6. Подтип Жгутиковые. Общая характеристика (на примере Эвглены Зеленой)
Они обитают в морях, пресных водах, в почве, а также в организмах животных и растений. Среди жгутиконосцев немало опасных паразитов животных и человека. Значение их в природе чрезвычайно велико. Многие жгутиконосцы составляют основу 40 планктона в водоемах и играют важную роль в биогенном круговороте в биоценозах. Жгутиконосцы — важное звено в цепях питания водных экосистем и служат объектом питания для более крупных организмов. Некоторые виды жгутиконосцев являются полезными симбионтами животных.
Подтип жгутиконосцев характеризуется следующими морфофизиологическими особенностями:
1.
Органеллами движения им служат жгутики — выросты цитоплазмы. Их может быть 1,
2, 4, 8 или множество.
Каждый
жгутик снаружи покрыт трехслойной
цитоплазматической мембраной. Внутри
жгутика имеются фибриллы: две центральные
и девять двойных периферических. Жгутик
крепится в цитоплазме при помощи
базального тельца — кинетосомы. Обычно
жгутики производят вращающее движение.
Жгутиковый
аппарат разнообразен по числу и форме
жгутиков, а также их расположению.
Движение жгутика обычно
Обычно
жгутики производят вращающее движение.
Жгутиковый
аппарат разнообразен по числу и форме
жгутиков, а также их расположению.
Движение жгутика обычно
Жгутик состоит из наружной части — бича и базальной части — кинетосомы, находящейся в эктоплазме клетки. Снаружи жгутик покрыт трехслойной мембраной, а внутри его располагаются 11 фибрилл. В центре жгутика располагаются две центральные фибриллы, а по периферии размещаются девять фибрилл, каждая из которых состоит из двух спаянных микротрубочек. Центральные фибриллы выполняют опорную функцию, а периферические — локомоторную.
Кинетосома
цилиндрической формы, покрыта мембраной.
В кинетосоме имеется особая аксиальная
гранула,
к которой прикрепляются центральные
фибриллы жгутика. Периферические девять
фибрилл продолжаются в кинетосому,
только становятся более сложными и
состоят уже не из двух, а из трех
спаянных микротрубочек. Центральные фибриллы в кинетосоме ниже
аксиальной гранулы отсутствуют. Рядом
с кинетосомой (базальным тельцем) может
располагаться особая органелла — кинетопласт,
который по своей функции соответствует
митохондрии и обеспечивает генерацию
энергии жгутику. В состав кинетопласта
входит дополнительно значительное
количество ДНК.
У части видов жгутиконосцев у основания
жгутика может находиться еще и парабазальное
тельце или блефаропласт,
содержащий запас резервных веществ,
расходуемых жгутиком при движении. По
своему строению парабазальное тельце
близко к аппарату Гольджи.
Центральные фибриллы в кинетосоме ниже
аксиальной гранулы отсутствуют. Рядом
с кинетосомой (базальным тельцем) может
располагаться особая органелла — кинетопласт,
который по своей функции соответствует
митохондрии и обеспечивает генерацию
энергии жгутику. В состав кинетопласта
входит дополнительно значительное
количество ДНК.
У части видов жгутиконосцев у основания
жгутика может находиться еще и парабазальное
тельце или блефаропласт,
содержащий запас резервных веществ,
расходуемых жгутиком при движении. По
своему строению парабазальное тельце
близко к аппарату Гольджи.
2. В отличие от саркодовых, у жгутиконосцев имеется пелликула, или панцирь. Поэтому у большинства жгутиковых форма тела постоянная.
3. Жгутиконосцам свойственны разнообразные способы питания. Среди них имеются

По характеру питания жгутиконосцев подразделяют на два класса:
4. Размножение чаще бесполое, путем продольного деления, реже наблюдается половое размножение (гамогамия) с образованием гамет и последующей копуляцией. Им свойственна зиготическая редукция хромосом.
Размеры жгутиконосцев варьируют от 1—2 мкм до нескольких миллиметров. Форма тела может быть овальной.
Клетка некоторых жгутиконосцев покрыта только мембраной, и они способны образовывать псевдоподии. Однако у большинства жгутиковых имеются пелликула, а у некоторых видов образуется панцирь, состоящий из клетчатки или из хитиноидного органического вещества.
Внутри
клетки жгутиконосцев имеется ядро,
а также другие органеллы. У зеленых
жгутиконосцев в цитоплазме располагаются хроматофоры,
содержащие хлорофилл. В результате
происходящего фотосинтеза в клетке
автотрофных жгутиконосцев накапливаются
резервные питательные вещества: зерна
парамила, близкого к крахмалу, капельки
жироподобных веществ.
У пресноводных жгутиконосцев часто имеется сократительная вакуоль, иногда с большим резервуаром, открывающимся наружу порой. Зеленые жгутиконосцы, как правило, имеют красный «глазок» — стигму, представляющую собой светочувствительную органеллу.
Размножение. Большинство жгутиковых размножается бесполым способом, путем продольного деления клетки на две дочерние. При этом ядро делится путем митоза. Делятся надвое базальные и парабазальные тельца, жгутик же переходит к одной из дочерних клеток, а у другой образуется заново.
У
колониальных жгутиконосцев бесполое
размножение колоний может происходить
двумя способами. При монотомическом
делении образующиеся дочерние клетки
сразу вырастают до размеров материнских
клеток. В делящейся колонии число клеток
увеличивается, а затем она перешнуровывается
надвое. При палинтомическом делении из
каждой материнской клетки колонии путем
многократного деления возникает новая
дочерняя колония, состоящая из мелких
клеток. В дальнейшем каждая дочерняя
колония растет и достигает размеров
материнской колонии.
В дальнейшем каждая дочерняя
колония растет и достигает размеров
материнской колонии.
| Главная / Сотрудники
| ||||||||||||||||||||||||||||||||||||||||||||||||||
 gif»>
gif»> gif»>
gif»>
 В., Верзилин Н.Н., Маслов Ю.И. Влияние различных источников азота на рост и накопление массы у Chlorella pyrenoidosa. Вестник ЛГУ,N 9, вып.2,1961, с.16-25.
В., Верзилин Н.Н., Маслов Ю.И. Влияние различных источников азота на рост и накопление массы у Chlorella pyrenoidosa. Вестник ЛГУ,N 9, вып.2,1961, с.16-25. В сб.: Развитие и значение почвенных водорослей в почвах Нечерноземной 6. Зоны и некоторые общие вопросы почвенной альгологии. Киров,1977, с.147-158.
В сб.: Развитие и значение почвенных водорослей в почвах Нечерноземной 6. Зоны и некоторые общие вопросы почвенной альгологии. Киров,1977, с.147-158. БиНИИ №39, 1988, с.153-163.
БиНИИ №39, 1988, с.153-163. Вестник С.-Петерб. ун-та, сер.3,2005 г., вып.3, с.121-128.
Вестник С.-Петерб. ун-та, сер.3,2005 г., вып.3, с.121-128. Простая английская Википедия, бесплатная энциклопедия
Переключить оглавлениеИз простой английской Википедии, бесплатной энциклопедии
(перенаправлено с Protista)
Различные виды протистов: . По часовой стрелке сверху слева: красные водоросли, бурые водоросли, инфузории, золотистые водоросли, фораминиферы; паразитический жгутиконосец; патогенная амеба; амебозойная слизевая плесень Протисты — одноклеточные эукариоты (организмы с ядром). Срок Protista впервые был использован Эрнстом Геккелем в 1866 году. [1] Протисты не являются естественной группой или кладой, поскольку они не имеют общего происхождения. Подобно водорослям или беспозвоночным, их часто для удобства группируют вместе.
Срок Protista впервые был использован Эрнстом Геккелем в 1866 году. [1] Протисты не являются естественной группой или кладой, поскольку они не имеют общего происхождения. Подобно водорослям или беспозвоночным, их часто для удобства группируют вместе.
Термин «протисты» включает микроорганизмы из нескольких отдаленно родственных типов. Некоторые из них автотрофны (что означает, что они производят себе пищу путем фотосинтеза), а другие гетеротрофны (что означает, что они едят органический материал).
Большинство простейших очень маленькие. Они состоят из одной или максимум нескольких клеток — они микроскопические и обычно невидимы невооруженным глазом. Некоторые водоросли являются протистами, если они одноклеточные. Многие протисты являются частью планктона и очень важны для экосистемы. Клетки, обнаруженные у протистов, могут быть чрезвычайно сложными и часто малоизученными. Теперь можно проводить секвенирование ДНК, и было проанализировано несколько простейших.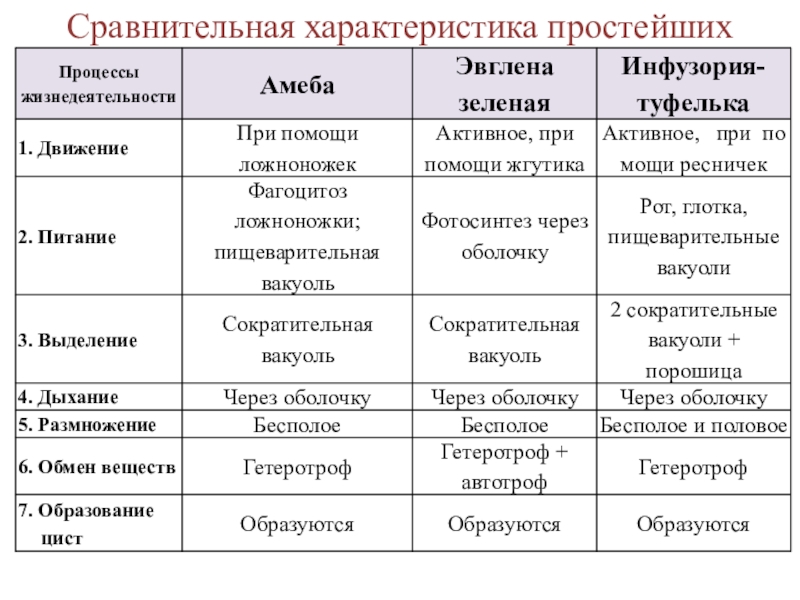 Результаты показывают, что Protista не является монофилетической группой. Это парафилетический, а не одна клада. Таким образом, таксономия Protista довольно запутана.
Результаты показывают, что Protista не является монофилетической группой. Это парафилетический, а не одна клада. Таким образом, таксономия Protista довольно запутана.
Некоторые простейшие вызывают болезни. Plasmodium falciparum вызывает малярию; сонная болезнь также вызывается простейшими.
Примером одноклеточного организма в царстве протистов является Paramecium или «тапочка анималистика». Paramecium передвигается с помощью своих маленьких волосовидных волокон, называемых ресничками, и ест, используя реснички, чтобы подметать пищу в пищевую вакуоль. Другими протистами могут быть амебы, которые передвигаются, вытягивая ложноножки и втекая в них, или обтекая частицы пищи и поглощая их.
| Тип питания | Источник энергии | Источник углерода | Примеры |
|---|---|---|---|
| Фототрофы | Солнечный свет | Органические соединения или фиксация углерода | Водоросли, динофлагелляты или эвглены |
| Органотрофы Гетеротрофы | Органические соединения | Органические соединения | Апикомплексы, трипаносомы или амебы |
Протисты не являются монофилетической кладой. Этот термин является удобным для обозначения примерно 20 различных видов одноклеточных эукариот. [2] Они отличаются своими клеточными органеллами, специализированными единицами, выполняющими четко определенные функции, такими как митохондрии и пластиды. Это доказывает, что они совершили переход от прокариот разными путями. Теперь совершенно ясно, что все или большинство этих органелл происходят от некогда независимых прокариот (бактерий или архей) и что эукариотическая клетка представляет собой «сообщество микроорганизмов», работающих вместе в «браке по расчету». [3] [4] [5] [6] По общему признанию, Protista представляет собой набор разрозненных одноклеточных форм, но в то время как более сложная таксономия находится в постоянном движении (изменении), Protista все еще остается полезный термин.
Этот термин является удобным для обозначения примерно 20 различных видов одноклеточных эукариот. [2] Они отличаются своими клеточными органеллами, специализированными единицами, выполняющими четко определенные функции, такими как митохондрии и пластиды. Это доказывает, что они совершили переход от прокариот разными путями. Теперь совершенно ясно, что все или большинство этих органелл происходят от некогда независимых прокариот (бактерий или архей) и что эукариотическая клетка представляет собой «сообщество микроорганизмов», работающих вместе в «браке по расчету». [3] [4] [5] [6] По общему признанию, Protista представляет собой набор разрозненных одноклеточных форм, но в то время как более сложная таксономия находится в постоянном движении (изменении), Protista все еще остается полезный термин.
- ↑ Haeckel, E. Das Protistenreich. Лейпциг, 1878 г. [1]
- ↑ Шехтер М. 2012. Эукариотические микробы .
 Амстердам, Академическая пресса.
Амстердам, Академическая пресса. - ↑ Маргулис Л. и Макменамин 1990. Брак по расчету. Науки 30 , 31-36.
- ↑ Маргулис Л. и Долан М.Ф. 2002. Ранняя жизнь: эволюция на докембрийской Земле . 2-е изд., Джонс и Бартлетт, Бостон. р89
- ↑ Маргулис Л. Шварц К.В. & Долан М. 1999. Разнообразие жизни: иллюстрированный путеводитель по пяти королевствам . Джонс и Бартлетт, Бостон, стр. 94. В этой работе авторы предлагают 19 типов для Протистов и называют это «Царство» «Протоктистой», термин, который, к сожалению, почти непроизносим.
- ↑ Дайер Б.Д. и Обар Р.А. 1994. Прослеживая историю эукариотических клеток . Колумбия, штат Нью-Йорк
Простая английская Википедия, бесплатная энциклопедия
Типичная животная клетка Эукариот — это организм со сложными клетками или отдельная клетка со сложной структурой. В этих клетках генетический материал организован в виде хромосом в ядре клетки.
Животные, растения, водоросли и грибы — все это эукариоты. Среди одноклеточных протистов есть и эукариоты. Напротив, более простые организмы, такие как бактерии и археи, не имеют ядер и других сложных клеточных структур. Такие организмы называются прокариотами.
Эукариот часто рассматривают как надцарство или домен.
Эукариоты эволюционировали в протерозойскую эру. Самый старый из известных вероятных эукариот — это Grypania , спиральная неразветвленная нить длиной до 30 мм. [3] Самые старые окаменелости Grypania происходят из железного рудника недалеко от Негони, штат Мичиган. Окаменелости были первоначально датированы 2100 миллионами лет назад, [4] , но более поздние исследования показали дату около 1874 миллионов лет назад. [5] Грипания просуществовала до мезопротерозойской эры.
Еще одна древняя группа — акритархи, которые считаются кистами или репродуктивными стадиями водорослевого планктона.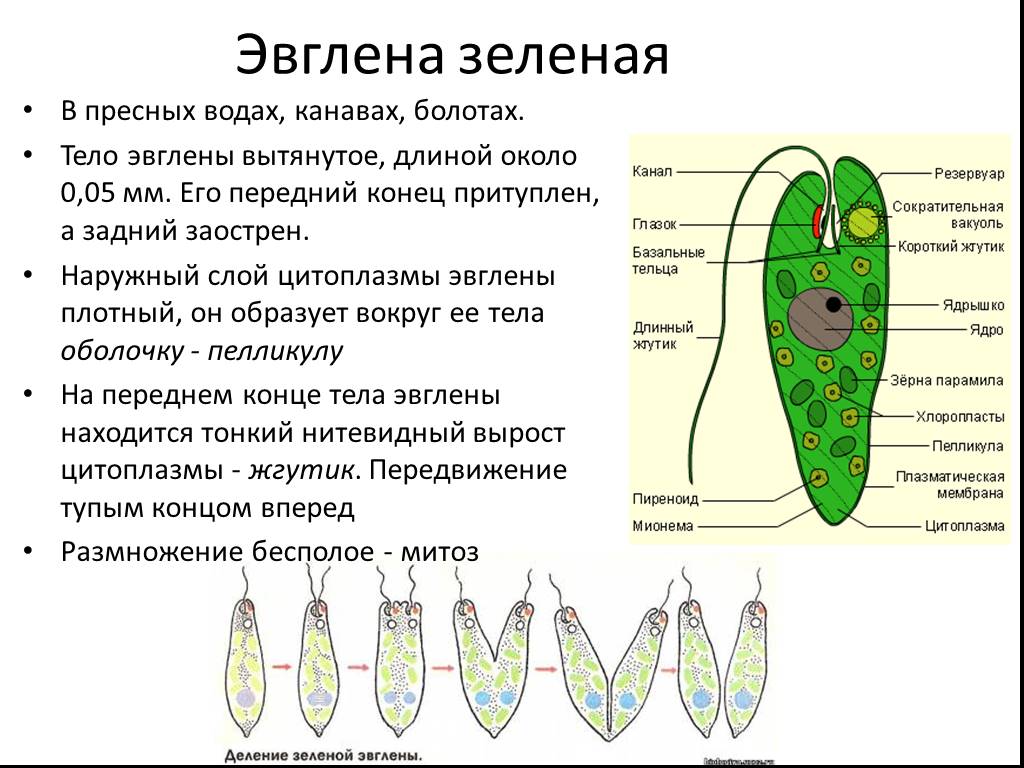 Они обнаружены 1400 миллионов лет назад, в мезопротерозойскую эру. [6] p57
Они обнаружены 1400 миллионов лет назад, в мезопротерозойскую эру. [6] p57
Классификация эукариот активно обсуждается, и было предложено несколько таксономий. Современные версии расходятся во мнениях относительно количества королевств. [7] Он несет ДНК так же, как и организм.
Эукариотические клетки обычно намного крупнее прокариот. Они могут быть в 10 раз больше. Эукариотические клетки имеют множество различных внутренних мембран и структур, называемых органеллами. Они также имеют цитоскелет. Цитоскелет состоит из микротрубочек и микрофиламентов. Эти части очень важны для формы клетки. ДНК эукариот собрана в пучки, называемые хромосомами, которые во время клеточного деления разделены микротрубочками. У большинства эукариот есть своего рода половое размножение посредством оплодотворения, которое прокариоты не используют.
Прокариоты не имеют пола, но могут передавать ДНК другим бактериям. Деление их клеток бесполое. Бактериальная конъюгация — это когда бактерии перемещают генетический элемент (часто плазмиду или транспозон) из одного в другой. [8] [9]
[8] [9]
Эукариоты имеют наборы линейных хромосом, расположенных в ядре, и число хромосом обычно типично для каждого вида.
Деталь эндомембранной системы и ее компонентовВнутренняя мембрана[изменить | изменить источник]
В эукариотических клетках есть много вещей, окруженных мембранами. Все они вместе называются эндомембранной системой. Простые мешочки, называемые везикулами или вакуолями, иногда образуются путем отпочковывания других мембран, точно так же, как дети делают пузыри своими игрушками. Многие клетки принимают пищу и другие вещества с помощью так называемого эндоцитоза. При эндоцитозе ближайшая к внешней стороне мембрана изгибается внутрь, а затем отщипывается, образуя везикулу. Многие другие органеллы, имеющие мембраны, вероятно, начинались как везикулы.
Ядро окружено двумя мембранными мембранами, в которых есть отверстия, через которые вещи могут входить и выходить. Из ядерной оболочки торчат вещи, похожие на трубки и листы. Они называются эндоплазматическим ретикулумом, который часто сокращается до ER. ER работает с перемещением белков и позволяет им созревать.
Они называются эндоплазматическим ретикулумом, который часто сокращается до ER. ER работает с перемещением белков и позволяет им созревать.
ER состоит из двух частей: грубой ER и гладкой ER. К шероховатой ЭР прикреплены рибосомы. Белки, образуемые рибосомами, прикрепленными к шероховатой ЭПР, попадают внутрь шероховатой ЭПР, называемой просветом. После этого они обычно переходят в везикулы, которые разрастаются и отщипываются от гладкого ЭПР. У большинства эукариот везикулы с белками внутри сливаются с грудами уплощенных везикул, называемых тельцами Гольджи, где белки внутри снова изменяются.
Везикулы иногда изменяются, поэтому они могут делать что-то очень хорошо. Это называется специализацией или дифференциацией. Например, в лизосомах есть ферменты, которые расщепляют пищу, поступающую из пищевых вакуолей, а в пероксисомах есть ферменты, которые расщепляют пероксид, яд, поэтому он больше не ядовит.
Многие простейшие имеют сократительные вакуоли, которые могут сливаться или отщипываться от наружной мембраны.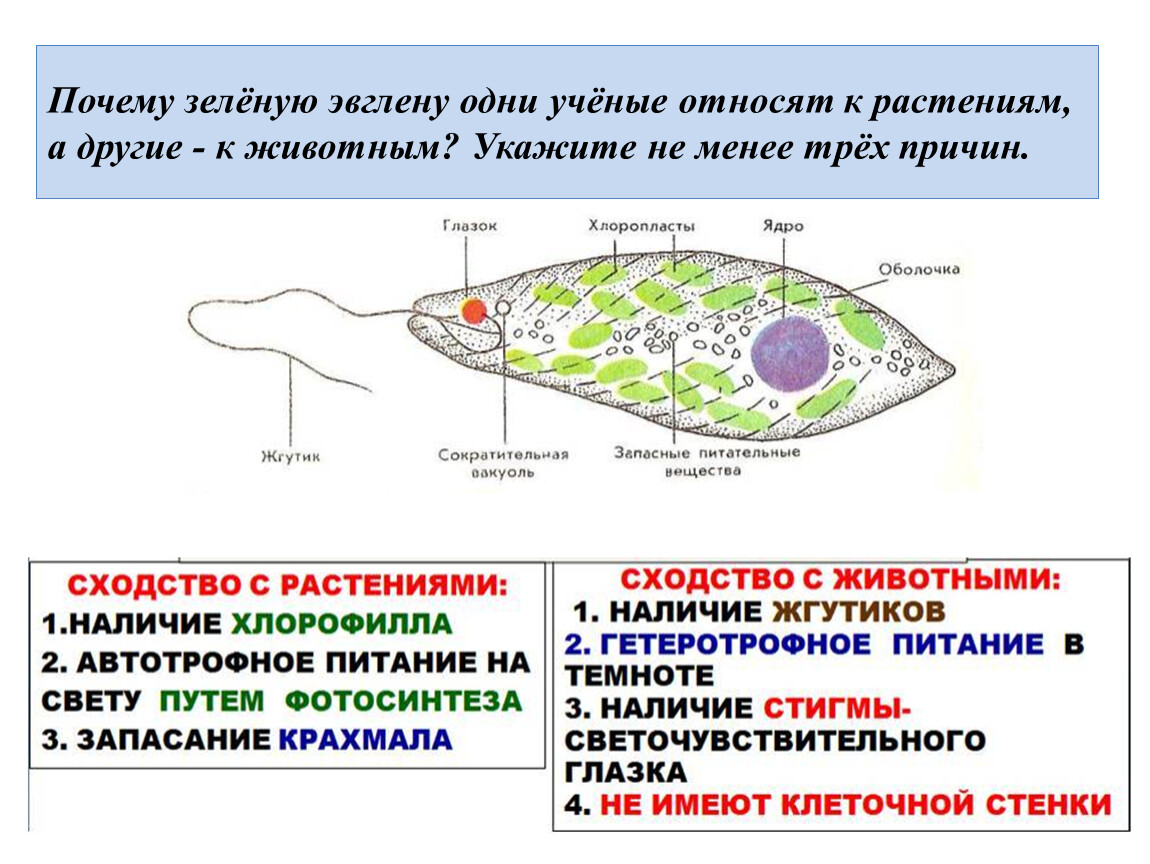 Сократительные пузырьки часто используются для получения и избавления от ненужной воды. Экструсомы выбрасывают вещества, которые заставляют хищников уходить или ловить пищу. У многоклеточных организмов гормоны часто вырабатываются в везикулах. У сложных растений большая часть внутренней части растительной клетки занята центральной вакуолью. Эта центральная вакуоль — главное, что поддерживает осмотическое давление, чтобы клетка могла сохранять свою форму.
Сократительные пузырьки часто используются для получения и избавления от ненужной воды. Экструсомы выбрасывают вещества, которые заставляют хищников уходить или ловить пищу. У многоклеточных организмов гормоны часто вырабатываются в везикулах. У сложных растений большая часть внутренней части растительной клетки занята центральной вакуолью. Эта центральная вакуоль — главное, что поддерживает осмотическое давление, чтобы клетка могла сохранять свою форму.
Поскольку клеточные органеллы эукариот имеют различное (полифилетическое) происхождение, возникает вопрос, является ли группа единой кладой или нет. [10] Определенно, что протисты — нет. [11] [12] Органеллы клетки представляют собой специализированные единицы, выполняющие четко определенные функции, такие как митохондрии и пластиды. Теперь совершенно ясно, что все или большинство этих органелл происходят от некогда независимых прокариот (бактерий или архей) и что эукариотическая клетка представляет собой «сообщество микроорганизмов», работающих вместе в «браке по расчету».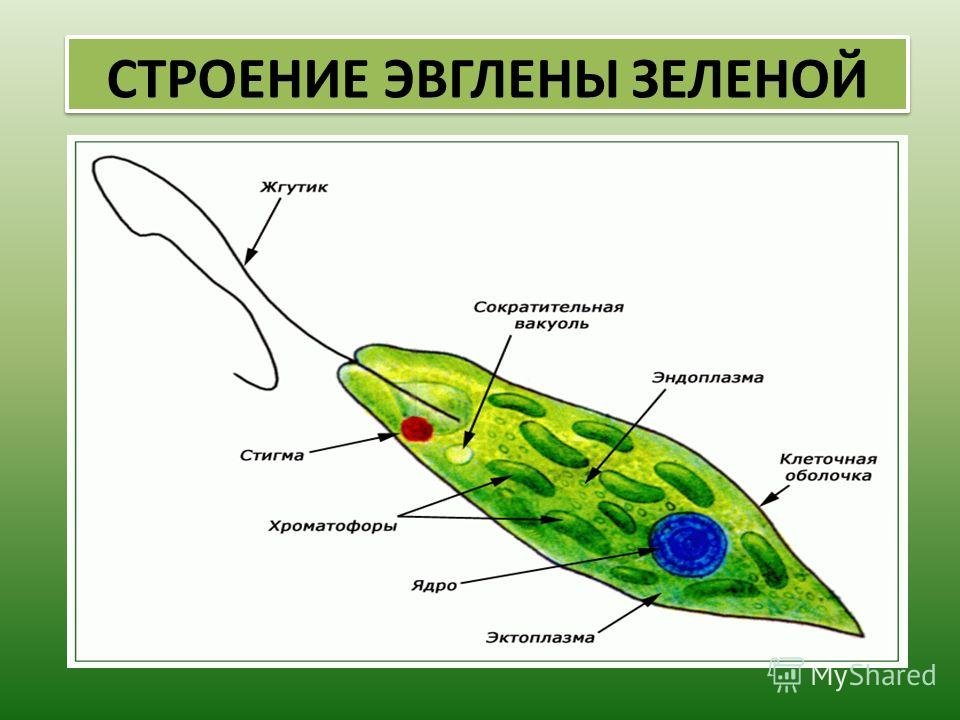 [13] Первые подобные события произошли между древними бактериями и привели к образованию двухмембранного класса, известного как грамотрицательные бактерии. [14] Поскольку к грамотрицательным бактериям относятся цианобактерии, это было первое из нескольких подобных событий в истории эукариот.
[13] Первые подобные события произошли между древними бактериями и привели к образованию двухмембранного класса, известного как грамотрицательные бактерии. [14] Поскольку к грамотрицательным бактериям относятся цианобактерии, это было первое из нескольких подобных событий в истории эукариот.
Роль архей[изменить | изменить источник]
Недавние исследования показывают, что «известный репертуар« специфичных для эукариот »белков архей [указывает], что клетка-хозяин археи уже содержит много ключевых компонентов, которые определяют сложность эукариотической клетки». [15]
Таксономия[изменить | изменить источник]
Protista — это группа различных одноклеточных организмов. Были предложены более точные таксономии, но ученые все еще обсуждают их. По этой причине Protista все еще полезно говорить об этих организмах. Одна современная схема классификации эукариот выглядит следующим образом: [16] [17]
| Опистоконты | Животные, грибы, хоанофлагелляты и др. |
| Амебозоа | Большинство лопастных амебоидов и слизевиков |
| Ризария | Foraminifera, Radiolaria и другие амебоидные простейшие. |
| Раскопки | Различные жгутиковые простейшие |
| Archaeplastida (или Primoplantae) | Наземные растения, зеленые водоросли, красные водоросли и глаукофиты |
| Хромальвеолаты | Гетероконты, гаптофиты, криптомонады и альвеоляты. |
Однако в 2005 г. были высказаны сомнения относительно того, являются ли некоторые из этих супергрупп монофилетическими, особенно Chromalveolata, [18] , а обзор 2006 г. отметил отсутствие доказательств для нескольких из предполагаемых шести супергрупп. [19]
Эукарии могут быть унифицированы только в том смысле, что клетки представляют собой сообщество, происходящее от бактерий и архей; мнения разнятся. Как и Protista, Eukarya может быть полифилетической сборкой, хотя и полезной.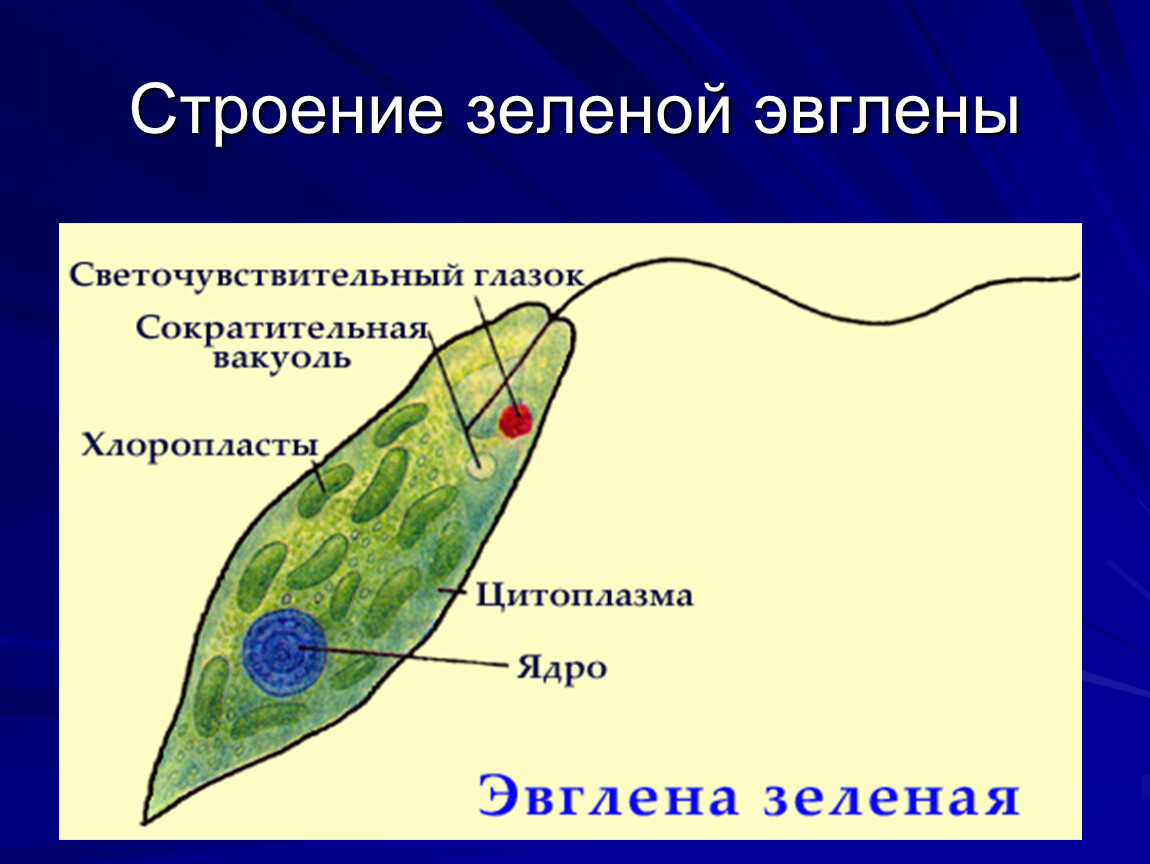 Однако, как было сказано выше, все ветви эукариот имеют половое размножение. [20] Это и общая организация ядра являются определяющими чертами. Эти два момента являются главным доказательством монофилетического происхождения. [21]
Однако, как было сказано выше, все ветви эукариот имеют половое размножение. [20] Это и общая организация ядра являются определяющими чертами. Эти два момента являются главным доказательством монофилетического происхождения. [21]
- Хронология жизни
- ↑ Сакагути М., Такисита К., Мацумото Т., Хашимото Т., Инагаки Й. (июль 2009 г.). «Отслеживание эволюции генов EFL в сообществе криптомонад-гаптофитов: отдельное происхождение генов EFL у гаптофитов, фотосинтетических криптомонад и гониомонад». Ген . 441 (1–2): 126–31. doi:10.1016/j.gene.2008.05.010. PMID 18585873.
- ↑ Adl SM, Simpson AG, Lane CE, Lukeš J, Bass D, Bowser SS, et al. (сентябрь 2012 г.). «Пересмотренная классификация эукариот» (PDF) . Журнал эукариотической микробиологии . 59 (5): 429–93. doi:10.1111/j.1550-7408.2012.00644.x. PMC 3483872. PMID 23020233. Архивировано из оригинала (PDF) 16 июня 2016 г. .

- ↑ Нолл, Эндрю Х. и др. 2006. Эукариотические организмы в протерозойских океанах. Философские труды Королевского общества B 361 (1470): 1023–38. [1]
- ↑ Хан Т.М. и Раннегар Б. 1992. Мегаскопические эукариотические водоросли из железной формации Негауни возрастом 2,1 миллиарда лет, Мичиган. Наука 257 (5067): 232–235. [2]
- ↑ Шнайдер Д.А. et al 2006. Возрастные ограничения для палеопротерозойского оледенения в районе озера Верхнее: возраст детритового циркона и гидротермального ксенотима для группы Чоколей, надгруппы хребта Маркетт. Canadian Journal of Earth Sciences 43 , 571-591. [3]
- ↑ Кларксон Э.Н.К. 1998. Палеонтология и эволюция беспозвоночных . Блэквелл, Оксфорд.
- ↑ Лукета, Стефан (2012). «Новые взгляды на мегаклассификацию жизни». Протистология . 7 (4): 218–237. ISSN 1680-0826.
- ↑ Райан К.Дж., Рэй К.
 Г., ред. (2004). Sherris Medical Microbiology (4-е изд.). Макгроу Хилл. стр. 60–4. ISBN 978-0-8385-8529-0 .
Г., ред. (2004). Sherris Medical Microbiology (4-е изд.). Макгроу Хилл. стр. 60–4. ISBN 978-0-8385-8529-0 . - ↑ Холмс Р.К. и Джоблинг М.Г.; и другие. (1996). Генетика: конъюгация. В: Медицинская микробиология Барона (Барон С. (4-е изд.). Университет Техасского медицинского отделения. ISBN 0-9631172-1-1 .
- ↑ Дайер Б.Д. и Обар Р.А. 1994. Прослеживая историю эукариотических клеток . Колумбия, штат Нью-Йорк
- ↑ Маргулис Л. и Долан М.Ф. 2002. Ранняя жизнь: эволюция на докембрийской Земле . 2-е изд., Джонс и Бартлетт, Бостон. р89
- ↑ Маргулис Л. Шварц К.В. & Долан М. 1999. Разнообразие жизни: иллюстрированный путеводитель по пяти королевствам . Джонс и Бартлетт, Бостон, стр. 94. В этой работе авторы предлагают 19 типов для Протистов и называют это «Царство» «Протоктистой», термин, который, к сожалению, почти непроизносим.
- ↑ Маргулис Л. и Макменамин 1990.
 Брак по расчету. Науки 30 , 31-36.
Брак по расчету. Науки 30 , 31-36. - ↑ Лейк, Джеймс А. 2009. Доказательства раннего прокариотического эндосимбиоза. Природа 460 : p967.
- ↑ Zaremba-Niedzwiedzka, Katarzyna et al 2017. Асгардские археи освещают происхождение эукариотической клеточной сложности. Природа (журнал) . 541 (7637): 353–358. [4]
- ↑ Адл С.М. и др. 2005. Новая классификация эукариот более высокого уровня с упором на таксономию простейших. Дж. Эукариот. Микробиол . 52 (5): 399–451. [5]
- ↑ Бурки Ф.; и другие. (2007). «Филогеномика перетасовывает эукариотические супергруппы». ПЛОС ОДИН . 2 (8): e790. Бибкод: 2007PLoSO…2..790B. doi:10.1371/journal.pone.0000790. PMC 1949142. PMID 17726520.
- ↑ Харпер Дж.Т.; Вандерс Э. и Килинг П. Дж. 2005. О монофилии хромальвеолятов с использованием шестибелковой филогении эукариот. Междунар. Дж. Система.
