ИБВВ РАН
П.П. Уморин
Роль хищников в устойчивом существовании нескольких видов водорослей.
Институт биологии внутренних вод им. И.Д. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
e-mail: [email protected]
Проведены эксперименты по совместному выращиванию зеленой Ankistrodesmus arcuatus Кorsch и диатомовой Diatoma elongatum (Lyngb.) водорослей с инфузориями Cyclidium glaucoma Ehrenberg и Paramecium caudatum Ehrenberg и без них в накопительных и проточных культурах.
В присутствии инфузорий оба вида водорослей успешно развивались, несмотря на потребление обоих видов инфузориями, тогда как в их отсутствие водоросль Ankistrodesmus arcuatus была подавлена и не развивалась.
Предполагается, что инфузории способствуют стабилизации сообщества конкурирующих жертв не только как хищники, но и выделяя прижизненно растворенный восстановленный азот в окружающую среду.
Ключевые слова: конкуренция, сосуществование, водоросли. .
.
Показать список литературы
М.М. Леонов
Фауна солнечников водоемов и водотоков лесостепи Cреднерусской возвышенности.
Воронежский государственный университет, 394000 Воронеж, Университетская площадь, 1
e-mail: [email protected]
Исследован видовой состав солнечников пресных вод лесостепи Среднерусской возвышенности на примере разнотипных водоемов Усманского бора Воронежской обл. и р. Усмань.
Обнаружено девять видов и две формы солнечников из четырех таксономических групп, пять видов оказались новыми для пресноводной фауны России.
Приведены иллюстрированные описания выявленных видов.
Наибольшее видовое разнообразие исследуемых протистов отмечено в болотах, что свидетельствует о более благоприятных для данной группы организмов условиях по сравнению с озерами и малой рекой.
Ключевые слова: солнечники, Heliozoa, Actinophryida, Centrohelida, Desmothoracida, Rotosphaerida, разнотипные водоемы, малая река..
Показать список литературы
Е. И. Миронова*, И.В. Телеш**, С.О. Скарлато*
И. Миронова*, И.В. Телеш**, С.О. Скарлато*
Планктонные инфузории Балтийского моря (обзор).
*Институт цитологии РАН, 194064 Санкт-Петербург, Тихорецкий проспект, 4
**Зоологический институт РАН, 199034 Санкт-Петербург, Университетская набережная, 1
e-mail: [email protected]
Представлены данные по видовому составу инфузорий Балтийского моря – солоновато-водного полузамкнутого водоема, значительную часть планктонной фауны которого составляют пресноводные виды.
За период исследования выявлено 789 видов инфузорий, из них 160 – типичные планктонные формы.
Даны экологические характеристики планктонных инфузорий и оценка их роли в продуктивности пелагических сообществ Балтийского моря.
Ключевые слова: инфузории, видовой состав, продукция, Балтийское море..
Показать список литературы
В.Г. Гагарин
Свободноживущие нематоды малой реки Центральной России.
Институт биологии внутренних вод им. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
Борок, Ярославская обл., Некоузский р-н
e-mail: [email protected]
Изучены видовой состав и количественное развитие нематод нижнего течения р. Сутка (притока Рыбинского водохранилища).
Обнаружены 54 вида из 11 отрядов.
Доминировали Dorylaimus stagnalis, Tobrilus gracilis и Eumonhystera filiformis.
Приведены описание и рисунки нового для науки вида Tridentulus pseudolemani sp. n.
Ключевые слова: малая река, свободноживущие нематоды, видовой состав..
Показать список литературы
Ж.П. Селифонова
Оithona brevicornis Giesbrecht (Copepoda, Cyclopoida) в акваториях портов северо-восточной части шельфа Черного моря.
Мурманский морской биологический институт Кольского научного центра РАН, 183010 г. Мурманск, ул. Владимирская, 17
e-mail: [email protected]
В пробах зоопланктона, собранных осенью 2005 и 2006 гг. в Новороссийском и Туапсинском портах северо-восточного шельфа Черного моря, в массовых количествах обнаружена новая для Черного моря циклопоида Oithona brevicornis Giesbrecht.
Предполагается, что данный вид попал в Черное море с балластными водами судов.
Ключевые слова: Oithona brevicornis, Сopepoda, портовые акватории, северо-восточный шельф, Черное море..
Показать список литературы
Л.В. Петрожицкая, В.И. Родькина
Пространственное распределение мошек (Diptera: Simuliidae) в бассейне горной реки Сема Северного Алтая.
Институт систематики и экологии животных СО РАН, 630091 Новосибирск, ул. Фрунзе, 11
e-mail: [email protected]
Проанализировано распределение преимагинальных фаз мошек в бассейне горной р. Сема (Северный Алтай) на основании количественных данных.
Исследования проведены на высотах 300?2 500 м над уровнем моря.
Выделены структурные элементы сообществ мошек в высотных поясах, обсуждается распределение мошек на ландшафтно-экологических профилях, рассмотрена последовательность изменений структуры сообществ мошек по течению реки.
Дана оценка значимости факторов среды, определяющих неоднородность населения мошек в бассейне р. Сема.
Сема.
Ключевые слова: бассейн горной реки, мошки, фауна, структура сообществ..
Показать список литературы
Н.Г. Косолапова, Д.Б. Косолапов
Разнообразие и распределение гетеротрофных наннофлагеллят в эвтрофном озере Неро.
Институт биологии внутренних вод им. И.Д. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
e-mail: [email protected]
Изучена фауна и обилие планктонных гетеротрофных наннофлагеллят (ГНФ), а также количественное распределение их основных пищевых объектов – бактерий в высокоэвтрофном мелководном оз. Неро (Ярославская обл.).
Обнаружены 70 видов и форм ГНФ, среди которых по числу видов доминировали воротничковые жгутиконосцы из отряда Choanoflagellida.
Жгутиконосцы и бактерии достигали высокого уровня количественного развития, характерного для продуктивных вод.
В сезонной динамике ГНФ выделялось два пика численности (в июне и сентябре) и один минимум – (в августе), который совпадал с максимальным развитием бактериопланктона.
Ключевые слова: гетеротрофные жгутиконосцы, бактериопланктон, видовой состав, численность, биомасса, эвтрофное мелководное озеро..
Показать список литературы
Е.И. Зуйкова, Н.А. Бочкарев
Характеристика пелагического зоопланктона крупных озер Тоджинской котловины (бассейн реки Большой Енисей, Тува).
Институт систематики и экологии животных СО РАН, 630091 Новосибирск, ул. Фрунзе, 11
e-mail: [email protected]
Исследована структура зоопланктона крупных озер Тоджинской котловины – Нойон-Холь, Шурам-Холь, Борзу-Холь, Маны-Холь, Кадыш и Тоджа.
Проведено сравнение видового состава, численности и биомассы зоопланктона в озерах.
Наибольшим видовым разнообразием характеризуются коловратки – от 10 до 16 видов, массового развития достигают Conochilus sp., Collotheca sp., Kellicottia longispina (Kellicott) и Asplanchna priodonta Gosse.
Веслоногие ракообразные представлены преимущественно Arctodiaptomus paulseni (Sars), Eudiaptomus graciloides (Lilljeborg), Cyclops scutifer Sars и Mesocyclops leuckarti Claus.
Среди ветвистоусых ракообразных доминируют Daphnia galeata Sars и Bosmina longispina Leydig.
Показано, что зоопланктонные сообщества озер Тоджинской котловины характеризуются высокой степенью сходства видового состава.
Выявлены факторы, формирующие таксономическую структуру пелагического зоопланктона в этой системе озер.
Ключевые слова: пелагический зоопланктон, видовой состав, численность, биомасса, озера Тоджинской котловины, Тува..
Показать список литературы
N.A. Yevdokimov*, M.V. Yermokhin**
Crustaceans of Zooplankton in Temporary Waterbodies of Saratov District on Territory of Different Natural Zones.
*N.I. Vavilov Saratov State Agrarian University, 410600 Saratov, ul. Sovetskaya, 60, Russia
**N.G. Tchernischevsky Saratov State University, 410026 Saratov, ul. Astrakhanskaya, 83, Russia
The zooplankton of temporary waterbodies of Saratov district include 68 crustacean species: fairy shrimps – 9, Cladocera – 25, Calanoida – 10, Cyclopoida – 24.
Regional and local patterns of formation of high species richness in temporary waterbodies are discussed.
The peculiarity of crustacean species composition in temporary waterbodies in intrazonal landscape of river valleys of the Don basin, steppe and desert steppe of Zavolzhiye are considered.
The number of crustacean species increases from north-west to south-east in temporary waterbodies of forest-steppe (33 species), steppe (39 species) and desert steppe (52 species).
The effect of irrigative systems on zooplankton species composition formation in temporary waterbodies is found.
Ключевые слова: temporary waterbodies, zooplankton, crustacean..
В.Б. Вербицкий, Т.И. Вербицкая, О.А. Малышева
Влияние различных температурных режимов на динамику численности и теплоустойчивость ветвистоусых ракообразных Ceriodaphnia quadrangula (O.F. Muller, 1785).
Институт биологии внутренних вод им. И.Д. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
e-mail: verb@ibiw. yaroslavl.ru
yaroslavl.ru
На основании экспериментальных исследований динамики численности и теплоустойчивости ветвистоусых ракообразных Ceriodaphnia quadrangula (O.F. Muller, 1785) в зависимости от величины и характера действия температурного фактора, а также полевых наблюдений и математических расчетов получены значения температурной зоны нормальной жизнедеятельности (17.0 – 25.0°С), а также зоны температурного «статического» оптимума (21.4 – 25.0°С) C. quadrangula.
На примере экспериментальных популяций C. quadrangula показано, что для создания оптимальных условий для роста и развития необходимо не только поддерживать диапазон оптимальных значений фактора на шкале толерантности, т.е «статический» оптимум, но и задавать оптимальные параметры динамических изменений фактора, или «динамический» оптимум.
Для C. quadrangula таким «динамическим» температурным оптимумом будет температура воды 24.0 + 1.1°С с непериодическим (ступенчатым) прогревом до 24.7 – 25.3°С в течение 5 – 8 сут.
Ключевые слова: зоопланктон, температурный оптимум, критический тепловой максимум, теплоустойчивость. .
.
Показать список литературы
Е.Н. Науменко
Зоопланктон в эстуариях разного типа (на примере Куршского и Вислинского заливов Балтийского моря).
Атлантический научно-исследовательский институт рыбного хозяйства и океанографии, 236000 Калининград, ул. Дм. Донского, 5
e-mail: [email protected]
На основании многолетнего мониторинга зоопланктона, проводившегося автором в Куршском и Вислинском заливах Балтийского моря, рассмотрены особенности структуры и функционирования сообществ зоопланктона.
Показано, что сообщества зоопланктона в эстуариях открытого (Вислинский залив) и закрытого (Куршский залив) типов различаются по видовой структуре, численности, биомассе и функционированию.
Сходство между сообществами зоопланктона этих эстуариев выражается в общности трофической структуры.
Сообщество зоопланктона Вислинского залива значительно больше подвержено влиянию градиента абиотических факторов, чем Куршского.
Механизмом, обеспечивающим устойчивость сообщества зоопланктона в эстуариях открытого типа, служит перенос энергии через всеядные виды и увеличение количества факультативных хищников в конечных звеньях трофической цепи.
Показано, что сообщество зоопланктона Вислинского залива характеризуется более высокой удельной скоростью продукции и скоростью ее оборота и использования.
Ключевые слова: Куршский и Вислинский заливы, зоопланктон, видовой состав, численность, биомасса, продукция, трофическая сеть..
Показать список литературы
М.В. Степанов*, В.Г. Терещенко**
Пространственное распределение тюльки в предзимовальный период в Рыбинском водохранилище.
*Управление Россельхознадзора по Вологодской области, 162602 г. Череповец, Вологодская область, ул. Парковая, д. 7
e-mail: [email protected], [email protected]
В предзимовальный период тюлька скапливается в северной части Шекснинского плеса.
Максимальные концентрации вида зафиксированы в верховьях плеса в местах с наибольшими глубинами и скоростями течений.
Отмечен пятнистый характер распределения рыб.
В пелагических скоплениях молоди рыб преобладала тюлька, составившая 95 – 100% численности (по уловам пелагическим тралом).
Доминировали две размерные группы тюльки: 40 – 60 мм и 70 – 80 мм. Местами наблюдалaсь обособленность этих размерных групп.
Ключевые слова: тюлька, пространственная структура, размерная структура, предзимовальный период..
Показать список литературы
Л.В. Балабанова, Д.В. Микряков, В.Р. Микряков
Реакция лейкоцитов карпа Cyprinus carpio L. на гормониндуцированный стресс.
**Институт биологии внутренних вод им. И.Д. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
e-mail: [email protected]
Изучено влияние кортизона на состав лейкоцитов крови карпа Cyprinus carpio L.
В лейкограмме опытных рыб после введения гормона зафиксировано уменьшение относительного количества лимфоцитов и увеличение нейтрофилов и бластных форм клеток.
Ключевые слова: кортизон, карп, лейкоциты..
Показать список литературы
В.А. Гремячих, И.И. Томилина, Л.П. Гребенюк
Влияние хлорида ртути на морфофункциональные показатели личинок Chironomus riparius Meigen (Dipterа, Сhironomidae).
Институт биологии внутренних вод им. И.Д. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
e-mail: [email protected]
Изучено действие различных концентраций хлорида ртути (HgCl2), ассоциированного с донными отложениями, на накопление металла личинками хирономид и изменение морфофункциональных показателей (смертность, рост, строение ротового аппарата).
Показано, что за период экспозиции животные накапливали Нg в количествах, превышающих ее содержание в грунте.
Cмертность и продолжительность метаморфоза личинок с увеличением концентрации металла в среде возрастала, линейные размеры фактически не менялись.
Наибольший процент деформаций общего числа исследованных структур приходился на долю антенн.
Ключевые слова: ртуть, личинки Chironomus riparius, накопление, деформации структур ротового аппарата..
Показать список литературы
И.В. Рыбакова*, Н.Л. Белькова**, Н.А. Лаптева*, Е.В. Суханова**
Адаптация молекулярно-генетических методов для изучения таксономического разнообразия микробных сообществ, ассоциированных с макрофитами.
*Институт биологии внутренних вод им. И.Д. Папанина РАН, 152742 пос. Борок, Ярославская обл., Некоузский р-н
**Лимнологический институт СО РАН, 664033 г. Иркутск, ул. Улан-Баторская, 3, а/я 278
e-mail: [email protected]
Использованный в работе комплекс молекулярно-генетических методов применен для изучения микробных сообществ, ассоциированных с макрофитами.
Метод ферментативного лизиса с фенол-хлороформной экстракцией оптимален для выделения суммарной бактериальной ДНК из проб перифитонных микроорганизмов и накопительных культур бактерий.
Рекомендована амплификация суммарной ДНК на высококонсервативных праймерах с использованием режима двухстадийной ПЦР.
Проведен анализ таксономического разнообразия микробного сообщества из смыва с тростника и накопительных культур, полученных на разных питательных средах.
Результаты показали большое разнообразие перифитонных микроорганизмов, ассоциированных с тростником, включающих представителей таких филогенетических линий как протеобактерии (α-, β-, γ- и δ-подгруппы), Bacteroidetes/Chlorobi, Chlamydiae/Verrucomicrobia и цианобактерии.
Невысокое разнообразие последовательностей в накопительных культурах представлено доминирующими с высоким процентом гомологии генотипами Cellvibrio и некультивируемых бацилл.
Ключевые слова: молекулярно-генетический подход, бактериоперифитон, таксономия, ген 16S рРНК, филогения..
Показать список литературы
Инфузории (CILIOPHORA, CILIATA) Среднего Приамурья (фауна, распространение, биология, морфо-экологическая эволюция) + «
Автореферат диссертации по теме «Инфузории (CILIOPHORA, CILIATA) Среднего Приамурья (фауна, распространение, биология, морфо-экологическая эволюция)»
Санкт-Петербургский государственный университет
На правах рукописи УДК 593.17
НИКИТИНА Людмила Ивановна
ИНФУЗОРИИ (СПЛОРНСШЛ, СИЛАХ А) почв СРЕДНЕГ О ПРИАМУРЬЯ
(фауна, распространение, биология, морфо-экологическая эволюция)
03.00.08 — Зоодсм ия
АВТОРЕФЕРАТ диссертации па соискание учетгой с1сисии доктора биологических наук
Сапкг-Пеюрбург — 1998
Работа выполнена в Хабаровском государственном педагогическом университете и Российском государственном педагогическом университете им. А. И. Герцена
А. И. Герцена
Доктор биологических наук , проф. В. В. Хлебович Доктор биологических наук, проф. Д. В. Осипов Доктор биологических наук, проф. Л. С. Колабская
Ведущее учреждение: Институт проблем экологии и эволюции РАН, лаборатория почвенной зоологии и экспериментальной энтомологии
Защита состоится февраля 1998 г. в 60 на заседании Диссертационного Совета Д. 063.57.22 по защите диссертаций на соискание ученой степени доктора биологических наук в Санкт-Петербургском государственном университете по адресу: 199034, Санкт — Петербург, Университетская набережная, 7/9, ауд. 133
С диссертацией можно ознакомиться в научной библиотеке СПбГУ.
Официальные оппоненты:
Автореферат разослан
Ученый секретарь Диссертационного Совета, кандидат биологических наук
Д. К. Обухов
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Почвенные инфузории (СШор1юга) — это неотъемлемый компонент естественных и антропогенных биогеоценозов. Они вляются пионерами заселения различных безжизненных пространств, ;след за микроорганизмами. Известно нахождение их в суглинистом суб-траге отвалов молодых техногенных систем в лесостепной зоне Мордкович,1993), в такырах на острове Барсакельмес в Аральском море Никитина, 1986), в почвах пустыни, неосвоенных полупустынных ссрозс-1ах Г олодной степи (Николюк, 1965), в гравийной пыли карьеров и других >иогеоценозах.
Они вляются пионерами заселения различных безжизненных пространств, ;след за микроорганизмами. Известно нахождение их в суглинистом суб-траге отвалов молодых техногенных систем в лесостепной зоне Мордкович,1993), в такырах на острове Барсакельмес в Аральском море Никитина, 1986), в почвах пустыни, неосвоенных полупустынных ссрозс-1ах Г олодной степи (Николюк, 1965), в гравийной пыли карьеров и других >иогеоценозах.
СШа’.а играют немаловажную роль в жизни почвы. Известна деятель-юсть почвенных инфузорий в формировании пищевых звеньев различных >иогеоценозов как естественных, так и антропогенных ландшафтов. Они финиматот активное участие в процессах продукции и деструкции органи-[есасого вещества на первых трофических уровнях. Это способствует естест-1енному восстановлению плодородия почв, которое в основном зависит от :корости круговорота веществ в них. В этом заключается очень важная функция, которую выполняют цилиаты, т.к. с их помощью и при их участии интенсивно протекают процессы почвообразования: почвенные биохимические превращения, заканчивающиеся подзолообразованием: изменялся дисперсность почвы; стимулируется биологическая фиксация азота и )асщспление целлюлозы (Гельцер, 1993;Гельцер, Ибадов, Мордкопич, 1980; Яковлев, 1997).
Как показывают результаты исследований, биомасса инфузорий может доставлять несколько десятков килограммов на гектар почвы, что немно-им меньше биомассы микроорганизмов (Николюк, Мавлянова, 1976; Ни-ситина,1987; Мирза-заде, 1989; Гельцер, 1993 и другие). Инфузории принимают участие в создании и обеспечении биогенных процессов. Они создают 1 почве большое количество биомассы, которая является составной частью )бщего живого вещества литосферы, накапливаемого микроорганизмами, -рибами, водорослями, высшими растениями и беспозвоночными живог-1ыми. Отмирая, ресничные непосредственно способствуют концентрации )рганических веществ в верхних горизонтах почвы, где они в основном и обитают. Несмотря на явную актуальность изучения простейших, как на-сопителей органики в почве, исследований по этому вопросу до сих пор яв-ю недостаточно.
Инфузории, входа в почвенные биоценозы, вступают в сложнейшие (заимоотношения с другими микроорганизмами и растениями, чаще всего 5лагоприятно влияя на них (Николюк, 1980; Николюк, Гельцер, 1972, 1993;
Николюк, Тапильская, 1976; Гельцер, 1984, 1993; Палеева, Верниченко, 1991; Модкович, 1993; Bamforth, 1980 и другие). Ресничные способствуют формированию микробных ценозов; взаимодействуя с микроорганизмами, инфузории активизируют их деятельность, в результате чего в почвенную среду выделяется целый ряд ферментов, которые активно участвуют в синтезе органических веществ, уже доступных растениям (Мавлянова, 1975).
Ресничные способствуют формированию микробных ценозов; взаимодействуя с микроорганизмами, инфузории активизируют их деятельность, в результате чего в почвенную среду выделяется целый ряд ферментов, которые активно участвуют в синтезе органических веществ, уже доступных растениям (Мавлянова, 1975).
Инфузории, среди которых важное место занимают представители рода Colpoda, являются перспективными тест-объектами для изучения влияния разнообразных биологически активных соединений (Гельцер, 1967; Гельцер, Гептнер, 1980; Гептнер, 1977; Ласкаускайте, 1982; Багданавичене и др., 1990; Сисакян и др., 1993; Никитина, Ножкина, 1996; Ekelund et al., 1992; Pratt, Xu, 1995 и другие).
Инфузории играют важную роль в методах биологической диагностики почв, составной частью которых является зоологический метод диагностирования почв, разработанный основателем российской почвенно-зоологической школы академиком М. С. Гиляровым (1965).
Зооиндикация с помощью инфузорий дает возможность проследить зависимость структуры цилиофауны от абиотических условий почвенной среды: гидротермического режима, наличия органического вещества, механического состава и физических свойств почвы, определяющих стратиграфию распределения простейших по профилю (Гельцер, 1991 и др. ). Кроме того, изучая комплексы инфузорий, морфо-физиологические характеристики доминантных видов, можно уточнить степень сапробности почв.
). Кроме того, изучая комплексы инфузорий, морфо-физиологические характеристики доминантных видов, можно уточнить степень сапробности почв.
Благодаря интенсивным исследованиям протозоологов уже выделено свыше 300 видов простейших из почв разных регионов бывшего СССР. Однако почвы Сибири, севера и Дальнего Востока остаются еще недостаточно изученными.
К началу наших исследований в литературе практически не было публикаций о почвенных инфузориях Приамурья. Совершенно недостаточно был исследован видовой состав и экология инфузорий, отсутствовали сведения о закономерностях распределения этих простейших в почвах Среднего Приамурья, ничего не было известно о численности и биомассе инфузорий в почвах естественных и антропогенных ландшафтов. Необходимо отметить, что Дальний Восток является центром фаунистического разнообразия России. Это связано с тем, что на юге Дальнего Востока не было сплошного ледового покрова даже в периоды максимального развития ледников, поэтому сохранялись условия для существования фаун, различных по своей экологии и генезису (Куренцов, 1965; Рябинин, Криволуцкий, 1989).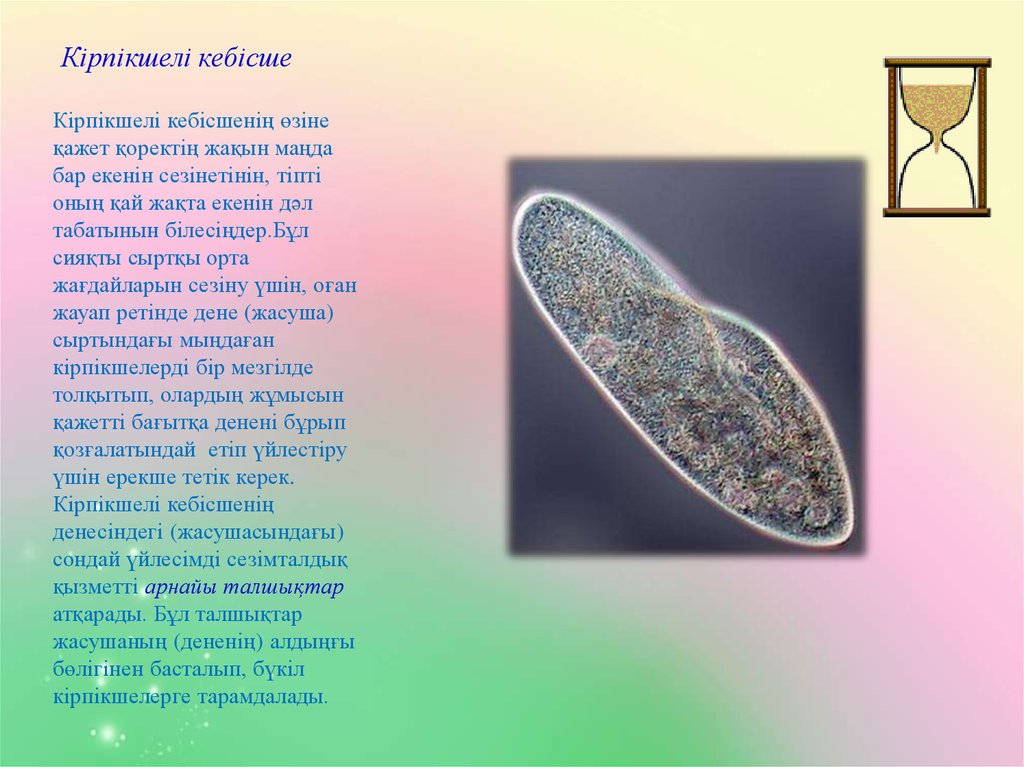 Изучение почвенных животных юга Дальнего Востока, и простейших
Изучение почвенных животных юга Дальнего Востока, и простейших
в частности, может дать ключ к пониманию того, каким было почвенное население страны до наступления ледникового периода.
Цель и задачи исследования. Исходя из большой практической и теоретической значимости почвенных инфузорий, а также их малой изученностью в почвах Дальневосточного региона, мы поставили своей целыо пополнить. критически пересмотреть и теоретически осмыслить оригинальные и литературные данные но видовому составу, географическому распространению и адаптивным особенностям инфузории из почв разных биогеоценозов как естественных, гак и антропогенных ландшафтов Среднего Приамурья. На этом основании обсудить вопросы, касающиеся закономерностей распределения фаунистического состава комплексов инфузорий в биогеоценозах данной территории. Реализация намеченной цели представлена следующими задачами:
1. Изучить видовой состав почвенных инфузорий и закономерности их распределения в разных биогеоценозах Среднего Приамурья.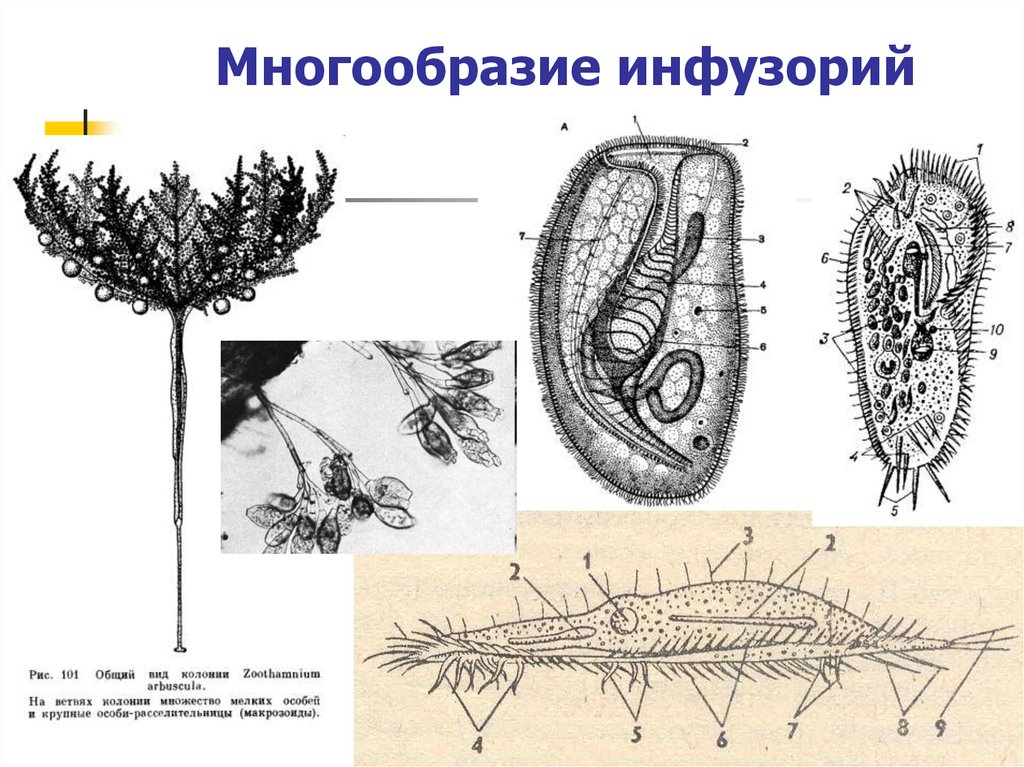
2. Произвести сравнительный анализ цилиофауны из почв разных регионов бывшего СССР, с применением методов фаунистического сходства, и выявить комплексы видов, специфичных для почв Приамурья.
3. Исследовать взаимосвязь инфузорий с ризосферой ряда дальневосточных растений.
4. Изучи 1Ь адагпивные особенности инфузорий, обитающих в разных ¡ппах почв Среднего Приамурья.
5. Обосновать возможные нуги морфо-экологической эволюции почв ен н ы х и н ф у з о р и й.
Защищаемые положения:
1. Видовой состав почвенных инфузорий Среднего Приамурья значительно богаче и разнообразнее, чем это дано в существующей литературе, что свидетельствует о недостаточной степени изученности этой группы почвенных протестов.
2. Особые почвенно-климатические условия Дальневосточного региона оказали влияние на закономерности жгпнсдеятельносш популяций почвенных инфузорий: сезонную динамику, глубину проникновения и распространение по почвенным горизонтам.
3. Для каждого типа почв существуют свои комплексы инфузорий с набором видов г еобионтов. 1 еофилов и геоксенов.
4. Результаты изучения комплексов видов инфузорий почвы и форм их адаптации к факторам среды обитания, в условиях разных биоклиматических зон, показали существование географических рас у целого ряда ви-
дов. Эти данные позволяют нам предложить гипотезу о возможных путях морфо-экологической эволюции ресничных инфузорий.
Научная новизна. Впервые осуществлена инвентаризация видового состава почвенных инфузорий из разных биогеоценозов Среднего Приамурья. В результате было выявлено 102 вида и разновидностей инфузорий, из которых 32 вида являются новыми для почвенной фауны бывшего СССР. На основании применения методов фаунистического сходства проведен сравнительный анализ видового состава цилиат из почв различных регионов и республик бывшего СССР. Установлено, что в состав почвенной фауны Приамурья входят виды, обитающие в самых разных географических и климатических районах.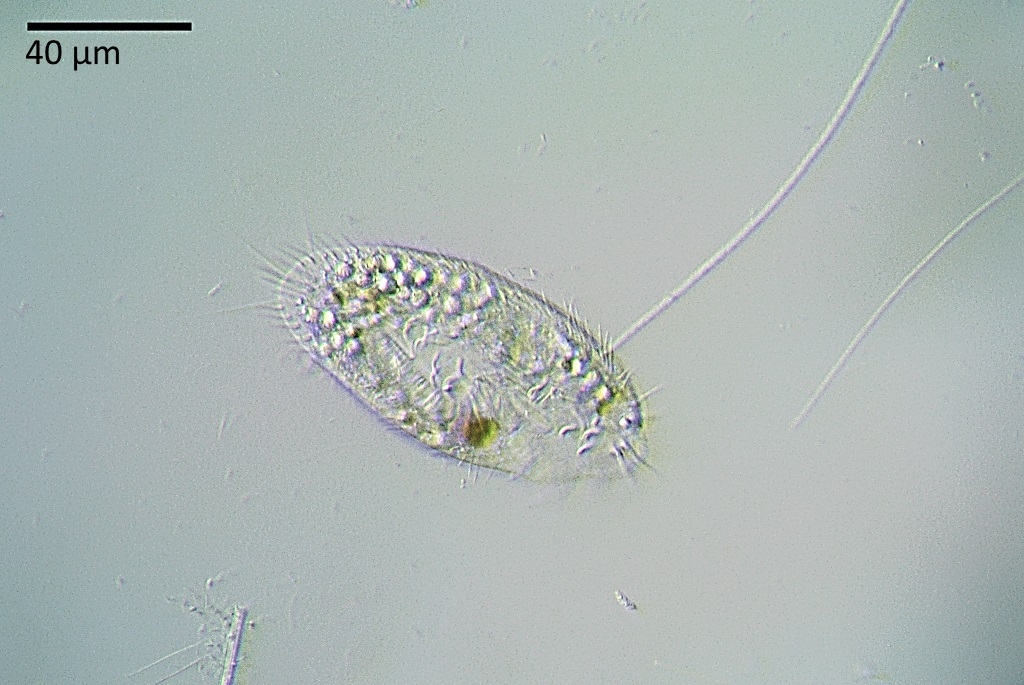 Это положение соответствует разнообразию животного и растительного мира, возникшего в результате отсутствия сплошного ледового покрова в четвертичном периоде, на данной территории.
Это положение соответствует разнообразию животного и растительного мира, возникшего в результате отсутствия сплошного ледового покрова в четвертичном периоде, на данной территории.
Определен комплекс видов инфузорий, характеризующий бурые почвы Среднего Приамурья. Данный комплекс является специфичным для исследуемых почв. Установлено, что наиболее богаты по видовому разнообразию почвы Большехехцирского заповедника, которые можно использовать в качестве резервата генофонда и биоразнообразия цилиат.
Впервые изучена сезонная и глубинная динамика популяций почвенных инфузорий в естественных и антропогенных ландшафтах юга Дальнего Востока.
При изучении видового разнообразия инфузорий и их плотности в прикорневой системе ряда сельскохозяйственных и эндемичных растений (соя, рододендрон даурский, аралия маньчжурская и др.) и в почве, удаленной от корней, отмечаем наличие хорошо выраженного ризосферного эффекта (аккумуляция инфузорий в ризоплане и ризосфере корней), однако не для всех культур.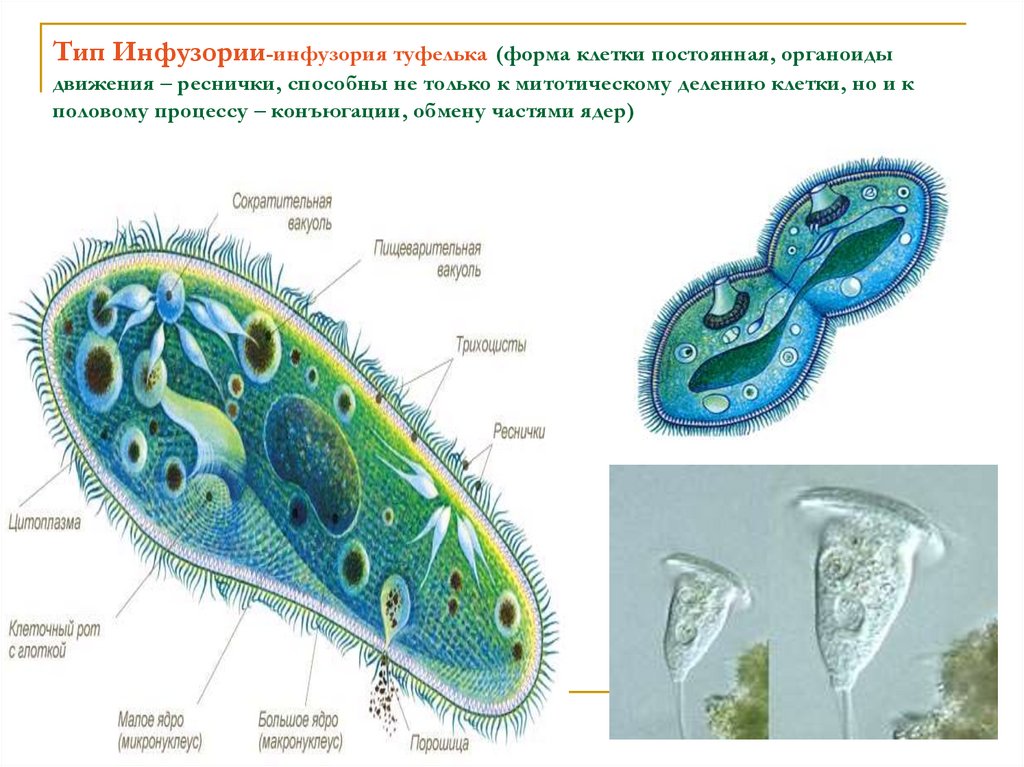 Ингибирование развития инфузорий, уменьшение их численности, происходит в корневой системе томатов, что связано со специфическими веществами (томатин), которые они выделяют в почву.
Ингибирование развития инфузорий, уменьшение их численности, происходит в корневой системе томатов, что связано со специфическими веществами (томатин), которые они выделяют в почву.
На основе изучения почвенных инфузорий, адаптивных особенностей видов, выделенных из разных типов почв и биогеоценозов Среднего Приамурья, а также сравнительного фаунистического анализа с почвенными инфузориями других регионов, мы предлагаем концепцию формирования у отдельных видов инфузорий географических рас и концепцию одного из путей морфо — экологической эволюции инфузорий: переход от обитания в водоемах Земли к обитанию в микроводоемах почвы. Данные гипотезы базируются на морфологических, физиологических и других отличиях между популяциями инфузорий из разных географических зон и наличии морфо-
логических адаптации, которые обеспечивают устойчивое существование популяций в условиях микроводоемов разных типов почв.
Теоретическое и практическое значение. Модифицированный нами метод количественного учета почвенных инфузории позволяет более быстро и эффективно производить анализ почвенных образцов на предмет выявления в них инфузории, их количественного определения и последующего культивирования для изучения жизненных циклов и морфологических показателей (Никитина. 1497). Полученные результаты по видовому разнообразию почвенной цилиофауны, особенно почв Большехехцирского заповедника, использованы в программе по изучению бпорадпообразия животного п растительного мира заповедников.
1497). Полученные результаты по видовому разнообразию почвенной цилиофауны, особенно почв Большехехцирского заповедника, использованы в программе по изучению бпорадпообразия животного п растительного мира заповедников.
Почвенные инфузории (Colpoda cucullas, Uroleptus piscis, Stylonychia sp. и другие) являются хорошими тест объектами для лабораторных, учебных и экологических исследований. Виды инфузорий и их комплексы можно использовать в качестве биоиндикаторов в системе экологического мониторинга.
В связи с тем, что в литературе отсутствует описание и каталог почвенных инфузорий Среднего Приамурья, большое практическое значение имеет диагностическое описание выявленных видов инфузорий н издание иллюстрированно!! монографии, которую могут использовать в своей работе цитологи, экологи, руководители детских экологических центров и преподаватели вузов и школ (Никитина, 1997).
Апробация работы. Основные результаты исследований докладывались на итоговых научных конференциях преподавателей ХГПУ с 1986 г. по 1997 г., на X Всесоюзном совещании по проблемам почвенной зоологии в 1991 г. (г. Новосибирск), на 5 Всесоюзном съезде протозоологов в 1992 г. (г. Витебск), а также на Межвузовской конференции «Герцетювские чтения» в ГТПУ им.А.И.1 ерпена и 1997 г. (г. Санкт-Петербург), на заседании лаборатории «Экология животных» Института водных и экологических проблем РАН в 1997 г. (Хабаровск), на конференции «Производственные и полевые практики — основа эколого-профессиональиой подготовки молодого специалиста» (Хабаровск. 1997).
по 1997 г., на X Всесоюзном совещании по проблемам почвенной зоологии в 1991 г. (г. Новосибирск), на 5 Всесоюзном съезде протозоологов в 1992 г. (г. Витебск), а также на Межвузовской конференции «Герцетювские чтения» в ГТПУ им.А.И.1 ерпена и 1997 г. (г. Санкт-Петербург), на заседании лаборатории «Экология животных» Института водных и экологических проблем РАН в 1997 г. (Хабаровск), на конференции «Производственные и полевые практики — основа эколого-профессиональиой подготовки молодого специалиста» (Хабаровск. 1997).
Внедрение. Результаты исследования видового состава почвенных инфузорий использованы в программе «Стратегия сохранения биологического разнообразия Сихоте-Алиня в пределах Хабаровского края».
Публикации. Основные результаты исследований по теме диссертации изложены в 31 опубликованной работе, из них одна монография «Почвенные инфузории Среднего Приамурья» (1997 года).
Структура и объем диссертации. Диссертация изложена на 361 страницах машинописного текста и состоит из введения, 8 глав, заключения, выводов, списка литературы ( 401 название) и приложения, которое содержит описание и рисунки выявленных видов и разновидностей инфузорий. В диссертации имеется 36 таблиц и 36 рисунков (не считая приложения).
В диссертации имеется 36 таблиц и 36 рисунков (не считая приложения).
Выражаю глубокую благодарность доктору биол. наук, проф. кафедры зоологии РГПУ им. А.И. Герцена — K.M. Сухановой за консультации во время работы над диссертацией.
СОДЕРЖАНИЕ РАБОТЫ
ПРИРОДНЫЕ УСЛОВИЯ РАЙОНА ИССЛЕДОВАНИЯ
Почвенные пробы отбирались в окрестностях г. Хабаровска (Болыыехехцирский заповедник, Воронеж-1, агробиологическая станция ХГПУ, пос. Некрасовка, пос. Березовка, левый берег Амура, Уссурийские острова, с. Петропавловка, устье реки Талги), в Нанайском районе (окрестности пос. Троицкое, пос. Славянка) и Комсомольском районе: оз. Болонь и окрестности серно-кислотного завода г. Комсомольска-на-Амуре (рис. 1).
По почвенно-климатическому районированию территория Средне-амурской низменности входит в состав особой Дальневосточной муссон-ной почвенно-биоклиматической зоны. Однако впоследствии было установлено, что данная территория имеет континентальный климат с муссон-ными чертами, т. к. находится под влиянием Евроазиатского материка и Тихого океана (Герасимов, 1933; Витвицкий, 1969). Огмечено, что климат Дальнего Востока существенно отличается от климата других частей России и не имеет аналогов (Колосков, 1962).
к. находится под влиянием Евроазиатского материка и Тихого океана (Герасимов, 1933; Витвицкий, 1969). Огмечено, что климат Дальнего Востока существенно отличается от климата других частей России и не имеет аналогов (Колосков, 1962).
Отличительным свойством климата является наличие в летние месяцы влажно-тропической погоды с суточными температурами воздуха более 22° С и влажностью воздуха свыше 80% (табл. 1).
Таблица 1
Средние месячные температуры воздуха (в градусах) и количество осадков (в мм)_
Месяцы 1 2 3 4 5 6 7 8 9 10 11 12 годовая
темпера -тура -12,3 -17,2 -8,5 +3,1 111 17.4 22,1 20,0 13,9 +4.7 -8.1 -18,5 +14
кол-во осадков 19 13 23 42 61 80 115 123 89 46 37 24 672
Рис. 1 Карта района исследований. • Указаны места взятия почвенных образцов
В конце лета и начале осени нередко из Японского моря приходят тропические циклоны — тайфуны, что приводит к летне-осенним разливам.
Метеорологический режим зимних месяцев обусловлен резким преобладанием северо-западных континентальных ветров, в результате чего зима оказывается очень холодной, сухой и малоснежной.
Из-за соседства с самым холодным районом Северного полушария и проникновения морского воздуха с полуполярного Охотского моря климат здесь более суров зимой, чем на территориях, расположенных на тех же широтах в Европейской части страны.
Почвенные образцы для выявления инфузорий были взяты из следующих типов почв:
1. Горные буро-таежные иллювиально-гумусовые, характерные для папоротниковых и зеленомошных пихтово-еловых лесов, 700 м над уровнем моря (Большехехцирский заповедник).
2. Бурые горно-лесные, характерные для елово-широколиственного леса с разнотравно-папоротниковым покровом (Большехехцирский заповедник, 400 м над уровнем моря).
3. Бурые лесные оподзоленные, характерные для смешанных лесов (Большехехцирский заповедник, Воронеж-1, агростанция ХГПУ, пос. Некрасовка, пос. Березовка, с. Троицкое, окрестности г. Комсомольска-на-Амуре).
4. Луговые глееватые почвы, характерные для пойменных лугов Приамурья (левый берег Амура, с. Петропавловка, устье реки Талги).
Петропавловка, устье реки Талги).
ФАУНА ИНФУЗОРИЙ БИОГЕОЦЕНОЗОВ
СРЕДНЕГО ПРИАМУРЬЯ
Наиболее подробно изучен видовой состав почвенных инфузорий в биогеоценозах трех ландшафтов: Большехехцирский заповедник, Воронежские высоты и агробиологическая станция ХГПУ.
Почвенные инфузории биогеоценозов Большехехцирского заповедника
Заповедные территории являются эталоном экосистем и резерватом генофонда животного и растительного мира (Криволуцкий, Покаржевский, 1985). Важнейшим компонентом наземных экосистем считается почва и, следовательно, организмы, ее населяющие. В Большехехцирском заповеднике еще не все элементы природных комплексов изучены равномерно, а исследование почвенной цилиофауны совсем не проводилось до начала наших изысканий.
Нами были выявлены инфузории из почвенных образцов трех биоценозов на северном склоне хребта Большой Хехцир: осиново-березовый лес,
хвойно-тироколиственный лес и пихтово-еловый лес. С южной стороны исследовались: ивово-вейниковая и дубово-березовая растительные ассоциации.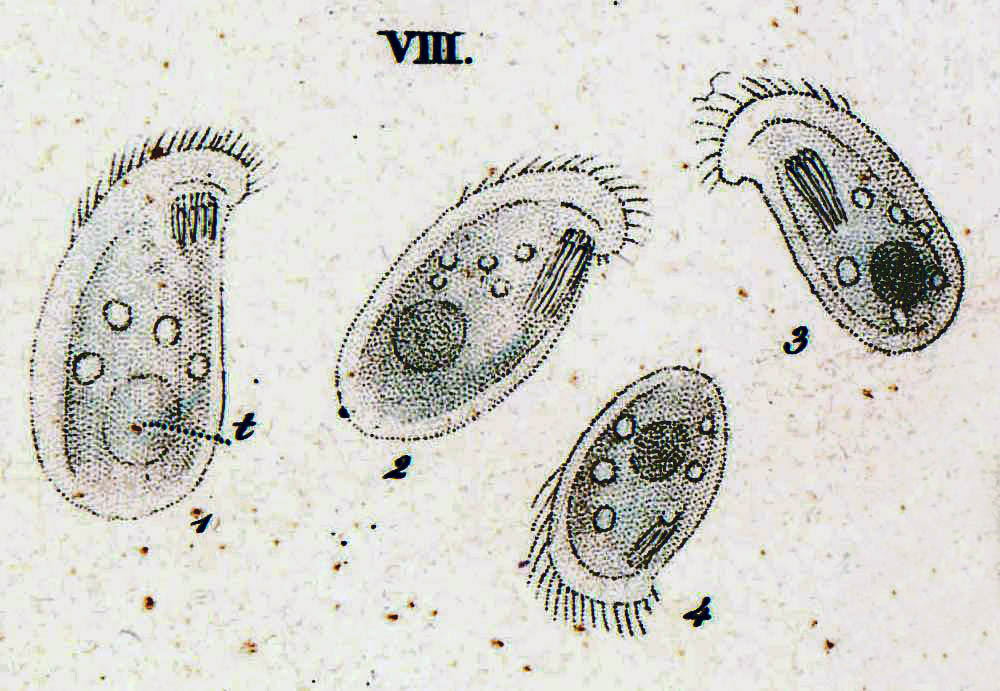
Анализируя видовой состав инфузорий, выделенных из разных биогеоценозов Большехехцирского заповедника, следует отмстить, что наиболее многочисленным оказался класс KINETOFRAGMINOPHORA — 31 вид цилиат, относящихся к 14 семействам и 8 отрядам. Этот класс является и самым разнообразным по числу отрядов, семейств и видов. Из столь большого числа семейств только одно семейство (Colpodidae) характеризовалось максимальным видовым разнообразием — 6 видов. Из семейства Enchelyidae было зарегистрировано 4 вида инфузорий. В трех семействах (Spathidiidae. Trachcliidae. Amphileptidae) было выявлено по 3 вида инфузорий. Остальные семейства были представлены 1 — 2 видами. Больше всего видов было выявлено из отряда Haptorida — 11, а из отряда Colpodida — 7 видов.
Класс OLIGOHYMENOPHORA, представленный 17 видами инфузорий из 7 семейств и 3 отрядов. Причем, из отряда Hymenostomatida было выявлено максимальное количество видов — 8, входящих в 3 семейства: Tetrahymenidae, Glaucomidae и Paramecidae. Из отряда Scuticociliatida выявлено 5 видов, a Peritrichida — 4 вида.
Из отряда Scuticociliatida выявлено 5 видов, a Peritrichida — 4 вида.
Класс POLYHYMENOPHORA представлен 22 видами инфузорий из 9 семейств и 3 отрядов. По количеству видов и семейств наиболее предеia»n-тельным оказался отряд H\potrichida — 3 семейства и 10 видов. Наибольшим видовым разнообразием характеризовалось семейство Oxytrichidae…..
6 видов инфузорий.
При анализе видового состава инфузорий из почвенных образцов разных биогеоценозов Большехехцирского заповедника выделен комплекс видов, которые являются общими для всех исследованных почвенных разностей: Colpoda cucullus. С. maupasi, С. sleini, Drcpanomonas revoluta. Chilodonella uncmata, Colpidium colpoda, Urouema marinum, Vorticella microstoma, Strombidium sp.l и Aspidisca costata. Всего 10 видов, которые составляют индикационный комплекс для почв Большехехцирского заповедника.
Видовой состав почвенных инфузорий Воронежа-1
Воронежские высоты (100 — 160 м над уровнем моря) занимают северные окрестности г. Хабаровска и являются лесопарковой зоной. Расположены они на правом высоком берегу Амура. Воронежские высоты сложены из триасовых осадочных и вулканогенных пород (Варнавский и др., 1991). В конце сороковых годов на месте вырубок были произведены лесопосадки, в основном, сосны обыкновенной. За прошедшие годы на сопках разросся широколиственно-хвойный лес. В настоящее время это место активно
Хабаровска и являются лесопарковой зоной. Расположены они на правом высоком берегу Амура. Воронежские высоты сложены из триасовых осадочных и вулканогенных пород (Варнавский и др., 1991). В конце сороковых годов на месте вырубок были произведены лесопосадки, в основном, сосны обыкновенной. За прошедшие годы на сопках разросся широколиственно-хвойный лес. В настоящее время это место активно
посещается хабаровчанами как место для отдыха, а также для сбора дико-росов и лекарственных растений (грибы, орех маньчжурский, виноград даурский, папоротник орляк, липа амурская и др.). Почвы Воронежских высот на склонах сопок бурые оподзоленные (Климат Хабаровска, 1981).
Видовой состав почвенных инфузорий выявлялся в следующих биотопах Воронежа-1: сосново-леспедецивый, рододендрово-тростниковый, липово-дубовый и орехово-разнотравный.
Наибольшее количество видов было выявлено из класса KINETOFRAGMINOPHORA — 22 вида инфузорий (51,1%). Из класса OLIGOHYMENOPHORA — только 8 видов (17,8 %), а из класса POLYHYMENOPHORA — 13 видов (32,3 %). В первом классе явное лидирование по числу выявленных видов принадлежит отряду Haptorida: 7 видов, что составляет 16,3 % от общего числа выявленных видов инфузорий. Этот отряд был представлен инфузориями из трех семейств: Enchelyidae (3 вида), Tracheliidae (3 вида) и Didiniidae (1 вид). Следующий по численности видов, отряд Colpodida (16,3 % от общего числа выявленных видов инфузорий): 6 видов из семейства Colpodidae и 1 вид из семейства Cyrtolophosididae. Из остальных отрядов было зарегистрировано по 1 — 2 вида.
В первом классе явное лидирование по числу выявленных видов принадлежит отряду Haptorida: 7 видов, что составляет 16,3 % от общего числа выявленных видов инфузорий. Этот отряд был представлен инфузориями из трех семейств: Enchelyidae (3 вида), Tracheliidae (3 вида) и Didiniidae (1 вид). Следующий по численности видов, отряд Colpodida (16,3 % от общего числа выявленных видов инфузорий): 6 видов из семейства Colpodidae и 1 вид из семейства Cyrtolophosididae. Из остальных отрядов было зарегистрировано по 1 — 2 вида.
В классе Oligohymenophora — 4 вида инфузорий относятся к отряду Scuticociliatida, 3 вида из отряда Hymenostomatida и 1 вид из отряда Peritrichida.
В классе Polyhymenophora — самым многовидовым оказался отряд Hypotrichida — 9 видов инфузорий (20,9 % от общего числа выявленных видов). Из семейства Oxytrichidae зарегистрировано 5 видов, из семейства Holostichidae — 3 вида инфузорий и 1 вид из семейства Aspidiscidae. Из остальных отрядов (Heterotrichida, Oligotrichida) было выявлено по 2 вида.
Сравнивая по видовому составу все исследуемые биотопы Воронежских высот, мы выделили комплекс видов инфузорий, которые встречаются во всех почвенных образцах биотопов. К ним относятся следующие виды: Colpoda fastigata Kahl, С. maupasi Enriquez, Colpoda steini Maupas, Chilodonella sp., Tetrahymena sp., Uronema marinum Dujardin, Vorticella microstoma Ehrenberg и Strombidium sp. Коэффициент видового сходства определяли по формуле Жаккара (Чернов, 1975) для всех исследованных биотопов Воронежских высот.
Необходимо отметить, что в почвенных образцах исследуемых биотопов Воронежских высот было выявлено видов инфузорий в 1,6 раза меньше, чем в биогеоценозах Большехехцирского заповедника. Уменьшение видового разнообразия почвенных инфузорий, по-видимому, связано в пер-
вую очередь со значительной антропогенной нагрузкой, какую испытывает широколиственно-хвойный лес Воронежских высот. Видовой состав почвенных инфузорий агробиологической станции ХГПУ
Агробиологическая станция Хабаровского педуниверситета находится в черте города. Заложена она в пятидесятые годы.
Заложена она в пятидесятые годы.
Почвы агробиологической станции буро-подзолистые. По агрохимическим показателям для них характерно рН солевое в кислом интервале 5 -5,5. Кислая реакция почвенной среды является одной из причин низкого содержания в буро-подзолистых почвах агростанции доступной для раае-ний фосфорной кислоты. Кроме того, периодическое переувлажнение почвы способствует переходу подвижных форм фосфорной кислоты в труднодоступные для растений соединения. Кислая реакция среды угнетает деятельность тарификаторов, от которых зависит образование и накопление в почве доступных для растений соединений азота (нитратов).
Мы анализировали почвенные образцы, на предмет выявления инфузорий, с полей трехрядного севооборота (томаты, капуста, огурцы) и сои. как с ризосферы растений, так и с удаленной от корней почвы
В почвенных образцах полей с разными сельскохозяйственными к>ль-|урами был выявлен 51 вид инфузорий, что составляет 50.0 % от общего числа видов, зарегистрированных для Среднего Приамурья.
Наибольшее количество Ридов инфузорий относится к классу К1\’1-ТО1711АС}Л’П.\»0Р11(ЖА — 25 видов или 49 % от зарегистрированных в данном биогеоценозе цилиат. Самыми представительными были отряды Со1росйс1а и Иар1отк1а — по 8 видов (по 32 % соответственно на каждый отряд). Из семейств этого класса наиболее многовидовыми были Со1росУс!ае -6 видов, НпсЬеМйае — 5 видов, а также СЫ1о(ЗопеШс1ае. представленное 4 видами. Впервые были выявлены виды рода СЬаепеа (семейство БпсЬе1у1Йае) и рода СгапоШспсПит (семейство БраШсШс1ае).
Класс ОЬЮОНУМЕМОРНОИА представлен 12 видами (23,5 % от выявленных видов в агроценозе). Самым представительным был отряд Нутпепо51:отаНс1а — 6 видов или 50 % от видового состава данного класса. Отряд Реп1пс1ш1а включал 4 вида инфузорий. Самыми многовидовыми семействами в этом классе были семейства ТсЧгаЬутепкке и УогйссШйае — т них было выявлено 4 и 3 вида соответственно. Впервые были зарегистрированы инфузории из рода Орегайапа (семейство Орегси1агпс1ае).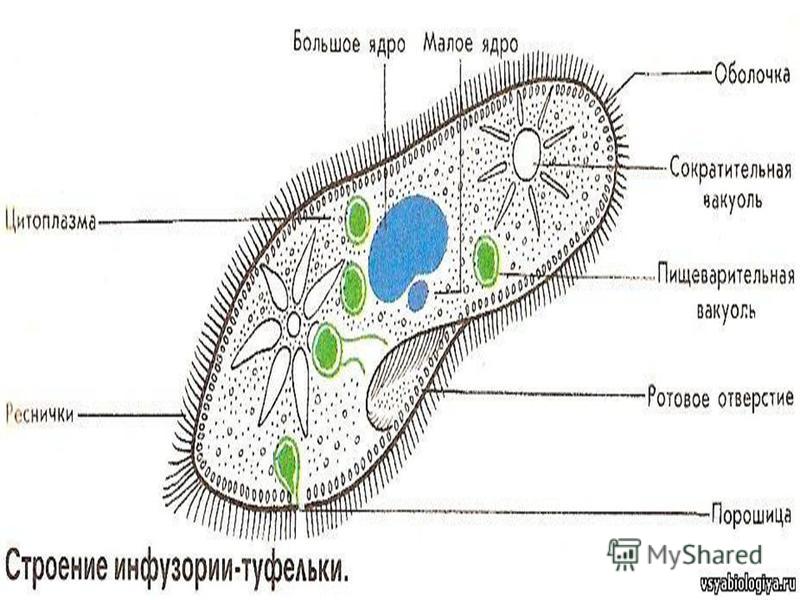
К классу Ро1уЬутпепор1юга относится 14 видов или 27,5 % от всего списка цилиат этого биогеоценоза. Соотношение видов к отрядам равно 14:2. Это самый высокий показатель из всех трех классов. Отряд Нуро1псЫс!а
был представлен 12 видами, что составляет 85,7 %от видов цилиат данного класса. Из них, на долю семейства Oxytrichidae приходится 50 % этого отряда (6 видов инфузорий). Новыми родами были Paruroleptus (семейство Holostichídae) и Lacazea (семейство Keronidae). Надо отметить, что представители семейства Keronidae впервые были зарегистрированы в почвах агроценоза, так что и само семейство является новым для почв Среднего Приамурья.
Отличительной особенностью видового состава почвенных инфузорий биоценозов агростанции является уникальность видового состава, т.к. в ризосфере сельскохозяйственных растений были выявлены инфузории не встреченные в других исследуемых почвенных образцах. К таким инфузориям относятся: Chaenea teres, Chaenea sp., Enchelys pupa, Cranotheridium sp. , Cyrtolophosis mucicola, Opercularia sp., Lacazea sp., Oxytricha fallax. Таким образом, мы можем говорить о специфичности почвенной цилиофауны агроценозов.
, Cyrtolophosis mucicola, Opercularia sp., Lacazea sp., Oxytricha fallax. Таким образом, мы можем говорить о специфичности почвенной цилиофауны агроценозов.
Наибольшее количество видов инфузорий было выделено из почвенных образцов Большехехцирского заповедника — 67 видов, или 74,4 % от всех видов, зарегистрированных в исследуемых трех биогеоценозах. Далее следуют почвы агробиологической станции — 51 вид, или 56,6 %. И на третьем месте по числу видов находятся почвы Воронежских высот — 43 вида, что составляет 47,7 %.
Общими видами для почв трех биогеоценозов были 20 видов. На их долю приходится 22,2 %. Наибольшая доля участия в почвенных процессах принадлежит инфузориям семейства Colpodidae (отряд Colpodída). Из 7 зарегистрированных видов данного таксона общими для почв исследуемых биогеоценозов являются 5 видов.
Нужно также отметить, что еще 29 видов инфузорий или 32,2 % являлись общими для почв двух «з трех исследуемых биогеоценозов. Таким образом, более 50 % видов почвенных инфузорий являются наиболее распространенными в почвах окрестностей г. Хабаровска. Ведущая роль здесь принадлежит инфузориям таких семейств как Colpodidae, Chilodonellidae, Uronematidae, Pleuronematidae, Vorticellidae, Strombidiidae, Holostichídae и Oxytrichidae.
Хабаровска. Ведущая роль здесь принадлежит инфузориям таких семейств как Colpodidae, Chilodonellidae, Uronematidae, Pleuronematidae, Vorticellidae, Strombidiidae, Holostichídae и Oxytrichidae.
Для сравнения видового состава почвенных инфузорий использовали коэффициент Жаккара (Чернов, 1975). Наибольшее значение данного показателя характерно для почвенной цилиофауны Большехехцирского заповедника и Воронежских высот. В почвенных образцах двух рассматриваемых ландшафтов общих видов оказалось 35. Соответственно индекс видового сходства составляет 0,46, т. е. почти 50 % . В почвах заповедника и аг-
росганции выявлено 33 общих вида инфузорий. Коэффициент видового сходства биогеоценозов заповедника и агростанцин несколько ниже — 0.39. Самый низкий показатель — между биогеоценозами Воронежских высот и агростанции. В них было зарегистрировано общих видов только 21. Индекс равен 0.28.
РАСПРОСТРАНЕНИЕ И ДИНАМИКА ПОЧВЕННЫХ
ИНФУЗОРИЙ В БИОГЕОЦЕНОЗАХ СРЕДНЕГО ПРИАМУРЬЯ
Задачей настоящего раздела исследования явилось изучение и выявление закономерностей количественного развития инфузорий как в почвах естественных биогеоценозов, так и подвергнутых антропогенному воздействию. характерные для разновидностей бурых лесных почв: в них нмеехся только один пик численности видов — осенний. Причем, если п почве хвошго-широколисгвенного леса численность видов весной и легом различается незначительно, то в почве пихтово-слового леса весной было зарегистрировано 14 видов инфузорий, летом — 18, а осенью 27.
характерные для разновидностей бурых лесных почв: в них нмеехся только один пик численности видов — осенний. Причем, если п почве хвошго-широколисгвенного леса численность видов весной и легом различается незначительно, то в почве пихтово-слового леса весной было зарегистрировано 14 видов инфузорий, летом — 18, а осенью 27.
Глубина проникновения инфузорий в почву в лесных биоценозах в основном ограничивается 10-15 см. Аккумуляция инфузорий весной и осенью наблюдается в подстилке и верхнем почвенном горизонте, а летом — в слое 2 — 10 см.
Влияние антропогенного фактора на почвенных инфузорий прослеживается в осиново-березовом биогеоценозе (осиновый биотоп) и связано в первую очередь с уменьшением видового разнообразия, а также наличием видов — полисапробов к которым относятся инфузории семейства Оху1:псЫ(1ае.
Рассматривая вертикальную зональность распределения почвенных инфузорий, необходимо отметить, что с подъемом местности над уровнем моря происходит постепенное увеличение видового состава, примерно, до 400 м, а затем наблюдается снижение видового разнообразия и плотности популяций.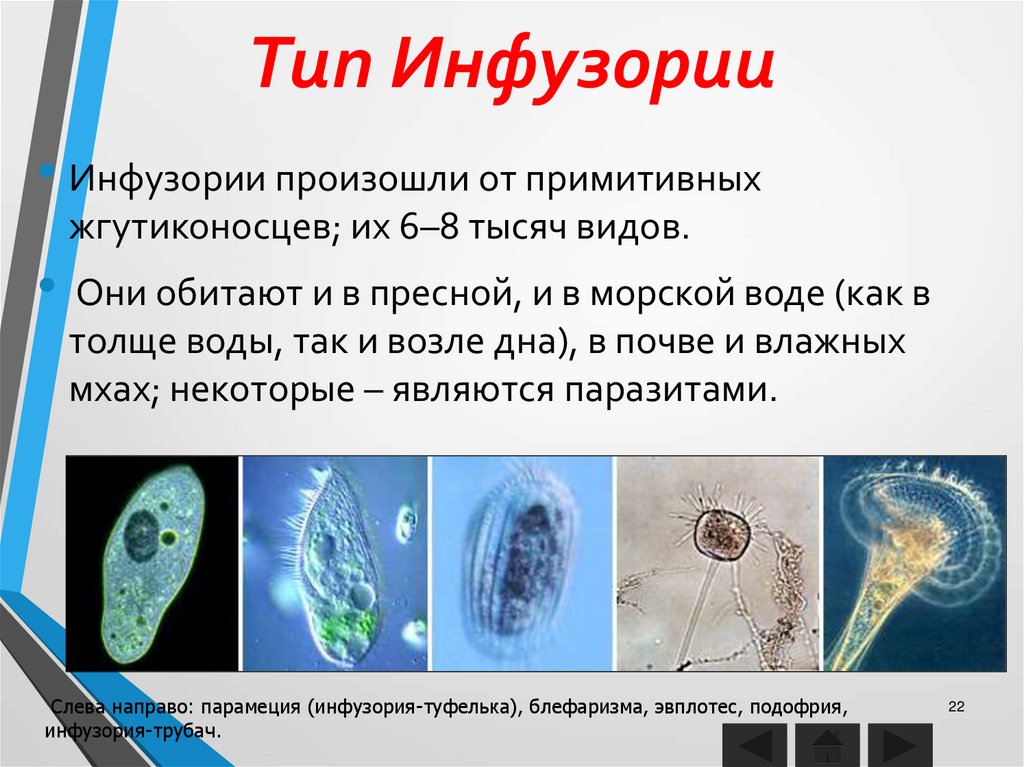 Кроме того, цилиофауна очень тесно связана с растительностью: чем разнообразнее видовой состав растений (хвойно-
Кроме того, цилиофауна очень тесно связана с растительностью: чем разнообразнее видовой состав растений (хвойно-
широколиственный лес) тем больше в почве обитает видов. В хвойных биотопах, в связи с закисленностью почв, разнообразие видового состава инфузорий уменьшается.
В разных биогеоценозах Воронежских высот, находящихся под прессом антропогенного воздействия, мы выявили следующие закономерности: плотность популяций инфузорий довольно низкая, не превышает 2800 экз./г в почвенных образцах рододендрово-тростникового биотопа и 600 экз./г в сосново-леспедецивом биоценозе; глубина проникновения инфузорий в почву ограничивается 10-15 см, максимальная численность видов и обилия инфузорий приходится на конец лета- начало осени.
Нами изучались закономерности распространения инфузорий в почвах нескольких полей агробиологической станции ХГПУ в течение вегетационного периода. Необходимо отметить, что в период исследования (лето 1992-1994 г.) в почву вносились как минеральные удобрения (мочевина, сульфат аммония и др. ) так и органические в виде торфокомпоста на основе птичьего помета. Однако в течение лета 1995 и 1996 годов удобрения в почву не вносились, в связи с их отсутствием.
) так и органические в виде торфокомпоста на основе птичьего помета. Однако в течение лета 1995 и 1996 годов удобрения в почву не вносились, в связи с их отсутствием.
Мы изучали также обилие инфузорий в почвенных образцах ризосферы некоторых сельскохозяйственных растений. В 1 г воздушно-сухой почвы, снятой с корневой системы сои, было зарегистрировано до 300000 клеток. Такая высокая плотность инфузорий обусловлена наличием разнообразных клубеньковых бактерий, которые и привлекали инфузорий — бактериофагов.
Необходимо также отметить, что почвы агроценоза характеризуются сильно выраженным цилиостазисом. Он проявлялся в очень низкой численности как видов инфузорий, так и их обилия в свежей почве по сравнению с воздушно-сухой. Возможно, что низкое содержание активных форм инфузорий вызвано кислотным характером почвенного раствора (рН 5 -5,5), поскольку кислая реакция среды угнетает деятельность многих бактерий. Некоторые авторы (Николюк, Гельцер, 1972) считают, что цилиоста-зис вызван бактериальным токсикозом почв, в результате чего большинство инфузорий находится в состоянии цист покоя. Скорее всего, что оба эти фактора, слагаясь, приводят к развитию цилиостазиса.
Скорее всего, что оба эти фактора, слагаясь, приводят к развитию цилиостазиса.
В пахотных почвах агроценоза наблюдается летнее увеличение численности видов инфузорий и их обилия, а со второй половины лета — снижение видового разнообразия и численности; глубина проникновения инфузорий в почвах агроценозов достигает 25 см, что обусловлено разными агротехническими мероприятиями. Аккумуляция инфузорий в весенний период происходит в верхних слоях почвы (0 — 2-5 см), летом в горизонте 5 -
10 см, а к осени наблюдается миграция цнлиат опять в верхние слои; внесение удобрений положительно сказывается на видовом разнообразии почвенных инфузорий и их обилии.
Таким образом, рассмотрев динамику и распространение инфузорий в почвах естественных и антропогенных биогеоценозов Среднего Приамурья, мы можем сделать следующее заключение: в связи с особыми природно-климатическими условиями Среднего Приамурья, закономерности распространения инфузорий в бурых лесных, бурых оподзоленных и лугово-I леевых почвах отличаются от закономерностей, установленных для почв европейской части бывшего СССР и других регионов. Специфичность закономерностей состоит в следующем:
Специфичность закономерностей состоит в следующем:
1 — в постоянном увеличении видового разнообразия инфузорий и их обилия от весны к лету, т.е. наличие только одного пика численности в почвах естественных биогеоценозов, в отличие от двух пиков численности (весеннего и осеннего), характерных для других регионов бывшего СССР;
2 — в агроценозах также наблюдается только один пик численности инфузорий, но летний. Отсутствие осеннего пика численности обусловлено сильным промывным режимом пахотного слоя почвы во второй половине лета в условиях своеобразного муссонного климата Среднего Приамурья:
3 — глубина проникновения инфузорий в почву ограничивается горизонтом 15 — 20 см, в агроценозах увеличивается до 25 см. Аккумуляция инфузорий весной наблюдается и верхнем почвенном слое (0 — 2-5 см), летом 1! более нижележащих слоях (5-10 см) а осенью — опять в верхнем почвенном горизонте:
4 — для почв Среднего Приамурья в большой степени характерен ци-лиостазис, который обусловлен кислотным характером почвенного раствора;
5 — биомасса, накапливаемая почвенными инфузориями, идет в общий баланс органики и способствует повышению плодородия почв.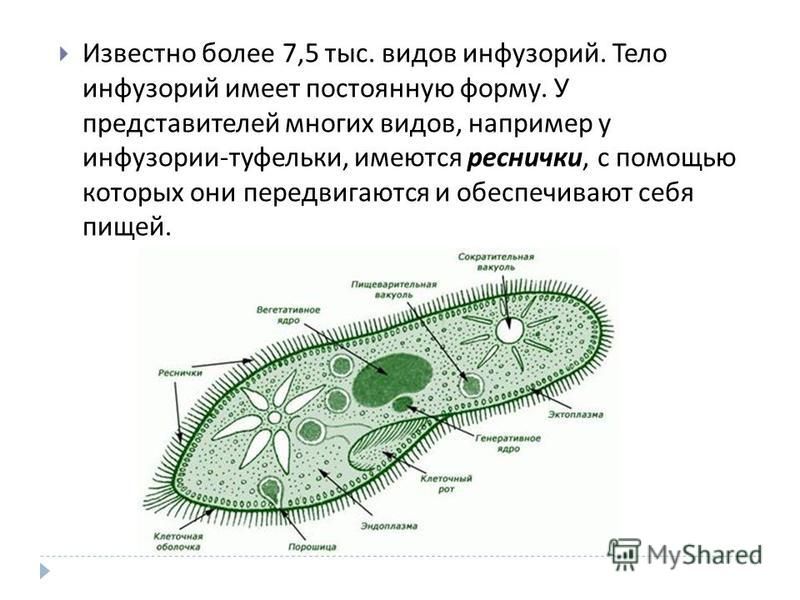
ЗООГЕОГРАФИЧЕСКИЙ АНАЛИЗ ФАУНЫ
ПОЧВЕННЫХ ИНФУЗОРИЙ СРЕДНЕГО ПРИАМУРЬЯ
Изучение состава фауны почвенных инфузории имеет большой зоо-географичсский интерес. Это вызвано тем, что данный ра йон характеризуется своеобразными природно-климатическими условиями, а также интересной геологической историей. В частности, отсутствием на территории Среднеамурской низменности сплошного ледового покрова даже в периоды максимального развития ледников.
В связи с тем, что Дальний Восток является центром фаунистического разнообразия животных и растений, а Среднее Приамурье характеризуется высокой степенью эндемичности как фауны ряда групп многоклеточных животных, так и флоры, можно было бы ожидать некоторой специфичности и фауны почвенных инфузорий. Однако сопоставление списка видов приамурских инфузорий и инфузорий других географических районов показывает, что видовой состав фауны не столь уж и специфичен, как мы предполагали. Поскольку видовой состав фауны почвенных инфузорий в Приамурье более изучен (Никитина, 1987, 1996, 1997), нежели в других регионах бывшего СССР, то отличительные особенности в цилиофауне имеются.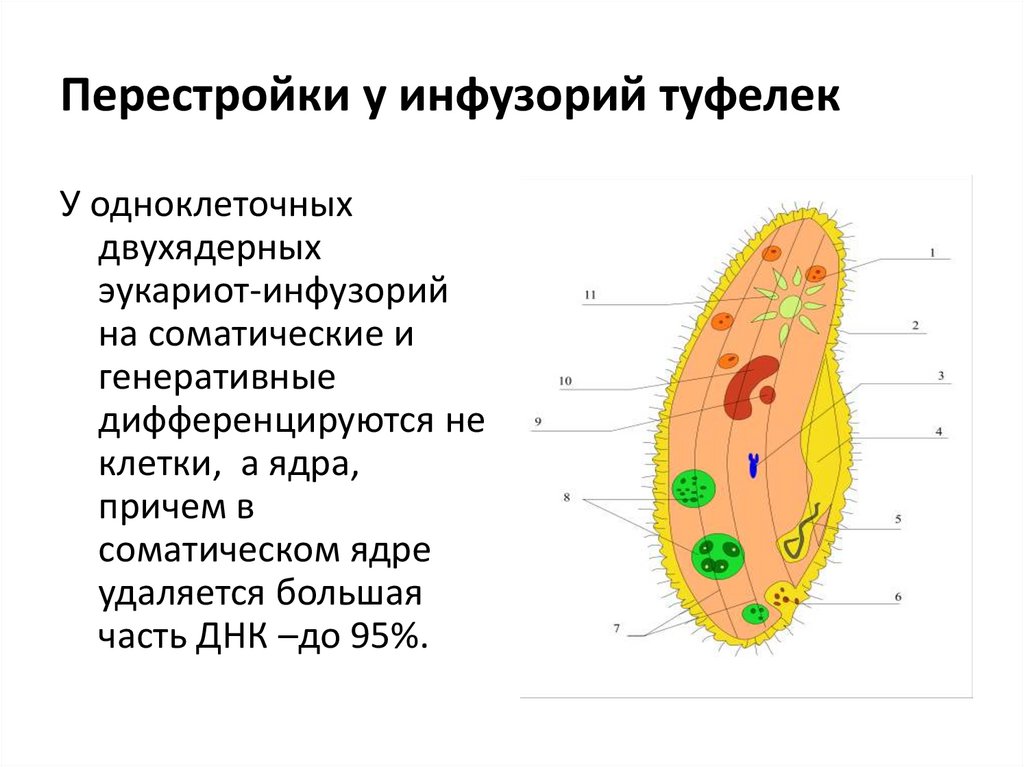
В результате многолетней работы по изучению видового состава почвенных инфузорий разных биогеоценозов Среднего Приамурья было выявлено 102 вида и разновидностей инфузорий, относящихся к 3 классам (Kinetofragminophora, Oligohymenophora и Polyhymenophora), 7 подклассам (Gymnostomata, Vestibulifera, Hypostomata, Suctoria, Hymenostomata, Peritricha, Spirotricha) и 14 отрядам, согласно классификации Corliss (1979).
Из 102 видов и разновидностей инфузорий нами точно определены — 67 видов, т.е. 65,7 % от всего списочного состава. Среднеамурской низменности. На сегодняшний день — это один из наиболее изученных районов в отношении почвенных инфузорий на территории бывшего СССР. Однако остается еще не определенными более 35 % видов инфузорий. Мы считаем, также, что и обнаруженные виды не являются пределом фаунистического разнообразия для почв Среднего Приамурья. Уверены, что дальнейшие исследования почвенных образцов еще пополнят список видов почвенных инфузорий.
Сравнивая видовой состав почвенной цилиофауны Среднего Приамурья с имеющимися литературными данными, можно сделать вывод о том, что в результате наших исследований значительно пополнился список видов инфузорий, обитающих в почвенных условиях.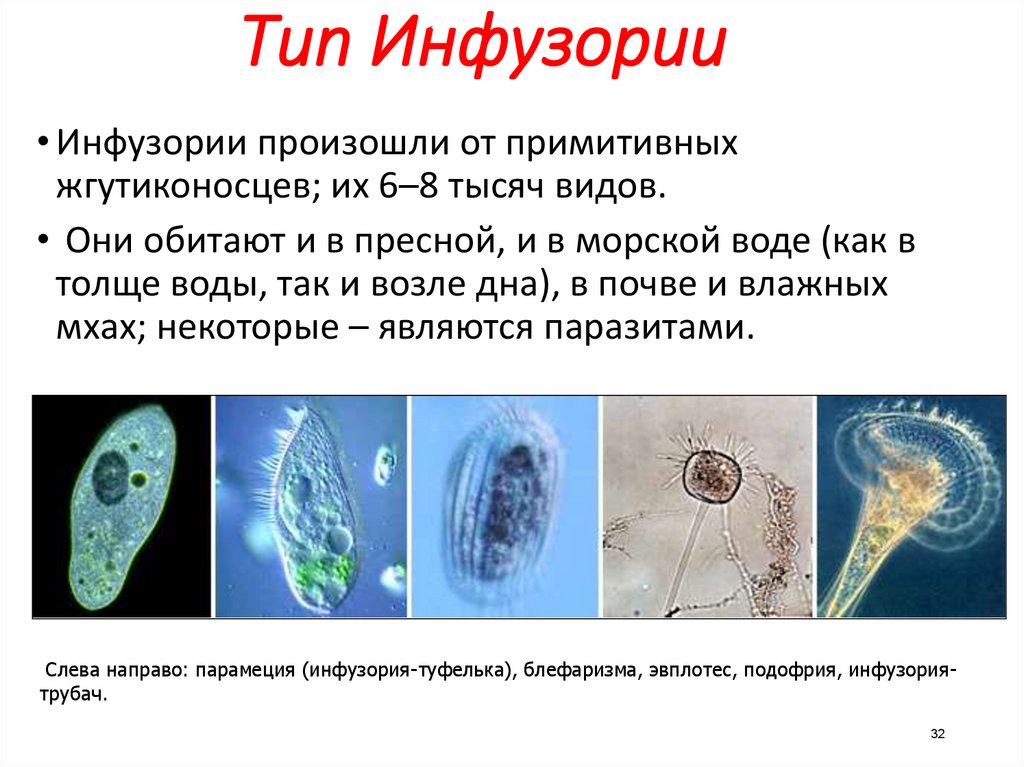 Новыми для почв бывшего СССР оказались 32 вида инфузорий. В основном выявленные виды ранее были описаны как пресноводные инфузории. Но поскольку почвы Среднего Приамурья характеризуются переувлажненностью, то почвенные микроводоемы стали для них естественной средой обитания.
Новыми для почв бывшего СССР оказались 32 вида инфузорий. В основном выявленные виды ранее были описаны как пресноводные инфузории. Но поскольку почвы Среднего Приамурья характеризуются переувлажненностью, то почвенные микроводоемы стали для них естественной средой обитания.
Нами выявлены 19 родов и 9 семейств инфузорий, не описанных ранее как почвенные. Выделены даже инфузории, входящие в отряды, не зарегистрированные для почв бывшего СССР. К таким отрядам относятся Karyorelictida Corliss, 1974 и Suctorida Claparede et Lachmann, 1858.
Также впервые нами отмечены в почвенных образцах Большехехцир-ского заповедника инфузории, относящиеся к подклассу Suctoria Claparede
et Lachmann,T858.
Результаты исследований почвенных инфузорий разными авторами (Бродский. 1935; Гельцер 1993; Лепинис и др., 1973; Chardcz, 1967; Foissner. 1988; Berger et Foissner, 1989 и другие) показали, что фауна инфузорий различных географических районов обладает некоторой степенью общности видового состава.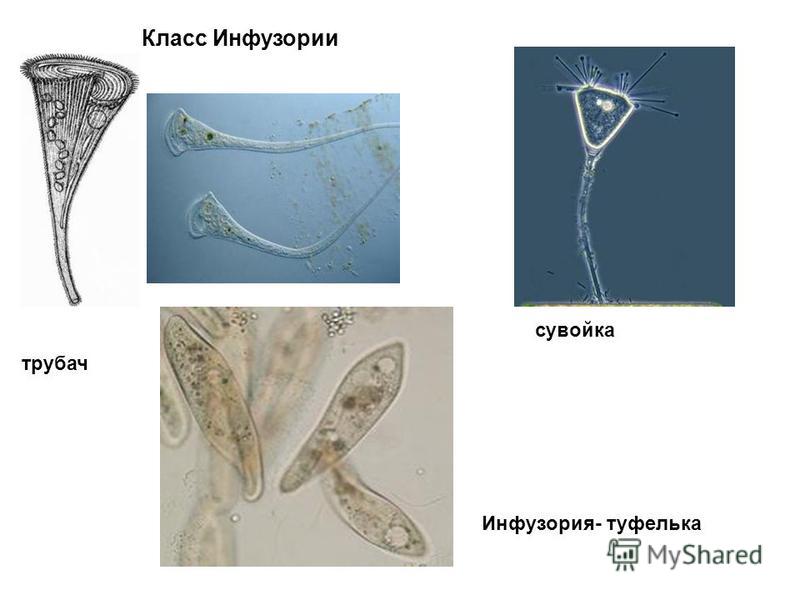 Еще в начале изучения почвенной фауны Л. Л. Бродский (1935) отмечает, что почвенные простейшие, по всей вероятности, являются космополитами.
Еще в начале изучения почвенной фауны Л. Л. Бродский (1935) отмечает, что почвенные простейшие, по всей вероятности, являются космополитами.
Следует отметить, что коэффициент общности как видового состава почвенных инфузорий, так и родов, как показали наши исследования, в большой мере зависит от степени изученности почвенной цилиофауны того или иного региона (табл. 2). Так, самый высокий коэффициент общности наблюдается между почвенной фауной инфузорий Среднего Приамурья и Европейской частью СССР, где число выявленных видов наибольшее. Самый низкий коэффициент — с наименее изученным районом — Алтаем.
Таблица 2
Общность фауны почвенных инфузорий Среднего Приамурья с фауной
других географическич районов
Районы Число ВИДОК Обшие вида с Приамурьем Коэффициент общности
Европейская часть СССР 48 28 32,2
Западная Сибирь 24 11 13,8
Алтай 10 7 10,0
Прибалтика 26 11 13,4
Украина 32 13 15.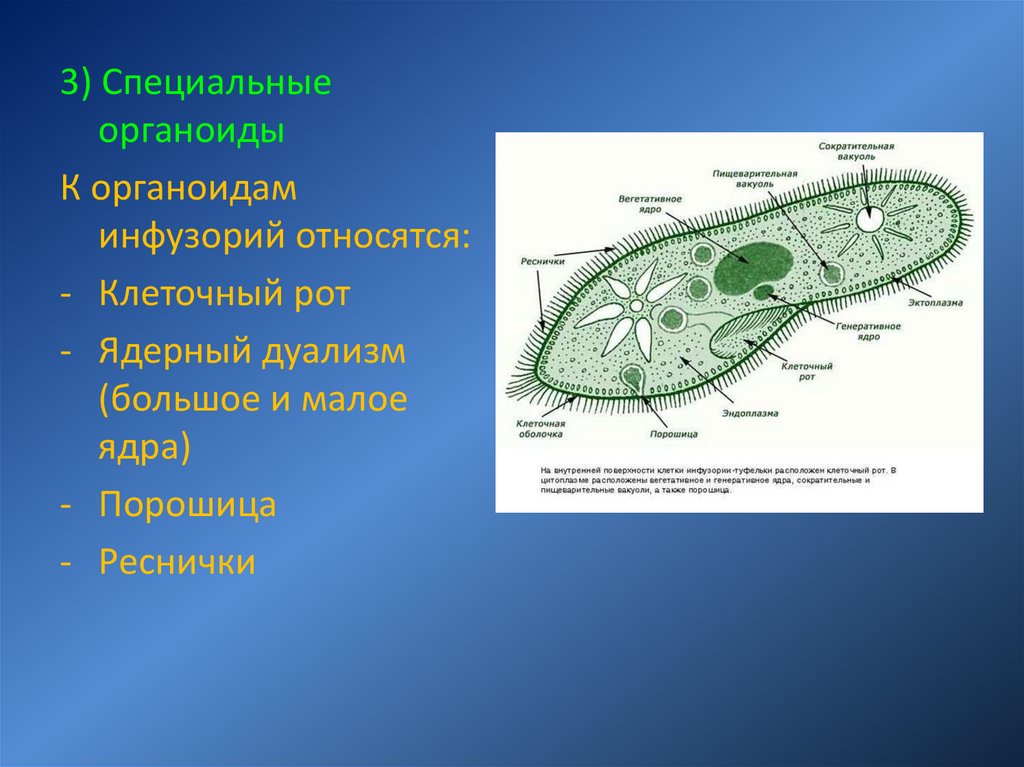 1
1
Закавказье 28 14 17,3
Средняя Азия 47 . 24 26,7
Эта закономерность делает маловероятной существование заметного эндемизма фауны почвенных инфузорий Среднего Приамурья. Если бы такой эндемизм существовал, то по мере изучения цилиофауны, коэффициент общности не увеличивался бы, а наоборот — уменьшался.
Наличие одинаковых видов инфузорий в разных типах почв и различных биоценозах как Среднего Приамурья, так и других географических районов бывшего СССР, является одним из важных аргументов в пользу гипотезы о космополитном характере распространения почвенных инфузорий.
ЖИЗНЕННЫЕ ЦИКЛЫ ПОЧВЕННЫХ ИНФУЗОРИЙ
«Наличие определенного индивидуального цикла развития представляет собою одно из основных проявлений жизни. Изучение сложных жизненных циклов животных организмов имеет для биологии глубокий теоретический и практический интерес. Всякое развитие протекает на основе взаимодействия внутренних и внешних факторов среды. Необходимо отметить, что познание факторов, определяющих сложные циклы развития животных, представляет не только теоретический интерес. Возможность управления циклами развития организмов, воздействия на них внешними факторами, очень важна в практическом отношении» (Полянский, 1940, с. 5).
Возможность управления циклами развития организмов, воздействия на них внешними факторами, очень важна в практическом отношении» (Полянский, 1940, с. 5).
Нами рассмотрены жизненные циклы инфузорий Со1рос1а сисиПш, 51у1опусЫа ер., Со1р1сНит со!рос!а и иго1ер1из рхказ.
Мы выбрали этих инфузорий в связи с тем, что они имеют большое биологическое, экологическое и практическое значение. Биологическое значение их заключается в увеличении почвенной органики, поскольку они являются одними из самых крупных и многочисленных инфузорий, способных быстро размножаться. Экологическое значение заключается в том. что они являются организмами-индикаторами сапробности как водоемов (Мамаева, 1979; Локоть, 1987 и др.), так и почв. Практическое значение состоит в использовании инфузорий в качестве тест-объектов для выявления влияния на организмы разнообразных экологических факторов.
Влияние минеральных удобрений на жизненные циклы
почвенных инфузорий
Данных по изучению воздействия разных минеральных удобрений на жизненные циклы конкретных видов почвенных инфузорий, на их биологические и физиологические особенности в литературе очень мало. Поэтому мы поставили своей целью изучить ответную реакцию нескольких видов инфузорий на разные концентрации ряда, широко применяемых в сельскохозяйственной практике, удобрений.
Поэтому мы поставили своей целью изучить ответную реакцию нескольких видов инфузорий на разные концентрации ряда, широко применяемых в сельскохозяйственной практике, удобрений.
Нами было проанализировано действие таких азотосодержащих минеральных удобрений как мочевина, сульфат аммония, аммиачная селитра. Объектами исследования мы выбрали виды почвенных инфузорий, жизненные циклы которых подробно изучены, В основном работа проводилась на инфузориях Со1рос1а сисиНш и БГуЬпуеЫа Бр. Использовали растворы в концентрациях, которые были рекомендованы в справочнике по агрохимии для применения в сельском хозяйстве (Артюшин, Державин, 1971)).
Мочевина содержит вредную примесь биурет, которая при высоком содержании может быть токсична как для растений , так и для животных (Артюшин, Державин, 1971). Внесенный в почву карбамид, под влиянием
уробактерий в течение двух — трех дней аммонизируется, превращаясь в углекислый аммоний. Затем ионы аммония под действием нитрифицирующих бактерий переходят в нитраты, подкисляющие почву. Следовательно, мочевина является потенциально кислым удобрением (Аристовская, 1458: Федоров, 1954; Мишустин, Емцев, 1987 и др.). Ее растворы увеличивают продолжительность активной фазы трофозоитов и блокируют процесс размножения (табл. 3). Поэтому в почве, после внесения мочевины, происходит заметное снижение численности инфузорий. Это отмечалось многими исследователями (Николюк, Гельцер, 1972; Гельцер, 1993 и др.).
Следовательно, мочевина является потенциально кислым удобрением (Аристовская, 1458: Федоров, 1954; Мишустин, Емцев, 1987 и др.). Ее растворы увеличивают продолжительность активной фазы трофозоитов и блокируют процесс размножения (табл. 3). Поэтому в почве, после внесения мочевины, происходит заметное снижение численности инфузорий. Это отмечалось многими исследователями (Николюк, Гельцер, 1972; Гельцер, 1993 и др.).
Таблица 3.
Влияние разных концентраций мочевины на фазы жизненного цикла Colpoda cucullus
I Фазы жизненного цикла
2 3 4 5 6 7 8
307 20-22 21,7 1,0-1,2 1,1 6,0-8,0 4-2 3-4
384 23-25 23,4 0,8-1,0 0,9 7,0-10 2 3-4
615 48 -70 61.5 — — — 2-3
Контроль 16- 19 15,8 1.3 1,5 1.4 3.5-5,0 4
Примечание: ¡ — концентрация мочевины (мп’л). 2 — активная фаза трофозоитов (в час), 3 — среднее значение, 4 — теми деления, 5 — среднее значение. 6 — деление в цистах размножения (в час). 7 — количество клеток к цистах размножения, 8 — образование цист покоя (в сутках).
7 — количество клеток к цистах размножения, 8 — образование цист покоя (в сутках).
Сульфат’ аммония является физиологически кислым удобрением. Использование в опыте растворов сульфата аммония в таких же концентрациях на инфузорий Colpoda cucullus показало, что при воздействии этого удобрения совсем не образуются цисты размножения, а формируются сразу же цисты покоя. Эксцистирования клеток в дальнейшем не происходит, даже при полной замене минеральной среды на сенной отвар. Окрашивание цист покоя раст вором эозина показало, что произошла 100 % гибель клеток.
Таким образом, используя почвенных инфузорий в качестве tcci-обт.сктов, можно уточнить концентрации минеральных удобрений и других веществ, применяемых в сельском хозяйстве, и внести практические предложения по их внесению в почву. Нами было установлено, что более щадящее действие на почвенных инфузорий оказывает одновременное внесение органических и минеральных удобрений или извести и минеральных
удобрений. Это способствует снижению кислотного характера минеральных удобрений, что в меньшей степени отражается на почвенных инфузориях.
Это способствует снижению кислотного характера минеральных удобрений, что в меньшей степени отражается на почвенных инфузориях.
Влияние разных концентраций солей тяжелых металлов на жизненные циклы почвенных инфузорий
В научной литературе все больше появляется данных о том, что почва загрязняется солями тяжелых металлов, инсектицидами и другими вредными для жизнедеятельности организмов веществами
В многочисленных работах, посвященных автотранспортному загрязнению почв, наибольшее внимание уделяется свинцу, который в значительном количестве поступает в атмосферу с отработанными газами автомобилей, а также оседает на поверхности почв (Макарова, 1988). Наиболее удобно изучать влияние всех поступающих в почву веществ на одноклеточных организмах. Их легко культивировать в лабораторных условиях, они быстро размножаются и дают немедленную реакцию на токсичные вещества.
В своей работе мы попытались экспериментальным путем определить адаптацию почвенных инфузорий Colpoda cucullus, Stylonychia sp. и Uroleptus piscis к различным концентрациям ацетата свинца (0,015 %, 0,03 %, 0,05 %, 0,1 %, 0,125 %), а также выяснить как влияют эти концентрации на жизненные циклы инфузорий.
и Uroleptus piscis к различным концентрациям ацетата свинца (0,015 %, 0,03 %, 0,05 %, 0,1 %, 0,125 %), а также выяснить как влияют эти концентрации на жизненные циклы инфузорий.
Результаты исследования показали, что даже малые концентрации ацетата свинца губительно действуют на развитие и жизненные циклы инфузорий Uroleptus piscis . При внесении клеток в растворы с концентрацией 0,1 % инфузории «съёживались», нарушалась целостность их оболочки. Большинство клеток погибало мгновенно (93,2 %). Оставшиеся в живых клетки находились все время в S-образно изогнутом состоянии. Передвигались очень медленно, поскольку у них была парализована соматическая ци-лиатура, а некоторые клетки были полностью обездвижены. Однако пери-сгомальная цилиатура работала энергично. Цитоплазма заполнялась пищеварительными вакуолями. В концентрации 0,5 % инфузории Uroleptus piscis уже через 2-3 минуты останавливались на одном месте. Тело клеток сильно утолщалось в ширину. На заднем конце и в середине тела появлялись огромные сократительные вакуоли, которые сокращались медленно. Инфузории погибали через 20 — 30 мин.
Инфузории погибали через 20 — 30 мин.
Раствор ацетата свинца в концентрации 0,125 % и выше приводил к лизису клеток Colpoda cucullus. Однако, концентрация ацетата свинца 0,1 % оказывала стимулирующее действие на инфузорий Colpoda cucullus. В течение всего периода наблюдения кольпода в массе размножались в концентрации раствора 0.1 %. В дальнейшем, мы адаптированных инфузорий перенести в микроаквариумы, содержащие 0,5 % раствор ацетата свинца. Ин-
фузории прожили чуть более суток, а затем сформировали цисты покоя. Эти цисты покоя были переведены в солонки с сенным отваром, но оксцис-тирования не произошло. Окрасив их раствором эозина, мы убедились, что все цисты были мертвы.
Stvlonvchia sp.. в огличие от Uroleptus piscisu Colpoda cucullus, оказались более резистентными к растворам разных концентраций ацетата свинца. Необходимо огметигь, что низкие концентрации растворов ацетата свинца (0,015 %, 0.03г 5 и 0, 05 %) были токсичнее, чем концентрация 0.125 %.
Таким образом, растворы ацетата свинца нарушают жизненный цикл инфузорий ра зных видов, и это приводит к гибели все популяции.
Мы считаем, что в популяции инфузорий Stylonycliia sp., культивируемых в растворе ацетата свинца с концентрацией 0,125 и 0,015%, оказались особи генетически устойчивые к воздействию данного фактора и из их потомков и сложились резистентные популяции стилоних. В измененных условиях быстрее размножаются особи, более устойчивые к действующему фактору среды, и в результате происходит постепенный сдвиг в направлении адаптации к действующему фактору (Полянский, 1978; Бурковскпй. 1983; Гроздов, 1987; Моравцева, 1988; Бойкова, 1990; Вронский. 1996 и др.).
МОРФО-ЭКОЛОГИЧЕСКАЯ ЭВОЛЮЦИЯ ПОЧВЕННЫХ ИНФУЗОРИ Й
Особенности почвы, как среды обитания, повлияли на морфологические показатели простейших п на некоторые процессы их жизнедеятельности. Например, в о глпчие от видов, обитающих в водных биоценозах, мните почвенные инфузории имеют размеры в 5-10 раз меньше. Так, если водные формы инфузорий Prorodon teres имеют следующие размеры: длина 100 — 200 мкм, ширина 60 — 115 мкм. то почвенные формы имеют длину 70 -100 мкм, а ширину 38 — 50 мкм. Инфузории Spirosiomum teres, обитающие в водоемах, имели длину клеток 150 — 400 мкм, в то время как длина тела, выявленных из почвенных образцов особей данного вида, составляла Srt — 90 мкм. Кроме того, частота пульсации сократительных вакуолей у почвенных инфузорий, примерно, н 1.5 — 2 раза медленнее, чем у их аналогов из пресных водоемов. Некоторые авторы считают, что измельчение инфузорий, обитающих в почве, произошло в результате своеобразия почвенных условий. а именно: низкого содержания кислорода и недостатка пищи (Корганова, 1977 и др.). Мелкие размеры клеток обеспечивают быструю диффузию кислорода к центру инфузории (Kubiena, 1938, цит. по Коргано-вой, 1977). Карликовость почвенных цшшат возможна еще и потому, что
Так, если водные формы инфузорий Prorodon teres имеют следующие размеры: длина 100 — 200 мкм, ширина 60 — 115 мкм. то почвенные формы имеют длину 70 -100 мкм, а ширину 38 — 50 мкм. Инфузории Spirosiomum teres, обитающие в водоемах, имели длину клеток 150 — 400 мкм, в то время как длина тела, выявленных из почвенных образцов особей данного вида, составляла Srt — 90 мкм. Кроме того, частота пульсации сократительных вакуолей у почвенных инфузорий, примерно, н 1.5 — 2 раза медленнее, чем у их аналогов из пресных водоемов. Некоторые авторы считают, что измельчение инфузорий, обитающих в почве, произошло в результате своеобразия почвенных условий. а именно: низкого содержания кислорода и недостатка пищи (Корганова, 1977 и др.). Мелкие размеры клеток обеспечивают быструю диффузию кислорода к центру инфузории (Kubiena, 1938, цит. по Коргано-вой, 1977). Карликовость почвенных цшшат возможна еще и потому, что
из-за недостатка кислорода инфузории начинают делиться до достижения нормальной величины (Корганова, 1977).
Кроме уменьшения размеров клеток, во многих случаях наблюдается заметное снижение колебания целого ряда других морфометрических показателей у почвенных форм. В частности, инфузории Loxodes rostrum, выделенные из почвы, имели амплитуду длины клеток от 50 до 80 мкм, т.е. 30 мкм. Колебания длины клеток у водных форм этого же вида намного больше: от 150 до 300 мкм, т.е. 150 мкм. Это в 5 раз превышает показатель почвенных форм. Или Spirostomum teres. Размах колебания длины клеток водных форм составляет 250 мкм, в то время как у почвенных форм всего лишь 10 мкм. Мы считаем, что это один из путей морфо-экологической эволюции почвенных инфузорий. В данном случае небольшие размеры почвенных микроводоемов ограничивают вариабельность длины клеток цилиат, что закрепилось в ходе длительной эволюции.
Одним из проявлений адаптации инфузорий к почвенным условиям является уплощение тела и наличие хвостовых придатков. Длинные хвостовые реснички мы отмечали у Pleuronema coronatum, Microthorax pusillus, Leptopharynx sp. , Holophrya hexatricha, а также у всех выявленных видов рода Uronema: и многих других видов инфузорий. Кроме того, нами было выявлено прикрепление инфузорий некоторых видов (например, Pleuronema coronatum) к почвенным частицам.
, Holophrya hexatricha, а также у всех выявленных видов рода Uronema: и многих других видов инфузорий. Кроме того, нами было выявлено прикрепление инфузорий некоторых видов (например, Pleuronema coronatum) к почвенным частицам.
Необходимо отметить, зафиксированное нами у ряда видов почвенных инфузорий (Oxytricha sp., Colpidium colpoda, Uroleptus piscis и др.), утолщение в виде ампул на концах соматических ресничек По всей вероятности, такая модификация цилиатуры обеспечивает прикрепление инфузорий на какое-то время к почвенному субстрату. Прикрепление к субстрату является полезной биологической адаптацией, которая препятствует вымыванию почвенных инфузорий из грунта во время муссонных дождей. Инфузории прикрепляются к субстрату посредством каудальных ресничек или цирр. На временное прикрепление интерстициальных инфузорий к поверхности песчинок указывал Сведмарк (Swedmark, 1964, цит. по Агамалиеву, 1977).
Одной из важных адаптаций инфузорий к почвенным условиям является очень подвижная пелликула , благодаря которой клетки довольно части приобретают амебоидную форму. К таким инфузориям, с очень пластичной оболочкой, относятся Enchelys pupa, Blepharisma steini и lateritium, Colpoda maupasi, Dileptus anser и многие другие. Эластичная пелликула дает возможность клеткам передвигаться между почвенными частицами.
К таким инфузориям, с очень пластичной оболочкой, относятся Enchelys pupa, Blepharisma steini и lateritium, Colpoda maupasi, Dileptus anser и многие другие. Эластичная пелликула дает возможность клеткам передвигаться между почвенными частицами.
Формой биологических адаптаций к почвенным условиям инфузорий является также и подвижность клеток. Из почвенных образцов были выделены в основном активно передвигающиеся особи и очень мало — прикрепленных. При этом, прикрепленные виды инфузорий лаже в воздушно-сухой почве имели очень низкую плотность, за исключением УогйсеНа писголюта.
У некоторых видов рода Со1рос!а изменения затронули продолжительность жизненного цикла (Никитина, 1994). Так, если водные формы Со!роёа сисиНич в активном состоянии находятся 5 6 дней и за это время дают, примерно, 9 новых генераций, а затем образуют цисты покоя, то у почвенных форм цисты покоя появляются уже на 3-4 день. При этом, толщина цистной оболочки почвенных форм в 1,5-2 раза больше, чем у водных форм, что обеспечивает гораздо большую резистентность этих простейших к почвенным условиям и даже к целому ряду экстремальных факторов.
Изучая морфометрические, физиологические и другие характеристики многих видов инфузорий, выделенных из почв разных географических регионов бывшего СССР, мы можем также обратить внимание еще на одп\ адаптивную особенность — наличие географических рас (Никитина. 1984). Существование географических рас привело к стойким различиям в размерах клеток и их жизненных циклах (табл. 4).
Таблица 4
Морфо-физнологнческие показатели Со1рос1а сисиНия. выделенных
из почв р а з н ы х г с о г р а ф и ч е ск и х юн (Никитина. 1984|
Показатели гп¡фузорий Популяции
Мурманская Ленинграда*: Барсакельм. Хабаровска
Среднее время активной фазы трофозоитов 20,3+0,2 1.2+0.05 15,8+0,4 10,2+0,2 23.4+0.4
Темп деления 1,5+0,04 2,4+0,09 1,0+0,04
Средняя длина трофозоитов (в мкм) 58,7+0,7 97,3+1,0 70,1+0,6 113,1+1.0
Диаметр цист покоя (мкм) 33.4+0.2 39.3+0.3 32,1+0,2 54,8+0,2
Ус гойчивосз ь трофозоитов к 5-мииутному действию температуры 40° С (% живых клеток) 0 7,2+0,1 95,9+1,2 0
Сравнивая морфометрические показатели некоторых видов инфузорий, выделенных из почв Среднего Приамурья и Европейской части Рос-
сии, отмечаем, наличие существенных различий (табл.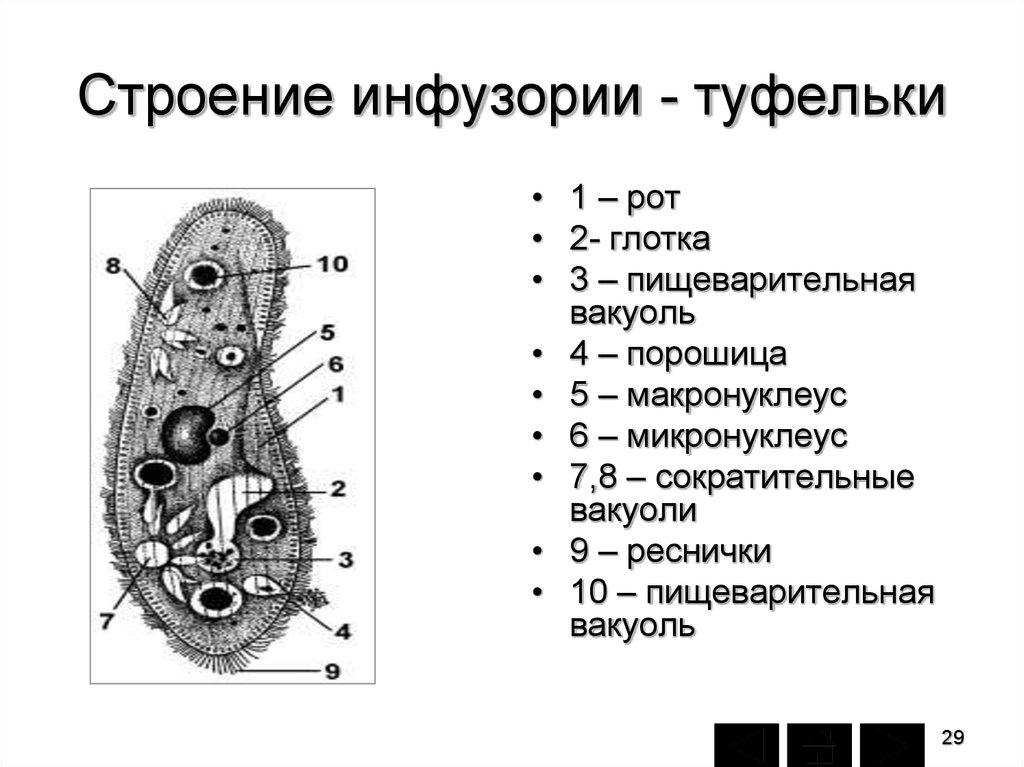 5). Это свидетельствует в пользу гипотезы существования географических рас у почвенных инфузорий. Поскольку одним из критериев географических рас являются различия по морфометрическим показателям (Яблоков, Юсуфов, 1989; Тыщенко, 1992 и др.).
5). Это свидетельствует в пользу гипотезы существования географических рас у почвенных инфузорий. Поскольку одним из критериев географических рас являются различия по морфометрическим показателям (Яблоков, Юсуфов, 1989; Тыщенко, 1992 и др.).
Таблица 5
Сравнительный анализ морфометрических показателей инфузорий,
выделенных из почв разных регионов
Виды инфузорий Морфометр. показатели иш зузорий разных регионов
Среднее Приамурье Европейская часть
1 2 j 4 5 6
Prorodon teres 70-100 38-50 2 : 1 80-200 50-140 1,5 : 1
Chilodonella unci-nata 30-50 17-28 1,8:1 50-90 — -
Colpidium colpoda 70-90 35-50 2 : 1 80 50 1,6: 1
Glaucoma piriformis 8-20 6-10 1,5:1 38-80 — -
Cyrtolophosis mu-cicola 59-62 27-30 2 : 1 25-28 — -
Blepharisma lateritium 90-120 50-80 1,6 : 1 130-200 — -
Примечание: 1 — длина клеток (мкм), 2- ширина (мкм), 3 — соотношение дины к ширине, 4 — длина (мкм). 5 — ширина (мкм), 6 — соотношение длины к ширине.
5 — ширина (мкм), 6 — соотношение длины к ширине.
Таким образом, почвенные условия наложили значительный отпечаток на морфологию, биологию и физиологию почвенных цилиат. Морфо-экологическая эволюция инфузорий от водных форм к почвенному образу жизни шла в направлении:
1. Уменьшение размеров тела инфузорий;
2. Уменьшение вариабельности размеров клеток;
3. Увеличение пластичности пелликулы инфузорий;
4. Развитие разнообразных прикрепительных структур (ресничек, цирр и др.) для удержания на поверхности почвенных частиц во время мус-сонных дождей;
5. Большая подвижность клеток;
6. Сокращение жизненного цикла инфузорий, по сравнению с водными формами;
7. Формирование географических рас у некоторых видов почвенных инфузорий.
Рассмотрев адаптации инфузорий к почвенным условиям, которые выработались в течение длительной эволюции, в результате перехода их о г морских форм к почвенным, мы можем выделить группы инфузорий гео-бионтов, геофилов и геоксенов для почв Среднего Приамурья (табл. 6).
6).
Таблица 6
Видовой состав жизненных форм почвенных инфузорий
Среднего Приаму рья
Геобионты Геофилы Геоксены
Colpoda steini Holophrva hexatncha Loxodes magnus
С. maupasi II. snnplex L. rostrum
С. cucullus Plagiocampa mutabihis Prorodon teres
С. aspera Clwenca teres Coleps hirtus
Uronema marinum Enchelys farcimen Enchelys microstomata
Strombidium sp. E. gasterosteus Dileptus gigas
Enchelus pupa Lacrymaria vermicularis Didinium balbiani
Colpidium colpoda Spathidium porculus Amphileptus ilexilis
Cyrtolophosis elondata S. spatula Litonotus fasciola
Drepanononas revoluta Dileptus anser L. varsaviensis
Glaucoma pyriformis D. nistroviensis Loxopliylltim helus
11 видов Cy rtolophosis mucicola Chilodonella uncmata
Chilodonclla cucullulus Vorticella campanula
Pleuronema coronatum V. convallaria
Vonicclla microstoma V. putrina
putrina
Blephansma lateritium Spirostomum minus
B. steini S. teres
Paruroleptus piscis и др. Meseres cordiformts и др.
33 вида 24 вида
ВЫВОДЫ
1. Изучен видовой состав и экология инфузорий из основных чипов почв (бурые лесные, буро-подзолистые, лугово-глеевые и др.) и различных биоценозов Среднего Приамурья. Выявлено 102 вида и разновидности! почвенных инфузорий.
2. Наиболее богат и разнообразен видовой состав почвенной цилио-фауны в биогеоценозах Большехехцирского заповедника (67 видов) и очень беден видовой состав в почвах Воронежских высот (43 вида), которые находятся под сильным антропогенным прессом.
3. Новыми для почв бывшего СССР являются 32 вида инфузорий, 19 родов, 9 семейств, 2 отряда (Karyorelictida Corliss, 1974 и Suctorida Claparede et Lachmann, 1858) и подкласс Suctoria Claparede et Lachmann, 1858.
4. Сравнение видового состава фауны почвенных инфузорий Среднего Приамурья с другими регионами показало, что индекс видового сходства инфузорий зависит от степени изученности фауны инфузорий в этих районах.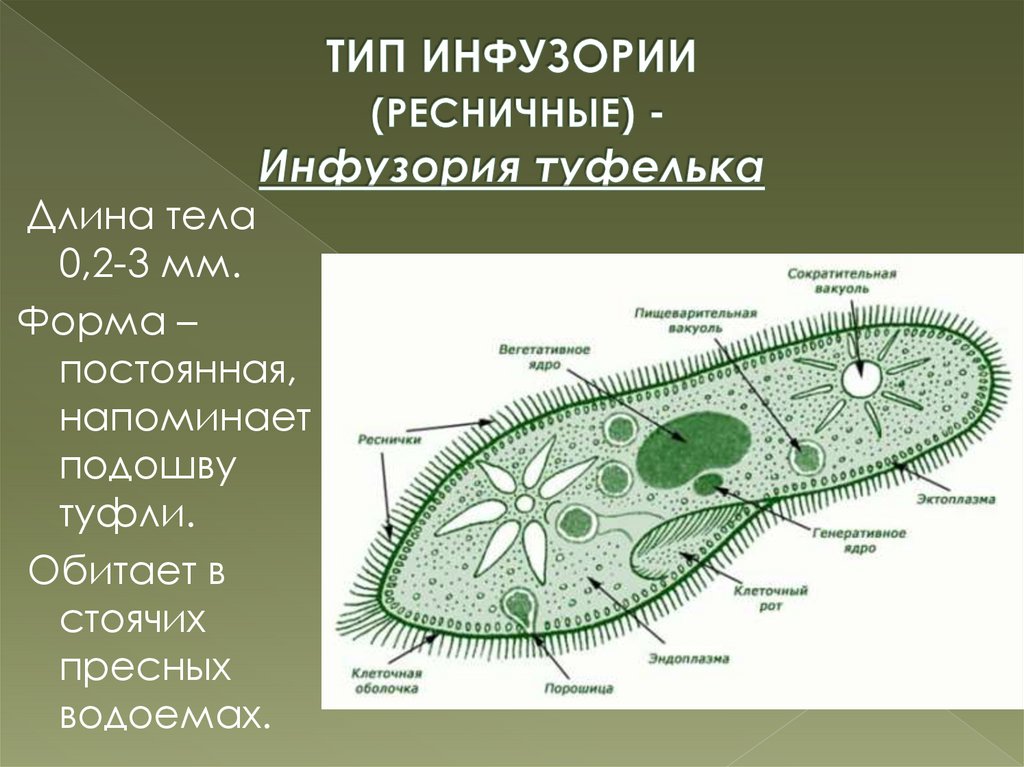 Коэффициент общности варьирует по числу видов от 10,0 (Алтай) до 32,2 (Европейская часть России) и числу родов — от 11,5 (Алтай) до 38,5 (Европейская часть России). Наличие общих видов подтверждает гипотезу A. JI. Бродского о космополитном характере географического распространения почвенных инфузорий.
Коэффициент общности варьирует по числу видов от 10,0 (Алтай) до 32,2 (Европейская часть России) и числу родов — от 11,5 (Алтай) до 38,5 (Европейская часть России). Наличие общих видов подтверждает гипотезу A. JI. Бродского о космополитном характере географического распространения почвенных инфузорий.
5. Выявлены закономерности существования почвенных инфузорий в условиях Среднего Приамурья, которое имеет уникальные природно-климатические особенности. Отличительной чертой динамики популяций является наличие в почвах естественных биоценозов только одного пика численности инфузорий — осеннего, по сравнению с двумя пиками (весенним и осенним) характерными для почв Европейской части России и Средней Азии.
6. В почвах агроценозов также выявлен только один пик численности инфузорий — летний. Это объясняется промывным режимом пахотных почв во второй половине лета и начале осени в связи с муссонными дождями. Установлено, что внесение в почву органических удобрений вместе с известью способствует задержанию инфузорий в почве пахотного горизонта.
7. Почва ризосферы и ризопланы как сельскохозяйственных, так и лесных растений содержит почвенных инфузорий значительно больше, чем удаленная от корней, что связано, в первую очередь, с наличием в данных зонах бактерий, являющихся пищей для многих инфузорий.
8. Аккумуляция почвенных инфузорий весной происходит в верхнем почвенном слое (0 — 2-5 см) или подстилке, летом инфузории мигрируют вглубь почвы, а осенью концентрируются в верхних слоях. Глубина проникновения почвенных инфузорий в основном составляет 15-20 см.
9. Установлено, что буро-подзолистые почвы обладают сильно выраженным цилиостазисом, который обусловлен кислотным характером почвенного раствора.
10. Определены комплексы почвенных инфузорий для бурых лесных и буро-подзолистых почв Большехехцирского заповедника, буро-подзолистых почв Воронежских высот и буро-подзолистых почв агроцено-
за. Каждый биогеоценоз обладает набором своих собственных видов почвенных инфузорий, многие из которых являются общими.
11. Определен комплекс видов инфузорий, характеризующий бурые почвы Среднего Приамурья: Colpoda steini. С. maupasi, С. cucullus, Uronema marinum, Aspidisca costata, Vorticella microstoma, Strombidium sp., Holosticha sp., Oxytricha pcllionella, Drepanomonas revoluta, Chilodonella uncinata, Colpidium colpoda.
12. Фауна почвенных инфузорий подтвердила «зрелость» Хехцирекой свиты, поскольку в почвах Болыпехехцирского заповедника наблюдается наибольшее сходство цилиофауны но разным биогеоценозам и «молодость» Хабаровской и Воронежской свит, в почвах которых выявлена значительная вариабельность видового состава инфузорий.
13. Фауна почвенных инфузорий Среднего Приамурья не выявила высокой степени эндемичности, характерной для многоклеточных животных и растений этого региона. В почвах Приамурья обитают инфузории, встречающиеся в самых разнообразных почвах и климатических зонах.
14. Используя некоторые виды почвенных инфузорий в качестве тест-объектов для определения влияния минеральных удобрений и шлеп гяже-лых металлов на их жизненные циклы, было установлено, что минеральные удобрения блокируют процесс размножения инфузорий, а соли свинца нарушают процесс цистообразования у инфузорий.
15. Наиболее устойчивыми к воздействию ацетата свинца оказались инфузории Stylonyehia sp. Концентрация 0,125 % ацетата свинца оказала стимулирующее воздействие на популяцию стнлотпгаш, поскольку темп деления увеличился в 1,2 раза, по сравнению с контролем. Инфуюрии Stylonyehia sp. можно отнести к индикаторам сапробности почв.
16. Морфо-экологическая эволюция инфузорий к условиям обитания в почве осуществлялась по следующим направлениям:
а) уменьшение размеров клеток;
б) уменьшение вариабельности размеров клеток;
в) укорочение жизненного цикла;
г) формирование своеобразной соматической цилиатуры. Реснички многих видов почвенных инфузорий заканчиваются небольшими ампулами;
д) более редкая пульсация сократительных вакуолей у почвенных инфузорий по сравнению с аналогами пресноводных форм:
е) формирование толстых защитных оболочек цист покоя;
ж) повышение жизнестойкости цист покоя к изменили условий окружающей среды;
з) формирование географических рас у почвенных инфузорий.
Список основных работ автора, опубликованных
по теме диссертации:
1. Никитина Л.И: К вопросу о влиянии некоторых абиотических факторов на природные популяции инфузорий Со1рос1а сисиНш // ВИНИТИ, N 6662-84 Дел. 12.10.1984r.-13 с.
2. Никитина Л.И. Эколого-фаунистическая характеристика инфузорий рода Со1рос1а некоторых географических зон //ВИНИТИ, N 6663-84 Деп. 12.10.1984г.-17 с.
3. Никитина Л.И. Влияние длительного культивирования на физиологические показатели инфузорий // Проблемы зоокультивирования.-М., 1986.-С. 97
4. Никитина Л.И. Эколого-фаунистическая характеристика инфузорий рода Со1ро<1а некоторых биоценозов Приамурья // Актуальные проблемы преподавания биологов в ВУЗЕ в условиях Приамурья. Хабаровск: ХГПИ.-1987.-С. 17-23
5. Никитина Л.И. Содержание резервных веществ в цитоплазме клеток, выделенных из почв разных географических зон // Современные проблемы протозоологии. — Л., 1987,- С. 74-75.
6. Никитина Л.И. Особенности распространения инфузорий в лесных и окультуренных почвах окр. г. Хабаровска // Изменение растительного и животного мира под влиянием хозяйственной деятельности в условиях Д.В. -Хабаровск 1988.-С.132-138.
Никитина Л.И. Особенности распространения инфузорий в лесных и окультуренных почвах окр. г. Хабаровска // Изменение растительного и животного мира под влиянием хозяйственной деятельности в условиях Д.В. -Хабаровск 1988.-С.132-138.
7. Никитина Л.И. Инфузории из буроподзолистых почв агробиологической станции //там же. — С. 138-142.
8. Никитина Л.И. Видовой состав и количественная характеристика инфузорий в почвах некоторых лесных биоценозов окр. г. Хабаровска // Почвенные беспозвоночные юга Дальнего Востока. Хабаровск: ДВО АН СССР, 1989,- С. 48-54.
9. Никитина Л.И. Распространение инфузорий в окультуренных почвах окр. г. Хабаровска //там же. — С. 54-57.
10. Никитина Л.И. Эколого-фаунистическая характеристика инфузорий лесных и окультуренных почв окр. г. Хабаровска // Региональные эко-лого-фаунистические исследования как научная основа мониторинга, охраны и рационального использования животных. — Курск, 1990,- С. 47.
11. Никитина Л.И.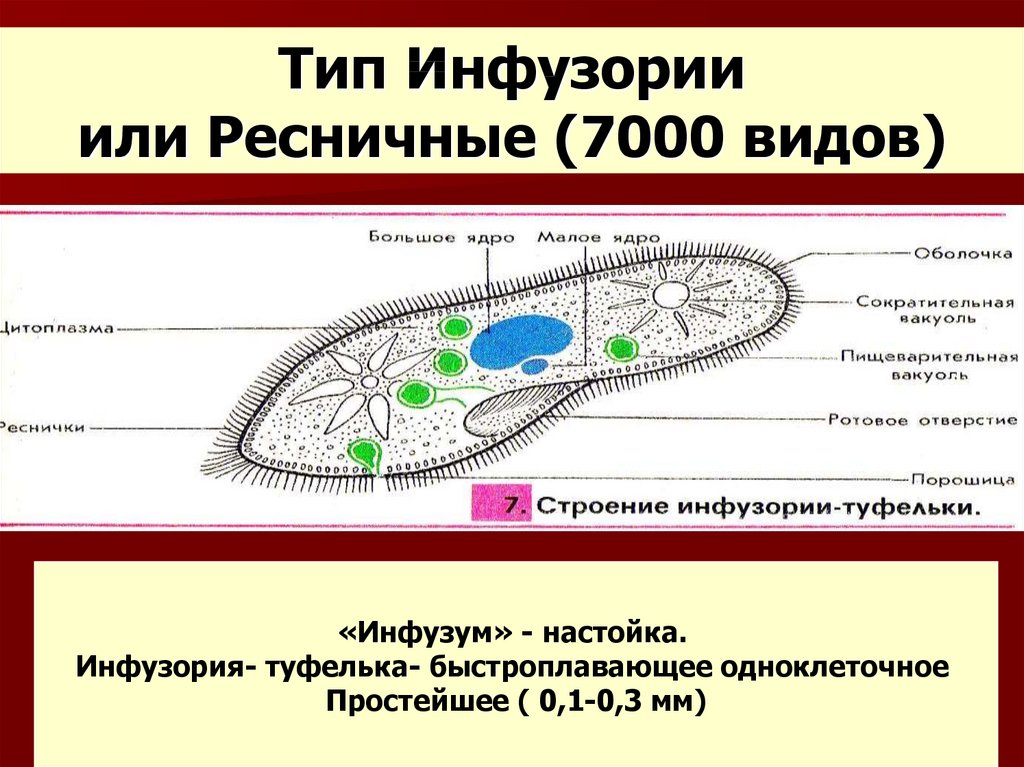 Фауна инфузорий рода Со1рос1а серо-бурых почв острова Барсакельмес // Проблемы почвенной зоологии. — Новосибирск, 1991,- С. 79.
Фауна инфузорий рода Со1рос1а серо-бурых почв острова Барсакельмес // Проблемы почвенной зоологии. — Новосибирск, 1991,- С. 79.
12. Никитина Л.И. Фауна и экологические закономерности существования почвенных инфузорий в биоценозах Большехехцирского заповедника
// Охрана и рациональное использование экосистем Дальнего Востока. -Хабаровск: ХГПИ. 1992,- С. 96-99.
13. Никитина Л.И. Влияние мочевины на жизненный цикл Со1рос1а сосиНиз// Цитология. — 1992, т.34,- С. 107.
14. Никитина Л.И. Изучение инфузорий стоячих водоемов в курсе ■зоологии // Экологическое образование в школах и педагогических институтах. Хабаровск: ХГПИ, 1992,- С, 72-79.
15. Никитина Л.И. Инфузории стоячих водоемов окр. г. Хабаровска // Состояние и динамика природных и социальных систем Приамурья. — Ком-сомольска-на-Лмуре. — 1992. С. 91-92.
16. Никитина Л.И. Лисицкая О. Протозоологические иследования водоемов окр. г. Хабаровска » Съезд сведущих людей Дальнего Востока. -Хабаровск, 1994,-С. 145-148.
-Хабаровск, 1994,-С. 145-148.
17. Никитина Л.И., Кутырева Л.Т. Педобионты Приамурья, их мониторинг // Фундаментальная и методическая подготовка будущего специалиста по экологии и охране природы. — Орел, 1994,- С.73-74.
18. Никитина Л.И., Кутырева Л.Т. Педобионты — индикаторы почвенных условий //Экология и охрана окружающей среды. — Пермь. 1995,-С. 77-78.
19. Никитина Л.И. Инфузории стоячих водоемов окр. г. Хабаровска // Материалы 41 итоговой научной конф. Преподавателей ХГПУ. — Хабаровск. 1995,- С. 23-24.
20. Никитина Л.И,. Кутырева Л.Т. Педобионты и их роль в определении почвенных условий /У Экология и охрана окружающей среды. — Владимир, 1996,- С. 233-234.
21. Никитина Л.И. Почвенные инфузории — объекты экологических исследований ¡1 там же. — С. 174-175.
22. Никитина Л.И. Эколог ические особенности почвенных инфузорий // Материалы 42 научной конференции ХГПУ. — Хабаровск, 1996,- С. 121123,
23. Никитина Л.И. Сезонная динамика почвенных инфузорий некоторых биоценозов Большехехцирского заповедника //там же.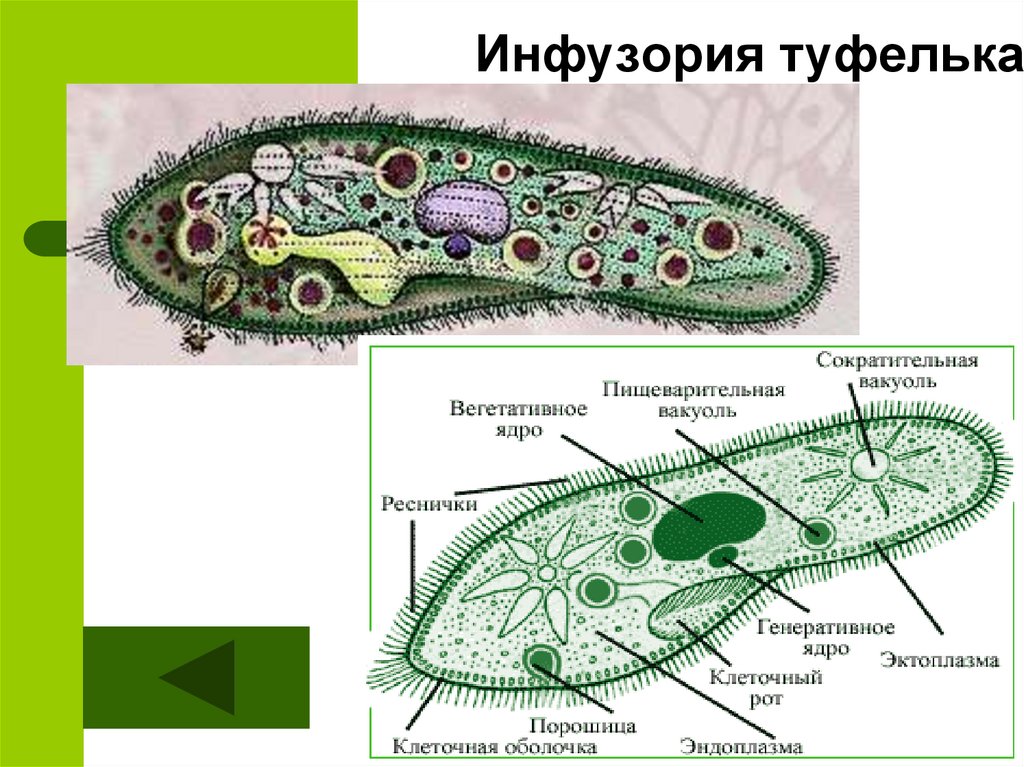 — С. 123-125.
— С. 123-125.
24. Никитина Л.П. Фауна и динамика инфузорий в бурых лесных почвах Большехехцирского заповедника >! Проблемы почвенной зоологии. -Ростов-на-Дону. 1996,-С. 108-109,
25. Никитина Л.И., Ножкина Л.В. Влияние азотосодержащих минеральных удобрений на жизненные циклы некоторых почвенных инфузорий И тли же.- С. 109-110.
26. Никитина Л.И. Почвенные инфузории Среднего Приамурья И Монография. Хабаровск: Изд-во ХГПУ, 1997 г. — 102 с.
27. Никитина Л.И., Лукашук Н.П., Красненкова Е. Инфузории пахотного горизонта агробиологической станции ХГПУ // Материалы 43 научной конференции ХГПУ. — Хабаровск, 1997.- С. 57-60.
28. Никитина Л.И. Комплексы инфузорий как показатель почвенных условий смешанного леса Большехехцирского заповедника и Воронежа-1 // там же. — С. 60-63.
29. Никитина Л.И., Титова Г., Белова Т. Почвенные инфузории как тест-объекты для оценки токсичности некоторых веществ // там же. — С. 63-66.
30. Никитина Л.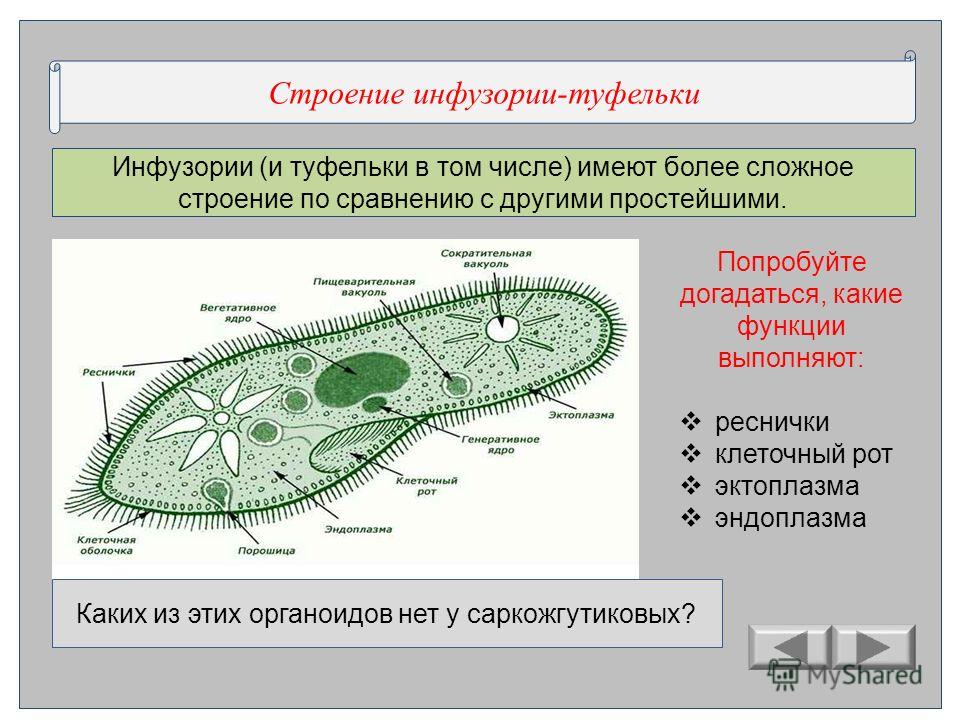 И. Почвенные инфузории Со1рос1а сисиИш как модельные объекты в биотестировании // Инфузории в биотестировании. -Санкт-Петербург, 1997 — С. 87.
И. Почвенные инфузории Со1рос1а сисиИш как модельные объекты в биотестировании // Инфузории в биотестировании. -Санкт-Петербург, 1997 — С. 87.
31. Никитина Л.И. Изучение почвенных инфузорий в период полевой практики по зоологии // Производственные и полевые практики — основа эколого-профессиональной подготовки молодого специалиста. — Хабаровск: ХГПУ, — 1997,- С. 17-18.
Инфузории пробуют пищу
Е. Г. Черемных, кандидат технических наук,
Е. И. Симбирева, кандидат химических наук
«Химия и жизнь» №1, 2009
Если зачерпнуть воды из пруда у берега и посмотреть на нее под микроскопом, то откроется удивительная картина. Циклопы и дафнии, нематоды и коловратки поражают фантастичностью и разнообразием форм. Эти организмы и более мелкие — бактерии, грибы, одноклеточные водоросли, инфузории и другие многочисленные простейшие — необходимое звено круговорота жизни. Особенно важную работу они выполняют сегодня, когда в окружающую среду поступают огромные количества техногенных отходов.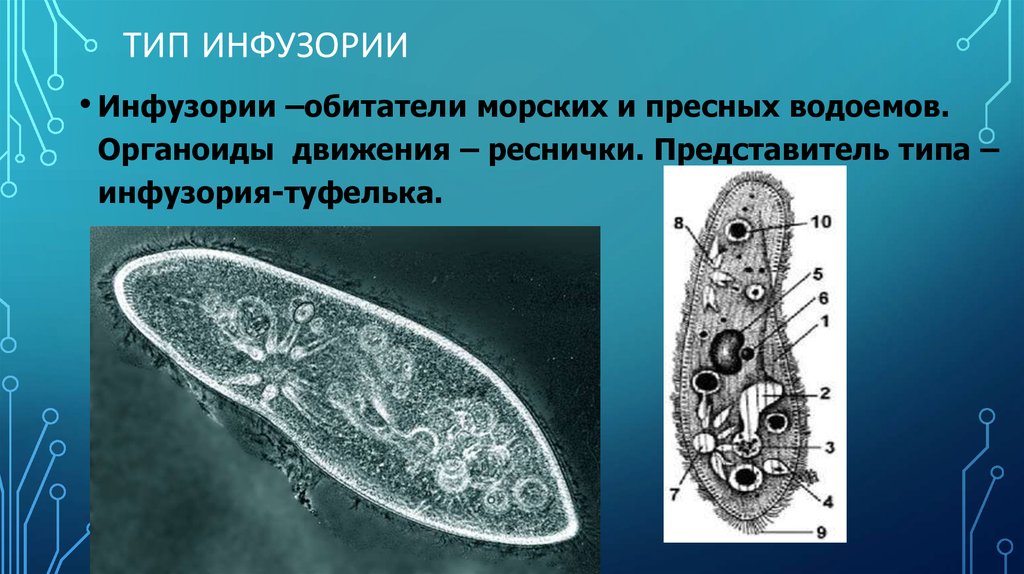 Главная роль в переработке и обезвреживании разнообразных искусственных соединений принадлежит именно микроскопическим организмам.
Главная роль в переработке и обезвреживании разнообразных искусственных соединений принадлежит именно микроскопическим организмам.
Удивительно: с одной стороны, простейшие очень хорошо приспосабливаются к изменениям, иначе они не смогли бы выжить в меняющемся мире. С другой стороны, каждый отдельный организм очень тонко чувствует изменения среды, поэтому их можно использовать в биологических исследованиях безопасности почвы, природных и сточных вод. Такие исследования называют биотестами.
О биотесте подробнее
Биотестирование — это оценка реакции тест-организмов на ту или иную субстанцию. В качестве тест-организмов в экологии обычно используют низшие организмы, в том числе и одноклеточные, поскольку проводить опыты с ними гораздо удобнее, чем с высшими животными. Лучше всего подходят инфузории. Их легко выращивать, и оценить результат несложно — достаточно сосчитать их до начала опыта и в конце.
В экологических целях биотесты применяют и как отдельные методы, и в комплексных исследованиях. Биотесты также используют для оценки токсичности кормов, лекарств, полимерных материалов и питьевой воды. А вот безопасность пищевых продуктов и пищевых добавок с помощью биотестирования не определяют. Но если б определяли, была бы от этого польза? Прежде чем ответить на этот вопрос, рассмотрим состав современных продуктов.
Биотесты также используют для оценки токсичности кормов, лекарств, полимерных материалов и питьевой воды. А вот безопасность пищевых продуктов и пищевых добавок с помощью биотестирования не определяют. Но если б определяли, была бы от этого польза? Прежде чем ответить на этот вопрос, рассмотрим состав современных продуктов.
Что мы едим
Допустим, мелким шрифтом на упаковке написано, что пищевой продукт содержит загадочные Е102, Е214, Е342, Е621. Технолог знает, что Е с номерами от 100 до 182 — красители, от 200 до 299 — консерванты, от 300 до 399 — антиоксиданты и регуляторы кислотности, от 600 до 699 — ароматизаторы и усилители вкуса. В частности, Е102 — краситель тартразин, Е214 — этиловый эфир парагидроксибензойной кислоты, Е342 — фосфаты аммония, Е621 — глутамат натрия. Все эти добавки разрешены в РФ, но, если говорить о мире в целом, на каждое из этих веществ имеется «компромат»: тартразин запрещен в ряде стран, глутамат (о котором мы еще поговорим подробнее) не рекомендован к применению в детском питании. С химической точки зрения (рис. 1) это самые разные соединения.
С химической точки зрения (рис. 1) это самые разные соединения.
Поскольку один продукт обычно содержит множество добавок, он может оказаться более вредным для здоровья, чем каждый компонент в отдельности.
Возможно, именно поэтому список нежелательных соединений постоянно растет; в него попадают и те, которые первоначально разрешили. Медики уже обнаружили канцерогенное, гепатотоксическое, неиротоксическое и аллергическое действие некоторых пищевых красителей, консервантов, антиокислителей, усилителей вкуса и подсластителей. Один из любимых аргументов у сторонников искусственных химических компонентов — их незначительные концентрации в пище при значительной эффективности. Однако, несмотря на малые количества каждой добавки, совокупное их действие в одном продукте может быть опасным для человека.
Патологические изменения метаболизма начинаются уже с пищеварения. Некоторые из этих веществ блокируют пищеварительные ферменты, из-за чего организм человека не может эффективно использовать питательные компоненты пищи. В результате начинает бурно размножаться гнилостная микрофлора толстого кишечника. Это происходит еще и потому, что пищевые добавки из группы консервантов угнетают полезные бифидум- и лактобактерии. А метаболиты гнилостных микроорганизмов (триптамин, спермидин, индол и скатол), попадая в кровь, отравляют организм.
Сами низкомолекулярные пищевые добавки тоже хорошо всасываются в желудочно-кишечном тракте и с кровью попадают в клетки тканей и органов. Их биохимические эффекты так же разнообразны, как и химическое строение. Так, например, пищевые азокрасители провоцируют свободно-радикальное окисление липидов, которое, в свою очередь, нарушает структуру и проницаемость мембран клеток. Свободные радикалы, участвуя в лавинообразном процессе пероксидного окисления, могут быть причиной опухолей и преждевременного старения.
Доказано, что пищевые добавки гетероциклического строения метаболизируют ферменты системы цитохрома Р450. Организм откликается тем, что начинает производить больше таких ферментов. От их активности зависит скорость образования канцерогенных веществ из проканцерогенов, то есть чем больше таких ферментов, тем быстрее происходит это нежелательное превращение. Например, при метаболизме бенз(а)пирена, который есть во всех копченых продуктах, образуется канцерогенный дигидроксиэпоксид.
Поскольку метаболизм любых чужеродных веществ сложен и неоднозначен, существующая система санитарно-гигиенической оценки пищи не справляется со своей задачей. Сегодня определение отдельных чужеродных веществ и ограниченные испытания на крысах уже не гарантируют безопасность продукта.
Получается парадоксальная ситуация: экологи, имеющие разнообразный и надежный инструментарии для контроля окружающей среды, стараются обеспечить безопасность пищевого сырья, а технологи добавляют в это сырье ксенобиотики и представляют результат как улучшенный продукт.
Пора менять систему санитарно-гигиенической оценки, причем и для вновь вводимых пищевых добавок, и для уже существующих. Здесь пригодились бы разнообразные цитотоксикологические методы, среди которых могут быть и биотесты на инфузориях.
Инфузории как тест-объекты
Аргументирование последнего утверждения начнем издалека — с описания биологических особенностей этих тест-организмов.
Инфузории — это одноклеточные эукариотические организмы, то есть у них есть ядро (а у многих видов инфузорий даже два ядра). Почти все клетки человека и других высших животных имеют ядра, а бактерии — прокариоты (безъядерные). Инфузории были одним из любимых объектов биологов еще лет сто назад, но и сейчас они остаются в центре внимания. Совсем недавно расшифровали геном инфузорий из рода Tetrahymena (рис. 2), состоящий из 27 тысяч генов. Кстати, геном человека содержит примерно столько же генов — 25 тысяч.
Эти одноклеточные — в основном свободноживущие организмы, которые в воде и почве поедают бактерии и водоросли. Они — необходимое звено в трансформации органики, их вклад в биологический круговорот веществ не менее важен, чем у бактерий. Живут инфузории практически везде: в пресных и соленых водах, почве, и число их видов, по разным источникам от 7 до 10 тысяч.
Характерная особенность инфузорий — относительно быстрая изменчивость, которая позволяет им адаптироваться к самым разным условиям. Инфузории живут и в тундровых озерах, и в тропиках, и даже в горячих источниках с температурой до 50°С. Они приспосабливаются и к разному минеральному и органическому составу среды, а также к присутствию растворенных газов. Например, Paramecium caudatum (рис. на заставке) может жить как в аэробных, так и в анаэробных условиях. Эту высокую адаптивную способность обеспечивает колоссальное количество генов, большинство которых в повседневной жизни инфузориям не нужны, но включаются, когда условия меняются.
По мере того как простейшие адаптируются к условиям среды, перестраиваются все их жизненные функции, изменяются скорость движения, темп размножения и способность поглощать пищу, а также форма и размеры тела. Но если среда не меняется, то свойства инфузорий остаются стабильными, это и позволяет использовать их как тесты. Конечно, это относится только к инфузориям, культивируемым в лабораторных условиях.
В биотестах на инфузориях проще всего фиксировать изменение подвижности, гибель и скорость размножения. Например, можно смотреть изменение подвижности за 15–30 мин; гибель отдельных клеток за 1–4 часа; снижение скорости размножения за 1–3 суток; гибель популяции за 4–30 суток. Зачем так много параметров? Дело в том, что наблюдать за изменением подвижности недостаточно для оценки токсичности. Поскольку на движение у простейших расходуется всего 1% энергии общего обмена, то подвижность только незначительно отражает те изменения, которые происходят при отравлении токсичными агентами. Гибель отдельных клеток — достаточно надежный показатель, но с его помощью невозможно выявить низкие концентрации токсикантов. Оценка скорости размножения — биотест с большей чувствительностью, по нему можно определять и небольшие концентрации вредных веществ. Если сочетать все тесты, то результат получается надежный.
В большинстве методик биотестирования просто подсчитывают клетки до начала и в конце опыта. Как правило, это делают визуально под микроскопом, что весьма утомительно. Естественно, такая технология не нашла широкого применения.
На кафедре биохимии МГУ Прикладной биотехнологии мы разработали прибор (рис. 3), в котором клетки подсчитывает специальная компьютерная программа. Такая автоматизация существенно облегчает биотестирование, тем более что прибор позволяет использовать два вида инфузорий — Tetrahymena pyriformis и Paramecium caudatum (их размеры 0,03 × 0,05 мм и 0,06 × 0,35 мм). Тест проводят в два этапа: Paramecium caudatum, также известную как инфузория-туфелька, используют для предварительного выбора концентраций, а по динамике роста тетрахимен судят о воздействии на клетку какого-то вещества или пищевого продукта.
Двухэтапный биотест (рис. 4 и 5) подсластителей показал, что если добавлять ацесульфам калия, цикламат натрия и сахарин в концентрации 2%, то уже через 20 минут инфузории-туфельки погибают. Если уменьшить концентрацию вдвое, то все инфузории выживают во всех подсластителях. В 2%-ных растворах трех других подсластителей — глюкозы, сорбита и сукралозы выживают все инфузории в течение двух часов.
На втором этапе биотестирования, с инфузориями Tetrahymena pyriformis, использовали 1%-ные растворы. Оказалось, что самое плохое действие на тетрахимен оказывают ацесульфам калия и сахарин. Сукралоза и цикламат менее вредны, а сорбит вызывает даже больший прирост клеток, чем глюкоза. С учетом первого этапа, где цикламат действовал на клетку так же, как сахарин, этот подсластитель относим к группе добавок, негативно влияющих на клетки.
Мы попробовали исследовать также натуральные пряности. Добавление экстрактов кардамона и перца душистого стимулирует рост культуры, а экстракт мускатного ореха инфузориям не очень понравился.
Что же с усилителями вкуса? Среди них самый опасный — глутамат натрия (рис. 6): во всех пробах с этой добавкой прирост культуры клеток значительно меньше, чем в контроле.
Ранее мы говорили, что современный подход к определению безопасности пищи не позволяет точно узнать, насколько опасны смеси химических веществ. Поэтому мы предлагали инфузориям не только отдельные вещества, но и продукты (рис. 7).
Мы посмотрели, как растут тетрахимены на четырех видах мясных продуктов: колбаса вареная, буженина, ветчина варено-копченая и вареное мясо. Интересно, что буженина с таким же содержанием белка, как в вареном мясе, дала такой же маленький прирост клеток, как колбаса. Это говорит о том, что в ее состав входили пищевые добавки (скорее всего, консерванты), которые ингибируют рост и размножение клеток. В дополнение к результатам биотестирования мы посмотрели, как меняется активность пищеварительных ферментов пепсина и химотрипсина в присутствии этих мясных продуктов. Тут тоже оказалось, что активность ферментов при «переваривании» буженины меньше, чем в ветчине и вареном мясе. Получается, что качественная коптильная жидкость, которую добавляют в ветчину, да и традиционное копчение менее опасны, чем консерванты в буженине.
На инфузориях мы проверили консерванты, красители и загустители. Одновременно мы оценили активность пищеварительных ферментов в пробах, содержащих эти добавки. Результаты всех опытов хорошо соотносились друг с другом. Мы показали, что те добавки, которые вызывали уменьшение прироста культуры, как правило, действовали угнетающе на ферменты человека.
Конечно, одноклеточные тест-организмы имеют гораздо меньше ферментных систем, чем человек, и в целом устроены проще. Но без сомнения, инфузорию можно рассматривать как интегральный биологический датчик с определенными параметрами, подходящими для пищевых биотестов. Например, кислые пептидазы есть и у человека, и у инфузории. Это известно давно, а мы доказали, что различные пищевые добавки ингибируют и пепсин, и рост культуры. Системы, общие для инфузории и человека, как правило, дают схожие ответы на компоненты пищи, поэтому оценка с помощью инфузорий довольно достоверна. Выявить всю совокупность ферментных систем инфузорий, соответствующих системам человека в токсикологическом плане, — задача, которую мы сейчас решаем.
Краснокнижный вид | ООПТ России
Главная
Краснокнижный вид
Классификация: Основные группы > Algae and other protists (Водоросли и другие простейшие) > Ciliophora (Инфузории) > Oligohymenophorea > Peniculida > Stokesiidae > Stokesia
| Таксон | Stokesia vernalis |
Правовой статус
| Документ | Приложение | Дата | Номер таксона | Указан как | Статус | Категория | Дополнительно |
|---|---|---|---|---|---|---|---|
| Об утверждении перечней видов беспозвоночных животных, высших растений, грибов, лишайников и водорослей, занесенных в Красную книгу Нижегородской области | 1999-03-15 | 1 | добавлен | Д |
Описания
| Издание |
(2003) Красная книга Нижегородской области. Том 1. Животные. Нижний Новгород 380 |
| Таксон указан как | Stokesia vernalis |
| Категория | Д: Неопределенный вид, для которого нет достаточных данных, чтобы конкретизировать его статус. Один из двух видов рода. |
| Морфологическое описание |
Достаточно крупная инфузория, имеющая форму асимметричного конуса. Размеры стокезии варьируют, наиболее крупными оказываются весенние формы: диаметр клетки 175–200 мкм, длина (высота конуса) 100–150 мкм; летние формы несколько мельче – 120 и 80 мкм, соответственно. Клеточный рот расположен на плоской брюшной стороне, соответствующей основанию конуса, в виде изогнутой щели и способен округляться во время приема пищи. Величина рта составляет 1/3–1/2 длины клетки. Ресничный предротовой аппарат представлен мерцательной мембраной, тремя рядами мембранелл со свободными несклеенными ресничками и погружен внутрь тела. |
| Распространение | Известна в водоемах Западной Европы, Северной Америки. Описана в составе планктонных сообществ крупных холодноводных озер европейской части России и Сибири (Онежское, Ладожское, Отрадное, Байкал), а также верхневолжских водохранилищ и р. Волги. В центральной части Европейской России зарегистрирована в планктоне оз. Светлояр, Пустынских озер Нижегородской области, оз. Открытого и оз. Сосновского Чувашии. Более южное распространение ограничивается указанием стокезии в составе придонного комплекса инфузорий в оз. Кагул (Молдавия). |
| Образ жизни |
Места обитания. В составе планктонного сообщества пелагиали слабо проточных озер, водохранилищ, в реках как результат сноса планктона из расположенных выше по течению водоемов. Особенности биологии. Характеризуется как холодолюбивый стенотермный вид с оптимумом температуры в пределах 0–10°С. Питается бактериями и водорослями (Stephanodiscus hantzchii Grun.), часто характеризуется как активный фитофаг, по экспериментальным данным способна поедать детрит и поглощать растворенное органическое вещество. В свою очередь служит пищей для коловраток рода Asplanchna, хищных инфузорий, например, Bursaria truncatella O. F. M.. ll. В весеннем планктоне встречается в смешанных популяциях вместе с Amphileptus trachelioidesZach. Предпочитает чистые насыщенные кислородом воды (90–100% насыщения) с низким содержанием органических веществ (13–18 мг O |
| Численность |
Сведения малочисленны, тенденции изменения оценить трудно. В водоемах Нижегородской области и Чувашии является малочисленным видом инфузорного планктона: средние показатели численности составляют примерно 0,4 млн. экз./м3. Наибольшее количество инфузорий (1,2 млн. экз./м 3) отмечено в оз. Долгом при температуре воды 14°С. Появляется обычно в майском планктоне, реже в осеннем. В летние месяцы стокезия исчезает из состава инфузорных сообществ проточных озер. В глубоководных водоемах, характеризующихся наличием летней термической стратификации (залив Некрасова, Святое, Кругленькое), встречается в относительно холодном слое металимниона на протяжении всего сезона. Численность стокезии в этом случае в 4–10 раз ниже, чем весной. В составе августовского планктона оз. Светлояр стокезия отмечена при температуре 16,3°С в слое температурного скачка в количестве 0,08 млн. экз./м3. |
| Лимитирующие факторы |
Особенности биологии вида, связанные с повышенной требовательностью к условиям обитания (содержание кислорода, РОВ, скорость течения и пр. ). Эвтрофикация водоемов и нарушение гидрологического режима в результате деятельности человека. |
| Охранные меры | Принятые меры охраны. Местообитания охраняются в составе 2 ПП: «Озеро Светлояр» и «Пустынские озера». Необходимые меры охраны. Реализация мер охраны олиготрофных озер, обеспечивающих сохранение гидрологического режима данных водоемов и запрещающих загрязнение воды, что способствует сохранению данного комплекса инфузорий. |
| Ссылки | 1. Арсланова, 1983. 2. Арсланова, 1989. 3. Гаевская, 1949. 4. Лазарева, 1980. 5. Мамаева, 1979. 6. Чорик, 1967. 7. Шень Юнь Фень, 1960. 8. Noland, 1959. 9. Orr, 1954. 10. Pace, 1982. |
| Составители | Т. П. Станковска |
Как вырастить инфузорию туфельку в домашних условиях
Инфузория-туфелька – вид инфузорий, относящийся к группе альвеолят. Получила свое название за счет необычной формы, напоминающей подошву туфли. Обитает во всех пресных водоемах.
Содержание:
- 1 Описание
- 2 Зачем выращивать?
- 3 Как найти инфузорию?
- 4 Как культивировать?
- 5 Чем кормить
- 6 Использование в качестве корма
Описание
Размеры туфелек крохотные, но в то же время, относительно других одноклеточных, достаточно большие. Взрослая туфелька может достигать размеров до 0.3 мм, однако, некоторым удавалось вырастить особи в 0.6 мм . Тельце вытянутое, полукруглой формы в разрезе. Верхней оболочкой для организма служит наружная мембрана. Она прозрачна, поэтому сквозь нее можно рассмотреть все внутреннее строение инфузорий. Самым выделяющимся на фоне остальных органов является макроядро. Оно проявляется жирной точкой на теле. На поверхности туфелек располагаются реснички, при помощи которых инфузория двигается и охотится. Их количество может варьироваться от 10 до 15 тысяч.
Зачем выращивать?
Дело в том, что мальки рыб не могут есть пищу, которую с легкостью проглатывают большие особи. Для их разведения нужен особый, стартовый корм. В качестве такого корма подходят инфузории туфельки. Их разведение не составит особого труда, однако мальки, питаясь ими, будут расти более здоровыми и крепкими.
Как найти инфузорию?
Есть легкий и занятный способ найти, а главное отделить туфельку от остальных микроорганизмов:
- Возьмите кусочек стекла и поместите на него 2 капли воды, причем одна должна быть взята из аквариума, а вторая из водопроводного крана, и отстоявшаяся некоторое время.
- В каплю из аквариума добавьте несколько крупинок соли.
- Постройте тонкую «дорожку» из воды между каплями. Для этого может подойти любая иголка или зубочистка, достаточно провести ей между каплями. Все пресные микроорганизмы ринутся к чистой, несоленой воде.
- Туфелька, за счет своих ресничек, гораздо проворнее своих собратьев. Именно поэтому в водном мостике первой окажется никто иная, как породистая инфузория.
- При помощи пипетки отправляем ее в резервуар с чистой водой, для дальнейшего разведения.

Как культивировать?
Для того, чтобы развести культуру туфелек, не нужны какие-либо специальные условия, поэтому их культивация очень проста и под силу многим заводчикам рыб.
Для создания большой колонии туфелек достаточно обзавестись одной. Примерно через месяц содержания, эта туфелька разродится, и в банке будет уже колония инфузорий – более 40 тысяч экземпляров на сантиметр кубический. Это число является максимальной концентрацией туфелек в воде.
Особь инфузории нужно поместить в стеклянную банку (желательно 3-х литровую) отстоявшейся пресной воды. Стекло пропускает свет, что улучшает рост колонии. Комнатная температура отлично подходит для того, чтобы начать разведение микроорганизмов, однако идеалом для инфузорий будет 22-26 градусов. При такой температуре удастся вырастить колонию с наибольшим числом туфелек. Банку желательно поставить в проветриваемое место или снабдить продувкой. Это связано с тем, что при наличии кислорода в воде, инфузории опускаются на дно, а при его недостатке всплывают, что помогает при отслеживании и дальнейшем сборе.
Чем кормить
В еде туфельки так же неприхотливы. Прокормить их можно в домашних условиях. Для питания им нужны субстраты для развития бактерий. Едят любую растительную пищу, рыбий корм, молоко и печень. Для удобства продукты высушивают, а затем в марле опускают в резервуар с инфузориями. Чтобы их не перекармливать, достаточно будет кусочка примерно в 2-3 см.
Так же для кормежки можно использовать сенный настой. Приготовить его очень просто. В кипящую воду опустите сено, в расчете 10 г на 1 л и оставьте вариться на медленном огне в течение 20 минут. Высокая температура убьет все микроорганизмы, однако бактерии сохранятся, именно ими в дальнейшем будет питаться инфузория. Готовый раствор переливаем в любую удобную тару и оставляем при комнатной температуре на 2-3 дня, за это время бактерии размножатся, и их можно будет скармливать инфузориям. Такой вид питания называется – гидролизные дрожжи, добавлять их в воду надо из расчета 1 г на 10 л раз в неделю-полторы.
Самый легкий способ прокормить инфузорию – молоко и молочные продукты. Лучше всего подойдет обезжиренное молоко или простая сгущенка. В раствор вносить по 2 капли в неделю. Инфузории питаются не самим молоком, а кисломолочными бактериями.
При подкармливании культуры нужно помнить, что при перенасыщении раствора бактериями инфузории начнут погибать от недостаточного количества воздуха. Для избегания такой ситуации нужно тщательно следить за порциями бактерий, попадающими в резервуар к туфелькам.
Использование в качестве корма
После успешного разведения можно приступать к сбору инфузорий. Для удобства всю колонию желательно переместить на поверхность воды. Рассмотрим 2 самых удобных и легких способа это сделать:
- Сбор при помощи молока.
В воду влить молочную смесь и отключать продувку. После этого остается подождать 2 часа и инфузории сами всплывут на поверхность.
- Сбор при помощи соли.
В банку вносится раствор соли, заставляя инфузорий всплывать на поверхность.
Теперь можно приступать к самому сбору. Собрать их можно с использованием шланга. Также можно соорудить конструкцию, которая будет постоянно подкармливать мальков свежими инфузориями. Для этого понадобится обычная трубка для капельниц, которую можно приобрести в аптеке. Поставьте банку с инфузориями над аквариумом, вставьте в нее шланг, опустите и настройте подачу воды из банки при помощи зажима. В идеале вода должна подаваться каплями с интервалом в 2-3 секунды.
Такую мини-ферму по разведению инфузорий может сделать каждый в домашних условиях. Питаясь инфузориями, мальки будут расти здоровыми и крепкими, а, значит, смогут прожить долгую жизнь.
Инфузория туфелька: строение, форма тела, обитание
Жизнь на нашей планете отличается невероятным многообразием всевозможных живых организмов, имеющих подчас невероятно сложное строение. Все это многообразие жизни: от простейших насекомых и растений до нас, людей (пожалуй, самых «сложных организмов») состоит из клеток, этих маленьких кирпичиков живой материи. И если человек – венец биологической эволюции, то весьма любопытным будет рассмотреть ее начало: простейшие одноклеточные организмы, которые, по сути, на заре истории стали родоначальниками всего живого. Инфузория туфелька (наряду с
амебойи эвгленой зеленой) является одним из самых известных простых одноклеточных существ. Какое строение инфузории туфельки, среда обитания, как она питается и размножается, обо всем этом читайте далее.
Инфузория туфелька: описание и характеристика. Как выглядит инфузория туфелька?
На самом деле инфузория туфелька это вовсе не один простейший одноклеточный организм, за этим названием скрывается более 7 тысяч разных видов инфузорий. Всех их объединяет форма, которая чем-то напоминает подошву туфли, отсюда и «туфелька» в названии. (Впрочем, «туфелька» в названии прижилась только у нас, в английском языке «инфузория туфелька» значится под латинским названием «Paramecium caudatum», что переводится как «парамеция хвостатая»).
Также все инфузории обладают способностью к осморегуляции, то есть могут регулировать
давлениевнутренней среды своего организма. В этом деле им помогают две сократительные вакуоли, они сжимаются и разжимаются, таким образом, выталкивая излишки жидкости из тела инфузории.
Размеры инфузории туфельки составляют от 1 до 5 десятых миллиметра.
Фото инфузории туфельки.
Хотя инфузория туфелька и является простейшим одноклеточным существом, то есть все ее тело состоит только из одной клетки, тем не менее, она имеет способность самостоятельно дышать, питаться, размножаться, передвигаться. Иными словами, обладает всеми теми функциями и способностями, которые имеет всякое другое животное. Более того среди других простейших одноклеточных организмов именно инфузория туфелька является самой сложной. В частности среди ее
органоидов(элементов клетки) есть такие, которых нет у других ее одноклеточных «коллег»: амеб и эвглен.
Среди «передовых» органоидов инфузории можно отметить:
Обладая ртом, порошицей, пищеварительными вакуолями, инфузории практикуют голозойный тип питания, то есть захватывают органические частицы внутрь своего тела.
Так выглядит инфузория туфелька под
микроскопом.
Интересный факт: дыхание инфузории туфельки осуществляется не с помощью рта, а всем телом:
кислородчерез покровы клетки поступает в цитоплазму, где при его помощи происходит окисление органических веществ, превращение их в углекислый газ, воду и другие соединения.
Еще одной удивительной особенностью инфузории, которая ее делает «самой сложной из простейших» является наличие в ее клетке целых двух ядер. Одно из ядер большое, его зовут макронуклеусом, а второе маленькое соответственно зовется микронуклеусом. Оба ядра хранят одинаковую информацию, однако если большое ядро постоянно пребывает в работе и его информация постоянно эксплуатируется, а значит, может быть повреждена (подобно ходовым книгам в библиотеке). Если такое повреждение случается, то на этот случай как раз и предусмотрено второе маленькое ядро, служащее чем-то вроде резерва на случай сбоя основного ядра.
Как видите наша сегодняшняя героиня, инфузория туфелька, является самым совершенным среди простейших одноклеточных организмов.
Внешнее строение
По внешнему строению этот представитель фауны обладает тонкой эластичной оболочкой, именуемой в природе мембраной. Она на всем цикле жизни способна фиксировать свою сформировавшуюся форму.
Это происходит благодаря наличию цитоплазменного слоя с развитыми опорными волокнами. Такие виды волокон располагаются тесным образом к поверхности оболочки. Для инфузории характерно два ядра, одно из которых ответственно за процесс пищеварения, а другое за процесс размножения.
Вся поверхность инфузории с особыми ресничками несет ответственность за процесс передвижения. Этих ресничек больше 15 тыс. Их движения схожи с веслами. Перемещение происходит со скоростью 3 миллиметра в секунду. Инфузория передвигается с помощью волнообразных движений ресничек.
Строение инфузории туфельки
Несмотря на внешнюю простоту строение инфузории отнюдь не простое. Снаружи она защищена тонкой эластичной оболочкой, которая также помогает телу инфузории сохранять постоянную форму. Защитные опорные волокна инфузории расположены в слое плотной цитоплазмы, которая прилегает к оболочке.
Помимо этого в цитоскелет инфузории входят различные микротрубочки, цистерны альвеолы, базальные тельца с ресничками, фибриллы и филамены и другие органоиды.
По причине наличия цитоскелета инфузория в отличие от амебы не может произвольно менять форму своего тела.
Схематический рисунок строения инфузории.
Инфузория туфелька: строение
- Простейший организм инфузории-туфельки имеет и другое название – парамеция хвостатая. Ее размер от 0,1 мм до 0,5 мм. Инфузория имеет бесцветный окрас тельца состоящего из основных органоидов – внутренних двух ядер.
- Малое ядро инфузории-туфельки может быть не в одном экземпляре — исполняет роль ответственного за половую деятельность животного.
- А большое ядро – регулирует функцию приема пищи, усвоение кислорода, метаболизм и систему передвижения. Внешний край поверхности оснащен мелкими ресничками.
- Реснитчатые отростки выполняют функцию движения инфузории. Их количество достигает — около 15 тысяч. Реснитчатая ножка имеет базальное тельце у своего основания, рядом находится парасональный мешок, который заглатывается мембраной.
Строение
- Наружная часть клетки имеет тонкую оболочку, выполняющую функцию защиты и сохранности целостности формы. Кроме вышеуказанных составляющих, инфузория содержит: цистерны альвеолы, филомены, микротрубочки, фибриллы.
- Цитоскелет позволяет инфузории сохранять форму тельца в первоначальном виде.
- У туфельки существуют: ротовое отверстие, глотка, и зона для выведения фрагментов переработки пищи – порошица. Ее сократительные вакуоли имеют приводящие каналы.
Класс инфузории туфельки
Также строение инфузории зависит от ее класса. Так различают два класса инфузории туфельки:
- ресничные инфузории,
- сосущие инфузории.
Далее подробно остановимся на них.
Ресничные инфузории
Названы так, поскольку их тело покрыто маленькими ресницами, которые также именуются цилиями. Длина ресницы составляет не более 0,1 микрометра. Ресницы могут, как распределятся равномерно по телу нашей простейшей красавицы, так и собираться в пучки, которые биологи называют «цирры». Сами ресницы представляют собой пучок фибрилл, которые являются нитевидными белками.
Каждая ресничная инфузория может иметь несколько тысяч таких вот ресниц. Передвижение инфузории также осуществляется при помощи ресниц.
Сосущие инфузории
Сосущие инфузории совсем не имеют не только ресничек, но и рта, глотки и пищеварительных вакуолей, столь характерных для их «волосатых» сородичей. Зато у них есть своеобразные щупальца, представляющие собой плазматические трубочки. Именно эти щупальца-трубочки у сосущих инфузорий выполняют функцию рта и глотки, так как захватывают и проводят питательные вещества в эндоплазму клетки.
Не имея ресниц сосущие инфузории не способны передвигаться. Впрочем, им это и не нужно, имея особую ножку-присоску, они прикрепляются к коже какого-нибудь краба или рыбы и на них живут. Сосущих инфузорий всего лишь несколько десятков видов, против тысячи видов их ресничных собратьев.
Способы размножения инфузории туфельки
Обычный способ – бесполый, происходит путем деления клетки на две равные части. Во время активного движения, тело туфельки вытягивается в длину и строго по его середине образуется углубляющая перетяжка – место, в котором и происходит в дальнейшем деление инфузории на два организма. С появлением перетяжки изменяется и строение инфузории туфельки – образуются по две глотки, ротовых отверстия и перистомы. В каждой новой клетке имеется по одному большому и малому ядру, часть органоидов материнской клетки, недостающие образуются заново. Бесполое размножение происходит довольно быстро – всего за сутки из одной инфузории 2 – 4 одноклеточных животных.
Половое размножение – не часто используемый способ, но возможный в случае температурных изменений и недостатке пищи. По завершении процесса, инфузория туфелька зачастую перевоплощается в цисту. Две инфузории на время соединяются, в месте соприкосновения растворяется оболочка, возникает своеобразный мостик. В это время каждая инфузория лишается большого ядра, малое делится дважды, образуя 4 дочерних ядра, три из которых исчезают, последнее снова разделяется на равные части. В завершении этого в теле каждой инфузории имеется по 2 ядра. Каждая из клеток обменивается с другой одним ядром, которые перемещаются по соединительному мостику. Попав в тело другой инфузории, ядро сливается там с еще одним – «родным» этому организму. Получившиеся ядра снова разделяются на большое и малое ядро, как это и должно быть, инфузории рассоединяются. Данный способ размножения имеет название конъюгация, длительность его составляет 12 часов. Несмотря на то, что новой клетки в этом случае не образуется, польза от такого процесса —, обмен генетического материала между особями, что ведет к перераспределению наследственности, обновлению и увеличению жизнестойкости организмов.
Среда обитания инфузории туфельки
Инфузории туфельки обычно живут в мелких пресных водоемах со стоячей водой и гниющей органикой. Стоячая вода им необходима, чтобы не преодолевать силу течения, которая их снесет, поэтому инфузорий нет в реках. В мелких водоемах Солнце достаточно прогревает воду, и гниющая органика служит источником их пищи. К слову по насыщенности того или иного водоема инфузориями можно судить о степени его загрязнения, чем их больше, тем более грязный водоем.
А вот соленую воду инфузории не любят, поэтому их нет в морях и океанах.
Среда обитания и строение инфузории-туфельки
Инфузория туфелька обитает в небольших пресных водоемах с загрязненной, застоявшейся водой — в прудах, канавах, лужах, то есть там же, где обитают и другие простейшие — амеба и эвглена зеленая. Она постоянно находится в движении, плавая в поисках пищи. Передвигаться в воде ей помогают специальные органы движения — реснички, которые покрывают все ее вытянутое, похожее на туфельку, тело. Ресничек у инфузории около 15 тысяч, в основании каждой реснички лежит базальное тельце. Реснички колеблются синхронно, примерно 30 раз в одну секунду, напоминая весла лодки.
Благодаря согласованной работе ресничек инфузория-туфелька плывет со скоростью до 2,5 мм в секунду, одновременно вращаясь вокруг продольной оси. Если учесть, что размер инфузории всего 0,5 мм, это довольно высокая скорость. На замедленной съемке видно, что движение каждой реснички состоит из резкого взмаха в одном направлении и более плавного возвращения назад.
Сохранять форму инфузории туфельке помогает эластичная оболочка, а также опорные волоконца, располагающиеся в прилегающем к оболочке слое цитоплазмы. Кроме того, под оболочкой у инфузории туфельки имеются специальные органы защиты — трихоциты. Трихоциты — это упругие длинные нити, сжатые до состояния коротких палочек и расположенные перпендикулярно поверхности тела инфузории. Если на инфузорию нападают, трихоциты резко расправляются и «выстреливают» в сторону нападения. На месте использованных трихоцитов образуются новые.
Еще один отличительный признак инфузории-туфельки — это наличие у нее двух ядер: большого и маленького. Большое ядро отвечает за питание, дыхание, обмен веществ, а также за движения. Малое ядро участвует в половом размножении инфузории-туфельки.
У инфузории-туфельки имеется также клеточный рот, две сократительные вакуоли на переднем и заднем концах тела и пищеварительные вакуоли.
Питание инфузории туфельки
Чем питается инфузория туфелька? Питание инфузории зависит от ее класса. Так сосущие инфузории являются подлинными хищниками одноклеточного мира: источником их пищи служат другие более мелкие одноклеточные организмы, на свою беду проплывающие мимо. Своими щупальцами сосущие инфузории хватают других одноклеточных. Изначально жертва захватывается одним щупальцем, а потом «к столу» подходят и другие «собратья». Щупальца растворяют клеточную оболочку жертвы и поглощают ее внутрь.
А вот ресничная инфузория в этом плане «вегетарианка», источником ее пищи обычно служат одноклеточные водоросли, которые захватываются ротовым углублениями, оттуда они попадают в пищевод, а потом к пищеварительным вакуолям. Переработанная пища выбрасывается через порошицу.
Интересный факт: во рту ресничной инфузории также имеются реснички, которые колышась, создают течение, чем увлекают частицы пищи в ротовую область.
Питание инфузории
Питание инфузории зависит от ее класса. Хищные сосальщики орудуют щупальцами. К ним прилипают, присасываются, проплывающие мимо одноклеточные. Питание инфузории туфельки осуществляется за счет растворения клеточной оболочки жертвы. Пленка разъедается в местах контакта со щупальцами. Изначально жертва, как правило, захватывается одним отростком. Прочие щупальца «подходят к уже накрытому столу».
Реснитчатая форма инфузории туфельки питается одноклеточными водорослями, захватывая их ротовым углублением. Оттуда еда попадает в пищевод, а затем, в пищеварительную вакуоль. Она закрепляется на коне «глотки», отцепляясь от нее каждые несколько минут. После, вакуоль проходит по часовой стрелке к заду инфузории. Во время пути цитоплазмой усваиваются полезные вещества пищи. Отходы выбрасываются в порошицу. Это отверстие, подобное анальному.
Во рту инфузории тоже есть реснички. Колышась, они создают течение. Оно увлекает частицы пищи в ротовую полость. Когда пищеварительная вакуоль перерабатывает еду, образуется новая капсула. Она тоже стыкуется с глоткой, получает пищу. Процесс цикличен. При комфортной для инфузории температуре, а это около 15 градусов тепла, пищеварительная вакуоль образуется каждые 2 минуты. Это указывает на скорость обмена веществ туфельки.
Размножение инфузории туфельки
Размножение инфузории может быть как половым, так и бесполым – посредством деления клетки.
- Половое размножение: при нем две инфузории сливаются боковыми поверхностями, при этом оболочки между слитыми поверхностями растворяются, и образуется своеобразный цитоплазматический мостик. Через этот мостик клетки обмениваются ядрами. Большие ядра при этом вовсе растворяются, а маленькие дважды делятся. Затем из полученных четырех ядер, три исчезает, а оставшееся ядро снова делится надвое. Обмен оставшимися ядрами происходит по цитоплазматическому мостику. Из полученного материала возникают вновь рожденные ядра, и большие, и маленькие. Затем инфузории расходятся друг с другом.
- Бесполое размножение инфузории посредством деления намного проще. При нем оба ядра клетки делятся на два, как и другие органоиды. Таким образом, из одной инфузории образуется две, каждая с полным набором необходимых органоидов.
Размножение
Бесполое
Инфузория обычно размножается бесполым путём — делением надвое. Ядра делятся на две части, и в каждой новой инфузории оказывается по одному большому и по одному малому ядру. Каждая из двух дочерних получает часть органоидов, а другие образуются заново.
Размножение инфузории-туфельки
Половое
При недостатке пищи или изменении температуры инфузории переходят к половому размножению, а затем могут превратиться в цисту.
При половом процессе увеличения числа особей не происходит. Две инфузории временно соединяются друг с другом. На месте соприкосновения оболочка растворяется, и между животными образуется соединительный мостик. Большое ядро каждой инфузории исчезает. Малое ядро дважды делится. В каждой инфузории образуются четыре дочерних ядра. Три из них разрушаются, а четвёртое снова делится. В результате в каждой остаётся по два ядра. По цитоплазматическому мостику происходит обмен ядрами, и там сливается с оставшимся ядром. Вновь образовавшиеся ядра формируют большое и малое ядра, и инфузории расходятся. Такой половой процесс называется конъюгацией. Он длится около 12 часов. Половой процесс ведёт к обновлению, обмену между особями и перераспределению наследственного (генетического) материала, что увеличивает жизнестойкость организмов.
Жизненный цикл инфузории-туфельки
Рекомендованная литература и полезные ссылки
- Ehrenberg C. G. Dritter Beitrag zur Erkenntniss grosser Organisation in der Richtung des kleinsten Raumes (нем.) // Abhandlungen der Koniglichen Akademie der Wissenschaften zu Berlin. Aus dem Jahre 1833 : magazin. — Leipzig, 1835. — S. 268—269, 323.
- Ehrenberg C. G. 502. Paramecium caudatum, geschwanztes Pantoffelthierchen // Die Infusionsthierchen als volkommene Organismen. — Leipzig, 1838. — P. 351—352.
- Полянский Ю. И. Подцарство Простейшие, или Одноклеточные (Protozoa) // Жизнь животных / под ред. Ю. И. Полянского, гл. ред. В. Е. Соколов. — 2-е изд. — М.: Просвещение, 1987. — Т. 1. Простейшие. Кишечнополостные. Черви. — С. 95—101. — 448 с.
- Warren, A. (2015). Paramecium caudatum Ehrenberg, 1833. In: Warren, A. (2015) World Ciliophora Database. — WoRMS — World Register of Marine Species
определение инфузории в The Free Dictionary
Также найдено в: Тезаурус, Медицина, Энциклопедия, Википедия.
Связанные с инфузориями: Ciliophora, paramecium, Sporozoa
cil·i·ate
(sĭl′ē-ĭt, -āt′)прил.
Реснитчатый.
н.
Любое из различных простейших филума Ciliophora, характеризующееся многочисленными ресничками.
секция н.
Словарь английского языка American Heritage®, пятое издание. Авторские права © 2016, издательство Houghton Mifflin Harcourt Publishing Company. Опубликовано издательством Houghton Mifflin Harcourt Publishing Company. Все права защищены.
реснитчатый
(ˈsɪlɪɪt; -eɪt)прил
1. (зоология) Также: ресничный обладающий ресничным эпителием или относящийся к нему: реснитчатый эпителий.
2. (Animals) of or relating to protozoans of the phylum Ciliophora , which have an outer layer of cilia
n
(Animals) a protozoan of the phylum Ciliophora
ˌciliˈation нет
Collins English Dictionary – Complete and Unabridged, 12th Edition 2014 © HarperCollins Publishers 1991, 1994, 1998, 2000, 2003, 2006, 2007, 2009, 2011, 2014
cil•i•ate
(ˈɪs)н.
1. любое простейшее из филума Ciliophora, характеризующееся ресничками, покрывающими все тело или его часть.
прил. 2. Также cil•i•at•ed (ˈsɪl iˌeɪ tɪd) с ресничками.
[1785–95; < Новая латиница ciliātus = cili(a) cilia + -ātus -ate 1 ]
cil`i•ation, сущ.
Рэндом Хаус Словарь колледжа Кернермана Вебстера, © 2010 K Dictionaries Ltd. Авторские права Random House, Inc., 2005, 1997, 1991. Все права защищены.
ТезаурусАнтонимыРодственные словаСинонимы Легенда:
Перейти к новому тезаурусу0109
инфузории — любой представитель подкласса Infusoria
парамеции, paramecium – любой представитель рода Paramecium
tetrahymena – родственники paramecium; часто используется в генетических исследованиях
stentor – любой из нескольких трубкообразных простейших инфузорий, входящих в род Stentor
vorticella – любой из различных простейших, имеющих прозрачное бокаловидное тело с втягивающимся стеблем
. cilial, ciliary
ciliary
реснитчатый
шероховатый — края листовидной формы; с обрезанным, бахромчатым или зубчатым краем
На основе WordNet 3.0, коллекции клипартов Farlex. © 2003-2012 Принстонский университет, Farlex Inc.
Переводы
ripsieläin
trilhaardiertje
Упоминается в ?
- breakstone
- cilial
- ciliary
- Ciliata
- ciliated
- ciliated protozoan
- Ciliophora
- ciliophoran
- class Ciliata
- class Ciliophora
- class Flagellata
- class Mastigophora
- class Sarcodina
- класс Sporozoa
- конъюгация
- конъюгация
- зубчато-реснитчатые
- жгутиковые
- род Paramecium
Ссылки в архиве периодических изданий ?
Эта некультивируемая инфузория показала 98,9% идентичность и 100% покрытие запросов согласно опубликованному P. менее вероятно, что их упустят из виду, Трибби обнаружил зеленую водоросль Ankistrodesmus и инфузорию Uronema, ни одна из которых не была зарегистрирована в 2010 г.
Протисты и беспозвоночные во временных водоемах на Enchanted Rock, округ Ллано, Техас: 1965 и 2010
Несмотря на желание подавляющего большинства афганцев вести переговоры с Талибаном, США все еще не готовы к примирению с бескомпромиссными талибами и в лучшем случае соглашается на их реинтеграцию, и то тоже при определенных условиях.
Обзор войны с террором в декабре 2010 г.
Диагноз: голова в белых и черных чешуйках, щупики в коричневатых и беловатых смешанных чешуйках, с ворсинчатыми чешуйками по внешней границе, внутренняя кайма с белыми ворсинчатыми чешуйками, усики двупильчатые, глаза реснитчатые, темя с беловатым, черноватым и с чешуйками и черной полосой поперек, тегулы и патагии покрыты белым цветом с примесью темно-коричневых и коричневатых чешуек и ворсинчатых чешуек.
Dos nuevas especies del genero Argyrana (Lepidoptera: Noctuidae)
Использование описательной системы мионемных волокон, разработанной для другой перитриховой инфузории Campanella umbellaria (Shi et al.
Субклеточная локализация центрина в отдельных компартментах Vorticella convallaria coli, a
3 90 , размножается бесполым путем делением и половым путем конъюгации
Глава 6 Паразитические зоонозы
Инфузионные эпибионты, паразитирующие на планктонных ракообразных (Fernandez-Leborans & Tato-Porto, 2000), также были обнаружены в Балтийском море (Hirche, 1974; Виктор и Краевская-Солтыс, 1994).
Пагубное влияние перитриховых инфузорий (Epistylis sp.) как эпибионтов на выживаемость копепод Acartia bifilosa/Aerjalalise Acartia bifilosa isenditel esinevate ripsloomadest epibiontide (Epistylis sp.) kahjustav moju
Определитель видов 1 Клипеальные боковые края неклиновидные; переднеспинка грубо и довольно густо пунктирована; междоузлия надкрылий слегка выпуклые, микросетчатые, поэтому мало блестящие, тонко отчетливо и довольно густо пунктированные.
Систематический пересмотр рода Cephalocyclus с описанием десяти новых видов из Мексики и Коста-Рики (scarabaeoidea: aphodiidae)
Инфузория Toona, принадлежащая к семейству Meliaceae, является полезным местным деревом в районе Гималаев.
Морфогенез in vitro Toona ciliata a partir de raquis de hojas jovenes con tidiazuron. Tetrahymena pyriformis (Niculescu et al.
Валидация моделей (Q)SAR
sp., инфузория с наружным слоем органических чешуек (Ciliophora, Litostomatea, Haptoria).
Пресноводные гелиозоа (protista, heliozoa) из Индианы
Лечение EOM 0,2 увеличило кратность изменения инфузорий простейших (p<0,001), Butyrivibrio fibrisolvens (лечение, линейный и квадратичный эффект: p<0,001, 0,002 и 0,007 соответственно) , грибы (лечение и линейный эффект: p = 0,001 и <0,001 соответственно) и Ruminococcus flavefaciens (лечение, линейный и квадратичный эффект: p<0,001, <0,0001 и 0,002 соответственно).
Смесь эфирных масел на ферментацию рубца и микробное сообщество — исследование in vitro
Браузер словарей ?
- ▲
- cigarette case
- cigarette end
- cigarette holder
- cigarette lighter
- cigarette paper
- cigarette smoker
- cigarfish
- cigarillo
- cigarlike
- cigar-shaped
- cigar-store Indian
- ciggie
- ciggy
- cigrinophily
- CIGS
- ciguatera
- ciguatoxin
- cilantro
- cilia
- cilial
- ciliary
- ciliary artery
- ciliary body
- ciliary veins
- Ciliata
- ciliate
- реснитчатые
- реснитчатые простейшие
- реснитчатые
- киликийские
- киликийские
- киликийские ворота
- реснитчатые
- Cilicism
- Ciliform
- Cilioflagellata
- Ciliograde
- ciliolate
- Ciliophora
- ciliophoran
- cilium
- cill
- Çiller
- Cillosis
- cilostazol
- CIM
- Cima
- Cimabue
- Cimabue Giovanni
- Cimar
- Cimarosa
- ▼
Полный браузер ?
FA108/FA108: Общие паразиты пресноводных рыб Иллюстрированное руководство: подвижные инфузории
Дебора Б. Паудер, Эрик В. Кертис и Рой П. Э. Янонг 2
Введение
Эта публикация является одной из серии иллюстрированных руководств, предназначенных для помощи в идентификации распространенных паразитов пресноводных рыб. Публикации, включенные в эту серию:
Иллюстрированное руководство по распространенным паразитам пресноводных рыб: Сидячие инфузории
Иллюстрированное руководство по паразитам распространенных пресноводных рыб: подвижные инфузории
Иллюстрированное руководство по распространенным паразитам пресноводных рыб: жгутиконосцы
Иллюстрированное руководство по распространенным паразитам пресноводных рыб: динофлагелляты, кокцидии, микроспоридии и миксозои
Иллюстрированный справочник по распространенным паразитам пресноводных рыб: моногенеи
Иллюстрированное руководство по распространенным паразитам пресноводных рыб: дигенейские трематоды
Иллюстрированное руководство по паразитам распространенных пресноводных рыб: нематоды
Иллюстрированное руководство по паразитам распространенных пресноводных рыб: скребни, цестоды, пиявки и пентастомы
Иллюстрированное руководство по паразитам распространенных пресноводных рыб: ракообразные
Информация, представленная в этом руководстве, не предназначена для полного и подробного описания каждого паразита или группы паразитов и их характеристик, а скорее предназначена для помощи в визуальной идентификации некоторых из наиболее распространенных видов или групп паразитов, встречающихся в пресноводная рыба. Для получения дополнительной информации о каждом паразите обратитесь к публикациям в разделах «Рекомендуемая литература» и «Справочник» ниже.
Информация о руководстве
Следующая информация предоставляется для каждого паразита в руководстве.
Ткань-мишень: указывает местонахождение на/внутри рыбы, где чаще всего встречается паразит.
Характеристика: дает краткое описание внешнего вида паразита.
Размер: указывает размер или диапазон размеров паразита. (1 мкм = 0,001 мм = 0,0001 см) (мкм = микрон или микрометр; мм = миллиметр; см = сантиметр)
Движение: указывает тип движения паразита, если таковой имеется.
Примечание: содержит краткий интересный комментарий о паразите.
Подвижные инфузории
Хилодонелла
Целевые ткани: Кожа, плавники, жабры
Внешний вид: Яйцевидная или почковидная форма с полосами ресничек вдоль длинной оси; дорсовентрально уплощенный; кажется полупрозрачным
Размер: Прибл. 30–80 мкм x 20–60 мкм
Движение: Свободное перемещение
Примечание: Часто вызывает чрезмерное выделение слизи у рыб
Ихтиофтириус многофилис
Целевые ткани: Кожа, плавники, жабры
Внешний вид: Амебоид; С-образное ядро встречается только у «взрослых»
№Размер: Trophont ок. 50 мкм–1 мм; ок. 30 мкм x 50 мкм
Механизм: Свободный ход; перекатывание, амебоидное движение
Примечание: Жизненный цикл включает стадии на рыбе и вне ее
Тетрагимена
Целевые ткани: Кожа, плавники, жабры, глаза, внутренние органы
Внешний вид: Яйцевидная форма с равномерно распределенными ресничками
Размер: Прибл. 55 мкм x 30 мкм
Механизм: Свободно движущийся Примечание: Обычно встречается на мертвой рыбе или детрите или в воде с высокой концентрацией органических веществ
Триходина
Целевые ткани: Кожа, плавники, жабры
Внешний вид: Форма «летающая тарелка» или «скребущие пузыри»
Размер: Прибл. 35–135 мкм в диаметре
Механизм: Свободный ход; вихревое движение
Примечание: Обычен в воде с высокой концентрацией органических веществ
Благодарности
Авторы благодарят Грега Вермеера за фотографию, которую он предоставил для этой публикации.
Рекомендуемое чтение
UF/IFAS Циркуляр 91 Заражение рыб нематодами (аскаридами). https://edis.ifas.ufl.edu/fa091
UF/IFAS Циркуляр 120 Вопросы управления здоровьем рыб в замкнутых системах аквакультуры – Часть 1: Введение и общие принципы. https://edis.ifas.ufl.edu/fa099
UF/IFAS Циркуляр 121 Вопросы управления здоровьем рыб в замкнутых системах аквакультуры – Часть 2: Патогены. https://edis.ifas.ufl.edu/fa100
UF/IFAS Циркуляр 122 Вопросы управления здоровьем рыб в замкнутых системах аквакультуры – Часть 3: Общие рекомендации и подходы к решению проблем. https://edis.ifas.ufl.edu/fa101
UF/IFAS Циркуляр 920 Ichthyophthirius multifiliis (белая пятнистость) Инфекции у рыб. https://edis.ifas.ufl.edu/fa006
Циркуляр UF/IFAS 921 Введение в управление здоровьем рыб. https://edis.ifas.ufl.edu/fa004
Информационный бюллетень UF/IFAS FA-13 Использование меди в пресноводной аквакультуре и фермерских прудах. https://edis.ifas.ufl.edu/fa008
Информационный бюллетень UF/IFAS FA-90 Пентастомидные инфекции у рыб. https://edis.ifas.ufl.edu/fa090
Информационный бюллетень UF/IFAS FA-107 Иллюстрированное руководство по распространенным паразитам пресноводных рыб: Сидячие инфузории. https://edis.ifas.ufl.edu/fa107
Информационный бюллетень UF/IFAS FA-109 Иллюстрированное руководство по распространенным паразитам пресноводных рыб: жгутиконосцы. https://edis.ifas.ufl.edu/fa109
Информационный бюллетень UF/IFAS FA-110 Иллюстрированное руководство по распространенным паразитам пресноводных рыб: динофлагелляты, кокцидии, микроспоридии и миксозои. https://edis.ifas.ufl.edu/fa110
Информационный бюллетень UF/IFAS FA-111 Иллюстрированный справочник по распространенным паразитам пресноводных рыб: моногенеи. https://edis.ifas.ufl.edu/fa111
Информационный бюллетень UF/IFAS FA-112 Иллюстрированное руководство по распространенным паразитам пресноводных рыб: дигенейские трематоды. https://edis.ifas.ufl.edu/fa112
Информационный бюллетень UF/IFAS FA-113 Справочник по распространенным паразитам пресноводных рыб: Нематоды. https://edis.ifas.ufl.edu/fa113
Информационный бюллетень UF/IFAS FA-114 Иллюстрированный справочник по распространенным паразитам пресноводных рыб: скребни, цестоды, пиявки и пентастомы. https://edis.ifas.ufl.edu/fa114
Информационный бюллетень UF/IFAS FA-115 Иллюстрированный справочник по распространенным паразитам пресноводных рыб: Ракообразные. https://edis.ifas.ufl.edu/fa115
UF/IFAS Fact Sheet VM-104 Cryptobia iubilans у цихлид. https://edis.ifas.ufl.edu/vm077
Ссылки
Hoffman, G.L. 1999. Паразиты североамериканских пресноводных рыб . Итака, Нью-Йорк: Издательство Корнельского университета.
Лонгшоу М. и С. В. Файст. 2001. Паразитарные болезни. Страницы 167–183 в W.H. Уайлдгус, редактор. Руководство BSAVA по декоративным рыбам, второе издание. Британская ветеринарная ассоциация мелких животных, Глостер, Англия.
Noga, E. J. 1996. Болезни рыб: диагностика и лечение . Сент-Луис, Миссури: Mosby-Yearbook, Inc.
Stoskopf, M.K. 1993. Fish Medicine . Филадельфия, Пенсильвания: Компания WB Saunders.
Ву, П. Т. К., редактор. 1995. Болезни и расстройства рыб , том 1: инфекции простейших и многоклеточных животных. CAB International, Уоллингфорд, Великобритания.
Сноски
1. Это документ FA108, один из серии Школы лесных ресурсов и охраны, Программы рыболовства и водных наук, Расширение UF/IFAS. Исходная дата публикации — июль 2005 г. Пересмотрен в марте 2020 г. Посетите веб-сайт EDIS по адресу https://edis.ifas.ufl.edu, чтобы узнать о поддерживаемой в настоящее время версии этой публикации.
2. Дебора Б. Паудер, координатор исследовательских программ и услуг; Эрик В. Кертис, бывший ученый-биолог; и Рой П. Э. Янонг, профессор; Лаборатория тропической аквакультуры, Школа лесных ресурсов и программы сохранения в области рыболовства и водных наук, UF/IFAS Extension, Gainesville, FL 32611.
инфузорий — они пришли из пруда
ноябрь 182015
Я наконец опознал предмет, который нашел у себя во рту почти 25 лет назад. К сожалению, решение этой старой загадки привело меня к новой, касающейся простейших, обычно встречающихся в задней кишке тараканов и термитов.
Я наткнулся на него осенью 1990 года. Моя жена была студенткой первого курса медицинского факультета и одолжила микроскоп для изучения образцов тканей человека для курса гистологии. В те дни меня больше интересовала поэзия, чем простейшие, но еще больше меня интересовала прокрастинация. Итак, однажды утром я отложил в сторону свою докторскую диссертацию и попробовал одолженный прицел.
Ее подготовленные гистологические препараты — фиксированные клетки, окрашенные в бледно-розовые и бледно-голубые цвета — ненадолго привлекли мое внимание. Я хотел увидеть что-то живое и извивающееся. Я читала о пылевых клещах, поэтому нанесла на предметное стекло пух с пола, но это меня разочаровало: ничего, кроме волос, чешуек кожи и цветных волокон ткани. Затем я попробовал немного воды из суповой кастрюли, которая стояла в раковине всю ночь, и она была немного лучше. Там были бактерии, тысячи их, толкающиеся и трясущиеся в молочном бульоне. Но бактерии скучны.
Реконструкция Существа. (Стрелки должны показать, как она двигалась. Да, похоже, что у клетки есть руки, но мне не хочется это снова рисовать).
Наконец, я вытер немного слюны с задней части рта, и это было больше похоже на то. Если вы когда-нибудь этим занимались, то знаете, что я обнаружил: большие рваные эпителиальные клетки ротовой полости, водовороты интригующей органики, пухлые лейкоциты, которые недавно съели бактериальную муку. Затем в поле зрения всплыло что-то действительно странное. Оно было круглым и пестрым, как счастливое лицо дегенерата с пучком длинных волос, торчащих с одной стороны. Он вращался взад-вперед с гипнотическим, раскачивающимся движением: по часовой стрелке, затем против часовой стрелки, затем снова по часовой стрелке. С каждой сменой направления пряди двигались вместе по ленивой дуге.
Удивительно! Паразит жил у меня во рту, и у него были волосы, как у рок-звезды. Что бы это могло быть? Я поймал автобус до Библиотеки наук о жизни Макгилла и вытащил из стопки несколько текстов по паразитологии. Из них я определил, что это существо должно быть каким-то «жгутиконосцем» (это слово было для меня новым). Я пролистал фотографии, размытые микрофотографии Trichomonas , Chilomastix , Giardia , Retortamonas и всех их проблемных собратьев, но нигде не мог найти своего парня. Все известные жгутиковые паразиты были либо слишком заостренными, либо слишком заостренными, и ни у одного из них не было пышной шевелюры, которую я видел на своем существе.
Через пару ночей к нам вломился грабитель и украл микроскоп, что положило конец моей любительской паразитологии на пару десятков лет. Я вернулся к своей настоящей работе, но никогда не забывал, что внутри моего рта живут миниатюрные глэм-рокеры. Много лет спустя, когда я вернулся к микроскопии (подняв прокрастинацию на совершенно новый уровень), первое, что я сделал, это снова попытался найти этих жгутиконосцев. Я провел тампоном по нёбу, поскреб языком и сплевывал слайд за слайдом, но безуспешно. К тому времени я узнал гораздо больше о протистах и приобрел хорошую библиотеку полевых справочников и текстов по протозоологии, ни в одном из которых не было ничего похожего на тот организм, который я видел. Я начал сомневаться в своей памяти.
Затем, всего два дня назад, друг из Уругвая прислал мне ссылку на видео из Мексики, показывающее предполагаемого простейшего, найденного в бронхиальном лаваже пациента, который чуть не утонул в реке. И вот оно снова, Вещь из моих уст!
Это раскачивающееся движение — именно то, что я видел в 1990 году. Видео было размещено на YouTube как неизвестное «ciliado» (инфузория), и внешне оно действительно похоже на него. Сначала он показался мне перезрелым олиготрихом, возможно, раздутым от осмотического давления и плохо двигающимся. Но путь его движения выглядел совершенно не так, даже для нездорового экземпляра. Это повторяющееся, неэффективное раскачивающееся движение не было похоже на знакомое движение сросшихся ресничек (адоральные поликинетиды), которые окружают клеточный рот олиготриховых инфузорий, или на какие-либо венчики и бахромы ресничек, которые я знал у других свободноживущих цилиофор. Раскачивание предполагало, что что-то сидячее оторвалось от субстрата, что напомнило мне, что эта «инфузория» была обнаружена в жидкости бронхоальвеолярного лаважа (БАЛ). Как это бывает, у всех нас есть реснитчатые клетки, выстилающие некоторые из наших носовых и бронхиальных полостей, которые выполняют полезную задачу по очистке легких от слизи и мусора. Реснитчатые клетки остаются неподвижными на одном месте, и отходы как бы «толпой» скользят по ним, переходя от одной клетки к другой на пути к выходу. Может ли это быть отслоившейся бронхиальной клеткой?
Поиск на YouTube дал другое видео, на этот раз из лаборатории диагностической бактериологии Сингапурской больницы общего профиля. На нем показан большой скопление бронхиального эпителия с активно работающими ресничками:
. плавающая ячейка. Тем не менее, меня убедили, что клетки на обоих видео, а также на том, что я видел еще в 1990, все реснитчатые носовые или бронхиальные клетки.
И это подводит меня к новой загадке. Комментарий ко второму видео отправил меня на интригующую запись в патологоанатомическом блоге под названием Microcosm, где я узнал кое-что захватывающее и, извините, немного отвратительное. Кажется, были медицинские сообщения о случаях заражения людей простейшими с пышными жгутиками под названием Lophomonas blattarum , которые обычно живут комменсально в кишечнике тараканов или термитов.
Lophomonas blattarum, из William Saville Kent, A Manual of Infusoria, 1881.
L. blattarum известен с 1860 года, когда он был обнаружен и описан великим Самуэлем Фридрихом Штейном. Сообщения о респираторных инфекциях у людей появились совсем недавно, начиная с 1993 года, когда первый случай был зарегистрирован в Китайском журнале паразитологии и паразитарных заболеваний. С тех пор было сообщено о многих десятках подобных случаев, что привело к растущему убеждению некоторых исследователей в том, что Lophomonas blattarum представляет собой «потенциально важную причину бронхолегочной инфекции и респираторных симптомов». (Мартинес-Хирон и ван Вурден, 2013а).
Проблема в том, что это может быть вовсе не настоящая инфекция. Во всех опубликованных сообщениях о случаях инфекции Lophomonas микроорганизмы были идентифицированы с помощью световой микроскопии либо из живых образцов, либо из окрашенных препаратов. На сегодняшний день молекулярных исследований предполагаемого паразита не проводилось. Идентификация была полностью основана на морфологических особенностях и симптомах.
Было проведено одно ультраструктурное исследование, но оно не подтверждает теорию о том, что лофомониаз является настоящим инфекционным заболеванием. Статья с 12 авторами была опубликована на китайском языке, но из аннотации на английском языке (и путем кропотливой загрузки текста в Google Translate по одному абзацу за раз) я понял, что исследователи использовали оптическую и электронную микроскопию для изучения подвижных клеток из ЖБАЛ 6 пациентов, которые оказались морфологически непохожими на Lophomonas . Они также проанализировали имеющуюся литературу о случаях лофомониаза в Китае и пришли к следующему выводу: «За последние 20 лет все диагностированные случаи легочного Инфекция Lophomonas blattarum , зарегистрированная в нашей стране, была неправильно диагностирована. В настоящее время нет доказательств того, что Lophomonas blattarum является патогеном, вызывающим легочную инфекцию». (Mu XL, et al., 2013) Согласно этому исследованию, Lophomonas инфекция s , зарегистрированная в литературе, на самом деле была ошибочно идентифицирована бронхиальными клетками.
Типично нечеткая микрофотография клеток, идентифицированных как Lophomonas blattarum в He et al, 2011. Martinez-Girón и van Woerden считают, что это ошибочно идентифицированные бронхиальные эпителиальные клетки.
Другие исследователи, соглашаясь с тем, что произошла путаница между эпителиальными клетками и Lophomonas, продолжают утверждать, что многие случаи инфекции являются подлинными. Мартинес-Хирон и ван Вурден, опубликовавшие о лофомониазе больше, чем кто-либо, ответили своим китайским коллегам, подтвердив свое мнение о том, что «наблюдение под световой микроскопией этого многожгутикового простейшего у пациентов с симптомами, которые положительно реагируют на антипротозойную терапию, может разумно охарактеризовать как бронхолегочный лофомониаз». (Мартинес-Хирон и ван Вурден, 2013b)
Тонкая структура Lophomonas blattarum (Beams et al, 1961, адаптировано из Kudo). Обратите внимание на чашечку в форме бокала (CY), аксиальную нить (AXF) и парабазальный «воротник» канальцев (PNT), все они видны в световой микроскоп.
Как человека, который обычно больше интересуется таксономией организмов, чем патологиями, которые они вызывают, я поражен тем, что клиницисты делают акцент на болезни и симптомах, а не на самом существе. С клинической точки зрения дело в инфекции веское: у данного пациента имеются респираторные симптомы, в жидкости бронхиального лаважа наблюдаются явно жгутиковые клетки, и состояние улучшается при лечении метронидазолом, который, в конце концов, является предпочтительное лечение от другого паразита метомонады, Вагинальная трихомонада . Все это важные факты, от которых трудно отмахнуться, если подойти к этому вопросу с клинической точки зрения. Однако с чисто протистологической точки зрения мы могли бы отметить, что идентификация с помощью световой микроскопии часто ненадежна, особенно с такими маленькими клетками; микрофотографии в опубликованной литературе обычно довольно плохие и могут изображать клетки в неестественном состоянии; морфологические признаки, используемые в этих тематических исследованиях, имеют тенденцию быть слабыми, в то время как более сильные диагностические признаки, такие как тонкие структуры, показанные на диаграмме справа, не упоминаются; и, наконец, рассматриваемый организм представляет собой глубоко ветвящийся анаэроб, приспособленный к жизни в кишечнике насекомых отряда Blattodea, и несколько удивительно, что он процветает в легком человека. Среда обитания организма — важный ключ к его идентичности; этот обычно находится в безвоздушном заднем кишечнике некоторых жуков и, возможно, в экскрементах некоторых птиц, питающихся жуками. Учитывая, что эти жгутиконосцы, если это так, были обнаружены в нетипичной среде, внимательный систематик мог бы дождаться более подробной морфологической информации или, что еще лучше, молекулярных данных, прежде чем остановиться на идентификации на уровне вида. Где чашечка и осевая нить, которые Ричарду Кудо удалось увидеть в 1926? Где трубочки парабазального тельца, которые должны опоясывать клетку до верха, как взъерошенный воротник елизаветинского джентльмена? Из того, что я видел, они почти не упоминаются в литературе по лофомониазу.
Итак, у нас есть сильный клинический профиль в сочетании с невпечатляющими таксономическими данными, что оставляет нас с вопросом, который не может быть решен, пока кому-нибудь не удастся секвенировать эти клетки. Как заключают Мартинес-Хирон и ван Вурден, «для решения проблемы требуется разработка метода культивирования организма или использование молекулярных методов». Если кто-то этим занимается, будет интересно посмотреть на результаты.
——————
ССЫЛКИ (НАЖМИТЕ ЗДЕСЬ)
13 февраля 2015 г.
Несколько лет назад я заглянул в пробу воды из болотного озера и увидел что-то вроде гиперактивного авокадо, шевелящегося внутри в крошечной керосиновой лампе:
Архитектором этого симпатичного жилища является инфузория Calyptotricha pleuronemoides . Вид и род были обнаружены в 1882 году в образцах из пруда недалеко от Хертфорда, Англия, натуралистом-любителем по имени Фредерик У. Филлипс. О нем мало что известно. В 1880-х годах он был активным членом Хартфордширского общества естествознания и полевого клуба, которым он время от времени читал эссе о «Простейших животных Хартфордшира», основанные в основном на схеме классификации из «9» Уильяма Сэвилла Кента. 0418 Руководство инфузории . Он был членом Лондонского Линнеевского общества, нашел и назвал несколько новых таксонов.
Впервые увидев это существо, Филлипсу посчастливилось поймать его в процессе постройки лорики. «С первого взгляда, — пишет он, — я подумал, что это зачаточная или инцистированная стадия какой-то монады; но, применив увеличение примерно в 900 раз, я заметил, что он обладает своеобразной вибрирующей мембраной, очень похожей на ту, которая характерна для членов семейства Pleuronemidae». Неделю спустя Филлипс снова посмотрел на него и обнаружил, что «лорика увеличилась в размерах, а один конец удлинился в форме соска». На этом этапе он случайно позволил образцу высохнуть, оставив пустую, наполовину готовую лорику организма, все еще прикрепленную к пряди водорослей. Он сделал хороший рисунок того, что он видел.
A. Первый этап B. То же, доработанный C. Вид лорики с торца D. Совершенное животное E. Вентральный вид (по Филипсу)
Современному читателю, привыкшему к безличному, пассивному стилю научного письма– «были собраны образцы», «живые клетки были выделены и исследованы» — есть что-то приятно откровенное в том, как викторианские натуралисты сообщают о своих открытиях. Филипс не просто описывает свой новый род, он рассказывает нам историю его открытия, включая несчастный случай, который уничтожил его первый образец, и его первоначальное неправильное понимание овальной раковины, после чего он переносит нас в тот самый момент открытия, когда он раскрыл истинную природу существа, «применив увеличение в 900 диаметров». Что-то в этом мне напоминает исследовательскую литературу того же периода. Вероятно, это не просто случайность стиля: викторианские микроскописты 90 418 были 90 419 исследователями. Превосходные линзы и красители открыли миниатюрный Темный Континент на их лабораторных столах, и джентльмен-авантюрист откуда-нибудь вроде Хартфордшира теперь мог проникнуть в эти скрытые царства, возвращаясь с затаившим дыхание отчетом о том, что он видел. Сеанс у микроскопа был экспедицией в неизвестность.
В наше время от исследователей ожидается накопление каких-то данных перед тем, как отправиться в печать, и никто не будет пытаться возводить новый род инфузорий на основе краткого наблюдения за несколькими экземплярами. Без сомнения, это хорошо: XIX век оставил большое наследие плохо определенных таксонов, многие из которых до сих пор отчаянно нуждаются в пересмотре. Но такая полевая работа, какой бы схематичной и дилетантской она ни казалась сейчас, в значительной степени отошла в сторону, не заменившись ничем лучшим. За пределами нескольких очагов активности полевые работы с инфузориями замедлились до минимума. Примите во внимание тот факт, что через 132 года после того, как Филлипс написал свою трехстраничную заметку о Calyptotricha pleuronemoides это все еще один из двух существенных трактовок этого вида и единственный источник, описывающий строение его любопытного лорика. Тому, кто хочет узнать об этой инфузории больше, чем ее название, придется перенестись в 19 век.
Соломенная шляпка, начало 19 века. (Нажмите, чтобы увидеть источник)
Излишне говорить, что старая информация не всегда достоверна.
Phillips сразу и правильно понял, что Calyptotricha тесно связана с более распространенной инфузорией Pleuronema . Как и его двоюродный брат, он оснащен большой вздымающейся мембраной, которая проходит вдоль правой стороны его ротового отверстия. Однако Филипс совершенно неправильно понял форму этой структуры, описав ее как «перепончатую ловушку, или velum , которая по форме напоминала старомодный чепчик».
Когда я впервые прочитал этот отрывок, сравнение с «чепчиком» смутило меня. Волнистая мембрана плевронематидных инфузорий имеет форму паруса или флага: лист сросшихся ресничек проходит вдоль одной стороны рта организма. Филлипс, однако, интерпретировал это строение (которое, правда, очень трудно ясно увидеть в световой микроскоп) как своего рода капюшон или навес, закрывающий ротовое отверстие инфузории. Если вы внимательно посмотрите на его иллюстрацию, то увидите, что он нарисовал ее в виде мешковатой трубки.
Истинная форма ундулирующей мембраны Calyptotricha (изображение из Colin R. Curds, British and Other Freshwater Ciliated Protozoa, со стрелками) любопытное название, Calyptotricha , составленное из греческого calyptos («завуалированный» или «покрытый») и trich ( «волосы»). Кажется, «волосатая» голотриховая инфузория напомнила ему женскую голову, на макушке которой как старомодная шляпа сидит перепонка!
Это пример того, как ожидание формирует наблюдение. Интерпретируя эту мембрану как закрытый капюшон, он полагался на более раннюю ошибку своего прославленного современника Уильяма Сэвилла Кента. Описывая Pleuronema , Кент говорит: «[Э]то его перепончатую ловушку можно уместно сравнить с раскладывающимся капотом кареты или наружной оконной шторой, образующей в раскрытом виде вместительный тент в форме капюшона, а когда он не используется, он упаковывается. Аккуратными складками вокруг рта анималистики.
«Растяжимый капюшон», о котором упоминает Кент, был обычным удобством в вагонах того времени и представлял убедительную механическую аналогию «аккуратных складок», которыми он представлял Pleuronema , оттягивающих свой велум.
Вот, для сравнения, иллюстрация Кента Pleuronema chrysalis , которую я перевернул, чтобы продемонстрировать ее «растяжимый капюшон».
С колесами из него получилась бы хорошая тележка для патронов.
Для современных специалистов, знакомых с морфологией перепончаторотых инфузорий, обнаруженной в образцах, окрашенных серебром, такой дизайн кажется неправдоподобным. Однако Кенту, проделавшему новаторскую работу по изучению хоанофлагеллят, казалось разумным предположить, что Капюшон Pleuronema может иметь «отдалённое гомологическое родство» с «нежными воронкообразными мембранами», обнаруженными у жгутиконосцев с воротничком, которые действительно носят что-то вроде соломенной шляпки (но на заднем конце макушки). клетка).
Наконец, раз уж мы заговорили о Pleuronema и ее сестрах, я опубликую несколько видеозаписей, на которых одна из них тихо просматривает бактерии в воде, взятой из приливного бассейна на побережье штата Мэн:
ССЫЛКИ
Декабрь 182013
В начале прошлого года мэр Зальцбурга с гордостью объявил о создании новой природоохранной зоны вокруг этого «уникального в мировом масштабе природного памятника»: , или крепость 11-го века, которая находится на его вершине. Это длинная узкая лужа на переднем плане. Это пруд Краутхюгель, эфемерный водоем глубиной всего 30 см, где исследователи обнаружили 121 вид инфузорий, десять из которых ранее не были описаны. Благодаря этим организмам, пять из которых больше нигде не были обнаружены, Зальцбург теперь обладает 9-ю мировыми0852 первый второй «Памятник природы для одноклеточных организмов». Заповедник протистов!
Пруд появляется и исчезает в течение года, появляясь после проливных дождей на приподнятом сельскохозяйственном участке, известном как Краутхюгель, или «капустная гора». Русло, в котором он лежит, считается остатками старого ручья, естественное русло которого могло быть изменено сельским хозяйством в средние века. С тех пор дорожные работы и городское развитие изолировали тело от других окружающих каналов.
С 1789 по 1960 год поле использовалось для выращивания овощей. После этого оно стало пастбищем. Около тридцати лет коровы вытаптывали мягкий дерн и кормили местных микробов своим навозом, создавая то, что экологи называют «эвтрофированным прудом». Другими словами, крупный рогатый скот валяется или топится.
Это не то, что можно назвать нетронутой природой. Здесь нет приюта для крупных харизматичных животных. Он не особенно живописен, если его вообще можно увидеть (большую часть года «пруд» пересыхает). Короче говоря, трудно представить себе участок земли, который с меньшей вероятностью будет выделен для сохранения.
Однако у пруда Краутхюгель есть кое-что, чего не хватает вашим местным канавам и грязевым ямам: близость к Вильгельму Фойснеру, удивительно продуктивному цилиатологу, который живет и работает в Зальцбурге.
Вильгельм Фойсснер (Щелкните изображение, чтобы открыть источник)
Возможно, «чудом природы» здесь является не столько пруд Краутхюгель, сколько профессор Фойсснер, чьи обширные работы возвышаются над современной систематикой инфузорий, такой как сам Фестунгсберг. С пятью или шестью сотнями публикаций на его имя — не менее трехсот в рецензируемых журналах — Фойснер, работая в одиночку или в сотрудничестве, открыл и описал более 500 новых видов простейших. Если бы на вашем коровьем поле появились новые инфузории, он был бы человеком, который нашел бы их.
На самом деле, увидеть новый вид не так уж и необычно. Вероятно, все мы время от времени сталкиваемся с неописанными организмами, сами того не зная. Маленький красный жучок, садящийся вам на руку, может быть чем-то, о чем никогда раньше не упоминалось в литературе, если бы вы только знали. Поместите образцы местной грязи под микроскоп, и вполне вероятно, что вы найдете организмы, у которых еще нет названий. Конечно, одно дело видеть что-то новое, когда оно проплывает мимо, и совсем другое — знать, что ты уже видел. Чтобы должным образом задокументировать свое открытие и опубликовать новости о нем, требуются навыки и технологии, которых крайне не хватает.
Итак, этим инфузориям очень повезло родиться в Зальцбурге, недалеко от одного из немногих людей в мире, имеющих возможность (и склонность) видеть их такими, какие они есть, и лоббировать их защиту. Это поднимает некоторые интересные вопросы.
Во-первых, насколько исключительным является разнообразие микробов, сохранившееся в Краутхюгеле? В отчете о пруде, опубликованном ранее в этом году, Fenton P.D. Коттерилл и его соавторы сравнивают количество видов инфузорий в их местонахождении с различными «хорошо изученными эфемерными водами» в других частях мира: два пруда с талой водой на юго-западе Онтарио, придорожная лужа в Намибии, каменный бассейн в Венесуэле, луг в Венгрии и несколько других популярных мест. Они обнаружили, что Зальцбургский пруд находится в «верхнем диапазоне» по общему количеству видов, но только в «среднем диапазоне» по количеству новых видов.
Очевидно, что поле старой капусты поддерживает высокое, но далеко не уникальное, разнообразие, и при тщательном исследовании лучшими протистологами в этом бизнесе оно дает примерно ожидаемое количество новых организмов. Богатый, но довольно обычный водоем, кажется. Зачем выделять его для защиты?
Три вида, встречающиеся только в Краутхюгеле e) Semispathidium pulchrum f) Papillorhabdos multinucleatus g) Fuscheria nodosa salisburgensis
На это есть несколько причин. Во-первых, как отмечают авторы, призывы к сохранению обычно основаны «на узком распространении одного или нескольких видов и мест их обитания, либо видов и мест обитания, находящихся под угрозой исчезновения в результате деятельности человека». Если лес поддерживает единственную известную популяцию карликового лемура Сибри, у нас есть причина сохранить его, потому что, если мы этого не сделаем, мы можем ожидать, что этот вид исчезнет навсегда. По этому стандарту — при условии, что мы подавляем предубеждение о размере, которое может сделать нас равнодушными к судьбе микробного вида — аргументы в пользу защиты Краутхюгеля довольно веские. По состоянию на апрель 2013 года пять из десяти новых видов, обнаруженных там, «не были зарегистрированы ни в каком другом месте». Пока они не появятся в другом месте, эти пять видов считаются «эндемичными» для Зальцбурга (то есть ограниченными этой территорией). Учитывая нехватку компетентных цилиатологов в других частях мира, они могут оставаться таковыми еще некоторое время.
Независимо от того, окажутся ли они действительно эндемичными или нет, бесспорно, что организмы в пруду были «подвержены опасности в результате деятельности человека». В 2010 году в рамках арт-проекта кто-то засыпал его землей. Представьте тревогу исследователей, изучавших это место в течение десятилетий, когда они узнали, что их протисты были похоронены! Именно это событие побудило следователей вызвать охрану, в результате чего пруд был восстановлен до прежнего состояния и вокруг него была создана буферная зона:
Охраняемая зона вокруг пруда Краутхюгель
И это подводит нас ко второй причине сохранения этой лужи: благодаря уже проделанной работе она стала «типовой местностью» для примерно восемнадцати видов (восемь новых видов, и десять переописанных таксонов). Значение этого может потребовать небольшого пояснения.
Amblyodus taurus (щелкните изображение, чтобы перейти к источнику)
Когда биолог дает название новому таксону, обычная практика состоит в том, чтобы выбрать определенный фиксированный образец или группу образцов в качестве «типа» и (в идеале) депонировать этот образец где-то в постоянной коллекции, доступной для других исследователей. Это обеспечивает постоянную конкретную ссылку, поэтому не может быть двусмысленности в отношении того, что мы на самом деле имеем в виду, когда говорим 9.0418 Utricularia floridana (вид хищного растения) или Amblyodus (род жуков). Если нужно, мы можем указать на конкретную ошибку, нанизанную на конкретную булавку, и сказать: « Вот ! Amblyodus означает , что ».
Участок, на котором был собран типовой экземпляр, становится «типовым местом», где можно было бы ожидать найти другие особи той же породы. Это место особенно важно для таксономистов протистов. Протисты маленькие и хрупкие, а экземпляры фиксированного типа организмов с более старыми названиями редко доступны. Даже если существуют постоянные слайды, они могут быть потеряны или просто испортятся со временем. Если мы знаем тип места, где изначально были найдены наши ребята, мы можем пойти поискать их там. Теоретически. Но если место, где велась работа, осушено или заасфальтировано, а типографский материал отсутствует, идентичность найденных там видов может быть потеряна в таксономическом шуме.
То, что сохраняется в Краутхюгеле, является, по крайней мере частично, уже проделанной там научной работой. Это совокупность приобретенных биологических знаний, а не только сами организмы, которые подлежат охране. С этой точки зрения охрана окружающей среды может быть аналогична задаче, которую выполняют реставраторы музеев и произведений искусства, сохраняя лучшие продукты человеческого труда для будущих поколений.
Куда делись все пруды, которые не были и, вероятно, никогда не будут изучены? Весной, когда я еду по сельской местности, где живу, я вижу сотни и тысячи эфемерных луж. Они мелькают в окне машины, миля за милей: бобровые пруды, канавы, мельничные заводи, поймы и широкие неглубокие лужи в полях, куда коровы опускают морды и сбрасывают свои питательные какашки. У некоторых протистов будет меньше разнообразия, чем у Краутхюгеля, у некоторых может быть больше, но ни один из них никогда не будет пользоваться благосклонным надзором Вильгельма Фойснера.
Но что, если бы в этих водоемах проводилось больше исследований — по одному протистологу на каждую лужу! — и больше водоемов было бы признано достойным сохранения? Непонятно, куда ведет эта дорога. Если инициатива Krauthügel вызвала какие-либо разногласия в Зальцбурге, об этом не говорится ни в статье, ни в пресс-релизе, но нетрудно предвидеть, какой отпор мы бы увидели, если бы аналогичные инициативы были опробованы здесь. Попытки контролировать использование частной земли вызывают глубокое и невероятно долгоживущее негодование. Спустя двадцать пять лет после усилий по сохранению среды обитания северной пятнистой совы в PNW, антиэкологи все еще кипят и насмехаются. В некоторых кругах слова «пятнистая сова» стали своего рода сокращением для «назойливых цепляющихся за деревья идиотов, которые ценят глупую птицу выше, чем жизнь и средства к существованию трудолюбивых людей». Представьте себе вулкан возмущения, который может извергнуться из-за обязательной защиты одноклеточного организма! Мы никогда не услышим его конца.
Тем не менее, идея защиты среды обитания простейших меня очень привлекает. Вниз по дороге от моего дома на холмах Гатино есть группа эфемерных прудов, где я люблю собирать образцы. В течение нескольких лет они почти наверняка будут засыпаны, так как земля разбита под новое жилье. Я наблюдал за этими прудами в течение нескольких сезонов и ненавижу мысль потерять их удивительное микроскопическое разнообразие. Если, что статистически вероятно, они содержат несколько новых видов, может быть, даже есть основания для их сохранения. Однако совершенно очевидно, что бульдозеры доберутся до любых новых организмов до того, как я получу компетентность и ресурсы для их поиска и описания.
Тем не менее, мне немного утешительно напомнить себе об очень разных масштабах и скорости жизни на микроскопическом уровне. Когда вы длиной от кончика до хвоста в сто микрон, лужа — это озеро, а пруд — это океан. Час — это год!
Через несколько дней залитая дождем колея шин может превратиться в поразительное разнообразие, подобно миниатюрному коралловому рифу. Виды расцветают в быстрой последовательности, сменяя и вытесняя один другой. Каждое из них изменяет химический состав, светопроницаемость и питательную нагрузку воды, настраивая окружающую среду в соответствии с определенными организмами, которые, в свою очередь, изменяют воду вокруг себя. Случайности географии (плавающий лист, шарик навоза) открывают возможности для одних организмов и уничтожают всякую надежду для других. Дела развиваются быстро. Если каждый день возвращаться к колее и следить за ее ходом с помощью микроскопа, может показаться, что смотришь цейтраферную пленку. В «большом» мире окружающая среда меняется почти так же, но в течение более длительных периодов времени: леса вторгаются в прерии, а затем отступают; водно-болотные угодья формируются, заиляются и исчезают; животные приходят и уходят. На микроскопическом уровне изменения в популяциях могут происходить за часы, а не за годы, и изменения не прекращаются. «Экологический баланс» никогда не достигается: то происходит, то происходит больше. Одни виды процветают, другие исчезают из поля зрения. Затем однажды жаркое солнце высушивает все это, и маленькие существа забираются обратно в свои покоящиеся цисты, а их среда обитания возвращается к траве.
———————————
Ссылки:
Cotterill, Fenton PD, et al. «Сохранение протистов: пруд Краутхюгель в Австрии». Разнообразие 5.2 (2013): 374-392.
Австрийский научный фонд (FWF) Пресс-релиз
ноябрь 292013
Как я уже говорил, мы, позвоночные, обычно высокого мнения о большом скоплении нейронов, расположенном на одном из концов нашего спинного мозга. Так и должно быть: в таких существах, как мы, происходит волшебство. Чем больше скопление, тем более когнитивно разносторонний организм; уберите его, и весь организм остановится (хотя сначала может побегать какое-то время).
Но, как мы знаем, даже одноклеточные существа способны ощущать и координировать движения и могут демонстрировать сложное, даже целенаправленное поведение. Так что, вероятно, было неизбежно, что какой-нибудь благонамеренный ученый попытается оснастить их собственной центральной нервной системой.
Stylonychia mytilus ползает по нитям водорослей. Изображение Actinophrys на Flickr. Нажмите для ссылки.
В 1880 году немецкий исследователь по имени Th. В. Энгельманн провел тщательный осмотр тканей инфузории Stylonychia mytilus. Как и у других представителей группы инфузорий, широко известных как «гипотрихи», у Stylonychia на «брюшке» имеется пучок толстых, подвижных cirri : пучки сросшихся ресничек, которые он использует как маленькие ножки, когда карабкается по твердым объектам в своем затонувшем мире. Эти псевдоноги работают очень хорошо, и ползающий гипотрих может быть очень похож на жука или таракана.
В отличие от конечностей насекомых, усики гипотрихов не имеют заметных прикреплений, таких как мышцы, сухожилия и нервы. Однако, внимательно изучив окрашенные образцы, Энгельманн смог обнаружить в основании усиков Stylonychia группы волокон, которые он назвал «Wimperwurzeln», или «ресничными корешками». Он предположил, что эти волокна могут функционировать точно так же, как нервы у «высших» организмов, служа проводящими дорожками, по которым могут двигаться стимулы, приводящие в движение реснички!
Это была новаторская идея; впрочем, особого восторга это не вызвало. Несмотря на его впечатляюще четкое описание этих небольших структур, предположение, что он обнаружил животноподобные органы у одноклеточных существ, могло прийти в неподходящее время. К 1880 г. представление о простейших как о миниатюрных животных, несущих полный набор репродуктивных и пищевых органов, было давно дискредитировано. В конце 19 века простейшие обычно рассматривались как примитивные организмы, не более чем мешочки с универсальной протоплазмой, составлявшей, по памятному выражению Томаса Хаксли, «физическую основу жизни».
Но времена меняются, и идея о том, что свободноживущие клетки могут состоять из дифференцированных тканей, вернется в моду (действительно, в начале 20 века некоторые протозоологи полностью отвергли клеточную природу своих организмов).
Чарльз Этвуд Кофоид
В 1914 году Роберт Дж. Шарп, работавший в Калифорнии под руководством выдающегося протозоолога Чарльза Этвуда Кофойда, опубликовал статью, в которой он смело заявил, что нашел инфузорию, у которой были не только нервные волокна, но полноценный центрально расположенный «нейромоторный аппарат». Он обнаружил эту структуру в организме под названием Diplodinium ecudatum (теперь известный как Epidinium ecudatum ), симбионт в желудках крупного рогатого скота. То, что обнаружил Шарп, нигде не фиксировалось: небольшая масса волокон, расположенная рядом с подвижными органеллами клетки и служащая «общим центром двигательных влияний». Он открыл орган, который контролировал и координировал подвижные части инфузории, по сути, цитоплазматический «мозг» клетки. Он назвал это «моториум».
Работа Шарпа вызвала небольшой вихрь научной активности, особенно среди его коллег из лаборатории Кофойда. В следующем году Кофоид сам описал «нейромоторный аппарат» у кишечного паразита, Лямблии . Вскоре после этого Гарри Б. Йоком, еще один сотрудник группы Кофоида, опубликовал информацию об открытии полноценного нейромотория у гипотриха Euplotes patella («ходячей инфузории», подобной Stylonychia ).
Но зачем приписывать нейронную функцию структурам, которые он видел в Euplotes ? Yocom подробно рассказывает об этом. Во-первых, эти волокна окрашиваются в ярко-красный цвет при окраске фуксином по Мэллори, точно так же, как нервные клетки у многоклеточных животных; во-вторых, поясняет он, эти волокна находятся в связи с подвижными частями инфузории: ротовыми перепонками и локомоторными усиками.
Иллюстрация нейромоторного аппарата Euplotes patella, сделанная Yocom, с «моториумом», обведенным красным.
Наконец, поведение Euplotes patella подтверждает эту интерпретацию. Это, пожалуй, классический пример гипотезы, ведущей наблюдение по носу. Йокому казалось очевидным, что E. patella использует свою «переднюю губу» (выпуклость наверху клетки, на которой виднеется окрашенная в красный цвет решетка волокон) в качестве органа чувств. Более того, поперечные усики, которые были связаны с «моториумом», двигались скоординировано, в то время как неприкрепленные усики двигались беспорядочно. Как выразился Йоком: «Вихревые нерегулярные движения передних вентральных и маргинальных усиков никоим образом не скоординированы с регулярными ритмическими движениями мембранелл или с отбрасыванием усиков назад».
Или так ему показалось.
До этого момента функция «нейромоториума» как «координационного центра» движений клетки была просто гипотезой, не подтвержденной экспериментом. Но подтверждение не заставило себя ждать. В 1919 году К. В. Тейлор — еще один работник кружка Беркли — провел эксперимент на Euplotes patella , инфузории, которую Йоком так подробно изучал годом ранее. Используя кварцевые микроскальпели, он перерезал волокна, идущие между моториумом и анальными усиками (заметная группа из пяти тяжелых усиков в задней половине клетки, теперь обычно известная как «поперечные усики»). Эксперимент убедительно подтвердил гипотезу; отделенные от моториума, усики, по-видимому, вообще не работали должным образом: «Отсечение волокон анальных усиков влияет как на ползание, так и на плавание… Разрушение моториума или перерезание прикрепленных к нему волокон нарушает координацию движений адоральных мембран и анальных усиков. ». (Тейлор, 19 лет19)
Paramecium от Rees (1920), показывающий узор из тонких «нейрофибрилл» (предположительно, скелетных микротрубочек), сходящихся к «нервному центру» клетки, обведенных здесь красным.
С тех пор нейромоторный аппарат появлялся у одной инфузории за другой. Он был обнаружен у Paramecium (1920), Balantidium (1922), Tintinnopsis (1926), Dileptus (1927), Chlamydodon (1928), 4 Uroleptus.0418 Окситриха (1935). По ходу дела сомнения неуклонно угасали, и к 1927 г. стало возможным прямо утверждать, что «убедительно продемонстрировано существование сложной нейромоторной системы у некоторых инфузорий с ресничками». (Visscher, 1927) Это стало, по крайней мере, в Северной Америке, твердым фактом.
Но даже твердые факты могут снова превратиться в жидкость, а затем испариться, как каналы Марса. Это может происходить медленно, а может и сразу. Иногда гипотеза умирает быстрой смертью, когда ее опровергает эксперимент; иногда его сметает поток лучшей теории; а иногда его просто откусывают, в то время как мир вокруг него медленно меняется, пока в один прекрасный день никто не может вспомнить, почему это когда-то казалось правдой.
Нейромоторная концепция имела долгую жизнь, опередив самого Кофоида на полтора десятилетия. Даже появление электронной микроскопии не убило его сразу. Еще в 1957 г. ЭМ, по-видимому, выявила у Euplotes «массу переплетающихся корешковых филаментов», соответствующих нейромоториуму, а также целую систему внутриклеточных волокон, которые, как утверждали авторы, главным образом предназначались для «координации биение ресничек». (Roth, 1957) Но когда Р. Глиддон снова поставил Euplote s под электронным микроскопом в 1966 году он не смог найти ничего, что можно было бы назвать моториумом. Оно исчезло.
В том же году два исследователя в Японии наконец смогли воспроизвести классический эксперимент Тейлора на E. patella — эксперимент, который, возможно, положил начало золотой лихорадке нейромоториума. (Okajima and Kinosita, 1966) Но теперь результаты были совсем другими. Исследователи разрезали «нейрофибриллы» точно так же, как это сделал Тейлор, и запечатлели движения ресничек на пленку; однако на этот раз цирри просто продолжали двигаться в своей обычной скоординированной манере, как будто ничего не произошло. Неизвестно, почему 9 Тейлора0418 Euplotes вел себя иначе. Возможно, он искалечил несчастных, рассекая их «нейрофибриллы», и сделал свои знаменитые наблюдения за организмами в их предсмертной агонии.
В 1970 году Дороти Пителька была готова объявить игру, объявив об общем «отказе от старой концепции фибриллярной нейромоторной системы у инфузорий». Беглый поиск в Google Книгах находит последнюю краткую ссылку на «нервные фибриллы» Paramecium в школьном учебнике биологии, опубликованном в 1919 году.83. После этого ничего.
Так что же это были за «нейрофибриллы» в конце концов? Они не были результатом плохого снаряжения или причудливой наблюдательности, а скорее наоборот. Примечательно, что микроскописты того времени, по-видимому, уже обнаруживали части цитоскелета — каркас из микротрубочек и филаментов, придающих клетке ее форму, — а также пелликулярную «решетку», так называемую ресничную «систему серебристых линий», которая позднее полностью обнажаются в препаратах азотнокислого серебра и протаргола. Интерпретация отставала от наблюдений на поколение или два. Видение превзошло понимание; затем пришла новая информация, и старые уверенности просто канули в воду.
Щелкните здесь, чтобы просмотреть список ссылок
ноябрь 232013
Когда Рене Декарт искал в человеческом мозгу Местонахождение Души, он остановился на шишковидной железе, маленьком эндокринном органе, обнаруженном почти у всех позвоночных. Он казался хорошим кандидатом, потому что это единственный орган, не продублированный в каждой половине мозга, а расположенный прямо на средней линии, где он мог бы, как предполагал Декарт, получать сенсорные впечатления через маленькие трубки и напрягать мышцы мозга. тела, выдыхая «животные духи» по системе шлангов, ведущих к рукам и ногам.
Иллюстрация из Traité de l’Homme (1644 г.), показывающая каплевидную шишковидную железу, расположенную в черепе, как маленький меха-пилот.
Душа Декарта сидит в центре управления мясной марионеткой, собирая новости мира через глаза, уши и нос и воздействуя на мир своей волей через пневматические механизмы тела.
В его концепции легко распознать структуру, аналогичную тому, что мы знаем как нервную систему. Мы больше не думаем о «животных духах», а скорее о «нейронных сигналах», и мы больше не помещаем разум в какую-то конкретную железу в мозгу. Но для большинства из нас, кто имеет лишь приблизительное представление о том, как все это работает, эта схема «сигнал/передатчик/приемник» — всего лишь еще одна метафора, чуть более изощренная, чем та, которую использовал Декарт. По большому счету, мы все еще оперируем представлением о нематериальном разуме, каким-то образом локализованном в «центральной» нервной системе и осуществляющем свои дела с миром через разветвленную сеть телесных нитей.
В самом деле, многим из нас трудно даже представить разум, который не сконцентрирован на вершине иерархии, идущей от мозга (где находится мыслящее я) к кончикам периферических нервов, где лежит мир ждет, когда его почувствуют, услышат и увидят.
Итак, распределенный интеллект нервной системы осьминога — половина его нейронов находится в его лапах! — кажется нам чем-то причудливым и удивительным, требующим дальнейшего изучения и объяснения. Бросая вызов наивному ожиданию, что когнитивные процессы должны быть «центральными», а органы чувств — «периферийными», исследователи осьминогов разработали остроумные эксперименты, чтобы показать, что руки моллюска продолжают выполнять целенаправленные действия, такие как поиск добычи, исследование и избегание опасности, даже когда они отделены от так называемого мозга.
И если кажется удивительным, что оторванная рука осьминога все еще может ощущать воспринимаемую опасность и реагировать на нее, насколько же более странным может быть то, что одна клетка может делать все эти вещи, вообще без помощи какой-либо нервной системы ? Но это совершенно обычное дело в мире протистов, где одноклеточные охотники плавают вокруг, выслеживая одноклеточных жертв, которые отшатываются, когда чувствуют беду, укрываются в своих панцирях или прячутся от врагов в грудах обломков.
Только представьте, если бы хоть одна клетка вашего тела могла вырваться на свободу и отправиться вот так бродить, охотясь и пожирая добычу! О, точно… они делают. На самом деле, клетки эукариот, независимо от того, находятся ли они внутри более крупных организмов или блуждают по «миру», совершенно обычно демонстрируют сложное поведение, которое бывает очень трудно отличить от поведения, которое мы наблюдаем у крупных многоклеточных животных. .
Возьмите инфузорию типа Stichotricha aculeata . Обычно он обитает в ветхом самодельном трубчатом жилище (лорике), состоящем из слизи, смешанной с мусором. Он живет, заталкивая более мелкие организмы — бактерии и некоторых более изящных простейших — в отверстие, похожее на рот, где они упаковываются в маленькие пищевые пузырьки (вакуоли) и перевариваются. Подобно кошке или раку, он достаточно велик, чтобы быть хищником, и достаточно мал, чтобы быть добычей. Следовательно, это осторожное существо, полагающееся на свою лорику для защиты. Когда он чувствует беду, он прячется в свою трубу, а когда берег свободен, он высовывает свой длинный кормовой аппарат. Как и другие инфузории, он может чувствовать движение (вероятно, с помощью сенсорных ресничек, которые распространены в эукариотических клетках), и его легко пугать. Если вы постучите пальцем по предметному столику микроскопа, пока вы смотрите на него, он обнаружит вибрации и отодвинется. Спрятавшись некоторое время в своей желейной трубке, он предварительно вытягивает хоботок. Если он больше не столкнется с проблемами, он снова начнет питаться.
Однако, если разрушение достаточно сильное или продолжительное, он может полностью отказаться от своей лорики, как это делает близкородственная Chaetospira в видео, которое я записал в прошлом году. Stichotricha во встроенных кадрах, которые приведены ниже, уже покинули свой первоначальный «дом» и укрылись в небольшом замкнутом пространстве, между коралловидным трубчатым жилищем колониального жгутиконосца под названием Rhipidodendron huxleyi и мукоидной колонией какой-то другой организм (Spongomonas?). В этом импровизированном замене лорики она ведет себя точно так же, как обычно дома: скользит вперед и назад, кормится, отступает, продвигается, всегда голодна и всегда бдительна:
Легко понять, почему ранние микроскописты называли этих существ «анималькулами»: «очень маленькими животными». Без нервной системы инфузории, такие как Stichotricha и Chaetospira , демонстрируют поведение, поразительно напоминающее рыбу, затаившуюся в расщелине, или насекомое, обитающее в футляре, например, личинку ручейника. Если они часто напоминают нам «настоящих» животных, полезно напомнить себе, что, в конце концов, их образ жизни очень мало отличается от образа жизни других обитающих в полостях водных существ, таких как те, которые мы могли бы найти на кораллах. рифе или, как эта личиночная мошка, в гравии пресноводного ручья:
определение инфузории и синонимы инфузории (английский)
Из Википедии, свободной энциклопедии
Перейти к: навигация, поиск
Чтобы узнать о типе края листа, см. Leaf#Margins (край).
Инфузории представляют собой группу простейших, характеризующихся наличием волосовидных органелл, называемых ресничками, которые идентичны по структуре жгутикам, но обычно короче и присутствуют в гораздо большем количестве с волнообразным рисунком, отличным от жгутиков. Реснички встречаются у всех членов группы (хотя у своеобразных сукторий они есть только в течение части жизненного цикла) и по-разному используются при плавании, ползании, прикреплении, питании и ощущении.
Термин «Ciliophora» используется в классификации как тип. [1] Ciliophora можно отнести к Protista [2] или Protozoa. [3] Также используется термин «Цилиата», [4] как класс. [5] (Однако этот последний термин может также относиться к типу рыб.) Классификация протистов быстро развивается, и нередко эти термины используются для описания других иерархических уровней.
Инфузории — одна из важнейших групп протистов, распространенная практически везде, где есть вода — в озерах, прудах, океанах, реках и почвах. У инфузорий есть много эктосимбиотических и эндосимбиотических членов, а также некоторые облигатные и условно-патогенные паразиты. Инфузории, как правило, являются крупными простейшими, некоторые из них достигают 2 мм в длину и являются одними из самых сложных простейших по строению.
Contents
|
Строение клетки
Строение клетки цилиофоры: 1-сократительная вакуоль, 2-пищеварительная вакуоль, 3-макроядерная, цитостом, 8-ресничный.
В отличие от других эукариот, инфузории имеют два разных типа ядер: маленькое диплоидное микроядро (репродукция) и большое полиплоидное макронуклеус (общее регулирование клетки). Последний создается из микроядра путем амплификации генома и жесткого редактирования. Деление макронуклеуса происходит путем амитоза, расхождение хромосом происходит в результате процесса, механизм которого неизвестен. Этот процесс отнюдь не совершенен, и примерно через 200 поколений клетка проявляет признаки старения. Периодически макронуклеусы необходимо регенерировать из микроядер. В большинстве случаев это происходит в течение спряжение . Здесь две клетки выстраиваются в линию, микроядра подвергаются мейозу, некоторые гаплоидные дочерние клетки обмениваются и затем сливаются, образуя новые микроядра и макронуклеусы.
Пищевые вакуоли образуются в результате фагоцитоза и обычно следуют по определенному пути через клетку, поскольку их содержимое переваривается и расщепляется лизосомами, поэтому вещества, содержащиеся в вакуоли, становятся достаточно маленькими, чтобы диффундировать через мембрану пищевой вакуоли в клетку. Все, что остается в пищевой вакуоли, к тому времени, когда оно достигает цитопрокта (ануса), выводится путем экзоцитоза. У большинства инфузорий также есть одна или несколько заметных сократительных вакуолей, которые собирают воду и выталкивают ее из клетки для поддержания осмотического давления или в какой-либо функции для поддержания ионного баланса. Они часто имеют характерную звездообразную форму, каждая точка которой представляет собой собирательную трубку.
Питание
Большинство инфузорий питаются более мелкими организмами (гетеротрофными), такими как бактерии и водоросли, и детритом, заносимым в ротовую канавку (рот) модифицированными ротовыми ресничками. Это обычно включает ряд мембранелл слева от рта и пароральную мембрану справа от него, обе из которых возникают из поликинетид , групп многих ресничек вместе с ассоциированными структурами. Пища перемещается ресничками через ротовое отверстие в пищевод, где образуются пищевые вакуоли.
Однако это значительно варьируется. Некоторые инфузории не имеют рта и питаются путем всасывания, в то время как другие являются хищниками и питаются другими простейшими и, в частности, другими инфузориями. Сюда входят суктории, питающиеся несколькими специализированными щупальцами.
Размножение
Инфузории могут размножаться как бесполым, так и половым путем. Бесполое размножение происходит путем бинарного деления. Микронуклеус подвергается митозу, а макронуклеус удлиняется и делится пополам. Каждая из новых клеток получает копию микронуклеуса и макронуклеуса. Половое размножение включает конъюгацию, в которой участвуют две клетки. После конъюгации две клетки делятся, образуя четыре новые клетки.
Специализированные структуры
У некоторых форм есть также поликинетиды тела, например, у спиротрихов, где они обычно образуют щетинки, называемые cirri . Чаще реснички тела располагаются в моно- и дикинетид , которые включают соответственно одну и две кинетосомы (базальные тельца), каждая из которых может поддерживать ресничку. Они организованы в ряды, называемые кинетиками , которые проходят от передней части клетки к задней. Тело и ротовые кинетиды составляют infraciliature , организация, уникальная для инфузорий и важная для их классификации, и включает различные фибриллы и микротрубочки, участвующие в координации ресничек.
Инфракилиатура является одним из основных компонентов клеточной коры. Другие альвеолы , маленькие везикулы под клеточной мембраной, которые упакованы против нее, чтобы сформировать пелликулу, поддерживающую форму клетки, которая варьируется от гибкой и сократительной до жесткой. Обычно также присутствуют многочисленные митохондрии и экструсомы. Наличие альвеол, строение ресничек, форма митоза и другие различные детали указывают на близкое родство инфузорий, апикомплексов и динофлагеллят. Эти внешне непохожие группы составляют альвеоляты.
Ископаемая летопись
До недавнего времени самыми древними известными окаменелостями инфузорий были тинтинниды ордовикского периода. В 2007 г. Li et al. опубликовал описание ископаемых инфузорий из формации Доушантуо около 580 миллионов лет назад, в эдиакарский период. К ним относятся два типа тинтиннидов и возможный предковый сукториан. [6]
Классификация
Stentor roeseli
Подтип Postciliodesmatophora
- Class Karyorelictea
- Class Heterotrichea (e. g. Stentor )
Subphylum Intramacronucleata
Oxytricha trifallax
- Class Spirotrichea
- Subclass Choreotrichia (e.g. Tintinnidium )
- Subclass Oligotrichia (e.g. Halteria )
- Подкласс Stichotrichia (например, Stylonychia )
- Подкласс Hypotrichia (например, Euplotes )
File:Balantidium trophB.JPG
A trophozoite of Balantidium coli
- Class Litostomatea
- Subclass Haptoria (e.g. Didinium )
- Subclass Trichostomatia (e.g. Balantidium )
- Class Phyllopharyngea
- Подкласс Phyllopharyngia
- Подкласс Rhynchodia
- Подкласс Chonotrichia
- Подкласс Suctoria (например, Podophrya )
- Class Nassophorea
- Class Colpodea (e.g. Colpoda )
- Class Prostomatea (e. g. Coleps )
- Class Oligohymenophorea
- Subclass Peniculia (e.g. Paramecium )
- Subclass Hymenostomatia (e.g. Tetrahymena )
- Подкласс Scuticociliatia
- Подкласс Peritrichia (например, Vorticella )
- Подкласс Astromatia
- Подкласс Apostomatia 9 Yi Z, Song W, Clamp JC, Chen Z, Gao S, Zhang Q (декабрь 2008 г.). являются protistainkinghub.elsevier.com/retrieve/pii/S1055-7903(08)00581-2 «Пересмотр систематических взаимоотношений внутри отряда Euplotida (Protista, Ciliophora) с использованием новых последовательностей гена, кодирующего малую субъединицу рРНК, и тестирование использования комбинированных наборов данных для построения филогений комплекса Диофрис». Мол. Филогенет. Эвол. . doi:10.1016/j.ympev.2008.12.006. PMID 1
- 02. http://ltheprotistainkinghub.elsevier.com/retrieve/pii/S1055-79 Ли, К.-В.; и др. (2007 г.). [ Ошибка выражения: Отсутствует операнд для > «Реснитчатые простейшие из докембрийской формации Доушантуо, Венган, Южный Китай»].
Геологическое общество, Лондон, специальные публикации 286 : 151–156. дои: 10.1144/SP286.11. - Подкласс Coccidiomo
- Подкласс Gregarinomorpha
- Губки
- мезозои
- Подкласс Spirotricha
- Класс Реснички
- Класс корненожки
- Класс Actinopoda
- Класс споровиков
Класс Мастигофора
Подкласс Зоомастигина
Инфузории (класс Реснички) Примеры, отряды и характеристики
Инфузории (класс Реснички)
Эти простейшие имеют множество ресничек и ядер двух размеров. Поскольку у мастигофоры Opalina также много жгутиков (или ресничек), именно ядра действуют как критерий и определяют настоящие инфузории как отдельную группу. Другие особенности включают кинетическую систему фибрилл и гранул для координации ресничек, характерное поперечное деление и характерную форму полового размножения, называемую conjuga-don.
Диапазон форм этого класса настолько велик, что предполагает очень долгую независимую эволюционную историю. В то время как относительно легко отнести простейших к инфузориям, часто бывает довольно трудно определить их отряд, если род уже не известен, потому что в каждом отряде происходили разработки и слияние одних ресничек, часто сопровождаемые потерей других. Этот тип параллельной эволюции производит роды, которые внешне похожи.
Единственным надежным методом идентификации является применение одного из методов окрашивания серебряной линии, с помощью которого можно показать части кинетической системы и таким образом получить необходимую характеристику, на которой основана их таксономия. Но это метод, который нелегко или не легко осуществить.
Среда обитания разнообразна: во многих отрядах встречаются паразитические, комменсальные и свободноживущие виды. Примечательна степень морфологической специализации у некоторых спиротрих как пример развития в одиночной клетке: адаптация к необычным местам обитания, таким как интерстициальные пространства в песке, привела к большому удлинению до 5 мм. у некоторых видов, в то время как некоторые свободноживущие пресноводные виды имеют длину всего 15 микрон.
Голотрихи (Подкласс Голотриха)
Эта группа считается более примитивной, чем другие подклассы, из-за отсутствия развития перистомиальных мембран. Особенностью, которая исключает все другие усложнения и упрощения структуры, является способ слияния ресничек с образованием дополнительных органелл в воронкообразной цитофаринксе или клеточном устье. Для этого подкласса характерно отсутствие сросшихся структур и открытый рот. Обычно реснички тела (соматические) имеют примерно одинаковую длину и присутствуют по всей клетке.
Gymnostomes (Отряд Gymnostomatida) Как следует из названия, во рту нет ресничных органелл. Это самая разнообразная группа структурно простых инфузорий. Но разнообразие привело к значительным изменениям в классификации в последние годы. Сравнение существующих классификаций показывает, что общего согласия пока не достигнуто. Большинство видов отряда содержат одно мегануклеус и одно или несколько микроядер. Рот может быть передним и конечным или боковым. Если рот расположен сбоку, то тело превращается в тонкое расширение, образующее новый передний конец.
Это основа для классификации подотряда Rhabdo-phorina, у которого цитофаринкс (пространство непосредственно внутри клеточного отверстия, цитостом) является растяжимым. Другой подотряд — Cyrtophorina, у которого жесткая нерастяжимая цитофаринкс.
Подотряд Rhabdophorina В этой группе встречаются удивительно прожорливые кормушки, плотоядные во всех смыслах этого слова, несмотря на свои небольшие размеры. Простая форма, такая как голофрия, имеет овальную форму и длину около 160 микрон. Встречается на поверхности пресной воды, где питается диножгутиковыми.
В нескольких пищевых вакуолях в цитоплазме отчетливо видны остатки прошлой еды. Сократительная вакуоль частично поддерживает водный баланс в клетке. Didinium похож по форме, но имеет значительно уменьшенные ресничные поля. Остались только две полосы, одна вокруг устья и одна у экватора. Этот организм может проглотить целый Paramecium, хотя добыча в несколько раз крупнее его самого.
Энергия, с которой жертва атакует, парализована, а затем втягивается в рот, который кажется слишком маленьким, завораживает. Другие роды включают Dileptus и Lionotus, которые имеют расширенные передние отростки с трихоцистами и ресничками, придающие гребенчатый вид области перед ртом. Эта форма является приспособлением к жизни между песчинками и средами, где свободное плавание невозможно.
инфузории
У высших инфузорий есть кольцо усиков для сбора пищи, известное как адоральный венок, вокруг пищевода, и на этой фотографии его хорошо видно вместе с ядром и пищевыми вакуолями (x 1680).
Подотряд Cyrtophorina
Считается, что вентрально расположенный рот имеет эволюционное значение ввиду того факта, что большинство инфузорий похожи на него, но имеют дополнительные усовершенствования. Это небольшая группа. Типичным родом является Chilodonella. Вентральный рот виден с дорсальной стороны через клетку. Передний отросток значительно тоньше основной задней части. Линия ресничек, дорсальная щеточка, проходит от края клетки к области над ртом, и ее не следует путать с пищевыми органеллами других инфузорий, которые сметают пищу в рот.
Сукторианцы (Орден Сукторид)
Сукторианцы — это компактная группа, которая совсем недавно считалась голотрихами. Взрослая форма сидячая, без ресничек. Пища захватывается длинными щупальцами с выступами на концах, которые способны парализовать добычу.
После поимки добыча не притягивается к телу, ибо рта нет, а высасывается насухо щупальцами. Недавние исследования фильмов показывают постоянный поток цитоплазмы из организма, такого как Tetrahymena, в сукторию. Размножение несколькими способами.
Наиболее распространенным является внутреннее почкование. После деления ядра передняя пелликула по окружности погружается в цитоплазму, и вокруг нового ядра развивается молодая реснитчатая суктория. Созревая, дочерняя клетка покидает родительскую и свободно плавает. Позже он оседает на каком-нибудь предмете, выделяет несокращающийся стебель и теряет свои реснички, чтобы стать маленькой взрослой особью. Окрашенные препараты сукториан показывают, что на всех стадиях развития
кинетическая система присутствует даже при отсутствии ресничек. Типичными родами являются Tokophyra, Podophyra и Adult:.
Трихостомы (Отряд Trichostomatida) Эта группа имеет много общего с голосеменными, но отличается от них наличием преддверия. Этот орган представляет собой углубление пелликулы с ресничками, образующими трубку, ведущую ко рту. Реснички преддверия проталкивают пищу в рот.
Цитоплазма, примыкающая ко рту, кажется сильно вакуолизированной, и здесь перевариваются водоросли или бактерии, которыми питается свободноживущий род Coelosomides. По строению кишечный паразит амфибий и млекопитающих Balantidium colt внешне напоминает некоторых гименоротых или гетеротрихов.
Однако рот питается вестибулярными ресничками, которые происходят из нормальных соматических ресничек, покрывающих клетку, и развиваются таким же образом. Обычный пресноводный род — Colpoda, имеет характерную почковидную форму и длину около 75 микрон. Рот находится на зазубренной стороне, а вестибулярные реснички, хорошо заметные на окрашенных препаратах, немного короче соматических ресничек.
Гименостомы (Отряд Hymenostomatida)
Этот отряд является отправной точкой для усовершенствований, обнаруженных у остальных инфузорий. Питающее устройство подробно описано здесь, а названия структур применимы позже. Первой из этих структур является щечная полость между цитостомом и щечной увертюрой, которая является демаркационной линией между соматическими ресничками и щечными ресничками.
Ротовая полость имеет два типа цилиарных органелл: адоральная зона мембран (AZM) слева от цитостома и ундулирующая мембрана (UM) справа. (Левый и правый используются для организмов, чей рот считается вентральным, когда наблюдатель смотрит с дорсальной поверхности на вентральную, как левое и правое на наши собственные тела.
Для получения четких изображений и из-за при стандартном способе изготовления микроскопических препаратов организмы всегда рассматриваются с вентральной поверхности, поэтому на рисунках и фотографиях левое и правое положение переставлено, как если бы два человека смотрели друг на друга лицом к лицу. )
UM представляет собой ряд ресничек, сросшихся вместе и бьющихся в координации. AZM изменчив и может представлять собой ряд сросшихся ресничек, как у спиротрихов, или может быть разделен на несколько слоев сросшихся ресничек, как у Tetrahymena. Ротовая полость не имеет ресничек, кроме тех, которые содержатся в UM и AZM.
Ротовая ресничка развита в два поля ресничек в щечной полости, которая открывается в преддверие, лежащее в неглубокой ротовой бороздке. Paramecium питается в основном бактериями, которые собираются питательным током, создаваемым ресничками в ротовой бороздке. Частицы сметаются по этой канавке в неглубокое преддверие и, если они попадают в ротовую полость, в цитостом. Частицы могут быть отброшены, и в этом случае они проходят через ротовую полость под прямым углом к нормальному ритму ресничек. Передвижение у Paramecium, как и у большинства инфузорий, осуществляется с помощью соматических ресничек, покрывающих тело. Реснички бьются в скоординированном порядке, создавая волны движения, бегущие по поверхности.
Реснички располагаются рядами, называемыми меридианами, идущими в передне-заднем направлении. Каждый ряд или кинетий имеет реснички, соединенные фибриллами, проходящими под пелликулой. Кинетики перекрестно связаны поперечными фибриллами на более глубоком уровне, чем продольные фибриллы. Все эти взаимосвязи под пленкой известны как инфрацилиатура.
Paramecium хорошо известен своей реакцией избегания. Нормальный курс плавания вперед и вращение вокруг своей длинной оси. Но при достижении неприятного раздражителя организм останавливается, реверсирует под небольшим углом на два-четыре длины, а затем снова движется вперед под небольшим углом в обратном направлении. Конечным результатом является то, что организм сворачивает со своего первоначального пути и уходит от неприятного раздражителя. Кажется, что он не питается при быстром движении.
, а макронуклеус делится путем расщепления без характерного митотического аппарата. Способы полового размножения у инфузорий — конъюгация и автогамия. При конъюгации два человека сходятся и соединяются на своих оральных поверхностях. Оба макронуклеуса частично дегенерируют, увеличиваясь и становясь диффузными, а затем вытягиваясь в нити, которые позже распадаются, образуя многочисленные слабо окрашивающиеся остатки исходных ядер. Микроядра, которых у Paramecium aurelia два, делятся мейотически, давая восемь дочерних ядер. Семь из восьми дегенерируют, а оставшийся снова делится с образованием двух гомозиготных ядер гамет.
Одно гаметическое ядро от каждого партнера мигрирует к другому партнеру и сливается с неподвижным гаметическим ядром. Зиготическое ядро делится дважды, и пока эти деления происходят, конъюганты расходятся. Из четырех образовавшихся микроядер два становятся макронуклеусами, а два — микроядрами.
Последние снова делятся, а эксконъюгант делится, причем один макронуклеус идет к одной дочери, а другой — ко второй. Таким образом, каждая дочь получает два митотически полученных микронуклеуса и один цельный макронуклеус.
Второй формой полового размножения является автогамия. Здесь один организм совершает движения сопряжения без другого сопряжения. Образуются два гомозиготных гаметных ядра, которые сливаются, образуя зиготу. После этого образуются деления и новый макронуклеус, как при конъюгации. Генетическая автогамия приводит к расточительной рекомбинации генетического материала, поскольку гены, содержащиеся в дочерних микроядрах, теряются навсегда. Этот процесс подобен эндомейозу, о котором недавно сообщалось у тлей. Недавние генетические исследования показали, что виды Paramecium имеют классическое менделевское наследование многих признаков.
Хотя цитологические исследования. После того, как мы предложили приведенное выше описание полового размножения в течение примерно восьмидесяти лет, только благодаря генетическому анализу некоторые деления можно было с большей уверенностью определить как мейотические. Кроме того, некоторые признаки оказались непостоянными, в частности, свойства поверхностного белка ресничек. Эти белки развивались в ответ на температуру окружающей среды. Неизвестно, является ли изменение в структуре белка случайным или имеет какую-то функцию, поскольку каждое изменение является новым явлением даже у паразитических простейших.
Peritrichs (Отряд Peritrichida)
К ним относятся сидячие инфузории (Sessilina), которые живут на сокращающихся стеблях и часто встречаются у поверхности заросших аквариумов, и подвижные формы (Mobilina), которые, как полагают, произошли от сидячих форм. Характерными чертами отряда являются три специализированных ряда ресничек вокруг рта и общее отсутствие соматических ресничек. При взгляде спереди видно, что оральные реснички закручиваются в рот против часовой стрелки. Vorticella — типичный и распространенный род. Тело имеет форму колокола и закреплено заостренным концом на сократительной ножке. Во время питания стебель удлиняется, а коронка перистомиальных ресничек расширяется.
Paramecium является примером инфузорий (класс Ciliata)
Классы, подклассы и отряды простейших
#реснитчатые | Исследуйте сообщения и блоги Tumblr
adelicateculturecell · год назад
Текст
КЧ: «Вы когда-нибудь смотрели на микроба, который что-то делал, и думали: «Хотел бы я сделать это?» Я не знаю… может быть, нет. Не наблюдайте, как вортицелла небрежно берет еду издалека, и не думаете: «Вау, как бы мне не пришлось вставать, чтобы перекусить». , «Хм, разве не было бы здорово, если бы я мог просто охладить созданный мной пузырь, когда что-то соприкоснется?» И это нормально, хорошо быть довольным человеческим существованием».
Путешествие в микрокосмос. Могут ли водоросли питать наши автомобили?
Изображения, изначально сделанные Jam’s Germs
Цитата, озвученная Jam’s Germs
#Путешествие в микрокосмос#Могут ли водоросли питать наши машины? #hydra#vorticella#ciliate#science quotes
400 заметок · Посмотреть заметки
планктонная королева · год назад
Photo
Mesodinium rubrum
Произношение: Mesodin-ee-um rue-brum
Mesodinium — это планктонная инфузория, имеющая форму подсолнуха. Желтые пучки вокруг Мезодиния на самом деле представляют собой крошечные волоски, похожие на выросты, называемые ресничками. Реснички помогают ему передвигаться по поверхности океана. Кроме того, он может производить себе пищу с помощью фотосинтеза.
#планктон#mesodinium rubrum#инфузория#подсолнух#реснички#фотосинтез#биология#природа#морская биология#океанжизнь#планета земля#морская жизнь#морская#морская жизнь#океанская экосистема#эстетика#морские животные#образование#любитель океана#знание#не мой#knowlegde is power#море#животное#странный#нечетный
93 заметки · Посмотреть заметки
субботаxiii · год назад
Видео
youtube
Мне нравятся философские выводы, вытекающие из изучения клеточной жизни.
#микрокосм#смерть#жизнь#глубокая усадка#инфузория#шоу#рекомендуется
1 примечание · Посмотреть примечание
turquoisewaves07 · 11 месяцев назад
Текст
Лакримария инфузория — жестокий, микроскопический червь на ниточке
#червь на ниточке#червь сорванный с ниточки#lacrymaria#инфузория#инфузория#путешествие в микрокосмос#микрокосмос#крошечное существо
0 заметок
меризм · 6 месяцев назад
Видео
Вечеринка инфузорий, которую обычно выбрасывают в пробирки со старой прудовой водой, наполненные мертвыми и умирающими вещами (которые инфузории любят). Хотел бы я знать больше о передвижении инфузорий, например о том, почему одни выбирают медленный и равномерный темп, в то время как другие дергаются, казалось бы, наугад. Кстати говоря, посмотрите, сможете ли вы обнаружить Аспидиску, парня, похожего на медузу, который может практически телепортироваться.
#инфузории#микроскопия
94 заметки · Просмотр заметок
странных организмов · год назад
Текст
Блестящие инфузории
От @tardibabe в IG:
Эта блестящая инфузория, скорее всего, из класса Prostomatea и, возможно, из рода Holophrya (напишите мне, если у вас есть идеи!) . Как и его двоюродный брат нассула, он обладает ротовой корзиной, предназначенной для поедания цианобактерий и других нитчатых водорослей!
Инфузории Prostomateans живут как в пресноводной, так и в морской среде и, как известно, цветут, когда их добыча, например та, что вызывает красные приливы, в изобилии. Известно, что некоторые другие простоматеи, такие как раздражители Cryptocaryon, вызывают кожные заболевания у морских рыб.
Инфузории Prostomatida, такие как Holophrya, могут быть многочисленными в популяциях планктона и были обнаружены по всему миру, от небольших временных прудов до образцов почвы, рисовых полей и даже соленых озер в Европе, Австралии и Чаде!
Поскольку их можно найти в почве, они должны иметь способность образовывать спящие цисты, как и наши маленькие моховые поросята! Формируя эти покоящиеся цисты, они могут выживать в течение длительного периода времени при отсутствии воды. Когда идет дождь или затапливается почва, эти инфузории могут распуститься и снова стать активными 😄
Если вы когда-нибудь захотите поддержать мою работу, сделав небольшое пожертвование (любая сумма считается) и просто запустив страницу ко-фи: http://ko-fi.com/tardibabe 💜
Саундтрек предоставлен @accelerator.jengold ✨
Видео снято на мой iPhone, установленный на микроскоп BA310E Motic с адаптером @ilabcam 🔬
Использованная литература:
Фойсснер, В., и Хоксворт, Д.Л. (ред.). (2009 г.). Разнообразие простейших и географическое распространение. Темы биоразнообразия и сохранения.
Х., Линн, Денис (2010). Реснитчатые простейшие: характеристика, классификация и справочник по литературе. Нью-Йорк: Спрингер.
#странные организмы#инфузории#микроскопия#образовательный
2 заметки · Посмотреть заметки
rattyexplores · год назад
Фото
Исследование чешуекрылых — 01.03.21 — Anthene lycaenoides, “Pale Ciliate Blue” Надсемейство Papilionoidea; Семейство Lycaenidae; Подсемейство Polyommatinae; Племя Lycaenesthini
Гусеница еще не сфотографирована, но ее цвет варьируется от желтого до зеленого и фиолетового в зависимости от цвета бутонов, которые она потребляет.
Пищевые растения представляют собой бутоны и цветки различных растений, в том числе Cassia fistula «Golden Shower», Flagellaria indica «Supplejack», Bridelia tomentosa «Pop-gun Seed», Pongamia pinnata «Pongam», Clerodendrum floribundum «Lolly Bush», Rhyssopterys timorensis «Timor Liana» и Cupaniopsis anacardioides «Carrotwood».
Муравьи, которые сопровождают эту гусеницу, включают Oecophylla smaragdina «Цитрусовые муравьи», Crematogaster «Муравьи-акробаты», Iridomyrmex anceps «Черные муравьи» и Odontomachus ruficeps «Ржавые муравьи-люки»
Этот вид встречается в QLD, WA и NT. За пределами Австралии: Индонезия, Малайзия, Новая Гвинея и Таиланд.
(Источник). #бабочка#взрослая бабочка#взрослая форма#бабочки#членистоногие#артропода
2 примечания · Посмотреть заметки
синингатдива · год назад
Фото
Микробы, обнаруженные в воде пруда в городе Пуэрто-Принсеса, Палаван, Филиппины
2 примечания · Посмотреть примечания
бабочкакерала · 8 месяцев назад
Текст
Anthene lycaenina lycaenina (R. Felder, 1868) – Даханская остроконечная ресничная синяя
വനകോകിലൻ
#Синяя остроконечная ресничная#Anthene lycaenina#butterfly 09 09 mmph 09 09003
4 0
0
വനകോകിലൻ · год назад
Текст
АННОТИРОВАННЫЙ КОНТРОЛЬНЫЙ СПИСОК ЭПИБИОНТ ЦИЛИАТЫ _ Zoothamnium | UTTAR PRADESH JOURNAL OF ZOOLOGY
На основе изучения существующей литературы по Zoothamnium на всех континентах был составлен контрольный список одной из эпибионтных инфузорий рода Zoothamnium со всей необходимой информацией. Этот контрольный список пригодится, если вы хотите больше учиться в этой области. В этом списке 57 видов Zoothamnium.
См. ссылку: — http://mbimph.com/index.php/UPJOZ/article/view/2147
#Zoothamnium#checklist#epibionts#ciliates
0 примечаний
adelicatculturecell · год назад
Текст
куб.см: «Когда дело доходит до движения, на инфузорий приятно смотреть. Их похожие на волосы реснички бьются волнами, направляя их по микрокосму, и это здорово, если вы пытаетесь попасть из Точки А в точку Б. Но иногда вы на самом деле ничего не пытаетесь достичь. Вы просто пытаетесь быть немного меньше в том месте, где вы сейчас находитесь».
Journey to the Microcosmos- Flinching Saves Lives in the Microcosmos
Изображения, изначально полученные Jam’s Germs
Цитата, озвученная Хэнком Грином #микробиология#микробы#микроорганизмы#стентор#инфузория#научные цитаты
198 заметок · Посмотреть заметки
xignis · год назад
Текст
еще микроорганизмы, на этот раз гидра
#кажется, ранена?#все должно быть в порядке, так как у гидры сумасшедшая регенерация#я люблю их#микробиология#микроорганизм#гидриды#книдарии#хигсштуки#я думаю, что это инфузории как колодец
0 нот
бдительность · год назад
Видео
youtube
Стать собственным ребенком через спряжение | Путешествие в микрокосмос
#sfw#путешествие в микрокосмос#микробиология#микробы#инфузории#размножение
0 нот
чакитрон · год назад
Текст
Сукторианы: Гадкий утенок инфузорий
Сукторианы: Гадкий утенок инфузорий
Сукторианы: Гадкий утенок инфузорий
Категория Основная
Описание:
Этот выпуск представляет вам альбом «Музыка для ученых»! Прослушайте альбом на основных музыкальных сервисах здесь: https://biglink. to/music-for-scientists. Проверить …
TopTrengingTV Охота за самым трендовым видео на данный момент, каждый час, каждый день 24/7.
Видеоданные Youtube
Опубликовано: 2021-01-25T17:22:54Z
Теги: …
Посмотреть на WordPress популярные видео на YouTube#ВИДЕО#водяной медведь#видео на YouTube
0 примечаний
montereybayaquarium · год назад
Текст
Не позволяйте их нежной красоте обмануть вас — гребневики-гребенники, с которыми нужно считаться!
Лопастные гребенчатые желеобразные, такие как Bolinopsis infundibulum, собирают планктон с воронкообразными лопастями в паре с ушными раковинами, в то время как морские грецкие орехи Hormiphora californiensis используют двойные щупальца, покрытые липкими нитями, для траления в поисках микроскопических закусок.
Но берегитесь цветения берое! Эти студенистые рты-с-повесткой дня выслеживают свою добычу (часто других гребешков!)
Гребневики оказывают огромное влияние на доступный планктон, когда они рядом, и настолько плодовиты эти хорошо причесанные обедающие, что вторжение гребневиков в новые места обитания может вызвать экологическую катастрофу!
Так что в следующий раз, когда вы будете очарованы завораживающими рядами радуг рассеянного света, танцующих вдоль их сторон, подумайте о том, насколько грозными являются эти трясущиеся безжалостные силы в своем водном образе!
#аквариум в монтерей-бей#несанкционированная неделя желе#радужная связь, которую мы нашли#расчешите здесь и проверьте это#гребенчатый пластинчатый удивительный#гребневик#гребенчатое желе
2K note · Посмотреть заметки
statecryptids · 7 дней назад
Текст
Еще одна страница космического планктона из моего полевого справочника по Сатурну.
Это всего лишь небольшая часть существ, дрейфующих в разреженной атмосфере колец Сатурна. Эвриалы — это медузоподобные организмы со стрекательными клетками на гофрированных щупальцах, которые они используют для захвата пищи. Группа названа в честь Эвриалы, сестры Горгоны Медузы.
Инфузории представляют собой разнообразную группу одноназываемых организмов, обладающих волосовидными ресничками.
#спекулятивная фантастика#спекулятивная биология#инопланетный дизайн#планктон
27 заметок · Просмотр примечаний
чарлидон · 9 месяцев назад
Примечание
Что семья Ганнибала подумала бы о читателе, страдающем аутизмом, который интересуется энтомологией (набивкой жуков)?
«Это потрясающе! Вы сами их нашли?»
Он обязательно успокоит вас с самого начала и похвалит вашу коллекцию жуков.
Его любимое насекомое — бабочка, так как у нее так много цветов, и она попытается поговорить с вами об этом, завязав разговор, который вам обоим будет удобен.
Он никогда не сделает ничего, что может причинить вам дискомфорт, и постарается заставить вас раскрыться как можно шире.
Не любитель жуков..
Но он понимает вашу страсть, как и свою.
У тебя может и не быть такого хобби, но он как-нибудь попытается заставить его работать ?
«Крутое насекомое.. Может, мне попробовать его покрасить ?»
Тогда вы будете проводить часы рядом друг с другом в приятной тишине; ты пронзаешь своих насекомых, а он рисует их.
Он поддерживает.
Возможно, ему не особенно нравится тот факт, что вы приходите домой весь в грязи после того, как большую часть дня провели в погоне за летающими жуками всех видов… но он понимает.
Он знает, что это твой способ развлечься, и не будет осуждать тебя за это.
Он даже будет участвовать, когда придет время приколоть их к доске или помочь вам найти их имена в некоторых книгах по энтомологии.
Почему-то ему нравится время, которое вы проводите вместе; вместе ищем и любуемся жуками.
«Этот называется…Anthene Seltuttus? Более известный как Темно-Ресничный-синий. Мне он нравится. Цвет великолепный, такой глубокий и насыщенный фиолетовый. Молодец !»
Ганнибал-старший сидит в тюрьме, так что особо помочь не может.
Но он всегда очень рад, когда вы приходите и показываете ему свою коллекцию, а также новинки.
«Очень хорошо. Молодец, жук-обнимашка.»
Вы даже дали ему в прошлом то, что до сих пор лежит в его камере. К сожалению, булавки пришлось снять из соображений безопасности, но он держит их в безопасности в маленькой деревянной коробочке и смотрит на них каждый раз, когда хочет чувствовать себя как дома.
Он находит их утешительными и всегда спит рядом с маленькой коробочкой.
Вы нравитесь Моргану, потому что вы так же увлечены природой, как и он, это позволяет ему сопровождать вас в лесу.
Одним из любимых занятий Моргана является долгая прогулка по лесу, это помогает ему расслабиться.
Он закрывает глаза и слышит почти незаметный голос леса; из всех звуков, которые издают его обитатели.