Значение простейших. Какие функции она выполняет
Стр. 46
Вспомните
1.Какими чертами строения эвглена зеленая и инфузория — туфелька отличаются от амебы протея?
Эвглена зеленая способна менять характер питания в зависимости от условий среды. В клетке инфузории два ядра, большое ядро отвечает за дыхание, питание, движение, обмен веществ, а малое за размножение.
2.В каких средах жизни обитают простейшие?
Простейшие живут только в жидкой среде — в воде разнообразных водоемов — от морей до болот, внутри растений и животных.
3.Какие организмы называют паразитами?
Паразиты — это организмы, питающиеся за счет особей другого вида и временно или постоянно пребывающие на поверхности их тела или внутри организма.
Стр. 49
Вопросы после параграфа
1.
Клетка простейших является самостоятельным организмом.
2.Назовите меры, предупреждающие заболевания амебной дизентерией и малярией.
Меры предупреждением большинства болезней служит, элементарное соблюдение гигиены, мытье рук перед едой, хорошая и качественная обработка продуктов питания перед использованием в пищу.
3.Сформулируйте вывод о роли простейших в природе и их влиянии на человека.
Простейшие — это питание для других животных. Они нужны для образования горных пород, могут быть показателем загрязнения в водоеме. Большинство ведут паразитический образ жизни, потому как селятся в организмах животных и людей и приносят вред.
Подведем итоги
1.Объясните, почему клетка простейших является самостоятельным организмом.
Клетки простейших выполняют все функции организма.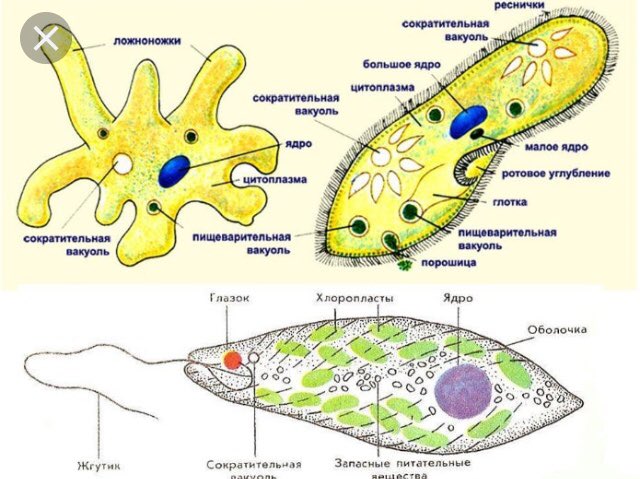
2.Охарактеризуйте среды обитания одноклеточных. Какое условие является обязательным для их существования?
Простейшие могут жить только в жидкой среде — в воде разнообразных водоёмов — от морей до капелек, держащихся на моховых «подушках» болот, в увлажнённой почве, внутри растений и животных.
3.Объясните, в чем заключаются функции вакуолей в организме одноклеточных.
Пищеварительная вакуоль отвечает за захват пищи и переваривание, после она выносит ненужные останки из организма. В сократительной вакуоли скапливаются вредные вещества, которые образуются в теле простейшего, достигнув определённого размера вакуоль, подплывает к поверхности тела и выбрасывает «мусор» наружу.
4.Установите взаимосвязь строения и способов движения одноклеточных.
Двусторонняя симметрия — активно передвигающиеся. Радиальная симметрия — неподвижный образ жизни.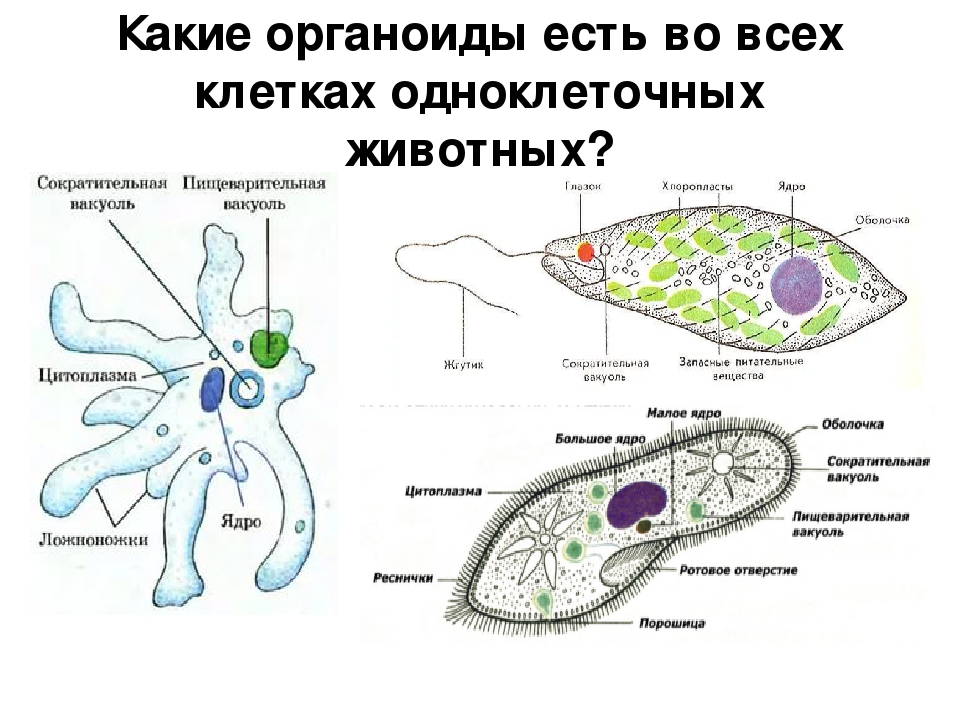
5.Назовите черты приспособленности простейших к неблагоприятным условиям.
Циста. При неблагоприятных условиях простейшие «прячутся» в неё и находятся там, до наступления благоприятных условий жизни.
6.Опишите роль в природе двух — трех представителей простейших, обитающих в водной среде.
Основная роль, которую играют простейшие, обитающие в водной среде (например, инфузория туфелька или амеба) заключается в том, что они служат пищей для более крупных и развитых организмов (например, моллюсков). Некоторые простейшие (например, радиолярии) принимают участие в образовании горных пород, которые состоят из остатков их скелетов.
7.Назовите меры предупреждения заболеваний, вызываемых простейшими.
Избегать контакта с водой ( имеется ввиду не пить воду из родников и озёр), а также не употреблять в пищу организмов, заражённых этими же простейшими и паразитами.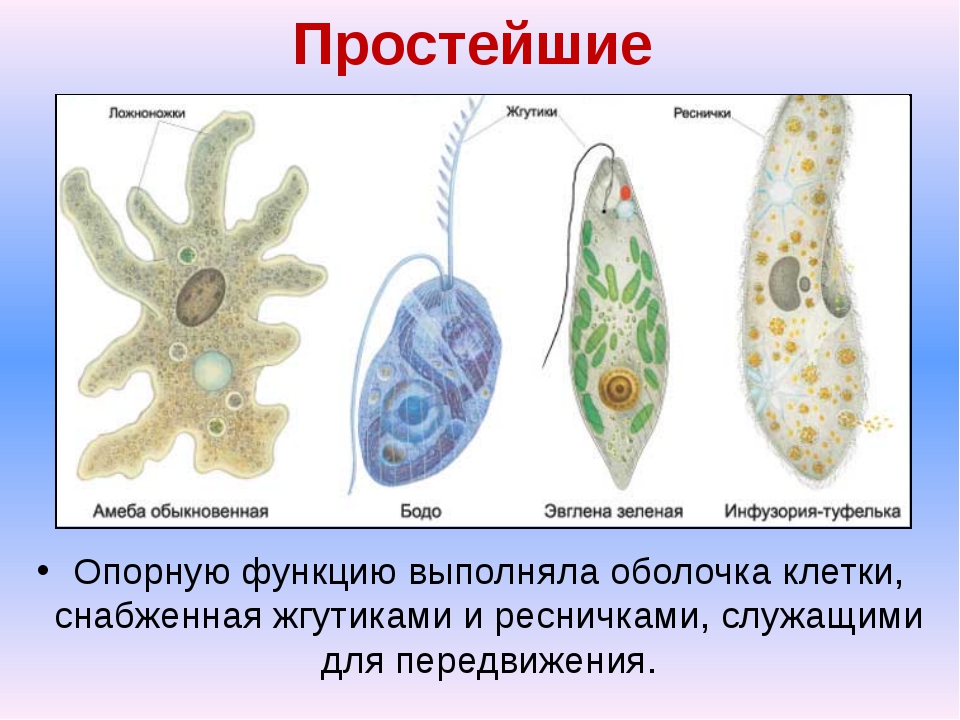 Меры предупреждением большинства болезней служит, элементарное соблюдение гигиены, мытье рук перед едой, хорошая и качественная обработка продуктов питания перед использованием в пищу.
Меры предупреждением большинства болезней служит, элементарное соблюдение гигиены, мытье рук перед едой, хорошая и качественная обработка продуктов питания перед использованием в пищу.
Какие утверждения верны?
1. Клетка простейших выполняет роль самостоятельного организма.
2. Размножение у амёбы бесполое, а у инфузории — туфельки и бесполое, и половое.
4. Эвглена зелёная является переходной формой от растений к животным: имеет хлорофилл, как у растений, а питается гетеротрофно и передвигается, как животные.
6. Малое ядро у инфузории участвует в половом размножении, а большое отвечает за жизнедеятельность.
Какое строение имеет клетка простейших? Подробное описание
Знаете ли вы, какое строение имеет клетка простейших? Если нет, то эта статья для вас.
Какая наука изучает клетку?
Эта наука называется цитологией. Она является отраслью биологии. Она и может ответить на вопрос, какое строение имеет клетка простейших. Также данная наука изучает не только структуру, но и процессы, которые происходят в клетке. Это клеточное дыхание, обмен веществ, размножение и фотосинтез. Способ размножения простейших — простое деление клетки. Некоторые клетки простейших способны осуществлять фотосинтез — выработку органических веществ из неорганических. Клеточное дыхание происходит при расщеплении глюкозы. В этом и заключается главная функция простых углеводов в клетке. При их окислении клетка получает энергию.
Она и может ответить на вопрос, какое строение имеет клетка простейших. Также данная наука изучает не только структуру, но и процессы, которые происходят в клетке. Это клеточное дыхание, обмен веществ, размножение и фотосинтез. Способ размножения простейших — простое деление клетки. Некоторые клетки простейших способны осуществлять фотосинтез — выработку органических веществ из неорганических. Клеточное дыхание происходит при расщеплении глюкозы. В этом и заключается главная функция простых углеводов в клетке. При их окислении клетка получает энергию.
Кто такие простейшие?
Перед тем как рассматривать вопрос о том, какое строение имеет клетка простейших, давайте разберемся, что из себя представляют эти «существа».
Это организмы, которые состоят из одной клетки. Они называются еще эукариотами, так как в их клетках есть ядро. Клетка простейших во многом похожа на клетку многоклеточного организма.
Классификация
Существует шесть типов простейших:
- инфузории;
- радиолярии;
- солнечники;
- споровики;
- саркожгутиконосцы;
- жгутиковые.

Представители первого типа населяют соленые водоемы. Некоторые виды также могут жить в почве.
Споровики в основном представлены паразитами позвоночных животных.
Радиолярии, как и инфузории, обитают в океанах. Они имеют твердые оболочки из диоксида кремния, из которых формируются некоторые горные породы.
Особенность солнечников заключается в том, что они передвигаются с помощью псевдоподий.
Саркожгутиконосцы также используют такой способ передвижения. К этому типу относятся амебы и многие другие простейшие.
Жгутиковые представлены множеством разнообразных организмов, которые используют для передвижения жгутики. Некоторые виды таких простейших могут жить в водоемах, а некоторые являются паразитами. Кроме того, у многих представителей данного типа в клетках присутствуют хлоропласты. Такие простейшие сами вырабатывают необходимые для жизни питательные вещества с помощью фотосинтеза.
Какое строение имеет клетка простейших?
Структуру клетки можно разделить на три основных части: плазматическую мембрану, цитоплазму и ядро.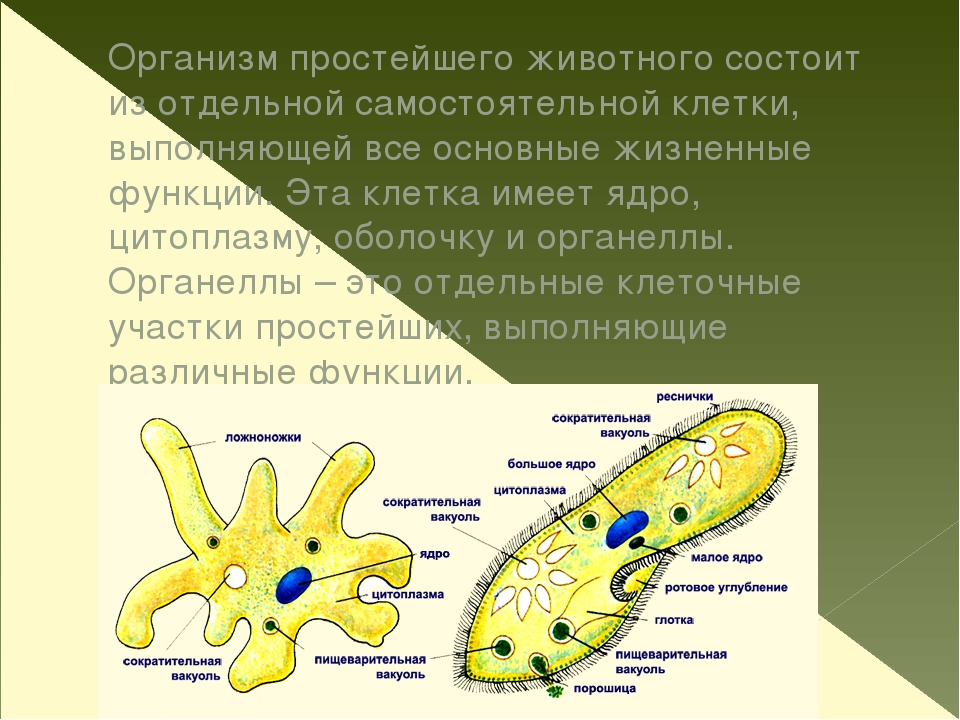 Количество ядер в клетках простейших равняется одному. Этим они отличаются от клеток бактерий, которые вообще не имеют ядер. Итак, рассмотрим детально каждый из трех компонентов клетки.
Количество ядер в клетках простейших равняется одному. Этим они отличаются от клеток бактерий, которые вообще не имеют ядер. Итак, рассмотрим детально каждый из трех компонентов клетки.
Плазматическая мембрана
Строение клетки простейших обязательно предусматривает наличие этой составляющей. Она отвечает за поддержание гомеостаза клетки, защищает ее от воздействий внешней среды. Плазматическая мембрана состоит из липидов трех классов: фосфолипидов, гликолипидов и холестерола. Преобладают в структуре мембраны фосфолипиды.
Цитоплазма: как она устроена?
Это вся та часть клетки, за исключением ядра, которая находится внутри плазматической мембраны. Она состоит из гиалоплазмы и органоидов, а также включений. Гиалоплазма — это внутренняя среда клетки. Органоиды являются постоянными структурами, которые выполняют определенные функции, а включения —это непостоянные структуры, которые выполняют в основном запасающую функцию.
Строение клетки простейших: органоиды
В клетке простейших присутствуют многие органоиды, которые свойственны для животных клеток.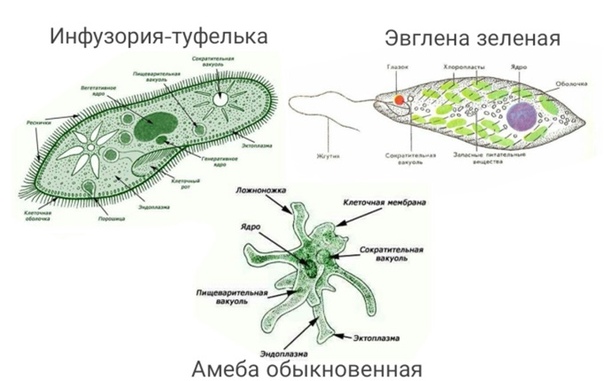 Кроме того, в отличие от клеток многоклеточных организмов большинство клеток простейших обладают органоидами движения — всевозможными жгутиками, ресничками и другими структурами. Наличием таких образований могут похвастаться очень немногие клетки многоклеточных животных — только сперматозоиды.
Кроме того, в отличие от клеток многоклеточных организмов большинство клеток простейших обладают органоидами движения — всевозможными жгутиками, ресничками и другими структурами. Наличием таких образований могут похвастаться очень немногие клетки многоклеточных животных — только сперматозоиды.
К органоидам, которые присутствуют в клетках простейших, относятся митохондрии, рибосомы, лизосомы, эндоплазматический ретикулум, комплекс Гольджи. В клетках некоторых простейших также находятся хлоропласты, которые характерны для растительных клеток. Рассмотрим строение и функции каждого из них в таблице.
| Органоид | Строение | Функции |
| Митохондрии | Обладают двумя мембранами: внешней и внутренней, между которыми присутствует межмембранное пространство. Внутренняя мембрана обладает выростами — кристами или гребнями. На них и происходят все основные химические реакции. То, что находится внутри обоих мембран, называется матриксом. В нем у этих органоидов присутствуют свои рибосомы, включения, митохондриальная РНК и митохондриальная ДНК. То, что находится внутри обоих мембран, называется матриксом. В нем у этих органоидов присутствуют свои рибосомы, включения, митохондриальная РНК и митохондриальная ДНК. | Выработка энергии. В этих органоидах происходит процесс клеточного дыхания. |
| Рибосомы | Состоят из двух субъединиц. Не имеют мембран. Одна из субъединиц обладает большим размером, чем вторая. Рибосомы объединяются только в процессе функционирования. Когда органоид не функционирует, две субъединицы находятся раздельно. | Синтез белков (процесс трансляции). |
| Лизосомы | Обладают округлой формой. Имеют одну мембрану. Внутри мембраны находятся ферменты, которые необходимы для расщепления сложных органических веществ. | Клеточное пищеварение. |
| Эндоплазматический ретикулум | Трубчатая форма. | Участвует в обмене веществ, отвечает за синтез липидов. |
| Комплекс Гольджи | Стопка дискообразных цистерн. | Служит для синтеза гликозамингликанов, гликолипидов. Модифицирует и классифицирует белки. Модифицирует и классифицирует белки. |
| Хлоропласты | Обладают двумя мембранами с межмембранным пространством между ними. В матриксе находятся тилакоиды, объединенные в стопки (граны ламеллами. Кроме того, в матриксе находятся рибосомы, включения, РНК и ДНК. | Фотосинтез (происходит в тилакоидах). |
| Вакуоли | Многие простейшие, населяющие пресные водоемы, обладают сократительными вакуолями (шарообразными органоидами с одной мембраной) | Выкачивание из организма лишней жидкости. |
Кроме того, клетки простейших снабжены органоидами движения. Это могут быть жгутики и реснички. В зависимости от вида, организм может обладать как одним, так и несколькими жгутиками.
29. Обзор простейших. Их строение и жизнедеятельность . Общая биология
Простейшие – это одноклеточные организмы, тело которых состоит из цитоплазмы и одного или нескольких ядер. Клетка простейшего – это самостоятельная особь, проявляющая все основные свойства живой материи. Она выполняет функции всего организма.
Она выполняет функции всего организма.
Одна клетка умеет делать все: и питаться, и двигаться, и нападать, и спасаться от врагов, и переживать неблагоприятные условия среды, и размножаться, и избавляться от продуктов обмена, и защищаться от высыхания и от чрезмерного проникновения воды внутрь клетки.
Размеры простейших от 3—150 мкм до 2–3 см в диаметре.
Известно около 100 000 видов простейших. Среда их обитания – вода, почва, организм хозяина (для паразитических форм).
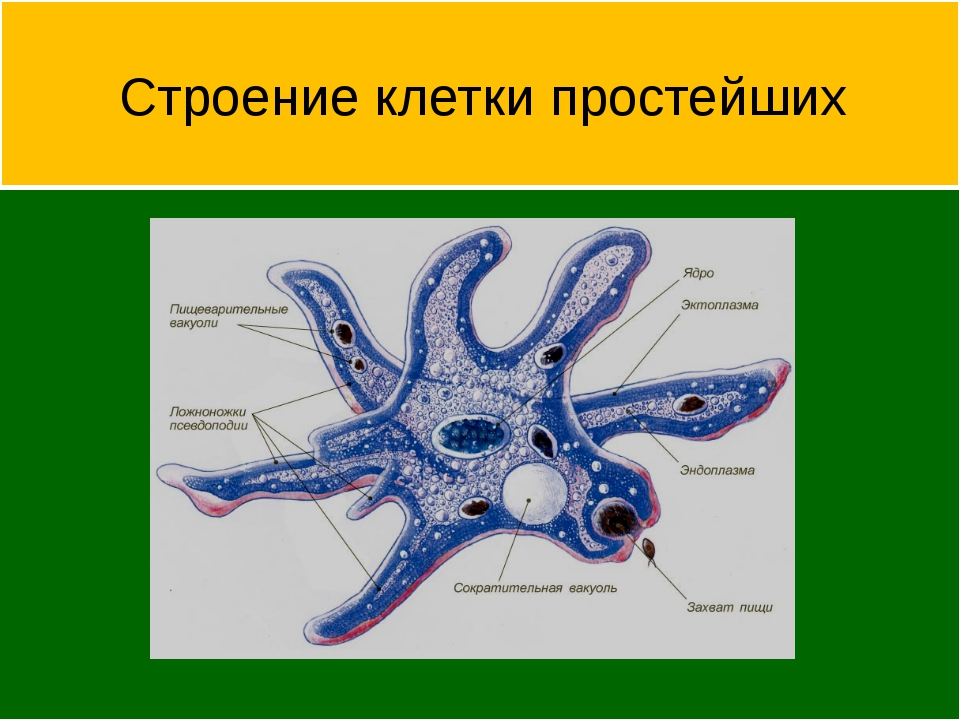
Простейшие имеют органеллы общего (митохондрии, рибосомы, клеточный центр, ЭПС и др.) и специального назначения. Органы движения: ложноножки, жгутики, реснички, пищеварительные и сократительные вакуоли.
Большинство простейших имеет одно ядро, но есть представители с несколькими ядрами. Ядра характеризуются полиплоидностью.
Цитоплазма неоднородна. Она подразделяется на более светлый и гомогенный наружный слой, или эктоплазму, и зернистый внутренний слой, или эндоплазму. Наружные покровы представлены либо ци-топлазматической мембраной (у амебы), либо пелликулой (у эвглены).
Подавляющее большинство простейших – гетеро-трофы. Их пищей могут служить бактерии, детрит, соки и кровь организма хозяина (для паразитов). Непереваренные остатки удаляются через порошицу или через любое место клетки. Через сократительные вакуоли осуществляется осмотическая регуляция, удаляются продукты обмена.
Дыхание происходит через всю поверхность клетки.
Раздражимость представлена таксисами.
Размножение простейших
Бесполое – митозом ядра и делением клетки надвое (у амебы, эвглены, инфузории), а также путем шизогонии – многократного деления (у споровиков).
Половое – копуляция. Клетка простейшего становится функциональной гаметой; в результате слияния гамет образуется зигота.
Для инфузорий характерен половой процесс – конъюгация. Клетки обмениваются генетической информацией, но увеличения числа особей не происходит.
Простейшие способны существовать в двух формах – трофозоита (вегетативной формы, способной к активному питанию и передвижению) и цисты, которая образуется при неблагоприятных условиях.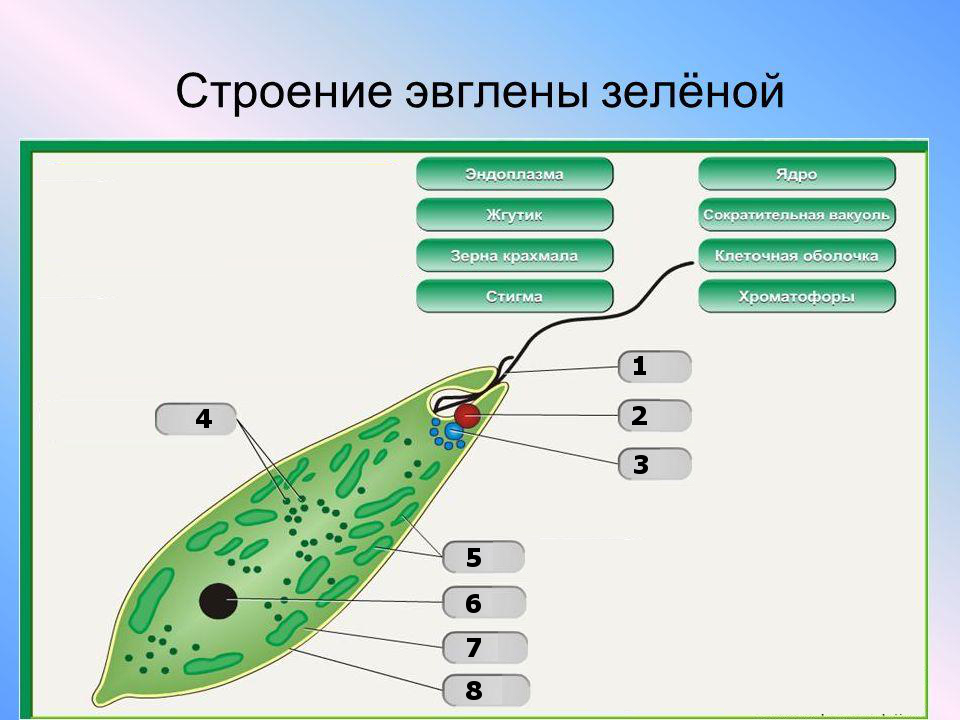 При попадании в благоприятные условия обитания происходит эксцистирование, клетка начинает функционировать в состоянии трофозоита.
При попадании в благоприятные условия обитания происходит эксцистирование, клетка начинает функционировать в состоянии трофозоита.
Для многих представителей типа Protozoa характерно наличие жизненного цикла.
Время генерации для простейших составляет 6– 24 ч.
Заболевания, вызываемые простейшими, называются протозойными.
Эти загадочные Простейшие
Дидактическая цель: cоздать условия для закрепления и осмысления блока информации посредством использования исследовательского метода обучения.
Учебная цель: способствовать осознанию, взаимосвязи между строением живой системы и ее функциями; развивать самостоятельность мышления, умение устанавливать причинно-следственные связи, обобщать, делать выводы; воспитывать потребность в бережном отношении к жизни.
Средства обучения: дидактические материалы
для организации самостоятельной работы
учащихся, микроскопы, готовые микропрепараты,
мультимедийный проектор.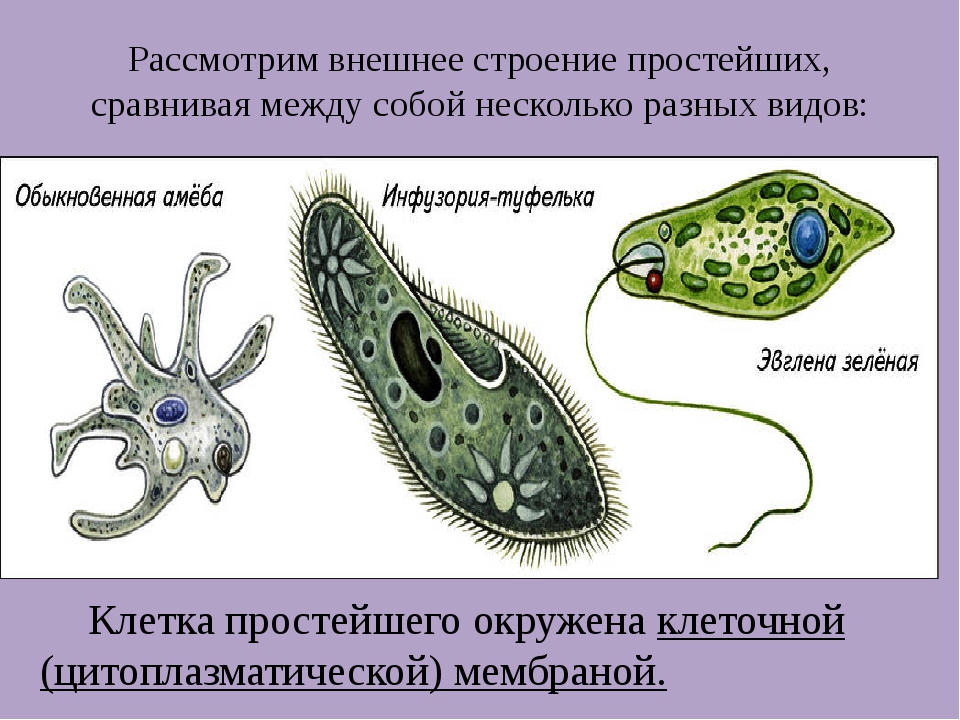
Ход мастерской
I. Актуализация знаний. Введение в тему
Учитель: Мы с вами изучили особенности подцарства Простейшие. Это особенное подцарство животных. В чем же особенности представителей этого подцарства (3 мин.)
Высказываются мнения:
- Организм простейших состоит из одной клетки.
- Клетка выполняет все функции многоклеточного организма.
- Клетка состоит из органоидов, которые выполняют определенные функции.
II. Ребята рассаживаются в группы по 5-6 человек
Вопрос учителя: Что вы знаете, а хотели бы узнать о Простейших? (3 мин.)
Учащиеся по группам на листе бумаги записывают свои вопросы. Учитель собирает листочки и раскладывает их на группы: тип Саркодовые, тип Жгутиконосцы, тип Инфузории.
III. Каждая группа получает задание на отдельном
листе бумаги. На выполнение этого задания
выделяется 10 минут.
На выполнение этого задания
выделяется 10 минут.
Задание для группы № 1
1. Каковы общие признаки простейших. Какие функции выполняют органоиды клетки Простейших?
Заполните таблицу “Функции органоидов клетки простейших”
Органоиды клетки |
Функции органоидов |
Задание для группы № 2
1. Какую роль играют простейшие в природе и жизни человека?
Паразитические одноклеточные животные.
Значение простейших
| Значение | Корненожки | Жгутиковые | Инфузории |
| Положительное | |||
| Отрицательное |
Задание для группы № 3
1. Какими способами могут размножаться
простейшие? Охарактеризуйте эти способы
размножения. Составьте схемы размножений.
Какими способами могут размножаться
простейшие? Охарактеризуйте эти способы
размножения. Составьте схемы размножений.
Задание для группы № 4
1. На примере вольвокса поясните, в чем появляется усложнение организации колониальных форм животных по сравнению с одноклеточными.
Задание для группы № 5
Составьте кроссворд к теме “Простейшие” (10-15 слов)
IV. Каждая группа вывешивает результаты работы и старший от группы выступает по результатам работы, отвечает на поставленные учащимися класса вопросы (группа помогает) – 15 мин.
V. Сообщение заранее подготовленного ученика о паразитических простейших. – 3 мин.
VI. Выполнение тестовой работы (индивидуально)
Выпишите по порядку цифры, вслед за которыми даны сведения об инфузории – туфельке (I – вариант), об амебе обыкновенной (II- вариант) – 5 мин.
1. Форма тела постоянная.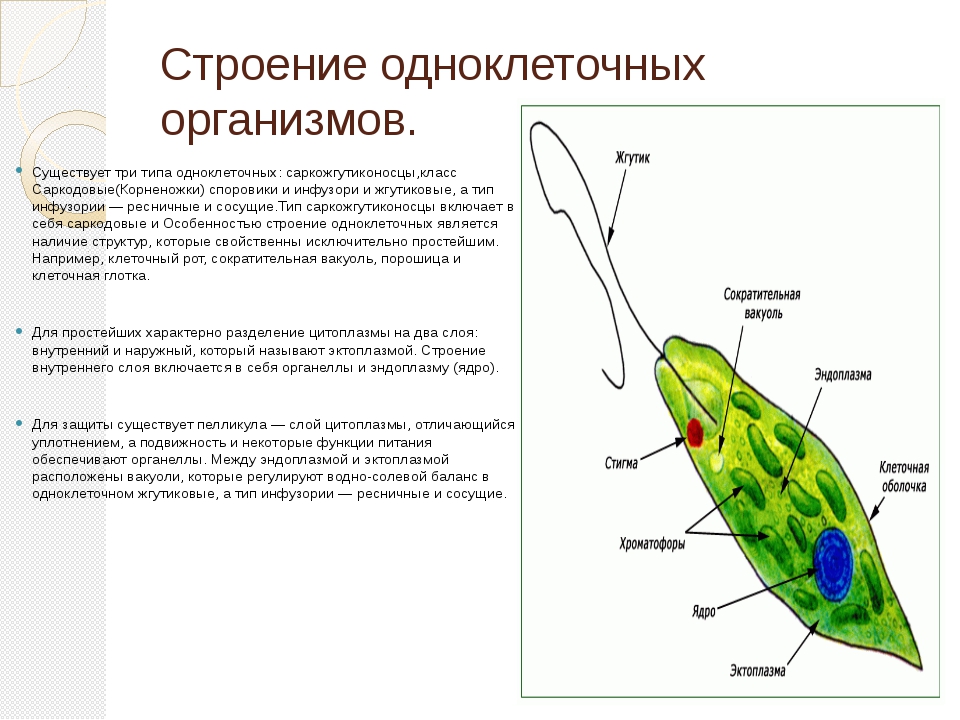
2. Передвигается при помощи многочисленных ресничек тупым концом вперед.
3. Питается бактериями, мельчайшими водорослями, простейшими.
4. Может питаться растворенными в воде органическими и неорганическими веществами. В питании принимают участие хлоропласты.
5. Жидкие продукты жизнедеятельности и избыток воды удаляются через две сократительные вакуоли.
6. Переваривание пищи происходит в пищеварительных вакуолях.
7 При неблагоприятных условиях превращается в цисту.
8. В цитоплазме одно ядро.
9. В цитоплазме два ядра — большое и малое.
VII. Учащиеся работают с готовыми микропрепаратами по заданию: “Определить вид простейшего по микропрепарату” — 3 мин.
Учитель проверяет работу учеников, консультирует их.
VIII. Подведение итогов урока, 2-3 мин.
Вопросы учащимися:
— Что нового вы узнали на уроке?
— Что нового вы узнали о себе?
— Ваши пожелания на будущее.
IX. Задание на дом (по выбору учащихся) — 2-3 мин.
- Написать сочинение на тему “Природа без простейших”. (Что изменилось бы в окружающей среде, если бы исчезли простейшие?).
- Составьте кроссворд к теме “Простейшие”.
- Составить игру “Лишнее слово”.
Дидактический материал для учителя
Задание для группы № 1
| Органоиды клетки | Функции органоидов |
| Ядро | Регулирует все процессы жизнедеятельности |
| Цитоплазма | В ней притекают все процессы обмена веществ |
| Сократительная вакуоль | Собирает вредные вещества и избыток воды, которые выталкиваются наружу |
| Цитоплазматическая мембрана | Отграничивает клетку от окружающей среды |
| Ложноножки | Органоиды передвижение амебы |
| Респики | Органоиды передвижение инфузории – туфельки |
| Жгутики | Органоиды передвижение эвглены зелено |
| Пищеварительная вакуоль | Переваривание пищи |
Задание для группы № 2
Значение простейших
| Значение | Корненожки | Жгутиковые | Инфузории |
| Положительное | Компонент биоценоза в цепи питания, морские корненожки имеют известковую раковину – образуют осадочные горные породы – мел, известняк; по некоторым видам корненожек судят о присутствии нефти | Компонент биоценоза в цепи питания, имеет познавательное значение для изучения общих признаков растений и животных | Компонент биоценоза в цепи питания |
| Отрицательное | Дизентерийная амеба вызывает
амебную дизентерию. Малярийный плазмодий вызывает малярию. |
Вызывают цветение воды в
водоемах. Трипаносомы паразитируют в крови и
спинномозговой жидкости позвоночных. Вызывают
сонную болезнь, а также нагану – болезнь
крупного рогатого скота. Лямблии паразитируют в кишечнике млекопитающих, вызывая болезнь лямблиоз. |
Паразитическая инфузория балантидий вызывает у человека тяжелые, напоминающее дизентерию расстройства. Она также может паразитировать в кишечнике свиней, которая не приносит вреда, но ими может заразиться человек. |
Задание для группы № 3
Для животных подцарства Простейшие характерны два способа размножения: бесполое и половое.
При бесполом размножении (характерно для типов
(Саркодовые, Жгутиконосцы, Инфузорий) происходит
деление клетки надвое. Сначала делится ядро
клетки, а затем на теле появляются перетяжка,
которая делит его на две примерно равные части, в
каждой из которых оказывается по одному ядру.
Сначала делится ядро
клетки, а затем на теле появляются перетяжка,
которая делит его на две примерно равные части, в
каждой из которых оказывается по одному ядру.
Для представителей типа Инфузорий характерен и половой способ размножения – конъюгация. При половом процессе увеличения числа особей не происходит. Две инфузории временно соединяются друг с другом. На месте соприкосновения пелликула растворяется, и между животными образуется соединительный мостик из цитоплазмы. Большое ядро каждой инфузории исчезает. Малое ядро дважды делится, в результате чего в каждой инфузории получается 4 дочерних ядра. Три из них размножаются, а четвертое снова делится. В результате в каждой инфузории остается по два ядра.
Одно из этих ядер по цитоплазматическому
мостику переходит в другую инфузорию и там
сливается с неподвижным ядром. Затем в каждой
инфузории из этого вновь образовавшегося ядра
формируются большое и малое ядро и инфузории
расходятся.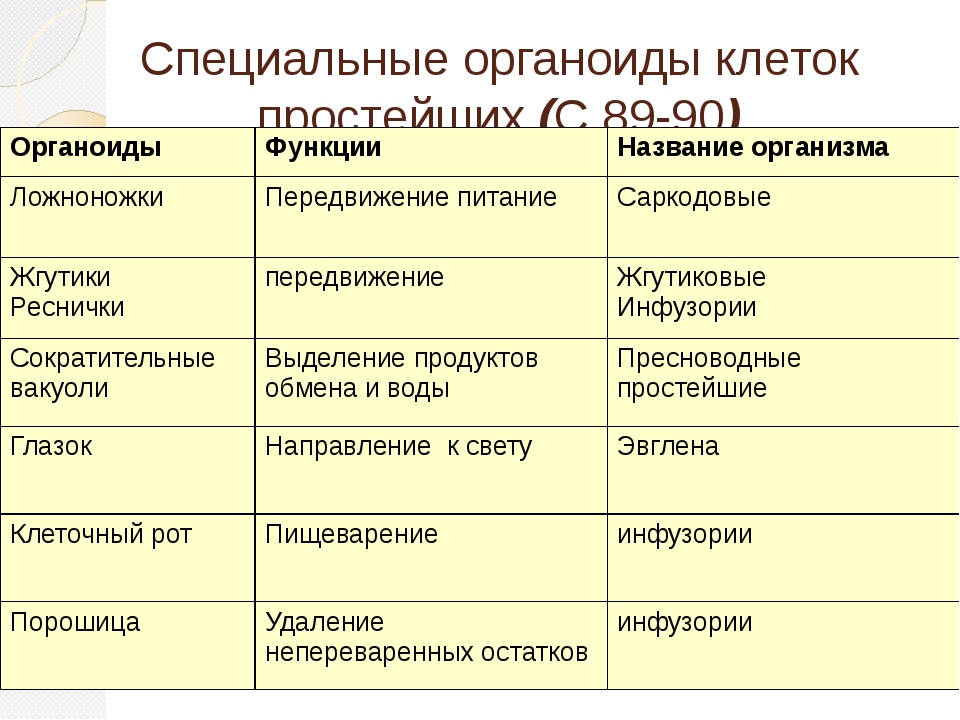 Половой процесс ведет к обновлению
генетического материала, что увеличивает
жизнестойкость организмов.
Половой процесс ведет к обновлению
генетического материала, что увеличивает
жизнестойкость организмов.
Задание для группы № 4
Вольвокс может служить моделью, показывающей, как из одноклеточных организмов могли произойти многоклеточные.
Тело колониальных жгутиконосцев состоят из многих клеток. Первые колонии возникают, вероятно, вследствие того, что после деления клетки не расходятся, а остаются вместе. Число клеток в колонии вольвокса может достигать более 60 тысяч, внутренняя полость его шара занята жидкой слизью.
В колонии вольвокса существуют различные типы клеток (отвечающие за питание, движение, размножение), что характерно для многоклеточных.
.
Задание для группы № 5
Рекомендуемые для кроссворда слова: амеба,
клетка, цитоплазма, ядро, циста, органоид,
конъюгация, колония, жгутик, ложноножка,
раздражимость, выделение, эвглена, вольвокс,
радиолярии, лучевики, пелликула, эндоплазма,
эктоплазма.
Сообщение учащегося о болезнетворных простейших
В организме человека паразитирует более 30 видов простейших.
В человеке может паразитировать несколько видов плазмодия, вызывающих разные формы малярии. Попадая в кровь человека со слюной комара из рода Anopheles, возбудитель малярии разносится по всему телу, задерживаясь в печени, а затем переселяются в эритроциты, где питается гемоглобином. Сразу во многих красных клетках крови идет размножение паразитов. Затем все они одновременно разрывают оболочку эритроцитов и выходят в кровяное русло. Это и вызывает приступ лихорадки. После того как паразиты внедряются в новые эритроциты, температура тела больного нормализуется до их следующего массового выхода.
Возбудителями ряда опасных болезней человека и
животных являются паразитические жгутиконосцы.
Один из видов трипаносом вызывает “сонную
болезнь”, от которой в Африке ежегодно умирают
тысячи людей. Природный резервуар этого паразита
– африканские антилопы, совершенно не
страдающие от присутствия в их крови
жгутиконосцев. Переносчиками трипаносом
являются кровососущие мухи цеце. Трипаносомы
проникают в кровь, лимфатические железы, а затем
и в спинномозговую жидкость. При отсутствии
лечения “сонная болезнь” всегда приводит к
смерти.
Природный резервуар этого паразита
– африканские антилопы, совершенно не
страдающие от присутствия в их крови
жгутиконосцев. Переносчиками трипаносом
являются кровососущие мухи цеце. Трипаносомы
проникают в кровь, лимфатические железы, а затем
и в спинномозговую жидкость. При отсутствии
лечения “сонная болезнь” всегда приводит к
смерти.
Ответы к тестовой работе.
I вариант (инфузория туфелька) – 2, 3, 5, 8, 9, 11, 13
II вариант (амеба обыкновенная) – 1, 4, 5, 7, 9, 11, 12.
Какие функции выполняют органеллы выделения простейших. Органеллы простейших, их строение и основные функции. Что такое ропалий
Знаете ли вы, какое строение имеет клетка простейших? Если нет, то эта статья для вас.
Какая наука изучает клетку?
Эта наука называется цитологией. Она является отраслью биологии. Она и может ответить на вопрос, какое строение имеет клетка простейших. Также данная наука изучает не только структуру, но и процессы, которые происходят в клетке. Это обмен веществ, размножение и фотосинтез. Способ размножения простейших — простое деление клетки. Некоторые клетки простейших способны осуществлять фотосинтез — выработку органических веществ из неорганических. Клеточное дыхание происходит при расщеплении глюкозы. В этом и заключается главная функция простых углеводов в клетке. При их окислении клетка получает энергию.
Это обмен веществ, размножение и фотосинтез. Способ размножения простейших — простое деление клетки. Некоторые клетки простейших способны осуществлять фотосинтез — выработку органических веществ из неорганических. Клеточное дыхание происходит при расщеплении глюкозы. В этом и заключается главная функция простых углеводов в клетке. При их окислении клетка получает энергию.
Кто такие простейшие?
Перед тем как рассматривать вопрос о том, какое строение имеет клетка простейших, давайте разберемся, что из себя представляют эти «существа».
Это организмы, которые Они называются еще эукариотами, так как в их клетках есть ядро. Клетка простейших во многом похожа на клетку многоклеточного организма.
Классификация
Существует шесть типов простейших:
- инфузории;
- радиолярии;
- солнечники;
- споровики;
- саркожгутиконосцы;
- жгутиковые.
Представители первого типа населяют соленые водоемы. Некоторые виды также могут жить в почве.
Радиолярии, как и инфузории, обитают в океанах. Они имеют твердые оболочки из диоксида кремния, из которых формируются некоторые горные породы.
Особенность солнечников заключается в том, что они передвигаются с помощью псевдоподий.
Саркожгутиконосцы также используют такой способ передвижения. К этому типу относятся амебы и многие другие простейшие.
Какое строение имеет клетка простейших?
Структуру клетки можно разделить на три основных части: плазматическую мембрану, цитоплазму и ядро. Количество ядер в клетках простейших равняется одному. Этим они отличаются от клеток бактерий, которые вообще не имеют ядер. Итак, рассмотрим детально каждый из трех компонентов клетки.
Плазматическая мембрана
Простейших обязательно предусматривает наличие этой составляющей. Она отвечает за поддержание гомеостаза клетки, защищает ее от воздействий внешней среды. Плазматическая мембрана состоит из липидов трех классов: фосфолипидов, гликолипидов и холестерола. Преобладают в структуре мембраны фосфолипиды.
Преобладают в структуре мембраны фосфолипиды.
Цитоплазма: как она устроена?
Это вся та часть клетки, за исключением ядра, которая находится внутри плазматической мембраны. Она состоит из гиалоплазмы и органоидов, а также включений. Гиалоплазма — это внутренняя среда клетки. Органоиды являются постоянными структурами, которые выполняют определенные функции, а включения —это непостоянные структуры, которые выполняют в основном запасающую функцию.
Строение клетки простейших: органоиды
В клетке простейших присутствуют многие органоиды, которые свойственны для животных клеток. Кроме того, в отличие от клеток большинство клеток простейших обладают органоидами движения — всевозможными жгутиками, ресничками и другими структурами. Наличием таких образований могут похвастаться очень немногие клетки многоклеточных животных — только сперматозоиды.
К органоидам, которые присутствуют в клетках простейших, относятся митохондрии, рибосомы, лизосомы, эндоплазматический ретикулум, комплекс Гольджи.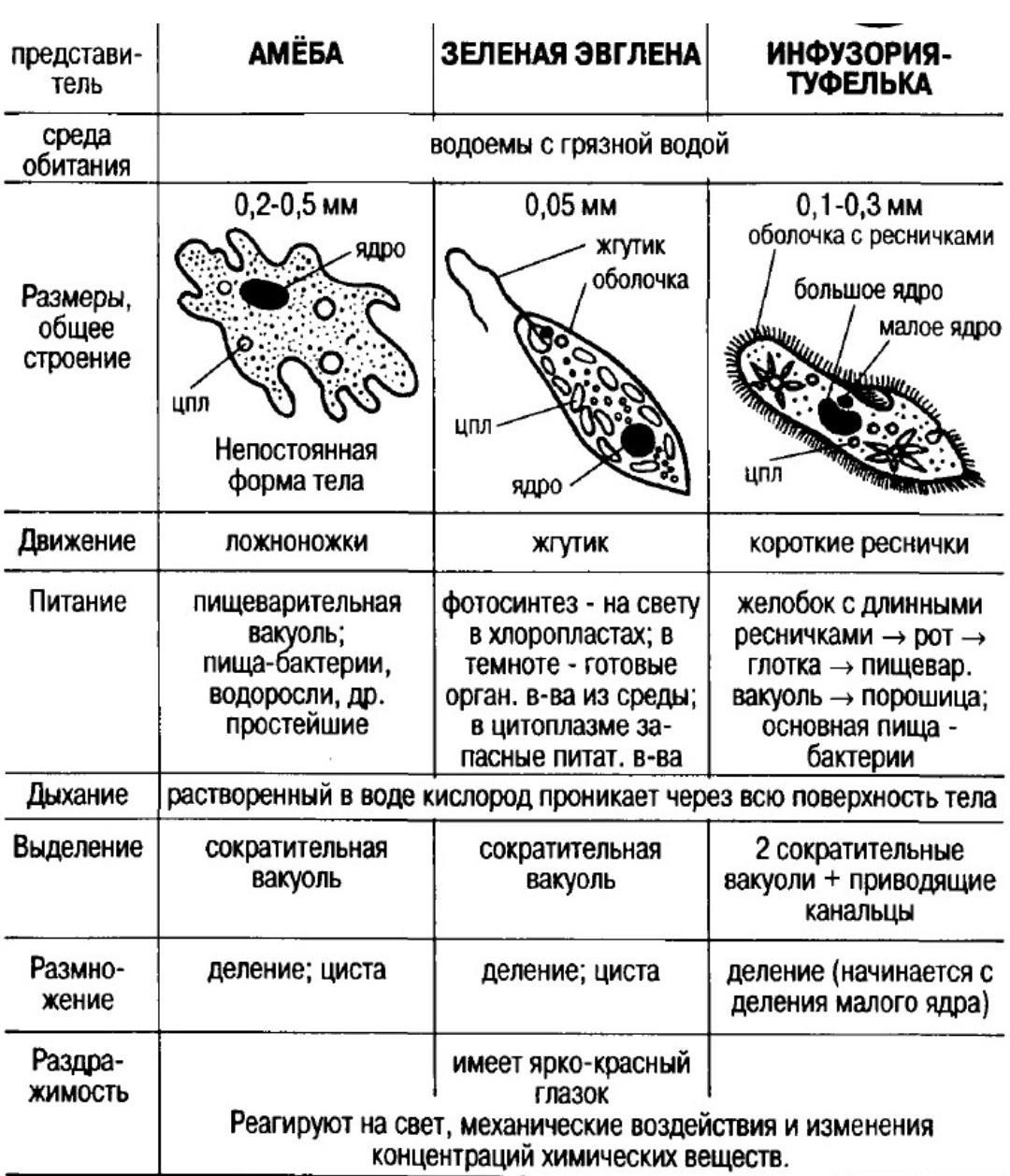 В клетках некоторых простейших также находятся хлоропласты, которые характерны для растительных клеток. Рассмотрим строение и функции каждого из них в таблице.
В клетках некоторых простейших также находятся хлоропласты, которые характерны для растительных клеток. Рассмотрим строение и функции каждого из них в таблице.
| Органоид | Строение | Функции |
| Митохондрии | Обладают двумя мембранами: внешней и внутренней, между которыми присутствует межмембранное пространство. Внутренняя мембрана обладает выростами — кристами или гребнями. На них и происходят все основные химические реакции. То, что находится внутри обоих мембран, называется матриксом. В нем у этих органоидов присутствуют свои рибосомы, включения, митохондриальная РНК и митохондриальная ДНК. | Выработка энергии. В этих органоидах происходит процесс клеточного дыхания. |
| Рибосомы | Состоят из двух субъединиц. Не имеют мембран. Одна из субъединиц обладает большим размером, чем вторая. Рибосомы объединяются только в процессе функционирования.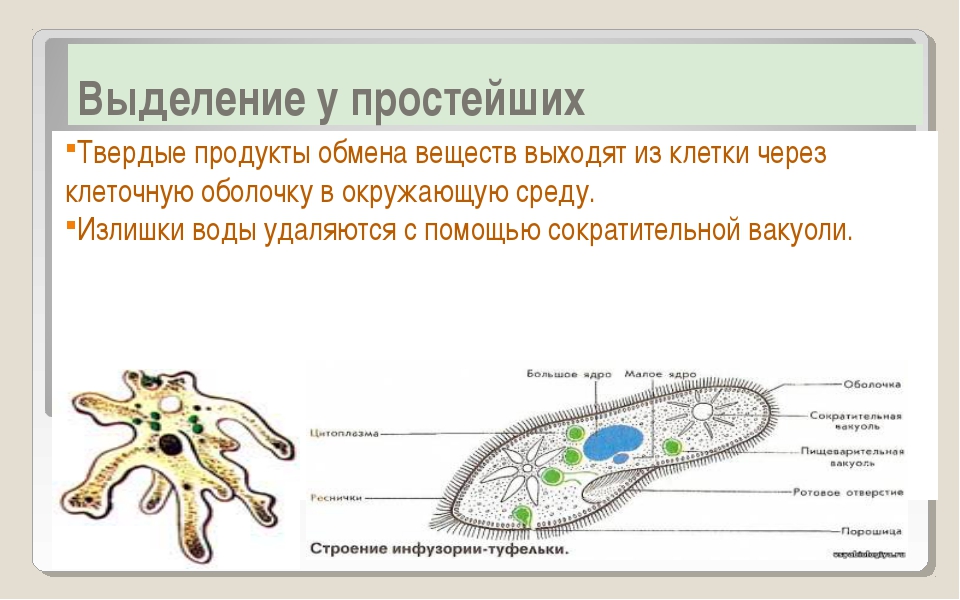 Когда органоид не функционирует, две субъединицы находятся раздельно. Когда органоид не функционирует, две субъединицы находятся раздельно. | Синтез белков (процесс трансляции). |
| Лизосомы | Обладают округлой формой. Имеют одну мембрану. Внутри мембраны находятся ферменты, которые необходимы для расщепления сложных органических веществ. | Клеточное пищеварение. |
| Эндоплазматический ретикулум | Трубчатая форма. | Участвует в обмене веществ, отвечает за синтез липидов. |
| Комплекс Гольджи | Стопка дискообразных цистерн. | Служит для синтеза гликозамингликанов, гликолипидов. Модифицирует и классифицирует белки. |
| Хлоропласты | Обладают двумя мембранами с межмембранным пространством между ними. В матриксе находятся тилакоиды, объединенные в стопки (граны ламеллами. Кроме того, в матриксе находятся рибосомы, включения, РНК и ДНК. | Фотосинтез (происходит в тилакоидах). |
| Вакуоли | Многие простейшие, населяющие пресные водоемы, обладают (шарообразными органоидами с одной мембраной) | Выкачивание из организма лишней жидкости. |
Кроме того, клетки простейших снабжены органоидами движения. Это могут быть жгутики и реснички. В зависимости от вида, организм может обладать как одним, так и несколькими жгутиками.
Органелла — это постоянное образование в клетке, которое выполняет определенные функции. Их еще называют органоидами. Органелла — это то, что позволяет клетке жить. Точно так же, как животное и человек состоят из органов, так каждая клетка состоит из органоидов. Они разнообразны и выполняют все функции, обеспечивающие жизнь клетки: это и обмен веществ, и их запасание, и деление.
Какие бывают органоиды?
Органелла — это сложная структура. Некоторые из них могут даже иметь свою собственную ДНК и РНК. Во всех клетках присутствуют митохондрии, рибосомы, лизосомы, клеточный центр, аппарат (комплекс) Гольджи, эндоплазматическая сеть (ретикулум). Растения имеют также специфические клеточные органеллы: вакуоли и пластиды. Некоторые относят к органоидам также микротрубочки и микрофиламенты.
Органелла — это и рибосома, и вакуоль, и клеточный центр, и многие другие. Давайте рассмотрим подробнее строение и функции органелл.
Митохондрии
Эти органоиды обеспечивают клетку энергией — они отвечают за Они есть и у растений, и у животных, и у грибов. Данные клеточные органеллы имеют две мембраны: внешнюю и внутреннюю, между которыми есть межмембранное пространство. То, что находится внутри оболочек, называется матриксом. В нем находятся разнообразные ферменты — вещества, необходимые для ускорения химических реакций. Внутренняя мембрана обладает складками — кристами. Именно на них и происходит процесс клеточного дыхания. Кроме того, в матриксе митохондрий находится митохондриальная ДНК (мДНК) и мРНК, а также рибосомы, практически аналогичные тем, которыми обладают
Рибосома
Этот органоид отвечает за процесс трансляции, при котором из отдельных аминокислот синтезируется белок. Строение органеллы рибосомы проще, чем митохондрии, — она не обладает мембранами. Данный органоид состоит из двух частей (субъединиц) — малой и большой. Когда рибосома бездействует, они находятся раздельно, а когда она начинает синтезировать белок — объединяются. Также собираться вместе могут и несколько рибосом, если полипептидная цепочка, синтезируемая ими, очень длинная. Такая структура называется «полирибосома».
Данный органоид состоит из двух частей (субъединиц) — малой и большой. Когда рибосома бездействует, они находятся раздельно, а когда она начинает синтезировать белок — объединяются. Также собираться вместе могут и несколько рибосом, если полипептидная цепочка, синтезируемая ими, очень длинная. Такая структура называется «полирибосома».
Лизосомы
Функции органелл этого вида сводятся к осуществлению клеточного пищеварения. Лизосомы обладают одной мембраной, внутри которой находятся ферменты — катализаторы химических реакций. Иногда эти органоиды не только расщепляют но и переваривают целые органоиды. Такое может происходить при длительной голодовке клетки и позволяет ей жить еще некоторое время. Хотя если питательные вещества все еще не начнут поступать, клетка умирает.
и функции
Эта органелла состоит из двух частей — центриолей. Это образования в форме цилиндров, состоящие из микротрубочек. Клеточный центр — очень важный органоид. Он участвует в процессе формирования веретена деления.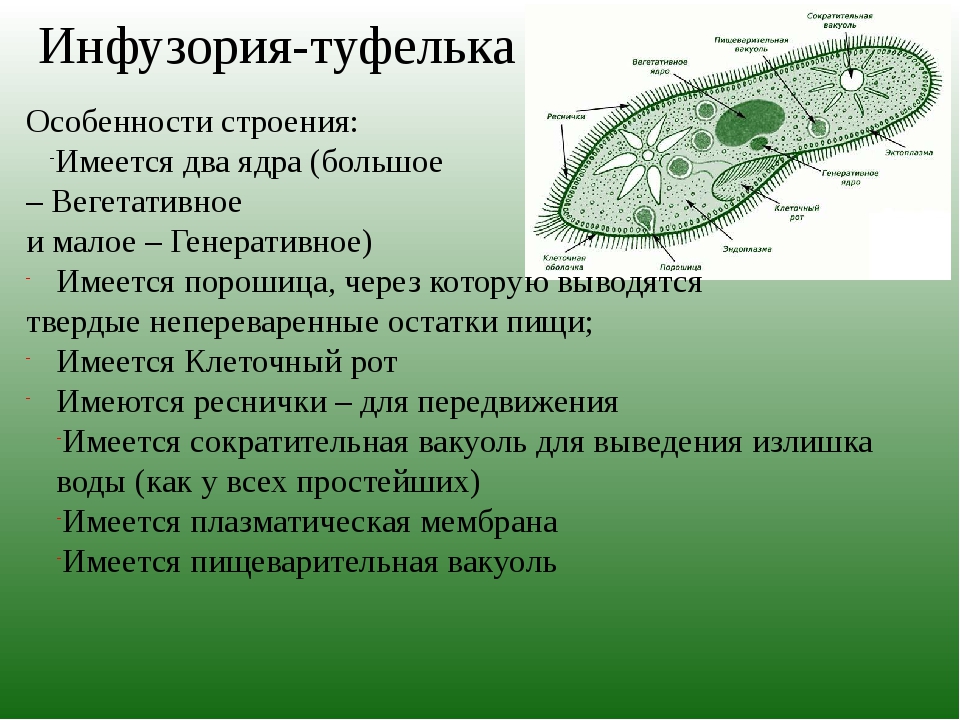 Кроме того, он является центром организации микротрубочек.
Кроме того, он является центром организации микротрубочек.
Аппарат Гольджи
Это комплекс дискообразных мембранных мешочков, называемых цистернами. Функции этого органоида заключаются в сортировке, запасании и превращении некоторых веществ. Синтезируются здесь в основном углеводы, которые входят в состав гликокаликса.
Строение и функции эндоплазматического ретикулума
Это сеть трубочек и карманов, окруженных одной мембраной. Существует два вида эндоплазматического ретикулума: гладкий и шероховатый. На поверхности последнего расположены рибосомы. Гладкий и шероховатый ретикулумы выполняют различные функции. Первый отвечает за синтез гормонов, хранение и преобразование углеводов. Кроме того, в нем формируются зачатки вакуолей — органоидов, характерных для растительных клеток. Шероховатый эндоплазматический ретикулум содержит на своей поверхности рибосомы, которые производят полипептидную цепочку из аминокислот. Дальше она попадает в эндоплазматическую сеть, и здесь формируется определенная вторичная, третичная и четвертичная структура белка (цепочка правильным образом закручивается).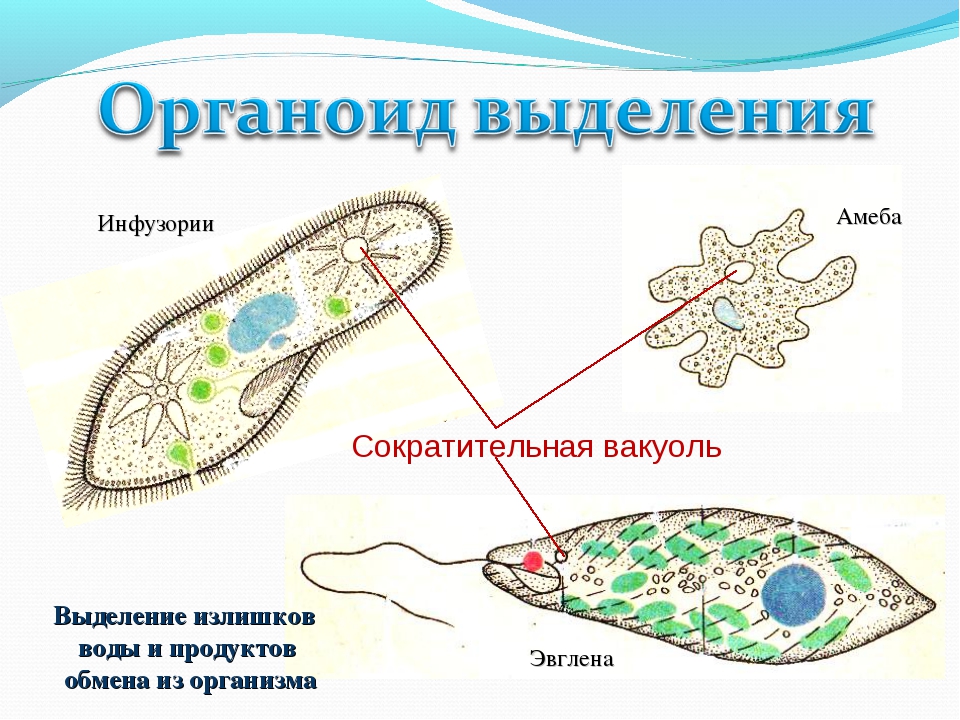
Вакуоли
Это органеллы Они обладают одной мембраной. В них накапливается клеточный сок. Вакуоль необходима для поддержания тургора. Также она участвует в процессе осмоса. Кроме того, существуют Они содержатся в основном в одноклеточных организмах, живущих в водоемах, и служат в качестве насосов, выкачивающих из клетки лишнюю жидкость.
Пластиды: разновидности, строение и функции
Это также органеллы Они бывают трех видов: лейкопласты, хромопласты и хлоропласты. Первые служат для хранения запасных питательных веществ, в основном это крахмал. Хромопласты содержат в себе различные пигменты. Благодаря им лепестки растений разноцветные. Это нужно организму в первую очередь для того, чтобы привлекать насекомых-опылителей.
Хлоропласты — самые важные пластиды. Самое большое их количество находится в листьях и стеблях растений. Они отвечают за фотосинтез — цепь химических реакций, в процессе которых из неорганических веществ организм получает органические. Эти органоиды обладают двумя мембранами.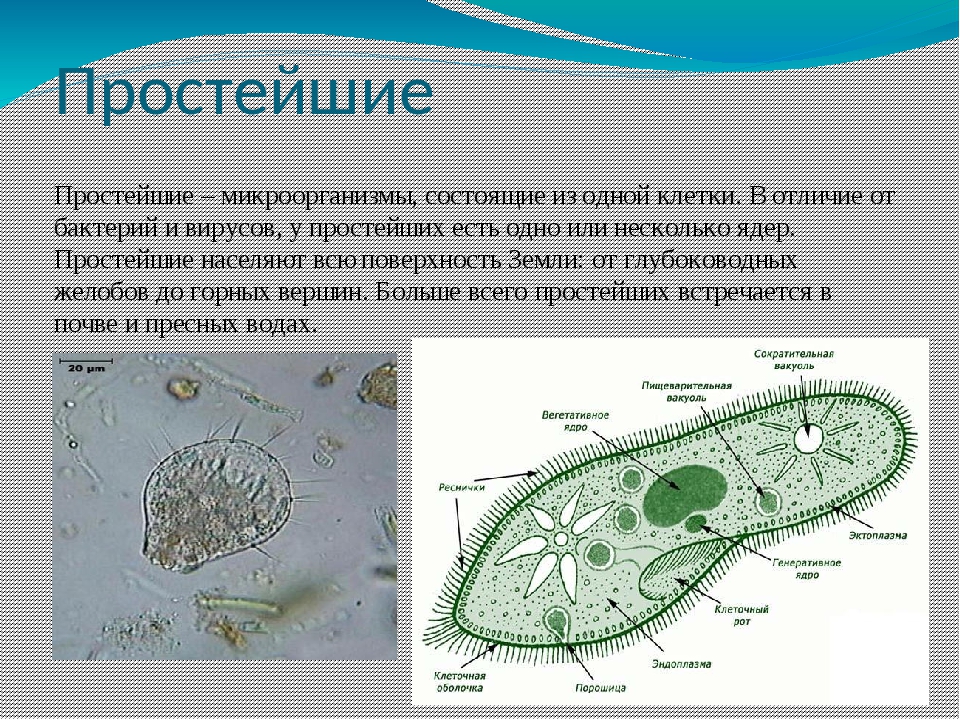 Матрикс хлоропластов называется «строма». В ней находятся пластидная ДНК, РНК, ферменты, а также крахмальные включения. В хлоропластах находятся тилакоиды — мембранные образования в виде монеты. Внутри их и происходит фотосинтез. Здесь же содержится и хлорофилл, служащий катализатором для химических реакций. Тилакоиды хлоропластов объединяются в стопки — граны. Также в органоидах находятся ламеллы, которые соединяют между собой отдельные тилакоиды и обеспечивают связь между ними.
Матрикс хлоропластов называется «строма». В ней находятся пластидная ДНК, РНК, ферменты, а также крахмальные включения. В хлоропластах находятся тилакоиды — мембранные образования в виде монеты. Внутри их и происходит фотосинтез. Здесь же содержится и хлорофилл, служащий катализатором для химических реакций. Тилакоиды хлоропластов объединяются в стопки — граны. Также в органоидах находятся ламеллы, которые соединяют между собой отдельные тилакоиды и обеспечивают связь между ними.
Органеллы движения
Они характерны в основном для одноклеточных организмов. К ним относятся жгутики и реснички. Первые присутствуют у эвглен, трипаносом, хламидомонад. Также жгутики присутствуют у сперматозоидов животных. Реснички есть у инфузорий и других одноклеточных.
Микротрубочки
Они обеспечивают транспорт веществ, а также постоянную форму клетки. Некоторые ученые не относят микротрубочки к органеллам.
Делит все клетки (или живые организмы ) на два типа: прокариоты и эукариоты . Прокариоты — это безъядерные клетки или организмы, к которым относятся вирусы, прокариот-бактерии и сине-зеленые водоросли, у которых клетка состоит непосредственно из цитоплазмы, в которой расположена одна хромосома — молекула ДНК (иногда РНК).
Прокариоты — это безъядерные клетки или организмы, к которым относятся вирусы, прокариот-бактерии и сине-зеленые водоросли, у которых клетка состоит непосредственно из цитоплазмы, в которой расположена одна хромосома — молекула ДНК (иногда РНК).
Эукариотические клетки имеют ядро , в котором находятся нуклеопротеиды (белок гистон + комплекс ДНК), а также другие органоиды . К эукариотам относятся большинство современных известных науке одноклеточных и многоклеточных живых организмов (в том числе, и растений).
Строение ограноидов эукариотов.
Название органоида | Строение органоида | Функции органоида |
|---|---|---|
Цитоплазма | Внутренняя среда клетки, в которой находится ядро и другие органоиды. Имеет полужидкую, мелкозернистую структуру. |
|
Рибосомы | Мелкие органоиды сферической или эллипсоидной формы диаметром от 15 до 30 нанометров. | Обеспечивают процесс синтеза молекул белка, их сборку из аминокислот. |
Митохондрии | Органоиды, имеющие самую разнообразную форму — от сферической до нитевидной. Внутри митохондрий имеются складки от 0,2 до 0,7 мкм. Внешняя оболочка митохондрий имеет двухмембранную структуру. Наружная мембрана гладкая, а на внутренней имеются выросты крестообразной формы с дыхательными ферментами. |
|
Эндоплазматическая сеть (ЭПС) | Система оболочек в цитоплазме, которая образует каналы и полости. |
|
Пластиды (органоиды, свойственные только растительным клеткам) бывают трех видов: | Двухмембранные органоиды | |
Лейкопласты | Бесцветные пластиды, которые содержатся в клубнях, корнях и луковицах растений. | Являются дополнительным резервуаром для хранения питательных веществ. |
Хлоропласты | Органоиды овальной формы, имеющие зеленый цвет. От цитоплазмы отделяются двумя трехслойными мембранами. Внутри хлоропластов находится хлорофилл. | Преобразуют органические вещества из неорганических, используя энергию солнца. |
Хромопласты | Органоиды, от желтого до бурого цвета, в которых накапливается каротин. | Способствуют появлению у растений частей с желтой, оранжевой и красной окраской. |
Лизосомы | Органоиды округлой формы диаметром около 1 мкм, имеющие на поверхности мембрану, а внутри — комплекс ферментов. | Пищеварительная функция. Переваривают питательные частицы и ликвидируют отмершие части клетки. |
Комплекс Гольджи | Может быть разной формы. Состоит из полостей, разграниченных мембранами. Из полостей отходят трубчатые образования с пузырьками на концах. |
|
Клеточный центр | Состоит из центросферы (уплотненного участка цитоплазмы) и центриолей — двух маленьких телец. | Выполняет важную функцию для деления клетки. |
Клеточные включения | Углеводы, жиры и белки, которые являются непостоянными компонентами клетки. | Запасные питательные вещества, которые используются для жизнедеятельности клетки. |
Органоиды движения | Жгутики и реснички (выросты и клетки), миофибриллы (нитевидные образования) и псевдоподии (или ложноножки). | Выполняют двигательную функцию, а также обеспечивают процесс сокращения мышц. |

 Бывает двух типов: гранулированная, на которой имеются рибосомы и гладкая.
Бывает двух типов: гранулированная, на которой имеются рибосомы и гладкая.
Ядро клетки является главным и самым сложным органоидом клетки, поэтому его мы рассмотрим
Простейшие — одноклеточные животные, и морфологически тело их соответствует одной клетке многоклеточного организма. Физиологически же каждая особь простейших представляет самостоятельный организм в отличие от клетки, являющейся частью многоклеточного организма.
Каждому простейшему животному присущи все основные жизненные функции, характерные для целого организма: обмен веществ с ассимиляцией и диссимиляцией, раздражимость, движение, размножение, приспособление к среде и т. д. В теле простейших дифференцируются отдельные клеточные участки, выполняющие различные жизненные функции и в этом смысле аналогичные органам многоклеточных животных. Но так как эти приспособления не имеют клеточного строения и не состоят из тканей, они называются не органами, а органеллами, или органоидами.
Органеллами движения служат либо ложноножки (псевдоподии), представляющие собой временные выступы протоплазмы, возникающие по мере необходимости в различных местах тела, либо постоянно существующие образования в виде длинных и немногочисленных жгутов или же коротких многочисленных ресничек. У некоторых форм под оболочкой расположены сократительные тяжи (мионемы), у ряда простейших имеется сложно устроенная система скелетных образований.
Органеллами питания служат у некоторых простейших клеточный рот, различно устроенная глотка, пищеварительные вакуоли. У многих форм имеются сократительные (пульсирующие) вакуоли, служащие для экскреции и регулирования осмотического давления внутри организма.
У многих форм имеются сократительные (пульсирующие) вакуоли, служащие для экскреции и регулирования осмотического давления внутри организма.
Организация простейших может в связи с наличием тех или других органелл достигать значительной сложности (особенно у некоторых инфузорий).
Споровики: кокцидии, малярийный плазмодий.
Инфузории: балантидии.
Саркодовые: амеба дизентерийная.
Простейшие — древнейшая группа живых организмов. Когда возникли первые простейшие, неизвестно. В западной литературе простейших не рассматривают в качестве животных и относят к царству протистов. А согласно новейшим системам среди простейших выделяют несколько царств.
Каждый жгутик снаружи покрыт трехслойной цитоплазматической
мембраной. Внутри жгутика имеются фибриллы: две центральные
и девять двойных периферических. Жгутик крепится в
цитоплазме при помощи базального тельца — кинетосомы. Обычно
жгутики производят вращающее движение, а реснички — гребное.
Жгутики свойственны жгутиконосцам, а реснички — инфузориям.
Некоторые простейшие способны к быстрому сокращению тела за
счет особых сократительных фибрилл — мионем. Например,
сидячие инфузории-сувойки способны резко сокращать свой
длинный стебелек и сворачивать его в спираль. Радиолярии
способны то растягивать тело клетки на радиальных иглах, то
сокращать его за счет сократительных волокон. Это
обеспечивает им регуляцию свободного плавания в толще воды.
ПОДЦАРСТВО ПРОСТЕЙШИЕ, ИЛИ ОДНОКЛЕТОЧНЫЕ (PROTOZOA)
Двигательные органеллы. Наиболее примитивным способом движения у простейших можно
считать амебоидное движение при помощи ложных ножек, или
псевдоподий. При этом образуются особые выступы клетки, в
которые перетекает цитоплазма. Такие органеллы движения
присущи одноклеточным с непостоянной формой тела.
Более сложное движение свойственно простейшим, имеющим в
качестве органелл движения жгутики или реснички. Строение
жгутика и ресничек сходно.
Основные органеллы простейших. Органоиды клетки и их функции Какие органеллы выполняют дыхательную функцию у простейших
Знаете ли вы, какое строение имеет клетка простейших? Если нет, то эта статья для вас.
Какая наука изучает клетку?
Эта наука называется цитологией. Она является отраслью биологии. Она и может ответить на вопрос, какое строение имеет клетка простейших. Также данная наука изучает не только структуру, но и процессы, которые происходят в клетке. Это обмен веществ, размножение и фотосинтез. Способ размножения простейших — простое деление клетки. Некоторые клетки простейших способны осуществлять фотосинтез — выработку органических веществ из неорганических. Клеточное дыхание происходит при расщеплении глюкозы. В этом и заключается главная функция простых углеводов в клетке. При их окислении клетка получает энергию.
Кто такие простейшие?
Перед тем как рассматривать вопрос о том, какое строение имеет клетка простейших, давайте разберемся, что из себя представляют эти «существа».
Это организмы, которые Они называются еще эукариотами, так как в их клетках есть ядро. Клетка простейших во многом похожа на клетку многоклеточного организма.
Классификация
Существует шесть типов простейших:
- инфузории;
- радиолярии;
- солнечники;
- споровики;
- саркожгутиконосцы;
- жгутиковые.

Представители первого типа населяют соленые водоемы. Некоторые виды также могут жить в почве.
Радиолярии, как и инфузории, обитают в океанах. Они имеют твердые оболочки из диоксида кремния, из которых формируются некоторые горные породы.
Особенность солнечников заключается в том, что они передвигаются с помощью псевдоподий.
Саркожгутиконосцы также используют такой способ передвижения. К этому типу относятся амебы и многие другие простейшие.
Какое строение имеет клетка простейших?
Структуру клетки можно разделить на три основных части: плазматическую мембрану, цитоплазму и ядро. Количество ядер в клетках простейших равняется одному. Этим они отличаются от клеток бактерий, которые вообще не имеют ядер. Итак, рассмотрим детально каждый из трех компонентов клетки.
Плазматическая мембрана
Простейших обязательно предусматривает наличие этой составляющей. Она отвечает за поддержание гомеостаза клетки, защищает ее от воздействий внешней среды.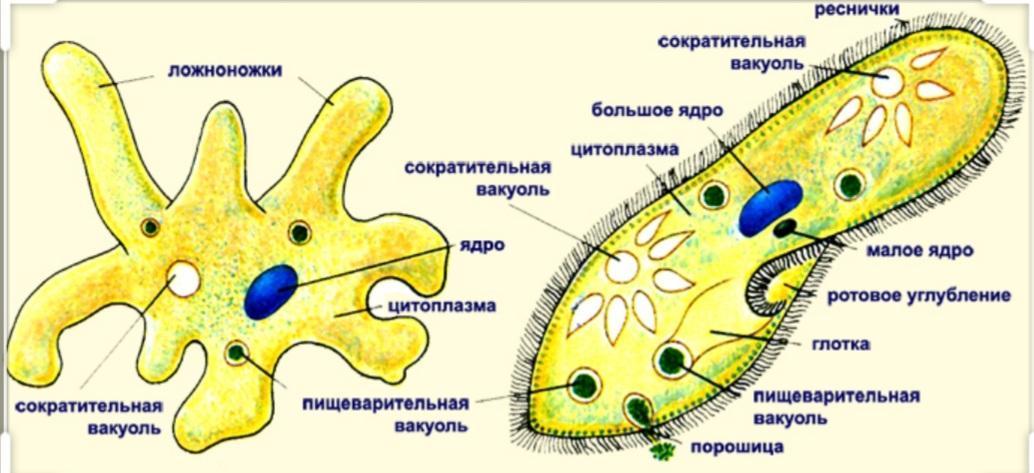 Плазматическая мембрана состоит из липидов трех классов: фосфолипидов, гликолипидов и холестерола. Преобладают в структуре мембраны фосфолипиды.
Плазматическая мембрана состоит из липидов трех классов: фосфолипидов, гликолипидов и холестерола. Преобладают в структуре мембраны фосфолипиды.
Цитоплазма: как она устроена?
Это вся та часть клетки, за исключением ядра, которая находится внутри плазматической мембраны. Она состоит из гиалоплазмы и органоидов, а также включений. Гиалоплазма — это внутренняя среда клетки. Органоиды являются постоянными структурами, которые выполняют определенные функции, а включения —это непостоянные структуры, которые выполняют в основном запасающую функцию.
Строение клетки простейших: органоиды
В клетке простейших присутствуют многие органоиды, которые свойственны для животных клеток. Кроме того, в отличие от клеток большинство клеток простейших обладают органоидами движения — всевозможными жгутиками, ресничками и другими структурами. Наличием таких образований могут похвастаться очень немногие клетки многоклеточных животных — только сперматозоиды.
К органоидам, которые присутствуют в клетках простейших, относятся митохондрии, рибосомы, лизосомы, эндоплазматический ретикулум, комплекс Гольджи. В клетках некоторых простейших также находятся хлоропласты, которые характерны для растительных клеток. Рассмотрим строение и функции каждого из них в таблице.
В клетках некоторых простейших также находятся хлоропласты, которые характерны для растительных клеток. Рассмотрим строение и функции каждого из них в таблице.
| Органоид | Строение | Функции |
| Митохондрии | Обладают двумя мембранами: внешней и внутренней, между которыми присутствует межмембранное пространство. Внутренняя мембрана обладает выростами — кристами или гребнями. На них и происходят все основные химические реакции. То, что находится внутри обоих мембран, называется матриксом. В нем у этих органоидов присутствуют свои рибосомы, включения, митохондриальная РНК и митохондриальная ДНК. | Выработка энергии. В этих органоидах происходит процесс клеточного дыхания. |
| Рибосомы | Состоят из двух субъединиц. Не имеют мембран. Одна из субъединиц обладает большим размером, чем вторая. Рибосомы объединяются только в процессе функционирования. Когда органоид не функционирует, две субъединицы находятся раздельно. Когда органоид не функционирует, две субъединицы находятся раздельно. | Синтез белков (процесс трансляции). |
| Лизосомы | Обладают округлой формой. Имеют одну мембрану. Внутри мембраны находятся ферменты, которые необходимы для расщепления сложных органических веществ. | Клеточное пищеварение. |
| Эндоплазматический ретикулум | Трубчатая форма. | Участвует в обмене веществ, отвечает за синтез липидов. |
| Комплекс Гольджи | Стопка дискообразных цистерн. | Служит для синтеза гликозамингликанов, гликолипидов. Модифицирует и классифицирует белки. |
| Хлоропласты | Обладают двумя мембранами с межмембранным пространством между ними. В матриксе находятся тилакоиды, объединенные в стопки (граны ламеллами. Кроме того, в матриксе находятся рибосомы, включения, РНК и ДНК. | Фотосинтез (происходит в тилакоидах). |
| Вакуоли | Многие простейшие, населяющие пресные водоемы, обладают (шарообразными органоидами с одной мембраной) | Выкачивание из организма лишней жидкости.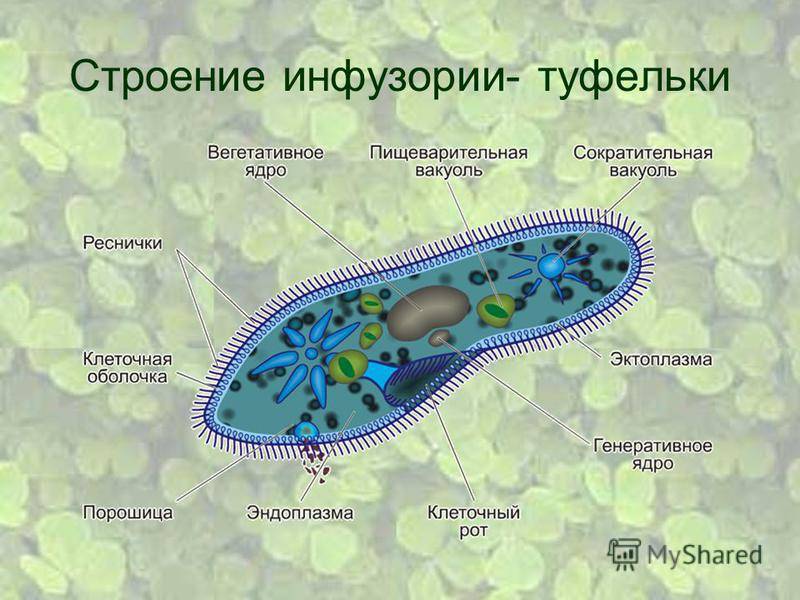 |
Кроме того, клетки простейших снабжены органоидами движения. Это могут быть жгутики и реснички. В зависимости от вида, организм может обладать как одним, так и несколькими жгутиками.
Органелла — это постоянное образование в клетке, которое выполняет определенные функции. Их еще называют органоидами. Органелла — это то, что позволяет клетке жить. Точно так же, как животное и человек состоят из органов, так каждая клетка состоит из органоидов. Они разнообразны и выполняют все функции, обеспечивающие жизнь клетки: это и обмен веществ, и их запасание, и деление.
Какие бывают органоиды?
Органелла — это сложная структура. Некоторые из них могут даже иметь свою собственную ДНК и РНК. Во всех клетках присутствуют митохондрии, рибосомы, лизосомы, клеточный центр, аппарат (комплекс) Гольджи, эндоплазматическая сеть (ретикулум). Растения имеют также специфические клеточные органеллы: вакуоли и пластиды. Некоторые относят к органоидам также микротрубочки и микрофиламенты.
Органелла — это и рибосома, и вакуоль, и клеточный центр, и многие другие. Давайте рассмотрим подробнее строение и функции органелл.
Митохондрии
Эти органоиды обеспечивают клетку энергией — они отвечают за Они есть и у растений, и у животных, и у грибов. Данные клеточные органеллы имеют две мембраны: внешнюю и внутреннюю, между которыми есть межмембранное пространство. То, что находится внутри оболочек, называется матриксом. В нем находятся разнообразные ферменты — вещества, необходимые для ускорения химических реакций. Внутренняя мембрана обладает складками — кристами. Именно на них и происходит процесс клеточного дыхания. Кроме того, в матриксе митохондрий находится митохондриальная ДНК (мДНК) и мРНК, а также рибосомы, практически аналогичные тем, которыми обладают
Рибосома
Этот органоид отвечает за процесс трансляции, при котором из отдельных аминокислот синтезируется белок. Строение органеллы рибосомы проще, чем митохондрии, — она не обладает мембранами.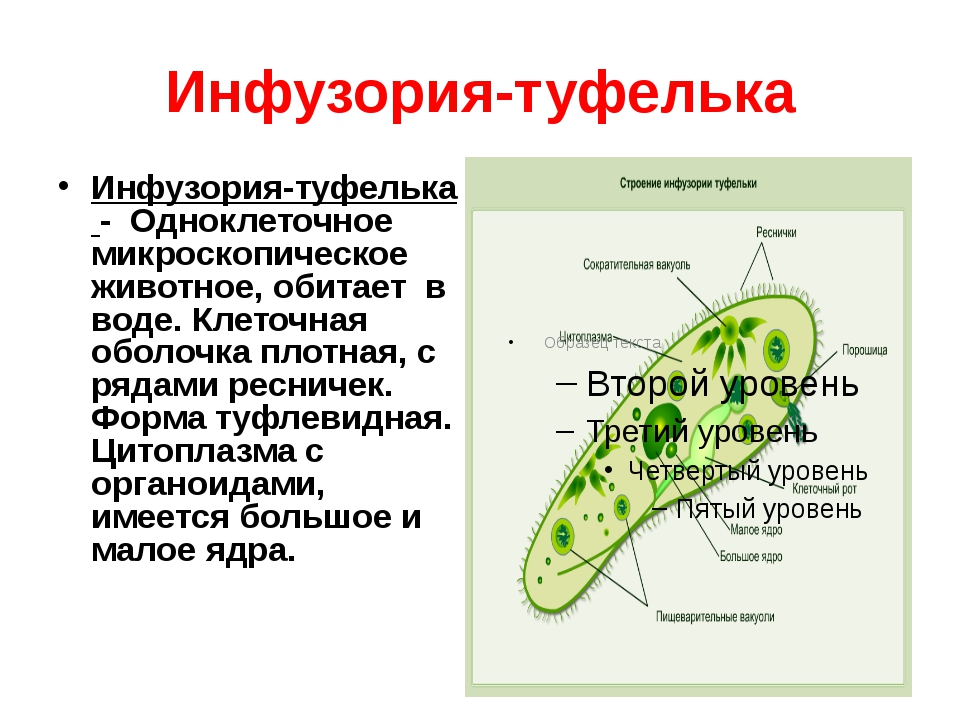 Данный органоид состоит из двух частей (субъединиц) — малой и большой. Когда рибосома бездействует, они находятся раздельно, а когда она начинает синтезировать белок — объединяются. Также собираться вместе могут и несколько рибосом, если полипептидная цепочка, синтезируемая ими, очень длинная. Такая структура называется «полирибосома».
Данный органоид состоит из двух частей (субъединиц) — малой и большой. Когда рибосома бездействует, они находятся раздельно, а когда она начинает синтезировать белок — объединяются. Также собираться вместе могут и несколько рибосом, если полипептидная цепочка, синтезируемая ими, очень длинная. Такая структура называется «полирибосома».
Лизосомы
Функции органелл этого вида сводятся к осуществлению клеточного пищеварения. Лизосомы обладают одной мембраной, внутри которой находятся ферменты — катализаторы химических реакций. Иногда эти органоиды не только расщепляют но и переваривают целые органоиды. Такое может происходить при длительной голодовке клетки и позволяет ей жить еще некоторое время. Хотя если питательные вещества все еще не начнут поступать, клетка умирает.
и функции
Эта органелла состоит из двух частей — центриолей. Это образования в форме цилиндров, состоящие из микротрубочек. Клеточный центр — очень важный органоид. Он участвует в процессе формирования веретена деления. Кроме того, он является центром организации микротрубочек.
Кроме того, он является центром организации микротрубочек.
Аппарат Гольджи
Это комплекс дискообразных мембранных мешочков, называемых цистернами. Функции этого органоида заключаются в сортировке, запасании и превращении некоторых веществ. Синтезируются здесь в основном углеводы, которые входят в состав гликокаликса.
Строение и функции эндоплазматического ретикулума
Это сеть трубочек и карманов, окруженных одной мембраной. Существует два вида эндоплазматического ретикулума: гладкий и шероховатый. На поверхности последнего расположены рибосомы. Гладкий и шероховатый ретикулумы выполняют различные функции. Первый отвечает за синтез гормонов, хранение и преобразование углеводов. Кроме того, в нем формируются зачатки вакуолей — органоидов, характерных для растительных клеток. Шероховатый эндоплазматический ретикулум содержит на своей поверхности рибосомы, которые производят полипептидную цепочку из аминокислот. Дальше она попадает в эндоплазматическую сеть, и здесь формируется определенная вторичная, третичная и четвертичная структура белка (цепочка правильным образом закручивается).
Вакуоли
Это органеллы Они обладают одной мембраной. В них накапливается клеточный сок. Вакуоль необходима для поддержания тургора. Также она участвует в процессе осмоса. Кроме того, существуют Они содержатся в основном в одноклеточных организмах, живущих в водоемах, и служат в качестве насосов, выкачивающих из клетки лишнюю жидкость.
Пластиды: разновидности, строение и функции
Это также органеллы Они бывают трех видов: лейкопласты, хромопласты и хлоропласты. Первые служат для хранения запасных питательных веществ, в основном это крахмал. Хромопласты содержат в себе различные пигменты. Благодаря им лепестки растений разноцветные. Это нужно организму в первую очередь для того, чтобы привлекать насекомых-опылителей.
Хлоропласты — самые важные пластиды. Самое большое их количество находится в листьях и стеблях растений. Они отвечают за фотосинтез — цепь химических реакций, в процессе которых из неорганических веществ организм получает органические. Эти органоиды обладают двумя мембранами. Матрикс хлоропластов называется «строма». В ней находятся пластидная ДНК, РНК, ферменты, а также крахмальные включения. В хлоропластах находятся тилакоиды — мембранные образования в виде монеты. Внутри их и происходит фотосинтез. Здесь же содержится и хлорофилл, служащий катализатором для химических реакций. Тилакоиды хлоропластов объединяются в стопки — граны. Также в органоидах находятся ламеллы, которые соединяют между собой отдельные тилакоиды и обеспечивают связь между ними.
Матрикс хлоропластов называется «строма». В ней находятся пластидная ДНК, РНК, ферменты, а также крахмальные включения. В хлоропластах находятся тилакоиды — мембранные образования в виде монеты. Внутри их и происходит фотосинтез. Здесь же содержится и хлорофилл, служащий катализатором для химических реакций. Тилакоиды хлоропластов объединяются в стопки — граны. Также в органоидах находятся ламеллы, которые соединяют между собой отдельные тилакоиды и обеспечивают связь между ними.
Органеллы движения
Они характерны в основном для одноклеточных организмов. К ним относятся жгутики и реснички. Первые присутствуют у эвглен, трипаносом, хламидомонад. Также жгутики присутствуют у сперматозоидов животных. Реснички есть у инфузорий и других одноклеточных.
Микротрубочки
Они обеспечивают транспорт веществ, а также постоянную форму клетки. Некоторые ученые не относят микротрубочки к органеллам.
Тип простейшие включает примерно 25 тыс. видов одноклеточных животных, обитающих в воде, почве или организмах других животных и человека.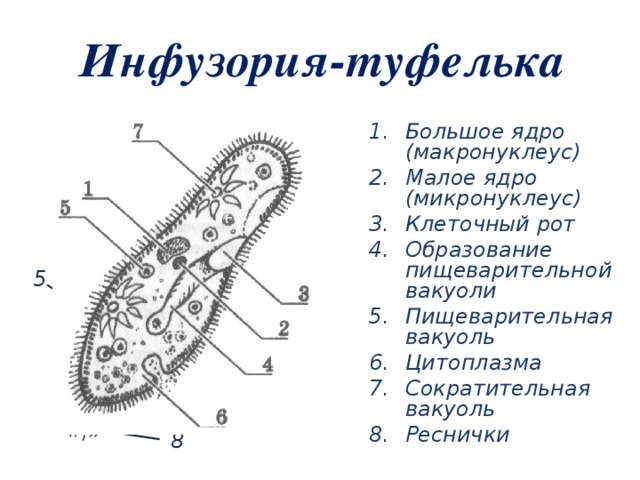 Имея морфологическое сходство в строении клеток с многоклеточными организмами, простейшие существенно отличаются от них в функциональном отношении.
Имея морфологическое сходство в строении клеток с многоклеточными организмами, простейшие существенно отличаются от них в функциональном отношении.
Если клетки многоклеточного животного выполняют специальные функции, то клетка простейшего является самостоятельным организмом, способным к обмену веществ, раздражимости, движению и размножению.
Простейшие — это организмы на клеточном уровне организации. В морфологическом отношении простейшее равноценно клетке, но в физиологическом представляет собой целый самостоятельный организм. Подавляющее большинство их — микроскопически малых размеров (от 2 до 150 мкм). Однако некоторые из ныне живущих простейших достигают 1см, а раковины ряда ископаемых корненожек имеют в диаметре до 5-6 см. Общее количество известных видов превышает 25 тыс.
Строение простейших чрезвычайно разнообразно, но все они обладают чертами, характерными для организации и функции клетки. Общим в строении в строении простейших являются два основных компонента тела — цитоплазма и ядро.
Цитаплазма
Цитоплазма ограничена наружной мембраной, которая регулирует поступление веществ в клетку. У многих простейших она усложняется дополнительными структурами, увеличивающими толщину и механическую прочность наружного слоя. Таким образом возникают образования типа пелликулы и оболочки.
Цитоплазма простейших обычно распадается на 2 слоя — наружный более светлый и плотный — эктоплазму и внутренний, снабженный многочисленными включениями,- эндоплазму.
В цитоплазме локализуются общеклеточные органоиды. Кроме того, в цитоплазме многих простейших могут присутствовать разнообразные специальные органеллы. Особенно широко распространены различные фибриллярные образования — опорные и сократимые волоконца, сократительные вакуоли, пищеварительные вакуоли и др.
Ядро
Простейшие обладают типичным клеточным ядром, одним или несколькими. Ядро простейших имеет типичную двухслойную ядерную оболочку. В ядре распределен хроматиновый материал и ядрышки.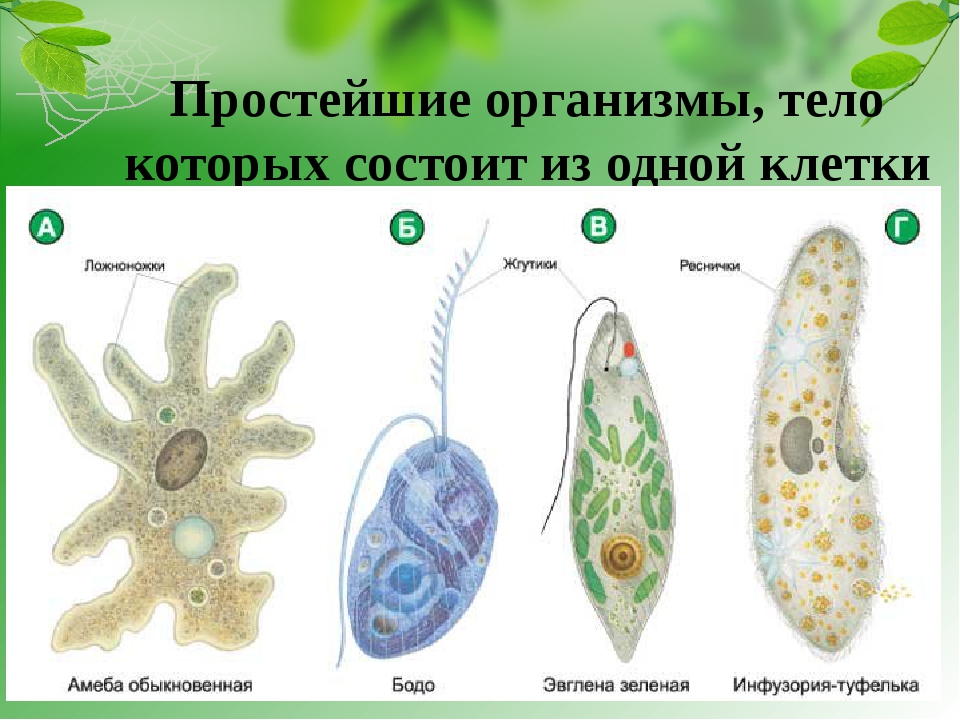 Ядра простейших характеризуются исключительным морфологическим многообразием по размерам, числу ядрышек, количеству ядерного сока и т.д.
Ядра простейших характеризуются исключительным морфологическим многообразием по размерам, числу ядрышек, количеству ядерного сока и т.д.
Особенности жизнедеятельности простейших
В отличие от соматических клеток многоклеточные простейшие характеризуются наличием жизненного цикла. Он слагается из ряда следующих друг за другом стадий, которые в существовании каждого вида повторяются с определенной закономерностью.
Чаще всего цикл начинается стадией зиготы, отвечающей оплодотворенному яйцу многоклеточных. За этой стадией следует однократно или многократно повторяющееся бесполое размножение, осуществляемое путем клеточного деления. Затем образуются половые клетки (гаметы), попарное слияние которых вновь дает зиготу.
Важной биологической особенностью многих простейших является способность к инцистированию. При этом животные округляются, сбрасывают или втягивают органеллы движения, выделяют на своей поверхности плотную оболочку и впадают в состояние покоя.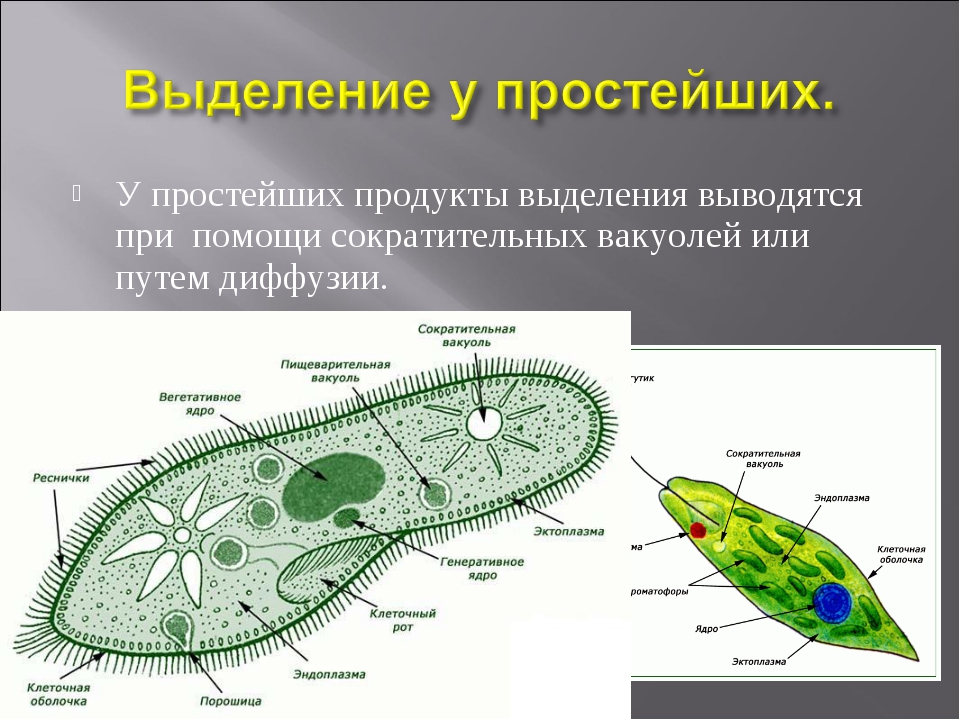 В инцистированном состоянии простейшие могут переносить резкие изменения окружающей среды, сохраняя жизнеспособность. При возвращении благоприятных для жизни условий цисты раскрываются и простейшие выходят из них в виде активных, подвижных особей.
В инцистированном состоянии простейшие могут переносить резкие изменения окружающей среды, сохраняя жизнеспособность. При возвращении благоприятных для жизни условий цисты раскрываются и простейшие выходят из них в виде активных, подвижных особей.
По строению органоидов движения и особенностей размножения тип простейшие делится на 6 классов. Основные 4 класса: Саркодовые, Жгутиковые, Споровики и Инфузории.
Клетки могут перемещаться при помощи специализированных органоидов, к которым относятся реснички и жгутики. Реснички клеток всегда многочисленны (у простейших их количество исчисляется сотнями и тысячами), а длина составляет 10-15мкм. Жгутиков же чаще всего 1-8, длина их — 20-50мкм.
Строение и функции органоидов движения
Строение ресничек и жгутиков, как у растительных, так и животных клеток сходно. Под электронным микроскопом обнаружено, что реснички и жгутики это немембранные органоиды, состоящие из микротрубочек. Две из них располагаются в центре, а вокруг них по периферии лежат еще 9 пар микротрубочек. Вся эта структура покрыта цитоплазматической мембраной, являющейся продолжением клеточной мембраны.
Вся эта структура покрыта цитоплазматической мембраной, являющейся продолжением клеточной мембраны.
Жгутики и реснички обеспечивают не только передвижение клеток в пространстве, но и перемещение различных веществ на поверхности клеток, а также попадание пищевых частиц в клетку. У основания ресничек и жгутиков находятся базальные тельца, которые тоже состоят из микротрубочек.
Предполагают, что базальные тельца являются центром формирования микротрубочек жгутиков и ресничек. Базальные тельца, в свою очередь, нередко происходят из клеточного центра.
Большое количество одноклеточных организмов и некоторые клетки многоклеточных не имеют специальных органоидов движения и передвигаются при помощи псевдоподий (ложноножек), которое получило название амебоидного. В основе его лежит движение молекул особых белков, называемых сократимыми.
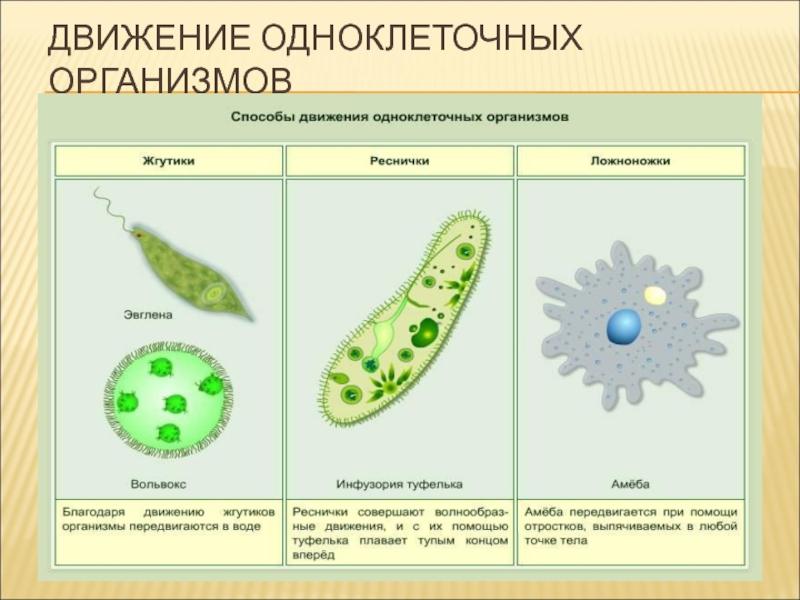
Особенности движения простейших
Одноклеточные организмы также способны передвигаться (инфузория туфелька, эвглена зеленая, амеба обыкновенная).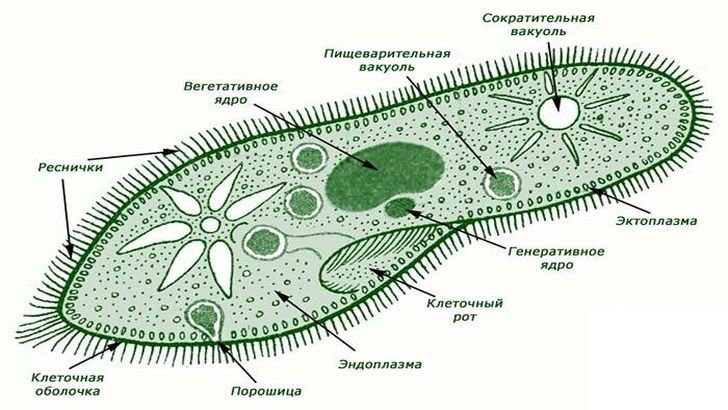 Для перемещения в толще воды каждая особь наделена специфическими органоидами. У простейших такими органоидами являются реснички, жгутики, ложноножки.
Для перемещения в толще воды каждая особь наделена специфическими органоидами. У простейших такими органоидами являются реснички, жгутики, ложноножки.
Эвглена зелёная
Эвглена зелёная — представитель простейших из класса жгутиковых. Тело эвглены веретенообразной формы, удлиненное с заостренным концом. Органоиды движения эвглены зеленой представлены жгутиком, который находится на тупом конце. Жгутики — это тонкие выросты тела, число которых варьирует от одного до десятков.
Механизм движения при помощи жгутика отличается у разных видов. В основном это вращение в виде конуса, вершина которого обращена к телу. Перемещение наиболее эффективно при достижении углом вершины конуса 45°. Скорость колеблется в пределах от 10 до 40 оборотов за секунду. Часто наблюдается помимо вращательного движения жгутика, также его волнообразные покачивания.
Такой характер движения свойствен для одножгутиковых видов. У многожгутиковых нередко жгутики располагаются в одной плоскости и не формируют конуса вращения.
Микроскопическое строение жгутиков довольно сложное. Они окружены тонкой оболочкой, которая является продолжением наружного слоя эктоплазмы — пелликулы. Внутреннее пространство жгутика заполнено цитоплазмой и продольно расположенными нитями — фибриллами.
Периферически расположенные фибриллы отвечают за осуществление движения, а центральные выполняют опорную функцию.
Инфузория туфелька
Передвигается инфузория туфелька за счет ресничек, осуществляя ими волнообразные движения. Направляется вперед тупым концом.
Реснички двигаются в одной плоскости и делают прямой удар после полного выпрямления, а возвратный — в выгнутом положении. Удары идут последовательно один за другим с небольшой задержкой. Во время плаванья, инфузория осуществляет вращательные движения вокруг продольной оси.
Перемещается туфелька со скоростью до 2,5мм/c. Направленность меняется за счёт перегибов тела. Если на пути будет преграда, то после столкновения инфузория начинает двигаться в противоположную сторону.
Все реснички инфузорииимеют сходное строение с жгутиками эвглены зеленой. Ресничка у основания образует базальное зерно, которое играет важную роль в механизме движения организма.
У некоторых инфузорий реснички соединяются между собой и таким образом позволяют развить большую скорость.
Инфузории относятся к высокоорганизованным простейшим и свою двигательную активность они осуществляют с помощью сокращений. Форма тела простейшего может меняться, а после возвращаться в прежнее состояние. Быстрые сократительные движения возможны благодаря наличию особых волокон — мионем.
Амеба обыкновенная
Амеба — простейшее довольно крупных размеров (до 0,5мм). Форма тела полиподиальная, обусловлена наличием множественных псевдоподий — это выросты с внутренней циркуляцией цитоплазмы.
У амебы обыкновенной псевдоподии еще называют ложноножками. Направляя ложноножки в разные стороны, амёба развивает скорость в 0,2 мм/минуту.
К органоидам движения простейших не относятся цитоплазма, ядро, вакуоли, рибосомы, лизосомы, ЭПР, Аппарат Гольджи.
Тело простейшего состоит из цитоплазмы и одного или нескольких ядер. Ядро окружено двойной мембраной и содержит хроматин, в состав которого входит дезаксирибонуклеиновая кислота (ДНК), определяющая генетическую информацию клетки. Большинство простейших имеет пузырьковидное ядро с небольшим содержанием хроматина, собранного по периферии ядра или во внутриядерном тельце, кариосоме. Микронуклеусы инфузорий относятся к ядрам массивного типа с большим количеством хроматина. К обычным компонентам клетки большинства простейших относятся митохондрии и аппарат Гольджи.
Поверхность тела амебоидных форм (саркодовые, а также некоторые стадии
жизненного никла других групп) одета клеточной мембраной толщиной около 100 А. У
большинства простейших имеется более плотная, но эластичная оболочка, пелликула.
Тело многих жгутиконосцев одето перипластом, образованным серией слитых с
пелликулой продольных фибрилл. У многих простейших имеются специальные опорные
фибриллы, как, например, опорная фибрилла ундулирующей мембраны у трипаносом и
трихомонад.
Плотные и жесткие оболочки имеют покоящиеся формы простейших, цисты. Раковинные амебы, фораминиферы и некоторые другие простейшие заключены в домики или раковинки.
В отличие от клетки многоклеточного организма клетка простейшего представляет собой целостный организм. Для выполнения многообразных функций организма в теле простейшего могут специализироваться структурные образования, органеллы. По своему назначению органеллы простейших делятся на органеллы движения, питания, выделения и др.
Весьма разнообразны органеллы движения простейших. Амебоидные формы
перемещаются посредством образования выпячиваний цитоплазмы, псевдоподии. Этот
тип движения носит название амебовидного а встречается у многих групп простейших
(саркодовые, бесполые формы споровиков и др.). Специальными органеллами движения
служат жгутики и реснички. Жгутики свойственны классу жгутиконосцев, а также
гаметам представителей других классов. Они у большинства форм немногочисленны
(от 1 до 8). Количество ресничек, являющихся органеллами движения инфузорий,
может достигать нескольких тысяч у одной особи. Электронномикроскопическое
изучение показало, что жгутики и реснички у Protozoa, Metazoa и растительных
клеток построены по единому типу. Основой их является пучок фибрилл, состоящий
из двух центральных и девяти парных, периферических.
Электронномикроскопическое
изучение показало, что жгутики и реснички у Protozoa, Metazoa и растительных
клеток построены по единому типу. Основой их является пучок фибрилл, состоящий
из двух центральных и девяти парных, периферических.
Жгут окружен оболочкой, являющейся продолжением клеточной мембраны.
Центральные фибриллы имеются лишь в свободной части жгута, а периферические
заходят в глубь цитоплазмы, образуя базальное зерно — блефаропласт. Жгут может
на значительном протяжении соединяться с цитоплазмой тонкой перепонкой —
ундулирующей мембраной. Ресничный аппарат инфузорий может достигать значительной
сложности и дифференцироваться на зоны, выполняющие самостоятельные функции.
Реснички часто сливаются группами, образуя шипы и мембранеллы. Каждая ресничка
начинается от базалытого зерна, кинетосомы, залегающей в поверхностном слое
цитоплазмы. Совокупность кинетосом образует инфрацилиатуру. Кннетосомы
воспроизводятся только делением надвое и не могут возникать заново. При
частичной или полной редукции жгутикового аппарата инфрацилиатура остается и в
дальнейшем дает начало новым ресничкам.
Движение простейших происходит при помощи временных или постоянных органоидов движения. К первым относятся псевдоподии, или ложноножки,- временно образующиеся выросты эктоплазмы, например, у амебы, в которые как бы «переливается» эндоплазма, благодаря чему само простейшее как бы «перетекает» с места на место. Постоянными органоидами движения являются бичи, или жгутики, и реснички.
Все эти органоиды являются выростами протоплазмы простейшего. Жгут имеет по оси более плотную эластическую нить, одетую как бы футляром из более жидкой плазмы. В теле простейшего основание жгута соединяется с базальным зерном, которое считается гомологом центрозомы. Свободным концом жгут бьет по окружающей жидкости, описывая кругообразные движения.
Реснички, в противоположность бичам, очень коротки и чрезвычайно
многочисленны. Реснички быстро наклоняются в одну сторону и затем медленно
выпрямляются; движение их происходит последовательно, благодаря чему глаз
наблюдателя получает впечатление мерцания пламени, и самое движение называется
мерцательным.
Некоторые простейшие могут одновременно иметь псевдоподии и жгут или псевдоподии
и реснички. У других же простейших могут наблюдаться различные способы
передвижения в разных стадиях жизненного цикла.
У некоторых простейших в протоплазме диференцируются сократительные волокна, или
мионемы, благодаря работе которых тело простейших может быстро изменять форму.
В первом случае заглатывание пищи производится работой псевдоподий так называемое фагоцитарное питание,- например заглатывание цист простейших и бактерий кишечной амебой или же ресничками, загоняющими частицы в клеточный рот (цитостом например, инфузории Balantidium coll и зерна крахмала). Эндосмотическое питание свойственно простейшим, не имеющим органоидов питания, например, трипанозомам, лейшманиям, грегаринам, некоторым инфузориям и мн. др. Питание в таких случаях происходит за счет всасывания органических растворенных веществ из окружающей среды; такая форма питания называется также сапрофитной.
Заглатываемые пищевые вещества поступают в эндоплазму где и происходит их
переваривание.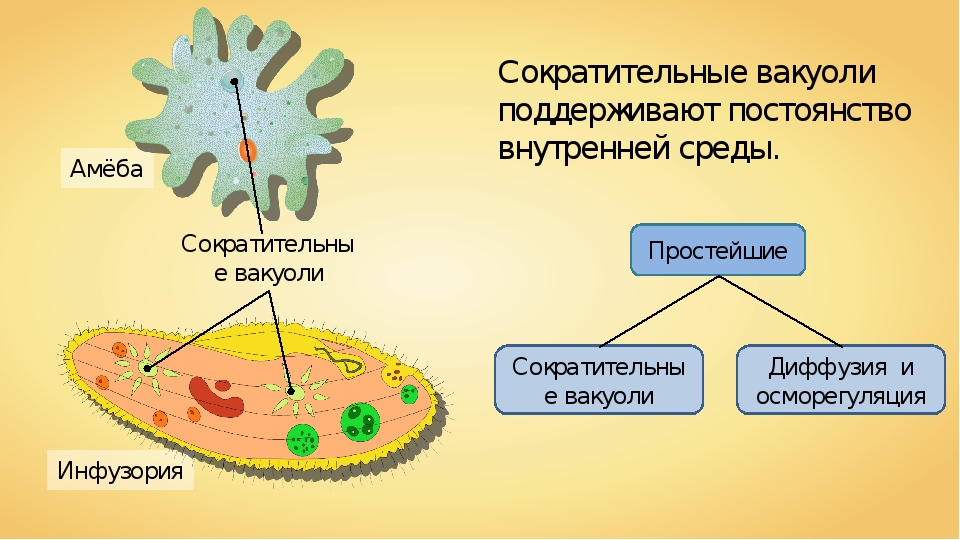 Неиспользованные остатки выбрасываются наружу или в любом месте
поверхности тела простейшего или в определенном его участке (аналогия процесса
дефекации).
Неиспользованные остатки выбрасываются наружу или в любом месте
поверхности тела простейшего или в определенном его участке (аналогия процесса
дефекации).
В эндоплазме простейшего отлагаются запасные питательные вещества в виде
гликогена, парагликогена (нерастворимого в холодной воде и в спирту), жира и
других веществ.
В эндоплазме заложен также и экскреторный аппарат, если он вообще морфологически
выражен у данного вида простейшего. Органоидами экскреции, а также осморегуляции,
частично и дыхания являются пульсирующие вакуоли, которые, ритмически
сокращаясь, опорожняют наружу свое жидкое содержимое, снова набирающееся в
вакуоль из прилежащих частей эндоплазмы. В эндоплазме же заложено ядро
простейшего. Многие простейшие имеют два или более ядер, обладающих у различных
Protozoa разнообразным строением.
Ядро является необходимой составной частью простейшего, ибо все жизненные
процессы могут протекать только при его наличии; безъядерные участки протоплазмы
простейшего в условиях эксперимента могут лишь некоторое время переживать.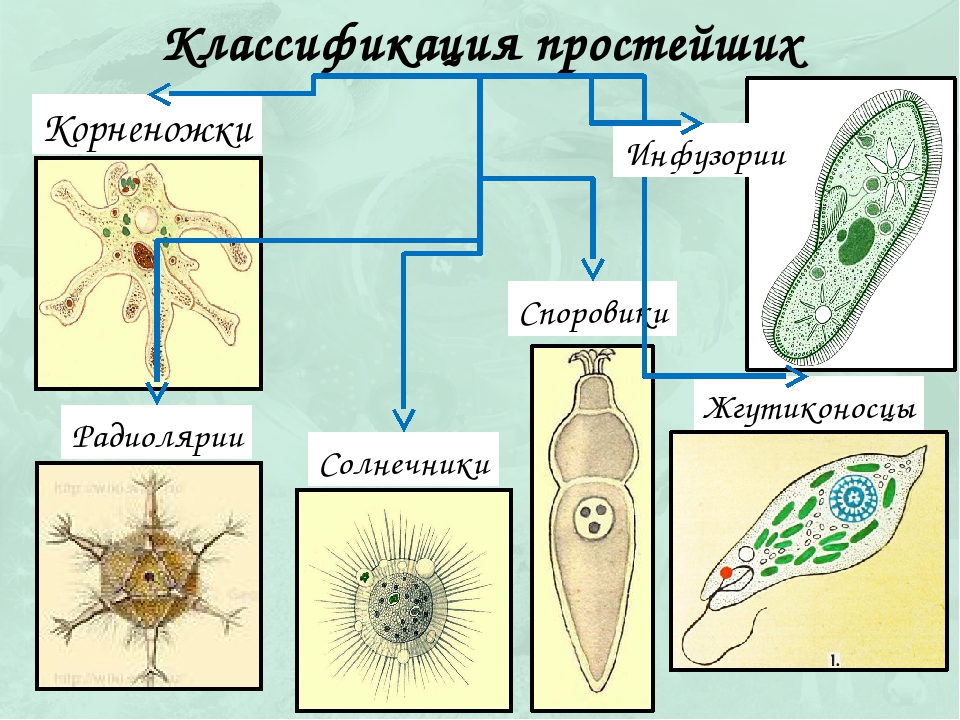
У простейших отмечается специфичность и к переносчикам. Некоторые виды их адаптируются лишь к одному определенному переносчику, для других-переносчиками могут быть несколько видов, относящихся часто к какому-либо одному классу.
Строение простейших одноклеточных организмов
Клетка простейшего — это самостоятельный организм
, которому свойственны все жизненные функции: обмен веществ, раздражимость, передвижение в пространстве.
Все простые относятся к эукариот, их клетки имеют оформленное ядро, в котором находится генетический материал. От окружающей среды клетка отделена клеточной мембраны, которая выкапывает барьерную и защитную функции. В цитоплазме различают два слоя: эктоплазмы (внешний) и ендоплазму (внутренняя). К органелл, типичных для эукариотических клеток, принадлежащих митохондрии, эндоплазматический ретикулум, рибосомы, аппарат Гольджи, лизосомы, ядро. Наряду с ними в цитоплазме простейших присутствуют специфические органеллы — мае и сократительные вакуоли, общее тельце (в жгутиковых), ячейки.
Сократительная вакуоль — это органеллы осморегуляции, контролирует поступление воды в клетку. Вода скапливается в скорачивающейся вакуоли, а вследствие ее сокращение выливается наружу сквозь время. Другие функции сократительной вакуоли: выделение – вместе с водой наружу выводятся продукты обмена веществ дыхания — с водой поступает растворенный кисень.Травна вакуоль выполняет функцию травлення.У ее полость выделяются ферменты, перетравливают поглощены частицы пищи (органические остатки, бактерии и проч.).
Как и всем живым организмам, простым присуща раздраженность — способность реагировать на различные изменения окружающей среды. Раздражительность одноклеточных носит характер. Различают положительные таксисы — движение к источнику раздражения,и отрицательные — избежание воздействия раздражителя.
По типу питания простейшие могут быть фототрофы и гетеротрофами. Некоторые одноклеточные, например эвглена зеленая, в условиях яркого освещения себя как фототрофы, а в темноте, при наличии органических веществ, переходят на гетеротрофный тип питания. Среди простейших встречаются свободноживущие (Прикрепленные, свободноплавающие) и паразитические формы. Вольные способны к активному движению, которое обеспечивается меняющимися выростами клетки — псевдособытиями, (амебы, радиолярии), жгутиками (хламидомонада, трипаносома), ресничками (инфузории). Многие из простейших образуют колонии (вольвокс). Некоторые одноклеточные приспособились к паразитическому образу жизни (дизентерийная амеба, малярийный плазмодий).
Среди простейших встречаются свободноживущие (Прикрепленные, свободноплавающие) и паразитические формы. Вольные способны к активному движению, которое обеспечивается меняющимися выростами клетки — псевдособытиями, (амебы, радиолярии), жгутиками (хламидомонада, трипаносома), ресничками (инфузории). Многие из простейших образуют колонии (вольвокс). Некоторые одноклеточные приспособились к паразитическому образу жизни (дизентерийная амеба, малярийный плазмодий).
Простейшие распространены повсеместно. Они освоили пресные, морские воды и почву. Многие простых, живущих в Мировом океане, должны минеральный скелет (фораминиферы, радиолярии) с кальций карбоната или кремний оксида. После отмирания организмов эти скелеты образуют мощные донные залежи.
На сегодня описано более 30 тыс. видов простейших. Тип простых делят на классы: саркодовые, Жгутиковые, Споровики, Инфузории.
Урок простейших для детей: определение и факты — видео и расшифровка урока
Части простейших
Точно так же, как у вас есть разные части тела, у простейших они тоже есть! Кожа простейших называется пелликулой. У простейших есть цитосома, которая во многом напоминает рот, принимающий пищу, и вакуоль, похожая на желудок, помогающая переваривать пищу. Как и у большинства других клеток, у простейших есть ядро, похожее на мозг клетки. У некоторых их даже два: большой мозг, называемый макронуклеусом, и маленький мозг, называемый микронуклеусом.
У простейших есть цитосома, которая во многом напоминает рот, принимающий пищу, и вакуоль, похожая на желудок, помогающая переваривать пищу. Как и у большинства других клеток, у простейших есть ядро, похожее на мозг клетки. У некоторых их даже два: большой мозг, называемый макронуклеусом, и маленький мозг, называемый микронуклеусом.
Как мы уже говорили, простейшие могут двигаться, но разные виды простейших передвигаются, используя разные части тела. У некоторых простейших есть псевдоподии, которые, по сути, являются фальшивыми ногами — простейшие могут создавать временные бугорки, которые выходят из нижней части клетки и помогают ей двигаться. Большинство других видов простейших используют для передвижения постоянные части тела: у некоторых есть реснички (короткие структуры, такие как волоски), а у других есть жгутики (более длинные структуры, такие как хлысты).
Protozoa Среда обитания и диета
Простейшие обитают в самых разных местах, но обычно им нравится оставаться во влажной среде. Их можно найти как в соленой, так и в пресноводной среде обитания, включая океаны, пруды, реки, болота и озера. Простейшие могут жить на самом дне моря, а могут жить у поверхности воды. Их также часто можно встретить в разных типах почвы.
Их можно найти как в соленой, так и в пресноводной среде обитания, включая океаны, пруды, реки, болота и озера. Простейшие могут жить на самом дне моря, а могут жить у поверхности воды. Их также часто можно встретить в разных типах почвы.
Эти влажные среды являются идеальным домом для крошечных простейших, потому что они являются питательной средой для водорослей и бактерий, которыми простейшие любят питаться. Простейшие также поедают других простейших. Иногда эти организмы теряют маленькие кусочки себя, и простейшие тоже поедают эти крошечные кусочки!
Итоги урока
Хорошо, теперь, когда мы рассмотрели всю информацию о простейших, давайте повторим то важное, что мы узнали. Простейшие — это крошечные одноклеточные животные, которые живут во влажной среде, такой как пруды, болота и почва. Они могут жить сами по себе или жить внутри более крупного растения или животного, действуя как паразитов , что означает, что они живут и питаются растениями и животными (включая людей), чтобы выжить. Они питаются бактериями, водорослями и другими простейшими и передвигаются с помощью ресничек, жгутиков или псевдоподий. Теперь вы должны быть полностью осведомлены об этих маленьких существах, и вы сможете идентифицировать их, если посмотрите на них в микроскоп!
Они питаются бактериями, водорослями и другими простейшими и передвигаются с помощью ресничек, жгутиков или псевдоподий. Теперь вы должны быть полностью осведомлены об этих маленьких существах, и вы сможете идентифицировать их, если посмотрите на них в микроскоп!
Клеточная биология патогенных простейших и их взаимодействие с клетками-хозяевами
Царство Protista включает большое количество эукариотических микроорганизмов, которые являются возбудителями важных паразитарных заболеваний.Некоторые из этих заболеваний, такие как болезнь Шагаса, вызываемая Trypanosoma cruzi , в основном ограничены регионами Латинской Америки. Другие, такие как токсоплазмоз, вызываемый Toxoplasma gondii , распространены по всему миру.
В этом специальном выпуске собраны десять оригинальных статей и два всесторонних обзора, которые, хотя и не являются полным представлением в данной области, представляют собой важную смесь многогранных знаний, которыми мы с удовольствием делимся с читателями.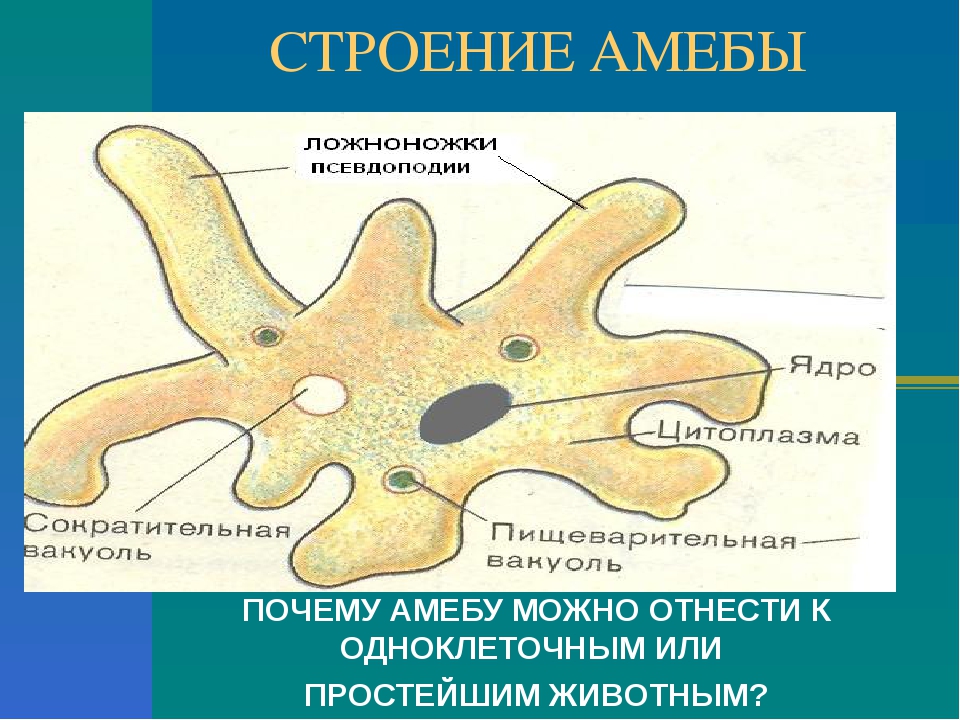 Эти статьи охватывают соответствующие аспекты биологии и взаимодействия между клетками-хозяевами и такими видами, как Trypanosoma cruzi, Trypanosoma brucei, Toxoplasma gondii, Plasmodium falciparum, Trichomonas vaginalis, Entamoeba histolytica, и Blastocystis видов.
Эти статьи охватывают соответствующие аспекты биологии и взаимодействия между клетками-хозяевами и такими видами, как Trypanosoma cruzi, Trypanosoma brucei, Toxoplasma gondii, Plasmodium falciparum, Trichomonas vaginalis, Entamoeba histolytica, и Blastocystis видов.
О. Шериф и др. используйте Trypanosoma brucei в качестве модели для анализа динамики субпелликулярных микротрубочек, визуализируемых с помощью индуцируемой экспрессии YFP- α -тубулина, особенно во время биогенеза и роста клеток нового жгутика / зоны прикрепления жгутика (FAZ).Модификации цитоскелета на заднем конце клеток также наблюдались с использованием белка EB1, связывающего плюс-конец микротрубочек, особенно во время митоза. Результаты предполагают тесную связь между образованием новых микротрубочек и сборкой новых FAZ.
C.M. Batista et al. описывают приобретение и характеристику моноклонального антитела, которое распознает крузипаин, основную цистеиновую протеиназу, обнаруженную в Trypanosoma cruzi , особенно в резервосоме, особой органелле эндоцитарного пути этого простейшего.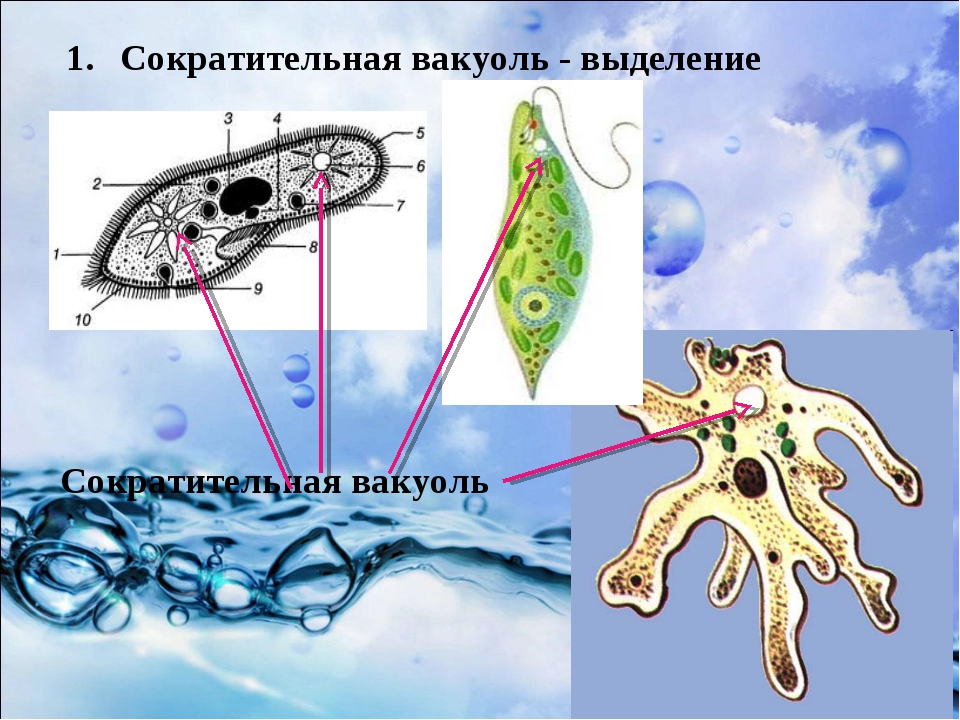
С. Чимадан и др. описывают анализ роли передачи сигналов кальция в регуляции транскрипции генома апикопласта Plasmodium falciparum . Были проанализированы транскрипционные ответы этого простейшего на два ионофора кальция, а также остановка развития на стадии шизонта. Кроме того, снижение стационарных уровней мРНК наблюдалось практически во всех транскриптах, кодируемых геномами апикопластов клеток, обработанных ионофорами. Кроме того, был идентифицирован и локализован апикопласт, кодируемый ядром белок с кальций-связывающим доменом.
J. S. Calla-Choque et al. анализировать процессы регуляции генов на транскрипционном и посттранскрипционном уровнях, опосредованные концентрацией железа в Trichomonas vaginalis . Белок был идентифицирован и обозначен как TvACTN3, цитоплазматический белок, который специфически связывается со шпилечными РНК-структурами трихомонад, когда паразиты выращиваются в условиях дефицита железа. Т.о., TvACTN3 может участвовать в регуляции экспрессии генов железом у T. vaginalis .
vaginalis .
К. Хименес и др. описывают сравнительный анализ роли кальретикулина (CRT), обнаруженного у патогенных видов Entamoeba histolytica и непатогенных видов Entamoeba dispar , которые взаимодействуют с человеческим C1q, ингибируя активацию классического пути комплемента. Показано, что CRT и C1q человека совместно локализуются в цитоплазматических везикулах, расположенных вблизи поверхностной мембраны паразита. Уровень экспрессии CRT был проанализирован in situ в поражениях, связанных с амебным абсцессом печени у хомяка, и результаты показали, что CRT может модулировать некоторые функции в ранние моменты взаимоотношений паразит-хозяин.
J. Pacheco-Yepez и коллеги анализируют молекулярные механизмы, участвующие в формировании амебного абсцесса печени, вызванного Entamoeba histolytica . Также исследовалась важность пероксинитрита (ONOO — ) как основного агента образования абсцесса печени во время амебной инвазии, так и в качестве объяснения превосходной способности амеб защищаться от этого токсического агента через системы пероксиредоксина и тиоредоксин.
Z. Wu et al.описывают анализ процесса взаимодействия Blastocystis , нового протистанового паразита, колонизирующего кишечник человека, с поляризованной линией эпителиальных клеток толстой кишки человека Caco-2. Было показано, что протисты индуцируют апоптоз эпителиальных клеток путем активации каспаз 3 и 9 клетки-хозяина, но не 8.
K. D. Cruz et al. представляют экспериментальные доказательства того, что липидные рафты из клеток-хозяев (макрофаги и эпителиальные клетки) играют определенную роль в процессе инвазии клеток-хозяев патогенными простейшими Toxoplasma gondii .
М. Л. Чирибао и др. сообщают об исследовании взаимодействия Trypanosoma cruzi с эпителиальными клетками in vitro, где было идентифицировано до 1700 значительно измененных генов, регулируемых непосредственным инфицированием. Это указывает на то, что клетки-хозяева перепрограммированы с помощью T . cruzi , который влияет на реакцию клеток на стресс (хемотаксис нейтрофилов и реакцию на повреждение ДНК), большое количество факторов транскрипции (включая большинство членов семейства NF-B) и метаболизм хозяина (холестерин, жирные кислоты и фосфолипиды).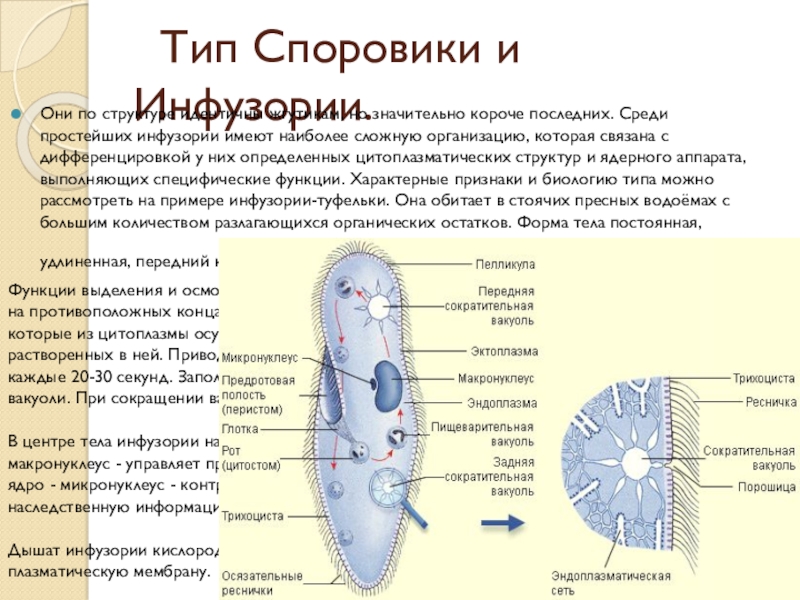
M. R. Garcia-Silva et al. далее проанализируйте роль, которую играют малые РНК, обнаруженные в микровезикулах, происходящих из эндоцитарного пути и секретируемых во внеклеточную среду патогенными простейшими Trypanosoma cruzi . Сообщалось, что большой набор генов клетки-хозяина экспрессировался при включении внеклеточных везикул, происходящих из T. cruzi , модифицируя цитоскелет клетки-хозяина, внеклеточный матрикс и пути иммунного ответа.
П. Флорентино и др.обзор с исторической точки зрения, как достижения в области микроскопии способствовали пониманию Leishmania spp. и Trypanosoma cruzi отношения хозяин-паразит.
Р. Ф. С. Менна-Баррето и С. Л. де Кастро делают обзор ключевой роли, которую играют митохондрии простейших семейства Trypanosomatidae в окислительном стрессе и биоэнергетике. Эти метаболические процессы представляют собой важную цель для разработки новых лекарств против этих паразитов, особенно если мы сможем лучше понять процессы митохондриальной окислительной регуляции.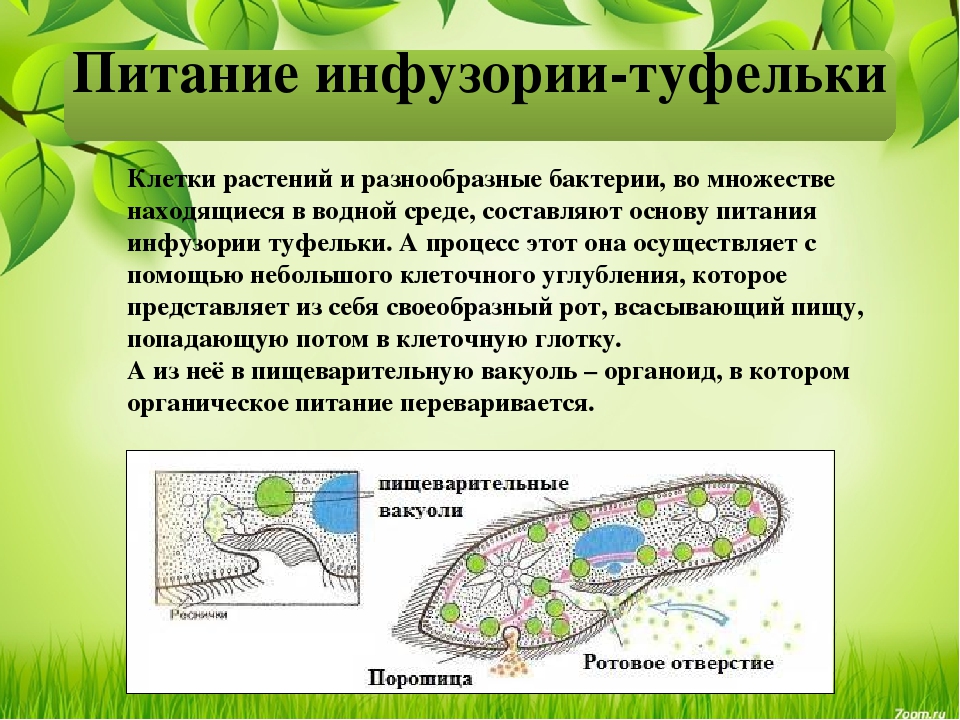
Marlene Benchimol
Juan C. Engel
Kevin S. W. Tan
Wanderley de Souza
Copyright
Copyright © 2014 Marlene Benchimol et al. Это статья с открытым доступом, распространяемая в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии надлежащего цитирования оригинальной работы.
Количество и регулирование простейших аквапоринов отражают сложность окружающей среды
Abascal, F. , И. Ирисарри и Р. Зардоя . 2014 . Разнообразие и эволюция мембранных внутренних белков. Биохим. Биофиз. Acta 1840 : 1468–1481.
, LS King , M. Yasui , WB Guggino , Op ottersen , Y. Fujiyoshi , А.Энгель и С. Нильсен . 2002 . Аквапориновые водные каналы — от атомной структуры до клинической медицины. Дж. Физиол . 542 : 3–16.
2002 . Аквапориновые водные каналы — от атомной структуры до клинической медицины. Дж. Физиол . 542 : 3–16.
Аллен, Р. Д. и Ю. Найтох . 2002 . Осморегуляция и сократительные вакуоли простейших. Междунар. Преподобный Цитол . 215 : 351–394.
Алмасалмех, А. , Д.Krenc , B. Wu и E. Beitz . 2014 . Структурные детерминанты проницаемости аквапоринов для перекиси водорода. ФЕБС J . 281 : 647–656.
Baker, N. , L. Glover , JC Munday , , D. aguinaga andres , MP Barrett , HP de Koning и Д.Рог . 2012 . Акваглицеропорин 2 контролирует чувствительность африканских трипаносом к меларсопролу и пентамидину. Проц. Натл. акад. науч. США 109 : 10996–11001.
Баккер, Б. М. , П. А. Михельс , Ф. Р. Оппердоус и Х. Вестофф 90 1997 . Гликолиз в кровотоке формы Trypanosoma brucei можно понять с точки зрения кинетики гликолитических ферментов. Дж. Биол. Химия . 272 : 3207–3215.
Baldauf, S. L. , A.J. Roger , I. Wenk-Siefert , and W. Dottle 90 2000 . Филогенез эукариот на уровне царства, основанный на комбинированных данных о белках. Наука 290 : 972–977.
Бассарак, Б. , Н. Л. Узкатеги , С.Шёнфельд и М. Душенко . 2011 . Функциональная характеристика трех акваглицеропоринов из Trypanosoma brucei в осморегуляции и транспорте глицерина. Сотовый. Физиол. Биохим . 27 : 411–420.
Бейтц, Э. 2005 . Аквапорины патогенных простейших паразитов: структура, функция и потенциал для химиотерапии. Биол. Сотовый 97 : 373–383.
Биол. Сотовый 97 : 373–383.
Бейтц, Э. 2006 . Аквапорин водные и растворенные каналы от малярийных паразитов и других патогенных простейших. ХимМедХим 1 : 587–592.
Бейтц, Э. 2007 . Застрявший трафик препятствует росту паразита. Проц. Натл. акад. науч. США 104 : 13855–13856.
Бейтц, Э. , С. Павлович-Джуранович , М.Ясуи , П. Агре и Дж. Э. Шульц . 2004 . Молекулярный анализ проницаемости для воды и глицерина акваглицеропорина из Plasmodium falciparum с помощью мутационного анализа. Проц. Натл. акад. науч. США 101 : 1153–1158.
Biyani, N. , S. Mandal , C. Seth , M. Saint , K. K.Натараджан , И. Гош и Р. Мадхубала . 2011 . Характеристика аквапоринов Leishmania donovani показывает наличие субклеточных аквапоринов, сходных с собственными белками тонопластов растений. PLoS One 6 : e24820.
Мадхубала . 2011 . Характеристика аквапоринов Leishmania donovani показывает наличие субклеточных аквапоринов, сходных с собственными белками тонопластов растений. PLoS One 6 : e24820.
Боззаро С. 2013 . Модельный организм Dictyostelium discoideum . Методы Мол. Биол . 983 : 17–37.
Bülow, J. Von , , A. Müller-Lunks , L. KAI , F. Bernhard и E. Beitz . 2012 . Функциональная характеристика нового аквапорина из Dictyostelium discoideum амеб подразумевает уникальный механизм ворот. Дж. Биол. Химия . 287 : 7487–7494.
Бюлов, Й. фон , А.Голдэк , Т. Альберс и Э. Бейтц . 2015 . Амебоидный аквапорин AqpB Dictyostelium запирается через Tyr216, и делеция гена aqpB влияет на случайную подвижность клеток. Биол. Сотовый 107 : 78–88.
Биол. Сотовый 107 : 78–88.
1
Contreras-Ochoa, C. O. , A. Lagunas-Martinez , J. Belkind-Gerson , J. Diaz-Chavez и D.Корреа . 2013 . Инвазия и репликация Toxoplasma gondii в астроцитах новорожденных мышей и изменения в молекулах, связанных с апоптозом. Экспл. Паразитол . 134 : 256–265.
Коттер, Д. А. и К. Б. Рапер . 1968а . Свойства прорастающих спор Dictyostelium discoideum . Дж. Бактериол . 96 : 1680–1689.
Коттер Д.А. и К. Б. Рапер . 1968б . Прорастание спор у штаммов Dictyostelium discoideum и других представителей Dictyosteliaceae. Дж. Бактериол . 96 : 1690–1695.
Куйяр, П. , Ф. Потье и П. Майерс . 1989 .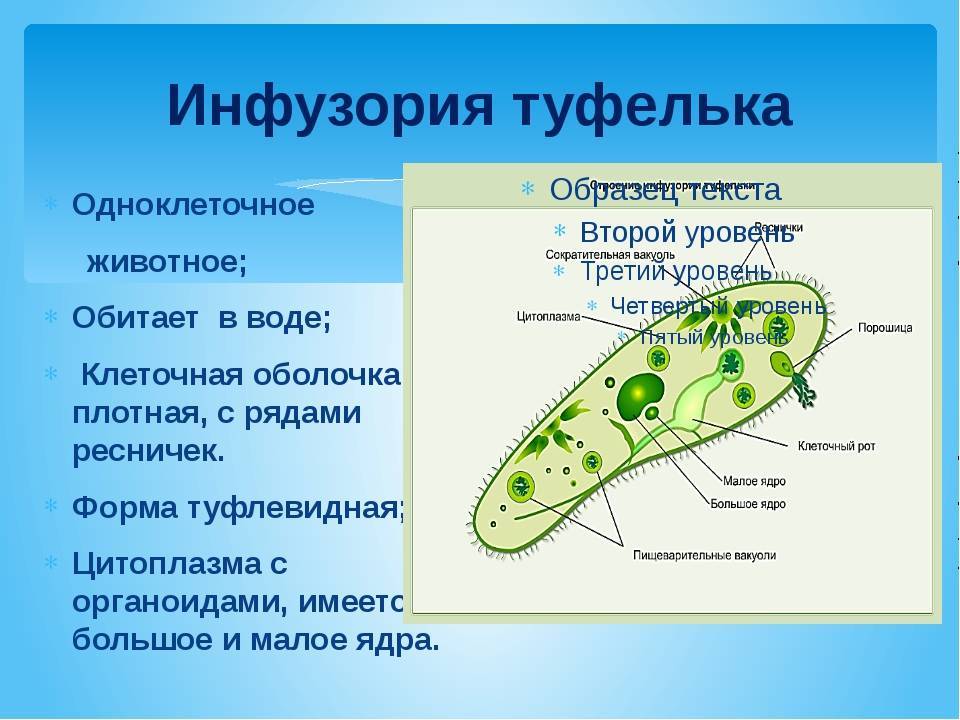 Влияние вазопрессина и родственных пептидов на осморегуляцию у Amoeba proteus . Ген. комп. Эндокринол . 76 : 106–113.
Влияние вазопрессина и родственных пептидов на осморегуляцию у Amoeba proteus . Ген. комп. Эндокринол . 76 : 106–113.
de Groot, B. L. и H. Grubmüller . 2001 . Проникновение воды через биологические мембраны: механизм и динамика аквапорина-1 и GlpF. Наука 294 : 2353–2357.
Эйзенталь, Р. и А. Корниш-Боуден . 1998 . Перспективы противопаразитарных препаратов — случай Trypanosoma brucei , возбудителя африканской сонной болезни. Дж. Биол. Химия . 273 : 5500–5505.
0Fadiel, A. , RD Isokpehi , N. Stumbouli , , A. Hamza , A. Benammar-Elgaaied и TJ Scleise . 2009 . Аквапорины простейших паразитов. Expert Rev. Proteomics 6 : 199–211.
Fichera, ME и D. С. Роос . 1997 . Пластидная органелла как лекарственная мишень у апикомплексановых паразитов. Природа 390 : 407–409.
С. Роос . 1997 . Пластидная органелла как лекарственная мишень у апикомплексановых паразитов. Природа 390 : 407–409.
Figarella, K. , NL UzCategui , Y. Zhou , , A. Lefurgey , M. OUELLETTE , H. Bhattacharjee , и Р. Мухопадхьяй . 2007 . Биохимическая характеристика акваглицеропорина LmAQP1 Leishmania major : возможная роль в регуляции объема и осмотаксиса. Мол. Микробиол . 65 : 1006–1017.
Fischer, G. , U. Kosinska-eriksson , C. aponte-Santamaría , M. Palmgren , C. Geijer , К. Хедфальк , С. Хохманн , Б.Л. де Гроот , Р. Нойтце , и Кетсон-П.00018 7 2009 . Кристаллическая структура дрожжевого аквапорина при 1,15 ангстрем раскрывает новый механизм ворот.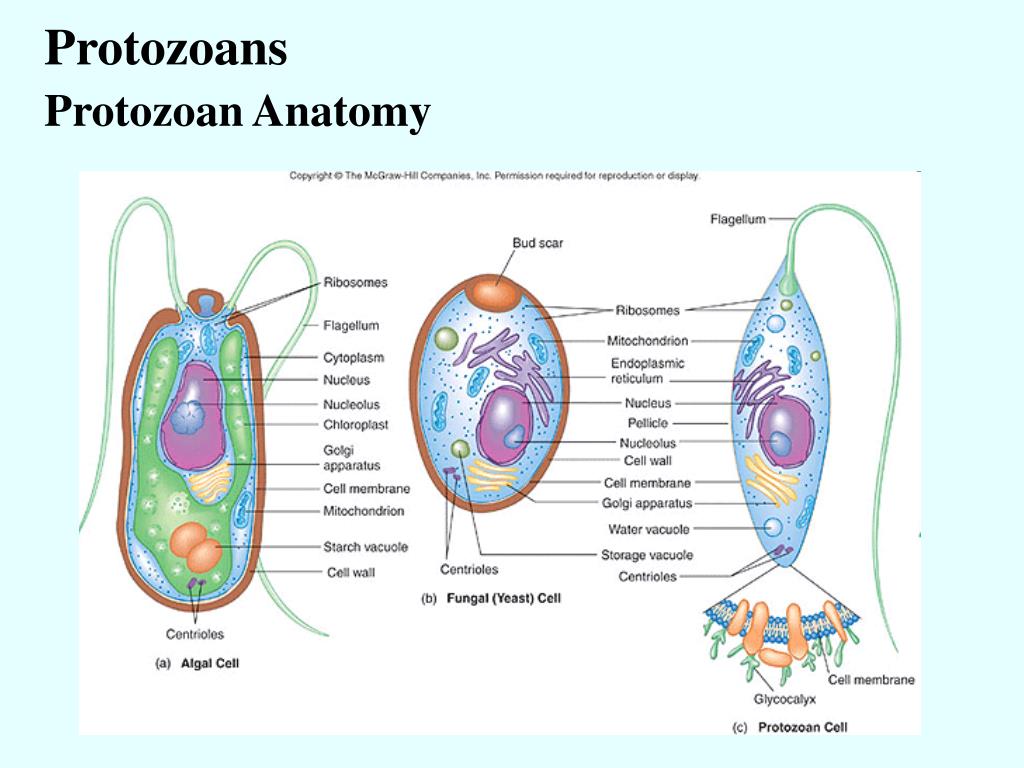 ПЛОС Биол . 7 : e1000130.
ПЛОС Биол . 7 : e1000130.
Флик, К. М. , Г. Шаульский и В. Ф. Лумис . 1997 . Ген wacA Dictyostelium discoideum является регулируемым в развитии членом семейства MIP. Ген 195 : 127–130.
Фу, Д. , A. libson , , , , C. weitzman , P. nollert , J. Krucinski и R. M. Proud . 2000 . Структура глицеринпроводящего канала и основа его селективности. Наука 290 : 481–486.
Гарднер, М. Дж. , Н. Холл , Э.FUNG , O. White , M. Berriman , RW Hyman , JM Carlton , A. Pain , Ke Nelson , С. Боумен и др. . 2002 . Последовательность генома малярийного паразита человека Plasmodium falciparum . Природа 419 : 498–511.
Природа 419 : 498–511.
Джинджер М.Л. 2006 . Метаболизм ниши у паразитических простейших. Филос. Транс. Р. Соц. Лонд. Б биол. Наука . 361 : 101–118.
1
, , , , H. Bhattacharjee , D. Legare , S. Sundar , M. ouellette , BP Rosen и Р. Мухопадхьяй . 2004 . Поглощение лекарств и модуляция лекарственной устойчивости у Leishmania с помощью акваглицеропорина. Дж. Биол. Химия . 279 : 31010–31017.
Хансен, М. , Дж. Ф. Кун , Дж. Э. Шульц , и
Беитц . 2002 . Один бифункциональный акваглицеропорин в крови паразитов малярии Plasmodium falciparum . Дж. Биол.Химия . 277 : 4874–4882.
Hedfalk, K. , RM Bill , JG Mullins , S. Karlgren , C. Filipsson , J. Bergstrom , М.Дж. Тамаш , Дж. Ридстрем и С. Хохманн . 2004 . Регуляторный домен в С-концевом удлинении дрожжевого глицеринового канала Fps1p. Дж. Биол. Химия . 279 : 14954–14960.
, RM Bill , JG Mullins , S. Karlgren , C. Filipsson , J. Bergstrom , М.Дж. Тамаш , Дж. Ридстрем и С. Хохманн . 2004 . Регуляторный домен в С-концевом удлинении дрожжевого глицеринового канала Fps1p. Дж. Биол. Химия . 279 : 14954–14960.
Хуанг, Г. , Дж. Фанг , С. Сант-Анна , Z.-H Ли , Д. Л. Веллемс , П. Рохлофф и Р. Докампо . 2011 . Комплекс адапторного белка-3 (AP-3) опосредует биогенез ацидокальцисом и необходим для роста и вирулентности Trypanosoma brucei . Дж. Биол. Химия . 286 : 36619–36630.
Хуанг, Дж. , Н. Муллапуди , Т. Зихериц-Понтен , и Дж. Киссинг 90 2004 . Первый взгляд на характер и масштаб переноса генов в апикомплексах.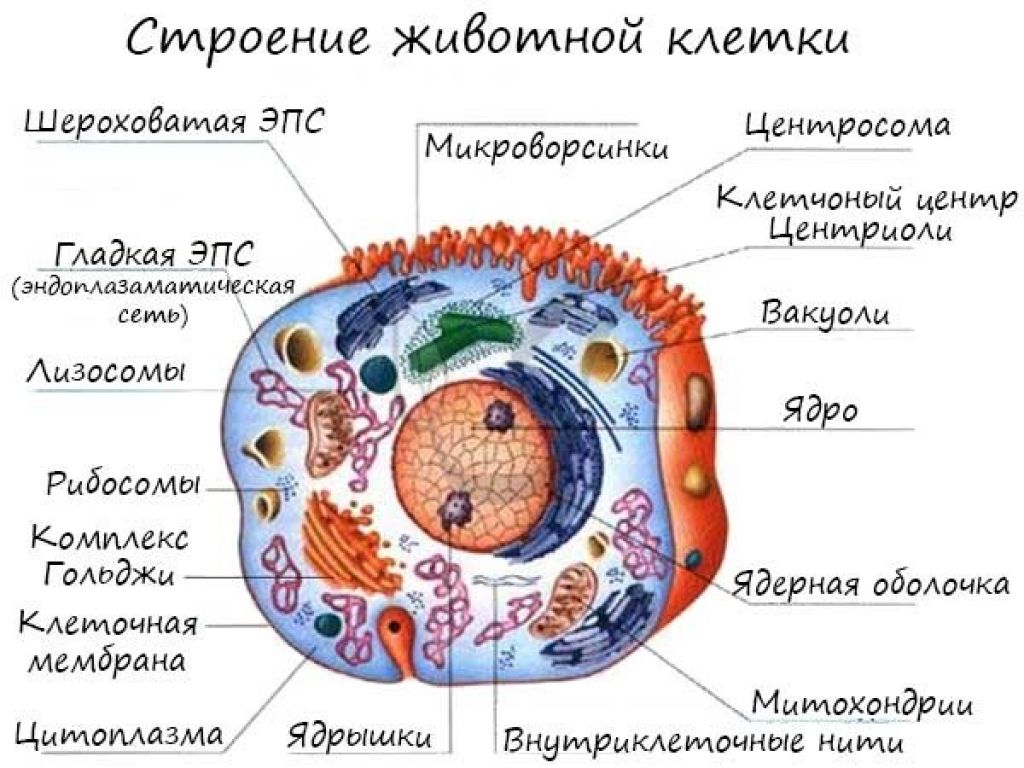 Междунар. Дж. Паразитол . 34 : 265–274.
Междунар. Дж. Паразитол . 34 : 265–274.
Хвиид, Л. , Л. Барфод и Ф.Дж. И. Фоукс . 2015 . Пытаюсь вспомнить: иммунологическая В-клеточная память на малярию. Тренды Паразитол . 31 : 89–94.
Чон, К. В. 1995 . Крупные свободноживущие амебы: прекрасные клетки для биологических исследований. Дж. Эукариот. Микробиол . 42 : 1–7.
Йоханссон И. , М. Карлссон , В. К. Шукла , М.Дж. Криспилс , К. Ларссон и П. Кьеллбом . 1998 . Воднотранспортная активность аквапорина PM28A плазматической мембраны регулируется фосфорилированием. Растительная клетка 10 : 451–459.
Karlgren, S. , C. FILIPSSON ,
 M. Bill , M. J. Tamás и S.Хохманн . 2004 . Идентификация остатков, контролирующих транспорт через дрожжевой акваглицеропорин Fps1, с помощью генетического скрининга. евро. Дж. Биохим . 271 : 771–779.
M. Bill , M. J. Tamás и S.Хохманн . 2004 . Идентификация остатков, контролирующих транспорт через дрожжевой акваглицеропорин Fps1, с помощью генетического скрининга. евро. Дж. Биохим . 271 : 771–779.Кумар, Ю. и Р. Х. Вальдивия . 2009 . Защищенный образ жизни: внутриклеточные патогены и поддержание вакуолярных компартментов. Микроб-хозяин клетки 5 : 593–601.
Кун, Дж.F. и EG de Carvalho . 2009 . Новые терапевтические мишени в Plasmodium falciparum: акваглицеропоринов. Экспертное заключение. тер. Цели 13 : 385–394.
Leirião, P. , C.D. Rodrigues , S.S. Albuquerque 901 M. , и 9001 Mo 2004 . Выживание простейших внутриклеточных паразитов в клетках-хозяевах. EMBO Реп. . 5 : 1142–1147.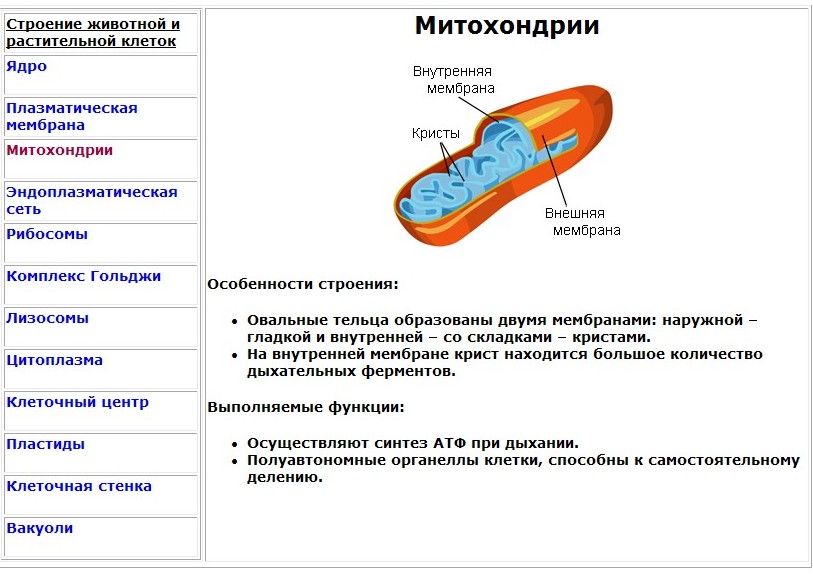
Luyten, K. , J. Albertyn , WF Skibbe , BA предыдущий , J. Ramos , JM Thevelin С. Хохманн . 1995 . Fps1, дрожжевой член семейства белков каналов MIP, способствует поглощению и оттоку глицерола и неактивен при осмотическом стрессе. EMBO J . 14 : 1360–1371.
Mandal, G. , M. Sharma , M. Kruse , C. Sander-Juelch , La Munro , Y. Wang , СП VILG , MJ TAMAS , H. Bhattacharjee , M. Wiese и R. Mukhopadhyay . 2012 . Модуляция активности акваглицеропорина Leishmania major с помощью митоген-активируемой протеинкиназы. Мол. Микробиол . 85 : 1204–1218. Мандал, Г. , Дж.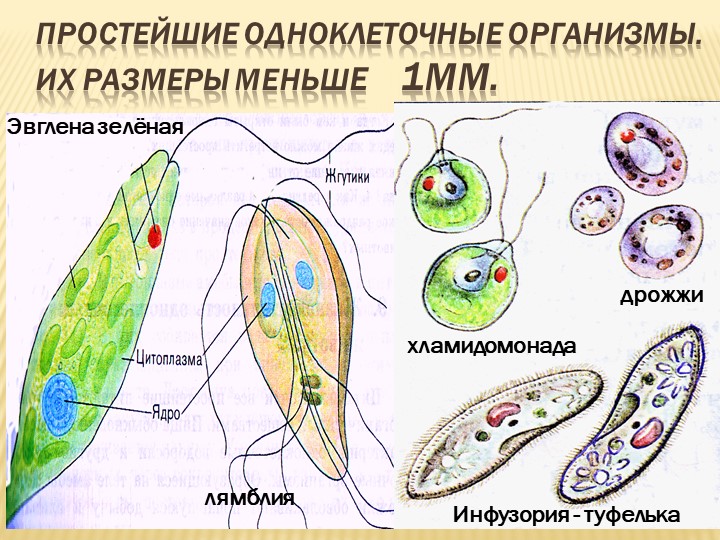 Орта , М. Шарма , и
Орта , М. Шарма , и
Мандал, Г. , S. Mandal , M. Sharma , K. S. Charrez , , B. Papadopulou , H. Bhattachariee и R. Mukhopadhyay . 2015 . Видоспецифическая чувствительность к сурьме у Leishmania обусловлена посттранскрипционной регуляцией AQP1. PLoS Негл. Троп. Дис . 9 : e0003500.
Морел, К. , H. Javot , , , P. Gerbeau , C. Tournaire , V. Santoni и J. Heyes . 2002 . Молекулярная физиология аквапоринов в растениях. Междунар. Преподобный Цитол . 215 : 105–148.
Миранда, К. , Д. А. Пейс , Р. Цинтрон , Дж.C. Rodrigues , , J. Fang , A. Smith , P. Rohloff , E. Coelho , F. de haas , W де Соуза и др. . 2010 . Характеристика новой органеллы Toxoplasma gondii с аналогичным составом и функцией растительной вакуоли. Мол. Микробиол . 76 : 1358–1375.
, Д. А. Пейс , Р. Цинтрон , Дж.C. Rodrigues , , J. Fang , A. Smith , P. Rohloff , E. Coelho , F. de haas , W де Соуза и др. . 2010 . Характеристика новой органеллы Toxoplasma gondii с аналогичным составом и функцией растительной вакуоли. Мол. Микробиол . 76 : 1358–1375.
Mitra, BN , R. yoshino , T. Morio , M. yokoyama , M. Maeda , H. urushihara , и Ю. Танака . 2000 . Потеря члена семейства генов аквапоринов, aqpA, влияет на состояние покоя спор у Dictyostelium . Ген 251 : 131–139.
Монтальветти, А. , П. Рохлофф и Р. Докампо . 2004 .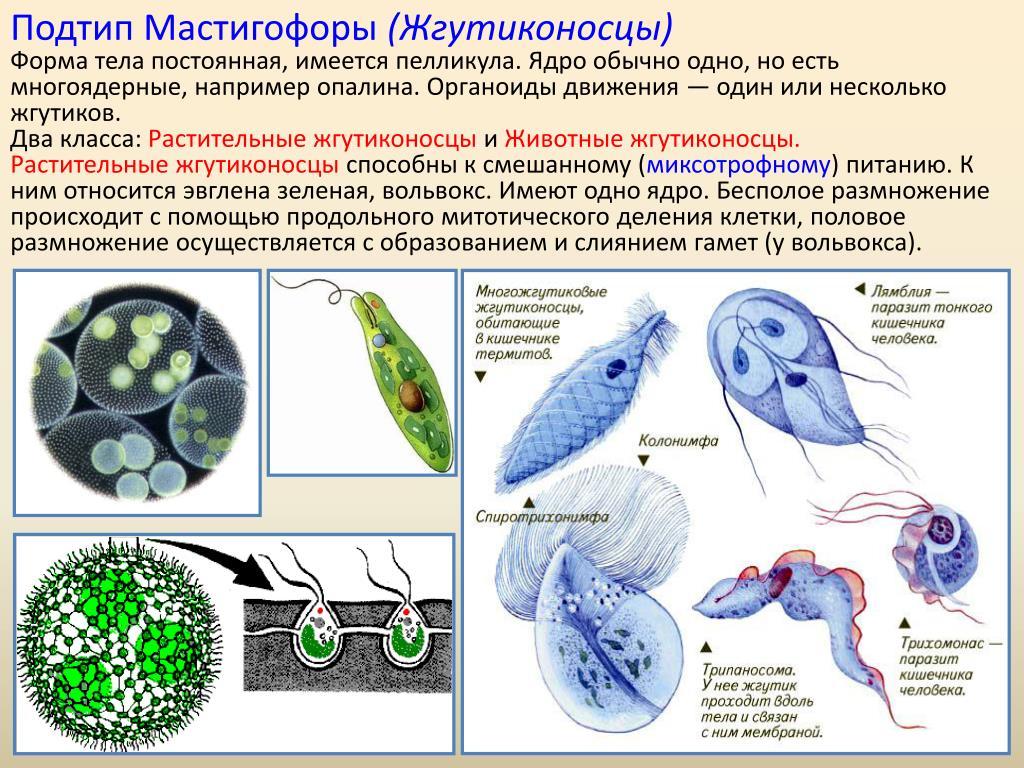 Функциональный аквапорин совместно локализуется с вакуолярной протонной пирофосфатазой в ацидокальцисомах и комплексе сократительных вакуолей Trypanosoma cruzi . Дж. Биол. Химия . 279 : 38673–38682.
Функциональный аквапорин совместно локализуется с вакуолярной протонной пирофосфатазой в ацидокальцисомах и комплексе сократительных вакуолей Trypanosoma cruzi . Дж. Биол. Химия . 279 : 38673–38682.
1
Munday, J. C. , A. A. EZE , , N. Baker , L. Glover , C.CLUCAS , D. AGUINAGA ANDRÉS , , , I. A. Teka , J. McDonald , R. S. Lee et al . 2014 . Trypanosoma brucei Акваглицеропорин 2 является высокоаффинным переносчиком препаратов пентамидина и меламинофенилмышьяка и основной генетической детерминантой резистентности к этим препаратам. J. Антимикроб. Чемотер . 69 : 651–663.
Murata, K. , K. Mitsuoka , , , , , T.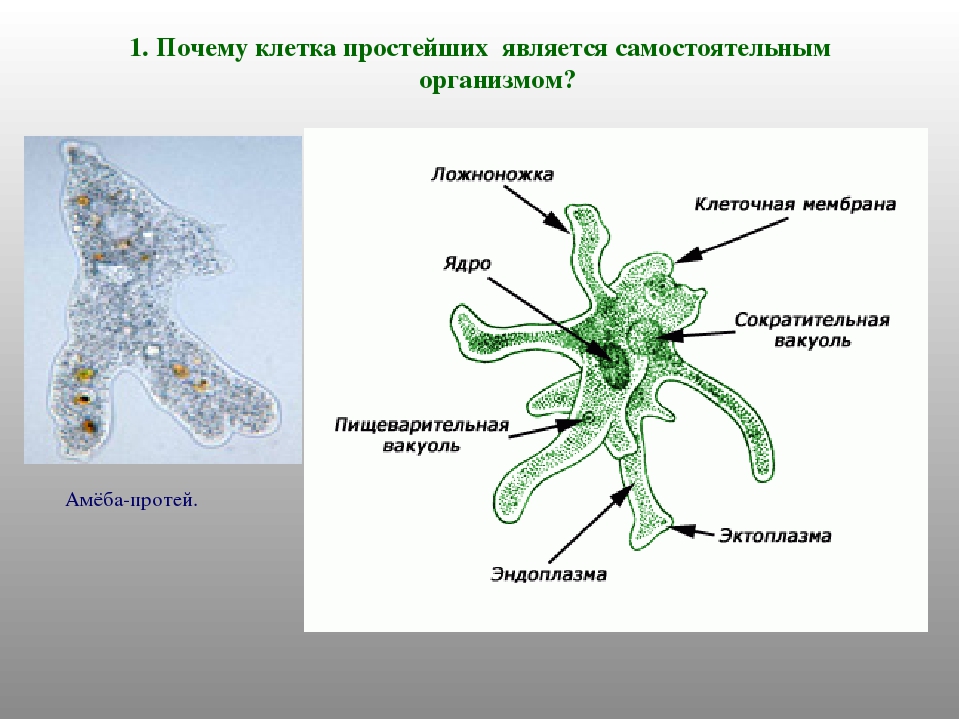 Walz , P.uy , JB Heymann , А. Энгель и Ю. Фудзиёси . 2000 . Структурные детерминанты проникновения воды через аквапорин-1. Природа 407 : 599–605.
Walz , P.uy , JB Heymann , А. Энгель и Ю. Фудзиёси . 2000 . Структурные детерминанты проникновения воды через аквапорин-1. Природа 407 : 599–605.
Ньюэлл, П.C. и IA Mullens . 1978 . Рецепторы цАМФ на клеточной поверхности у Dictyostelium . Симп. соц. Эксп. Биол . 32 : 161–171.
Нишихара, Э. , Т. Шиммен и С. Сонобе . 2004 . Функциональная характеристика сократительной вакуоли, выделенной из Amoeba proteus . Структура ячейки. Функция . 29 : 85–90.
Nishihara, E. , E. Yokota , A. Tazaki , H. Orii , M. Katsuhara , K. Kataoka , H. Igarashi , Y. Moriyama , T.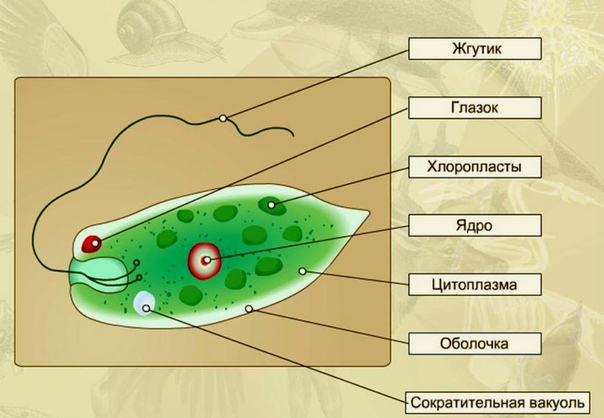 Shimmen , and S. Sonobe . 2008 . Presence of aquaporin and V-ATPase on the contractile vacuole of Amoeba proteus . Биол. Сотовый 100 : 179–188.
Shimmen , and S. Sonobe . 2008 . Presence of aquaporin and V-ATPase on the contractile vacuole of Amoeba proteus . Биол. Сотовый 100 : 179–188.
Нолта, К. В. и Т. Л. Стек . 1994 . Выделение и первоначальная характеристика двудольного сократительного комплекса вакуолей из Dictyostelium discoideum . Дж. Биол. Химия . 269 : 2225–2233.
Павлович-Джуранович, С. , Дж. Э. Шульц и Э. Бейтц . 2003 . Единственный ген аквапорина кодирует посредник воды/глицерина/мочевины в Toxoplasma gondii со сходством с внутренними белками тонопластов растений. ФЭБС Письмо . 555 : 500–504.
Павлович-Джуранович С. , Дж. Ф. Кун , Дж. Э. Шульц 0 7 Be 1001 8 , 900 900 900 2006 . Дигидроксиацетон и метилглиоксаль, проникающие в акваглицеропорин Plasmodium , ингибируют пролиферацию паразита.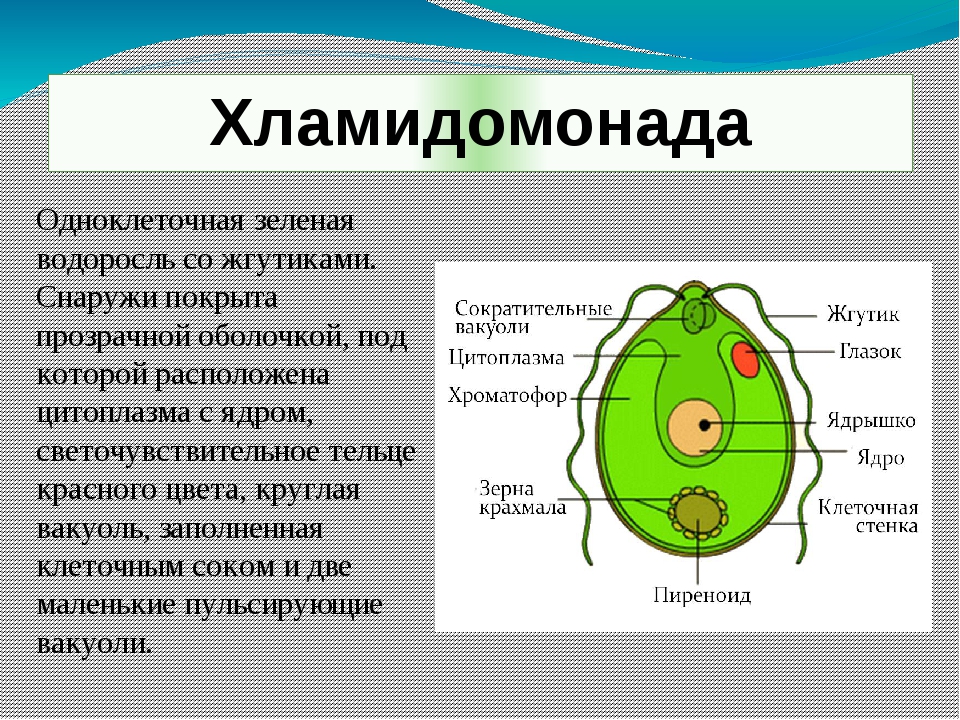 Биохим. Биофиз. Acta 1758 : 1012–1017.
Биохим. Биофиз. Acta 1758 : 1012–1017.
Pearson, R.D. и A.Q. Sousa . 1996 . Клинический спектр лейшманиоза. клин. Заразить. Дис . 22 : 1–13.
Платтнер Х. 2013 . Сократительный комплекс вакуолей — его расширяющийся запас белков. Междунар. Преподобная ячейка. Мол. Биол . 306 : 371–416.
Променёр, Д. , Y. LIU , J. Maciel , P. CORM , , L. S. King и N. KUMAR . 2007 . Акваглицеропорин PbAQP во время внутриэритроцитарного развития малярийного паразита Plasmodium berghei . Проц. Натл. акад. науч. США 104 : 2211–2216.
Расси, А. и Ж. Маркондес де Резенде . 2012 . Американский трипаносомоз (болезнь Шагаса). Заразить. Дис. клин. Северная Ам . 26 : 275–291.
26 : 275–291.
Рохлофф, П. и Р. Докампо . 2008 . Комплекс сократительных вакуолей участвует в осморегуляции у Trypanosoma cruzi . Экспл. Паразитол . 118 : 17–24.
Рохлофф, П. , А. Монтальветти и Р.Докампо . 2004 . Ацидокальцисомы и комплекс сократительных вакуолей участвуют в осморегуляции у Trypanosoma cruzi . Дж. Биол. Химия . 279 : 52270–52281.
ROUDIER, N. , P. PIPOCHE , P. GANE , PY LE PENNEC , G. Daniels , JP Cartron , и П.Байи . 2002 . Дефицит AQP3 у людей и молекулярная основа новой системы групп крови GIL. Дж. Биол. Химия . 277 : 45854–45859.
Саадун, С. , М. К. Пападопулос , М. Хара-Чикума А. Веркман
Веркман
Шапиро, Т. А. и П. Т. Инглунд . 1995 . Структура и репликация кинетопластной ДНК. год. Ред. Микробиол . 49 : 117–143.
1
Smith, B. L. , Г. М. Престон , Ф. А. Весна , , D. J. Anstee и P. CORM . 1994 . ЧИП с аквапорином эритроцитов человека.I. Молекулярная характеристика антигенов групп крови ABH и Colton. Дж. Клин. Инвестировать . 94 : 1043–1049.
Сонг, Дж. , Э. Мак , Б. Ву и Э. Бейтц 90. 2014 . Аквапорины паразитов: современные разработки в области облегчения и устойчивости к лекарственным средствам. Биохим. Биофиз. Acta 1840 : 1566–1573.
Acta 1840 : 1566–1573.
Стейнборн, К. , А. Саллис , Д. Мекке и М. Душенко . 2000 . Клонирование, гетерологичная экспрессия и кинетический анализ глицеролкиназы (TbGLK1) из Trypanosoma brucei . Биол. Химия . 381 : 1071–1077.
Тайхоршид, Е. , П. Ноллерт , М. Ø. Дженсен , Л. Дж. В. Мирке , Дж.О’Коннелл , Р. М. Страуд и К. Шультен . 2002 . Контроль селективности семейства водных каналов аквапоринов путем глобальной настройки ориентации. Наука 296 : 525–530.
Tamás, M. J. , , S. Karlgren , R. M. M. Bill , K. Hedfalk , L. Allegri , М.Ferreira , JM Thevelein , J. Rydström , JG Mullins , и S.0 Homann 8 9 2003 . Короткий регуляторный домен ограничивает транспорт глицерина через Fps1p дрожжей. Дж. Биол. Химия . 278 : 6337–6345.
Rydström , JG Mullins , и S.0 Homann 8 9 2003 . Короткий регуляторный домен ограничивает транспорт глицерина через Fps1p дрожжей. Дж. Биол. Химия . 278 : 6337–6345.
Törnroth-Horsefield, S. , Y. Wang , K. Hedfalk , U 9.Йохансон , М. Карлссон , Э. Тайхоршид , Р. Нойтце и П. . 2006 . Структурный механизм аквапоринового гейтирования растений. Природа 439 : 688–694.
Uzcagei, N. L. , A. Szallies , , S. Pavlovic-Djuranovic , M. Palmada , K.Figarella , C. Boehmer , F. Lang , E. Beitz , и 90 Душенко М. 2004 . Клонирование, гетерологичная экспрессия и характеристика трех акваглицеропоринов из Trypanosoma brucei . Дж. Биол. Химия . 279 : 42669–42676.
Дж. Биол. Химия . 279 : 42669–42676.
Узкатеги, Н. Л. , К. Фигарелла , Б.Бассарак , С.З. Меза , Р. Мухопадхьяй , Ж.Л. 2013 . Trypanosoma brucei акваглицеропорины облегчают поглощение арсенита и антимонита в зависимости от рН. Сотовый. Физиол. Биохим . 32 : 880–888.
Ван Хаастерт, П. Дж. 1983 . Сенсорная адаптация клеток Dictyostelium discoideum к хемотаксическим сигналам. J. Cell Biol . 96 : 1559–1565.
Винсент, И. М. и М. П. Барретт . 2015 . Стратегии, основанные на метаболоме, для открытия противопаразитарных препаратов. Дж. Биомол. Экран . 20 : 44–55.
1
Уокер, Д. М. , S. Oghumu , G. Gupta , B. S. Mcgwire , M. Э. Дрю и А. Р. Сатоскар . 2014 . Механизмы клеточной инвазии внутриклеточными паразитами. Сотовый. Мол. Науки о жизни . 71 : 1245–1263.
Э. Дрю и А. Р. Сатоскар . 2014 . Механизмы клеточной инвазии внутриклеточными паразитами. Сотовый. Мол. Науки о жизни . 71 : 1245–1263.
Walz, T. , Y. Fujiyoshi и A. Engel . 2009 . Структура AQP и функциональные последствия. Справочник. Эксп. Фармакол . 190 : 31–56.
Весселс, Д. , D. R. Soll , D. Knecht , , W. F. loomis , A. de lozanne и J. Spudich . 1988 . Подвижность клеток и хемотаксис у амеб Dictyostelium , лишенных тяжелой цепи миозина. Дев. Биол . 128 : 164–177.
Ву, Б. , К. Стейнбронн , М.Альстерфьорд , Т. Цойтен и Э. Бейтц . 2009 . Согласованное действие двух катионитовых фильтров в водном канале аквапорина. EMBO J . 28 : 2188–2194.
EMBO J . 28 : 2188–2194.
Ву, Б. , Дж. Сонг и Э. Бейтц . 2010 . Новые слитые белки канальных ферментов придают устойчивость к арсенатам. Дж. Биол. Химия . 285 : 40081–40087.
1
Yamamoto, E. , T. Akimoto , , Y. Hirano , M. Yasui и K. Yasuoka . 2014 . 1/ f колебания аминокислот регулируют транспорт воды в аквапорине 1. Phys. Преп. E Стат. Нонлин. Физика мягкой материи . 89 : 022718.
Zeuthen, T. , B. Wu , S.Павловик-Djuranovic , Л. М. Холм , Н. Л. Uzcategui , М. Duszenko , Дж Ф. Куна , Дж Е. Шульц и Е. Beitz . 2006 . Аммиачная проницаемость акваглицеропоринов из Plasmodium falciparum, Toxoplasma gondii и Trypansoma brucei. Мол. Микробиол . 61 : 1598–1608.
Мол. Микробиол . 61 : 1598–1608.
Функция Bax при отсутствии митохондрий у примитивных простейших Giardia lamblia
Abstract
Bax-индуцированная пермеабилизация наружной мембраны митохондрий и высвобождение цитохрома с являются ключевыми событиями апоптоза.Хотя Bax может скомпрометировать митохондрии в примитивных одноклеточных организмах, у которых отсутствует классический механизм апоптоза, до сих пор неясно, достаточно ли для этого одного Bax или требуются дополнительные митохондриальные компоненты. Протозойный паразит Giardia lamblia является одним из самых ранних ветвящихся эукариот и содержит сильно дегенерированные остатки митохондриальных органелл (митосомы), у которых отсутствует геном. Здесь мы проверили, можно ли использовать Bax человека, экспрессированный в Giardia , для удаления митосом.Мы демонстрируем, что эти органеллы не являются мишенями и не скомпрометированы Bax. Однако Bax полностью уничтожает специализированные отделы регулируемого секреторного пути. Как следствие, созревающие белки стенки кисты, которые сортируются в этих органеллах, высвобождаются в цитоплазму, вызывая остановку развития и гибель клеток. Интересно, что это эктопическое высвобождение груза зависит от карбоксиконцевых 22 аминокислот Bax и может быть предотвращено Bax-ингибирующим пептидом Ku70.Укороченный на С-конце вариант Bax все еще локализуется в секреторных органеллах, но неспособен проницать эти мембраны, разобщая нацеливание на мембраны и высвобождение груза. Несмотря на то, что митосомы слишком дивергированы, чтобы их можно было распознать с помощью Bax, нецелевая пермеабилизация мембран, по-видимому, сохраняется и приводит к гибели клеток совершенно независимо от митохондрий.
Как следствие, созревающие белки стенки кисты, которые сортируются в этих органеллах, высвобождаются в цитоплазму, вызывая остановку развития и гибель клеток. Интересно, что это эктопическое высвобождение груза зависит от карбоксиконцевых 22 аминокислот Bax и может быть предотвращено Bax-ингибирующим пептидом Ku70.Укороченный на С-конце вариант Bax все еще локализуется в секреторных органеллах, но неспособен проницать эти мембраны, разобщая нацеливание на мембраны и высвобождение груза. Несмотря на то, что митосомы слишком дивергированы, чтобы их можно было распознать с помощью Bax, нецелевая пермеабилизация мембран, по-видимому, сохраняется и приводит к гибели клеток совершенно независимо от митохондрий.
Образец цитирования: Hehl AB, Regos A, Schraner E, Schneider A (2007) Функция Bax при отсутствии митохондрий у примитивных простейших Giardia lamblia .ПЛОС ОДИН 2(5): е488. https://doi.org/10.1371/journal.pone.0000488
Академический редактор: Джулиан Резерфорд, Ньюкаслский университет, Соединенное Королевство
Получено: 26 апреля 2007 г. ; Принято: 5 мая 2007 г .; Опубликовано: 30 мая 2007 г.
; Принято: 5 мая 2007 г .; Опубликовано: 30 мая 2007 г.
Авторские права: © 2007 Hehl et al. Это статья с открытым доступом, распространяемая в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания оригинального автора и источника.
Финансирование: Швейцарский национальный научный фонд, Гранты №. 31-100270/1 и 3100A0-112327
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Одноклеточный кишечный паразит Giardia lamblia (син. G.intestalis, G.duodenalis ) является основной причиной диарейных заболеваний у людей и животных во всем мире [1]. Giardia принадлежит к одной из самых ранних ветвей эукариотической линии, известной на сегодняшний день [2], [3], и не имеет классического пути апоптоза; как каспазы, так и белки, подобные Bcl-2, отсутствуют.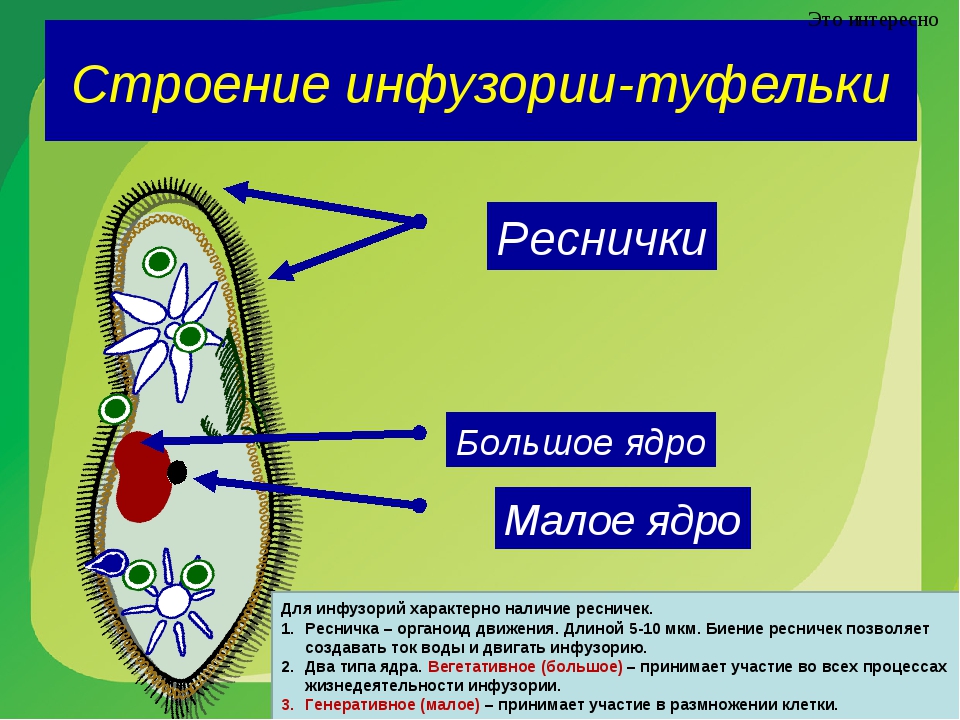 Кроме того, у Giardia отсутствуют добросовестные митохондрии, участвующие в окислительном фосфорилировании [1]. Недавно были идентифицированы и охарактеризованы митосомы, т.е. предполагаемые рудиментарные органеллы митохондриального происхождения [4]. Митосомы представляют собой небольшие, лишенные ДНК органеллы с двойной мембраной, которые участвуют в созревании железо-серных белков [5], [6], но не участвуют в энергетическом обмене. Митосомы лямблий лишены генома, но в ядерной ДНК обнаружены гены митохондриального происхождения [4]–[8].Все доступные данные подтверждают гипотезу о том, что митосомы являются рудиментарными органеллами, происхождение которых восходит к единственному эндосимбионтическому событию, приведшему к установлению митохондрий в эукариотической линии. Митосомы значительно уменьшились в размерах и утратили свои энергогенерирующие функции в ходе длительной независимой эволюции дипломонад [4]–[9], предположительно вследствие адаптации предка к микроаэрофильным нишам.
Кроме того, у Giardia отсутствуют добросовестные митохондрии, участвующие в окислительном фосфорилировании [1]. Недавно были идентифицированы и охарактеризованы митосомы, т.е. предполагаемые рудиментарные органеллы митохондриального происхождения [4]. Митосомы представляют собой небольшие, лишенные ДНК органеллы с двойной мембраной, которые участвуют в созревании железо-серных белков [5], [6], но не участвуют в энергетическом обмене. Митосомы лямблий лишены генома, но в ядерной ДНК обнаружены гены митохондриального происхождения [4]–[8].Все доступные данные подтверждают гипотезу о том, что митосомы являются рудиментарными органеллами, происхождение которых восходит к единственному эндосимбионтическому событию, приведшему к установлению митохондрий в эукариотической линии. Митосомы значительно уменьшились в размерах и утратили свои энергогенерирующие функции в ходе длительной независимой эволюции дипломонад [4]–[9], предположительно вследствие адаптации предка к микроаэрофильным нишам. Морфологически органеллы можно разделить на две отдельные популяции: небольшие сферические периферические митосомы (Pm), беспорядочно распределенные в цитоплазме, и удлиненные центральные митосомы (Cm), локализация которых зафиксирована в комплексе базальных телец в центре клетки [6]. ].Только Cm делится во время митоза и активно разделяется на дочерние клетки. Однако неизвестно, как возникают эти органеллы и как поддерживается их постоянный размер и количество [6].
Морфологически органеллы можно разделить на две отдельные популяции: небольшие сферические периферические митосомы (Pm), беспорядочно распределенные в цитоплазме, и удлиненные центральные митосомы (Cm), локализация которых зафиксирована в комплексе базальных телец в центре клетки [6]. ].Только Cm делится во время митоза и активно разделяется на дочерние клетки. Однако неизвестно, как возникают эти органеллы и как поддерживается их постоянный размер и количество [6].
Синтез и секреция защитного внеклеточного матрикса (стенки кисты), а также сопутствующие морфологические изменения необходимы для выживания инфекционных стадий Giardia в окружающей среде и, следовательно, для передачи [1]. Инцистирование in vitro или in vivo вызывается сигналами окружающей среды, например, концентрацией холестерина в содержимом кишечника.Секретируемый материал стенки кисты состоит как минимум из трех паралогичных белков стенки кисты (CWP1–3) [10]–[12] между 26 и 39 кДа и гомополимеров β(1–3) GalNAc [13]. CWP синтезируются в эндоплазматическом ретикулуме (ER) и концентрируются в больших, регулируемых развитием, подобных Гольджи компартментах, называемых энцистоспецифическими везикулами (ESV), где они подвергаются процессингу и олигомеризации перед секретированием. ESV возникают, по-видимому, de novo и содержат только предварительно отсортированный груз, предназначенный для стенки кисты [14]–[16].Учитывая их способность рекрутировать маркерные белки Гольджи и их чувствительность к брефельдину А, ESV, возможно, являются эволюционно ранней версией цистерн Гольджи, которые функционально ограничены посттрансляционным созреванием белков [17].
CWP синтезируются в эндоплазматическом ретикулуме (ER) и концентрируются в больших, регулируемых развитием, подобных Гольджи компартментах, называемых энцистоспецифическими везикулами (ESV), где они подвергаются процессингу и олигомеризации перед секретированием. ESV возникают, по-видимому, de novo и содержат только предварительно отсортированный груз, предназначенный для стенки кисты [14]–[16].Учитывая их способность рекрутировать маркерные белки Гольджи и их чувствительность к брефельдину А, ESV, возможно, являются эволюционно ранней версией цистерн Гольджи, которые функционально ограничены посттрансляционным созреванием белков [17].
Члены семейства белков Bcl-2 жестко контролируют митохондриальный путь апоптоза у высших эукариот и могут быть разделены на про- и антиапоптотические члены. Во время апоптоза проапоптотический член семейства Bcl-2 Bax перемещается из цитозоля в митохондрии и вызывает активацию каскада каспаз путем высвобождения цитохрома с из межмембранного пространства [18], [19]. Сигнал, запускающий активацию Bax, еще не идентифицирован. Тем не менее, сверхэкспрессии Bax в клетках млекопитающих было достаточно для нацеливания на митохондрии и высвобождения цитохрома с [20]. Кроме того, было показано, что Bax человека нацелен на митохондрии и приводит к гибели клеток у дрожжей [21] и трипаносом [22], а экспрессия меченного myc Bax вызывает цитоплазматическое высвобождение цитохрома с у дрожжей, подобное эффекту, наблюдаемому у млекопитающих. клетки [23]. Хотя Bax действует специфически на митохондрии in vivo, Bax сам по себе способен образовывать поры в искусственных липидных бислоях, подвергаемых воздействию рекомбинантного белка Bax, что было выявлено с помощью атомно-силовой микроскопии [24].Однако вопрос о том, достаточно ли одного Bax in vivo для индукции высвобождения цитохрома с, или он нуждается в дополнительных митохондриальных компонентах, является предметом споров [25], [26].
Сигнал, запускающий активацию Bax, еще не идентифицирован. Тем не менее, сверхэкспрессии Bax в клетках млекопитающих было достаточно для нацеливания на митохондрии и высвобождения цитохрома с [20]. Кроме того, было показано, что Bax человека нацелен на митохондрии и приводит к гибели клеток у дрожжей [21] и трипаносом [22], а экспрессия меченного myc Bax вызывает цитоплазматическое высвобождение цитохрома с у дрожжей, подобное эффекту, наблюдаемому у млекопитающих. клетки [23]. Хотя Bax действует специфически на митохондрии in vivo, Bax сам по себе способен образовывать поры в искусственных липидных бислоях, подвергаемых воздействию рекомбинантного белка Bax, что было выявлено с помощью атомно-силовой микроскопии [24].Однако вопрос о том, достаточно ли одного Bax in vivo для индукции высвобождения цитохрома с, или он нуждается в дополнительных митохондриальных компонентах, является предметом споров [25], [26].
Поскольку неясно, выполняют ли митосомы Giardia важные функции для паразита или они достаточно редуцированы, чтобы быть незаменимыми, мы условно экспрессировали Bax человека в Giardia в попытке скомпрометировать эти органеллы.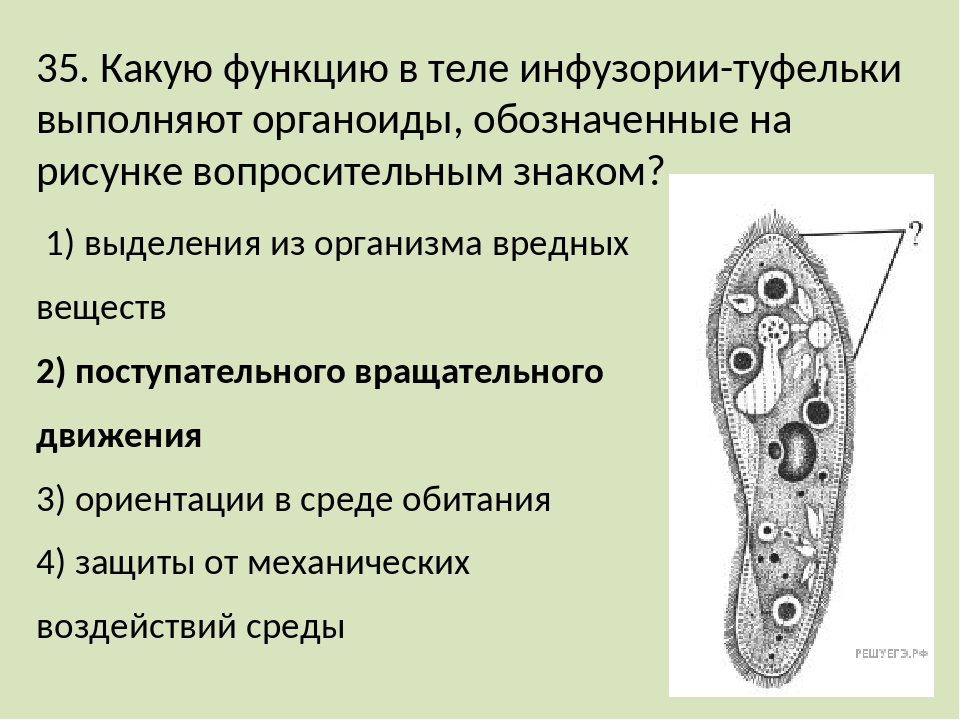 Эта стратегия основана на предположении, что органеллы будут достаточно консервативными, чтобы сделать возможным специфическое распознавание гетерологичного белка Bax.Однако мы обнаружили, что Bax физически не взаимодействует с митосомами и не влияет на них, что указывает на высокую степень дегенерации. Вместо этого случайное наблюдение показало, что в клетках, подвергающихся стадии дифференцировки, органеллы ESV были повреждены и потеряли свои грузовые белки. Здесь мы анализируем это необычное мембранное взаимодействие, которое впервые демонстрирует в живом организме общую способность белка Bax нацеливаться на компартментные мембраны и подвергать их риску в отсутствие дополнительных митохондриально-специфических факторов.
Эта стратегия основана на предположении, что органеллы будут достаточно консервативными, чтобы сделать возможным специфическое распознавание гетерологичного белка Bax.Однако мы обнаружили, что Bax физически не взаимодействует с митосомами и не влияет на них, что указывает на высокую степень дегенерации. Вместо этого случайное наблюдение показало, что в клетках, подвергающихся стадии дифференцировки, органеллы ESV были повреждены и потеряли свои грузовые белки. Здесь мы анализируем это необычное мембранное взаимодействие, которое впервые демонстрирует в живом организме общую способность белка Bax нацеливаться на компартментные мембраны и подвергать их риску в отсутствие дополнительных митохондриально-специфических факторов.
Материалы и методы
Giardia Культура, трансфекция и анализ трансгенных клеток Трофозоиты Giardia lamblia штамма WBC6 (каталог АТСС № 50803) выращивали в среде TYI-S-33 с добавлением 10% эмбриональной бычьей сыворотки и бычьей желчи. Для индукции экспрессии генов, находящихся под контролем промотора CWP1, применяли двухэтапный метод инцистирования, как описано ранее [27], путем повышения рН среды и добавления свиной желчи после культивирования в течение примерно 44 ч в среде без желчи.Для исследований ингибирования Bax пептидный ингибитор Ku70 (Calbiochem, Сан-Диего, Калифорния) добавляли вместе со средой для инцистирования в конечной концентрации 200 мкМ.
Для индукции экспрессии генов, находящихся под контролем промотора CWP1, применяли двухэтапный метод инцистирования, как описано ранее [27], путем повышения рН среды и добавления свиной желчи после культивирования в течение примерно 44 ч в среде без желчи.Для исследований ингибирования Bax пептидный ингибитор Ku70 (Calbiochem, Сан-Диего, Калифорния) добавляли вместе со средой для инцистирования в конечной концентрации 200 мкМ.
Плазмидную ДНК, содержащую интересующие гены и селектируемые маркеры, электропорировали в трофозоиты, а стабильные трансгенные клетки отбирали с использованием антибиотика G418 (Sigma, Сент-Луис, Миссури), как описано ранее [27]. Плазмиды сохраняли эписомально при непрерывном отборе антибиотиков.
Все экспрессионные конструкции были основаны на кассете C1-CWP для экспрессии под контролем индуцибельного промотора CWP1 [27].Полноразмерную или укороченную с С-конца ОРС Bax человека амплифицировали с помощью ПЦР из плазмиды [22] с использованием праймеров (ориентация от 5′ к 3′) ATGCATGACGGGTCCGGGGAG (смысловой), TTAATTAATCAGCCCATCTTCTTCCAGAT (полноразмерный антисмысловой), CCTTAATTAATCACCACGTGGGCGTCCCAAAG (укороченный антисмысловой) вырезали и лигировали в сайты Nsi I и Pac I кассеты экспрессии.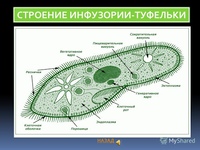
Подготовка клеток к микроскопии – конфокальная микроскопия:
инцистирующих трофозоита через четыре часа после индукции фиксировали для иммунофлуоресцентной микроскопии в 3% формальдегиде (все химические вещества, приобретенные в Fluka, Buchs, Швейцария, если не указано иное), пермеабилизировали с помощью 0.2% Triton X-100 и блокировали в 2% растворе бычьего сывороточного альбумина/фосфатно-солевого буфера (PBS). CWP был обнаружен с помощью конъюгированного с Texas Red моноклонального антитела против CWP (Waterborne Inc., Новый Орлеан, Луизиана) в качестве маркера для ESV. Рекомбинантный Bax был обнаружен с помощью моноклонального антитела (NeoMarkers, Fremont, CA), а затем вторичного конъюгированного с Alexa488 антитела против IgG мыши (Invitrogen AG, Базель, Швейцария). Для мечения митосом использовали кроличью антисыворотку против IscS (любезный подарок доктора Дж. Товара), а затем вторичное антитело, конъюгированное с Alexa594 (Invitrogen, Базель, Швейцария).Конфокальную микроскопию выполняли на установке Leica SP2 AOBS (Leica Microsystems, Wetzlar, Германия).
Электронная микроскопия:
Клетки регистрировали при увеличении в 5000 раз с использованием электронного микроскопа СМ 12 (Philips, Нидерланды). Инцистирующие трофозоиты готовили для электронной микроскопии, как описано ранее [14], и позволяли активно прикрепляться к сапфировым дискам для получения однородно ориентированных паразитов для фиксации и последующего разрезания в дорсо-вентральной плоскости.Прикрепленные трофозоита Giardia были подвергнуты сверхбыстрому замораживанию перед фиксацией для оптимального сохранения мембранных структур.
Результаты и обсуждение
Митосомы Giardia не являются мишенью для рекомбинантного BaxНаличие и поддержание митосом в Giardia остается загадкой. В частности, неясно, выполняют ли они все еще какую-либо важную функцию или фактически устарели в ходе их значительного эволюционного сокращения.Мы попытались нацелиться на эти органеллы и удалить их, используя порообразующий белок Bax, и оценить жизнеспособность паразитов. Эта стратегия оказалась успешной у эволюционного базального простейшего Trypanosoma brucei , у которого также отсутствует механизм апоптоза, и привела к высвобождению цитохрома с, фрагментации митохондрий и гибели клеток [22]. Для экспрессии человеческого гена BAX в Giardia мы сконструировали плазмидный вектор, содержащий полноразмерную открытую рамку считывания под контролем индуцибельного промотора CWP1, который жестко репрессирован и активируется только в трофозоитах, индуцированных для инцистации [27]. ].Субклеточную локализацию рекомбинантного белка определяли с помощью конфокальной лазерной сканирующей микроскопии трансгенных паразитов после фиксации и пермеабилизации детергентом. Рекомбинантный Bax был обнаружен с помощью специфического антитела в виде точечной картины в цитоплазме, напоминающей периферическое распределение митосом (рис. 1А, левая панель). Удивительно, но исследования двойной иммунофлуоресценции не выявили совместной локализации с маркерным белком митосомного матрикса IscS (рис. 1А, средняя панель).Что наиболее важно, Bax не локализуется в сигнатурной структуре митосом, центральной митосоме (Cm), в базальном комплексе тельца в центре клетки (стрелка). Это свидетельствует о том, что Bax вообще не взаимодействует с этими органеллами, а либо образует агрегаты в цитоплазме, либо прикрепляется к неизвестным структурам клетки.
Рисунок 1. Экспрессия Bax человека в индуцированных трофозоитах Giardia не влияет на митосомы.
A) Анализ конфокальной микроскопии внутриклеточного распределения Bax в трансгенных клетках.Рекомбинантный Bax (левая панель, зеленый цвет) не локализуется в митосомах (средний график, красный цвет), помеченных кроличьей антисывороткой против IscS в качестве митосомного маркера. Характерная структура центральной митосомы указана стрелкой. Объединенные изображения показывают явное отсутствие совместной локализации. Ядерная ДНК окрашивается DAPI (синий). Вставка: дифференциально-интерференционно-контрастное (ДИК) изображение. Масштабная линейка: 5 мкм. B) Центральная структура митосомы (Cm, наконечник стрелки) определяется как плотно упакованный массив сферических органелл при электронной микроскопии прикрепленных клеток.Субъединицы неотличимы от отдельных периферических митосом (Pm, стрелка). Эта структура органелл неизменна у трансгенных трофозоитов, экспрессирующих Bax. Н, ядро; Bb, базальные тела; топор, аксонемы; Vd, вентральный диск.
https://doi.org/10.1371/journal.pone.0000488.g001
Центральная митосома (Cm) представляет собой характерную структуру, которая выглядит как удлиненная органелла между ядрами на иммунофлуоресцентных микрофотографиях и точно делится и разделяется во время клеточное деление [6].Хотя экспрессия Bax, по-видимому, не влияла на Cm (рис. 1А), незначительные изменения могли остаться незамеченными. Морфология центральной митосомы не была определена с помощью электронной микроскопии, потому что очень трудно получить подходящим образом ориентированный срез клеточного центра в произвольно разделенных клеточных осадках. Чтобы ответить на эти вопросы, мы использовали ранее применявшуюся технику срезов равномерно ориентированных прилипших трофозоитов в плоскости, перпендикулярной дорсально-вентральной оси [28], для анализа с помощью электронной микроскопии.При таком подходе можно часто находить продольные срезы через клеточный центр, что позволяет систематически исследовать митосомные структуры в диком типе Giardia и трансгенных клетках, экспрессирующих Bax. На рис. 1В показано репрезентативное изображение, на котором центральная митосома видна как плотно упакованный кластер небольших сферических органелл (стрелка), очень похожий на периферические митосомы в цитоплазме (стрелка). Сгруппированные органеллы встроены в базальное тело — пучок аксонем, что согласуется с наблюдаемым верным разделением на дочерние клетки, поскольку это одна из первых клеточных структур, разделяющихся в делящихся клетках [29], [30].В клетках, экспрессирующих Bax, морфология центральной митосомы неразличима (данные не показаны). Вместе с отсутствием нацеливания Bax на эти органеллы это свидетельствует против прямого взаимодействия Bax с митосомами и указывает на высокую степень дегенерации во время эволюционной редукции. Как и у Giardia , классические каспазозависимые пути апоптоза и белки семейства Bcl-2 отсутствуют у других базальных эукариот, таких как простейшие Trypanosoma brucei .Тем не менее, трипаносомы и родственный Leishmania могут подвергаться форме запрограммированной гибели клеток [31]–[33] в ответ на внешние раздражители. Кроме того, условная экспрессия Bax человека в T. brucei приводит к динамин-зависимой фрагментации одиночной митохондрии и высвобождению цитохрома с из межмембранного пространства [22], [34]. Это указывает на то, что факторы, необходимые для нацеливания Bax на цитоплазматическую сторону митохондриальной мембраны и для высвобождения цитохрома с, остаются консервативными у Т.brucei , даже если остальные механизмы отсутствуют. Запрограммированная гибель клеток также была постулирована для Giardia , хотя прямые доказательства отсутствуют, а физиологическая роль неизвестна. Тот факт, что Bax неспособен нацеливаться на митосомы лямблий, согласуется с сильно дивергированным статусом этих органелл.
Экспрессия Bax влияет на жизнеспособность клеток и развитие секреторных органелл
Несмотря на то, что экспрессия Bax не оказывала явного влияния на митосомы, большая часть индуцированных трансгенных паразитов Giardia не смогла завершить дифференцировку и погибла.Через несколько часов после индукции клетки теряли подвижность, способность прикрепляться к поверхности культуральных пробирок и лизировались. Микроскопическая количественная оценка этого эффекта через 6 часов после индукции показала, что количество жизнеспособных трансгенных клеток было снижено примерно на 85% по сравнению с идентично индуцированным родительским штаммом дикого типа (рис. 2А). Клетки оценивали по морфологии, подвижности и способности прикрепляться к пластиковому субстрату. Этот эффект коррелировал с повышенной экспрессией мРНК Bax (рис. 2А, вставка).
Рисунок 2. Экспрессия Bax летальна для Giardia.
А) Индуцированные клетки, экспрессирующие Bax, перестают делиться и погибают в течение 6 часов. Врезка: полуколичественный RT-PCR анализ уровней мРНК Bax в трансгенных клетках до индукции (PI) и через 4 часа после индукции. B) Субклеточная локализация CWP в ESV инцистирующих клеток дикого типа, меченных моноклональным антителом против CWP, через 4 часа после индукции. C) Bax подавляет образование ESV в инцистирующих клетках и вызывает накопление CWP в цитоплазме и нуклеоплазме.Вставка: дифференциально-интерференционно-контрастное (ДИК) изображение. Масштабная линейка: 5 мкм.
https://doi.org/10.1371/journal.pone.0000488.g002
В индуцированных трофозоитах Giardia , подвергающихся стадии дифференцировки, белки стенки кисты накапливаются и посттрансляционно созревают в больших ESV перед регулируемой секрецией на поверхность клетки на завершающей стадии этого процесса. Чтобы определить, была ли гибель трансгенных клеток напрямую связана с инцистированием, мы исследовали, влияет ли Bax на образование ESV.Чтобы оценить инцистацию в присутствии Bax, мы сравнили образование ESV у дикого типа и трансгенных Giardia с помощью конфокальной иммунофлуоресцентной микроскопии с использованием антитела против CWP1, основного грузового белка ESV. Локализация и появление нормальных ESV в клетках дикого типа через 4 часа после индукции показаны на рисунке 2B. На этой стадии в индуцированных клетках отчетливо проявляется синтез в ER и концентрация белков стенки кисты в растущих ESV. Удивительно, но в трансгенных клетках, экспрессирующих Bax, мы не обнаружили ESV.Вместо этого CWP был обнаружен в цитозоле и, по-видимому, накапливался также в ядрах (рис. 2C). Самое простое объяснение заключалось в том, что Bax нацелился на ESV и скомпрометировал их, выпустив при этом свой груз. На какой стадии развития ESV становятся восприимчивыми, определить не удалось. Однако отсутствие этих органелл в Bax-экспрессирующих клетках даже на ранних стадиях инцистации (2-4 ч) указывает на то, что возникающие ESV становятся мишенями Bax очень рано.
Вредные эффекты на дифференцировку
Giardia связаны с консервативными функциональными доменами BaxМы попытались определить, были ли очевидная абляция органелл ESV и сопутствующее высвобождение груза следствием Bax-специфического механизма или просто следствием избыточной экспрессии.Чтобы проверить это, мы обработали трансгенные клетки Bax-ингибирующим пептидом Ku70 [35] и проанализировали влияние на образование ESV после индукции. В отличие от полной абляции ESV в необработанном контроле, органеллы, содержащие материал стенки кисты, развивались в присутствии пептида, и проникновение CWP в цитоплазму и ядра было значительно уменьшено (рис. 3A). Это указывало на то, что ESV были, по крайней мере, частично защищены, указывая на специфическое ингибирование Bax, аналогичное специфическим эффектам Ku70 у высших эукариот.Что еще более важно, результирующая стабилизация ESV теперь выявила локализацию рекомбинантного Bax в этих органеллах. Это также подтверждает идею о том, что быстрая и полная абляция ESV в необработанных клетках исключает прямое наблюдение за нацеливанием Bax. Таким образом, смягчение этого эффекта добавлением Ku70 свидетельствует о способности рекомбинантного Bax связываться с мембранами ESV. Однако, в отличие от клеток млекопитающих, пептид-ингибитор не отменяет нацеливания Bax на мембрану [35]. Чтобы более подробно исследовать специфичность Bax для мембран ESV, мы экспрессировали мутантный вариант, названный BaxΔ22, в котором отсутствуют 22 C-концевые аминокислоты белка, который, как считается, участвует в образовании пор.Клетки, экспрессирующие BaxΔ22, были полностью компетентны в инцистировании и способны образовывать ESV. Без своего карбокси-конца Bax все еще локализован на мембранах ESV, но высвобождения CWP в цитоплазму не наблюдалось (рис. 3B). В совокупности это продемонстрировало полное разобщение нацеливания на мембрану и высвобождение груза в этой системе, что согласуется с результатами, полученными в Bax-экспрессирующих дрожжах и в клетках млекопитающих, демонстрируя, что нацеливание Bax на органеллы сохраняется в отсутствие его C-конца [36]. , [37].Последнее, однако, все еще остается спорным, так как другое исследование показало, что гидрофобный С-конец Bax действует как митохондриально-нацеленный сигнал [38].
Рисунок 3. Высвобождение CWP от ESV зависит от Bax.
A) ESV в инцистирующих Bax-экспрессирующих клетках, обработанных мембранопроницаемым Bax-ингибирующим пептидом Ku70, по крайней мере частично защищены. Обратите внимание на совместную локализацию Bax (зеленый) с (частично интактными) ESV и значительно меньший цитоплазматический или нуклеоплазматический сигнал CWP, чем в необработанных клетках (сравните с фиг.2C).16-May B) Делеция С-конца Bax разъединяет нацеливание Bax на мембраны ESV и высвобождение CWP. В BaxΔ22 отсутствуют С-концевые 22 аминокислоты Bax, и он локализуется на мембранах интактного ESV. C, D) Электронные микрофотографии репрезентативных клеток, экспрессирующих Bax или BaxΔ22, через четыре часа после индукции. C) Инцистирующий трофозоит из популяции, экспрессирующей BaxΔ22, демонстрирующий многочисленные ESV с электронно-плотным материалом (стрелки). ESV и общая морфология компартментов неотличимы от клеток дикого типа (не показаны).Увеличенная область показывает отдельный ESV. Хорошо видны периферические везикулы, лежащие под плазматической мембраной. D) ESV или остатки органелл отсутствуют в выживших клетках, экспрессирующих Bax. Видны ядра и части структур микротрубочек передних жгутиков и значительно увеличенные ЛВ. Н, ядра. Шкала баров: 2 мкм.
https://doi.org/10.1371/journal.pone.0000488.g003
Хотя данные конфокальной микроскопии предполагают, что груз ESV высвобождается в цитозоль в результате экспрессии Bax, судьба этих органелл остается неясной.Чтобы более подробно изучить влияние Bax на целостность компартмента ESV, мы провели просвечивающую электронную микроскопию тонких срезов, сравнивая клетки через 4 часа после индукции, экспрессирующие либо Bax, либо BaxΔ22 (рис. 3). Обширное исследование однородно ориентированных и срезов Giardia показало, что >80% клеток, экспрессирующих BaxΔ22, содержат ESV с ультраструктурой, идентичной таковой в клетках дикого типа (рис. 3C, D) [28]. Напротив, в клетках, экспрессирующих полноразмерный Bax, практически не было ESV (рис. 3D).Кроме того, не наблюдалось необычных мембранных структур, которые можно было бы интерпретировать как «пустые» ESV. Единственной аномалией было значительное увеличение размера периферических пузырьков (ПВ). PVs составляют отчетливо локализованную эндосомально-лизосомальную систему в Giardia , которая располагается ниже плазматической мембраны и участвует в эндоцитарных и экзоцитарных процессах [39]. Клетки, экспрессирующие Bax, содержали увеличенные PV (диаметр: ~ 0,5–1 мкм) (рис. 3D), в отличие от инцистирующих паразитов дикого типа [28] или трансгенных клеток, экспрессирующих BaxΔ22 (диаметр: ~ 100–150 нм).Это говорит о том, что PV могут участвовать в удалении лизированных ESV. Т.о., одним из объяснений полного отсутствия ESVs в трансгенных клетках, экспрессирующих Bax, может быть то, что процесс, подобный аутофагии, устраняет эти компартменты после потери груза.
Таким образом, мы показываем, что в отсутствие bona fide митохондрий в ранней дивергенции Giardia условно экспрессируемый Bax человека нацелен на ESVs. Одного Bax достаточно, чтобы скомпрометировать эти органеллы, по-видимому, по тому же механизму, что и для митохондриальных мембран у высших эукариот, т.е.д., образование пор. В соответствии с этим мы наблюдали высвобождение CWP в цитоплазму. Подобно высвобождению цитохрома с из митохондрий, этот процесс зависит от карбоксиконца Bax [38] и может частично ингибироваться пептидом Ku70. Т.о., эктопическая экспрессия Bax при инцистировании Giardia приводит к нацеливанию на специфические органеллы, высвобождению груза и гибели клеток совершенно независимо от митохондрий.
Благодарности
Мы благодарны доктору Дж.Товара за подарок антитела против IscS, доктора П. Уайлда за подготовку клеток для ЭМ-анализа и Т. Мишеля за отличную техническую помощь.
Авторские взносы
Идея и дизайн экспериментов: AS AH AR. Выполнены опыты: АХ АР ЭС. Проанализированы данные: АХ АР. Написал статью: AS AH.
Каталожные номера
- 1. Адам Р.Д. (2001) Биология Giardia lamblia. Clin Microbiol Rev 14: 447–475.
- 2. Хеджес С.Б., Чен Х., Кумар С., Ван Д.Ю., Томпсон А.С. и др.(2001) Геномная шкала времени происхождения эукариот. БМС Эвол Биол 1:4.
- 3. Согин М.Л., Гандерсон Дж.Х., Элвуд Х.Дж., Алонсо Р.А., Питти Д.А. (1989) Филогенетическое значение концепции царства: необычная рибосомная РНК из Giardia lamblia. Наука 243: 75–77.
- 4. Товар Дж., Леон-Авила Г., Санчес Л.Б., Сутак Р., Тачези Дж. и др. (2003) Митохондриальные остатки органелл Giardia функционируют при созревании железо-серного белка. Природа 426: 172–176.
- 5.Долезал П., Смид О., Рада П., Зубакова З., Бурсак Д. и соавт. (2005) Митосомы Giardia и гидрогеносомы трихомонад имеют общий способ нацеливания на белок. Proc Natl Acad Sci USA 102: 10924–10929.
- 6. Регоес А., Зурмпану Д., Леон-Авила Г., ван дер Гизен М., Товар Дж. и др. (2005)Импорт белка, репликация и наследование рудиментарной митохондрии. J Biol Chem 280: 30557–30563.
- 7. Роджер А.Дж., Свард С.Г., Товар Дж., Кларк К.Г., Смит М.В. и др.(1998) Митохондриальноподобный ген шаперонина 60 у Giardia lamblia: свидетельство того, что дипломонады когда-то содержали эндосимбионт, связанный с прародителем митохондрий. Proc Natl Acad Sci U S A 95: 229–234.
- 8. Арисуэ Н., Санчес Л.Б., Вайс Л.М., Мюллер М., Хашимото Т. (2002)Гены hsp70 митохондриального типа амитохондриальных протистов, лямблии кишечной, Entamoeba histolytica и двух микроспоридий. Паразитол Инт 51: 9–16.
- 9. Зильберман Дж. Д., Симпсон А. Г., Кулда Дж., Чепицка И., Хэмпл В. и др.(2002) Жгутиконосцы ретортамонад тесно связаны с дипломонадами — последствия для истории митохондриальной функции в эволюции эукариот. Мол Биол Эвол 19: 777–786.
- 10. Sun CH, McCaffery JM, Reiner DS, Gillin FD (2003) Изучение генома Giardia lamblia для новых белков стенки кисты. J Biol Chem 278: 21701–21708.
- 11. Lujan HD, Mowatt MR, Conrad JT, Bowers B, Nash TE (1995)Идентификация нового белка стенки кисты Giardia lamblia с повторами, богатыми лейцином.Влияние на образование секреторных гранул и сборку белков в стенке кисты. J Biol Chem 270: 29307–29313.
- 12. Моватт М.Р., Лужан Х.Д., Коттен Д.Б., Бауэрс Б., Йи Дж. и др. (1995)Регулируемая развитием экспрессия гена белка стенки кисты Giardia lamblia. Мол микробиол 15: 955–963.
- 13. Gerwig GJ, van Kuik JA, Leeflang BR, Kamerling JP, Vliegenthart JF, et al. (2002) Стенка нитчатой кисты Giardia кишечная содержит новый полимер бета (1-3)-N-ацетил-D-галактозамин: структурное и конформационное исследование.Гликобиология 12: 499–505.
- 14. Марти М., Ли Ю., Шранер Э.М., Уайлд П., Колер П. и др. (2003) Секреторный аппарат древнего эукариота: сортировка белков по отдельным путям экспорта происходит до образования временных Golgi-подобных компартментов. Мол Биол Ячейка 14: 1433–1447.
- 15. Reiner DS, McCaffery M, Gillin FD (1990)Сортировка белков стенки кисты по регулируемому секреторному пути во время дифференцировки примитивного эукариота Giardia lamblia.Eur J Cell Biol 53: 142–153.
- 16. Lujan HD, Marotta A, Mowatt MR, Sciaky N, Lippincott-Schwartz J, et al. (1995) Индукция развития структуры и функции Гольджи у примитивных эукариот Giardia lamblia. J Biol Chem 270: 4612–4618.
- 17. Марти М., Хель А.Б. (2003)Везикулы, специфичные для инцистирования в Giardia: изначальный Гольджи или просто еще один секреторный отсек? Тенденции Параситол 19: 440–446.
- 18. Desagher S, Martinou JC (2000)Митохондрии как центральная контрольная точка апоптоза.Тенденции Cell Biol 10: 369–377.
- 19. Ferri KF, Kroemer G (2001)Органелл-специфическое инициирование путей гибели клеток. Nat Cell Biol 3: E255–263.
- 20. Эскес Р., Антонссон Б., Осен-Санд А., Монтессуит С., Рихтер С. и др. (1998) Bax-индуцированное высвобождение цитохрома C из митохондрий не зависит от поры перехода проницаемости, но сильно зависит от ионов Mg2+. J Cell Biol 143: 217–224.
- 21. Жа Х., Фиск Х.А., Яффе М.П., Махаджан Н., Герман Б. и др.(1996) Сравнение структуры и функции проапоптотического белка Bax в клетках дрожжей и млекопитающих. Мол Селл Биол 16: 6494–6508.
- 22. Esseiva AC, Chanez AL, Bochud-Allemann N, Martinou JC, Hemphill A, et al. (2004)Временное рассечение Bax-индуцированных событий, приводящих к делению одиночной митохондрии у Trypanosoma brucei. EMBO Rep 5: 268–273.
- 23. Manon S, Chaudhuri B, Guerin M (1997) Высвобождение цитохрома с и снижение активности цитохрома с оксидазы в Bax-экспрессирующих дрожжевых клетках и предотвращение этих эффектов путем коэкспрессии Bcl-xL.FEBS Lett 415: 29–32.
- 24. Epand RF, Martinou JC, Montessuit S, Epand RM, Yip CM (2002) Прямые доказательства образования мембранных пор апоптотическим белком Bax. Biochem Biophys Res Commun 298: 744–749.
- 25. Замзами Н., Кремер Г. (2001) Митохондрия в апоптозе: как открывается ящик Пандоры. Nat Rev Mol Cell Biol 2: 67–71.
- 26. Мартину Дж. К., Грин Д. Р. (2001) Преодоление митохондриального барьера. Nat Rev Mol Cell Biol 2: 63–67.
- 27. Хель А.Б., Марти М., Колер П. (2000)Этап-специфическая экспрессия и нацеливание на химеры белка стенки кисты и зеленого флуоресцентного белка в Giardia. Мол Биол Ячейка 11: 1789–1800.
- 28. Марти М., Регос А., Ли И., Шранер Э.М., Уайлд П. и др. (2003)Наследственный секреторный аппарат простейшего паразита Giardia кишечная. J Biol Chem 278: 24837–24848.
- 29. Нохынкова Е., Тумова П., Кулда Дж. (2006) Деление клеток Giardia кишечной: цикл развития жгутиков включает трансформацию и обмен жгутиками между мастигонтами клетки дипломонады.Эукариотическая клетка 5: 753–761.
- 30. Саголла М.С., Доусон С.К., Манкузо Дж.Дж., Канде В.З. (2006)Трехмерный анализ митоза и цитокинеза двуядерного паразита Giardia кишечная. J Cell Sci 119: 4889–4900.
- 31. Ameisen JC, Idziorek T, Billaut-Mulot O, Loyens M, Tissier JP, et al. (1995) Апоптоз у одноклеточного эукариота (Trypanosoma cruzi): последствия для эволюционного происхождения и роли запрограммированной гибели клеток в контроле клеточной пролиферации, дифференцировки и выживания.Различие клеточной смерти 2: 285–300.
- 32. Ли Н., Бертолет С., Дерабант А., Мюллер Дж., Дункан Р. и др. (2002)Запрограммированная гибель клеток у одноклеточного простейшего паразита Leishmania. Отличие клеточной смерти 9: 53–64.
- 33. Фигарелла К., Равер М., Ускатеги Н.Л., Кубата Б.К., Лаубер К. и соавт. (2005)Простагландин D2 индуцирует запрограммированную гибель клеток в форме кровотока Trypanosoma brucei. Cell Death Differ 12: 335–346.
- 34. Chanez AL, Hehl AB, Engstler M, Schneider A (2006) Абляция одиночного динамина T.brucei блокирует деление митохондрий и эндоцитоз и приводит к точной остановке цитокинеза. J Cell Sci 119: 2968–2974.
- 35. Савада М., Хейс П., Мацуяма С. (2003)Цитопротекторные мембранопроницаемые пептиды, сконструированные из Bax-связывающего домена Ku70. Nat Cell Biol 5: 352–357.
- 36. Priault M, Cartron PF, Camougrand N, Antonsson B, Vallette FM и др. (2003) Исследование роли С-конца Bax и tc-Bid во взаимодействии Bax с митохондриями дрожжей.Cell Death Differ 10: 1068–1077.
- 37. Картрон П.Ф., Моро С., Оливер Л., Маят Э., Мефлах К. и др. (2002) Участие N-конца Bax в его внутриклеточной локализации и функции. FEBS Lett 512: 95–100.
- 38. Nechushtan A, Smith CL, Hsu YT, Youle RJ (1999) Конформация С-конца Bax регулирует субклеточное расположение и гибель клеток. Эмбо J 18: 2330–2341.
- 39. Lanfredi-Rangel A, Attias M, de Carvalho TM, Kattenbach WM, De Souza W (1998) Периферические везикулы трофозоитов примитивных простейших Giardia lamblia могут соответствовать ранним и поздним эндосомам и лизосомам.J Struct Biol 123: 225–235.
Каковы функции простейших? – idswater.com
Каковы функции простейших?
Клетка простейших осуществляет все процессы, включая питание, рост, размножение, выделение и движение, необходимые для поддержания и размножения жизни. Клетка заключена в мембрану, называемую плазматической мембраной.
Каковы структуры простейших?
Структура простейших Это одноклеточные эукариоты.И точно так же, как у других эукариотических клеток, ядро окружено мембраной. Кроме того, ядро везикулярное, с рассеянным хроматином, что придает ядру диффузный вид. Кроме того, все ядра в отдельном организме выглядят одинаково.
Что характерно для простейших?
Характеристики простейших:
- Не имеют клеточной стенки; однако некоторые из них обладают гибким слоем, пленкой или жесткой оболочкой из неорганических материалов снаружи клеточной мембраны.
- Они обладают способностью в течение всего своего жизненного цикла или его части передвигаться с помощью локомоторных органелл или механизма скольжения.
Каковы преимущества простейших?
Полезные простейшие:
- Простейшие служат пищей для многих мелких водных организмов. Зоопланктон — это крошечные простейшие, обитающие в море.
- Они являются первоклассными редуцентами в природе, так как питаются бактериями и грибами, разлагающими мертвые органические вещества.
- Некоторые простейшие живут в теле других организмов и помогают им.
Что такое простейшие короткий ответ?
Простейшие — одноклеточные организмы. Они бывают разных форм и размеров: от амебы, которая может менять свою форму, до парамеция с фиксированной формой и сложной структурой. Они живут в самых разнообразных влажных средах обитания, включая пресную воду, морскую среду и почву.
Что такое простейшие простыми словами?
Простейшие — мелкие (но не простые) организмы. Это одноклеточные гетеротрофные эукариоты, которые питаются бактериями и другими источниками пищи.Многие виды простейших являются симбионтами, некоторые — паразитами, а некоторые — хищниками почвенных бактерий и водорослей.
Какие 4 типа простейших существуют?
Для наших целей здесь будут рассмотрены только 4 группы простейших: эти группы разделены по подвижности и клеточной структуре.
- Амебы (представитель: Ameba proteus)
- Жгутиконосцы (представители: Trypanosoma, Euglena)
- Инфузории (представитель: Paramecium)
- Apicomplexa (представитель: Plasmodium)
Кому полезны простейшие?
Некоторые простейшие живут в теле других организмов и помогают им.У термитов, например, в теле живут простейшие. Простейшие переваривают целлюлозу из древесины, поедаемой термитами, и превращают ее в углеводы, которые термиты могут использовать.
В чем преимущество и недостаток простейших?
✔В отличие от бактерий, простейшие могут изменять белки. ✔ Их легко изучать, потому что они принадлежат к одной классификации/группе. ✔ С ними легко работать в лаборатории. Их можно легко сравнить и противопоставить другим видам.
| |
%PDF-1.4 % 925 0 объект > эндообъект внешняя ссылка 925 120 0000000016 00000 н 0000004338 00000 н 0000004567 00000 н 0000004603 00000 н 0000005356 00000 н 0000005494 00000 н 0000005642 00000 н 0000005778 00000 н 0000005928 00000 н 0000006065 00000 н 0000006213 00000 н 0000006351 00000 н 0000006501 00000 н 0000006639 00000 н 0000006789 00000 н 0000006926 00000 н 0000007076 00000 н 0000007214 00000 н 0000007364 00000 н 0000007501 00000 н 0000007649 00000 н 0000007786 00000 н 0000007934 00000 н 0000008074 00000 н 0000008224 00000 н 0000008364 00000 н 0000008512 00000 н 0000008652 00000 н 0000008801 00000 н 0000008941 00000 н 0000009091 00000 н 0000009228 00000 н 0000009376 00000 н 0000009515 00000 н 0000009662 00000 н 0000009801 00000 н 0000009948 00000 н 0000010106 00000 н 0000010255 00000 н 0000010781 00000 н 0000011042 00000 н 0000011483 00000 н 0000011770 00000 н 0000011807 00000 н 0000012193 00000 н 0000012919 00000 н 0000013671 00000 н 0000013879 00000 н 0000014317 00000 н 0000014377 00000 н 0000014741 00000 н 0000017593 00000 н 0000017893 00000 н 0000018281 00000 н 0000018759 00000 н 0000019246 00000 н 0000022457 00000 н 0000022805 00000 н 0000023041 00000 н 0000026186 00000 н 0000026237 00000 н 0000029841 00000 н 0000033020 00000 н 0000033084 00000 н 0000033143 00000 н 0000036047 00000 н 0000039192 00000 н 0000041661 00000 н 0000044354 00000 н 0000046748 00000 н 0000050930 00000 н 0000058213 00000 н 0000061998 00000 н 0000063164 00000 н 0000068041 00000 н 0000068476 00000 н 0000071927 00000 н 0000078170 00000 н 0000084120 00000 н 0000084176 00000 н 0000084223 00000 н 0000084269 00000 н 0000084323 00000 н 0000084868 00000 н 0000085004 00000 н 0000134672 00000 н 0000134713 00000 н 0000135266 00000 н 0000135409 00000 н 0000170318 00000 н 0000170359 00000 н 0000170803 00000 н 0000170915 00000 н 0000171430 00000 н 0000171542 00000 н 0000171617 00000 н 0000171742 00000 н 0000171873 00000 н 0000172036 00000 н 0000172205 00000 н 0000172358 00000 н 0000172515 00000 н 0000172662 00000 н 0000172839 00000 н 0000173095 00000 н 0000173353 00000 н 0000173530 00000 н 0000173729 00000 н 0000174011 00000 н 0000174200 00000 н 0000174415 00000 н 0000174665 00000 н 0000174906 00000 н 0000175079 00000 н 0000175266 00000 н 0000175455 00000 н 0000175690 00000 н 0000175899 00000 н 0000176124 00000 н 0000002696 00000 н трейлер ]/предыдущая 1144450>> startxref 0 %%EOF 1044 0 объект >поток h-TLSW>h/—uRP( (UBo=@$PdɄშs 8adD,(*.