Процесс питания реснитчатых инфузорий
Процесс питания реснитчатых инфузорий вместе с анализом всей пищевой системы подробно описан Догелем и Полянским.
По характеру пищи инфузории делятся на: 1) бактериофагов и детритофагов, 2) сапрозоев и 3) хищников и питающихся смешанной пищей. К типичным бактериофагам среди свободноживущих инфузорий относится туфелька Paramaecium caudatum, хотя она иногда заглатывает и одноклеточные водоросли, и бесцветных жгутиковых. Последних поедают также Stentor, Spirostomum, Frontonia и другие инфузории. На способность инфузорий питаться водорослями, даже синезелеными, указывают Винберг, Сорокин и др., Мордухай-Болтовская и Сорокин, хотя опыты с кормлением инфузорий Microcystis показали пищевую неполноценность последнего для инфузорий.
Характер заглатывания пищи у инфузорий зависит от устройства околоротового аппарата. Различают «глотателей» – хищников, иногда прожорливых, как, например, Coleps hirtus, Didynium nasutum, которые захватывают крупную добычу и у которых рот в покоящемся состоянии закрыт, и «подгонятелей», у которых рот постоянно открыт.
Среди хищных инфузорий некоторые обладают хорошо выраженной избирательной способностью при питании. Например, Didynium nasutum питается преимущественно инфузориями, Nassula – синезелеными. Эберли обнаружил убыль в металимнионе одного озера популяции Oscillatoria agardhii в результате выедания ее Nassula aurea.
Проведенный выше обзор различных аспектов питания у представителей зоопланктона, позволяет прийти к следующему важному в практическом отношении заключению. Ввиду наличия тесного трофического взаимодействия между зоопланктоном и фитопланктоном, зная скорость питания зоопланктона, степень усвоения потребленных водорослей и пищевые потребности животных, можно определить роль фитопланктона в удовлетворении пищевых потребностей зоопланктона в тех водоемах, где планктон изучен с достаточной полнотой. А из сопоставления величин первичной продукции в водоеме и рыбопродукции можно также рассчитать, какой процент первичного органического вещества воспроизводится в теле конечного трофического звена – рыбы.
Ввиду наличия тесного трофического взаимодействия между зоопланктоном и фитопланктоном, зная скорость питания зоопланктона, степень усвоения потребленных водорослей и пищевые потребности животных, можно определить роль фитопланктона в удовлетворении пищевых потребностей зоопланктона в тех водоемах, где планктон изучен с достаточной полнотой. А из сопоставления величин первичной продукции в водоеме и рыбопродукции можно также рассчитать, какой процент первичного органического вещества воспроизводится в теле конечного трофического звена – рыбы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите
ГДЗ биология 7 класс Константинов, Бабенко Вентана-Граф Задание: § Стр 43
На данной странице представлено детальное решение задания § Стр. 43 по биологии для учеников 7 классa автор(ы) Константинов, Бабенко
§ Стр. 43
Вопросы после параграфа
№ 1. Докажите на конкретных примерах, что инфузории имеют более сложное строение, чем амебовые и жгутиковые.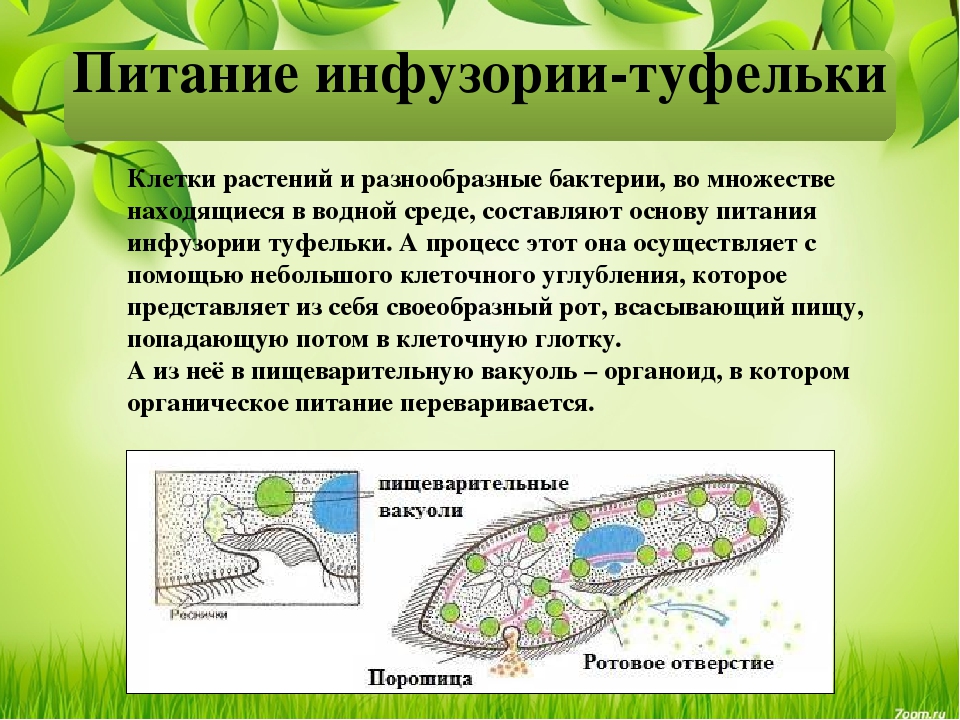
У инфузории-туфельки более сложное строение, чем у амёбовых и жгутиковых, которое выражено в наличии в их строении двух ядер. Одно ядро – большое и отвечает за такие процессы, как дыхание, питание, обмен веществ, передвижение. Второе ядро – маленькое, принимающее участие в половом размножении инфузорий.
Также постоянная форма у инфузории поддерживается при помощи тонкой эластичной оболочки, которая покрывает ее снаружи. А прилегающий к оболочке слой цитоплазмы дополнительно укреплен опорными волокнами. Движение инфузории-туфельки осуществляется благодаря нескольким тысячам ресничек, которые слаженно колеблются и за счет этого толкают ее тело вперед.
№ 2. Установите связь между усложнением строения инфузории-туфельки и процессами питания и выделения.
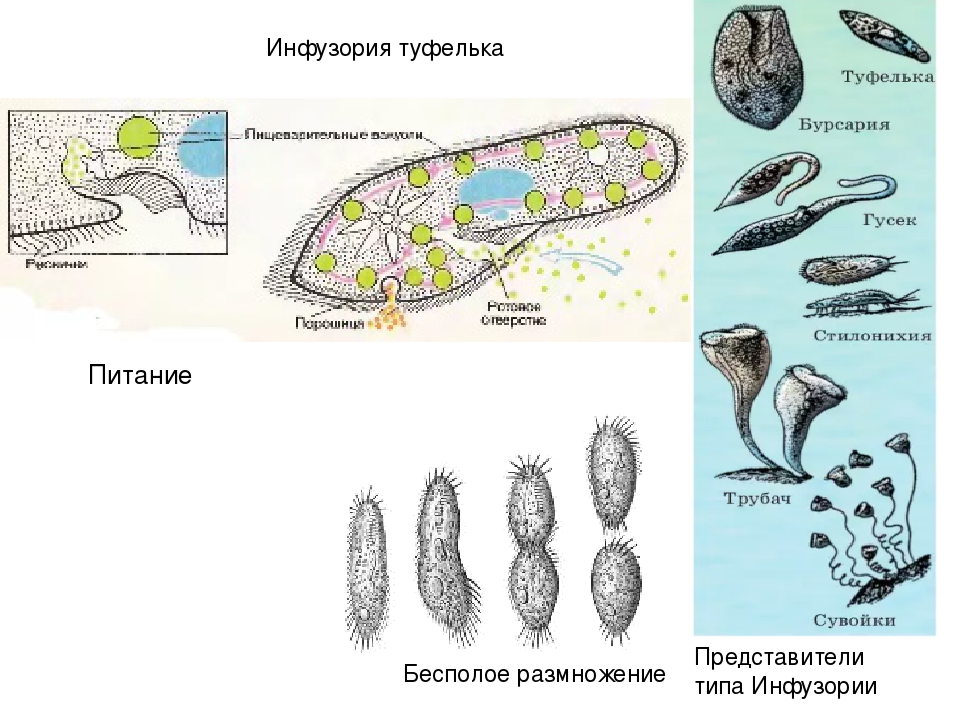
У инфузории-туфельки гетеротрофный тип питания, потому как питается она готовыми органическими веществами. Попадают они сначала в клеточный рот – углубление на теле инфузории, около которого располагается большое количество длинных и толстых ресничек.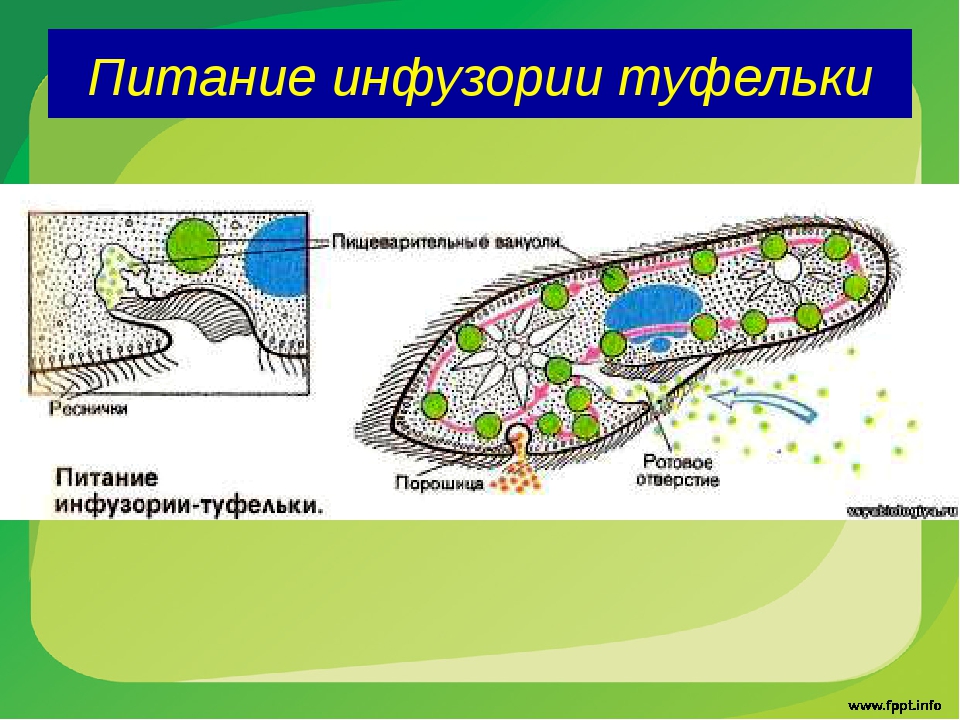 Именно движением этих ресничек с током воды бактерии и мельчайшие частички загоняются в клеточную глотку. На ее дне пища попадает в пищеварительные вакуоли, которые постоянно перемещаются в теле инфузории благодаря току цитоплазмы. Остатки пищи, которые не переварились, выбрасываются через порошицу – особая структура, которая располагается на заднем конце инфузории-туфельки.
Именно движением этих ресничек с током воды бактерии и мельчайшие частички загоняются в клеточную глотку. На ее дне пища попадает в пищеварительные вакуоли, которые постоянно перемещаются в теле инфузории благодаря току цитоплазмы. Остатки пищи, которые не переварились, выбрасываются через порошицу – особая структура, которая располагается на заднем конце инфузории-туфельки.
Органом выделения служат две сократительные вакуоли, которые находятся у разных концов тела инфузории. Каждая из них состоит из центрального резервуара и нескольких каналов, направленных к нему. Лишняя жидкость сначала попадает в каналы, а потом в центральный резервуар. Примерно 2 – 4 раза в минуту вакуоли поочередно сокращаются, выталкивая накопленную жидкость и растворенные в ней вредные продукты жизнедеятельности во внешнюю среду.
№ 3. Охарактеризуйте особенности процесса размножения инфузории-туфельки.
Инфузории-туфельки размножаются обычно бесполым способом – делением клетки надвое при помощи образования перетяжки. Но, в отличие от жгутиковых, у которых тело делится вдоль, у инфузории оно делится поперек. Первым начинает делиться ядро – большое и малое. Таким образом, у каждой образовавшейся клетки есть по два ядра и часть органоидов. Недостающие органоиды образуются заново. При этом деление может происходить несколько раз в сутки при благоприятных условиях.
Но, в отличие от жгутиковых, у которых тело делится вдоль, у инфузории оно делится поперек. Первым начинает делиться ядро – большое и малое. Таким образом, у каждой образовавшейся клетки есть по два ядра и часть органоидов. Недостающие органоиды образуются заново. При этом деление может происходить несколько раз в сутки при благоприятных условиях.
Половое размножение у инфузорий-туфелек заключается в контакте двух клеток, в результате которого происходит обмен содержимым ядер через мостик цитоплазмы, образовавшийся в месте соприкосновения оболочек. Большие ядра в теле клеток исчезают, а малые начинают дважды делиться, образуя в каждой дочерней клетке по четыре маленьких ядра. Далее три ядра разрушаются, а одно начинает снова делиться. Таким образом, в каждой клетке образуется два малых ядра, одно из которых переходит по цитоплазматическому мостику к другой особи, где и сливается с оставшимся там ядром. Данный процесс называется конъюгацией.
№ 4. Объясните, почему половой процесс не является половым размножением.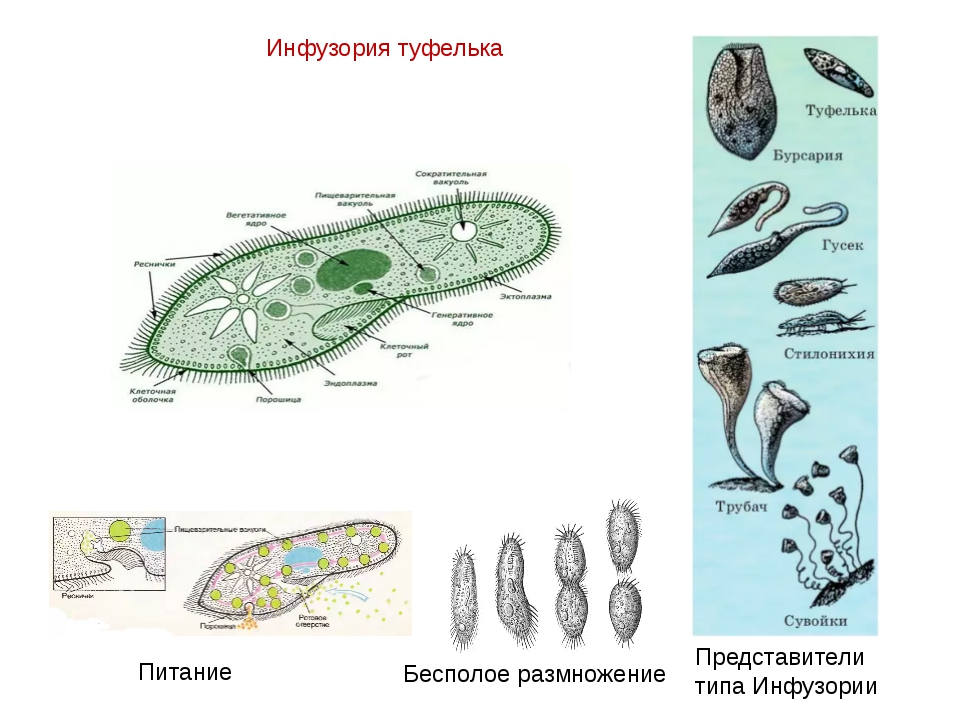 В чем его биологическое значение?
В чем его биологическое значение?
При половом процессе между инфузориями-туфельками, который называется конъюгацией, не происходит увеличение числа особей. В месте соприкосновения двух организмов оболочки растворяются, образуя своеобразный мостик из цитоплазмы. В результате большие ядра исчезают, а малые начинают делиться.
Таким образом, в каждой инфузории-туфельке образуется по четыре маленьких ядра. Далее по цитоплазматическому мостику происходит обмен ядрами – от каждой особи одно из ядер переходит к другой особи, где и сливается с оставшимся ядром.
Данный процесс происходит для того, чтобы клетки могли обменяться генетическим материалом. Его биологическое значение состоит в обновлении генетического материала и, как следствии, возможном появлении у клеток новых признаков.
Параграф Значение простейших
Стр. 43
Вспомните
№ 1. Какими чертами строения эвглена зеленая и инфузория-туфелька отличаются от амебы протей?
Эвглена зеленая является активно передвигающимся простейшим, у которого есть органоиды для движения – жгутики.
Инфузория-туфелька является наиболее сложноорганизованным простейшим, на поверхности тела которого есть такие органоиды движения, как реснички. Также ее отличительной особенностью является наличие в ее составе двух ядер – малого и большого, каждое из которых отвечает за определённые функции. Форма тела у инфузории-туфельки веретеновидная, напоминающая туфлю, постоянная за счет тонкой эластичной оболочки, которая покрывает его снаружи. Также к оболочке прилегает слой цитоплазмы, который укреплен опорными волокнами.
Внешне амеба протей напоминает небольшой студеный комочек. Тело ее не имеет постоянной формы, потому что лишено плотной оболочки. Оно образует выросты или ложноножки, при помощи которых амеба может медленно передвигаться из одного места в другое, захватывая добычу. В самостоятельном организме амебы протей содержится цитоплазма, которая покрыта клеточной мембраной. Наружный слой ее плотный и прозрачный, внешний – более текучий и зернистый. Также в цитоплазме находится ядро, пищеварительная и сократительная вакуоли.
№ 2. В каких средах жизни обитают простейшие?Простейшие могут обитать в самых разнообразных условиях среды. Большинство среди них – водные организмы, которые широко распространены в морских и пресных водоемах. Некоторые из них живут даже в придонных слоях, а также входят в состав бентоса.
Небольшое число видов простейших приспособилось к выживанию в почве, а именно в тончайших оболочках воды, которые окружают почвенные частицы и заполняют капиллярные просветы в структуре почвы.
Есть простейшие, которые ведут паразитический образ жизни – используют другие живые организмы в качестве своей среды обитания и источника пищи. Это могут быть растения, животные и даже человек.
№ 3. Какие организмы называют паразитами?
Паразитами называют организмы, которые выбирают другие организмы в качестве своей среды обитания (внутри или на поверхности тела) и питаются за их счет, отравляя и нанося вред продуктами своей жизнедеятельности.
Более 2 млн видов животных, обитающих повсеместно, отличаются
от растений гетеротрофностью питания, подвижностью, развитой опорно-двигательной
системой, сложностью поведенческих реакций. Согласно принятой точке зрения,
ближе всего к общему предку животных и растений находятся жгутиковые одноклеточные
— эвглены, которые могут питаться как автотрофно, так и гетеротрофно, и
активно передвигаться, отвечая на раздражения.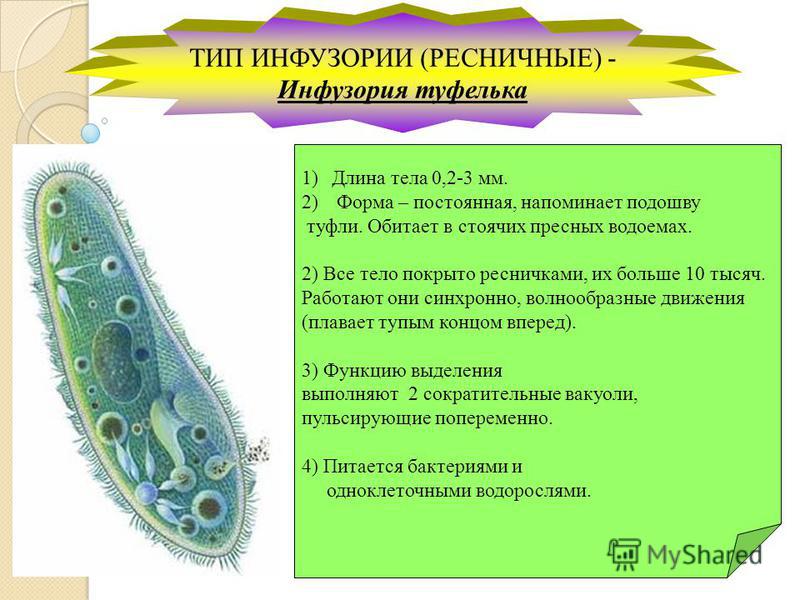 |
| В систематику животных включены следующие таксоны: царство, тип, подтип, класс, отряд, семейство, род, вид. |
| Одноклеточные животные (Простейшие) |
| Тип Простейшие — процветающая и разнообразная группа организмов. Известно около 50 000 видов простейших, которых в природе можно обнаружить повсюду, где есть вода. Каждое простейшее представляет собой самостоятельный организм, способный выполнять все необходимые для жизни функции. |
| Подавляющее большинство простейших обладает аэробным типом обмена. Для дыхания они используют кислород, растворенный в воде. Окисление происходит в митохондриях. Виды, паразитирующие в кишечнике своих животных-хозяев — амебы, жгутиконосцы — анаэробны, поэтому утратили свои митохондрии. |
Амеба — представитель класса Саркодовые. Обитает в небольших
мелких прудах или проточных канавах с илистым дном. Тело амебы достигает
0,1 мм и ограничено тончайшей плазмолемой (плазматической мембраной).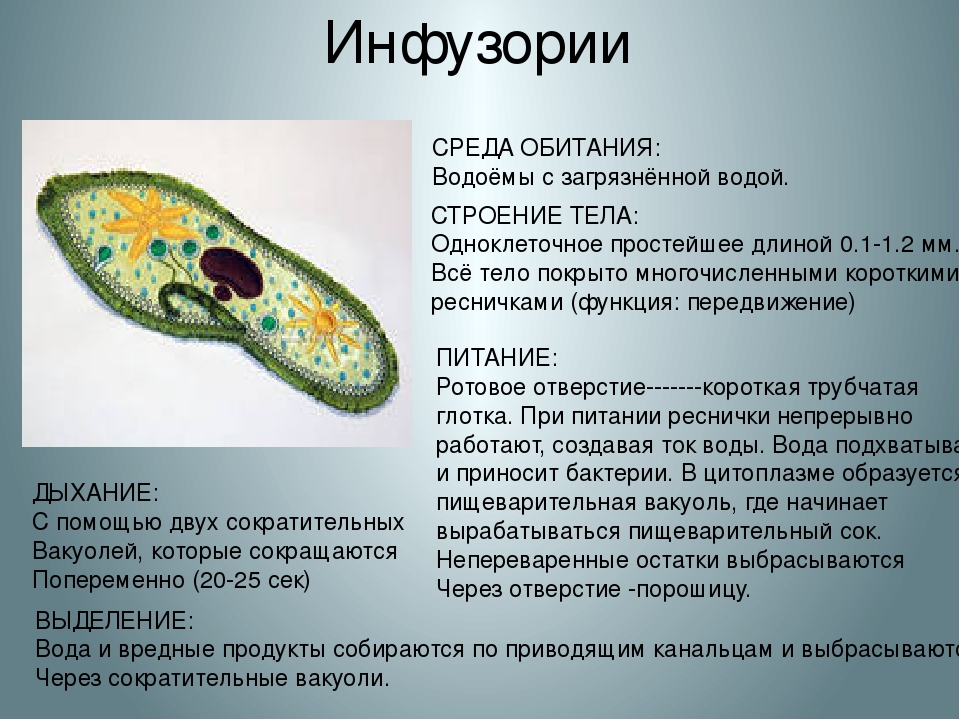 Протоплазма
подразделяется на экто- и эндоплазму. Ядро в клетке регулирует процессы
метаболизма и деления клеток, не занимает определенного положения. Цитоплазма
содержит пищеварительные вакуоли, формирующиеся в разных участках клетки
вокруг пищевых комочков, путем выделения пищеварительного сока из цитоплазмы.
Пищей для нее служат одноклеточные водоросли, жгутиковые, инфузории. Сократительная
вакуоль, периодически сокращаясь, выделяет наружу избыток воды с растворенными
ненужными веществами в любой точке тела амебы. Кислород поступает через
всю поверхность тела амебы. Форма тела амебы постоянно меняется из-за образующихся
цитоплазматических выростов — псевдоподий (ложноножек), служащих для захвата
пищи (фагоцитоза) и передвижения. Протоплазма
подразделяется на экто- и эндоплазму. Ядро в клетке регулирует процессы
метаболизма и деления клеток, не занимает определенного положения. Цитоплазма
содержит пищеварительные вакуоли, формирующиеся в разных участках клетки
вокруг пищевых комочков, путем выделения пищеварительного сока из цитоплазмы.
Пищей для нее служат одноклеточные водоросли, жгутиковые, инфузории. Сократительная
вакуоль, периодически сокращаясь, выделяет наружу избыток воды с растворенными
ненужными веществами в любой точке тела амебы. Кислород поступает через
всю поверхность тела амебы. Форма тела амебы постоянно меняется из-за образующихся
цитоплазматических выростов — псевдоподий (ложноножек), служащих для захвата
пищи (фагоцитоза) и передвижения. |
Класс Инфузории насчитывает более 10 000 видов. Тело инфузорий
имеет постоянную форму, вся его поверхность несет многочисленные реснички,
которые; по строению являются укороченными жгутиками. Ядерный аппарат инфузорий
устроен более сложно, чем у других классов простейших.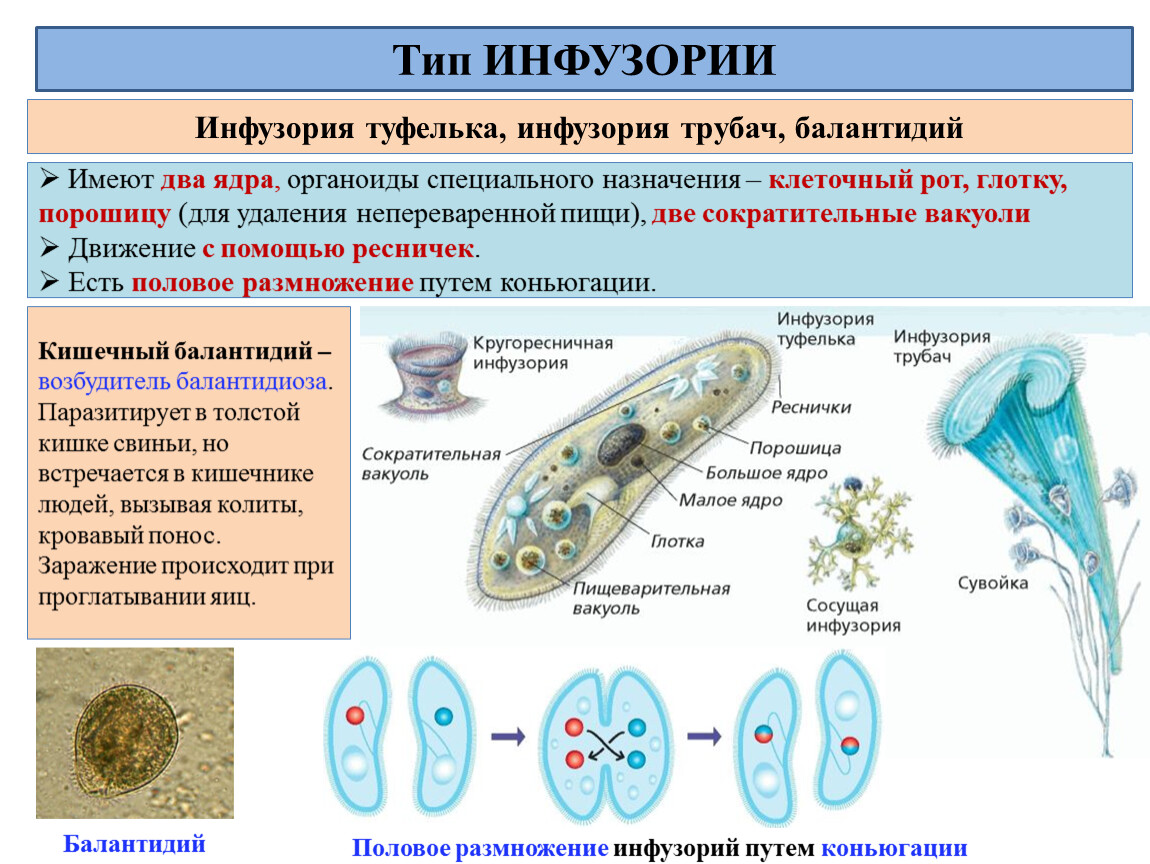 У инфузорий два ядра:
большое бобовидное ядро — макронуклеус — контролирует процессы метаболизма
и дифференцировки клетки; мелкое — микронуклеус — регулирует размножение. У инфузорий два ядра:
большое бобовидное ядро — макронуклеус — контролирует процессы метаболизма
и дифференцировки клетки; мелкое — микронуклеус — регулирует размножение. |
| Инфузория-туфелька обитает в стоячих водоемах с большим количеством разлагающейся органики. На ее вентральной поверхности находится постоянное углубление — околоротовая воронка. Сужаясь, она переходит в глотку, которая заканчивается клеточным ртом или цитостомом. Реснички околоротовой воронки загоняют бактерии и частички пищи в глотку. На конце глотки формируются пищеварительные вакуоли, которые движутся в цитоплазме по постоянной траектории. Непереваренные частицы выводятся через порошицу наружу. Две сократительные вакуоли занимают постоянное место. Вокруг каждой из них находятся радиальные каналы, в которые поступает вода из цитоплазмы. |
Для инфузорий характерно в основном бесполое размножение
путем поперечного деления тела надвое. При недостатке пищи они переходят
к половому процессу путем конъюгации, который возможен только между совместимыми
особями одного вида.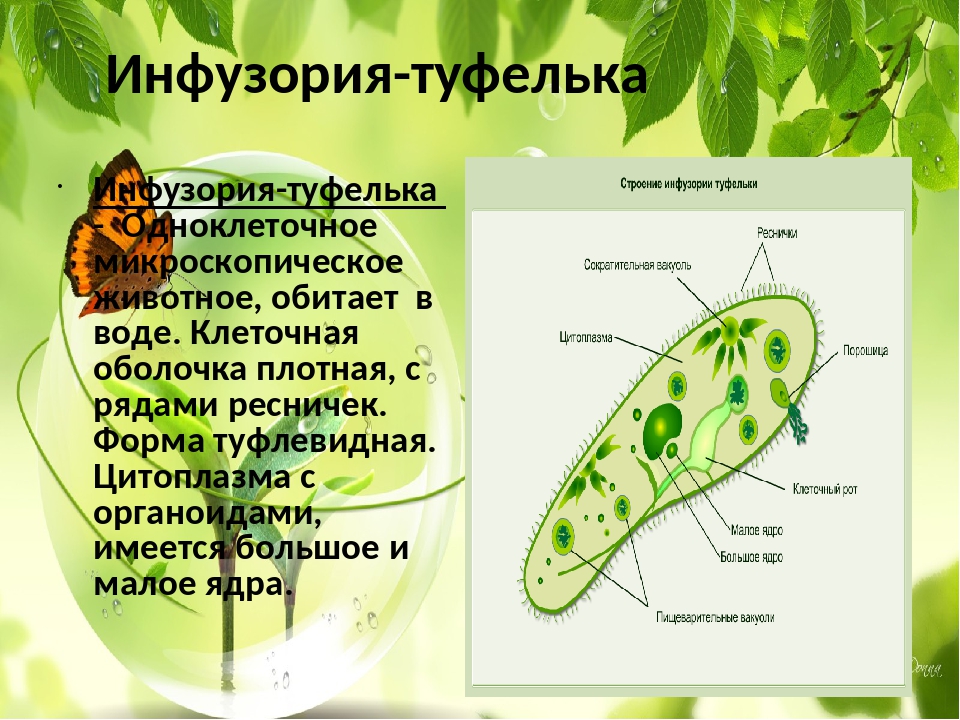 Этот процесс обеспечиваетобмен генетической информацией
при участий микронуклбусов. Этот процесс обеспечиваетобмен генетической информацией
при участий микронуклбусов. |
| Жгутиконосцы — самая многочисленная группа простейших, в состав которых входят автотрофные и гетеротрофные организмы. Характерной их особенностью является наличие жгутиков, количество которых варьирует. Жгутиконосцы обладают постоянной формой тела и размножаются преимущественно делением. Поддерживание постоянной формы тела у жгутиковых и инфузорий возможно за счет того, что самый поверхностный слой их клеточного тела сильно уплотняется и становится упругим. Кроме того, здесь концентрируются многочисленные нитевидные белковые структуры, образующие цитоскелет. Автотрофные жгутиковые обладают хроматофорами, содержащими хлорофилл, благодаря чему они способны к фотосинтезу. Гетеротрофное питание у жгутиконосцев осуществляется через клеточный рот или поверхность тела пиноцитозом. Среди Жгутиконосцев и Инфузорий широко распространена колональность. |
Таблица «Инфузория — туфелька» (для учащихся 7 классов)
МКОУ «Новокаякентская СОШ»
с. Новокаякент
Новокаякент
Каякентский район Республика Дагестан
Таблица «Инфузория — туфелька»
(для учащихся 7 классов)
Автор: учитель биологии
МКОУ «Новокаякентская СОШ»
Умалатова Равганият Бийбулатовна
с.Новокаякент
2018 г.
Пояснительная записка
Данная таблица «Инфузория — туфелька» рекомендуется для учащихся
7 классов. Материал можно использовать при прохождении темы
«Подцарство Простейшие» в 7 классах. В таблице дана характеристика инфузории — туфельки. Характеристика включает: систематику, строение, размеры, среду обитания, питание, дыхание, выделение, размножение, образование цисты и значение инфузории — туфельки. Таблицу можно использовать при подготовке к ОГЭ и ЕГЭ по биологии.
Задачи: ознакомление учащихся с систематикой, строением, размерами, со средой обитания, питанием, дыханием, выделением, размножением, со значением инфузории — туфельки.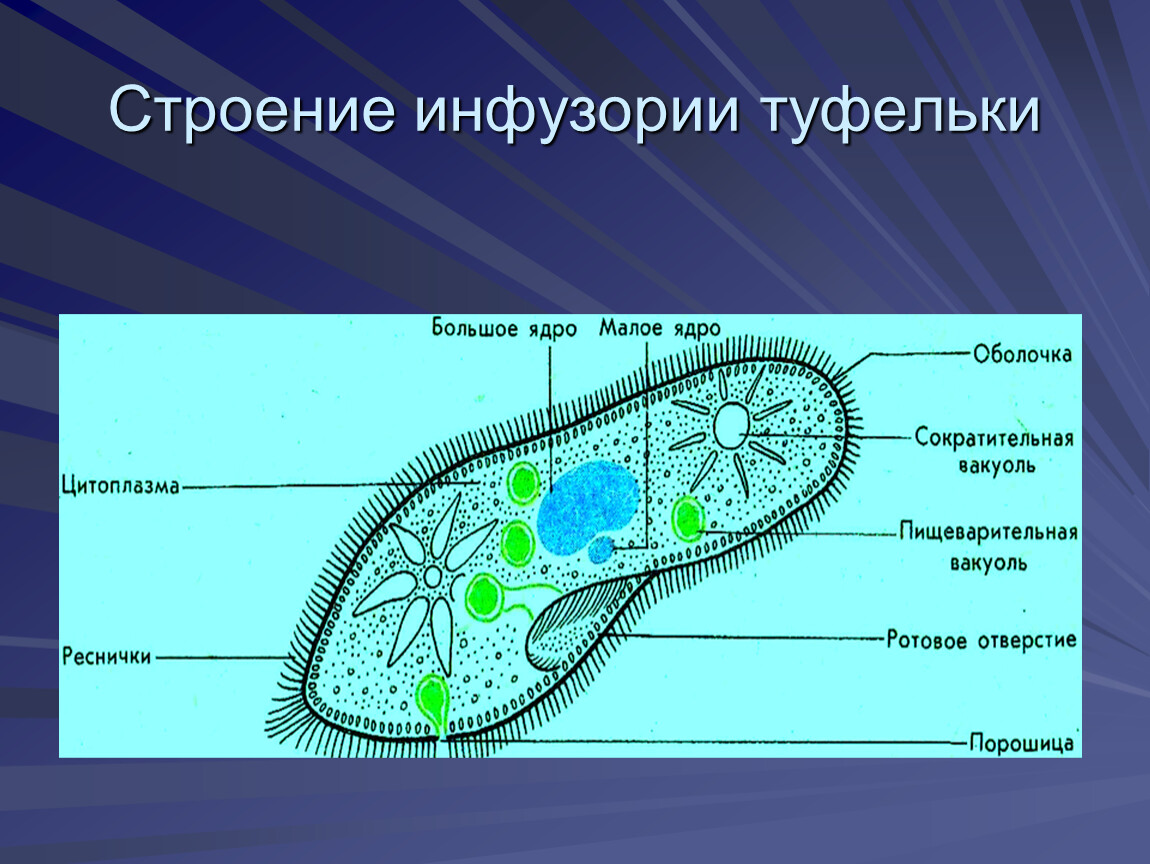
Таблица «Инфузория — туфелька»
Инфузория — туфелька
Систематика инфузории — туфельки
Империя: Клеточные
Надцарство: Ядерные
Царство: Животные
Подцарство: Одноклеточные (простейшие)
Тип: Инфузории
Класс: Ресничные инфузории
Подкласс: Равноресничные инфузории
Род: Парамеции
Вид: Инфузория – туфелька
Среда обитания
Обитатель стоячих вод, встречается также в пресноводных водоемах с очень слабым течением, содержащих разлагающийся органический материал.
Размеры
Размер инфузории туфельки составляет
0,1—0,3 мм.
Форма тела
Тело (клетка) Инфузории напоминает след человеческой туфельки (отсюда название)
Строение
Питание
На середине брюшной стороны тела туфельки расположен клеточный рот, окруженный длинными ресничками. С помощью ресничек инфузория загоняет в рот пищу — бактерий и другие мелкие организмы.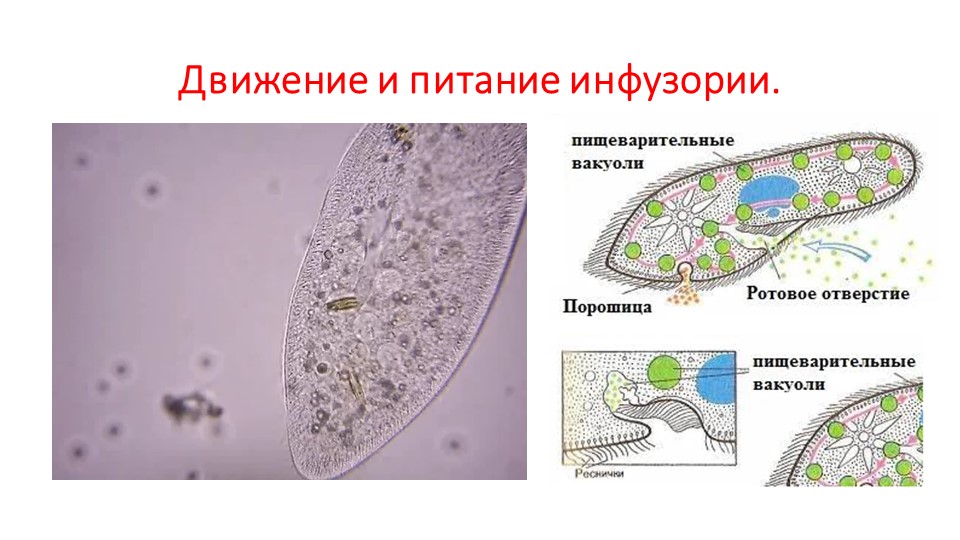 Рот ведет в глотку, на дне которой образуются пищеварительные вакуоли. Токами цитоплазмы вакуоли перемещаются в теле инфузории. В это время в них происходит переваривание и перенос питательных веществ в цитоплазму. Непереваренные остатки пищи выбрасываются из вакуолей через специальное отверстие в пелликуле — порошицу, расположенную на брюшной стороне задней части тела.
Рот ведет в глотку, на дне которой образуются пищеварительные вакуоли. Токами цитоплазмы вакуоли перемещаются в теле инфузории. В это время в них происходит переваривание и перенос питательных веществ в цитоплазму. Непереваренные остатки пищи выбрасываются из вакуолей через специальное отверстие в пелликуле — порошицу, расположенную на брюшной стороне задней части тела.
Движение
Совершая ресничками волнообразные движения, туфелька передвигается (плывёт тупым концом вперёд). Ресничка движется в одной плоскости и совершает прямой (эффективный) удар в выпрямленном состоянии, а возвратный — в изогнутом
Дыхание
Газообмен осуществляется через всю поверхность тела путем диффузии. Кислород проникает в клетку и используется в процессе дыхания, а образующийся углекислый газ выходит наружу.
Размножение
Размножение инфузории происходит бесполым путем, продольным делением клетки.
Половое размножение у инфузории – туфельки происходит между двумя животными. Между двумя особями образуется цитоплазматический мостик, макронуклеусы разрушаются, а микронуклеусы делятся мейозом с образованием четырех гаплоидных ядер, три из которых погибают, а четвертое делится пополам, но уже митозом, образуя два ядра: одно — стационарное, другое — мигрирующее. Затем между инфузориями происходит обмен мигрирующими ядрами. Потом стационарное и мигрировавшее ядра сливаются, особи расходятся, и в них снова образуются микро- и макронуклеусы.
Между двумя особями образуется цитоплазматический мостик, макронуклеусы разрушаются, а микронуклеусы делятся мейозом с образованием четырех гаплоидных ядер, три из которых погибают, а четвертое делится пополам, но уже митозом, образуя два ядра: одно — стационарное, другое — мигрирующее. Затем между инфузориями происходит обмен мигрирующими ядрами. Потом стационарное и мигрировавшее ядра сливаются, особи расходятся, и в них снова образуются микро- и макронуклеусы.
Процесс у инфузорий не приводит к увеличению числа особей.
Выделение
Непереваренные остатки удаляются через порошицу. Избыток воды и продукты обмена веществ через две сократительные вакуоли расположены в противоположных концах тела.
Раздражимость
Инфузории туфельки обладают раздражимостью. При столкновении с посторонними предметами они способны мгновенно менять направление движения на противоположное. У них хорошо развит хемотаксис: инфузории активно движутся в сторону пищи и, наоборот, избегают вредных химических воздействий.
Циста инфузории — туфельки
В неблагоприятных условиях (низкая температура, высыхание водоема) эвглена зеленая образует цисту. При образовании цисты происходит отпадание жгутика, клетка приобретает округлую форму и покрывается плотной оболочкой.
Значение инфузории- туфельки
Инфузория туфелька является элементом биологического разнообразия на Земле. Она участвует в круговороте веществ в природе. Она является составной частью пищевых цепей: Инфузория питается бактериями и детритом, ею питаются мальки рыб, гидры, какие-то черви, мелкие ракообразные.
Источники информации:
1.Захаров В.Б., Сонин Н.И. Биология. Многообразие живых организмов.
М.: Дрофа, 2005
2.Виды, особенности инфузорий..http://biofile.ru/bio/15750.html (дата обращения: 30.01.2018)
3.https://videouroki.net/videouroki/conspekty/bio7bespozv/7-tip-infuzorii-ili-riesnichnyie.files/image003.jpg
4.https://studfiles.net/html/2706/394/html_7m95jKZIP8.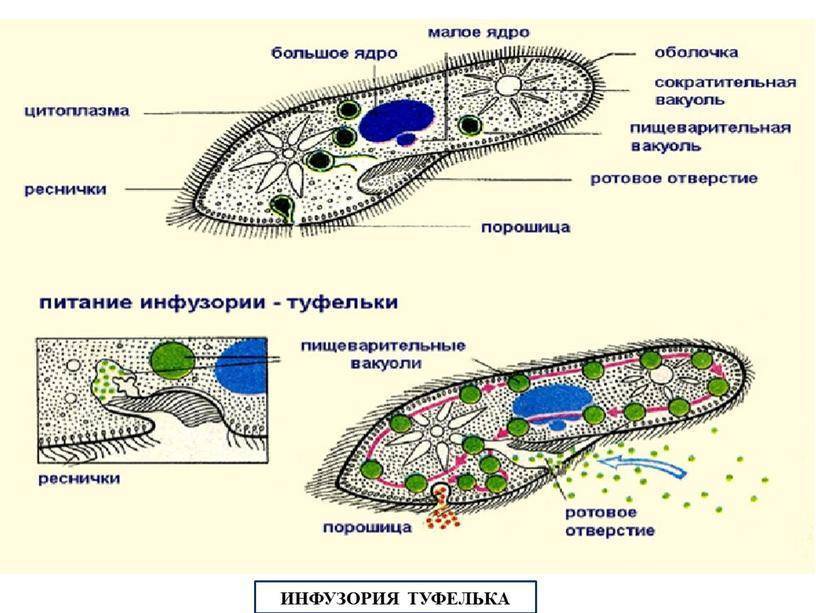 +mi7z#=nj C8,»pݠ4
aN((aGc_0!IJ5__UIvqQ_ս[m!&%m»9v7a81Ԋ E A0D[c@/~-z}-n/zm’In MTL$N.6y s9Na9m C)!GW ᐣoQW~mzOkҵnMfF9A@D#~8W\B`}IVx`t_’FJʃa;DA֗n?8G䆭ij_%5_i8ɸBDAG$A4ôU»znz[AA]l!!8Y6~__Z9!x»?K]%I}!#»»»»»![‘_nutZ[>w%Er~BA\WTHKcEsF90%#QIiKUvבNH»Ge !
odp璄Tе:
j}?{VuDcGinMEJȃa!t4&x0p?KuﯿKi{0M#m:ZӶ»»c{?mo/ҴK
%p9CDlGVIj-)*`[a’-gWui?-}xIo8Bۋ
7EJRC=aI:a5A44cO㤸l..[뭴4’_U[KTtAOwٌRe
Lr$L& 4 lBIUL0:$wJ}_鱗!8#,(X»=uX»Ј*$ئFvzKzj+KI_rtcZ/,s#EՑڋb4؈e8BA MH M
i8zSU
.W/ֿ{‘Pp?Hm!» `S A4064ҶؤzBPUk#98xt؉ AWPADDCIebblBXPi4ki0JV,utRn# 1P@T»F»»»»BR,pD}L ail%»E(بUJ=»?]l *1-Fj»»»»! `R7 a!!4m4n4GHF=q~GODM/
ga9@L
A0 MaS9;A菶ЊM07pq0S(sq9ð !Aqa?@B:»»»»»» .,As»>»H9I> za!qǸS#$1X;B #on #a_*4GJ0c}3\UqTG `$G[oW&INEwP@Ap{eL/]A!T1m#T.
I4qlN$y7 ،{HV#tEIA#фtQ»: I5q
1X0A-
F#T»»Dtos2BJH$¤DzC݄X1b) @mBG0″ GX ) S%A{lE)7Q`pD|0BˌAApH»H]B{tBBGFPY6aDtF»Dڄ&BPZքDfa43afKxDutCHK&A0r͇.KYaf]Jzbm6~HL&nD8g
\9#GEDaF P[Ȗntqi&pݠz 0)R|!da»Fэ{[ml*Da»pd\HHp ذA0BKPÛ\) B9UG;ˠ;:>D?tWWפh&M6’]CaMa5YR:0E0″>a%~-~*{j»۫MȐAh5″:r1;wf»{_~M+INީx,EzjuTBjW{^g;»AnA#!H~Wiߺ];ݽ*PB1Џ4 KmQTuj^Dt6″!8㴂ҏY_G^:y7Wdb (=A8^FH»?kai}*Ȯq#!28ע1ȣOB#Aw߱
Ua!ɸX|0N;C-9}7U*]Bq^F:#$#zդAt»oՄL}j\t.
+mi7z#=nj C8,»pݠ4
aN((aGc_0!IJ5__UIvqQ_ս[m!&%m»9v7a81Ԋ E A0D[c@/~-z}-n/zm’In MTL$N.6y s9Na9m C)!GW ᐣoQW~mzOkҵnMfF9A@D#~8W\B`}IVx`t_’FJʃa;DA֗n?8G䆭ij_%5_i8ɸBDAG$A4ôU»znz[AA]l!!8Y6~__Z9!x»?K]%I}!#»»»»»![‘_nutZ[>w%Er~BA\WTHKcEsF90%#QIiKUvבNH»Ge !
odp璄Tе:
j}?{VuDcGinMEJȃa!t4&x0p?KuﯿKi{0M#m:ZӶ»»c{?mo/ҴK
%p9CDlGVIj-)*`[a’-gWui?-}xIo8Bۋ
7EJRC=aI:a5A44cO㤸l..[뭴4’_U[KTtAOwٌRe
Lr$L& 4 lBIUL0:$wJ}_鱗!8#,(X»=uX»Ј*$ئFvzKzj+KI_rtcZ/,s#EՑڋb4؈e8BA MH M
i8zSU
.W/ֿ{‘Pp?Hm!» `S A4064ҶؤzBPUk#98xt؉ AWPADDCIebblBXPi4ki0JV,utRn# 1P@T»F»»»»BR,pD}L ail%»E(بUJ=»?]l *1-Fj»»»»! `R7 a!!4m4n4GHF=q~GODM/
ga9@L
A0 MaS9;A菶ЊM07pq0S(sq9ð !Aqa?@B:»»»»»» .,As»>»H9I> za!qǸS#$1X;B #on #a_*4GJ0c}3\UqTG `$G[oW&INEwP@Ap{eL/]A!T1m#T.
I4qlN$y7 ،{HV#tEIA#фtQ»: I5q
1X0A-
F#T»»Dtos2BJH$¤DzC݄X1b) @mBG0″ GX ) S%A{lE)7Q`pD|0BˌAApH»H]B{tBBGFPY6aDtF»Dڄ&BPZքDfa43afKxDutCHK&A0r͇.KYaf]Jzbm6~HL&nD8g
\9#GEDaF P[Ȗntqi&pݠz 0)R|!da»Fэ{[ml*Da»pd\HHp ذA0BKPÛ\) B9UG;ˠ;:>D?tWWפh&M6’]CaMa5YR:0E0″>a%~-~*{j»۫MȐAh5″:r1;wf»{_~M+INީx,EzjuTBjW{^g;»AnA#!H~Wiߺ];ݽ*PB1Џ4 KmQTuj^Dt6″!8㴂ҏY_G^:y7Wdb (=A8^FH»?kai}*Ȯq#!28ע1ȣOB#Aw߱
Ua!ɸX|0N;C-9}7U*]Bq^F:#$#zդAt»oՄL}j\t. L;bK؈8U%q#FҶJ!*
?&]tc#DDH
4(sXDr,rDNˢB.;#Hf/DtyDDDm nG#фaQ>]l2:0″»‘e8″ȏl)xG(Fa#qH$|GˣE|G0#9:#DDDD]G菗E:.#8ˢ;/h._ڒ5″:7E>].)
(菛>∈»膈y쎋;#8p5dxDtG菑:#}F2F6f|Dxp6″»»»»&Z#8:#:.|f
1-ԑ#tGdw|eX
xlUQ)GEьDR»» NB$tytGRA!ʴ25D}!.SX)b2¬Ѝt$I4RtDE&R:8E:*)L)BHUʉ!DDBB,`R?,h,!#YVRELDDDA:eqE4- $DDBKHIJFj&]»%Z!`#:)cDq.PP(AL,VNB»JB]KeG!5,N.`»EQ t$t#E %rh5H) HDK#B@EJtaB&]a»&A!3BB9B$
hI4GB’Љ;1KQYBbX2PLą4Ў_GIZ>B(rbGLi5″8DtGDv»jB,W;8DQ tGB%iDB$!!2RXGBUQ*ФTЂ%SB#»
L;bK؈8U%q#FҶJ!*
?&]tc#DDH
4(sXDr,rDNˢB.;#Hf/DtyDDDm nG#фaQ>]l2:0″»‘e8″ȏl)xG(Fa#qH$|GˣE|G0#9:#DDDD]G菗E:.#8ˢ;/h._ڒ5″:7E>].)
(菛>∈»膈y쎋;#8p5dxDtG菑:#}F2F6f|Dxp6″»»»»&Z#8:#:.|f
1-ԑ#tGdw|eX
xlUQ)GEьDR»» NB$tytGRA!ʴ25D}!.SX)b2¬Ѝt$I4RtDE&R:8E:*)L)BHUʉ!DDBB,`R?,h,!#YVRELDDDA:eqE4- $DDBKHIJFj&]»%Z!`#:)cDq.PP(AL,VNB»JB]KeG!5,N.`»EQ t$t#E %rh5H) HDK#B@EJtaB&]a»&A!3BB9B$
hI4GB’Љ;1KQYBbX2PLą4Ў_GIZ>B(rbGLi5″8DtGDv»jB,W;8DQ tGB%iDB$!!2RXGBUQ*ФTЂ%SB#»
$#D ZAH计ОT-q14HD%{S@h83Gc4&q%DgeYeyHci (@mD»*@R)AAp!#tuB»ihqDIq;*FS@HEC*q:$!d!
C)Iq»sQ0D!F!#e)\#I,&&e(DEM
hv#fHRH8t’_t#Q93&h»2:)ԦB%4>GTqD6F%»@3XA҆qEHhO&S)ՔPWRHL#E&PB_e
d}Qģ%G>3XI
BDzXUMb.[/ ڒLb B% H莌4$ttB2″V»»*]xT9U%QG~DR3Q`X]*Gɚʢ@»8y :p@JhDG`MXL!F(&GFhDA
4%ZD$t8:OB»%QCĺ#Z#Fd»RJFh5D# X)Y@PNQ`B$b#lK%emCy
GXrDH!
«@Љ?uB&hE)MָY؎GY!HփKhFhND’ЊThDt؎$t$t2u#~ʹ5!A$I(dKpE[uEt,!ʢBVV%d4GQ0G/Rtjd ԑt»#)y=6#٪7hӒ&R!YL)6tTo>%Ha%’ \%eU&S(($fπ$»PBUGB»BGMЩUBI>I&K»؎ t&R;0 AHDDK:%(R r82# HGb»%BRmchYpfh0:(*GTj_0S0TqA4″-L#mG\/T*»nSaNn2C[eOiY E=D[>,XʆGLtKm)0F퐵?ЉқI6Er[G~5^fmsz/N];W+$~S%(BZhSm]X)cή,{q-:UXuQ48DǺB)L147g3V»avGA݈»:0nu E:cr?C1ͪ[lԊt»dJ)dìRvSA8tAl6R+G#p@u=jHYśI !JS h5uO6SZD7}Z݊_i#Dy{di:]E;_˭I! Ȳ%aC
^K}Pt#)4Am$$u₰NE!UMmmi»L2P). >)Cm׳Si]Vڶ7[b1(«(xxi[Bs߰R᭤U]E%OƗEluOJ+a&aFtavE|I.}pI1>8j?hG\2I/zCۄa;ڰU-@\xdq 3iXHi GT]>=+iizIX&8AB_ L%$)*[[\{*{82$C8(aAB3
>)Cm׳Si]Vڶ7[b1(«(xxi[Bs߰R᭤U]E%OƗEluOJ+a&aFtavE|I.}pI1>8j?hG\2I/zCۄa;ڰU-@\xdq 3iXHi GT]>=+iizIX&8AB_ L%$)*[[\{*{82$C8(aAB3
Относительная пищевая ценность реснитчатых простейших и водорослей в качестве корма для дафний
Относительное значение автотрофных жгутиконосцев, десмидий, цианобактерий и инфузорий в качестве пищи для Daphnia magna изучалось с помощью групповых таблиц выживаемости. Каждой когорте давали один тип пищи в заданной концентрации, и между каждым типом проводились сравнения. Кормление водорослями включало три уровня молодых (в возрасте от 7 до 14 дней) Chlamydomonas reinhardi (Chlorophyta, Chlamydomonadacae) и два уровня стареющих (> 14 дней) C.reinhardi, два уровня Cryptomonas sp. (Chlorophyta, Cryptomonadacae), два уровня Staurastrum sp. (Chlorophyta, Desmidacae), четыре уровня молодых (в возрасте от 7 до 15 дней) или стареющих (в возрасте > 15 дней) Microcystis aeruginosa (Cyanophyta, Chlorococcacae) и лечение без пищи.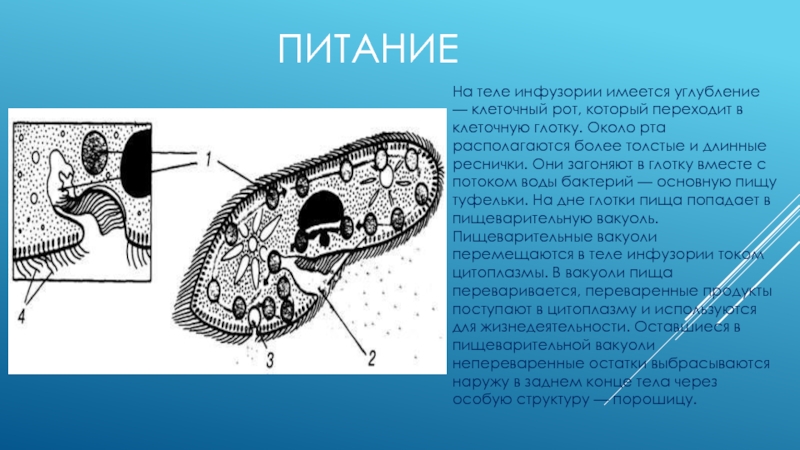 Инфузории Cyclidium sp. и Paramecium caudatum также были представлены в концентрациях 1 или 10(2) клеток/мл, а также смеси C. reinhardi (10(3)/мл) и Cyclidium (1/мл) или P. caudatum (1/мл). Рост, размножение и выживаемость дафний были самыми высокими, когда C.reinhardi или Cryptomonas были источником пищи, в то время как голодающие или кормившиеся M. aeruginosa имели более короткую выживаемость и более низкий рост и размножение. Daphnia росла и имела высокую выживаемость при кормлении P. caudatum, но, несмотря на то, что яйца были произведены, большинство из них были абортированы через 2 или 3 дня. Staurastrum и Cyclidium давали промежуточный рост и выживаемость, но размножение наблюдалось только при обработке 10(3) Staurastrum/мл. Содержание углерода и азота было общим показателем пищевой ценности. Тем не менее, рост, размножение и выживаемость были выше в некоторых когортах, получавших лечение, содержащее относительно низкие уровни углерода и азота.Другие когорты жили недолго и не размножались, несмотря на то, что их кормили гораздо более высоким уровнем углерода и азота.
Инфузории Cyclidium sp. и Paramecium caudatum также были представлены в концентрациях 1 или 10(2) клеток/мл, а также смеси C. reinhardi (10(3)/мл) и Cyclidium (1/мл) или P. caudatum (1/мл). Рост, размножение и выживаемость дафний были самыми высокими, когда C.reinhardi или Cryptomonas были источником пищи, в то время как голодающие или кормившиеся M. aeruginosa имели более короткую выживаемость и более низкий рост и размножение. Daphnia росла и имела высокую выживаемость при кормлении P. caudatum, но, несмотря на то, что яйца были произведены, большинство из них были абортированы через 2 или 3 дня. Staurastrum и Cyclidium давали промежуточный рост и выживаемость, но размножение наблюдалось только при обработке 10(3) Staurastrum/мл. Содержание углерода и азота было общим показателем пищевой ценности. Тем не менее, рост, размножение и выживаемость были выше в некоторых когортах, получавших лечение, содержащее относительно низкие уровни углерода и азота.Другие когорты жили недолго и не размножались, несмотря на то, что их кормили гораздо более высоким уровнем углерода и азота.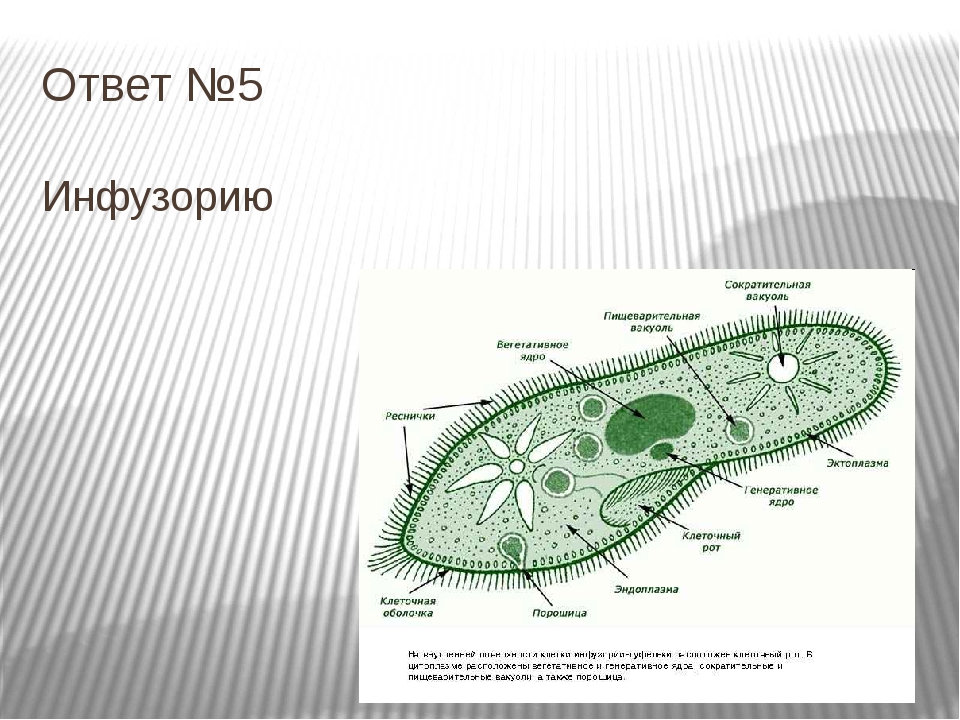 Результаты также свидетельствуют о том, что зеленые водоросли имеют питательную ценность для дафний, тогда как цианобактерии — нет. Пищевая ценность инфузорий, измеренная по параметрам таблицы жизни, была переменной, причем некоторые из них были плохими источниками пищи. Таким образом, потенциал инфузорий как трофического звена между микробной продукцией и более высокими трофическими уровнями может варьироваться в зависимости от структуры сообщества инфузорий. Наши результаты показывают, что одних только инфузорий в качестве источника пищи недостаточно для поддержки роста популяции дафний.
Результаты также свидетельствуют о том, что зеленые водоросли имеют питательную ценность для дафний, тогда как цианобактерии — нет. Пищевая ценность инфузорий, измеренная по параметрам таблицы жизни, была переменной, причем некоторые из них были плохими источниками пищи. Таким образом, потенциал инфузорий как трофического звена между микробной продукцией и более высокими трофическими уровнями может варьироваться в зависимости от структуры сообщества инфузорий. Наши результаты показывают, что одних только инфузорий в качестве источника пищи недостаточно для поддержки роста популяции дафний.
Относительная пищевая ценность инфузорий простейших и водорослей в качестве пищи для дафний на JSTOR
Абстрактный Относительная важность автотрофных жгутиконосцев, десмидий, цианобактерий и инфузорий в качестве пищи для Daphnia magna была изучена с использованием когортных таблиц выживаемости. Каждой когорте давали один тип пищи в заданной концентрации, и между каждым типом проводились сравнения. Кормление водорослями включало три уровня молодых (возраст от 7 до 14 дней) Chlamydomonas reinhardi (Chlorophyta, Chlamydomonadacae), два уровня стареющих (старше 14 дней) C.reinhardi, два уровня Cryptomonas sp. (Chlorophyta, Cryptomonadacae), два уровня Staurastrum sp. (Chlorophyta, Desmidacae), четыре уровня молодых (в возрасте от 7 до 15 дней) или стареющих (в возрасте > 15 дней) Microcystis aeruginosa (Cyanophyta, Chlorococcacae) и лечение без пищи. Инфузории Cyclidium sp. и Paramecium caudatum также были представлены в концентрациях 1 или 102 клеток/мл, а также смеси C. reinhardi (103/мл) и Cyclidium (1/мл) или P. caudatum (1/мл). Рост, размножение и выживаемость дафний были самыми высокими, когда C.reinhardi или Cryptomonas были источником пищи, в то время как те, кто голодал или кормился M. aeruginosa, имели более короткую выживаемость и более низкий рост и размножение. Дафния росла и имела высокую выживаемость при кормлении P. caudatum, но, несмотря на то, что яйца были произведены, большинство из них были абортированы через 2 или 3 дня.
Кормление водорослями включало три уровня молодых (возраст от 7 до 14 дней) Chlamydomonas reinhardi (Chlorophyta, Chlamydomonadacae), два уровня стареющих (старше 14 дней) C.reinhardi, два уровня Cryptomonas sp. (Chlorophyta, Cryptomonadacae), два уровня Staurastrum sp. (Chlorophyta, Desmidacae), четыре уровня молодых (в возрасте от 7 до 15 дней) или стареющих (в возрасте > 15 дней) Microcystis aeruginosa (Cyanophyta, Chlorococcacae) и лечение без пищи. Инфузории Cyclidium sp. и Paramecium caudatum также были представлены в концентрациях 1 или 102 клеток/мл, а также смеси C. reinhardi (103/мл) и Cyclidium (1/мл) или P. caudatum (1/мл). Рост, размножение и выживаемость дафний были самыми высокими, когда C.reinhardi или Cryptomonas были источником пищи, в то время как те, кто голодал или кормился M. aeruginosa, имели более короткую выживаемость и более низкий рост и размножение. Дафния росла и имела высокую выживаемость при кормлении P. caudatum, но, несмотря на то, что яйца были произведены, большинство из них были абортированы через 2 или 3 дня. Staurastrum и Cyclidium давали промежуточный рост и выживаемость, но размножение наблюдалось только при обработке 103 Staurastrum/мл. Содержание углерода и азота было общим показателем пищевой ценности. Тем не менее, рост, размножение и выживаемость были выше в некоторых когортах, получавших лечение, содержащее относительно низкие уровни углерода и азота.Другие когорты жили недолго и не размножались, несмотря на то, что их кормили гораздо более высоким уровнем углерода и азота. Результаты также показывают, что зеленые водоросли имеют питательную ценность для дафний, тогда как цианобактерии — нет. Пищевая ценность инфузорий, измеренная по параметрам таблицы жизни, была переменной, причем некоторые из них были плохими источниками пищи. Таким образом, потенциал инфузорий как трофического звена между микробной продукцией и более высокими трофическими уровнями может варьироваться в зависимости от структуры сообщества инфузорий.Наши результаты показывают, что одних только инфузорий было недостаточно в качестве источника пищи для поддержания роста популяции дафний.
Staurastrum и Cyclidium давали промежуточный рост и выживаемость, но размножение наблюдалось только при обработке 103 Staurastrum/мл. Содержание углерода и азота было общим показателем пищевой ценности. Тем не менее, рост, размножение и выживаемость были выше в некоторых когортах, получавших лечение, содержащее относительно низкие уровни углерода и азота.Другие когорты жили недолго и не размножались, несмотря на то, что их кормили гораздо более высоким уровнем углерода и азота. Результаты также показывают, что зеленые водоросли имеют питательную ценность для дафний, тогда как цианобактерии — нет. Пищевая ценность инфузорий, измеренная по параметрам таблицы жизни, была переменной, причем некоторые из них были плохими источниками пищи. Таким образом, потенциал инфузорий как трофического звена между микробной продукцией и более высокими трофическими уровнями может варьироваться в зависимости от структуры сообщества инфузорий.Наши результаты показывают, что одних только инфузорий было недостаточно в качестве источника пищи для поддержания роста популяции дафний.
Микробная экология лежит в основе функционирования почти каждой экосистемы на планете, от глубоководных жерл и подземных систем до благополучия людей и животных; от нетронутой морской и наземной среды до функционирования промышленных биореакторов.
Экология микробов представляет собой специализированный международный форум для презентации высококачественных научных исследований того, как микроорганизмы взаимодействуют с биотической и абиотической средой, друг с другом, а также со своими соседями и хозяевами для выполнения своих разнообразных функций.Microbial Ecology предлагает статьи об оригинальных исследованиях в формате полных статей и заметок, а также краткие обзоры и тематические документы с изложением позиции.
Охват включает экологию микроорганизмов в естественных и искусственных средах; геномные и молекулярные достижения в понимании микробных взаимодействий и филогении; микробные драйверы биогеохимических процессов; меж- и внутривидовая микробная коммуникация; экологические исследования микробиологии и болезней животных, растений и насекомых; микробные процессы и взаимодействия в экстремальных или необычных условиях; микробная популяция и экология сообщества и многое другое.
Springer — одно из ведущих международных научных издательств, выпускающее более 1200 журналов и более 3000 новых книг ежегодно, охватывающих широкий круг предметов, включая биомедицину и науки о жизни, клиническую медицину, физика, инженерия, математика, информатика и экономика.
Инфузории в планктонных пищевых сетях: общение и адаптивный ответ
Монофилум Oxytrichidae (Ciliophora, Hypotrichia) включает пресноводные, морские и почвенные виды.Некоторые таксоны чрезвычайно широко распространены в пресноводных (например, Oxytricha, Stylonychia) или наземных местообитаниях (например, Gonostomum). Oxytricha и Stylonychia использовались для многих молекулярных, ультраструктурных и генетических исследований.
Это первая книга после монументальной работы Эренберга (1838 г.), в которой обобщены морфологические, морфогенетические, фаунистические и экологические данные за последние 220 лет, разбросанные по более чем 2500 ссылкам со всего мира. Один центральный дихотомический ключ ведет непосредственно к 32 родам, включающим около 169 видов, которые проиллюстрированы более чем 2400 рисунками, включая оригинальные.Таксономисты, экологи, лимнологи, почвоведы и клеточные биологи найдут список синонимов, обсуждение номенклатурных и таксономических проблем, подробное морфологическое и экологическое описание, включая почти все опубликованные фаунистические записи. Это позволяет как специалисту, так и практикующему врачу определять виды, не обращаясь к оригинальной литературе. Книга предлагает заинтересованному ученому тщательный и современный обзор окситрихид. Рецензия на книгу SONG Weibo (2000): Монография Oxytrichidae (Ciliophora, Hypotrichia).Х. Бергер. Опубликовано Kluwer Academic Publishers (1999). NLG880.00/доллар США528. стр. 1079. ISBN: 0-7923-5795-7. — Микробиология сегодня, август 2000 г.
Oxytrichidae — богатое видами, хорошо известное, но весьма запутанное семейство инфузорий — всегда привлекало интерес протозоологов, работающих в области экологии, таксономии и клеточной биологии.
Один центральный дихотомический ключ ведет непосредственно к 32 родам, включающим около 169 видов, которые проиллюстрированы более чем 2400 рисунками, включая оригинальные.Таксономисты, экологи, лимнологи, почвоведы и клеточные биологи найдут список синонимов, обсуждение номенклатурных и таксономических проблем, подробное морфологическое и экологическое описание, включая почти все опубликованные фаунистические записи. Это позволяет как специалисту, так и практикующему врачу определять виды, не обращаясь к оригинальной литературе. Книга предлагает заинтересованному ученому тщательный и современный обзор окситрихид. Рецензия на книгу SONG Weibo (2000): Монография Oxytrichidae (Ciliophora, Hypotrichia).Х. Бергер. Опубликовано Kluwer Academic Publishers (1999). NLG880.00/доллар США528. стр. 1079. ISBN: 0-7923-5795-7. — Микробиология сегодня, август 2000 г.
Oxytrichidae — богатое видами, хорошо известное, но весьма запутанное семейство инфузорий — всегда привлекало интерес протозоологов, работающих в области экологии, таксономии и клеточной биологии.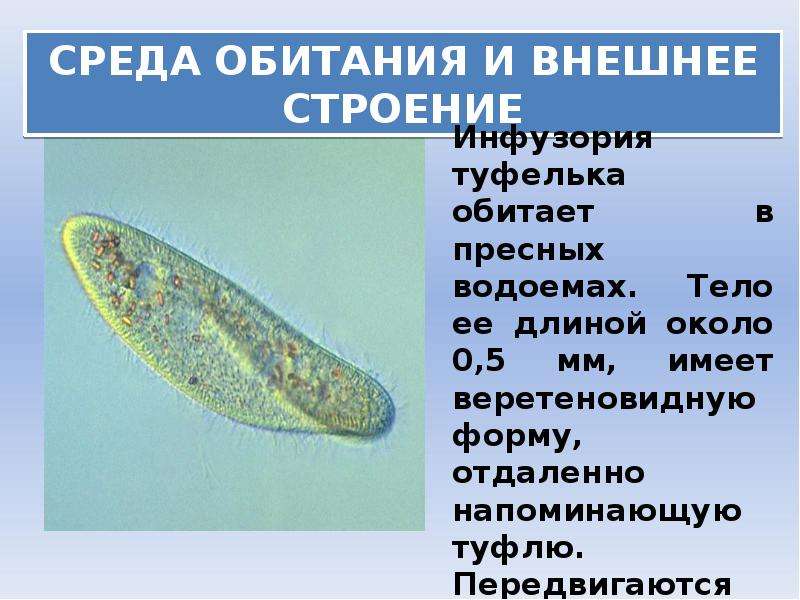 Эта актуальная книга содержит исчерпывающую информацию как об исторических, так и о недавних исследованиях, включая тщательно определенный список синонимов для каждого вида, обсуждение (и исправления) номенклатурных проблем, подробные морфологические и таксономические описания почти всех опубликованных фаунистических записей, а также в виде таблиц, содержащих полные доступные морфометрические данные.В качестве еще одной заметной особенности упоминается почти вся экологическая, физиологическая и фаунистическая информация по каждому виду, разбросанная в литературе за последние 300 лет, что позволяет пользователю определить вид, не обращаясь к исходной литературе. Таким образом, эта книга является не только суперполевым справочником, но и незаменимым справочником как для начинающих, так и для опытных систематиков, и должна быть на полках каждой цилиатологической лаборатории.
Weibo SONG, Музей естественной истории, Лондон Ключевые слова: Африка, Allotricha, Alveolata, Америка, Ancystropodium, Apoamphisiella, Азия, Австралия, Australocirrus, Австрия, Bestimmungsschlüssel, библиография, биоразнообразие, Biodiversität, биогеография, Biogeographie, биология, Bodenbiologie, Boden, клеточное деление, Китай, инфузории, Ciliophora, кладистика , клон, Coniculostomum, конъюгация, сократительная вакуоль, кортикальные гранулы, культивирование, Cyrtohymena, киста, цитоплазма, детерминация, Diversität, разнообразие, днк, экология, Эренберг, Европа, Европа, эволюция, Gastrostyla, Gonostomum, направляющая, Hemigastrostyla, Histriculus, Hypotricha , hypotriche, Hypotrichia, Hypotrichida, гипотрихи, идентификация, Идентификация, пропитка, Каль, Керона, ключ, Клон, Клювер, Лаурентелла, жизненный цикл, лимнетик, Limnologie, лимнология, морская биология, микробиология, молекулярная биология, Морфогенез, морфогенез, движение, номенклатура, Номенклатура, Notohymena, питание, Onychodromopsis, Onychodromus, Oxytricha, окситрихиды, Oxytrichinae, Paramecium, Para stylonychia, Paraurostyla, Parentocirrus, Parurosoma, Pattersonella, Phylogenie, филогенез, Plankton, Pleurotricha, Protista, Protozoa, Pseudostrombidium, Pseudouroleptus, покоящаяся киста, обзор, ревизия, Rigidocortex, rna, Rubrioxytricha, Salzburg, море, см.
Эта актуальная книга содержит исчерпывающую информацию как об исторических, так и о недавних исследованиях, включая тщательно определенный список синонимов для каждого вида, обсуждение (и исправления) номенклатурных проблем, подробные морфологические и таксономические описания почти всех опубликованных фаунистических записей, а также в виде таблиц, содержащих полные доступные морфометрические данные.В качестве еще одной заметной особенности упоминается почти вся экологическая, физиологическая и фаунистическая информация по каждому виду, разбросанная в литературе за последние 300 лет, что позволяет пользователю определить вид, не обращаясь к исходной литературе. Таким образом, эта книга является не только суперполевым справочником, но и незаменимым справочником как для начинающих, так и для опытных систематиков, и должна быть на полках каждой цилиатологической лаборатории.
Weibo SONG, Музей естественной истории, Лондон Ключевые слова: Африка, Allotricha, Alveolata, Америка, Ancystropodium, Apoamphisiella, Азия, Австралия, Australocirrus, Австрия, Bestimmungsschlüssel, библиография, биоразнообразие, Biodiversität, биогеография, Biogeographie, биология, Bodenbiologie, Boden, клеточное деление, Китай, инфузории, Ciliophora, кладистика , клон, Coniculostomum, конъюгация, сократительная вакуоль, кортикальные гранулы, культивирование, Cyrtohymena, киста, цитоплазма, детерминация, Diversität, разнообразие, днк, экология, Эренберг, Европа, Европа, эволюция, Gastrostyla, Gonostomum, направляющая, Hemigastrostyla, Histriculus, Hypotricha , hypotriche, Hypotrichia, Hypotrichida, гипотрихи, идентификация, Идентификация, пропитка, Каль, Керона, ключ, Клон, Клювер, Лаурентелла, жизненный цикл, лимнетик, Limnologie, лимнология, морская биология, микробиология, молекулярная биология, Морфогенез, морфогенез, движение, номенклатура, Номенклатура, Notohymena, питание, Onychodromopsis, Onychodromus, Oxytricha, окситрихиды, Oxytrichinae, Paramecium, Para stylonychia, Paraurostyla, Parentocirrus, Parurosoma, Pattersonella, Phylogenie, филогенез, Plankton, Pleurotricha, Protista, Protozoa, Pseudostrombidium, Pseudouroleptus, покоящаяся киста, обзор, ревизия, Rigidocortex, rna, Rubrioxytricha, Salzburg, море, см. , почва, Spirotricha, Spirotrichea , ssrdna, Stein, Steinia, Sterkiella, Stichotrichia, стихотрихи, Stylonychia, Stylonychinae, стилонихиды, систематика, Systematik, Tachysoma, Taxonomie, таксономия, Teilung, Territricha, Tetrahymena, Trichoda, Urosoma, Urosomoida, Wimpertierchen, Ziliaten
, почва, Spirotricha, Spirotrichea , ssrdna, Stein, Steinia, Sterkiella, Stichotrichia, стихотрихи, Stylonychia, Stylonychinae, стилонихиды, систематика, Systematik, Tachysoma, Taxonomie, таксономия, Teilung, Territricha, Tetrahymena, Trichoda, Urosoma, Urosomoida, Wimpertierchen, Ziliaten
Еда для всехОдно из самых важных растений в освещенной солнцем зоне и самое маленькое. Фитопланктон — это организмы, которые плавают на поверхности воды или вблизи нее. Большинство из них округлые и одноклеточные. Весь фитопланктон использует фотосинтез для получения энергии, но некоторые получают дополнительную энергию, потребляя другие организмы. Наиболее распространенным фитопланктоном являются диатомовые водоросли и динофлагелляты . Диатомеи – одноклеточные водоросли. Они часто объединяются в длинные цепочки. Динофлагелляты представляют собой небольшие организмы с двумя хвостами или жгутиками. Phaeophyta или бурые водоросли являются еще одним видом фитопланктона. Сине-зеленые водоросли или цианобактерии тоже относятся к фитопланктону, хотя они весьма уникальны. Они фотосинтезируют, но некоторые также используют азот для получения энергии. Это азотфиксаторы .Они превращают свободный азот в нитраты, которые используются цианобактериями и другими растениями в океане. Фитопланктон является основой пищевой сети океана и источником пищи для зоопланктона. Плыть по течениюЗоопланктон — это океанические животные, которые вообще не умеют плавать или плавают очень слабо, и они дрейфуют или движутся вместе с океанскими течениями. Их можно встретить в освещенной солнцем зоне и в глубоких океанских водах. Размер зоопланктона варьируется от крошечных микробов до медуз, хотя большинство зоопланктона представляют собой крошечные одноклеточные организмы.Различают два вида зоопланктона. Постоянный голопланктон или всегда будет зоопланктоном. Временный меропланктон или состоит из личинок рыб, ракообразных и других морских животных. Если они выживут, то вырастут в нектона или в свободно плавающие организмы. Их можно встретить в освещенной солнцем зоне и в глубоких океанских водах. Размер зоопланктона варьируется от крошечных микробов до медуз, хотя большинство зоопланктона представляют собой крошечные одноклеточные организмы.Различают два вида зоопланктона. Постоянный голопланктон или всегда будет зоопланктоном. Временный меропланктон или состоит из личинок рыб, ракообразных и других морских животных. Если они выживут, то вырастут в нектона или в свободно плавающие организмы. Есть место для ростаФораминиферы — крошечные одноклеточные, покрытые панцирем организмы, обычно диаметром от миллиметра до сантиметра. По мере роста они добавляют камеры в свои раковины.В зависимости от вида раковина может состоять из песка, кальцита или органических веществ. Они передвигаются и ловят пищу с помощью тонких волосовидных отростков, называемых псевдоподиями. Когда фораминиферы умирают, их раковины опускаются на дно океана и образуют ил.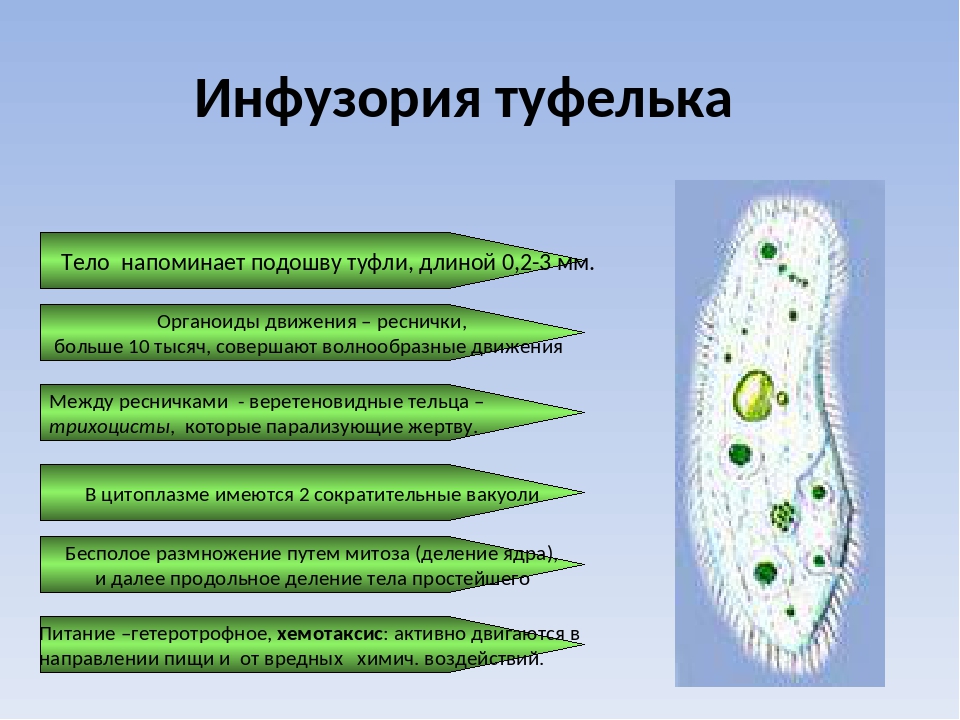 Подсчитано, что 30 процентов дна океана состоит из раковин фораминифер. И известняк, и мел происходят из фораминифер! Подсчитано, что 30 процентов дна океана состоит из раковин фораминифер. И известняк, и мел происходят из фораминифер!Как каменьРадиолярии — маленькие, круглые, покрытые панцирем организмы.Они делают свои раковины из кремнезема. Кремнезем используется в производстве стекла и может быть найден в таких минералах, как кварц. Они получают кремнезем из океана. Радиолярии имеют длинные, липкие, похожие на щупальца руки, называемые псевдоподиями . Они высовывают свои псевдоподии из отверстий в панцирях, чтобы ловить проплывающий мимо фитопланктон. Когда радиолярии умирают, их раковины опускаются на дно океана. Со временем, если достаточное количество раковин утонет вместе, останки скелета могут превратиться в осадочную породу! | Волосатые движенияИнфузории имеют реснички или маленькие волосовидные отростки по всему телу, которые они используют для передвижения и ловли пищи. Существует более 8000 видов инфузорий, в том числе парамеции. Живут в соленой и пресной воде. Некоторые свободно плавают, другие прикрепляются к организмам или объектам, а некоторые являются паразитами. Только свободно плавающие считаются зоопланктоном! Существует более 8000 видов инфузорий, в том числе парамеции. Живут в соленой и пресной воде. Некоторые свободно плавают, другие прикрепляются к организмам или объектам, а некоторые являются паразитами. Только свободно плавающие считаются зоопланктоном!Размахивая флагомЗоофлагелляты , как и динофлагелляты, имеют длинные жгутики. Они либо поглощают свою пищу, либо поглощают ее пищевыми вакуолями или карманами. Они живут в самых разных условиях.Некоторые из них являются паразитами и могут быть обнаружены в пищеварительном тракте животных, таких как таракан и термит; другие свободно плавают. Свободноплавающие – это зоопланктон.Желе БеллиМедузы также являются зоопланктоном. Медузы — это в основном большие желудки и длинные щупальца! На их щупальцах есть жала, и они используют их, чтобы ловить и парализовать пищу и переносить ее в желудок. Они передвигаются в воде, накачивая свои желудки.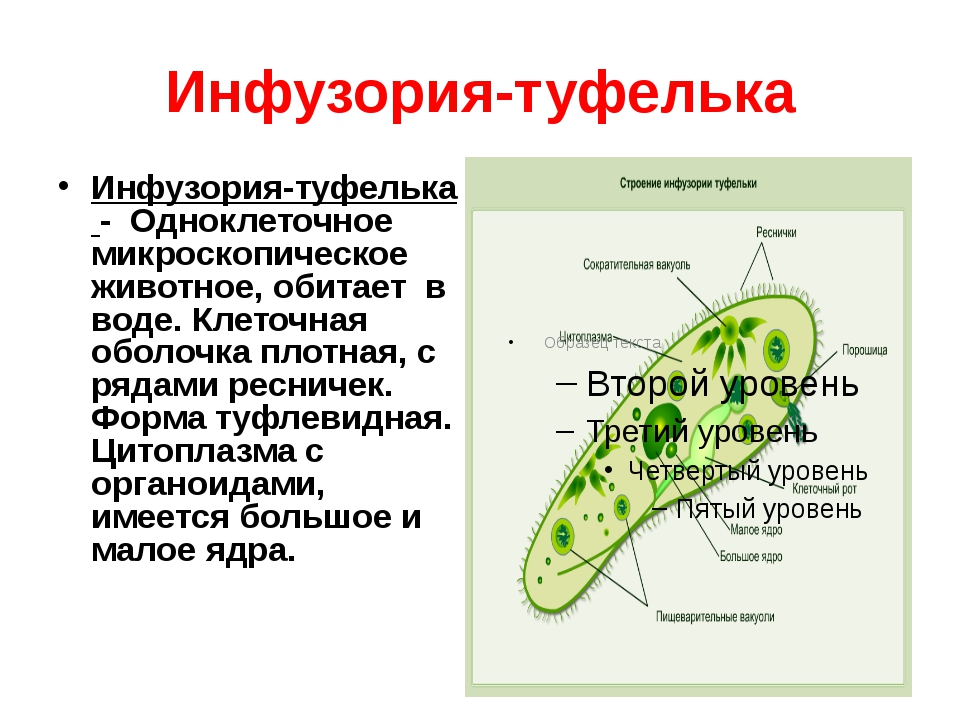 В основном они двигаются вверх и вниз в воде и позволяют течениям нести их из стороны в сторону. В основном они двигаются вверх и вниз в воде и позволяют течениям нести их из стороны в сторону.Чем больше, тем веселееСифонофоры , как у португальского военного корабля, выглядят как медузы, но это не так. На самом деле это группы или колонии животных. У каждого организма в колонии есть особая ниша или роль. Некоторые образуют щупальца, а некоторые образуют рот и желудок.БаггиВеслоногих рачков иногда называют морскими насекомыми, потому что их так много — около 10 000 видов! Их можно найти в пресной и соленой воде.Копеподы очень маленькие, обычно не более нескольких миллиметров в длину. Самая крупная копепода, Pennella balaenopterae, , живет на спинном ките и может вырасти до фута в длину! Веслоногие ракообразные. Они имеют два усика, раковину и сегментированное тело. Питаются фитопланктоном и зоопланктоном. Веслоногие раки — крупнейший источник белка в океане!Хорошей еды!Криль , одно из самых маленьких морских животных, является обедом для одного из его самых больших китов! Существует около 82 видов криля размером от менее четверти дюйма до двух дюймов в длину.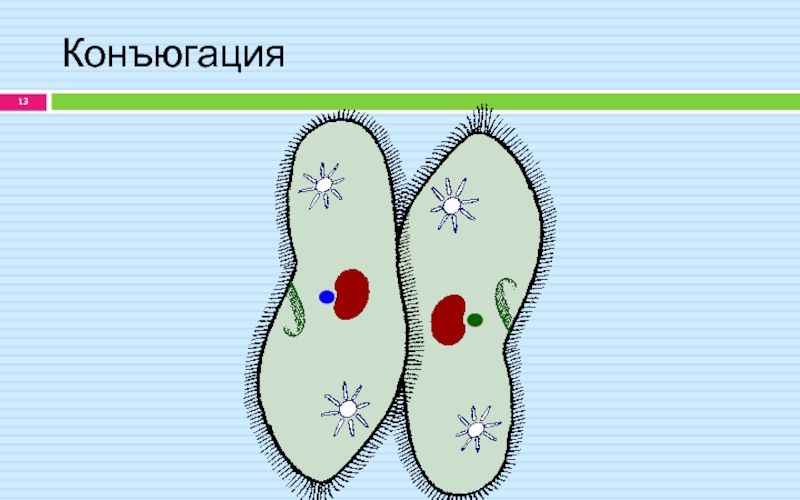 Криль — это ракообразные, похожие на веслоногих. У них часто есть биолюминесцентные органы. Их можно встретить в освещенной солнцем зоне и в сумеречной зоне. Криль — это ракообразные, похожие на веслоногих. У них часто есть биолюминесцентные органы. Их можно встретить в освещенной солнцем зоне и в сумеречной зоне. Изображения CC BY 2.0 CC BY 3.0 CC BY-SA 3.0. |
 Динофлагелляты бывают самых разных форм и размеров.У кого-то есть раковины, у кого-то нет. Не все динофлагелляты полагаются только на фотосинтез всей своей энергии. Некоторые оборачивают себя вокруг еды и поглощают ее. Некоторые динофлагелляты могут излучать свет с помощью биолюминесценции.
Динофлагелляты бывают самых разных форм и размеров.У кого-то есть раковины, у кого-то нет. Не все динофлагелляты полагаются только на фотосинтез всей своей энергии. Некоторые оборачивают себя вокруг еды и поглощают ее. Некоторые динофлагелляты могут излучать свет с помощью биолюминесценции. Плавание инфузорий при геометрических ограничениях: Journal of Applied Physics: Vol 125, No 20
В этом разделе мы исследуем силы смазки, действующие на клетку вблизи жесткой стенки, твердой сферы и свободной поверхности. , чтобы лучше понять поведение инфузорий в последующих разделах.
A. Модель сквирмера
Модель, которую мы здесь используем, является самой простой, в которой сквирмер плавает и имеет конечный размер, так что объемные эффекты и гидродинамические взаимодействия могут быть проанализированы нетривиально. 39 39. Т. Исикава, М. П. Симмондс и Т. Дж. Педли, «Гидродинамическое взаимодействие двух моделей микроорганизмов для плавания», J. Fluid Mech. 568 , 119–160 (2006). https://doi.org/10.1017/S0022112006002631 Мы предполагаем, что модель инфузории имеет сферическую форму с радиусом a .Он имеет нейтральную плавучесть и не является броуновским, а окружающее поле течения можно рассматривать как течение Стокса. Предполагается, что зигзагообразный двигается, создавая поверхностные извивающиеся скорости. Эта модель была впервые предложена Лайтхиллом 40 40. М. Дж. Лайтхилл, «О колебательном движении почти сферических деформируемых тел в жидкости при очень малых числах Рейнольдса», Комм. Чистое приложение Мат. 5 , 109–118 (1952). https://doi.org/10.1002/cpa.3160050201 и расширено Блейком. 41 41. Дж. Р. Блейк, «Сферический подход оболочки к цилиарному движению», J. Fluid Mech. 46 , 199–208 (1971). https://doi.org/10.1017/S002211207100048X Модель изначально была предложена для инфузорий и микроводорослей, таких как Opalina и Volvox ; 42,43 42.
Fluid Mech. 568 , 119–160 (2006). https://doi.org/10.1017/S0022112006002631 Мы предполагаем, что модель инфузории имеет сферическую форму с радиусом a .Он имеет нейтральную плавучесть и не является броуновским, а окружающее поле течения можно рассматривать как течение Стокса. Предполагается, что зигзагообразный двигается, создавая поверхностные извивающиеся скорости. Эта модель была впервые предложена Лайтхиллом 40 40. М. Дж. Лайтхилл, «О колебательном движении почти сферических деформируемых тел в жидкости при очень малых числах Рейнольдса», Комм. Чистое приложение Мат. 5 , 109–118 (1952). https://doi.org/10.1002/cpa.3160050201 и расширено Блейком. 41 41. Дж. Р. Блейк, «Сферический подход оболочки к цилиарному движению», J. Fluid Mech. 46 , 199–208 (1971). https://doi.org/10.1017/S002211207100048X Модель изначально была предложена для инфузорий и микроводорослей, таких как Opalina и Volvox ; 42,43 42. Т. Дж. Педли, «Сферические сквирмеры: модели плавающих микроорганизмов», IMA J. Appl. Мат. 81 , 488–521 (2016). https://doi.org/10.1093/imamat/hxw03043. Т. Дж. Педли, Д.Р. Брамли и Р. Э. Гольдштейн, «Сквирмеры с вихрем: модель плавания вольвоксов», J. Fluid Mech. 798 , 165–186 (2016). https://doi.org/10.1017/jfm.2016.306 однако он также использовался для моделирования частиц Януса 44 44. А. Вюргер, «Самодиффузиофорез частиц Януса в околокритических смесях», Phys. Преподобный Летт. 115 , 188304 (2015). https://doi.org/10.1103/PhysRevLett.115.188304 и самодвижущиеся капли жидкости. 45 45. С. Тутупалли, Р.Зееманн и С. Хермингхаус, «Роевое поведение простых модельных сквирмеров», New J. Phys. 13 , 073021 (2011). https://doi.org/10.1088/1367-2630/13/7/073021 Мы предполагаем, что поверхность сферы движется чисто тангенциально по отношению к телу клетки и что тангенциальное движение осесимметрично и не зависит от времени. Тангенциальная поверхностная скорость u с определяется выражением
Т. Дж. Педли, «Сферические сквирмеры: модели плавающих микроорганизмов», IMA J. Appl. Мат. 81 , 488–521 (2016). https://doi.org/10.1093/imamat/hxw03043. Т. Дж. Педли, Д.Р. Брамли и Р. Э. Гольдштейн, «Сквирмеры с вихрем: модель плавания вольвоксов», J. Fluid Mech. 798 , 165–186 (2016). https://doi.org/10.1017/jfm.2016.306 однако он также использовался для моделирования частиц Януса 44 44. А. Вюргер, «Самодиффузиофорез частиц Януса в околокритических смесях», Phys. Преподобный Летт. 115 , 188304 (2015). https://doi.org/10.1103/PhysRevLett.115.188304 и самодвижущиеся капли жидкости. 45 45. С. Тутупалли, Р.Зееманн и С. Хермингхаус, «Роевое поведение простых модельных сквирмеров», New J. Phys. 13 , 073021 (2011). https://doi.org/10.1088/1367-2630/13/7/073021 Мы предполагаем, что поверхность сферы движется чисто тангенциально по отношению к телу клетки и что тангенциальное движение осесимметрично и не зависит от времени. Тангенциальная поверхностная скорость u с определяется выражением| us(θ)=32U0sinθ+34βsin(2θ), | (1) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Рис. 1(а)], U 0 — скорость плавания одиночного извильщика в бесконечной жидкости, β — сила извивающейся второй моды. Здесь мы рассматриваем извивающиеся моды до второй моды, так как первая мода определяет скорость плавания, а вторая мода является ведущим членом при рассмотрении гидродинамических взаимодействий. Сквирмер с положительным (отрицательным) β является тянущим (толкающим), аналогом клетки, у которой тяга создается в основном впереди (за) телом.Сквирмер с β = 0 является нейтральным пловцом, аналогом клетки, у которой центры тяги и сопротивления совпадают.
Рис. 1(а)], U 0 — скорость плавания одиночного извильщика в бесконечной жидкости, β — сила извивающейся второй моды. Здесь мы рассматриваем извивающиеся моды до второй моды, так как первая мода определяет скорость плавания, а вторая мода является ведущим членом при рассмотрении гидродинамических взаимодействий. Сквирмер с положительным (отрицательным) β является тянущим (толкающим), аналогом клетки, у которой тяга создается в основном впереди (за) телом.Сквирмер с β = 0 является нейтральным пловцом, аналогом клетки, у которой центры тяги и сопротивления совпадают.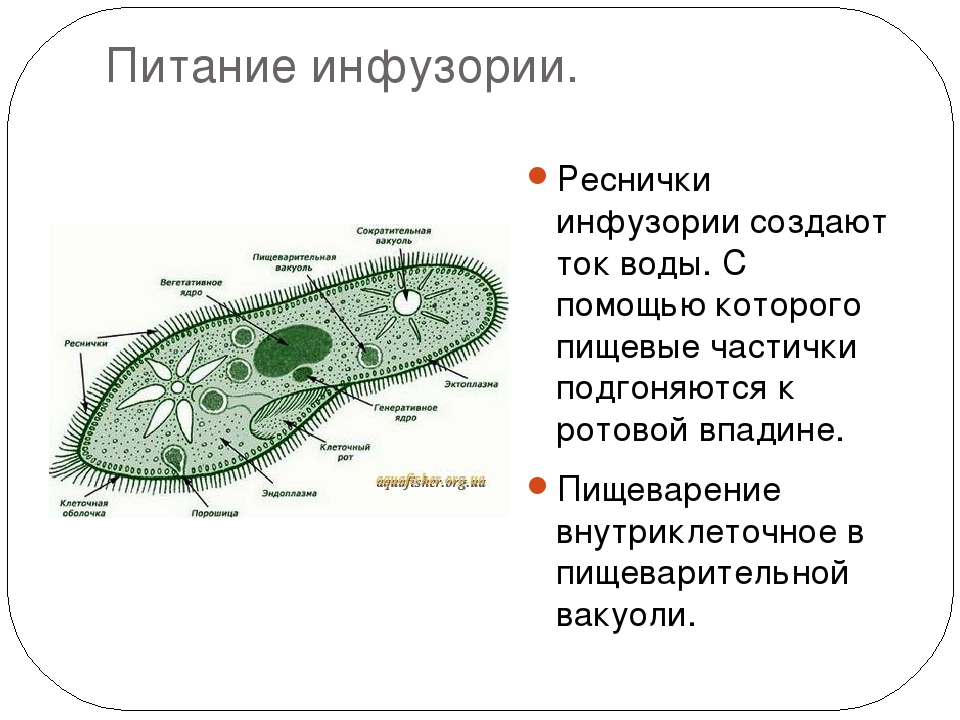 Сквирмер размещается под стеной, а ɛ — минимальное расстояние между стеной и поверхностью сквирмера. Предполагается, что сквирмер неподвижен в пространстве, т. е. в нем нет ни поступательного, ни вращательного движения. Скорость изгиба поверхности воздействует на неподвижный вибратор силой смазки F и крутящим моментом T . В случае твердого шара [ср. Инжир.1(b)], межцентровый вектор сферы и сквирмера установлен параллельно оси z . Предполагается, что и сфера, и сквирмер неподвижны в пространстве и имеют одинаковый радиус a . Для поверхности шара задано граничное условие прилипания, а для поверхности сквирмера – скорость изгиба. ɛ — минимальное расстояние между двумя поверхностями. Вектор ориентации сквирмера e установлен в направлении x . В случае свободной поверхности [ср.1(c)], поверхность считается плоской и расположена на z = 0. Предполагается, что нормальная скорость и касательное напряжение равны нулю на свободной поверхности.
Сквирмер размещается под стеной, а ɛ — минимальное расстояние между стеной и поверхностью сквирмера. Предполагается, что сквирмер неподвижен в пространстве, т. е. в нем нет ни поступательного, ни вращательного движения. Скорость изгиба поверхности воздействует на неподвижный вибратор силой смазки F и крутящим моментом T . В случае твердого шара [ср. Инжир.1(b)], межцентровый вектор сферы и сквирмера установлен параллельно оси z . Предполагается, что и сфера, и сквирмер неподвижны в пространстве и имеют одинаковый радиус a . Для поверхности шара задано граничное условие прилипания, а для поверхности сквирмера – скорость изгиба. ɛ — минимальное расстояние между двумя поверхностями. Вектор ориентации сквирмера e установлен в направлении x . В случае свободной поверхности [ср.1(c)], поверхность считается плоской и расположена на z = 0. Предполагается, что нормальная скорость и касательное напряжение равны нулю на свободной поверхности.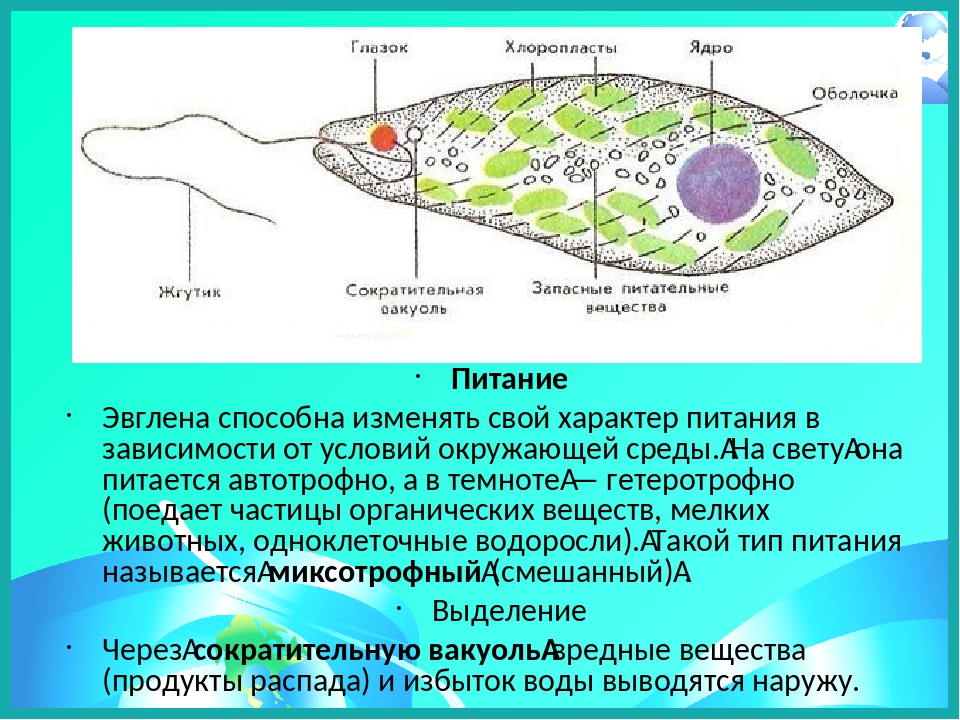 Сквирмер помещается ниже поверхности, где ε — минимальное расстояние между двумя поверхностями. Вектор ориентации сквирмера e установлен в направлении x , и предполагается, что сквирмер зафиксирован в пространстве.
Сквирмер помещается ниже поверхности, где ε — минимальное расстояние между двумя поверхностями. Вектор ориентации сквирмера e установлен в направлении x , и предполагается, что сквирмер зафиксирован в пространстве.| Fx=85πμaα(2α2+α+2)(1+α)3Δu[log(1/ε)+O(1)], | (2) |
| Ty=45πμa2α(α+4)(1+α)2Δu[log(1/ε)+O(1)], | (3) |
 Здесь мы используем общие выражения для наших конкретных задач. В случае жесткой стенки α можно считать бесконечным. Поскольку скорость на стенке равна нулю, а скорость изгиба поверхности определяется уравнением (1) имеем Δu=us(π/2)=3U0/2. С другой стороны, в случае твердой сферы α = 1 и Δu=3U0/2. Для свободной поверхности α и Δ u можно считать бесконечными и нулевыми соответственно. Вставляя значения α и Δ u , мы получаем члены старшего порядка для наших настроек задачи, как показано в таблице I.Мы видим, что член главного порядка исчезает для свободной поверхности. Это указывает на то, что вблизи свободной поверхности силы смазки малы по сравнению с жесткой стенкой; такая тенденция будет обсуждаться в гл. . ТАБЛИЦА I. Основные условия сил смазки, действующих на сквирмер с поверхностной скоростью, заданной уравнением. (1). Ячейка закреплена параллельно у жесткой стенки, твердой сферы или свободной поверхности.
Здесь мы используем общие выражения для наших конкретных задач. В случае жесткой стенки α можно считать бесконечным. Поскольку скорость на стенке равна нулю, а скорость изгиба поверхности определяется уравнением (1) имеем Δu=us(π/2)=3U0/2. С другой стороны, в случае твердой сферы α = 1 и Δu=3U0/2. Для свободной поверхности α и Δ u можно считать бесконечными и нулевыми соответственно. Вставляя значения α и Δ u , мы получаем члены старшего порядка для наших настроек задачи, как показано в таблице I.Мы видим, что член главного порядка исчезает для свободной поверхности. Это указывает на то, что вблизи свободной поверхности силы смазки малы по сравнению с жесткой стенкой; такая тенденция будет обсуждаться в гл. . ТАБЛИЦА I. Основные условия сил смазки, действующих на сквирмер с поверхностной скоростью, заданной уравнением. (1). Ячейка закреплена параллельно у жесткой стенки, твердой сферы или свободной поверхности.| Жесткая стена | Жесткая сфера | Бесплатная поверхность | бесплатная поверхность | ||
|---|---|---|---|---|---|
| F x | 245πμau0 [Log (1 / ε) + O (1)] | 32πμaU0[log(1/ε)+O(1)] | μaU0⋅O(1) | ||
| T y | 65πμa2+O(εlog)/U+O(1ε/a2+O) ] | 32πμa2U0[log(1/ε)+O(1)] | μa2U0⋅O(1) |
Старший член для твердого объекта равен log(1/ε), и, таким образом, он доминирует над решением только в математическом смысле. Даже если ε привести к молекулярному размеру, log (1 / ε) может быть недостаточно большим, чтобы доминировать над членом следующего порядка O (1). Поэтому для полного уточнения сил смазки необходимо численное моделирование.
Даже если ε привести к молекулярному размеру, log (1 / ε) может быть недостаточно большим, чтобы доминировать над членом следующего порядка O (1). Поэтому для полного уточнения сил смазки необходимо численное моделирование.
 01 и . Чтобы принять во внимание жесткую стенку и свободную поверхность, функции ядра, полученные Blake 46 46. J. Blake, «О движении слизи в легких», J. Biomech. 8 , 179–190 (1975). https://doi.org/10.1016/0021-9290(75)-8. Численные результаты F x показаны как функция ɛ на рис. 2(а); линия 6π указывает силу, создаваемую извивающейся скоростью в отсутствие каких-либо геометрических ограничений.Наибольшее значение F x было создано для жесткой стенки, тогда как наименьшее значение F x было создано для свободной поверхности; F x для твердой сферы находился между двумя случаями. F x для свободной поверхности меньше 6π, что указывает на то, что сила смазки качественно отличается от двух других случаев. Численные результаты T y показаны как функция ɛ на рис.2(б). В области ɛ / a T y было сгенерировано для жесткой стенки, тогда как наименьшее T y было сгенерировано для свободной поверхности.
01 и . Чтобы принять во внимание жесткую стенку и свободную поверхность, функции ядра, полученные Blake 46 46. J. Blake, «О движении слизи в легких», J. Biomech. 8 , 179–190 (1975). https://doi.org/10.1016/0021-9290(75)-8. Численные результаты F x показаны как функция ɛ на рис. 2(а); линия 6π указывает силу, создаваемую извивающейся скоростью в отсутствие каких-либо геометрических ограничений.Наибольшее значение F x было создано для жесткой стенки, тогда как наименьшее значение F x было создано для свободной поверхности; F x для твердой сферы находился между двумя случаями. F x для свободной поверхности меньше 6π, что указывает на то, что сила смазки качественно отличается от двух других случаев. Численные результаты T y показаны как функция ɛ на рис.2(б). В области ɛ / a T y было сгенерировано для жесткой стенки, тогда как наименьшее T y было сгенерировано для свободной поверхности.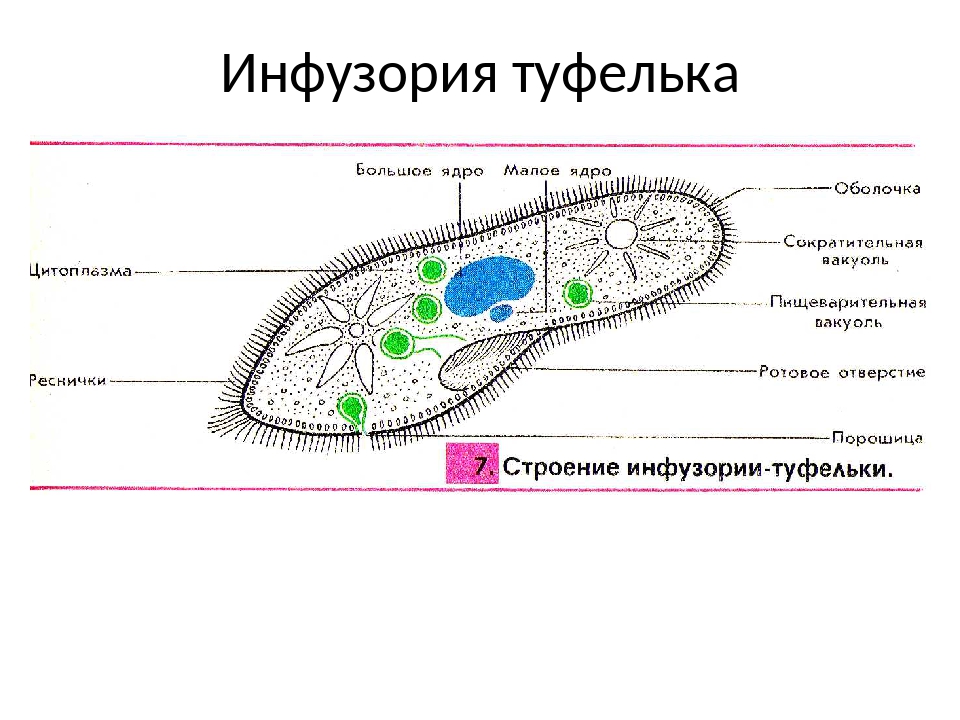 T y для твердой сферы находился между двумя случаями. Знак T y был положительным в области ɛ / a T y жесткой стенки был намного больше, чем свободной поверхности, клетки могут выйти из жесткой стенки гораздо больше без труда.Мы обсудим такие тенденции в пп. и . В области ɛ / a > 0,3 наибольшее значение T y было получено для твердой сферы, а не для жесткой стены. Более того, T y для свободной поверхности стали отрицательными, направляя клетку к свободной поверхности. Учитывая, что главные члены теории смазки (таблица 1) не объясняют прямо эти тенденции, для полного понимания момента смазки требуется более подробный численный анализ.
T y для твердой сферы находился между двумя случаями. Знак T y был положительным в области ɛ / a T y жесткой стенки был намного больше, чем свободной поверхности, клетки могут выйти из жесткой стенки гораздо больше без труда.Мы обсудим такие тенденции в пп. и . В области ɛ / a > 0,3 наибольшее значение T y было получено для твердой сферы, а не для жесткой стены. Более того, T y для свободной поверхности стали отрицательными, направляя клетку к свободной поверхности. Учитывая, что главные члены теории смазки (таблица 1) не объясняют прямо эти тенденции, для полного понимания момента смазки требуется более подробный численный анализ.C. Силы смазки, действующие на вибратор с углом входа
В этом подразделе мы численно рассчитываем силы смазки, действующие на вибратор, используя метод граничных элементов 39 39. T. Ishikawa, MP Simmonds и TJ Pedley, “ Гидродинамическое взаимодействие двух плавающих модельных микроорганизмов», J.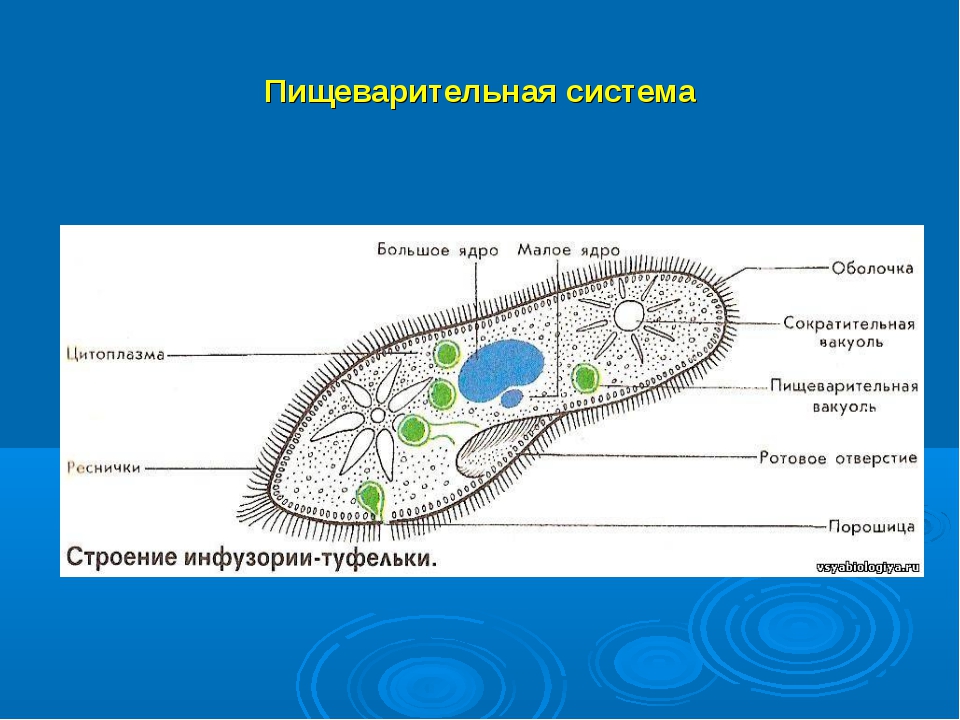 Fluid Mech. 568 , 119–160 (2006). https://doi.org/10.1017/S0022112006002631 в двух условиях: вблизи жесткой стенки и свободной поверхности, как показано на рис.3. Угол входа ϕ менялся от 0 (параллельно поверхности раздела) до π/2 (перпендикулярно поверхности раздела). Мы также варьировали прочность второй моды β , так как на силы смазки влияет β , когда ϕ≠0Z. Минимальное расстояние ɛ было установлено как 0,01 a . Остальные настройки задачи были такими же, как и в подразделе . На рис. 4(а) показаны численные результаты T y в случае жесткой стенки с различными условиями ϕ и β ; серые стрелки на рисунке показывают, как ϕ изменяется во времени.Когда β был положительным, т. е. пуллером, T y был положительным в диапазоне 0 ϕ β ≤ −2, т. е. сильным пушером, 9 y был отрицательным, 9 T большой режим ϕ . Отрицательное T y позволило ϕ сходиться к ϕ = π/2, как показано серыми стрелками.
Fluid Mech. 568 , 119–160 (2006). https://doi.org/10.1017/S0022112006002631 в двух условиях: вблизи жесткой стенки и свободной поверхности, как показано на рис.3. Угол входа ϕ менялся от 0 (параллельно поверхности раздела) до π/2 (перпендикулярно поверхности раздела). Мы также варьировали прочность второй моды β , так как на силы смазки влияет β , когда ϕ≠0Z. Минимальное расстояние ɛ было установлено как 0,01 a . Остальные настройки задачи были такими же, как и в подразделе . На рис. 4(а) показаны численные результаты T y в случае жесткой стенки с различными условиями ϕ и β ; серые стрелки на рисунке показывают, как ϕ изменяется во времени.Когда β был положительным, т. е. пуллером, T y был положительным в диапазоне 0 ϕ β ≤ −2, т. е. сильным пушером, 9 y был отрицательным, 9 T большой режим ϕ . Отрицательное T y позволило ϕ сходиться к ϕ = π/2, как показано серыми стрелками. Этот результат показывает, что сильный толкающий сквирмер может быть захвачен жесткой стенкой, когда угол входа достаточно велик.Мы обсудим эти тенденции в гл. . На рис. 4(б) показаны результаты T y в случае свободной поверхности с различными условиями ϕ и β ; серые стрелки на рисунке показывают, как ϕ изменяется со временем. Хотя рисунок похож на рис. 4(а), эффект β был совершенно другим. В случае свободной поверхности толкающий сквирмер, т. е. β β ≥ 2, т. е. сильный тянущий T y , на большом режиме ϕ становился отрицательным.Таким образом, при достаточно большом входном угле может захватиться свободная поверхность сильного пуллера. Эти результаты показывают, что характеристики геометрических ограничений оказывают существенное влияние на поведение клеток.
Этот результат показывает, что сильный толкающий сквирмер может быть захвачен жесткой стенкой, когда угол входа достаточно велик.Мы обсудим эти тенденции в гл. . На рис. 4(б) показаны результаты T y в случае свободной поверхности с различными условиями ϕ и β ; серые стрелки на рисунке показывают, как ϕ изменяется со временем. Хотя рисунок похож на рис. 4(а), эффект β был совершенно другим. В случае свободной поверхности толкающий сквирмер, т. е. β β ≥ 2, т. е. сильный тянущий T y , на большом режиме ϕ становился отрицательным.Таким образом, при достаточно большом входном угле может захватиться свободная поверхность сильного пуллера. Эти результаты показывают, что характеристики геометрических ограничений оказывают существенное влияние на поведение клеток.Структура сообщества и биоразнообразие почвенных инфузорий в мангровом лесу Дунчжайган на острове Хайнань, Китай
Было исследовано распределение почвенных инфузорий в трех различных местах обитания в типичном мангровом лесу в Дунчжайган, Хайнань, Китай. Проанализированы численность, биоразнообразие и сходство сообществ инфузорий в свежей и воздушно-сухой почве с различными физико-химическими свойствами. Было обнаружено три класса, 11 отрядов, 34 рода и 70 видов инфузорий, причем первой доминирующей группой были Hypotrichida. Биоразнообразие инфузорий соответствовало Участку B < Участку A < Участку C как в свежих, так и в высушенных образцах. Обилие инфузорий положительно коррелировало с влажностью почвы, соленостью, органическим веществом (OM), общим азотом (TN), общим фосфором (TP) и сульфатом (), но отрицательно с pH и общим калием (TK).Сайт A и сайт B и сайт B и сайт C показали наибольшее сходство в свежих и высушенных образцах соответственно. Повсеместные характеристики распространения инфузорий предполагают их важную роль в пищевых сетях и круговороте питательных веществ. Присутствие Colpodida было связано с мангровыми растениями.
Проанализированы численность, биоразнообразие и сходство сообществ инфузорий в свежей и воздушно-сухой почве с различными физико-химическими свойствами. Было обнаружено три класса, 11 отрядов, 34 рода и 70 видов инфузорий, причем первой доминирующей группой были Hypotrichida. Биоразнообразие инфузорий соответствовало Участку B < Участку A < Участку C как в свежих, так и в высушенных образцах. Обилие инфузорий положительно коррелировало с влажностью почвы, соленостью, органическим веществом (OM), общим азотом (TN), общим фосфором (TP) и сульфатом (), но отрицательно с pH и общим калием (TK).Сайт A и сайт B и сайт B и сайт C показали наибольшее сходство в свежих и высушенных образцах соответственно. Повсеместные характеристики распространения инфузорий предполагают их важную роль в пищевых сетях и круговороте питательных веществ. Присутствие Colpodida было связано с мангровыми растениями.
1. Введение
Мангровые экосистемы представляют собой типичные системы водно-болотных угодий в прибрежных отложениях ила и ила в тропиках и субтропических широтах. Они играют важную роль в поддержании и улучшении биологической среды, очистке воздуха и противодействии загрязнению сточными водами и стихийным бедствиям в бухтах или устьях рек.Они рассматриваются как «морские леса» из-за их особой экологической, экономической и туристической ценности [1]. Значение мангровых зарослей привлекает все большее внимание в последние годы, особенно после недавнего сильного цунами в Индийском океане [2]. В мангровых экосистемах проводится все больше и больше исследований.
Они играют важную роль в поддержании и улучшении биологической среды, очистке воздуха и противодействии загрязнению сточными водами и стихийным бедствиям в бухтах или устьях рек.Они рассматриваются как «морские леса» из-за их особой экологической, экономической и туристической ценности [1]. Значение мангровых зарослей привлекает все большее внимание в последние годы, особенно после недавнего сильного цунами в Индийском океане [2]. В мангровых экосистемах проводится все больше и больше исследований.
Хотя все основные группы мангровых растений и животных в той или иной степени изучены [3–7], мало информации об одноклеточных почвенных простейших, за исключением нескольких описанных видов фораминифер, амебоидных и реснитчатых простейших [8–7]. 11].Исследования почвенных инфузорий в местообитаниях мангровых лесов проводятся редко.
Являясь важной группой в круговороте питательных веществ, потоках энергии и пищевых цепях [12, 13], почвенные инфузории участвовали в разложении бентосных остаточных отложений, формировании и развитии мангровых почв и ускоряли процессы минерализации углерода, азота , и другие элементы минерального питания [14]. Как основные потребители бактерий, почвенные инфузории также обладают особыми характеристиками, такими как сильное дыхание, короткое время генерации и быстрое размножение.В ризосфере живых растений простейшие играют важную роль в минерализации элементов минерального питания. Горшечные эксперименты Экелунда и Ронна [14] показали, что органические вещества, выделяемые растениями, могут стимулировать активность бактерий и инфузорий в корневой зоне, что приводит к минерализации органического почвенного азота и его ассимиляции растениями. Заметный эффект, оказываемый почвенными инфузориями, может иметь важное значение для питания мангровых растений; с другой стороны, рост растений также может существенно влиять на качество почвы и сообщество инфузорий.Корни растений и сообщество почвенных инфузорий взаимозависимы.
Как основные потребители бактерий, почвенные инфузории также обладают особыми характеристиками, такими как сильное дыхание, короткое время генерации и быстрое размножение.В ризосфере живых растений простейшие играют важную роль в минерализации элементов минерального питания. Горшечные эксперименты Экелунда и Ронна [14] показали, что органические вещества, выделяемые растениями, могут стимулировать активность бактерий и инфузорий в корневой зоне, что приводит к минерализации органического почвенного азота и его ассимиляции растениями. Заметный эффект, оказываемый почвенными инфузориями, может иметь важное значение для питания мангровых растений; с другой стороны, рост растений также может существенно влиять на качество почвы и сообщество инфузорий.Корни растений и сообщество почвенных инфузорий взаимозависимы.
Кроме того, они являются хорошими биоиндикаторами почвенной среды [15–17]. Важно изучить структуру сообщества почвенных инфузорий и их значение в почвенной среде, чтобы лучше понять функцию мангровых экосистем и, таким образом, помочь в защите мангровых ресурсов. Настоящее исследование направлено на изучение обилия видов, биоразнообразия и индекса сходства сообщества почвенных инфузорий в типичном мангровом лесу в Дунчжайгане, остров Хайнань в Китае.В исследовании также делается попытка сравнить разницу в сообществе инфузорий между тремя различными средами обитания: голой землей, а также посаженными и естественными мангровыми зарослями в лесу, и связать обилие инфузорий со свойствами почвы.
Настоящее исследование направлено на изучение обилия видов, биоразнообразия и индекса сходства сообщества почвенных инфузорий в типичном мангровом лесу в Дунчжайгане, остров Хайнань в Китае.В исследовании также делается попытка сравнить разницу в сообществе инфузорий между тремя различными средами обитания: голой землей, а также посаженными и естественными мангровыми зарослями в лесу, и связать обилие инфузорий со свойствами почвы.
2. Материалы и методы
2.1. Описание области исследования и отбора проб почвы
Дунчжайган (110°3–110°3 в.д., 19°5–20°0 с.ш.) представляет собой залив с мелководьем, образованный континентальным стоком во время Великого землетрясения в Цюнчжоу в 1605 г. Общая площадь Эверглейд составляет 5400 м 2 , из них 2065 м 2 мангровых лесов и 3335 м 2 илистых и мелководных акваторий.Дунчжайган имеет типичный субтропический муссонный морской климат. среднегодовая температура воздуха колеблется от 23,3°С до 23,8°С; Среднегодовое количество осадков достигает 1676,4 мм, а среднее количество солнечного света – 2200 ч [18].
Образцы почвы были отобраны в трех различных местах обитания в национальном мангровом заповеднике Дунчжайган в апреле и сентябре 2006 г. (рис. 1). Участок А представлял собой голую землю без какой-либо растительности; Участок B представлял собой искусственно посаженный мангровый лес Sonneratia apetala (на расстоянии между двумя деревьями 2.5 м друг от друга) заповедником три года назад и достигал 2 м в высоту; Участок C представлял собой естественную среду обитания мангровых зарослей, в которой преобладали Bruguiera gymnorrhiza с несколькими особями Ceriops tagal . Три участка располагались в треугольнике на расстоянии около 100 м друг от друга. Площадь опробования каждой пробной площадки составляла около 10 м 2 , а десять повторных проб почвы (0–5 см) с тонкими корнями растений были отобраны с использованием «метода параллельного скачка» [15, 19]. Поскольку требовались только поверхностные образцы почвы (0–5 см), вместо пробоотборника для отбора образцов почвы здесь использовалась лопата. Десять повторных образцов поверхностной почвы были равномерно перемешаны и объединены в составной образец в полевых условиях, затем немедленно собрано около 3 кг этого составного образца и немедленно помещено в стерилизованный пластиковый пакет, запечатан и доставлен обратно в лабораторию [19]. . Свежие образцы были немедленно проанализированы; остальные порции сушили на воздухе не менее одного месяца, а затем анализировали.
Десять повторных образцов поверхностной почвы были равномерно перемешаны и объединены в составной образец в полевых условиях, затем немедленно собрано около 3 кг этого составного образца и немедленно помещено в стерилизованный пластиковый пакет, запечатан и доставлен обратно в лабораторию [19]. . Свежие образцы были немедленно проанализированы; остальные порции сушили на воздухе не менее одного месяца, а затем анализировали.
2.2. Анализ физико-химических параметров образцов почвы
Следующие свойства почвы каждого образца почвы были определены в соответствии со стандартными процедурами [20].Каждый объединенный образец вручную очищали от фрагментов растений и камешков, взвешивали, сушили при 60°С в течение 72 ч и повторно взвешивали для определения влажности. Значения pH и солености были получены с использованием раствора почва/вода 1 : 5 по электропроводности (автоматический кондуктометр с температурной компенсацией). Анализы органического вещества проводили кислотно-дихроматным методом окисления, общего азота (ОN) – полумикрометодом Кьельдаля, общего фосфата (ОФ) – методом спектрофотометрии молибдата аммония, общего калия (ТК) – методом пламенной фотометрии, и с использованием помутнения сульфата бария. Механический состав почвы каждого высушенного образца анализировали ареометрическим методом.
Механический состав почвы каждого высушенного образца анализировали ареометрическим методом.
2.3. Качественные исследования почвенных инфузорий
Все образцы, как свежие, так и воздушно-сухие, были качественно проанализированы по «методу незатопляемых чашек Петри», описанному Фойсснером [15]. Образец почвы массой 100–150 г помещали в чашку Петри (диаметром 15–20 см), насыщенную, но не заливаемую дистиллированной водой. На 2, 4, 7, 14, 21 и 28 сут для определения видов инфузорий отбирали около 2 мл стока из каждой культуры [15, 19].Живые образцы наблюдали под мощным масляным иммерсионным объективом с помощью светлопольной, фазово-контрастной или дифференциальной интерференционно-контрастной микроскопии (увеличение 40–1000; Nikon, YS2-H и E800) и дополняли окрашиванием серебряной линией [21]. . Идентификация, номенклатура и терминология видов инфузорий были выполнены в соответствии со следующим: Berger [22], Carey [23], Foissner [21], Kahl [24], Lee et al. [25], Shen and Gong [26] Shen et al. [27] и Песня [28].
[27] и Песня [28].
2.4. Количественные исследования почвенных инфузорий
Количественный анализ почвенных инфузорий был основан на модифицированном методе «наиболее вероятного числа» (MPN), использованном Darbyshire et al. [29]. Согласно нашему предварительному эксперименту, как для свежих, так и для воздушно-сухих образцов почвы были приняты коэффициенты разбавления 10 2 –10 4 .
2.5. Обработка данных и статистический анализ
Для расчета индекса разнообразия сообщества инфузорий использовали формулу Маргалефа [30]: где – индекс разнообразия; – номер вида; это общее количество особей всех видов.
Доминирование – отношение числа доминирующих групп к общему числу видов [31]. Формула Жаккара [32] использовалась для расчета анализа сходства сообществ инфузорий из разных образцов почвы: где – индекс подобия; и – общее количество видов на участках 1 и 2 соответственно; – число видов, общих для обоих участков 1 и 2.
Для выявления комплексного влияния мультифакторов на обилие почвенных инфузорий и степени корреляции между этими факторами и обилием инфузорий был проведен пошаговый регрессионный анализ [33].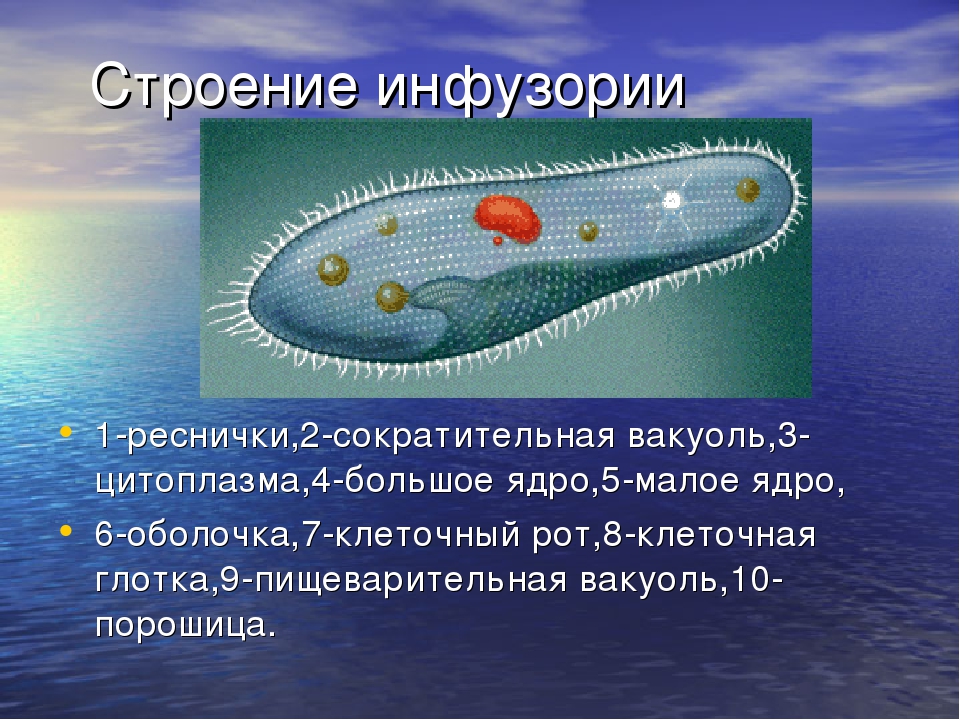 Иерархический кластер был проведен для анализа сходства физико-химических признаков 3-х мест отбора проб. Все статистические анализы проводились с использованием программного обеспечения SPSS 13.0.
Иерархический кластер был проведен для анализа сходства физико-химических признаков 3-х мест отбора проб. Все статистические анализы проводились с использованием программного обеспечения SPSS 13.0.
3. Результаты
3.1. Физические/химические параметры образцов почвы мангровых лесов
Основные физико-химические параметры в трех местообитаниях мангровых лесов, измеренные с использованием образцов, собранных в апреле 2006 г. в качестве фона окружающей среды, перечислены в таблице 1. Некоторые физические/химические факторы, такие как как процент воды, ОВ, TN, и сильно различаются на разных участках отбора проб.Другие, такие как pH, соленость, TP и TK, немного отличаются. Иерархическая кластерная дендрограмма показала, что физико-химические свойства почвы на голой земле и в антропогенных мангровых зарослях более схожи; в остальном естественные мангровые заросли отличаются от других участков (рис. 2).
3.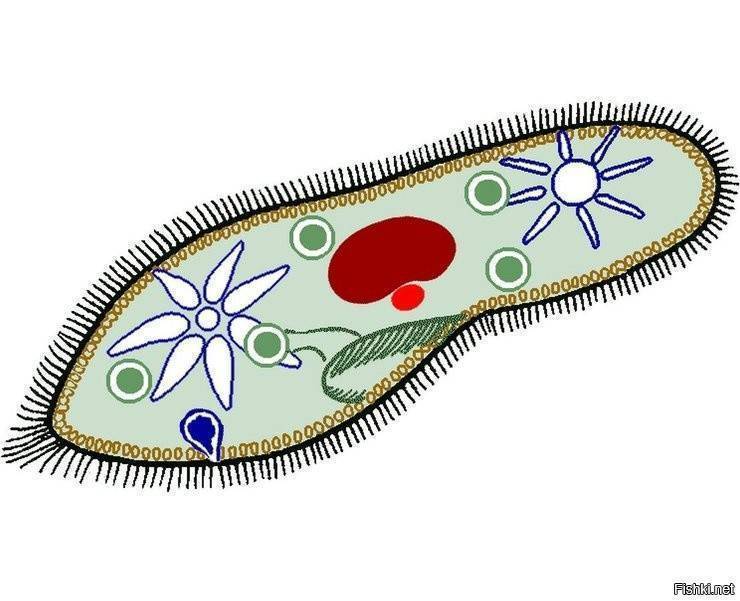 3. Численность инфузорий и биоразнообразие 3. Численность инфузорий и биоразнообразиеТаблица 3 показывает, что численность почвенных инфузорий в трех местообитаниях снижалась в порядке Участок A < Участок B < Участок C. Численность инфузорий была самой высокой в свежем образце с участка C, достигая 13 770 особей. инд. g -1 , в то время как самый низкий был обнаружен в высушенном образце с площадки А, всего 114 инд. г -1 . Результаты также показали, что индекс биоразнообразия Маргалефа для всех образцов колебался от 2 до 3, за исключением свежего образца из Зоны C.Индекс биоразнообразия инфузорий как в свежих, так и в высушенных образцах также различался между тремя местообитаниями в порядке Участок Б < Участок А < Участок С. Разница была более очевидной в свежих образцах (расхождение составило 3,12), чем в высушенных образцах.
FS: образцы свежей почвы; DS: высушенные образцы почвы. | 3.4. Индексы сходства сообщества инфузорий Индексы сходства сообщества инфузорий варьировались от 0.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 51
51 2. Структура сообщества почвенных инфузорий в мангровом лесу
2. Структура сообщества почвенных инфузорий в мангровом лесу
 Г -1 )
Г -1 )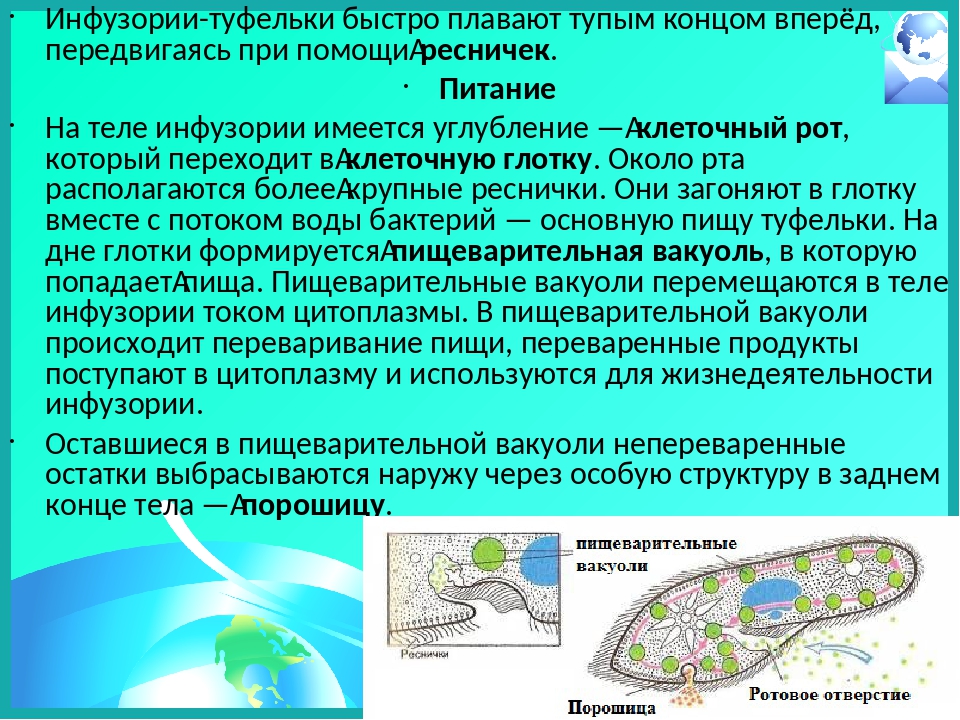 от 18 до 0,26 (свежие образцы) и от 0,38 до 0,54 (высушенные образцы) (табл. 4). У высушенных образцов она была больше, чем у свежих. Порядок индекса подобия в свежих образцах следовал за Участком A и Участком C < Участком B и Участком C < Участком A и Участком B, в то время как порядок был немного другим в высушенных образцах (Участок A и Участок C < Участок A и Участок B < Участок). B и сайт C).
от 18 до 0,26 (свежие образцы) и от 0,38 до 0,54 (высушенные образцы) (табл. 4). У высушенных образцов она была больше, чем у свежих. Порядок индекса подобия в свежих образцах следовал за Участком A и Участком C < Участком B и Участком C < Участком A и Участком B, в то время как порядок был немного другим в высушенных образцах (Участок A и Участок C < Участок A и Участок B < Участок). B и сайт C). 54
54 934
934
 002
002 005
005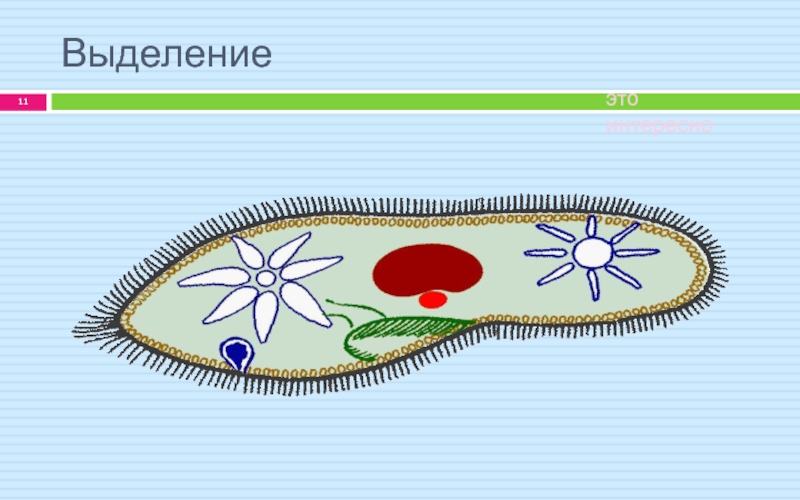 Такие биологические характеристики позволяют этим двум группам инфузорий адаптироваться к литоральной мангровой почве, где среда обитания попеременно находится в условиях погружения и воздействия. Это также помогает объяснить, почему Hypotrichida и Colpodida, которые распространены в земных почвах, также доминируют в местообитаниях мангровых почв.Однако доминирующей группой на участке А были Oligotrichida, а не Colpodida, что позволяет предположить, что среда обитания на участке А отличалась от земных почв. Инфузории Oligotrichida, особенно Halteria spp., обычно считались планктонными видами инфузорий. Участок А, голая земля в наиболее обращенном к морю месте, имел более низкий рельеф, и почва чаще покрывалась приливной морской водой, чем на других участках. Также возможно, что некоторые виды планктонных инфузорий были занесены на Участок А приливами.Инфузории-олиготрихиды были основными потребителями водорослей и бактерий в водной среде и играли очень важную роль в водных пищевых цепях из-за их обширного питания, быстрого роста, быстрого перемещения и сильной активности [40].
Такие биологические характеристики позволяют этим двум группам инфузорий адаптироваться к литоральной мангровой почве, где среда обитания попеременно находится в условиях погружения и воздействия. Это также помогает объяснить, почему Hypotrichida и Colpodida, которые распространены в земных почвах, также доминируют в местообитаниях мангровых почв.Однако доминирующей группой на участке А были Oligotrichida, а не Colpodida, что позволяет предположить, что среда обитания на участке А отличалась от земных почв. Инфузории Oligotrichida, особенно Halteria spp., обычно считались планктонными видами инфузорий. Участок А, голая земля в наиболее обращенном к морю месте, имел более низкий рельеф, и почва чаще покрывалась приливной морской водой, чем на других участках. Также возможно, что некоторые виды планктонных инфузорий были занесены на Участок А приливами.Инфузории-олиготрихиды были основными потребителями водорослей и бактерий в водной среде и играли очень важную роль в водных пищевых цепях из-за их обширного питания, быстрого роста, быстрого перемещения и сильной активности [40].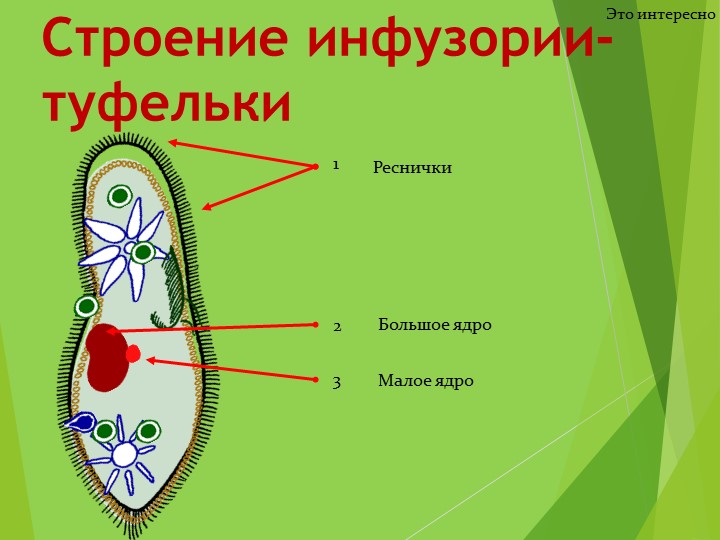
 7
7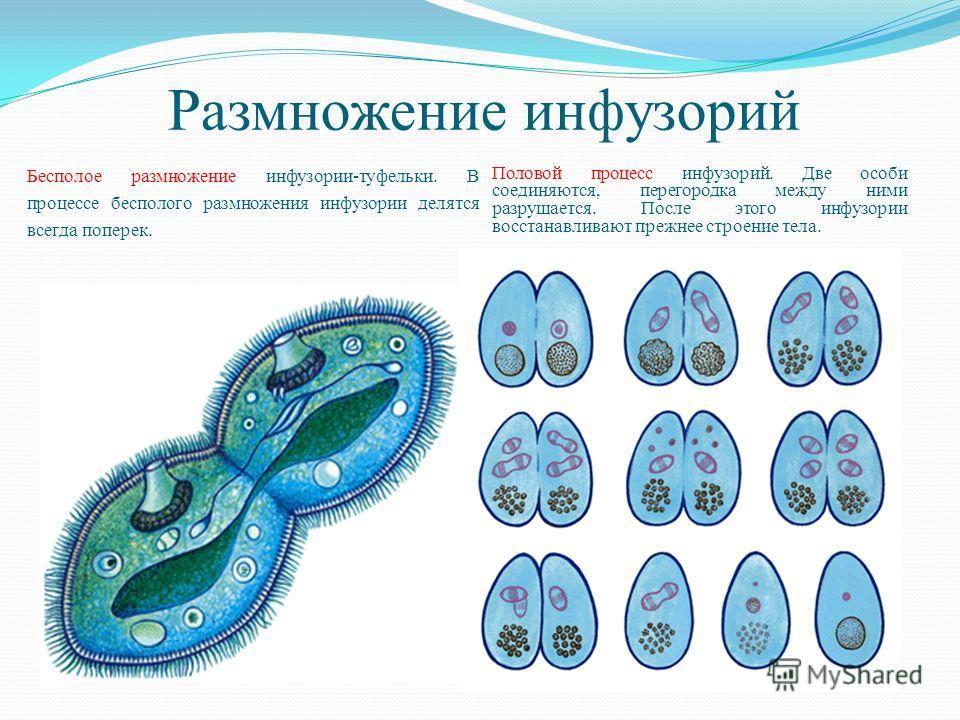 Частоты Colpoda cucullus и Colpoda inflata также были высокими в Антарктиде [37]. Как указывали Finlay et al. [41, 42], виды инфузорий, которые были локально редкими или многочисленными, были столь же редкими или многочисленными в глобальном масштабе.Например, Gonostomum affine и Colpoda spp. могут рассматриваться как космополитные виды инфузорий во всех почвенных образцах с различным местонахождением и средой из-за их морфологических особенностей и особенностей распространения. В настоящем исследовании эти инфузории также доминировали в почвах мангровых лесов, что позволяет предположить, что вывод Финлея можно применить к такому типу почвенной среды обитания.
Частоты Colpoda cucullus и Colpoda inflata также были высокими в Антарктиде [37]. Как указывали Finlay et al. [41, 42], виды инфузорий, которые были локально редкими или многочисленными, были столь же редкими или многочисленными в глобальном масштабе.Например, Gonostomum affine и Colpoda spp. могут рассматриваться как космополитные виды инфузорий во всех почвенных образцах с различным местонахождением и средой из-за их морфологических особенностей и особенностей распространения. В настоящем исследовании эти инфузории также доминировали в почвах мангровых лесов, что позволяет предположить, что вывод Финлея можно применить к такому типу почвенной среды обитания. Их взаимодействие установило основные условия среды для почвенных инфузорий [14]. Варгас и Хаттори [43] показали, что агрегатная структура почвы будет эффективно ограничивать передвижение инфузорий, и только при достаточно высокой влажности почвы возможен их переход от одного агрегата к другому. Поэтому фауна инфузорий из разных агрегатов в одной и той же почве может сильно различаться.
Их взаимодействие установило основные условия среды для почвенных инфузорий [14]. Варгас и Хаттори [43] показали, что агрегатная структура почвы будет эффективно ограничивать передвижение инфузорий, и только при достаточно высокой влажности почвы возможен их переход от одного агрегата к другому. Поэтому фауна инфузорий из разных агрегатов в одной и той же почве может сильно различаться. Напротив, участок А (голая земля) состоял только из плотного ила, и около 50% гранул почвы имели диаметр менее 0.002 мм. Такая структура почвы была тонкой и не имела многих подходящих пор для подвижных инфузорий и жизнедеятельности инфузорий [44]. Во-вторых, влажность почвы на участке С была значительно выше, чем на участке А; значения на участках A, B и C составили 24,2%, 35,6% и 45,4% соответственно. Сообщалось, что почвенная влага была основным фактором, ограничивающим выживание, размножение и распространение почвенных простейших на полуострове Файлдс в Антарктиде [45]. Высокая влажность почвы на участке C объясняет, почему виды инфузорий были более многочисленными, а биоразнообразие здесь выше, чем на двух других участках.В-третьих, в донном иле на участке С (естественные мангровые заросли) обнаружено большое количество органических остатков и растительных остатков. Пометы не только обеспечивали достаточное количество органического вещества, гумуса и других питательных веществ для почвенных инфузорий, но также позволяли им значительно перемещаться (особенно для инфузорий Hypotrichida).
Напротив, участок А (голая земля) состоял только из плотного ила, и около 50% гранул почвы имели диаметр менее 0.002 мм. Такая структура почвы была тонкой и не имела многих подходящих пор для подвижных инфузорий и жизнедеятельности инфузорий [44]. Во-вторых, влажность почвы на участке С была значительно выше, чем на участке А; значения на участках A, B и C составили 24,2%, 35,6% и 45,4% соответственно. Сообщалось, что почвенная влага была основным фактором, ограничивающим выживание, размножение и распространение почвенных простейших на полуострове Файлдс в Антарктиде [45]. Высокая влажность почвы на участке C объясняет, почему виды инфузорий были более многочисленными, а биоразнообразие здесь выше, чем на двух других участках.В-третьих, в донном иле на участке С (естественные мангровые заросли) обнаружено большое количество органических остатков и растительных остатков. Пометы не только обеспечивали достаточное количество органического вещества, гумуса и других питательных веществ для почвенных инфузорий, но также позволяли им значительно перемещаться (особенно для инфузорий Hypotrichida).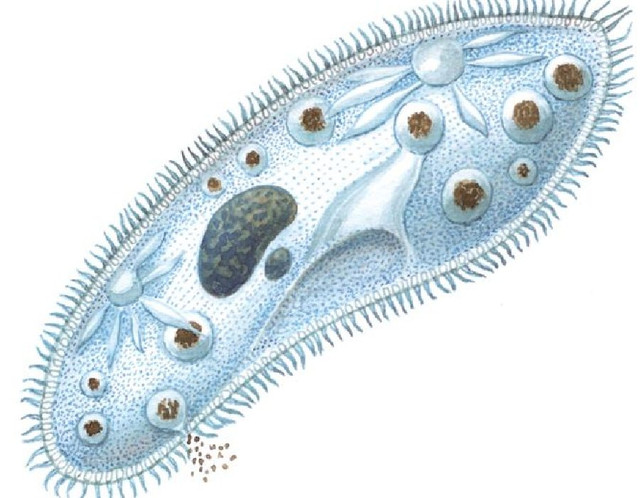 Наконец, синергетический эффект нескольких факторов, таких как pH, соленость и температура, усилит различия между различными агрегатами/местами обитания мангровых почв.
Наконец, синергетический эффект нескольких факторов, таких как pH, соленость и температура, усилит различия между различными агрегатами/местами обитания мангровых почв.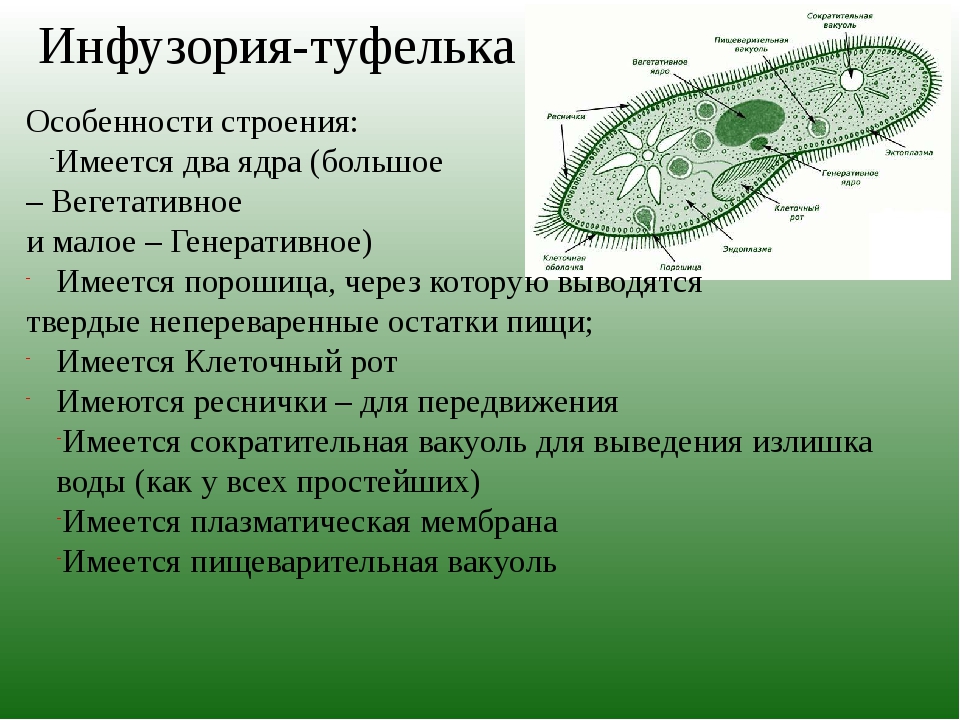 Образцы почвы из трех разных местообитаний были определены с помощью MPN и сопоставлены методом прямого подсчета. Численность инфузорий в свежих образцах была намного выше, чем в высушенных образцах, что свидетельствует о том, что некоторые инфузории не инцистировались или эксцистировались после сушки на воздухе, что приводило к прямому снижению численности инфузорий. С другой стороны, уменьшение количества бактерий (служащих пищей для инфузорий) в высушенных образцах может косвенно влиять на обилие инфузорий. В этом исследовании виды инфузорий, которые были в изобилии в свежих образцах, но исчезли в высушенных образцах, были Euplotes , Diphrys , Cyrtolophosis, и Plagiocampa , что указывает на то, что эти виды имели более низкую способность к инцистированию или эксцистированию, чем другие виды. инфузории, такие как Colpoda и Gonostomum .Подобные результаты были также получены Foissner [19]. Имеющиеся данные о составе и численности инфузорий показали, что для получения более детальной информации о сообществе почвенных инфузорий необходимо совместное использование МПН и других методов анализа почвенных инфузорий.
Образцы почвы из трех разных местообитаний были определены с помощью MPN и сопоставлены методом прямого подсчета. Численность инфузорий в свежих образцах была намного выше, чем в высушенных образцах, что свидетельствует о том, что некоторые инфузории не инцистировались или эксцистировались после сушки на воздухе, что приводило к прямому снижению численности инфузорий. С другой стороны, уменьшение количества бактерий (служащих пищей для инфузорий) в высушенных образцах может косвенно влиять на обилие инфузорий. В этом исследовании виды инфузорий, которые были в изобилии в свежих образцах, но исчезли в высушенных образцах, были Euplotes , Diphrys , Cyrtolophosis, и Plagiocampa , что указывает на то, что эти виды имели более низкую способность к инцистированию или эксцистированию, чем другие виды. инфузории, такие как Colpoda и Gonostomum .Подобные результаты были также получены Foissner [19]. Имеющиеся данные о составе и численности инфузорий показали, что для получения более детальной информации о сообществе почвенных инфузорий необходимо совместное использование МПН и других методов анализа почвенных инфузорий.
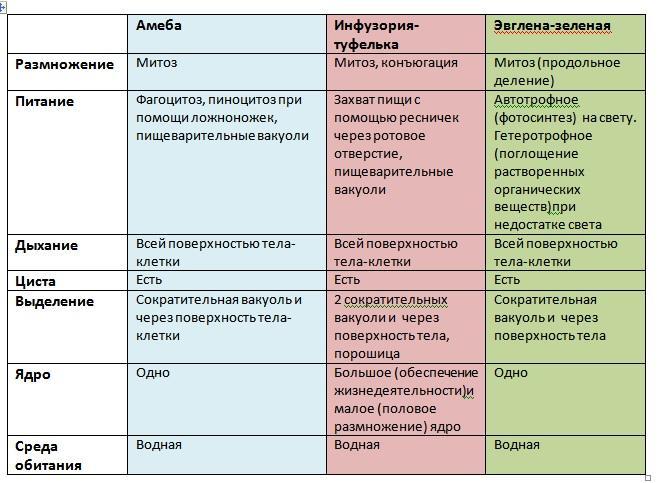
 Фактически, участок B (искусственно посаженные мангровые заросли) был переходным этапом от участка A (голая земля) к участку C (естественный мангровые заросли) в мангровом лесу.Таким образом, мы можем вывести эволюцию различных мангровых почв и восстановление мангровой растительности через их показатели сходства. Сообщество почвенных инфузорий можно использовать для оценки восстановления экосистемы мангровых зарослей.
Фактически, участок B (искусственно посаженные мангровые заросли) был переходным этапом от участка A (голая земля) к участку C (естественный мангровые заросли) в мангровом лесу.Таким образом, мы можем вывести эволюцию различных мангровых почв и восстановление мангровой растительности через их показатели сходства. Сообщество почвенных инфузорий можно использовать для оценки восстановления экосистемы мангровых зарослей. Рональд Эдвард Перлман
Рональд Эдвард Перлман 
 Белковые феромоны, которые контролируют само-/не-самораспознавание и спаривание у инфузорий, считаются эволюционными предшественниками факторов роста животных.
Белковые феромоны, которые контролируют само-/не-самораспознавание и спаривание у инфузорий, считаются эволюционными предшественниками факторов роста животных.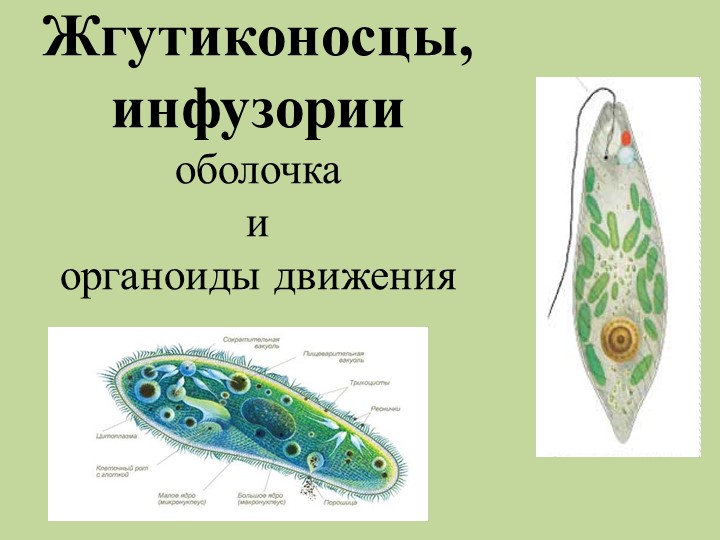 Рукописи можно подавать до указанного срока.Все статьи будут рецензироваться. Принятые статьи будут постоянно публиковаться в журнале (как только они будут приняты) и будут перечислены вместе на веб-сайте специального выпуска. Приглашаются исследовательские статьи, обзорные статьи, а также короткие сообщения. Для планируемых статей в редакцию можно отправить название и краткую аннотацию (около 100 слов) для размещения на сайте.
Рукописи можно подавать до указанного срока.Все статьи будут рецензироваться. Принятые статьи будут постоянно публиковаться в журнале (как только они будут приняты) и будут перечислены вместе на веб-сайте специального выпуска. Приглашаются исследовательские статьи, обзорные статьи, а также короткие сообщения. Для планируемых статей в редакцию можно отправить название и краткую аннотацию (около 100 слов) для размещения на сайте.