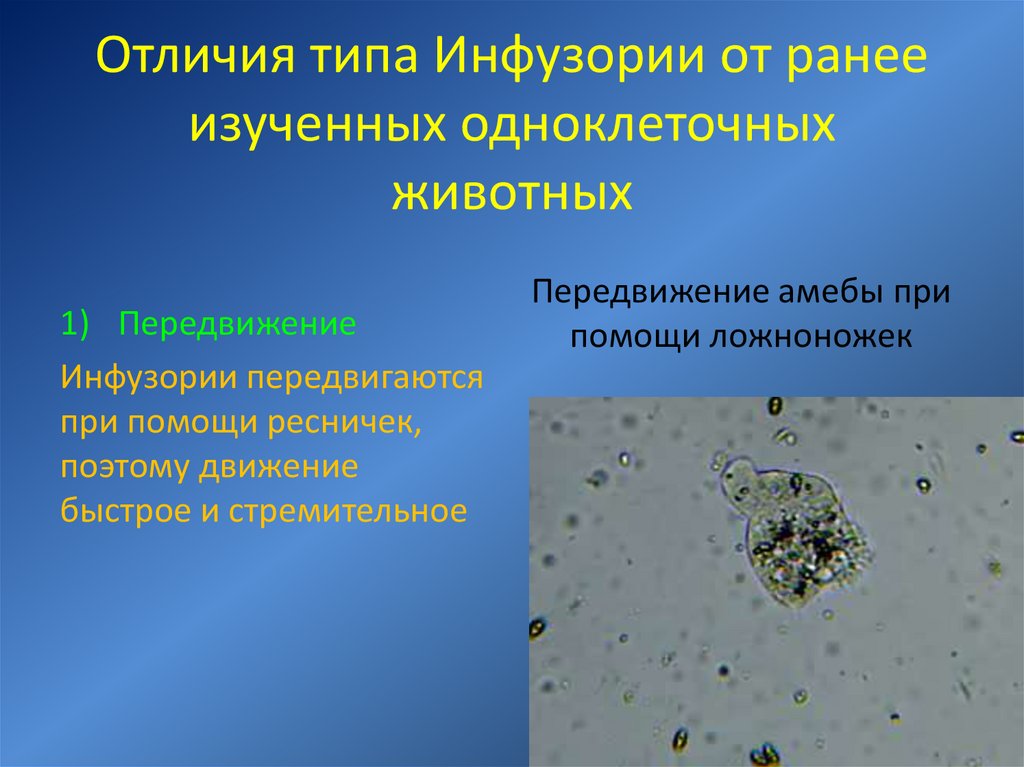
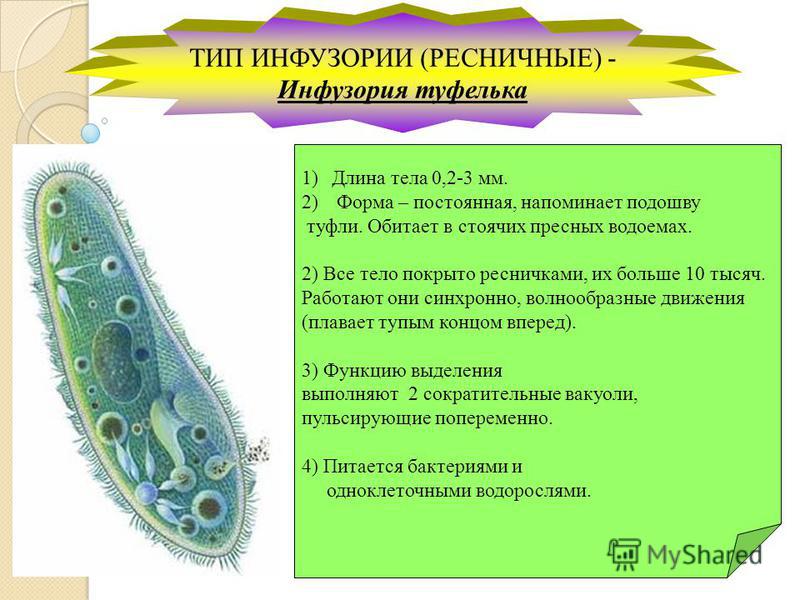
Инфузория туфелька это бактерия или животное или растение или гриб
Инфузория туфелька – самая сложная из простейших
Жизнь на нашей планете отличается невероятным многообразием всевозможных живых организмов, имеющих подчас невероятно сложное строение. Все это многообразие жизни: от простейших насекомых и растений до нас, людей (пожалуй, самых «сложных организмов») состоит из клеток, этих маленьких кирпичиков живой материи. И если человек – венец биологической эволюции, то весьма любопытным будет рассмотреть ее начало: простейшие одноклеточные организмы, которые, по сути, на заре истории стали родоначальниками всего живого. Инфузория туфелька (наряду с амебой и эвгленой зеленой) является одним из самых известных простых одноклеточных существ. Какое строение инфузории туфельки, среда обитания, как она питается и размножается, обо всем этом читайте далее.
Инфузория туфелька: описание и характеристика. Как выглядит инфузория туфелька?
На самом деле инфузория туфелька это вовсе не один простейший одноклеточный организм, за этим названием скрывается более 7 тысяч разных видов инфузорий. Всех их объединяет форма, которая чем-то напоминает подошву туфли, отсюда и «туфелька» в названии. (Впрочем, «туфелька» в названии прижилась только у нас, в английском языке «инфузория туфелька» значится под латинским названием «Paramecium caudatum», что переводится как «парамеция хвостатая»).
Всех их объединяет форма, которая чем-то напоминает подошву туфли, отсюда и «туфелька» в названии. (Впрочем, «туфелька» в названии прижилась только у нас, в английском языке «инфузория туфелька» значится под латинским названием «Paramecium caudatum», что переводится как «парамеция хвостатая»).
Также все инфузории обладают способностью к осморегуляции, то есть могут регулировать давление внутренней среды своего организма. В этом деле им помогают две сократительные вакуоли, они сжимаются и разжимаются, таким образом, выталкивая излишки жидкости из тела инфузории.
Размеры инфузории туфельки составляют от 1 до 5 десятых миллиметра.
Фото инфузории туфельки.
Хотя инфузория туфелька и является простейшим одноклеточным существом, то есть все ее тело состоит только из одной клетки, тем не менее, она имеет способность самостоятельно дышать, питаться, размножаться, передвигаться. Иными словами, обладает всеми теми функциями и способностями, которые имеет всякое другое животное.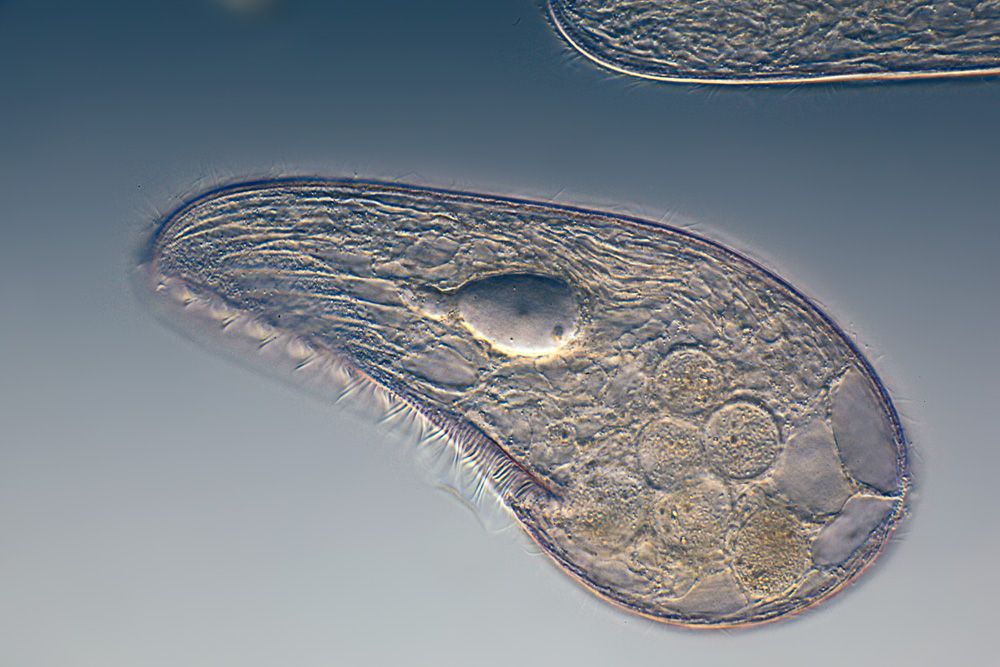 Более того среди других простейших одноклеточных организмов именно инфузория туфелька является самой сложной. В частности среди ее органоидов (элементов клетки) есть такие, которых нет у других ее одноклеточных «коллег»: амеб и эвглен.
Более того среди других простейших одноклеточных организмов именно инфузория туфелька является самой сложной. В частности среди ее органоидов (элементов клетки) есть такие, которых нет у других ее одноклеточных «коллег»: амеб и эвглен.
Среди «передовых» органоидов инфузории можно отметить:
- Уже упомянутые нами сократительные вакуоли, отвечающие за осморегуляцию, уровень давления внутри клетки.
- Пищеварительные вакуоли, они ответственны за переработку пищи. По сути, служат желудком для инфузории.
- Порошица, это отверстие в задней конечности инфузории, отвечающее за выход пищеварительных отходов. Догадайтесь сами аналогом, какого места нашего тела является порошица.
- Рот, представляющий собой углубление в оболочки клетки. С его помощью инфузория захватывает бактерии и прочую пищу, которая затем попадает в специальный канал цитофаринкс (аналог нашей глотки).
Обладая ртом, порошицей, пищеварительными вакуолями, инфузории практикуют голозойный тип питания, то есть захватывают органические частицы внутрь своего тела.
Так выглядит инфузория туфелька под микроскопом.
Интересный факт: дыхание инфузории туфельки осуществляется не с помощью рта, а всем телом: кислород через покровы клетки поступает в цитоплазму, где при его помощи происходит окисление органических веществ, превращение их в углекислый газ, воду и другие соединения.
Еще одной удивительной особенностью инфузории, которая ее делает «самой сложной из простейших» является наличие в ее клетке целых двух ядер. Одно из ядер большое, его зовут макронуклеусом, а второе маленькое соответственно зовется микронуклеусом. Оба ядра хранят одинаковую информацию, однако если большое ядро постоянно пребывает в работе и его информация постоянно эксплуатируется, а значит, может быть повреждена (подобно ходовым книгам в библиотеке). Если такое повреждение случается, то на этот случай как раз и предусмотрено второе маленькое ядро, служащее чем-то вроде резерва на случай сбоя основного ядра.
Как видите наша сегодняшняя героиня, инфузория туфелька, является самым совершенным среди простейших одноклеточных организмов.
Строение инфузории туфельки
Несмотря на внешнюю простоту строение инфузории отнюдь не простое. Снаружи она защищена тонкой эластичной оболочкой, которая также помогает телу инфузории сохранять постоянную форму. Защитные опорные волокна инфузории расположены в слое плотной цитоплазмы, которая прилегает к оболочке.
Помимо этого в цитоскелет инфузории входят различные микротрубочки, цистерны альвеолы, базальные тельца с ресничками, фибриллы и филамены и другие органоиды.
По причине наличия цитоскелета инфузория в отличие от амебы не может произвольно менять форму своего тела.
Схематический рисунок строения инфузории.
Класс инфузории туфельки
Также строение инфузории зависит от ее класса. Так различают два класса инфузории туфельки:
- ресничные инфузории,
- сосущие инфузории.
Далее подробно остановимся на них.
Ресничные инфузории
Названы так, поскольку их тело покрыто маленькими ресницами, которые также именуются цилиями.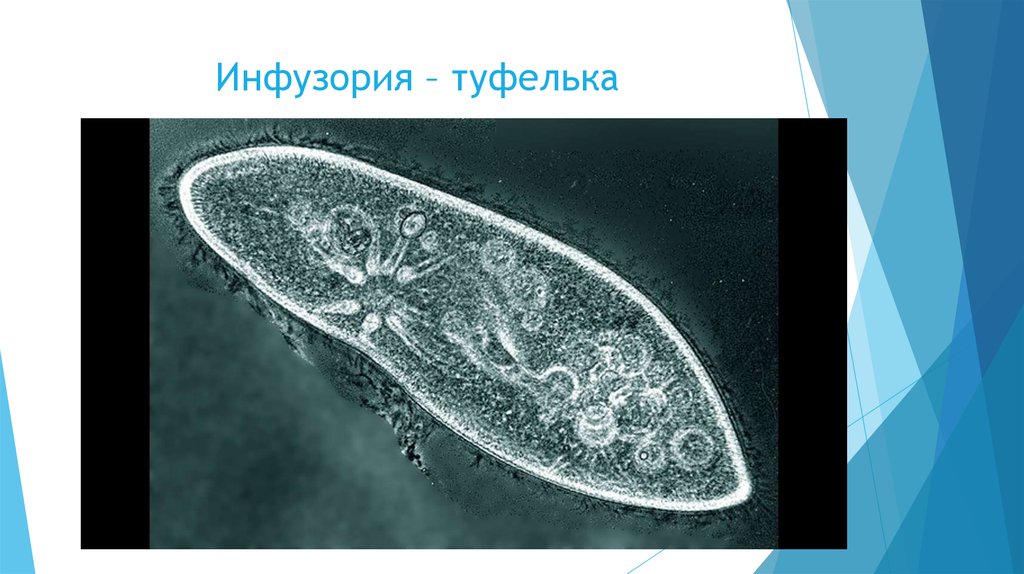 Длина ресницы составляет не более 0,1 микрометра. Ресницы могут, как распределятся равномерно по телу нашей простейшей красавицы, так и собираться в пучки, которые биологи называют «цирры». Сами ресницы представляют собой пучок фибрилл, которые являются нитевидными белками.
Длина ресницы составляет не более 0,1 микрометра. Ресницы могут, как распределятся равномерно по телу нашей простейшей красавицы, так и собираться в пучки, которые биологи называют «цирры». Сами ресницы представляют собой пучок фибрилл, которые являются нитевидными белками.
Каждая ресничная инфузория может иметь несколько тысяч таких вот ресниц. Передвижение инфузории также осуществляется при помощи ресниц.
Сосущие инфузории
Сосущие инфузории совсем не имеют не только ресничек, но и рта, глотки и пищеварительных вакуолей, столь характерных для их «волосатых» сородичей. Зато у них есть своеобразные щупальца, представляющие собой плазматические трубочки. Именно эти щупальца-трубочки у сосущих инфузорий выполняют функцию рта и глотки, так как захватывают и проводят питательные вещества в эндоплазму клетки.
Не имея ресниц сосущие инфузории не способны передвигаться. Впрочем, им это и не нужно, имея особую ножку-присоску, они прикрепляются к коже какого-нибудь краба или рыбы и на них живут.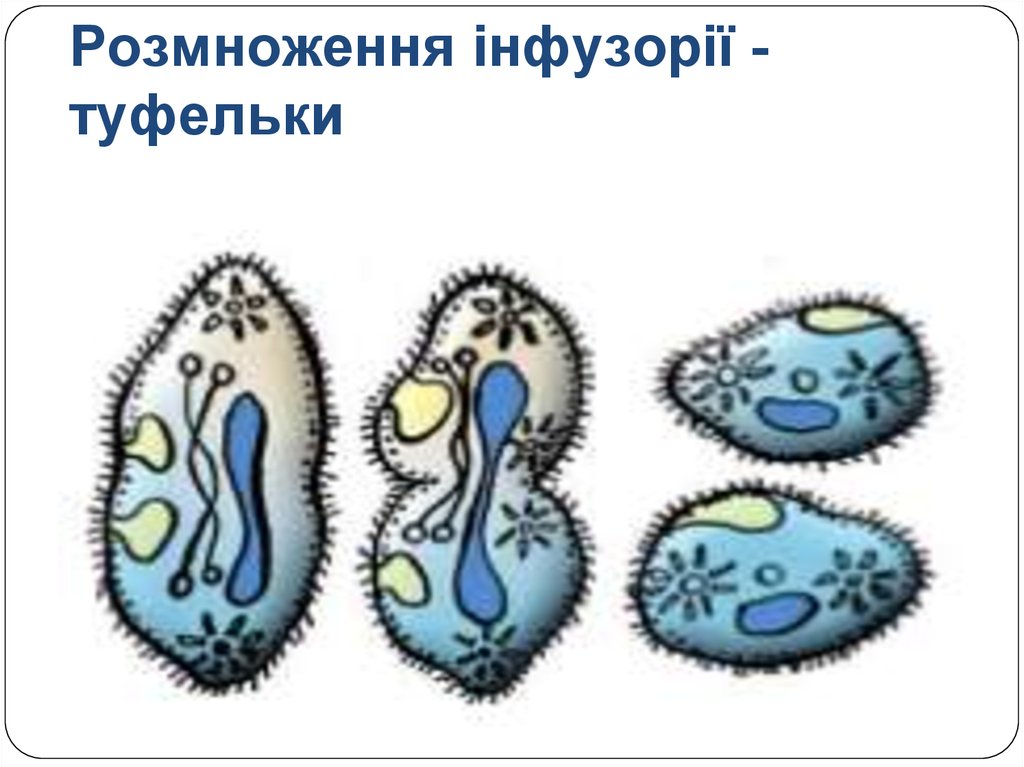 Сосущих инфузорий всего лишь несколько десятков видов, против тысячи видов их ресничных собратьев.
Сосущих инфузорий всего лишь несколько десятков видов, против тысячи видов их ресничных собратьев.
Среда обитания инфузории туфельки
Инфузории туфельки обычно живут в мелких пресных водоемах со стоячей водой и гниющей органикой. Стоячая вода им необходима, чтобы не преодолевать силу течения, которая их снесет, поэтому инфузорий нет в реках. В мелких водоемах Солнце достаточно прогревает воду, и гниющая органика служит источником их пищи. К слову по насыщенности того или иного водоема инфузориями можно судить о степени его загрязнения, чем их больше, тем более грязный водоем.
А вот соленую воду инфузории не любят, поэтому их нет в морях и океанах.
Питание инфузории туфельки
Чем питается инфузория туфелька? Питание инфузории зависит от ее класса. Так сосущие инфузории являются подлинными хищниками одноклеточного мира: источником их пищи служат другие более мелкие одноклеточные организмы, на свою беду проплывающие мимо. Своими щупальцами сосущие инфузории хватают других одноклеточных. Изначально жертва захватывается одним щупальцем, а потом «к столу» подходят и другие «собратья». Щупальца растворяют клеточную оболочку жертвы и поглощают ее внутрь.
Изначально жертва захватывается одним щупальцем, а потом «к столу» подходят и другие «собратья». Щупальца растворяют клеточную оболочку жертвы и поглощают ее внутрь.
А вот ресничная инфузория в этом плане «вегетарианка», источником ее пищи обычно служат одноклеточные водоросли, которые захватываются ротовым углублениями, оттуда они попадают в пищевод, а потом к пищеварительным вакуолям. Переработанная пища выбрасывается через порошицу.
Интересный факт: во рту ресничной инфузории также имеются реснички, которые колышась, создают течение, чем увлекают частицы пищи в ротовую область.
Размножение инфузории туфельки
Размножение инфузории может быть как половым, так и бесполым – посредством деления клетки.
- Половое размножение: при нем две инфузории сливаются боковыми поверхностями, при этом оболочки между слитыми поверхностями растворяются, и образуется своеобразный цитоплазматический мостик. Через этот мостик клетки обмениваются ядрами. Большие ядра при этом вовсе растворяются, а маленькие дважды делятся.
 Затем из полученных четырех ядер, три исчезает, а оставшееся ядро снова делится надвое. Обмен оставшимися ядрами происходит по цитоплазматическому мостику. Из полученного материала возникают вновь рожденные ядра, и большие, и маленькие. Затем инфузории расходятся друг с другом.
Затем из полученных четырех ядер, три исчезает, а оставшееся ядро снова делится надвое. Обмен оставшимися ядрами происходит по цитоплазматическому мостику. Из полученного материала возникают вновь рожденные ядра, и большие, и маленькие. Затем инфузории расходятся друг с другом. - Бесполое размножение инфузории посредством деления намного проще. При нем оба ядра клетки делятся на два, как и другие органоиды. Таким образом, из одной инфузории образуется две, каждая с полным набором необходимых органоидов.
Функции инфузории туфельки
Инфузории, как впрочем, и другие простейшие организмы выполняют ряд важных биологических функций. Они уничтожают многие виды бактерий, и сами в свою очередь служат пищей для мелких беспозвоночных организмов. Порой их специально разводят в качестве корма для мальков некоторых аквариумных рыбок.
Источник
Инфузории (ресничные)
Инфузории (ресничные) — наиболее сложноорганизованный, развитый тип простейших. Среди инфузорий можно встретить как свободноживущие (в морских и пресных водах), прикрепленные формы, так и паразитические — балантидий. Представители свободноживущих форм: инфузория-туфелька, инфузория-трубач.
Представители свободноживущих форм: инфузория-туфелька, инфузория-трубач.
Инфузория-туфелька
Инфузория-туфелька — вид инфузорий, который получил свое названия благодаря форме тела (клетки) в виде туфельки. Это связано с наличием у клетки плотной наружной оболочки — пелликулы. Излюбленное место обитания — пресные водоемы со стоячей водой, ее легко можно обнаружить и в обычном аквариуме, взяв пробу воды на микроскопию.
Органы движения у инфузории — реснички, которые покрывают тело полностью или частично. Совершая ими волнообразные движения, инфузория начинает вращаться и подобно винту вкручивается в толщу воду (штопорообразное движение).
За счет наличия плотной пелликулы, у инфузории имеется достаточно сложноустроенная пищеварительная система — по сравнению с амебой, у которой нет плотной оболочки, а вещества могут захватываться и выделяться в любом участке поверхности клетки. У инфузории такого хаоса, как у амебы, нет — для всего отведено свое место.
Ближе к переднему концу тела на поверхности инфузории имеется углубление — клеточный рот, также называемый цитостом (др. -греч. κύτος «вместилище» и στόμα — «рот»), служит местом поступления твердых пищевых частиц, бактерий.
-греч. κύτος «вместилище» и στόμα — «рот»), служит местом поступления твердых пищевых частиц, бактерий.
Сужаясь, клеточный рот переходит в клеточную глотку (цитофаринкс — от греч. kytos – вместилище, клетка и pharyngos – глотка). На дне глотки пищевые частицы попадают в пищеварительные вакуоли (фагосомы), в которых благодаря ферментам перевариваются. Расщепленные пищевые частицы поступают в цитоплазму и используются клеткой для своих нужд.
Непереваренные остатки пищи удаляются с помощью экзоцитоза в специально отведенном месте, где прерывается пелликула — порошица (цитопиг).
Дыхательная система отсутствует, поэтому дыхание (поглощение кислорода и выделение углекислого газа) у инфузории-туфельки осуществляется диффузно всей поверхностью клетки. При низкой концентрации кислорода в воде, инфузория способна существовать за счет гликолиза (от греч. glykys-сладкий и lysis — разложение) — бескислородного расщепления глюкозы.
Продукты азотистого обмена удаляются с помощью сократительных вакуолей.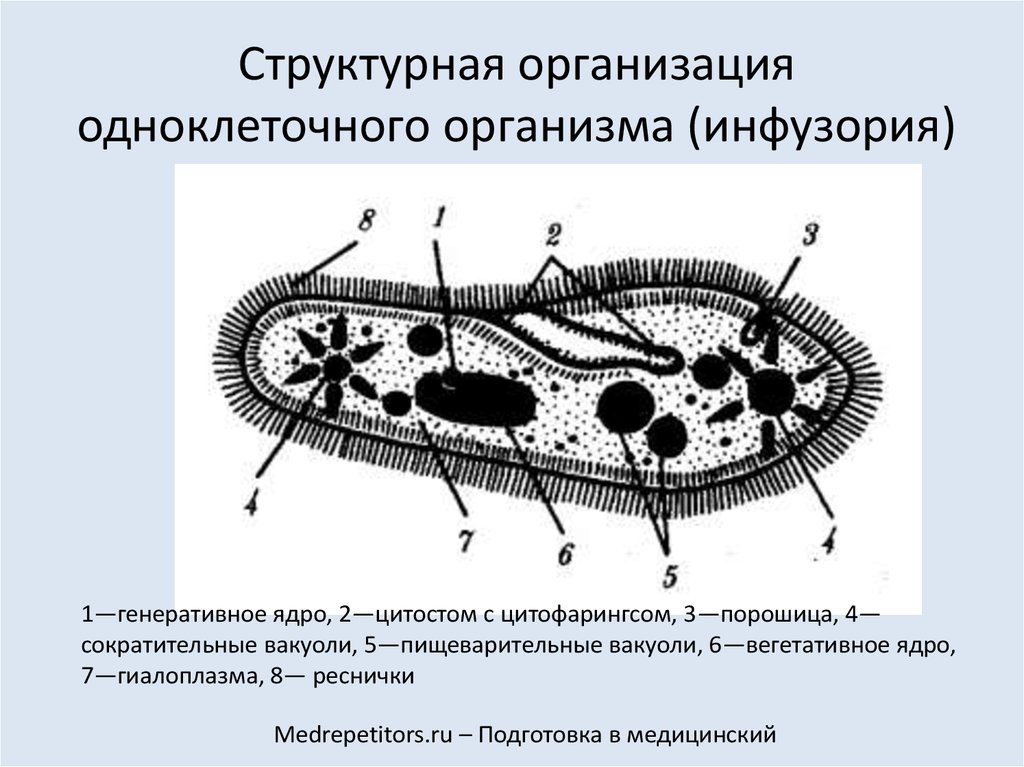 Этим же вакуолям принадлежит крайне важная функция: регуляция осмотического давления клетки — поддержание гомеостаза. В процессе работы сократительной вакуоли из клетки удаляется избыток воды, что препятствует разрыву клетки.
Этим же вакуолям принадлежит крайне важная функция: регуляция осмотического давления клетки — поддержание гомеостаза. В процессе работы сократительной вакуоли из клетки удаляется избыток воды, что препятствует разрыву клетки.
Если бы не сократительные вакуоли, удаляющие избыток воды, клетка лопнула, как переполненный воздушный шарик.
Эта тема заслуживает нашего особенного, пристального внимания. У инфузории-туфельки имеются два ядра: большое — вегетативное (макронуклеус), которое отвечает за процессы жизнедеятельности в клетке, и малое — генеративное (микронуклеус), основная функция которого заключается в процессе размножения клетки.
Для инфузорий характерно бесполое размножение, путем поперечного деления надвое. Заметьте, именно — поперечного, а не продольного, которое присуще эвглене зеленой. Под действием неблагоприятных факторов у инфузорий запускается механизм конъюгации — полового процесса.
Конъюгация не является в привычном смысле «половым размножением», так как в результате конъюгации не увеличивается число особей вида, а происходит только перекомбинирование генетического материала и обмен им между двумя инфузориями.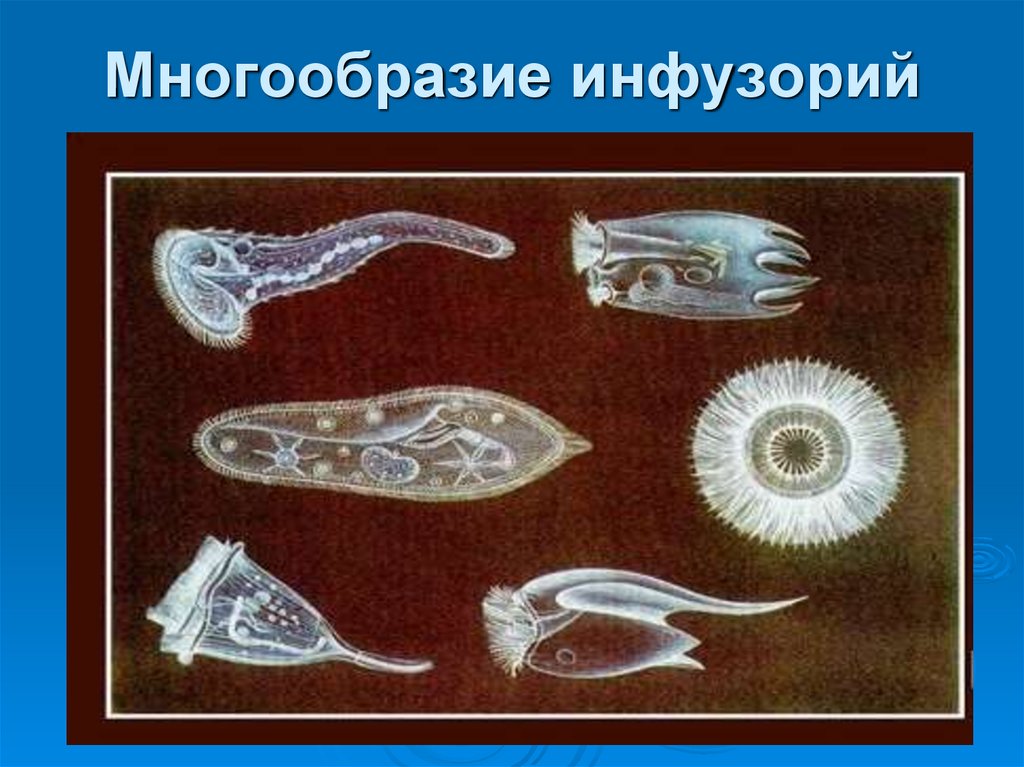 В ходе конъюгации не образуются гаметы, и уже очевидно — не образуется зиготы.
В ходе конъюгации не образуются гаметы, и уже очевидно — не образуется зиготы.
При конъюгации две инфузории соединяются в области клеточных ртов (цитостом), между ними возникает цитоплазматический мостик. Вегетативное ядро (полиплоидное) каждой клетки растворяется, а генеративное (2n) мейотически делится, в результате образуется 4 ядра (n), 3 из которых растворяются, а одно оставшееся (n) делится митотически на мужское (n) и женское (n) ядро.
Женское ядро каждой инфузории остается на месте, а мужское (n) по цитоплазматическому мостику перемещается в клетку партнера, где сливается с женским (n) ядром клетки-партнера.
В результате в каждой клетке сливается женское ядро (которое никуда не перемещалось) с мужским ядром клетки-партнера, переместившимся по цитоплазматическому мостику. При слиянии образуется синкарион.
Это и есть половой процесс у инфузорий, в результате него происходит обмен генетической информацией между клетками.
Балантидий
Балантидий — вид инфузорий, являющийся самым крупным из патогенных кишечных простейших. Возбудитель балантидиаза. Форма клеток яйцевидная, покрыты ресничками. Ядерный аппарат типичен для инфузорий, состоит из вегетативного и генеративного ядер.
Возбудитель балантидиаза. Форма клеток яйцевидная, покрыты ресничками. Ядерный аппарат типичен для инфузорий, состоит из вегетативного и генеративного ядер.
Паразитирует балантидий в толстой кишке, клинически заболевание протекает по типу колита (от греч. kolon — толстая кишка) — воспаления толстой кишки, и энтерита (от греч. enteron — кишка) — воспаления тонкой кишки.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Инфузория-туфелька
Инфузория-туфелька обитает в мелких стоячих водоёмах. Это одноклеточное животное длиной 0,5 мм имеет веретеновидную форму тела, отдалённо напоминающую туфлю. Инфузории все время находятся в движении, плавая тупым концом вперёд. Скорость передвижения этого животного достигает 2,5 мм в секунду. На поверхности тела у них имеются органоиды движения — реснички. В клетке два ядра: большое ядро отвечает за питание, дыхание, движение, обмен веществ; малое ядро участвует в половом процессе.
Инфузории все время находятся в движении, плавая тупым концом вперёд. Скорость передвижения этого животного достигает 2,5 мм в секунду. На поверхности тела у них имеются органоиды движения — реснички. В клетке два ядра: большое ядро отвечает за питание, дыхание, движение, обмен веществ; малое ядро участвует в половом процессе.
Строение инфузории туфельки
Организм инфузории устроен сложнее. Тонкая эластичная оболочка, покрывающая инфузорию снаружи, сохраняет постоянную форму её тела. Этому же способствуют хорошо развитые опорные волоконца, которые находятся в прилегающем к оболочке слое цитоплазме. На поверхности тела инфузории расположено около 15 000 колеблющихся ресничек. У основания каждой реснички лежит базальное тельце. Движение каждой реснички состоит из резкого взмаха в одном направлении и более медленного, плавного возвращения к исходному положению. Реснички колеблются примерно 30 раз в секунду и, словно вёсла, толкают инфузорию вперёд. Волнообразное движение ресничек при этом согласованно. Когда инфузория-туфелька плывёт, она медленно вращается вокруг продольной оси тела.
Когда инфузория-туфелька плывёт, она медленно вращается вокруг продольной оси тела.
Процессы жизнедеятельности
Питание
Туфелька и некоторые другие свободно живущие инфузории питаются бактериями и водорослями.
Реакция инфузории-туфельки на пищу
Тонкая эластичная оболочка, (клеточная мембрана) покрывающая инфузорию снаружи, сохраняет постоянную форму тела. На поверхности тела расположено около 15 тысяч ресничек. На теле имеется углубление — клеточный рот, который переходит в клеточную глотку. На дне глотки пища попадает в пищеварительную вакуоль. В пищеварительной вакуоле пища переваривается в течение часа, вначале при кислой, а затем при щелочной реакции. Пищеварительные вакуоли перемещаются в теле инфузории током цитоплазмы. Не переваренные остатки выбрасываются наружу в заднем конце тела через особую структуру — порошицу, расположенную позади ротового отверстия.
Дыхание
Дыхание происходит через покровы тела. Кислород поступает в цитоплазму через всю поверхность тела и окисляет сложные органические вещества, в результате чего они превращаются в воду, углекислый газ и некоторые другие соединения. При этом освобождается энергия, которая необходима для жизни животного. Углекислый газ в процессе дыхания удаляется через всю поверхность тела.
При этом освобождается энергия, которая необходима для жизни животного. Углекислый газ в процессе дыхания удаляется через всю поверхность тела.
Выделение
В организме инфузории-туфельки находятся две сократительные вакуоли, которые располагаются у переднего и заднего концов тела. В них собирается вода с растворёнными веществами, образующимися при окислении сложных органических веществ. Достигнув предельной величины, сократительные вакуоли подходят к поверхности тела, и их содержимое изливается наружу. У пресноводных одноклеточных животных через сократительные вакуоли удаляется избыток воды, постоянно поступающей в их тело из окружающей среды.
Раздражимость
Инфузории-туфельки собираются к скоплениями бактерий в ответ на действие выделяемых ими веществ, но уплывают от такого раздражителя, как поваренная соль.
Раздражимость — свойство всех живых организмов отвечать на действия раздражителей — света, тепла, влаги, химических веществ, механических воздействий. Благодаря раздражимости одноклеточные животные избегают неблагоприятных условий, находят пищу, особей своего года.
Благодаря раздражимости одноклеточные животные избегают неблагоприятных условий, находят пищу, особей своего года.
Размножение
Бесполое
Инфузория обычно размножается бесполым путём — делением надвое. Ядра делятся на две части, и в каждой новой инфузории оказывается по одному большому и по одному малому ядру. Каждая из двух дочерних получает часть органоидов, а другие образуются заново.
Половое
При недостатке пищи или изменении температуры инфузории переходят к половому размножению, а затем могут превратиться в цисту.
При половом процессе увеличения числа особей не происходит. Две инфузории временно соединяются друг с другом. На месте соприкосновения оболочка растворяется, и между животными образуется соединительный мостик. Большое ядро каждой инфузории исчезает. Малое ядро дважды делится. В каждой инфузории образуются четыре дочерних ядра. Три из них разрушаются, а четвёртое снова делится. В результате в каждой остаётся по два ядра. По цитоплазматическому мостику происходит обмен ядрами, и там сливается с оставшимся ядром. Вновь образовавшиеся ядра формируют большое и малое ядра, и инфузории расходятся. Такой половой процесс называется конъюгацией. Он длится около 12 часов. Половой процесс ведёт к обновлению, обмену между особями и перераспределению наследственного (генетического) материала, что увеличивает жизнестойкость организмов.
Вновь образовавшиеся ядра формируют большое и малое ядра, и инфузории расходятся. Такой половой процесс называется конъюгацией. Он длится около 12 часов. Половой процесс ведёт к обновлению, обмену между особями и перераспределению наследственного (генетического) материала, что увеличивает жизнестойкость организмов.
Источник
Инфузория туфелька это бактерия или животное или растение или гриб
Инфузория-туфелька – вид простейших одноклеточных животных из класса ресничных инфузорий типа инфузории. Свое название данный вид получил за внешнее сходство с подошвой туфельки.
Инфузории-туфельки обитают в пресных водоемах любого типа со стоячей водой и наличием в воде массы разлагающихся органических веществ. Также данные организмы встречаются в аквариумах. В этом можно убедиться, отобрав пробы воды с илом из аквариума и рассмотрев их под микроскопом.
В строении инфузории-туфельки отмечаются характерные особенности. Это относительно крупный организм, размеры тела достигают 0,5 мм. Минимальные размеры особей – от 0,1 мм. Форма тела, как уже было отмечено, напоминает туфельку. Внешней оболочкой этого простейшего является наружная мембрана. Под ней находится пелликула – плотный слой цитоплазмы с уплощенными мембранными цистернами (альвеолами), микротрубочками и другими составляющими цитоскелета.
Минимальные размеры особей – от 0,1 мм. Форма тела, как уже было отмечено, напоминает туфельку. Внешней оболочкой этого простейшего является наружная мембрана. Под ней находится пелликула – плотный слой цитоплазмы с уплощенными мембранными цистернами (альвеолами), микротрубочками и другими составляющими цитоскелета.
Передвижение инфузории-туфельки возможно, благодаря волнообразным движениям ресничек. Так она плывет притупленным краем вперед со скоростью примерно 2 мм/с. В основном, инфузория-туфелька передвигается в одной плоскости, при этом в толще одной массы особь может вращаться вокруг продольной оси. Простейшие меняют направления движения, благодаря изгибам своего тела. Если инфузория сталкивается с препятствием, она моментально начинает двигаться в противоположную сторону.
Чем питается инфузория-туфелька? Питание данного простейшего имеет характерные особенности. Основой пищевого рациона инфузории-туфельки являются бактерии, скопления которых привлекают инфузорию выделением особых химических веществ. Также инфузории могут проглатывать другие взвешенные в воде частицы, даже не имеющие особой питательной ценности. В организме простейшего различают клеточный рот, переходящий в клеточную глотку. Возле рта находятся специальные реснички, собранные в сложные комплексы. При волнообразных движениях ресничек данного типа пища с потоком воды попадает в глотку. У основания глотки формируется крупная пищеварительная вакуоль. Эта вакуоль, как и все последующие новообразованные, мигрируют в цитоплазме организма особи по определенному «пути» — спереди назад, а затем сзади кпереди (как бы по кругу), при этом крупная вакуоль распадается на более мелкие. Таким образом, ускоряется всасывание питательных веществ. Переваренные вещества поступают в цитоплазму, где используются для нужд организма. Ненужные вещества выводятся в окружающую среду через порошицу в задней части клетки – участок с недоразвитой пелликулой.
Также инфузории могут проглатывать другие взвешенные в воде частицы, даже не имеющие особой питательной ценности. В организме простейшего различают клеточный рот, переходящий в клеточную глотку. Возле рта находятся специальные реснички, собранные в сложные комплексы. При волнообразных движениях ресничек данного типа пища с потоком воды попадает в глотку. У основания глотки формируется крупная пищеварительная вакуоль. Эта вакуоль, как и все последующие новообразованные, мигрируют в цитоплазме организма особи по определенному «пути» — спереди назад, а затем сзади кпереди (как бы по кругу), при этом крупная вакуоль распадается на более мелкие. Таким образом, ускоряется всасывание питательных веществ. Переваренные вещества поступают в цитоплазму, где используются для нужд организма. Ненужные вещества выводятся в окружающую среду через порошицу в задней части клетки – участок с недоразвитой пелликулой.
В клетке инфузории-туфельки имеются две сократительные вакуоли спереди и сзади тела.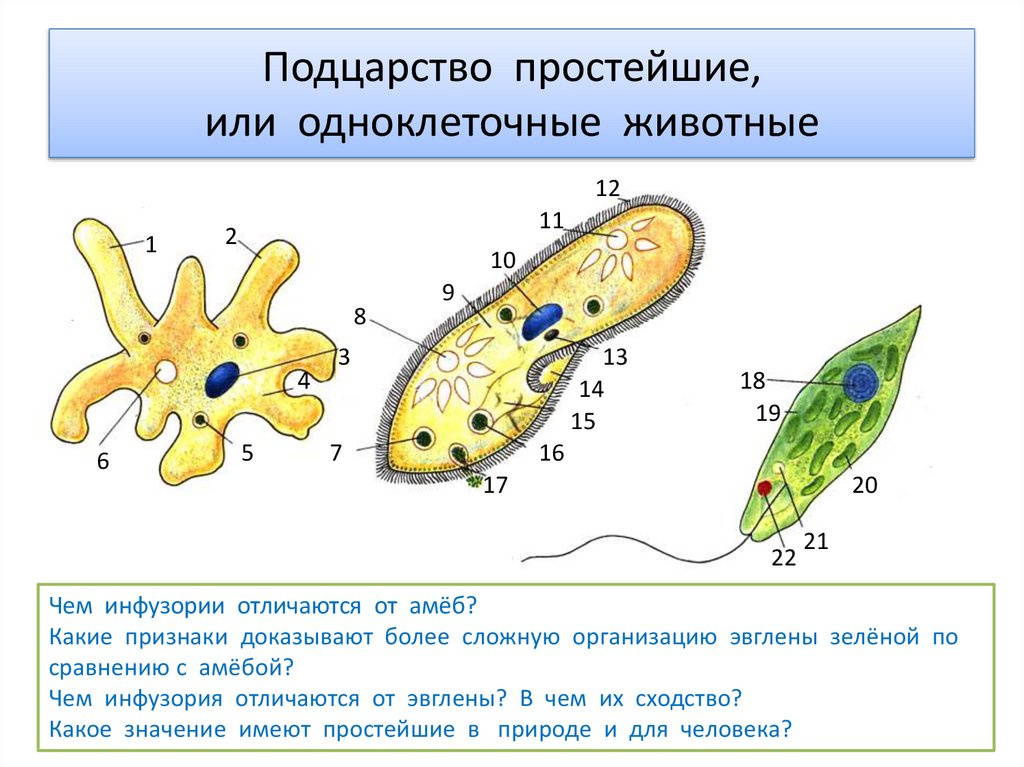 В структуре такой вакуоли различают резервуар и канальцы. Через канальцы вода поступает из цитоплазмы в резервуар, из которого выталкивается наружу через пору. Благодаря цитоскелету из микротрубочек весь данный комплекс постоянно находится в определенном участке клетки. Главная функция сократительных вакуолей – осморегуляторная. Черех них из клетки удаляется избыточное количество воды, а также продукты азотистого обмена.
В структуре такой вакуоли различают резервуар и канальцы. Через канальцы вода поступает из цитоплазмы в резервуар, из которого выталкивается наружу через пору. Благодаря цитоскелету из микротрубочек весь данный комплекс постоянно находится в определенном участке клетки. Главная функция сократительных вакуолей – осморегуляторная. Черех них из клетки удаляется избыточное количество воды, а также продукты азотистого обмена.
Дыхание инфузории-туфельки происходит через всю поверхность тела. А при пониженной концентрации кислорода в воде инфузория живет за счет гликолиза.
Два ядра инфузории-туфельки имеют разное строение и выполняют различные функции. Малое ядро диплоидное, имеет округлую форму; большое ядро полиплоидное, имеет бобовидную форму. Малое ядро отвечает за половое размножение, а большое ядро руководит синтезом всех белков клетки инфузории-туфельки.
Бесполое размножение происходит путем деления клетки пополам. Половое размножение осуществляется посредством конъюгации. Две туфельки соединяются и при сложных превращениях ядер образуются новые особи.
Две туфельки соединяются и при сложных превращениях ядер образуются новые особи.
Источник
Инфузория туфелька. Описание, особенности, строение и размножение инфузории туфельки
Инфузория туфелька — обобщающее понятие. За названием скрываются 7 тысяч видов. У всех постоянная форма тела. Она напоминает подошву туфли. Отсюда и название простейшего. Еще все инфузории владеют осморегуляцией, то есть регулируют давление внутренней среды организма. Для этого служат две сократительные вакуоли. Они сжимаются и разжимаются, выталкивая излишки жидкости из туфельки.
Описание и особенности организма
Инфузория туфелька — простейшее животное. Соответственно, оно одноклеточное. Однако в клетке этой есть все, чтобы дышать, размножаться, питаться и выводит отходы наружу, двигаться. Это список функций животных. Значит, к ним относятся и туфельки.
Простейшими одноклеточных называют за примитивное в сравнение с прочими животными устройство. Среди одноклеточных даже есть формы, относимые учеными как к животным, так и к растениям. Пример — эвглена зеленая. В ее теле есть хлоропласты и хлорофилл — пигмент растений. Эвглена осуществляет фотосинтез и почти неподвижна днем. Однако ночью одноклеточное переходит на питание органикой, твердыми частицами.
Среди одноклеточных даже есть формы, относимые учеными как к животным, так и к растениям. Пример — эвглена зеленая. В ее теле есть хлоропласты и хлорофилл — пигмент растений. Эвглена осуществляет фотосинтез и почти неподвижна днем. Однако ночью одноклеточное переходит на питание органикой, твердыми частицами.
Инфузория туфелька и эвглена зеленая стоят на разных полюсах цепи развития простейших. Героиня статьи признана среди них наиболее сложным организмом. Организмом, кстати, туфелька является, поскольку имеет подобие органов. Это элементы клетки, отвечающие за те или иные функции. У инфузории есть отсутствующие у прочих простейших. Это и делает туфельку передовиком среди одноклеточных.
К передовым органеллам инфузории относятся:
- Сократительные вакуоли с проводящими канальцами. Последние служат своеобразными сосудами. По ним в резервуар, коим является сама вакуоль, поступают вредные вещества. Они перемещаются из протоплазмы — внутреннего содержимого клетки, включающего цитоплазму и ядро.
Тело инфузории туфельки содержит две сократительные вакуоли. Накапливая токсины, они выбрасывают их вместе с излишками жидкости, попутно поддерживая внутриклеточное давление.
- Пищеварительные вакуоли. Они, подобно желудку, перерабатывают пищу. Вакуоль при этом движется. В момент подхода органеллы к задней оконечности клетки, полезные вещества уже усвоены.
- Порошица. Это отверстие в задней оконечности инфузории, подобное анальному. Функция у порошицы такая же. Через отверстие из клетки выводятся отходы пищеварения.
- Рот. Это углубление в оболочке клетки захватывает бактерии и прочую пищу, проводя в цитофаринкс — тонкий каналец, заменяющий глотку. Имея ее и рот, туфелька практикует голозойный тип питания, то есть захват органических частиц внутрь тела.
Еще совершенным простейшим инфузорию делают 2 ядра. Одно из них большое, именуется макронуклеусом. Второе ядро малое — микронуклеус. Информация, хранящаяся в обоих органеллах идентична. Однако в микронуклеусе она не тронута. Информация макронуклеуса рабочая, постоянно эксплуатируется. Поэтому возможны повреждения каких-то данных, как книг в читальном зале библиотеки. В случае таких сбоев резервом служит микронуклеус.
Однако в микронуклеусе она не тронута. Информация макронуклеуса рабочая, постоянно эксплуатируется. Поэтому возможны повреждения каких-то данных, как книг в читальном зале библиотеки. В случае таких сбоев резервом служит микронуклеус.
Большое ядро инфузории имеет форму боба. Малая органелла шаровидная. Органоиды инфузории туфельки хорошо видны под увеличением. Все простейшее в длину не превышает 0,5 миллиметра. Для простейших это гигантизм. Большинство представителей класса не превышают в длину 0,1 миллиметра.
Строение инфузории туфельки
Строение инфузории туфельки отчасти зависит от ее класса. Их два. Первый называется ресничным, поскольку его представители покрыты ресничками. Это волосковидные структуры, иначе именуются цилиями. Их диаметр не превышает 0,1 микрометра. Реснички на теле инфузории могут распределяться равномерно или собираться в своеобразные пучки — цирры. Каждая ресничка — пучок фибрилл. Это нитевидные белки. Два волокна являются стержнем реснички, еще 9 располагаются по периметру.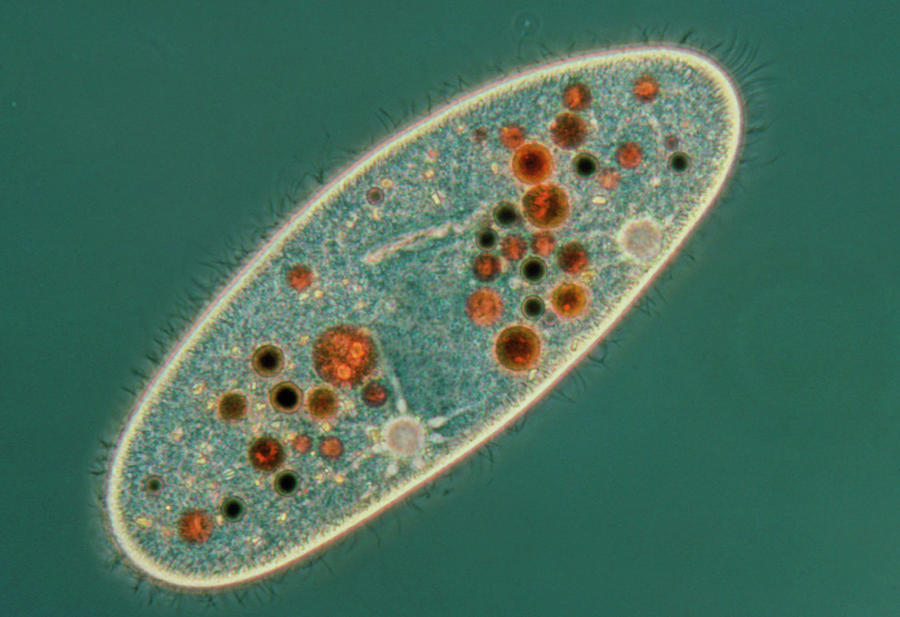
Когда обсуждается реснитчатый класс, инфузории туфельки могут иметь несколько тысяч ресничек. В противовес встают сосущие инфузории. Они представляют отдельный класс, лишены ресничек. Нет у сосущих туфелек и рта, глотки, пищеварительных вакуолей, характерных для «волосатых» особей. Зато, у сосущих инфузорий есть подобие щупалец. Таковых видов несколько десятков против многих тысяч реснитчатых.
Щупальца сосущих туфелек — полые плазматические трубочки. Они проводят питательные вещества в эндоплазму клетки. Питанием служат другие простейшие. Иначе говоря, сосущие туфельки — хищники. Ресничек сосущие инфузории лишены, поскольку не двигаются. У представителей класса есть особая ножка-присоска. С ее помощью одноклеточные закрепляются на ком-то, к примеру, крабе или рыбе, или внутри их и других простейших. Реснитчатые же инфузории активно передвигаются. Собственно за этим и нужны цилии.
Среда обитания простейшего
Обитает героиня статьи в пресных, мелких водоемах со стоячей водой и обилием разлагающейся органики. Во вкусах сходятся инфузория туфелька, амеба. Стоячая вода им нужна, дабы не преодолевать течение, которое попросту снесет. Мелководье гарантирует прогрев, необходимый для активности одноклеточных. Обилие же гниющей органики — пищевая база.
Во вкусах сходятся инфузория туфелька, амеба. Стоячая вода им нужна, дабы не преодолевать течение, которое попросту снесет. Мелководье гарантирует прогрев, необходимый для активности одноклеточных. Обилие же гниющей органики — пищевая база.
По насыщенности воды инфузориями, можно судить о степени загрязненности пруда, лужи, старицы. Чем больше туфелек, тем больше питательной базы для них — разлагающейся органики. Зная интересы туфелек, их можно разводить в обычных аквариуме, банке. Достаточно положить туда сено и залить прудовой водой. Скошенная трава послужит той самой разлагающейся питательной средой.
Нелюбовь инфузорий к соленой воде наглядна, при помещении в обычную частиц поваренной соли. Под увеличением видно, как одноклеточные уплывают подальше от нее. Если же простейшие засекают скопление бактерий, напротив, направляются к ним. Это именуется раздражимостью. Сие свойство помогает животным избегать неблагоприятных условий, находить пищу и других особей своего рода.
Питание инфузории
Питание инфузории зависит от ее класса. Хищные сосальщики орудуют щупальцами. К ним прилипают, присасываются, проплывающие мимо одноклеточные. Питание инфузории туфельки осуществляется за счет растворения клеточной оболочки жертвы. Пленка разъедается в местах контакта со щупальцами. Изначально жертва, как правило, захватывается одним отростком. Прочие щупальца «подходят к уже накрытому столу».
Реснитчатая форма инфузории туфельки питается одноклеточными водорослями, захватывая их ротовым углублением. Оттуда еда попадает в пищевод, а затем, в пищеварительную вакуоль. Она закрепляется на коне «глотки», отцепляясь от нее каждые несколько минут. После, вакуоль проходит по часовой стрелке к заду инфузории. Во время пути цитоплазмой усваиваются полезные вещества пищи. Отходы выбрасываются в порошицу. Это отверстие, подобное анальному.
Во рту инфузории тоже есть реснички. Колышась, они создают течение. Оно увлекает частицы пищи в ротовую полость. Когда пищеварительная вакуоль перерабатывает еду, образуется новая капсула. Она тоже стыкуется с глоткой, получает пищу. Процесс цикличен. При комфортной для инфузории температуре, а это около 15 градусов тепла, пищеварительная вакуоль образуется каждые 2 минуты. Это указывает на скорость обмена веществ туфельки.
Когда пищеварительная вакуоль перерабатывает еду, образуется новая капсула. Она тоже стыкуется с глоткой, получает пищу. Процесс цикличен. При комфортной для инфузории температуре, а это около 15 градусов тепла, пищеварительная вакуоль образуется каждые 2 минуты. Это указывает на скорость обмена веществ туфельки.
Размножение и продолжительность жизни
Инфузория туфелька на фото может быть в 2 раза больше, чем по стандарту. Это не зрительная иллюзия. Дело в особенностях размножения одноклеточного. Процесс бывает двух типов:
- Половой. В этом случае две инфузории сливаются боковыми поверхностями. Оболочка здесь растворяется. Получается соединительный мостик. Через него клетки меняются ядрами. Большие растворяются вовсе, а малые дважды делится. Три из полученных ядер исчезают. Оставшееся снова делится. Два получившихся ядра переходят в соседнюю клетку. Из нее тоже выходят две органеллы. На постоянном месте одна из них преобразуется в большое ядро.
- Бесполый.
 Иначе именуется делением. Ядра инфузории членятся, каждое на два. Клетка делится. Получается две. Каждая — с полным набором ядер и частичным прочих органелл. Они не делятся, распределяются меж вновь образовавшимися клетками. Недостающие органоиды образуются уже после отсоединения клеток друг от друга.
Иначе именуется делением. Ядра инфузории членятся, каждое на два. Клетка делится. Получается две. Каждая — с полным набором ядер и частичным прочих органелл. Они не делятся, распределяются меж вновь образовавшимися клетками. Недостающие органоиды образуются уже после отсоединения клеток друг от друга.
Как видно, при половом размножении число инфузорий остается прежним. Это называется конъюгацией. Происходит лишь обмен генетической информацией. Число клеток остается прежним, но сами простейшие по факту получаются новыми. Генетический обмен делает инфузорий живучее. Поэтому к половому размножению туфельки прибегают в неблагоприятных условиях.
Источник
Инфузории туфельки | Мир аквариума
Инфузории туфельки простейшие одноклеточные реснитчатые микроорганизмы. Распространены повсеместно, а также повсеместно являются важнейшим звеном пищевой цепочки любого биоценоза. Для аквариумиста эти простейшие представляют интерес как кормовая база для только что появившихся из икринок мальков.
| классификация | |
|---|---|
| Домен | Эукариоты |
| Тип | Infusoria |
| Класс | Ciliatea |
| Отряд | Hymenostomatida |
| Семейство | Parameciidae |
| Род | Парамеции |
| Вид | Paramecium caudatum |
Латынь: Paramecium caudatum
Безусловно, на сегодняшний день существует большое количество производителей корма для домашних животных и аквариумных рыбок в том числе. [sam_ad id=»15″ codes=»true»] Среди их продукции есть так же корма, которые позиционируются как корма для мальков. Но все же не всех мальков удовлетворят комбикорма производимые промышленностью. Для мальков некоторых видов необходим живой пищевой объект определенных размеров. Такая мелкая живая пища, как Paramecium caudatum новорожденным малькам нужна всего несколько первых дней их жизни. Но наличие такого корма в данном случае является вопросом жизни и смерти.
Инфузории туфельки получили свое название за внешнее сходство формы их тела с дамской туфелькой. Длина тела достигает 0,3 миллиметра. Все тело их покрыто сократительными ресничками, выполняющими роль своеобразного движителя, благодаря которому животное перемещается очень быстро. За 1 секунду оно преодолевает расстояние равное
Внутреннее строение2-2,5 миллиметрам. Это составляет расстояние приблизительно в 10 – 15 раз больше чем длина тела животного. Эта же подвижность может быть причиной неудач выкармливания ими мальков некоторых видов, которые из-за своей медлительности не могут угнаться за столь быстрой едой. Сама героиня нашего повествования питается бактериями, грибами и одноклеточными водорослями. Пищевые частицы не заглатываются в обычном смысле этого слова, а направляются слаженными движениями ресничек к ротовому отверстию, переходящему в глотку.
Еще из особенностей поведения, которые необходимо учитывать при размножении этого вида корма необходимо отметить чувствительность к свету. Электрическому току. Оптимальная температура для данного вида 22-26 градусов по Цельсию. Жесткость воды желательна до 10°dH.
Что же нам предстоит сделать?
- 1. Найти водоем, населенный нужными нам микроорганизмами.
- 2. Выделить из этого водоема чистую культуру.
- 3. Создать оптимальные условия для размножения.
- 4. Поддерживать жизнеспособное состояние колонии микроорганизмов.
- 5. Перед кормлением мальков очистить культуру от балласта.
Поскольку интересующее нас живое существо – пресноводное то идем к ближайшему пресноводному водоему… к аквариуму. Воду набираем у дна, желательно чтобы, где то неподалеку был слегка разложившийся листочек водного растения. Если вам повезет, то на улицу выходить не придется. Проверить свое везение и ускорить получение ответа на вопрос: «повезло или нет?», можно с помощью микроскопа или мощной лупы. Paramecium caudatum хорошо видны при малом увеличении микроскопа. Берем трехлитровый бутыль, наполняем его на половину кипяченой отстоянной водой. Капаем туда 1 – 3 капли молока. Вносим в бутыль воду из аквариума, которая предположительно содержит Paramecium caudatum. Внутрь бутыли опускаем распылитель от компрессора и делаем очень слабую аэрацию. Температуру Воды стараемся держать на уровне 24 — 26 градусов по Цельсию. Впрочем, при комнатной температуре тоже можно получить вполне удовлетворительные результаты. Примерно через двое – трое суток скопления животных можно будет заметить невооруженным глазом.
Если вам повезет, то на улицу выходить не придется. Проверить свое везение и ускорить получение ответа на вопрос: «повезло или нет?», можно с помощью микроскопа или мощной лупы. Paramecium caudatum хорошо видны при малом увеличении микроскопа. Берем трехлитровый бутыль, наполняем его на половину кипяченой отстоянной водой. Капаем туда 1 – 3 капли молока. Вносим в бутыль воду из аквариума, которая предположительно содержит Paramecium caudatum. Внутрь бутыли опускаем распылитель от компрессора и делаем очень слабую аэрацию. Температуру Воды стараемся держать на уровне 24 — 26 градусов по Цельсию. Впрочем, при комнатной температуре тоже можно получить вполне удовлетворительные результаты. Примерно через двое – трое суток скопления животных можно будет заметить невооруженным глазом.
Если не повезло, ваш аквариум оказался стерилен, то за Парамецией придется отправиться к ближайшему пресноводному стоячему водоему (озеро, пруд, большая редко пересыхающая лужа). Не забудьте прихватить с собой три небольших стеклянных сосуда. Почему три? Вам придется взять три пробы из мест наиболее вероятного нахождения искомого объекта. А именно.
Почему три? Вам придется взять три пробы из мест наиболее вероятного нахождения искомого объекта. А именно.
Первую порцию аккуратно зачерпнем у самой поверхности воды.
Вторую банку заполним водой из лужи и положим в нее несколько поднятых со дна органических остатков – подгнившие веточки, прошлогодние листья.
С третьей банкой поступим так же как со второй только вместо веточек и листьев насыплем в нее пригоршню ила взятого со дна пруда.
Теперь домой. Ставим на несколько дней эти три образца в подходящие условия. Через несколько дней Paramecium caudatum размножившись, образуют скопления заметные невооруженным глазом. Взяв воду из мест наибольшего их скопления пипеткой, переносим ее в заранее подготовленную банку, как описано выше.
Перед тем как зарядить культуру желательно все же ее проконтролировать под микроскопом. Если набранная вами капля воды содержит посторонние микроорганизмы, то желательно от них избавиться. Для этого на предметное стекло нужно капнуть еще одну каплю чистой воды. Две капли соединяются водяным мостиком, созданным с помощью зубочистки или заточенной спички. Более проворные инфузории туфельки опередят своих конкурентов в марафоне к чистой воде. После этого мостик можно разрушить, аккуратно вытерев первую каплю салфеткой. Вторую каплю пипеткой переносим в заранее подготовленный сосуд с бактериями.
Две капли соединяются водяным мостиком, созданным с помощью зубочистки или заточенной спички. Более проворные инфузории туфельки опередят своих конкурентов в марафоне к чистой воде. После этого мостик можно разрушить, аккуратно вытерев первую каплю салфеткой. Вторую каплю пипеткой переносим в заранее подготовленный сосуд с бактериями.
Если микроскопа нет, то необходимо хотя бы проследить чтобы образец не содержал мелких ракообразных которые тоже не прочь полакомится Парамециями.
Для размножения бактерий, которыми собственно и питается Парамеция нужно молоко. Молочная культура самая продуктивная, но и самая неустойчивая. Связано это с очень бурным развитием кисломолочных бактерий, которые конкурируют с Парамециями за кислород. Если бактерий слишком много инфузории туфельки задохнуться. Поэтому работая с молочной линией нужно быть очень умеренным с подкормками. 1-3 капли молока 1-2 раза на неделю вполне достаточно.
Более устойчивой является культура на банановой кожуре.
Так же широко известен метод получения большого количества бактерий на сенном отваре. 10 – 20 граммов сена кипятят на протяжении 20 минут. Затем отвар остужают и цедят. Хранят его на нижней полке холодильника внутри плотно закрытой тары. Для использования его разбавляют чистой отстоянной водой в два – три раза.
Вообще же бактериальную массу можно получить на чем угодно. Любые органические остатки станут субстратом для размножения этих микроорганизмов. Так на страницах книг можно встретить упоминания о культуре, полученной на брюкве, моркови, сушеных листьях салата.
Здесь же описаны способы проверенные многими поколениями аквариумистов. Какая из бактериальных культур больше нравится инфузориям — судить не берусь.
Культуру Парамеции не рекомендуется использовать дольше двадцати дней. Это связано с тем что, как и все живые организмы — Парамеции, а также их пищевая база – бактерии выделяют продукты своей жизнедеятельности в окружающую среду. Поскольку наша колония пищевых микроорганизмов находится в замкнутом объеме, то постепенно этот объем заполняется шлаками на столько, что происходит самоотравление живых организмов собственными продуктами жизнедеятельности. Культура постепенно увядает.
Учитывая, что малькам нужен столь мелкий корм всего 3-5 дней после их рождения. То двадцати дневного интервала вполне достаточно чтобы загодя зарядить культуру и получить достаточное количество корма к нужному сроку. Хранить чистую культуру лучше при температуре +3-+10 градусов по Цельсию. Для этого вполне подойдет нижняя полка холодильника. Если вы решили хранить чистую культуру у себя дома, то необходимо помнить, что ее следует пересевать несколько раз за год, чтобы она не погибла.
И вот все готово.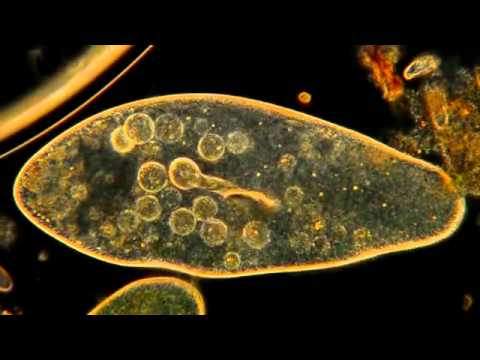 В банке видны характерные скопления живой массы. На предметном стекле микроскопа или под мощной лупой видны только инфузории туфельки. Ждем появления мальков.
В банке видны характерные скопления живой массы. На предметном стекле микроскопа или под мощной лупой видны только инфузории туфельки. Ждем появления мальков.
И вот свершилось. Заботливый аквариумист прямо таки СЛЫШИТ, как его подопечные просят есть. Что делать дальше?
Дальше в самом простом случае берем пипеткой несколько капель воды из места наибольшего скопления живой массы и капаем ее в нерестилище с голодными мальками.
Все просто и понятно… Какие могут быть подводные камни во всей этой простоте?
Камень первый. Харациновые виды очень чувствительны к бактериальному загрязнению. Поэтому чтобы избежать неприятностей берем Paramecium caudatumи помещаем их в литровую банку с чистой водой без пищевых субстратов для бактерий. Выдерживаем таким образом еду около суток. Лишенные пищи бактерии перестанут размножаться, а тех которые попали внутрь банки — съедят инфузории туфельки, которые на воле тоже играют роль санитаров. После такой самоочистки от бактерий инфузориями можно смело кормить изголодавшихся мальков.
[sam_ad id=»22″ codes=»true»]
Камень второй. Мальки существа прожорливые. Им необходимо большое количество питательных веществ для обеспечения нужд быстро растущего организма. За один раз суточную дозу корма они съесть не могут. Значит, поступление корма должно быть дробным – несколько раз за день. А что делать, если вам на работу или в школу и отпуск по уходу за мальками не дадут? Тогда следует наладить медленное, но постоянное поступление корма. Самый простой, на мой взгляд, способ – это использование капельницы. Еще один старый проверенный способ — поступление инфузорий внутрь аквариума с мальками по льняной увлажненной нитке.
Вот такая вот статейка получилась. Написал, перечитал. Вроде все понятно. Но это мне и тем, кто уже не раз это все проделывал. Самое темное пятно во всей этой почти детективной истории — это работа с невидимками. Хорошо если у вас есть доступ к микроскопу. А что делать, если нет? Главное не отчаиваться! Опыт, который приходит со временем и после нескольких неудачных попыток, и микроскоп заменить может. Могу лишь посоветовать проделать все выше описанное задолго до предполагаемого нереста, чтобы, когда понадобиться — быть уверенным в своих силах и точно знать, что в каждый конкретный момент делают ваши руки. И что видят ваши глаза.
А что делать, если нет? Главное не отчаиваться! Опыт, который приходит со временем и после нескольких неудачных попыток, и микроскоп заменить может. Могу лишь посоветовать проделать все выше описанное задолго до предполагаемого нереста, чтобы, когда понадобиться — быть уверенным в своих силах и точно знать, что в каждый конкретный момент делают ваши руки. И что видят ваши глаза.
Инфузория (Paramecium caudatum) | АкваДомик
Среди множества видов инфузорий, входящих в состав «прудовой пыли», основное место занимает инфузория-туфелька (Paramecium caudatum), размеры которой колеблются от 0,1 до 0,3 мм.
Туфелька относится к инфузориям среднего размера и имеет стройное веретенообразное тело, по форме действительно напоминающее подошву туфли.
Ее существование зависит от наличия в воде разлагающегося органического вещества. Движется она очень быстро, при поступательном движении животное вращается вокруг своей оси. Невооруженному глазу инфузории кажутся просто мелкими движущимися точками, но при небольшом увеличении с помощью хорошей лупы можно разглядеть, что это живые существа. Новорожденные мальки их хорошо видят и поедают в несметных количествах. Для совсем крошечных мальков некоторых видов, например гурами, инфузории на начальном этапе жизни просто незаменимы, и если их нет, большая часть мальков погибнет.
Новорожденные мальки их хорошо видят и поедают в несметных количествах. Для совсем крошечных мальков некоторых видов, например гурами, инфузории на начальном этапе жизни просто незаменимы, и если их нет, большая часть мальков погибнет.
Добыть туфельку можно в любом стоячем водоеме. Обильно населены ею небольшие, непроточные пруды с опавшей листвой, маленькие торфяные болота, загородные ямы с гниющими органическими веществами.
Такие водоемы часто издают характерный сероводородный запах и обычно кишат множеством различных инфузорий, в особенности у берега. Если набрать в бутылочку или пробирку воды из такого водоема, то через лупу можно просмотреть ее население. Инфузории отличаются от ракообразных своими плавными движениями и округленными очертаниями. У большинства ракообразных движения скачкообразны и под лупой уже вполне заметны их членистые конечности.
Врагами простейших, в том числе туфельки, являются все мелкие животные, питающиеся ими, — хищные инфузории, коловратки, дафнии, циклопы, насекомые и их личинки. Поэтому, учитывая предпочтительность использования для корма именно парамеции, нужно выбрать культуру, где есть только инфузории, — в большинстве туфельки.
Поэтому, учитывая предпочтительность использования для корма именно парамеции, нужно выбрать культуру, где есть только инфузории, — в большинстве туфельки.
Инфузория легко разводится в домашних условиях на различных питательных средах (корки банана, сено, молоко, листья салата, очистки картофеля, горох, фасоль и т. д.). Основные трудности заключаются в получении чистой культуры, а также в очистке от бактерий перед скармливанием. Разведение инфузорий следует приурочить приблизительно к началу нереста. Ниже мы приведем наилучшие способы разведения инфузорий. Все они базируются на едином принципе: наличии в воде какого-то разлагающегося растительного материала, что обеспечивает бактериальную среду для питания инфузорий.
Трехлитровую банку на две трети заполняют остуженной кипяченой или «старой» аквариумной водой. В банку помещают пятую часть свежей или подсушенной корки банана. Питательная среда для инфузорий готова уже через сутки, когда вода в банке помутнеет. Если вода в банке слишком мутная и имеет сильный запах сероводорода, что свидетельствует о том, что в банке слишком высокая концентрация культуральной среды (положено слишком много банановых корок), воду необходимо слить и заложить новую культуру, поскольку в таких условиях инфузории размножаться не будут. Именно для этого рекомендуется подготавливать питательную среду для инфузорий в нескольких банках: если в одной культура окажется негодной, можно будет использовать остальные.
Именно для этого рекомендуется подготавливать питательную среду для инфузорий в нескольких банках: если в одной культура окажется негодной, можно будет использовать остальные.
Для получения чистой культуры туфелек берется немного воды из природной культуры, в которой были замечены туфельки, и просматривается под лупой. Если в стекле окажется много инфузорий различных видов, то культуру нужно разбавить водой, чтобы инфузории распределились реже, затем под лупой можно вылавливать отдельных инфузорий-туфелек, что делается с помощью пипетки, с тонко оттянутым концом стеклянной трубки. Выловленных в числе одного-двух десятков туфелек пускают в заготовленный питательный раствор.
Если инфузорий нет, из пруда, где обитают инфузории, берут немного подгнивших водных растений и помещают их в банку с питательной средой.
Банку с инфузориями ставят в теплое место. Сосуд с культурой не следует ставить под прямые солнечные лучи, он может стоять в темном месте. Не следует бояться понижения температуры даже до 10 °С и ниже. Но хорошо размножаются инфузории только при комнатной температуре 18—22 °С.
Но хорошо размножаются инфузории только при комнатной температуре 18—22 °С.
Туфельки размножаются быстро, деление происходит не реже раза в сутки и через неделю при благоприятных условиях их можно увидеть даже невооруженным глазом в виде массы беловатых подвижных точек , в верхней части сосуда.
Для постоянного разведения инфузорий необходим 1 запас засушенных банановых корок, сушеного салата 1 или сена. Остаток раствора, в котором содержались инфузории, выливать не следует, достаточно добавить свежей воды. Когда инфузории потребуются вновь, в банку с раствором достаточно бросить корку банана, и через 3—5 дней в ней снова появятся инфузории.
Для постоянного поддержания культуры добавляют кипяченое молоко не чаще двух раз в месяц, исходя из пропорции 5 капель на 100 см3 воды.
Существует также способ разведения инфузорий на сыром молоке. Вместо банановых корок в банку добавляют 3—4 капли молока на 2 л воды каждые 2—3 суток. Далее инфузорий можно подкармливать или банановыми корками, или одной-двумя каплями молока, не забывая подливать воду.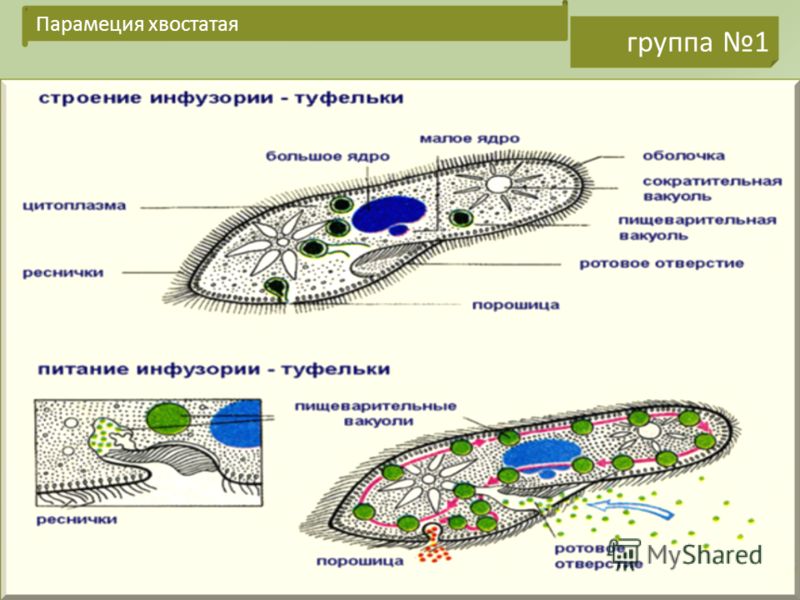
Кроме того, инфузорий разводят на сенном настое (1—2 г сена на 10 л воды), кормовых дрожжах (1 г на 100 л воды), салате, на кожуре тыквы или дыни и др.
Вода, в которой содержится культура инфузорий, имеет неприятный запах. Если вылить ее прямо в аквариум, это приведет к порче воды и гибели мальков. Поэтому, как уже говорилось выше, инфузорий надо очищать. Для этого используют фильтровальную или промокательную бумагу, которую закладывают в воронку. Затем через нее тщательно процеживают инфузорий, чтобы освободить от бактерий, после чего бумага обмывается в местах скопления мальков.
Поскольку вода в сосудах, где находятся инфузории, часто портится, они погибают. Чтобы сохранить разведенную культуру, лучше содержать их в нескольких банках.
Для солоноватоводных рыб подходят морские инфузории (Euplotes vannus) размером 0,05—0,08 мм. В 10-литровую емкость помещается в качестве питательного субстрата 50 г икры морских рыб (соленость воды должна быть не ниже 12 %, температура — 23—25 °С). Перед скармливанием морских инфузорий малькам ее обрабатывают 0,001 %-ным раствором фуразолидона.
Перед скармливанием морских инфузорий малькам ее обрабатывают 0,001 %-ным раствором фуразолидона.
В отличие от коловраток инфузории — простейшие одноклеточные животные, то есть все их тело — это одна-единственная клетка.
Среди инфузорий, наловленных в естественном водоеме, могут оказаться и опасные для рыб (ловят инфузорий, как и коловраток, очень частым сачком, ведь инфузории, коловратки— это и есть «живая пыль»). Поэтому инфузорий, как и коловраток, лучше всего развести дома в стеклянной банке.
Наиболее подходящие для разведения дома — инфузории-туфельки. Туфелек просто отличить от других мельчайших животных по их быстрому характерному движению и по оригинальной форме. Тело у туфелек веретенообразное, и, двигаясь, они вращаются вокруг своей оси. Размер туфельки 0,25—0,3 мм.
Разводить инфузорий, как и коловраток, можно на сенном настое. Готовят сенной настой так же: кипятят воду с сеном (литр воды и 10 г сена), чтобы погибли все простейшие организмы, но кипятят недолго (не более 10—15 мин), чтобы сохранились в таком сенном настое споры бактерий — сенной палочки, которой и будет питаться инфузория-туфелька. После того как настой будет готов (он должен настояться после кипения в течение двух-трех дней), в него и помещают инфузорий. В этом случае настой не разбавляют водой, как при разведении коловраток.
После того как настой будет готов (он должен настояться после кипения в течение двух-трех дней), в него и помещают инфузорий. В этом случае настой не разбавляют водой, как при разведении коловраток.
Добыть инфузорий-туфелек можно либо из естественного водоема, либо из аквариума.
Если вы добываете инфузорий из естественного водоема, то их следует отделить от других живых существ, попавшихся вместе с ними.
«Если в пробе не обнаружится никаких животных, кроме туфельки, то культура пригодна для массового разведения. В противном случае большая капля воды с максимальной концентрацией инфузорий располагается на чистом стекле, рядом с ней, со стороны света, располагается капля свежей, отстоявшейся воды. Обе капли соединяются с помощью отточенной спички водным мостиком; туфельки устремляются в сторону свежей воды и света с большей скоростью, чем все остальные микроорганизмы»— так советует отделять туфелек от прочих животных, находящихся в капле воды, Михаил Николаевич Ильин в своей книге «Аквариумное рыбоводство». Это очень нужная книга для серьезного аквариумиста. Издана она в Москве.
Это очень нужная книга для серьезного аквариумиста. Издана она в Москве.
Если вы будете добывать инфузорий из аквариума, то поступить надо так: взять из аквариума отмирающие части растений и, сдавив пальцами лист или стебелек, выдавить из них капельки воды. В этих капельках обязательно будут инфузории-туфельки, которых можно тут же поместить в сенной настой.
Если инфузории и в сенном настое будут развиваться медленно, то в банку, где они разводятся, можно добавить одну-две капли кипяченого молока, и тогда через два-три дня вы увидите в своей банке легкие белые облачка — полоски мельчайших живых существ.
Если банку закрыть чем-нибудь и таким образом ограничить доступ воздуха к сенному настою, то инфузории поднимутся к поверхности воды, где их легко собрать пипеткой, трубочкой или очень частым сачком. Так же поступают обычно и с коловратками. Помните, что «живая пыль» собирается тоже у наиболее освещенной части сосуда, в котором ее выращивают. Здесь относительно просто и собрать урожай прекрасного корма для мальков.
Можно собрать урожай «живой пыли» и с помощью колбы с высоким и узким горлом. В колбу наливают сенной настой вместе с «живой пылью». Наливают с таким расчетом, чтобы настой поднялся в колбе до середины горлышка. После этого на раствор опускают кусочек ваты так, чтобы он примерно на треть был опущен в раствор. Затем осторожно, пипеткой или трубочкой, в колбу доливают свежей, отстоявшейся воды — доливают воду так, чтобы она не смешалась с раствором и осталась светлым чистым столбиком над ваткой. В этот столбик чистой, свежей воды тут же станет подниматься «живая пыль»— она устремится к свежей воде. Очень хорошо поместить над колбой лампу, чтобы горлышко освещалось, а нижнюю часть колбы можно даже затенить. Через некоторое время почти вся «живая пыль» пройдет через вату и поднимется в столбик воды над ватой. Теперь эту воду вместе с «живой пылью» можно собрать и вылить в аквариум, где мальки поджидают корм.
Конспект урока по биологии «Инфузория – туфелька как более сложное простейшее животное.
 Половой процесс. Ползающие и сидячие инфузории»
Половой процесс. Ползающие и сидячие инфузории»Цель урока:
Познакомиться с зеленой эвгленой, инфузорией туфелька и вольвоксом как колониальным простейшим.
Задачи урока:
1) Образовательные — на основе повторения и обобщения знаний по простейшим углубить знания по жгутиконосцам и инфузориям; развивать умение высказывать свою точку зрения о значении простейших; продолжить формирование навыков работы с учебником, микроскопом; раскрыть особенности строения, процессов жизнедеятельности эвглены зеленой, а также вольвокса как колониального простейшего; выяснить различия эвглены от амебы; формировать умения выделять главное.
2) Воспитательная — воспитывать правильное отношение к окружающей действительности.
3) Развивающие – развивать мировоззренческие позиции при обращении к теме « Жгутиконосцы. Инфузории»; развивать умение вести учебный диалог; развивать способности правильно формулировать свои мысли в процессе обобщения по ранее изученным простейшим.
Оборудование к уроку: микропрепараты эвглены зеленой, инфузории туфельки, таблицы «Тип Простейшие», микроскопы.
Ход урока:
1. Организационный момент
Деятельность учителя
Сообщение темы, цели, задачи урока (на доске – записано)
1. Вызывает к доске ученика и дает ему задание (изобразить амебу и назвать органойды).
2. Предлагает ученику приготовить микропрепарат амебы, используя ранее подготовленную культуру простейших, рассмотреть под микроскопом.
3. Производит фронтальный опрос по вопросам;
(1. Какие живые организмы относятся к простейшим?
2. Какое строение имеет амеба обыкновенная?
3. Как передвигается и питается амеба?
4. Как происходит размножение амебы?
5. Какова роль пищеварительной и сократительной вакуолей?
6. Как амеба переносит неблагоприятные условия жизни?
7. Что вам известно о дизентерийной амебе? 8.Чем опасен малярийный плазмодий?)
Что вам известно о дизентерийной амебе? 8.Чем опасен малярийный плазмодий?)
Рассказ учителя. Эвглена зеленая- очень необычное существо. Ее описание можно встретить в учебниках ботаники и в учебниках зоологии. Эвглена живет в пресноводных водоемах, богатых органическими соединениями. Тело ее вытянутое, длиной около 0,05 мм. Ее передний конец притуплен, задний заострен. У эвглены, в отличии от растительных клеток, нет клеточной стенки. Наружный слой цитоплазмы плотный, он образует вокруг тела эвглены оболочку.
На переднем конце тела эвглены находится жгутик, при помощи которого она передвигается. Эвглена обладает положительным фототаксисом – ее цитоплазма содержит светочувствительный глазок. В цитоплазме эвглены содержится около 20 хлоропластов, придающих ей зеленый цвет. В хлоропластах находится хлорофилл. На свету эвглена питается как растения – путем фотосинтеза. В темноте она усваивает готовые органические вещества, образующиеся при разложении различных отмерших организмов, т. е. питается как животное.
е. питается как животное.
Задает вопросы классу:
— Почему ботаники относят эвглену к одноклеточным водорослям?
— Почему зоологи относят эвглену к простейшим животным?
— О чем свидетельствует существование таких промежуточных форм, как эвглена?
Продолжение рассказа учителя.
Размножение эвглены происходит также, как и размножение амебы, — деление клетки пополам. Рассмотрите рисунок в учебнике.
Продолжение рассказа учителя.
Вольвокс по-русски называется шаровницей…(И т. д. Сведения о вольвоксе даются по образцу из работы, без таблицы).
Инфузория туфелька.
Туфелька – обитатель стоячих водоемов с большим количеством органического материала. Она имеет постоянную, удлиненную форму тела, длина которого достигает 0,1-0,3мм Все тело инфузории покрыто продольными рядами многочисленных коротких ресничек, при помощи которых туфелька плавает тупым концом вперед. Инфузория туфелька отличается от других простейших сложностью внутриклеточной организации. В клетке имеется два ядра: макронуклеус, регулирующий процессы питания, движения, выделения, и микронуклеус, координирующий процесс размножения.
Инфузория туфелька отличается от других простейших сложностью внутриклеточной организации. В клетке имеется два ядра: макронуклеус, регулирующий процессы питания, движения, выделения, и микронуклеус, координирующий процесс размножения.
Ближе к переднему концу тела инфузории находится углубление- перистом( ротовая воронка), которое ведет в глотку. Реснички желобка постоянно работают, создавая ток воды. Вода подхватывает и подносит ко рту основную пищу туфельки- бактерий. Через глотку бактерии попадают внутрь тела инфузории. В цитоплазме вокруг них образуется пищеварительная вакуоль. Переваривание пищи и усвоение питательных веществ у туфельки происходит так же, как и у амебы. Непереваренные остатки выбрасываются наружу через отверстие — порошицу.
Весь материал — в документе.
КЛАСС ИНФУЗОРИИ (INFUSORIA или CILIATA)
Простейшие этого обширного по количеству видов — около 6 тыс.— класса широко распространены в природе. (Эта цифра приводится в сводке Корлисса, 1961 г.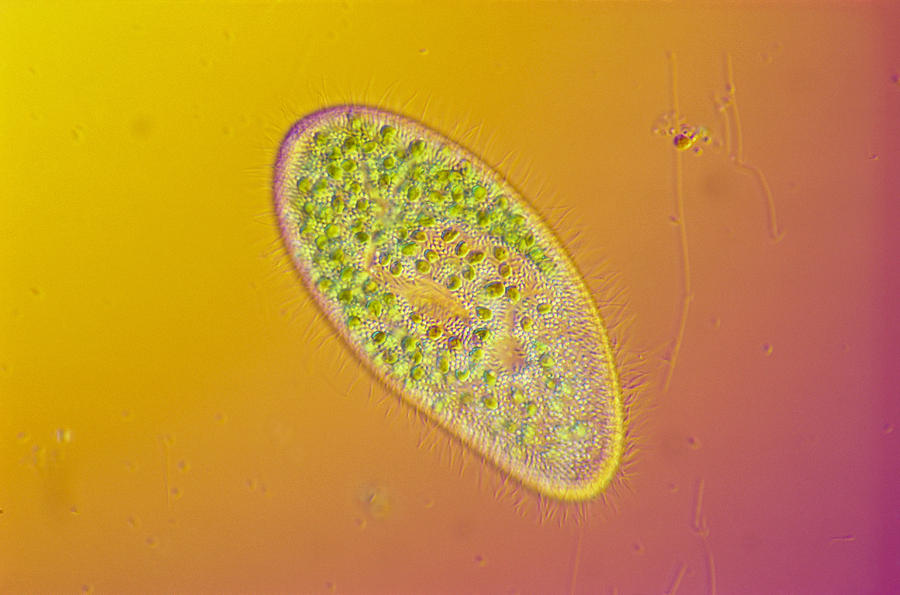 ). К ним относятся многочисленные обитатели морских и пресных вод. Некоторые виды приспособились к жизни во влажной почве. Немалое количество видов инфузорий ведет паразитический образ жизни. Хозяевами для паразитических инфузорий являются самые разнообразные беспозвоночные и позвоночные животные до высших обезьян и человека включительно.
). К ним относятся многочисленные обитатели морских и пресных вод. Некоторые виды приспособились к жизни во влажной почве. Немалое количество видов инфузорий ведет паразитический образ жизни. Хозяевами для паразитических инфузорий являются самые разнообразные беспозвоночные и позвоночные животные до высших обезьян и человека включительно.
По сравнению с другими группами простейших инфузории имеют наиболее сложное строение, что связано с разнообразием и сложностью их функций.
Откуда взялось название «инфузория туфелька»? Вы не будете удивлены, если взглянете под микроскопом на живую инфузорию или даже на ее изображение (рис. 85).
Действительно, форма тела этой инфузории напоминает изящную дамскую туфельку.
Инфузория туфелька находится в непрерывном довольно быстром движении. Скорость его (при комнатной температуре) около 2, 0—2, 5 мм/сек. Для такого маленького животного это большая скорость! Ведь это означает, что за секунду туфелька пробегает расстояние, превышающее длину ее тела в 10—15 раз.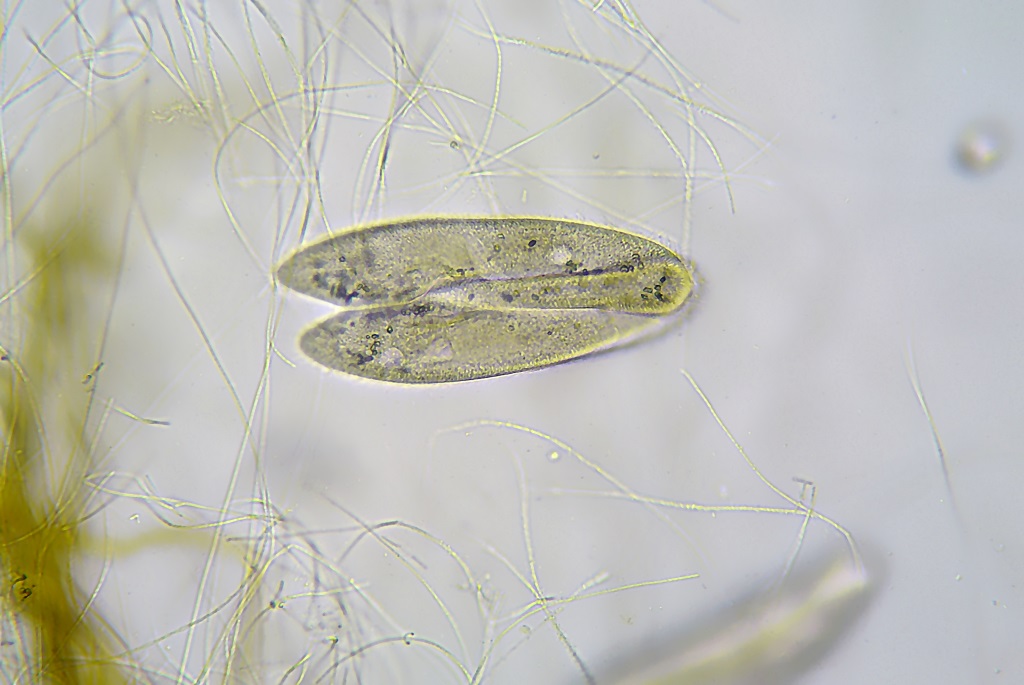 Траектория движения туфелькд довольно сложна. Она движется передним концом прямо вперед
Траектория движения туфелькд довольно сложна. Она движется передним концом прямо вперед
ИНФУЗОРИЯ ТУФЕЛЬКА (PARAMECIUM CAUDATUM)
Чтобы ознакомиться со строением и образом жизни этих интересных одноклеточных организмов, обратимся сначала к одному характерному примеру. Возьмем широко распространенных в мелких пресноводных водоемах инфузорий туфелек (виды рода Paramecium). Этих инфузорий очень легко развести в небольших аквариумах, если залить прудовой водой обычное луговое сено. В таких настойках развивается множество различных видов простейших и почти всегда развиваются инфузории туфельки. При помощи обычного учебного микроскопа можно рассмотреть многое из того, о чем будет дальше рассказано.
Среди простейших инфузории туфельки являются довольно крупными организмами. Длина тела их около 1/6—1/3 мм. и при этом вращается вправо вдоль продольной оси тела.
Столь активное движение туфельки зависит от работы большого количества тончайших волосковидных придатков — ресничек, которые покрывают все тело инфузории. Количество ресничек у одной особи инфузории туфельки равняется 10—15 тыс.!
Количество ресничек у одной особи инфузории туфельки равняется 10—15 тыс.!
Каждая ресничка совершает очень частые веслообразные движения — при комнатной температуре до 30 биений в секунду. Во время удара назад ресничка держится в выпрямленном положении. При возвращении же ее в исходную позицию (при движении вниз) она движется в 3—5 раз медленнее и описывает полукруг.
При плавании туфельки движения многочисленных покрывающих ее тело ресничек суммируются. Действия отдельных ресничек оказываются согласованными, в результате чего получаются правильные волнообразные колебания всех ресничек. Волна колебания начинается у переднего конца тела и распространяется назад. Одновременно вдоль тела туфельки проходят 2—3 волны сокращения. Таким образом, весь ресничный аппарат инфузории представляет собой как бы единое функциональное физиологическое целое, действия отдельных структурных единиц которого (ресничек) тесно связаны (координированы) между собой.
Строение каждой отдельной реснички туфельки, как показали электронно микроскопические исследования, является весьма сложным.
Направление и быстрота движения туфельки не являются величинами постоянными и неизменными. Туфелька, как и все живые организмы (мы видели это уже на примере амебы), реагирует на изменение внешней среды изменением направления движения.
Изменение направления движения простейших под влиянием различных раздражителей называют таксисами. У инфузорий легко наблюдать различные таксисы. Если в каплю, где плавают туфельки, поместить какое-либо неблагоприятно действующее на них вещество (например, кристаллик поваренной соли), то туфельки уплывают (как бы убегают) от этого неблагоприятного для них фактора (рис. 86).
Перед нами пример отрицательного таксиса на химическое воздействие (отрицательный хемотаксис). Можно наблюдать у туфельки и положительный хемотаксис. Если, например, каплю воды, в которой плавают инфузории, прикрыть покровным стеклышком и подпустить под него пузырек углекислого газа (С02), то большая часть инфузорий направится к этому пузырьку и расположится вокруг него кольцом.
Очень наглядно явление таксиса проявляется у туфелек под влиянием электрического тока. Если через жидкость, в которой плавают туфельки, пропустить слабый электрический ток, то можно наблюдать следующую картину: все инфузории ориентируют свою продольную ось параллельно линии тока, а затем, как по команде, двинутся в направлении катода, в области которого и образуют густое скопление. Движение инфузорий, определяемое направлением электрического тока, носит название гальванотаксиса. Различные таксисы у инфузорий могут быть обнаружены под влиянием самых разнообразных факторов внешней среды.
Все цитоплазматическое тело инфузории отчетливо распадается на 2 слоя: наружный — более светлый (эктоплазма) и внутренний —’ более темный и зернистый (эндоплазма). Самый поверхностный слой эктоплазмы образует наружную очень тонкую и вместе с тем прочную и эластичную оболочку — пелликулу, которая играет важную роль в сохранении постоянства формы тела инфузории.
В наружном слое (в эктоплазме) тела живой туфельки хорошо видны многочисленные коротенькие палочки, расположенные перпендикулярно к поверхности (рис. 85, 7). Эти образования носят название трихоцисты. Функция их очень интересна и связана с защитой простейшего. При механическом, химическом или каком-либо ином сильном раздражении трихоцисты с силой выбрасываются наружу, превращаясь в тонкие длинные нити, которые поражают хищника, нападающего на туфельку. Трихоцисты представляют собой мощную защиту. Они располагаются закономерно между ресничками, так что число трихоцист приблизительно соответствует числу ресничек. На месте использованных («выстреленных») трихоцист в эктоплазме туфельки развиваются новые.
85, 7). Эти образования носят название трихоцисты. Функция их очень интересна и связана с защитой простейшего. При механическом, химическом или каком-либо ином сильном раздражении трихоцисты с силой выбрасываются наружу, превращаясь в тонкие длинные нити, которые поражают хищника, нападающего на туфельку. Трихоцисты представляют собой мощную защиту. Они располагаются закономерно между ресничками, так что число трихоцист приблизительно соответствует числу ресничек. На месте использованных («выстреленных») трихоцист в эктоплазме туфельки развиваются новые.
На одной стороне, приблизительно по середине тела (рис. 85, 5), у туфельки имеется довольно глубокая впадина. Это ротовая впадина, или перистом. По стенкам перистома, так же как и по поверхности тела, расположены реснички. Они развиты здесь гораздо более мощно, чем на всей остальной поверхности тела. Эти тесно расположенные реснички собраны в две группы. Функция этих особо дифференцированных ресничек связана не с движением, а с питанием (рис. 87).
87).
Как и чем питаются туфельки, как осуществляется у них пищеварение?
Туфельки относятся к числу инфузорий, основную пищу которых составляют бактерии. Наряду с бактериями они могут заглатывать и любые другие взвешенные в воде частицы независимо от их питательности. Околоротовые реснички создают непрерывный ток воды со взвешенными в ней частицами в направлении ротового отверстия, которое расположено в глубине перистома. Мелкие пищевые частицы (чаще всего бактерии) проникают через рот в небольшую трубковидную глотку и скапливаются на дне ее, на границе с эндоплазмой. Ротовое отверстие всегда открыто. Пожалуй, не будет ошибкой сказать, что инфузория туфелька — одно из самых прожорливых животных: она непрерывно питается. Этот процесс прерывается только в определенные моменты жизни, связанные с размножением и половым процессом.
Скопившийся на дне глотки пищевой комочек в дальнейшем отрывается от дна глотки и вместе с небольшим количеством жидкости поступает в эндоплазму, образуя пищеварительную вакуолю.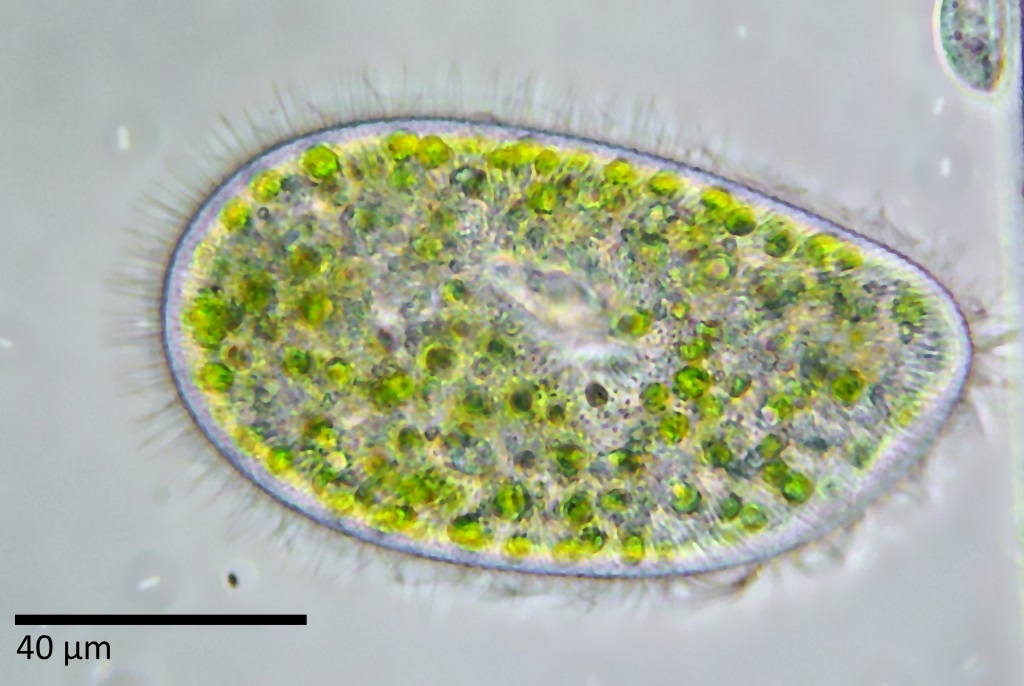 Последняя не остается на месте своего образования, а, попадая в токи эндоплазмы, проделывает в теле туфельки довольно сложный и закономерный путь, называемый циклозом пищеварительной вакуоли (рис. 88). Во время этого довольно длительного (при комнатной температуре занимающего около часа) путешествия пищеварительной вакуоли внутри ее происходит ряд изменений, связанных с перевариванием находящейся в ней пищи.
Последняя не остается на месте своего образования, а, попадая в токи эндоплазмы, проделывает в теле туфельки довольно сложный и закономерный путь, называемый циклозом пищеварительной вакуоли (рис. 88). Во время этого довольно длительного (при комнатной температуре занимающего около часа) путешествия пищеварительной вакуоли внутри ее происходит ряд изменений, связанных с перевариванием находящейся в ней пищи.
Здесь, так же как у амеб и некоторых жгутиконосцев, происходит типичное внутриклеточное пищеварение. Из окружающей пищеварительную вакуолю эндоплазмы в нее поступают пищеварительные ферменты, которые воздействуют на пищевые частицы. Продукты переваривания пищи всасываются через стгнку пищеварительной вакуоли в эндоплазму.
По ходу циклоза пищеварительной вакуоли в ней сменяется несколько фаз пищеварения. В первые моменты после образования вакуоли заполняющая ее жидкость мало отличается от жидкости окружающей среды. Вскоре начинается поступление из эндоплазмы в вакуолю пищеварительных ферментов и реакция среды внутри нее становится резко кислой. Это легко обнаружить, добавляя к пище какой-либо индикатор, цвет которого меняется в зависимости от реакции (кислой, нейтральной или щелочной) среды. В этой кислой среде проходят первые фазы пищеварения. Затем картина меняется и реакция внутри пищеварительных вакуолей становится слабощелочной. В этих условиях и протекают дальнейшие этапы внутриклеточного пищеварения. Кислая фаза обычно более короткая, чем щелочная; она длится примерно 1/6—1/4 часть всего срока пребывания пищеварительной вакуоли в теле инфузории. Однако соотношение кислой и щелочной фаз может варьировать в довольно широких пределах в зависимости от характера пищи.
Это легко обнаружить, добавляя к пище какой-либо индикатор, цвет которого меняется в зависимости от реакции (кислой, нейтральной или щелочной) среды. В этой кислой среде проходят первые фазы пищеварения. Затем картина меняется и реакция внутри пищеварительных вакуолей становится слабощелочной. В этих условиях и протекают дальнейшие этапы внутриклеточного пищеварения. Кислая фаза обычно более короткая, чем щелочная; она длится примерно 1/6—1/4 часть всего срока пребывания пищеварительной вакуоли в теле инфузории. Однако соотношение кислой и щелочной фаз может варьировать в довольно широких пределах в зависимости от характера пищи.
Путь пищеварительной вакуоли в эндоплазме заканчивается тем, что она приближается к поверхности тела и через пелликулу содержимое ее, состоящее из жидкости и непереваренных остатков пищи, выбрасывается наружу — происходит дефекация. Этот процесс, в отличие от амеб, у которых дефекация может происходить в любом месте, у туфелек, как и у других инфузорий, строго приурочен к определенному участку тела, расположенному на брюшной стороне (брюшной условно называют ту поверхность животного, на которой помещается околоротовое углубление), примерно посередине между перистомом и задним концом тела.
Таким образом, внутриклеточное пищеварение представляет собой сложный процесс, слагающийся из нескольких последовательно сменяющих друг друга фаз.
Кроме пищеварительных вакуолей, в теле инфузории туфельки имеются выполняющие также очень важные жизненные функции сократительные вакуоли. У туфельки их две, они расположены в передней и задней трети тела. Каждая из вакуолей состоит из центрального резервуара и приводящих каналов (5—7), которые расположены радиально вокруг центрального резервуара. Цикл работы сократительной вакуоли начинается с того, что приводящие каналы заполняются жидкостью и становятся хорошо видимыми (рис. 89). Затем жидкое содержимое их изливается в центральный резервуар, сами каналы после опорожнения становятся на некоторое время невидимыми. Последний этап цикла работы сократительной вакуоли заключается в том, что жидкость из центрального резервуара изливается через особую пору в пелликуле наружу. После этого центральный резервуар на короткий срок перестает быть видимым.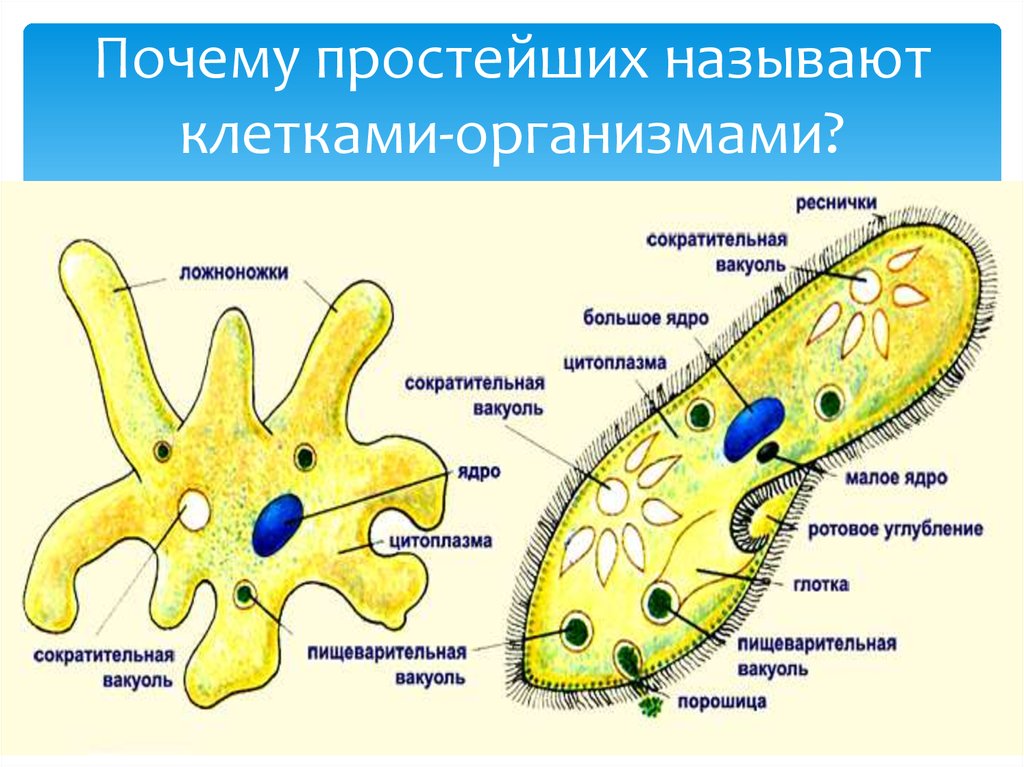 В это время приводящие каналы вновь начинают заполняться жидкостью и весь цикл начинается сначала. Обычно передняя и задняя сократительные вакуоли работают последовательно, как бы по очереди. Каков темп пульсации вакуолей? Какое количество жидкости выводится ими наружу? Частота сокращения сократительной вакуоли в большой степени зависит от условий внешней среды, и особенно от температуры и осмотического давления. При комнатной температуре у туфельки сократительная вакуоля проделывает весь цикл пульсации за 10—15 секунд. У морских и паразитических инфузорий темп пульсации сократительных вакуолей обычно значительно ниже, чем у пресноводных.
В это время приводящие каналы вновь начинают заполняться жидкостью и весь цикл начинается сначала. Обычно передняя и задняя сократительные вакуоли работают последовательно, как бы по очереди. Каков темп пульсации вакуолей? Какое количество жидкости выводится ими наружу? Частота сокращения сократительной вакуоли в большой степени зависит от условий внешней среды, и особенно от температуры и осмотического давления. При комнатной температуре у туфельки сократительная вакуоля проделывает весь цикл пульсации за 10—15 секунд. У морских и паразитических инфузорий темп пульсации сократительных вакуолей обычно значительно ниже, чем у пресноводных.
Подсчеты показывают, что примерно за 30—45 минут у туфельки через сократительные вакуоли выводится объем жидкости, равный объему тела инфузории. Таким образом, благодаря деятельности сократительных вакуолей через тело инфузории осуществляется непрерывный ток воды, поступающей снаружи через ротовое отверстие (вместе с пищеварительными вакуолями), а также осмотически непосредственно через пелликулу. Сократительные вакуоли играют важную роль в регулировании тока воды, проходящего через тело инфузории, в регулировании осмотического давления. Этот процесс здесь протекает в принципе так же, как у амеб, только структура сократительной вакуоли намного сложнее.
Сократительные вакуоли играют важную роль в регулировании тока воды, проходящего через тело инфузории, в регулировании осмотического давления. Этот процесс здесь протекает в принципе так же, как у амеб, только структура сократительной вакуоли намного сложнее.
В течение долгих лет среди ученых, занимающихся изучением простейших, шел спор по вопросу о том, имеются ли в цитоплазме какие-нибудь структуры, связанные с появлением сократительной вакуоли, или же она образуется всякий раз заново. На живой инфузории никаких особых структур, которые предшествовали бы ее образованию, наблюдать не удается. После того как произойдет сокращение вакуоли — систола, в цитоплазме на месте бывшей вакуоли не видно решительно никаких структур. Затем заново появляются прозрачный пузырек или приводящие каналы, которые начинают увеличиваться в размерах. Однако никакой связи вновь возникающей вакуоли с существовавшей ранее не обнаруживается. Создается впечатление, что преемственности между следующими друг за другом циклами сократительной вакуоли нет и всякая новая сократительная вакуоля образуется в цитоплазме заново. Однако специальные методы исследования показали, что на самом деле это не так. Применение электронной микроскопии, дающей очень большое увеличение (до 100 тыс. раз), убедительно показало, что у инфузории на том участке, где формируются сократительные вакуоли, имеется особо дифференцированная цитоплазма, состоящая из переплетения тончайших трубочек. Таким образом, оказалось, что сократительная вакуоля возникает в цитоплазме не на «пустом месте», а на основе предшествующего особого органоида клетки, функцией которого является формирование сократительной вакуоли.
Однако специальные методы исследования показали, что на самом деле это не так. Применение электронной микроскопии, дающей очень большое увеличение (до 100 тыс. раз), убедительно показало, что у инфузории на том участке, где формируются сократительные вакуоли, имеется особо дифференцированная цитоплазма, состоящая из переплетения тончайших трубочек. Таким образом, оказалось, что сократительная вакуоля возникает в цитоплазме не на «пустом месте», а на основе предшествующего особого органоида клетки, функцией которого является формирование сократительной вакуоли.
Как и у всех простейших, у инфузорий имеется клеточное ядро. Однако по строению ядерного аппарата инфузории резко отличаются от всех других групп простейших.
Ядерный аппарат инфузорий характеризуется своим дуализмом. Это означает, что у инфузорий имеется два разных типа ядер — большие ядра, или макронуклеусы, и малые ядра, или микронуклеусы. Посмотрим, какое строение имеет ядерный аппарат у инфузории туфельки (рис. 85).
85).
В центре тела инфузории (на уровне перистома) помещается большое массивное ядро яйцевидной или бобовидной формы. Это макронуклеус. В тесном соседстве с ним расположено второе ядро во много раз меньших размеров, обычно довольно тесно прилежащее к макронуклеусу. Это микронуклеус. Различие между этими двумя ядрами не сводится только к размерам, оно более значительно, глубоко затрагивает их структуру.
Макронуклеус по сравнению с микронуклеусом гораздо богаче особым ядерным веществом (хроматином, или, точнее, дезоксирибонуклеиновой кислотой, сокращенно ДНК), входящим в состав хромосом.
Исследования последних лет показали, что макронуклеус обладает в несколько десятков (а у некоторых инфузорий и сотен) раз большим числом хромосом, чем микронуклеусы. Макронуклеус является совсем своеобразным типом многохромосомных (полиплоидных) ядер. Таким образом, различие между микро- и макронуклеусами затрагивает их хромосомный состав, отчего и зависит большее или меньшее богатство их ядерным веществом — хроматином.
У одного из наиболее обычных видов инфузории — туфельки (Paramecium caudatum) — имеется один макронуклеус (сокращенно Ма) и один микронуклеус (сокращенно Mi). Такая структура ядерного аппарата свойственна многим инфузориям. У других может быть по нескольку Ма и Mi. Но характерная особенность всех инфузорий — дифференцировка ядер на две качественно различные группы, на Ма и Mi, или, другими словами, явление ядерного дуализма.
Как размножаются инфузории? Обратимся в качестве примера опять к инфузории туфельке. Если отсадить в небольшой сосуд (микроаквариум) один-единственный экземпляр туфельки, то уже через сутки там окажется две, а нередко и четыре инфузории. Как это происходит? После некоторого периода активного плавания и питания инфузория несколько вытягивается в длину. Затем точно по середине тела появляется все углубляющаяся поперечная перетяжка (рис. 90). В конце концов инфузория как бы перешнуровывается пополам и из одной особи получаются две, первоначально несколько меньших размеров, чем материнская особь. Весь процесс деления занимает при комнатной температуре около часа. Изучение внутренних процессов показывает, что еще до того, как появляется поперечная перетяжка, начинается процесс деления ядерного аппарата. Первым делится Mi и лишь после него — Ма. Мы не будем здесь останавливаться на детальном рассмотрении процессов ядерного деления и лишь отметим, что Mi делится путем митоза, тогда как деление Ма по внешности напоминает прямое деление ядра — амитоз. Этот бесполый процесс размножения инфузории туфельки, как мы видим, сходен с бесполым размножением амеб и жгутиконосцев. В отличие от них инфузории в процессе бесполого размножения делятся всегда поперек, тогда как у жгутиконосцев плоскость деления параллельна продольной оси тела.
Весь процесс деления занимает при комнатной температуре около часа. Изучение внутренних процессов показывает, что еще до того, как появляется поперечная перетяжка, начинается процесс деления ядерного аппарата. Первым делится Mi и лишь после него — Ма. Мы не будем здесь останавливаться на детальном рассмотрении процессов ядерного деления и лишь отметим, что Mi делится путем митоза, тогда как деление Ма по внешности напоминает прямое деление ядра — амитоз. Этот бесполый процесс размножения инфузории туфельки, как мы видим, сходен с бесполым размножением амеб и жгутиконосцев. В отличие от них инфузории в процессе бесполого размножения делятся всегда поперек, тогда как у жгутиконосцев плоскость деления параллельна продольной оси тела.
Во время деления происходит глубокая внутренняя перестройка тела инфузории. Образуется два новых перистома, две глотки и два ротовых отверстия. К этому же времени приурочено деление базальных ядер ресничек, за счет которых образуются новые реснички.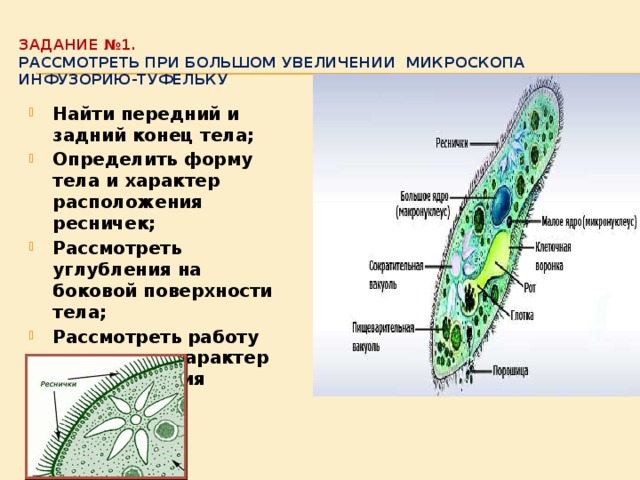 Если бы при размножении число ресничек не возрастало, то в результате каждого деления дочерние особи получили бы примерно половину числа ресничек материнской особи, что привело бы к полному «облысению» инфузорий. На самом деле этого не происходит.
Если бы при размножении число ресничек не возрастало, то в результате каждого деления дочерние особи получили бы примерно половину числа ресничек материнской особи, что привело бы к полному «облысению» инфузорий. На самом деле этого не происходит.
Время от времени у большинства инфузорий, в том числе и у туфелек, наблюдается особая и чрезвычайно своеобразная форма полового процесса, которая получила название конъюгации. Мы не будем здесь детально разбирать все те сложные ядерные изменения, которые сопровождают этот процесс, а отметим только самое главное. Конъюгация протекает следующим образом (рис. 91), Две инфузории сближаются, тесно прикладываются друг к другу брюшными сторонами и в таком виде плавают довольно длительное время вместе (у туфельки примерно в течение 12 часов при комнатной температуре). Затем конъюганты расходятся. Что же происходит в теле инфузории во время конъюгации? Сущность этих процессов сводится к следующему (рис. 91). Большое ядро (макронуклеус) разрушается и постепенно растворяется в цитоплазме. Микронуклеусы сначала делятся, часть ядер, образующихся в результате деления, разрушается (см. рис. 91). В каждом из конъюгантов сохраняется по два ядра. Одно из этих ядер остается на месте в той особи, в которой оно образовалось (стационарное ядро), другое же активно перемещается в партнера по конъюгации (мигрирующее ядро) и сливается с его стационарным ядром. Таким образом, в каждом из конъюгантов на этой стадии имеется по одному ядру, образовавшемуся в результате слияния стационарного и мигрирующего ядер. Это сложное ядро получает название синкарион. Образование синкариона представляет собой не что иное, как процесс оплодотворения. И у многоклеточных существенным моментом оплодотворения является слияние ядер половых клеток. У инфузорий половые клетки не образуются, имеются лишь половые ядра, которые и сливаются между собой. Таким образом происходит взаимное перекрестное оплодотворение.
Микронуклеусы сначала делятся, часть ядер, образующихся в результате деления, разрушается (см. рис. 91). В каждом из конъюгантов сохраняется по два ядра. Одно из этих ядер остается на месте в той особи, в которой оно образовалось (стационарное ядро), другое же активно перемещается в партнера по конъюгации (мигрирующее ядро) и сливается с его стационарным ядром. Таким образом, в каждом из конъюгантов на этой стадии имеется по одному ядру, образовавшемуся в результате слияния стационарного и мигрирующего ядер. Это сложное ядро получает название синкарион. Образование синкариона представляет собой не что иное, как процесс оплодотворения. И у многоклеточных существенным моментом оплодотворения является слияние ядер половых клеток. У инфузорий половые клетки не образуются, имеются лишь половые ядра, которые и сливаются между собой. Таким образом происходит взаимное перекрестное оплодотворение.
Вскоре после образования синкариона конъюганты расходятся. По строению своего ядерного аппарата они на этой стадии еще очень существенно отличаются от обычных так называемых нейтральных (не конъюгирующих) инфузорий, так как у них имеется лишь по одному ядру. В дальнейшем за счет синкариона происходит восстановление нормального ядерного аппарата. Синкарион делится (один или несколько раз). Часть продуктов этого деления путем сложных преобразований, связанных с увеличением числа хромосом и обогащением хроматином, превращается в макронуклеусы. Другие сохраняют структуру, характерную для микронуклеусов. Таким путем восстанавливается характерный и типичный для инфузорий ядерный аппарат, после чего инфузории приступают к бесполому размножению путем деления.
В дальнейшем за счет синкариона происходит восстановление нормального ядерного аппарата. Синкарион делится (один или несколько раз). Часть продуктов этого деления путем сложных преобразований, связанных с увеличением числа хромосом и обогащением хроматином, превращается в макронуклеусы. Другие сохраняют структуру, характерную для микронуклеусов. Таким путем восстанавливается характерный и типичный для инфузорий ядерный аппарат, после чего инфузории приступают к бесполому размножению путем деления.
Таким образом, процесс конъюгации включает в себя два существенно важных биологических момента: оплодотворение и восстановление нового макронуклеуса за счет синкариона.
В чем заключается биологическое значение конъюгации, какую роль играет она в жизни инфузорий? Мы не можем назвать ее размножением, ибо при этом увеличения числа особей не происходит. Поставленные выше вопросы послужили материалом для многочисленных экспериментальных исследований, осуществлявшихся во многих странах. Основной результат этих исследований сводится к следующему. Во-первых, конъюгация, как и всякий другой половой процесс, при котором происходит объединение в одном организме двух наследственных начал (отцовского и материнского), ведет к повышению наследственной изменчивости, наследственного многообразия. Повышение наследственной изменчивости увеличивает приспособительные возможности организма к условиям окружающей среды. Второй биологически важной стороной конъюгации является развитие нового макронуклеуса за счет продуктов деления синкариона и одновременно с этим разрушение старого. Экспериментальные данные показывают, что именно макронуклеус играет исключительно важную роль в жизни инфузорий. Им контролируются все основные жизненные процессы и определяется важнейший из них — образование (синтез) белка, составляющего основную часть протоплазмы живой клетки. При длительном бесполом размножении путем деления происходит как бы своеобразный процесс «старения» макронуклеуса, а вместе с тем и всей клетки: снижается активность процесса обмена веществ, снижается темп деления.
Основной результат этих исследований сводится к следующему. Во-первых, конъюгация, как и всякий другой половой процесс, при котором происходит объединение в одном организме двух наследственных начал (отцовского и материнского), ведет к повышению наследственной изменчивости, наследственного многообразия. Повышение наследственной изменчивости увеличивает приспособительные возможности организма к условиям окружающей среды. Второй биологически важной стороной конъюгации является развитие нового макронуклеуса за счет продуктов деления синкариона и одновременно с этим разрушение старого. Экспериментальные данные показывают, что именно макронуклеус играет исключительно важную роль в жизни инфузорий. Им контролируются все основные жизненные процессы и определяется важнейший из них — образование (синтез) белка, составляющего основную часть протоплазмы живой клетки. При длительном бесполом размножении путем деления происходит как бы своеобразный процесс «старения» макронуклеуса, а вместе с тем и всей клетки: снижается активность процесса обмена веществ, снижается темп деления. После конъюгации (в процессе которой, как мы видели, старый макронуклеус разрушается) происходит восстановление уровня обмена веществ и темпа деления. Поскольку при конъюгации происходит процесс оплодотворения, который у большинства других организмов связан с размножением и появлением нового поколения, у инфузорий особь, образовавшуюся после конъюгации, тоже можно рассматривать как новое половое поколение, которое возникает здесь как бы за счет «омолаживания» старого.
После конъюгации (в процессе которой, как мы видели, старый макронуклеус разрушается) происходит восстановление уровня обмена веществ и темпа деления. Поскольку при конъюгации происходит процесс оплодотворения, который у большинства других организмов связан с размножением и появлением нового поколения, у инфузорий особь, образовавшуюся после конъюгации, тоже можно рассматривать как новое половое поколение, которое возникает здесь как бы за счет «омолаживания» старого.
На примере инфузории туфельки мы познакомились с типичным представителем обширного класса инфузорий. Однако класс этот характерен чрезвычайным разнообразием видов как по строению, так и по образу жизни. Познакомимся ближе с некоторыми наиболее характерными и интересными формами.
У инфузории туфельки реснички равномерно покрывают всю поверхность тела. Это характерная черта строения отряда равноресничных инфузорий (Holotricha). Многим инфузориям свойствен иной характер развития ресничного покрова. Дело в том, что реснички инфузорий способны, соединяясь вместе, образовывать более сложные комплексы. Например, нередко наблюдается, что расположенные в один или два ряда близко друг к другу реснички соединяются (слипаются) вместе, образуя пластинку, которая, так же как и реснички, способна к биению. Такие пластинчатые сократимые образования получили название мембранелл (если они короткие) или мембран (если они длинные). В других случаях соединяются вместе реснички, расположенные тесным пучком. Эти образования — цирры — напоминают кисточку, отдельные волоски которой слеплены друг с другом. Разного рода сложные ресничные образования характерны для многих инфузорий. Очень часто ресничный покров развивается не равномерно, а лишь на некоторых участках тела.
Дело в том, что реснички инфузорий способны, соединяясь вместе, образовывать более сложные комплексы. Например, нередко наблюдается, что расположенные в один или два ряда близко друг к другу реснички соединяются (слипаются) вместе, образуя пластинку, которая, так же как и реснички, способна к биению. Такие пластинчатые сократимые образования получили название мембранелл (если они короткие) или мембран (если они длинные). В других случаях соединяются вместе реснички, расположенные тесным пучком. Эти образования — цирры — напоминают кисточку, отдельные волоски которой слеплены друг с другом. Разного рода сложные ресничные образования характерны для многих инфузорий. Очень часто ресничный покров развивается не равномерно, а лишь на некоторых участках тела.
ИНФУЗОРИЯ ТРУБАЧ (STENTOR POLYMORPHIC)
В пресных водах очень часто встречаются виды крупных красивых инфузорий, относящихся к роду трубачей (Stentor). Это название вполне соответствует форме тела этих животных, которая действительно напоминает трубу (рис. 92), широко раскрывшуюся на одном конце. При первом же знакомстве с живыми трубачами можно заметить одну особенность, которая не свойственна туфельке. При малейшем раздражении, в том числе механическом (например, постукивание карандашом по стеклу, где имеется капля воды с трубачами), тело их резко и очень быстро (в долю секунды) сокращается, принимая почти правильную шарообразную форму. Затем довольно медленно (время измеряется секундами) трубач расправляется, принимая характерную для него форму. Эта способность трубача быстро сокращаться обусловлена наличием особых мускульных волоконец, расположенных вдоль тела и в эктоплазме. Таким образом, и в одноклеточном организме может развиться мышечная система.
92), широко раскрывшуюся на одном конце. При первом же знакомстве с живыми трубачами можно заметить одну особенность, которая не свойственна туфельке. При малейшем раздражении, в том числе механическом (например, постукивание карандашом по стеклу, где имеется капля воды с трубачами), тело их резко и очень быстро (в долю секунды) сокращается, принимая почти правильную шарообразную форму. Затем довольно медленно (время измеряется секундами) трубач расправляется, принимая характерную для него форму. Эта способность трубача быстро сокращаться обусловлена наличием особых мускульных волоконец, расположенных вдоль тела и в эктоплазме. Таким образом, и в одноклеточном организме может развиться мышечная система.
В роде трубачей имеются виды, некоторые из которых характеризуются довольно яркой окраской. Очень обычен в пресных водах голубой трубач (Stentor coeruleus), который окрашен в ярко-голубой цвет. Эта окраска трубача обусловлена тем, что в эктоплазме его расположены мельчайшие зерна синего пигмента.
Другой вид трубача (Stentor polymorphus) нередко бывает окрашен в зеленый цвет. Причина этой окраски совсем иная. Зеленый цвет обусловлен тем, что в эндоплазме инфузории живут и размножаются мелкие одноклеточные зеленые водоросли, которые и придают телу трубача характерную окраску. Stentor polymorphus — типичный пример взаимно полезного сожительства — симбиоза. Трубач и водоросли находятся во взаимно симбиотических отношениях: трубач защищает водоросли, живущие в его теле, и снабжает их углекислотой, образующейся в результате дыхания; со своей стороны водоросли дают трубачу кислород, освобождающийся в процессе фотосинтеза. По-видимому, часть водорослей переваривается инфузорией, являясь пищей для трубача.
Трубачи медленно плавают в воде широким концом вперед. Но они могут также временно прикрепляться к субстрату задним узким концом тела, на котором при этом образуется небольшая присоска.
В теле трубача можно различить расширяющийся сзади наперед туловищный отдел и почти перпендикулярно к нему расположенное широкое околоротовое (перистомальное) поле. Это поле напоминает асимметричную плоскую воронку, с одного края которой имеется углубление — глотка, ведущая в эндоплазму инфузории. Тело трубача покрыто продольными рядами коротких ресничек. По краю перистомального поля по кругу располагается мощно развитая околоротовая (адоральная) зона мембранелл (рис. 92). Эта зона состоит из большого числа отдельных мерцательных пластинок, каждая из которых в свою очередь слагается из множества слипшихся друг с другом, расположенных двумя тесно сближенными рядами ресничек.
Это поле напоминает асимметричную плоскую воронку, с одного края которой имеется углубление — глотка, ведущая в эндоплазму инфузории. Тело трубача покрыто продольными рядами коротких ресничек. По краю перистомального поля по кругу располагается мощно развитая околоротовая (адоральная) зона мембранелл (рис. 92). Эта зона состоит из большого числа отдельных мерцательных пластинок, каждая из которых в свою очередь слагается из множества слипшихся друг с другом, расположенных двумя тесно сближенными рядами ресничек.
В области ротового отверстия околоротовые мембранеллы заворачивают в сторону глотки, образуя левозакрученную спираль. Ток воды, вызываемый колебанием околоротовых мембранелл, направлен в сторону ротового отверстия (в глубину воронки, образуемой передним концом тела). Вместе с водой в глотку попадают и взвешенные в воде пищевые частицы. Пищевые объекты у трубача разнообразнее, чем у туфельки. Наряду с бактериями он поедает мелких простейших (например, жгутиковых), одноклеточные водоросли и т. п.
п.
У трубача хорошо развита сократительная вакуоля, имеющая своеобразное строение. Центральный резервуар расположен в передней трети тела, несколько ниже ротового отверстия. От него отходят два длинных приводящих канала. Один из них идет от резервуара к заднему концу тела, второй расположен в области перистомального поля параллельно околоротовой зоне мембранелл.
Очень своеобразно устроен ядерный аппарат трубача. Макронуклеус здесь поделен на четки (их около 10), связанные друг с другом тонкими перемычками. Микронуклеусов несколько. Они очень мелкие и обычно тесно прилегают к четкам макронуклеуса.
Инфузория трубач — излюбленный объект для экспериментальных исследований по регенерации. Многочисленными опытами была доказана высокая регенеративная способность трубачей. Инфузорию тонким скальпелем можно разрезать на множество частей, и каждая из них через короткое время (несколько часов, иногда сутки и более) превратится в пропорционально построенного, но маленького трубача, который затем в результате энергичного питания достигает типичного для данного вида размера. Для завершения восстановительных процессов в регенерирующем кусочке должен быть хотя бы один сегмент четковидного макронуклеуса.
Для завершения восстановительных процессов в регенерирующем кусочке должен быть хотя бы один сегмент четковидного макронуклеуса.
Трубач, как мы видели, обладает разными ресничками: с одной стороны короткими, покрывающими все тело, а с другой — околоротовой зоной мембранелл. В соответствии с этой характерной чертой строения отряд инфузорий, к которому относится трубач, получил название разноресничных инфузорий (Heterotricha).
ИНФУЗОРИЯ БУРСАРИЯ (BURSARIA TRUNCATELLA)
Второй интересный представитель разноресничных инфузорий — часто встречающаяся в пресных водах бурсария (Bursaria truncatella, рис. 93). Это гигант среди инфузорий: ее размеры могут достигать 2 мм, наиболее обычные величины — 0, 5—1, 0 мм. Бурсарию хорошо видно невооруженным глазом. В соответствии со своим названием бурсария имеет форму мешка, широко открытого на переднем конце (bursa — латинское слово, в переводе на русский язык означает «кошель», «мешок») и несколько расширенного на заднем конце.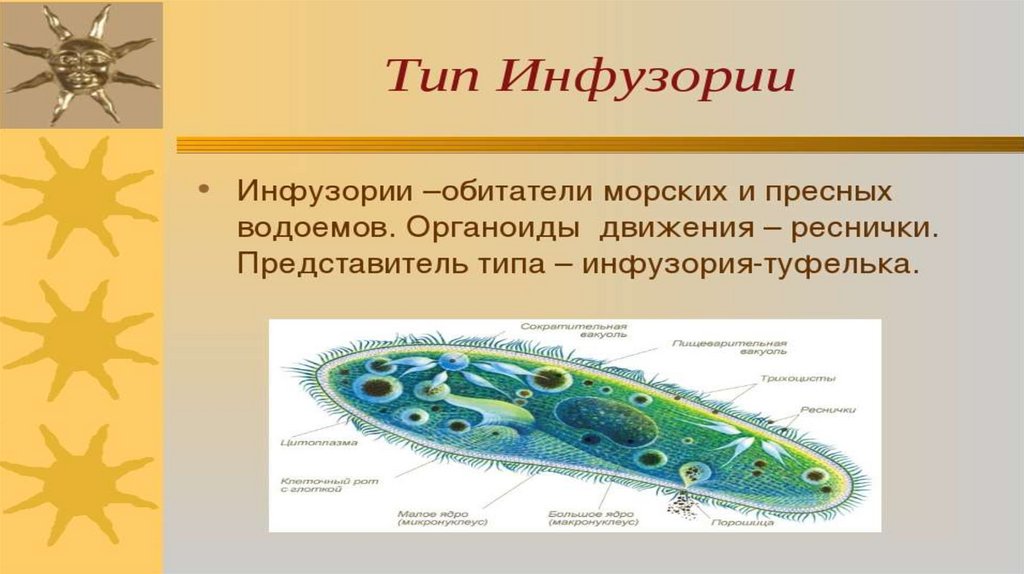 Все тело инфузории покрыто продольно идущими рядами коротких ресничек. Биение их обусловливает довольно медленное поступательное движение животного. Бурсария плывет как бы «переваливаясь» с бока на бок.
Все тело инфузории покрыто продольно идущими рядами коротких ресничек. Биение их обусловливает довольно медленное поступательное движение животного. Бурсария плывет как бы «переваливаясь» с бока на бок.
От переднего конца в глубь тела (примерно на 2/3 его длины) вдается околоротовое углубление — перистом. С брюшной стороны оно сообщается с наружной средой узкой щелью, на спинной стороне полость перистома с наружной средой не сообщается. Если посмотреть на поперечный разрез верхней трети тела бурсарии (рис. 93, Б), то видно, что полость перистома занимает большую часть тела, тогда как цитоплазма окружает ее в виде ободка.
На переднем конце тела, слева берет начало очень мощно развитая у бурсарии зона околоротовых (адоральных) мембранелл (рис. 93, 4). Она спускается в глубину полости перистома, заворачивая налево. Адоральная зона доходит до самой глубокой части перистома. Кроме околоротовых мембранелл, других ресничных образований в полости перистома нет, за исключением ресничной полоски, идущей по брюшной стороне полости перистома (рис. 93, 10). На внутренней задней стенке перистомальной полости почти по всей ее длине проходит узкая щель (рис. 93, 7), края которой обычно тесно прилегают друг к другу. Это ротовая щель. Ее края раздвигаются только в момент приема пищи.
93, 10). На внутренней задней стенке перистомальной полости почти по всей ее длине проходит узкая щель (рис. 93, 7), края которой обычно тесно прилегают друг к другу. Это ротовая щель. Ее края раздвигаются только в момент приема пищи.
Бурсарии не обладают узкой пищевой специализацией, но в основном это хищники. При поступательном движении они наталкиваются на различных мелких животных. Благодаря работе мембранелл околоротовой зоны добыча с силой втягивается в обширную перистомальную полость, откуда уже не может выплыть наружу. Пищевые объекты прижимаются к спинной стенке перистомальной полости и через раздвигающуюся ротовую щель проникают в эндоплазму. Бурсарии очень прожорливы, они могут проглатывать довольно крупные объекты: например, излюбленной пищей их являются инфузории туфельки. Бурсария способна подряд проглотить 6—7 туфелек. В результате в эндоплазме бурсарии образуются очень крупные пищеварительные вакуоли.
Ядерный аппарат бурсарии устроен довольно сложно. Они имеют один длинный колбасовидный макронуклеус и большое (примерно до 30) количество мелких микронуклеусов, беспорядочно разбросанных в эндоплазме инфузории.
Они имеют один длинный колбасовидный макронуклеус и большое (примерно до 30) количество мелких микронуклеусов, беспорядочно разбросанных в эндоплазме инфузории.
Бурсарии относятся к числу немногих видов пресноводных инфузорий, у которых отсутствует сократительная вакуоля. Как осуществляется у этой крупной инфузории осморегуляция, до сих пор остается не вполне ясным. Под эктоплазмой бурсарии в разных участках тела можно наблюдать различной формы и размеров пузырьки жидкости— вакуоли, которые меняют свой объем. По-видимому, эти неправильной формы вакуоли и соответствуют по своей функции сократительным вакуолям других инфузорий.
Интересно наблюдать за последовательными стадиями бесполого размножения бурсарии. На начальных стадиях деления происходит полная редукция всей полости перистома и околоротовой зоны мембранелл (рис. 94). Сохраняется лишь наружный ресничный покров. Инфузория приобретает форму яйца. После этого тело перешнуровывается поперечной бороздой на две половины. В каждой из образовавшихся дочерних инфузорий путем довольно сложных преобразований развивается типичный перистом и околоротовая зона мембранелл. Весь процесс деления бурсарии протекает быстро и занимает немногим больше часа.
В каждой из образовавшихся дочерних инфузорий путем довольно сложных преобразований развивается типичный перистом и околоротовая зона мембранелл. Весь процесс деления бурсарии протекает быстро и занимает немногим больше часа.
Очень легко наблюдать у бурсарии еще один важный жизненный процесс, наступление которого связано с неблагоприятными для инфузории условиями, — процесс образования цист (инцистирование). Это явление свойственно, например, амебам. Но оказывается, что и такие сложно организованные простейшие, как инфузории, способны переходить в недеятельное состояние. Если культуру, где живут бурсарии, вовремя не покормить или сильно охладить, то уже через несколько часов начнется массовое инцистирование. Протекает этот процесс следующим образом. Бурсарид, так же как перед делением, теряют перистом и околоротовую зону мембранелл. Затем они становятся совершенно шаровидными, после чего выделяют характерной формы двойную оболочку (рис. 94, Г).
В состоянии цист бурсарии могут находиться месяцами. При наступлении благоприятных условий оболочка цисты лопается, бурсарии выходят из нее, развивают перистом и переходят к активной жизни.
При наступлении благоприятных условий оболочка цисты лопается, бурсарии выходят из нее, развивают перистом и переходят к активной жизни.
СТИЛОНИХИЯ (STYLONICHIA MYTILUS)
Очень сложно и разнообразно дифференцированный ресничный аппарат имеют инфузории, относящиеся к отряду брюхоресничных (Hypotricha), многочисленные виды которых обитают как в пресной, так и в морской воде. Одним из наиболее обычных, часто встречающихся представителей этой интересной группы можно назвать стилонихию (Stylonichia mytilus). Это довольно крупная инфузория (длина до 0, 3 мм), живущая на дне пресноводных водоемов, на водной растительности (рис. 95). В отличие от туфельки, трубача и бурсарии у стилонихии отсутствует сплошной ресничный покров, а весь ресничный аппарат представлен ограниченным числом строго определенно расположенных ресничных образований.
Тело стилонихии (как и большинства других брюхоресничных инфузорий) сильно сплющено в спинно-брюшном направлении, и у нее ясно различимы спинная и брюшная стороны, передний и задний концы. Спереди тело несколько расширено, сзади — сужено. При рассматривании животного с брюшной стороны ясно видно, что в передней трети слева расположен сложно устроенный перистом и ротовое отверстие.
Спереди тело несколько расширено, сзади — сужено. При рассматривании животного с брюшной стороны ясно видно, что в передней трети слева расположен сложно устроенный перистом и ротовое отверстие.
На спинной стороне довольно редко расположены реснички, которые не способны к биению. Их скорее можно назвать тонкими эластичными щетинками. Они неподвижны и не имеют отношения к функции движения. Этим ресничкам приписывают обычно осязательную, чувствительную, функцию.
Все ресничные образования, связанные с движением и захватом пищи, сосредоточены на брюшной стороне животного (рис. 95). Здесь имеется небольшое количество толстых пальцевидных образований, расположенных несколькими группами. Это брюшные цирры. Каждая из них — сложное ресничное образование, результат тесного соединения (слипания) многих десятков отдельных ресничек. Таким образом, цирры представляют собой как бы кисточки, отдельные волоски которых тесно сближены и соединены между собой.
При помощи цирр животное довольно быстро передвигается, «бегает» по субстрату. Кроме «ползания» и «бегания» по субстрату стилонихия способна производить довольно резкие и сильные скачки, сразу отрываясь при этом от субстрата. Эти резкие движения осуществляются при помощи двух мощных хвостовых цирр (рис. 95), которые в обычном «ползании» участия не принимают.
Кроме «ползания» и «бегания» по субстрату стилонихия способна производить довольно резкие и сильные скачки, сразу отрываясь при этом от субстрата. Эти резкие движения осуществляются при помощи двух мощных хвостовых цирр (рис. 95), которые в обычном «ползании» участия не принимают.
По краю тела справа и слева расположены два ряда краевых (маргинальных) цирр. С правого края животного они проходят вдоль всего тела, с левого же края доходят лишь до области перистома. Эти ресничные образования служат для поступательного движения животного, когда оно оторвано от субстрата и свободно плавает в воде.
Mы видим, таким образом, что разнообразный и специализированный ресничный аппарат стилонихии позволяет ей делать весьма разнообразные движения в отличие, например, от простого скольжения в воде, как у туфельки или трубача.
Сложно устроен также ресничный аппарат, связанный с функцией питания. Мы видели уже, что околоротовое углубление (перистом), на дне которого помещается ротовое отверстие, ведущее в глотку, расположен в передней половине животного слева. По левому краю начиная с самого переднего конца тела проходит сильно развитая зона околоротовых (адоральных) мембранелл. Своим биением они направляют ток воды в сторону ротового отверстия. Кроме того, в области перистомального углубления расположены еще три сократимые перепонки (мембраны), своими внутренними концами заходящие в глотку, и ряд особых околоротовых ресничек (рис. 95). Весь этот сложный аппарат служит для улавливания и направления пищи в ротовое отверстие.
По левому краю начиная с самого переднего конца тела проходит сильно развитая зона околоротовых (адоральных) мембранелл. Своим биением они направляют ток воды в сторону ротового отверстия. Кроме того, в области перистомального углубления расположены еще три сократимые перепонки (мембраны), своими внутренними концами заходящие в глотку, и ряд особых околоротовых ресничек (рис. 95). Весь этот сложный аппарат служит для улавливания и направления пищи в ротовое отверстие.
Стилонихия относится к числу простейших с очень широким диапазоном пищевых объектов. Ее с полным правом можно назвать всеядным животным. Она может питаться, как и туфелька, бактериями. В число ее пищевых объектов входят жгутиконосцы, одноклеточные водоросли (нередко диатомовые). Наконец, стилонихия может быть и хищником, нападая на другие, более мелкие виды инфузорий и поглощая их.
У стилонихии имеется сократительная вакуоля. Она состоит из центрального резервуара, расположенного у левого заднего конца перистома, и одного направленного назад приводящего канала.
Ядерный аппарат, как всегда у инфузорий, состоит из макронуклеуса и микронуклеуса.
Макронуклеус слагается из двух половинок, соединенных тонкой перетяжкой; микронуклеусов два, они расположены непосредственно около обеих половинок Ма.
Стилонихия, отчасти бурсария, трубач — это все инфузории с широким диапазоном пищевых объектов. Способность поглощать различную пищу свойственна большинству инфузорий. Однако среди них можно найти и такие виды, которые в отношении характера пищи строго специализированы.
ИНФУЗОРИИ-ХИЩНИКИ
Среди инфузорий есть хищники, которые очень «разборчивы» в отношении своей жертвы. Хорошим примером может служить инфузория дидинии (Didinium nasutum). Дидиний — относительно небольшая инфузория, длиной в среднем около 0, 1—0, 15 мм. Передний конец вытянут в виде хоботка, на конце которого помещается ротовое отверстие. Ресничный аппарат представлен двумя венчиками ресничек (рис. 96). Дидиний быстро плавает в воде, часто меняя направление движения. Предпочитаемая пища дидиниев — инфузории туфельки. В данном случае хищник оказывается меньше своей жертвы. Дидиний внедряется в добычу хоботом, а затем, постепенно все более и более расширяя ротовое отверстие, проглатывает туфельку целиком! В хоботке имеется особый, так называемый палочковый, аппарат. Он состоит из ряда эластичных прочных палочек, располагающихся в цитоплазме по периферии хоботка. Предполагают, что этот аппарат увеличивает прочность стенок хоботка, который не разрывается при проглатывании такой огромной по сравнению с дидинием добычи, как туфелька. Дидиний — пример крайнего случая хищничества среди простейших. Если сравнить заглатывание дидинием своей добычи — туфельки —с хищничеством у высших животных, то аналогичные примеры найти трудно.
96). Дидиний быстро плавает в воде, часто меняя направление движения. Предпочитаемая пища дидиниев — инфузории туфельки. В данном случае хищник оказывается меньше своей жертвы. Дидиний внедряется в добычу хоботом, а затем, постепенно все более и более расширяя ротовое отверстие, проглатывает туфельку целиком! В хоботке имеется особый, так называемый палочковый, аппарат. Он состоит из ряда эластичных прочных палочек, располагающихся в цитоплазме по периферии хоботка. Предполагают, что этот аппарат увеличивает прочность стенок хоботка, который не разрывается при проглатывании такой огромной по сравнению с дидинием добычи, как туфелька. Дидиний — пример крайнего случая хищничества среди простейших. Если сравнить заглатывание дидинием своей добычи — туфельки —с хищничеством у высших животных, то аналогичные примеры найти трудно.
Дидиний, проглотивший парамецию, разумеется, очень сильно раздувается. Процесс переваривания протекает очень быстро, при комнатной температуре он занимает всего около двух часов. Затем непереваренные остатки выбрасываются наружу и дидиний начинает охотиться за очередной жертвой. Специальными исследованиями было выяснено, что суточный «рацион» дидиния составляет 12 туфелек — поистине колоссальный аппетит! Нужно иметь в виду, что в промежутках между очередными «охотами» дидинии иногда делятся. При недостатке пищи дидинии очень легко инцистируются и так же легко вновь выходят из цист.
Затем непереваренные остатки выбрасываются наружу и дидиний начинает охотиться за очередной жертвой. Специальными исследованиями было выяснено, что суточный «рацион» дидиния составляет 12 туфелек — поистине колоссальный аппетит! Нужно иметь в виду, что в промежутках между очередными «охотами» дидинии иногда делятся. При недостатке пищи дидинии очень легко инцистируются и так же легко вновь выходят из цист.
РАСТИТЕЛЬНОЯДНЫЕ ИНФУЗОРИИ
Гораздо реже, чем хищничество, встречается среди инфузорий «чистое вегетарианство» — питание исключительно растительной пищей. Одним из немногих примеров инфузорий-«вегетарианцев» могут служить представители рода пассула (Nassula). Объектом питания их являются нитчатые сине-зеленые водоросли (рис. 97).
Они проникают в эндоплазму через рот, расположенный сбоку, а затем закручиваются инфузорией в плотную спираль, которая постепенно переваривается. Пигменты водорослей частично поступают в цитоплазму инфузории и окрашивают ее в яркий темно-зеленый цвет.
СУВОЙКА (VORTICELLA NEBULIFERA)
Интересную и довольно большую по числу видов группу инфузорий составляют сидячие, прикрепленные к субстрату формы, образующие отряд кругоресничных (Peritricha). Широко распространенными представителями этой группы являются сувойки (виды рода Vorticella).
Сувойки напоминают изящный цветок вроде колокольчика или ландыша, сидящий на длинном стебельке, который своим концом прикреплен к субстрату. Большую часть жизни сувойка проводит в прикрепленном к субстрату состоянии.
Рассмотрим строение тела инфузории. У разных видов их размеры варьируют в довольно широких пределах (примерно до 150 мк). Ротовой диск (рис. 98) расположен на расширенном переднем участке тела, которое совершенно лишено ресничек. Ресничный аппарат расположен лишь по краю ротового (перистомального) диска (рис. 98) в особой бороздке, снаружи от которой образуется валик (перистомальная губа). По краю валика идут три мерцательные мембраны, из которых две расположены вертикально, одна (наружная) — горизонтально. Они образуют несколько больше одного полного оборота спирали. Эти мембраны находятся в постоянном мерцательном движении, направляя ток воды к ротовому отверстию. Ротовой аппарат начинается довольно глубоко воронкой у края перистомального поля (рис. 98), в глубине которой располагается ротовое отверстие, ведущее в короткую глотку. Сувойки, так же как и туфельки, питаются бактериями. Их ротовое отверстие постоянно открыто, и возникает непрерывный ток воды в направлении рта.
По краю валика идут три мерцательные мембраны, из которых две расположены вертикально, одна (наружная) — горизонтально. Они образуют несколько больше одного полного оборота спирали. Эти мембраны находятся в постоянном мерцательном движении, направляя ток воды к ротовому отверстию. Ротовой аппарат начинается довольно глубоко воронкой у края перистомального поля (рис. 98), в глубине которой располагается ротовое отверстие, ведущее в короткую глотку. Сувойки, так же как и туфельки, питаются бактериями. Их ротовое отверстие постоянно открыто, и возникает непрерывный ток воды в направлении рта.
Одна сократительная вакуоля без приводящих каналов расположена недалеко от ротового отверстия. Макронуклеус имеет лентовидную или колбасовидную форму, к нему тесно примыкает единственный маленький микронуклеус.
Сувойка способна резко сокращать стебелек, который в долю секунды закручивается штопором. Одновременно с этим сокращается и тело инфузории: перистомальный диск и мембраны втягиваются внутрь и весь передний конец замыкается.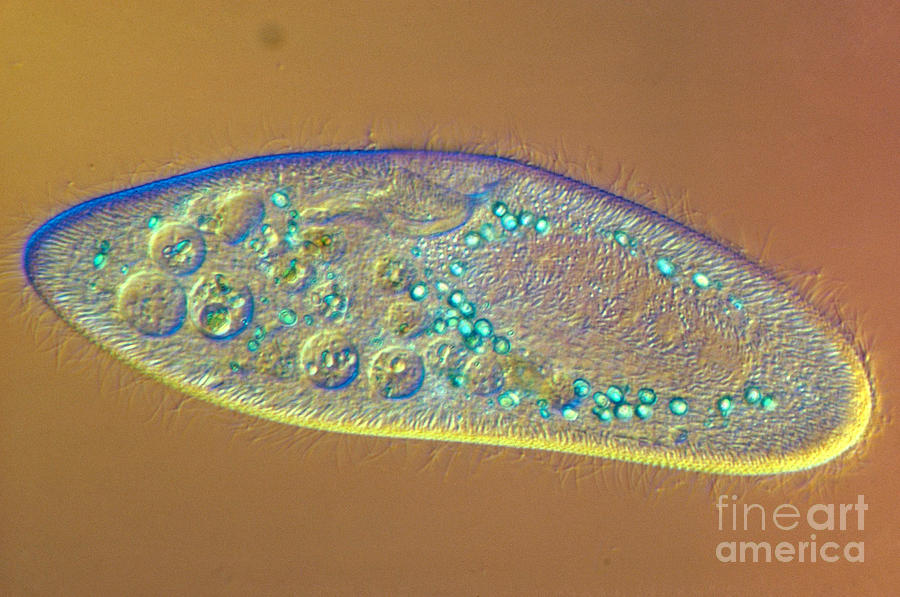
Естественно возникает вопрос: поскольку сувойки прикреплены к субстрату, каким путем осуществляется их расселение по водоему? Это происходит путем образования свободноплавающей стадии— бродяжки. На заднем конце тела инфузории возникает венчик ресничек (рис. 99). Одновременно перистомальный диск втягивается внутрь и инфузория отделяется от стебелька. Образовавшаяся бродяжка способна плавать в течение нескольких часов. Затем события разыгрываются в обратном порядке: инфузория прикрепляется к субстрату задним концом, вырастает стебелек, редуцируется задний венчик ресничек, на переднем конце расправляется перистомальный диск, начинают работать адоральные мембраны. Образование бродяжек у сувойки нередко связано с процессом бесполого размножения. Инфузория на стебельке делится, причем одна из дочерних особей (а иногда и обе) становится бродяжкой и уплывает.
Многие виды сувоек при неблагоприятных условиях способны инцистироваться.
Среди сидячих инфузорий, относящихся к отряду кругоресничных, лишь относительно немногие виды вроде рассмотренных выше сувоек являются одиночно живущими формами. Большая часть относящихся сюда видов — организмы колониальные.
Большая часть относящихся сюда видов — организмы колониальные.
Обычно колониальность возникает в результате не вполне завершенного бесполого или вегетативного размножения. Образующиеся в результате размножения особи в большей или меньшей степени сохраняют связь друг с другом и все вместе образуют органическую индивидуальность высшего порядка, объединяющую большие количества отдельных особей, которая и получает название колонии (с примерами колониальных организмов мы уже познакомились в классе жгутиконосцев.
Колонии кругоресничных инфузорий образуются в результате того, что разделившиеся особи не превращаются в бродяжек, а сохраняют связь друг с другом при помощи стебельков (рис. 100). При этом основной стебелек колонии, так же как и его первые разветвления, не может быть отнесен ни к одной из особей, а принадлежит всей колонии в целом. Иногда колония состоит лишь из небольшого числа особей, у других же видов инфузорий число отдельных особей колонии может достигать нескольких сотен. Однако рост любой колонии не беспределен. По достижении характерных для данного вида размеров колония перестает увеличиваться и образующиеся в результате деления особи развивают венчик ресничек, становятся бродяжками и уплывают, давая начало новым колониям.
Однако рост любой колонии не беспределен. По достижении характерных для данного вида размеров колония перестает увеличиваться и образующиеся в результате деления особи развивают венчик ресничек, становятся бродяжками и уплывают, давая начало новым колониям.
Колонии кругоресничных инфузорий бывают двух типов. У одних стебелек колонии несократим: при раздражении сокращаются лишь отдельные особи колонии, втягивая перистом, вся же колония в целом не претерпевает изменений (к такому типу колоний относятся, например, роды Epistylis, Opercularia). У других (например, род Carchesium) стебелек всей колонии способен сокращаться, так как цитоплазма проходит внутри всех веточек и связывает, таким образом, между собой всех особей колонии. При раздражении таких колоний они сокращаются целиком. Вся колония в данном случае реагирует как единое целое, как органическая индивидуальность.
Среди всех колониальных кругоресничных инфузорий особый интерес представляет, пожалуй, зоотамний (Zoothamnium arbuscula). Колонии этой инфузории отличаются особенной правильностью строения. Кроме того, в пределах колонии здесь намечается интересное биологическое явление полиморфизма.
Колонии этой инфузории отличаются особенной правильностью строения. Кроме того, в пределах колонии здесь намечается интересное биологическое явление полиморфизма.
Колония зоотамния имеет вид зонтика. На одном, главном, стебельке колонии располагаются вторичные ветви (рис. 101). Размер взрослой колонии 2—3 мм, так что они хорошо видны простым глазом. Живут зоотамний в небольших прудах с чистой водой. Колонии их обычно находят на подводных растениях, чаще всего на элодее (водяной чуме).
Стебельки колонии зоотамния обладают сократимостью, так как сократимая цитоплазма проходит через все ветви колонии, за исключением базальной части главного стебелька. При сокращении, которое происходит очень быстро и резко, вся колония собирается в комочек.
Для зоотамния характерно строго закономерное расположение ветвей. К субстрату прикрепляется один главный стебелек. От верхней части его в плоскости, перпендикулярной к стебельку, отходит девять главных ветвей колонии, строго закономерно расположенных друг относительно друга (рис.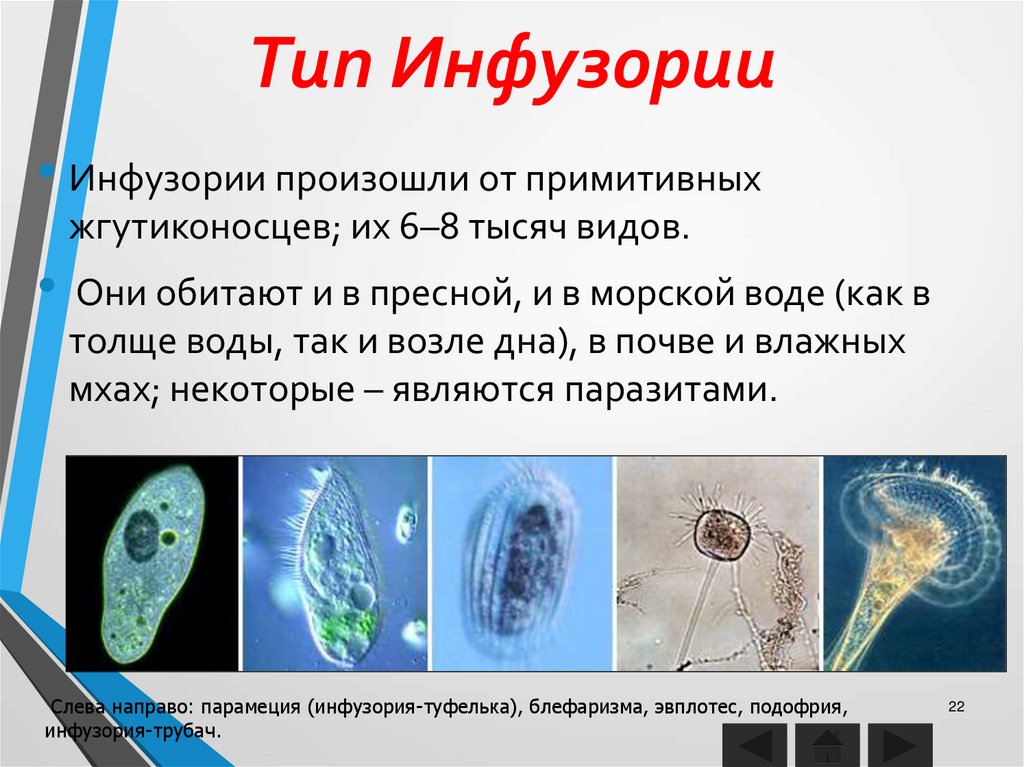 102, 6). От этих ветвей отходят вторичные веточки, на которых сидят отдельные особи колонии. На каждой вторичной веточке может быть до 50 инфузорий. Общее количество особей в составе колонии достигает 2—3 тыс.
102, 6). От этих ветвей отходят вторичные веточки, на которых сидят отдельные особи колонии. На каждой вторичной веточке может быть до 50 инфузорий. Общее количество особей в составе колонии достигает 2—3 тыс.
Большая часть особей колонии по своему строению напоминает небольших одиночных сувоек, размером 40—60 мк. Но кроме мелких особей, которые называются микрозоидами, на взрослых колониях, примерно на середине главных ветвей, развиваются особи совершенно другого вида и размера (рис. 102, 5). Это крупные шаровидные особи диаметром 200—250 мк, превосходящие по своей массе объем микрозоида в сотню и более раз. Крупные особи получили название макрозоидов.
По своему строению они существенно отличаются от мелких особей колонии. Перистом у них не выражен: он втягивается внутрь и не функционирует. Макрозоид с самого начала своего развития из микрозоида перестает самостоятельно принимать пищу. У него отсутствуют пищеварительные вакуоли. Рост макрозоида осуществляется, очевидно, за счет веществ, поступающих через цитоплазматические мостики, соединяющие между собой всех особей колонии. В участке тела макрозоида, которым он прикрепляется к стебельку, имеется скопление особых зернышек (гранул), которые, как увидим, в его дальнейшей судьбе играют существенную роль. Что же представляют собой эти крупные шаровидные макрозоиды, какова их биологическая роль в жизни колонии зоотамния? Наблюдение показывает, что макрозоиды — это будущие бродяжки, из которых развиваются новые колонии. Достигнув предельного размера, макрозоид развивает венчик ресничек, отделяется от колонии и уплывает. Его форма при этом несколько меняется, из шаровидного он становится коническим. Через некоторое время бродяжка прикрепляется к субстрату всегда той стороной, на которой расположена зернистость. Сразу же начинается образование и рост стебелька, причем на построение стебелька затрачиваются гранулы, которые локализованы на заднем конце бродяжки. По мере роста стебелька зернистость исчезает. После того как стебелек достигнет окончательной, характерной для зоотамния длины, начинается ряд быстро следующих друг за другом делений, ведущих к образованию колонии.
В участке тела макрозоида, которым он прикрепляется к стебельку, имеется скопление особых зернышек (гранул), которые, как увидим, в его дальнейшей судьбе играют существенную роль. Что же представляют собой эти крупные шаровидные макрозоиды, какова их биологическая роль в жизни колонии зоотамния? Наблюдение показывает, что макрозоиды — это будущие бродяжки, из которых развиваются новые колонии. Достигнув предельного размера, макрозоид развивает венчик ресничек, отделяется от колонии и уплывает. Его форма при этом несколько меняется, из шаровидного он становится коническим. Через некоторое время бродяжка прикрепляется к субстрату всегда той стороной, на которой расположена зернистость. Сразу же начинается образование и рост стебелька, причем на построение стебелька затрачиваются гранулы, которые локализованы на заднем конце бродяжки. По мере роста стебелька зернистость исчезает. После того как стебелек достигнет окончательной, характерной для зоотамния длины, начинается ряд быстро следующих друг за другом делений, ведущих к образованию колонии.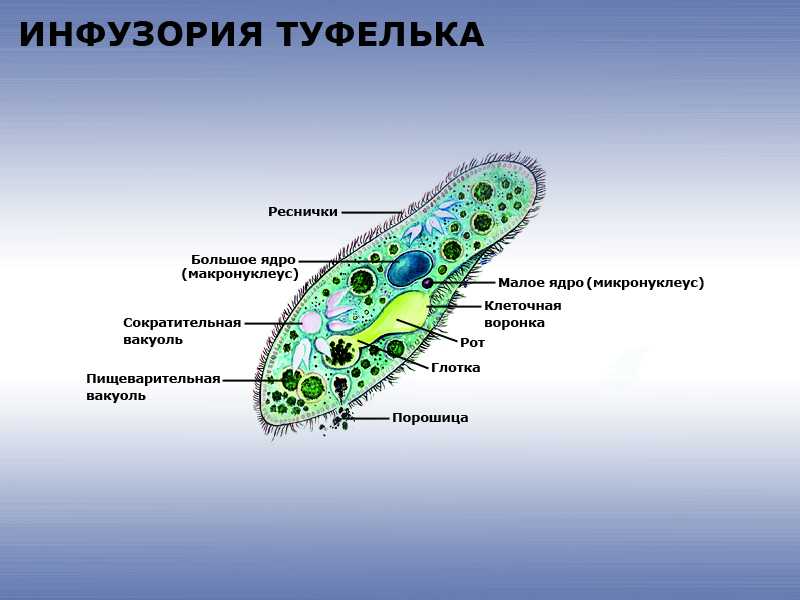 Деления эти совершаются в строго определенной последовательности (рис. 102).
Деления эти совершаются в строго определенной последовательности (рис. 102).
На рассмотрении деталей этого процесса мы останавливаться не будем. Обратим лишь внимание на следующее интересное явление. Во время первых делений бродяжки зоотамния при развитии колонии у образующих особей перистом и рот не функционируют. Питание начинается позже, когда молодая колония состоит уже из 12—16 особей. Таким образом, все первые стадии развития колонии осуществляются исключительно за счет тех запасов, которые образовались в теле макрозоида в период его роста и развития на материнской колонии. Существует бесспорное сходство между развитием бродяжки зоотамния и развитием яйца у многоклеточных животных. Это сходство выражается в том, что развитие в обоих случаях осуществляется за счет ранее накопленных запасов, без восприятия пищи из внешней среды.
При изучении сидячих кругоресничных инфузорий возникает вопрос: а как же осуществляется у них характерная для инфузорий форма полового процесса — конъюгация? Оказывается, что в связи с сидячим образом жизни она претерпевает некоторые существенные изменения. К началу полового процесса на колонии образуются особые, очень мелкие бродяжки. Активно двигаясь при помощи венчика ресничек, они ползают в течение некоторого времени по колонии, а затем вступают в конъюгацию с крупными нормальными сидячими особями колонии. Таким образом, здесь происходит дифференцировка конъюгантов на две группы особей: мелкие, подвижные (микроконъюганты) и более крупные, неподвижные (макроконъюганты). Эта дифференцировка конъюгантов на две категории, из которых одна (микроконъюганты) подвижная, явилась необходимым приспособлением к сидячему образу жизни. Без этого нормальный ход полового процесса (конъюгации) не мог быть, очевидно, обеспечен.
К началу полового процесса на колонии образуются особые, очень мелкие бродяжки. Активно двигаясь при помощи венчика ресничек, они ползают в течение некоторого времени по колонии, а затем вступают в конъюгацию с крупными нормальными сидячими особями колонии. Таким образом, здесь происходит дифференцировка конъюгантов на две группы особей: мелкие, подвижные (микроконъюганты) и более крупные, неподвижные (макроконъюганты). Эта дифференцировка конъюгантов на две категории, из которых одна (микроконъюганты) подвижная, явилась необходимым приспособлением к сидячему образу жизни. Без этого нормальный ход полового процесса (конъюгации) не мог быть, очевидно, обеспечен.
СОСУЩИЕ ИНФУЗОРИИ (SUCTORIA)
Весьма своеобразную в отношении способа питания группу представляют сосущие инфузории (Suctoria). Эти организмы, подобно сувойкам и другим кругоресничным инфузориям, сидячие. Количество относящихся к этому отряду видов измеряется несколькими десятками.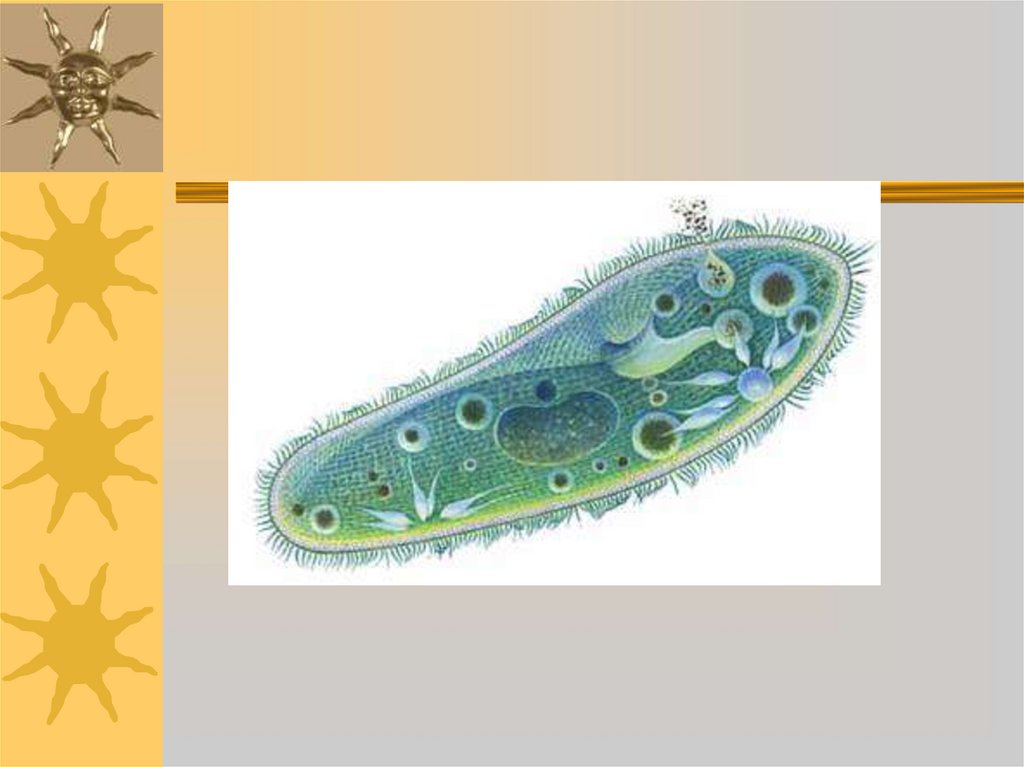 Форма тела сосущих инфузорий весьма разнообразна. Некоторые характерные виды их изображены на рисунке 103. Одни сидят на субстрате на более или менее длинных стебельках, другие не имеют стебельков, у некоторых тело довольно сильно ветвится и т. п. Однако, несмотря на разнообразие формы, все сосущие инфузории характеризуются следующими двумя признаками:
Форма тела сосущих инфузорий весьма разнообразна. Некоторые характерные виды их изображены на рисунке 103. Одни сидят на субстрате на более или менее длинных стебельках, другие не имеют стебельков, у некоторых тело довольно сильно ветвится и т. п. Однако, несмотря на разнообразие формы, все сосущие инфузории характеризуются следующими двумя признаками:
1) полным отсутствием (у взрослых форм) ресничного аппарата,
2) наличием особых придатков — щупалец, служащих для высасывания добычи.
У разных видов сосущих инфузорий количество щупалец неодинаково. Нередко они бывают собраны группами. При большом увеличении микроскопа можно рассмотреть, что на конце щупальце снабжено небольшим булавовидным утолщением.
Как функционируют щупальца? На этот вопрос нетрудно ответить, наблюдая в течение некоторого времени за сосущими инфузориями. Если какое-нибудь небольшое простейшее (жгутиконосец, инфузория) прикоснется к щупальцу суктории, то оно к нему мгновенно прилипнет.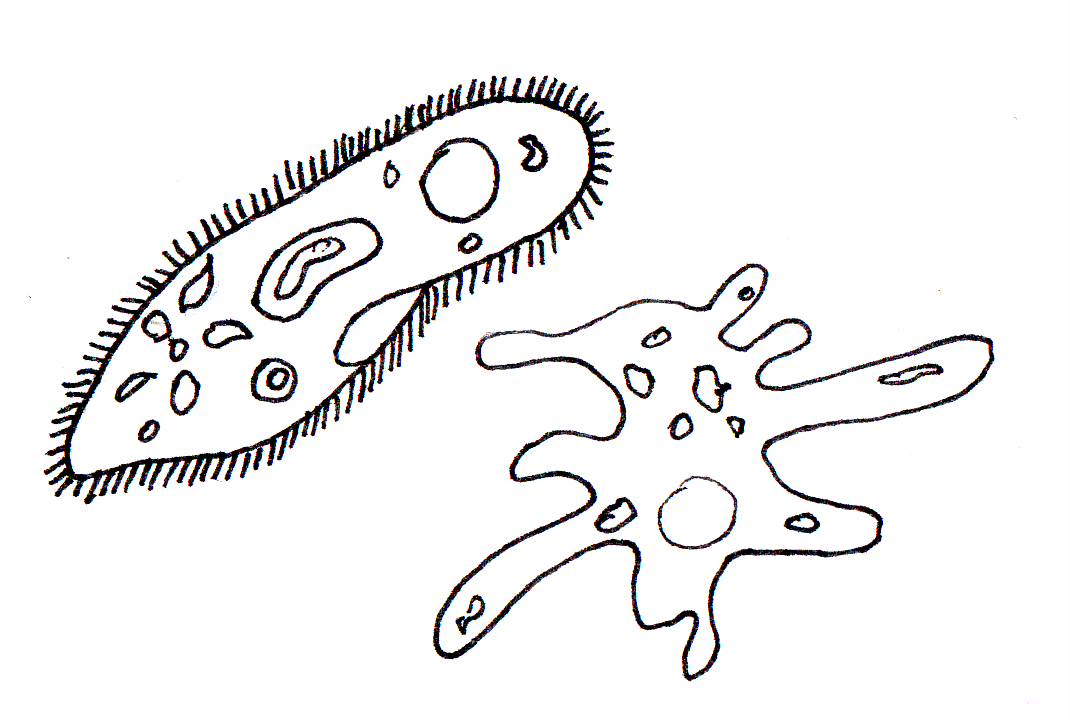 Все попытки жертвы оторваться бывают обычно тщетными. Если продолжить наблюдение за прилипшей к щупальцам жертвой, то можно видеть, что она постепенно начинает уменьшаться в размерах. Ее содержимое через шупальца постепенно «перекачивается» внутрь эндоплазмы сосущей инфузории до тех пор, пока от жертвы не останется одна пелликула, которая отбрасывается. Таким образом, щупальца сосущих инфузорий — это совершенно своеобразные, нигде больше в животном мире не встречающиеся органы улавливания и вместе с тем высасывания пищи (рис. 103).
Все попытки жертвы оторваться бывают обычно тщетными. Если продолжить наблюдение за прилипшей к щупальцам жертвой, то можно видеть, что она постепенно начинает уменьшаться в размерах. Ее содержимое через шупальца постепенно «перекачивается» внутрь эндоплазмы сосущей инфузории до тех пор, пока от жертвы не останется одна пелликула, которая отбрасывается. Таким образом, щупальца сосущих инфузорий — это совершенно своеобразные, нигде больше в животном мире не встречающиеся органы улавливания и вместе с тем высасывания пищи (рис. 103).
Сосущие инфузории — это неподвижные хищники, которые не гоняются за добычей, но мгновенно улавливают ее, если только неосторожная добыча к ним прикоснется сама.
Почему же эти своеобразные организмы мы относим к классу инфузорий? На первый взгляд они не имеют с ними ничего общего. О принадлежности сукторий к инфузориям говорят следующие факты. Во-первых, они обладают типичным для инфузорий ядерным аппаратом, состоящим из макронуклеуса и микронуклеуса. Во-вторых, во время размножения у них появляются реснички, отсутствующие «у взрослых» особей. Бесполое размножение и вместе с тем расселение сосущих инфузорий осуществляется путем образования бродяжек, снабженных несколькими кольцевыми венчиками ресничек. Образование бродяжек у сукторий может происходить по-разному. Иногда они образуются в результате не вполне равномерного деления (почкованием), при котором каждая отделяющаяся наружу почка получает участок макронуклеуса и один микронуклеус (рис. 104, Л). На одной материнской особи может образоваться сразу несколько дочерних почек (рис. 104, 5). У других видов {рис. 104, Г, Д) наблюдается очень своеобразный способ «внутреннего почкования». При этом внутри тела суктории-матери образуется полость, в которой и формируется почка-бродяжка. Наружу она выходит через специальные отверстия, сквозь которые с известным трудом «протискивается».
Во-вторых, во время размножения у них появляются реснички, отсутствующие «у взрослых» особей. Бесполое размножение и вместе с тем расселение сосущих инфузорий осуществляется путем образования бродяжек, снабженных несколькими кольцевыми венчиками ресничек. Образование бродяжек у сукторий может происходить по-разному. Иногда они образуются в результате не вполне равномерного деления (почкованием), при котором каждая отделяющаяся наружу почка получает участок макронуклеуса и один микронуклеус (рис. 104, Л). На одной материнской особи может образоваться сразу несколько дочерних почек (рис. 104, 5). У других видов {рис. 104, Г, Д) наблюдается очень своеобразный способ «внутреннего почкования». При этом внутри тела суктории-матери образуется полость, в которой и формируется почка-бродяжка. Наружу она выходит через специальные отверстия, сквозь которые с известным трудом «протискивается».
Такое развитие зародыша внутри тела матери, а затем акт деторождения — интересная аналогия простейшего с тем, что происходит у вышестоящих многоклеточных организмов.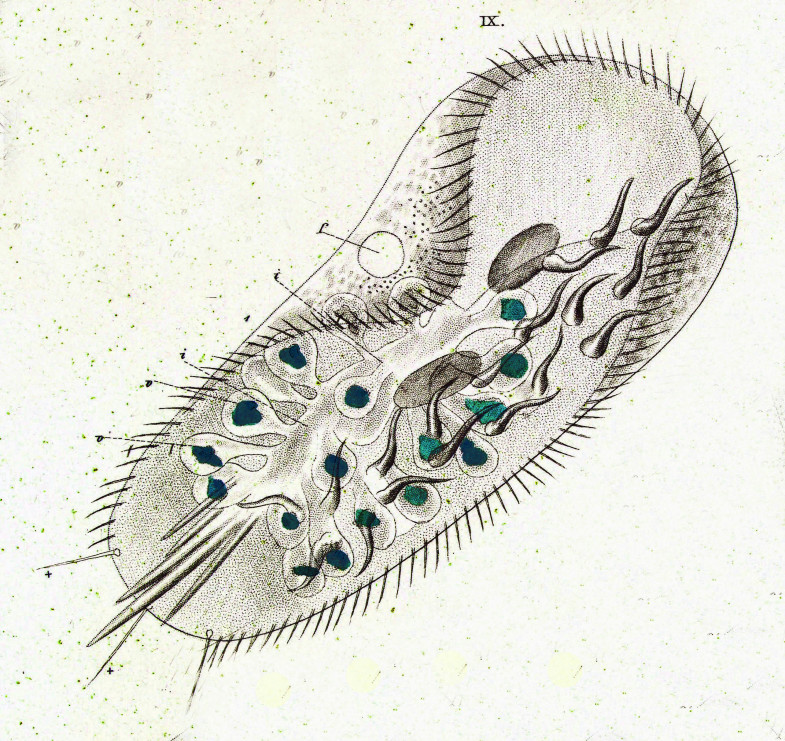
На предыдущих страницах было рассмотрено несколько типичных свободноживущих представителей класса инфузорий, по-разному приспособленных к различным условиям среды. Интересно подойти к вопросу приспособления инфузорий к условиям жизни и, с другой стороны, посмотреть, каковы характерные общие черты инфузорий, живущих в определенных, резко очерченных условиях среды.
В качестве примера возьмем две очень резко различающиеся среды обитания: жизнь в составе планктона и жизнь на дне в толще песка.
ПЛАНКТОННЫЕ ИНФУЗОРИИ
В составе как морского, так и пресноводного планктона встречается довольно большое число видов инфузорий.
Особенно ярко выражены черты приспособлений к жизни в толще воды у радиолярий. Основная линия приспособления к планктонному образу жизни сводится к выработке таких черт строения, которые способствуют парению организма в толще воды.
Типичным планктонным, к тому же почти исключительно морским семейством инфузорий является тинтинниды (Tintinnidae, рис. 105, 5). Общее число извест ных до сих пор видов тинтиннид около 300. Это мелкие формы, характерные тем, что протоплазматическое тело инфузории помещается в прозрачном, легком и вместе с тем прочном домике, состоящем из органического вещества. Из домика выдается наружу диск, несущий венчик ресничек, находящихся в постоянном мерцательном движении. В состоянии парения инфузорий в толще воды поддерживает здесь главным образом постоянная активная работа ресничного аппарата. Домик, очевидно, выполняет функцию защиты нижней части тела инфузории. В пресной воде обитает всего 2 вида тинтиннид (не считая 7 видов, характерных только для озера Байкал).
105, 5). Общее число извест ных до сих пор видов тинтиннид около 300. Это мелкие формы, характерные тем, что протоплазматическое тело инфузории помещается в прозрачном, легком и вместе с тем прочном домике, состоящем из органического вещества. Из домика выдается наружу диск, несущий венчик ресничек, находящихся в постоянном мерцательном движении. В состоянии парения инфузорий в толще воды поддерживает здесь главным образом постоянная активная работа ресничного аппарата. Домик, очевидно, выполняет функцию защиты нижней части тела инфузории. В пресной воде обитает всего 2 вида тинтиннид (не считая 7 видов, характерных только для озера Байкал).
У пресноводных инфузорий наблюдаются некоторые другие приспособления к жизни в планктоне. У многих из них цитоплазма очень сильно вакуолизирована (Loxodes, Condylostoma, Trachelius), так что напоминает пену. Это ведет к значительному уменьшению удельного веса. Все перечисленные инфузории обладают, кроме того, ресничным покровом, благодаря работе которого тело инфузории, по удельному весу лишь немногим превышающее удельный вес воды, легко поддерживается в состоянии «парения». У некоторых видов форма тела способствует увеличению удельной поверхности и облегчает парение в воде. Например, некоторые планктонные инфузории озера Байкал напоминают по форме зонтик или парашют (Liliomorpha, рис. 105, 2). Существует в озере Байкал одна планктонная сосущая инфузория (Mucophrya pelagica, рис. 105, 4), которая резко отличается от своих сидячих родичей. Этот вид лишен стебелька. Его протоплазматическое тело окружено широким слизистым футляром — приспособлением, ведущим к уменьшению веса. Наружу торчат длинные тонкие щупальца, которые наряду со своей прямой функцией, вероятно, выполняют и другую — увеличение удельной поверхности, способствующее парению в воде.
У некоторых видов форма тела способствует увеличению удельной поверхности и облегчает парение в воде. Например, некоторые планктонные инфузории озера Байкал напоминают по форме зонтик или парашют (Liliomorpha, рис. 105, 2). Существует в озере Байкал одна планктонная сосущая инфузория (Mucophrya pelagica, рис. 105, 4), которая резко отличается от своих сидячих родичей. Этот вид лишен стебелька. Его протоплазматическое тело окружено широким слизистым футляром — приспособлением, ведущим к уменьшению веса. Наружу торчат длинные тонкие щупальца, которые наряду со своей прямой функцией, вероятно, выполняют и другую — увеличение удельной поверхности, способствующее парению в воде.
Нужно, наконец, упомянуть еще об одной, так сказать, косвенной форме приспособления инфузорий к жизни в планктоне. Это прикрепление мелких инфузорий к другим организмам, ведущим планктонный образ жизни. Так, среди кругоресничных инфузорий (Peritricha) имеются довольно многочисленные виды, которые прикрепляются к планктонным веслоногим рачкам. Это обычный и нормальный для данных видов инфузорий образ жизни.
Это обычный и нормальный для данных видов инфузорий образ жизни.
Наряду с кругоресничными инфузориями и среди сосущих (Suctoria) имеются виды, поселяющиеся на планктонных организмах.
ИНФУЗОРИИ, ЖИВУЩИЕ В ПЕСКЕ
Чрезвычайно своеобразную среду обитания представляют песчаные пляжи и отмели. По побережью морей они занимают огромные пространства и характеризуются своеобразной фауной.
Проведенные за последние годы в различных странах многочисленные исследования показали, что толща многих морских песков очень богата разнообразной микроскопической или приближающейся по своим размерам к микроскопической фауной. Между частичками песка имеются многочисленные заполненные водой мелкие и мельчайшие пространства. Оказывается, что эти пространства богато заселены организмами, относящимися к самым различным группам животного мира. Здесь живут десятки видов ракообразных, кольчатые черви, круглые черви, особенно многочисленные плоские черви, некоторые моллюски, кишечнополостные.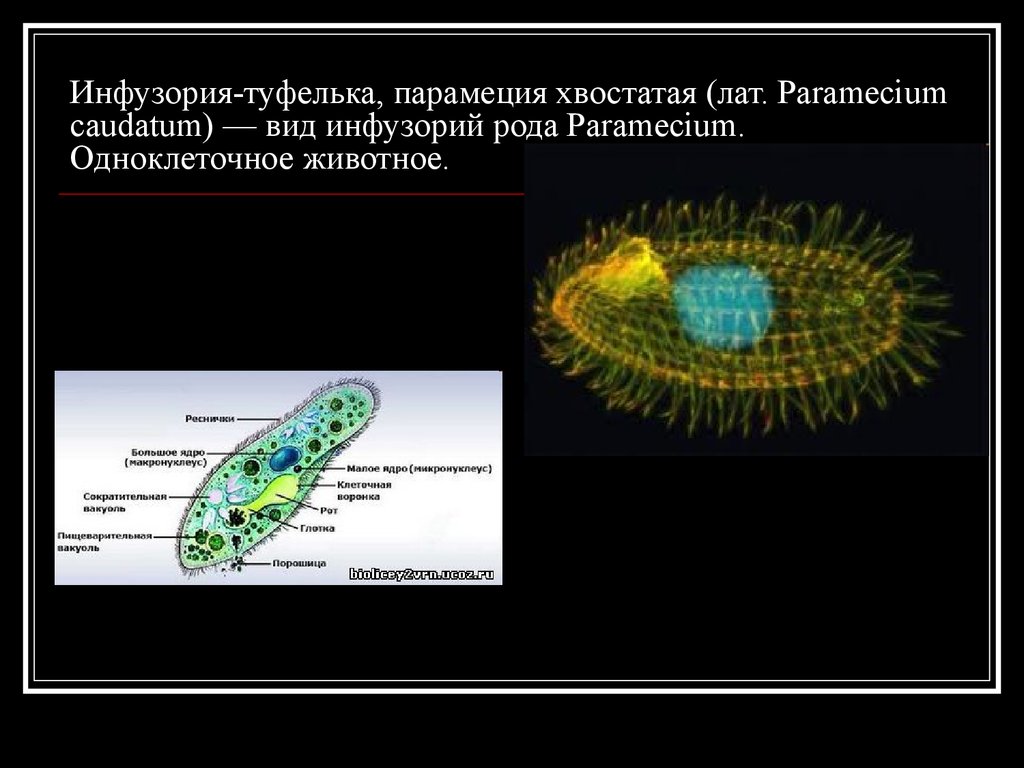 В большом количестве здесь встречаются и простейшие, главным образом инфузории. По современным данным, в состав фауны инфузорий, населяющих толщу морских песков, входит примерно 250—300 видов. Если иметь в виду не только инфузорий, а и другие группы населяющих толщу песка организмов, то общее число видов их будет очень велико. Всю совокупность животных, населяющих толщу песка, живущих в мельчайших просветах между песчинками, называют псаммофил ьной фауной.
В большом количестве здесь встречаются и простейшие, главным образом инфузории. По современным данным, в состав фауны инфузорий, населяющих толщу морских песков, входит примерно 250—300 видов. Если иметь в виду не только инфузорий, а и другие группы населяющих толщу песка организмов, то общее число видов их будет очень велико. Всю совокупность животных, населяющих толщу песка, живущих в мельчайших просветах между песчинками, называют псаммофил ьной фауной.
Богатство и видовой состав псаммофильной фауны определяется многими факторами. Среди них особенно важное значение имеет размер частиц песка. Крупнозернистые пески имеют бедную фауну. Бедна также фауна очень мелкозернистых заиленных песков (с диаметром частиц менее 0, 1 мм), где, очевидно, просветы между частицами слишком мелки для обитания в них животных. Наиболее богаты жизнью пески средне- и мелкозернистые.
Второй фактор, играющий важную роль в развитии псаммофильной фауны, — богатство песка органическими остатками, разлагающимися органическими веществами (так называемая степень сапробности). Пески, лишенные органических веществ, бедны жизнью. С другой стороны, почти безжизненны и пески, очень богатые органическими веществами, так как распад органических веществ ведет к обеднению кислородом. Нередко к этому добавляется анаэробное сероводородное брожение.
Пески, лишенные органических веществ, бедны жизнью. С другой стороны, почти безжизненны и пески, очень богатые органическими веществами, так как распад органических веществ ведет к обеднению кислородом. Нередко к этому добавляется анаэробное сероводородное брожение.
Наличие свободного сероводорода является крайне отрицательно действующим на развитие фауны фактором.
В поверхностных слоях песка иногда развивается довольно богатая флора одноклеточных водорослей (диатомовые, перидиниевые). Это фактор, благоприятствующий развитию псаммофильной фауны, так как многие мелкие животные (в том числе и инфузории) питаются водорослями.
Наконец, фактор, очень отрицательно действующий на псаммофильную фауну, — прибой. Это вполне понятно, так как прибой, перемывающий верхние слои песка, убивает здесь все живое. Наиболее богата псаммофильная фауна в защищенных, хорошо прогреваемых бухточках. Приливы и отливы не препятствуют развитию псаммофильной фауны. Когда в отлив вода временно уходит, обнажая песок, то в толще песка, в промежутках между песчинками, она сохраняется, и это не препятствует существованию животных.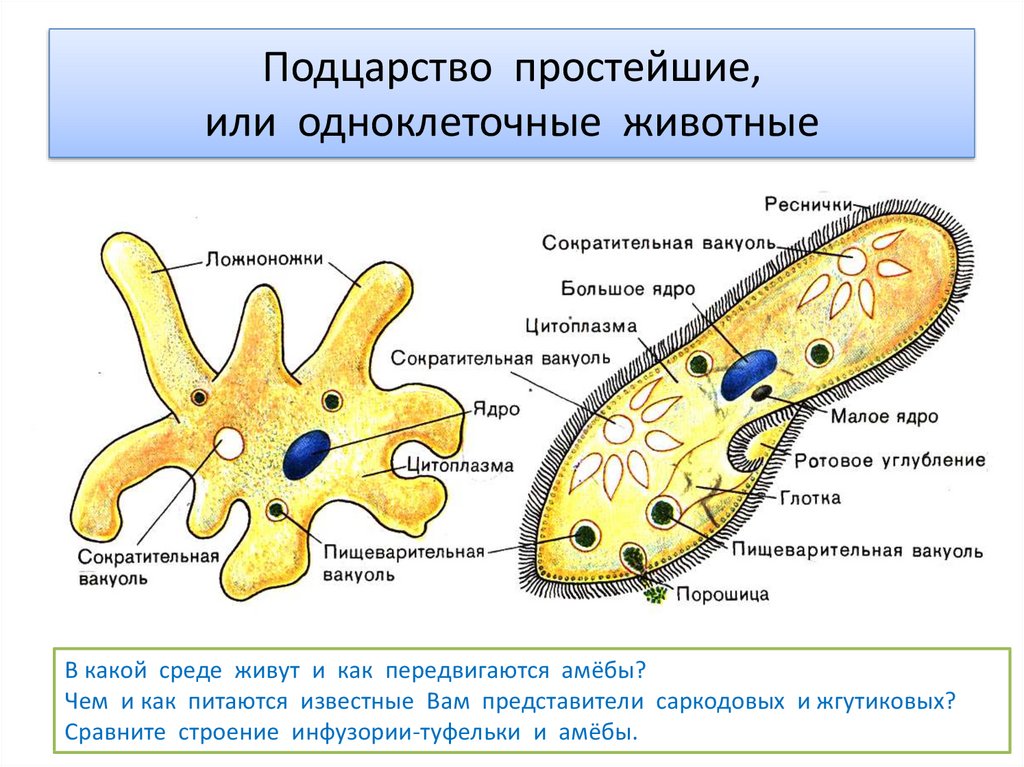
У инфузорий, входящих в состав псаммофильной фауны и относящихся к различным систематическим группам (отрядам, семействам), вырабатываются в процессе эволюции многие общие черты, являющиеся приспособлениями к своеобразным условиям существования между частицами песка.
На рисунке 106 изображены некоторые виды псаммофильной фауны инфузорий, относящиеся к разным отрядам и семействам. Между ними много сходства. Тело большинства их более или менее сильно вытянуто в длину, червеобразно. Это дает возможность легко «протискиваться» в мельчайшие отверстия между песчинками. У очень многих видов (рис. 106) удлинение тела сочетается с его уплощением. Ресничный аппарат всегда хорошо развит, что позволяет активно, с известной силой, двигаться в узких просветах. Нередко реснички развиваются на одной стороне червеобразного уплощенного тела, противоположная сторона оказывается голой. Эта особенность связана, вероятно, с резко выраженной у большинства псаммофильных в»идов способностью очень тесно и очень прочно при посредстве ресничного аппарата приставать (прикрепляться) к субстрату (явление, называемое тигмотаксисом). Это свойство позволяет животным оставаться на месте в тех случаях, когда Б узких просветах, где они живут, возникают токи воды. При этом, вероятно, выгоднее, чтобы сторона, противоположная той, которой животное прикрепилось к субстрату, была гладкой.
Это свойство позволяет животным оставаться на месте в тех случаях, когда Б узких просветах, где они живут, возникают токи воды. При этом, вероятно, выгоднее, чтобы сторона, противоположная той, которой животное прикрепилось к субстрату, была гладкой.
Чем питаются псаммофильные инфузории? Значительную часть «рациона» у многих видов составляют водоросли, в особенности диатомовые. Бактерии в меньшей степени служат им пищей. Это зависит в значительной мере и от того, что в песках, не сильно загрязненных, бактерий мало. Наконец, особенно среди наиболее крупных псаммофильных инфузорий имеется немалое количество хищных форм, которые поедают других инфузорий, относящихся к более мелким видам. Псаммофильные инфузории распространены, по-видимому, повсеместно.
ИНФУЗОРИИ АПОСТОМАТЫ
В процессе приспособления к условиям жизни очень сложные жизненные циклы, сопровождающиеся сменой нескольких форм строения, выработались у своеобразной морской группы инфузорий — апостомат (Apostomata). Сложность и своеобразие жизненного пути этих инфузорий зависит от того, что часть стадий цикла они проводят как свободноживущие организмы, другие же стадии являются паразитическими или полупаразитическими. Рассмотрим на конкретном примере жизненный цикл апостомат.
Сложность и своеобразие жизненного пути этих инфузорий зависит от того, что часть стадий цикла они проводят как свободноживущие организмы, другие же стадии являются паразитическими или полупаразитическими. Рассмотрим на конкретном примере жизненный цикл апостомат.
Инфузория спирофрия (Spirophrya subparasitica) в инцистированном состоянии часто может быть обнаружена сидящей на небольшой ножке на мелких планктонных морских ракообразных (особенно часто на рачках рода Idia). Пока рачок активно плавает в морской воде, сидящие на нем спирофрии не претерпевают никаких изменений. Для дальнейшего развития инфузорий необходимо, чтобы рачок был съеден морским гидроидным полипом, что случается нередко (рис. 107). Как только цисты спирофрии вместе с рачком проникнут в пищеварительную полость, из них сразу же выходят мелкие инфузории, которые начинают энергично питаться пищевой кашицей, образующейся в результате переваривания заглоченного рачка. В течение часа размеры инфузории увеличиваются в 3—4 раза.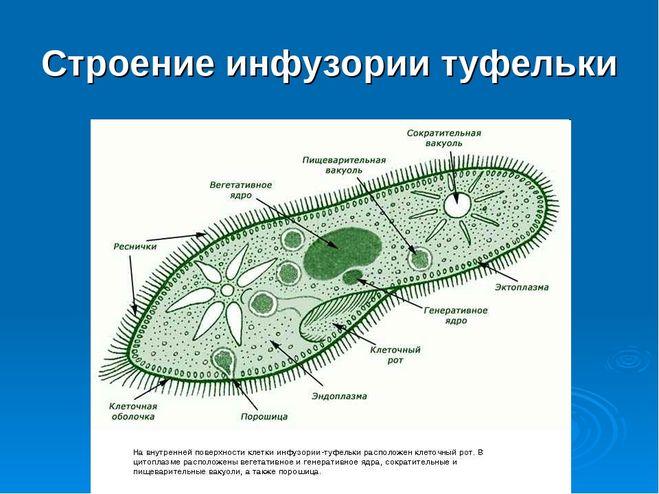 Однако размножения на этой стадии не происходит. Перед нами типичная стадия роста инфузории, которую называют трофонтом. Через некоторое время вместе с непереваренными остатками пищи трофонт выбрасывается полипом наружу в морскую воду. Здесь он, активно плавая, спускается по телу полипа к его подошве, где и прикрепляется, окружаясь при этом цистой. Эта стадия инцистированной, сидящей на полипе крупной инфузории получила название томонта. Это фаза размножения. Томонт не питается, но быстро последовательно несколько раз делится (рис. 107, 7). В результате получается целая группа очень мелких инфузорий. Количество их зависит от размеров томонта, который определяется в свою очередь величиной трофонта, давшего ему начало. Мелкие инфузории, образовавшиеся в результате деления томонта (они называются томитами или бродяжками), представляют собой стадию расселения.
Однако размножения на этой стадии не происходит. Перед нами типичная стадия роста инфузории, которую называют трофонтом. Через некоторое время вместе с непереваренными остатками пищи трофонт выбрасывается полипом наружу в морскую воду. Здесь он, активно плавая, спускается по телу полипа к его подошве, где и прикрепляется, окружаясь при этом цистой. Эта стадия инцистированной, сидящей на полипе крупной инфузории получила название томонта. Это фаза размножения. Томонт не питается, но быстро последовательно несколько раз делится (рис. 107, 7). В результате получается целая группа очень мелких инфузорий. Количество их зависит от размеров томонта, который определяется в свою очередь величиной трофонта, давшего ему начало. Мелкие инфузории, образовавшиеся в результате деления томонта (они называются томитами или бродяжками), представляют собой стадию расселения.
Они выходят из цисты, быстро плавают (не питаясь при этом, а используя имеющиеся у них в цитоплазме запасы). Если им «посчастливится» натолкнуться на веслоногого рачка, то они тотчас же прикрепляются к нему и инцистируются. Это стадия, с которой мы начали рассмотрение цикла.
Это стадия, с которой мы начали рассмотрение цикла.
В рассмотренном нами жизненном цикле спирофрии обращает на себя внимание резкая разграниченность стадий, имеющих разное биологическое значение. Трофонт — это стадия роста. Он только растет, накапливает большое количество цитоплазмы и всякого рода резервных веществ за счет энергичного и быстрого питания. К размножению трофонт не способен. Обратное явление наблюдается у томонта — неспособность питаться и энергичное быстрое размножение. После каждого деления не происходит роста, и поэтому размножение томонта сводится к быстрому распаду на множество бродяжек. Наконец, бродяжки выполняют свою специальную и только им свойственную функцию: это особи — расселительницы и распространительницы вида. Они не способны ни питаться, ни размножаться.
ЖИЗНЕННЫЙ ЦИКЛ ИХТИОФТИРИУСА
Среди инфузорий, часть жизненного цикла которых протекает в паразитическом, часть в свободноживущем состоянии, большой интерес, особенно в практическом отношении, представляет ихтиофтириус (Ichthyophthirius multifiliis).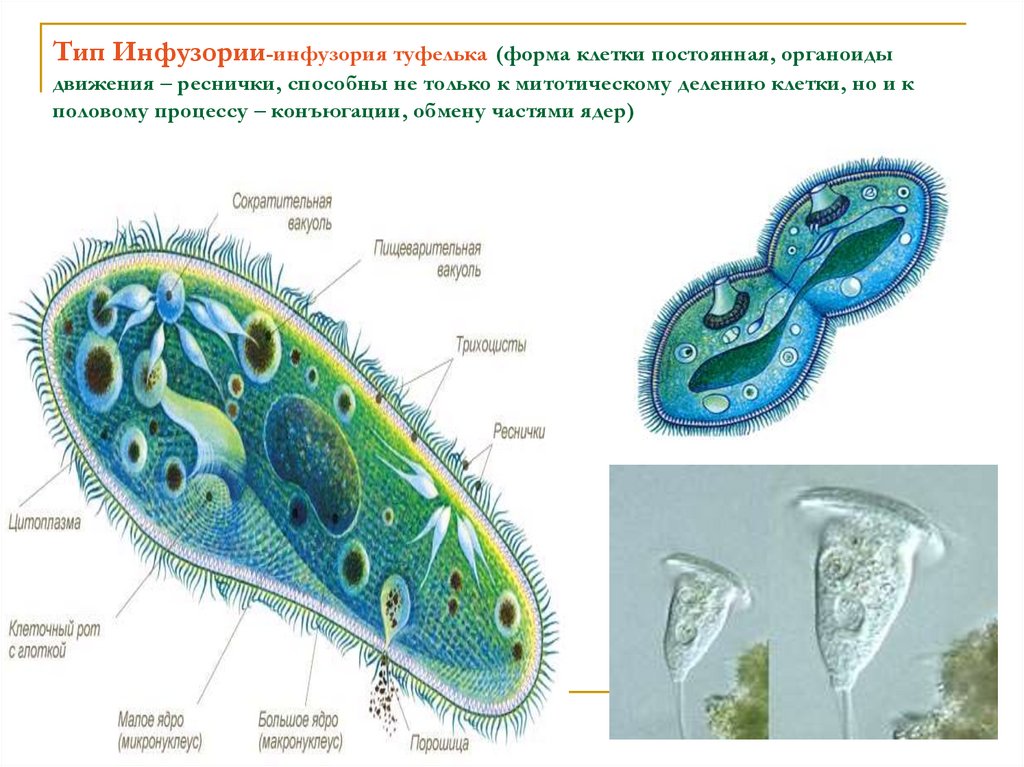 Эта инфузория в течение большей части своей жизни является паразитом рыб. Она может вызвать тяжелое заболевание, в особенности поражающее мальков, часто приводящее к гибели хозяина (рис. 108).
Эта инфузория в течение большей части своей жизни является паразитом рыб. Она может вызвать тяжелое заболевание, в особенности поражающее мальков, часто приводящее к гибели хозяина (рис. 108).
Паразитарные стадии развития ихтиофтириуса протекают непосредственно в покровах рыбы: в коже, на плавниках, очень часто на жабрах рыбы.
Рыба заражается очень мелкими свободноплавающими бродяжками (20—30 мк диаметром), плавающими в воде. Бродяжки при соприкосновении с рыбой немедленно прикрепляются к ее поверхности и сразу же активными движениями внедряются в ткани. Проникнув в живую ткань, инфузории начинают активно питаться и расти. Питание осуществляется путем заглатывания при помощи помещающегося на переднем конце тела ротового отверстия клеточных элементов хозяина. Растущие в тканях рыбы ихтиофтириусы вскоре становятся видны простым глазом. Рыба как бы обсыпана манной крупой (рис. 108). Продолжительность стадии роста и паразитирования ихтиофтириуса в субэпителиальном слое рыбы в высокой степени зависит от температуры. По исследованиям Бауера, при 28—30°С этот срок составляет 4 суток, при 19—21°С—7Г при 13—15°С-10-11, а при 8-9°С период этот затягивается до 40 дней.
По исследованиям Бауера, при 28—30°С этот срок составляет 4 суток, при 19—21°С—7Г при 13—15°С-10-11, а при 8-9°С период этот затягивается до 40 дней.
К концу периода роста ихтиофтириус по сравнению с бродяжками достигает очень большой величины: 0, 5—1 мм в диаметре. По достижении предельной величины инфузории активными движениями выходят из тканей рыбы в воду и некоторое время медленно плавают при помощи покрывающего все их тело ресничного аппарата. Вскоре крупные ихтиофтириусы оседают на каком-нибудь подводном предмете и выделяют цисту. Тотчас же вслед за инцистированием начинаются последовательные деления инфузории: сначала пополам, затем каждая дочерняя особь делится опять на две и т. д. до 10—11 раз. В результате внутри цисты образуется до 2000 мелких почти округлых особей, покрытых ресничками. Внутри цисты бродяжки активно двигаются. Они прободают оболочку и выходят наружу. Активно плавающие бродяжки заражают новых особей рыб.
Быстрота деления ихтиофтириуса в цистах, так же как и темп его роста в тканях рыб, в большой степени зависит от температуры.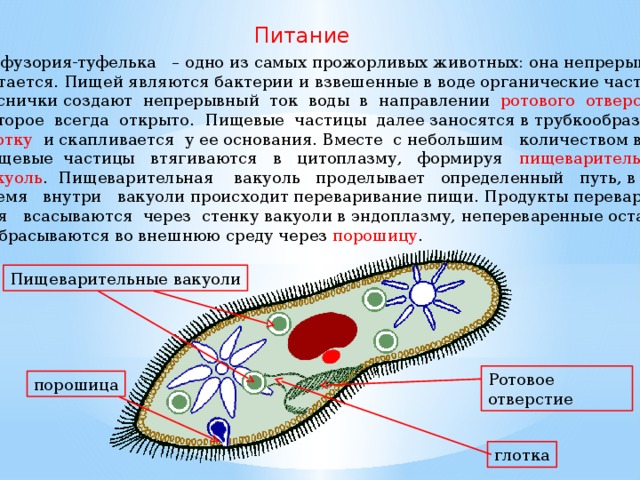 По исследованиям различных авторов, приводятся следующие цифры: при 26—27°С процесс развития бродяжек в цисте занимает 10—12 часов, при 15—16°С требуется 28—30 часов, при 4—5°С он тянется уже в течение 6—7 суток.
По исследованиям различных авторов, приводятся следующие цифры: при 26—27°С процесс развития бродяжек в цисте занимает 10—12 часов, при 15—16°С требуется 28—30 часов, при 4—5°С он тянется уже в течение 6—7 суток.
Ихтиофтириус — широко распространенный и очень опасный паразит рыб. Особенно большой вред он приносит в рыбоводческих хозяйствах, где происходит нерест и выращивание мальков. Ихтиофтириус, нападая на мальков, в короткий срок приводит их к гибели. Известны многочисленные случаи поголовного уничтожения мальков как карповых, так и лососевых рыб в прудовых хозяйствах.
Борьба с ихтиофтириусом представляет значительные трудности. Главное значение здесь имеют профилактические мероприятия, направленные на то, чтобы не допустить свободно плавающих в воде бродяжек проникнуть в ткани рыбы. Для этого полезно осуществлять частую пересадку больных рыб в новые водоемы или аквариумы, создавать условия проточности, что особенно эффективно в борьбе с ихтиофтириусом.
ИНФУЗОРИИ ТРИХОДИНЫ
Очень своеобразны паразитирующие главным образом на рыбах инфузории семейства урцелариид (Urcelariidae) отряда кругоресничных.
Особенно характерны многочисленные представители рода триходин (Trichodina) и некоторых других близких к нему родов (рис. 109). За немногими исключениями, все триходины и близкие к ним формы являются эктопаразитами, т. е. они живут на поверхности (а не в толще тканей и не внутри органов) своих хозяев (рыб). Место локализации триходин — кожные покровы, плавники, очень часто жабры рыб. Они встречаются как на морских, так и на пресноводных рыбах. Количество видов триходин измеряется несколькими десятками. Триходины не являются прикрепленными животными, а, напротив, активно передвигаются по поверхности своих хозяев. Вне организма хозяина, в воде, они долго жить не могут и, будучи отделены от рыбы, довольно быстро погибают.
Вся система приспособлений триходин к жизни на поверхности хозяина направлена на то, чтобы не оторваться от тела хозяина (что почти всегда равносильно гибели), сохраняя при этом подвижность. Приспособления эти весьма совершенны.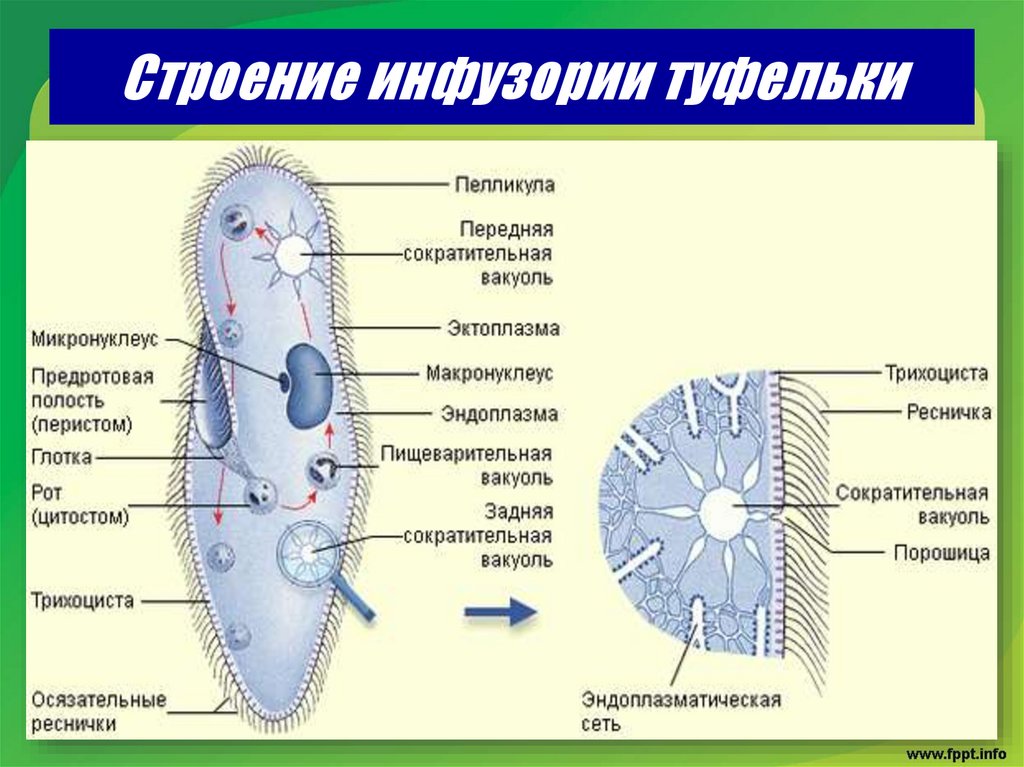 Тело большинства триходин имеет форму довольно плоского диска, иногда шапочки. Сторона, обращенная к телу хозяина, слегка вогнутая, она образует прикрепительную присоску. По наружному краю присоски расположен венчик хорошо развитых ресничек, при помощи которых главным образом и происходит передвижение (ползание) инфузории по поверхности тела рыбы. Этот венчик соответствует венчику, имеющемуся у бродяжек сидячих кругоресничных инфузории, рассмотренных выше. Таким образом, триходину можно сравнить с бродяжкой. На брюшной поверхности (на присоске) у триходин имеется очень сложный опорный и прикрепительный аппарат, способствующий удержанию инфузории на хозяине. Не вдаваясь в детали его строения, отметим, что основу его составляет сложной конфигурации кольцо, слагающееся из отдельных сегментов, несущих наружный и внутренний зубцы (рис. 109, Б). Это кольцо образует эластичную и вместе с тем прочную основу брюшной поверхности, выполняющей функцию присоски. Разные виды триходин отличаются друг от друга по количеству сегментов, образующих кольцо, и по конфигурации наружного и внутреннего крючьев.
Тело большинства триходин имеет форму довольно плоского диска, иногда шапочки. Сторона, обращенная к телу хозяина, слегка вогнутая, она образует прикрепительную присоску. По наружному краю присоски расположен венчик хорошо развитых ресничек, при помощи которых главным образом и происходит передвижение (ползание) инфузории по поверхности тела рыбы. Этот венчик соответствует венчику, имеющемуся у бродяжек сидячих кругоресничных инфузории, рассмотренных выше. Таким образом, триходину можно сравнить с бродяжкой. На брюшной поверхности (на присоске) у триходин имеется очень сложный опорный и прикрепительный аппарат, способствующий удержанию инфузории на хозяине. Не вдаваясь в детали его строения, отметим, что основу его составляет сложной конфигурации кольцо, слагающееся из отдельных сегментов, несущих наружный и внутренний зубцы (рис. 109, Б). Это кольцо образует эластичную и вместе с тем прочную основу брюшной поверхности, выполняющей функцию присоски. Разные виды триходин отличаются друг от друга по количеству сегментов, образующих кольцо, и по конфигурации наружного и внутреннего крючьев.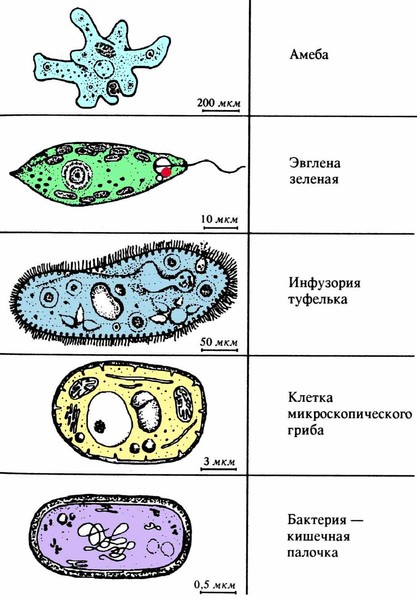
На противоположной от диска стороне тела триходины расположен перистом и ротовой аппарат. Строение его более или менее типично для кругоресничных инфузорий. Закрученные по часовой стрелке адоральные мембраны ведут в углубление, на дне которого расположен рот. Ядерный аппарат триходин устроен типично для инфузорий: один лентовидный макронуклеус и один расположенный рядом с ним микронуклеус. Имеется одна сократительная вакуоля.
Триходины широко распространены в водоемах всех типов. Особенно часто их находят на мальках разных видов рыб. При массовом размножении триходины наносят большой вред рыбе, в особенности если массами покрывают жабры. Это нарушает нормальное дыхание рыбы.
Для того чтобы очистить рыбу от триходин, рекомендуют делать лечебные ванны из 2-процентного раствора поваренной соли или 0, 01-процентного раствора марганцовокислого калия (для мальков — в течение 10—20 минут).
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА КОПЫТНЫХ
Среди инфузорий имеется множество видов, ведущих эндопаразитический образ жизни, т. е. живущих внутри (в различных органах и тканях) животных-хозяев. Особенно многочисленны и разнообразны инфузории, населяющие кишечный тракт копытных животных. Большинство их образует самостоятельный отряд инфузорий, называемый энтодиниоморфами (Еntodiniomorpha). У жвачных (рогатый скот, овцы, козы, антилопы, олени, лоси) эти инфузории в огромных количествах населяют передние отделы желудка. Желудок жвачных состоит из чэтырех отделов — рубца, сетки, книжки и сычуга. Из них только сычуг соответствует желудку других млекопитающих: он имеет пищеварительные железы, в него выделяются пищеварительные ферменты и соляная кислота. Реакция среды в сычуге резкокислая. Первые три отдела не имеют пищеварительных желез. Их внутренняя стенка частично ороговевает. Эти три отдела по происхождению следует рассматривать как нижние расширенные части пищевода. Названия сетки и книжки даны по форме и характеру складок их внутренней стенки. Самый вместительный отдел — рубец. Заглоченная и почти не пережеванная растительная пища сначала проходит в рубец.
е. живущих внутри (в различных органах и тканях) животных-хозяев. Особенно многочисленны и разнообразны инфузории, населяющие кишечный тракт копытных животных. Большинство их образует самостоятельный отряд инфузорий, называемый энтодиниоморфами (Еntodiniomorpha). У жвачных (рогатый скот, овцы, козы, антилопы, олени, лоси) эти инфузории в огромных количествах населяют передние отделы желудка. Желудок жвачных состоит из чэтырех отделов — рубца, сетки, книжки и сычуга. Из них только сычуг соответствует желудку других млекопитающих: он имеет пищеварительные железы, в него выделяются пищеварительные ферменты и соляная кислота. Реакция среды в сычуге резкокислая. Первые три отдела не имеют пищеварительных желез. Их внутренняя стенка частично ороговевает. Эти три отдела по происхождению следует рассматривать как нижние расширенные части пищевода. Названия сетки и книжки даны по форме и характеру складок их внутренней стенки. Самый вместительный отдел — рубец. Заглоченная и почти не пережеванная растительная пища сначала проходит в рубец. Сюда же попадает большое количество слюны. В рубце пища подвергается сложным процессам брожения, которые увеличивают ее усвояемость. В этом отделе в огромных количествах живут разнообразные микроорганизмы, вызывающие процессы брожения, которые протекают в условиях нейтральной или слабощелочной реакции. В рубце наряду с бактериями обитает масса простейших — очень мелких жгутиконосцев и инфузорий, так как здесь создаются весьма благоприятные условия для их развития.
Сюда же попадает большое количество слюны. В рубце пища подвергается сложным процессам брожения, которые увеличивают ее усвояемость. В этом отделе в огромных количествах живут разнообразные микроорганизмы, вызывающие процессы брожения, которые протекают в условиях нейтральной или слабощелочной реакции. В рубце наряду с бактериями обитает масса простейших — очень мелких жгутиконосцев и инфузорий, так как здесь создаются весьма благоприятные условия для их развития.
Из рубца через сетку пища отрыгивается в ротовую полость, где дополнительно пережевывается (жвачка). Проглатываемая вновь пищевая пережеванная масса по особой трубке, образуемой складками пищевода, идет уже не в рубец, а в книжку и оттуда в сычуг, где подвергается действию пищеварительных соков жвачного. В сычуге в условиях кислой реакции и наличия пищеварительных ферментов инфузории погибают. Попадая туда со жвачкой, они перевариваются.
Количество простейших в рубце (а также в сетке) может достигать колоссальных величин. Если взять каплю содержимого рубца и рассмотреть ее под микроскопом (при нагревании, так как при комнатной температуре инфузории останавливаются), то в поле зрения инфузории буквально кишат. Трудно даже в условиях культуры получить такую массу инфузорий. Количество инфузорий в 1 см3 содержимого рубца достигает миллиона, а нередко и более. В пересчете на весь объем рубца это дает поистине астрономические цифры! Богатство содержимого рубца инфузориями в большой степени зависит от характера пищи жвачного. Если пища богата клетчаткой и бедна углеводами и белками (трава, солома), то инфузорий в рубце относительно немного. При добавлении в пищевой рацион углеводов и белков (отруби) количество инфузорий резко увеличивается и достигает огромных цифр. Нужно иметь в виду, что существует постоянный отток инфузорий. Попадая вместе с жвачкой в сычуг, они погибают. Высокий уровень количества инфузорий поддерживается энергичным размножением их.
Если взять каплю содержимого рубца и рассмотреть ее под микроскопом (при нагревании, так как при комнатной температуре инфузории останавливаются), то в поле зрения инфузории буквально кишат. Трудно даже в условиях культуры получить такую массу инфузорий. Количество инфузорий в 1 см3 содержимого рубца достигает миллиона, а нередко и более. В пересчете на весь объем рубца это дает поистине астрономические цифры! Богатство содержимого рубца инфузориями в большой степени зависит от характера пищи жвачного. Если пища богата клетчаткой и бедна углеводами и белками (трава, солома), то инфузорий в рубце относительно немного. При добавлении в пищевой рацион углеводов и белков (отруби) количество инфузорий резко увеличивается и достигает огромных цифр. Нужно иметь в виду, что существует постоянный отток инфузорий. Попадая вместе с жвачкой в сычуг, они погибают. Высокий уровень количества инфузорий поддерживается энергичным размножением их.
У непарнокопытных (лошадь, осел, зебра) в пищевом тракте тоже имеется большое количество инфузорий, однако локализация их в хозяине иная. У непарнокопытных нет сложного желудка, благодаря чему возможность развития простейших в передних отделах пищевого тракта отсутствует. Зато у непарнокопытных очень сильно развиты толстая и слепая кишка, которые обычно забиты пищевыми массами и играют существенную роль в пищеварении. В этом отделе кишечника, подобно тому как в рубце и сетке жвачных, развивается очень богатая фауна простейших, преимущественно инфузорий, большинство которых также относится к отряду энтодиниоморф. Однако по видовому составу фауна рубца жвачных и фауна толстого кишечника непарнокопытных не совпадают.
У непарнокопытных нет сложного желудка, благодаря чему возможность развития простейших в передних отделах пищевого тракта отсутствует. Зато у непарнокопытных очень сильно развиты толстая и слепая кишка, которые обычно забиты пищевыми массами и играют существенную роль в пищеварении. В этом отделе кишечника, подобно тому как в рубце и сетке жвачных, развивается очень богатая фауна простейших, преимущественно инфузорий, большинство которых также относится к отряду энтодиниоморф. Однако по видовому составу фауна рубца жвачных и фауна толстого кишечника непарнокопытных не совпадают.
ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА ЖВАЧНЫХ
Наибольший интерес представляют инфузории семейства офриосколецид (Ophryoscolecidae), относящегося к отряду энтодиниоморф. Характерный признак этого отряда — отсутствие сплошного ресничного покрова. Сложные ресничные образования — цирры — расположены на переднем конце тела инфузорий в области ротового отверстия. К этим основным элементам ресничного аппарата могут прибавиться еще дополнительные группы цирр, расположенные либо на переднем, либо на заднем конце тела. Общее количество видов инфузорий семейства офриосколецид — около 120.
К этим основным элементам ресничного аппарата могут прибавиться еще дополнительные группы цирр, расположенные либо на переднем, либо на заднем конце тела. Общее количество видов инфузорий семейства офриосколецид — около 120.
На рисунке 110 изображены некоторые наиболее типичные представители офриосколецид из рубца жвачных. Наиболее просто устроены инфузории рода энтодиний (Entodinium, рис. 110, Л). На переднем конце тела их имеется одна околоротовая зона цирр. Передний конец тела, на котором расположено ротовое отверстие, может втягиваться внутрь. Резко разграничены эктоплазма и эндоплазма. Хорошо видна на заднем конце анальная трубка, служащая для выведения наружу непереваренных остатков пищи. Несколько сложнее строение аноплодиния (Anoplodinium, рис. 110, Б). У них имеются две зоны ресничного аппарата — околоротовые цирры и спинные цирры. Обе они расположены на переднем конце. На заднем конце тела изображенного на рисунке вида имеются длинные острые выросты — это довольно типично для многих видов офриосколецид. Высказывалось предположение, что эти выросты способствуют «проталкиванию» инфузорий между растительными частицами, заполняющими рубец.
Высказывалось предположение, что эти выросты способствуют «проталкиванию» инфузорий между растительными частицами, заполняющими рубец.
Виды рода эудиплодиний (Eudiplodinium, рис. 110, Б) сходны с аноплодинием, но, в отлпчие от них, имеют скелетную опорную пластинку, расположенную но правому краю вдоль глотки. Эта скелетная пластинка состоит из вещества, близкого по химической природе к клетчатке, т. е. к веществу, из которого состоят оболочки растительных клеток.
У рода полипластрон (Polyplastron, рис. 110, Г, Д) наблюдается дальнейшее усложнение скелета. Строение этих инфузорий близко к эудиплодиниям. Отличия сводятся прежде всего к тому, что вместо одной скелетной пластинки эти инфузории имеют пять. Две из них, наиболее крупные, расположены по правой стороне, а три, более мелкие, — по левой стороне инфузории. Вторая особенность полипластрона — увеличение количества сократительных вакуолей. У энтодиниев одна сократительная вакуоля, у аноплодиниев и эудиплодиниев две сократительные вакуоли, у полипластрона их около десятка.
У эпидиниев (Epidinium, рис. 110), обладающих хорошо развитым углеводным скелетом, расположенным по правой стороне тела, спинная зона цирр смещается с переднего конца на спинную сторону. На заднем конце инфузорий этого рода часто развиваются шипы.
Наиболее сложное строение обнаруживает род офриосколекс (Ophryoscolex), по которому названо и все семейство инфузорий (рис. 110, Е). У них хорошо развита спинная зона цирр, охватывающая около 2/3 окружности тела и скелетные пластинки. На заднем конце образуются многочисленные шипы, из которых один бывает обычно особенно длинным.
Знакомство с некоторыми типичными представителями офриосколецид показывает, что в пределах этого семейства произошло значительное усложнение организации (от энтодиниев до офриосколекса).
Очень часто, как мы это увидим, например, при знакомстве с червями, паразитизм, особенно эндопаразитизм, приводит к значительному упрощению организации, к регрессивным изменениям многих систем органов. У инфузорий, живущих в кишечнике копытных, этого не произошло. Напротив, здесь имело место прогрессивное развитие, не упрощение, а усложнение организации. Это связано, вероятно, с особенностями среды обитания инфузорий рубца. Они живут как бы в своеобразном водоеме с очень сложными условиями (наличие многочисленных механических препятствий, сложный бактериальный состав среды и многое другое), приспособление к которым было связано с усложнением строения инфузорий.
Кроме инфузорий семейства офриосколецид, в рубце жвачных встречаются в небольших количествах представители уже известного нам отряда равноресничных инфузорий. Они представлены небольшим числом видов. Их тело равномерно покрыто продольными рядами ресничек, скелетные элементы отсутствуют. В общей массе инфузорного населения рубца они не играют заметной роли, и поэтому мы не будем здесь останавливаться на их рассмотрении.
Чем и как питаются инфузории офриосколециды? Этот вопрос подробно изучался многими учеными, особенно детально профессором В. А. Догелем.
Пища офриосколецид довольно разнообразна, и у разных видов их наблюдается известная специализация. Самые мелкие виды рода энтодиниев питаются бактериями, зернами крахмала, грибками и другими небольшими частицами. Очень многие средние и крупные офриосколециды поглощают частицы растительных тканей, которые составляют главную массу содержимого рубца. Эндоплазма некоторых видов бывает буквально забита растительными частицами. Можно видеть, как инфузории набрасываются на обрывки растительных тканей, буквально разрывают их на кусочки и затем заглатывают, нередко закручивая их в своем теле спиралью (рис. 111, 4). Иногда приходится наблюдать такие картины (рис. 111, 2), когда само тело инфузории оказывается деформированным благодаря заглоченным крупным частицам.
У офриосколецид наблюдается иногда хищничество. Более крупные виды пожирают более мелкие. Хищничество (рис. 112) сочетается со способностью тех же видов питаться растительными частицами.
Каким путем инфузории проникают в рубец жвачного? Каковы пути заражения офриосколецидами? Оказывается, что новорожденные жвачные не имеют еще в рубце инфузорий. Они отсутствуют также, пока животное питается молоком. Но как только жвачное переходит на растительную пищу, сразу же в рубце и сетке появляются инфузории, количество которых быстро нарастает. Откуда они берутся? Долгое время предполагали, что инфузории рубца образуют какие-то покоящиеся стадии (вероятнее всего, цисты), которые широко рассеиваются в природе и, будучи проглоченными, дают начало активным стадиям инфузорий. Дальнейшие исследования показали, что у инфузорий рубца жвачных никаких покоящихся стадий нет. Удалось доказать, что заражение происходит активными подвижными инфузориями, которые проникают в ротовую полость при отрыгивании жвачки. Если исследовать под микроскопом взятую из ротовой полости жвачку, го в ней всегда имеется большое количество активно плавающих инфузорий. Эти активные формы легко могут проникнуть в рот и далее в рубец других особей жвачного из общего сосуда для пойла, вместе с травой, сеном (на которых может оказаться слюна с инфузориями) и т. п. Этот путь заражения был доказан экспериментально.
Если покоящиеся стадии у офриосколецид отсутствуют, то, очевидно, легко получить «безинфузорных» животных, изолировав их, когда они еще питаются молоком. Если не допускать прямого контакта между растущим молодняком и имеющими инфузорий жвачными, то молодые животные могут остаться без инфузорий в рубце. Такие опыты были проведены несколькими учеными в разных странах. Результат был однозначен. При отсутствии контакта между молодняком (отнятым от матери еще в период кормления молоком) и имеющими в рубце инфузорий жвачными, животные вырастают стерильными в отношении инфузорий. Однако достаточно даже кратковременного контакта с животными, имеющими инфузорий (общая кормушка, общее ведро для питья, общее пастбище), чтобы в рубце стерильных животных появилась фауна инфузорий.
Массовое развитие фауны простейших в рубце и сетке, естественно, ставит вопрос о характере взаимоотношений между простейшими и их хозяевами — жвачными. Мы уже видели на примере жгутиконосцев, паразитирующих в кишечнике термитов, какие тесные симбиотические взаимоотношения могут возникать между простейшими, населяющими кишечный тракт, и их хозяевами. Не существует ли между инфузориями, населяющими рубец и сетку, аналогичных симбиотических взаимоотношений? Этот вопрос ставился многими исследователями. Однако до сих пор окончательный ответ на него не получен, и требуются дальнейшие исследования. Ввиду большого биологического и практического интереса этого вопроса рассмотрим его несколько подробнее.
Выше были приведены результаты опытов по содержанию жвачных, совершенно лишенных инфузорий в рубце и сетке. Это достигается, как мы видели, путем ранней изоляции молодняка. Опыты проводились на овцах и на козах.
Таким путем удавалось проводить наблюдения за «безинфузорными» животными в течение значительного промежутка времени (свыше года). Как отражается отсутствие инфузорий в рубце на жизни хозяина? Отрицательно или положительно влияет отсутствие инфузорий на хозяина? Для ответа на этот вопрос на козах провели следующие опыты. Были взяты козлята-близнецы (одного помета и одного пола), чтобы иметь более сходный материал. Затем один из близнецов данной пары воспитывался без инфузорий в рубце (ранняя изоляция), другой же с самого начала питания растительной пищей обильно заражался множеством видов инфузорий. Оба получали совершенно одинаковый рацион и воспитывались в одинаковых условиях. Единственное различие между ними сводилось к наличию или отсутствию инфузорий. На нескольких изученных таким образом парах козлят не было обнаружено каких-либо различий в ходе развития обоих членов каждой пары («инфузорного» и «безинфузорного»). Таким образом, можно утверждать, что сколько-нибудь резкого влияния на жизненные функции животного-хозяина инфузории, живущие в рубце и сетке, не оказывают.
Приведенные выше результаты опытов не позволяют, однако, утверждать, что инфузории рубца совершенно безразличны для хозяина. Эти опыты проводились при нормальном пищевом рационе хозяина. Не исключена возможность, что при других условиях, при ином пищевом режиме (например, при недостаточном кормлении) удастся выявить влияние на хозяина фауны простейших, населяющих рубец.
В литературе высказывались различные предположения о возможном положительном влиянии протозойной фауны рубца на пищеварительные процессы хозяина. Указывалось, что многие миллионы инфузорий, активно плавающие в рубце и размельчающие растительные ткани, способствуют брожению и перевариванию пищевых масс, находящихся в передних отделах пищеварительного тракта. Значительное количество инфузорий, попадающее вместе с жвачкой в сычуг, переваривается, и белок, составляющий значительную часть тела инфузорий, усваивается. Инфузории, таким образом, могут быть дополнительным источником белка для хозяина. Высказывались также предположения, что инфузории способствуют перевариванию клетчатки, составляющей основную массу пищи жвачных, переводу ее в более усвояемое состояние.
Все эти предположения не являются доказанными, и против некоторых из них выдвигались возражения. Указывалось, например, что инфузории строят протоплазму своего тела из белков, которые поступают в рубец с пищей хозяина. Поглощая белок растительный, онп переводят его в животный белок своего тела, который затем переваривается в сычуге. Дает ли это какие-нибудь преимущества хозяину, остается неясно. Все эти вопросы имеют большой практический интерес, так как речь идет о пищеварении жвачных — основных объектов животноводства. Дальнейшие исследования о роли инфузорий рубца в пищеварении жвачных очень желательны.
Насколько строго приурочены определенные виды офриосколецид к определенным видам хозяев? Имеются ли, например, в рогатом скоте свои виды инфузорий, в овцах — свои и т. д.? Приспособленность паразита к определенному кругу хозяев называют его специфичностью. Если паразит строго приурочен к одному виду хозяев, это говорит об узкой специфичности. Если данный вид паразита может жить в разных видах хозяев, то это проявление широкой специфичности.
Офриосколециды жвачных обладают, как правило, широкой специфичностью. В видовом отношении население рубца и сетки рогатого скота, овец, коз очень близко между собой. Если сравнить видовой состав рубца африканских антилоп с рогатым скотом, то и здесь около 40% от общего числа видов окажется общим. Однако существует немало видов офриосколецид, которые встречаются только в антилопах или только в оленях. Таким образом, на фоне общей широкой специфичности офриосколецид можно говорить об отдельных более узко специфичных видах их.
ИНФУЗОРИИ КИШЕЧНИКА НЕПАРНОКОПЫТНЫХ
Обратимся теперь к краткому знакомству с инфузориями, населяющими толстую и слепую кишку непарнокопытных.
В видовом отношении эта фауна, как и фауна рубца жвачных, тоже весьма разнообразна. В настоящее время описано около 100 видов инфузорий, обитающих в толстом кишечнике животных семейства лошадиных. Встречающиеся здесь инфузории в смысле их принадлежности к разным систематическим группам более разнообразны, чем инфузории рубца жвачных.
В кишечнике лошадиных обитает довольно много видов инфузорий, относящихся к отряду равноресничных, т. е. инфузорий, у которых ресничный аппарат не образует около ротовой зоны мембранелл или цирр (рис. 113, 1).
Отряд энтодиниоморф (Entodiniomorpha) тоже богато представлен в кишечнике лошади. В то время как в рубце жвачных встречается лишь одно семейство энтодиниоморф (семейство офриосколецид), в кишечнике лошади живут представители трех семейств, на характеристике которых мы, однако, останавливаться здесь не будем, ограничившись лишь несколькими рисунками типичных для лошади видов (рис. 113).
Детальными исследованиями А.Стрелкова было показано, что разные виды инфузорий далеко не равномерно распределяются по ходу толстого кишечника лошади. Имеются две разные группы видов, как бы две фауны. Одна из них населяет слепую кишку и брюшной отдел большой ободочной кишки (начальные отделы толстого кишечника), а другая—спинной отдел большой ободочной и малую ободочную. Эти два комплекса видов довольно резко разграничены. Видов, общих для этих двух разделов, немного — менее десятка.
,
Интересно отметить, что среди многочисленных видов инфузорий, населяющих толстый кишечник непарнокопытных, имеются представители одного рода, относящегося к сосущим инфузориям. Как мы видели выше, сосущие инфузории (Suctoria) — это типичные свободноживущие сидячие организмы с совершенно особым способом питания при помощи щупалец (рис. 103). Один из родов сукторий приспособился к такой, казалось бы, необычной среде обитания, как толстый кишечник лошади, например несколько видов аллантозом (Allantosoma). Эти очень своеобразные животные (рис. 114) не имеют стебелька, реснички отсутствуют, булавовидные, утолщенные на концах щупальца хорошо развиты.
При помощи щупалец аллантозомы присасываются к разным видам инфузорий и высасывают их. Нередко при этом добыча во много раз превосходит хищника.
Неясен еще вопрос о характере взаимоотношений инфузорий толстого кишечника непарнокопытных с хозяевами. Количество инфузорий может быть столь же велико, а иногда даже больше, чем в рубце жвачных. Имеются данные, показывающие, что количество инфузории в толстых кишках лошади может достигать 3 млн. в 1 см3. Симбиотическое значение, предполагавшееся некоторыми учеными, еще менее вероятно, чем для инфузорий рубца.
Наиболее вероятно мнение, что они наносят некоторый вред хозяину, поглощая значительное количество пищи. Часть инфузорий выносится наружу с фекальными массами, и таким образом органические вещества (в том числе и белок), входящие в состав их тела, остаются не использованными хозяином.
Вопрос о путях заражения непарнокопытных инфузориями, населяющими толстый кишечник, до сих пор не разрешен.
БАЛАНТИДИЙ-ПАРАЗИТ КИШЕЧНИКА ЧЕЛОВЕКА
Один вид паразитических инфузорий из отряда равноресничных — балантидий коли (Balantidium coli) — паразитирует в толстом кишечнике человека. Случаи заражения человека балантидиями очень редки. В этом отношении балантидий несравним с кишечными амебами, которые, как мы видели, встречаются часто. Паразитирование балантидия в кишечнике человека обычно вызывает тяжело протекающий колит. Поэтому паразит этот требует к себе пристального внимания, несмотря на то что встречается он относительно редко.
Тело балантидия, паразитирующего в кишечнике человека, имеет яйцевидную форму (рис. 115). Размеры варьируют в очень широких пределах: длина 30—150, ширина 20—110 мк. Эти вариации величины в значительной степени зависят от того, в какой мере инфузория набита пищей. Все тело густо покрыто короткими ресничками, расположенными продольными рядами от переднего конца к заднему. На переднем конце имеется углубление в виде неглубокой щели. Это околоротовое углубление (перистом), на дне которого помещается ротовое отверстие. По краю перистома расположены особые сильно развитые реснички, направляющие пищу в глубину его. Имеются две сократительные вакуоли (одна в передней трети тела, вторая на заднем конце). Ядерный аппарат типичный для инфузорий: бобовидный, расположенный примерно в центре тела макронуклеус и в непосредственном соседстве с ним один маленький микронуклеус.
Балантидий захватывает разнообразные пищевые частицы из содержимого толстой кишки. Особенно охотно он питается крахмальными зернами. Если балантидий живет в просвете толстой кишки человека, то он питается содержимым кишечника и не оказывает никакого вредного влияния. Это типичное «носительство», с которым мы познакомились уже при рассмотрении дизентерийной амебы. Однако балантидий реже, чем дизентерийная амеба, остается таким «безобидным квартирантом».
Он активно внедряется в слизистую кишечника и проникает в толщу его стенки. Здесь балантидий переключается на питание клетками хозяина и особенно «охотно» заглатывает красные кровяные тельца (эритроциты). В результате нарушения целостности внутренней стенки кишечник изъязвляется, и это сопровождается тяжелой формой кровавого поноса (колита). Поскольку паразит внедряется в самую толщу тканей кишечника, лечение балантидиоза очень затруднительно.
В настоящее время в медицине известны некоторые специфические лекарства, убивающие балантидия. Однако успех лечения не всегда бывает положительным именно в силу внутритканевой локализации паразита.
Может ли человеческий балантидий паразитировать в других хозяевах, кроме человека? Оказывается, что эта паразитическая инфузория часто встречается в кишечнике свиней. Но, в отличие от человеческого, этот балантидий обычно не вызывает у свиней болезненных явлений и живет в кишечнике их как «безобидный квартирант». Основным хозяином балантидия следует считать свинью, тогда как человек заражается редко, причем почти всегда источником заражения являются свиньи.
Каким же путем осуществляется заражение балантидием? У этой инфузории имеются покоящиеся стадии — цисты. Инцистирование происходит при проникновении паразита в заднюю кишку. Цисты выводятся наружу с фекальными массами. Они долгое время (свыше двух месяцев при комнатной температуре) сохраняют жизнеспособность. Если циста будет случайно проглочена свиньей или человеком, то она проходит в неизменном виде через желудок и тонкие кишки. Когда циста достигнет толстой кишки, оболочка ее разрушается и из нее выходит активная стадия балантидия.
Из сказанного выше следует, что персонал, обслуживающий свиней, должен соблюдать элементарные гигиенические правила, а главное не приступать к еде, не вымыв предварительно руки. Помимо кишечника человека и свиней, балантидий коли может паразитировать (при непосредственном заражении) в кишечнике крыс и человекообразных обезьян. У обезьян, так же как и у человека, балантидий вызывают тяжелую форму колита.
В настоящее время специалистами хорошо разработаны различные методы, позволяющие культивировать балантидиев в искусственной среде — вне организма хозяина.
ИНФУЗОРИИ-ПАРАЗИТЫ ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯН
Мы видели, что человеческий балантидий может паразитировать в обезьянах, вызывая колиты. Однако в человекообразных обезьянах, кроме того, встречаются очень интересные и сложно устроенные инфузории, которые в организме человека не паразитируют. Они относятся к отряду энтодиниоморф (Entodiniomorpha), т. е. к тому же отряду, который, как мы видели, богато представлен в кишечном тракте копытных млекопитающих.
В человекообразных обезьянах (горилле и шимпанзе) встречаются виды рода троглодителла (Troglodytella), паразитирующие в толстой и слепой кишке (рис. 116). В природной обстановке в Африке как гориллы, так и шимпанзе заражены, по-видимому, поголовно. В условиях неволи, в зоологических садах, инфузории постепенно исчезают из организма обезьян.
Как видно из рисунка, троглодптелла относится к числу сложно устроенных энтодиниоморф. У нее, кроме околоротовой зоны цирр (на переднем конце тела), имеются еще три зоны хорошо развитых цирр, кольцеобразно охватывающих тело инфузории. Троглодителлы обладают хорошо развитым, охватывающим почти весь передний конец тела скелетным аппаратом, состоящим из углеводов. Размеры этих своеобразных инфузорий довольно значительны. В длину они достигают 200—280 мк.
БЕЗРОТЫЕ ИНФУЗОРИИ АСТОМАТЫ
На предыдущих страницах было рассказано о различных паразитических инфузориях, хозяевами которых являются позвоночные животные. Наряду с этим большое количество видов паразитических инфузорий живет в самых различных беспозвоночных животных. Рассмотрим кратко наиболее интересных представителей их.
Глубоко приспособленной к паразитизму и выработавшей ряд интересных приспособлений является обширная группа безротых инфузорий — астомат (Astomata). Самая характерная особенность этого отряда отражена в его названии — инфузории астоматы лишены ротового отверстия. Их питание осуществляется осмотическим путем — всасыванием питательных веществ всей поверхностью тела. Поскольку оформленной пищи астоматы не поглощают, пищеварительные вакуоли у них не образуются.
Общее число видов астомат превышает сотню. Встречаются они в беспозвоночных, относящихся к самым различным группам (типам и классам) животного мира: в ресничных червях, кольчатых червях, моллюсках, иглокожих. Небольшое число видов астомат паразитирует и в позвоночных животных, а именно в земноводных. Однако астоматы распределены по разным группам хозяев совсем неравномерно. Существует одна группа «излюбленных» хозяев астомат, в которой сосредоточено свыше 75% их видов. Такой группой являются малощетинковые кольчатые черви (Oligochaeta), класс животных, к которому относится всем известный дождевой червь. Однако лишь часть видов малощетпнковых кольчецов живет в почве. Большая часть их — обитатели пресных вод, где они образуют заметную (а иногда и преобладающую) часть донного населения. Эти пресноводные малощетинковые кольчецы и являются «излюбленными» хозяевами астомат. Небольшое число видов олигохет встречается и в море, почти исключительно в прибрежной, литоральной (осушаемой во время отлива) зоне, но в морской фауне они не играют сколько-нибудь заметной роли.
Органом, в котором локализуются в олигохетах инфузории астоматы, является кишечник. Поэтому многие черты строения астомат связаны с приспособлением к жизни в кишечнике. Через кишечник постоянно осуществляется движение пищи, иногда довольно активное. Паразиты кишечника, чтобы не быть вынесенными наружу (что равнозначно гибели), должны обладать приспособлениями, направленными на то, чтобы удержаться в кишечнике. Эти приспособления выражены не только у инфузорий, но и у всех других групп кишечных паразитов, к какой бы систематической группе они ни относились и в каких бы хозяевах ни паразитировали. Мы увидим это в дальнейшем, при знакомстве с разными группами паразитических червей.
Тело всех паразитирующих в кишечнике олигохет астомат вытянуто в длину, что способствует их движению в просвете кишечника. Ресничный покров всегда хорошо развит, реснички покрывают все тело инфузорий и расположены тесными продольными рядами.
В простейшем случае, например у представителей рода аноплофрий (Anoplophrуа, рис. 117, Б), никаких особых прикрепительных и опорных образований не развивается. Но у большинства видов других родов имеются разнообразные и подчас довольно сложные структуры двух категорий — опорные и прикрепительные, являющиеся формами приспособления к кишечному паразитизму.
Опорные скелетные образования развиваются преимущественно на переднем конце тела, которому приходится испытывать механические воздействия и преодолевать препятствия, проталкиваясь в просвете кишечника между частицами пищи. У видов рода радиофрий (Radiophrya) на переднем конце с одной стороны тела (которую условно считают брюшной) располагаются очень прочные эластичные ребра (спикулы), лежащие в поверхностном слое эктоплазмы (рис. 117, Б, Г, Д). У видов рода менилелла (Mesnilella) тоже имеются опорные лучи (спикулы), которые на большей части своего протяжения лежат в более глубоких слоях цитоплазмы (в эндоплазме, рис. 117, А). Аналогично устроенные опорные образования развиты и у видов некоторых других родов астомат.
У многих представителей этой интересной группы инфузорий имеются специальные структуры, позволяющие паразиту прочно прикрепляться к стенке кишечника. У известной уже нам радиофрии на самом переднем конце торчит наружу острый зубец, подвижно сочленяющийся с особыми опорными образованиями, имеющими форму наконечника стрелы. При помощи этого зубца радиофрия может прочно прикрепляться (зацепляться) к стенке кишечника.
Другим типом прикрепительных образований астомат, но менее распространенным, являются присоски на переднем конце тела. Такого рода прикрепительный аппарат развит у видов рода гаптофрий (Haptophrya). Разные виды этого рода паразитируют в очень несхожих друг с другом хозяевах: одни — в ресничных червях (турбелляриях), другие — в земноводных.
При изучении прикрепительных аппаратов паразитирующих в кишечнике различных хозяев инфузорий астомат обращает на себя внимание их большое сходство с аналогичными органами у других групп паразитов. Например, у ленточных червей основными формами органов прикрепления являются присоски и крючья. У инфузорий астомат — одноклеточных организмов — возникают структуры, весьма напоминающие органы прикрепления сложных многоклеточных животных (ленточные черви). Возникновение сходных структур, выполняющих аналогичную функцию у животных, относящихся к совершенно различным группам животного мира, связано с приспособлением к сходным условиям существования, в данном случае с приспособлением паразитов к жизни в кишечнике хозяев. Это явление носит название конвергенции.
Бесполое размножение у некоторых инфузорий астомат протекает своеобразно. Вместо поперечного деления надвое, свойственного большинству инфузорий, у многих астомат происходит неравномерное деление (почкование). При этом почки, отделяющиеся на заднем конце, некоторое время остаются связанными с материнской особью (рис. 117, В). В результате получаются цепочки, состоящие из передней крупной и задних более мелких особей (почек). В дальнейшем почки постепенно отделяются от цепочки и переходят к самостоятельному существованию. Эта своеобразная форма размножения широко распространена, например, у известной уже нам радиофрий. Возникающие в результате почкования цепочки некоторых астомат напоминают по внешнему виду цепочки ленточных червей. Здесь мы вновь встречаемся с явлением конвергенции.
Ядерный аппарат астомат имеет характерную для инфузорий структуру: макронуклеус, чаще всего лентовидной формы (рис. 117), и один микронуклеус. Сократительные вакуоли обычно хорошо развиты. У большинства видов имеется несколько (иногда свыше десятка) сократительных вакуолей, расположенных в один продольный ряд.
Изучение распределения видов астомат по разным видам хозяев показывает, что большая часть астомат приурочена к строго определенным видам хозяев. Большинству астомат свойственна узкая специфичность: хозяином для них может служить лишь один вид животного.
Несмотря на большое количество исследований, посвященных изучению инфузорий астомат, одна очень важная сторона их биологии остается совершенно неясной: каким образом происходит передача этих инфузорий от одной особи хозяина к другой? Никогда еще не удавалось наблюдать образование цист у этих инфузорий.
Поэтому высказывают предположение, что заражение происходит активно — подвижными стадиями.
Инфузории астоматы представляют собой яркий пример глубокого приспособления инфузорий к паразитическому образу жизни. Это приспособление отразилось на различных сторонах их организации (скелет, органоиды прикрепления) и жизненных отправлений. Они утеряли характерный для инфузорий способ питания путем заглатывания оформленных частиц пищи и перешли к осмотическому питанию.
ИНФУЗОРИИ КИШЕЧНИКА МОРСКИХ ЕЖЕЙ
Разнообразный комплекс паразитических инфузорий обитает в кишечнике морских ежей. Морские ежи—богатый видами класс морских беспозвоночных животных, относящихся к типу иглокожих. Размеры морских ежей различны, но у большинства встречающихся в наших водах видов поперечник их почти шаровидного тела составляет 5—10 см.
Очень многочисленны морские ежи в прибрежной зоне наших северных (Баренцево) и дальневосточных морей (Японского моря, тихоокеанского побережья Курильских островов). Большинство морских ежей питается растительной пищей, главным образом водорослями, которые они соскабливают с подводных предметов особыми острыми «зубами», окружающими ротовое отверстие. В кишечнике этих растительноядных ежей имеется богатая фауна инфузорий. Нередко они развиваются здесь в массовом количестве, и содержимое кишечника морского ежа под микроскопом почти так же «кишит» инфузориями, как содержимое рубца жвачных. Нужно сказать, что, кроме глубоких различий в условиях жизни инфузорий кишечника морского ежа и рубца жвачного, имеются и некоторые черты сходства. Они заключаются в том, что и там и здесь инфузории живут в среде, очень богатой растительными остатками. В настоящее время известно свыше 50 видов инфузорий, обитающих в кишечнике морских ежей, которые встречаются только в прибрежной зоне, где ежи питаются водорослями. На больших глубинах, где водоросли уже не растут, инфузорий в морских ежах нет.
Что же представляет собой фауна паразитических инфузорий кишечника морских ежей в отношении своего систематического состава (принадлежности их к определенным систематическим группам)? Оказывается, что в кишечнике морских ежей встречаются представители разных отрядов и семейств инфузорий. На рисунке 118 изображены некоторые характерные представители этой фауны из кишечника морских ежей, обитающих в прибрежных водах Японского моря в окрестностях Владивостока. Здесь имеются различные равноресничные инфузории (Holotricha), брюхоресничные (Hypotrycha), малоресничные (Oligotricha). Интересно отметить, что некоторые из видов инфузорий из кишечника морских ежей относятся к родам, имеющим свободноживущие виды. У них нет тех глубоких приспособлений, которые можпо видеть у энтодиниоморф (Entodiniomorpha) к жизни в кишечнике копытных или у астомат (Astomata) к жизни в кишечнике малощетинковых кольчецов. Лишь у некоторых видов, например у энторипидиум (Entorhipidium), развиваются скелетные лучи (подобно тому как это мы видели у радиофрий, рис. 117). Другие же виды мало чем отличаются от свободноживущих инфузорий.
По образу жизни и характеру питания большинство инфузорий кишечника морских ежей растительноядны. Они питаются водорослями, которые в большом количестве наполняют кишечник хозяина. Некоторые виды довольно «привередливы» в выборе пищи. Например, стробилидий (Strobilidium, рис. 118, А) питается почти исключительно крупными диатомовыми водорослями. Имеются здесь и хищники, поедающие представителей других, более мелких видов инфузорий.
У инфузорий из кишечника морских ежей, в отличие от астомат, не наблюдается какой-либо строгой приуроченности к определенным видам хозяев. Они живут в самых разных видах морских ежей, питающихся водорослями.
Как уже говорилось, простейшие из кишечника морских ежей хотя и ведут эндо-паразитический образ жизни, но почти не отличаются от свободноживущих родичей. Вероятно, эти инфузории относительно недавно перешли к паразитизму. Этот переход осуществлялся независимо для разных видов. В новых условиях паразитической жизни они нашли среду, не очень отличающуюся от морской воды. В кишечнике морских ежей всегда много заглоченной вместе с пищей воды и много водорослей на разных стадиях переваривания. Эти условия оказались весьма благоприятными для жизни инфузорий, которые и размножаются здесь в больших количествах.
Пути заражения морских ежей инфузориями не изучены. Однако здесь с большой долей вероятности можно допустить, что оно происходит активными свободноплавающими формами. Дело в том, что инфузории из кишечника морских ежей могут длительное время (многие часы) жить в морской воде. Однако они уже настолько приспособились к жизни в кишечнике ежей, что вне тела их, в морской воде, рано или поздно погибают.
Заканчивая знакомство с инфузориями, следует еще раз подчеркнуть, что они представляют собой богатую видами, обширную и процветающую группу (класс) животного мира. Оставаясь на уровне клеточной организации, инфузории достигли по сравнению с другими классами простейших наибольшей сложности строения и функций.
Особенно существенную роль в этом прогрессивном развитии (эволюции) сыграло, вероятно, преобразование ядерного аппарата и возникновение ядерного дуализма (качественной неравноценности ядер). Богатство макронуклеуса нуклеиновыми веществами связано с активными процессами обмена веществ, с энергично протекающими процессами синтеза белков цитоплазмы и ядер.
Организация инфузорий оказалась очень пластичной, что позволило им приспособиться к различным средам обитания в морской и пресной воде и дать разнообразные паразитические формы.
Заключение
Мы подошли к концу нашего обзора строения и образа жизни обширного типа животного мира — простейших. Характерной особенностью их, как уже неоднократно подчеркивалось выше, является одноклеточность. В отношении своего строения простейшие — клетки. Однако они несравнимы с клетками, входящими в состав тела многоклеточных организмов, потому что сами являются организмами. Таким образом, простейшие — это организмы на клеточном уровне организации. Некоторые высокоорганизованные простейшие, обладающие многими ядрами, уже как бы выходят за морфологические пределы строения клетки, что дает основание некоторым ученым называть таких простейших «надклеточными». Это мало меняет существо дела, ибо типичной для Protozoa является все же одноклеточная организация.
В пределах одноклеточности простейшие прошли длинный путь эволюционного развития и дали огромное разнообразие форм, приспособленных к самым различным условиям жизни. В основе родословного ствола простейших стоят два класса: саркодовые и жгутиконосцы. До сих пор в науке дебатируется вопрос, какой из этих классов более примитивный. С одной стороны, низшие представители саркодовых (амебы) обладают самым примитивным строением. Но жгутиконосцы обнаруживают наибольшую пластичность типа обмена веществ и стоят как бы на границе между животным и растительным миром. В жизненном цикле некоторых саркодовых (например, фораминифер) имеются жгутиковые стадии (гаметы), что указывает на их связь со жгутиконосцами. Очевидно, что ни современные саркодовые, ни современные жгутиконосцы не могут быть исходной группой эволюции животного мира, ибо они сами прошли большой путь исторического развития и выработали многочисленные приспособления к современным условиям жизни на Земле. Вероятно, оба этих класса современных простейших следует рассматривать как два ствола в эволюции, берущих свое начало от древних, не сохранившихся до наших дней форм, которые жили на заре развития жизни на нашей планете.
В дальнейшей эволюции простейших произошли изменения различного характера. Некоторые из них привели к общему повышению уровня организации, повышению активности, интенсивности жизненных процессов. К числу таких филогенетических (эволюционных) преобразований следует, например, отнести развитие органоидов движения и захвата пищи, которое достигло высокого совершенства в классе инфузорий. Бесспорно, что реснички представляют собой органоиды, соответствующие (гомологичные) жгутикам. В то время как у жгутиконосцев, за немногими исключениями, число жгутиков невелико, у инфузорий число ресничек достигло многих тысяч. Развитие ресничного аппарата резко повысило активность простейших, сделало более многообразными и сложными формы их взаимоотношений со средой, формы реакций на внешние раздражения. Наличие дифференцированного ресничного аппарата, бесспорно, явилось одной из основных причин прогрессивной эволюции в классе инфузорий, где возникло большое многообразие форм, приспособленных к разным средам обитания.
Развитие ресничного аппарата инфузорий — пример такого рода эволюционных изменений, которые были названы акад. Северцовым ароморфозами. Ароморфозы характеризуются общим повышением организации, развитием приспособлений широкого значения. Под повышением организации понимают изменения, которые вызывают усиление жизнедеятельности организма; они связаны с функциональной дифференцировкой его частей, ведут к более многообразным формам связи организма с окружающей средой. Развитие ресничного аппарата инфузорий как раз и относится к такого рода преобразованиям строения в процессе эволюции. Это типичный ароморфоз.
У простейших, как это было подчеркнуто В. А. Догелем, изменения типа ароморфозов обычно бывают связаны с увеличением числа органоидов. Происходит полимеризация органоидов. Развитие ресничного аппарата у инфузорий— типичный пример такого рода изменений. Вторым примером ароморфоза в эволюции инфузорий может служить их ядерный аппарат. Мы рассмотрели выше особенности строения ядра инфузорий. Ядерный дуализм инфузорий (наличие микронуклеуса и макронуклеуса) сопровождался увеличением в составе макронуклеуса числа хромосом (явление полиплоидии). Поскольку хромосомы связаны с основными синтетическими процессами в клетке, в первую очередь с синтезом белков, этот процесс привел к общему повышению интенсивности основных жизненных функций. И здесь имела место полимеризация, затронувшая хромосомные комплексы ядра.
Инфузории — одна из наиболее многочисленных и прогрессивных групп простейших, происходит от жгутиконосцев. Об этом говорит полное морфологическое сходство их органоидов движения. Этот этап эволюции был связан с двумя большими ароморфозами: один из них затрагивал органоиды движения, второй — ядерный аппарат. Оба эти типа изменений связаны между собой, так как оба ведут к повышению жизнедеятельности и усложнению форм взаимосвязей с внешней средой.
Наряду с ароморфозами существует и другой тип эволюционных изменений, выражающихся в выработке приспособлений (адаптации) к определенным, резко очерченным условиям существования. Этот тип эволюционных изменений Северцовым был назван идиоадаптациями. В эволюции простейших этот тип изменений играл очень большую роль. Выше, при рассмотрении разных классов простейших, приведепы многочисленные примеры идиоадаптивных изменений. Приспособления к планктонному образу жизни у разных групп простейших, приспособления к жизни в песке у инфузорий, образование защитных оболочек ооцист у кокцидий и многое другое — все это идиоадаптации, игравшие большую роль в возникновении и развитии отдельных групп, но не связанные с общими прогрессивными изменениями организации.
Приспособления к различным конкретным средам обитания у простейших носят весьма разнообразный характер. Они обеспечили широкое распространение этого типа в самых разных средах обитания, что подробно рассматривалось выше, при описании отдельных классов.
Следует еще остановиться на вопросе о паразитизме у простейших. В типе простейших переход от свободного образа жизни к паразитизму распространен очень широко. Паразитические формы мы встречаем среди всех классов Protozoa. Кроме того, два класса — споровики и книдоспоридии — целиком состоят из паразитов. Один из них (споровики), как вы видели, взял свое начало от жгутиконосцев, второй (книдоспоридий) —от саркодовых. По ходу эволюции паразитических групп простейших у них выработались весьма сложные жизненные циклы, обеспечивающие как размножение паразитов, так и их широкое распространение в природе. Многие паразитические простейшие имеют большое практическое значение для человека как возбудители тяжелых заболеваний самого человека, домашних и промысловых млекопитающих, птиц, рыб, а также полезных насекомых. Поэтому борьба с протозойными паразитарными заболеваниями, профилактика и лечение их играют большую роль в медицинской и ветеринарной практике.
Жизнь животных: в 6-ти томах. — М.: Просвещение.
Под редакцией профессоров Н.А.Гладкова, А.В.Михеева.
1970.
Инфузория-туфелька
- Животные
Содержание
- Внешний вид инфузории туфельки
- Характер и образ жизни инфузории туфельки
- Где живет инфузория?
- Питание инфузории
- Размножение
- Опасна ли для человека?
- Разведение инфузории туфельки в домашних условиях
- Значение инфузорий в природе и жизни человека
Внешний вид инфузории туфельки
За свое сходство с подошвой женской обуви этот вид инфузорий приобрел второе название – «туфелька». Форма этого одноклеточного организма постоянна и не меняется с ростом или другими факторами. Все тело покрыто мельчайшими ресничками, похожими на жгутики эвглены. Удивительно, но этих ресничек на каждой особи насчитывается около 10 тысяч! С их помощью клетка передвигается в воде и захватывает пищу.
Инфузория туфелька, строение которой так знакомо по учебникам биологии, не видна невооруженным глазом. Инфузории представляют собой мельчайшие одноклеточные организмы, но при большом скоплении их можно увидеть и без увеличительных приборов. В мутной воде они будут выглядеть как продолговатые белые точки, находящиеся в постоянном движении.
Характер и образ жизни инфузории туфельки
Эти микроскопические существа обычно находятся в постоянном волнообразном движении, набирая скорость около двух с половиной миллиметров в секунду, что для таких ничтожно малых созданий в 5-10 раз превышает длину их тела. Передвижение инфузории туфельки осуществляется тупым концов вперёд, при этом она имеет обыкновение поворачиваться вокруг оси собственного тела.
Туфелька, резко взмахивая ресничками-ножками и плавно возвращая их на место, работает такими органами передвижения словно вёслами в лодке. Причём количество подобных взмахов имеет частоту около трёх десятков раз за одну секунду. Что же касается внутренних органоидов туфельки, большое ядро инфузории участвует в обмене веществ, движении, дыхании и питании, а малое отвечает за процесс воспроизводства.
Дыхание этих простейших созданий осуществляется следующим образом: кислород через покровы тела поступает в цитоплазмы, где с помощью данного химического элемента происходит окисление органических веществ и превращение их в углекислых газ, воду и прочие соединения. А в результате указанных реакций образуется энергия, употребляемая микроорганизмом для своей жизнедеятельности. После всего, вредный углекислый газ удаляется из клетки через её поверхности.
Особенность инфузории туфельки, как микроскопической живой клетки, состоит в способности этих крошечных организмов реагировать на внешнюю среду: механические и химические воздействия, влагу, тепло и свет. С одной стороны, они стремятся передвигаться к скоплениям бактерий для осуществления своей жизнедеятельности и питания, но с другой, вредные выделения этих микроорганизмов, заставляют инфузорий уплывать от них подальше. Также туфельки реагируют и на солёную воду, от которой спешат удалиться, зато с охотой передвигаются в сторону тепла и света, но в отличие от эвглены, инфузория туфелька настолько примитивна, что не имеет светочувствительного глазка.
Где живет инфузория?
Проживают инфузории в водоемах и очень часто становятся пищей для рыб и других обитателей морей и океанов. Основная среда обитания туфельки — пресные водоемы со стоячей водой. Питанием служат водоросли и бактерии. Встретить ее можно и в домашних аквариумах. Волнообразное движение ресничек позволяет ей передвигаться со скоростью до 2 мм/с.
Направление движения может меняться двумя способами:
- изгиб самой клетки — обычный вариант;
- столкновение с каким-то препятствием.
В последнем случае туфелька может развернуться на 180 градусов. Реснички туфельки помогают ей не только в передвижении. Они отвечают также за питание, создавая ток жидкости в направлении ротового отверстия инфузории. Часть ресничек прогоняет бактерии вдоль тела инфузории. Часть, склеенная в более сложные формы, помогает «заглатывать» еду. Ротовое отверстие, или клеточный рот, инфузории находится примерно посередине вогнутой части.
Внимание! Разводят туфельку и искусственным путём. Опытные аквариумисты знают, что идеальным кормом для мальков рыб является именно инфузория-туфелька. Более того, среди новорожденных существуют привереды, которые, кроме нее, ничем не питаются. На множестве интернет-проектов, посвященных аквариумистике, люди рассказывают о способах ее разведения.
Питание инфузории
Питание инфузории зависит от ее класса. Хищные сосальщики орудуют щупальцами. К ним прилипают, присасываются, проплывающие мимо одноклеточные. Питание инфузории туфельки осуществляется за счет растворения клеточной оболочки жертвы. Пленка разъедается в местах контакта со щупальцами. Изначально жертва, как правило, захватывается одним отростком. Прочие щупальца «подходят к уже накрытому столу». Реснитчатая форма инфузории туфельки питается одноклеточными водорослями, захватывая их ротовым углублением. Оттуда еда попадает в пищевод, а затем, в пищеварительную вакуоль. Она закрепляется на коне «глотки», отцепляясь от нее каждые несколько минут. После, вакуоль проходит по часовой стрелке к заду инфузории.
35 фактов про жирафов
Животные Крыма
36 фактов про слонов
Грызуны
35 фактов про лисиц
Животные Бразилии
Во время пути цитоплазмой усваиваются полезные вещества пищи. Отходы выбрасываются в порошицу. Это отверстие, подобное анальному. Во рту инфузории тоже есть реснички. Колышась, они создают течение. Оно увлекает частицы пищи в ротовую полость. Когда пищеварительная вакуоль перерабатывает еду, образуется новая капсула. Она тоже стыкуется с глоткой, получает пищу. Процесс цикличен. При комфортной для инфузории температуре, а это около 15 градусов тепла, пищеварительная вакуоль образуется каждые 2 минуты. Это указывает на скорость обмена веществ туфельки.
Размножение
Существует два типа размножения микроорганизмов:
- Бесполое, являющееся обычным делением.
 Этот процесс происходит как раздел одной инфузории туфельки надвое, причем новые организмы обладают своим большим и малым ядром. При этом в новую жизнь переходит только малая часть «старых» органоидов, все остальные быстро образуются заново.
Этот процесс происходит как раздел одной инфузории туфельки надвое, причем новые организмы обладают своим большим и малым ядром. При этом в новую жизнь переходит только малая часть «старых» органоидов, все остальные быстро образуются заново. - Половое. Этот тип применяется только при появлении температурных колебаний, недостаточности пищи и других неблагоприятных условиях. Именно тогда животные могут разделиться полами и затем превратиться в цисту.
Именно второй вариант размножения наиболее интересен:
- Две особи временно сливаются в одну;
- На месте слияния образуется некий канальчик, соединяющий пару;
- Большое ядро полностью исчезает (у обоих особей), а малое разделяется два раза.
Таким образом, каждая инфузория туфелька становится обладательницей двух ядер дочернего типа. Причем три ядра должны полностью разрушиться, а последнее снова поделиться. Из оставшихся двух ядер, которые снова обмениваются местами по мостику из цитоплазмы, формируется большое и малое. На этом процесс заканчивается и животные расходятся. Коньюгация позволяет перераспределить генетический материал между организмами, тем самым увеличивая жизненную силу и стойкость особей. И теперь они снова могут спокойно делиться на две новые жизни.
Опасна ли для человека?
Существует множество инфузорий, которые могут паразитировать в организме рыб и даже человека. Например, заболевание балантидиаз связано с присутствием балантидия кишечного в организме человека. А ихтиофтириус паразитирует в аквариумных рыбах, вызывая их массовую гибель. Однако инфузория-туфелька не представляет никакой угрозы для их здоровья. Она служит основной пищей для беспозвоночных и мальков рыб.
Разведение инфузории туфельки в домашних условиях
Необходимый инвентарь: трехлитровые банки, спринцовка (пластиковый наконечник следует заменить на стеклянную часть пипетки), линза с сильным увеличением, кусочек стекла. Быстрее всего размножение происходит на молоке, но и погибают микроорганизмы в нем за короткое время. Более выгодна высушенная кожура спелого банана (достаточно 3-х кв. см).
Культуру инфузории можно приобрести у любителей аквариумов либо добыть самостоятельно, зачерпнув воду со дна стоячего водоема. В последнем варианте капля воды помещается на стекло под микроскоп, где среди простейших хорошо видны инфузории. Рядом с каплей из водоема следует капнуть чистую воду. При помощи спички протягивается соединительный канал от одной капли к другой, по нему инфузории сами переплывут в более свежую воду. Переводя инфузорий снова и снова в каплю новой воды, получают чистую культуру этих микроорганизмов.
Дальше животных нужно отправить в инкубатор – набирается половина 3-х литровой банки чистой воды, добавляется 3 капли молока и помещается культура инфузорий. Банка помещается в теплом освещенном месте, но не под прямыми лучами солнца. Кормом для инфузорий служат бактерии, потому обычно они скапливаются у частичек органического материала.
Чтобы использовать туфельки на корм малькам, в емкость опускается водоросль, вокруг которой сразу появляются инфузории, собираются они при помощи пипетки и отправляются в аквариум, но не сразу. Следует немного подождать, пока животные съедят бактерий, обитающих в данных каплях воды, а уж после помещать жидкость с инфузориями в аквариум с мальками.
Значение инфузорий в природе и жизни человека
- Установлено, что инфузории играют значительную роль в круговороте веществ в природе. Инфузориями питаются различные виды более крупных животных (мальки рыб).
- Они служат регуляторами численности одноклеточных водорослей и бактерий, тем самым очищая водоемы.
- Инфузории могут служить индикаторами степени загрязнения поверхностных вод — источников водоснабжения.
- Инфузории, проживающие в почве, улучшают ее плодородие.
- Человек разводит инфузорий в аквариумах для кормления рыб и их мальков.
- В ряде стран широко встречаются заболевания человека и животных, вызываемые инфузориями. Особую опасность представляет инфузория балантидиум, обитающая в кишечнике свиньи и передающаяся человеку от животного.
Видео
Источники
- http://aquariumax.ru/drugie/infuzoriya.html
https://uchim.guru/biologiya/infuzoriya-tufelka-sreda-obitaniya.html
https://www.syl.ru/article/176084/new_infuzoriya-tufelka-stroenie-i-jiznedeyatelnost
https://givotniymir.ru/infuzoriya-tufelka-obraz-zhizni-i-sreda-obitaniya-infuzorii-tufelki/
https://givnost.ru/infuzoriya-tufelka-opisanie-osobennosti-stroenie-i-razmnozhenie-infuzorii-tufelki/
https://parazits.
ru/infuzoriya-tufelka-gde-obitaet-stroenie-i-funkcii/#infuz_tuf5
http://yachist.ru/parazityi/infuzoriya-tufelka-stroenie-forma-tela-obitanie/
https://animals-world.ru/klass-infuzorii/#i-4Жгутики и реснички — определение, строение и функции
Делиться — значит заботиться!
Эта статья охватывает
Что такое жгутики и реснички? – краткий обзор
Жгутики (единственное число: жгутик ) и реснички (единственное число: ресничка ) представляют собой два типа клеточных структур, которые обеспечивают движение в большинстве микроорганизмов и клеток животных, но не в клетках высоких растений. И жгутики, и реснички выглядят как бьющиеся волосовидные придатки, растущие на поверхности клеток. Обычно жгутики значительно длиннее ресничек. Однако одна клетка может иметь тысячи ресничек, но только один или два жгутика.
[В этом видео] Методы перемещения микроорганизмов: Euglena жгутиками, Amoeba псевдоподиями и Paramecium ресничками.
У одноклеточных микроорганизмов реснички и жгутики являются основными единицами движения. Например, реснички, покрывающие кожу Paramecium, помогают ему плавать, размахивая этими крошечными волосками. Один длинный, похожий на хлыст жгутик на голове эвглены может заставить клетку двигаться как пропеллер.
[На этом изображении] Структуры клеток включают реснички и жгутики у Paramecium и Euglena.
Обратите внимание, что направление движения эвглены определяется положением ее жгутика. С другой стороны, жгутик сперматозоида функционирует как хвост, который толкает клетку вперед, как подводную лодку.
В многоклеточных организмах реснички и жгутики также играют жизненно важную роль. Например, наши дыхательные пути выстланы множеством ресничек, которые удерживают вдыхаемую пыль, смог и потенциально вредные микроорганизмы от попадания в легкие. Мужские половые клетки, сперматозоиды, используют свои хвостовидные жгутики, чтобы двигаться к яйцеклетке.
[На этом рисунке] Клеточные структуры, обеспечивающие движение клеток животных: жгутик (хвост сперматозоида) и реснички (волнистые волоски на поверхности клеток дыхательных путей).
[На этом изображении] Сканирующая электронная микрофотография (СЭМ) реснитчатых эпителиальных клеток.
Источник изображения: Steve Gschmeissner/Science Photo Library
Часть 1. Жгутик
Жгутик (множественное число: жгутик) представляет собой длинную хлыстообразную структуру, выступающую из тела клетки. Обычно клетка имеет один или несколько (менее 5-10) жгутиков. Основная функция жгутика – движение клеток. Но некоторые специализированные жгутики могут действовать как сенсорные органеллы (например, антенны насекомых), будучи чувствительными к химическим веществам и температуре вне клетки.
У кого есть жгутики?
Клетки, имеющие жгутики, называются « жгутиковыми ». Жгутики можно найти среди трех доменов жизни — бактерий, архей и эукариот. Все три вида жгутиков могут использоваться для плавания, но они сильно различаются по структуре, белковому составу и механизму движения.
Примеры жгутиконосцев:
1. Helicobacter pylori – бактерия, вызывающая язву. Helicobacter pylori имеют несколько жгутиков, чтобы продвигаться через слизистую оболочку, чтобы достичь желудка.
[На этом изображении] Несколько жгутиков на одной клетке Helicobacter pylori (слева) и Helicobacter felis (справа).
Источник изображения: Электронные микрофотографии предоставлены Люси Томпсон, Школа биотехнологии и биомолекулярных наук, Университет Нового Южного Уэльса это изображение.
2. Жгутики архей имеют особое название, archaellum , чтобы подчеркнуть их отличие от бактериальных жгутиков.
[На этом изображении] Электронная микрофотография архей (слева: M. maripaludis и справа: S. acidocaldarius) , на которой видны многочисленные археи.
Источник изображения: Jarrell KF., et. al., Life , 2013
3. Примером жгутиковой клетки эукариот является сперматозоид. Он использует свой жгутик, чтобы продвигаться по половым путям самки.
[На этом изображении] Сперматозоиды кабана с длинными хвостовидными жгутиками под фазово-контрастным микроскопом.
Источник изображения: Компьютерное моделирование объектов, представленных в изображениях, Proc. CompImage Symposium 2006
Структура жгутика и принцип его работы
Бактериальные и архейные жгутики
Бактериальные жгутики представляют собой спиральных нитей , каждая из которых имеет вращательный двигатель в основании, который может вращаться по часовой стрелке или против часовой стрелки. Между двигателем и нитью находится сегмент, называемый «крючком», который функционирует как гибкое соединение и передает вращающую силу от двигателя к жгутику. Движение бактерий можно разделить на два вида:
- Бег – в результате вращения жгутика против часовой стрелки
- Кувыркание – в результате вращения жгутика по часовой стрелке
[На этом изображении] Подробная структура бактериального жгутика.
Бактериальный жгутик представляет собой длинный тонкий отросток тела клетки. Он вращается, как пропеллер, когда бактерия плывет.
Источник изображения: wiki
[На этом изображении] Крюк действует как динамическое соединение, передающее крутящий момент изнутри клетки для вращения внешнего хвоста, позволяя бактериям двигаться.
Источник изображения: доктор Сатоси Шибата
[На этом изображении] Бактериальные жгутики — одна из самых тонких биологических микромашин.
Примеры, показанные на этом изображении, представляют собой жгутиковый базовый комплекс двух видов бактерий — Salmonella enterica и Vibrio alginolyticus . Ученые использовали передовую криоэлектронную томографию (Cryo-ET), чтобы увидеть, как работают эти фантастические микромашины.
Источник изображения: Эхазаррета М. А. и Клозе К. Э., Фронт. Клетка. Заразить. микробиол. , 2019
[В этом видео] Посмотрите, как бактерии движутся под микроскопом.
Жгутики архей (archaella) похожи на жгутики бактерий и также работают как вращательный двигатель. Однако белки, образующие жгутики архей, отличаются от жгутиков бактерий.
Примечание. Бактерии имеют другой тип длинных волосков на своей клетке, называемый pili (единственное число: pilus). Пили представляют собой тонкие жесткие волокна, состоящие из белков, которые выступают из клеточных стенок. Пили помогают бактериям прикрепляться к определенным поверхностям или другим клеткам. Некоторые бактерии могут переносить ДНК из одной бактериальной клетки в другую с помощью своих специальных пили. Однако пили, реснички и жгутики представляют собой разные структуры.
Жгутики эукариот
Эукариотические клетки (клетки животных, растений и протистов) имеют свои жгутики совсем по-другому. Одно из основных отличий состоит в том, что прокариотические (бактериальные и архейные) жгутики совершают вращательных движений , в то время как эукариотические жгутики совершают изгибающих движений (которые качаются вперед и назад).
[На этом изображении] Прокариотические (спиральные) и эукариотические (хлыстовые) жгутики принципиально различаются. Вы можете видеть вращающийся двигатель бактериального жгутика (А) с шириной основания около 45 нм. (B) Бактерия, S. typhimurium , плавает с полярным пучком жгутиков. И наоборот, (C) жгутики эукариот питаются от пучков микротрубочек (диаметр поперечного сечения ~ 200 нм). Примером эукариотических жгутиков являются плавающие сперматозоиды на (D).
Источник изображения: Schwarz L., et. al., Applied Physics Reviews , 2017
Структурно эукариотические жгутики и реснички неразличимы. Оба обладают центральным пучком белков филаментов, называемым аксонемой . Каждая аксонема содержит девять пар по микротрубочек (дуплет), образующих внешнее кольцо и две центральные микротрубочки, известные как «9+2». Микротрубочки удерживаются вместе сшивающими белками.
В жгутиках/ресничках имеются моторные белки, называемые Dynein , расположенные поперек каждого парного волокна микротрубочек. Dynein использует АТФ в качестве энергии, чтобы ползти по микротрубочкам. Когда белки динеина движутся вверх с одной стороны и вниз с другой стороны, жгутик/ресничка изгибается. Повторение циклов сгибания-расслабления заставляет жгутики/реснички действовать как весла, взмахивая вперед и назад, создавая движение.
Обычно жгутики намного длиннее. Одна клетка может иметь только один или два жгутика в головной или хвостовой областях. С другой стороны, реснички короче по длине, но больше по количеству, обычно покрывая значительную площадь клеточной поверхности.
[На этом рисунке] Организация жгутиков и ресничек эукариот.
Каждый жгутик/ресничка содержит девять пар микротрубочек, образующих внешнее кольцо, и две центральные микротрубочки. Эта структура известна как аксонема. Микротрубочки удерживаются вместе сшивающими белками. Есть моторные белки, называемые динеином, расположенные поперек каждого парного волокна микротрубочек. 900:30 Фотография изменена из LadyofHats на вики.
Щелкните здесь, чтобы узнать, что такое микротрубочки и другие белки цитоскелета.
Практический пример – Эвглена
Эвглена (греч. eu = истина, glene = глазное яблоко) — род одноклеточных эукариот, обитающих в пресноводных прудах и канавах. Эвглена разделяет некоторые характеристики как растений, так и животных. Например, эвглена содержит хлоропласты; в результате они могут производить себе пищу, характерную для растений. Напротив, эвглена также может двигаться с помощью своих жгутиков и потреблять пищу посредством фагоцитоза, что характерно для животных. У эвглены также отсутствует клеточная стенка.
[На этом изображении] Анатомия Euglena и ее органеллы, включая два жгутика.
Один из двух жгутиков длинный и виден под световым микроскопом, а другой очень короткий и не выступает из клеток. Функция жгутиков — помогать эвглене плавать. Узнайте больше о строении эвглены здесь.
Практический пример – Volvox
Volvox – это род зеленых водорослей. Volvoxes — это свободно плавающие одноклеточные водоросли, но обычно они держатся вместе в виде сферических колоний (или шаров) из 500-50 000 клеток. Они могут жить в различных пресноводных средах обитания, включая пруды, бассейны и канавы. Под микроскопом вольвокс выглядит как зеленый шарик, медленно вращающийся, что делает его одним из самых очаровательных микроскопических организмов.
Эти клетки вольвокса, расположенные на поверхности сферы, несут два жгутика. Эти жгутики обращены в сторону окружающей воды и бьются, чтобы продвигать всю колонию по воде. Вот почему вольвокс движется как катящийся мяч.
[На этом изображении] Volvox представляет собой скопление множества одноклеточных зеленых водорослей. Каждая клетка, сидящая на поверхности, имеет два жгутика. Узнайте больше о Volvox.
Практический пример – Сперматозоиды
Единственными клетками человека со жгутиками являются мужские гаметы, то есть сперматозоиды. Сперматозоид (также называемый сперматозоидом) представляет собой мужскую репродуктивную клетку животных, которая несет в ядре гаплоидный набор хромосом. Сперматозоид состоит из головки, шейки, средней части и хвоста. Его ядро расположено в головной части. Остальная часть сперматозоида в основном представляет собой мощный пропеллер жгутика.
[На этом изображении] Структура сперматозоида.
За исключением головной части, сперматозоид образован своим жгутиком. В шейке сперматозоида находится центриоль, которая является корнем пучков микротрубочек. Эти пучки микротрубочек собираются в аксонему и удлиняются в длинный хвост. Оболочка покрывает осевые нити в качестве защиты в хвостовой части. В средней части осевая нить окружена множеством митохондрий, которые обеспечивают энергию для движения жгутика.
Источник изображения: BYJU’S
Практический пример — стадии жизненного цикла жгутиковых
Многие организмы не имеют жгутиков в зрелых формах. Однако на определенном этапе своего жизненного цикла у них развиваются специализированные жгутиковые формы. Например, сперматозоиды — это мужские половые клетки животных, многие из этих жгутиковых стадий жизненного цикла связаны с размножением. Примерами являются зооспоры (подвижные споры используют жгутик для движения) и мужские гаметы (мужские половые клетки) большинства зеленых/бурых водорослей, некоторых грибов, низших растений (печеночников, роголистников и мхов), саговников и гинкго.
Часть 2. Реснички
Реснички (множественное число: реснички) — структура, встречающаяся только в эукариотических клетках. Он имеет форму тонкого выступа, выступающего из тела клетки. На бактериях есть похожие структуры (то есть пили и фимбрии), но они не являются ресничками.
Многие микроорганизмы имеют реснички. Примеры включают Paramecium, Stentor, Vorticella, Lacrymaria, Coleps и Dileptus . Они классифицируются как группа под названием «инфузории», разделяя эту общую характеристику.
[В этом видео] Видеоколлекция нескольких обычных инфузорий под микроскопом.
Более крупные эукариоты, такие как животные, также имеют реснички. Реснички обычно присутствуют на поверхности клетки в большом количестве и бьются скоординированными волнами. У человека реснички находятся на эпителиальных клетках, выстилающих дыхательные пути. Эти реснички постоянно двигаются, очищая легкие от слизи и грязи.
Структура реснички и принципы ее работы
Реснички выглядят как множество крошечных волосков, растущих на поверхности клеток. Их корни представляют собой цитоскелетоподобные структуры, называемые базальным телом, расположенным под клеточной мембраной. Пучки микротрубочек отходят от базального тела, образуя осевое ядро ресничек. Как мы упоминали выше, и эукариотические реснички, и жгутики состоят из «9+2» расположения пучков микротрубочек. Движение создается за счет изгиба пучков микротрубочек с помощью моторных белков динеина.
[На этом рисунке] Ученые использовали передовые микроскопы, чтобы ответить на свои вопросы о ресничках парамеций.
Слева: Сканирующая электронная микроскопия показывает нам морфологию ресничек (Источник: Джудит Л. Ван Хаутен). В центре: Трансмиссионная электронная микроскопия дает подробное изображение поперечного сечения ресничек (Фото: Ричард Аллен). Справа: флуоресцентный микроскоп показывает нам, как реснички закрепляются на поверхности клетки.
Микротрубочки представляют собой белковые волокна внутри клеток, выполняющие множество функций. Микротрубочки также могут служить межклеточной магистралью для транспортировки молекул и органелл. Во время клеточного деления волокна микротрубочек, отходящие от двух центросом, растягивают хромосомы в новые ядра.
Два типа ресничек
Существует два основных типа ресничек: подвижные и неподвижные реснички. Подвижные реснички движутся, потребляя энергию для создания постоянного движения — для подвижности отдельных клеток (например, Paramecium) или транспортировки жидкости за пределы клеток (например, эпителий дыхательных путей).
Неподвижные реснички также называются первичными ресничками , которые служат сенсорными органеллами. Многие клетки имеют по крайней мере одну первичную ресничку на клетку. Например, наши обонятельные нейроны в носу обладают несколькими неподвижными ресничками, которые функционируют как клеточные антенны для восприятия молекул запаха.
[На этом изображении] Иллюстрация первичных ресничек на обонятельном сенсорном нейроне.
Обонятельный эпителий содержит обонятельные нейроны с множественными неподвижными отростками ресничек (обонятельные реснички). Реснички улавливают молекулы запаха, когда они проходят. Затем информация о молекулах запаха передается от обонятельных рецепторов к обонятельной луковице в головном мозге.
Источник изображения: Berbari N.F., et. al., Текущая биология , 2009
Практический пример – Paramecium
Paramecium (pair-ah-me-see-um; множественное число, Paramecia) — одноклеточный (одноклеточный) живой организм, по форме напоминающий башмачок. Paramecium естественным образом встречается в водной среде обитания. Вам нужен микроскоп, чтобы увидеть парамеции, потому что они имеют длину всего от 50 до 300 мкм (микрометров).
Кожа Paramecium покрыта множеством крошечных ресничек, похожих на тонкие волоски на нашем теле. Одна клетка Paramecium может иметь 5000-6000 ресничек. Каждая ресничка очень крошечная – примерно 0,25 мкм в диаметре и до 20 мкм в длину. Реснички Paramecium можно разделить на два типа: ротовые реснички и реснички тела. Оральные реснички расположены на поверхности ротовой бороздки. Они помогают собирать продукты питания. Реснички тела находятся на поверхности тела и облегчают его передвижение. Они действуют как микроскопические весла, двигая организм в одном направлении.
Реснички тела расположены продольными рядами (вдоль оси голова-хвост) с довольно равномерной длиной по всей клетке. Есть также несколько более длинных ресничек на заднем конце клетки (совершенно очевидно у P. caudatum ). Они образуют хвостовой пучок ресничек (отсюда и название caudatum).
[На этом рисунке] Различные типы ресничек на клетке парамеция – оральные реснички, реснички тела и каудальный пучок.
Если реснички Paramecium будут точно так же махать туда-сюда, клетки никуда не денутся. Удары вперед и назад должны быть в разных фазах, чтобы создать значимую движущую силу.
[В этом видео] Движение ресничек Paramecium, записанное высокоскоростной видеокамерой под микроскопом.
Чтобы ответить на этот вопрос, ученые использовали микроскоп с высокоскоростной видеокамерой, чтобы запечатлеть, как бьются реснички, приводя в движение все тело парамеция. Анализируя кадр за кадром высокоскоростное видео, ученые обнаружили, что парамеций плавает так же, как мы плаваем кролем вперед.
[На этом рисунке] Обводка ресничек на Paramecium.
Движение ресничек можно разделить на Эффективное (вперед) и Восстановительное (назад) взмахи. Два вида взмахов попеременно повторяются, чтобы привести в движение тело парамеция, когда мы плывем кролем на груди.
Узнайте больше о том, как реснички Paramecium создают волну ниже. У них роговидное тело с ресничками, равномерно покрывающими большую часть поверхности. Некоторые виды Stentors являются одними из крупнейших известных одноклеточных организмов на Земле, которые могут достигать 2 мм в длину.
Наиболее заметной частью Stentor является «корона» из ресничек, окружающих трубчатый «колокол». Эти реснички могут скоординировано колебаться взад и вперед, создавая поток или вихрь, который притягивает пищу ближе. Рядом с ротовым отверстием происходит слияние нескольких ресничек, образующих «суперреснички» или мембранеллы. Эти гигантские реснички образуют спираль, которая ведет в устье стентора. Время от времени стентор смыкает коронку реснички и сжимается, опуская пищу в тело.
[В этом видео] Под микроскопом вы можете увидеть эти более длинные реснички, образующие «корону» трубы Стентора. Также имеется множество более коротких ресничек, покрывающих все тело Stentor для свободного плавания.
Практический пример – Респираторный эпителий
Респираторный эпителий (или эпителий дыхательных путей) представляет собой тип реснитчатого столбчатого эпителия, который выстилает большую часть дыхательных путей. Его функции заключаются в увлажнении и защите дыхательных путей.
Каждая реснитчатая эпителиальная клетка имеет около 200 ресничек, которые постоянно сокращаются с частотой от 10 до 20 раз в секунду. Направление их ударов удаляет микробы и мусор вверх и из дыхательных путей.
[На этом изображении] Респираторный эпителий защищает наши легкие слизистой оболочкой и просветом ресничек.
Источник изображения: wiki
Часть 3. Каковы различия между жгутиками и ресничками?
В этой таблице приведены основные различия между жгутиками и ресничками.
| Символ | Жгутики | Реснички |
| Определение0010 | Жгутики представляют собой длинные нитевидные клеточные структуры, присутствующие на поверхности живой клетки. | Реснички представляют собой короткие, похожие на волоски клеточные структуры, отходящие от поверхности живой клетки. |
| Наличие | Жгутики встречаются как в прокариотических, так и в эукариотических клетках. | Встречается только в эукариотических клетках. |
| Длина | Длинный; длиной до 150 мкм. | Короткий; 5-10 мкм в длину. |
| Позиция | Обычно располагается на обоих или одном из концов ячейки. | Присутствует на всей поверхности клеток. |
| Плотность | Несколько (менее 10) на ячейку. | Много (от сотен до тысяч) на ячейку. |
| Движение | Вращательное движение у прокариот и изгибающее движение у эукариот; оба генерируют пропеллерное движение. | Быстрые волнообразные движения взад-вперед. |
| Биение | Жгутики бьются независимо друг от друга. | Реснички бьют согласованно или друг за другом. |
| Функции | В основном только при передвижении. | Передвижение, транспортировка, питание, восприятие, кровообращение и т. д. |
| Структура | Прокариотические: белки спиральных филаментов с вращающимся моторным основанием. Эукариотические: базальное тело, пучки микротрубочек (9+2), моторные белки. | То же, что и жгутики: базальное тело, пучки микротрубочек (9+2), моторные белки. |
| Примеры | Жгутиковые бактерии, подобные Helicobacter pylori, Salmonella ; некоторые археи. Euglena, Dinoflagellates, Зеленые водоросли Мужские гаметы, Сперматозоиды | Простейшие инфузории (Paramecium, Stentors и др.) Респираторный эпителий дыхательных путей человека |
Резюме
1. Клетки, помогающие жгутикам и ресничкам двигаться.
2. Жгутики представляют собой длинные нитевидные клеточные структуры, которые можно найти как в прокариотических, так и в эукариотических клетках. Однако их структура и белковый состав различны.
3. Реснички представляют собой короткие волосовидные клеточные структуры, отходящие от поверхности эукариотических клеток.
4. Жгутики и реснички эукариот одинаковы по своему строению. Оба они состоят из пучков микротрубочек в качестве ядра.
Делиться — значит заботиться!
Инфузории и их бактериальные партнеры
Путь к странице:
- Пресс-служба
- Инфузории и их бактериальные партнеры
12 июля 2017 г.
Глобальное партнерство
Инфузории, как и люди, заселены огромным разнообразием бактерий. Некоторые инфузории и их бактериальные симбионты стали друзьями на всю жизнь, как продемонстрировали исследователи Макса Планка и их коллеги, сравнив группу этих одноклеточных инфузорий и их бактериальных партнеров из Карибского и Средиземного морей. Они опубликовали свои выводы в Proceedings of the Royal Society B. Бактерии обеспечивают своих реснитчатых хозяев питанием, окисляя серу. Удивительно, но они обнаружили, что это партнерство возникло когда-то, от одного предка инфузорий и одного предка бактерий, хотя места отбора проб разделяет целый океан.
Изящные пловцы
Инфузории — крошечные одноклеточные организмы с несколькими ядрами, которых много в пресной воде, океанах и почве. Название «инфузория» происходит от «реснички», крошечных волосовидных структур, которые покрывают эти организмы и используются для движения и транспортировки пищи к ротовому отверстию. Хорошо известная инфузория – башмачок анималкуле 9.0059 Парамеций .
Под микроскопом становится очевидной изящность и красота инфузорий. Некоторые виды вырастают довольно большими и даже видны невооруженным глазом в виде маленьких точек в капле воды.
В своем исследовании Брэндон Сих из Института морской микробиологии Макса Планка (MPIMM) и его коллеги описывают партнерство между инфузориями рода Kentrophoros , которые потеряли ротовое отверстие, и симбиотическими сероокисляющими бактериями, от которых они зависят. Этот тип симбиоза называется мутуализмом, то есть оба партнера зависят друг от друга.
Хемосинтез и симбиоз как стратегия
Известно множество организмов, использующих в качестве источника энергии сероокисляющие бактерии. Первые были обнаружены по чистой случайности возле гидротермальных источников в глубоком море в 1970-х годах. Двумя примерами являются глубоководная мидия Bathymodiolus и трубчатый червь Riftia . До сих пор не было известно, кто такие симбионты Kentrophoros : связаны ли они с другими симбионтами или это совершенно новые виды бактерий?
Несколько инфузорий Kentrophoros из Средиземного моря под микроскопом. Сера в бактериях отражает свет и делает их белыми. Один из объектов — не инфузория, а пришелец, многоклеточный плоский червь. Каждая инфузория имеет длину до 3 мм. Источник: Oliver Jäckle, MPI MM. Поперечный срез инфузории Kentrophoros, окрашенной цветным красителем, показывает, как тело клетки инфузории складывается вокруг тысяч палочковидных бактерий, увеличивая площадь поверхности. Источник: МПИММ
В своем исследовании исследователи сравнили Kentrophoros видов из Карибского и Средиземного морей. Хотя общий вид (фенотип и морфология) различался, анализ последовательности ДНК показал, что все инфузории произошли от одного общего предка. То же самое было и с бактериями, которые все принадлежали к одной группе близких родственников из новой для науки линии. Это означает, что в какой-то момент миллионы лет назад первый Kentrophoros и предок этих бактерий сформировали партнерство, которое сохранилось на протяжении многих лет, и их потомки теперь можно найти по всему миру.
«Бактериальные симбионты растут только на одной стороне тела инфузории. Некоторые инфузории имеют специальные складки, чтобы увеличить площадь для оптимального роста. Эти инфузории несут свои личные огороды, которые они собирают путем фагоцитоза», — объясняет Брэндон Си, аспирант MPIMM.
Профессор, доктор Николь Дюбилье, директор MPIMM, говорит: «Один из удивительных результатов нашего исследования заключался в том, что партнерство между инфузориями и их симбионтами было очень стабильным и специфичным на протяжении очень длительного периода эволюции, возможно, от десятков до сотен лет. миллионов лет. Мы предположили, что, поскольку симбионты сидят снаружи своих хозяев и могут быть легко потеряны, когда инфузории передвигаются по воде или песку, эти симбиозы могут быть не такими специфичными, как те, в которых симбионты живут внутри своих хозяев. Но оказывается, что физическое расположение партнеров не обязательно связано с их близостью».
Исследователи обнаружили 17 видов Kentrophoros , которые все связаны друг с другом, имеют один и тот же базовый план тела, хотя каждый из них имеет свои уникальные особенности.
Перспективы
Следующим шагом на повестке дня является секвенирование генома бактериальных симбионтов и их хозяев. Кроме того, выращивание инфузорий и их симбионтов откроет двери для будущих исследований того, какой вклад каждый партнер в этой симбиотической команде вносит в их отношения.
Оригинальная публикация
Seah BKB, Schwaha T, Volland J-M, Huettel B, Dubilier N, Gruber-Vodicka HR. 2017 Специфичность в разнообразии: единое происхождение широко распространенного симбиоза инфузорий и бактерий. проц. Р. Соц. B 20170764.
http://dx.doi.org/10.1098/rspb.2017.0764
Институты
— Институт морской микробиологии им. и биоокеанография, Вена, Австрия
— Центр генома Макса Планка, Кельн, Институт исследований селекции растений им. Макса Планка, Кельн, Германия
— MARUM, Центр морских наук об окружающей среде, Бремен, Германия
Для получения дополнительной информации
Инфузории
Инфузории – одноклеточные организмы размером до нескольких миллиметров.
Название Kentrophoros означает «носитель шипов», поскольку исследователи в 1920-х годах ошибочно интерпретировали бактерии как шипы. Kentrophoros встречаются по всему миру, включая Средиземное море, Карибский бассейн и Тихий океан. Они могут размножаться бесполым путем.
Инфузории обычно имеют несколько клеточных ядер двух разных типов. Один тип мал и несет генетическую информацию, которой обмениваются во время полового размножения. Другие контролируют функции клетки и ее внешний вид посредством экспрессии генов.
Кентрофорос Фильм –Видеоролики живых инфузорий под микроскопом. Источник: Брэндон Сих
Директор
Проф. д-р Николь Дюбилье
MPI для морской микробиологии
Celsiusstr. 1
D-28359 Бремен
Германия
Комната: | 3241 |
Телефон: | +49 421 2028-9320 |
[Большая активация Javascript]
или свяжитесь с пресс-службой
Начальник отдела прессы и коммуникаций
Пресс-служба
Доктор Фанни Аспетсбергер
MPI для морской микробиологии
Celsiusstr. 1
D-28359 Бремен
Германия
Комната: | 1345 |
Телефон: | +49 421 2028-9470 |
[Большая активация Javascript]
Редкая пресноводная инфузория Paramecium chlorelligerum Kahl, 1935 и ее крупноядерная симбиотическая бактерия Candidatus Holospora parva
1. Кренек С., Берендонк Т.Ю., Фокин С.И. Новые конгенеры Paramecium (Ciliophora, Oligohymenophorea) формируют наш взгляд на его биоразнообразие. Разнообразие и эволюция организмов
2015 г.; 15: 215–233. [Академия Google]
2. Вихтерман Р. Биология Paramecium 2-е изд. Нью-Йорк: Пленум; 1986. [Google Scholar]
3. Фокин С.И. Род Paramecium : биоразнообразие, некоторые морфологические признаки и ключ к различению основных морфовидов. Протистология 2010 г.; 6: 227–235. [Google Scholar]
4. Мюллер О.Ф. Animalcula infusoria fluviatilia et marina. 1786.
5. Вихтерман Р. Биология Paramecium Нью-Йорк: Макгроу-Хилл; 1953. [Google Scholar]
6. Фокин С.И., Пшибош Е., Чивилев С.М., Бейер С.Л., Хорн М., Скотарчак Б. и соавт.
Морфологические и молекулярные исследования Paramecium schewiakoffi sp. ноябрь (Ciliophora, Oligohymenophorea) и текущий статус распространения и таксономии Paramecium spp.
Европейский журнал протистологии
2004 г. ; 40: 225–243. [Google Scholar]
7. Chatton E, Brachon S. Sur une paramécie à deux races: Paramecium duboscqui , n. сп. Comptes Rendus Societe Biolgie 1933; 114: 988–991. Французский. [Google Scholar]
8. Shi X, Jin M, Liu G. Повторное открытие Paramecium duboscqui Chatton and Brachon, 1933, и описание его характеристик. Журнал эукариотической микробиологии 1997 год; 44: 134–141. [Google Scholar]
9. Gelei J. Uj Paramecium szeged kornyekerol. Paramecium nefridiatum нов. сп. Allat Kozlony Zoology Mitteilungen 1925 год; 22: 121–162. Венгерский с немецким резюме. [Google Scholar]
10. Фокин С.И., Стоек Т., Шмидт Х.Ю. Новое открытие Paramecium nefridiatum Gelei, 1925 и его характеристика. Журнал эукариотической микробиологии 1999 г.; 46: 416–426. [Google Scholar]
11. Kahl A. Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 4. Peritricha und Chonotricha. Tierwelt Dtl. 1935 год; 30: 651–886. Немецкий. [Google Scholar]
12. Kreutz M, Stoeck T, Foissner W. Морфологическая и молекулярная характеристика Paramecium (Viridoparamecium nov. subgen.) chlorelligerum Kahl (Ciliophora). Журнал эукариотической микробиологии 2012 г.; 59: 548–563. 10.1111/j.1550-7408.2012.00638.x [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Фокин С.И., Герц Х.Д. Разнообразие бактерий Holospora в Paramecium и их характеристика В: Эндосимбионты в Paramecium . Спрингер Берлин Гейдельберг; 2009 г.; стр. 161–199. [Google Scholar]
14. Феррантини Ф., Фокин С.И., Ваннини С., Верни Ф., Петрони Г. Характеристика романа Голоспора — подобный симбионт из Frontonia (Ciliophora, Oligohymenophorea). Журнал эукариотической микробиологии 2007 г.; 54:75а. [Google Scholar]
15. Фокин С.И. Бактериальные эндоцитобионты Ciliophora и их взаимодействие с клеткой-хозяином. Международный обзор цитологии
2004 г.; 181–249. [PubMed] [Google Scholar]
16. Millot L, Kaltz O. Криоконсервация Holospora undulata , бактериального паразита инфузории Paramecium caudatum . Криобиология 2006 г.; 52: 161–165. 10.1016/ж.криобиол.2005.10.014 [PubMed] [CrossRef] [Академия Google]
17. Кастелли М., Ланцони О., Фокин С.И., Шраллхаммер М., Петрони Г. Реакция бактериального симбионта Holospora caryophila на различные условия роста его хозяина. Европейский журнал протистологии 2015 г.; 51: 98–108. 10.1016/j.ejop.2014.11.006 [PubMed] [CrossRef] [Google Scholar]
18. Кастелли М., Ланцони О., Росси Л., Потехин А., Шраллхаммер М., Петрони Г. Оценка протоколов обогащения бактериальных эндосимбионтов инфузорий методом ПЦР в реальном времени. Текущая микробиология 2016; 72: 723–732. 10.1007/с00284-016-1006-з [PubMed] [CrossRef] [Академия Google]
19. Dohra H, Tanaka K, Suzuki T, Fujishima M, Suzuki H. Черновые последовательности генома трех видов Holospora ( Holospora obtusa , Holospora undulata , Holospora undulata , Holosporanubiotic бактерий 9006 elegans), инфузория Paramecium caudatum . Письма по микробиологии FEMS
2014; 359: 16–18. 10.1111/1574-6968.12577
[PubMed] [CrossRef] [Google Scholar]
20. Banerji A, Duncan AB, Griffin JS, Humphries S, Petchey OL, Kaltz O. Опосредованные плотностью и признаками эффекты паразита и хищника в тритрофическом пищевой сети. Журнал экологии животных 2015 г.; 84: 723–733. 10.1111/1365-2656.12317 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Dusi E, Gougat-Barbera C, Berendonk TU, Kaltz O. Длительный эксперимент по отбору приводит к нарушению горизонтальной трансмиссивности у паразита со смешанным способом передачи. Эволюция 2015 г.; 69: 1069–1076. 10.1111/эво.12638 [PubMed] [CrossRef] [Google Scholar]
22. Дункан А.Б., Гонсалес А., Кальц О. Расселение, внешнее воздействие и паразиты в совокупности влияют на синхронность и стабильность метапопуляции. Экология 2015 г.; 96: 284–290. [PubMed] [Google Scholar]
23. Боскаро В., Фокин С. И., Шраллхаммер М., Швейкерт М., Петрони Г. Пересмотренная систематика Holospora -подобные бактерии и характеристика « Candidatus Gortzia infectiva», нового макроядерного симбионта Paramecium jenningsi . Микробная экология
2013; 65: 255–267. 10.1007/s00248-012-0110-2
[PubMed] [CrossRef] [Google Scholar]
24. Аманн Р.И., Спрингер Н., Людвиг В., Герц Х.Д., Шлейфер К.Х. Идентификация in situ и филогения некультивируемых бактериальных эндосимбионтов. Природа 1991 год; 351: 161–164. 10.1038/351161а0 [PubMed] [CrossRef] [Академия Google]
25. Hori M, Fujii K, Fujishima M. Micronucleus-специфичная бактерия Holospora elegans необратимо усиливает экспрессию гена стресса хозяина Paramecium caudatum . Журнал эукариотической микробиологии 2008 г.; 55: 515–521. 10.1111/j.1550-7408.2008.00352.x [PubMed] [CrossRef] [Google Scholar]
26. Ваккерок-Коузова Н.Д., Раутиан М.С. Получение и характеристика « Holospora curviuscula » и Holospora obtusa , бактериальных симбионтов макронуклеусов Paramecium bursaria и Paramecium caudatum , микробиология 2011 г.; 80: 728–732. [PubMed] [Google Scholar]
27. Rautian MS, Wackerow-Kouzova ND. Филогенетическое размещение двух ранее описанных внутриядерных бактерий из инфузории Paramecium bursaria (Protozoa, Ciliophora): « Holospora acuminata » и « Holospora curviuscula ». Международный журнал систематической и эволюционной микробиологии
2013; 63: 1930–1933. 10.1099/ижс.0.046631-0
[PubMed] [CrossRef] [Академия Google]
28. Каракашян С.Ж., Каракашян М.В. Эволюция и симбиоз у рода Chlorella и родственных водорослей. Эволюция 1965 год; 19: 368–377. [Google Scholar]
29. Nichols HW, Bold HC. Trichosarcina polymorpha род. и др. ноябрь Журнал психологии 1965 год; 1: 34–38. [Google Scholar]
30. Сковородкин И.Н. Устройство для иммобилизации биологических объектов при исследованиях в световом микроскопе. Цитология 1990 г.; 32: 301–302. Русский с аннотацией на английском. [Академия Google]
31. Корлисс Дж.О. Импрегнация серебром реснитчатых простейших по методике Чаттона-Льоффа. Технология окрашивания
1953 год; 28: 97–100. [PubMed] [Google Scholar]
32. Фойсснер В. Основные методы световой и сканирующей электронной микроскопии для таксономических исследований реснитчатых простейших. Европейский журнал протистологии 1991 год; 27: 313–330. 10.1016/С0932-4739(11)80248-8 [PubMed] [CrossRef] [Google Scholar]
33. Фокин С.И., Герц Х.Д. Caedibacter macronucleorum sp. nov., бактерия, обитающая в макронуклеусе Paramecium duboscqui . Архив Протистенкунде 1993 год; 143: 319–324. [Google Scholar]
34. Приер Л.Б. Альфа, инфекционный макронуклеарный симбионт Paramecium aurelia . Журнал протозоологии 1969 год; 16: 570–578. [PubMed] [Google Scholar]
35. Андреоли И., Манджини Л., Феррантини Ф., Сантанджело Г., Верни Ф., Петрони Г. Молекулярная филогения некультивируемых кариореликтей (Alveolata, Ciliophora). Зоологическая письменность 2009 г.; 38: 651–662. [Google Scholar]
36. Медлин Л., Элвуд Х.Дж., Стикель С., Согин М.Л. Характеристика ферментативно амплифицированных 16S-подобных рРНК-кодирующих областей. Ген
1988; 71: 491–499. [PubMed] [Google Scholar]
37. Петрони Г., Дини Ф., Верни Ф., Розати Г. Молекулярный подход к запутанным внутриродовым отношениям, лежащим в основе филогении Euplotes (Ciliophora, Spirotrichea). Молекулярная филогенетика и эволюция 2002 г.; 22: 118–130. 10.1006/mpev.2001.1030 [PubMed] [CrossRef] [Google Scholar]
38. Фокин С.И., Андреоли И., Верни Ф., Петрони Г. Apofrontonia dohrni sp. н. и филогенетические отношения в пределах Peniculia (Protista, Ciliophora, Oligohymenophorea). Зоологическая письменность 2006 г.; 35: 289–300. [Google Scholar]
39. Strüder-Kypke MC, Wright ADG, Fokin SI, Lynn DH. Филогенетические отношения рода Paramecium выведены из последовательностей генов малых субъединиц рРНК. Молекулярная филогенетика и эволюция 2000 г.; 14: 122–130. 10.1006/mpev.1999.0686 [PubMed] [CrossRef] [Google Scholar]
40. Rosati G, Modeo L, Melai M, Petroni G, Verni F. Междисциплинарный подход к описанию протистов: морфологическое, ультраструктурное и молекулярное исследование Peritromus kahli Villeneuve-Brachon, 1940 (Ciliophora, Heterotrichea). Журнал эукариотической микробиологии
2004 г.; 51: 49–59. [PubMed] [Google Scholar]
41. Боскаро В., Фокин С.И., Верни Ф., Петрони Г. Исследование Paramecium duboscqui с использованием трех маркеров и оценка молекулярной изменчивости в роде Paramecium . Молекулярная филогенетика и эволюция 2012 г.; 65: 1004–1013. 10.1016/ж.ымпев.2012.09.001 [PubMed] [CrossRef] [Академия Google]
42. Strüder-Kypke MC, Lynn DH. Сравнительный анализ митохондриального гена субъединицы I цитохром с-оксидазы (COI) у инфузорий (Alveolata, Ciliophora) и оценка его пригодности в качестве маркера биоразнообразия. Систематика и биоразнообразие 2010 г.; 8: 131–148. [Google Scholar]
43. Vannini C, Rosati G, Verni F, Petroni G. Идентификация бактериальных эндосимбионтов морской инфузории Euplotes magnicirratus (Ciliophora, Hypotrichia) и предложение « Candidatus 9″0060 Devosia euplotis». Международный журнал систематики и эволюционной микробиологии
2004 г. ; 54: 1151–1156. [PubMed] [Google Scholar]
44. Лейн DJ. Секвенирование 16S/23S рРНК В: Stackebrandt E., Goodfellow M. (eds.), Nucleic Acid Techniques in Bacterial Systematics, Wiley, New York: 1991 год; стр. 115–147. [Google Scholar]
45. Cole JR, Wang Q, Fish JA, Chai B, McGarrell DM, Sun Y и другие. Проект базы данных рибосом: данные и инструменты для высокопроизводительного анализа рРНК. Исследования нуклеиновых кислот 2014; [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P и другие. Проект базы данных генов рибосомной РНК SILVA: улучшенная обработка данных и веб-инструменты. Исследования нуклеиновых кислот 2013; [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Greuter D, Loy A, Horn M, Rattei T. ProbeBase — онлайн-ресурс для олигонуклеотидных зондов и праймеров, нацеленных на рРНК: новые функции. Исследования нуклеиновых кислот 2016; [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Manz W, Amann RI, Ludwig W, Wagner M, Schleifer KH. Филогенетические олигодезоксинуклеотидные зонды для основных подклассов протеобактерий: проблемы и решения. Систематическая и прикладная микробиология
1992; 15: 593–600. [Google Scholar]
49. Аманн Р.И., Биндер Б.Дж., Олсон Р.Дж., Чизхолм С.В., Деверо Р., Шталь Д.А. Комбинация олигонуклеотидных зондов, нацеленных на 16S рРНК, с проточной цитометрией для анализа смешанных микробных популяций. Прикладная и экологическая микробиология 1990 г.; 56: 1919–1925. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O. Новые алгоритмы и методы оценки филогений максимального правдоподобия: оценка производительности PhyML 3.0. Систематическая биология 2010 г.; 59: 307–21. 10.1093/sysbio/syq010 [PubMed] [CrossRef] [Google Scholar]
51. Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar, et al.
ARB: программная среда для данных последовательности. Исследования нуклеиновых кислот
2004 г.; 32: 1363–1371. 10.1093/нар/гх393
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Ronquist F, Teslenko M, Van der Mark P, Ayres DL, Darling A, Höhna D, et al. MrBayes 3.2: Эффективный байесовский филогенетический вывод и выбор модели в большом модельном пространстве. Систематическая биология 2012 г.; 61: 539–542. 10.1093/sysbio/sys029 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: больше моделей, новая эвристика и параллельные вычисления. Природные методы 2012 г.; 9: 772. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Цукер М. Веб-сервер Mfold для прогнозирования укладки нуклеиновых кислот и гибридизации. Исследования нуклеиновых кислот 2003 г.; 31: 3406–3415. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Coleman AW. Повторное посещение Paramecium aurelia . Журнал эукариотической микробиологии
2005 г. ; 52: 68–77. 10.1111/ж.1550-7408.2005.3327р.х
[PubMed] [CrossRef] [Google Scholar]
56. Эдгар Р.С. MUSCLE: множественное выравнивание последовательностей с высокой точностью и высокой пропускной способностью. Исследования нуклеиновых кислот 2004 г.; 32: 1792–1797. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Фучикова К., Льюис П.О., Льюис Л.А. Широко распространенная пустынная принадлежность требуксиофитовых водорослей (Trebouxiophyceae, Chlorophyta), включая открытие трех новых пустынных родов. Психологические исследования 2014; 62: 292–305. [Google Scholar]
58. Fawley MW, Fawley KP, Owen HA. Разнообразие и экология мелких кокковидных зеленых водорослей из озера Итаска, штат Миннесота, США, включая Meyerella planktonica , gen. и др. ноябрь Психология 2005 г.; 44: 35–48. [Google Scholar]
59. Rathinasamy K, Preeti R, Kalimuthu JR, Varalakshmi P. Биосинтез жирных кислот из нового изолята Meyerella sp. N4: молекулярная характеристика, голодание по питательным веществам и профилирование жирных кислот для повышения уровня липидов. Энергия и топливо
2015 г.; 29: 143–149. [Google Scholar]
60. Holzinger A, Karsten U. Стресс высыхания и толерантность у зеленых водорослей: последствия для ультраструктуры, физиологических и молекулярных механизмов. Границы в науке о растениях 2013; [бесплатная статья PMC] [PubMed] [Google Scholar]
61. Андреева В.М. Почвенные и аэрофильные зеленые водоросли (Chlorophyta: Tetrasporales, Chlorococcales, Chlorosarcinales) Наука, Санкт-Петербург: 1998 год; 352 стр. Русский. [Google Scholar]
62. Фокин С.И., Бригге Т., Бреннер Дж., Герц Х.Д. 9Виды 0059 Holospora , поражающие ядра Paramecium , по-видимому, принадлежат к двум группам бактерий. Европейский журнал протистологии 1996 год; 32: 19–24. [Google Scholar]
63. Гиллеспи Дж.Дж., Нордберг Э.К., Собрал Б.В., Азад А.Ф. Филогения и сравнительная геномика: меняющийся ландшафт в эпоху геномики. Внутриклеточные патогены II; риккетсиозы . Американское общество микробиологии;
2012, стр. 84–141. [Google Scholar]
64. Дрисколл Т., Гиллеспи Дж. Дж., Нордберг Э. К., Азад А. Ф., Собрал Б. В. Бактериальная ДНК, просеянная из Trichoplax adhaerens (Animalia: Placozoa) Геномный проект выявил предполагаемого риккетсиозного эндосимбионта. Геномная биология и эволюция 2013; 5: 621–645. 10.1093/gbe/evt036 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
65. Montagna M, Sassera D, Epis S, Bazzocchi C, Vannini C, Lo N и другие. « Candidatus Midichloriaceae» сем. ноябрь ( Rickettsiales ), экологически широко распространенная клада внутриклеточных альфапротеобактерий. Прикладная и экологическая микробиология 2013; 79: 3241–3248. [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Соколи Ф., Сабанеева Э., Кастелли М., Кренек С., Шраллхаммер М., Соарес КАГ и другие. « Candidatus Fokinia solitaria», новая «автономная» симбиотическая линия Midichloriaceae ( Rickettsiales ). ПЛОС ОДИН 2016; [Бесплатная статья PMC] [PubMed] [Google Scholar]
67. Hess S, Suthaus A, Melkonian M. « Candidatus Finniella» ( Rickettsiales , Alphaproteobacteria ), новые эндосимбионты Viridiraptorid Amoeboflagellates (Cercozoa, Rhizaria). Прикладная экологическая микробиология
2016; 82: 659–670. [бесплатная статья PMC] [PubMed] [Google Scholar]
68. Фокин С.И. Частота и биоразнообразие симбионтов у представителей основных классов Ciliophora. Европейский журнал протистологии 2012 г.; 48: 138–148. 10.1016/j.ejop.2011.12.001 [PubMed] [CrossRef] [Google Scholar]
69. Finlay BJ, Maberly SC, Esteban GF. Впечатляющее обилие инфузорий в бескислородной воде пруда: вклад симбионтного фотосинтеза в потребности хозяина в кислороде для дыхания. FEMS Микробиология Экология 1996; 20: 229–235. [Google Scholar]
70. Summerer M, Sonntag B, Sommaruga R. Инфузорно-симбионтная специфичность пресноводного эндосимбиотика Chlorella . Журнал психологии 2008 г.; 44: 77–84. [PubMed] [Google Scholar]
71. Esteban GF, Fenchel T, Finlay BJ. Миксотрофия инфузорий. Протист
2010 г.; 161: 621–641. 10.1016/ж.протис.2010.08.002
[PubMed] [CrossRef] [Google Scholar]
72. Быкова С.В. Развитие миксотрофных инфузорий водоемов нижнего и среднего течения рек Волги и Камы как индикатор особых условий обитания. Новости Самарский научный центр РАН 2013; 15: 2224–2233. Русский с аннотацией на английском. [Академия Google]
73. Кройц М., Фойснер В. Сфагновые пруды Зиммельрида в Германии: очаг биологического разнообразия микроскопических организмов. Протозоогические монографии 2006 г.; 3: 1–267. [Google Scholar]
74. Vuxanovici A. Noi contributii la studiul ciliatelor dulcicole din Republica Populara Romina (примечание 1). Студии Церк. биол. 1960 г.; 12: 353–381. Румынский с русским и французским резюме. [Google Scholar]
75. Berninger U-G, Finlay BJ, Canter HM. Пространственное распределение и экология инфузорий, несущих зоохлореллы, в продуктивном водоеме. Журнал протозоологии 1986; 33: 557–563. [Google Scholar]
76. Петрони Г., Розати Г., Ваннини К., Модео Л., Дини Ф., Верни Ф. Идентификация in situ с помощью флуоресцентно меченных олигонуклеотидных зондов морфологически сходных, близкородственных видов инфузорий. Микробная экология
2003 г.; 45: 156–162. 10.1007/s00248-002-2016-х
[PubMed] [CrossRef] [Google Scholar]
77. Fried J, Foissner W. Дифференциация двух очень похожих глаукомидных морфовидов инфузорий (Ciliophora, Tetrahymenida) по флуоресценции гибридизация in situ с олигонуклеотидными зондами-мишенями 18S рРНК. Журнал эукариотической микробиологии 2007 г.; 54: 381–387. 10.1111/j.1550-7408.2007.00277.x [PubMed] [CrossRef] [Google Scholar]
78. Хошина Р., Хаяши С., Имамура Н. Внутривидовая генетическая дивергенция Paramecium bursaria и реконструкция филогенетического древа Paramecian. Acta Protozoologica 2006 г.; 45: 377–386. [Google Scholar]
79. Прешольд Т., Дариенко Т., Сильва П.С., Рейссер В., Криниц Л. Систематика Зоохлорелла пересмотрена с использованием комплексного подхода. Экологическая микробиология
2011 г.; 13: 350–364. 10.1111/j.1462-2920.2010.02333.x
[PubMed] [CrossRef] [Google Scholar]
80. Neustupa J, Němcová Y, Vesela J, Steinová J, Škaloud P. Leptochlorella corticola gen. и др. ноябрь и Kalinella apyrenoidosa sp. nov.: две новые хлореллоподобные зеленые микроводоросли (Trebouxiophyceae, Chlorophyta) из субаэральных местообитаний. Международный журнал систематики и эволюционной микробиологии 2013; 63: 377–387. [PubMed] [Академия Google]
81. Питч Г., Адамек Л., Диррен С., Ницше Ф., Симек К., Сирова Д. и другие. Зеленый Tetrahymena utriculariae n. сп. (Ciliophora, Oligohymenophorea) с ее эндосимбиотическими водорослями ( Micractinium sp.), обитающими в ловушках хищного водного растения. Журнал эукариотической микробиологии 2016; [PubMed] [Google Scholar]
82. Хошина Р., Кусуока Ю. Анализ ДНК водорослевых эндосимбионтов инфузорий выявляет состояние интеграции водорослей и удивительную специфичность симбиоза. Протист
2016; 167: 174–184. 10.1016/ж.протис.2016.02.004
[PubMed] [CrossRef] [Академия Google]
83. Van Etten JL, Meints RH, Kuczmarski D, Burbank DE, Lee K. Вирусы симбиотических Chlorella -подобных водорослей, выделенных из Paramecium bursaria и Hydra viridis . Труды Национальной академии наук Соединенных Штатов Америки 1982 год; 79: 3867–3871. [PMC free article] [PubMed] [Google Scholar]
84. Kawakami H, Kawakami N. Поведение вируса в симбиотической системе Paramecium bursaria -Zoochlorella. Журнал протозоологии 1978; 25: 217–225. [Google Scholar]
85. Ященко ВВ, Гаврилова ОВ, Раутиан М.С., Якобсен К.С. Ассоциация вирусов Paramecium bursaria Chlorella с клетками Paramecium bursaria : ультраструктурные исследования. Европейский журнал протистологии 2012 г.; 48: 149–159. 10.1016/j.ejop.2011.05.002 [PubMed] [CrossRef] [Google Scholar]
86. Görtz HD, Dieckmann J. Жизненный цикл и инфекционность Holospora elegans Haffkine, микроядерного симбионта Paramecium caudatum (Эренберг). Протистология
1980 г.; 16: 591–603. [Google Scholar]
87. Осипов Д.В., Скобло И.И., Борхсениус О.Н., Раутиан М.С. Holospora acuminata– новый вид симбиотической бактерии из микроядра инфузории Paramecium bursaria Focke. Цитология (Санкт-Петербург) 1980 г.; 22: 922–929. Русский с аннотацией на английском. [Google Scholar]
88. Громов БВ, Осипов ДВ. Holospora (ex Hafkine 1890) ном. rev., род бактерий, населяющих ядра парамеций. Международный журнал систематической бактериологии 1981; 31: 348–352. [Google Scholar]
89. Preer JR, Preer LB. Возрождение названий эндосимбионтов простейших и предложение Holospora caryophila nom. ноябрь Международный журнал систематической бактериологии 1982 год; 32: 140–141. [Google Scholar]
90. Борхсениус О.Н., Скобло И.И., Осипов Д.В. Holospora curviuscula — новый вид крупноядерных симбиотических бактерий Paramecium bursaria . Цитология
1983 год; 25: 91–97. Русский с аннотацией на английском. [Академия Google]
91. Фокин С.И. Бактериальные эндобионты инфузории Paramecium woodruffi . I. Эндобионты макронуклеуса. Цитология 1989 год; 31: 839–844. Русский с аннотацией на английском. [Google Scholar]
92. Фокин С.И. Holospora recta sp. nov. – микроядерный эндобионт инфузории Paramecium caudatum . Цитология 1991 год; 33: 135–141. Русский с аннотацией на английском. [Google Scholar]
93. Фокин С.И., Сабанеева Е.В. Бактериальные эндоцитобионты инфузории Paramecium calkinsi . Европейский журнал протистологии 1993 год; 29: 390–395. 10.1016/С0932-4739(11)80401-3 [PubMed] [CrossRef] [Google Scholar]
94. Фокин С., Бригге Т., Герц Х-Д. Инфекционная бактерия, обитающая в макронуклеусе Paramecium putrinum . Журнал эукариотической микробиологии 1999 г.; 46: 11а. [Google Scholar]
95. Ferla MP, Thrash JC, Giovannoni SJ, Patrick WM. Новые филогении Alphaproteobacteria , основанные на генах рРНК, дают представление об основных группах, митохондриальном происхождении и филогенетической нестабильности. ПЛОС ОДИН
2013; [Бесплатная статья PMC] [PubMed] [Google Scholar]
96. Георгиадес К., Мадуи М.А., Ле П., Роберт С., Роулт Д. Филогеномный анализ Odyssella thessalonicensis подтверждает общее происхождение Rickettsiales , Pelagibacter ubique и Reclinomonas americana. ПЛОС ОДИН 2011 г.; [Бесплатная статья PMC] [PubMed] [Google Scholar]
97. Schulz F, Martijn J, Wascher F, Lagkouvardos I, Kostanjšek R, Ettema TJG, et al. Симбионт амеб Rickettsiales с древними чертами. Экологическая микробиология 2015 г.; [PubMed] [Академия Google]
98. Соколи Ф., Кастелли М., Сабанеева Е., Шраллхаммер М., Кренек С., Доак Т.Г. и другие. Разделение таксономии Rickettsiales и описание двух новых симбионтов (« Candidatus Bealeia paramacronuclearis» и « Candidatus Fokinia cryptica»), разделяющих цитоплазму инфузорного простейшего Paramecium biaurelia . Прикладная и экологическая микробиология 2016; [Бесплатная статья PMC] [PubMed] [Google Scholar]
99. Серра В., Фокин С., Кастелли М., Басури К.К., Нитла В.М., Верни Ф.
и другие. Candidatus Gortzia shahrazadis», новый эндосимбионт Paramecium multimicronucleatum и пересмотр биогеографического распространения Holospora -подобных бактерий. Границы микробиологии
2016; [Бесплатная статья PMC] [PubMed] [Google Scholar]
100. Wiemann M, Görtz HD. Высвобождение эндонуклеобиотической бактерии Holospora elegans из клетки-хозяина Paramecium caudatum . Европейский журнал протистологии 1989 год; 25: 100–108. 10.1016/С0932-4739(89)80021-5 [PubMed] [CrossRef] [Google Scholar]
101. Фокин С.И. Выделение голоспороподобных бактерий у разных видов инфузорий. Резюме VII Европейского конгресса протистологов, Севилья, Испания, 5-10 сентября 2015 г.
102. Ярза П., Йилмаз П., Прюссе Э., Глекнер Ф.О., Людвиг В., Шлейфер К.Х.
и другие.
Объединение классификации культивируемых и некультивируемых бактерий и архей с использованием последовательностей генов 16S рРНК. Природа Обзоры Микробиология
2014; 12: 635–645. 10.1038/нрмикро3330
[PubMed] [CrossRef] [Академия Google]
Что такое парамеций? (с картинками)
`;
С. Митра
Paramecium — это одноклеточный организм, принадлежащий к царству Protista, так что это не совсем растение или животное. Организм может переваривать пищу, передвигаться по воде с помощью ресничек и размножаться. Как один из древнейших организмов на Земле, он эволюционировал и развил очень простые методы защиты, генетического обмена и мобильности.
Существует несколько различных видов парамеций, но все они относятся к реснитчатым простейшим. Это означает, что они используют реснички, чтобы плавать в воде, а их единственная сложная клетка, эукариот, выполняет все основные функции организма. Он не разделяет труд между различными тканями или клетками, как животное. Вместо этого каждый парамеций способен к аэробному обмену, подобному дыханию, бесполому размножению, поглощению питательных веществ и удалению отходов.
Внешний вид
При хорошем зрении можно разглядеть пятнышко парамеция, поскольку его длина составляет около 0,02 дюйма (0,5 мм). Их лучше рассматривать под микроскопом, где можно увидеть клетку в форме башмачка или почки. Снаружи его мембраны выстилают крошечные бьющиеся волоски, называемые ресничками. Реснички движутся вместе, как ряд весел на корабле, чтобы перемещать парамеций через жидкость. Эукариот даже достаточно умен, чтобы объезжать препятствия и приближаться к еде.
Когда парамеций сталкивается с едой, он поворачивается, чтобы переместить пищу в пищевод. Пищевод представляет собой небольшое отверстие, похожее на рот. Он выстлан другими ресничками, чтобы помочь «проглотить» кусочки органического или разлагающегося вещества, которое он ест, например, других одноклеточных организмов или бактерий. Пища будет продолжать спускаться по пищеводу, чтобы храниться в пищевых вакуолях до тех пор, пока клетке не понадобится энергия.
Возможно, также удастся различить другие округлые структуры, называемые органеллами, которые функционируют подобно органам животного. Одной из таких органелл являются сократительные вакуоли. Парамеций должен поддерживать осмотическое равновесие, а это означает, что давление воды вне его кожи и внутри его тела должно быть постоянно одинаковым. Сократительные вакуоли пропускают воду изнутри клетки наружу и наоборот.
Репродукция
В большинстве случаев парамеции размножаются, разделяя себя пополам и давая каждому новому организму половину органелл. Это называется бинарным делением и представляет собой простую форму бесполого размножения. Иногда парамеций встречается с другим и обменивается генетическим материалом во время своего рода примитивного полового размножения. Мембранные оболочки объединяются в один гигантский парамеций, после чего крошечные микроядра, содержащие весь генетический материал, меняются местами. Когда он делится на четыре меньшие парамеции, у них появляются новые комбинации ДНК.
Цитоплазматическое наследование: инфузории | Микробные модели молекулярной биологии: от генов к геномам
Фильтр поиска панели навигации Oxford AcademicМикробные модели молекулярной биологии: от генов к геномамБиохимияМолекулярная и клеточная биологияКнигиЖурналы Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Oxford AcademicМикробные модели молекулярной биологии: от генов к геномамБиохимияМолекулярная и клеточная биологияКнигиЖурналы Термин поиска на микросайте
Расширенный поиск
Иконка Цитировать Цитировать
Разрешения
- Делиться
- Твиттер
- Подробнее
CITE
Davis, Rowland H. ,
‘Цитоплазматическое наследование: реснички
,
Микробные модели молекулярной биологии: от генов до геном
(
, Нью -Йорк,
2003; онлайн-издание,
Oxford Academic
, 1 апреля 2010 г.
), https://doi.org/10.1093/acprof:oso/9780195154368.003.0013,
, по состоянию на 24 сентября 2022 г.
Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Фильтр поиска панели навигации Oxford AcademicМикробные модели молекулярной биологии: от генов к геномамБиохимияМолекулярная и клеточная биологияКнигиЖурналы Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Oxford AcademicМикробные модели молекулярной биологии: от генов к геномамБиохимияМолекулярная и клеточная биологияКнигиЖурналы Термин поиска на микросайте
Advanced Search
Abstract
В этой главе рассматривается основная проблема биологии начала 20-го века: роль цитоплазмы, особенно в развитии. В нем обсуждаются исследования Paramecium и других инфузорий, жизненный цикл Paramecium, проблема спаривания Paramecium и модельный статус Paramecium.
Ключевые слова: цитоплазма, развитие, модельные организмы, Paramecium, цитоплазматическое наследование
Тема
БиохимияМолекулярная и клеточная биология
В настоящее время у вас нет доступа к этой главе.
Войти
Получить помощь с доступом
Получить помощь с доступом
Доступ для учреждений
Доступ к контенту в Oxford Academic часто предоставляется посредством институциональных подписок и покупок. Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту одним из следующих способов:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения. Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Нажмите Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- Находясь на сайте учреждения, используйте учетные данные, предоставленные вашим учреждением. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Войти с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Доступ члена общества к журналу достигается одним из следующих способов:
Войти через сайт сообщества
Многие общества предлагают единый вход между веб-сайтом общества и Oxford Academic. Если вы видите «Войти через сайт сообщества» на панели входа в журнале:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Вход через личный кабинет
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам. Смотри ниже.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам.
Просмотр ваших зарегистрированных учетных записей
Щелкните значок учетной записи в правом верхнем углу, чтобы:
- Просмотр вашей личной учетной записи и доступ к функциям управления учетной записью.
- Просмотр институциональных учетных записей, предоставляющих доступ.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Ведение счетов организаций
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т. д.
Покупка
Наши книги можно приобрести по подписке или приобрести в библиотеках и учреждениях.
Информация о покупке
вопросов с несколькими вариантами ответов для экзамена 1, биология 250
вопросов с несколькими вариантами ответов для экзамена 1, биология 250Примеры вопросов с несколькими вариантами ответов для экзамена 1 по биологии 250
Буквы для правильных ответов заглавные.
- 1. Скользкое внешнее покрытие у некоторых бактерий, которое защищает их от фагоцитоза клетками-хозяевами,
- A. капсула b. клеточная стенка в. жгутик д. пептидогликан
- 2. Когда жгутики распределены вокруг бактериальной клетки, такое расположение называется
- а. полярный б. случайный C. peritrichous d. инкапсулированный
- 3. Блестящая, липкая колония Streptococcus pneumoniae, вероятно,
- A. инкапсулированные и патогенные c. неинкапсулированные и непатогенные
- б. неинкапсулированные и патогенные d. инкапсулированные и непатогенные
- 4. Клеточная стенка бактерий выполняет все следующие функции, кроме
- а. придает форму и жесткость клетке б. является местом действия некоторых антибиотиков
- с. связан с некоторыми симптомами заболевания D. защищает клетку от фагоцитоза
- 5. Минимальное расстояние, на котором микроскоп способен различить две точки как отдельные, является его
- а. увеличение б. освещенность C. разрешающая способность
- 6. Грамотрицательная клеточная стенка __________, чем грамположительная.
- а. толще Б. тоньше
- 7. Поскольку пенициллин препятствует синтезу пептидогликана, он более эффективен в отношении _______________ клеток.
- A. Грамположительные b. грамотрицательный
- 8. Жгутики и пили состоят из
- а. липиды б. углеводы в. нуклеиновые кислоты D. белок
- 9. Генетическая информация бактерий хранится в _______, в одной кольцевой хромосоме, расположенной в цитоплазме.
- А. ДНК б. белок в. фосфолипиды д. РНК
- 10. Различия между эукариотическими и прокариотическими клетками включают все перечисленное, кроме
- а. эукариотические клетки имеют митохондрии c. прокариотические клетки имеют более сложную клеточную стенку
- б. эукариотические клетки имеют реснички и жгутики сложной структуры D. прокариотические клетки не имеют генетического материала
- 11. Тот факт, что вирусы являются облигатными внутриклеточными паразитами, означает, что для размножения им требуется ____________________.
- а. культуральная чашка B. клетка-хозяин c. феноловый красный бульон d. вторичный вирус
- 12. Одним из способов определения размера, клеточной морфологии и группировки бактериальных клеток является использование __________ методики.
- а. полосовая пластина б. феноловый красный C. простая окраска d. питательная бульонная культура
- 13. Помутнение является признаком роста бактерий в -_______________ после инокуляции и инкубации.
- а. пластинка с штрихами B. пробирка с питательным бульоном c. окраска по Граму д. простое пятно
- 14. Бактериальная оболочка включает все следующие структуры, кроме
- а. капсула б. клеточная стенка в. клеточная мембрана D. эндоспора
- 15. Все следующие структуры бактерий содержат белок (или состоят из него), кроме
- A. плазмиды b. рибосомы в. пили д. клеточная мембрана
- 16. Что из следующего содержит полисахарид?
- A. Стенка грамотрицательных клеток b. пили c. жгутики д. плазмиды
- 17. Что из перечисленного содержит ДНК?
- а. Грамположительная клеточная стенка б. капсула в. пили D. плазмиды
- 18. При окрашивании по Граму бактерии с грамположительными клеточными стенками обесцвечиваются менее легко, чем бактерии с грамотрицательными клеточными стенками.
А. верно б. ЛОЖЬ
- 19. Тип клеточной стенки с меньшим содержанием пептидогликана – это
- A. Грамотрицательные b. грамположительный
- 20. Наиболее уязвимым для действия пенициллина типом клеточной стенки является
- а. Грамотрицательные B. Грамотрицательные
- 21. Когда жгутики расположены вокруг всей бактериальной клетки, такое расположение называется
- а. полярный б. случайный в. биполярный Д. перитрихоз
- 22. Инкапсулированная клетка будет размножаться, образуя колонии, которые появляются
- а. непатогенный б. полупрозрачный в. розовый Д. гладкий
- 23. Эндоспорами являются все перечисленные, кроме ___________ по сравнению с вегетативными клетками.
- г. н.э. больше шансов выжить после обработки дезинфицирующими средствами c. более устойчив к окрашиванию
- B. больше шансов умереть в условиях плохого питания d. более устойчивы к перепадам температуры
- 24. Фагоцитоз бактерий, вызывающих пневмонию, ____ вероятен, если бактерии защищены капсулой.
- А. меньше b. более
- 25. Если вы используете микроскоп видимого света для изучения живой культуры бактерий, обладающих жгутиками, вы сможете увидеть, как жгутики двигают бактерии.
- а. правда Б. ложь
- 26. В какой фазе кривой роста находится культура (для ответов используйте буквы фаз кривой роста):
- наиболее чувствительны к антибиотикам? а, В-экспоненциальный рост, в, г
- имеют максимальное количество питательных веществ? А-начальный стационарный, б, в, г
- имеют максимальное количество накопленных продуктов жизнедеятельности? а, б, в, Д-экспоненциальная смерть
- имеют одинаковое количество делящихся и умирающих клеток? A-начальная стационарность, b, C-максимальная стационарность, d
- имеют максимальное количество питательных веществ? А-начальный стационарный, б, в, г
- 27.