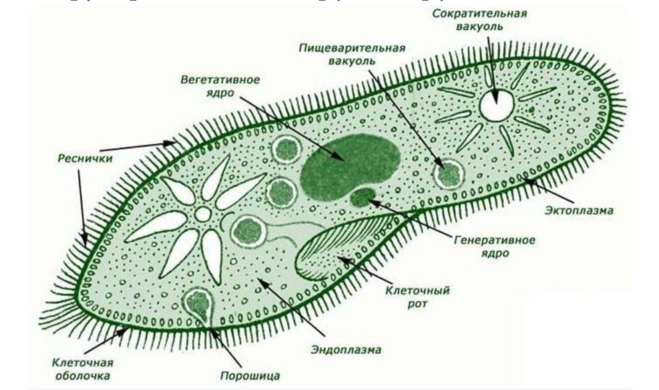
• Инцистирование. Засушливые периоды и низкие температуры одноклеточные животные переживают в виде особенных стадий покоя — цист. В процессе образования цист (инцистирования) может происходить сложная перестройка клетки. Например, у инфузорий во время инцистирования исчезают структуры, связанные с активной жизнью (реснички, органеллы захвата пищи, вакуоли и др.), цитоплазма обезвоживается, уплотняется, а вокруг клетки формируются защитные оболочки (обычно их не менее двух) (рис. 38.8). Эти оболочки характеризуются низкой проницаемостью и осуществляют механическую и химическую защиту клетки, которая сохраняет жизнеспособность в инцистированном состоянии многие месяцы и даже годы. Цисты обеспечивают распространение протист ветром, водой или при участии живых существ.
Рис. 38.8. Цисты амебы (1) и инфузории (2)
Ключевые термины и понятия. Инцистирование, таксис.
Кратко о главном
- Движения протист обеспечивают жгутики, реснички, ложные ножки (псевдоподии), волнообразные сокращения поверхности клетки или направленное стекание слизи.
- Среди протист по типу питания различают автотрофов, гетеротрофов и миксотрофов. Непереваренные твердые остатки пищи амебообразные организмы могут выбрасывать в любом месте своей поверхности, а у имеющих постоянную форму тела (например, инфузория-туфелька) для этого служит участок особого строения — порошица. Функцию выделения из клетки избытка воды вместе с растворенными в ней продуктами обмена веществ осуществляют сократительные вакуоли, присущие в основном пресноводным протистам. У паразитических видов и обитателей соленых водоемов эта вакуоль обычно отсутствует, а выделение происходит через всю поверхность клетки.
- У протист известно бесполое и половое размножение. Бесполое размножение происходит в виде разновидностей митоза: деления пополам, множественного деления, почкования и др. Половой процесс осуществляется в виде слияния гамет с образованием зиготы. Для многих видов характерны сложные жизненные циклы в результате смены поколений с разными формами размножения.
 Для переживания неблагоприятных условий протисты образуют споры (водоросли и грибы) или цисты (одноклеточные животные).
Для переживания неблагоприятных условий протисты образуют споры (водоросли и грибы) или цисты (одноклеточные животные). - Протистам присуща раздражимость — способность воспринимать раздражители окружающей среды и определенным образом на них отвечать. Двигательные реакции на раздражители имеют название таксисов.
Вопросы для самоконтроля
1. Какие формы движения известны у протист? Какие органеллы их обеспечивают? 2. Как у протист удаляются остатки пищи и выделяются конечные продукты обмена веществ? 3. Как размножаются протисты? 4. Что такое конъюгация? Каково ее биологическое значение? 5. Как протисты переживают неблагоприятные условия?
Подумайте
Что общего и отличного в размножении прокариот и одноклеточных эукариот?
Попередня
СторінкаНаступна
СторінкаЗміст
Цей контент створено завдяки Міністерству освіти і науки України
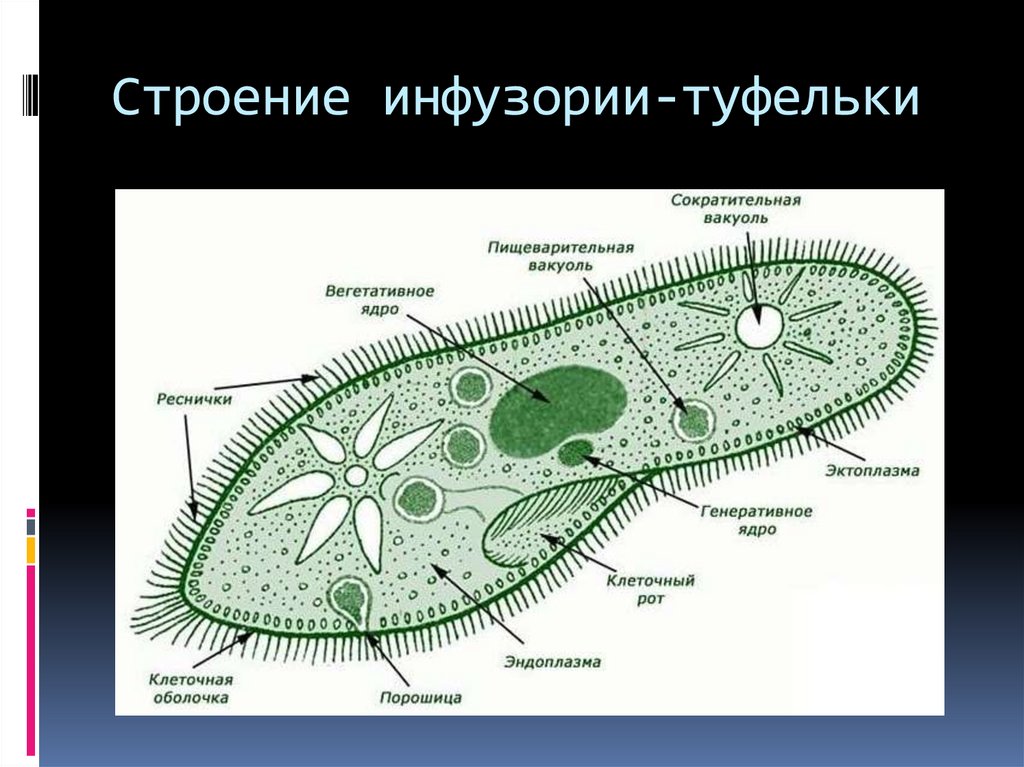
Инфузория туфелька.
 Многообразие инфузорий
Многообразие инфузорийМюнхгаузен расписал быт инфузорий в таких очаровательных красках,
Что его слушатель пришел в восторг; он нарисовал ему
Целые идиллии, эпопеи и трагедии,
Которые, по его заверению, происходят в каждой капле воды.
К. Иммерман.
Инфузории покрыты ресничками в течение всей жизни. Питаются через цитостом или осмотически. Размножение – преимущественно бесполое, поперечным делением. Имеет место половой процесс – конъюгация и половое размножение.
Представитель – инфузория туфелька. Свободноживущее, пресноводное простейшее. Ее цитоплазма богата сложными органеллами, выполняющими специфические функции. Характеризуется высоким уровнем протоплазматической дифференцировки.
Вся клетка покрыта тонкой и гибкой пелликулой, под которой располагается прозрачный слой плотной эктоплазмы. В ней находятся кинетосомы – базальные тела ресничек, связанные с комплексом фибрилл, регулирующих работу ресничек.
На границе эктоплазмы и зернистой эндоплазмы расположены продольные тяжи микрофибрилл (мионем). Их сокращение меняет форму тела инфузрии.
Их сокращение меняет форму тела инфузрии.
По краю тела в эктоплазме видны мелкие колбообразные структуры – трихоцисты. Выполняют защитную роль. А также, при действии раздражителя, способны выбрасывать тонкие остроконечные нити, поражающие добычу.
Околоротовая воронка (перистом) является постоянным образованием и находится ближе к переднему концу. Сужаясь, она переходит в клеточную глотку – цитофаринкс, которая заканчивается участком эндоплазмы, не имеющим пелликулу – клеточным ртом – цитостом.
Реснички околоротовой воронки загоняют в глотку вместе с водой бактерии и другие объекты питания. В цитостоме, в эндоплазме вокруг них формируется пищеварительная вакуоль. Питание туфкльки проходит способом фагоцитоза – коетка заглатывает частицы, заключая их в мембранный пузырек. Сливаясю с пищеварительной вакуолью лизосомы вводят в нее пищеварительные фенрменты. Через некоторое время крупные молекулы разбиваются на более мелкие. Эти мелкие молекулы проходят через мембрану вакуоли в цитоплазму. Это – внутриклеточное пищеварение. Непереваренные остатки выходят через порошицу (анальную пору) путем эзкоцитоза.
Это – внутриклеточное пищеварение. Непереваренные остатки выходят через порошицу (анальную пору) путем эзкоцитоза.
Осморегуляция осуществляется парными сократительными вакуолями. Вокруг каждой из них располагается несколько радиальных каналов (резервуаров), в которые поступает вода из цитоплазмы. Периодически радиальные каналы опорожняются в сократительные вакуоли, выводящие воду во внешнюю среду.
Для инфузорий характерно наличие двух ядер макронуклеуса (полиплоидное ядро, контролирующее обменные процессы, процессы дифференцировки) и микронуклеус (диплоидное ядро, регулирующее процесс размножения).
Размножение преимущественно поперечным делением пополам. При этом макронуклеус делится амитотически, и микронуклеус – митозом. При недостатке пищи инфузории переходят к конъюгации.
Процесс конъюгации характеризуется тем, что:
1. Две совместимые особи (конъюганты) прикрепляются друг к другу перистомиальными областями.
2. Пелликула разрушается, и между особями образуется цитоплазматический мостик.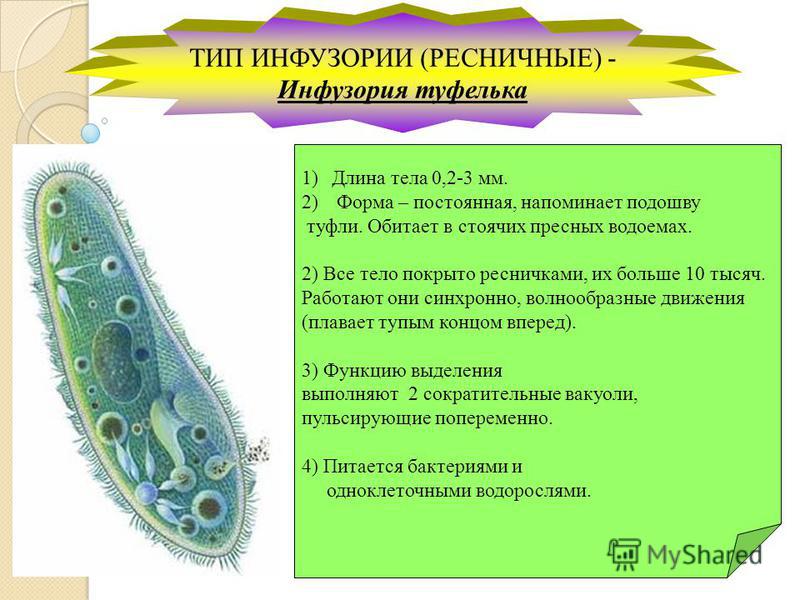 Происходит дезынтеграция (растворение) макронуклеусов. Микронуклеусы делятся митотически, образуя по четыре дочерних ядра.
Происходит дезынтеграция (растворение) макронуклеусов. Микронуклеусы делятся митотически, образуя по четыре дочерних ядра.
3. Три микронуклеуса разрушаются и исчезают.
4. Оставшееся ядро в каждом из конъюгантов делится митозом, образуя два пронуклеуса. Одно из пронуклеусов стационарное (женское), а другое перемещается по цитоплазматическому мостику в клетку партнера (подвижное, мужское ядро).
5. Ядра сливаются с образованием синкариона. В результате происходит обмен генетической информацией.
6. Конъюганты расходятся. Синкарион делится митозом с образованием 8 ядер.
7. Из этих ядер четыре становятся макронуклеусами, а четыре – микронуклеусами. Затем три микронуклеуса разрушаются.
8. Каждый из конъюгатов делится надвое. При этом макронуклеусы расходятся попарно в каждую клетку, а микронуклеус делится митотически.
9. Затем следует второе деление клетки надвое. Макронуклеусы расходятся по одному в каждую клетку, микронуклеус делится митозом. Таким образом каждый родительский конъюгант дает начало четырем дочерним особям.
Однако, при длительном бесполом размножении, когда макронуклеус делится амитозом и хромосомы произвольно распределяются между дочерними особями, жизнедеятельность инфузорий может нарушаться, наступает «депрессия». Процесс, направленный на ликвидацию этих последствий и сходный с половым называется аутогамия. Он приводит к образованию нового макронуклеуса, содержащего полный набор хромосом и повторяется 3 – 4 недели:
1. Микронуклеус делится, образуя восемь гаплоидных ядер. Шесть из них разрушаются. Макронуклеус тоже лизируется.
2. Два оставшихся ядра образуют синкарион.
3. Синкарион дважды делится, и из четырех образовавшихся ядер два становятся макронуклеусами, а два – микронуклеусами.
4. Инфузория делится пополам. Теперь макронуклеусы у дочерних инфузорий имеют нормальный набор хромосом.
В каждой щелочке,
В каждом узоре
Жизнь богата и многогранна.
Всюду – даже среди инфузорий
Лилипуты
И великаны
М.Светлов.
Среди видов туфелек привлекает внимание зеленая Paramecium bursaria.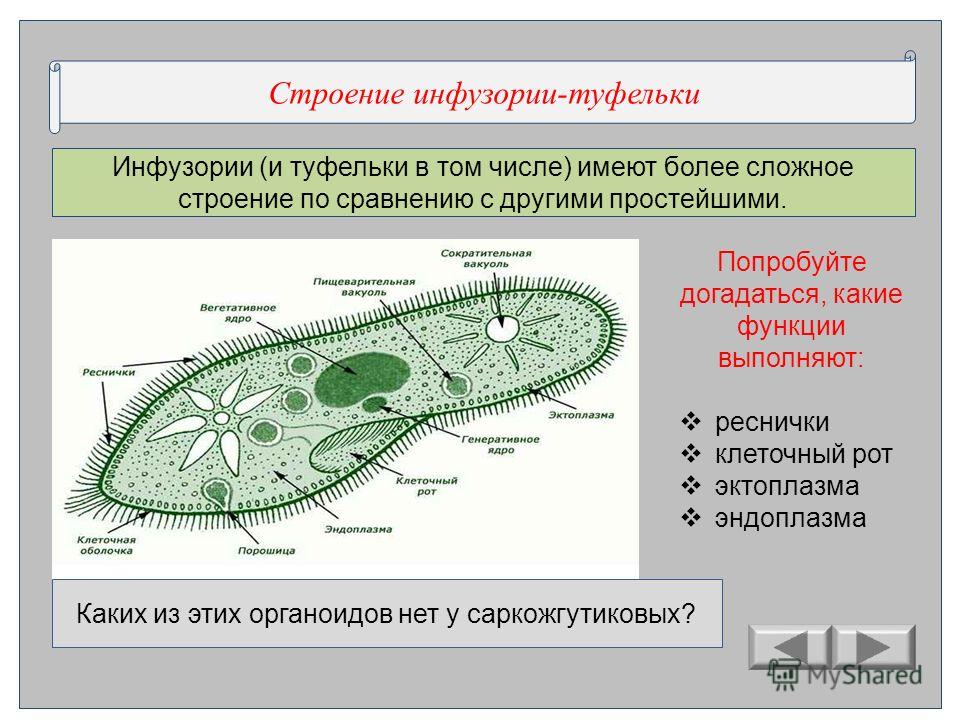 В ее цитоплазме обитают одноклеточные водоросли зоохлореллы. Это симбионты туфельки. В процессе фотосинтеза она образуют сахара, что является кормом для клетки-хозяина. А туфелька не перерваривает зоохлорелл. Они живут в вакуолях, подобных пищеварительным. Размножаются зоохлореллы также внутри туфельки. В пресных водоемах встречаются также инфузории – трубачи, бегающие по дну брюхоресничные инфузории. Напоминающие колокольчик сувойки.
В ее цитоплазме обитают одноклеточные водоросли зоохлореллы. Это симбионты туфельки. В процессе фотосинтеза она образуют сахара, что является кормом для клетки-хозяина. А туфелька не перерваривает зоохлорелл. Они живут в вакуолях, подобных пищеварительным. Размножаются зоохлореллы также внутри туфельки. В пресных водоемах встречаются также инфузории – трубачи, бегающие по дну брюхоресничные инфузории. Напоминающие колокольчик сувойки.
Некоторые сувойки связаны между собой стебельком. Это – колониальные инфузории. Колонии возникают в результате деления клеток. Например, бродяжек у вида зоотамния. Инфузории могут быть хищниками. Например дидиниум – пожиратель туфелек. Вокруг цитостома у него есть мембранные пузырьки я ядом. При столкновении с туфелькой яд парализует жертву. Сильно растягивая клеточный рот дидиниум заглатывает добычу. Хищниками являются сосущие инфузории – суктории. Они ловят добычу при помощь щупалец. Прилипшая добыча парализуется, а затем суктория высасывает ее содержимое.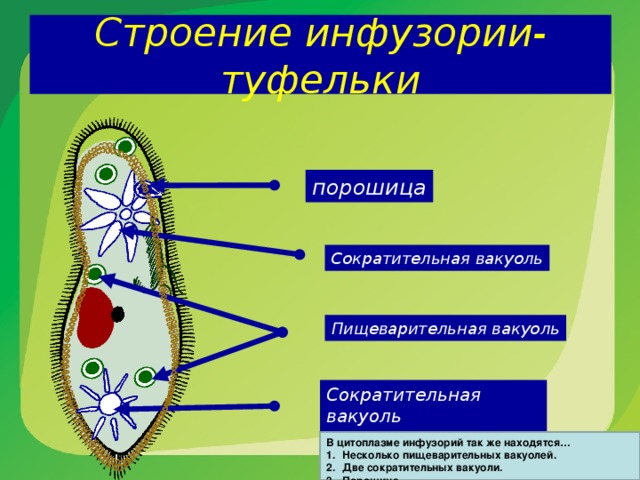 Есть морские инфузории.
Есть морские инфузории.
Немало инфузорий обитают в макрообранизмах. Например, в рубце у коровы. Участвуют в переваривании клетчатки. В другие организмы попадают через воду при питье коровы.
У споровиков нет ни рта, ни глотки, и они питаются осмотически.
Токсоплазма – возбудитель токсоплазмоза человека, грызунов, других видов млекопитающих, птиц. Жизненный цикл состоит из двух фаз: кишечной и тканевой. Если в цикле развития присутствуют промежуточные хозяева, то у них наблюдается только тканевая фаза.
Кишечная фаза цикла развития проходит у представителей семейства кошачьих, которые являются окончательными хозяевами. Состоит из шизогонии, гаметогонии и спорогонии.
1. Объект исследования больной человек
2. Диагностический материал – мазок фекалий.
3. Способ диагностики – микроскопирование нативного мазка фекалий.
4. Анализ микропрепарата позволяет отнести обнаруженных простейших к классу Саркодовые, вид – дизентерийная амеба.
5. Стадия
развития
Entamoeba histolytica f. magna.
Стадия
развития
Entamoeba histolytica f. magna.
Тип: Простейшие (Protozoa)
Подтип: Ciliophora
Класс: Инфузории (Infusoria)
Вид: Заболевание:
Balantidium coli Балантидиаз
КЛАСС ИНФУЗОРИИ (INFUSORIA).
По сравнению с
другими группами простейших инфузории
имеют наиболее высоко дифференцированное
строение. Связано это с разнообразием
и сложностью функций. Тело инфузорий
покрыто плотной пелликулой, что сохраняет
им постоянную форму. Характерными
признаками класса инфузорий являются
волосовидные выросты цитоплазмы-
реснички- органоиды движения. Реснички,
покрывающие всю инфузорию или часть
ее, морфологически сходны со жгутиками,
но относительно короче. Основание каждой
реснички заканчивается в эктоплазме
базальным тельцем. В физиологическом
отношении реснички отличаются от
жгутиков тем, что движение их состоит
из резкого взмаха в одном направлении
и более медленного, плавного возвращения
к исходному положению.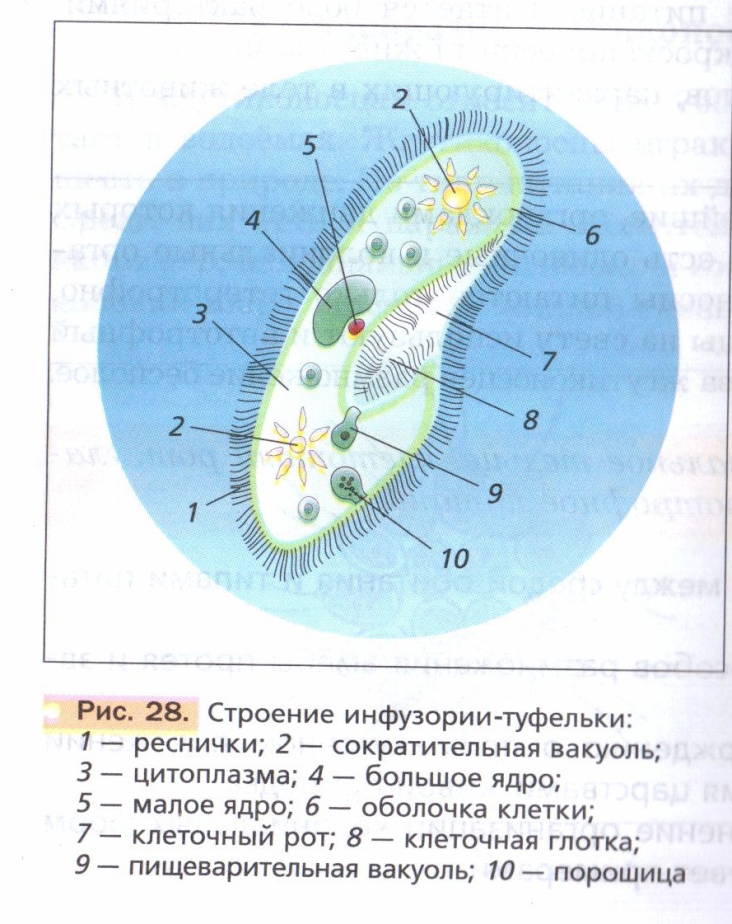
Около рта реснички, в связи с функцией захвата пищи, нередко становятся более мощными, длинными и даже склеиваются по нескольку вместе в прозрачные пластинки, мембранеллы. Если таким образом сливается друг с другом длинный ряд ресничек, то на месте его получается хлопающая волнообразная перепонка.
Тело делится на наружный слой — эктоплазму, и внутреннюю часть — эндоплазму. Эктоплазма имеет сложное строение и часто содержит в себе пробегающие в продольном направлении сократительные волоконца, мионемы. Благодаря последним, многие инфузории обладают очень сильной сократимостью и сжимаются при раздражении в маленький комок. Эндоплазма, занимающая всю центральную часть тела, представляет собой зернистую полужидкую массу, которая находится в постоянном круговом движении.
Вторым
важным общим признаком инфузорий
является присутствие в их теле по меньшей
мере двух ядер, и притом различных по
своим свойствам. Одно из них (макронуклеус)
всегда значительно крупнее другого
(микронуклеуса).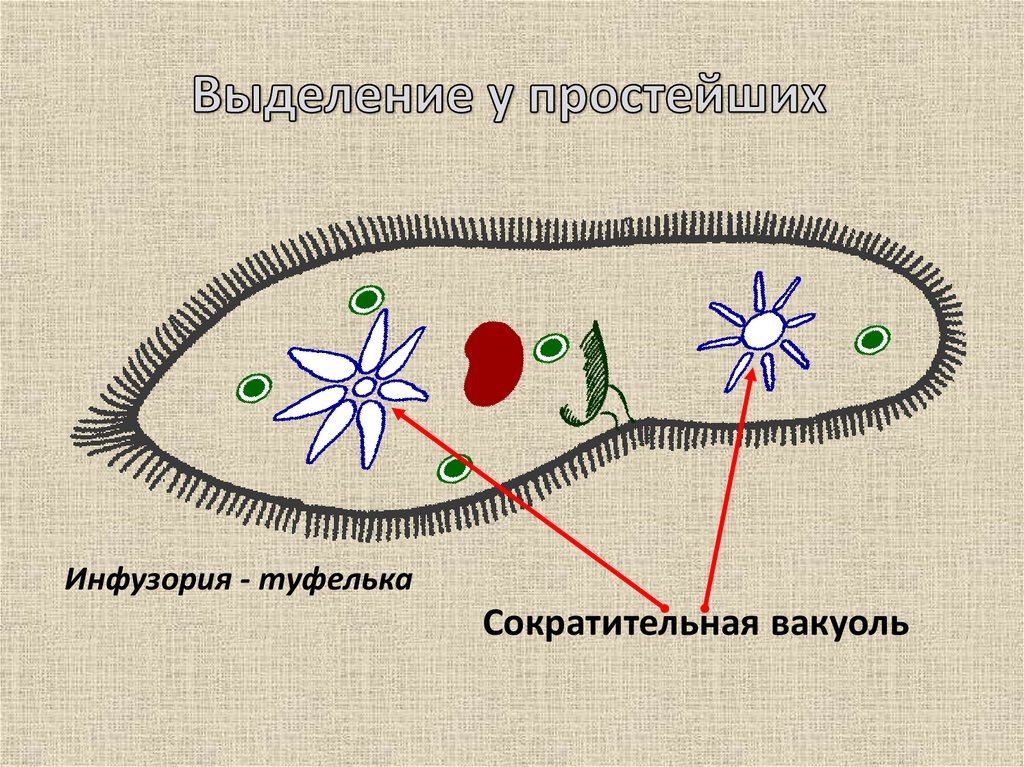 Крупный макронуклеус состоит
из множества тесно сближенных зёрен
хроматина.
Микронуклеус в виде маленького
шарика прилегает вплотную к большому
ядру. Макронуклеус
контролирует обмен веществ и
движение. Микронуклеус играет доминирующую
роль в размножении.
Крупный макронуклеус состоит
из множества тесно сближенных зёрен
хроматина.
Микронуклеус в виде маленького
шарика прилегает вплотную к большому
ядру. Макронуклеус
контролирует обмен веществ и
движение. Микронуклеус играет доминирующую
роль в размножении.
Бесполое размножение происходит посредством деления тела надвое в поперечном направлении. Это сопровождается делением обоих ядер, причем макронуклеус делится путем простой перешнуровки (прямое деление, или амитоз). Микронуклеус может делиться с помощью митоза или мейоза.
Время от времени в жизненном цикле инфузорий наблюдается половой процесс, который носит у них характер коньюгации, главное отличие коньюгации инфузорий от раньше описанных нами половых процессов заключается в том, что она состоит во временном, преходящем соединении двух инфузорий; последние обмениваются в это время частями своего ядерного аппарата, после чего расходятся.
Во
время коньюгации инфузории
сходятся попарно и прикладываются
друг к другу
брюшной стороной, далее на месте
соприкосновения пелликула обеих особей
растворяется
и между ними
получается соединительный
мостик из плазмы.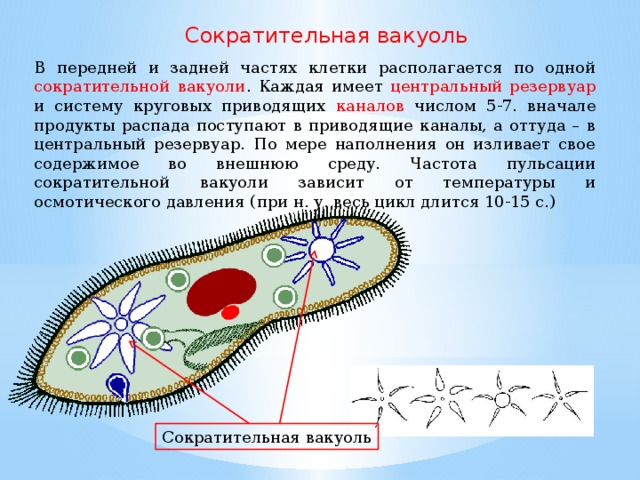 Самые сильные изменения
испытывает во время конъюгации
ядерный аппарат.
Макронуклеус
конъюгантов постепенно
рассасывается
в
плазме. Микронуклеус
делится мейозом в результате
получаются 4 дочерних
ядра, 3 из них гибнут, а четвертое снова
делится
митозом. В результате каждый
конъюгант обладает
двумя маленькими ядрами,
стационарным и мигрирующим. Мигрирующее
ядро(условно мужское) переходит через
цитоплазматичексий мостик в тело
другого коньюганта и сливается там со
стационарным ядром (условно женским).
К
концу конъюгации каждый конъюгант имеет
по одному ядру
двойственного происхождения или
синкариону.
К
этому времени обе инфузории отрываются,
друг от друга и снова приобретают полную
самостоятельность.
Самые сильные изменения
испытывает во время конъюгации
ядерный аппарат.
Макронуклеус
конъюгантов постепенно
рассасывается
в
плазме. Микронуклеус
делится мейозом в результате
получаются 4 дочерних
ядра, 3 из них гибнут, а четвертое снова
делится
митозом. В результате каждый
конъюгант обладает
двумя маленькими ядрами,
стационарным и мигрирующим. Мигрирующее
ядро(условно мужское) переходит через
цитоплазматичексий мостик в тело
другого коньюганта и сливается там со
стационарным ядром (условно женским).
К
концу конъюгации каждый конъюгант имеет
по одному ядру
двойственного происхождения или
синкариону.
К
этому времени обе инфузории отрываются,
друг от друга и снова приобретают полную
самостоятельность.
У
всех инфузорий происходит усложнение
пищеварительной системы. На передней
части находится ротовое отверстие-
цитостом, которое ведет в
глубокий канал —
глотку. На
дне глотки образуется
маленькая
пищеварительная вакуоль. Наполнившись
пищей, вакуоль отрывается
от глотки и увлекается током эндоплазмы,
описывая в теле животного
определенный путь.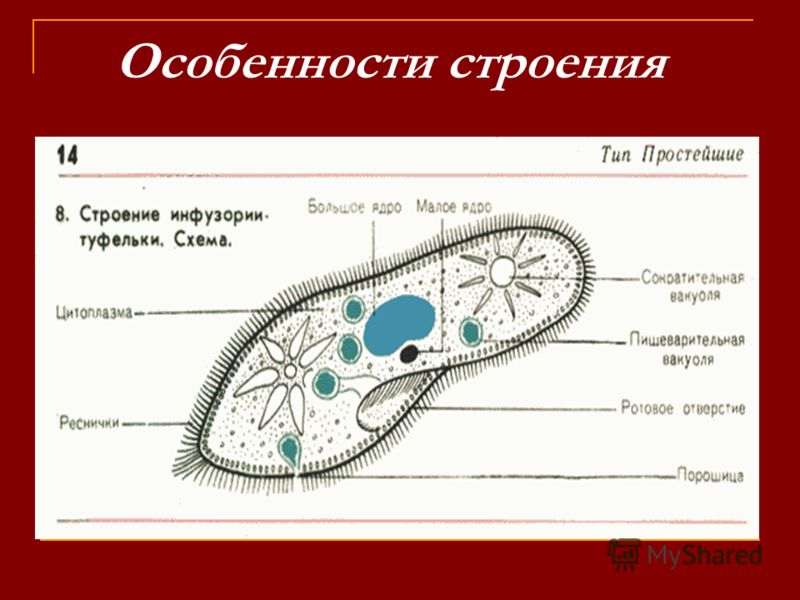 Оставшиеся
внутри
вакуоли непереваренные остатки
пищи выталкиваются вместе с
ней через особое, находящееся неподалеку
от заднего конца тела отверстие —
порошицу. Пищеварительные
вакуоли образуются на дне глотки через
каждые 1,5 – 2 минуты. Интересно, что
первые стадии пищеварения
протекают при кислой, последующие
— при щелочной реакциях.
Эта смена реакций аналогична двум
фазам пищеварения (пепсинному
в желудке и трипсинному в тонком
кишечнике) у высших животных. Жидкие
продукты выделения, удаляются, через
2 пульсирующие вакуоли. Они имеют
довольно сложное строение.
Типично этот органоид состоит из
собственно вакуоли, окруженной
венчиком приводящих каналов. Кроме
того, вакуоль при помощи тонкого выводного
канальца сообщается с
наружной средой. Выделяемая жидкость
собирается из протоплазмы сначала
в каналы; последние сокращаются и
опоражнивают свое содержимое
в вакуоль, которая при этом раздувается
(стадия диастолы). Далее происходит
сокращение самой вакуоли (систола),
выталкивающее жидкость
наружу, между тем как приводящие каналы
вновь наполняются жидкостью
и т.
Оставшиеся
внутри
вакуоли непереваренные остатки
пищи выталкиваются вместе с
ней через особое, находящееся неподалеку
от заднего конца тела отверстие —
порошицу. Пищеварительные
вакуоли образуются на дне глотки через
каждые 1,5 – 2 минуты. Интересно, что
первые стадии пищеварения
протекают при кислой, последующие
— при щелочной реакциях.
Эта смена реакций аналогична двум
фазам пищеварения (пепсинному
в желудке и трипсинному в тонком
кишечнике) у высших животных. Жидкие
продукты выделения, удаляются, через
2 пульсирующие вакуоли. Они имеют
довольно сложное строение.
Типично этот органоид состоит из
собственно вакуоли, окруженной
венчиком приводящих каналов. Кроме
того, вакуоль при помощи тонкого выводного
канальца сообщается с
наружной средой. Выделяемая жидкость
собирается из протоплазмы сначала
в каналы; последние сокращаются и
опоражнивают свое содержимое
в вакуоль, которая при этом раздувается
(стадия диастолы). Далее происходит
сокращение самой вакуоли (систола),
выталкивающее жидкость
наружу, между тем как приводящие каналы
вновь наполняются жидкостью
и т. д. Вакуоли выполняют не только
выделительную, но и осморегулирующую
функцию.
д. Вакуоли выполняют не только
выделительную, но и осморегулирующую
функцию.
4.3. Медицинское значение класса Инфузории . Морфологическая характеристика, жизненный цикл, диагностика и профилактика балантидиаза.
БАЛАНТИДИЙ — Balantidium coli — возбудитель балантидиаза — антропозооноза.
Географическое распространение — повсеместно.
Локализация — толстый кишечник, особенно часто слепая кишка.
Цикл развития.
Источником распространения балантидиаза по современным представлениям чаще всего являются домашние и дикие свиньи, реже — больной человек или цистоноситель. В отличие от человека балантидий у свиней не вызывает болезненных явлений. Наблюдение показывает, что большой процент носителей балантидиаза среди рабочих свиноводческих ферм. Заражение происходит при уходе за животными, уборке помещений и т.д. Заражение балантидиазом возможно в колбасном производстве и при заготовке кишечного сырья.
Заражение
происходит алиментарным путем,
перорально. Под действием ферментом
желудочно-кишечного тракта оболочка
цисты растворяется и в кишечнике выходят
вегетативные формы. В течении длительного
времени балантидий может не вызывать заболевания,
то есть развивается носительство. Он
обитает в просвете толстого кишечника,
питается бактериями и крахмальными
зернами. Однако, при заболеваниях
желудочно-кишечного тракта, вирусных
инфекциях, приводящих снижению защитных
функций организма, при злоупотреблениях
жирной пищей, при дефиците углеводов в
пище и т.д. балантидий начинает выделять
протеолитический фермент, за счет
которого внедряется в стенку кишечника.
Под действием ферментом
желудочно-кишечного тракта оболочка
цисты растворяется и в кишечнике выходят
вегетативные формы. В течении длительного
времени балантидий может не вызывать заболевания,
то есть развивается носительство. Он
обитает в просвете толстого кишечника,
питается бактериями и крахмальными
зернами. Однако, при заболеваниях
желудочно-кишечного тракта, вирусных
инфекциях, приводящих снижению защитных
функций организма, при злоупотреблениях
жирной пищей, при дефиците углеводов в
пище и т.д. балантидий начинает выделять
протеолитический фермент, за счет
которого внедряется в стенку кишечника.
Патогенное действие. При внедрении в слизистую кишечника образуются гангренозные язвы 3-4 см в диаметре. Развиваются кровавые поносы, приводящие к истощению организма. Общетоксическое действие балантидия в виде тошноты, рвоты, головной боли, повышения температуры.
Диагностика. Обнаружение цист и вегетативных форм
в фекалиях, ядра которых имеют характерную
бобовидную форму.
Профилактика: а) общественная — обследование, выявление и лечение больных и носителей, которые наиболее часто встречаются среди рабочих свиноводческих ферм и работников колбасного производства; б) личная — соблюдение правил личной гигиены (мытье рук, овощей, фруктов, кипячение воды).
Кугарчинский район
Инфузория-туфелька. Многообразие инфузорий
Учитель биологии
Мраково-2011
Тема: Инфузория туфелька. Многообразие инфузорий.
Цели:
Познавательные: Изучить особенности строения, поведения и процессы жизнедеятельности у инфузории туфельки.
Развивающие: Развивать умения сравнивать и анализировать процессы жизнедеятельности простейших, и переносить знания в новую ситуацию.
Воспитательные: Формирование научного мировоззрения; воспитание эстетического отношения к окружающему миру.
Оборудование: Мультимедийный проектор, экран, учебный диск « Виртуальная школа Кирилла и Мефодия.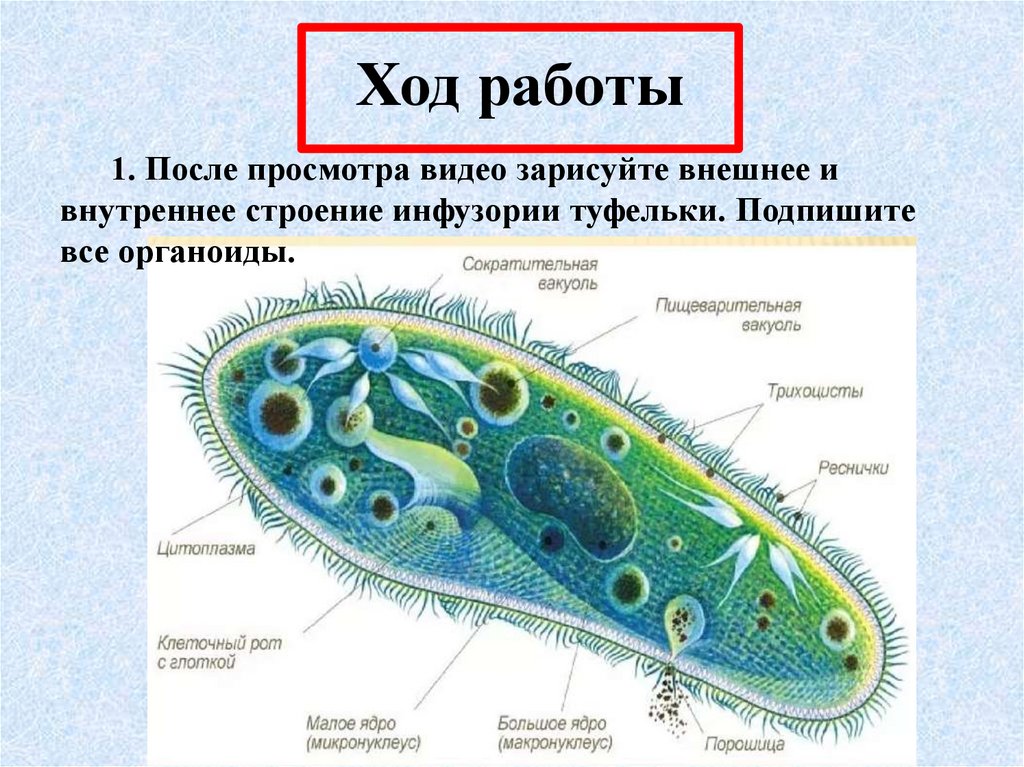 Уроки биологии», таблица «Тип Простейшие». Микроскоп, готовый микропрепарат «Инфузория-туфелька», цветной картон.
Уроки биологии», таблица «Тип Простейшие». Микроскоп, готовый микропрепарат «Инфузория-туфелька», цветной картон.
Тип урока: комбинированный.
Вид урока: смешанный.
Формы работы : индивидуальная, групповая.
Методы и методические приемы: беседа, рассказ, частично — поисковый метод.
Ход урока:
׀) Организационный момент.
Сегодня наш урок мне хотелось бы начать со стихотворения
Человек рождается на свет,
Чтоб творить, дерзать – и не иначе,
Чтоб оставить в жизни добрый след
И решить все трудные задачи.
На предыдущем уроке мы изучили тип Саркожгутиконосцы, и мне хотелось бы выяснить, как вы усвоили этот материал. Для этого проведем интеллект – шоу «Эврика».
׀׀) Актуализация знаний
1 Конкурс «Третий – лишний» (Презентация. Приложение 1).
Вы должны найти смысловое несоответствие и пояснить его.
а) Амеба, радиолярия, фораминифера (Амеба и фораминифера
2 Конкурс «Интересные слова» (Приложение 2).
Объясните значение следующих слов:
а) Циста (плотная защитная оболочка, образуется при наступлении холодов)
б) Пиноцитоз (поглощение воды клеткой)
в) Фагоцитоз (активное захватывание и поглощение живых объектов и твердых частиц)
г) Миксотрофы (смешанный тип питания: автотрофное и гетеротрофное у зеленой эвглены)
д) Стигма (светочувствительный глазок у зеленой эвглены)
е) Таксис (двигательная реакция в ответ на односторонне действующий раздражитель)
ж) Сапрофит (организм, питающийся продуктами распада органических веществ)
з) Псевдоподии (выступы цитоплазмы клетки, выполняющие функцию органелл движения)
3 Конкурс «Найди ошибку» (Приложение 3).
Перед вами таблица: Сравнительная характеристика простейших, в ней есть ошибки. Ваша задача выявить их. Работу ведете индивидуально, а затем проведем самооценку и взаимооценку в парах.
Структура | Эвглена зеленая | |
1. | ||
2.Цитоплазма | ||
4.Ложноножки | ||
5.Жгутики | ||
6.Пищеварительная вакуоль | ||
7.Сократительная вакуоль | ||
8.Хлоропласт | ||
Ошибки: У амебы: 4,5,6,8.
У эвглены зеленой 2,4,5,6.
4 конкурс «Верны ли утверждения? » (Приложение 4).
При верном суждении вы должны показать зеленый сигнал светофора, если утверждение неверно, то красный.
1. Кислород в цитоплазму амебы и эвглены зеленой поступает через всю поверхность тела. (з)
2. Продукты обмена веществ и избытки воды из тела простейших удаляются через сократительную вакуоль (з)
3. При неблагоприятных условиях большинство простейших переходят в состояние цисты. (з)
4. Эвглена зеленая участвует в образовании известняка. (к)
5. Размножение у амебы половое и бесполое (к)
Верные утверждения: 1,2,3.
С конкурсными с заданиями вы справились, а сейчас мне хочется поиграть с вами.
У меня на столе лежит конверт, а в нем разорванная загадка. Попытайтесь ее восстановить и разгадать (изначально загадка записывается на картоне, затем разрезается)
Здравствуйте, мои друзья.
Вы же знаете меня!
Мне «реснички» очень просто
Заменяют микровесла
За секунду 30 взмахов
Могу ими совершать
И на четверть сантиметра
Вперед тело продвигать
Так кто же я?
(Инфузория – туфелька) (Приложение 5).
׀׀׀) Изучение новой темы:
Откройте тетради и запишите тему урока: «Инфузория – туфелька. Многообразие инфузорий». А какие цели мы поставим перед собой?
План урока: (Приложение 6).
1) Систематическое положение инфузории – туфельки;
2) Знакомство со средой обитания, внешнее строение инфузории – туфельки;
3) Внутреннее строение инфузории туфельки;
4) Питание;
5) Дыхание и обмен веществ;
6) Бесполое размножение;
7) Половое размножение;
8) Многообразие инфузорий;
9) Происхождение и родственные связи.
Каждое животное имеет свое определенное место в системе органического мира. Вот и наша инфузория – туфелька, которая только что появилась на свет имеет право получить свидетельство о рождении. Ребята, подскажите ее систематическое положение.
1) Систематика инфузории – туфельки .(Приложение 7).
Империя – клеточные
Надцарство – ядерные
Царство – животные
Подцарство – одноклеточные (простейшие)
Тип – инфузории (7,5 тыс. видов)
Класс –Ресничные инфузории, подкласс Равноресничные инфузории
Род – Парамеции
Вид — инфузория – туфелька
Ответьте на вопрос: « Кто впервые открыл простейших?»
(В 1675г. Голландский естествоиспытатель Антуан Ван Левенгук). (Приложение 8). Левенгук выяснил, что инфузорий можно разводить в настоях трав. Инфузория — от латинского слова «инфузум», что значит настой, наливка.
2)Знакомство со средой обитания, внешним строением инфузории – туфельки.
Этот организм можно встретить в водоёмах с опавшими листьями, где много органических останков.
Пресноводное простейшее длиной 0,2 – 0,3 мм. За 1 секунду проплывает путь, в 8 – 10 раз превышающий длину ее тела. Рекордсмен среди простейших. В процессе эволюции возникло важное приспособление для передвижения — реснички. Заостренный конец является — задним, а тупой конец – передним. Передвигается задним концом вперед. У неё постоянная форма тела. Поверхность клетки покрыта ресничками их количество около 10 тысяч. Клеточная оболочка из клеточной мембраны и эктоплазмы. Между ресничками находятся веретеновидные тельца трихоцисты-органоиды защиты и нападения. (Приложение 9).
3) Изучим особенности внутреннего строения инфузории-туфельки (Приложение 10). Вы должны заполнить в тетрадях таблицу « Особенности строения инфузории»
Таблица Особенности строения
Структура | Инфузория — туфелька |
Оболочка | |
Цитоплазма | |
2+ большое, малое | |
Реснички | |
Пищев. вакуоль | |
Сократ. вакуоль | |
Рот. отверствие | |
Порошица |
Проведём проверку вашей памяти и внимания, используя тренажер. Внимательно смотрите на экран. Читая вопрос, заполните пропуски органелл методом подстановки. После выполнения этого задания подготовьте микроскоп к лабораторной работе , рассмотрите микропрепарат «Инфузория-туфелька». Ответьте на вопрос: « Что вы увидели, изучая микропрепарат?» (Приложение 11).
4)Выясним питание инфузории — туфельки (Приложение 12). От переднего конца до середины тела туфельки проходит желобок с более длинными ресничками. На заднем конце желобка имеется рот – ротовое отверстие, ведущее в трубчатую глотку. Реснички желобка непрерывно работают, создавая ток воды. Вода подхватывает и подносит ко рту основную пищу туфельки-бактерии. Через глотку бактерии попадают внутрь тела инфузории. Здесь, в цитоплазме, вокруг них образуется пищеварительная вакуоль, в которую выделяется пищеварительный сок. Цитоплазма у туфельки находится в постоянном движении. Вакуоль открывается от глотки и подхватывается течением цитоплазмы.
Переваривание пищи и усвоение питательных веществ у инфузории происходит так же, как у амёбы. Непереваренные остатки выбрасываются наружу через отверстие-порошицу.
5) Дыхание и обмен веществ (Приложение 13).
Дыхание через всю поверхность тела. Обмен веществ, происходит за счёт двух попеременно сокращающихся сократительных вакуолей. Они сокращаются через 20-25 секунд каждая. Вода и вредные продукты жизнедеятельности собираются у туфельки из всей цитоплазмы по приводящим канальцам, которые подходят к сократительным вакуолям.
Твёрдые продукты обмена удаляются через порошицу.
6)Наша инфузория с наступлением лета, после того как вырастит, готовится к размножению. Она может размножаться бесполым и половым путем. Половой путь наиболее прогрессивный. (Приложение 14).
Этот процесс происходит с периодичностью в одни сутки. Все начинается с деления генеративного ядра. Оно делится на две части и каждая часть отходит к переднему и заднему концу. Затем происходит удвоение клеточного рта, и образование зачатков 2 новых сократительных вакуолей. Потом делится вегетативное ядро, новые сократительные вакуоли образуют систему канальцев, а цитоплазма в центре клетки образуют глубокую перетяжку и материнская клетка делится на 2 молодые инфузории.
7)Посмотрим (Приложение 15). «Половое размножение» (конъюгация)
Две инфузории сходятся поверхностями в области клеточного рта. В области их контакта, между клетками образуются цитоплазматические мостики. Вегетативное ядро исчезает. Генеративное ядро каждой клетки делится дважды, после этого 3 из 4 ядер исчезают. Оставшееся ядро делится еще раз. Одно из образовавшихся ядер остается в клетке неподвижно, а другое перетекает по цитоплазматическому мостику в соседнюю клетку и происходит обмен ядер (генетической информацией) и их слияние.
Инфузории восстанавливают свои оболочки и расходятся. В результате деления нового ядра в клетке образуется вегетативное и генеративное ядро. Усложнение ядерного аппарата – ароморфоз. За один год потомство инфузорий может достичь 75*10¹º³ особей. Но этого не происходит, как вы думаете почему?
4)Закрепление.
Посмотрите на экран, здесь показано родословное древо животного мира (Приложение 17), и ответьте на вопрос: « От кого произошли инфузории?»
(От жгутиковых) Для проверки знаний по теме каждый ученик получит карточку-рассказ «Путешествие в мир инфузорий», которую должен заполнить, затем проводится взаимоконтроль.
Карточка – задание (Приложение 18).
«Путешествие в мир инфузорий».
Впишите пропущенные слова в предложениях:
1. На встречу с вами приплыла инфузория — туфелька, она активно работала своими органами передвижения………………
2. От врагов она защищалась веретенообразными тельцами ………….
3. Инфузория – туфелька торопилась к нам и едва не обожгла свое тело опасными химическими веществами, но успела применить поведенческую реакцию -……………., благодаря чему осталась жива.
4. По дороге к нам она успела позавтракать лишь бактериями, то есть тип ее питания…………………..
5. В пути она встретила жвачных животных, которые приглашали ее на взаимовыгодное сожительство…………………….
6. Она рассказала о новообразованиях, которыми наделила ее природа, в отличие от всех простейших:
а) пища в нее попадает через …………………, непереваренные остатки выбрасываются наружу через ……………………
б) лишняя вода удаляется за счет двух……………………вакуолей
в) два способа размножения — ………………. и………………….
г) при конъюгации основная роль падает на ………………ядро.
д) в геноме человека 25 тысяч генов, а у инфузории — туфельки на 15 больше……………….
7. Инфузория – более ……устроена, чем амеба, эвглена зеленая.
Ответы к карточке «Путешествие в мир инфузорий»:
1 ресничками
2 трихоцистами
3 хемотаксис
4 гетеротрофный
5 симбиоз
6 а) рот, порошицу
б) сократительных
в) половое и бесполое
д) 40 тысяч
5) Итоги урока
а) Оценивание
б) Домашнее задание.
Подготовить сообщения по темам:
Инфузории — классический объект биохимических исследований.
Очистка сточных вод от азота и фосфора микроорганизмами (активный ил).
Аквариумные рыбки и инфузории.
Инфузории, встречающиеся в республике Башкортостан.
Составить кроссворды, шарады
| Вид: | Инфузория-туфелька | ||
| Латинское название | |||
|---|---|---|---|
| Paramecium caudatum Ehrhart | |||
|
Функции цитоплазматического динеина в планарной поляризации базальных телец внутри реснитчатых клеток
. 2020 26 июня; 23 (6): 101213.
doi: 10.1016/j.isci.2020.101213. Epub 2020 1 июня.
Маки Такагиши 1 , Нобутоши Эсаки 2 , Кунихико Такахаши 3 , Масахидэ Такахаши 4
Принадлежности
- 1 Кафедра патологии, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония. Электронный адрес: [email protected]. jp.
- 2 Кафедра патологии, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония.
- 3 Кафедра биостатистики, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония; Департамент биостатистики, Центр научных исследований данных M&D, Токийский медицинский и стоматологический университет, Бункё-ку, Токио 113-8510, Япония.
- 4 Кафедра патологии, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония; Международный центр клеточной и генной терапии, Университет здоровья Фудзита, Тойоаке, Аити 470-1192, Япония.
- PMID: 32535020
- PMCID: PMC7300155
- DOI:
10. 1016/j.isci.2020.101213
Бесплатная статья ЧВК
Маки Такагиши и др. iНаука. .
Бесплатная статья ЧВК
. 2020 26 июня; 23 (6): 101213.
doi: 10.1016/j.isci.2020.101213. Epub 2020 1 июня.
Авторы
Маки Такагиши 1 , Нобутоши Эсаки 2 , Кунихико Такахаши 3 , Масахидэ Такахаши 4
Принадлежности
- 1 Кафедра патологии, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония. Электронный адрес: [email protected].
- 2 Кафедра патологии, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония.
- 3 Кафедра биостатистики, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония; Департамент биостатистики, Центр научных исследований данных M&D, Токийский медицинский и стоматологический университет, Бункё-ку, Токио 113-8510, Япония.
- 4 Кафедра патологии, Высшая школа медицины Университета Нагоя, Нагоя, Аити 466-8550, Япония; Международный центр клеточной и генной терапии, Университет здоровья Фудзита, Тойоаке, Аити 470-1192, Япония.
- PMID: 32535020
- PMCID: PMC7300155
- DOI:
10. 1016/j.isci.2020.101213
Абстрактный
Несмотря на общепринятое мнение о важности белков планарной клеточной полярности (PCP) в тканевой ориентации, мало что известно о механизмах, используемых белками PCP для обеспечения планарной поляризации цитоскелета внутри отдельных клеток. Один белок РСР Fzd6 асимметрично локализуется на апикальной клеточной мембране многореснитчатых эпендимальных клеток, выстилающих стенку бокового желудочка (ЛЖ) на стороне, контактирующей с током спинномозговой жидкости. Индивидуальные эпендимальные клетки имеют плоские поляризованные микротрубочки, которые соединяют базальные тела ресничек (BBs) с клеточным кортексом стороны Fzd, чтобы координировать ориентацию ресничек. Здесь мы сообщаем, что цитоплазматический dynein прикрепляется к клеточному кортексу со стороны Fzd через адапторный белок Daple, который регулирует динамику микротрубочек. Асимметричная локализация коркового динеина генерирует тянущую силу на динамические микротрубочки, связанные с BB, что, в свою очередь, ориентирует BB в сторону Fzd. Это необходимо для координированной ориентации ресничек на стенке ЛЖ.
Ключевые слова: Биологические науки; клеточная биология; функциональные аспекты клеточной биологии; Специализированные функции клеток.
Copyright © 2020 Авторы. Опубликовано Elsevier Inc. Все права защищены.
Заявление о конфликте интересов
Заявление об интересах Авторы не заявляют о конкурирующих интересах.
Цифры
Графический реферат
Графический реферат
Рисунок 1
Произошли динамические изменения микротрубочек…
Рисунок 1
Динамические изменения микротрубочек произошли на стороне Fzd клеточной коры…
Фигура 1 Динамические изменения микротрубочек произошли на стороне Fzd клеточной коры для модуляции позиционирования BB в эпендимальных клетках (A) Иллюстрация направленного потока спинномозговой жидкости в желудочках головного мозга. ЦСЖ (красная стрелка) вырабатывается в сосудистом сплетении (красная область) в задней области бокового желудочка (ЛЖ) и оттекает через своды Монро в передне-вентральной области ЛЖ по направлению к третьему желудочку (3V). и четвертый желудочек (4V). На рисунке ниже представлена поверхность рассеченной правой дистальной стенки ЛЖ. Для всех экспериментов показаны микроскопические изображения выделенной области. А, передний; П, задний; Д, спинной; V, вентральные стороны мозга. (B) Конфокальные изображения стенки ЛЖ в горизонтальной плоскости показывают апикальную клеточную кору эпендимальных клеток. Цельные препараты ткани стенки ЛЖ окрашивали антителами против Fzd6 (левая панель, зеленый), Vangl2 (правая панель, зеленый), γ-тубулина (базальные тельца, красные на левой панели или синие на правой панели) и β- катенин (граница клетки, красный). Стрелки указывают на асимметричную локализацию Fzd6 на передне-вентральной стороне (левая панель) или Vangl2 на задне-дорсальной стороне (правая панель) апикальной клеточной мембраны эпендимальных клеток. (C) Представление эпендимальных клеток, выстилающих стенку ЛЖ, ориентация множественных ресничек (желтый) и асимметричная локализация Fzd6 (красный) и Vangl2 (синий) на апикальной клеточной мембране, параллельно поверхностному току спинномозговой жидкости (красная стрелка). BF (базальная ножка, зеленый) и BB (базальное тело, светло-зеленый), расположенные у основания ресничек, ориентированы ротационно в сторону Fzd. (D) Окрашивание стенки левого желудочка антителами против Fzd6 (зеленый) и тирозинированного (Tyr)-тубулина (красный). Стрелки указывают места накопления Fzd6 с тирозинированным тубулином, расположенным на вентральной стороне апикальной клеточной мембраны. (E) Изображение эпендимальной клетки на стенке ЛЖ, полученное с помощью электронной микроскопии, показывает филаменты (стрелки), связанные со структурой BF/BB. (F) Окрашивание стенки ЛЖ целиком антителами против EB3, которые маркируют плюс-конец микротрубочки (зеленый), γ-тубулин (красный) и β-катенин (синий). (G) Окрашивание стенки ЛЖ целиком антителами против CAMSAP2, которые маркируют минус-конец микротрубочки (зеленый), антитела к γ-тубулину (красный) и β-катенину (синий). (H) Тканевые эксплантаты из стенки ЛЖ инкубировали с ДМСО или нокодазолом (10 мкМ) в течение 24 часов и окрашивали антителами против тирозинированного тубулина (белый), γ-тубулина (зеленый) и β-катенина (красный). (I) Иллюстрация (слева) показывает измерение расстояния между клеточной корой и центром кластера BB ( d ) или клеточной поверхности ( D ). Отношение d/D представляли как относительное расстояние между кластером BB и клеточной корой. Количественная оценка (справа) представлена как среднее значение ± SEM для 54 клеток от трех мышей в каждой группе лечения (репрезентативные данные микроскопии показаны на [H]). Данные представлены для d/D с обработкой ДМСО и нокодазолом.
Рисунок 2
Daple локализует на Fzd/Dvl…
Рисунок 2
Daple локализуется на стороне Fzd/Dvl коры клеток и регулирует микротрубочки…
Фигура 2 Daple локализуется на стороне Fzd/Dvl клеточного кортекса и регулирует динамику микротрубочек (A) Окрашивание стенки левого желудочка антителами против Dvl1 (зеленый) и Daple (красный). Увеличенное изображение указанной области окна показано ниже. (B) Окрашивание стенки ЛЖ целиком антителами против Fzd6 (зеленый), Dvl1 (красный), Daple (белый/синий) и тирозинированного тубулина (белый/пурпурный). На каждой панели показана апикальная поверхность эпендимальных клеток. Стрелки указывают места совместной локализации Fzd6, Dvl1 и Daple с тирозинированным тубулином в клеточном кортексе. (C) Изображения конфокальной микроскопии апикальной поверхности Daple +/+ или Daple −/− Эпендимальные клетки на стенке ЛЖ, окрашенные антителами против Fzd6 (зеленый), тирозинированным тубулином (красный) и γ-тубулином или β-катенином (белый). Стрелки указывают на накопление Fzd6 или тирозинированного тубулина в клеточном кортексе. Штриховые линии обозначают скопления BB. (D) Накопление тирозинированного тубулина на стороне Fzd6 клеточной коры было количественно определено для Daple +/+ и Daple −/− эпендимальные клетки. Среднюю интенсивность тирозинированного тубулина в области Fzd6 клеточной коры нормализовали по отношению к средней интенсивности тирозинированного тубулина для всей площади клеточной поверхности. Нормализованные интенсивности нанесены как средние значения ± стандартная ошибка среднего для 30 клеток эпендимы, взятых у трех мышей в каждой группе. (E) Кимограммы одиночных микротрубочек из Daple +/+ или Daple -/- Эпендимальные клетки. Кончики EMTB-EGFP на краю клетки контролировали с помощью покадровой флуоресцентной микроскопии, см. также видео S1 и S2. (F) Количественная оценка состояния динамики микротрубочек в Daple +/+ или Daple -/- эпендимальных клеток. Сокращение, рост и пауза (пауза определяется как > 0,1 мкм/2 с) состояния 30 микротрубочек измерялись в течение 2,55 мин для каждой группы. Значения представлены в таблице S1.
Рисунок 3
Цитоплазматический динеин был прикреплен к…
Рисунок 3
Цитоплазматический динеин был прикреплен к клеточной коре с помощью Daple в эпендимальных клетках…
Рисунок 3 Цитоплазматический динеин был прикреплен к клеточному кортексу с помощью Daple в эпендимальных клетках (A) Окрашивание всего препарата стенки ЛЖ антителами против промежуточной цепи динеина (IC, зеленый), тирозинированного тубулина (красный) и γ-тубулина или β- катенин (синий). Панели ниже представляют собой большое увеличение окна, показанного на правой панели. Стрелки указывают места накопления динеина IC с тирозинированным тубулином в корковом слое клеток. (B) Лизаты ткани стенки ЛЖ подвергали иммунопреципитации с помощью IgG или антител против Daple. Общий клеточный лизат (входной) и иммунопреципитаты блоттировали антителами против динеина IC, анти-динеина HC, анти-Dvl1 или анти-Daple, как указано слева на каждой панели. Звездочками отмечены полосы ко-иммунопреципитированных белков. (C) Тканевые лизаты из стенки ЛЖ были иммунопреципитированы с помощью IgG или антитела против динеина IC и подвергнуты иммуноблотингу с соответствующими антителами, как показано слева. Звездочками отмечены полосы белков, ко-иммунопреципитировавшихся вместе. (D) Цельное крепление Daple +/+ и Daple −/− ткань, окрашенная антителами против динеина IC (зеленый), Daple (красный) и γ-тубулина (синий) на стенке ЛЖ. Стрелки указывают места совместной локализации динеина IC с Daple в коре клеток. См. также рисунок S1 и таблицу S2.
Рисунок 4
Расположение и ориентация…
Рисунок 4
Позиционирование и ориентация BB были снижены за счет ингибирования цитоплазматического динеина…
Рисунок 4 Положение и ориентация BB были уменьшены за счет ингибирования цитоплазматического динеина (A) Тканевые эксплантаты из стенки ЛЖ обрабатывали в течение 24 ч ДМСО или динаррестином (20 мкМ), ингибитором цитоплазматического динеина. Тканевые эксплантаты окрашивали антителами против γ-тубулина (зеленый), β-катенина (красный) и тирозинированного тубулина (белый). (B) Количественная оценка отношения относительного расстояния (d/D) между кластером BB и клеточной корой. Данные представлены как среднее значение ± SEM для 58 клеток от трех мышей в каждой группе из данных микроскопии, представленных в (A). (C) Тканевые эксплантаты стенки ЛЖ обрабатывали в течение 12 ч ДМСО или динаррестином (20 мкМ). Показаны репрезентативные изображения эксплантатов LV, окрашенных антителами против FOP (зеленый) и против γ-тубулина (красный). (D) Углы от FOP до точек γ-тубулина (n = 20), показанные на (C), были нанесены на круговую диаграмму в каждой ячейке. Красный вектор — это средний результирующий вектор данных отдельных углов. R¯ — длина среднего результирующего вектора; csd, круговое стандартное отклонение. (E) Циркулярная дисперсия (V=1-R¯) в пределах отдельных эпендимальных клеток была рассчитана для представления распределения углов BB ( n = 17–22 BB в каждой из 43 клеток эпендимы от трех мышей на группу лечения). Данные на диаграммах представлены в виде медиан, 25-75-го процентилей, диапазонов и выбросов. (F) Предлагаемая модель молекулярного позиционирования BB. Dvl связывается с Fzd на клеточной мембране и захватывает адапторный белок Daple через свой PBM, чтобы закрепить цитоплазматический динеин на клеточном кортексе. Дапле/динеин в клеточном кортексе может способствовать динамическому циклированию микротрубочек и создает тянущую силу на микротрубочках, связанных с BB.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Плоская полярность клеток индуцирует локальное связывание микротрубочек для координированного биения ресничек.
Накаяма С., Яно Т., Намба Т., Кониси С., Такагиши М., Херавати Э., Нисида Т., Имото Ю., Исихара С., Такахаши М., Фурута К., Оива К., Тамура А., Цукита С. Накаяма С. и др. Джей Селл Биол. 2021 5 июля; 220(7):e202010034. doi: 10.1083/jcb.202010034. Джей Селл Биол. 2021. PMID: 33
- 5 Бесплатная статья ЧВК.
Daple координирует динамику плоскополяризованных микротрубочек в эпендимальных клетках и способствует развитию гидроцефалии.
Такагиши М., Савада М., Охата С., Асаи Н., Эномото А., Такахаши К., Венг Л., Усида К., Ара Х., Мацуи С., Кайбути К., Савамото К., Такахаши М. Такагиши М. и соавт. Cell Rep. 25 июля 2017 г .; 20 (4): 960-972. doi: 10.1016/j.celrep.2017.06.089. Представитель ячейки 2017. PMID: 28746879
p73 регулирует полярность эпендимальных плоских клеток путем модуляции актина и цитоскелета микротрубочек.
Фуэртес-Альварес С., Маесо-Алонсо Л., Виллох-Фернандес Дж., Вильдунг М., Мартин-Лопес М., Маршалл С., Вильена-Кортес А.Дж., Диес-Прието И., Питенпол Дж.А., Тиссир Ф., Лизе М., Маркес М.М., Марин МС. Фуэртес-Альварес С. и соавт. Клеточная смерть Дис. 2018 5 декабря; 9 (12): 1183. doi: 10.1038/s41419-018-1205-6. Клеточная смерть Дис. 2018. PMID: 30518789Бесплатная статья ЧВК.
Плоскостная полярность эпендимальных ресничек.
Кисимото Н., Савамото К. Кисимото Н. и др. Дифференциация. 2012 Февраль;83(2):S86-90. doi: 10.1016/j.diff.2011.10.007. Epub 2011 17 ноября. Дифференциация. 2012. PMID: 22101065 Обзор.
Расположение центриолей в эпителиальных клетках и его тесная связь с плоскостной полярностью клеток.
Карвахаль-Гонсалес Х.М., Мулеро-Наварро С., Млодзик М. Карвахаль-Гонсалес Дж. М. и соавт. Биоэссе. 2016 дек;38(12):1234-1245. doi: 10.1002/bies.201600154. Epub 2016 24 октября. Биоэссе. 2016. PMID: 27774671 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Цитируется
DAPLE организует сборку апикального актомиозина из комплексов соединительной полярности.
Маривин А., Хо Р.С., Гарсия-Маркос М. Маривин А. и др. Джей Селл Биол. 2022 2 мая; 221(5):e202111002. doi: 10.1083/jcb.202111002. Epub 2022 7 апр. Джей Селл Биол. 2022. PMID: 35389423
Анализ транскриптома мозга мышиной модели CLN2 в зависимости от прогрессирования заболевания.
Домович М.С., Чан В.К., Клаудио-Васкес П., Гонсалес Т., Шварц Н.Б. Домович М.С. и соавт. J Нейровоспаление. 2021 8 ноября; 18 (1): 262. дои: 10.1186/с12974-021-02302-з. J Нейровоспаление. 2021. PMID: 34749772 Бесплатная статья ЧВК.
Плоская полярность клеток индуцирует локальное связывание микротрубочек для координированного биения ресничек.
Накаяма С., Яно Т., Намба Т., Кониси С., Такагиши М.
, Херавати Э., Нисида Т., Имото Ю., Исихара С., Такахаши М., Фурута К., Оива К., Тамура А., Цукита С.
Накаяма С. и др.
Джей Селл Биол. 2021 5 июля; 220(7):e202010034. doi: 10.1083/jcb.202010034.
Джей Селл Биол. 2021.
PMID: 33- 5 Бесплатная статья ЧВК.
использованная литература
- Аснар Н., Мидде К.К., Дункель Ю., Лопес-Санчес И., Павлова Ю., Маривин А., Барбазан Дж., Мюррей Ф., Ницше У., Янссен К.П. Daple представляет собой новый нерецепторный GEF, необходимый для активации тримерного G-белка в передаче сигналов Wnt. Элиф. 2015;4:e07091. — ЧВК — пабмед
- Бутин С. , Лабедан П., Димидшштейн Дж., Ричард Ф., Кремер Х., Андре П., Ян Ю., Монкукиоль М., Гоффине А.М., Тиссир Ф. Двойная роль генов плоскостной клеточной полярности в реснитчатых клетках. проц. Натл. акад. науч. США. 2014;111:E3129–E3138.
—
ЧВК
—
пабмед
- Бутин С.
- Батлер М.Т., Уоллингфорд Дж.Б. Плоская клеточная полярность в развитии и болезни. Нац. Преподобный Мол. Клеточная биол. 2017;18:375–388. — ЧВК — пабмед
- Карвахаль-Гонсалес Дж. М., Мулеро-Наварро С., Млодзик М. Позиционирование центриолей в эпителиальных клетках и его тесная связь с плоскостной клеточной полярностью. Биоэссе. 2016; 38:1234–1245.
—
ЧВК
—
пабмед
- Карвахаль-Гонсалес Дж.
- Чиен Ю.Х., Келлер Р., Кинтнер С., Шук Д.Р. Механическая деформация определяет ось планарной полярности мерцательного эпителия. Курс. биол. 2015;25:2774–2784. — ЧВК — пабмед
Особенности строения и передвижения туфельки инфузории.
Биология в лицее. Симптомы и осложненияК инфузориям относят около 6000 видов простейших, органоидами которых является большое количество ресничек. Для большинства инфузорий характерно наличие двух ядер: крупного вегетативного — макронуклеуса — и более мелкого генеративного — микронуклеуса. Макронуклеус имеет полиплоидный набор хромосом и регулирует метаболические процессы. Микронуклеус содержит диплоидный набор хромосом и участвует в половом процессе.
Свободноживущие инфузории включают инфузорию-туфельку . Размеры клеток 0,1-0,3 мм. Самый простой имеет постоянную форму, так как эктоплазма уплотняется и образует пленку. Тело инфузории покрыто ресничками. Их насчитывается от 10 до 15 тысяч. В эктоплазме у инфузорий имеются защитные образования — трихоцисты. При раздражении трихоцисты выстреливают наружу, превращаясь в длинные нити, парализующие пострадавшего. После использования одних трихоцист на их месте в эктоплазме развиваются новые.
Пищевые органеллы включают ротовое отверстие, расположенное на вентральной стороне и ведущее в клеточный рот, переходящий в клеточный глоток. Вода с бактериями через рот попадает в эндоплазму, где образуются пищеварительные вакуоли. Вакуоли передвигаются по телу инфузории.
Непереваренные остатки пищи, оставшиеся внутри вакуоли, выводятся наружу через порох — отверстие, расположенное вблизи заднего конца тела инфузории.
Инфузория туфелька имеет две сократительные вакуоли, расположенные в передней и задней части тела. Каждая вакуоль состоит из округлого резервуара и 5-7 трубочек, подходящих к нему в виде звезды. Жидкие продукты и вода из цитоплазмы сначала поступают в приводящие канальцы, затем канальцы все сразу сокращаются и изливают свое содержимое в резервуар, после чего последний сокращается и выбрасывает жидкость через отверстие наружу, а канальцы в это время пополнение. Вакуоли сокращаются одна за другой.
Бесполое размножение инфузорий осуществляется поперечным делением и сопровождается делением макро- и микроядер. Размножение повторяют 1 — 2 раза в день. Через несколько поколений в жизненном цикле инфузорий происходит половой процесс, который называется конъюгацией. Две инфузории сближаются вентральными сторонами, раковина растворяется в месте соприкосновения, и между ними образуется цитоплазматический мостик. При этом макронуклеусы разрушаются, а микроядра делятся мейозом на четыре ядра, три из которых разрушаются, а четвертое снова делится митозом пополам.
В результате у каждой инфузории образуются мужские (мигрирующие) и женские (неподвижные) ядра. Затем между особями происходит обмен мигрирующими ядрами с последующим слиянием неподвижных и мигрирующих ядер, после чего особи рассредоточиваются. Вскоре в каждом из них происходит деление ядра и в последующем образуются микро- и макронуклеусы. Таким образом, во время полового процесса численность инфузорий не увеличивается, а обновляются наследственные свойства макронуклеуса и возникают новые сочетания генетической информации.
Диагноз ставится при обнаружении балантидиев в фекалиях. Профилактика такая же, как и при других кишечных заболеваниях.
Строение
Наиболее типичным распространенным представителем цилиарных является инфузория туфелька (Paramecium). Обитает в стоячей воде, а также в пресноводных водоемах с очень слабым течением, содержащих разлагающийся органический материал.
Сложность строения клетки у парамеций объясняется тем, что ей приходится выполнять все функции, присущие всему организму, а именно питание, осморегуляцию и движение. Тело парамеция имеет характерную форму: его передний конец тупой, а задний несколько заострен.
Реснички инфузории туфельки расположены попарно по всей поверхности клетки. Расположенные продольными диагональными рядами, они, совершая биения, заставляют инфузорию вращаться и двигаться вперед. Между ресничками имеются отверстия, ведущие в специальные камеры, называемые трихоцистами. Из этих камер под влиянием определенных раздражителей могут выбрасываться тонкие заостренные нити, служащие, вероятно, для удерживания добычи.
Под пленкой инфузории туфельки располагается эктоплазма — прозрачный слой плотной цитоплазмы гелеобразной консистенции. В эктоплазме имеются базальные тельца (идентичные центриолям), от которых отходят реснички, а между базальными тельцами имеется сеть тонких фибрилл, участвующих, по-видимому, в координации биения ресничек.
Объемная цитоплазма инфузории туфельки представлена эндоплазмой, имеющей более жидкую консистенцию, чем эктоплазма. Именно в эндоплазме находится большинство органелл. На вентральной (нижней) поверхности туфельки, ближе к ее переднему концу, имеется периоральная воронка, на дне которой находится рот, или цитостом.
Ротовая часть инфузории туфельки ведет в короткий канал — цитофаринкс, или глотку. Как околоротовая воронка, так и глотка могут быть выстланы ресничками, движения которых направляют струю воды в сторону цитостома, увлекающую за собой различные пищевые частицы, например бактерии. Пищевая вакуоль образуется вокруг пищевых частиц, попавших в цитоплазму путем эндоцитоза. Эти вакуоли движутся по эндоплазме к так называемому порошку, через который путем экзоцитоза выводятся наружу непереваренные остатки.
В цитоплазме инфузории туфельки имеются также две сократительные вакуоли, расположение которых в клетке строго фиксировано. Эти вакуоли отвечают за осморегуляцию, то есть поддерживают определенный водный потенциал в клетке. Жизнь в пресной воде осложняется тем, что вода постоянно поступает в клетку в результате осмоса; эту воду необходимо постоянно выталкивать из клетки, чтобы она не лопнула.
Это происходит с помощью процесса активного транспорта, требующего затрат энергии. Примерно каждые сократительная вакуоль инфузории туфельки имеется ряд излучающих каналов, которые собирают воду, прежде чем выпустить ее в центральную вакуоль.
В клетке инфузории парамеции туфельки находятся два стержня. Более крупный из них — макронуклеус — полиплоидный; он имеет более двух наборов хромосом и контролирует метаболические процессы, не связанные с размножением. Микронуклеус представляет собой диплоидное ядро. Он контролирует размножение и образование макронуклеусов во время деления ядра.
Туфельки Paramecium infusoria Может размножаться как бесполым путем (путем поперечного деления надвое), так и половым путем (путем конъюгации).
Движение
Совершая волнообразные движения ресничками, туфелька движется (плавает тупым концом вперед). Ресница движется в одной плоскости и совершает прямой (эффективный) удар в расправленном состоянии, а ответный удар — в изогнутом. Каждая следующая ресничка в ряду прокрашивается с небольшой задержкой по сравнению с предыдущей. Плавая в толще воды, башмак вращается вокруг продольной оси. Скорость движения около 2 мм/с. Направление движения может меняться за счет изгиба тела. При столкновении с препятствием направление прямого удара меняется на противоположное, и ботинок отскакивает назад. Затем некоторое время «раскачивается» туда-сюда, а затем снова начинает двигаться вперед. При столкновении с препятствием клеточная мембрана деполяризуется, и внутрь клетки поступают ионы кальция. В фазе «качания» кальций выкачивается из клетки
Дыхание, выделение, осморегуляция
Обувь дышит всей поверхностью клетки. Он способен существовать за счет гликолиза при низкой концентрации кислорода в воде. Продукты азотистого обмена также выводятся через клеточную поверхность и частично через сократительную вакуоль. Основная функция сократительных вакуолей – осморегуляторная. Они выводят из клетки лишнюю воду, проникающую туда за счет осмоса. Сначала набухают ведущие каналы, затем вода из них перекачивается в водохранилище. При сжатии резервуара он отделяется от подводящих каналов, и вода выбрасывается через поры. Две вакуоли работают в противофазе, каждая в нормальных физиологических условиях сокращается один раз в 10-15 с. За час вакуоли выбрасывают из клетки объем воды, примерно равный объему клетки.
размножение
Туфелька имеет бесполое и половое размножение (половой процесс). Бесполое размножение — поперечное деление в активном состоянии. Сопровождается сложными регенерационными процессами. Например, одна из особей переформирует клеточный рот периоральными ресничками, каждая достраивает недостающую сократительную вакуоль, базальные тельца размножаются и образуются новые реснички и т. д.
Половой процесс, как и у других инфузорий, происходит у форма сопряжения. Туфельки, принадлежащие к разным клонам, временно «склеиваются» ротовой стороной, и между клетками образуется цитоплазматический мостик. Затем макронуклеусы конъюгирующих инфузорий разрушаются, а микроядра делятся мейозом. Из четырех образовавшихся гаплоидных ядер три погибают, а оставшееся одно делится митозом. Каждая инфузория теперь имеет два гаплоидных пронуклеуса — один женский (неподвижный), а другой мужской (мигрирующий). Инфузории обмениваются мужскими пронуклеусами, а самки остаются в «своей» клетке. Затем у каждой инфузории «свой» женский и «чужой» мужской пронуклеусы сливаются, образуя диплоидное ядро — синкарион. При делении синкариона образуются два ядра. Один из них становится диплоидным микронуклеусом, а второй превращается в полиплоидный макронуклеус. В действительности этот процесс более сложен и сопровождается особыми постконъюгационными делениями.
Сувойка
Род простейших из подкласса реснитчатых инфузорий (Peritricha). Включает более 100 широко распространенных видов, обитающих в морской и пресной воде. С. — сидячие животные, прикрепляющиеся к субстрату (в отличие от других родов Peritricha) с помощью неветвящегося сократительного стебля. Тело С., по форме напоминающее колокольчик, лишено ресничек. На ее расширенном переднем конце (адоральная зона) имеется двойной ряд ресниц (обычно сливающихся в меморанеллы), закрученных влево (в отличие от спиральных ресничных инфузорий, у которых адоральная зона перепончатых ресничок закручена вправо). Периоральная спираль ведет к ротовому отверстию. С. питаются взвешенными в воде мелкими органическими частицами (например, бактериями, детритом). При бесполом размножении в результате деления образуются свободно плавающие «бродяги», снабженные венчиком ресничек, которые затем образуют стебель и прикрепляются к субстрату. Половой процесс — по типу анизогамной конъюгации (крупные неподвижные макроконъюганты и малые подвижные микроконъюганты).
Инфузория трубач
Род простейших класса инфузорий подотряда ресничных. Длина до 1 мм . Более 10 видов. Тело воронкообразной формы. На расширенном переднем конце имеется мощно развитая зона периоральных перепонок, направляющих поток пищевых частиц к ротовому отверстию. Остальное тело покрыто продольными рядами мелких ресничек. Они способны резко сокращаться, приобретая шаровидную форму за счет сократительных нитей — мионем. Они могут свободно плавать или прикрепляться к субстрату зауженным задним концом. Они обитают в морях и пресных водах. У некоторых видов в цитоплазме имеются многочисленные симбионты — одноклеточные водоросли.
Инфузория-стилохония
Есть такая инфузория — стилонихия. Под действием летучих луковых фитонцидов он распадается на мельчайшие крупинки и даже растворяется. Микробиологи называют это явление лизисом. Тело инфузории «исчезает». То же самое происходит и с инфузориями, называемыми «loxodes rostrum». В течение 10-15 секунд все ее тело растворяется в окружающей жидкой среде!
Точно в таких же условиях другая инфузория — spirostomum teres — под действием тех же фитонцидов распадается на зерна, но растворения всего тела не происходит. Мы называем эту смерть зернистым распадом.
Тип Инфузории также принято называть Реснитчатыми — органами движения этих простейших являются реснички . Ресничная клетка имеет два ядра, их называют малым и большим. Первый регулирует процесс размножения, а второй отвечает за процессы питания, движения и дыхания.
Особенности жизнедеятельности этого вида следует рассмотреть на примере инфузории-туфельки.
Движение и дыхание
Инфузория-туфелька, длина которой составляет примерно 0,5 мм, местом обитания выбирает водоемы. О форме тела простейшего легко догадаться по названию – оно напоминает туфельку. Скорость движения составляет примерно 2,5 мм в секунду.
Наличие внешней эластичной оболочки обеспечивает устойчивую форму тела.
В цитоплазме, прилежащей к оболочке, имеются опорные волокна, их развитие является гарантией сохранения постоянной формы инфузории.
На поверхности инфузории 15 тысяч ресничек, у их основания находится базальное тельце. Движение происходит с помощью колебаний ресничек: они производят около 30 взмахов в секунду, выталкивая тем самым инфузорию-туфельку вперед.
Дышит через поверхность тела.
Питание
Особенностью инфузории является наличие ротовой полости , возле которой расположены особенно длинные и густые реснички. Клеточный рот продолжается ячеистым зевом: реснички проталкивают в него воду и реснитчатую пищу — бактерии.
Инфузория чувствует химические вещества, которые выделяют скопления бактерий. Таким образом, она ищет добычу.
Пища затем попадает в пищеварительную вакуоль, где она переваривается. Отсюда он вытекает уже в цитоплазму.
Отбор
Отбор осуществляется с помощью двух сократительных вакуолей , одна расположена на переднем конце, а другая — на заднем. Вакуоли состоят из резервуара и каналов.
Жидкость заполняет каналы, затем следует по центральному резервуару, а затем выходит из инфузорий. Процесс сокращения вакуолей занимает 10-20 секунд.
размножение
Инфузория размножается бесполым путем – делится надвое. Его особенностью является деление поперек тела.
Ядра инфузорий делятся на две части: новообразованные инфузории имеют малое и большое ядра. Дочерние инфузории обладают частями органоидов, а недостающие образуются самостоятельно. Размножение происходит несколько раз в сутки.
У инфузорий-туфельок также возможно половое размножение, но в этом случае увеличения численности особей не происходит. Временно простейшие соединяются, образуя соединительный мостик из цитоплазмы.
У каждой особи большое ядро исчезает, а малые делятся дважды — появляются четыре ядра. Из них остается только одно ядро, которое также делится. Особь содержит два ядра, затем происходит обмен ядрами — одно из ядер переходит к другой особи.
Там он сливается с оставшимся ядром, и таким образом у каждой из особей образуется маленькое и большее ядро. Этот процесс, называемый конъюгацией , необходим для обновления генетического материала между особями.
Виды инфузорий
Инфузории — сложноорганизованные простейшие, их насчитывается около 7000 видов.
Инфузория-туфелька — простейший одноклеточный микроорганизм, получивший свое название за сходство с подошвой обуви. Его размеры колеблются от 10 мкм до 4,5 мм , но такие крупные экземпляры встречаются крайне редко. В основном встречается в пресных и стоячих водах.
Этот микроорганизм невозможно увидеть невооруженным глазом. Однако при большом скоплении в загрязненной и мутной воде можно увидеть продолговатые точки белого цвета – это инфузории-туфельки. Они находятся в постоянном движении.
Инфузория туфелька — это бактерия или нет?
Бактерия – одноклеточный организм, характеризующийся отсутствием ядра, а инфузория-туфелька имеет два ядра. Отсюда можно сделать вывод, что данный представитель фауны не является бактерией .
Где обитает инфузория-туфелька?
Как было сказано выше, инфузория-туфелька обитает в пресноводных водоемах . Изучив воду из домашнего аквариума под микроскопом, можно заметить большое количество микроорганизмов, в том числе и инфузорий.
Вы можете самостоятельно создать искусственный водоем, в котором будет жить этот простейший одноклеточный организм, для этого достаточно будет залить водой обычное сено и дать настояться несколько дней.
Строение и функции
Внешний покров этого представителя фауны представляет собой тонкую эластичную оболочку, называемую мембраной . Сохраняет форму тела на протяжении всего жизненного цикла. Это связано с наличием в цитоплазматическом слое развитых опорных волокон. Эти волокна расположены близко к оболочке. Инфузория-туфелька имеет два ядра . Большое ядро отвечает за пищеварение, а малое — за размножение.
На всей поверхности инфузории-туфельки расположены органы, отвечающие за ее передвижение. Эти органы называются ресничек , а их число превышает 15000 . Их движения напоминают движения весла. Движение инфузории осуществляется тупым концом вперед со скоростью до 3 мм/с . Во время движения этот микроорганизм вращается вокруг продольной оси своего тела. Это связано с медленными волнообразными движениями ресничек.
Инфузория-туфелька – высокоорганизованный простой организм, осуществляющий множество процессов для поддержания своей жизнедеятельности.
Дыхание организма осуществляется за счет поступления кислорода в цитоплазму через мембрану. Благодаря двум сократительным вакуолям происходит газообмен, который осуществляется за счет специальных канальцев. Выведение лишней жидкости, являющейся результатом жизненного процесса, происходит каждые 30 секунд. В неблагоприятной среде работа сократительных вакуолей замедляется, и инфузория-туфелька перестает питаться.
Размножение этого высокоорганизованного микроорганизма может быть как половым, так и бесполым.
Бесполое размножение У инфузории-туфельки нормальный процесс деления клеток. Примерно раз в сутки большое и малое ядра расходятся в разные стороны тела и делятся на два. В результате деления образуются две инфузории-туфельки с таким же набором органов, как и в родительском организме.
Половое размножение Характерно только для тех инфузорий, которые неоднократно подвергались бесполому размножению или в неблагоприятных условиях. В результате такого размножения две особи не образуются. Два микроорганизма соединяются, создавая между собой соединительный мост.
Крупные ядра инфузорий бесследно исчезают, а мелкие делятся на два. Ученые дали этому процессу название – конъюгация. Он может длиться более одиннадцати часов. При понижении температуры воды или смене освещения две инфузории-туфельки могут превратиться в цисту и просуществовать около десяти лет в состоянии анабиоза, хотя в среднем при благоприятных условиях продолжительность их жизни длится не более суток.
Что он ест?
Рацион инфузории-туфельки состоит из бактерий и микроводорослей, которые в большом количестве встречаются в мутной стоячей воде. Питание происходит с помощью клеточного рта, вокруг которого расположены реснички. С их помощью микроорганизм может легко захватывать в рот как можно больше пищи. Изо рта пища проходит через клетчаточную глотку, попадая в вакуоли, в которых происходит процесс пищеварения. Он может возникать сразу в нескольких вакуолях и длиться более часа.
Инфузория туфелька может питаться постоянно, особенно при температуре воды более 17 градусов, прерываясь только для размножения.
Опасно ли это для человека?
Заключение
Строение и внешний вид инфузории-туфельки одинаковы у каждой особи. Они могут различаться по размеру. Жизненный цикл при благоприятных условиях у них также одинаков. Эти микроорганизмы остро реагируют на температуру воды, освещение и содержание солей в водоеме. При неблагоприятных условиях они впадают в анабиоз, и продолжительность их жизни увеличивается в сотни и даже тысячи раз.
По сравнению с другими группами простейших инфузории имеют наиболее сложное строение, что связано с разнообразием и сложностью их функций.
Откуда произошло название «инфузория туфелька»? Вы не удивитесь, если посмотрите под микроскопом на живую инфузорию или даже на ее изображение (рис. 85).
Действительно, форма тела этой инфузории напоминает изящный женский башмачок.
Туфелька инфузория находится в непрерывном достаточно быстром движении. Его скорость (при комнатной температуре) составляет около 2,0-2,5 мм/сек. Это большая скорость для такого маленького животного! Ведь это означает, что за секунду ботинок пробегает расстояние, превышающее длину его тела в 10-15 раз. Траектория движения обуви довольно сложная. Она двигает переднюю часть прямо вперед
ИНФУЗОРИЙ ТУФЕЛЬНЫЙ (PARAMECIUM CAUDATUM)
Чтобы познакомиться со строением и образом жизни этих интересных одноклеточных организмов, обратимся сначала к одному характерному примеру. Возьмем инфузории-туфельки (виды рода Paramecium), широко распространенные в мелководных пресноводных водоемах. Этих инфузорий очень легко разводить в небольших аквариумах, если наполнить пруд обычным луговым сеном. В таких настойках развивается множество различных видов простейших, почти всегда развиваются инфузории-туфельки. С помощью обычного учебного микроскопа можно увидеть многое из того, о чем пойдет речь дальше.
Среди простейших инфузорий туфельки являются достаточно крупными организмами. Длина их тела составляет около 1/6-1/3 мм. и вращается вправо вдоль продольной оси корпуса.
Такое активное движение туфельки зависит от работы большого количества тончайших волосовидных придатков — ресничек, покрывающих все тело инфузории. Количество ресничек у одной особи инфузории туфельки составляет 10-15 тысяч!
Каждая ресничка совершает очень частые веслообразные движения — при комнатной температуре до 30 ударов в секунду. Во время удара ресничка удерживается в выпрямленном положении. При возвращении в исходное положение (при движении вниз) движется в 3-5 раз медленнее и описывает полукруг.
При плавании туфельки суммируются движения многочисленных ресничек, покрывающих ее тело. Действия отдельных ресничек координируются, в результате чего возникают правильные волнообразные колебания всех ресничек. Волна колебаний начинается на переднем конце тела и распространяется назад. При этом по телу туфельки проходят 2-3 волны сокращения. Таким образом, весь ресничный аппарат инфузории представляет собой как бы единое функциональное физиологическое целое, действия отдельных структурных единиц которого (реснички) тесно связаны (скоординированы) друг с другом.
Строение каждой отдельной реснички туфельки, как показали электронно-микроскопические исследования, очень сложное.
Направление и скорость движения обуви не являются величинами постоянными и неизменными. Туфелька, как и все живые организмы (это мы уже видели на примере амебы), реагирует на изменение внешней среды изменением направления движения.
Изменение направления движения простейших под влиянием различных раздражителей называется таксисом. У инфузорий легко наблюдать различные таксисты. Если в каплю, где плавают туфли, поместить какое-либо неблагоприятно воздействующее на них вещество (например, кристаллик поваренной соли), то туфли уплывают (как бы убегают) от этого неблагоприятного для них фактора (рис. 86). ).
Перед нами пример отрицательного таксиса на химическом воздействии (отрицательный хемотаксис). Можно наблюдать туфельку и положительный хемотаксис. Если, например, каплю воды, в которой плавают инфузории, накрыть покровным стеклом и подпустить под нее пузырек углекислого газа (СО2), то большая часть инфузорий пойдет к этому пузырю и поселится вокруг него в звенеть.
Феномен такси очень ярко проявляется в обуви под действием электрического тока. Если через жидкость, в которой плавают туфли, пропустить слабый электрический ток, то можно наблюдать следующую картину: все инфузории ориентируют свою продольную ось параллельно линии тока, а затем, как по команде, двигаются к катоду, в площадь, на которой они образуют плотное скопление. Движение инфузорий, определяемое направлением электрического тока, называется гальванотаксисом. Различные таксисы у инфузорий могут быть обнаружены под влиянием самых разнообразных факторов внешней среды.
Все цитоплазматическое тело инфузории четко разделено на 2 слоя: наружный более светлый (эктоплазма) и внутренний более темный и зернистый (эндоплазма). важную роль в поддержании постоянства формы тела инфузории.
В наружном слое (в эктоплазме) тела живой туфельки хорошо видны многочисленные короткие палочки, расположенные перпендикулярно поверхности (рис. 85, 7). Эти образования называются трихоцистами. Их функция очень интересна и связана с защитой простейших. При механическом, химическом или каком-либо другом сильном раздражении трихоцисты с силой выбрасываются наружу, превращаясь в тонкие длинные нити, которые поражают атакующего туфельку хищника. Трихоцисты являются мощной защитой. Они равномерно расположены между ресничками, так что число трихоцист примерно соответствует числу ресничек. На месте использованных («расстрелянных») трихоцист в эктоплазме туфельки развиваются новые.
С одной стороны, примерно посередине туловища (рис. 85, 5), туфелька имеет довольно глубокое углубление. Это ротовая полость, или перистом. Реснички располагаются вдоль стенок перистома, а также по поверхности тела. Они здесь развиты гораздо мощнее, чем на остальной поверхности тела. Эти близко расположенные реснички расположены в две группы. Функция этих высокодифференцированных ресничек связана не с движением, а с питанием (рис. 87).
Как и чем питается обувь, как она переваривается?
Туфельки относятся к числу инфузорий, основной пищей которых являются бактерии. Наряду с бактериями они могут заглатывать и любые другие частицы, взвешенные в воде, вне зависимости от их пищевой ценности. Периоральные реснички создают непрерывный поток воды с взвешенными частицами в направлении ротового отверстия, расположенного в глубине перистома. Мелкие пищевые частицы (чаще всего бактерии) проникают через рот в небольшую трубчатую глотку и скапливаются на дне ее, на границе с эндоплазмой. Ротовое отверстие всегда открыто. Пожалуй, не будет ошибкой сказать, что инфузория-туфелька — одно из самых прожорливых животных: она непрерывно питается. Этот процесс прерывается только в определенные моменты жизни, связанные с размножением и половым процессом.
Скопившийся на дне глотки пищевой комок затем отрывается от дна глотки и вместе с небольшим количеством жидкости попадает в эндоплазму, образуя пищеварительную вакуоль. Последний не остается на месте своего образования, а, попадая в токи эндоплазмы, проделывает в теле туфельки довольно сложный и закономерный путь, называемый циклозом пищеварительной вакуоли (рис. 88). Во время этого довольно длительного (при комнатной температуре, занимающего около часа) путешествия пищеварительной вакуоли внутри нее происходит ряд изменений, связанных с перевариванием в ней пищи.
Здесь, как и у амеб и некоторых жгутиковых, происходит типичное внутриклеточное пищеварение. Из эндоплазмы, окружающей пищеварительную вакуоль, в нее поступают пищеварительные ферменты, действующие на пищевые частицы. Продукты пищеварения всасываются через пищеварительные вакуоли в эндоплазму.
В ходе циклоза пищеварительной вакуоли в ней происходит смена нескольких фаз пищеварения. В первые моменты после образования вакуоли заполняющая ее жидкость мало отличается от жидкости окружающей среды. Вскоре в вакуоль из эндоплазмы начинают поступать пищеварительные ферменты, и реакция среды внутри нее становится резко кислой. Это легко обнаружить, добавляя в пищу какой-либо индикатор, цвет которого меняется в зависимости от реакции (кислой, нейтральной или щелочной) среды. В этой кислой среде проходят первые фазы пищеварения. Затем картина меняется и реакция внутри пищеварительных вакуолей становится слабощелочной. В этих условиях протекают дальнейшие этапы внутриклеточного пищеварения. Кислая фаза обычно короче щелочной; она длится примерно 1/6-1/4 всего пребывания пищеварительной вакуоли в теле инфузории. Однако соотношение кислой и щелочной фаз может изменяться в довольно широких пределах в зависимости от характера пищи.
Путь пищеварительной вакуоли в эндоплазме заканчивается тем, что она подходит к поверхности тела и через пелликулу ее содержимое, состоящее из жидкости и непереваренных остатков пищи, выбрасывается наружу — происходит дефекация. Этот процесс, в отличие от амеб, у которых дефекация может происходить где угодно, у туфельки, как и у других инфузорий, строго приурочен к определенному участку тела, расположенному на брюшной стороне (брюшной условно называют поверхность животного на котором находится периоральное углубление), примерно посередине между перистомом и задним концом тела.
Таким образом, внутриклеточное пищеварение представляет собой сложный процесс, состоящий из нескольких фаз, последовательно сменяющих друг друга.
Расчеты показывают, что примерно через 30-45 минут из туфельки через сократительные вакуоли выделяется объем жидкости, равный объему реснитчатого тела. Таким образом, благодаря деятельности сократительных вакуолей через тело инфузории осуществляется непрерывный ток воды, поступающей извне через ротовое отверстие (вместе с пищеварительными вакуолями), а также осмотически непосредственно через пелликулу. Сократительные вакуоли играют важную роль в регуляции тока воды, проходящей через тело инфузории, и в регуляции осмотического давления. Этот процесс протекает здесь в принципе так же, как и у амеб, только строение сократительной вакуоли значительно сложнее.
В течение многих лет среди ученых, изучающих простейшие, шел спор по вопросу о том, существуют ли какие-либо структуры в цитоплазме, связанные с появлением сократительной вакуоли, или же она образуется каждый раз заново. На живой инфузории не наблюдается никаких особых структур, которые предшествовали бы ее образованию. После того, как происходит сокращение вакуоли — систола, в цитоплазме на месте бывшей вакуоли совершенно не видно никаких структур. Затем вновь появляются прозрачный пузырь или приводящие каналы, которые начинают увеличиваться в размерах. Однако никакой связи между вновь возникшей вакуолью и существовавшей ранее вакуолью не обнаружено. По-видимому, между последовательными циклами сократительной вакуоли нет преемственности и любая новая сократительная вакуоль образуется заново в цитоплазме. Однако специальные методы исследования показали, что на самом деле это не так. Применение электронной микроскопии, дающей очень большое увеличение (до 100 тыс. раз), убедительно показало, что инфузория имеет особо дифференцированную цитоплазму в области образования сократительных вакуолей, состоящую из переплетения тончайших трубочек. Таким образом, оказалось, что сократительная вакуоль возникает в цитоплазме не на пустом месте, а на основе предшествующего особого клеточного органоида, функцией которого является образование сократительной вакуоли.
Как и все простейшие, инфузории имеют клеточное ядро. Однако по строению ядерного аппарата инфузории резко отличаются от всех других групп простейших.
Ядерный аппарат инфузорий характеризуется двойственностью. Это означает, что у инфузорий есть два разных типа ядер — большие ядра, или макронуклеусы, и малые ядра, или микроядра. Посмотрим, какое строение имеет ядерный аппарат у инфузории туфельки (рис. 85).
В центре тела инфузории (на уровне перистома) расположено крупное массивное ядро яйцевидной или бобовидной формы. Это макронуклеус. В непосредственной близости от него находится второе ядро, во много раз меньшее, обычно довольно тесно примыкающее к макронуклеусу. Это микронуклеус. Разница между этими двумя ядрами не только в размерах, она более существенна, глубоко затрагивая их структуру.
Макронуклеус, по сравнению с микронуклеусом, значительно богаче особым ядерным веществом (хроматином, или, точнее, дезоксирибонуклеиновой кислотой, сокращенно ДНК), входящим в состав хромосом.
Недавние исследования показали, что макронуклеус имеет в несколько десятков (а у некоторых инфузорий и сотни) раз больше хромосом, чем микроядра. Макронуклеус представляет собой весьма своеобразный тип мультихромосомных (полиплоидных) ядер. Таким образом, различие между микро- и макронуклеусами сказывается на их хромосомном составе, определяющем большее или меньшее богатство их ядерного вещества — хроматина.
У одного из наиболее распространенных видов инфузорий — туфельки (Paramecium caudatum) — имеется один макронуклеус (сокращенно Ма) и один микронуклеус (сокращенно Ми). Такое строение ядерного аппарата характерно для многих инфузорий. У других может быть несколько Ма и Ми. Но характерной особенностью всех инфузорий является дифференциация ядер на две качественно различные группы, на Ма и Ми, или, другими словами, явление ядерного дуализма.
Как размножаются инфузории? Обратимся в качестве примера снова к инфузории туфельке. Если посадить единственный экземпляр туфельки в небольшой сосуд (микроаквариум), то через сутки там будет две, а нередко и четыре инфузории. Как это произошло? После определенного периода активного плавания и питания инфузория несколько вытягивается в длину. Затем ровно посередине тела появляется все более углубляющаяся поперечная перетяжка (рис. 9).0). В конце концов инфузории как бы сплетаются пополам, и из одной особи получается две особи, изначально несколько мельче родительской особи. Весь процесс деления занимает около часа при комнатной температуре. Изучение внутренних процессов показывает, что еще до появления поперечной перетяжки начинается процесс деления ядерного аппарата. Сначала делится Ми, и только после этого Ма. Мы не будем здесь останавливаться на подробном рассмотрении процессов деления ядер и лишь отметим, что Ми делится митозом, тогда как деление Ма по внешнему виду напоминает прямое деление ядер — амитоз. Этот процесс бесполого размножения у инфузорий-туфельок, как мы видим, сходен с бесполым размножением амеб и жгутиконосцев. Напротив, инфузории в процессе бесполого размножения всегда делятся поперек, а у жгутиковых плоскость деления параллельна продольной оси тела.
При делении происходит глубокая внутренняя перестройка тела инфузории. Образуются два новых перистома, два зева и два ротовых отверстия. К этому же времени приурочено деление базальных ядер ресничек, благодаря которому образуются новые реснички. Если бы число ресничек не увеличивалось при размножении, то в результате каждого деления дочерние особи получали бы примерно половину числа ресничек материнской особи, что приводило бы к полному «облысению» инфузории. На самом деле этого не происходит.
Время от времени у большинства инфузорий, в том числе и у туфель, возникает особая и крайне своеобразная форма полового процесса, которая называется конъюгацией. Мы не будем здесь подробно разбирать все сложные ядерные изменения, сопровождающие этот процесс, а отметим лишь самые важные. Конъюгация протекает следующим образом (рис. 91). Две инфузории сближаются, тесно прилегают друг к другу брюшными сторонами и в таком виде плавают вместе довольно продолжительное время (у туфельки около 12 часов при комнатной температуре). Затем конъюгаты разделяются. Что происходит в организме инфузорий при конъюгации? Суть этих процессов заключается в следующем (рис. 91). Большое ядро (макронуклеус) спадается и постепенно растворяется в цитоплазме. Микроядра сначала делятся, часть образовавшихся в результате деления ядер разрушается (см. рис. 91). Каждая из конъюгантов сохраняет два ядра. Одно из этих ядер остается на месте у особи, у которой оно образовалось (неподвижное ядро), а другое активно переходит к партнеру по конъюгации (мигрирующее ядро) и сливается с его неподвижным ядром. Таким образом, в каждой из конъюгантов на этой стадии имеется по одному ядру, образовавшемуся в результате слияния стационарного и мигрирующего ядер. Это сложное ядро называется синкарионом. Образование синкариона есть не что иное, как процесс оплодотворения. А у многоклеточных существенным моментом оплодотворения является слияние ядер половых клеток. У инфузорий половые клетки не образуются, имеются только половые ядра, которые сливаются между собой. Таким образом, происходит взаимное перекрестное оплодотворение.
Вскоре после образования синкариона конъюганты расходятся. По строению своего ядерного аппарата на этой стадии они еще очень существенно отличаются от обычных так называемых нейтральных (неконъюгирующих) инфузорий, так как имеют только по одному ядру. В дальнейшем за счет синкариона восстанавливается нормальный ядерный аппарат. Синкарион делится (один или несколько раз). Часть продуктов этого деления путем сложных превращений, связанных с увеличением числа хромосом и обогащением хроматином, превращается в макронуклеусы. Другие сохраняют структуру, характерную для микроядер. Таким путем восстанавливается характерный и типичный для инфузорий ядерный аппарат, после чего инфузории приступают к бесполому размножению делением.
Таким образом, процесс конъюгации включает в себя два существенных биологических момента: оплодотворение и восстановление нового макронуклеуса за счет синкариона.
Какое биологическое значение имеет конъюгация, какую роль она играет в жизни инфузорий? Мы не можем назвать это размножением, потому что нет увеличения количества особей. Вышеуказанные вопросы послужили материалом для многочисленных экспериментальных исследований, проведенных во многих странах. Основной результат этих исследований состоит в следующем. Во-первых, конъюгация, как и всякий другой половой процесс, при котором два наследственных начала (отцовский и материнский) соединяются в одном организме, приводит к увеличению наследственной изменчивости, наследственного разнообразия. Увеличение наследственной изменчивости увеличивает приспособительные возможности организма к условиям внешней среды. Второй биологически важной стороной конъюгации является развитие нового макронуклеуса за счет продуктов деления синкариона и одновременно разрушение старого. Экспериментальные данные показывают, что именно макронуклеус играет чрезвычайно важную роль в жизни инфузорий. Он контролирует все основные жизненные процессы и определяет важнейший из них — образование (синтез) белка, составляющего основную часть протоплазмы живой клетки. При длительном бесполом размножении делением происходит своеобразный процесс «старения» макронуклеуса, а вместе с тем и всей клетки: снижается активность метаболического процесса, снижается скорость деления. После конъюгации (при которой, как мы видели, старый макронуклеус разрушается) скорость метаболизма и скорость деления восстанавливаются. Поскольку процесс оплодотворения происходит при конъюгации, которая у большинства других организмов связана с размножением и появлением нового поколения, то у инфузорий образовавшуюся после конъюгации особь также можно рассматривать как новое половое поколение, которое появляется здесь как бы вследствие к «омоложению» старого.
На примере инфузории туфельки мы познакомились с типичным представителем обширного класса инфузорий. Однако для этого класса характерно чрезвычайное разнообразие видов как по строению, так и по образу жизни. Рассмотрим подробнее некоторые из наиболее характерных и интересных форм.
У инфузорий реснички туфельки равномерно покрывают всю поверхность тела. Это характерная черта строения (голотриха). Для многих инфузорий характерен различный характер развития ресничного покрова. Дело в том, что реснички инфузорий способны, соединяясь вместе, образовывать более сложные комплексы. Например, часто наблюдается, что реснички, расположенные в один или два ряда близко друг к другу, соединяются (слипаются) между собой, образуя пластинку, которая, как и реснички, способна биться. Такие пластинчатые сократительные образования называются мембранеллами (если они короткие) или мембранами (если они длинные). В других случаях реснички соединены между собой, располагаются в плотный пучок. Эти образования — цирры — напоминают кисточку, отдельные волоски которой слиплись. Для многих инфузорий характерны различные сложные ресничные образования. Очень часто ресничный покров развивается неравномерно, а лишь на отдельных участках тела.
ИНФУЗОРИЯ трубач (STENTOR POLYMORPHIC)
В пресных водах виды крупных красивых инфузорий, относящихся к роду трубачей (Stentor). Это название вполне соответствует форме тела этих животных, которое действительно напоминает трубу (рис. 92), широко открытую с одного конца. При первом знакомстве с живыми трубачами можно заметить одну особенность, не свойственную туфельке. При малейшем раздражении, в том числе и механическом (например, постукивание карандашом по стеклу, где у трубачей капля воды), их тело резко и очень быстро (за доли секунды) сокращается, принимая почти правильную форму. сферическая форма. Затем довольно медленно (время измеряется секундами) трубач выпрямляется, принимая характерную для него форму. Эта способность трубача быстро сокращаться обусловлена наличием особых мышечных волокон, расположенных вдоль тела и в эктоплазме. Таким образом, мышечная система может развиваться и в одноклеточном организме.
В роду трубачей есть виды, некоторые из которых отличаются довольно яркой окраской. Очень часто встречается в пресных водах голубой трубач (Stentor coeruleus), который имеет ярко-голубой цвет. Такая окраска трубача обусловлена тем, что в его эктоплазме расположены мельчайшие зерна синего пигмента.
Другой вид трубача (Stentor polymorphus) часто окрашен в зеленый цвет. Причина такого окрашивания совсем в другом. Зеленый цвет обусловлен тем, что в эндоплазме инфузории живут и размножаются мелкие одноклеточные зеленые водоросли, придающие телу трубача характерную окраску. Stentor polymorphus — типичный пример взаимовыгодного сожительства — симбиоза. Трубач и водоросли находятся во взаимно-симбиотических отношениях: трубач защищает живущие в его теле водоросли и снабжает их углекислым газом, образующимся в результате дыхания; со своей стороны, водоросли обеспечивают трубача кислородом, выделяющимся в процессе фотосинтеза. По-видимому, часть водорослей переваривается инфузориями, являясь пищей для трубача.
Трубачи медленно плавают в воде широким концом вперед. Но они также могут временно прикрепляться к субстрату задним узким концом тела, на котором образуется небольшая присоска.
В теле трубача можно различить туловищный отдел, расширяющийся сзади кпереди, и широкое периоральное (перистомальное) поле, расположенное почти перпендикулярно ему. Это поле напоминает асимметричную плоскую воронку, на одном краю которой имеется углубление — зев, ведущий в эндоплазму инфузории. Тело трубача покрыто продольными рядами коротких ресничек. По краю перистомального поля по окружности располагается мощно развитая околоротовая (адоральная) зона мембранелл (рис. 9).2). Эта зона состоит из большого количества отдельных реснитчатых пластинок, каждая из которых, в свою очередь, состоит из множества слипшихся друг с другом ресничек, расположенных в два близко расположенных ряда.
В области ротового отверстия периоральные оболочки завернуты в сторону глотки, образуя левозакрученную спираль. Поток воды, вызванный колебаниями околоротовой перепонки, направлен к ротовому отверстию (в глубину воронки, образованной передним концом тела). Вместе с водой в глотку попадают и взвешенные в воде частицы пищи. Пищевые объекты трубача более разнообразны, чем у башмачка. Наряду с бактериями поедает мелких простейших (например, жгутиковых), одноклеточных водорослей и др.
У трубача хорошо развита сократительная вакуоль, имеющая своеобразное строение. Центральный резервуар расположен в передней трети тела, немного ниже ротового отверстия. От него отходят два длинных приводящих канала. Один из них идет от резервуара к заднему концу тела, второй располагается в области перистомального поля параллельно периоральной зоне перепонки.
Инфузория трубач — излюбленный объект экспериментальных исследований регенерации. Многочисленные эксперименты доказали высокую регенеративную способность трубачей. Инфузорию тонким скальпелем можно разрезать на множество частей, и каждая из них через короткое время (несколько часов, иногда сутки и более) превратится в пропорционально сложенного, но небольшого трубача, который затем, в результате энергичного питания, достигает типичных для этого вида размеров. Для завершения восстановительных процессов регенерирующий фрагмент должен содержать хотя бы один сегмент бусообразного макронуклеуса.
У трубача, как мы видели, реснички разные: с одной стороны, они короткие, покрывающие все тело, а с другой — околоротовая зона мембранеллы. В соответствии с этой особенностью строения отряд инфузорий, к которому относится трубач, получил название инфузорий (Heterotricha).
ИНФУЗОРИЯ БУРСАРИЯ (BURSARIA TRUNCATELLA)
Второй интересный представитель ресничных инфузорий часто встречается в пресных водах бурсария (Bursaria truncatella, рис. 93). Это гигант среди инфузорий: его размеры могут достигать 2 мм, наиболее распространены размеры 0,5-1,0 мм. Бурсария хорошо видна невооруженным глазом. В соответствии со своим названием бурсария имеет форму мешка, широко открытого на переднем конце (bursa — латинское слово, в переводе на русский язык означает «кошелек», «мешок») и несколько расширенного на заднем конце. Все тело инфузории покрыто продольными рядами коротких ресничек. Их биение вызывает довольно медленное движение животного вперед. Бурсария плавает как бы «переваливаясь» из стороны в сторону.
От переднего конца в глубь тела (примерно на 2/3 его длины) выдается околоротовое углубление — перистом. На вентральной стороне сообщается с внешней средой через узкую щель; с дорсальной стороны полость перистома не сообщается с внешней средой. Если посмотреть на поперечное сечение верхней трети тела бурсарии (рис. 93, Б), то можно увидеть, что перистомальная полость занимает большую часть тела, а цитоплазма окружает ее в виде ободка.
На переднем конце тела слева от бурсарий берет начало очень мощно развитая зона периоральной (адоральной) мембранеллы (рис. 93, 4). Она спускается в глубь перистомальной полости, поворачивая влево. Адоральная зона простирается до самой глубокой части перистома. Других ресничных образований в перистомальной полости, кроме периоральных перепонок, нет, кроме ресничной полоски, проходящей вдоль вентральной стороны перистомальной полости (рис. 93, 10). На внутренней задней стенке перистомальной полости почти на всем ее протяжении имеется узкая щель (рис. 9).3, 7), края которых обычно плотно прилегают друг к другу. Это открывание рта. Его края раздвигаются только в момент еды.
Бурсарии не имеют узкой пищевой специализации, но в основном являются хищниками. Продвигаясь вперед, они сталкиваются с различными мелкими животными. Благодаря работе мембран околоротовой зоны добыча с силой втягивается в обширную перистомальную полость, откуда уже не может выплыть. Пищевые объекты прижимаются к дорсальной стенке перистомальной полости и проникают в эндоплазму через расширяющуюся ротовую щель. Бурсарии очень прожорливы, они могут проглатывать довольно крупные предметы: например, их излюбленная пища — инфузории-туфельки. Бурсария способна проглотить 6-7 ботинок подряд. В результате в эндоплазме бурсарии образуются очень крупные пищеварительные вакуоли.
Ядерный аппарат бурсарии довольно сложен. Они имеют один длинный колбасовидный макронуклеус и большое (примерно до 30) количество мелких микроядер, беспорядочно разбросанных в эндоплазме инфузории.
Bursaria — один из немногих видов пресноводных инфузорий, у которых отсутствует сократительная вакуоль. Как осуществляется осморегуляция у этой крупной инфузории, до сих пор не совсем ясно. Под эктоплазмой бурсариев на разных участках тела можно наблюдать различные по форме и размерам пузырьки жидкости — вакуоли, изменяющие свой объем. По-видимому, эти вакуоли неправильной формы по своей функции соответствуют сократительным вакуолям других инфузорий.
Интересно наблюдать за последовательными стадиями бесполого размножения бурсарии. На начальных стадиях деления происходит полная редукция всей полости перистома и околоротовой зоны мембранеллы (рис. 94). Сохраняется только наружная ресничка. Инфузория имеет форму яйца. После этого корпус шнуруется поперечным пазом на две половинки. У каждой из образовавшихся дочерних инфузорий путем довольно сложных преобразований развиваются типичный перистом и периоральная зона мембранелл. Весь процесс деления бурсарии протекает быстро и занимает чуть больше часа.
Очень легко наблюдать еще один важный жизненный процесс у бурсариев, начало которого связано с неблагоприятными для инфузорий условиями, — процесс цистообразования (энцистации). Это явление характерно, например, для амебы. Но оказывается, даже такие сложноорганизованные простейшие, как инфузории, способны переходить в неактивное состояние. Если культуру, в которой живут бурсарии, вовремя не подкормить или не охладить, то через несколько часов начнется массовое инцистирование. Этот процесс протекает следующим образом. Бурсариды, как и до деления, теряют перистом и периоральную зону мембранелл. Затем они становятся полностью шаровидными, после чего выделяют двойную оболочку характерной формы (рис. 94, Д).
В состоянии кисты бурсарии могут находиться месяцами. При наступлении благоприятных условий оболочка цисты лопается, из нее выходят бурсарии, развиваются перистом и переходят к активной жизни.
STYLONICHIA MYTILUS
Весьма сложный и разнообразно дифференцированный ресничный аппарат имеют инфузории, относящиеся к отряду желудочно-кишечных (Hypotricha), многочисленные виды которых обитают как в пресной, так и в морской воде. Одними из самых распространенных, часто встречающихся представителей этой интересной группы можно назвать стилонихия (Stylonichia mytilus). Это довольно крупная инфузория (длиной до 0,3 мм), обитающая на дне пресноводных водоемов, на водной растительности (рис. 95). В отличие от башмачка, трубача и бурсарии у стилонихий отсутствует сплошной ресничный покров, и весь ресничный аппарат представлен ограниченным числом строго определенных ресничных образований.
Тело Stilonychia (как и большинства других вентральных инфузорий) сильно уплощено в дорсо-абдоминальном направлении, и его дорсальная и вентральная стороны, передний и задний концы хорошо различимы. Тело несколько расширено спереди, сужено сзади. При осмотре животного с брюшной стороны хорошо видно, что в передней трети слева находится сложно расположенное перисто-ротовое отверстие.
На спинной стороне довольно редко расположены реснички, которые не способны биться. Их скорее можно назвать тонкими эластичными щетинками. Они неподвижны и не имеют никакого отношения к функции движения. Этим ресничкам обычно отводят осязательную, чувствительную функцию.
Все ресничные образования, связанные с движением и захватом пищи, сосредоточены на брюшной стороне животного (рис. 95). Имеется небольшое количество толстых пальцевидных образований, расположенных несколькими группами. Это брюшные цирры. Каждая из них представляет собой сложное ресничное образование, результат тесного соединения (слипания) многих десятков отдельных ресничек. Таким образом, цирры представляют собой как бы кисти, отдельные волоски которых тесно сближены и соединены друг с другом.
С помощью цирроз животное передвигается довольно быстро, «бегает» по субстрату. Помимо «ползания» и «бега» по субстрату, стилонихии способны производить довольно резкие и сильные прыжки, сразу же отрываясь от субстрата. Эти резкие движения осуществляются с помощью двух мощных хвостовых усиков (рис. 95), не принимающих участия в обычном «ползании».
Два ряда краевых (маргинальных) цир расположены по краю тела справа и слева. От правого края животного они идут вдоль всего тела, а от левого достигают только области перистома. Эти ресничные образования служат для движения животного, когда оно отрывается от субстрата и свободно плавает в воде.
Мы видим, таким образом, что разнообразный и специализированный ресничный аппарат стилонихии позволяет ей совершать весьма разнообразные движения, в отличие, например, от простого скольжения в воде, как туфелька или трубач.
Ресничный аппарат, связанный с функцией питания, также сложен. Мы уже видели, что околоротовое углубление (перистом), на дне которого находится ротовое отверстие, ведущее в глотку, расположено в передней половине животного слева. По левому краю, начиная с самого переднего конца тела, проходит сильно развитая зона периоральных (адоральных) перепонок. Своим биением они направляют поток воды в сторону ротового отверстия. Кроме того, в области перистомального углубления имеются еще три сократительные перепонки (мембраны), уходящие своими внутренними концами в глотку, и ряд особых периоральных ресничек (рис. 9).5). Весь этот сложный аппарат служит для захвата и направления пищи в ротовое отверстие.
Stilonychia относится к простейшим с очень широким набором пищевых объектов. Его по праву можно назвать всеядным. Она может есть, как ботинок, бактерии. Среди его пищевых объектов жгутиконосцы, одноклеточные водоросли (часто диатомовые). Наконец, стилонихии могут быть и хищниками, нападающими на другие, более мелкие виды инфузорий и пожирающими их.
Stilonychia имеет сократительную вакуоль. Он состоит из центрального резервуара, расположенного в левом заднем конце перистома, и одного приводящего канала, направленного назад.
Ядерный аппарат, как всегда у инфузорий, состоит из макронуклеуса и микронуклеуса.
Макронуклеус состоит из двух половинок, соединенных тонкой перетяжкой; микроядра два, они расположены непосредственно возле обеих половинок Ма.
Стилонихия, отчасти бурсария, трубач — это все инфузории с широким набором пищевых объектов. Для большинства инфузорий характерна способность поглощать различную пищу. Однако среди них можно найти и такие виды, которые строго специализированы по отношению к характеру питания.
ИНФУЗОРИЙНЫЕ ХИЩНИКИ
Среди инфузорий есть хищники, которые очень «придирчивы» к своей добыче. Яркий пример — инфузория. дидиния (Didinium nasutum). Дидиния — относительно небольшая инфузория, длина которой составляет в среднем около 0,1–0,15 мм. Передний конец вытянут в виде хоботка, на конце которого размещено ротовое отверстие. Ресничный аппарат представлен двумя венчиками ресничек (рис. 96). Дидиниус быстро плавает в воде, часто меняя направление. Предпочтительной пищей дидинии являются инфузории туфельки. В этом случае хищник меньше своей жертвы. Дидиний проникает в добычу хоботом, а затем, постепенно все больше расширяя ротовое отверстие, заглатывает туфельку целиком! В хоботке имеется особый, так называемый стержневой, аппарат. Он состоит из ряда эластичных прочных палочек, расположенных в цитоплазме по периферии хоботка. Считается, что этот аппарат повышает прочность стенок хоботка, который не лопается при заглатывании такой огромной по сравнению с дидинием добычи, как башмак. Didinius является примером крайнего случая хищничества среди простейших. Если сравнить заглатывание дидинием своей добычи — туфельки — с хищничеством у высших животных, то подобных примеров найти трудно.
Дидиниус, проглотив парамецию, конечно, сильно опухает. Процесс переваривания очень быстрый, при комнатной температуре он занимает всего около двух часов. Затем непереваренные остатки выбрасываются и дидиния начинает охотиться за очередной жертвой. Специальные исследования установили, что дневной «рацион» дидинии составляет 12 туфель — поистине колоссальный аппетит! Нужно иметь в виду, что в промежутках между очередными «охотами» дидинии иногда делятся. При недостатке питания дидинии очень легко инцистируются и так же легко вновь выходят из цист.
растительноядные инфузории
Значительно реже, чем хищничество, среди инфузорий встречается «чистое вегетарианство» — питание исключительно растительной пищей. Одними из немногих примеров «вегетарианских» инфузорий могут быть представители рода passula (Nassula). Объектом их питания являются нитчатые сине-зеленые водоросли (рис. 97).
Они проникают в эндоплазму через рот, расположенный сбоку, а затем закручиваются инфузориями в тугую спираль, которая постепенно переваривается. Пигменты водорослей частично проникают в цитоплазму инфузории и окрашивают ее в яркий темно-зеленый цвет.
СУВОЙКА (VORTICELLA NEBULIFERA)
Интересная и довольно многочисленная по количеству видов группа инфузорий представляет собой прикрепленные к субстрату сидячие формы, образующие отряд круглоглазых (Peritricha). Наиболее распространены представители этой группы сувойки (виды рода Vorticella).
Сувойки напоминают изящный цветок, похожий на колокольчик или ландыш, сидящий на длинном стебле, который своим концом прикреплен к субстрату. Большую часть жизни сувойка проводит в прикрепленном к субстрату состоянии.
Рассмотрим строение тела инфузории. У разных видов их размеры варьируют в довольно широких пределах (примерно до 150 мкм). Ротовой диск (рис. 98) расположен на расширенной передней части тела, полностью лишенной ресничек. Ресничный аппарат располагается только по краю ротового (перистомального) диска (рис. 98) в специальной бороздке, снаружи которой образуется валик (перистомальная губа). По краю валика проходят три реснитчатые перепонки, две из которых расположены вертикально, одна (наружная) – горизонтально. Они образуют несколько больше одного полного витка спирали. Эти перепонки находятся в постоянном мерцающем движении, направляя поток воды к ротовому отверстию. Ротовой аппарат начинается довольно глубоко воронкой у края перистомального поля (рис. 9).8), в глубине которого находится ротовое отверстие, ведущее в короткую глотку. Сувойки, как и туфельки, питаются бактериями. Их ротовое отверстие постоянно открыто, и ко рту идет непрерывный поток воды.
Около ротового отверстия расположена одна сократительная вакуоль без приводящих каналов. Макронуклеус имеет лентовидную или колбасовидную форму, вплотную к нему прилегает одиночный мелкий микронуклеус.
Сувойка умеет резко укорачивать стебель, который за доли секунды выкручивается штопором. При этом тело инфузории также сокращается: перистомальный диск и мембраны втягиваются внутрь и весь передний конец смыкается.
Естественно возникает вопрос: поскольку сувои прикрепляются к субстрату, то каким образом осуществляется их оседание в водоеме? Это происходит за счет образования свободно плавающей стадии, бродяги. На заднем конце тела инфузории появляется венчик ресничек (рис. 99). При этом перистомальный диск втягивается внутрь и инфузория отделяется от ножки. Получившийся бродяга способен плавать несколько часов. Затем события разыгрываются в обратном порядке: инфузория задним концом прикрепляется к субстрату, стебелек растет, задний венчик ресничек редуцируется, перистомальный диск расправляется на переднем конце, начинают работать адоральные мембраны. . Формирование бродяг у сувойки часто связано с процессом бесполого размножения. Инфузория на стебле делится, и одна из дочерних особей (а иногда и обе) становится бродягой и уплывает.
Многие виды сувоков способны к инцистированию в неблагоприятных условиях.
Среди сидячих инфузорий, принадлежащих к группе реснитчатых, лишь относительно немногие виды, как и рассмотренные выше сувои, являются одиночными живыми формами. Большинство видов, включенных сюда, являются колониальными организмами.
Обычно колониальность является результатом незавершенного бесполого или вегетативного размножения. Образовавшиеся в результате размножения особи в большей или меньшей степени сохраняют связь друг с другом и все вместе образуют органическую индивидуальность более высокого порядка, объединяющую большое количество отдельных особей, которая называется колонией (мы уже встречались с примерами колониальных организмов в класс жгутиконосцев .
Колонии круглореснитчатых инфузорий образуются в результате того, что отделившиеся особи не превращаются в бродяг, а сохраняют связь друг с другом при помощи стеблей (рис. 100). При этом главный ствол колонии, как и ее первые ветви, не может быть отнесен ни к одной из особей, а принадлежит всей колонии в целом. Иногда колония состоит лишь из небольшого числа особей, тогда как у других видов инфузорий количество отдельных особей колонии может достигать нескольких сотен. Однако рост любой колонии не безграничен. При достижении характерных для этого вида размеров колония перестает увеличиваться и образующиеся в результате деления особи развивают венчик из ресничек, становятся залетными и уплывают, давая начало новым колониям.
Колонии круглоресничных инфузорий бывают двух типов. У одних стебель колонии нередуцируемый: при раздражении сокращаются лишь отдельные особи колонии, втягивая перистое, но вся колония в целом не претерпевает изменений (к этому типу колонии относятся, например, роды Epistylis, Opercularia). У других (например, рода Carchesium) стебель всей колонии способен сокращаться, так как цитоплазма проходит через все ветви и таким образом соединяет между собой всех особей колонии. При раздражении такие колонии полностью сморщиваются. Вся колония в этом случае реагирует как единое целое, как органическая индивидуальность.
Среди всех колониальных круглоресничных инфузорий особый интерес представляет, пожалуй, зоотамник (Zoothamnium arbuscula). Колонии этой инфузории отличаются особой правильностью строения. Кроме того, здесь намечается интересное биологическое явление полиморфизма внутри колонии.
Колония зоотамнии выглядит как зонтик. На одном, главном, стебле колонии находятся второстепенные ветви (рис. 101). Размер взрослой колонии составляет 2-3 мм, поэтому они хорошо видны невооруженным глазом. Зоотамнии обитают в небольших водоемах с чистой водой. Их колонии обычно встречаются на подводных растениях, чаще всего на элодее (водяной чуме).
Стебли колонии зоотамний сократительные, так как сократительная цитоплазма проходит через все ветви колонии, за исключением базальной части основного стебля. При сокращении, происходящем очень быстро и резко, вся колония собирается в комок.
Зоотамния характеризуется строго правильным расположением ветвей. Один главный стебель прикреплен к субстрату. От ее верхней части в плоскости, перпендикулярной стеблю, отходят девять основных ветвей колонии, строго правильно расположенных друг относительно друга (рис. 102, 6). От этих ветвей отходят вторичные ветви, на которых сидят отдельные особи колонии. В каждой вторичной ветви может быть до 50 инфузорий. Общая численность особей в колонии достигает 2-3 тысяч особей.
Большинство особей колонии по своему строению напоминают мелких одиночных сувоиков, размером 40-60 мкм. Но кроме мелких особей, называемых микрозоидами, на взрослых колониях, примерно посередине основных ветвей, развиваются особи совсем другого типа и размера (рис. 102, 5). Это крупные шаровидные особи диаметром 200-250 мкм, по массе превышающие объем микрозоида в сто и более раз. Крупных особей называют макрозоидами.
По своему строению они существенно отличаются от мелких особей колонии. Их перистом не выражен: он втянут внутрь и не функционирует. С самого начала своего развития из микрозоида макрозоид перестает самостоятельно принимать пищу. В нем отсутствуют пищеварительные вакуоли. Рост макрозоида, по-видимому, осуществляется за счет веществ, поступающих через цитоплазматические мостики, соединяющие между собой всех особей колонии. В той части тела макрозоя, которой он прикрепляется к стеблю, происходит скопление особых зерен (гранул), которые, как мы увидим, играют значительную роль в его дальнейшей судьбе. Что это за крупные шаровидные макрозоиды, какова их биологическая роль в жизни колонии зоотамний? Наблюдения показывают, что макрозоиды — это будущие бродяги, из которых развиваются новые колонии. Достигнув предельных размеров, макрозоид развивает венчик из ресничек, отделяется от колонии и уплывает. При этом несколько меняется его форма, из сферической она становится конической. Через некоторое время бродяга всегда прикрепляется к подложке той стороной, на которой расположена зернистость. Сразу начинается формирование и рост стебля, причем на строительство стебля расходуются гранулы, локализующиеся на заднем конце бродяги. По мере роста стебля зернистость исчезает. После того как стебель достигает конечной длины, характерной для зоотамний, начинается ряд быстро следующих друг за другом делений, приводящих к образованию колонии. Эти деления производятся в строго определенной последовательности (рис. 102).
Мы не будем останавливаться на деталях этого процесса. Обратим внимание только на следующее интересное явление. При первых делениях зоотамний-бродяг, при развитии колонии у формирующихся особей перистое и ротовое не функционируют. Питание начинается позже, когда молодая колония уже состоит из 12-16 особей. Таким образом, все первые этапы развития семьи осуществляются исключительно за счет тех резервов, которые образовались в организме макрозоида в процессе его роста и развития на материнской семье. Существует неоспоримое сходство между развитием зоотамний бродячих и развитием яйцеклетки у многоклеточных животных. Это сходство выражается в том, что развитие в обоих случаях осуществляется за счет ранее накопленных резервов, без восприятия пищи из внешней среды.
При изучении сидячих круглореснитчатых инфузорий возникает вопрос: как осуществляется характерная для инфузорий форма полового процесса — конъюгация? Оказывается, в связи с малоподвижным образом жизни она претерпевает существенные изменения. К началу полового процесса на колонии образуются особые, очень мелкие бродяги. Активно передвигаясь с помощью венчика ресничек, они некоторое время ползают по колонии, а затем вступают в конъюгацию с крупными нормальными малоподвижными особями колонии. Таким образом, здесь происходит дифференциация конъюгантов на две группы особей: мелкие, подвижные (микроконъюганты) и более крупные, неподвижные (макроконъюганты). Эта дифференциация конъюгантов на две категории, одна из которых (микроконъюганты) подвижна, была необходимой адаптацией к малоподвижному образу жизни. Без этого нормальное протекание полового процесса (конъюгации) заведомо не могло быть обеспечено.
сосущие инфузории (SUCTORIA)
Весьма своеобразную группу по способу питания представляют сосущие инфузории (Suctoria). Эти организмы, как и сувойка и другие реснитчатые инфузории, сидячие. Число видов, принадлежащих к этому отряду, измеряется несколькими десятками. Форма тела сосущих инфузорий очень разнообразна. Некоторые их характерные виды показаны на рис. 103. Одни сидят на субстрате на более или менее длинных стеблях, другие не имеют стеблей, у некоторых тело достаточно сильно ветвится и т. д. Однако, несмотря на разнообразие форм, все сосущие инфузории характеризуются следующими двумя особенностями:
1) полное отсутствие (у взрослых форм) ресничного аппарата,
2) наличие особых придатков — щупалец, служащих для высасывания добычи.
У разных видов сосущих инфузорий количество щупалец неодинаково. Часто они собираются группами. При большом увеличении микроскопа видно, что на конце щупальца имеется небольшое булавовидное утолщение.
Как функционируют щупальца? На этот вопрос нетрудно ответить, понаблюдав некоторое время за сосущими инфузориями. Если какое-нибудь мелкое простейшее (жгутиконосец, инфузория) коснется щупальца суктории, то оно моментально к нему прилипнет. Все попытки жертвы оторваться обычно оказываются тщетными. Если продолжать наблюдать за прилипшей к щупальцам добычей, то можно заметить, что она постепенно начинает уменьшаться в размерах. Его содержимое постепенно «закачивается» через щупальца в эндоплазму сосущей инфузории до тех пор, пока от жертвы не останется только одна пелликула, которую отбрасывают. Таким образом, щупальца сосущих инфузорий совершенно уникальны, больше нигде в животном мире нет органов для улавливания и одновременно всасывания пищи (рис. 103).
Сосущие инфузории — неподвижные хищники, которые не преследуют добычу, а мгновенно ее ловят, стоит только неосторожной добыче прикоснуться к ним самой.
Почему мы относим эти своеобразные организмы к классу инфузорий? На первый взгляд, они не имеют к ним никакого отношения. О принадлежности суктории к инфузориям говорят следующие факты. Во-первых, у них типичный для инфузорий ядерный аппарат, состоящий из макронуклеуса и микронуклеуса. Во-вторых, при размножении у них развиваются реснички, отсутствующие у «взрослых» особей. Бесполое размножение и одновременно расселение сосущих инфузорий осуществляется путем образования бродяг, снабженных несколькими кольцевидными венчиками ресничек. Формирование залетников у суктории может происходить по-разному. Иногда они образуются в результате не совсем равномерного деления (почкования), при котором каждая отделяющаяся наружу почка получает сегмент макронуклеуса и один микронуклеус (рис. 104, Л). На одной материнской особи может образоваться сразу несколько дочерних почек (рис. 104, 5). У других видов (рис. 104, Г, Д) наблюдается весьма своеобразный способ «внутреннего почкования». При этом внутри тела материнского суктора образуется полость, в которой формируется трамповая почка. Он выходит через специальные отверстия, через которые «продавливается» с известным трудом.
Это развитие зародыша внутри тела матери, а затем акт деторождения — интересная аналогия простейшего с тем, что происходит в высших многоклеточных организмах.
На предыдущих страницах были рассмотрены несколько типичных свободноживущих представителей класса инфузорий, по-разному приспособленных к разным условиям среды. Интересно подойти к вопросу приспособления инфузорий к условиям жизни и, с другой стороны, посмотреть, каковы характерные общие черты инфузорий, обитающих в определенных, резко определенных условиях среды.
В качестве примера возьмем две очень резко разные среды обитания: жизнь в составе планктона и жизнь на дне в толще песка.
ПЛАНКТОННАЯ ИНФУЗОРИЯ
Достаточно большое количество видов инфузорий встречается как в морском, так и в пресноводном планктоне.
Особенности приспособления к жизни в толще воды особенно ярко выражены у радиолярий. Основная линия приспособления к планктонному образу жизни сводится к выработке таких особенностей строения, которые способствуют парению организма в толще воды.
Типичным планктонным, причем почти исключительно морским семейством инфузорий является тинтинниды (Tintinnidae, рис. 105, 5). Общее количество известных на сегодняшний день видов тинтиннид составляет около 300. Это мелкие формы, характеризующиеся тем, что протоплазматическое тело инфузории помещено в прозрачный, светлый и в то же время прочный дом, состоящий из органического вещества. Из домика выступает диск, несущий венчик ресничек, находящихся в постоянном мерцающем движении. В состоянии зависания инфузорий в толще воды в основном поддерживается постоянная активная работа ресничного аппарата. Домик, очевидно, выполняет функцию защиты нижней части тела инфузории. В пресной воде обитает всего 2 вида тинтиннид (не считая 7 видов, характерных только для озера Байкал).
У пресноводных инфузорий есть и другие приспособления к жизни в планктоне. У многих из них цитоплазма очень сильно вакуолизирована (Loxodes, Condylostoma, Trachelius), так что напоминает пену. Это приводит к значительному снижению удельного веса. Все перечисленные инфузории, кроме того, имеют ресничный покров, благодаря работе которого тело инфузории, по удельному весу лишь немного превышающему удельный вес воды, легко поддерживается в состоянии «плавания». . У некоторых видов форма тела способствует увеличению удельной поверхности и облегчает парение в воде. Например, некоторые планктонные инфузории оз. Байкал по форме напоминают зонтик или парашют (Liliomorpha, рис. 105, 2). В оз. Байкал встречается одна планктонная сосущая инфузория (Mucophrya pelagica, рис. 105, 4), резко отличающаяся от своих сидячих сородичей. Этот вид лишен стебля. Его протоплазматическое тело окружено широкой слизистой оболочкой, приспособление, ведущее к снижению веса. Торчат длинные тонкие щупальца, которые наряду со своей прямой функцией, вероятно, выполняют и другую — увеличение удельной поверхности, что способствует парению в воде.
Наконец, необходимо упомянуть еще об одной, так сказать, косвенной форме приспособления инфузорий к жизни в планктоне. Это прикрепление мелких инфузорий к другим организмам, ведущим планктонный образ жизни. Да, среди обрезанных инфузорий (Peritricha) имеется довольно много видов, прикрепляющихся к планктонным веслоногим. Это нормальный и нормальный образ жизни для этих видов инфузорий.
Наряду с циркумцилиарными инфузориями и среди сосущих (Suctoria) встречаются виды, поселяющиеся на планктонных организмах.
ГРАЖДАНЕ, ЖИВУЩИЕ В ПЕСКЕ
Песчаные пляжи и отмели представляют собой крайне своеобразную среду обитания. Вдоль побережья морей они занимают огромные пространства и характеризуются своеобразной фауной.
Многочисленные исследования, проведенные в последние годы в различных странах, показали, что толща многих морских песков очень богата разнообразной микроскопической или приближающейся по размерам микроскопической фауной. Между частицами песка имеется множество мелких и крошечных пространств, заполненных водой. Оказывается, эти пространства обильно заселены организмами, принадлежащими к самым разнообразным группам животного мира. Здесь обитают десятки видов ракообразных, кольчатых червей, круглых червей, особенно многочисленны плоские черви, некоторые моллюски, кишечнополостные. В большом количестве встречаются и простейшие, в основном инфузории. По современным данным, фауна инфузорий, населяющих толщу морских песков, насчитывает примерно 250-300 видов. Если иметь в виду не только инфузорий, но и другие группы организмов, населяющих толщу песка, то общее число их видов будет очень велико. Всю совокупность животных, населяющих толщу песка, живущих в мельчайших промежутках между песчинками, называют псаммофильной фауной.
Богатство и видовой состав псаммофильной фауны определяется многими факторами. Среди них особое значение имеет размер частиц песка. Крупнозернистые пески имеют бедную фауну. Бедна и фауна очень мелкозернистых пылеватых песков (с диаметром частиц менее 0,1 мм), где, очевидно, промежутки между частицами слишком малы для проживания в них животных. Наиболее богаты жизнью пески средне- и мелкозернистые.
Вторым фактором, играющим важную роль в развитии псаммофильной фауны, является насыщенность песка органическими остатками и разлагающимся органическим веществом (так называемая степень сапробности). Пески, лишенные органического вещества, бедны жизнью. С другой стороны, пески также почти безжизненны и очень богаты органикой, так как разложение органики приводит к истощению кислорода. Часто к этому добавляется анаэробное сероводородное брожение.
Наличие свободного сероводорода является крайне негативным фактором, влияющим на развитие фауны.
В поверхностных слоях песка иногда развивается довольно богатая флора одноклеточных водорослей (диатомовые, перидиниевые). Это фактор, благоприятствующий развитию псаммофильной фауны, поскольку многие мелкие животные (в том числе и инфузории) питаются водорослями.
Наконец, очень негативно влияющим на псаммофильную фауну фактором является прибой. Это вполне объяснимо, так как прибой, омывая верхние слои песка, убивает здесь все живое. Псаммофильная фауна наиболее богата в защищенных, хорошо прогреваемых бухтах. Приливы и отливы не препятствуют развитию псаммофильной фауны. Когда вода во время отлива временно уходит, обнажая песок, то в толще песка, в промежутках между песчинками она остается, и это не мешает существованию животных.
У инфузорий, входящих в состав псаммофильной фауны и относящихся к различным систематическим группам (отрядам, семействам), в процессе эволюции вырабатывается много общих признаков, являющихся приспособлениями к своеобразным условиям существования между песчинками.
На рис. 106 показаны некоторые виды псаммофильной фауны инфузорий, принадлежащие к разным отрядам и семействам. Между ними много общего. Тело большинства из них более или менее сильно вытянуто в длину, червеобразно. Это позволяет легко «протиснуться» в мельчайшие дырочки между песчинками. У очень многих видов (рис. 106) удлинение тела сочетается с его уплощением. Цилиарный аппарат всегда хорошо развит, что позволяет активно, с определенной силой, двигаться в узких щелях. Довольно часто на одной стороне червеобразного уплощенного тела развиваются реснички, противоположная сторона голая. Эта особенность, вероятно, связана с ярко выраженной у большинства псаммофильных видов способностью очень плотно и очень прочно прилипать (прикрепляться) к субстрату через ресничный аппарат (явление, называемое тигмотаксисом). Это свойство позволяет животным оставаться на месте в тех случаях, когда в узких промежутках, где они обитают, возникают течения воды. В этом случае, вероятно, выгоднее, чтобы сторона, противоположная той, которой животное прикреплялось к субстрату, была гладкой.
Чем питаются псаммофильные инфузории? Значительную часть «рациона» у многих видов составляют водоросли, особенно диатомовые. Бактерии служат им в меньшей степени пищей. Это также в значительной степени зависит от того, что в песках, которые не сильно загрязнены, мало бактерий. Наконец, особенно среди самых крупных псаммофильных инфузорий, имеется значительное число хищных форм, поедающих других инфузорий, принадлежащих к более мелким видам. Псаммофильные инфузории распространены, по-видимому, повсеместно.
INFUSORIA APOSTOMATES
инфузории spirophria (Spirophrya subparasitica) в инцистированном состоянии часто можно встретить сидящими на небольшом стебельке на мелких планктонных морских рачках (особенно часто на onus Igendia ракообразных). Пока рачок активно плавает в морской воде, сидящие на нем спирофрии не претерпевают никаких изменений. Для дальнейшего развития инфузории необходимо, чтобы рачок был съеден морским гидроидным полипом, что бывает довольно часто (рис. 107). Как только цисты спирофрии вместе с рачком попадают в пищеварительную полость, из них тут же выходят мелкие инфузории, которые начинают энергично питаться пищевой кашицей, образовавшейся в результате переваривания проглоченного рачка. В течение часа размеры инфузорий увеличиваются в 3-4 раза. Однако размножения на этом этапе не происходит. Перед нами типичная стадия роста инфузории, которая называется трофонт. Через некоторое время вместе с непереваренными остатками пищи трофонт выбрасывается полипом в морскую воду. Здесь, активно плавая, она спускается по телу полипа к его подошве, где и прикрепляется, будучи окружена кистой. Эта стадия инцистированной большой инфузории, сидящей на полипе, называется томонтом. Это фаза размножения. Томонта не питается, но несколько раз подряд быстро делится (рис. 107, 7). В результате получается целая группа очень мелких инфузорий. Их количество зависит от размера томонта, который, в свою очередь, определяется размером трофонта, давшего ему начало. Стадию расселения представляют мелкие инфузории, образующиеся в результате деления томонта (их называют томитами или бродягами).
Выходят из кисты, быстро плавают (не питаясь при этом, а используя имеющиеся у них запасы в цитоплазме). Если им «посчастливилось» встретить копеподу, они тут же прикрепляются к ней и инцистируются. Это этап, с которого мы начали рассмотрение цикла.
В рассмотренном нами жизненном цикле спирофрии обращает на себя внимание резкое разграничение стадий, имеющих различное биологическое значение. Трофон – стадия роста. Он только растет, накапливает большое количество цитоплазмы и всевозможных запасных веществ за счет энергичного и быстрого питания. Трофонт не способен к размножению. У томонта наблюдается обратное явление — неспособность к питанию и энергичное быстрое размножение. После каждого деления роста нет, а потому размножение томонта сводится к быстрому распаду на множество залетных. Наконец, бродяги выполняют свою особую и единственно характерную функцию: они являются особями-расселенцами и распространителями вида. Они не могут ни есть, ни размножаться.
ЖИЗНЕННЫЙ ЦИКЛ ИХТИОФТИРИУСОВ
К концу периода роста ихтиофтириус достигает очень больших размеров по сравнению с залетными: 0,5-1 мм в диаметре. При достижении предельного значения инфузории активно перемещаются из тканей рыб в воду и некоторое время медленно плавают с помощью ресничного аппарата, покрывающего все их тело. Вскоре крупные ихтиофтириусы оседают на каком-нибудь подводном предмете и выделяют цисту. Сразу после инцистирования начинаются последовательные деления инфузории: сначала пополам, затем каждая дочерняя особь снова делится на две, и так до 10-11 раз. В результате внутри кисты образуется до 2000 мелких почти округлых особей, покрытых ресничками. Внутри кисты активно перемещаются бродяги. Они пробивают скорлупу и выходят наружу. Активно плавающие бродяги заражают новых рыб.
Скорость деления ихтиофтириуса в цистах, а также скорость его роста в тканях рыб в значительной степени зависят от температуры. По исследованиям разных авторов приводятся следующие цифры: при 26-27°С развитие залетников в цисте занимает 10-12 часов, при 15-16°С — 28-30 часов, при 4-5°С — 28-30 часов. °С сохраняется в течение 6-7 дней.
Борьба с ихтиофтириозом представляет значительные трудности. Первостепенное значение здесь имеют профилактические мероприятия, направленные на предотвращение проникновения свободноплавающих залетных насекомых в ткани рыб. Для этого полезно проводить частую пересадку больных рыб в новые водоемы или аквариумы, создавать проточные условия, что особенно эффективно при борьбе с ихтиофтириозом.
инфузории триходины
Вся система приспособлений триходин к жизни на поверхности хозяина направлена на то, чтобы не отрываться от тела хозяина (что почти всегда равносильно смерти), сохраняя при этом подвижность. Эти устройства очень совершенны. Тело большинства триходин имеет форму довольно плоского диска, иногда шляпки. Сторона, обращенная к телу хозяина, слегка вогнута, образует присоску для прикрепления. По наружному краю присоски проходит венчик хорошо развитых ресничек, с помощью которых происходит движение (ползание) инфузории в основном по поверхности тела рыбы. Этот венчик соответствует венчику, обнаруженному у обсуждавшихся выше сидячих реснитчатых бродяг с круглыми ресничками. Таким образом, триходину можно сравнить с бродягой. На брюшной поверхности (на присоске) триходины имеют очень сложный опорно-прикрепительный аппарат, помогающий удерживать инфузорию на хозяине. Не вдаваясь в подробности его строения, отметим, что его основу составляет кольцо сложной конфигурации, состоящее из отдельных сегментов, несущих наружные и внутренние зубья (рис. 109)., Б). Это кольцо образует упругую и в то же время прочную основу брюшной поверхности, выполняющую роль присоски. Разные виды триходинов отличаются друг от друга количеством сегментов, образующих кольцо, и конфигурацией наружных и внутренних крючков.
На противоположной от диска стороне тела триходины расположены перисто-ротовой аппарат. Его строение более или менее типично для обрезанных инфузорий . Закрученные по часовой стрелке адоральные перепонки ведут к углублению, на дне которого находится рот. Ядерный аппарат триходина устроен типично для инфузорий: один лентовидный макронуклеус и один микронуклеус, расположенный рядом с ним. Имеется одна сократительная вакуоль.
Триходины широко распространены в водоемах всех типов. Особенно часто они встречаются на мальках разных видов рыб. При массовом размножении триходины наносят большой вред рыбам, особенно если массово закрывают жабры. Это нарушает нормальное дыхание рыбы.
С целью очистки рыб от триходина рекомендуется делать лечебные ванночки из 2% раствора хлорида натрия или 0,01% раствора перманганата калия (малькам — на 10-20 мин).
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА УНГЕЙТС
Из рубца через сетку пища отрыгивается в полость рта, где дополнительно пережевывается (жвачка). Пережеванная пищевая масса, вновь проглоченная, через специальную трубку, образованную складками пищевода, поступает уже не в рубец, а в книгу и оттуда в сычуг, где подвергается воздействию пищеварительных соков жвачных. В сычужном ферменте в условиях кислой реакции и наличия пищеварительных ферментов погибают инфузории. Попадая туда с жевательной резинкой, они перевариваются.
Количество простейших в рубце (как и в сетке) может достигать колоссальных величин. Если взять каплю содержимого рубца и рассмотреть ее под микроскопом (при нагревании, так как при комнатной температуре инфузории останавливаются), то инфузории буквально роятся в поле зрения. Получить такую массу инфузорий сложно даже в культуре. Количество инфузорий в 1 см3 содержимого рубца достигает миллиона, а нередко и больше. В пересчете на весь объем рубца это дает поистине астрономические цифры! Богатство содержимого рубца инфузориями в значительной степени зависит от характера питания жвачного. Если пища богата клетчаткой и бедна углеводами и белками (трава, солома), то инфузорий в рубце относительно мало. При добавлении в рацион углеводов и белков (отрубей) численность инфузорий резко возрастает и достигает огромных размеров. Необходимо учитывать, что происходит постоянный отток инфузорий. Попадая вместе с жвачкой в сычуг, они погибают. Высокий уровень численности инфузорий поддерживается за счет их бурного размножения.
Непарнокопытные (лошадь, осел, зебра) также имеют большое количество инфузорий в пищеварительном тракте, но их локализация в организме хозяина иная. Непарнокопытные не имеют сложного желудка, в связи с чем возможность развития простейших в передних отделах пищеварительного тракта отсутствует. Зато у непарнокопытных очень хорошо развиты крупная и слепая кишка, обычно забитые пищевыми массами и играющие существенную роль в пищеварении. В этом отделе кишечника, так же как в рубце и сетке жвачных животных, развивается очень богатая фауна простейших, главным образом инфузорий, большая часть которых также относится к отряду эндодиниоморфных. Однако по видовому составу фауна рубца жвачных и фауна толстого кишечника непарнокопытных не совпадают.
ИНФОРСУНОЧКИ КИШЕЧНО-КИШЕЧНОГО ТРАКТА ЖВАЧИХ
Наибольший интерес представляют инфузории семейства офриосколецид (Ophryoscolecidae), родственные отряду эндодиниоморф . характерная особенность этой отслойки — отсутствие сплошного ресничного покрова. Сложные реснитчатые образования — цирры — располагаются на переднем конце тела инфузорий в области ротового отверстия. К этим основным элементам цилиарного аппарата могут присоединяться дополнительные группы цирр, располагающиеся либо на переднем, либо на заднем конце тела. Общее число видов инфузорий семейства офриосколецид около 120.
На рис. 110 показаны некоторые наиболее типичные офриосколециды из рубца жвачных животных. Наиболее просто устроены инфузории рода Энтодиниум (Entodinium, рис. 110, L). На переднем конце их тела имеется одна периоральная зона цирра. Передний конец тела, на котором расположено ротовое отверстие, может быть втянут внутрь. Эктоплазма и эндоплазма резко разграничены. На заднем конце хорошо видна анальная трубка, служащая для удаления непереваренных остатков пищи. Несколько более сложная структура аноплодиния (Anoplodinium, рис. 110, Б). Имеют две зоны цилиарного аппарата — периоральные цирры и дорсальные цирры. Оба расположены на переднем конце. На заднем конце тела вида, показанного на рисунке, имеются длинные острые выросты — это довольно типично для многих видов офриосколецид. Высказано предположение, что эти выросты способствуют «проталкиванию» инфузорий между растительными частицами, заполняющими рубец.
Виды Род Eudiplodynia (Eudiplodinium, рис. 110, Б) сходны с anoplodynia , но в отличие от них имеют скелетную базальную пластинку, расположенную на правом крае вдоль глотки. Эта скелетная пластина состоит из вещества, похожего по химической природе на волокно, то есть на вещество, из которого состоят оболочки растительных клеток.
У рода полипластрон (Polyplastron, рис. 110, Г, Д) наблюдается дальнейшее усложнение скелета. Строение этих инфузорий близко к эвдиплодиниям. Различия сводятся прежде всего к тому, что вместо одной скелетной пластинки у этих инфузорий их пять. Два из них, самые крупные, расположены с правой стороны, а три, более мелкие, — с левой стороны инфузории. Второй особенностью полипластрона является увеличение числа сократительных вакуолей. Энтодиния имеет одну сократительную вакуоль, аноплодиния и эвдиплодиния — две сократительные вакуоли, а полипластрон — около десятка.
У epidinium (Epidinium, рис. 110), имеющих хорошо развитый углеводный скелет, расположенный на правой стороне тела, дорсальная зона цирра смещается с переднего конца на дорсальную сторону. Шипы часто развиваются на заднем конце инфузорий этого рода.
Наиболее сложное строение обнаруживает род Ofrioscolex (Ophryoscolex), именем которого названо все семейство инфузорий (рис. 110, Д). У них хорошо развита дорсальная зона цирра, покрывающая около 2/3 окружности тела и скелетных пластинок. На заднем конце образуются многочисленные шипы, один из которых обычно особенно длинный.
Знакомство с некоторыми типичными представителями офриосколецид показывает, что внутри этого семейства произошло значительное усложнение организации (от энтодинии до офриосколекса).
Кроме инфузорий из семейства риосколецид , в рубце жвачных животных в небольшом количестве встречаются представители уже известных нам. отряд изоцилиарных инфузорий . Они представлены небольшим количеством видов. Их тело равномерно покрыто продольными рядами ресничек, элементы скелета отсутствуют. В общей массе инфузорного населения рубца они не играют существенной роли, и поэтому мы не будем здесь на них останавливаться.
Чем и как питаются инфузории ofrioscolecides? Этот вопрос подробно изучался многими учеными, особенно подробно профессором В. А. Догелем.
Питание офриосколецид достаточно разнообразно, и у разных видов наблюдается определенная специализация. Самые мелкие виды рода Entodynia питаются бактериями, крахмальными зернами, грибами и другими мелкими частицами. Очень многие средние и крупные отриосколециды поглощают частицы растительных тканей, составляющие основную часть содержимого рубца. Эндоплазма некоторых видов буквально забита растительными частицами. Можно увидеть, как инфузории набрасываются на обрывки растительных тканей, буквально разрывают их на куски, а затем проглатывают, часто закручивая в спираль в своем теле (рис. 111, 4). Иногда приходится наблюдать такие картины (рис. 111, 2), когда тело самой инфузории деформируется из-за проглоченных крупных частиц.
У офриосколецид иногда наблюдается хищничество. Более крупные виды поедают более мелких. Хищничество (рис. 112) сочетается со способностью того же вида питаться растительными частицами.
Каким образом инфузории попадают в рубец жвачных животных? Каковы пути заражения офриосколецидами? Оказывается, у новорожденных жвачных инфузорий в рубце еще нет. Отсутствуют они и во время кормления животного молоком. Но как только жвачное переходит на растительную пищу, в рубце и сетке тут же появляются инфузории, численность которых быстро растет. Откуда они? Долгое время предполагалось, что инфузории рубца образуют некие покоящиеся стадии (скорее всего, цисты), которые широко расселены в природе и при заглатывании дают начало активным стадиям инфузории. Дальнейшие исследования показали, что у жвачных инфузорий нет стадий покоя. Удалось доказать, что заражение происходит активными подвижными инфузориями, проникающими в ротовую полость при отрыгивании жвачки. Если рассмотреть под микроскопом жевательную резинку, взятую из полости рта, в ней всегда содержится большое количество активно плавающих инфузорий. Эти активные формы могут легко проникать в рот и далее в рубец других жвачных из общего сосуда для питья вместе с травой, сеном (в котором может содержаться слюна с инфузориями) и т. д. Такой путь заражения доказан экспериментально.
Если у офриосколецид нет покоящихся стадий, то, очевидно, легко получить «бесинфузорных» животных, изолируя их, когда они еще питаются молоком. Если не допускается непосредственный контакт растущего молодняка и жвачных животных с инфузориями, то молодняк может остаться без инфузорий в рубце. Такие эксперименты проводились несколькими учеными в разных странах. Результат был ясен. При отсутствии контакта молодняка (взятого от матери в период вскармливания молоком) и жвачных с инфузориями в рубце животные вырастают стерильными по отношению к инфузориям. Однако даже кратковременного контакта с животными с инфузориями (общая кормушка, общее ведро для поения, общее пастбище) достаточно, чтобы инфузории появились в рубце стерильных животных.
Выше были приведены результаты опытов по содержанию жвачных животных, полностью лишенных инфузорий, в рубце и сетке. Это достигается, как мы видели, ранней изоляцией молоди. Опыты проводились на овцах и козах.
Таким способом можно было проводить наблюдения за «безинфузорными» животными в течение значительного периода времени (более года). Как влияет отсутствие инфузорий в рубце на жизнь хозяина? Отрицательно или положительно влияет отсутствие инфузорий на хозяина? Чтобы ответить на этот вопрос, на козах были проведены следующие опыты. Детей-близнецов (одного помёта и одного пола) взяли для того, чтобы иметь больше похожего материала. Затем один из близнецов этой пары воспитывался без инфузорий в рубце (ранняя изоляция), а другой с самого начала питания растительной пищей был обильно заражен многими видами инфузорий. Оба получали одинаковую диету и воспитывались в одинаковых условиях. Единственное различие между ними заключалось в наличии или отсутствии инфузорий. На нескольких парах исследованных таким образом малышей не обнаружено различий в ходе развития обоих представителей каждой пары («инфузорных» и «неинфузорных»). Таким образом, можно утверждать, что живущие в рубце и сети инфузории не оказывают резкого влияния на жизнедеятельность животного-хозяина.
Приведенные выше результаты опытов не позволяют, однако, утверждать, что инфузории рубца совершенно безразличны к хозяину. Эти эксперименты проводились при обычном рационе хозяина. Возможно, что и в других условиях, при ином рационе (например, при недостаточном кормлении) удастся выявить влияние на фауну хозяина простейших, обитающих в рубце.
В литературе высказывались различные предположения о возможном положительном влиянии простейшей фауны рубца на процессы пищеварения хозяина. Было указано, что многие миллионы инфузорий, активно плавая в рубце и раздавливая растительные ткани, способствуют брожению и перевариванию пищевых масс, находящихся в передних отделах пищеварительного тракта. Значительное количество инфузорий, попадающих в сычуг вместе с жвачкой, переваривается, а белок, составляющий значительную часть тела инфузорий, усваивается. Таким образом, инфузории могут быть дополнительным источником белка для хозяина. Высказывались также предположения, что инфузории способствуют перевариванию клетчатки, составляющей основную часть пищи жвачных животных, и переводу ее в более усвояемое состояние.
Все эти предположения не доказаны, и против некоторых из них были выдвинуты возражения. Указывалось, например, что инфузории строят протоплазму своего тела из белков, попадающих в рубец с пищей хозяина. Поглощая растительный белок, они превращают его в животный белок своего организма, который затем переваривается в сычуге. Приносит ли это какую-либо пользу хозяину, остается неясным. Все эти вопросы представляют большой практический интерес, поскольку речь идет о пищеварении жвачных животных — основных объектов животноводства. Крайне желательны дальнейшие исследования роли инфузорий рубца в пищеварении жвачных животных.
Офриосколециды жвачных животных обладают, как правило, широкой специфичностью. По видовому составу население рубцового и сетчатого крупного рогатого скота, овец и коз очень близко друг к другу. Если сравнить видовой состав рубца африканских антилоп с крупным рогатым скотом, то и здесь общими будут около 40% от общего числа видов. Однако есть много видов офриосколецид, которые встречаются только у антилоп или только у оленей. Таким образом, на фоне общей широкой специфичности офриосколецидов можно говорить об отдельных, более узкоспецифических их видах.
Инфузории кишечника непарнокопытных
Обратимся теперь к краткому знакомству с инфузориями, населяющими толстую и слепую кишку непарнокопытных.
В видовом отношении эта фауна, как и фауна рубца жвачных животных, также очень разнообразна. В настоящее время описано около 100 видов инфузорий, обитающих в толстом кишечнике животных семейства лошадиных. Встречающиеся здесь инфузории в смысле принадлежности к разным систематическим группам более разнообразны, чем инфузории рубца жвачных.
В кишечнике лошадей встречается довольно много видов инфузорий, относящихся к отряду изоцилиарных, т. е. инфузорий, у которых ресничный аппарат не образует мембранелл или цирроза вблизи ротовой зоны (рис. 113, 1) .
Отряд энтодиниоморф (Entodiniomorpha) также богато представлен в кишечнике лошади. В то время как в рубце жвачных животных встречается только одно семейство эндодиниоморф (семейство офриосколецид), в кишечнике лошади обитают представители трех семейств, но на характеристике которых мы здесь останавливаться не будем, ограничившись лишь несколькими рисунками типичные виды лошадей (рис. 113).
Детальные исследования А. Стрелкова показали, что разные виды инфузорий далеко не равномерно распределены по толстому кишечнику лошади. Есть две разные группы видов, как бы две фауны. Один из них заселяет слепую кишку и брюшную часть толстой кишки (начальные отделы толстой кишки), а другой заселяет дорсальную часть толстой и тонкой кишки. Эти два комплекса видов довольно резко разграничены. Общих для этих двух секций видов немного – менее десятка.
,
Интересно отметить, что среди многочисленных видов инфузорий, населяющих толстый кишечник непарнокопытных, есть представители одного рода, относящегося к сосущим инфузориям. Как мы видели выше, сосущие инфузории (Suctoria) — типичные свободноживущие сидячие организмы с совершенно особым способом питания с помощью щупалец (рис. 103). Один из родов сукторов приспособился к такой, казалось бы, необычной среде обитания, как толстая кишка лошади, например, несколько видов аллантоз (аллантосомы). У этих очень своеобразных животных (рис. 114) нет стебля, реснички отсутствуют, хорошо развиты булавовидные утолщенные на концах щупальца.
С помощью щупалец аллантосомы присасываются к разным видам инфузорий и высасывают их. Зачастую добыча во много раз превосходит хищника.
До сих пор не ясен вопрос о характере взаимоотношений инфузорий толстого кишечника непарнокопытных и их хозяев. Численность инфузорий может быть столь же высокой, а иногда и больше, чем в рубце жвачных животных. Имеются данные, показывающие, что количество инфузорий в толстом кишечнике лошади может достигать 3 млн в 1 см3. Предполагаемое некоторыми учеными симбиотическое значение еще менее вероятно, чем для инфузорий рубца.
Наиболее вероятно мнение, что они наносят некоторый вред хозяину, поглощая значительное количество пищи. Часть инфузорий выводится с фекальными массами, и таким образом органические вещества (в том числе белок), входящие в их тело, остаются неиспользованными хозяином.
До сих пор не решен вопрос о путях заражения непарнокопытных инфузориями, обитающими в толстой кишке.
Балантидии захватывают различные частицы пищи из содержимого толстого кишечника. Особенно охотно он питается крахмальными зернами. Если балантидии обитают в просвете толстой кишки человека, то они питаются содержимым кишечника и не оказывают вредного воздействия. Это типичный «лафет», с которым мы встречались уже при рассмотрении дизентерийной амебы. Однако балантидии с меньшей вероятностью, чем дизентерийные амебы, останутся таким «безобидным жильцом».
В настоящее время специалистами хорошо разработаны различные методы, позволяющие культивировать балантидии в искусственной среде — вне организма хозяина.
Как видно из рисунка, троглодптелла относится к числу сложных эндодиниоморф. Она, кроме периоральной зоны цирроза (на переднем конце тела), имеет еще три зоны хорошо развитых цирр, кольцеобразно покрывающих тело инфузории. Троглодителлы имеют хорошо развитый скелетный аппарат, состоящий из углеводов, покрывающий почти весь передний конец тела. Размеры этих своеобразных инфузорий весьма значительны. В длину они достигают 200-280 мкм.
Безротые инфузории АСТОМАТЫ
Опорные скелетные образования развиваются преимущественно на переднем конце тела, которому приходится испытывать механические нагрузки и преодолевать препятствия, проталкиваясь в просвет кишечника между пищевыми частицами. Вид рода radiophria (Radiophrya) на переднем конце с одной стороны тела (условно считающейся брюшной) имеются очень сильные эластичные ребра (спикулы), залегающие в поверхностном слое эктоплазмы (рис. 117, Б). , Д, Е). Виды Род Menilella (Mesnilella) имеются также опорные лучи (спикулы), которые на большей части своей длины лежат в более глубоких слоях цитоплазмы (в эндоплазме, рис. 117, А). Аналогично устроенные опорные образования развиты и у видов некоторых других родов Astomat.
Бесполое размножение у некоторых инфузорий астомат протекает своеобразно. Вместо поперечного деления надвое, характерного для большинства инфузорий, у многих астоматов деление неравномерное (почкование). В то же время отделяющиеся на заднем конце почки некоторое время остаются связанными с материнской особью (рис. 117, Б). В результате получаются цепочки, состоящие из передней крупной и задней более мелких особей (почек). В дальнейшем почки постепенно отделяются от цепочки и переходят к самостоятельному существованию. Эта своеобразная форма размножения широко распространена, например, у уже известных нам радиофрий. Цепочки некоторых астоматов, образующиеся в результате почкования, по внешнему виду напоминают цепочки ленточных червей. Здесь мы снова сталкиваемся с явлением конвергенции.
Ядерный аппарат астомата имеет характерное для инфузорий строение: макронуклеус, чаще всего лентовидной формы (рис. 117), и один микронуклеус. Сократительные вакуоли обычно хорошо развиты. Большинство видов имеют несколько (иногда более десятка) сократительных вакуолей, расположенных в один продольный ряд.
Изучение распределения видов астомат по разным типам хозяев показывает, что большая часть астомат приурочена к строго определенным типам хозяев. Большинству астоматов свойственна узкая специфичность: хозяином для них может служить только один вид животного.
Несмотря на большое количество работ, посвященных изучению астоматных инфузорий, остается совершенно неясным один очень важный аспект их биологии: как происходит передача этих инфузорий от одной особи хозяина к другой? Никогда не удавалось наблюдать образование цист у этих инфузорий.
Поэтому предполагается, что заражение протекает активно — подвижные стадии.
ИНФУЗОРИИ КИШЕЧНИКА МОРСКОГО ЕЖА
Очень многочисленны морские ежи в прибрежной зоне наших северных (Баренцево) и дальневосточных морей (Японское море, тихоокеанское побережье Курильских островов). Большинство морских ежей питаются растительной пищей, преимущественно водорослями, которые они соскребают с подводных предметов особыми острыми «зубами», окружающими ротовое отверстие. В кишечнике этих травоядных ежей имеется богатая фауна инфузорий. Часто они развиваются здесь в массовом количестве, и содержимое кишечника морского ежа под микроскопом почти так же «кишит» инфузориями, как и содержимое рубца жвачных. Надо сказать, что, кроме глубоких различий в условиях жизни инфузорий кишечника морского ежа и рубца жвачного, есть и некоторые сходства. Они заключаются в том, что и там, и там инфузории обитают в среде, очень богатой растительными остатками. В настоящее время известно более 50 видов инфузорий, обитающих в кишечнике морских ежей, которые встречаются только в прибрежной зоне, где ежи питаются водорослями. На больших глубинах, где уже не растут водоросли, у морских ежей нет инфузорий.
По образу жизни и характеру питания большинство инфузорий кишечника морских ежей растительноядны. Питаются водорослями, которые в большом количестве заполняют кишечник хозяина. Некоторые виды довольно «привередливы» в выборе пищи. Например, стробилидиум (Strobilidium, рис. 118, А) питается почти исключительно крупными диатомеями. Есть и хищники, которые поедают представителей других, более мелких видов инфузорий.
У инфузорий из кишечника морских ежей, в отличие от астоматов, нет строгой привязки к определенным видам хозяев. Они живут в самых разнообразных морских ежей, питающихся водорослями.
Пути заражения морских ежей инфузориями не изучены. Однако здесь с большой долей вероятности можно предположить, что она встречается в активных свободноплавающих формах. Дело в том, что инфузории из кишечника морских ежей могут длительное время (многие часы) жить в морской воде. Однако они уже настолько приспособились к жизни в кишечнике ежей, что вне их тела, в морской воде, рано или поздно погибают.
Заканчивая знакомство с инфузориями, следует еще раз подчеркнуть, что они представляют собой богатую видами, обширную и процветающую группу (класс) животного мира. Оставаясь на уровне клеточной организации, инфузории достигли, по сравнению с другими классами простейших, наибольшей сложности строения и функций.
Особенно значительную роль в этом прогрессивном развитии (эволюции), вероятно, сыграли трансформация ядерного аппарата и возникновение ядерного дуализма (качественная неравноценность ядер). Богатство макронуклеуса нуклеиновыми веществами связано с активными метаболическими процессами, с бурными процессами синтеза белков в цитоплазме и ядрах.
Заключение
Подошел к концу наш обзор строения и образа жизни экстенсивного типа животного мира — простейших . Их характерной чертой, как неоднократно подчеркивалось выше, является одноклеточность. По своему строению простейшие представляют собой клетки. Однако они несравнимы с клетками, составляющими тело многоклеточных организмов, потому что сами являются организмами. Таким образом, простейшие – это организмы на клеточном уровне организации. Некоторые высокоорганизованные простейшие, обладающие множеством ядер, уже как бы выходят за морфологические пределы строения клетки, что дает некоторым ученым основание называть таких простейших «надклеточными». Это мало меняет сути дела, ибо для простейших по-прежнему характерна одноклеточная организация.
В пределах одноклеточности простейшие прошли долгий путь эволюционного развития и дали огромное разнообразие форм, приспособленных к самым разнообразным условиям жизни. В основе родословной простейших лежат два класса: саркодовые и жгутиковые. Вопрос о том, какой из этих классов более примитивен, до сих пор обсуждается в науке. С одной стороны, низшие представители саркодовых (амебы) имеют наиболее примитивное строение. Но жгутиковые проявляют наибольшую пластичность типа обмена веществ и стоят как бы на границе между животным и растительным миром. В жизненном цикле некоторых саркодовых (например, фораминифер) есть жгутиковые стадии (гаметы), что указывает на их родство с жгутиковыми. Очевидно, что ни современные саркоды, ни современные жгутиконосцы не могут быть исходной группой эволюции животного мира, поскольку сами прошли долгий путь исторического развития и выработали многочисленные приспособления к современным условиям жизни на Земле. Вероятно, оба эти класса современных простейших следует рассматривать как два ствола в эволюции, происходящие от не доживших до наших дней древних форм, живших на заре развития жизни на нашей планете.
В ходе дальнейшей эволюции простейших произошли изменения другого характера. Некоторые из них привели к общему повышению уровня организованности, повышению активности, интенсивности жизненных процессов. К числу таких филогенетических (эволюционных) преобразований следует, например, отнести развитие органелл движения и захвата пищи, достигшее высокого уровня совершенства в классе инфузорий. Бесспорно, что реснички являются органеллами, соответствующими (гомологичными) жгутикам. Если у жгутиковых, за немногими исключениями, число жгутиков невелико, то у инфузорий количество ресничек достигает многих тысяч. Развитие цилиарного аппарата резко повысило активность простейших, сделало более разнообразными и сложными формы их взаимоотношений с окружающей средой, формы реакций на внешние раздражители. Наличие дифференцированного ресничного аппарата, несомненно, явилось одной из главных причин прогрессивной эволюции в классе инфузорий, где возникло большое разнообразие форм, приспособленных к различным средам обитания.
Развитие ресничного аппарата инфузорий является примером такого эволюционного изменения, которое было названо акад. Северцова Ароморфозы. Ароморфозы характеризуются общим повышением организации, развитием приспособлений широкого значения. Под повышением организации понимают изменения, вызывающие повышение жизнедеятельности организма; они связаны с функциональной дифференциацией его частей, приводят к более разнообразным формам связи организма со средой. Развитие ресничного аппарата инфузорий относится именно к такого рода структурным преобразованиям в процессе эволюции. Это типичный ароморфоз.
У простейших, как подчеркивал В. А. Догель, изменение типа ароморфозов обычно связано с увеличением числа органелл. Происходит полимеризация органелл. Характерным примером таких изменений является развитие цилиарного аппарата у инфузорий. Вторым примером ароморфоза в эволюции инфузорий является их ядерный аппарат. Мы рассмотрели выше особенности строения сердцевины инфузорий. Ядерный дуализм инфузорий (наличие микронуклеуса и макронуклеуса) сопровождался увеличением числа хромосом в макронуклеусе (явление полиплоидии). Поскольку хромосомы связаны с основными синтетическими процессами в клетке, прежде всего с синтезом белков, этот процесс привел к общему повышению интенсивности основных жизненных функций. Здесь тоже происходила полимеризация, затрагивающая хромосомные комплексы ядра.
инфузории — одна из самых многочисленных и прогрессивных групп простейших, происходит от жгутиковых. Об этом свидетельствует полное морфологическое сходство их органоидов движения. Этот этап эволюции был связан с двумя крупными ароморфозами: один из них затронул органеллы движения, второй — ядерный аппарат. Оба эти вида изменений взаимосвязаны, так как оба ведут к повышению жизнедеятельности и усложнению форм взаимоотношений с внешней средой.
Наряду с ароморфозами существует и другой тип эволюционных изменений, выражающийся в развитии приспособлений (приспособлений) к определенным, четко определенным условиям существования. Этот тип эволюционного изменения Северцов назвал идиоадаптацией. В эволюции простейших этот тип изменения сыграл очень важную роль. Выше при рассмотрении разных классов простейших приведены многочисленные примеры идиоадаптивных изменений. Приспособления к планктонному образу жизни у разных групп простейших, приспособления к жизни в песке у инфузорий, образование защитных оболочек ооцист у кокцидий и многое другое — все это идиоадаптации, сыгравшие большую роль в возникновении и развитии отдельных групп, но не связаны с общими прогрессивными изменениями в организации.
Адаптации к различным специфическим средам обитания у простейших очень разнообразны. Они обеспечили широкое распространение этого типа в самых разнообразных местообитаниях, о чем подробно говорилось выше, при описании отдельных классов.
Большая Медицинская Энциклопедия
Инфузория Tetrahymena thermophila … Википедия
— (Инфузории) класс наиболее высокоразвитых простейших (Protozoa). Основные признаки И.: наличие ресничек (для движения и питания), двух типов ядер (полиплоидный макронуклеус и диплоидный микронуклеус, различных по строению и… … Большая советская энциклопедия
Или жидкие животные (инфузории) класс простейших животных, снабженных мерцающими жгутиками или ресничками, иногда сменяемыми в зрелом возрасте цилиндрическими присосательными выростами, с определенной формой тела, большей частью с… … Энциклопедический словарь Ф.А. Брокгауз и И.А. Эфрон
Таксономия классификация ифузорий и систематизация организмов, относящихся к роду инфузорий, в соответствии с общей концепцией систематики простейших в частности и животных вообще Содержание 1 Систематика 1. 1 Классические … Википедия
Или жидкие животные (инфузории) класс простейших животных, снабженных мерцающими жгутиками или ресничками, которые во взрослом состоянии иногда сменяются цилиндрическими присосательными выростами, с определенной формой тела, большей частью с….. Энциклопедия Брокгауза и Ефрона
Инфузория туфелька обитает в небольших стоячих водоемах. По форме тела он напоминает подошву ботинка, достигает в длину 0,1-0,3 мм, покрыт прочной эластичной оболочкой — пелликулой, под которой в экто- и эндоплазме находятся скелетные поддерживающие нити. Такое строение позволяет инфузориям сохранять постоянную форму тела.
Органеллы движения — волосовидные реснички (у инфузорий их 10-15 тысяч), покрывающие все тело. При исследовании ресничек с помощью электронного микроскопа было обнаружено, что каждая из них состоит из нескольких (около 11) волокон. В основе каждой реснички лежит базальное тельце, расположенное в прозрачной эктоплазме. Туфелька быстро передвигается благодаря слаженной работе ресничек, загребающих воду.
В цитоплазме инфузорий четко различают эктоплазму и эндоплазму. В эктоплазме, между основаниями ресничек парамеций, располагаются атакующие и защитные органеллы — мелкие веретенообразные тельца — трихоцисты. На фотографиях, сделанных с помощью электронного микроскопа, видно, что выброшенные трихоцисты снабжены гвоздевидными кончиками. При раздражении трихоцисты выбрасываются наружу, превращаясь в длинную эластичную нить, поражающую противника или добычу.
В эндоплазме расположены — два ядра (большое и малое) и системы пищеварительных и выделительных органелл.
Пищевые органеллы . На так называемой вентральной стороне находится предротовое углубление — перистом, ведущее к устью клетки, переходящее в глотку (цитофаринкс), открывающееся в эндоплазму. Вода с бактериями и одноклеточными водорослями, которыми питаются инфузории, выгоняется через рот и глотку специальной группой ресничек перистома в эндоплазму, где окружается пищеварительной вакуолью. Последний постепенно перемещается по телу инфузории. По мере движения вакуоли проглоченные бактерии в течение часа перевариваются сначала с кислой, а затем с щелочной реакцией. Непереваренный остаток выбрасывается наружу через специальное отверстие в эктоплазме — порошкообразную, или анальную пору.
Органоиды осморегуляции . На переднем и заднем концах тела, на границе экто- и эндоплазмы, имеется по одной пульсирующей вакуоли (центральный резервуар), вокруг которой в венчике располагаются 5-7 приводящих трубочек. Вакуоль наполняется жидкостью из этих афферентных каналов, после чего заполненная жидкостью вакуоль (диастолическая фаза) сокращается, изливает жидкость через небольшое отверстие и схлопывается (фаза систолы). Вслед за этим жидкость, вновь заполнившая приводящие каналы, изливается в вакуоль. Передние и задние вакуоли сокращаются попеременно. Пульсирующие вакуоли выполняют двойную функцию — возврат избыточной воды, необходимой для поддержания постоянного осмотического давления в организме парамеция, и выделение продуктов диссимиляции.
Тапочек ядерный аппарат представлен как минимум двумя качественно различными ядрами, расположенными в эндоплазме. Форма ядер обычно овальная.
Большое вегетативное ядро называется макронуклеусом. В ней происходит транскрипция — синтез на матрицах ДНК информационных и других форм РНК, которые уходят в цитоплазму, где на рибосомах осуществляется синтез белка.
Генератив малый — микронуклеус. Находится рядом с макронуклеусом. В нем перед каждым делением число хромосом удваивается, поэтому микронуклеус рассматривается как «депо» наследственной информации, передающейся из поколения в поколение.
Инфузория-туфелька размножается как бесполым, так и половым путем.
При бесполом размножении клетка скручивается пополам вдоль экватора, а размножение осуществляется поперечным делением. Этому предшествуют митотическое деление малого ядра и процессы, характерные для митоза в большом ядре.
После многократного бесполого размножения в жизненном цикле происходит половой процесс, или конъюгация.
Половой процесс заключается во временном соединении двух особей ротовыми отверстиями и обмене частями их ядерного аппарата с небольшим количеством цитоплазмы. При этом крупные ядра распадаются на части и постепенно растворяются в цитоплазме. Мелкие ядра сначала делятся дважды, число хромосом уменьшается, затем три из четырех ядер разрушаются и растворяются в цитоплазме, а четвертое снова делится. В результате этого деления образуются два гаплоидных половых ядра. Один из них — мигрирующий, или мужской — переходит в соседнюю особь и сливается с оставшимся в ней женским (неподвижным) ядром. Тот же процесс происходит и в другом конъюгате. После слияния мужского и женского ядер диплоидный набор хромосом восстанавливается и инфузории расходятся. После этого у каждой инфузории новое ядро делится на две неравные части, в результате чего образуется нормальный ядерный аппарат — большое и малое ядро.
Конъюгация не приводит к увеличению количества особей. Его биологическая сущность состоит в периодической перестройке ядерного аппарата, его обновлении и повышении жизнеспособности инфузории, ее приспособляемости к окружающей среде.
Туфелька и некоторые другие свободноживущие инфузории питаются бактериями и водорослями. В свою очередь инфузории служат пищей для мальков рыб и многих беспозвоночных. Иногда туфельки разводят для кормления мальков рыб, только что вылупившихся из икры.
Общая характеристика вида инфузорий краткая. Вид инфузории, или цилиарные (цилиофора, или инфузория). Внешнее и внутреннее строение инфузории туфельки
Туфелька инфузория — простейший одноклеточный организм размером около 0,1 мм. Встречается в тех же водоемах, что и эвглена, и простейшая амеба. Питается в основном бактериями и микроскопическими водорослями. Служит пищей для личинок, мелких рыб, ракообразных.
Внешний вид инфузории туфельки
За сходство с подошвой женской обуви этот вид инфузорий приобрел второе название – «туфелька». Форма этого одноклеточного организма постоянна и не меняется в зависимости от роста или других факторов. Все тело покрыто крошечными ресничками, похожими на жгутики эвглены. Удивительно, но таких ресничек на каждой особи около 10 тысяч! С их помощью клетка передвигается в воде и захватывает пищу.
Инфузория-туфелька, строение которой так знакомо по учебникам биологии, невооруженным глазом не видна. Инфузории – мельчайшие одноклеточные организмы, но при большой концентрации их можно увидеть без увеличительных приборов. В мутной воде они будут выглядеть как продолговатые белые точки, находящиеся в постоянном движении.
Строение инфузории туфельки
Особенности строения инфузории туфельки заключаются не только во внешнем сходстве с подошвой туфельки. Внутренняя организация этого, казалось бы, простого организма всегда представляла большой интерес для науки. Одиночная клетка покрыта плотной оболочкой, содержащей цитоплазму. Эта студенистая жидкость содержит два ядра, большое и маленькое. Крупный отвечает за питание и выделение клеток, мелкий – за размножение.
Отверстие, выполняющее функцию рта, расположено на широкой стороне клетки. Она ведет в глотку, на конце которой образуются пищеварительные вакуоли.
Строение тела инфузории туфельки также отличается очень интересной особенностью – наличием трихоцист. Это особые органы, а точнее органеллы, служащие клетке для питания и защиты. Заметив пищу, инфузория выбрасывает трихоцисты и удерживает ими добычу. Она выдвигает их, когда хочет защититься от хищников.
Питание инфузории туфельки
Одноклеточные организмы питаются бактериями, которые в изобилии живут в загрязненной, мутной воде. Не исключение и инфузория-туфелька, строение рта которой позволяет улавливать проплывающие мимо бактерии и быстро направлять их в пищеварительную вакуоль. Рот инфузории окружен ресничками, которые в этом месте длиннее, чем в других частях тела. Они образуют околоротовую воронку, позволяющую захватить как можно больше пищи. Вакуоли образуются в цитоплазме по мере необходимости. При этом пища может перевариваться сразу в нескольких вакуолях. Время переваривания составляет около часа.
Инфузория почти непрерывно питается, если температура воды выше 15 градусов. Кормление прекращается до начала размножения.
Дыхание и выделение туфельки инфузории
Что касается дыхания, то здесь туфелька инфузории имеет сходное с другими простейшими строение. Дыхание осуществляется всей поверхностью тела тела. Этот процесс обеспечивают две сократительные вакуоли. Отработанный газ проходит по специальным канальцам и выводится через одну из сократительных вакуолей. Выделение лишней жидкости, которая является результатом жизнедеятельности, происходит каждые 20-25 секунд, также за счет сокращения. При неблагоприятных условиях инфузория прекращает питание, а сократительные движения вакуолей значительно замедляются.
Размножение инфузории туфельки
Инфузория башмачок размножается делением. Примерно раз в сутки ядра, большие и малые, расходятся в разные стороны, вытягиваются и делятся надвое. У каждой новой особи остается одно ядро и одна сократительная вакуоль. Второй формируется за несколько часов. Каждая туфелька инфузории имеет идентичное строение родительской.
У инфузорий, подвергшихся множественным делениям, наблюдается такое явление, как половое размножение. Два человека соединяются друг с другом. Внутри образовавшейся большой клетки происходит деление ядер и обмен хромосомами. После завершения такого сложного химического процесса инфузории отсоединяются. Количество особей от этого не увеличивается, но они становятся более жизнеспособными в меняющихся внешних условиях.
Строение и жизнедеятельность инфузории туфельки мало зависит от внешних факторов. Вся обувь выглядит одинаково, имеет одинаковую форму и размер вне зависимости от условий. Жизнедеятельность также протекает по одному сценарию. Имеют значение только температурный и световой факторы. Инфузории очень чувствительны к изменениям освещенности. Можно провести небольшой эксперимент: затемнить сосуд, в котором живут инфузории, оставив небольшое светлое окошко. Все особи будут стянуты в эту дыру через пару часов. Также инфузории воспринимают перепады температуры. При понижении до 15°С башмачки перестают питаться и размножаться, впадая в своеобразный анабиоз.
Тип Инфузории, или инфузории, являются наиболее сложными простейшими. На поверхности тела у них есть органеллы движения — реснички. В клетке инфузории два ядра: большое ядро отвечает за питание, дыхание, движение, обмен веществ; малое ядро участвует в половом процессе.
Особенности строения и жизнедеятельности инфузорий рассмотрены на примере инфузории-туфельки.
Среда обитания, строение и передвижение. В тех же водоемах, где обитают амебы протей и эвглена зеленая, встречается и инфузория-туфелька (рис. 30). Это одноклеточное животное длиной 0,5 мм имеет веретеновидное тело, отдаленно напоминающее башмак. Инфузории-туфельки всегда в движении, плывут тупым концом вперед. Скорость передвижения этого животного достигает 2,5 мм в секунду.
Рис. 30. Строение инфузорий-туфельки: 1 — реснички; 2 — сократительная вакуоль; 3 — цитоплазма; 4 — большое ядро; 5 — малое ядро; б — клеточная мембрана; 7 — устье клетки; 8 — ячейка зева; 9 — пищеварительная вакуоль; 10 — порошок
Тело инфузории более сложное, чем у амебы и эвглены. Тонкая эластичная оболочка, покрывающая инфузорию снаружи, поддерживает постоянную форму ее тела. Этому также способствует захоронение развитых опорных волокон, располагающихся в прилежащем к мембране слое цитоплазмы. На поверхности тела инфузории насчитывается около 15 тысяч колеблющихся ресничек. В основании каждой реснички лежит базальное тельце. Движение каждой реснички состоит из резкого взмаха в одном направлении и более медленного плавного возврата в исходное положение. Реснички колеблются около 30 раз в секунду и, как весла, толкают инфузорий вперед, при этом волнообразное движение ресничек скоординировано. Когда инфузория-туфелька плавает, она медленно вращается вокруг продольной оси тела.
Под эластической оболочкой по всему телу разбросаны особые образования — трихоцисты (от греч. trichos — «волос» иcystis — «пузырь»). Это короткие «стержни», расположенные в один слой перпендикулярно поверхности тела. В случае опасности трихоцисты с силой выбрасываются наружу, превращаясь в тонкие длинные эластичные нити, заражающие нападающего на туфельку хищника. На месте использованных трихоцист со временем появляются новые.
Питание. На теле инфузории имеется углубление — клетчаточный рот, переходящий в клетчаточный зев. Около рта расположены более толстые и длинные реснички. Они гонят в горло бактерии вместе со струей воды – основной пищи туфельки. На дне глотки пища попадает в пищеварительную вакуоль. Пищеварительные вакуоли передвигаются в теле инфузории током цитоплазмы. В вакуоли происходит переваривание пищи, переваренные продукты попадают в цитоплазму и используются для жизнедеятельности. Непереваренные остатки, оставшиеся в пищеварительной вакуоли, выбрасываются в задний конец тела через особую структуру — порошок.
Инфузории-туфельки находят свою добычу, ощущая присутствие химических веществ, выделяющих скопления бактерий.
Выбор. В теле инфузории-туфельки имеются две сократительные вакуоли, которые расположены на переднем и заднем концах тела. Каждая вакуоль состоит из центрального резервуара и 5-7 каналов, направленных к этим резервуарам. Сначала каналы заполняются жидкостью, затем она поступает в центральный резервуар, после чего жидкость выбрасывается наружу. Весь цикл сокращения этих вакуолей происходит один раз в 10-20 секунд. Сократительные вакуоли выводят вредные вещества, образующиеся в организме, и избыток воды.
Дыхание. Как и другие свободноживущие одноклеточные животные, инфузории дышат через покровы тела.
Репродукция. Половой процесс. Инфузории-туфельки обычно размножаются бесполым путем — делением надвое (рис. 31, А). Однако, в отличие от жгутиконосцев, инфузории делятся поперек тела. Ядра делятся на две части, и в каждой новой инфузории имеется одно большое и одно маленькое ядро. Каждая из двух дочерних инфузорий получает часть органелл (например, сократительные вакуоли), а другие образуются заново. Инфузории-туфельки делят один или два раза в день.
Рис. 31. Бесполое размножение(А) и половой процесс (Б) у инфузории туфельки
При половом процессе увеличения числа особей не происходит. Две инфузории временно соединены друг с другом (рис. 31, Б). В месте контакта мембрана растворяется, и между животными образуется соединительный мостик из цитоплазмы. Большое ядро каждой инфузории исчезает. Малое ядро дважды делится, и у каждой инфузории образуется по четыре дочерних ядра. Три из них разрушаются, а четвертый вновь делится. В результате у каждой инфузории остается по два ядра. Одно из этих ядер каждой из двух особей переходит через цитоплазматический мостик в другую инфузорию (т. е. происходит обмен ядрами) и там сливается с оставшимся ядром. Затем у каждой инфузории из этого новообразованного ядра образуются большое и малое ядра, и инфузории расходятся. Этот половой процесс называется конъюгацией. Он длится около 12 часов.
Половой процесс приводит к обновлению, обмену между особями и перераспределению наследственного (генетического) материала, что повышает жизнеспособность организмов.
Рис. 32. Разновидности инфузорий: 1 — бурсарии; 2 — стентор; 3 — стилонихия; 4 — suvoy
Бурсария имеет одно крупное и длинное колбасовидное ядро, мелких ядер около 30. Большинство инфузорий активно плавают, но некоторые из них, например, стилонихии, передвигаются по дну водоема, по водным растениям, как бы ступая на специальные удлиненные реснички, расположенные на брюшной стороне тела… Другие инфузории , например сувойи, прикрепляются к донцу или к растениям на длинных стеблях, которые могут сокращаться за счет особых сократительных волокон. Многие сувои образуют колонии. Эти инфузории питаются в основном бактериями. Сосущие инфузории также ведут малоподвижный образ жизни. У них нет ресничек. Они снабжены сосательными щупальцами в виде тонких сократимых трубочек, служащих для захвата добычи (главным образом других простейших) и высасывания из нее содержимого. Простейшие, прикасающиеся к щупальцам, например жгутиконосцы, моментально прилипают к ним. А затем содержимое жертвы всасывается, как бы закачивается по щупальцу в сосущую инфузорию.
Рис. 33. Простейшие из желудка копытных
Некоторые инфузории обитают в кишечнике крупных травоядных копытных (рис. 33). У коров, овец, коз, антилоп, оленей инфузории в огромном количестве заселяют передние отделы желудка. Эти инфузории питаются бактериями, крахмальными зернами, грибами и частичками растительных тканей. Более крупные инфузории поедают более мелких. В других отделах желудка травоядных происходит переваривание инфузорий. Таким образом, эти инфузории приносят пользу тем животным, в чьих желудках они обитают. Заражение инфузориями происходит во время групповой подкормки или полива.
Лабораторная работа №1
- Тема. Строение и передвижение инфузории-туфельки. Цель. Изучить особенности строения и передвижения инфузорий-туфельок.
- Оборудование: микроскоп, штативная лупа, предметные и покровные стекла, пипетка, вата, культура инфузорий-туфельок в пробирке.
Прогресс
- Определите, видны ли инфузории невооруженным глазом в пробирке.
- Каплю воды с инфузориями-туфельками из пробирки нанести на предметное стекло. Рассмотрите форму тела с помощью лупы, внешнее строение, разницу между передней частью тела и задней частью, способ движения. Подсчитайте количество инфузорий в капле воды.
- Поместите две капли воды с инфузориями на предметное стекло, соедините их водным «мостиком». Поместите кристалл соли на край одной капли. Объясните, что происходит.
- В каплю воды с инфузориями опустите два-три волокна ваты (для замедления движения инфузорий). Аккуратно накройте покровным стеклом.
- Поместите образец под микроскоп. Рассмотрим сначала при малом, а затем при большом увеличении микроскопа, что происходит внутри тела инфузории.
- Зарисуйте внешнее и внутреннее строение инфузории-туфельки, используя большое увеличение микроскопа. Сделайте необходимое обозначение.
- На основании наблюдений перечислите признаки, характерные для инфузорий как представителей простейших.
Инфузории — сложноорганизованные простейшие. У них в клетке два ядра: большое и малое. Они размножаются бесполым и половым путем. Половое размножение способствует обновлению, обмену между особями и перераспределению наследственного (генетического) материала, что повышает жизнеспособность инфузорий.
Упражнения на пройденном материале
- Почему так назвали инфузорию-туфельку?
- Какие признаки доказывают более сложную организацию инфузории-туфельки по сравнению с амебой Proteus и зеленой эвгленой?
- Как проявляется более сложное строение инфузории-туфельки в процессах питания и выделения, чем у других простейших?
- Каковы особенности процесса размножения инфузории-туфельки?
- Почему половой процесс имеет большое биологическое значение в жизни инфузории туфельки?
Классификация инфузорий основана на строении ресничного аппарата всего тела, в том числе периорального. Тип инфузорий делится на два класса: класс ресничных инфузорий (Ciliata) и класс сосущих инфузорий (Suctoria).
Представители реснитчатых инфузорий имеют реснички на протяжении всех фаз развития, а сосущие инфузории большую часть жизненного цикла лишены ресничек.
Класс ресничных — центральный, самый многочисленный класс инфузорий, включающий 3 подкласса и около 20 отрядов.
I. Подкласс Равнополые инфузории (HOLOTRICHA) — тело эквилаций равномерно покрыто ресничками одинаковой длины. Мембраны вокруг рта обычно отсутствуют.
1. Отряд Простоматиды (Prostomatida) — тело инфузории покрыто толстой оболочкой, состоящей из многих рядов пластинок.
Coleps hirtus — клетки мелкие, бочкообразные, коричневого цвета. Тело покрыто многочисленными мелкими пластинками, создающими эффект панциря. Длина тела 20-25 мкм, ширина
10-15 мкм. На переднем полюсе клеток имеются едва заметные зубчики, закрывающие устье клетки. На задней части тела хорошо видна одна хвостовая ресничка, которая в несколько раз длиннее остальных. Сократительная вакуоль одна, расположена на заднем конце тела. Макронуклеус округлый, одиночный, расположен центрально. Обитатель альфа-мезосапробных и полисапробных водоемов (прил. 1, фото 1).
2. Отряд Гимностоматиды (Gymnostomatida) характеризуется расположением устья на переднем конце клетки или сбоку. В основном это плотоядные инфузории. Многие из них имеют хорошо развитый палочковидный аппарат в цитоплазме вблизи устья, что способствует перфорации жертвенной клетки.
Представитель этого отряда инфузории Dileptus anser с щупальцевым отростком на переднем конце и латеральным расположением рта. Dieptus anser — крупные инфузории: длина тела 70–9.0 мкм, ширина 14–20 мкм. Передний конец тела вытянут в виде хоботка, длина которого чуть меньше половины общей длины тела. Каудальная часть клетки не образует шиповатого выроста. Макронуклеус одиночный, четкий, расположен посередине тела. Сократительная вакуоль одна, расположена в задней части клетки. Инфузории проталкивают пищу в рот с помощью длинного переднего отростка. Обитают в водоемах среднего загрязнения (прил. 1, фото 2).
Спатидий поркулюс (Spathidium porculus) — крупные инфузории, длина тела 100–120 мкм. Форма клеток кувшинообразная. Клетчатый рот расположен спереди, широкий, с крупными ресничками по бокам. Макронуклеус колбасы расположен в центре тела. Сократительная вакуоль расположена в каудальной части тела. Инфузории передвигаются медленно. Обычно они обитают в загрязненных водоемах (прил. 1, фото 3).
3. Отряд Colpodida (Колподиды) — ячеек от мала до велика. Клеточное устье расположено посередине вентральной стороны, окаймлено длинными ресничками. Передняя часть тела образует киль.
Colpoda cucullus — имеют хорошо выраженную бобовидную форму тела: спинная сторона выпуклая, а на брюшной стороне имеется глубокое полукруглое углубление, на дне которого имеется ячеистое устье. Окрас инфузории темный: от коричневого до черного. Цитоплазма закупорена пищеварительными вакуолями. Реснички равномерно покрывают тело, образуя 18–20 рядов. Макронуклеус округлый, расположен в срединной части тела. Сократительная вакуоль расположена на заднем конце тела. Встречается в альфа-мезосапробных и полисапробных водоемах (прил. 1, фото 4).
Colpoda maupasi — клетки широкоовальные, темного цвета. Длина 35–70 мкм, ширина 20–40 мкм. На переднем конце тела имеется киль с хорошо заметными 6–7 зубцами. Длина киля составляет 1/3 длины тела. Задний конец тела клетки широко закруглен. Макронуклеус округлый, смещен на дорсальную сторону. Сократительная вакуоль расположена на заднем конце тела. Инфузории обитают в мезосапробных водоемах (прил. 1, фото 5).
Колпода штейни – мелкие инфузории, длина колеблется в пределах 20–35 мкм, ширина 15–30 мкм. Форма тела односторонне выпуклая, при этом спинная сторона выпуклая, а брюшная почти плоская. В средней части брюшной стороны в небольшом углублении расположен ячеистый рот, окруженный длинными ресничками, образующими «бороду». Передний киль с 6–7 отчетливыми ребрами. Макронуклеус овальной формы, расположен ближе к спинной стороне. Сократительная вакуоль одна, расположена на заднем конце тела. Населяет альфа-мезосапробные водоемы (прил. 1, фото 6).
Colpoda aspera — клетки овальной формы, слегка сжатые с боков, светлая цитоплазма. Длина клетки 30-50 мкм, ширина
15-25 мкм. Ресничные ряды 14-16. Передний киль с 5 зубьями. Клеточное устье расположено ближе к середине тела, окружено более длинными ресничками. Макронуклеус округлый, расположен ближе к спинной стороне. Сократительная вакуоль находится в задней части клетки. Обитатель мезосапробных водоемов (прил. 1, фото 7).
4. Отряд Гименостоматиды (Hymenostomatida) — самый многочисленный по количеству видов. Большинство видов отряда свободноживущие, например инфузория-туфелька (Paramecium caudatum). Для этого отряда характерно наличие ротовой воронки — перистома, который с одной стороны окружен длинной мембраной, напротив которой с другой стороны располагаются три мембранеллы. Обычно инфузории питаются бактериями.
Инфузория-туфелька (Paramecium caudatum) — крупные инфузории, длина тела колеблется в пределах 180-280 мкм. Форма тела овальная, вытянутая в длину, напоминает туфельку. Наибольшая ширина в задней трети. Задний конец несколько заострен и несет более длинные реснички, чем остальная часть тела. На одной стороне тела (брюшной) внутрь выступает глубокая борозда, ведущая в глотку. Все тело инфузории покрыто ресничками, их количество около 15 тысяч. Ядерный аппарат состоит из почковидного макронуклеуса и одного довольно крупного микронуклеуса. Инфузории-туфельки обитают в мезосапробных водоемах (прил. 1, фото 8).
Колпидиум колпода (Colpidium colpoda) — мелкие инфузории, длина тела колеблется в пределах 70-90 мкм., ширина — 35-50 мкм. Форма тела напоминает боб: брюшная сторона вогнутая, спинная – выпуклая. Ротовое отверстие треугольной формы, окаймлено рядами ресничек. Ресничный покров плотный и однородный, ресничных рядов много. Макронуклеус округлый, расположен посередине тела. Сократительная вакуоль расположена на заднем конце тела. Инфузории обитают в мезосапробных водоемах (прил. 1, фото 9).).
Uronema marinum — мелкие инфузории, длина тела колеблется в пределах 18-30 мкм., ширина — 7-12 мкм. Форма тела удлиненно-овальная, задняя часть немного расширена. Ресничный покров едва заметен. На заднем конце тела имеется длинная каудальная щетинка. Ротовое отверстие находится в передней части тела. Макронуклеус округлый, расположен посередине тела. Сократительная вакуоль расположена в нижней части тела. Обитатель умеренно загрязненных водоемов (прил. 1, фото 10).
II. Подкласс Реснитчатые инфузории (PERITRICHA) — реснички у инфузорий располагаются только вокруг ротовой воронки, образуя левозакрученную спираль. Большинство видов прикрепляются.
Типичный представитель — Vorticella microstomata , мелкие инфузории, длина тела
колеблется в пределах 30–35 мкм, ширина 25–28 мкм. Форма тела бокаловидная, равномерно сужающаяся кверху. От основания клетки отходит сократительный стебель, в котором проходит пучок мионем. С помощью стебля инфузория прикрепляется к субстрату. Резким выкручиванием стебля сувой моментально спасает от опасности. Одни перитрихиды живут в хижинах, другие образуют пальмовидные колонии (Zoothamnium). Сувой размножают почкованием. При этом образуется свободноплавающая форма – «бродяга». Позже, когда он опустится на дно, образуется стебель. Стебель в 3-4 раза крупнее клеток. Рот окружен венчиком длинных ресничек; от него отходит конусовидный зев. Макронуклеус крупный, С-образно изогнутый, лежит поперек тела. Обитатель полисапробной зоны (приложение 1, фото 11).
III. Подкласс Спиральные ресницы (SPIRITRICHA) — У представителей этого подкласса отсутствует ресничный аппарат. Оральные реснички сильно развиты.
1. Отряд Oligotrichidae — реснички практически полностью исчезли, сохранились лишь короткие ряды отдельных щетинок или очень мало ресничек.
Стромбидиум вирид (Strombidium viride) — мелкие инфузории, длина тела колеблется в пределах 34-50 мкм, ширина — 27-41 мкм. Форма тела ближе к шаровидной: передняя часть широко закругленная, задняя немного удлиненная. Ресничный покров отсутствует. Рот расположен у апикального полюса, окружен венчиком мощных перепонок. Макронуклеус имеет овальную форму и лежит на экваторе клетки. Сократительная вакуоль расположена в апикальной части клетки. Обитатель мезосапробной зоны (приложение 1, фото 12).
По сравнению с другими группами простейших инфузории имеют наиболее сложное строение, что связано с разнообразием и сложностью их функций.
Откуда произошло название «инфузория туфелька»? Вы не удивитесь, если посмотрите под микроскопом живую инфузорию или даже ее изображение (рис. 85).
Действительно, форма тела этой инфузории напоминает изящный женский башмачок.
Инфузория-туфелька находится в непрерывном достаточно быстром движении. Скорость его (при комнатной температуре) около 2,0-2,5 мм/сек. Для такого маленького зверька это отличная скорость! Ведь это означает, что за секунду ботинок проходит расстояние, превышающее длину его тела в 10-15 раз. Траектория движения обуви довольно сложная. Он движется передней частью прямо вперед.
ИНФУЗОРИЯ ТУШЕЧНАЯ (PARAMECIUM CAUDATUM)
Чтобы познакомиться со строением и образом жизни этих интересных одноклеточных организмов, обратимся сначала к одному типичному примеру. Возьмем широко распространенных в небольших пресноводных водоемах инфузорий башмачков (виды рода Paramecium). Этих инфузорий очень легко разводить в небольших аквариумах, если залить прудовой водой обычное луговое сено. В таких настойках развивается множество различных видов простейших и почти всегда развиваются инфузории-туфельки. С помощью обычного учебного микроскопа можно увидеть многое из того, о чем пойдет речь далее.
Среди простейших инфузории туфельки являются достаточно крупными организмами. Длина их тела составляет около 1/6-1/3 мм. и таким образом поворачивается вправо вдоль продольной оси корпуса.
Такое активное движение туфельки зависит от работы большого количества тончайших волосистых придатков — ресничек, которыми покрыто все тело инфузории. Количество ресничек у одной особи инфузории туфельки 10-15 тысяч!
Каждая ресничка совершает очень частые веслообразные движения — при комнатной температуре до 30 ударов в секунду. Во время удара ресничка удерживается в выпрямленном положении. При возвращении в исходное положение (при движении вниз) движется в 3-5 раз медленнее и описывает полукруг.
Когда туфелька плывет, движения многочисленных ресничек, покрывающих ее тело, суммируются. Действия отдельных ресничек оказываются согласованными, в результате чего получаются правильные волнообразные колебания всех ресничек. Волна колебания начинается в передней части тела и распространяется назад. При этом по телу туфельки проходят 2-3 волны сокращения. Таким образом, весь ресничный аппарат инфузорий представляет собой как бы единое функциональное физиологическое целое, действия отдельных структурных единиц которого (реснички) тесно связаны (скоординированы) друг с другом.
Строение каждой отдельной реснички туфельки, как показали электронно-микроскопические исследования, очень сложное.
Направление и скорость движения обуви не являются величинами постоянными и неизменными. Туфелька, как и все живые организмы (мы видели это уже на примере амебы), реагирует на изменение внешней среды изменением направления движения.
Изменение направления движения простейших под влиянием различных раздражителей называется таксисом. У инфузорий легко наблюдать различные таксисты. Если в каплю, где плавают туфли, положить какое-либо неблагоприятно действующее на них вещество (например, кристаллик поваренной соли), то туфли уплывают (как бы убегают) от этого неблагоприятного для них фактора (рис. 86). ).
Это пример отрицательного таксиса при химическом воздействии (отрицательный хемотаксис). Вы также можете наблюдать положительный хемотаксис в обуви. Если, например, каплю воды, в которой плавают инфузории, накрыть покровным стеклом и поместить под нее пузырек с углекислым газом (СО2), то большая часть инфузорий перейдет к этому пузырю и поселится вокруг него в кольцо.
Явление таксиса очень ярко проявляется в обуви под действием электрического тока. Если через жидкость, в которой плавает обувь, пропустить слабый электрический ток, то можно наблюдать следующую картину: все инфузории ориентируют свою продольную ось параллельно к линии тока, а затем, как по команде, двигаться в сторону катода, в районе которого они образуют плотное скопление. Движение инфузорий, определяемое направлением электрического тока, называется гальванотаксисом. Различные таксисы у инфузорий могут обнаруживаться под влиянием самых разнообразных факторов внешней среды.
Все цитоплазматическое тело инфузории четко разделено на 2 слоя: наружный более светлый (эктоплазма) и внутренний более темный и зернистый (эндоплазма). Самый поверхностный слой эктоплазмы образует наружный очень тонкий и на одновременно прочная и эластичная оболочка — пелликула, играющая важную роль в поддержании постоянства формы тела инфузории
В наружном слое (в эктоплазме) тела живого туфельки многочисленные короткие палочки хорошо видны, располагаются перпендикулярно поверхности (рис. 85, 7). Эти образования называются трихоцистами. Их функция очень интересна и связана с защитой простейших. При механическом, химическом или каком-либо другом сильном раздражении трихоцисты выбрасываются с силой, превращаясь в тонкие длинные нити, атакующие нападающего на туфельку хищника. Трихоцисты являются мощными защитными средствами.Они располагаются регулярно между ресничками, так что количество трихоцист примерно соответствует количеству ресничек. На месте использованных («расстрелянных») трихоцист в эктоплазме туфельки развиваются новые.
С одной стороны, примерно посередине туловища (рис. 85, 5), туфелька имеет довольно глубокое углубление. Это ротовая полость, или циррус. На стенках перистома, а также на поверхности тела расположены реснички. Здесь они развиты гораздо мощнее, чем на остальном теле. Эти близко расположенные реснички сгруппированы в две группы. Функция этих особо дифференцированных ресничек связана не с движением, а с питанием (рис. 87).
Как и чем питаются туфельки, как осуществляется их пищеварение?
Тапочки относятся к числу инфузорий, основной пищей которых являются бактерии. Наряду с бактериями они могут заглатывать любые другие частицы, взвешенные в воде, вне зависимости от их пищевой ценности. Периоральные реснички создают непрерывный поток воды с взвешенными в ней частицами в направлении ротового отверстия, расположенного в глубине перистома. Мелкие пищевые частицы (чаще всего бактерии) проникают через рот в малую трубчатую глотку и скапливаются на дне ее, на границе с эндоплазмой. Ротовое отверстие всегда открыто. Пожалуй, не будет ошибкой сказать, что инфузория-туфелька — одно из самых прожорливых животных: она непрерывно питается. Этот процесс прерывается лишь в определенные моменты жизни, связанные с размножением и половым актом.
Скопившийся на дне глотки пищевой комок в дальнейшем отрывается от дна глотки и вместе с небольшим количеством жидкости поступает в эндоплазму, образуя пищеварительную вакуоль. Последний не остается на месте своего образования, а, попадая в токи эндоплазмы, проделывает в теле туфельки довольно сложный и закономерный путь, называемый циклозом пищеварительной вакуоли (рис. 88). Во время этого довольно длительного (при комнатной температуре, занимающего около часа) путешествия пищеварительной вакуоли внутри нее происходит ряд изменений, связанных с перевариванием в ней пищи.
Здесь, как и у амеб и некоторых жгутиковых, происходит типичное внутриклеточное пищеварение. Из эндоплазмы, окружающей пищеварительную вакуоль, в нее поступают пищеварительные ферменты, действующие на пищевые частицы. Продукты переваривания пищи всасываются через пищеварительные вакуоли в эндоплазму.
В ходе циклоза пищеварительной вакуоли в ней происходит смена нескольких фаз пищеварения. В первые моменты после образования вакуоли заполняющая ее жидкость мало отличается от окружающей жидкости. Вскоре начинается поступление пищеварительных ферментов из эндоплазмы в вакуоль и реакция среды внутри нее становится резко кислой. Это легко обнаружить, добавив в пищу индикатор, цвет которого меняется в зависимости от реакции (кислой, нейтральной или щелочной) окружающей среды. В этой кислой среде проходят первые фазы пищеварения. Затем картина меняется и реакция внутри пищеварительных вакуолей становится слабощелочной. В этих условиях протекают дальнейшие этапы внутриклеточного пищеварения. Кислая фаза обычно короче щелочной; она длится примерно 1/6-1/4 всей продолжительности пребывания пищеварительной вакуоли в теле инфузории. Однако соотношение кислой и щелочной фаз может изменяться в довольно широких пределах в зависимости от характера пищи.
Путь пищеварительной вакуоли в эндоплазме заканчивается тем, что она подходит к поверхности тела и через пелликулу ее содержимое, состоящее из жидкости и непереваренных остатков пищи, выбрасывается наружу — происходит дефекация. Этот процесс, в отличие от амеб, у которых дефекация может происходить где угодно, у туфельки, как и у других инфузорий, строго приурочен к определенному участку тела, расположенному на брюшной стороне (абдоминальной условно называют поверхность животного, на которой помещается околоротовая полость), примерно посередине между перистомом и задним концом тела.
Таким образом, внутриклеточное пищеварение представляет собой сложный процесс, состоящий из нескольких последовательных фаз.
Расчеты показывают, что примерно за 30-45 минут из туфельки через сократительные вакуоли выделяется объем жидкости, равный объему тела инфузории. Таким образом, благодаря деятельности сократительных вакуолей через тело инфузории осуществляется непрерывный ток воды, поступающей извне через ротовое отверстие (вместе с пищеварительными вакуолями), а также осмотически непосредственно через пелликулу. . Сократительные вакуоли играют важную роль в регуляции тока воды через тело инфузории, в регуляции осмотического давления. Этот процесс здесь протекает в принципе так же, как и у амеб, только строение сократительной вакуоли значительно сложнее.
В течение многих лет среди ученых, изучающих простейшие, шел спор по поводу того, существуют ли в цитоплазме какие-либо структуры, связанные с появлением сократительной вакуоли, или она каждый раз образуется заново. На живой инфузории не наблюдается никаких особых структур, которые предшествовали бы ее образованию. После сокращения вакуоли – систолы, в цитоплазме на месте бывшей вакуоли совершенно не видно никаких структур. Затем вновь появляются прозрачный пузырь или приводящие каналы, которые начинают увеличиваться в размерах. Однако никакой связи между вновь возникшей вакуолью и существовавшей ранее не обнаружено. Создается впечатление, что между последовательными циклами сократительной вакуоли нет преемственности, и всякая новая сократительная вакуоль образуется заново в цитоплазме. Однако специальные методы исследования показали, что на самом деле это не так. Применение электронной микроскопии, дающей очень большое увеличение (до 100 тыс. раз), убедительно показало, что у инфузории в области образования сократительных вакуолей имеется особо дифференцированная цитоплазма, состоящая из переплетения тончайшие трубочки. Таким образом, оказалось, что сократительная вакуоль возникает в цитоплазме не на пустом месте, а на основе предшествующего особого органоида клетки, функцией которого является образование сократительной вакуоли.
Как и все простейшие, инфузории имеют клеточное ядро. Однако по строению ядерного аппарата инфузории резко отличаются от всех других групп простейших.
Ядерный аппарат инфузорий характеризуется двойственностью. Это означает, что у инфузорий есть два разных типа ядер — большие ядра, или макронуклеусы, и малые ядра, или микроядра. Посмотрим, какое строение имеет ядерный аппарат инфузории-туфельки (рис. 85).
В центре тела инфузории (на уровне перистома) расположено крупное массивное овоидное или бобовидное ядро. Это макронуклеус. В непосредственной близости от него находится второе ядро, во много раз меньшее по размерам, обычно довольно тесно примыкающее к макронуклеусу. Это микронуклеус. Разница между этими двумя ядрами не ограничивается размерами, она более существенна, глубоко затрагивая их строение.
Макронуклеус, по сравнению с микронуклеусом, значительно богаче особым ядерным веществом (хроматином, или, точнее, дезоксирибонуклеиновой кислотой, сокращенно ДНК), входящим в состав хромосом.
Исследования последних лет показали, что макронуклеус имеет в несколько десятков (а у некоторых инфузорий даже сотни) раз больше хромосом, чем микроядра. Макронуклеус представляет собой весьма своеобразный тип полихромосомных (полиплоидных) ядер. Таким образом, различие между микро- и макронуклеусами сказывается на их хромосомном составе, от чего зависит большее или меньшее богатство их ядерного вещества — хроматина.
Один из наиболее распространенных видов инфузорий — туфельки (Paramecium caudatum) — имеется один макронуклеус (сокращенно Ма) и один микронуклеус (сокращенно Ми). Такое строение ядерного аппарата характерно для многих инфузорий. У других может быть несколько Ма и Ми. Но характерной особенностью всех инфузорий является дифференциация ядер на две качественно различные группы, на Ма и Ми, или, другими словами, явление ядерного дуализма.
Как размножаются инфузории? Обратимся в качестве примера снова к инфузории-туфельке. Если посадить единичный экземпляр туфельки в небольшой сосуд (микроаквариум), то через сутки инфузорий будет две, а часто и четыре. Как это произошло? После определенного периода активного плавания и питания инфузория несколько вытягивается в длину. Затем ровно посередине тела появляется все более углубляющаяся поперечная перетяжка (рис. 9).0). В конце концов инфузории как бы сплетаются пополам и из одной особи получаются две, вначале несколько меньших размеров, чем мать. Весь процесс деления занимает около часа при комнатной температуре. Изучение внутренних процессов показывает, что еще до появления поперечной перетяжки начинается процесс деления ядерного аппарата. Первым делится Ми, и только после этого Ма. Мы не будем здесь останавливаться на подробном рассмотрении процессов деления ядер и лишь отметим, что Ми делится митозом, тогда как деление Ма по внешнему виду напоминает прямое деление ядер — амитоз. Этот процесс бесполого размножения инфузорий, как мы видим, сходен с бесполым размножением амеб и жгутиковых. Напротив, инфузории в процессе бесполого размножения всегда делятся поперек, а у жгутиковых плоскость деления параллельна продольной оси тела.
При делении происходит глубокая внутренняя перестройка тела инфузории. Образуются два новых перистома, два зева и два ротовых отверстия. К этому же времени приурочено деление базальных ядер ресничек, благодаря которому образуются новые реснички. Если бы количество ресничек не увеличивалось при размножении, то в результате каждого деления дочерние особи получали бы примерно половину количества ресничек материнской, что приводило бы к полному «облысению» инфузории. На самом деле этого не происходит.
Время от времени у большинства инфузорий, в том числе и у туфель, возникает особая и крайне своеобразная форма полового процесса, которая называется конъюгацией. Мы не будем здесь подробно разбирать все те сложные ядерные изменения, которые сопровождают этот процесс, а лишь укажем на самое главное. Конъюгация протекает следующим образом (рис. 91). Две инфузории сближаются, тесно прикрепляются друг к другу брюшными сторонами и в таком виде плавают вместе довольно продолжительное время (туфелькой около 12 часов при комнатной температуре). Затем конъюгаты расходятся. Что происходит в организме инфузории при конъюгации? Суть этих процессов заключается в следующем (рис. 91). Большое ядро (макронуклеус) разрушается и постепенно растворяется в цитоплазме. Микроядра сначала делятся, часть образовавшихся в результате деления ядер разрушается (см. рис. 91). Каждая из конъюгантов сохраняет два ядра. Одно из этих ядер остается на месте у особи, у которой оно образовалось (стационарное ядро), а другое активно движется к партнеру по конъюгации (мигрирующее ядро) и сливается с его неподвижным ядром. Таким образом, в каждой из конъюгантов на этой стадии имеется по одному ядру, образовавшемуся в результате слияния стационарного и мигрирующего ядер. его сложное ядро получает название синкарион. Образование синкариона есть не что иное, как процесс оплодотворения. А у многоклеточных организмов существенным моментом оплодотворения является слияние ядер половых клеток. У инфузорий половые клетки не образуются, имеются только половые ядра, которые сливаются между собой. Таким образом, происходит взаимное перекрестное оплодотворение.
Вскоре после образования синкариона конъюганты расходятся. По строению своего ядерного аппарата на этой стадии они еще очень существенно отличаются от обычных так называемых нейтральных (неконъюгирующих) инфузорий, так как имеют только по одному ядру. В дальнейшем за счет синкариона восстанавливается нормальный ядерный аппарат. Синкарион делится (один или несколько раз). Часть продуктов этого деления путем сложных превращений, связанных с увеличением числа хромосом и обогащением хроматином, превращается в макронуклеусы. Другие сохраняют структуру, характерную для микроядер. Таким путем восстанавливается характерный и типичный для инфузорий ядерный аппарат, после чего инфузории переходят к бесполому размножению делением.
Таким образом, процесс конъюгации включает в себя два существенных биологических момента: оплодотворение и восстановление нового макронуклеуса за счет синкариона.
Каково биологическое значение конъюгации, какую роль она играет в жизни инфузорий? Мы не можем назвать это размножением, ибо это не увеличивает количество особей. Вышеуказанные вопросы послужили материалом для многочисленных экспериментальных исследований, проведенных во многих странах. Основной результат этих исследований состоит в следующем. Во-первых, конъюгация, как и всякий другой половой процесс, при котором в одном организме сочетаются два наследственных начала (отцовский и материнский), приводит к увеличению наследственной изменчивости, наследственного разнообразия. Увеличение наследственной изменчивости увеличивает приспособительные возможности организма к условиям внешней среды. Второй биологически важной стороной конъюгации является развитие нового макронуклеуса за счет продуктов деления синкариона и одновременно разрушение старого. Экспериментальные данные показывают, что именно макронуклеус играет чрезвычайно важную роль в жизни инфузорий. Он контролирует все основные жизненные процессы и определяет важнейший из них — образование (синтез) белка, составляющего основную массу протоплазмы живой клетки. При длительном бесполом размножении делением происходит своеобразный процесс «старения» макронуклеуса, а вместе с тем и всей клетки: снижается активность метаболического процесса, снижается скорость деления. После конъюгации (при которой, как мы видели, старый макронуклеус разрушается) уровень метаболизма и скорость деления восстанавливаются. Поскольку при конъюгации происходит процесс оплодотворения, который у большинства других организмов связан с размножением и появлением нового поколения, то у инфузорий особь, образующаяся после конъюгации, также может рассматриваться как новое половое поколение, возникающее здесь как бы вследствие «омоложение» старого.
На примере инфузории туфельки мы познакомились с типичным представителем большого класса инфузорий. Однако для этого класса характерно чрезвычайное разнообразие видов, как по строению, так и по образу жизни. Рассмотрим подробнее некоторые из наиболее характерных и интересных форм.
У инфузорий туфельки для ресничек равномерно покрывают всю поверхность тела. Это характерная черта строения (голотриха). Для многих инфузорий характерен различный характер развития ресничного покрова. Дело в том, что реснички инфузорий способны соединяться между собой, образуя более сложные комплексы. Например, часто наблюдается, что реснички, расположенные в один или два ряда близко друг к другу, соединяются (слипаются) между собой, образуя пластинку, которая, как и реснички, способна биться. Такие пластинчатые сократительные образования называются мембранеллами (если они короткие) или мембранами (если они длинные). В других случаях реснички соединены между собой, располагаются в плотный пучок. Эти образования — циррусы — напоминают кисточку, отдельные волоски которой слиплись. Для многих инфузорий характерны разного рода сложные ресничные образования. Очень часто ресницы развиваются неравномерно, а только на некоторых участках тела.
ИНФУЗОРИЯ ТРУБОЧНАЯ (СТЕНТОР ПОЛИМОРФНАЯ)
В пресных водах очень распространены виды крупных красивых инфузорий, родственных разновидности трубачей (Стентор). Это название вполне соответствует форме тела этих животных, которое действительно напоминает трубу (рис. 92), широко открытую с одного конца. При первом же знакомстве с живыми трубачами можно заметить одну особенность, не свойственную туфельке. При малейшем раздражении, в том числе и механическом (например, постукивание трубами карандаша по стакану, где есть капля воды), их тело резко и очень быстро (за доли секунды) сокращается, принимая почти правильную шаровидную форму. Затем довольно медленно (время измеряется секундами) трубач выпрямляется, принимая характерную для него форму. Эта способность трубача быстро сокращаться обусловлена наличием особых мышечных волокон, расположенных вдоль тела и в эктоплазме. Таким образом, в одноклеточном организме может развиваться мышечная система.
В роду трубачей есть виды, некоторые из которых отличаются довольно яркой окраской. Очень часто встречается в пресных водах голубой трубач (Stentor coeruleus), который имеет ярко-голубой цвет. Такой окрас трубача обусловлен тем, что в его эктоплазме расположены мельчайшие зерна синего пигмента.
Другой вид трубача (Stentor polymorphus) часто окрашен в зеленый цвет. Причина такого окрашивания совсем в другом. Зеленый цвет обусловлен тем, что в эндоплазме инфузории живут и размножаются мелкие одноклеточные зеленые водоросли, придающие телу трубача характерную окраску. Stentor polymorphus — типичный пример взаимовыгодного сожительства — симбиоза. Трубач и водоросли находятся во взаимно-симбиотических отношениях: трубач защищает живущие в его теле водоросли и снабжает их углекислым газом, образующимся при дыхании; с другой стороны, водоросли обеспечивают трубача кислородом, который выделяется в процессе фотосинтеза. По-видимому, часть водорослей переваривается инфузориями, являясь пищей для трубача.
Трубачи медленно плавают в воде широким концом вперед. Но они также могут временно прикрепляться к субстрату задним узким концом тела, на котором образована небольшая присоска.
В теле трубача можно различить расширяющийся сзади вперед туловищный отдел и расположенное почти перпендикулярно ему широкое околоротовое (перистомальное) поле. Это поле напоминает асимметричную плоскую воронку, на одном краю которой находится углубление — зев, ведущий в эндоплазму инфузории. Тело трубача покрыто продольными рядами коротких ресничек. По краю перистомального поля по окружности располагается мощно развитая периоральная (адоральная) зона мембранелл (рис. 9).2). Эта зона состоит из большого количества отдельных реснитчатых пластинок, каждая из которых, в свою очередь, состоит из множества прилегающих друг к другу, расположенных в двух близко расположенных рядах ресничек.
В области ротового отверстия периоральные мембраны заворачиваются в направлении глотки, образуя левозакрученную спираль. Поток воды, вызванный колебаниями периоральной мембраны, направлен к ротовому отверстию (в глубину воронки, образованной передним концом тела). Вместе с водой в глотку попадают и взвешенные в воде частицы пищи. Продукты питания трубача более разнообразны, чем у туфельки. Наряду с бактериями поедает мелких простейших (например, жгутиковых), одноклеточных водорослей и др.
У трубача хорошо развита сократительная вакуоль, имеющая своеобразное строение. Центральный резервуар расположен в передней трети тела, немного ниже ротового отверстия. От него отходят два длинных ведущих канала. Один из них идет от резервуара к заднему концу тела, второй располагается в перистомальном поле параллельно периоральной зоне мембранеллы.
Инфузория-трубач — излюбленный объект экспериментальных исследований по регенерации. Многочисленные эксперименты доказали высокую регенеративную способность трубачей. Тонким скальпелем инфузорию можно разрезать на множество частей, и каждая из них через короткое время (несколько часов, иногда сутки и более) превратится в пропорционально сложенного, но небольшого трубача, который затем, в результате энергичного питания, достигает типичных для этого вида размеров. Для завершения восстановительных процессов регенерирующий фрагмент должен иметь хотя бы один сегмент бусообразного макронуклеуса.
У трубача, как мы видели, разные реснички: с одной стороны короткие, покрывающие все тело, а с другой периоральная зона мембранелл. По этому характерному признаку состав отряда инфузорий, к которому относится трубач, получил название пестрых инфузорий (Heterotricha).
ИНФУЗОРИЯ БУРСАРИЯ (BURSARIA TRUNCATELLA)
Второй интересный представитель инфузорий часто встречается в пресных водах. бурсария (Bursaria truncatella, рис. 93). Это гигант среди инфузорий: его размер может достигать 2 мм, самые распространенные значения 0,5-1,0 мм. Бурсария хорошо видна невооруженным глазом. В соответствии со своим названием бурсария имеет форму мешка, широко открытого на переднем конце (bursa — латинское слово, в переводе на русский язык означает «кошелек», «мешок») и несколько расширенного на заднем конце. Все тело инфузории покрыто продольно идущими рядами коротких ресничек. Их биение вызывает довольно медленное движение животного вперед. Бурсария плавает, как бы «переваливаясь» из стороны в сторону.
От переднего конца вглубь тела (около 2/3 его длины) выступает околоротовое углубление — перистое. На вентральной стороне сообщается с внешней средой узкой щелью; с дорсальной стороны полость перистома не сообщается с внешней средой. Если посмотреть на поперечное сечение верхней трети тела бурсарии (рис. 93, Б), то можно увидеть, что перистомальная полость занимает большую часть тела, а цитоплазма окружает ее в виде ободка.
На переднем конце тела слева от бурсарий берет начало очень мощно развитая зона периоральной (адоральной) мембранеллы (рис. 93, 4). Опускается в глубь перистомальной полости, поворачивая влево. Адоральная зона достигает самой глубокой части перистома. Кроме периоральной мембранеллы, других реснитчатых образований в перистомальной полости нет, за исключением ресничной полоски, идущей вдоль брюшной стороны перистомальной полости (рис. 9).3, 10). На внутренней задней стенке перистомальной полости почти на всем ее протяжении проходит узкая щель (рис. 93, 7), края которой обычно тесно прилегают друг к другу. Это ротовая щель. Его края раздвигаются только в момент еды.
Бурсарии не имеют узкой пищевой специализации, но в основном являются хищниками. Двигаясь вперед, они натыкаются на различных мелких животных. Благодаря работе мембранелл периоральной зоны добыча с силой втягивается в обширную перистомальную полость, откуда уже не может выплыть. Пищевые объекты прижимаются к дорсальной стенке перистомальной полости и проникают в эндоплазму через расширяющуюся ротовую щель. Бурсарии очень прожорливы, они могут проглатывать довольно крупные предметы: например, инфузории туфельки — их излюбленная еда. Бурсария способна проглотить 6-7 ботинок подряд. В результате в эндоплазме бурсарии образуются очень крупные пищеварительные вакуоли.
Ядерный аппарат бурсарии довольно сложен. Они имеют один длинный колбасный макронуклеус и большое (примерно до 30) количество мелких микроядер, беспорядочно разбросанных в эндоплазме инфузории.
Bursaria — один из немногих видов пресноводных инфузорий, у которых отсутствует сократительная вакуоль. Как осуществляется осморегуляция у этой крупной инфузории, до сих пор не совсем ясно. Под эктоплазмой бурсарии на разных участках тела можно наблюдать различной формы и размеров жидкостные пузырьки — вакуоли, изменяющие свой объем. По-видимому, эти вакуоли неправильной формы по своей функции соответствуют сократительным вакуолям других инфузорий.
Интересно наблюдать за последовательными стадиями бесполого размножения бурсарии. На начальных стадиях деления происходит полная редукция всей полости перистома и периоральной зоны мембранеллы (рис. 94). Сохраняется только наружный ресничный покров. Инфузория принимает форму яйца. После этого корпус шнуруется поперечным желобком на две половины. У каждой из образовавшихся дочерних инфузорий путем довольно сложных преобразований развивается типичный перистом и периоральная зона мембранелл. Весь процесс деления бурсарии протекает быстро и занимает чуть больше часа.
Очень легко наблюдать у бурсариев еще один важный жизненный процесс, начало которого связано с неблагоприятными для инфузории условиями, — процесс цистообразования (энцистации). Это явление характерно, например, для амебы. Но оказывается, такие сложноорганизованные простейшие, как инфузории, способны переходить в неактивное состояние. Если культуру, в которой обитают бурсарии, вовремя не подкормить или сильно охладить, то через несколько часов начнется массовое инцистирование. Этот процесс протекает следующим образом. Бурсариды, как и до деления, теряют перистом и периоральную зону мембранелл. Затем они становятся полностью сферическими, после чего выделяют двойную оболочку характерной формы (рис. 94, Д).
В состоянии кисты бурсарии могут находиться месяцами. При наступлении благоприятных условий оболочка кисты прорывается, бурсарии выходят из нее, развиваются перистым и переходят к активной жизни.
STYLONICHIA MYTILUS
Весьма сложный и разнообразно дифференцированный ресничный аппарат имеют инфузории, относящиеся к отряду желудочно-кишечных (Hypotricha), многочисленные виды которых обитают как в пресноводных, так и в морских водах. частыми представителями этой интересной группы можно назвать стилонихия (Stylonichia mytilus). Это довольно крупная инфузория (длиной до 0,3 мм), обитающая на дне пресноводных водоемов, на водной растительности (рис. 95). В отличие от туфельки, трубача и бурсарии у стилонихии отсутствует сплошной ресничный покров, и весь ресничный аппарат представлен ограниченным числом строго определенных ресничных образований.
Тело стилонихии (как и большинства других желудочных инфузорий) сильно уплощено в дорсально-брюшном направлении, при этом хорошо различимы дорсальная и вентральная стороны, передний и задний концы. Тело слегка расширено спереди, сужено сзади. При осмотре животного с вентральной стороны хорошо видно, что в передней трети слева имеются сложно расположенные пинн и ротовое отверстие.
На спинной стороне довольно редко расположены реснички, которые не способны биться. Скорее их можно назвать тонкими эластичными щетинками. Они неподвижны и не имеют никакого отношения к функции движения. Этим ресничкам обычно приписывают осязательную, чувствительную функцию.
Все реснитчатые образования, связанные с передвижением и захватом пищи, сосредоточены на брюшной стороне животного (рис. 95). Имеется небольшое количество толстых пальцевидных образований, расположенных несколькими группами. Это брюшные циррусы. Каждая из них представляет собой сложное ресничное образование, результат тесного соединения (слипания) многих десятков отдельных ресничек. Таким образом, циррусы представляют собой как бы кисти, отдельные волоски которых тесно сплетены и соединены друг с другом.
С помощью цирроз животное передвигается довольно быстро, «бегает» по субстрату. Помимо «ползания» и «бега» по субстрату, стилонихии способны производить довольно резкие и сильные прыжки, сразу же отрываясь от субстрата. Эти резкие движения осуществляются с помощью двух мощных хвостовых циррусов (рис. 95), не принимающих участия в обычном «ползании».
По краю тела справа и слева два ряда краевых (маргинальных) циррусов. От правого края животного они проходят вдоль всего тела, от левого края достигают только околоротовой области. Эти ресничные образования служат для поступательного движения животного, когда оно отрывается от субстрата и свободно плавает в воде.
Мы видим, таким образом, что разнообразный и специализированный ресничный аппарат стилонихии позволяет ей совершать весьма разнообразные движения, в отличие, например, от простого скольжения в воде, как туфелька или трубач.
Цилиарный аппарат, связанный с силовой функцией, также сложен. Мы уже видели, что околоротовая полость (перистом), на дне которой находится ротовое отверстие, ведущее в глотку, расположена в передней половине животного слева. На левом крае, начиная с переднего конца тела, имеется сильно развитая зона периоральной (адоральной) мембранеллы. Своим биением они направляют поток воды в сторону ротового отверстия. Кроме того, в области перистомальной полости имеются еще три сократительные перепонки (мембраны), уходящие своими внутренними концами в глотку, и ряд особых периоральных ресничек (рис. 9).5). Весь этот сложный аппарат служит для улавливания и направления пищи в ротовое отверстие.
Стилонихия — одна из самых простых с очень широким набором продуктов питания. Его по праву можно назвать всеядным животным. Он может питать, как обувь, бактерии. Его пищевые объекты включают жгутиконосцев, одноклеточные водоросли (часто диатомовые водоросли). Наконец, стилонихия может быть и хищником, нападающим на другие, более мелкие виды инфузорий и пожирающим их.
Stylonychia имеет сократительную вакуоль. Он состоит из центрального резервуара, расположенного в левом заднем конце перистома, и одного заднего аддукционного канала.
Ядерный аппарат, как всегда у инфузорий, состоит из макронуклеуса и микронуклеуса.
Макронуклеус состоит из двух половинок, соединенных тонкой перетяжкой; микроядра два, они расположены непосредственно возле обеих половинок Ма.
Стилонихия, отчасти бурсария, трубач – это все инфузории с широким набором пищевых объектов. Для большинства инфузорий характерна способность поглощать различную пищу. Однако среди них можно найти и такие виды, которые строго специализированы по отношению к характеру питания.
ИНФУЗОРИИ ХИЩНИКОВ
Среди инфузорий есть хищники, которые очень «придирчивы» к своей добыче. Инфузория является хорошим примером. дидиния (Didinium nasutum). Didinius – относительно небольшая инфузория, средней длиной около 0,1-0,15 мм. Передний конец вытянут в виде хоботка, на конце которого размещено ротовое отверстие. Ресничный аппарат представлен двумя венчиками ресничек (рис. 96). Дидиниус быстро плавает в воде, часто меняя направление. Предпочтительной пищей дидинии являются инфузории-туфельки. В этом случае хищник оказывается меньше своей жертвы. Дидиний вторгается в добычу своим хоботом, а затем, постепенно расширяя ротовое отверстие все больше и больше, заглатывает туфельку целиком! В хоботке имеется особый, так называемый стержневой, аппарат. Он состоит из ряда прочных эластичных стержней, расположенных в цитоплазме по периферии хоботка. Считается, что этот аппарат увеличивает прочность стенок хоботка, который не ломается при заглатывании такой огромной добычи, как башмак по сравнению с дидиниумом. Didinius является крайним примером хищничества простейших. Если сравнить заглатывание дидинием своей добычи — обуви — с хищничеством у высших животных, то аналогичные примеры найти трудно.
Дидиний, проглотивший Парамецию, конечно, сильно опухает. Процесс переваривания очень быстрый, при комнатной температуре он занимает всего около двух часов. Затем непереваренные останки выбрасываются, и Дидиний начинает охотиться за очередной жертвой. Специальные исследования установили, что дневной «рацион» дидиниума составляет 12 туфель — поистине колоссальный аппетит! Следует иметь в виду, что в промежутках между очередными «охотами» дидинию иногда делят. При недостатке питания дидинии очень легко инцистируются и так же легко вновь выходят из цист.
РАСТИТЕЛЬНЫЕ ИНФУЗОРИИ
Значительно реже, чем хищничество, среди инфузорий встречается «чистое вегетарианство» — употребление в пищу исключительно растительной пищи. Представители рода пассула (Nassula). Объектом их питания являются нитчатые сине-зеленые водоросли (рис. 97).
Они проникают в эндоплазму через рот, расположенный сбоку, а затем закручивают инфузорию в тугую спираль, которая постепенно переваривается. Пигменты водорослей частично проникают в цитоплазму инфузории и окрашивают ее в яркий темно-зеленый цвет.
SOUVIKA (VORTICELLA NEBULIFERA)
Интересную и довольно большую группу инфузорий по числу видов составляют сидячие формы, прикрепленные к субстрату, образующие отряд кольцевых ресничек (Peritricha). Широко распространенными представителями этой группы являются сувой (виды рода Vorticella).
Сувойки напоминают изящный цветок, похожий на колокольчик или ландыш, сидящий на длинном стебле, который своим концом прикреплен к субстрату. Сувой большую часть жизни проводит в прикрепленном к субстрату состоянии.
Рассмотрим строение тела инфузории. У разных видов их размеры варьируют в довольно широких пределах (примерно до 150 мкм). Ротовой диск (рис. 98) расположен на вытянутой передней части тела, полностью лишенной ресничек. Ресничный аппарат располагается только по краю ротового (перистомального) диска (рис. 98) в специальной бороздке, снаружи которой образуется валик (перистомальная губа). По краю валика расположены три реснитчатые перепонки, две из которых расположены вертикально, одна (наружная) — горизонтально. Они образуют чуть более одного полного витка спирали. Эти перепонки находятся в постоянном мерцающем движении, направляя поток воды к ротовому отверстию. Ротовой аппарат начинается довольно глубоко воронкой у края перистомального поля (рис. 9).8), в глубине которого расположено ротовое отверстие, ведущее в короткую глотку. Сувойки, как и туфельки, питаются бактериями. Их ротовое отверстие постоянно открыто, и ко рту идет непрерывный поток воды.
Рядом с ротовым отверстием расположена одна сократительная вакуоль без аддукционных каналов. Макронуклеус имеет лентовидную или колбасовидную форму; к нему вплотную прилегает одиночное мелкое микроядро.
Сувойка умеет резко укорачивать стебель, который за доли секунды скручивается в штопор. При этом тело инфузории сокращается: перистомальный диск и мембраны втягиваются внутрь и весь передний конец закрывается.
Естественно возникает вопрос: поскольку сувойи прикреплены к субстрату, то каким образом осуществляется их расселение по водоему? Это происходит за счет образования свободно плавающей бродячей стадии. На заднем конце тела инфузории появляется венчик ресничек (рис. 99). При этом перистомальный диск втягивается внутрь и инфузория отделяется от ножки. Получившийся бродяга способен плавать несколько часов. Затем события разыгрываются в обратном порядке: инфузория задним концом прикрепляется к субстрату, вырастает стебель, редуцируется задний венчик ресничек, на переднем конце расширяется перистомальный диск, начинают работать адоральные мембраны. Формирование бродяг у сувоя часто связано с процессом бесполого размножения. Инфузория на стебле делится, и одна из дочерних особей (а иногда и обе) становится залетной и уплывает.
При неблагоприятных условиях многие виды сувуков способны к инцистированию.
Среди малоподвижных инфузорий, принадлежащих к отряду инфузорий, лишь сравнительно немногие виды, как и рассмотренные выше сувои, являются одиночными живыми формами. Большинство видов, связанных с этим, являются колониальными организмами.
Колониальность обычно возникает в результате неполного бесполого или вегетативного размножения. Образовавшиеся в результате размножения особи в большей или меньшей степени сохраняют связь друг с другом и все вместе образуют органическую индивидуальность более высокого порядка, объединяющую большое количество отдельных особей, которая называется колонией (мы уже встречал образцы колониальных организмов в класс жгутиковых .
Колонии реснитчатых инфузорий образуются в результате того, что отделившиеся особи не превращаются в бродяг, а поддерживают связь друг с другом при помощи стеблей (рис. 100). При этом главный ствол колонии, как и ее первые ветви, не может быть отнесен ни к одной из особей, а принадлежит всей колонии в целом. Иногда колония состоит лишь из небольшого числа особей, тогда как у других видов инфузорий количество отдельных особей колонии может достигать нескольких сотен. Однако рост любой колонии не безграничен. При достижении размеров, характерных для этого вида, колония перестает расти и особи, образовавшиеся в результате деления, развивают венчик из ресничек, становятся залетными и уплывают, давая начало новым колониям.
Колонии инфузорий бывают двух типов. У одних стебель колонии нередуцируемый: при раздражении сокращаются лишь отдельные особи колонии, втягиваясь пером, вся колония в целом не претерпевает изменений (к этому типу колоний относятся, например, роды Epistylis, Opercularia). У других (например, рода Carchesium) стебель всей колонии способен сокращаться, так как цитоплазма проходит через все ветви и таким образом связывает между собой всех особей колонии. Когда такие колонии раздражены, они полностью сокращаются. Вся колония в этом случае реагирует как единое целое, как органическая индивидуальность.
Среди всех колониальных реснитчатых инфузорий особый интерес представляет, пожалуй, зоотамный (Zoothamnium arbuscula). Колонии этой инфузории отличаются особой правильностью строения. Кроме того, здесь намечается интересное биологическое явление полиморфизма внутри колонии.
Колония зоотамнии выглядит как зонтик. На одном, главном, стебле колонии имеются второстепенные ветви (рис. 101). Размер взрослой колонии 2-3 мм, так что они хорошо видны невооруженным глазом. Живут зоотамны в небольших водоемах с чистой водой. Их колонии обычно встречаются на подводных растениях, чаще всего на элодеях (водяная чума).
Стебли колонии зоотамний сократительные, так как сократительная цитоплазма проходит через все ветви колонии, за исключением базальной части основного стебля. При сокращении, которое происходит очень быстро и резко, вся колония собирается в комок.
Зоотамния характеризуется строго правильным расположением ветвей. К субстрату прикрепляется один главный стебель. От его верхней части в плоскости, перпендикулярной стеблю, отходят девять основных ветвей колонии, строго равномерно расположенных друг относительно друга (рис. 102, 6). От этих ветвей отходят второстепенные ветви, на которых сидят отдельные особи колонии. В каждой вторичной ветви может быть до 50 инфузорий. Общая численность особей в колонии достигает 2-3 тысяч.
Большинство особей колонии по своему строению напоминают мелкие одиночные сувои, размером 40-60 мкм. Но кроме мелких особей, называемых микрозоидами, на взрослых колониях, примерно посередине основных ветвей, развиваются особи совсем другого вида и размера (рис. 102, 5). Это крупные шаровидные особи диаметром 200-250 мкм, превышающие объем микрозоида в сто и более раз. Крупных особей называют макрозоидами.
По своему строению они существенно отличаются от мелких особей колонии. Их перистом не выражен: он втянут внутрь и не функционирует. С самого начала своего развития макрозоид из микрозоида перестает самостоятельно принимать пищу. У него отсутствуют пищеварительные вакуоли. Рост макрозоида, по-видимому, осуществляется за счет поступления веществ через цитоплазматические мостики, соединяющие всех особей колонии. В той части тела макрозоида, которой он прикрепляется к стеблю, имеется скопление особых зерен (гранул), которые, как мы увидим, в дальнейшей его судьбе играют существенную роль. Что это за крупные шаровидные макрозоиды, какова их биологическая роль в жизни колонии зоотамний? Наблюдения показывают, что макрозоиды — это будущие бродяги, из которых развиваются новые колонии. Достигнув предельных размеров, макрозоид развивает венчик ресничек, отделяется от колонии и уплывает. При этом несколько меняется его форма, из сферической она становится конической. Через некоторое время бродяга всегда прикрепляется к субстрату той стороной, на которой находится зерно. Сразу начинается формирование и рост стебля, и гранулы, локализующиеся на заднем конце трамплина, расходуются на строительство стебля. По мере роста стебля зернистость исчезает. После того, как стебель достигает конечной длины, характерной для зоотамнии, начинается ряд быстро следующих делений, приводящих к образованию колонии. Эти деления выполняются в строго определенной последовательности (рис. 102).
Мы не будем останавливаться на деталях этого процесса. Обратим внимание только на следующее интересное явление. При первых делениях зоотамнии залетной в период развития колонии у формирующихся особей перья и рот не функционируют. Питание начинается позже, когда молодая колония уже состоит из 12-16 особей. Таким образом, все первые этапы развития семьи осуществляются исключительно за счет тех резервов, которые образовались в организме макрозоида в период его роста и развития на материнской семье. Существует неоспоримое сходство между развитием зоотамнии залетной и развитием яйца у многоклеточных животных. Это сходство выражается в том, что развитие в обоих случаях осуществляется за счет ранее накопленных резервов, без восприятия пищи из внешней среды.
При изучении сидячих круговых инфузорий возникает вопрос: как у них осуществляется характерная для инфузорий форма полового процесса — конъюгация? Оказывается, в связи с малоподвижным образом жизни она претерпевает значительные изменения. К началу полового процесса на колонии образуются особые, очень мелкие бродяги. Активно передвигаясь с помощью венчика ресничек, они некоторое время ползают по колонии, а затем вступают в конъюгацию с крупными нормальными малоподвижными особями колонии. Таким образом, здесь происходит дифференциация конъюгантов на две группы особей: мелкие, подвижные (микроконъюганты) и более крупные, неподвижные (макроконъюганты). Эта дифференциация конъюгантов на две категории, одна из которых (микроконъюганты) подвижна, была необходимой адаптацией к малоподвижному образу жизни. Без этого нормальное протекание полового процесса (конъюгации) заведомо не могло быть обеспечено.
Инфузории сосущие (SUCTORIA)
Весьма своеобразную группу по способу питания представляют сосущие инфузории (Suctoria). Эти организмы, как и сувои и другие инфузории, сидячие. Число видов, принадлежащих к этому отряду, измеряется несколькими десятками. Форма тела сосущих инфузорий очень разнообразна. Некоторые их характерные виды показаны на рис. 103. Одни сидят на субстрате на более или менее длинных стеблях, другие не имеют стеблей, у некоторых довольно сильно ветвится тело и т. д. Однако, несмотря на разнообразие форм, все сосущие инфузории характеризуются следующими двумя особенностями:
1) полное отсутствие (у взрослых форм) ресничного аппарата,
2) наличие особых придатков — щупалец, служащих для высасывания добычи.
Количество щупалец неодинаково у разных видов сосущих инфузорий. Они часто собираются в группы. При большом увеличении микроскопа видно, что на конце щупальца имеется небольшое булавовидное утолщение.
Как работают щупальца? На этот вопрос легко ответить, понаблюдав некоторое время за сосущими инфузориями. Если какое-либо мелкое простейшее (жгутиконосец, инфузория) коснется щупальца суктории, оно моментально прилипнет к нему. Все попытки жертвы оторваться обычно тщетны. Если продолжать наблюдать за прилипшей к щупальцам жертвой, то можно заметить, что она постепенно начинает уменьшаться в размерах. Его содержимое постепенно «закачивается» через щупальца в эндоплазму сосущей инфузории до тех пор, пока от жертвы не останется только одна пелликула, которую отбрасывают. Таким образом, щупальца сосущих инфузорий совершенно своеобразны, больше нигде в животном мире не встречаются органы для захвата и одновременно сосания пищи (рис. 103).
Сосущие инфузории — неподвижные хищники, которые не гонятся за добычей, а мгновенно ее ловят, стоит только неосторожной добыче прикоснуться к ним самой.
Почему мы относим эти своеобразные организмы к классу инфузорий? На первый взгляд, они не имеют к ним никакого отношения. О принадлежности суктории к инфузориям свидетельствуют следующие факты. Во-первых, у них типичный для инфузорий ядерный аппарат, состоящий из макронуклеуса и микронуклеуса. Во-вторых, при размножении у них развиваются реснички, отсутствующие у «взрослых» особей. Бесполое размножение и одновременно расселение сосущих инфузорий осуществляется путем образования бродяг, снабженных несколькими кольцевидными венчиками ресничек. Формирование залетников в суккториях может происходить по-разному. Иногда они образуются в результате не совсем равномерного деления (почкования), при котором каждая отделяющаяся наружу почка получает участок макронуклеуса и один микронуклеус (рис. 104, Л). На одной матке может образоваться сразу несколько дочерних почек (рис. 104, 5). У других видов (рис. 104, Г, Д) наблюдается весьма своеобразный способ «внутреннего почкования». При этом внутри тела материнского суктория образуется полость, в которой формируется заблудшая почка. Он выходит через специальные отверстия, через которые «продавливается» с некоторым трудом.
Это развитие зародыша внутри тела матери, а затем акт родов представляет собой интересную аналогию простейшего с тем, что происходит у высших многоклеточных организмов.
На предыдущих страницах были рассмотрены несколько типичных свободноживущих представителей класса инфузорий, по-разному приспособленных к разным условиям среды. Интересно подойти к вопросу приспособления инфузорий к условиям жизни и, с другой стороны, посмотреть, каковы характерные общие черты инфузорий, обитающих в определенных, резко определенных условиях среды.
В качестве примера возьмем две очень резко разные среды обитания: жизнь в составе планктона и жизнь на дне в толще песка.
ПЛАНКТОННЫЕ ИНФУЗОРИИ
Достаточно большое количество видов инфузорий встречается как в морском, так и в пресноводном планктоне.
Особенности приспособления к жизни в толще воды особенно ярко выражены у радиолярий. Основная линия приспособления к планктонному образу жизни сводится к выработке таких особенностей строения, которые способствуют парению тела в толще воды.
Типичным планктонным, причем почти исключительно морским семейством инфузорий является тинтинниды (Tintinnidae, рис. 105, 5). Общее количество известных на сегодняшний день видов тинтиннид составляет около 300. Это мелкие формы, характеризующиеся тем, что протоплазматическое тело инфузории помещено в прозрачный, легкий и в то же время прочный домик, состоящий из органического вещества. Из домика выступает диск, несущий венчик ресничек, находящихся в постоянном мерцающем движении. В состоянии парения инфузории в толще воды в основном поддерживаются здесь постоянной активной работой ресничного аппарата. Домик, очевидно, выполняет функцию защиты нижней части тела инфузории. В пресной воде обитает всего 2 вида тинтиннид (не считая 7 видов, характерных только для озера Байкал).
У пресноводных инфузорий есть и другие приспособления к жизни в планктоне. У многих из них цитоплазма очень сильно вакуолизирована (Loxodes, Condylostoma, Trachelius), поэтому напоминает пену. Это приводит к значительному снижению удельного веса. Все эти инфузории, кроме того, имеют ресничку, благодаря работе которой тело инфузории, по удельному весу лишь немного превышающему удельный вес воды, легко поддерживается в состоянии «парения». У некоторых видов форма тела увеличивает удельную поверхность и облегчает плавание в воде. Например, некоторые планктонные инфузории оз. Байкал по форме напоминают зонтик или парашют (Liliomorpha, рис. 105, 2). В оз. Байкал встречается одна планктонная сосущая инфузория (Mcophrya pelagica, рис. 105, 4), резко отличающаяся от своих оседлых сородичей. У этого вида отсутствует стебель. Его протоплазматическое тело окружено широкой слизистой оболочкой, приспособлением для похудения. Наружу выдаются длинные тонкие щупальца, которые наряду со своей прямой функцией, вероятно, выполняют и другую — увеличение удельной поверхности, облегчающее плавание в воде.
Наконец, необходимо упомянуть еще об одной, так сказать, косвенной форме приспособления инфузорий к жизни в планктоне. Это прикрепление мелких инфузорий к другим организмам, ведущим планктонный образ жизни. Так, среди круглых инфузорий (Peritricha) есть довольно многочисленные виды, прикрепляющиеся к планктонным веслоногим. Это обычный и нормальный образ жизни для этих видов инфузорий.
Наряду с инфузориями и среди сосущих (Suctoria) есть виды, населяющие планктонные организмы.
ИНФУЗОРИЯ, ЖИВУЩАЯ В ПЕСКЕ
Песчаные пляжи и отмели представляют собой чрезвычайно своеобразную среду обитания. Вдоль побережья морей они занимают обширные площади и характеризуются своеобразной фауной.
Проведенные за последние годы в различных странах многочисленные исследования показали, что толща многих морских песков очень богата разнообразной микроскопической или близкой по размеру к микроскопической фауной. Между частицами песка имеется множество мелких и крошечных пространств, заполненных водой. Оказывается, эти пространства богато заселены организмами, принадлежащими к самым разнообразным группам животного мира. Десятки видов ракообразных, кольчатые черви, круглые черви, особенно многочисленны плоские черви, некоторые моллюски, кишечнополостные. Есть также большое количество простейших, в основном инфузорий. По современным данным, фауна инфузорий, обитающих на морском дне, насчитывает около 250-300 видов. Если иметь в виду не только инфузорий, но и другие группы организмов, населяющих толщу песка, то общее число их видов будет очень велико. Всю совокупность животных, населяющих толщу песка, живущих в мельчайших промежутках между песчинками, называют псаммофильной фауной.
Богатство и видовой состав псаммофильной фауны определяется многими факторами. Среди них особенно важен размер частиц песка. Крупные пески имеют бедную фауну. Бедна и фауна очень мелкозернистых заиленных песков (с диаметром частиц менее 0,1 мм), где, очевидно, промежутки между частицами слишком малы для заселения их животными. Наиболее богаты жизнью пески средне- и мелкозернистые.
Вторым фактором, играющим важную роль в развитии псаммофильной фауны, является богатство песка органическими остатками, разлагающими органическое вещество (так называемая степень сапробности). Пески, лишенные органического вещества, бедны жизнью. С другой стороны, пески, очень богатые органическим веществом, также почти безжизненны, так как разложение органического вещества приводит к истощению кислорода. К этому часто добавляется анаэробное сероводородное брожение.
Наличие свободного сероводорода является крайне негативным фактором, влияющим на развитие фауны.
В поверхностных слоях песка иногда развивается довольно богатая флора одноклеточных водорослей (диатомовые, перидиниевые). Это фактор, благоприятствующий развитию псаммофильной фауны, так как многие мелкие животные (в том числе и инфузории) питаются водорослями.
Наконец, очень негативно влияющим на псаммофильную фауну фактором является прибой. Это вполне объяснимо, так как прибой, омывая верхние слои песка, убивает здесь все живое. Наиболее многочисленна псаммофильная фауна в защищенных, хорошо прогреваемых бухтах. Приливы и отливы не мешают развитию псаммофильной фауны. Когда вода во время отлива временно уходит, обнажая песок, то в толще песка, в промежутках между песчинками она остается, и это не мешает существованию животных.
У инфузорий, входящих в состав псаммофильной фауны и принадлежащих к различным систематическим группам (отрядам, семействам), в процессе эволюции вырабатывается много общих признаков, являющихся приспособлениями к своеобразным условиям существования между песчинками.
На рис. 106 показаны некоторые виды псаммофильной фауны инфузорий, принадлежащие к разным отрядам и семействам. Между ними много общего. Тело большинства из них более или менее сильно вытянуто в длину, червеобразно. Это дает возможность легко «протиснуться» в мельчайшие дырочки между песчинками. У очень многих видов (рис. 106) удлинение тела сочетается с его уплощением. Цилиарный аппарат всегда хорошо развит, что позволяет активно, с определенной силой передвигаться в узких щелях. Часто на одной стороне червеобразного уплощенного тела развиваются реснички, противоположная сторона голая. Эта особенность, вероятно, связана с резко выраженной у большинства псаммофильных ядов способностью очень тесно и очень прочно прикрепляться к субстрату через ресничный аппарат (явление, называемое тигмотаксисом). Это свойство позволяет животным оставаться на месте в тех случаях, когда в узких промежутках, где они обитают, возникают течения воды. В этом случае, вероятно, выгоднее, чтобы сторона, противоположная той, которой животное прикрепляется к субстрату, была гладкой.
Чем питаются псаммофильные инфузории? Водоросли, особенно диатомовые, составляют значительную часть «рациона» многих видов. Бактерии служат им в меньшей степени пищей. Это также в значительной степени зависит от того, что в песках, которые не сильно загрязнены, мало бактерий. Наконец, особенно среди самых крупных псаммофильных инфузорий, имеется значительное число хищных форм, поедающих других инфузорий, принадлежащих к более мелким видам. Псаммофильные инфузории распространены повсеместно.
ИНФУЗОРИИ АПОСТОМАТЫ
Инфузории spirofria (Spirophrya subparasitica) в инцистированном состоянии часто можно встретить сидящими на небольшом стебельке на мелких планктонных морских рачках (особенно часто на рачках рода Idia). Пока рачок активно плавает в морской воде, сидящие на нем спирофрии не претерпевают никаких изменений. Для дальнейшего развития инфузорий необходимо, чтобы рачок был съеден морским гидроидным полипом, что часто бывает (рис. 107). Как только цисты спирофрии вместе с рачком проникают в пищеварительную полость, из них тут же выходят мелкие инфузории, которые начинают усиленно питаться пищевой кашицей, образовавшейся в результате переваривания проглоченного рачка. В течение часа размеры инфузории увеличиваются в 3-4 раза. Однако размножения на этом этапе не происходит. Перед нами типичная стадия роста инфузории, которая называется трофонт. Через некоторое время вместе с непереваренными пищевыми остатками трофонт выбрасывается полипом в морскую воду. Здесь он, активно плавая, спускается по телу полипа к его подошве, где и прикрепляется, будучи окружен кистой. Эта стадия инцистированной крупной инфузории, сидящей на полипе, называется томонт. Это фаза размножения. Томонт не питается, но быстро делится несколько раз подряд (рис. 107, 7). В результате получается целая группа очень мелких инфузорий. Их количество зависит от размера томонта, который, в свою очередь, определяется размером трофона, давшего ему начало. Мелкие инфузории, образующиеся в результате деления томонта (их называют томитами или залетными), представляют стадию расселения.
Они выходят из кисты, быстро плавают (не питаясь, но используя имеющиеся у них запасы в цитоплазме). Если им «повезет» наткнуться на копеподу, они тут же прикрепятся к ней и инцистируют себя. Это этап, с которого мы начали наш обзор цикла.
В рассмотренном нами жизненном цикле спирофрии обращает на себя внимание резкое разграничение стадий, имеющих различное биологическое значение. Трофонт – стадия роста. Он только растет, накапливает большое количество цитоплазмы и всевозможных запасных веществ за счет энергичного и быстрого питания. Трофонт не способен к размножению. У томонтов наблюдается обратное явление — неспособность к питанию и бурное размножение. После каждого деления роста не происходит, и поэтому размножение томонта сводится к быстрому распаду на множество залетных. Наконец, бродяги выполняют свою особую и уникальную функцию: это особи — расселители и распространители видов. Они не могут ни питаться, ни размножаться.
ЖИЗНЕННЫЙ ЦИКЛ ИХТИОФТИРИУСОВ
К концу периода роста ихтиофтириус, по сравнению с залетными, достигает очень больших размеров: 0,5-1 мм в диаметре. Достигнув предельного значения, инфузории активными движениями покидают ткани рыбы в воду и некоторое время медленно плавают с помощью ресничного аппарата, покрывающего все их тело. Вскоре крупные ихтиофтирии оседают на каком-нибудь подводном предмете и выделяют цисту. Сразу после инцистирования начинаются последовательные деления инфузории: сначала пополам, затем каждая дочка снова делится на две, и так до 10-11 раз. В результате внутри кисты образуется до 2000 мелких почти округлых особей, покрытых ресничками. Внутри кисты активно перемещаются бродяги. Они пробивают скорлупу и выходят наружу. Активно плавающие бродяги заражают новых рыб.
Скорость деления ихтиофтириуса в цистах, а также скорость его роста в тканях рыб в значительной степени зависят от температуры. По исследованиям разных авторов приведены следующие цифры: при 26-27°С процесс развития залетников в кисте занимает 10-12 часов, при 15-16°С — 28-30 часов, при 4 -5°С сохраняется в течение 6 -7 дней.
Борьба с ихтиофтириозом представляет значительные трудности. Основное значение здесь имеют профилактические мероприятия, направленные на то, чтобы свободно плавающие в воде залетные особи не проникли в ткани рыб. Для этого полезно проводить частую пересадку больных рыб в новые водоемы или аквариумы, создавать проточные условия, что особенно эффективно при борьбе с ихтиофтириозом.
ИНФУЗОРИЯ ТРИХОДИНА
Вся система приспособлений триходин к жизни на поверхности хозяина направлена на то, чтобы не отрываться от тела хозяина (что почти всегда равносильно смерти), сохраняя при этом подвижность. Эти устройства чрезвычайно совершенны. Тело большинства триходин имеет форму довольно плоского диска, иногда шляпки. Сторона, обращенная к телу хозяина, слегка вогнута; он образует присоску для прикрепления. По наружному краю присоски расположен венчик из хорошо развитых ресничек, с помощью которых инфузория передвигается (ползает) по поверхности тела рыбы. Этот венчик соответствует венчику, обнаруженному у ресничных малоподвижных инфузорий, о которых говорилось выше. Таким образом, триходину можно сравнить с бродягой. На брюшной поверхности (на присоске) триходины имеют очень сложный опорно-прикрепительный аппарат, помогающий удерживать инфузорию на хозяине. Не вдаваясь в подробности его строения, отметим, что в его основе лежит кольцо сложной конфигурации, которое составлено из отдельных сегментов, несущих наружный и внутренний зубья (рис. 109)., Б). Это кольцо образует эластичную, но прочную основу брюшной поверхности, которая действует как присоска. Разные виды триходин отличаются друг от друга количеством члеников, образующих кольцо, и конфигурацией наружных и внутренних крючков.
На противоположной от диска стороне тела триходины расположены перистом и ротовой аппарат. Его строение более или менее типично для круглых инфузорий … Закрученные по часовой стрелке адоральные перепонки ведут в углубление, на дне которого расположен рот. Ядерный аппарат триходина устроен типично для инфузорий: один лентовидный макронуклеус и один микронуклеус, расположенный рядом с ним. Имеется одна сократительная вакуоль.
Триходины широко распространены во всех типах водоемов. Особенно часто они встречаются на мальках различных видов рыб. При массовом размножении триходины наносят большой вред рыбам, особенно если массово закрывают жабры. Это нарушает нормальное дыхание рыбы.
Для очистки рыб от триходина рекомендуется делать лечебные ванночки из 2% раствора хлорида натрия или 0,01% раствора перманганата калия (малькам — на 10-20 минут).
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА UNGEALS
Из рубца через сетку пища срыгивается в полость рта, где дополнительно пережевывается (жвачка). Вновь проглоченная пережеванная пищевая масса по специальной трубке, образованной складками пищевода, поступает уже не в рубец, а в книгу и оттуда в сычуг, где подвергается действию пищеварительных соков жвачных. В сычуге в условиях кислой реакции и наличия пищеварительных ферментов инфузории погибают. Когда они попадают туда с жевательной резинкой, они перевариваются.
Количество простейших в рубце (как и в сетке) может достигать колоссальных значений. Если взять каплю содержимого рубца и рассмотреть ее под микроскопом (при нагревании, так как инфузории останавливаются при комнатной температуре), то инфузории буквально роятся в поле зрения. Трудно даже в культуре получить такую массу инфузорий. Количество инфузорий в 1 см содержимого рубца достигает миллиона, а нередко и более. В пересчете на весь объем рубца это дает поистине астрономические цифры! Богатство содержимого рубца у инфузорий в значительной степени зависит от характера питания жвачных. Если пища богата клетчаткой и бедна углеводами и белками (трава, солома), то инфузорий в рубце относительно мало. При добавлении в рацион углеводов и белков (отрубей) численность инфузорий резко возрастает и достигает огромных размеров. Необходимо учитывать, что происходит постоянный отток инфузорий. Попадая в сычуг вместе с жвачкой, они погибают. Высокий уровень численности инфузорий поддерживается их бурным размножением.
У непарнокопытных (лошадь, осел, зебра) также большое количество инфузорий в пищеварительном тракте, но их локализация в организме хозяина иная. У непарнокопытных отсутствует сложный желудок, в связи с чем возможность развития простейших в передних отделах пищеварительного тракта отсутствует. Но у непарнокопытных очень хорошо развиты ободочная и слепая кишки, которые обычно забиты пищевыми массами и играют существенную роль в пищеварении. В этом отделе кишечника, так же как в рубце и ретикулуме жвачных животных, развивается очень богатая фауна простейших, главным образом инфузорий, большая часть которых также относится к отряду эндодиниоморфных. Однако видовой состав фауны рубца жвачных и фауны толстого кишечника непарнокопытных не совпадает.
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА ЖВАЧИХ
Наибольший интерес представляют инфузории. из семейства офриосколецид (Ophryoscolecidae), относящегося к отряду эндодиниоморфов . Характерная черта этого отряда — отсутствие сплошного цилиарного покрова. Сложные реснитчатые образования — циррусы — располагаются на переднем конце тела инфузорий в области ротового отверстия. К этим основным элементам цилиарного аппарата могут добавляться дополнительные группы циррусов, располагающиеся либо на переднем, либо на заднем конце тела. Общее число видов инфузорий семейства офриосколецид около 120.
На рисунке 110 изображены некоторые из наиболее типичных представителей офриосколецид рубца жвачных. Наиболее простыми являются инфузории рода Entodinium (Энтодиниум, рис. 110, Л). На переднем конце их тела имеется одна периоральная зона, цирроз. Передний конец тела, на котором расположено ротовое отверстие, можно втянуть внутрь. Эктоплазма и эндоплазма резко дифференцированы. На заднем конце хорошо видна анальная трубка, служащая для выведения наружу непереваренных остатков пищи. Несколько более сложная структура аноплодиния (Anoplodinium, рис. 110, Б). Имеют две зоны цилиарного аппарата — периоральный циррус и дорсальный циррус. Оба расположены на переднем конце. На заднем конце тела у видов, изображенных на рисунке, имеются длинные острые выросты — это довольно типично для многих видов офриосколецид. Высказано предположение, что эти выросты способствуют «проталкиванию» инфузорий между растительными частицами, заполняющими рубец.
просмотров род eudiplodinium (Eudiplodinium, рис. 110, Б) сходны с anoplodynia , но в отличие от них имеют скелетную базальную пластинку, расположенную на правом крае вдоль глотки. Эта скелетная пластинка состоит из вещества, сходного по химической природе с целлюлозой, то есть с веществом, из которого состоят оболочки растительных клеток.
У рода полипластрон (Polyplastron, рис. 110, Г, Д) наблюдается дальнейшее усложнение скелета. Строение этих инфузорий близко к эвдиплодиниям. Различия сводятся прежде всего к тому, что вместо одной скелетной пластинки у этих инфузорий их пять. Два из них, самые крупные, расположены с правой стороны, а три, более мелкие, — с левой стороны инфузории. Второй особенностью полипластрона является увеличение числа сократительных вакуолей. Энтодиния имеет одну сократительную вакуоль, аноплодиния и эвдиплодиния — две сократительные вакуоли, а полипластрон — около десятка.
У эпидиний (Epidinium, рис. 110), имеющих хорошо развитый углеводный скелет, расположенных на правой стороне тела, дорсальная зона цирроза смещена с переднего конца на дорсальную сторону. Шипы часто развиваются на заднем конце инфузорий этого рода.
Наиболее сложное строение обнаруживает род ofrioscolex (Ophryoscolex), которым названо все семейство инфузорий (рис. 110, Д). У них хорошо развит спинной цирроз, покрывающий примерно 2/3 окружности тела и скелетные пластинки. На заднем конце образуются многочисленные шипы, из которых один обычно особенно длинный.
Знакомство с некоторыми типичными представителями офриосколецид показывает, что внутри этого семейства произошло значительное увеличение сложности организации (от эндодиния до офриосколекса).
Кроме инфузорий семейства офриосколецид , в рубце жвачных животных встречаются представители уже известного отряда равносторонних инфузорий … Они представлены небольшим числом видов. Их тело равномерно покрыто продольными рядами ресничек, элементы скелета отсутствуют. В общей массе реснитчатого населения рубца они не играют заметной роли, и поэтому мы на них здесь останавливаться не будем.
Чем и как питаются инфузории из рода офриосколецид? Этот вопрос подробно изучался многими учеными, особенно подробно профессором В. А. Догелем.
Питание офриосколецид весьма разнообразно, и у разных видов наблюдается определенная специализация. Самые мелкие виды рода endodynia питаются бактериями, крахмальными зернами, грибами и другими мелкими частицами. Многие средние и крупные офриосколециды поглощают частицы растительных тканей, составляющие основную часть содержимого рубца. Эндоплазма некоторых видов буквально забита растительными частицами. Можно видеть, как инфузории набрасываются на обрывки растительной ткани, буквально разрывают их на куски, а затем проглатывают, часто закручивая спиралью в своем теле (рис. 111, 4). Иногда приходится наблюдать такие картины (рис. 111, 2), когда тело самой инфузории деформируется из-за проглоченных крупных частиц.
У офриосколецид иногда наблюдается хищничество. Более крупные виды поедают более мелких. Хищничество (рис. 112) сочетается со способностью того же вида питаться растительными частицами.
Каким образом инфузории попадают в рубец жвачных животных? Каковы пути заражения офриосколецидами? Оказывается, у новорожденных жвачных инфузорий в рубце еще нет. Они также отсутствуют, пока животное питается молоком. Но как только жвачное переходит на растительную пищу, в рубце и сачке тут же появляются инфузории, численность которых стремительно увеличивается. Откуда они? Долгое время предполагалось, что инфузории рубца образуют некие покоящиеся стадии (вероятнее всего, цисты), которые широко рассеяны в природе и, заглатывая, дают начало активным стадиям инфузории. Дальнейшие исследования показали, что у инфузорий рубца жвачных нет покоящихся стадий. Удалось доказать, что заражение происходит активными подвижными инфузориями, проникающими в полость рта при отрыжке десны. Если рассмотреть жвачку, взятую из ротовой полости, под микроскопом, в ней всегда содержится большое количество активно плавающих инфузорий. Эти активные формы могут легко проникать в рот и далее в рубец других жвачных из общего сосуда для помоев вместе с травой, сеном (в котором может содержаться слюна с инфузориями) и т. д. Такой путь заражения доказан экспериментально.
Если покоящиеся стадии офриосколецид отсутствуют, то, очевидно, легко получить «бесинфузорных» животных, изолируя их, пока они еще питаются молоком. Если не допускать прямого контакта растущего молодняка и жвачных с инфузориями, то молодняк может остаться без инфузорий в рубце. Такие эксперименты проводили несколько ученых в разных странах. Результат был однозначным. При отсутствии контакта молодняка (отъема от матери в период молочного вскармливания) и жвачных животных с инфузориями в рубце животные вырастают стерильными по отношению к инфузориям. Однако даже кратковременного контакта с животными с инфузориями (общая кормушка, общее ведро для поения, общее пастбище) достаточно, чтобы реснитчатая фауна появилась в рубце стерильных животных.
Выше были приведены результаты опытов по содержанию жвачных животных полностью лишенными инфузорий в рубце и сетке. Это достигается, как мы видели, ранней изоляцией молоди. Опыты проводились на овцах и козах.
Таким образом, можно было проводить наблюдения за «беспрививочными» животными в течение значительного периода времени (более года). Как влияет отсутствие инфузорий в рубце на жизнь хозяина? Отрицательно или положительно влияет отсутствие инфузорий на хозяина? Чтобы ответить на этот вопрос, на козах были проведены следующие опыты. Были взяты близнецы (одного помета и одного пола), чтобы иметь больше похожего материала. Затем один из близнецов этой пары воспитывался без инфузорий в рубце (ранняя изоляция), а другой с самого начала употребления в пищу растительной пищи был обильно заражен многими видами инфузорий. Оба получали одинаковую диету и воспитывались в одинаковых условиях. Единственное различие между ними заключалось в наличии или отсутствии инфузорий. В нескольких парах исследованных таким образом малышей не обнаружено различий в ходе развития обоих представителей каждой пары («инфузорных» и «безинфузорных»). Таким образом, можно утверждать, что живущие в рубце и ретикулуме инфузории не оказывают существенного влияния на жизнедеятельность животного-хозяина.
Приведенные выше экспериментальные результаты не позволяют, однако, утверждать, что инфузории рубца совершенно безразличны к хозяину. Эти эксперименты проводились с обычным рационом хозяина. Не исключена возможность, что и в других условиях, при другом режиме питания (например, при недостаточном питании) удастся выявить влияние на хозяина фауны простейших, обитающих в рубце.
В литературе высказывались различные предположения о возможном положительном влиянии протозойной фауны рубца на процессы пищеварения хозяина. Указано, что многие миллионы инфузорий, активно плавая в рубце и раздавливая растительные ткани, способствуют брожению и перевариванию пищевых масс, находящихся в передних отделах пищеварительного тракта. Значительное количество инфузорий, попадающих в сычуг вместе с жвачкой, переваривается, а белок, составляющий значительную часть тела инфузорий, усваивается. Таким образом, инфузории могут быть дополнительным источником белка для хозяина. Высказывалось также предположение, что инфузории способствуют перевариванию клетчатки, составляющей основную часть пищи жвачных животных, переводя ее в более усвояемое состояние.
Все эти предположения не доказаны, а некоторые из них вызывают возражения. Указывалось, например, что инфузории строят протоплазму своего тела из белков, попадающих в рубец с пищей хозяина. Поглощая растительный белок, они превращают его в животный белок своего тела, который затем переваривается в сычуге. Приносит ли это какую-либо пользу хозяину, остается неясным. Все эти вопросы представляют большой практический интерес, так как речь идет о пищеварении жвачных животных — основных объектов животноводства. Крайне желательны дальнейшие исследования роли инфузорий рубца в пищеварении жвачных животных.
Офриосколециды жвачных, как правило, обладают широкой специфичностью. По видовому составу рубцовое и чистое поголовье крупного рогатого скота, овец, коз очень близко друг к другу. Если сравнить видовой состав рубца африканских антилоп с крупным рогатым скотом, то и здесь общими будут около 40% от общего числа видов. Однако существует множество видов офриосколецид, которые встречаются только у антилоп или только у оленей. Таким образом, на фоне общей широкой специфичности офриосколецид можно говорить о некоторых более узкоспецифических их видах.
ИНФУЗОРИИ КИШЕЧНЫЕ
Обратимся теперь к краткому знакомству с инфузориями, населяющими толстую и слепую кишку непарнокопытных.
В видовом отношении эта фауна, как и фауна рубца жвачных животных, также очень разнообразна. В настоящее время описано около 100 видов инфузорий, обитающих в толстом кишечнике лошадиных животных. Встречающиеся здесь инфузории в смысле принадлежности к разным систематическим группам более разнообразны, чем инфузории рубца жвачных.
В кишечнике лошадей встречается довольно много видов инфузорий, относящихся к отряду эквилациевых, то есть инфузорий, у которых ресничный аппарат не образует мембранелл или цирросов вблизи ротовой зоны (рис. 113, 1 ).
Энтодиниоморфы отряда (Entodiniomorpha) также обильно присутствуют в кишечнике лошади. В то время как в рубце жвачных животных находится только одно семейство эндодиниоморф (семейство офриосколецид), в кишечнике лошади обитают представители трех семейств, на характеристике которых мы, однако, не будем здесь останавливаться, ограничившись лишь несколькими рисунками. типичных пород лошадей (рис. 113)…
Детальные исследования А. Стрелкова показали, что разные виды инфузорий далеко не равномерно распределены по ходу толстого кишечника лошади. Есть две разные группы видов, как две фауны. Один из них обитает в слепой кишке и брюшном отделе толстой кишки (начальные отделы толстой кишки), а другой — в дорсальном отделе толстой и тонкой кишки. Эти два комплекса видов довольно резко разграничены. Общих типов для этих двух разделов немного – менее десятка.
,
Интересно отметить, что среди многочисленных видов инфузорий, населяющих толстый кишечник непарнокопытных, есть представители одного рода, относящиеся к сосущим инфузориям. Как мы видели выше, сосущие инфузории (Suctoria) — типичные свободноживущие сидячие организмы с совершенно особым способом питания с помощью щупалец (рис. 103). Один из родов сукорий приспособился к такой, казалось бы, необычной среде обитания, как толстая кишка лошади, например, несколько видов аллантоз (аллантосомы). У этих очень своеобразных животных (рис. 114) нет стебля, реснички отсутствуют, булавовидные, утолщенные на концах щупальца хорошо развиты.
С помощью щупалец аллантосомы присасываются к разным видам инфузорий и высасывают их. Зачастую добыча во много раз превосходит хищника.
До сих пор не ясен вопрос о характере взаимоотношений инфузорий толстого кишечника непарнокопытных и их хозяев. Количество инфузорий может быть столь же велико, а иногда и больше, чем в рубце жвачных животных. Имеются данные, что количество инфузорий в толстой кишке лошади может достигать 3 млн в 1 см3. Предполагаемое некоторыми учеными симбиотическое значение еще менее вероятно, чем для инфузорий рубца.
Наиболее вероятно мнение, что они наносят некоторый вред хозяину, поглощая значительное количество пищи. Часть инфузорий выносится с фекальными массами, и таким образом органические вещества (в том числе белок), входящие в их тела, остаются неиспользованными хозяином.
До сих пор не решен вопрос о путях заражения непарнокопытных инфузориями, обитающими в толстой кишке.
Balantidium захватывает различные частицы пищи из содержимого толстой кишки. Особенно охотно он питается крахмальными зернами. Если балантидий обитает в просвете толстой кишки человека, то он питается содержимым кишечника и не оказывает вредного действия. Это типичный «переносчик», с которым мы уже встречались при рассмотрении дизентерийной амебы. Однако таким «безобидным постояльцем» остается балантидия, реже, чем дизентерийная амеба.
В настоящее время специалистами хорошо разработаны различные методы, позволяющие культивировать балантидии в искусственной среде — вне организма хозяина.
Как видно из рисунка, троглодптелла относится к числу сложных эндодиниоморф. У нее, помимо периоральной зоны цирроза (на переднем конце тела), имеются еще три зоны хорошо развитого цирроза, кольцеобразно опоясывающие тело инфузории. Troglodytellae имеют хорошо развитый скелетный аппарат, покрывающий почти весь передний конец тела, состоящий из углеводов. Размеры этих своеобразных инфузорий весьма значительны. В длину они достигают 200-280 мкм.
БЕЗ СТОМАТИЧЕСКИХ ИНФУЗОРИЙ
Опорные скелетные образования развиваются преимущественно на переднем конце тела, которому приходится испытывать механические нагрузки и преодолевать препятствия, проталкиваясь в просвет кишечника между пищевыми частицами. У вида рода радиофрии (Radiophrya) на переднем конце с одной стороны тела (условно считающейся брюшной) имеются очень сильные эластичные ребра (спикулы), залегающие в поверхностном слое эктоплазмы (рис. 117). , Б, Г, Д). В видах рода memelella (Mesnilella) также имеют опорные лучи (спикулы), которые на большей части своей длины лежат в более глубоких слоях цитоплазмы (в эндоплазме, рис. 117, А). Аналогично устроенные опорные образования развиты у видов некоторых других родов Astomat.
Бесполое размножение у некоторых инфузорий астомат протекает своеобразно. Вместо поперечного деления надвое, характерного для большинства инфузорий, у многих астомат деление неравномерное (почкование). При этом почки, отделившиеся на заднем конце, некоторое время остаются связанными с матерью (рис. 117, В). В результате получаются цепочки, состоящие из передних крупных и задних более мелких особей (почек). В дальнейшем почки постепенно отделяются от цепочки и переходят к самостоятельному существованию. Эта своеобразная форма размножения широко распространена, например, у уже известных радиофрий. Цепочки некоторых астоматиков, возникающие в результате почкования, по внешнему виду напоминают цепочки ленточных червей… Здесь мы снова сталкиваемся с явлением конвергенции.
Ядерный аппарат астомат имеет характерное для инфузорий строение: макронуклеус, чаще всего лентовидный (рис. 117), и один микронуклеус. Сократительные вакуоли обычно хорошо развиты. Большинство видов имеют несколько (иногда более десятка) сократительных вакуолей, расположенных в один продольный ряд.
Изучение распределения видов астоматов по разным типам хозяев показывает, что большая часть астоматов приурочена к строго определенным типам хозяев. Для большинства астомат характерна узкая специфичность: хозяином для них может служить только один вид животного.
Несмотря на большое количество работ, посвященных изучению астоматных инфузорий, остается совершенно неясным один очень важный аспект их биологии: как происходит передача этих инфузорий от одной особи хозяина к другой? Никогда не удавалось наблюдать образование цист у этих инфузорий.
Поэтому предполагается, что заражение происходит активно — подвижными стадиями.
Инфузории кишечные морских ежей
Морские ежи очень многочисленны в прибрежной зоне наших северных (Баренцево) и дальневосточных морей (Японское море, Тихоокеанское побережье Курильских островов). Большинство морских ежей питается растительной пищей, преимущественно водорослями, которые они соскребают с подводных предметов особыми острыми «зубами», окружающими ротовое отверстие. В кишечнике этих травоядных ежей имеется богатая фауна инфузорий. Часто они развиваются здесь в массовом количестве, и содержимое кишечника морского ежа под микроскопом почти так же «кишит» инфузориями, как и содержимое рубца жвачных. Надо сказать, что, кроме глубоких различий в условиях жизни инфузорий кишечника морского ежа и рубца жвачного, есть и некоторые сходства. Они заключаются в том, что и там, и там инфузории обитают в среде, очень богатой растительными остатками. В настоящее время известно более 50 видов инфузорий, обитающих в кишечнике морских ежей, которые встречаются только в прибрежной зоне, где ежи питаются водорослями. На больших глубинах, где уже не растут водоросли, у морских ежей нет инфузорий.
По образу жизни и характеру питания большинство инфузорий кишечника морских ежей растительноядны. Питаются водорослями, которые в большом количестве заполняют кишечник хозяина. Некоторые виды довольно «привередливы» в выборе пищи. Например, стробилидиум (Strobilidium, рис. 118, А) питается почти исключительно крупными диатомеями. Встречаются и хищники, поедающие представителей других, более мелких видов инфузорий.
У инфузорий из кишечника морских ежей, в отличие от астоматов, нет строгой привязки к определенным видам хозяев. В них обитают самые разные виды морских ежей, которые питаются водорослями.
Пути заражения морских ежей инфузориями не изучены. Однако и здесь с большой долей вероятности можно предположить, что это происходит активными свободноплавающими формами. Дело в том, что инфузории из кишечника морских ежей могут длительное время (многие часы) жить в морской воде. Однако они уже настолько приспособились к жизни в кишечнике ежей, что рано или поздно погибают вне их тела, в морской воде.
Заканчивая знакомство с инфузориями, следует еще раз подчеркнуть, что они представляют собой видовую, обширную и процветающую группу (класс) животного мира. Оставаясь на уровне клеточной организации, инфузории достигли наибольшей сложности строения и функций по сравнению с другими классами простейших.
Особенно значительную роль в этом прогрессивном развитии (эволюции), вероятно, сыграла трансформация ядерного аппарата и возникновение ядерного дуализма (качественная неравноценность ядер). Богатство макронуклеуса нуклеиновыми веществами связано с активными метаболическими процессами, с бурными процессами синтеза белков цитоплазмы и ядер.
Заключение
Вот и подошёл к концу наш обзор строения и образа жизни обширного вида животного мира — простейших . Характерной особенностью их, как неоднократно подчеркивалось выше, является одноклеточность. По своему строению простейшие представляют собой клетки. Однако они несравнимы с клетками, составляющими тело многоклеточных организмов, потому что сами являются организмами. Таким образом, простейшие – это организмы на клеточном уровне организации. Некоторые высокоорганизованные простейшие, обладающие множеством ядер, уже как бы выходят за морфологические пределы строения клетки, что дает основание некоторым ученым называть таких простейших «надклеточными». Это мало меняет сути дела, так как одноклеточная организация еще типична для простейших.
В пределах одноклеточности простейшие прошли долгий путь эволюционного развития и дали огромное разнообразие форм, приспособленных к самым разнообразным условиям жизни. В основе родословного ствола простейших лежат два класса: саркодовые и жгутиковые. До сих пор в науке дискутируется вопрос о том, какой из этих классов более примитивен. С одной стороны, низшие представители саркодовых (амебы) имеют самое примитивное строение. Но жгутиковые проявляют наибольшую пластичность типа метаболизма и стоят как бы на границе между животным и флорой… В жизненном цикле некоторых саркодовых (например, фораминифер) есть жгутиковые стадии (гаметы), что указывает на их связь с жгутиконосцами. Очевидно, что ни современные саркоды, ни современные жгутиконосцы не могут быть исходной группой эволюции животного мира, ибо сами прошли долгий путь исторического развития и выработали многочисленные приспособления к современным условиям жизни на Земле. Вероятно, оба эти класса современных простейших следует рассматривать как два ствола в эволюции, происходящие от не доживших до наших дней древних форм, живших на заре развития жизни на нашей планете.
В ходе дальнейшей эволюции простейших произошли изменения иного характера. Некоторые из них привели к общему повышению уровня организованности, активности, интенсивности жизненных процессов. К числу таких филогенетических (эволюционных) преобразований следует, например, отнести развитие органелл движения и захвата пищи, достигшее высокого совершенства в классе инфузорий. Бесспорно, что реснички представляют собой органеллы, соответствующие (гомологичным) жгутикам. Если у жгутиковых, за немногими исключениями, число жгутиков невелико, то у инфузорий количество ресничек достигает многих тысяч. Развитие цилиарного аппарата резко повысило активность простейших, сделало более разнообразными и сложными формы их взаимоотношений с окружающей средой, формы реакций на внешние раздражители. Наличие дифференцированного ресничного аппарата, несомненно, явилось одной из главных причин прогрессивной эволюции в классе инфузорий, где возникло большое разнообразие форм, приспособленных к различным средам обитания.
Развитие ресничного аппарата инфузорий является примером такого рода эволюционных изменений, которые были названы акад. Северцова Ароморфозы. Ароморфозы характеризуются общим повышением организации, развитием приспособлений широкого значения. Под повышением организации понимают изменения, вызывающие повышение жизнедеятельности организма; они связаны с функциональной дифференциацией его частей и приводят к более разнообразным формам связи организма со средой. Именно с такого рода структурными преобразованиями в процессе эволюции связано развитие ресничного аппарата инфузорий. Это типичный ароморфоз.
У простейших, как подчеркивал В.А. Догель, изменения типа ароморфозов обычно связаны с увеличением числа органелл. Происходит полимеризация органелл. Типичным примером такого рода изменений является развитие цилиарного аппарата у инфузорий. Вторым примером ароморфоза в эволюции инфузорий является их ядерный аппарат. Выше мы рассмотрели особенности строения ядра инфузории. Ядерный дуализм инфузорий (наличие микронуклеуса и макронуклеуса) сопровождался увеличением числа хромосом в макронуклеусе (явление полиплоидии). Поскольку хромосомы связаны с основными синтетическими процессами в клетке, прежде всего с синтезом белков, этот процесс привел к общему повышению интенсивности основных жизненных функций. И здесь произошла полимеризация, затронувшая хромосомные комплексы ядра.
Инфузории — одна из самых многочисленных и прогрессивных групп простейших, произошедших от жгутиковых. Об этом свидетельствует полное морфологическое сходство их органоидов движения. Этот этап эволюции был связан с двумя крупными ароморфозами: один из них затронул органеллы движения, второй — ядерный аппарат. Оба эти вида изменений взаимосвязаны, так как оба ведут к повышению жизнедеятельности и усложнению форм взаимосвязи с внешней средой.
Наряду с ароморфозами существует и другой тип эволюционных изменений, выражающийся в развитии приспособлений (приспособлений) к определенным, резко очерченным условиям существования. Этот тип эволюционного изменения Северцов назвал идиоадаптацией. В эволюции простейших этот тип изменения сыграл очень большую роль. Выше при рассмотрении разных классов простейших приведены многочисленные примеры идиоадаптивных изменений. Приспособление к планктонному образу жизни у разных групп простейших, приспособление к жизни в песке у инфузорий, образование защитных оболочек ооцист у кокцидий и многое другое — все это идиоадаптации, сыгравшие большую роль в возникновении и развитии отдельных групп. , но не связано с общими прогрессивными изменениями в организации.
Адаптации к различным специфическим средам обитания у простейших очень разнообразны. Они обеспечили широкое распространение этого типа в самых разнообразных местообитаниях, о чем подробно говорилось выше при описании отдельных классов.
Большая медицинская энциклопедия
Infusoria Tetrahymena thermophila … Википедия
— (инфузории) класс наиболее высокоразвитых простейших (Protozoa). Основные признаки И.: наличие ресничек (для движения и питания), двух типов ядер (полиплоидный макронуклеус и диплоидный микронуклеус, различных по строению и… … Большая советская энциклопедия
Или комковатые (инфузории), класс простейших животных, снабженных реснитчатыми веревками или ресничками, которые во взрослом возрасте заменяются иногда цилиндрическими сосущими выростами, с определенной формой тела, большей частью с… … Энциклопедический словарь Ф.А. Брокгауз и И.А. Эфрон
Таксономия Ifusorium Классификация и систематизация организмов, относящихся к роду инфузорий, в соответствии с общей концепцией систематики простейших в частности и животных вообще Содержание 1 Систематика 1.1 Классические … Википедия
Или ликерные животные (инфузории), класс простейших животных, снабженных реснитчатыми веревками или ресничками, которые во взрослом состоянии замещаются иногда цилиндрическими присасывающимися выростами определенной формы тела, большей частью с… … Энциклопедия Брокгауза и Эфрон
Строение животной клетки одноклеточного организма инфузории туфельки.
Особенности строения и жизни инфузорииМесто обитания «Инфузория-туфелька»
В тех же водоемах с загрязненной водой, где встречаются амеба и эвглена, можно встретить быстроплавающее одноклеточное простейшее длиной 0,1-0,3 мм , чье тело по форме напоминает крошечный ботинок. Это инфузория туфелька. Она сохраняет постоянную форму тела благодаря тому, что наружный слой ее цитоплазмы плотный.
Инфузория-туфелька
Все тело инфузории покрыто продольными рядами многочисленных коротких ресничек, сходных по строению с жгутиками эвглены и вольвокса. Реснички совершают волнообразные движения, и с их помощью туфелька плавает тупым (передним) концом вперед.
Простейших, передвигающихся с помощью многочисленных ресничек, относят к инфузориям. Впервые инфузории были обнаружены в воде, настоянной на различных травах («инфузум» означает «настойка»).
Питание «Инфузория-туфелька»
От переднего конца до середины тела туфельки проходит желобок с более длинными ресничками. Задний конец желобка имеет устье , ведущее к короткому трубчатому горлу … Реснички желобка работают непрерывно, создавая поток воды. Вода подхватывает и подносит ко рту основную пищу туфельки – бактерии. Через зев бактерии попадают в тело инфузории. В цитоплазме вокруг них образуется пищеварительная вакуоль, в которую выделяется пищеварительный сок. Цитоплазма туфельки, как и у амебы, находится в постоянном движении. Пищеварительная вакуоль отрывается от глотки и подхватывается током цитоплазмы. Переваривание пищи и усвоение питательных веществ у инфузорий происходит так же, как у амеб. Через отверстие выбрасываются непереваренные остатки — порошок .
Дыхание и выделения у «инфузорий туфельки»
Дыхание и выделение у инфузории туфельки происходит так же, как и у других рассмотренных ранее простейших. Две сократительные вакуоли туфельки (передняя и задняя) сокращаются попеременно, через 20-25 секунд каждая. Вода и вредные продукты жизнедеятельности собираются из всей цитоплазмы туфельки приводящих канальцев , которые подходят к сократительным вакуолям.
В цитоплазме туфельки два ядра: большое и малое. Ядра имеют разное значение. Маленькое ядро играет главную роль в размножении. Большое ядро влияет на процессы движения, питания, выделения.
Репродукция «Инфузория туфелька»
Летом туфелька, интенсивно питаясь, растет и делится, подобно амебе, на две части. Малое ядро отходит от большого и делится на две части, расходящиеся к переднему и заднему концам тела. Затем большое ядро делится. Обувь перестает есть. Его натягивают посередине. Новообразованные ядра распространяются на переднюю и заднюю часть обуви. Сужение становится глубже, и, наконец, обе половинки отдаляются друг от друга — получаются две молодые инфузории. В каждой из них остается одна сократительная вакуоль, а вторая образуется заново со всей канальцевой системой. Начав питаться, молодые туфли растут. Через сутки деление повторяют снова.
Раздражительность «Инфузория-туфелька»
Проведем следующий опыт. Поместите на стакан каплю чистой воды и рядом с ней каплю воды с инфузориями. Соединим обе капли тонким водяным каналом. Положите в каплю с инфузориями небольшой кристалл соли. По мере растворения соли обувь будет плавать в капле чистой воды — для инфузорий раствор соли вреден.
Давайте изменим условия эксперимента. В каплю с инфузориями ничего добавлять не будем. Но добавьте в чистую каплю немного настоя с бактериями. Тогда вокруг обуви соберутся бактерии – их обычная пища. Эти опыты показывают, что инфузории могут определенным образом (например, движением) реагировать на воздействие (раздражение) окружающей среды, т. е. обладают раздражительностью. Это свойство свойственно всем живым существам.
Инфузория-туфелька — это одноклеточное животное, принадлежащее к подцарству простейших, типу Ciliophora или Ресничатые.
Среда обитания и строение инфузории-туфельки
Инфузория башмачок обитает в небольших пресных водоемах с загрязненной, стоячей водой — в прудах, канавах, лужах, то есть там же, где обитают другие простейшие — амеба и зеленая эвглена. Она постоянно в движении, плавает в поисках пищи. Передвигаться в воде ей помогают специальные органы движения — реснички, которые покрывают все ее удлиненное, башмаковидное тело. Ресничек у инфузории около 15 тысяч, в основании каждой реснички находится базальное тельце. Реснички колеблются синхронно, около 30 раз в секунду, напоминая весла лодки.
Благодаря слаженной работе ресничек инфузорий туфелька плавает со скоростью до 2,5 мм в секунду, одновременно вращаясь вокруг продольной оси. Учитывая, что размер инфузории всего 0,5 мм, это довольно высокая скорость. В замедленной съемке видно, что движение каждой реснички состоит из резкой волны в одну сторону и более плавного возврата назад.
Эластичная оболочка, а также опорные волокна, расположенные в прилежащем к оболочке слое цитоплазмы, помогают туфельке сохранять форму инфузории. Кроме того, под панцирем инфузории туфельки находятся особые защитные органы — трихоциты. Трихоциты представляют собой эластичные длинные нити, сжатые до состояния коротких палочек и располагающиеся перпендикулярно поверхности тела инфузории. При нападении инфузории трихоциты резко распрямляются и «выстреливают» в направлении атаки. На месте использованных трихоцитов образуются новые.
Еще одним отличительным признаком инфузории-туфельки является наличие у нее двух ядер: большого и малого. Большое ядро отвечает за питание, дыхание, обмен веществ и движение. Малое ядро участвует в половом размножении инфузории туфельки.
Инфузория-туфелька также имеет клеточный рот, две сократительные вакуоли на переднем и заднем концах тела и пищеварительные вакуоли.
Инфузория-туфелька питание
Инфузория, как и эвглена зеленая, имеет клеточный рот — углубление в цитоплазме. Однако у инфузории-туфельки она покрыта длинными толстыми ресничками, которые, колеблясь, загоняют в устье клетки бактерии — основную пищу инфузории — и переходят в глотку, на дне которой образуется пищеварительная вакуоль. Пища переваривается в пищеварительной вакуоли сначала в кислой среде, а затем в щелочной. Пищеварительная вакуоль движется по цитоплазме и открывается наружу на заднем конце тела, выбрасывая наружу непереваренные частицы пищи. Инфузория-туфелька способна улавливать в воде химические вещества, которые выделяют скопления бактерий, и плывет к ним.
Мелирование
Функцию органов выделения у инфузории туфельки, как и у других простейших, выполняют сократительные вакуоли. Инфузория туфелька имеет две сократительные вакуоли — на переднем и заднем концах тела. Каждая сократительная вакуоль состоит из центрального резервуара и направленных к нему 5-7 канальцев. Растворенные вредные вещества и продукты обмена сначала поступают в канальцы, а затем собираются в центральном резервуаре и выталкиваются наружу. Весь цикл накопления и выведения продуктов обмена, а также избытка воды происходит с частотой один раз примерно в 10-20 секунд.
Дыхание
Инфузория-туфелька дышит растворенным в воде кислородом. Так же, как и у других простейших, кислород поступает через всю поверхность тела.
Размножение
Инфузории-туфельки размножаются бесполым путем — делением клетки надвое. Сначала делятся ядра, затем образуется поперечный мостик, разделяющий тело инфузории на две части, каждая из которых содержит 1 большое и 1 малое ядро. На перемычке инфузория делится на 2 части. Каждая из дочерних инфузорий получает часть органелл, а недостающие органеллы, например сократительные вакуоли, возникают заново.
Деление у инфузорий происходит 1-2 раза в сутки.
Время от времени у инфузорий происходит конъюгация. Это половой процесс, при котором количество особей не увеличивается, а происходит обмен генетической информацией. Новые комбинации генов, возникающие при конъюгации, способствуют возникновению и отбору более жизнеспособных форм организмов.
Половой процесс состоит из следующей последовательности событий:
Сначала две инфузории сближаются и вступают в контакт друг с другом, в месте соприкосновения их оболочки растворяются, и возникает цитоплазматический мостик. Крупные ядра исчезают у инфузорий. Мелкие ядра делятся дважды, при этом образуется 4 дочерних ядра. 3 из них разрушаются, а четвертые ядра вновь делятся, в результате чего в каждой инфузории образуется по 2 ядра. Одно ядро женское, неподвижное. А второй – мужской, подвижный. Мужские ядра переходят через цитоплазматический мостик к другой инфузории. Далее мелкие ядра каждой инфузории вновь соединяются. Вновь появляются крупные ядра, исчезает цитоплазматический мостик, и инфузории расходятся.
После конъюгации инфузории интенсивно делятся бесполым путем.
Инфузория башмачок – довольно распространенный вид, относящийся к отряду. Обитает в пресных стоячих водоемах с достаточным количеством органики, которой и питается. Кстати, строение инфузории-туфельки считается самой сложной из этой группы организмов.
Общая характеристика
Инфузория башмачок — одноклеточный организм, форма которого действительно напоминает подошву обуви и сохраняется за счет плотного наружного слоя цитоплазмы. Все тело животного покрыто огромным количеством ресничек, которые расположены продольными рядами. Их основная функция – движение.
Туфелька с ресничками движется тупым концом вперед. Реснички двигаются с небольшим опозданием относительно друг друга. При движении тело также вращается вокруг оси.
Между ресничками располагаются так называемые трихоцисты — мелкие веретенообразные органеллы, выполняющие функцию защиты. Каждая трихоциста состоит из тела и верхушки, которые при наличии раздражителя (соударения, нагревания, охлаждения) резко стреляют.
Башмак инфузории: структура
Большую часть тела составляет эндоплазма или жидкая часть цитоплазмы. Эктоплазма ближе к цитоплазматической мембране, имеет более плотную консистенцию и образует пелликулу.
Пищеварение. Инфузория башмачок питается бактериями и имеет довольно своеобразное клеточное строение. Ближе к переднему концу тела находится периоральная воронка, внутренняя поверхность которой покрыта сложной системой ресничек. Движения ресничек создают поток, вместе с которым всасываются микроорганизмы. Далее питательные частицы попадают в глотку, также выстланную ресничками, и только потом в рот. Путем эндоцитоза они попадают в пищеварительные вакуоли. Остатки выводятся через специфическую органеллу — порошок.
Генетический материал. Инфузория башмачок имеет два ядра — большое (макронуклеус) и малое (микронуклеус). Микронуклеус содержит полный набор генетической информации и принимает участие в половом размножении организма. Макронуклеус отвечает за синтез белковых соединений.
Выделение и дыхание. Инфузория башмачок способна существовать даже при очень низкой концентрации кислорода в воде. Кислород поглощается всей поверхностью.
Как уже упоминалось, этот простейший организм обитает в пресной воде и из-за разницы в концентрации нуждается в системе осморегуляции. Инфузория имеет две сократительные вакуоли — переднюю и заднюю, к каждой из которых ведет разветвленная трубчатая система. Избыток жидкости и вторичные продукты обмена собираются в канальцах и вакуоли выводятся в окружающую среду. Обе органеллы сокращаются попеременно, через каждые 15-20 секунд.
Размножение инфузории-туфельки
Для этого организма характерно как половое, так и бесполое размножение.
Осуществляется поперечным делением клетки на две равные части. При этом тело остается активным. Далее следуют довольно сложные процессы регенерации, в ходе которых каждая часть тела достраивает необходимые органеллы.
Половая связь между двумя лицами осуществляется путем сопряжения. Инфузории временно слипаются, и между их поверхностями образуется своеобразный цитоплазматический мостик. Макронуклеусы обоих организмов разрушаются, а мелкие ядра делятся мейозом.
После этого образуются четыре ядра с гаплоидным набором хромосом. Затем три из них отмирают, а остальные делятся митозом, образуя два протонуклеуса — женский и мужской. Организмы обмениваются «мужскими» протонядрами. Затем в каждом происходит слияние двух ядер и образование синкариона. Затем происходит митоз, после которого одно из образовавшихся ядер становится макронуклеусом, а второе — микронуклеусом.
Это достаточно распространенный вид простейших, обитающий в пресных водоемах со стоячей водой. Главным условием обитания инфузорий являются именно стоячие водоемы с достаточным количеством в них органических материалов, служащих пищей для этих простейших. Второе название этого существа – Хвостатая Парамеция из рода Paramecium. Любопытно, что строение инфузории-туфельки наиболее сложное из всех представителей этой группы организмов.
Инфузория туфелька. Структура
Этот одноклеточный организм получил свое название из-за сходства с подошвой обуви. Любопытно, что столь необычная форма этого существа обусловлена плотным наружным слоем цитоплазмы. Все тело инфузории-туфельки покрыто мельчайшими ресничками (жгутиками), расположенными продольными рядами. Именно они помогают инфузориям передвигаться в водной среде: за 1 секунду простейшая может преодолеть расстояние в 15 раз большее, чем она сама. Инфузория-туфелька движется тупым концом вперед, постоянно вращаясь при движении вокруг собственной оси.
Между жгутиками у инфузорий располагаются трихоцисты — мелкие веретеновидные органеллы, обеспечивающие ей защиту от внешних раздражителей. Каждая такая трихоциста состоит из небольшого тела и кончика, который резким выстрелом реагирует на любой раздражитель (нагревание, столкновение, охлаждение). Рот этого простейшего организма имеет воронкообразную форму: при попадании в него пищи он окружается пищевой вакуолью, совершая с ней небольшое «путешествие», пока не переварится. Отходы выбрасываются через так называемый порошок (специфическая органелла).
Основную часть этих существ составляет эндоплазма (жидкая часть цитоплазмы). Эктоплазма располагается рядом с цитоплазматической мембраной, имеет более плотную консистенцию и образует пелликулу. Инфузория-туфелька поглощает кислород всей своей поверхностью, существуя даже при малой его концентрации в воде. Все это позволяет по праву называть инфузорий наиболее высокоорганизованными простейшими, вершиной их эволюции.
Инфузория туфелька. Размножение
Этот одноклеточный организм размножается двумя способами: бесполым и половым. Бесполое размножение происходит за счет поперечного деления клетки на две равные части. При этом тело инфузории сохраняет свою активность. Далее происходят сложные процессы регенерации, в результате которых каждая из частей тела «достраивает» все необходимые органеллы.
Половой способ размножения инфузории-туфельки по понятным причинам выглядит несколько иначе. Две особи временно «слипаются» друг с другом, образуя между собой своеобразный мостик из цитоплазмы. В это время макронуклеусы обоих организмов разрушаются, а мельчайшие ядрышки начинают делиться мейозом.
Через некоторое время появляются четыре ядра, три из которых обязательно погибнут. Оставшееся ядро делится митозом. В результате образуются два протонуклеуса – мужской и женский. Обе особи начинают обмениваться «мужскими» протонядрами, после чего у каждой из них происходит дополнительное слияние двух ядер, сопровождающееся образованием синкариона. В результате очередного митоза одно из новообразованных ядер становится микронуклеусом, а второе — макронуклеусом.
Инфузории — группа простейших организмов с более сложным строением. К ним относятся различные виды инфузорий, бурсариев, сувоек и др. Типичным представителем является вид инфузории-туфельки. Большинство инфузорий живут в пресной воде в ее толще или на дне и питаются органическими остатками, бактериями и одноклеточными водорослями. Ряд инфузорий могут образовывать колонии.
Инфузории сохраняют постоянную форму тела за счет наличия клеточной стенки. Однако форма и размеры тела у разных видов разные. Размеры от 10 мкм до 5 мм.
У инфузорий тело покрыто ресничками. У инфузории-туфельки реснички равномерно распределены по всему телу. С их помощью ботинок плавает, ввинчиваясь в воду. У стелонихий реснички собраны в пучки, это дает возможность двигаться, как бы ползая. Сувои ведут оседлый образ жизни, прикрепляясь спиралевидным стеблем к растениям и различным предметам, находящимся под водой. Стебель может уменьшиться.
Инфузории имеют большое и малое ядро (малых может быть больше одного). Инфузория имеет ячеистый рот, глотку и присыпку (место для удаления мусора). У сократительных вакуолей имеются ведущие каналы.
.
1 — пищеварительная вакуоль, 2 — реснички, 3 — кристаллы, 4 — ротовое отверстие, 5 — формирующая пищеварительная вакуоль, 6 — порошок, 7 — продольные реснички, 8 — пищеварительная вакуоль, 9 — ядрышко, 10 — ядро, 11 — сократительная vacuole
Инфузория-туфелька питается бактериями. С помощью согласованной вибрации ресничек, окружающих устье клетки, туда попадают бактерии. Далее через зев клетки бактерии попадают в цитоплазму, где происходит образование пищеварительной вакуоли. После переваривания остатки удаляются через порошок.
Инфузории могут реагировать на различные вещества в воде, свет, температуру и т.д. Инфузории-туфельки подплывают к скоплениям бактерий, выплывают из соленой воды, уплывают в более освещенные места.
Инфузории размножаются делением надвое. Сначала их ядра делятся, после чего родительская клетка вытягивается в поперечном направлении, и образуются две дочерние клетки.
У сувоев, ведущих малоподвижный образ жизни, клетки также делятся надвое. При этом дочерние клетки отделяются от стебля и свободно плавают, их называют «бродягами». Таким образом, сувои оседают. «Бродяга» привязывается к предметам на новом месте.
При наступлении неблагоприятных условий (например, при температуре ниже 0°С) инфузории могут образовывать цисты округлой формы. Цисты могут распространяться ветром и живыми организмами.
| Международное научное название | |
|---|---|
Paramecium caudatum Эренберг, 1838 | |
| Просмотр: | Башмак инфузории | ||
| Латинское название | |||
|---|---|---|---|
| Paramecium caudatum Ehrhart | |||
|