102. Обыкновенная амеба заслуживает необыкновенного памятника: albercul — LiveJournal
Ползёт амёба, да по субстрату
И машет псевдоподией.
Какое горе, какое горе
Такою быть уродиной.
…………………………………
Вот так ползла, ползла амеба,
Пугая всех вокруг себя,
И наконец, вдруг цистой стала —
Такая вот ее судьба.
Великий симбиоз,
или как призошла эукариотная клетка
Более ста лет назад российские биологи А.С. Фаминцын и К.С. Мережковский, а затем и Б.М. Козо-Полянский сформулировали симбигенетическую концепцию происхождения живой клетки. Согласно этой идее, эукариотная клетка произошла за счет симбиоза нескольких различных одноклеточных организмов. Но как, в силу каких причин и обстоятельств такое могло произойти в эволюции биосферы? Каким образом одни клетки обосновались внутри других?
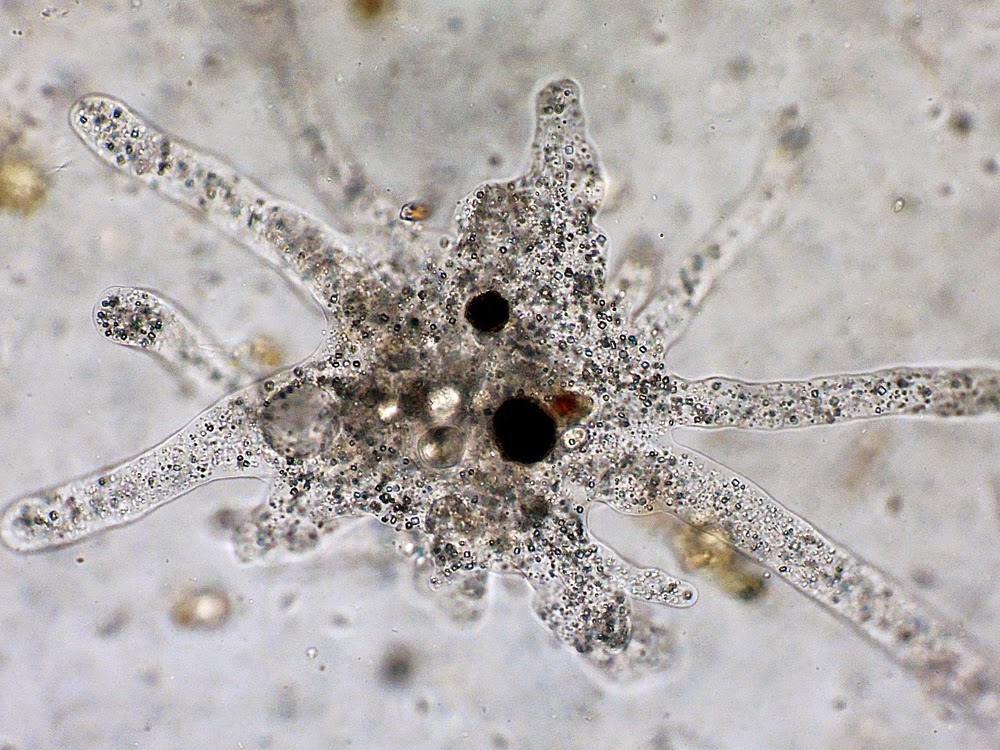
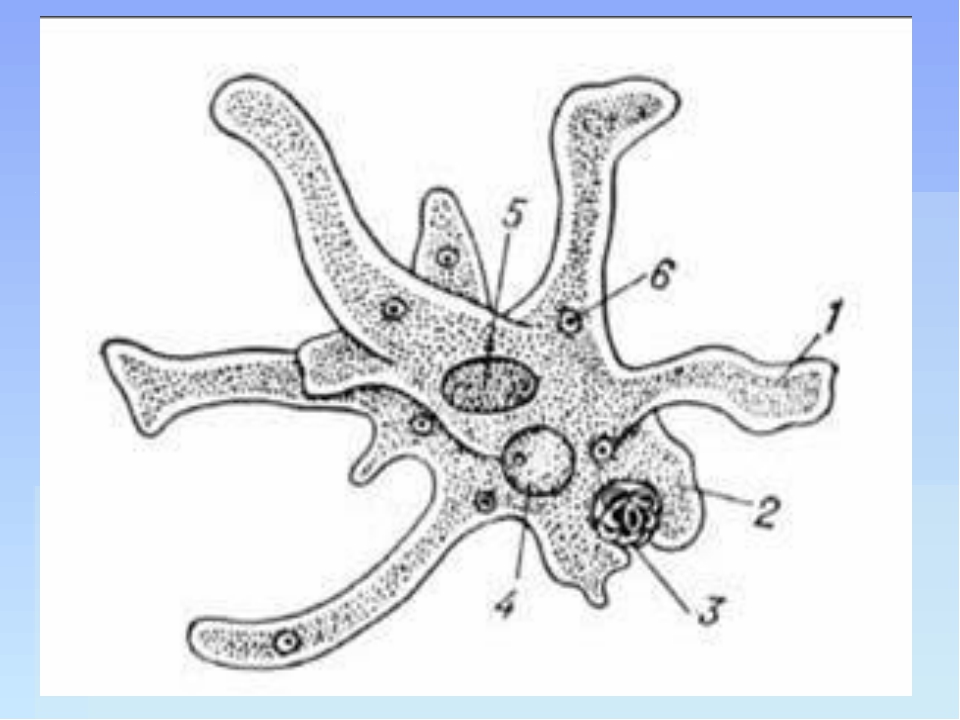
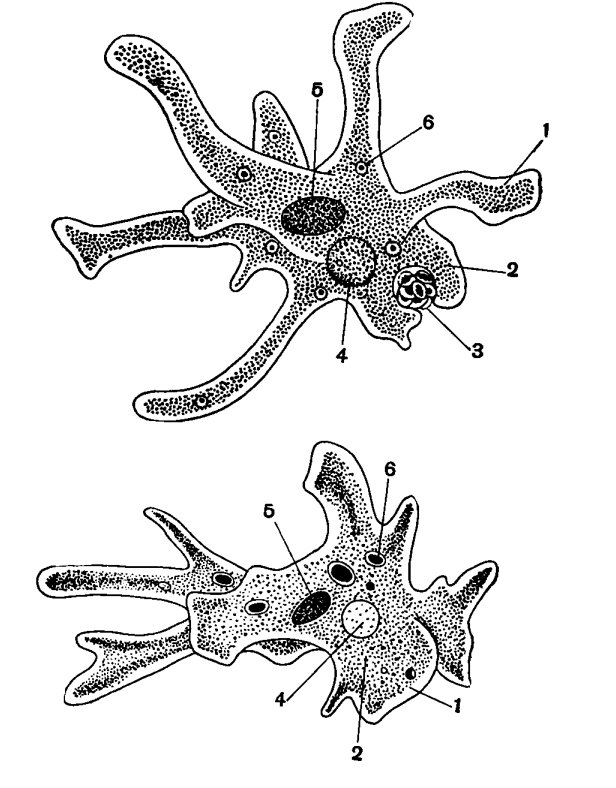
Класс одноклеточных животных, которому принадлежит обыкновенная амёба (Amoeba proteus), отнюдь не случайно назван в зоологической систематике латинским словом Rhizopoda, что значит Корненожки.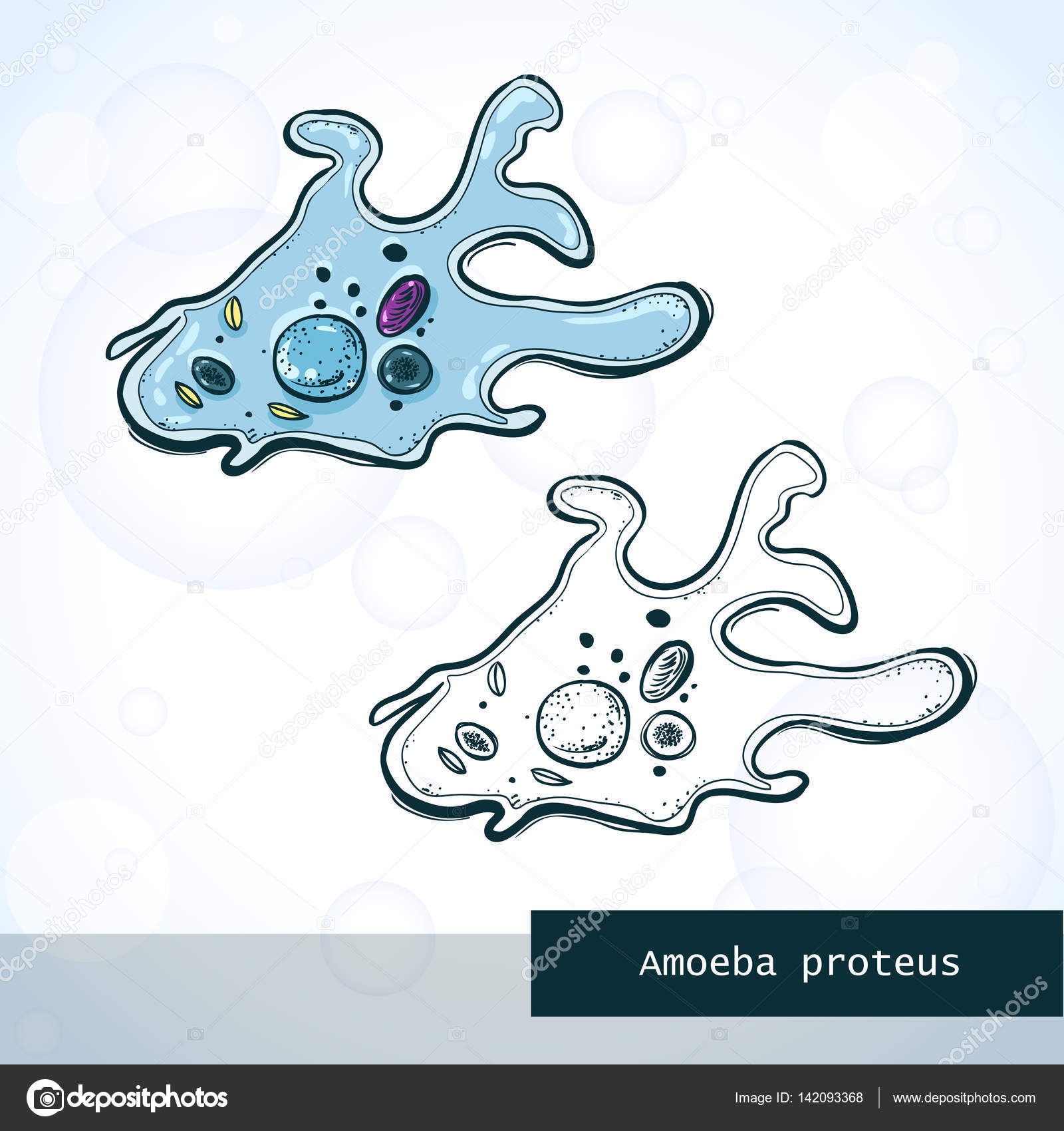 Тело амёбы (с ее плазматическими выростами-псевдоподиями, при помощи которых она не только передвигается, но и захватывает добычу), подобно ветвящимся корням растений. Кстати, это микроскопическое существо обитает в любом пресном водоеме, его несложно культивировать в лабораторных условиях.
Тело амёбы (с ее плазматическими выростами-псевдоподиями, при помощи которых она не только передвигается, но и захватывает добычу), подобно ветвящимся корням растений. Кстати, это микроскопическое существо обитает в любом пресном водоеме, его несложно культивировать в лабораторных условиях.
«Амёба — наиболее просто организованное одноклеточное животное», — читаем в учебниках по зоологии. Может, оно и так, но за этой внешней (и обманчивой) простотой скрывается глубочайший биологический смысл, скажу больше — великая тайна природы. Суть в том, что амёба с ее ветвистыми, непостоянными по своим очертаниям псевдоподиями — это живая модель эволюции эукариот.
Несомненные признаки существования жизни на Земле известны науке с периода 3,8–4 млрд. лет назад, то есть уже через 500 млн. лет после образования самой планеты). Также твердо установлено, что не менее 2 млрд. лет от начала своего рождения (вплоть до так называемой неопротерозойской революции) биосфера была прокариотной, т.е. в ней всецело господствовали одноклеточные безъядерные существа-прокариоты. Эукариоты заняли свое место в биосфере лишь около 1 млрд. лет назад.
лет после образования самой планеты). Также твердо установлено, что не менее 2 млрд. лет от начала своего рождения (вплоть до так называемой неопротерозойской революции) биосфера была прокариотной, т.е. в ней всецело господствовали одноклеточные безъядерные существа-прокариоты. Эукариоты заняли свое место в биосфере лишь около 1 млрд. лет назад.
Прокариотная клетка Эукариотная клетка
Прокариоты не имеют настоящего клеточного ядра. Генетические функции у них выполняют так называемые генофоры — кольцевые двухцепочечные молекулы ДНК, имеющие вид «клубка ниток», как правило, прикрепленные в одном месте к наружной клеточной мембране. Другая очень важная (в контексте рассматриваемой проблемы) особенность биологической организации прокариот, которыми являются и ныне существующие бактерии, состоит в том, что они принципиально неспособны образовывать ветвящиеся псевдоподии, а стало быть, и не обладают способностью захватывать, заглатывать и переваривать внутриклеточно (как это делает упомянутая амёба) свою добычу целиком или по частям. Гетеротрофные прокариоты выделяют пищеварительные ферменты на поверхность живой добычи, в результате чего происходит ее так называемое «наружное переваривание», завершающееся поглощением низкомолекулярных продуктов через цитоплазматическую мембрану. В силу этих обстоятельств среди прокариот нет настоящих хищников.
Гетеротрофные прокариоты выделяют пищеварительные ферменты на поверхность живой добычи, в результате чего происходит ее так называемое «наружное переваривание», завершающееся поглощением низкомолекулярных продуктов через цитоплазматическую мембрану. В силу этих обстоятельств среди прокариот нет настоящих хищников.
Трудно представить, как бы дальше сложилась судьба прокариотной биосферы и как бы она выглядела сейчас. Но (к счастью для всех нас!) получилось так (вероятно, в силу спонтанных генетических мутаций, случившихся «в одно прекрасное время»), что в некоторых клетках прокариот появилось очень важное новообразование — так называемая актин-миозиновая система. Актин и миозин — универсальные сократительные белки нынешних эукариот, составляющие основу цитоскелета. Эти белки обеспечивают амебоидную активность, движение органелл внутри клетки, мышечные сокращения и другие формы клеточной подвижности. Они-то и обусловили способность образовывать псевдоподии, захватывать ими жертву и формировать пищеварительные вакуоли.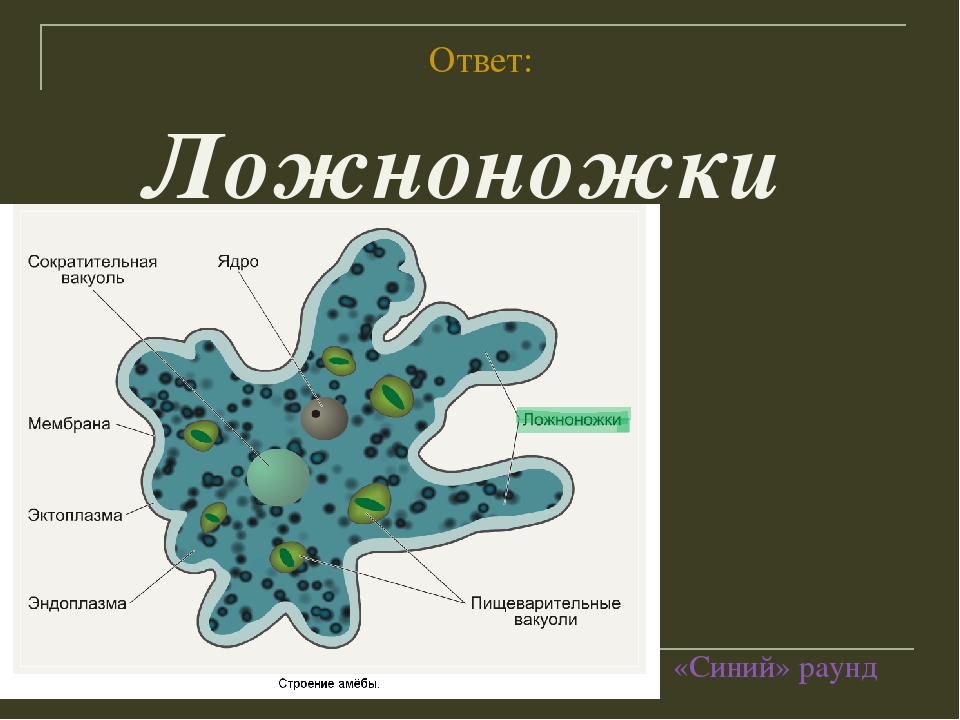 Такие перемены на молекулярно-биологическом уровне радикальнейшим образом изменили всю дальнейшую судьбу биосферы.
Такие перемены на молекулярно-биологическом уровне радикальнейшим образом изменили всю дальнейшую судьбу биосферы.
Цитоскелет эукариотной клетки
Тело живой клетки приобрело динамическую способность ветвиться, то есть образовывать (и, когда надо, убирать внутрь!) псевдоподии, которые стали активно захватывать бактерий, переводить их в цитоплазму для последующего внутриклеточного переваривания. На арену клеточной жизни выходит совершено новое явление — фагоцитоз (греч. phagos — пожирающий). В биосфере появляются, наконец, настоящие хищники-фагоциты. За этим революционным достижением последовала вереница новых событий и далеко идущих причинно-следственных отношений.
.
Фаготрофный образ жизни одноклеточных существ значительно активизировал подвижность цитоплазмы, особенно ее периферийных участков. В результате упомянутые генофоры оказались в зоне наиболее интенсивных токов цитоплазмы. Это обстоятельство способствовало погружению генофоров вглубь клетки, трансформации их в хромосомы, а затем и образованию на их основе обособленного клеточного ядра. А это уже первый шаг к эволюции настоящих ядерных организмов – эукариот.
А это уже первый шаг к эволюции настоящих ядерных организмов – эукариот.
Дальше — больше. Способность к фаготрофному питанию предопределила возможность появления у ветвящихся эукариот внутренних симбионтов. Некоторые бактерии, «проглоченные» новоявленными амёбообразными фагоцитами, оказались способными вступить с ними в симбиотические отношения (такое широко наблюдается и поныне в мире одноклеточных эукариот). В результате возникли клеточные органеллы митохондрии, которые, будучи окружены двойной мембраной (внутренняя — собственная и наружная — принадлежащая пищеварительной вакуоли хозяина), сохранили автономный генетический аппарат прокариотного типа и способность к самостоятельному размножению внутри хозяина, как это демонстрируют нам некоторые современные свободноживущие бактерии. Митохондрии взяли на себя задачу синтеза АТФ за счет окислительного фосфорилирования и стали универсальными энергетическими аккумуляторами всякой эукариотной клетки.
Данные современной науки убедительно свидетельствуют о том, что органеллы растительной клетки — хлоропласты также являются потомками способных к фотосинтезу прокариот, захваченных когда-то амёбообразным хищником. И, стало быть, симбиоз на клеточном уровне определил в дальнейшем прогрессивную эволюцию не только животных, но и огромного царства растений.
И, стало быть, симбиоз на клеточном уровне определил в дальнейшем прогрессивную эволюцию не только животных, но и огромного царства растений.
Так одноклеточный ветвящийся хищник, сильно похожий на нынешнюю амебу, положил в далеком прошлом начало великому симбиозу, который радикально, до неузнаваемости изменил направление всей дальнейшей эволюции биосферы.
Обыкновенная амеба: pritochnik — LiveJournal
Читая посты некоторых полит-блогеров и комментарии под ними… диву даюсь, когда в стране смогли вырастить столько «плакальщиков»?Ну ладно девушки, я понимаю…, им по статусу положено иной раз слезки кап… кап, но мужики туда же… что ни пост, то слезы или ругань, вперемешку с соплями и нытьем…
Чему учили их учителя в школе? Неужели тому как стать неудачником со слезами на глазах?
Куда смотрели родители, когда растили этих инфантильных недоносков — «амеб обыкновенных»?
Диву даешься, читая их опусы:
«Нам объявили очередные санкции — мы точно все умрем!!!», «Отдайте Крым, зачем он нам?», «Хакеры!!!», «Путин слил!!!».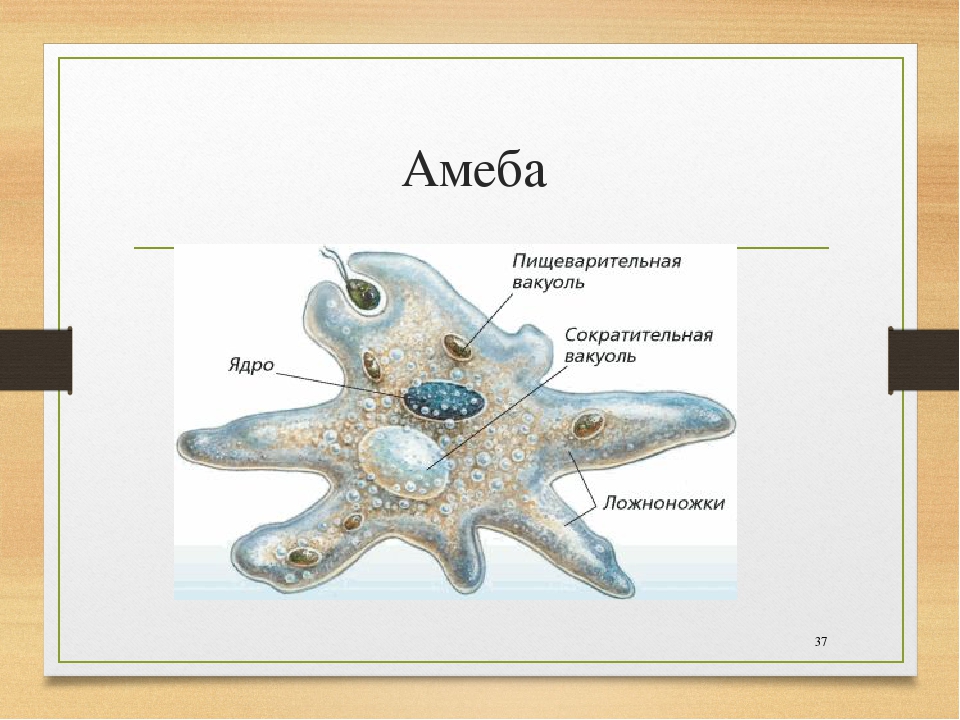 ………..
………..
…новые коммунисты требуют вернуть СССР, однопартийную систему и вновь начать строить коммунизм…
…новые монархисты требуют переименовать Путина в Николая II и пойти войной на Донбасс, Париж, Лондон и Вашингтон…
…новые националисты требуют переименовать Путина в Сталина и вернуть СССР, но! только из русских! и пойти войной на всех нерусских…
…либералы орут благим матом: «даешь в Президенты Навального», а Путина «демократично» расстрелять вместе с Шойгу… и требуют! распустить армию и раздать территорию страны Японии, США, Великобритании и Германии — ведь только их лидеры знают как, надо руководить странами…
и все вместе, дружно ругают Правительство, Президента, депутатов, министров …при этом сами мечтают взойти на Олимп власти…
…дабы там показать какие они «крутые» и насладится «плодами»…
а пока живут как амебы… ругают, ноют, опять ругают, опять ноют… как все плохо! при этом ничего не делая для создания собственной успешной жизни и ничего не предлагая конструктивного в обмен на ругань,
нытье. ..
..
и сплошные сопли…
замешанные на грязи информационных вбросов любезно подсунутых им зарубежными СМИ.
их великие гуру-аналитики год назад предрекали крах экономики России в результате падения цены на нефть,
сегодня спросил в комментарии — нефть 54…
все живы…
заводы работают…
скважины качают…
как быть с Вашим прогнозом? в ответ полная тишина… и невнятное бормотание о хакерах и грядущем импичменте Трампа…
и ни один из них, когда его спрашиваешь напрямую, не может ответить, а каковы их конкретные предложения…
ну кроме… свергнуть, вернуть и пр. ерунды?
все дружно молчат или переводят стрелки на очередное нытье про олигархов, недоказанных коррупционеров… и прочие «яхты» с «часами».
Хочется закричать на весь ЖЖ — мужики Вы в кого себя превратили?
Я удивлен… как еще с Вами девушки в постель ложатся?… и совершенно не удивляет возмущение женской половины ЖЖ вашим инфантилизмом и несостоятельностью…
Тьфу на ваши слезы
и тьфу три раза. .. на вашу брань в сторону политиков всех мастей, которые хоть что-то в этой жизни делают в отличии от Вас — нытиков.
.. на вашу брань в сторону политиков всех мастей, которые хоть что-то в этой жизни делают в отличии от Вас — нытиков.
Вы не политические оппозиционеры, вы — обыкновенные амебы.
фото: mypresentation.ru/presentation/rabota_v_
Конспект урока «Корненожки. Амеба обыкновенная» 7 класс
Пояснительная записка
Сценарий и презентация к уроку «Тип Саркодовые и Жгутиконосцы. Класс
Саркодовые. Амеба обыкновенная» предназначается для использования на уроке
биологии в 7 классе общеобразовательной школы. Это первый урок из цикла
уроков, посвященных изучению подцарства Простейшие. Обучение ведется по
учебнику Константинов В.М.,Бабенко В.Г., Кучменко В.С. Биология: Животные:
Учебник для 7 класса общеобразовательных учреждений / Под ред.
В.М.Константинова, И.Н.Пономаревой. – М.:Вентана – Граф, 2004.
Целью урока является создание условий для осмысления новой учебной
информации, повышение мотивации учащихся.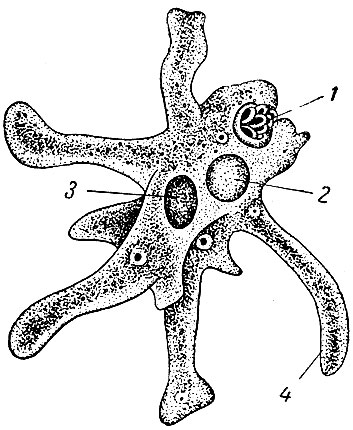 Презентация применяется на этапе объяснения нового материала, а так же на
этапе закрепления изученного на уроке. По ходу урока предполагается
самостоятельная работа учеников с текстом учебника, которая сопровождается
демонстрацией слайдов с заданиями. По окончании выполнения задания можно
осуществить проверку правильности выполнения, так как по щелчку мыши
происходит высвечивание правильных вариантов ответов. На этапе закрепления
нового материала проводится игра (слайды №16 №20). Система гиперссылок
позволяет моментально оценить ответ. Для этого необходимо навести мышь на
предполагаемый ответ, и щелкнуть по нему один раз. На экране появляются
надписи «Молодец!» или «Подумай!». Под надписями находятся кнопки, нажав
на которые мышью, можно вернуться к заданиям игры и продолжить ее. Данную
разработку можно использовать при наличии мультимедийного проектора при
фронтальной работе класса на уроке. Она проста в применении, технологична.
Презентация применяется на этапе объяснения нового материала, а так же на
этапе закрепления изученного на уроке. По ходу урока предполагается
самостоятельная работа учеников с текстом учебника, которая сопровождается
демонстрацией слайдов с заданиями. По окончании выполнения задания можно
осуществить проверку правильности выполнения, так как по щелчку мыши
происходит высвечивание правильных вариантов ответов. На этапе закрепления
нового материала проводится игра (слайды №16 №20). Система гиперссылок
позволяет моментально оценить ответ. Для этого необходимо навести мышь на
предполагаемый ответ, и щелкнуть по нему один раз. На экране появляются
надписи «Молодец!» или «Подумай!». Под надписями находятся кнопки, нажав
на которые мышью, можно вернуться к заданиям игры и продолжить ее. Данную
разработку можно использовать при наличии мультимедийного проектора при
фронтальной работе класса на уроке. Она проста в применении, технологична.
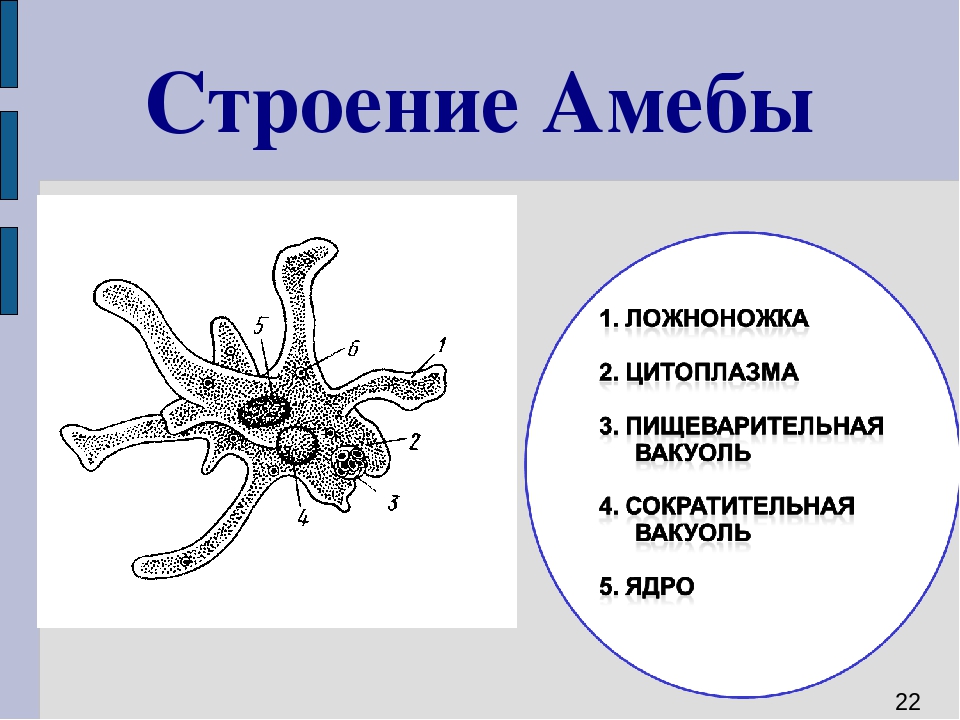 д.)
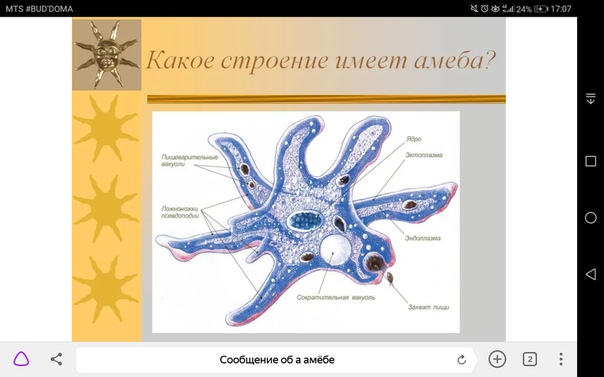
2.5.Процессы жизнедеятельности амебы
2.5.1.Питание,удаление непереваренных остатков и выделение избытка воды.
(Просмотр фрагмента слайд №6, с последующим самостоятельным выполнением задания на
слайде№7и №8 .Проверка качества выполнения работы осуществляется с помощью слайда№7,так
как правильные ответы появляются при щелчке мышью. Отвечавшие получают жетоны за свои
правильные ответы)
Задание слайда №7
Установи правильную последовательность процессов питания амебы
(По щелчку мышью, появляются ответы)
Задание слайда № 8
Выбери номера правильных утверждений
(правильные утверждения по щелчку мышью изменяют цвет)
2.5.2.Дыхание амебы
(организация самостоятельной работы учащихся с текстом учебника, взаимопроверка в паре)
2.5.3. Размножение амебы
(Просмотр фрагмента о размножении амебы на слайде№9,выполнение задания слайда №10, с
опорой на текст учебника. Фронтальная проверка выполненного по ответам ,появляющимся по
щелчку мышью)
Задание слайда №10 Вставь пропущенное слово в предложение.
д.)
2.5.Процессы жизнедеятельности амебы
2.5.1.Питание,удаление непереваренных остатков и выделение избытка воды.
(Просмотр фрагмента слайд №6, с последующим самостоятельным выполнением задания на
слайде№7и №8 .Проверка качества выполнения работы осуществляется с помощью слайда№7,так
как правильные ответы появляются при щелчке мышью. Отвечавшие получают жетоны за свои
правильные ответы)
Задание слайда №7
Установи правильную последовательность процессов питания амебы
(По щелчку мышью, появляются ответы)
Задание слайда № 8
Выбери номера правильных утверждений
(правильные утверждения по щелчку мышью изменяют цвет)
2.5.2.Дыхание амебы
(организация самостоятельной работы учащихся с текстом учебника, взаимопроверка в паре)
2.5.3. Размножение амебы
(Просмотр фрагмента о размножении амебы на слайде№9,выполнение задания слайда №10, с
опорой на текст учебника. Фронтальная проверка выполненного по ответам ,появляющимся по
щелчку мышью)
Задание слайда №10 Вставь пропущенное слово в предложение.
Ответ 19 Простейшие — Рабочая тетрадь по биологии 5 класс Н.И.Сонин, А.А.Плешаков
133. Дайте определение.
Простейшие — это
- Ответ: Одноклеточные микроскопические животные.
134. Используя материал учебника, заполните таблицу «Общая характеристика простейших».
-
Ответ:
Представитель Характерные особенности Амеба обыкновенная Тело не имеет постоянно формы, она передвигается и питается с помощью ложноножек. Инфузория туфелька Ее тело покрыто множеством коротких ресничек, с помощью которых она двигается.  Некоторые живут в симбиозе с жвачными.
Некоторые живут в симбиозе с жвачными.Молярийный плазмодий Паразит-возбудитель малярии Радиолярия Имеет скелет
135. Рассмотрите рисунок. Подпишите основные части клетки. Укажите ядро, цитоплазму, клеточную мембрану.
-
Ответ:
136.
- Ответ: Одноклеточные, имеют мембрану и цитоплазму, имеют средства передвижения.
137. Объясните, почему простейших можно назвать клетками-организмами.
- Ответ: Потому что органоиды простейших выполняют функцию подобно целому организму: простейшие дышат, питаются самостоятельно при помощи органоидов.
138. Используя дополнительные источники информации, подготовьте сообщение на тему по выбору: «Паразитические простейшие (2-3 примера), «Значение простейших в природе и жизни человека». Запишите план сообщения. Укажите источник использованной информации.
Используя дополнительные источники информации, подготовьте сообщение на тему по выбору: «Паразитические простейшие (2-3 примера), «Значение простейших в природе и жизни человека». Запишите план сообщения. Укажите источник использованной информации.
-
Ответ:
1) Полезные человеку простейшие
2) Значение простейших
Источник: энциклопедия
139. Вставьте пропущенные буквы.
- Ответ: ПрОстейшие, парАзит, Амеба, инФузория, мАлЯрийный плАзмоДий.
Составьте 2 — 3 предложения с этими словами.
- Ответ: Многочислены простейшие. Но кроме безобидных амеб и инфузорий существуют простейшие-паразиты. Например, малярийный плазмодий, передающий малярию.
Тренировочный вариант 2 ВПР 2021 по биологии 8 класс
Тренировочный вариант 2 ВПР 2021 по биологии 8 класс. Пробные варианты по биологии 8 класс ВПР 2021. ВСЕРОССИЙСКАЯ ПРОВЕРОЧНАЯ РАБОТА Биология 8 класс.
1.
Как называют научный метод, изображённый на фотографии?
1) метод наблюдения
2) метод моделирования
3) экспериментальный метод
4) метод измерения
Ответ:
Объясните свой ответ, воспользовавшись знанием научных методов биологии.
Ответ:
2.
Рассмотрите изображённое на фотографии животное и опишите его, выполнив задания.
2.1. Укажите тип симметрии животного.
Ответ:
2.2. Укажите среду обитания животного.
Ответ:
2.3. Установите последовательность расположения систематических групп изображённого животного, начиная с самой крупной. Используйте слова и словосочетания из предложенного перечня. Запишите в таблицу цифры, под которыми они указаны. Список слов и словосочетаний:
Список слов и словосочетаний:
1) Парнокопытные
2) Хордовые
3) Животные
4) Европейская косуля
5) Млекопитающие
Ответ:
2.4. Укажите одно из значений, которое имеют косули в природе.
Ответ:
3.
Известно, что чёрный аист – птенцовая перелётная птица, обитающая вблизи водоёмов. Используя эти сведения, выберите из приведённого ниже списка три утверждения, относящиеся к описанию данных признаков этого животного. Запишите в ответе цифры, соответствующие выбранным ответам.
1) Чёрный аист обитает в лесной зоне Евразии, а зимует в южной Азии и Африке.
2) Эта скрытная птица предпочитает жить вдали от человека.
3) Чёрный аист питается в основном рыбой, мелкими водными беспозвоночными животными.
4) Аисты устраивают массивные гнёзда из толстых веток и сучьев деревьев, скреплённых между собой при помощи дёрна, земли и глины на высоте 10-20 метров.
5) В кладке бывает от 4 до 7 яиц.
6) Птенцы сидят в гнёздах до двух месяцев, всё это время родители кормят своё потомство.
Ответ:
4.
4.1. Определите тип питания организмов, приведённых в перечне. Запишите цифры, под которыми указаны организмы, в соответствующую ячейку таблицы.
Список организмов:
1) черёмуха обыкновенная
2) лисичка ложная
3) африканский слон
4) ромашка лекарственная
5) большой прудовик
6) подорожник большой
Ответ:
4.2. Какой тип питания характерен для гадюки обыкновенной, изображённой на рисунке 1?
Ответ:
Обоснуйте свой ответ:
5.
Рассмотрите рисунок 2, на котором представлен цикл развития печёночного сосальщика, и ответьте на вопросы.
5.1. Какой цифрой обозначен на рисунке окончательный хозяин паразита?
Ответ:
5.2. Может ли человек заразиться печёночным сосальщиком, если он работает в хлеву и регулярно занимается уборкой коровьего навоза? Ответ обоснуйте.
Ответ:
6.
В приведённой ниже таблице между позициями первого и второго столбцов имеется взаимосвязь.
6.1. Какое понятие следует вписать на место пропуска в этой таблице?
1) грызущий
2) колюще-сосущий
3) лижущий
4) грызуще-лижущий
Ответ:
6.2. Чем питаются взрослые чешуекрылые?
Ответ:
7.
Схема какой системы органов животных показана на рисунке 3?
1) Выделительной
2) Нервной
3) Пищеварительной
4) Половой
Ответ:
8.
8.1. Установите соответствие между характеристиками и типами беспозвоночных: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
Запишите в таблицу выбранные цифры под соответствующими буквами.
Ответ:
8.2. Приведите по три примера животных, относящихся к указанным типам. Запишите их названия в таблицу.
9.
Вставьте в текст пропущенные термины из предложенного перечня, используя для этого их цифровые обозначения. Впишите номера выбранных терминов на места пропусков в тексте.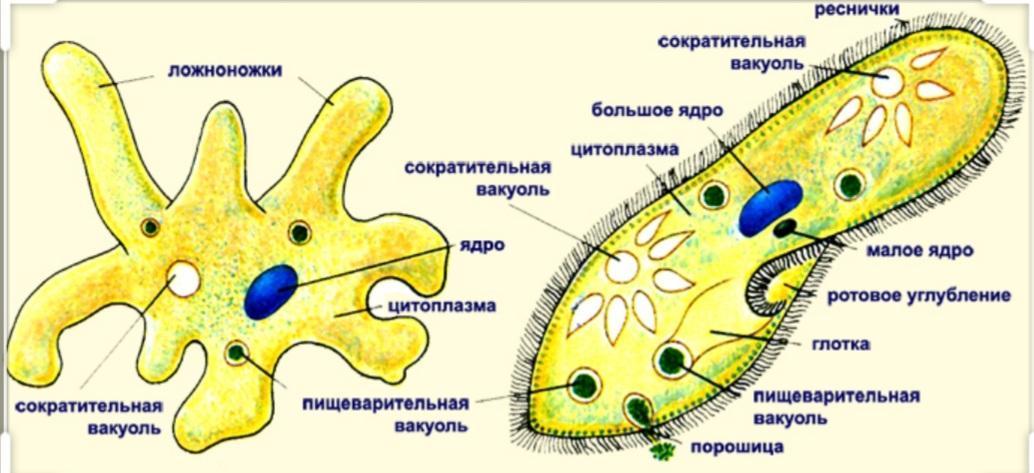
Дизентерийная амёба
Дизентерийная амёба относится к паразитическим ______(А) животным. Как и обыкновенная амёба, она имеет ______(Б) форму тела. Передвигается паразит с помощью ______(В). Заражение человека происходит при попадании в толстый кишечник ______(Г)
путём проглатывания загрязнённой воды или пищи. Для профилактики заболевания необходимо соблюдать правила личной и общественной гигиены.
СПИСОК ТЕРМИНОВ:
1) реснички
2) многоклеточные
3) спора
4) циста
5) одноклеточные
6) непостоянная
7) ложноножки
8) постоянная
Запишите в таблицу выбранные цифры под соответствующими буквами.
Ответ:
10.
10.1. Если животное способно «строить» конструкцию, изображённую на рисунке 4, то для этого животного, вероятнее всего, будут
характерны
1) кожно-мускульный мешок
2) развитие с неполным превращением
3) хитиновый покров
4) простые глаза
5) сложные глаза
Запишите в ответе цифры, под которыми указаны выбранные характеристики.
Ответ:
10.2. В описании животных зоологи часто употребляют термин «кожно-мускульный мешок». Что понимают под этим термином?
Ответ:
11.
Верны ли следующие суждения о моллюсках?
А. Тело моллюсков имеет мантию.
Б. Мантия моллюсков выделяет вещества, из которых образуется раковина.
1) верно только А
2) верно только Б
3) оба суждения верны
4) оба суждения неверны
Ответ:
12.
Изучите данные приведённой ниже таблицы и ответьте на вопросы.
У какого вида рыб отмечены самые низкие показатели оптимальных температур для развития икры?
Ответ:
У какого вида рыб икра может нормально развиваться при температуре выше 20 °С?
Ответ:
Какой орган чувств позволяет рыбам воспринимать движение и вибрацию воды?
Ответ:
13.
Рассмотрите фотографию собаки породы басенджи и выполните задания.
13.1. Выберите характеристики, соответствующие внешнему строению собаки, по следующему плану: окрас шерсти, форма ушей, форма хвоста.
А. Окрас
Б. Форма ушей
В. Форма хвоста
Запишите в таблицу выбранные цифры под соответствующими буквами.
Ответ:
13.2. Пётр решил выяснить, соответствует ли изображённая на фотографии собака породы басенджи стандартам для использования её в целях чистопородного размножения в клубе собаководства. Помогите Петру решить эту задачу, воспользовавшись фрагментом описания стандарта данной породы.
Стандарт породы басенджи (фрагмент)
1. Окрас: пятнистый.
2. Форма ушей: стоячие.
3. Форма хвоста: кольцом.
Сделайте заключение о соответствии изображённой на фотографии собаки указанным стандартам породы. Оцените возможность использования собаки этой породы для чистопородного размножения в клубе собаководства.
Ответ:
ОТВЕТЫ
1.
Правильный ответ должен содержать следующие элементы:
1) указание научного метода: 4 ИЛИ метод измерения;
2) объяснение, например: метод измерения позволяет получить количественные характеристики объекта (животного) с помощью измерительных приборов
2. 1
1
Двусторонне-симметричное животное
2.2
Наземно-воздушная
2.3
32514
2.4
В правильном ответе должно быть указано значение, например: косули являются звеном в цепи питания.
ИЛИ Крупные хищники охотятся на косуль.
ИЛИ Косули питаются травянистыми растениями, молодыми побегами и почками кустарников и деревьев, грибами и семенами
3.
136
4.1
автотрофный тип питания – 146
гетеротрофный тип питания – 235
(в любой последовательности)
4.2
Правильный ответ должен содержать следующие элементы:
1) тип питания животного: гетеротрофный;
2) обоснование, например: у гадюки обыкновенной отсутствуют хлоропласты, поэтому у неё гетеротрофный тип питания;
ИЛИ гадюка обыкновенная питается готовыми органическими веществами мелкими животными, то есть имеет гетеротрофный тип питания;
ИЛИ отсутствует фотосинтез
5. 1
1
1
5.2
Правильный ответ должен содержать следующие элементы:
1) ответ на вопрос: нет, заражение невозможно;
2) обоснование, например: человек заражается печёночным сосальщиком только при попадании в его организм цист вместе с травой или хвостатых личинок вместе с водой;
ИЛИ в навозе содержатся яйца сосальщика, которые не заразны для человека.
Элементы ответа могут быть приведены в иных, близких по смыслу формулировках
6.1
1
6.2
Правильный ответ должен содержать указание типа пищи, например: нектар / соки деревьев / гниющие или перезрелые фрукты / пыльца
7.
2
8.1
212111
8.2
Правильный ответ должен содержать по три примера беспозвоночных животных, относящихся к типам Моллюски и Плоские
9.
5674
10.1
35 (в любой последовательности)
10. 2
2
Правильный ответ должен содержать объяснение термина, например: кожно-мускульный мешок – покров тела червей, образованный кожным эпителием и расположенными под ним мышцами.
Объяснение термина может быть приведено в иной, близкой по смыслу формулировке
11.
3
12.
Правильный ответ должен содержать три элемента:
1) ручьевая форель;
2) карп;
3) боковая линия.
Элементы ответа могут быть приведены в иных, близких по смыслу формулировках
13.1
232
13.2
Правильный ответ должен содержать следующие элементы:
1) заключение, например: собака, изображённая на фотографии, соответствует стандарту породы по окрасу шерсти, форме хвоста, но не соответствует стандарту породы по форме ушей;
2) оценка, например: данная собака не подходит для чистопородного размножения в клубе собаководства
Какие прилагательные подобрать к слову «амеба»?
Вот такие прилагательные подберем к слову «амеба»:
-проклятая амеба,
-загадочная амеба,
-редкая амеба,
-жуткая амеба,
-местная амеба,
-неприятная амеба,
-сонная амеба,
-страшная амеба,
-дурная амеба,
-любая амеба,
-мерзкая амеба,
-неведомая амеба,
-опасная амеба,
-безнадёжная амеба,
-коварная амеба,
-красная амеба,
-мнимая амеба,
-модная амеба,
-неизвестная амеба,
-распространённая амеба,
-революционная амеба,
-скверная амеба,
-водяная амеба,
-мелкая амеба,
-отвратительная амеба,
-постыдная амеба,
-серьёзная амеба,
-смертельная амеба,
-смертоносная амеба,
-ужасная амеба,
-французская амеба,
-экзотическая амеба,
-грозная амеба,
-любая амеба,
-нехорошая амеба,
-опасная амеба,
-подобная амеба,
-роковая амеба,
-тропическая амеба,
-ушастая амеба,
-духовная амеба,
-профессиональная амеба,
-психическая амеба,
-чёрная амеба,
-большевистская амеба,
-воздушная амеба,
-генетическая амеба,
-гнусная амеба,
-жестокая амеба,
-зеркальная амеба,
-конкретная амеба,
-неизвестная амеба,
-обыкновенная амеба,
-позорная амеба,
-простая амеба,
-сильная амеба,
-другая амеба,
-коммунистическая амеба,
-маленькая амеба,
-непонятная амеба,
-новая амеба,
-прочая амеба,
-тяжёлая амеба,
-страшная амеба,
-таинственная амеба,
-чумная амеба,
-заразная амеба,
-неожиданная амеба,
-странная амеба,
-гигантская амеба,
-неведомая амеба.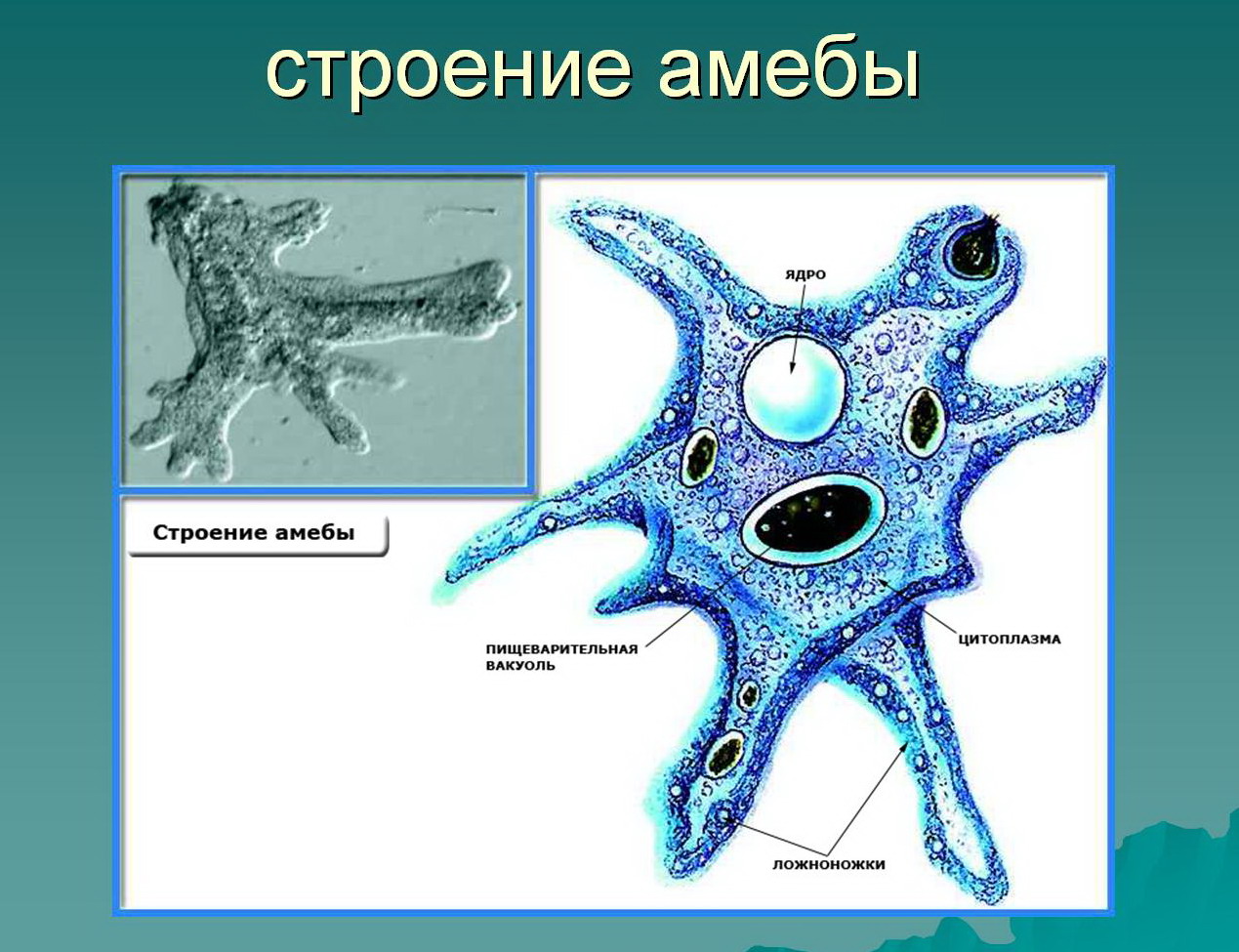
Entamoeba — обзор | ScienceDirect Topics
Характеристики видов
Entamoeba — псевдоподообразующие простейшие паразиты подтипа Sarcodina. Род Entamoeba состоит из пяти видов, которые заражают людей: E. histolytica , E. dispar , E. hartmanni , E. coli , E. polecki и Известно, что только E. histolytica вызывают заболевания у людей. E. dispar , E. hartmanni , E. coli и E. polecki являются комменсалами, обнаруживаемыми в толстой кишке, а E. gingivalis обнаруживаются в ротовой полости. Хотя E. histolytica и E. dispar морфологически идентичны, другие виды можно отличить друг от друга с помощью микроскопии. Большой трофозоит и цисту E. coli можно идентифицировать по осколочно-подобным хроматидным тельцам в цитоплазме и по тому факту, что цисты могут иметь до восьми ядер. E. hartmanni — самый мелкий представитель группы, размеры его цист и трофозоитов не превышают 10 мкм.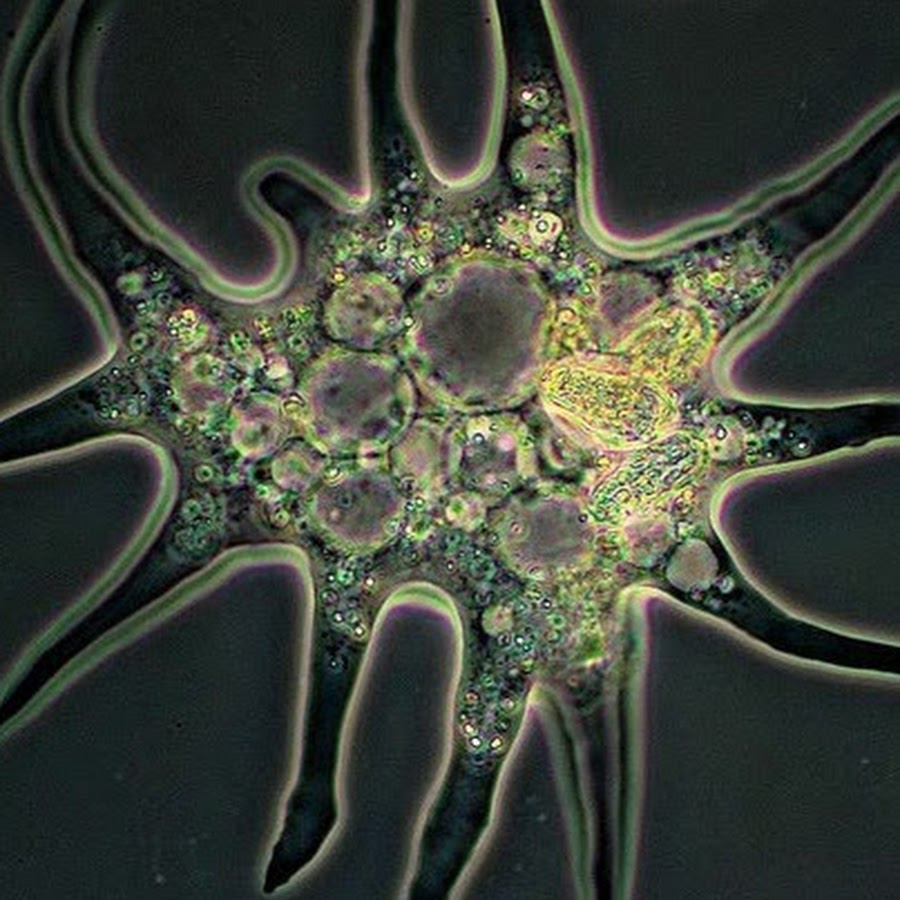
Амебиаз – это заболевание, вызываемое E. histolytica , которое состоит из двух основных синдромов: дизентерии и абсцессов печени. Цикл инфекции начинается, когда кистозная форма организма попадает в организм с загрязненной фекалиями пищей или водой. Кислотоустойчивая киста проходит через желудок невредимой, пока не достигает тонкой кишки, где эксцистирует с образованием восьми трофозоитов, подвижной и инвазивной формы этого вида (рис. 1А). Трофозоиты мигрируют в толстую кишку, где они могут либо колонизировать просвет кишечника в виде комменсальной флоры, либо внедряться в эпителий толстой кишки, вызывая воспаление и разрушение стенки кишечника.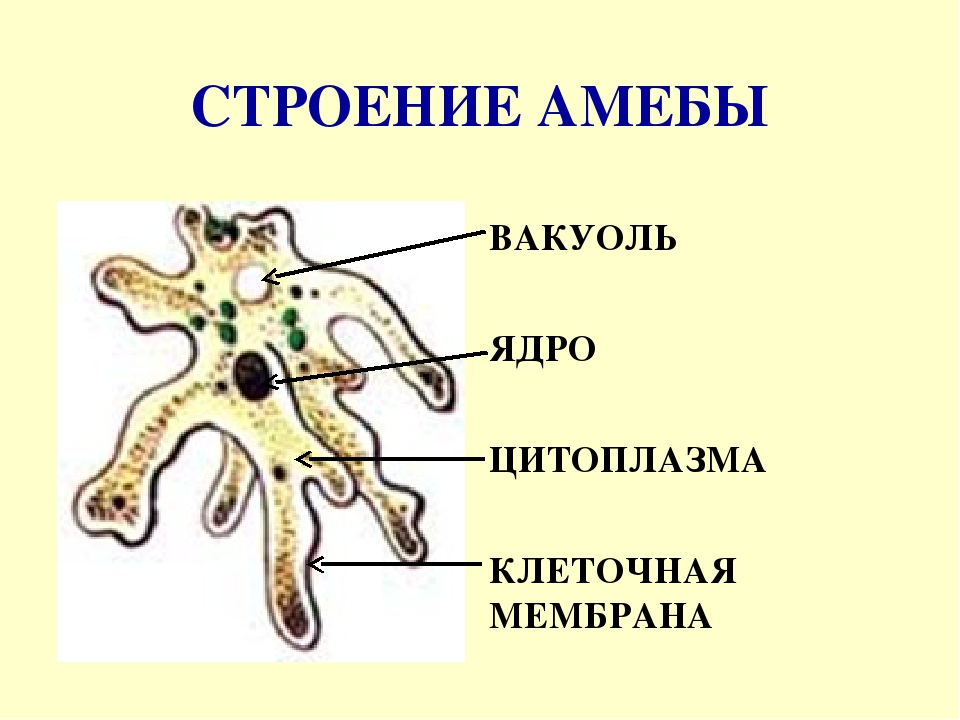 Что влияет на это решение об вторжении, пока неизвестно, но потенциальные факторы включают различия между штаммами амебы и вариациями хозяина, такие как генетический состав, кишечная флора, состояние питания и иммунокомпетентность. После инвазии амебы могут получить доступ к портальному кровообращению и транспортироваться к различным органам-мишеням, включая печень, мозг и легкие. Печень является наиболее частой локализацией внекишечного амебиаза. Жизненный цикл амебы завершается, когда трофозоиты в толстой кишке инцистируются и выделяются с фекалиями хозяина (рис.1Б).
Что влияет на это решение об вторжении, пока неизвестно, но потенциальные факторы включают различия между штаммами амебы и вариациями хозяина, такие как генетический состав, кишечная флора, состояние питания и иммунокомпетентность. После инвазии амебы могут получить доступ к портальному кровообращению и транспортироваться к различным органам-мишеням, включая печень, мозг и легкие. Печень является наиболее частой локализацией внекишечного амебиаза. Жизненный цикл амебы завершается, когда трофозоиты в толстой кишке инцистируются и выделяются с фекалиями хозяина (рис.1Б).
Рисунок 1. E. histolytica или E. dispar трофозоиты ( A , окраска трихомами) и цисты ( B , окраска йодом) в стуле. Различие между этими двумя видами невозможно только с помощью микроскопии. (Фотографии предоставлены Центрами по контролю за заболеваниями.)
Одноядерный трофозоит имеет диаметр 10–60 мкм, но четырехъядерная киста обычно меньше, в среднем 10 мкм в диаметре. Трофозоиты, вышедшие с калом, хрупки и быстро погибают вне хозяина.Кисты имеют стенку, которая делает их устойчивыми к хлору и высыханию, и могут жить в течение нескольких недель в тенистой и влажной среде при комнатной температуре и до нескольких месяцев в холодильнике. Однако кипячение или очень высокие уровни хлора могут разрушить цисты. Хотя наиболее распространенным способом передачи инфекции является прием зараженной пищи или воды, передача инфекции от человека к человеку также может происходить в условиях скопления людей и плохой личной гигиены, например, в психиатрических больницах и детских садах. Требуется всего 10–100 цист, чтобы вызвать амебную дизентерию на животных моделях, что является заразной дозой, сравнимой с печально известной контагиозностью Shigella sp., который может передаваться всего 10–100 организмами.
Трофозоиты, вышедшие с калом, хрупки и быстро погибают вне хозяина.Кисты имеют стенку, которая делает их устойчивыми к хлору и высыханию, и могут жить в течение нескольких недель в тенистой и влажной среде при комнатной температуре и до нескольких месяцев в холодильнике. Однако кипячение или очень высокие уровни хлора могут разрушить цисты. Хотя наиболее распространенным способом передачи инфекции является прием зараженной пищи или воды, передача инфекции от человека к человеку также может происходить в условиях скопления людей и плохой личной гигиены, например, в психиатрических больницах и детских садах. Требуется всего 10–100 цист, чтобы вызвать амебную дизентерию на животных моделях, что является заразной дозой, сравнимой с печально известной контагиозностью Shigella sp., который может передаваться всего 10–100 организмами.
Как следует из названия, E. histolytica вызывает заболевание путем лизиса тканей. Гибель клеток зависит от способности амебы прикрепляться к клеткам-мишеням с помощью связывающего галактозу адгезина, называемого лектином Gal/GalNAc. E. histolytica может убивать широкий спектр клеток тканевых культур, а также макрофаги, лимфоциты и нейтрофилы. Считается, что лектин Gal/GalNAc играет центральную роль в патогенезе, поскольку в дополнение к его роли в адгезии и уничтожении клеток он также участвует в резистентности к комплементу, важном навыке выживания, когда трофозоит перемещается по кровотоку.Другие факторы вирулентности включают амебопоры, которые создают отверстия в мембранах клеток-мишеней, и цистеиновые протеазы, разрушающие белки внеклеточного матрикса.
E. histolytica может убивать широкий спектр клеток тканевых культур, а также макрофаги, лимфоциты и нейтрофилы. Считается, что лектин Gal/GalNAc играет центральную роль в патогенезе, поскольку в дополнение к его роли в адгезии и уничтожении клеток он также участвует в резистентности к комплементу, важном навыке выживания, когда трофозоит перемещается по кровотоку.Другие факторы вирулентности включают амебопоры, которые создают отверстия в мембранах клеток-мишеней, и цистеиновые протеазы, разрушающие белки внеклеточного матрикса.
Когда амеба внедряется в стенку кишечника, она образует язвы в форме колб, и симптомы постепенно развиваются от легкой диареи до выраженной дизентерии. Амебная дизентерия характеризуется болями и болезненностью в животе, выделением стула с примесью крови (до 25 в сутки). Диагноз легко пропустить в странах развитого мира, где распространенными причинами кровавой диареи являются ротавирусы Shigella , Campylobacter , Salmonella , а также воспалительные заболевания кишечника.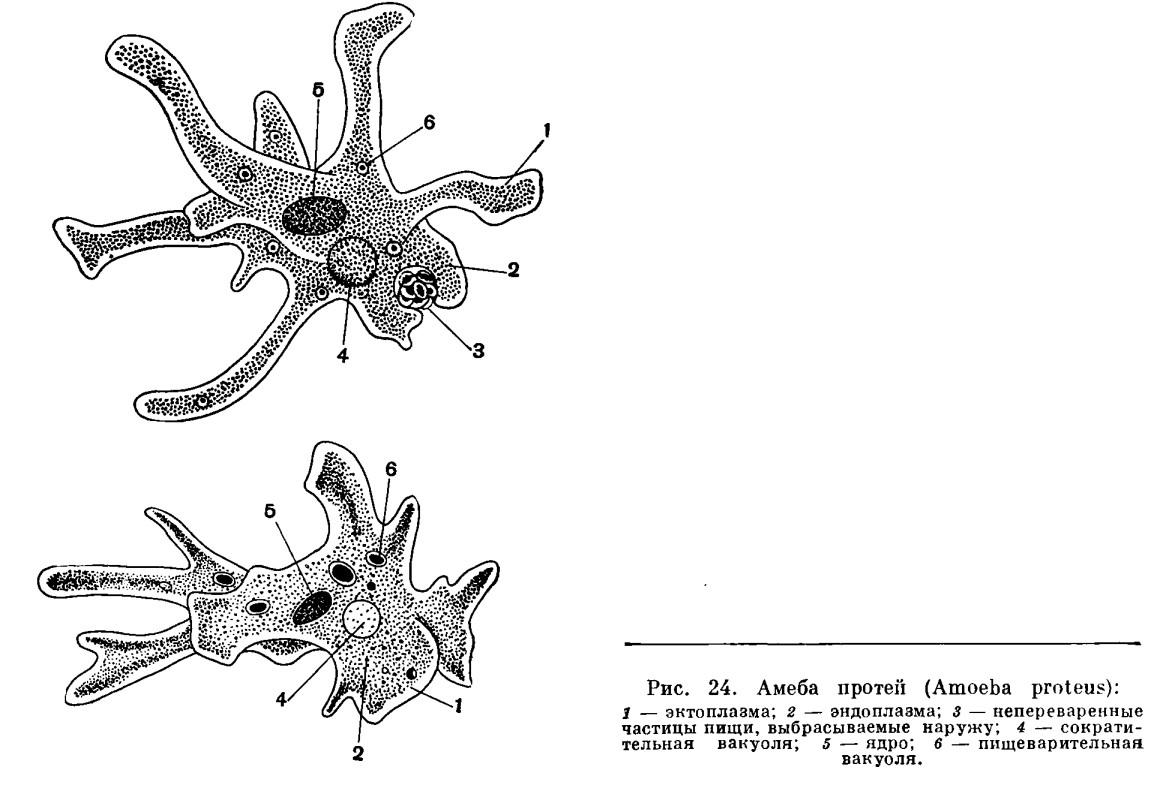 Реже кишечный амебиаз может перерасти в острый некротический колит. Это осложнение обычно лечится частичной или тотальной колэктомией и часто приводит к летальному исходу.
Реже кишечный амебиаз может перерасти в острый некротический колит. Это осложнение обычно лечится частичной или тотальной колэктомией и часто приводит к летальному исходу.
Амебные абсцессы печени, которые чаще встречаются у мужчин, чем у женщин, проявляются болью в животе и лихорадкой, иногда сопровождающейся потерей веса. Визуализирующие исследования печени, такие как УЗИ или компьютерная томография, выявляют дефект овальной формы, чаще всего в правой доле печени. Разорвавшиеся абсцессы печени могут распространяться через диафрагму в легкие и плевральную полость.Реже церебральные абсцессы могут возникать, когда амеба распространяется в мозг через кровоток. Это редкое осложнение амебиаза может привести к смерти через 12–72 часа.
Бессимптомную колонизацию E. histolytica можно лечить с помощью люминальных препаратов дилоксанида фуроата и паромомицина. Лечение всех форм инвазивного заболевания проводится метронидазолом или тинидазолом, а эффективность лечения для обычных схем лечения превышает 90%.
Поскольку люди являются единственным значительным резервуаром для E.histolytica , эффективная вакцина потенциально может полностью устранить болезнь. Способность определенных антигенов, таких как субъединицы лектина Gal/GalNAc, обеспечивать защиту от образования абсцесса печени у песчанок дает надежду на то, что в будущем будет разработана вакцина для человека.
Диагностика инфекций, вызванных патогенными свободноживущими амебами
Naegleria fowleri , Acanthamoeba spp. , Balamuthia mandrillaris и Sappinia sp.являются патогенными свободноживущими амебами. N. fowleri вызывает первичный амебный менингоэнцефалит, быстро смертельную болезнь центральной нервной системы, а Acanthamoeba spp. и B. mandrillaris вызывают хронический гранулематозный энцефалит. Acanthamoeba spp. также может вызывать кожные поражения и амебный кератит, угрожающую зрению инфекцию роговицы, связанную с использованием контактных линз или травмой роговицы. Sappinia pedata была идентифицирована как причина несмертельного случая амебного энцефалита.Ввиду потенциальных последствий для здоровья из-за заражения этими амебами быстрая диагностика имеет решающее значение для раннего лечения. В клинической лаборатории использовали микроскопическое исследование и посев образцов биопсии, спинномозговой жидкости (ЦСЖ) и соскобов с роговицы. При амебном кератите для успешной идентификации амеб в ткани роговицы использовалась конфокальная микроскопия. Совсем недавно были разработаны традиционные методы ПЦР и ПЦР в реальном времени, которые являются чувствительными и специфичными для амеб.Кроме того, доступны мультиплексные ПЦР для быстрой идентификации этих патогенов в биоптатах ткани, спинномозговой жидкости и образцах роговицы.
Sappinia pedata была идентифицирована как причина несмертельного случая амебного энцефалита.Ввиду потенциальных последствий для здоровья из-за заражения этими амебами быстрая диагностика имеет решающее значение для раннего лечения. В клинической лаборатории использовали микроскопическое исследование и посев образцов биопсии, спинномозговой жидкости (ЦСЖ) и соскобов с роговицы. При амебном кератите для успешной идентификации амеб в ткани роговицы использовалась конфокальная микроскопия. Совсем недавно были разработаны традиционные методы ПЦР и ПЦР в реальном времени, которые являются чувствительными и специфичными для амеб.Кроме того, доступны мультиплексные ПЦР для быстрой идентификации этих патогенов в биоптатах ткани, спинномозговой жидкости и образцах роговицы.
1. Введение
Свободноживущие амебы (FLA) встречаются в почве и воде по всему миру. Эти амебы потребляют бактерии, дрожжи и другие организмы в качестве источника пищи. В отличие от «настоящих» паразитов, патогенные FLA могут завершать свой жизненный цикл в окружающей среде, не попадая в организм человека или животного-хозяина. Из многих FLA, обнаруженных в окружающей среде, четыре рода FLA связаны с заболеваниями человека.Один вид Naegleria , N. fowleri, , один вид Balamuthia, B. mandrillaris и несколько видов Acanthamoeba могут вызывать смертельные инфекции центральной нервной системы (ЦНС). Помимо инфекции ЦНС, Acanthamoeba может вызывать кожные поражения и амебный кератит, угрожающую зрению инфекцию роговицы [1–8]. Заражение человека этими амебами описано во всем мире [3, 5]. Совсем недавно недавно обнаруженный патоген был обнаружен в мозговой ткани пациента с симптомами ЦНС, пережившего инфекцию.Хотя амеба не была выделена у пациента, с помощью световой и электронной микроскопии она была идентифицирована как вид Sappinia . Саппиния вид. летальных исходов у человека или экспериментальных животных не было показано [9, 10]. На рис. 1 представлена морфология трофозоитов N. fowleri (А), Acanthamoeba spp. (B) и B. mandrillaris (C) с помощью сканирующей электронной микроскопии.
Из многих FLA, обнаруженных в окружающей среде, четыре рода FLA связаны с заболеваниями человека.Один вид Naegleria , N. fowleri, , один вид Balamuthia, B. mandrillaris и несколько видов Acanthamoeba могут вызывать смертельные инфекции центральной нервной системы (ЦНС). Помимо инфекции ЦНС, Acanthamoeba может вызывать кожные поражения и амебный кератит, угрожающую зрению инфекцию роговицы [1–8]. Заражение человека этими амебами описано во всем мире [3, 5]. Совсем недавно недавно обнаруженный патоген был обнаружен в мозговой ткани пациента с симптомами ЦНС, пережившего инфекцию.Хотя амеба не была выделена у пациента, с помощью световой и электронной микроскопии она была идентифицирована как вид Sappinia . Саппиния вид. летальных исходов у человека или экспериментальных животных не было показано [9, 10]. На рис. 1 представлена морфология трофозоитов N. fowleri (А), Acanthamoeba spp. (B) и B. mandrillaris (C) с помощью сканирующей электронной микроскопии.
Патогенные ФЛА могут быть выделены из пресноводных озер, термически загрязненных вод, донных отложений, термальных источников, бассейнов, почвы, вентиляционных отверстий, воздуха и хозяйственно-питьевого водоснабжения [3, 6, 11–14].Помимо того, что FLA вызывает заболевания человека, они также могут содержать внутриклеточные патогенные бактерии, такие как Legionella pneumophila , и могут служить переносчиками бактериальных инфекций у людей [15, 16]. Было показано, что ряд клинических изолятов FLA из поражений роговицы и кожи содержит бактериальные эндосимбионты или патогены [16-20]. Лабораторные исследования также показали, что ряд патогенных бактерий, включая Mycobacterium avium, Burkholderia spp. , Escherichia coli O157:H7 и Vibrio cholerae , могут выживать и размножаться в FLA [6, 21–26].Было показано, что внутриклеточный рост бактерий внутри амеб повышает устойчивость бактерий к антибиотикам и биоцидам, а также увеличивает вирулентность бактерий [20, 23–26].
Поскольку большинство инфекций FLA являются смертельными и диагностируются посмертно, важно распознавать заболевания и разрабатывать более быстрые методы диагностики. Способ и патогенез инфекции различаются для каждого из FLA, вызывающих инфекции человека.
2.
Naegleria fowleri и первичный амебный менингоэнцефалит (ПАМ) Амебофлагеллят, N.fowleri, имеет три морфологические формы в своем жизненном цикле: стадию амебы или трофозоита (рис. 1(а)), которая питается и делится, плавающего жгутиконосца, который ищет новый источник пищи, и резистентную цисту, которая защищает амеб от неблагоприятных факторов. условия окружающей среды. N. fowleri вызывает первичный амебный менингоэнцефалит (ПАМ), острое молниеносное заболевание с быстрым летальным исходом, которое обычно возникает у ранее здоровых детей и молодых людей, которые в анамнезе занимались плаванием, нырянием и другими рекреационными видами деятельности в пресной воде и загрязненных плавательных бассейнах. 1–3, 27–29].Два других вида Naegleria , N. australiensis и N. italica могут вызывать инфекции у мышей, но никогда не были идентифицированы среди людей [5]. Случаи ПАМ, вызванные N. fowleri , также были связаны с загрязненной бытовой водой, используемой для купания [11, 12]. Заражение может произойти, когда амебы проникают в носовые ходы, прикрепляются к обонятельной слизистой оболочке и мигрируют через решетчатую пластинку вдоль обонятельных нервов. Попав в обонятельные луковицы головного мозга, амебы быстро делятся и гибель наступает в течение 7—10 дней. N. fowleri вызывает молниеносный геморрагический некроз головного мозга. Воспалительный инфильтрат, состоящий из нейтрофилов, эозинофилов и макрофагов, является гистопатологическим признаком инфицированной ткани головного мозга. В головном мозге встречаются только трофозоиты [3–5, 29]. Выживаемость при ПАМ зависит от быстрой диагностики и лечения заболевания и имеет место, когда заболевание было распознано на ранней стадии и лечение было начато быстро [30, 31].
1–3, 27–29].Два других вида Naegleria , N. australiensis и N. italica могут вызывать инфекции у мышей, но никогда не были идентифицированы среди людей [5]. Случаи ПАМ, вызванные N. fowleri , также были связаны с загрязненной бытовой водой, используемой для купания [11, 12]. Заражение может произойти, когда амебы проникают в носовые ходы, прикрепляются к обонятельной слизистой оболочке и мигрируют через решетчатую пластинку вдоль обонятельных нервов. Попав в обонятельные луковицы головного мозга, амебы быстро делятся и гибель наступает в течение 7—10 дней. N. fowleri вызывает молниеносный геморрагический некроз головного мозга. Воспалительный инфильтрат, состоящий из нейтрофилов, эозинофилов и макрофагов, является гистопатологическим признаком инфицированной ткани головного мозга. В головном мозге встречаются только трофозоиты [3–5, 29]. Выживаемость при ПАМ зависит от быстрой диагностики и лечения заболевания и имеет место, когда заболевание было распознано на ранней стадии и лечение было начато быстро [30, 31]. Выживаемость повышается при внутривенном применении комбинации амфотерицина В с интратекальным введением амфотерицина В и пероральным введением рифампина и других противогрибковых средств [30–34].Внутривенное введение амфотерицина В и флуконазола с последующим пероральным введением рифампицина привело к успешному лечению ребенка, у которого развился ПАМ [35]. Однако не все пациенты, получавшие амфотерицин В, выживают [36–38]. Другие предположили, что тройная комбинация низкой дозы амфотерицина В, вводимого внутривенно (в/в), с пероральным рифампацином и пероральным кетоконазолом приведет к более благоприятному исходу [39]. Было показано, что азитромицин эффективен против N. fowleri in vitro (культура клеток) и in vivo (мышиная модель инфекции) [8].Однако оптимальное лечение еще предстоит разработать.
Выживаемость повышается при внутривенном применении комбинации амфотерицина В с интратекальным введением амфотерицина В и пероральным введением рифампина и других противогрибковых средств [30–34].Внутривенное введение амфотерицина В и флуконазола с последующим пероральным введением рифампицина привело к успешному лечению ребенка, у которого развился ПАМ [35]. Однако не все пациенты, получавшие амфотерицин В, выживают [36–38]. Другие предположили, что тройная комбинация низкой дозы амфотерицина В, вводимого внутривенно (в/в), с пероральным рифампацином и пероральным кетоконазолом приведет к более благоприятному исходу [39]. Было показано, что азитромицин эффективен против N. fowleri in vitro (культура клеток) и in vivo (мышиная модель инфекции) [8].Однако оптимальное лечение еще предстоит разработать.
2.1. Симптомы инфекции PAM
PAM характеризуется внезапным началом сильной лобной головной боли, лихорадкой, тошнотой, рвотой и ринитом. За ними следуют ригидность затылочных мышц, диплопия, потеря обоняния, спутанность сознания и случайные судороги, быстро прогрессирующие до комы и смерти. Повышенное количество лейкоцитов обычно сопровождается заметным увеличением нейтрофилов. ЦСЖ содержит нейтрофилы, поэтому часто подозревают бактериальную инфекцию [3–5, 7].История, описывающая контакт с теплой водой (ныряние, вейкбординг, катание на водных лыжах), наводит на мысль о ЛАМ [40].
Повышенное количество лейкоцитов обычно сопровождается заметным увеличением нейтрофилов. ЦСЖ содержит нейтрофилы, поэтому часто подозревают бактериальную инфекцию [3–5, 7].История, описывающая контакт с теплой водой (ныряние, вейкбординг, катание на водных лыжах), наводит на мысль о ЛАМ [40].
2.2. Лабораторная диагностика
ЛАМ — редкое заболевание, но почти всегда смертельное. Поэтому для начала лечения важна ранняя диагностика. Заболевание часто неправильно диагностируют, потому что не существует отличительных различий в диагностике, позволяющих отличить ПАМ от бактериального менингоэнцефалита.
2.2.1. Методы визуализации
Компьютерная томография (КТ) или магнитно-резонансная томография (МРТ) показывают поражения, но они неспецифичны [41–43].КТ показывает облитерацию цистерн, окружающих средний мозг и субарахноидальное пространство [3].
2.2.2. Микроскопические методы
Предсмертная диагностика проводится редко, но при низком давлении спинномозговой жидкости можно выполнить люмбальную пункцию.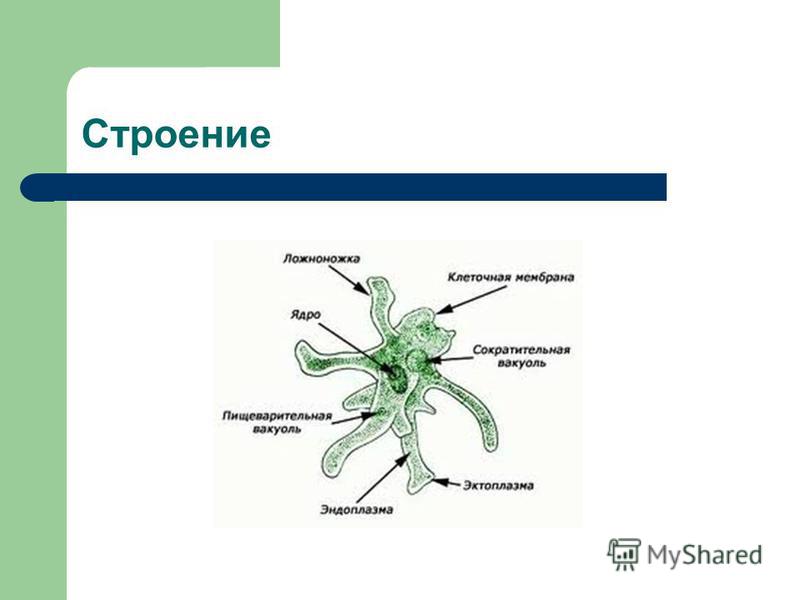 ЦСЖ имеет гнойный характер, и если бактерии не обнаружены, следует заподозрить амебный менингоэнцефалит. ЦСЖ мутная, слегка геморрагическая, с повышенной клеточностью, состоящая в основном из нейтрофилов. ЦСЖ характеризуется низким уровнем глюкозы и повышенным содержанием белка.Прямое микроскопическое исследование спинномозговой жидкости в виде влажного препарата является методом выбора при диагностике ПАМ, поскольку в спинномозговой жидкости содержатся подвижные амебы, которые можно распознать при исследовании под световым микроскопом [5, 44, 45]. N. fowleri можно отличить от других FLA, вызывающих инфекции ЦНС, поскольку амебы могут превращаться в плавающих жгутиконосцев, когда амебы помещаются в воду. Если амебы присутствуют в спинномозговой жидкости, их можно идентифицировать путем окрашивания фиксированных препаратов красителями Райта, Гимзы или гематоксилином и эозином (Н и Е).Хотя окраска по Граму используется в клинических лабораториях для обнаружения бактерий в спинномозговой жидкости, окраска по Граму бесполезна для диагностики амеб, поскольку она не отображает характерную морфологию ядер амеб.
ЦСЖ имеет гнойный характер, и если бактерии не обнаружены, следует заподозрить амебный менингоэнцефалит. ЦСЖ мутная, слегка геморрагическая, с повышенной клеточностью, состоящая в основном из нейтрофилов. ЦСЖ характеризуется низким уровнем глюкозы и повышенным содержанием белка.Прямое микроскопическое исследование спинномозговой жидкости в виде влажного препарата является методом выбора при диагностике ПАМ, поскольку в спинномозговой жидкости содержатся подвижные амебы, которые можно распознать при исследовании под световым микроскопом [5, 44, 45]. N. fowleri можно отличить от других FLA, вызывающих инфекции ЦНС, поскольку амебы могут превращаться в плавающих жгутиконосцев, когда амебы помещаются в воду. Если амебы присутствуют в спинномозговой жидкости, их можно идентифицировать путем окрашивания фиксированных препаратов красителями Райта, Гимзы или гематоксилином и эозином (Н и Е).Хотя окраска по Граму используется в клинических лабораториях для обнаружения бактерий в спинномозговой жидкости, окраска по Граму бесполезна для диагностики амеб, поскольку она не отображает характерную морфологию ядер амеб.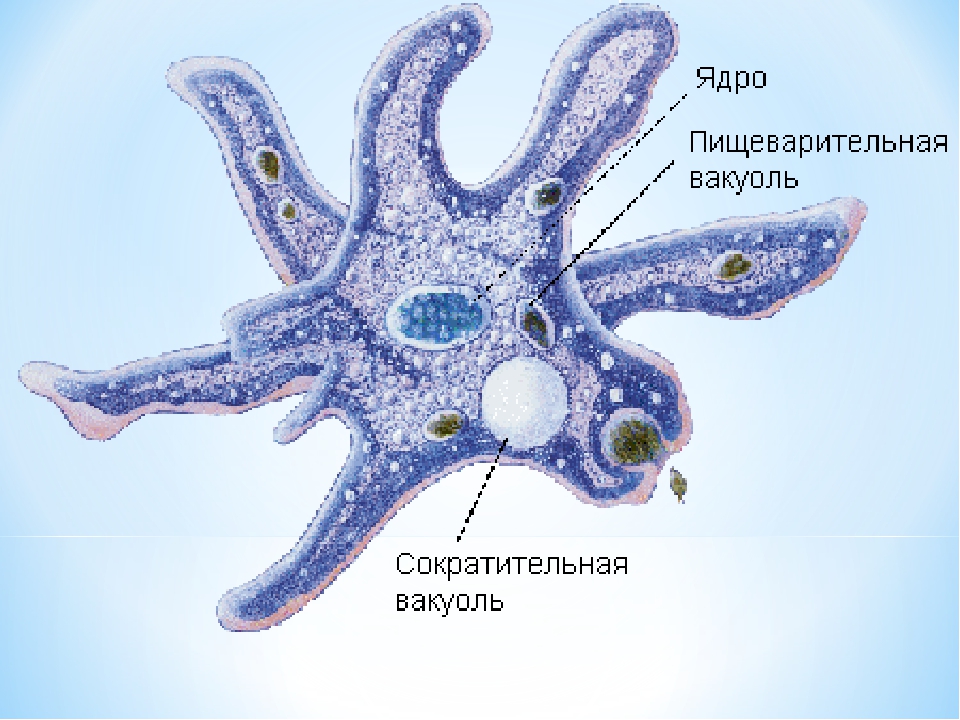 Амебы могут быть ошибочно приняты за макрофаги, но ядро N. fowleri содержит большое центральное круглое ядрышко, которое должно отличать его от клеток-хозяев [3, 27, 44, 45].
Амебы могут быть ошибочно приняты за макрофаги, но ядро N. fowleri содержит большое центральное круглое ядрышко, которое должно отличать его от клеток-хозяев [3, 27, 44, 45].
Поликлональные антитела, полученные у кроликов, или моноклональные антитела можно использовать для идентификации амеб в срезах тканей и ЦСЖ.Амебы в спинномозговой жидкости могут быть идентифицированы с помощью специфического непрямого иммунофлуоресцентного анализа антител с использованием поликлональных или моноклональных антител к амебе в сочетании с флуоресцеиновым вторичным антителом [3, 45, 46].
Материал биопсии следует зафиксировать в 10% нейтральном забуференном формалине для гистологического исследования. Амебы можно наблюдать в биопсии ткани головного мозга после окрашивания H & E или окрашивания иммунопероксидазой с использованием антител к амебам. В тканях головного мозга обнаружены только трофозоиты; кисты не наблюдаются [3].
Для диагностики инфекций можно использовать имеющийся в продаже иммуноферментный анализ (Indicia, Oulin, France), основанный на использовании моноклонального антитела (5D12), которое распознает гликозилированный эпитоп на N.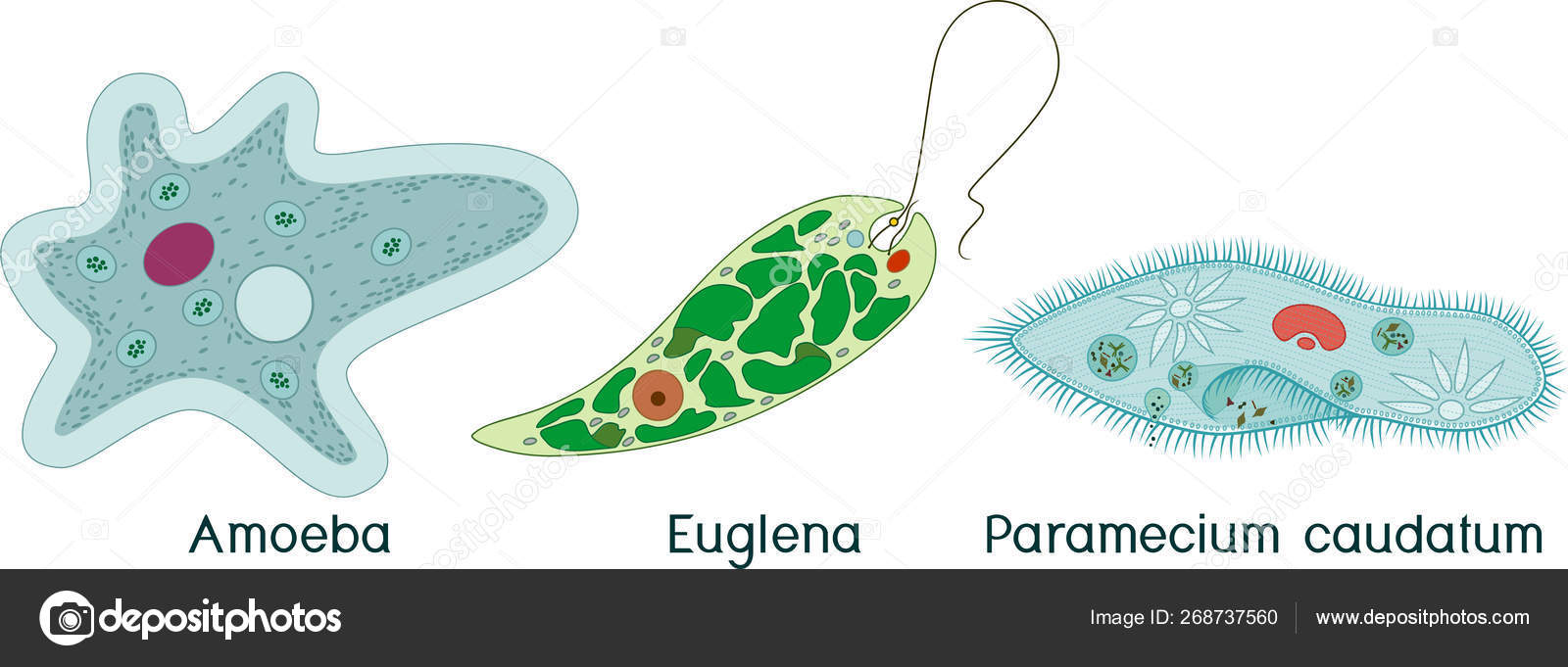 fowleri . Это моноклональное антитело можно использовать для отличия N. fowleri от других видов Naegleria и от других FLA в тканях и образцах окружающей среды [46, 47].
fowleri . Это моноклональное антитело можно использовать для отличия N. fowleri от других видов Naegleria и от других FLA в тканях и образцах окружающей среды [46, 47].
2.2.3. Методы культивирования
СМЖ или биоптат ткани головного мозга следует хранить и транспортировать при комнатной температуре в диагностическую лабораторию.Этот материал можно инокулировать на клетки культуры ткани (Vero, фибробласты) и инкубировать в присутствии антибиотиков пенициллин-стрептомицин. Фунгициды смертельны для амеб. Присутствующие амебы будут размножаться и разрушать монослой через 24–48 часов. Биоптатную ткань также можно поместить на 1,5% непитательный агар, покрытый слоем бактерий ( Escherichia coli ). Амебы выходят из ткани, поглощают бактерии и делятся. Затем амеб можно наблюдать на агаре с помощью инвертированного светового микроскопа [3, 44, 45, 48].
2.2.4. Серология
Антитела к Naegleria spp . были идентифицированы у здоровых людей [49, 50]. Поскольку ПАМ является быстропротекающим заболеванием, серологические тесты на повышение титра антител не всегда полезны. Как правило, повышения титра антител не наблюдается, хотя повышение уровня антител наблюдалось у пациентов, которые успешно лечились и перенесли инфекцию [30].
Поскольку ПАМ является быстропротекающим заболеванием, серологические тесты на повышение титра антител не всегда полезны. Как правило, повышения титра антител не наблюдается, хотя повышение уровня антител наблюдалось у пациентов, которые успешно лечились и перенесли инфекцию [30].
2.2.5. Анализы полимеразной цепной реакции (ПЦР)
В настоящее время в исследовательских лабораториях доступны более быстрые молекулярные методы, но эти методы, как правило, недоступны в большинстве клинических лабораторий.Для обнаружения N. fowleri в клинических образцах и образцах из окружающей среды были разработаны высокоспецифичные и чувствительные ПЦР и ПЦР в реальном времени [51–57]. Был разработан ПЦР-анализ с использованием праймеров для полной рибосомной внутренней транскрибируемой спейсерной области (ITS), который позволяет различать видов Naegleria , а видоспецифический анализ позволяет обнаруживать N. fowleri [51, 52]. Также сообщалось о ПЦР-анализе, который обнаруживает N. fowleri в свежей ткани мозга, а также в фиксированной формалином ткани мозга, залитой в парафин [56].Недавно Qvarnstrom et al. [57] разработали быстрый и чувствительный мультиплексный анализ ПЦР в реальном времени, основанный на использовании зондов, нацеленных на частичные или полноразмерные гены малых субъединиц ядерной рибосомы (ген 18S рРНК) для одновременного обнаружения Naegleria . , Balamuthia, и Acanthamoeba [57]. Этот анализ ПЦР видоспецифичен для N. fowleri и B. mandrillaris и родоспецифичен для Acanthamoeba. Таким образом, этот анализ может определить, какие амебы присутствуют в образце спинномозговой жидкости или образце биопсии головного мозга пациента с амебным энцефалитом.Было показано, что предел обнаружения для этого анализа составляет одну амебу на образец.
fowleri в свежей ткани мозга, а также в фиксированной формалином ткани мозга, залитой в парафин [56].Недавно Qvarnstrom et al. [57] разработали быстрый и чувствительный мультиплексный анализ ПЦР в реальном времени, основанный на использовании зондов, нацеленных на частичные или полноразмерные гены малых субъединиц ядерной рибосомы (ген 18S рРНК) для одновременного обнаружения Naegleria . , Balamuthia, и Acanthamoeba [57]. Этот анализ ПЦР видоспецифичен для N. fowleri и B. mandrillaris и родоспецифичен для Acanthamoeba. Таким образом, этот анализ может определить, какие амебы присутствуют в образце спинномозговой жидкости или образце биопсии головного мозга пациента с амебным энцефалитом.Было показано, что предел обнаружения для этого анализа составляет одну амебу на образец.
3.
Acanthamoeba spp. Acanthamoeba — одна из наиболее часто выделяемых амеб в пробах окружающей среды. Acanthamoeba распространена повсеместно и встречается в различных средах обитания, включая бытовое водоснабжение, больничную воду, стоматологические установки, воздух, почву и воду.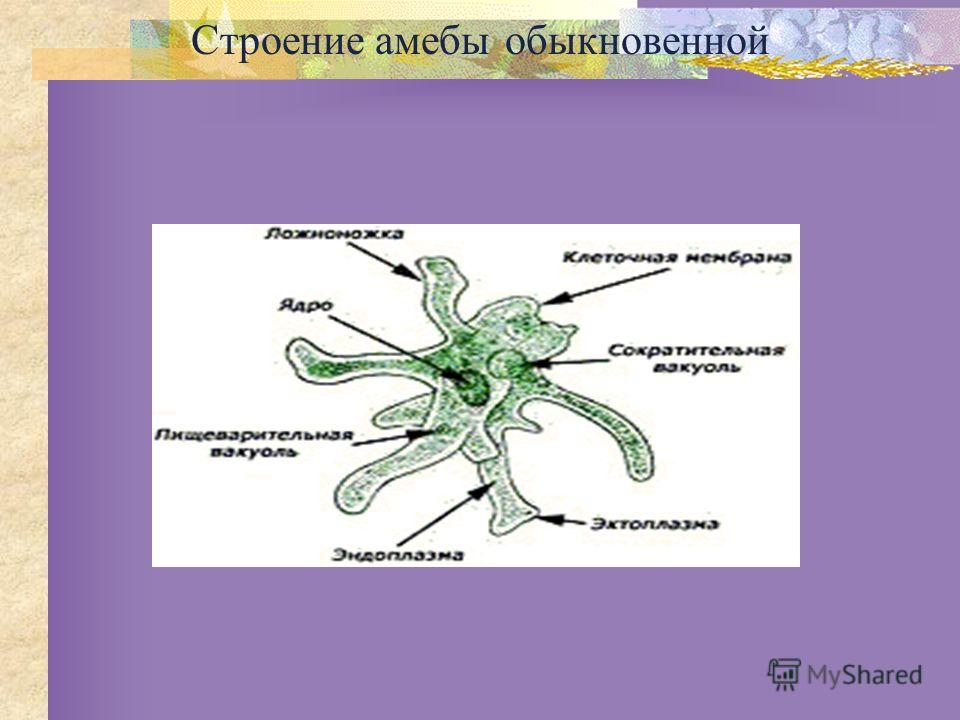 Acanthamoeba имеет две морфологические формы в своем жизненном цикле: трофозоит (рис. 1(b)) и стадию цисты.Обе стадии можно обнаружить в тканях инфицированных людей и в окружающей среде. Трофозоит является делящейся формой и считается инфекционной стадией. Кисты находятся в состоянии покоя и защищают амеб от вредных воздействий окружающей среды. Цисты устойчивы к биоцидам, хлорированию и антибиотикам. Несколько видов Acanthamoeba могут вызывать гранулематозный амебный энцефалит (ГАЭ), кожный акантамебиаз или амебный кератит (АК). АК представляет собой угрожающую зрению инфекцию роговицы, которая возникает у иммунокомпетентных лиц, в основном у пользователей контактных линз.GAE, также известный как Acanthamoeba Гранулематозный энцефалит (AGE), представляет собой редкую хроническую прогрессирующую инфекцию ЦНС, которая может поражать легкие [58]. ГЭ обычно связана с основным изнурительным заболеванием или у лиц с подавленным иммунитетом, включая пациентов с ВИЧ-СПИДом, диабетиков, лиц, перенесших трансплантацию органов или химиотерапию рака, и лиц, злоупотребляющих наркотиками [6, 7, 58, 59].
Acanthamoeba имеет две морфологические формы в своем жизненном цикле: трофозоит (рис. 1(b)) и стадию цисты.Обе стадии можно обнаружить в тканях инфицированных людей и в окружающей среде. Трофозоит является делящейся формой и считается инфекционной стадией. Кисты находятся в состоянии покоя и защищают амеб от вредных воздействий окружающей среды. Цисты устойчивы к биоцидам, хлорированию и антибиотикам. Несколько видов Acanthamoeba могут вызывать гранулематозный амебный энцефалит (ГАЭ), кожный акантамебиаз или амебный кератит (АК). АК представляет собой угрожающую зрению инфекцию роговицы, которая возникает у иммунокомпетентных лиц, в основном у пользователей контактных линз.GAE, также известный как Acanthamoeba Гранулематозный энцефалит (AGE), представляет собой редкую хроническую прогрессирующую инфекцию ЦНС, которая может поражать легкие [58]. ГЭ обычно связана с основным изнурительным заболеванием или у лиц с подавленным иммунитетом, включая пациентов с ВИЧ-СПИДом, диабетиков, лиц, перенесших трансплантацию органов или химиотерапию рака, и лиц, злоупотребляющих наркотиками [6, 7, 58, 59]. Также были описаны поражения кожи, вызванные Acanthamoeba . О кожном акантамебиазе чаще сообщалось у ВИЧ-позитивных пациентов, чем при других состояниях [60–66].Проявления кожной инфекции включают наличие многочисленных твердых эритематозных узелков, папул или язв по всему телу пациента [62–67]. Наличие как поражений кожи, так и симптомов со стороны ЦНС, возникающих одновременно, может указывать на инфекцию Acanthamoeba [58].
Также были описаны поражения кожи, вызванные Acanthamoeba . О кожном акантамебиазе чаще сообщалось у ВИЧ-позитивных пациентов, чем при других состояниях [60–66].Проявления кожной инфекции включают наличие многочисленных твердых эритематозных узелков, папул или язв по всему телу пациента [62–67]. Наличие как поражений кожи, так и симптомов со стороны ЦНС, возникающих одновременно, может указывать на инфекцию Acanthamoeba [58].
3.1. Симптомы GAE
Симптомы GAE включают головные боли, небольшую лихорадку, судороги, гемипарезы, параличи черепных нервов, изменения личности, тошноту, ригидность затылочных мышц, угнетение сознания и кому, типичные клинические признаки локализованной энцефалопатии [5–8].Клинические признаки ГАЭ неспецифичны. Так, заболевание часто ошибочно диагностируют как бактериальный лептоменингит, туберкулезный менингит, вирусный энцефалит, токсоплазмоз, грибковые инфекции, нейроцистицеркоз или опухоль головного мозга [5, 6, 68–71].
3.
 2. Методы диагностики
2. Методы диагностики3.2.1. Методы визуализации
Методы визуализации головного мозга, такие как КТ и МРТ, использовались для визуализации поражений головного мозга, вызванных Acanthamoeba , но сами по себе поражения не являются специфическими для GAE [41–43].Мультифокальные поражения низкой плотности как в корковых, так и в подкорковых областях головного мозга можно наблюдать с помощью компьютерной томографии. Усиленная КТ обычно показывает наличие прогрессирующей гидроцефалии с утолщением мозговых оболочек, псевдоопухолевыми поражениями, крупными изолированными поражениями или множественными овальными поражениями. Многоочаговые поражения, отек и множественные кольцевидные поражения обычно наблюдаются у пациентов с ГЭ при МРТ. Несмотря на эти характеристики, как КТ, так и МРТ имеют ограниченную диагностическую ценность для ЭГА [5, 7, 41–43, 68–73].
3.2.2. Микроскопические методы
Окончательным диагнозом ЭГА является обнаружение амебы в ткани или выделение амебы. Для визуального обнаружения как трофозоитов Acanthamoeba , так и цист в тканях головного мозга, поражениях кожи или спинномозговой жидкости (ЦСЖ) можно использовать как световую, так и электронную микроскопию. Трофозоиты Acanthamoeba можно отличить от воспалительных клеток-хозяев, таких как макрофаги, в основном по их ядерной структуре, поскольку Acanthamoeba обладает округлым ядром и большим центральным ядрышком, образующим гало [3, 6, 59].Однако отличить трофозоиты Acanthamoeba от патогенных трофозоитов B. mandrillaris с помощью световой микроскопии не представляется возможным, так как обе амебы имеют одинаковое строение ядра [7]. Образцы биопсии или аутопсии должны быть фиксированы формалином, залиты парафином и окрашены гематоксилин-эозином [74]. Использовались и другие типы гистологического окрашивания, в том числе периодическая кислота Shiff, метенаминовое серебро Гомори или трихом. Эти пятна оказались эффективными для идентификации кист [75, 76].
Для визуального обнаружения как трофозоитов Acanthamoeba , так и цист в тканях головного мозга, поражениях кожи или спинномозговой жидкости (ЦСЖ) можно использовать как световую, так и электронную микроскопию. Трофозоиты Acanthamoeba можно отличить от воспалительных клеток-хозяев, таких как макрофаги, в основном по их ядерной структуре, поскольку Acanthamoeba обладает округлым ядром и большим центральным ядрышком, образующим гало [3, 6, 59].Однако отличить трофозоиты Acanthamoeba от патогенных трофозоитов B. mandrillaris с помощью световой микроскопии не представляется возможным, так как обе амебы имеют одинаковое строение ядра [7]. Образцы биопсии или аутопсии должны быть фиксированы формалином, залиты парафином и окрашены гематоксилин-эозином [74]. Использовались и другие типы гистологического окрашивания, в том числе периодическая кислота Shiff, метенаминовое серебро Гомори или трихом. Эти пятна оказались эффективными для идентификации кист [75, 76]. Акридиновый оранжевый и калькофлюор белый успешно использовались для наблюдения цист Acanthamoeba в тканях [6, 77, 78].
Акридиновый оранжевый и калькофлюор белый успешно использовались для наблюдения цист Acanthamoeba в тканях [6, 77, 78].
Гранулемы головного мозга, некроз с наличием многоядерных клеток, воспалительные инфильтраты и амебы (как трофозоиты, так и кисты), окружающие кровеносные сосуды [3–5], могут наблюдаться в биопсийной ткани, окрашенной H&E. обнаружены в ЦСЖ во влажных препаратах или окрашенных по Гимзе предметных стеклах отложений ЦСЖ [29, 79, 80]. Для диагностики кожного акантамебиаза световая микроскопия окрашенных гематоксилин-эозином биоптатов кожи демонстрирует наличие гранулем, участков некроза, воспалительных инфильтратов и васкулитов, содержащих как трофозоиты, так и кисты Acanthamoeba [62, 64, 67, 75, 81].
Использование как флуоресцентной микроскопии, так и иммуногистохимии в срезах тканей головного мозга и кожи является эффективным методом специфического обнаружения Acanthamoeba [82–87]. Антитела к Acanthamoeba , полученные у кроликов, можно использовать для идентификации амеб в тканях.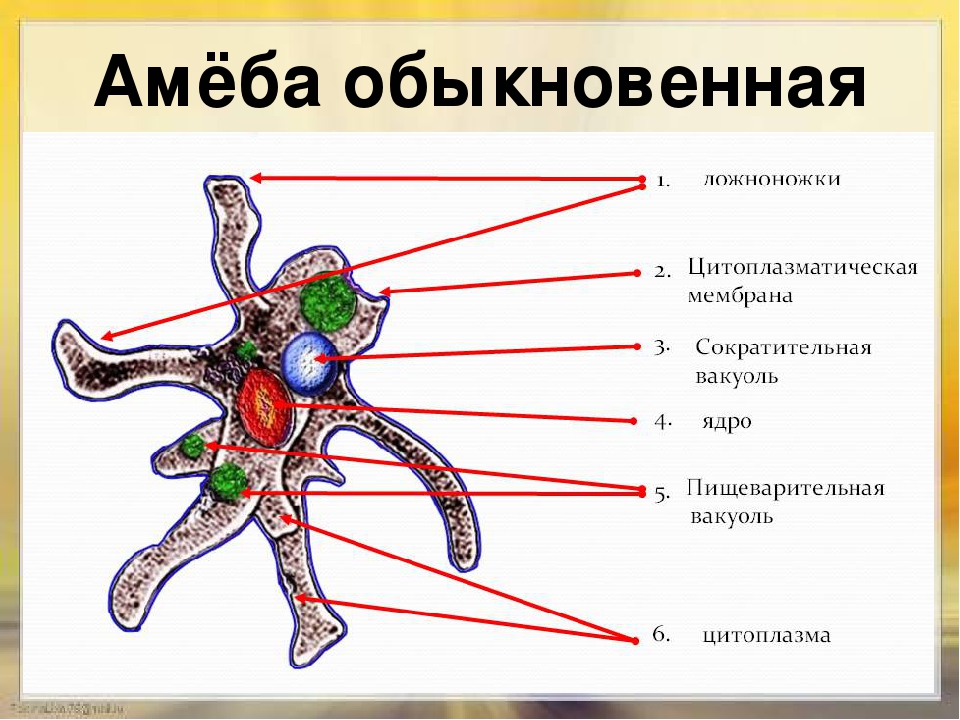 Образцы пациентов инкубируют с антителами против Acanthamoeba с последующим введением вторичного антикроличьего IgG, связанного с флуоресцентным маркером, для обнаружения как трофозоитов, так и цист [74]. Совсем недавно были разработаны видоспецифические моноклональные антитела для использования в качестве важного диагностического и эпидемиологического инструмента.Эти моноклональные антитела распознают A. castellanii, A. polyphaga, A. lenticulata и A. culbertsoni , реагируют с фиксированными формалином и залитыми парафином инфицированными тканями и распознают как трофозоиты, так и цисты амеб [88]. ].
Образцы пациентов инкубируют с антителами против Acanthamoeba с последующим введением вторичного антикроличьего IgG, связанного с флуоресцентным маркером, для обнаружения как трофозоитов, так и цист [74]. Совсем недавно были разработаны видоспецифические моноклональные антитела для использования в качестве важного диагностического и эпидемиологического инструмента.Эти моноклональные антитела распознают A. castellanii, A. polyphaga, A. lenticulata и A. culbertsoni , реагируют с фиксированными формалином и залитыми парафином инфицированными тканями и распознают как трофозоиты, так и цисты амеб [88]. ].
Просвечивающая электронная микроскопия (ПЭМ) также может быть использована в качестве инструмента для дифференциации цист и трофозоитов Acanthamoeba от клеток-хозяев и от других амеб, таких как B. mandrillaris [63, 70, 89, 90].Однако этот метод дорог; Подготовка проб занимает много времени и требует наличия квалифицированного персонала.
3.
 2.3. Методы культивирования
2.3. Методы культивированияВыделение и культивирование Acanthamoeba можно проводить путем помещения образцов биопсии/аутопсии головного мозга или кожи на чашки с 1,5% непитательным агаром, покрытые слоем Escherichia coli или Enterobacter aerogenes [3, 7, 48, 58, 83, 91]. В зависимости от плотности амеб Acanthamoeba можно наблюдать через 24 часа после инокуляции.Образцы также можно помещать на клетки тканевой культуры в присутствии антибиотиков (пенициллин-стрептомицин и гентамицин), в этом случае амебы разрушают монослой клеток за 24–48 часов в зависимости от количества присутствующих амеб [3, 6, 59]. ].
3.2.4. Серология
Для выявления инфекции Acanthamoeba увеличение титра антител может указывать на инфекцию. Для этой оценки проводят непрямую иммунофлуоресценцию (IIF) с использованием серийных разведений сыворотки человека с подозрением на акантамебиаз с последующей инкубацией сыворотки с предметными стеклами, содержащими фиксированные амебы или экстракты амеб. Обнаружение может быть достигнуто путем добавления анти-IgG-антитела, связанного с флуоресцентной меткой, такой как FITC, а обнаружение и титрование антител может быть определено с помощью флуоресцентной микроскопии [92]. Acanthamoeba инфицированные лица обладают высокими титрами антител (от 1 : 256 до 1 : 1024) в сыворотке крови, в то время как здоровые люди, подвергшиеся воздействию Acanthamoeba в окружающей среде, имеют низкие титры антител, обычно не выше 1 : 2 [8, 80]. 92–96]. Таким образом, IIF может быть полезным инструментом для подтверждения инфекции у пациентов с подозрением на инфекцию Acanthamoeba [96].Вестерн-иммуноблот-анализ также использовался для выявления антител к Acanthamoeba в сыворотке крови человека [97]. Был разработан метод ELISA с использованием цельных фиксированных трофозоитов, а не разрушенных амебных экстрактов в качестве источника антигена, который показал себя как эффективный инструмент для идентификации антител к Acanthamoeba в условиях клинической лаборатории [98].
Обнаружение может быть достигнуто путем добавления анти-IgG-антитела, связанного с флуоресцентной меткой, такой как FITC, а обнаружение и титрование антител может быть определено с помощью флуоресцентной микроскопии [92]. Acanthamoeba инфицированные лица обладают высокими титрами антител (от 1 : 256 до 1 : 1024) в сыворотке крови, в то время как здоровые люди, подвергшиеся воздействию Acanthamoeba в окружающей среде, имеют низкие титры антител, обычно не выше 1 : 2 [8, 80]. 92–96]. Таким образом, IIF может быть полезным инструментом для подтверждения инфекции у пациентов с подозрением на инфекцию Acanthamoeba [96].Вестерн-иммуноблот-анализ также использовался для выявления антител к Acanthamoeba в сыворотке крови человека [97]. Был разработан метод ELISA с использованием цельных фиксированных трофозоитов, а не разрушенных амебных экстрактов в качестве источника антигена, который показал себя как эффективный инструмент для идентификации антител к Acanthamoeba в условиях клинической лаборатории [98].
3.2.5. ПЦР
Обнаружение Acanthamoeba может быть быстро достигнуто с помощью молекулярных методов.В диагностических целях обнаружения Acanthamoeba на уровне рода достаточно, чтобы определить, инфицирован ли человек. Молекулярная идентификация Acanthamoeba может быть выполнена с помощью полимеразной цепной реакции [99–101]. Полная последовательность гена ДНК гена 18S рибосомной РНК (18S рДНК) позволила разработать надежную пару праймеров, специфичных для рода Acanthamoeba , названных JDP1 и JDP2. Использование JDP1 (прямой праймер) -GGCCCAGATCGTTTACCGTGAA и JDP2 (обратный праймер) -TCTCACAAGCTGCTAGGGAGTCA, соответственно, дает специфический амплимер размером 500 пар оснований, называемый ASA.С1. Преимущество этого ПЦР-анализа заключается в том, что он выявляет все известные подгрупп Acanthamoeba [100]. Этот ПЦР-анализ был успешно использован для обнаружения Acanthamoeba в окружающей среде, а также у пациентов с ГЭ и кожным акантамебиазом [83, 102–105]. Однако амплификация ДНК из цист представляет собой хлопотную задачу. Так, было предложено, чтобы обработка кист протеиназой К перед выделением ДНК увеличивала положительные результаты анализов ПЦР [106].
Однако амплификация ДНК из цист представляет собой хлопотную задачу. Так, было предложено, чтобы обработка кист протеиназой К перед выделением ДНК увеличивала положительные результаты анализов ПЦР [106].
Исследования показали, что большинство амеб, вызывающих ГАЭ и АК, имеют специфический продукт ПЦР (эти амебы включены в подгруппу, называемую Т4), при использовании праймеров для амплификации генов малых субъединиц рРНК (SSU рДНК) [107] .ПЦР митохондриальной ДНК также успешно применялась для обнаружения Acanthamoeba в срезах головного мозга и спинномозговой жидкости [108, 109]. ПЦР в реальном времени использовалась как быстрый инструмент для дифференциальной идентификации свободноживущих амеб и дифференциации Acanthamoeba от N. fowleri и B. mandrillaris [57, 110] . ПЦР в реальном времени, разработанный Rivière et al. [110], использует технологию Taqman для обнаружения 18S рибосомной ДНК (рДНК). Этот анализ, основанный на генотипе Acanthamoeba T4, не выявляет другие генотипы, такие как T10. ПЦР в реальном времени, разработанный Qvarnstrom et al. [57], представляет собой тройной анализ для различения Acanthamoeba от других патогенных FLA и является более полным, поскольку он был разработан для обнаружения более широкого диапазона генотипов Acanthamoeba . Оба анализа в реальном времени были проверены путем тестирования ряда положительных и отрицательных клинических образцов [111].
ПЦР в реальном времени, разработанный Qvarnstrom et al. [57], представляет собой тройной анализ для различения Acanthamoeba от других патогенных FLA и является более полным, поскольку он был разработан для обнаружения более широкого диапазона генотипов Acanthamoeba . Оба анализа в реальном времени были проверены путем тестирования ряда положительных и отрицательных клинических образцов [111].
4.
Acanthamoeba spp. и амебный кератит (АК)В отличие от ГЭ, который является хронической инфекцией, АК представляет собой острую болезненную инфекцию, которая может возникать у иммунокомпетентных лиц.Это заболевание связано с использованием контактных линз или предыдущей травмой роговицы. При несвоевременном лечении АК может наступить потеря остроты зрения и слепота [6, 7, 58, 59, 112–116].
4.1. Симптомы АК
Начальные симптомы АК неспецифичны и включают непропорциональную боль в глазах, светобоязнь, покраснение глаз и слезотечение, обычно поражающие один глаз [116].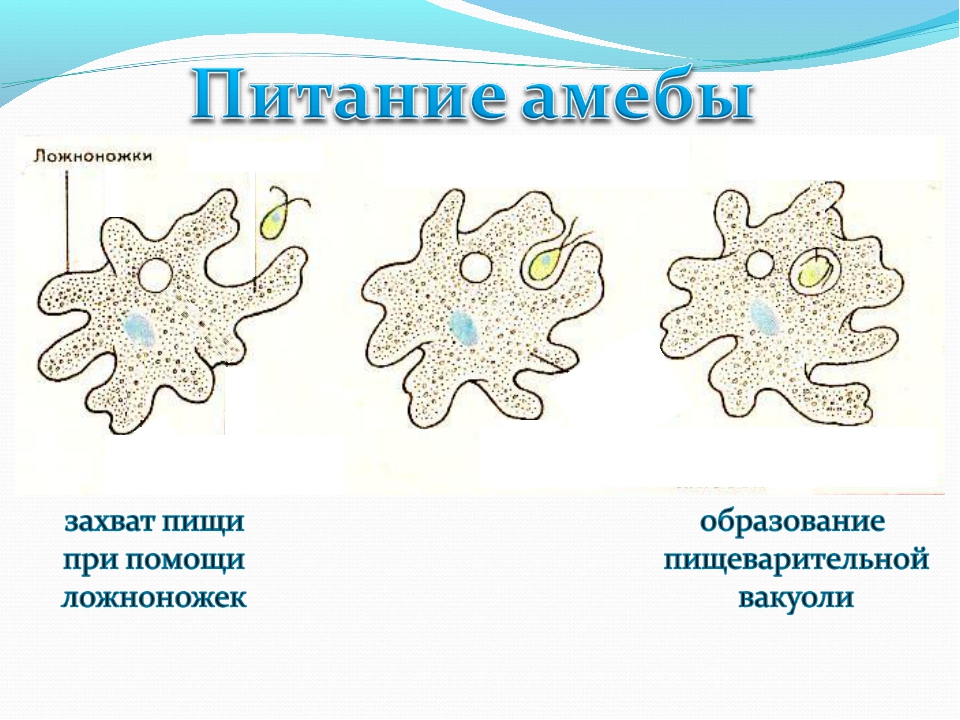 Однако двусторонний АК был описан как осложнение первичной инфекции [116]. С помощью щелевой лампы можно наблюдать воспаление роговицы, приводящее к образованию кольцевидного стромального инфильтрата.Кроме того, присутствуют эрозия эпителия роговицы, неровности и отек. Для АК характерно радиальное периневральное распространение инфильтрата (радиальный кератоневрит), сходное с типом инфильтрации, наблюдаемым при кератите Pseudomonas aeruginosa [112–115, 117–120]. Более поздние стадии инфекции могут привести к оголению эпителия и некрозу стромы. Использование контактных линз и/или случаи травмы роговицы являются сильными индикаторами АК [121–123]. Несмотря на клиническую картину, АК часто неправильно диагностируют как герпес или бактериальные кератиты, которые имеют сходную клиническую симптоматику [114, 124, 125].
Однако двусторонний АК был описан как осложнение первичной инфекции [116]. С помощью щелевой лампы можно наблюдать воспаление роговицы, приводящее к образованию кольцевидного стромального инфильтрата.Кроме того, присутствуют эрозия эпителия роговицы, неровности и отек. Для АК характерно радиальное периневральное распространение инфильтрата (радиальный кератоневрит), сходное с типом инфильтрации, наблюдаемым при кератите Pseudomonas aeruginosa [112–115, 117–120]. Более поздние стадии инфекции могут привести к оголению эпителия и некрозу стромы. Использование контактных линз и/или случаи травмы роговицы являются сильными индикаторами АК [121–123]. Несмотря на клиническую картину, АК часто неправильно диагностируют как герпес или бактериальные кератиты, которые имеют сходную клиническую симптоматику [114, 124, 125].
4.2. Диагностические методы
Диагноз АК может быть поставлен на основании анализа клинических проявлений роговицы и выявления амеб в роговице [112, 125].
4.2.1. Микроскопические методы
Обнаружение Acanthamoeba может быть достигнуто путем анализа биопсии роговицы. Однако соскоб с роговицы был эффективным и неинвазивным методом, используемым для выделения амеб и диагностики кератита Acanthamoeba [126]. После соскоба образцы можно размазать по предметным стеклам.Световая микроскопия является эффективным средством обнаружения Acanthamoeba в соскобах роговицы, в образцах биопсии и образцах кератопластики. Acanthamoeba можно обнаружить во влажных препаратах соскоба с роговицы с использованием 10% КОН [126–132]. Более того, импрессионная цитология позволила удалить образцы амеб у пациента с АК [132]. Окрашивание H & E и красители Гимзы успешно использовались для обнаружения как трофозоитов, так и цист Acanthamoeba . Кисты, окрашенные по Гимзе или Н&Е, прозрачные, яркие, с кистами полиэдрической или звездчатой формы, в то время как трофозоиты с центральным ядрышком и вакуолями обнаружить труднее, так как они могут напоминать воспалительные клетки [112, 115, 129, 130, 133–135]. Окрашивание PAS также использовалось для обнаружения кист [128, 136]. Как описано ранее, анализы IIF и иммунофлуоресцентную микроскопию можно использовать для обнаружения Acanthamoeba в тканях мозга и кожи, а также в образцах роговицы, контактных линзах и контейнерах для линз [134, 137, 138]. Иммунопероксидазный метод также эффективен для обнаружения Acanthamoeba в ткани хозяина [121, 134]. Цисты и трофозоиты легко выявляются с помощью просвечивающей электронной микроскопии [133].
Окрашивание PAS также использовалось для обнаружения кист [128, 136]. Как описано ранее, анализы IIF и иммунофлуоресцентную микроскопию можно использовать для обнаружения Acanthamoeba в тканях мозга и кожи, а также в образцах роговицы, контактных линзах и контейнерах для линз [134, 137, 138]. Иммунопероксидазный метод также эффективен для обнаружения Acanthamoeba в ткани хозяина [121, 134]. Цисты и трофозоиты легко выявляются с помощью просвечивающей электронной микроскопии [133].
Флуоресцентные красители использовались для обнаружения кист Acanthamoeba в образцах роговицы.Calcofluor white представляет собой флуоресцентное соединение, способное связывать целлюлозу в клеточной стенке цист Acanthamoeba , обнаруженных в соскобах роговицы и срезах ткани роговицы, залитых парафином [138, 139]. Кроме того, образцы, ранее окрашенные гематоксилин-эозином и другими красителями, могут быть впоследствии окрашены калькофлуором белым, что является быстрым и эффективным, но требует флуоресцентного микроскопа. Также важно отметить, что при смешанных грибковых инфекциях Acanthamoeba окрашиваются оба возбудителя, поскольку амебные цисты и клеточные стенки грибов являются возможными мишенями для calcofluor white.Акридиновый оранжевый, другой флуорохроматический краситель, использовался для экспресс-диагностики АК [78]. Использование флуоресцентных красителей может привести к ложноположительному окрашиванию клеточного дебриса [129, 130]; поэтому для правильной диагностики необходимы опытный наблюдатель и флуоресцентный микроскоп.
Также важно отметить, что при смешанных грибковых инфекциях Acanthamoeba окрашиваются оба возбудителя, поскольку амебные цисты и клеточные стенки грибов являются возможными мишенями для calcofluor white.Акридиновый оранжевый, другой флуорохроматический краситель, использовался для экспресс-диагностики АК [78]. Использование флуоресцентных красителей может привести к ложноположительному окрашиванию клеточного дебриса [129, 130]; поэтому для правильной диагностики необходимы опытный наблюдатель и флуоресцентный микроскоп.
Использование конфокальной микроскопии in vivo для обнаружения Acanthamoeba в ткани роговицы на определенной глубине в режиме реального времени без какой-либо инвазивной процедуры успешно применялось для диагностики АК [140–148].Во время недавней вспышки в районе Чикаго в сочетании с другими методами, такими как культивирование или световая микроскопия, конфокальная микроскопия использовалась для обнаружения Acanthamoeba [148]. Обычно наблюдаются высококонтрастные округлые тела, свидетельствующие об амебных кистах [143, 146, 147]. Были представлены ограничения конфокальной микроскопии как окончательного диагностического инструмента для АК [149]. Однако исследования показали, что при использовании конфокальной микроскопии опытными операторами этот метод является чувствительным и специфичным для обнаружения Acanthamoeba в ткани роговицы [148].
Были представлены ограничения конфокальной микроскопии как окончательного диагностического инструмента для АК [149]. Однако исследования показали, что при использовании конфокальной микроскопии опытными операторами этот метод является чувствительным и специфичным для обнаружения Acanthamoeba в ткани роговицы [148].
4.2.2. Методы культивирования
Как описано ранее для GAE, культивирование амеб из биоптатов роговицы или соскобов и смывов с контактных линз или контейнеров для линз по-прежнему является наиболее распространенным, дешевым и эффективным методом обнаружения Acanthamoeba . Образцы инфицированной роговицы инокулируют на чашки с 1,5% непитательным агаром, покрытые E. coli или другими немукоидными бактериями. Чашки можно инкубировать при 28–30°С от нескольких дней до нескольких недель, что зависит от количества амеб в образцах.Наличие амеб можно проверить с помощью инвертированного микроскопа [48, 150].
4.3. PCR
Как описано ранее для GAE и кожных поражений, ПЦР-зонды также использовались для подтверждения присутствия амеб в биоптатах и соскобах роговицы, контактных линзах, контейнерах для линз, растворах для линз, а также в окружающей среде [57, 100 , 110, 151, 152].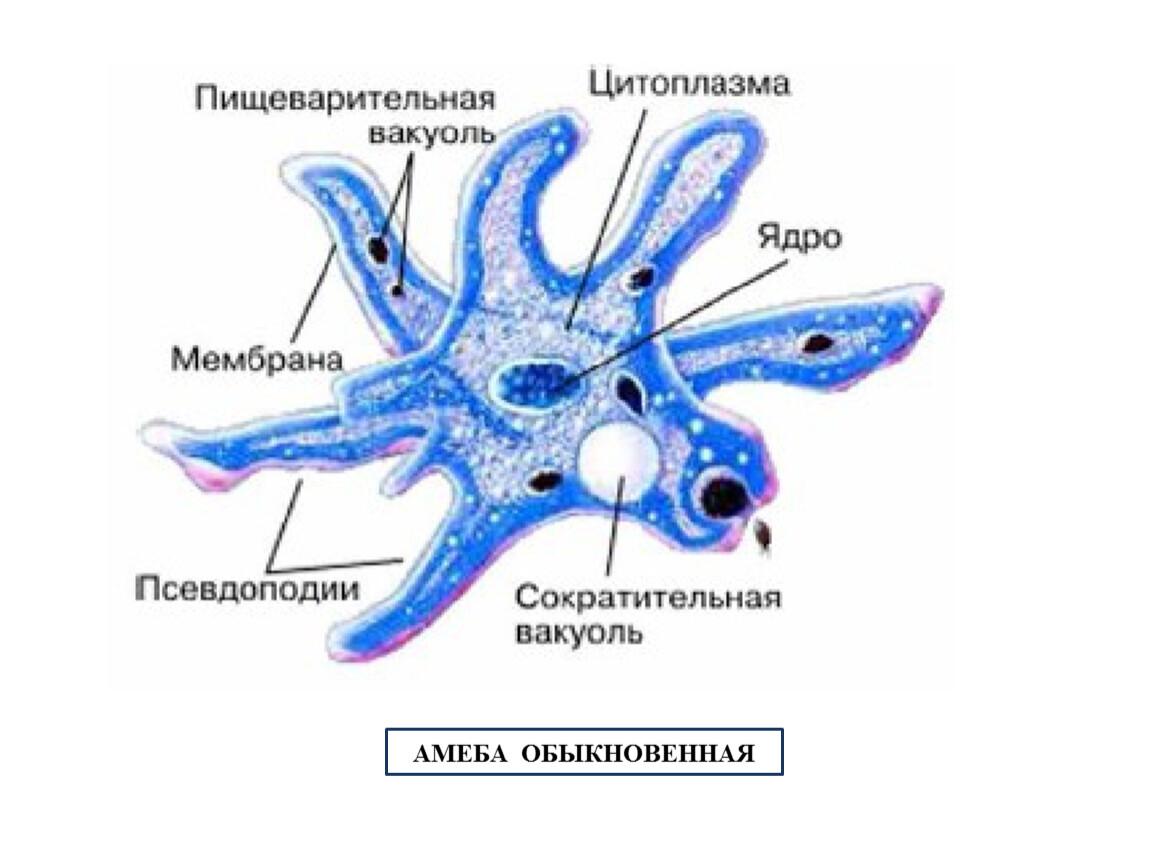 Кроме того, ПЦР является эффективным методом обнаружения Acanthamoeba в образцах слезы [153], полностью неинвазивным методом диагностики АК.Чувствительность методов ПЦР для диагностики АК сравнивали с прямым микроскопическим исследованием и культурой, и сообщалось, что ПЦР более чувствительна, чем морфологическое обнаружение [154]. Два метода ПЦР в реальном времени были утверждены для использования в качестве диагностических тестов на АК [57, 110]. Однако было показано, что ряд широко используемых местных офтальмологических препаратов оказывает ингибирующее действие на ПЦР-анализ [155]. Таким образом, важно, чтобы офтальмологи тщательно промывали поверхность глаза, чтобы удалить любые ингибирующие вещества, чтобы свести к минимуму риск ошибочного диагноза из-за ложноотрицательных результатов ПЦР.
Кроме того, ПЦР является эффективным методом обнаружения Acanthamoeba в образцах слезы [153], полностью неинвазивным методом диагностики АК.Чувствительность методов ПЦР для диагностики АК сравнивали с прямым микроскопическим исследованием и культурой, и сообщалось, что ПЦР более чувствительна, чем морфологическое обнаружение [154]. Два метода ПЦР в реальном времени были утверждены для использования в качестве диагностических тестов на АК [57, 110]. Однако было показано, что ряд широко используемых местных офтальмологических препаратов оказывает ингибирующее действие на ПЦР-анализ [155]. Таким образом, важно, чтобы офтальмологи тщательно промывали поверхность глаза, чтобы удалить любые ингибирующие вещества, чтобы свести к минимуму риск ошибочного диагноза из-за ложноотрицательных результатов ПЦР.
5.
Balamuthia mandrillaris и Balamuthia Амебный энцефалит (ВАЭ) B. mandrillaris — единственный вид Balamuthia , который, как известно, вызывает инфекцию у людей и животных. Balamuthia была впервые выделена из мозга мандрилового бабуина, умершего в зоопарке Сан-Диего от менингоэнцефалита. Амебы сначала были описаны как лептомиксидные амебы, но позже идентифицированы и названы Balamuthia mandrillairs [156–159].Жизненный цикл Balamuthia состоит из трофозоита (рис. 1(c)) и состояния цисты. Амеба встречается в почве, но ее присутствие в воде было предложено на основании случаев БЭ, возникающих у животных и людей, которые в анамнезе плавали в стоячей воде [160–163]. Хотя Balamuthia считается оппортунистическим патогеном, он может вызывать заболевание как у людей с ослабленным иммунитетом, так и у иммунокомпетентных лиц. Инфекции могут возникать у детей и взрослых. Инкубационный период БАЭ удлиняется, поэтому источник и способ заражения окончательно не установлены.Было высказано предположение, что ворота проникновения могут быть через кожные поражения, носовые ходы или вдыхание через дыхательные пути с последующим гематогенным распространением в мозг и другие органы [159].
Balamuthia была впервые выделена из мозга мандрилового бабуина, умершего в зоопарке Сан-Диего от менингоэнцефалита. Амебы сначала были описаны как лептомиксидные амебы, но позже идентифицированы и названы Balamuthia mandrillairs [156–159].Жизненный цикл Balamuthia состоит из трофозоита (рис. 1(c)) и состояния цисты. Амеба встречается в почве, но ее присутствие в воде было предложено на основании случаев БЭ, возникающих у животных и людей, которые в анамнезе плавали в стоячей воде [160–163]. Хотя Balamuthia считается оппортунистическим патогеном, он может вызывать заболевание как у людей с ослабленным иммунитетом, так и у иммунокомпетентных лиц. Инфекции могут возникать у детей и взрослых. Инкубационный период БАЭ удлиняется, поэтому источник и способ заражения окончательно не установлены.Было высказано предположение, что ворота проникновения могут быть через кожные поражения, носовые ходы или вдыхание через дыхательные пути с последующим гематогенным распространением в мозг и другие органы [159].
5.1. Симптомы BAE
Энцефалит, вызываемый B. mandrillaris , также является редким заболеванием с неспецифическими симптомами. Симптомы BAE являются хроническими и включают головные боли, тошноту, рвоту, лихорадку, миалгию, судороги, потерю веса, гемипарез и трудности с речью, обычно связанные с предшествующими гранулематозными поражениями кожи.Вышеуказанные симптомы сбивают с толку, так как они сходны с другими инфекциями головного мозга, включая туберкулез, токсоплазмоз, цистицеркоз, менингит, а также с опухолями головного мозга [7, 158, 164–169].
5.2. Методы диагностики
5.2.1. Методы визуализации
Поражения головного мозга, вызванные B. mandrillaris , можно обнаружить с помощью нейровизуализации, такой как компьютерная томография и МРТ [170–172]. Могут наблюдаться очаговые усиливающие поражения, кистозные поражения, отек и гидроцефалия [172, 173].Поражения могут имитировать другие типы заболеваний, такие как глиомы, абсцессы головного мозга и гематомы [7]. Таким образом, отсутствие специфичности затрудняет правильную диагностику методами визуализации.
Таким образом, отсутствие специфичности затрудняет правильную диагностику методами визуализации.
5.2.2. Микроскопические методы
Световая микроскопия может использоваться для обнаружения присутствия B. mandrillaris в ткани хозяина. Balamuthia часто идентифицируют в тканях как Acanthamoeba , поскольку обе амебы вызывают гранулематозный амебный энцефалит. Амебы можно наблюдать при биопсии и вскрытии головного мозга и кожи [174–176].В отличие от инфекции N. fowleri , B. mandrillaris обычно не обнаруживают в препаратах ЦСЖ, хотя он был выделен из ЦСЖ 33-летнего пациента, умершего от БАЭ [7, 177]. В большинстве описанных случаев B. mandrillaris наблюдали в образцах биопсии головного мозга, залитых в парафин и обработанных для H&E [158, 170, 175, 178]. Наблюдаются участки воспаления, гранулемы, наличие трофозоитов и кист B. mandrillaris , особенно вокруг кровеносных сосудов [165, 169, 179].Трофозоиты имеют форму от овальной до круглой, с одним ядром и большим ядрышком, а цисты имеют округлую форму с толстой стенкой.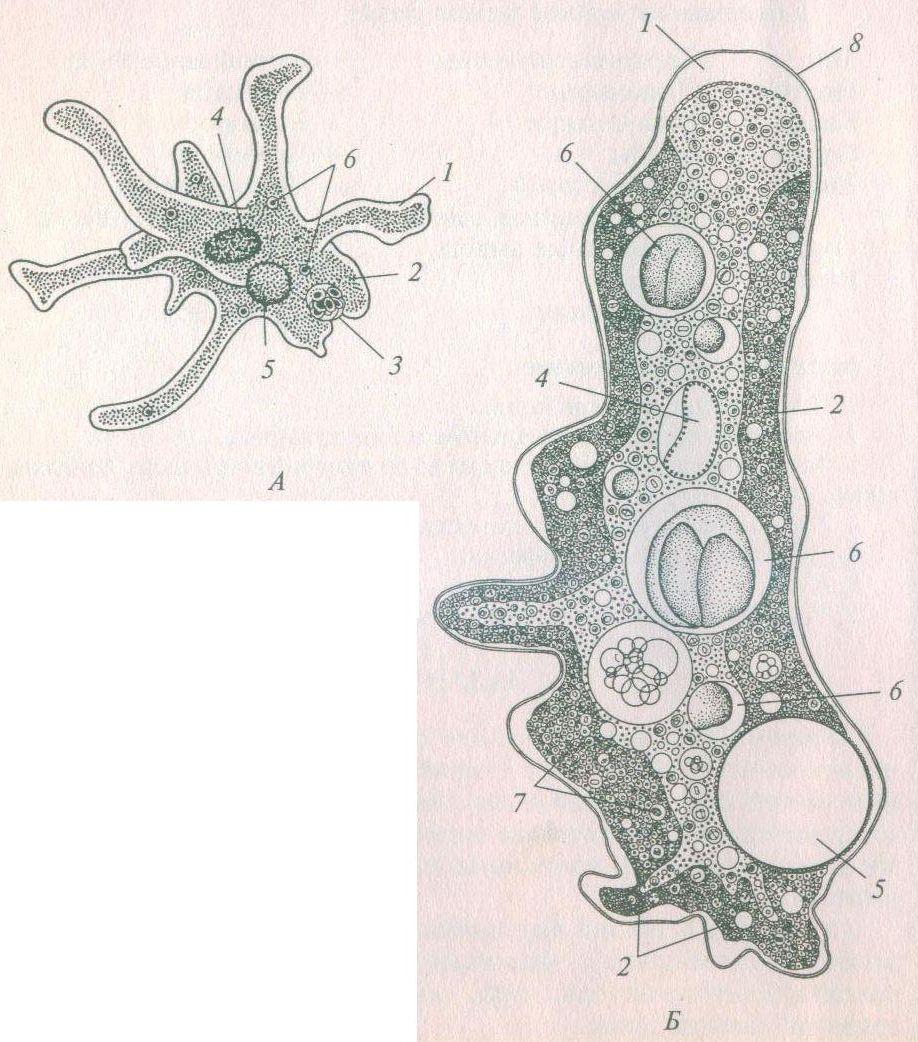 Однако этих морфологических характеристик недостаточно, чтобы отличить B. mandrillaris от Acanthamoeba spp. , а также способность дифференцировать этих амеб от макрофагов хозяина требует специальных знаний [7, 84].
Однако этих морфологических характеристик недостаточно, чтобы отличить B. mandrillaris от Acanthamoeba spp. , а также способность дифференцировать этих амеб от макрофагов хозяина требует специальных знаний [7, 84].
Антитела имеют решающее значение для специфического обнаружения B. mandrillaris в тканях. Исследования подтвердили, как в биоптатах, так и в аутопсиях, присутствие B.mandrillaris в тканях мозга и кожи [84, 157, 170, 174, 178]. Обычно образцы, залитые парафином, разрезают и инкубируют с кроличьей сывороткой против Balamuthia , а FITC-конъюгированные вторичные антитела против кроличьего IgG используют для обнаружения амеб с высокой степенью специфичности [159].
Можно идентифицировать B. mandrillaris в биоптатах с помощью просвечивающей электронной микроскопии (ПЭМ). В отличие от клеток-хозяев трофозоиты содержат характерное плотное ядрышко, а в цитоплазме много пузырьков.Кроме того, с помощью ПЭМ можно отличить B.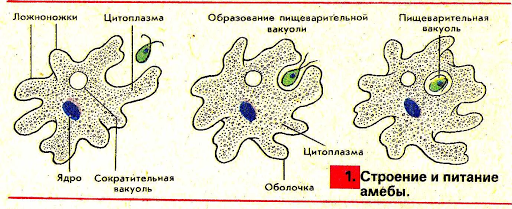 mandrillaris от Acanthamoeba , поскольку B. mandrillaris обладает кистой с тройными стенками, что является отличительной чертой по сравнению с кистой с двойными стенками Acanthamoeba [158, 159].
mandrillaris от Acanthamoeba , поскольку B. mandrillaris обладает кистой с тройными стенками, что является отличительной чертой по сравнению с кистой с двойными стенками Acanthamoeba [158, 159].
5.2.3. Культуральные методы
Возможно выделение B. mandrillaris из образцов биопсии; однако амебный рост медленный и также требует использования клеток тканевой культуры в качестве источника пищи, поскольку B.mandrillaris не питается бактериями [48, 157–159]. Это не рекомендуемый метод.
5.2.4. Серология
Одной из характеристик инфекции B. mandrillaris является высокая концентрация антител к амебам в сыворотке хозяина [180–182]. Инфекцию у людей с подозрением на БАЭ можно определить по наличию антител против B. mandrillaris в их сыворотке с помощью иммуноферментного анализа (ИФА). Эта методология была использована для скрининга группы пациентов с энцефалитом в Калифорнии, и было показано, что 7 отдельных образцов из 290 были положительными на B. mandrillaris [181]. Антитела к B. mandrillaris не реагируют перекрестно с другими амебами [180]. Таким образом, метод ELISA может быть полезен для скрининга образцов, содержащих большое количество людей. Совсем недавно проточная цитометрия была успешно использована для обнаружения и количественного определения антител против B. mandrillaris как у здоровых, так и у больных людей [182].
mandrillaris [181]. Антитела к B. mandrillaris не реагируют перекрестно с другими амебами [180]. Таким образом, метод ELISA может быть полезен для скрининга образцов, содержащих большое количество людей. Совсем недавно проточная цитометрия была успешно использована для обнаружения и количественного определения антител против B. mandrillaris как у здоровых, так и у больных людей [182].
5.2.5. ПЦР
Быстрое и высокоспецифичное обнаружение B. mandrillaris может быть достигнуто с помощью методологии полимеразной цепной реакции. B. mandrillaris является четко определенным филогенетическим видом с отсутствием вариаций последовательности SSU рДНК между изолятами и низким уровнем вариабельности митохондриальной ДНК. Booton и коллеги [183, 184] разработали специфические праймеры из части митохондриального гена рРНК (rns). ПЦР-реакция этих праймеров привела к продукту размером 1075 п.н., где продукт специфичен для B. mandrillaris , а не для Acanthamoeba . Обнаружение B. mandrillaris было успешно выполнено на клинических образцах с использованием этих праймеров, как в ткани головного мозга, так и в спинномозговой жидкости [177, 185, 186].Была проверена чувствительность обнаружения ПЦР, и было обнаружено, что для амплификации достаточно 0,2 амебы, поскольку использовалась митохондриальная ДНК [186], что указывает на то, что использование ПЦР является эффективным диагностическим инструментом. Эффективность метода ПЦР сравнивали с IIF в качестве диагностического инструмента на архивной ткани головного мозга, и сообщалось о высокой степени согласия [109]. С использованием различных праймеров была разработана мультиплексная ПЦР в реальном времени для обнаружения B. mandrillaris с быстрым временем завершения теста и высокой чувствительностью, обнаруживая одну амебу в образце [57].Этот мультиплексный анализ был утвержден и рекомендован для обнаружения FLA в клинических образцах [111]. Совсем недавно для B. mandrillaris был разработан анализ ПЦР в реальном времени, нацеленный на ген РНКазы P [187].
Обнаружение B. mandrillaris было успешно выполнено на клинических образцах с использованием этих праймеров, как в ткани головного мозга, так и в спинномозговой жидкости [177, 185, 186].Была проверена чувствительность обнаружения ПЦР, и было обнаружено, что для амплификации достаточно 0,2 амебы, поскольку использовалась митохондриальная ДНК [186], что указывает на то, что использование ПЦР является эффективным диагностическим инструментом. Эффективность метода ПЦР сравнивали с IIF в качестве диагностического инструмента на архивной ткани головного мозга, и сообщалось о высокой степени согласия [109]. С использованием различных праймеров была разработана мультиплексная ПЦР в реальном времени для обнаружения B. mandrillaris с быстрым временем завершения теста и высокой чувствительностью, обнаруживая одну амебу в образце [57].Этот мультиплексный анализ был утвержден и рекомендован для обнаружения FLA в клинических образцах [111]. Совсем недавно для B. mandrillaris был разработан анализ ПЦР в реальном времени, нацеленный на ген РНКазы P [187]. Наблюдались чувствительность и специфичность, при этом зонды могли обнаруживать небольшие количества амебной ДНК.
Наблюдались чувствительность и специфичность, при этом зонды могли обнаруживать небольшие количества амебной ДНК.
6.
Sappinia pedata и Sappinia Амебный энцефалит (SAE) Свободноживущая амеба , Sappinia , недавно обнаруженный человеческий патоген центральной нервной системы (ЦНС), может вызывать амебный энцефалит у людей [ 9, 10].Существует два вида Sappinia , S. pedata и S. diploidea . Sappinia имеет всемирное распространение и была выделена из фекалий лосей и буйволов, почвы, загрязненной фекалиями крупного рогатого скота, разлагающейся наземной растительной подстилки и пресной воды [188, 189]. Жизненный цикл Sappinia состоит из двух стадий: трофозоита и цисты [190]. Первый и единственный случай амебного энцефалита, вызванного Sappinia sp. произошел у ранее здорового иммунокомпетентного взрослого мужчины, пережившего инфекцию.Инкубационный период и путь заражения неизвестны, но считается, что это происходит при вдыхании через носоглотку или гематогенном распространении в головной мозг [9]. Амеба, вызвавшая этот энцефалит, первоначально была идентифицирована как S. diploidea , но теперь идентифицирована как S. pedata с использованием молекулярных методов для идентификации амебы (191, 192). Неизвестно, могут ли другие виды ( S. diploidea ) вызывать инфекции у людей или животных-хозяев.
Амеба, вызвавшая этот энцефалит, первоначально была идентифицирована как S. diploidea , но теперь идентифицирована как S. pedata с использованием молекулярных методов для идентификации амебы (191, 192). Неизвестно, могут ли другие виды ( S. diploidea ) вызывать инфекции у людей или животных-хозяев.
6.1. Симптомы SAE
В одном зарегистрированном случае Sappinia амебного энцефалита перед появлением симптомов произошла инфекция носовых пазух. У человека развилась тошнота, рвота, бифронтальная головная боль, светобоязнь и нечеткость зрения. На короткий период наступала потеря сознания [7, 9, 10, 191]. Сообщалось об успешном исходе у этого пациента после хирургического удаления опухолевидного образования в головном мозге и лечения с использованием азитромицина, пентамидина внутривенно, итраконазола и флуцитозина [10].
6.2. Диагноз
Солидное опухолевидное образование без стенки абсцесса в головном мозге может наблюдаться на МРТ при Sappinia энцефалите.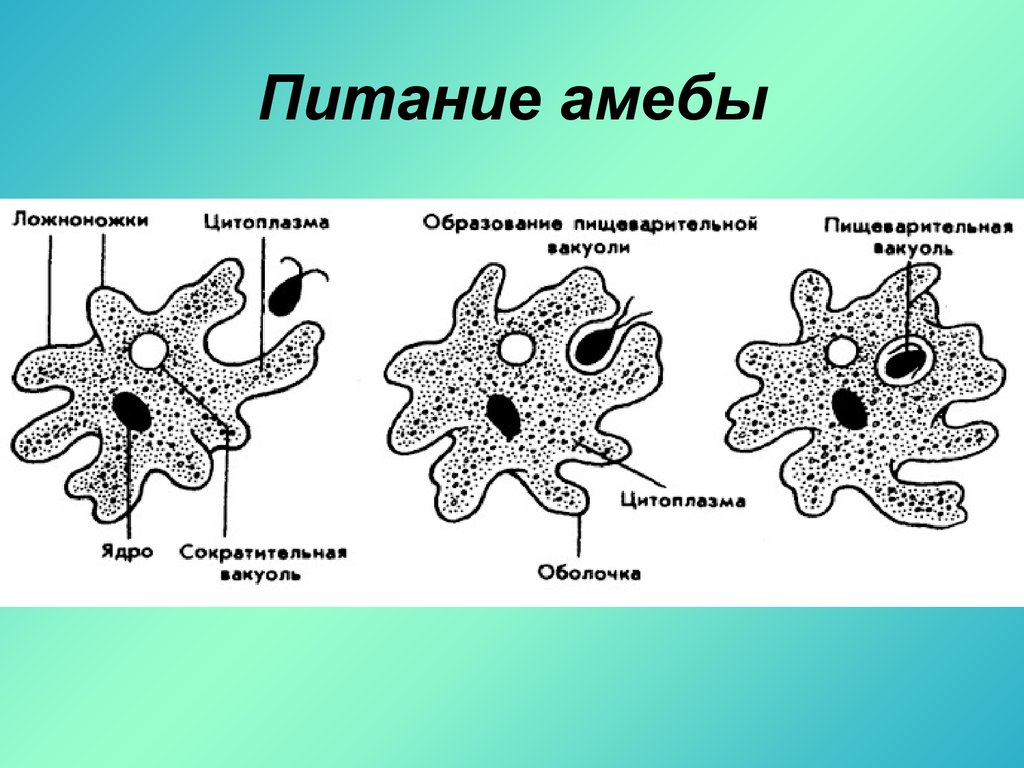 Иссеченную массу или ткань биопсии можно зафиксировать и залить парафином. Срезы головного мозга, окрашенные H&E, могут демонстрировать некротизирующее геморрагическое воспаление, содержащее амебы. Эозинофилы и гранулемы отсутствуют. Амебы Sappinia можно отличить от других FLA по наличию характерного двойного ядра, в котором 2 ядра близко прилегают друг к другу с уплощением в центре [188, 189].В двойном ядре обнаруживаются два ядрышка. Эти структуры можно наблюдать на парафиновых срезах, окрашенных H&E, Гимза или периодической кислотой Шиффа. Амебы легко обнаруживаются на криостатных срезах, окрашенных гематоксилин-эозином [7, 10, 191]. Ткань мозга можно зафиксировать в глутаровом альдегиде и подготовить к просвечивающей электронной микроскопии для визуализации амеб в ткани.
Иссеченную массу или ткань биопсии можно зафиксировать и залить парафином. Срезы головного мозга, окрашенные H&E, могут демонстрировать некротизирующее геморрагическое воспаление, содержащее амебы. Эозинофилы и гранулемы отсутствуют. Амебы Sappinia можно отличить от других FLA по наличию характерного двойного ядра, в котором 2 ядра близко прилегают друг к другу с уплощением в центре [188, 189].В двойном ядре обнаруживаются два ядрышка. Эти структуры можно наблюдать на парафиновых срезах, окрашенных H&E, Гимза или периодической кислотой Шиффа. Амебы легко обнаруживаются на криостатных срезах, окрашенных гематоксилин-эозином [7, 10, 191]. Ткань мозга можно зафиксировать в глутаровом альдегиде и подготовить к просвечивающей электронной микроскопии для визуализации амеб в ткани.
6.3. ПЦР
Sappinia pedata и S. diploidea можно идентифицировать и отличить путем амплификации рДНК SSU с использованием универсальных эукариотических праймеров SSU с последующим ПЦР-тестом с внутренним транскрибируемым спейсером. Праймерами, используемыми в ITS-ПЦР, являются ITS1-P1F-GTA ACA AGG TATCCG TAG GTG AAC C- и ITS2-P4R: TCC TCC GCT TAT TGA TAT GC— [190]. Амебы, первоначально идентифицированные как S. diploidea в единственном зарегистрированном случае амебного энцефалита Sappinia , позже были идентифицированы как S. pedata с использованием недавно разработанных анализов ПЦР в реальном времени, основанных на последовательностях гена 18S рРНК [191]. Анализ, специфичный для Sappinia , может быть включен в анализ мультиплексной ПЦР, описанный Qvarnstrom et al., [57], которые различают Acanthamoeba, B. mandrillaris и N. fowleri для одновременного обнаружения четырех родов FLA, вызывающих инфекции у людей [191].
Праймерами, используемыми в ITS-ПЦР, являются ITS1-P1F-GTA ACA AGG TATCCG TAG GTG AAC C- и ITS2-P4R: TCC TCC GCT TAT TGA TAT GC— [190]. Амебы, первоначально идентифицированные как S. diploidea в единственном зарегистрированном случае амебного энцефалита Sappinia , позже были идентифицированы как S. pedata с использованием недавно разработанных анализов ПЦР в реальном времени, основанных на последовательностях гена 18S рРНК [191]. Анализ, специфичный для Sappinia , может быть включен в анализ мультиплексной ПЦР, описанный Qvarnstrom et al., [57], которые различают Acanthamoeba, B. mandrillaris и N. fowleri для одновременного обнаружения четырех родов FLA, вызывающих инфекции у людей [191].
6.4. Культура
Sappinia может культивироваться на непитательном агаре, покрытом бактериями, и на клетках тканевой культуры [7].
7. Заключение: важность ранней диагностики
Хотя инфекции FLA считаются редкими, в последние годы отмечается рост числа зарегистрированных случаев.
Число пользователей контактными линзами во всем мире увеличилось, и, следовательно, вспышки АК стали более частыми, что требует более быстрых ответных мер общественного здравоохранения.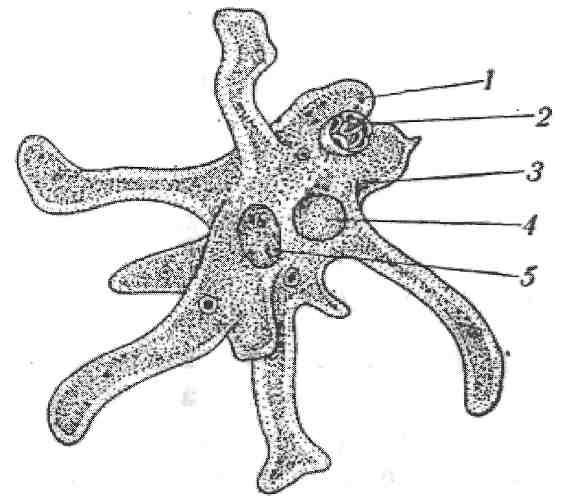 ГЭА и кожный акантамебиаз могут быть одними из наиболее частых вторичных заболеваний у больных СПИД-ВИЧ и у других лиц с иммуносупрессией, поскольку Acanthamoeba встречается во всем мире. Резкий рост BAE может отражать вероятность того, что эта инфекция станет распространенным заболеванием во всем мире. Увеличение заболеваемости PAM, вызванное N. fowleri в последние годы, может быть связано с большей рекреационной активностью в озерах и парках с теплой водой. ПАМ можно было бы избежать путем большей осведомленности о заболевании, использования программ общественного просвещения, закрытия бассейнов с ненадлежащим хлорированием и ношения зажимов для носа во время ныряния и занятий водными видами спорта, когда нос погружен в воду.Недавнее обнаружение S. pedata в качестве возбудителя ЦНС у людей позволяет предположить, что другие свободноживущие амебы, еще не идентифицированные, могут быть причиной амебного или амебного энцефалита.
ГЭА и кожный акантамебиаз могут быть одними из наиболее частых вторичных заболеваний у больных СПИД-ВИЧ и у других лиц с иммуносупрессией, поскольку Acanthamoeba встречается во всем мире. Резкий рост BAE может отражать вероятность того, что эта инфекция станет распространенным заболеванием во всем мире. Увеличение заболеваемости PAM, вызванное N. fowleri в последние годы, может быть связано с большей рекреационной активностью в озерах и парках с теплой водой. ПАМ можно было бы избежать путем большей осведомленности о заболевании, использования программ общественного просвещения, закрытия бассейнов с ненадлежащим хлорированием и ношения зажимов для носа во время ныряния и занятий водными видами спорта, когда нос погружен в воду.Недавнее обнаружение S. pedata в качестве возбудителя ЦНС у людей позволяет предположить, что другие свободноживущие амебы, еще не идентифицированные, могут быть причиной амебного или амебного энцефалита.
Более быстрый клинический ответ может обеспечить более высокие показатели выживаемости, поскольку лечение доступно. Таким образом, быстрые и эффективные диагностические тесты имеют решающее значение для успеха лечения. Наличие ПЦР-зондов является многообещающей процедурой для получения быстрой и специфической, подтверждающей диагностики PAM, GAE, AK, BAE и SAE своевременно для эффективного лечения.С этой целью доступен анализ мультиплексной ПЦР, который был утвержден как важный и специфический инструмент для идентификации N. fowleri, Acanthamoeba spp., и B. mandrillaris в клинических образцах [57, 111], но только лишь немногие референс-лаборатории способны проводить диагностическое выявление FLA [191]. Кроме того, был разработан анализ ПЦР для обнаружения Sappinia в клинических образцах [191]. В заключение, распознавание этих заболеваний и специальные диагностические тесты могут привести к более раннему лечению и снижению тяжести и летальности этих инфекций у человека-хозяина.
Таким образом, быстрые и эффективные диагностические тесты имеют решающее значение для успеха лечения. Наличие ПЦР-зондов является многообещающей процедурой для получения быстрой и специфической, подтверждающей диагностики PAM, GAE, AK, BAE и SAE своевременно для эффективного лечения.С этой целью доступен анализ мультиплексной ПЦР, который был утвержден как важный и специфический инструмент для идентификации N. fowleri, Acanthamoeba spp., и B. mandrillaris в клинических образцах [57, 111], но только лишь немногие референс-лаборатории способны проводить диагностическое выявление FLA [191]. Кроме того, был разработан анализ ПЦР для обнаружения Sappinia в клинических образцах [191]. В заключение, распознавание этих заболеваний и специальные диагностические тесты могут привести к более раннему лечению и снижению тяжести и летальности этих инфекций у человека-хозяина.
Благодарность
Этот обзор посвящен памяти доктора Фредерика Л. Шустера, пионера изучения свободноживущих амеб, опубликовавшего значительный вклад в эту область за последние 35 лет.
Акантамёбный кератит: симптомы, лечение и профилактика
Обзор
Что такое акантамёбный кератит?
Акантамёбный кератит — это серьезная инфекция, поражающая роговицу (прозрачную наружную оболочку глаза). Его вызывает амеба — крошечный одноклеточный организм.Амеба проникает в ваш глаз и проедает слои вашей роговицы. Он разрушает ткани, когда проходит через ваш глаз. Акантамеба распространена в воздухе, почве, озерах и океанах. Большинство инфекций происходит из пресной воды, такой как водопроводная вода, плавательные бассейны, джакузи, душевые и канализационные системы.
Акантамёбный кератит встречается редко, но может привести к боли в глазах, необратимой потере зрения или даже полной слепоте. Инфекция может поразить один или оба глаза. Это не заразно.
Кто подвержен риску заболеть акантамёбным кератитом?
Акантамёбный кератит может возникнуть у любого человека, но чаще всего он встречается у людей, которые носят мягкие контактные линзы. У вас есть больший риск этого состояния, если вы:
У вас есть больший риск этого состояния, если вы:
- Чистые ваши контактные линзы с водопроводной водой или физиологическим раствором вместо дезинфицирующего раствора.
- Зайди в контакт с водой, содержащей амебы.
- Не хранить и обрабатывать ваши контактные линзы правильно.
- имели в прошлом повреждения вашей роговицы.
- плавать, душ или использовать гидромассажную ванну при ношении контактных линз.
Как часто Acanthamoeba кератит?
Acanthamoeba кератит встречается редко.В США только один-два человека на миллион носителей контактных линз ежегодно заболевают этим заболеванием. Это еще реже встречается у людей, которые не носят контактные линзы.
Симптомы и причины
Что вызывает акантамёбный кератит?
Акантамеба вызывает эту глазную инфекцию. Амеба прикрепляется к клеткам на внешней поверхности роговицы. Он также может поражать глаз, проникая через небольшие ссадины роговицы (царапины).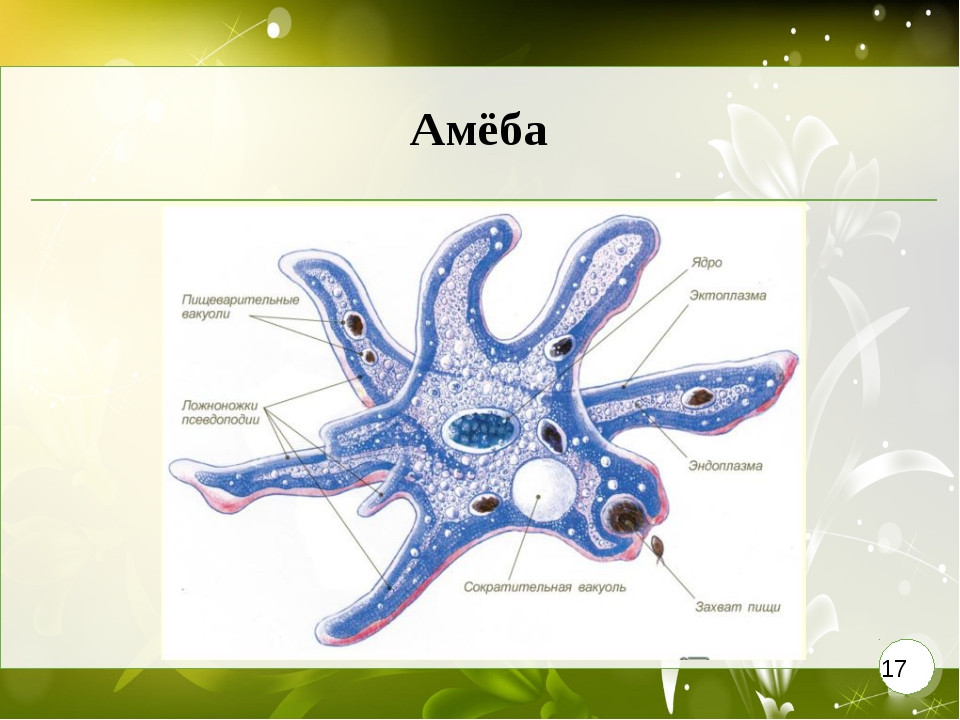 Инфекция разрушает клетки и продвигается дальше в роговицу.
Инфекция разрушает клетки и продвигается дальше в роговицу.
Каковы симптомы акантамёбного кератита?
Симптомы акантамёбного кератита на ранней стадии могут включать:
- Затуманенное зрение или потеря зрения.
- Мутная или грязная роговица.
- Боль в глазах.
- Покраснение глаз.
- Ощущение чего-то в глазу.
- Чувствительность к свету.
- Слезятся глаза.
- Белесые кольца на поверхности глаза.
Проявление симптомов после попадания акантамебы в глаз может занять от нескольких дней до нескольких недель.
Каковы осложнения акантамёбного кератита?
При отсутствии лечения инфекция может привести к необратимой потере зрения или полной слепоте. Другие осложнения акантамёбного кератита на поздних стадиях могут включать:
- Передний увеит (воспаление сосудистой оболочки или среднего слоя глаза).
- Атрофия радужки (смещенный зрачок).
- Зрелая катаракта (прозрачный хрусталик глаза полностью затуманен).

- Склерит (воспаление склеры или белой части глаза).
- Вторичная глаукома (поражение зрительного нерва).
Диагностика и тесты
Как диагностируется акантамёбный кератит?
Медицинский работник, специализирующийся на лечении глаз, например, окулист или офтальмолог, может диагностировать акантамёбный кератит. Они проведут осмотр глаз, изучат ваши симптомы и оценят гигиену ваших контактных линз (если вы носите контактные линзы).
Эту инфекцию легко поставить неверный диагноз, потому что ее симптомы схожи с симптомами других заболеваний роговицы.Поэтому очень важно сообщить своему лечащему врачу, если вы были в контакте с какой-либо потенциально загрязненной водой или другими веществами.
Анализы на акантамёбный кератит могут включать:
- Соскоб с роговицы: Соскоб с роговицы, также называемый поверхностной кератэктомией, представляет собой процедуру удаления клеток с поверхности роговицы.
 Ваш лечащий врач обезболивает поверхность вашего глаза и использует небольшое лезвие или щетку для удаления клеток. Они изучают клетки под микроскопом или проводят лабораторные анализы клеток, чтобы проверить наличие инфекции.
Ваш лечащий врач обезболивает поверхность вашего глаза и использует небольшое лезвие или щетку для удаления клеток. Они изучают клетки под микроскопом или проводят лабораторные анализы клеток, чтобы проверить наличие инфекции. - Конфокальная микроскопия: В этом исследовании используется лазерный свет для создания высокодетализированных изображений каждого слоя роговицы. Ваш лечащий врач использует специальный микроскоп, чтобы увидеть клетки роговицы, нервы и любых паразитов, таких как акантамёба.
Управление и лечение
Как лечится акантамёбный кератит?
Местный антисептик является наиболее распространенным методом лечения акантамёбы. Антисептик борется с микроорганизмами. Он наносится непосредственно на поверхность вашего глаза.Вам могут понадобиться эти процедуры от шести месяцев до года. Иногда ваш лечащий врач соскребает часть вашей роговицы перед нанесением лекарства. Это помогает лекарству проникнуть глубже в глаз.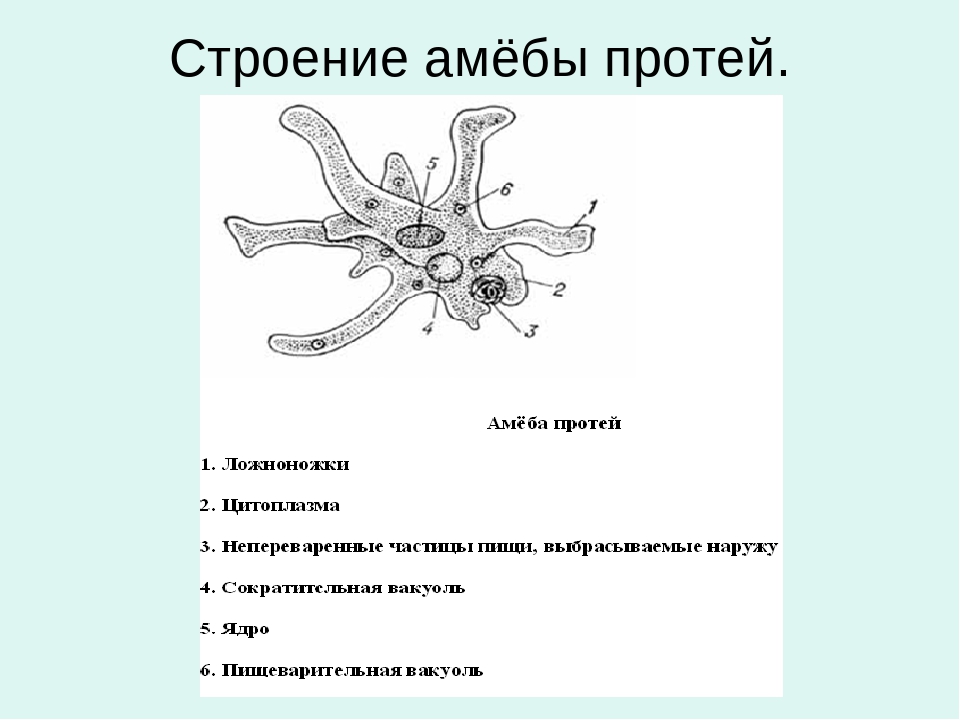
Ваш лечащий врач может также порекомендовать антибиотики или противогрибковые препараты. Стероиды или обезболивающие препараты могут помочь уменьшить боль и воспаление.
Вам может потребоваться операция по поводу прогрессирующего акантамебного кератита, который не улучшается при местной терапии. Это может включать сквозную кератопластику (ПК).ПК – пересадка роговицы на всю толщину. Роговица от донора заменяет вашу поврежденную роговицу.
Профилактика
Как предотвратить акантамёбный кератит?
Вы можете снизить риск акантамёбного кератита с помощью:
- Избегайте контакта с загрязненной водой, которая может включать пресноводные озера, реки и горячие источники.
- Наполнение футляра для хранения контактных линз свежим раствором каждый раз, когда вы его открываете.
- Никогда не спите в контактных линзах.
- Не принимать душ, плавать или пользоваться джакузи при ношении контактных линз.
- Не пользоваться чужими контактными линзами.

- Регулярная замена контактных линз в соответствии с рекомендациями вашего лечащего врача.
- Использование только дезинфицирующего раствора (не солевого раствора) для ополаскивания и хранения контактных линз — никогда не используйте водопроводную воду.
- Посещение окулиста или офтальмолога для регулярных осмотров.
- Мойте руки перед тем, как прикоснуться к глазам или контактным линзам.
Перспективы/прогноз
Каковы перспективы для людей с акантамёбным кератитом?
Чем раньше вы получите диагноз и лечение акантамебы, тем лучше будет ваш прогноз. Исследования показывают, что люди, которые обращаются за лечением при первых признаках инфекции, в 10 раз чаще выздоравливают со зрением 20/25 (легкая потеря зрения) или выше. Перспектива плохая, если инфекция уже находится в глубоких отделах глаза.
Если вы действительно потеряли зрение из-за этой инфекции, вы можете найти поддержку различными способами.Возможно, вы сможете лучше справиться с потерей зрения с помощью:
- Узнайте у своего поставщика медицинских услуг о вспомогательных средствах для слабовидящих, таких как телескопические очки или ручные лупы.

- Обратитесь за консультацией, если вы чувствуете себя подавленным или подавленным.
- Разговор с близкими и опекунами о вашем состоянии.
Жить с
Когда следует вызвать врача?
Обратитесь к врачу при первых признаках боли, покраснения или повышенной чувствительности глаз.Не ждите, пока вы почувствуете потерю зрения.
Какие вопросы я должен задать своему врачу?
Вы можете спросить своего поставщика медицинских услуг:
- Как справиться с симптомами во время лечения?
- Как далеко зашла инфекция?
- Стоит ли ожидать осложнений?
- Должен ли я следить за признаками осложнений?
- Каковы шансы, что я потеряю зрение?
Записка из клиники Кливленда
Акантамёбный кератит — это редкая серьезная глазная инфекция, вызываемая крошечным микроорганизмом, обитающим в пресной воде.Заразиться может любой, но чаще всего это происходит у людей, которые носят контактные линзы. Симптомы могут варьироваться от боли в глазах и покраснения до слепоты. Раннее выявление и лечение значительно улучшают прогноз для людей с этой инфекцией.
Симптомы могут варьироваться от боли в глазах и покраснения до слепоты. Раннее выявление и лечение значительно улучшают прогноз для людей с этой инфекцией.
Кровь: руководство по гистологии
Лейкоциты
Лейкоциты встречаются гораздо реже, чем эритроциты. Существует пять типов лейкоцитов (лейкоцитов). Они делятся на два основных класса
- Гранулоциты (включая нейтрофилы, эозинофилы и базофилы)
- Агранулоциты (включая лимфоциты и моноциты).
Эта классификация зависит от того, можно ли различить гранулы в их цитоплазме с помощью светового микроскопа и обычных методов окрашивания).
Все лейкоциты способны двигаться, как амебы, и могут мигрировать из кровеносных сосудов в окружающие ткани.
Обратите внимание — разные лейкоциты в мазках крови легко спутать. Чтобы идентифицировать их, вам нужно посмотреть на форму ядра и сравнить их размер с размером эритроцита. После того, как вы просмотрели приведенные ниже примеры — попробуйте сами — нажмите здесь и посмотрите, сможете ли вы идентифицировать «загадочные» ячейки.
После того, как вы просмотрели приведенные ниже примеры — попробуйте сами — нажмите здесь и посмотрите, сможете ли вы идентифицировать «загадочные» ячейки.
Гранулоциты:
Показывает нейтрофилы в мазке крови. Нейтрофилы имеют диаметр 12-14 мкм и поэтому выглядят больше, чем окружающие эритроциты. Имеется одно ядро, которое является многодольчатым и может иметь от 2 до 5 долей.
Хроматин в ядре конденсирован.Это означает, что нет синтеза белка. В цитоплазме мало органоидов.
Нейтрофилы
Нейтрофилы являются наиболее распространенным типом лейкоцитов, обнаруживаемым в мазке крови. Они составляют 60-70% от общего количества лейкоцитов.
Нейтрофилы имеют 3 типа гранул:
- лазурные гранулы (лизосомы),
- секреторных гранул в цитоплазме лососево-розового цвета, антимикробные ферменты.
- имеют гликопротеины и желатиназу.

Функция:
Нейтрофилы рождаются в костном мозге. Они циркулируют в крови в течение 6-10 часов, а затем попадают в ткани. Они подвижны и фагоцитируют и разрушают поврежденные ткани и бактерии. Они самоуничтожаются после одного всплеска активности.
Они важны при воспалительных реакциях.
На этом снимке показан эозинофил в мазке крови.Эти клетки имеют диаметр 12–17 мкм — больше, чем нейтрофилы, и примерно в 3 раза больше размера эритроцита. Вы можете видеть, что эозинофилы имеют только две доли ядра.
Эти клетки имеют крупные специфические ацидофильные гранулы, которые окрашиваются в ярко-красный или красновато-фиолетовый цвет. Эти гранулы содержат белки, которые являются «разрушительными» и токсичными.
Эозинофилы
Эозинофилы в мазках крови обнаруживаются довольно редко — составляют 1-6% от общего числа лейкоцитов.
Функция :
Эти клетки рождаются в костном мозге и через несколько часов мигрируют из системы периферической крови в рыхлую соединительную ткань дыхательных и желудочно-кишечных путей. Они фагоцитируют комплексы антиген-антитело. Они также продуцируют гистаминазу и арилсульфатазу В, два фермента, которые инактивируют два воспалительных агента, выделяемых тучными клетками. Высокий уровень эозинофилов в крови может свидетельствовать об аллергической реакции.
Эозинофилы также играют важную роль в уничтожении паразитических червей.
На этой фотографии изображен базофил. Они имеют диаметр 14-16 мкм, содержат большое количество темно-синих окрашенных гранул (основные) и двудольное ядро. Гранулы содержат гепарин, гистамин и серотонин. простагландины и лейкотриены.
Базофилы
Базофилы — самый редкий тип лейкоцитов, составляющий всего 1% лейкоцитов, обнаруживаемых в мазке крови.
Функция :
Эти клетки участвуют в иммунных реакциях на паразитов. У них есть рецепторы IgE, и гранулы высвобождаются, когда клетки связывают IgE. Эти клетки также накапливаются в местах инфекции, а высвобождение простагландинов, серотонина и гистамина способствует увеличению притока крови к области повреждения в рамках воспалительной реакции. Дегрануляция — высвобождение гистамина также играет роль в аллергических реакциях, таких как сенная лихорадка.
Агранулоциты:
Это фото лимфоцита в мазке крови.Большинство лимфоцитов мелкие; немного больше, чем эритроциты, около 6-9 мкм в диаметре,
Остальные (около 10%) крупнее, около 10-14 мкм в диаметре. Эти более крупные клетки имеют больше цитоплазмы, больше свободных рибосом и митохондрий. Лимфоциты могут выглядеть как моноциты, за исключением того, что лимфоциты не имеют ядра в форме фасоли, а лимфоциты обычно меньше. Большие лимфоциты обычно являются активированными лимфоцитами.
Большие лимфоциты обычно являются активированными лимфоцитами.
Они имеют маленькое сферическое ядро и обильное темное окрашивание конденсированного хроматина.Цитоплазмы не так много, и она базофильная (бледно-голубое/фиолетовое окрашивание).
Лимфоцит
Это вторые по распространенности лейкоциты (20-50%), и их легко обнаружить в мазках крови.
Хотя клетки выглядят одинаково, существует два основных типа: В-клетки и Т-клетки.
В-клетки развиваются в костном мозге. Т-клетки рождаются в костном мозге, но созревают в тимусе. Подробнее об этом будет рассказано в разделе об иммунной системе.
Функция :
В-клетки превращаются в плазматические клетки, которые вырабатывают антитела. Т-клетки атакуют вирусы, раковые клетки и трансплантаты.
Это фотография моноцита в мазке крови.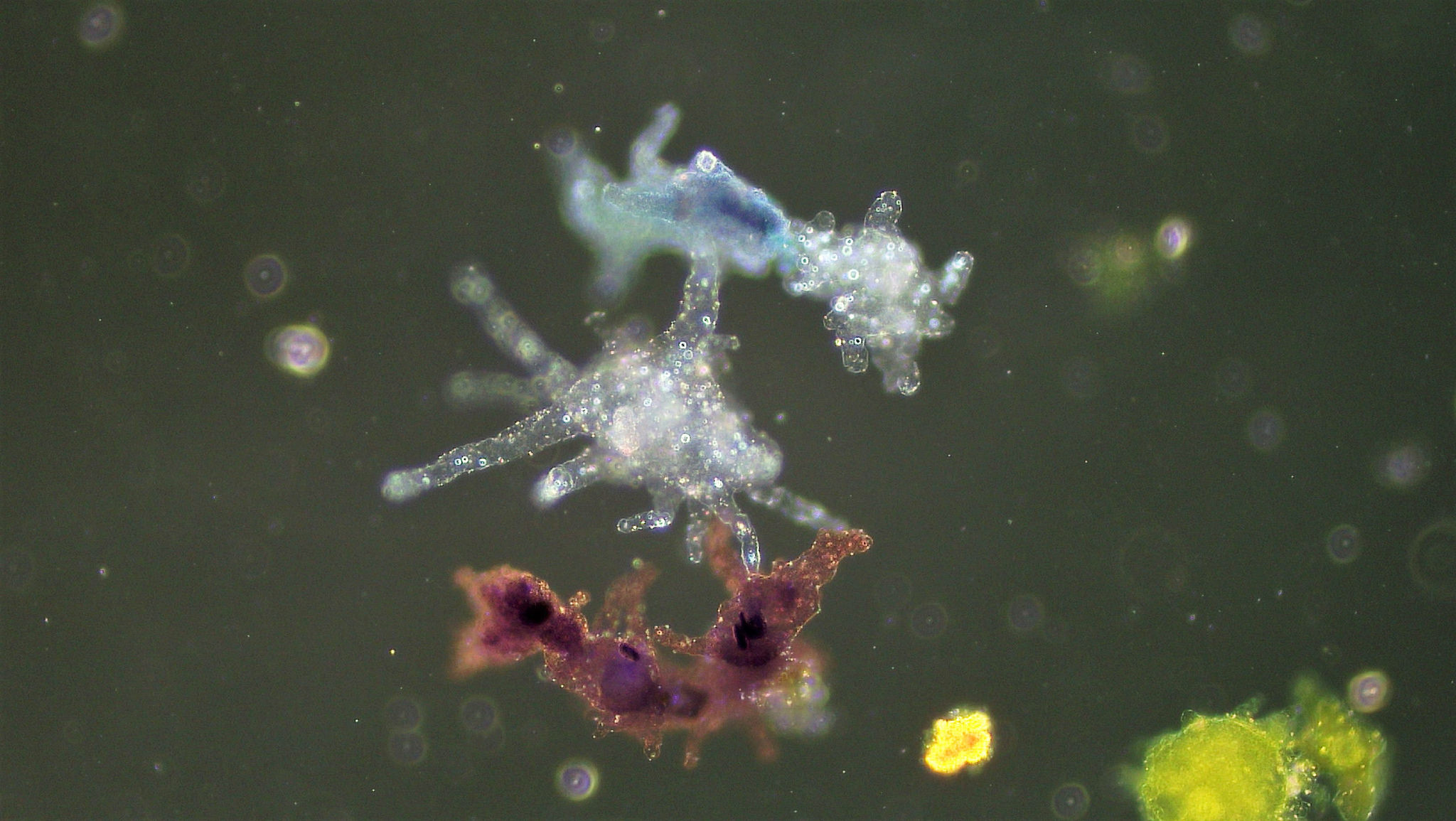
Это самый крупный тип лейкоцитов, их диаметр может достигать 20 мкм.
У них большое эксцентрично расположенное ядро, имеющее форму фасоли.
Они имеют обильную цитоплазму и несколько мелких розово-фиолетовых гранул в цитоплазме.
Моноцит
Моноциты являются третьим наиболее распространенным типом лейкоцитов; около 2-10% лейкоцитов составляют моноциты.
Функция:
Моноциты в кровотоке являются предшественниками тканевых макрофагов, которые активно фагоцитируют. Моноциты циркулируют в крови 1-3 дня, затем мигрируют в ткани организма, где трансформируются в макрофаги.Они будут фагоцитировать мертвые клетки и бактерии. Некоторые моноциты также могут трансформироваться в остеокласты.
Моноциты играют важную роль в воспалительной реакции.
Амеба-инфекция у собак – симптомы, причины, диагностика, лечение, выздоровление, лечение, стоимость
Entamoeba Histolytica
Ветеринар начнет диагностику амебы Entamoeba histolytica с тщательного исследования фекалий.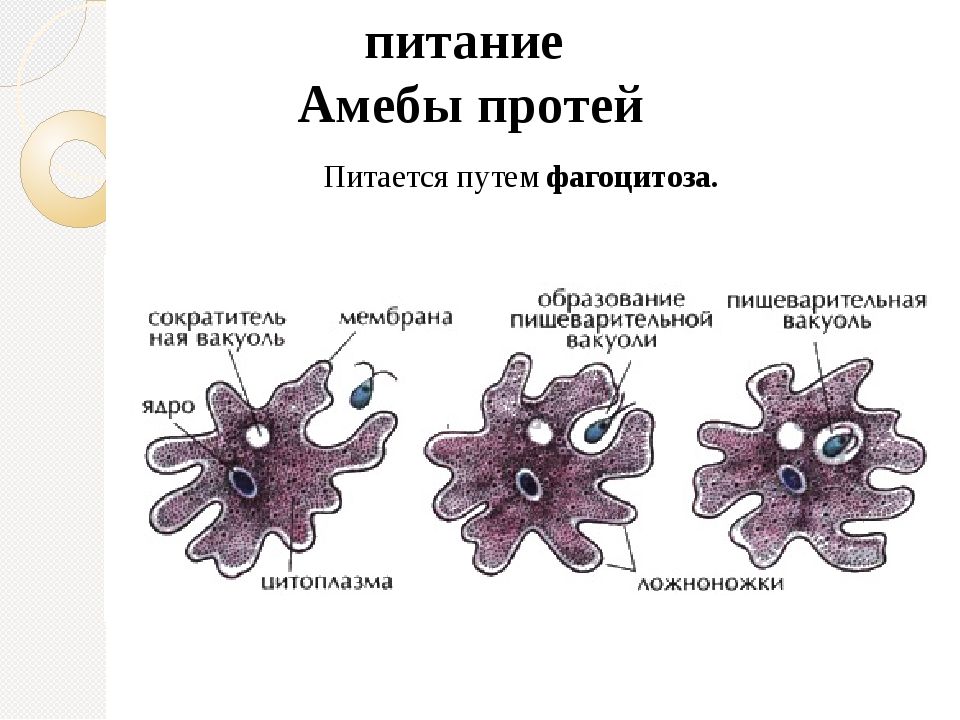 Свежий образец может показать присутствие микроорганизма в течение 30 минут после начала диагностического теста.Мазок кала может показать существующий трофозоит (стадия роста, когда питательные вещества поглощаются хозяином), который может содержать проглоченные эритроциты. Вот почему ветеринары регулярно наблюдают анемию у собак, зараженных этим паразитическим организмом. Часто для окончательного диагноза требуется более одного образца, поскольку амеба может выделяться периодически. Соскоб или биопсия могут показать присутствие амебы в тканевых поражениях. Гистологию можно использовать для поиска амебы в ткани.
Свежий образец может показать присутствие микроорганизма в течение 30 минут после начала диагностического теста.Мазок кала может показать существующий трофозоит (стадия роста, когда питательные вещества поглощаются хозяином), который может содержать проглоченные эритроциты. Вот почему ветеринары регулярно наблюдают анемию у собак, зараженных этим паразитическим организмом. Часто для окончательного диагноза требуется более одного образца, поскольку амеба может выделяться периодически. Соскоб или биопсия могут показать присутствие амебы в тканевых поражениях. Гистологию можно использовать для поиска амебы в ткани.
Акантамеба
К сожалению, у ветеринаров будет возможность диагностировать инвазию акантамебы только на поздних стадиях болезни и чаще всего в качестве патологоанатомического вмешательства. Часто состояние собаки будет прогрессировать без внешних симптомов, а признаки болезни проявляются только на стадии кризиса. Поскольку этот микроорганизм процветает у собак с подавленной иммунной системой, следует учитывать возможность заражения акантамебой у больного животного при наличии подобных симптомов.Поначалу поставить диагноз акантамеба может быть трудно. Однако ветеринар может предложить общий анализ крови, чтобы проверить уровень лейкоцитов в организме. Неврологическое обследование может показать, что у вашей собаки есть признаки нестабильности и инвазии паразитарных организмов. Выполнение центральной пункции спинномозговой жидкости может показать признаки неврологических изменений, в то время как МРТ может подтвердить утолщение головного мозга. Респираторный дистресс может быть обнаружен при биопсии ткани и может указывать на агрессивное воздействие на легочную ткань, что очень типично для этого заболевания.
Часто состояние собаки будет прогрессировать без внешних симптомов, а признаки болезни проявляются только на стадии кризиса. Поскольку этот микроорганизм процветает у собак с подавленной иммунной системой, следует учитывать возможность заражения акантамебой у больного животного при наличии подобных симптомов.Поначалу поставить диагноз акантамеба может быть трудно. Однако ветеринар может предложить общий анализ крови, чтобы проверить уровень лейкоцитов в организме. Неврологическое обследование может показать, что у вашей собаки есть признаки нестабильности и инвазии паразитарных организмов. Выполнение центральной пункции спинномозговой жидкости может показать признаки неврологических изменений, в то время как МРТ может подтвердить утолщение головного мозга. Респираторный дистресс может быть обнаружен при биопсии ткани и может указывать на агрессивное воздействие на легочную ткань, что очень типично для этого заболевания.
Амеба-инфекция у собак | Собачий амебиаз | Причина диареи собак
Амебиаз собак
Амебиаз — паразитарная инфекция, вызываемая одноклеточным организмом, известным как амеба. Амебиаз может поражать людей, а также собак и кошек. Чаще всего он встречается в тропических районах и его можно увидеть в Северной Америке.
Амебиаз может поражать людей, а также собак и кошек. Чаще всего он встречается в тропических районах и его можно увидеть в Северной Америке.
Симптомы и типы
Существует два типа паразитарных амеб, поражающих собак: Entamoeba histolytica и Acanthamoeba.
Entamoeba histolytica:
- Обычно бессимптомное заболевание
- Тяжелые инфекции могут вызывать колит, приводящий к кровавой диарее
- Гематогенное распространение (распространение по организму с током крови) вызывает повреждение и отказ основных систем органов.Симптомы зависят от пораженной системы органов, но обычным исходом является смерть.
Акантамеба:
- Вызывает гранулематозный амебный менингоэнцефалит (воспаление головного мозга), проявляющийся отсутствием аппетита, лихорадкой, вялостью, выделениями из глаз и носа, затрудненным дыханием и неврологическими симптомами (нарушение координации, судороги и др.)
Причины
Entamoeba histolyticus чаще всего распространяется при проглатывании инфицированных человеческих фекалий.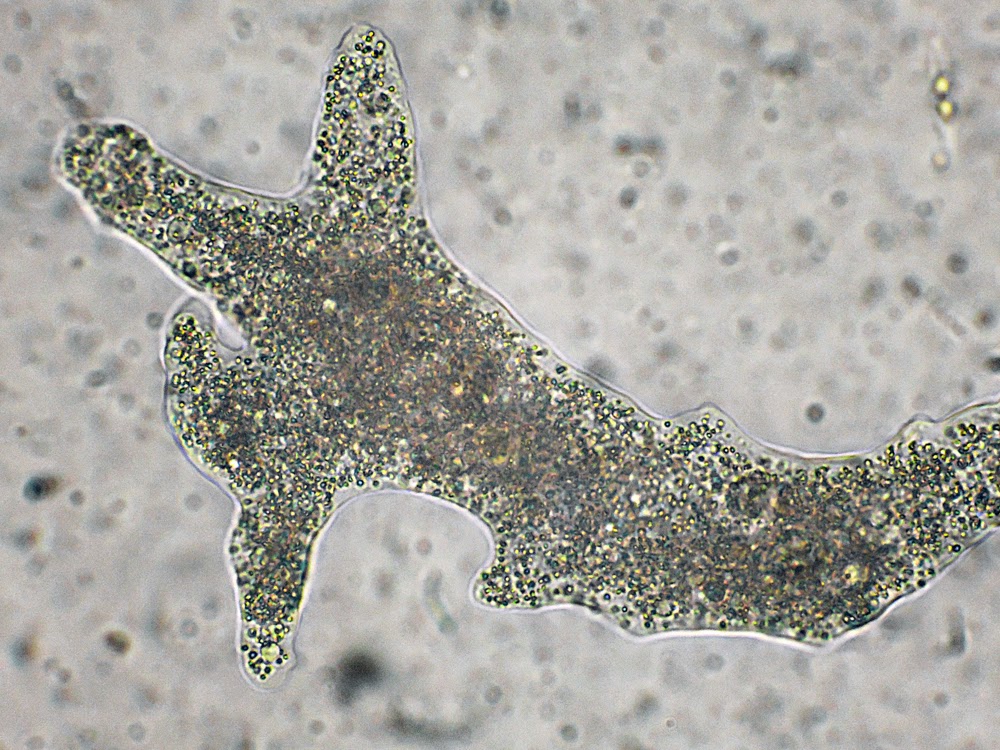 Есть два свободноживущих вида Acanthamoeba: A. castellani и A. culbertsoni. Эти виды можно найти в пресной воде, соленой воде, почве и сточных водах.
Есть два свободноживущих вида Acanthamoeba: A. castellani и A. culbertsoni. Эти виды можно найти в пресной воде, соленой воде, почве и сточных водах.
- Собаки могут заразиться при проглатывании или вдыхании загрязненной воды, почвы или сточных вод.
- Может произойти колонизация кожи собаки Acanthamoeba, которая может стать причиной инфекции.
- Может произойти колонизация роговицы глаза Acanthamoeba, что может стать причиной инфекции.
- Инфекция может передаваться с током крови (гематогенное распространение.)
- Инфекция из носа может распространиться в мозг.
Молодые собаки и собаки с ослабленным иммунитетом чаще всего заболевают.
Диагностика
Анализ крови (общий анализ клеток крови и биохимический профиль крови) и анализ мочи (анализ мочи) обычно выполняются и часто являются нормальными, хотя в этих тестах можно увидеть признаки обезвоживания, если они присутствуют.
Другие лабораторные анализы, которые может порекомендовать ваш ветеринар, включают:
- Биопсия толстой кишки, полученная при колоноскопии (осмотр толстой кишки длинным цилиндрическим эндоскопом с подсветкой.
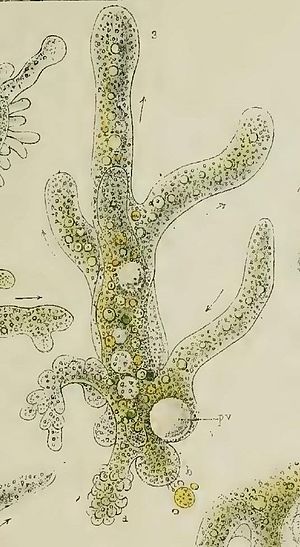 ) Биопсия может выявить повреждение слизистой оболочки кишечника, а также трофозоиты (стадия жизненного цикла инфекционного организма).
) Биопсия может выявить повреждение слизистой оболочки кишечника, а также трофозоиты (стадия жизненного цикла инфекционного организма). - исследование кала на трофозоиты. Трофозоиты трудно обнаружить в фекалиях. Для повышения их видимости часто используются специальные красители.
- центральные краны спинномозговой жидкости (ЦСЖ). Инфекции, связанные с менингоэнцефалитной формой заболевания, могут проявлять аномалии, включая повышенное количество лейкоцитов, аномальные уровни белка и ксантохромию.
- МРТ головного мозга может выявить гранулемы в менингоэнцефалитной форме.
- биопсии головного мозга.
Лечение
Метронидазол используется для контроля симптомов колита и обычно дает положительный эффект. Тем не менее, системные формы заболевания (т. е. инфекции, которые распространяются через кровоток) обычно приводят к летальному исходу, несмотря на лечение, хотя можно попытаться провести симптоматическое лечение.
Родственная дискриминация увеличивается с увеличением генетического расстояния у социальной амебы
Abstract
У социальной амебы Dictyostelium discoideum тысячи клеток объединяются при голодании, образуя многоклеточное плодовое тело, и примерно 20% из них умирают, образуя стебель, который приносит пользу другим.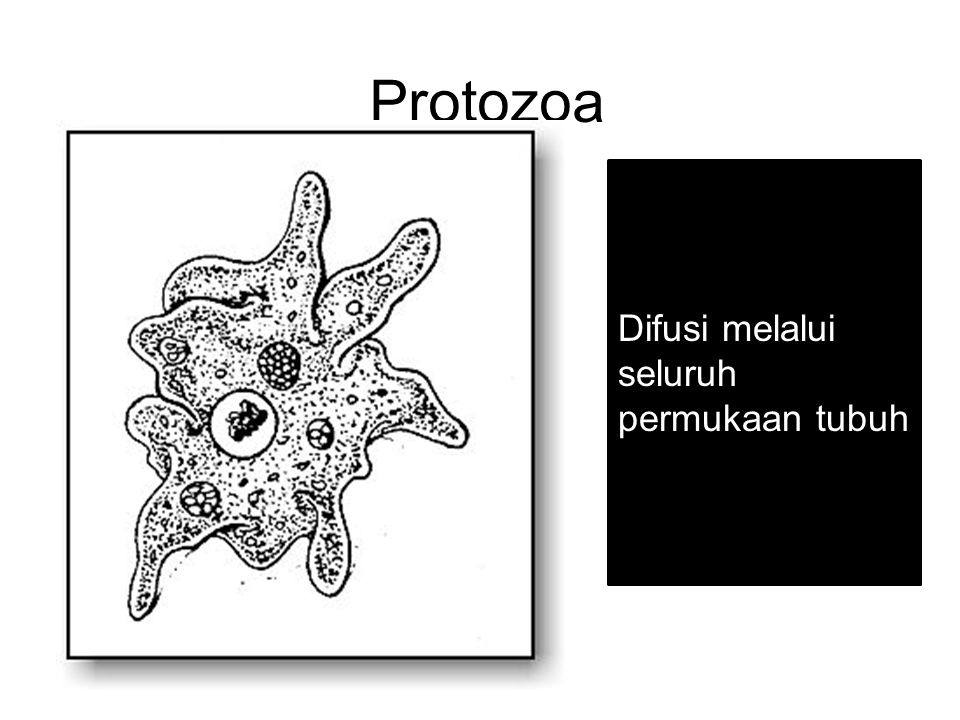 Агрегативный характер многоклеточного развития делает клетки уязвимыми для эксплуатации мошенниками, а потенциал для обмана действительно высок. Клетки могли бы избежать преследования, если бы они могли различать людей и избегать тех, кто генетически отличается. Мы проверили, насколько широко социальные амебы сотрудничают, смешивая изоляты из разных мест, которые покрывают большую часть их естественного ареала. Здесь мы показываем, что разные изоляты частично исключают друг друга при агрегации, и существует положительная связь между степенью этого исключения и генетической дистанцией между штаммами.Наши результаты показывают, что клетки D. discoideum больше коагрегируются с генетически сходными, чем с разнородными особями, что свидетельствует о существовании механизма, определяющего степень генетического сходства между особями в этом социальном микроорганизме.
Агрегативный характер многоклеточного развития делает клетки уязвимыми для эксплуатации мошенниками, а потенциал для обмана действительно высок. Клетки могли бы избежать преследования, если бы они могли различать людей и избегать тех, кто генетически отличается. Мы проверили, насколько широко социальные амебы сотрудничают, смешивая изоляты из разных мест, которые покрывают большую часть их естественного ареала. Здесь мы показываем, что разные изоляты частично исключают друг друга при агрегации, и существует положительная связь между степенью этого исключения и генетической дистанцией между штаммами.Наши результаты показывают, что клетки D. discoideum больше коагрегируются с генетически сходными, чем с разнородными особями, что свидетельствует о существовании механизма, определяющего степень генетического сходства между особями в этом социальном микроорганизме.
Резюме автора
У социальных амеб, таких как Dictyostelium discoideum, клетки объединяются, образуя многоклеточную слизь, которая мигрирует, а затем формирует плодовое тело, содержащее живые споры (из которых впоследствии образуются новые амебы) и мертвые клетки стебля. В отличие от животных, у которых все клетки происходят из одной оплодотворенной яйцеклетки, плодовые тела социальных амеб могут содержать клетки с разными генотипами. Этот потенциал для химеризма создает концептуальную проблему в том, что могут возникать «мошеннические» клетки, которые преимущественно становятся репродуктивными спорами и вынуждают жертв становиться клетками стебля и умирать. Один из способов, с помощью которого амебы могут избежать обмана, заключается в том, что они распознают и предпочтительно объединяются с генетически сходными клетками, избегая при этом генетически отдаленных клеток — процесс, называемый родственной дискриминацией.Мы проверили, могут ли клетки D. discoideum различать таким образом. Мы смешали клетки генетически различных штаммов и обнаружили, что они разделяются во время многоклеточного развития. Степень сегрегации постепенно увеличивается с увеличением генетического расстояния между штаммами. Наши результаты демонстрируют существование родственной дискриминации у D.
В отличие от животных, у которых все клетки происходят из одной оплодотворенной яйцеклетки, плодовые тела социальных амеб могут содержать клетки с разными генотипами. Этот потенциал для химеризма создает концептуальную проблему в том, что могут возникать «мошеннические» клетки, которые преимущественно становятся репродуктивными спорами и вынуждают жертв становиться клетками стебля и умирать. Один из способов, с помощью которого амебы могут избежать обмана, заключается в том, что они распознают и предпочтительно объединяются с генетически сходными клетками, избегая при этом генетически отдаленных клеток — процесс, называемый родственной дискриминацией.Мы проверили, могут ли клетки D. discoideum различать таким образом. Мы смешали клетки генетически различных штаммов и обнаружили, что они разделяются во время многоклеточного развития. Степень сегрегации постепенно увеличивается с увеличением генетического расстояния между штаммами. Наши результаты демонстрируют существование родственной дискриминации у D. discoideum, способности, которая, вероятно, снижает вероятность мошенничества и гарантирует, что смерть клеток стебля дает преимущество в приспособленности родственным особям.
discoideum, способности, которая, вероятно, снижает вероятность мошенничества и гарантирует, что смерть клеток стебля дает преимущество в приспособленности родственным особям.
Образец цитирования: Островски Э.А., Катох М., Шаульский Г., Квеллер Д.К., Штрассманн Дж.Е. (2008) Родственная дискриминация увеличивается с генетическим расстоянием у социальной амебы. ПЛОС Биол 6(11): е287. https://doi.org/10.1371/journal.pbio.0060287
Академический редактор: Nick H. Barton, Эдинбургский университет, Соединенное Королевство
Получено: 23 июня 2008 г.; Принято: 10 октября 2008 г.; Опубликовано: 25 ноября 2008 г.
Copyright: © 2008 Ostrowski et al.Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.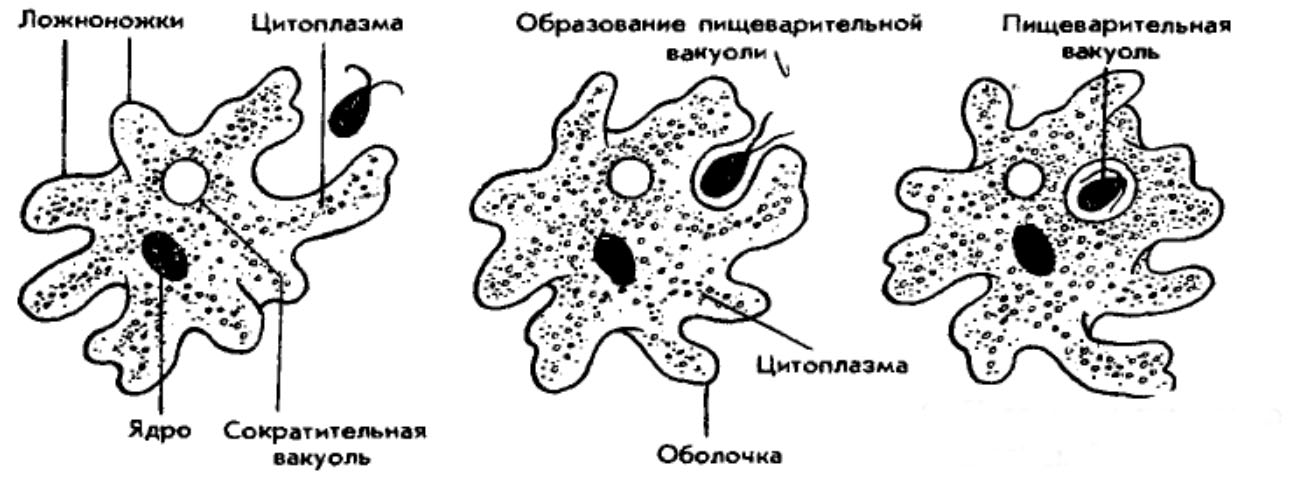
Финансирование: Этот материал основан на работе, поддержанной программой Национального научного фонда (NSF) в рамках гранта EF-0626963. EAO была поддержана постдокторской стипендией от Центра междисциплинарной подготовки биологических наук Кека консорциумов побережья Мексиканского залива (номер гранта NLM 5T15LM07093).
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Сокращения: ГФП, зеленый флуоресцентный белок
Введение
Способность распознавать родственников и предпочтительно взаимодействовать с ними может способствовать развитию альтруистических или кооперативных черт [1,2]. Микроорганизмы демонстрируют сложное социальное поведение [3-6], но мало что известно о генетическом и географическом масштабе их сотрудничества [3].Социальные черты, в частности, могут быть склонны к возникновению несовместимости: отбор, направленный на избежание потенциальных издержек сотрудничества, включая мошенничество, может привести к быстрой эволюции дискриминации или других локусов и ограничить сотрудничество близкородственными штаммами.
Социальная амеба D. discoideum (ранее известная как клеточная слизевики) предлагает уникальную возможность изучить взаимосвязь между генетической дистанцией и дискриминацией у кооперативного микроба. Он гаплоидный, и его геном содержит многочисленные микросателлитные локусы, позволяющие количественно оценить генетические различия между особями.Он имеет географически ограниченный ареал и встречается преимущественно в лесных почвах восточной части Северной Америки и Восточной Азии [7]. При голодании одноклеточные амебы собираются группами примерно по 10 4 –10 5 клеток, образуя многоклеточный агрегат. Совокупность может мигрировать к свету и теплу и в конечном итоге превратиться в плодовое тело, состоящее из шара спор, удерживаемого в воздухе жесткой клеточной ножкой. Приблизительно 70–80 % клеток исходного агрегата образуют споры, тогда как 20–30 % клеток погибают и образуют стебель.Формирование стебля считается альтруистическим, потому что клетки стебля умирают, чтобы принести пользу спорам, поднимая их над землей, что может увеличить их шансы на распространение и защитить их от опасностей в почве, пока они спорообразуют [8-11].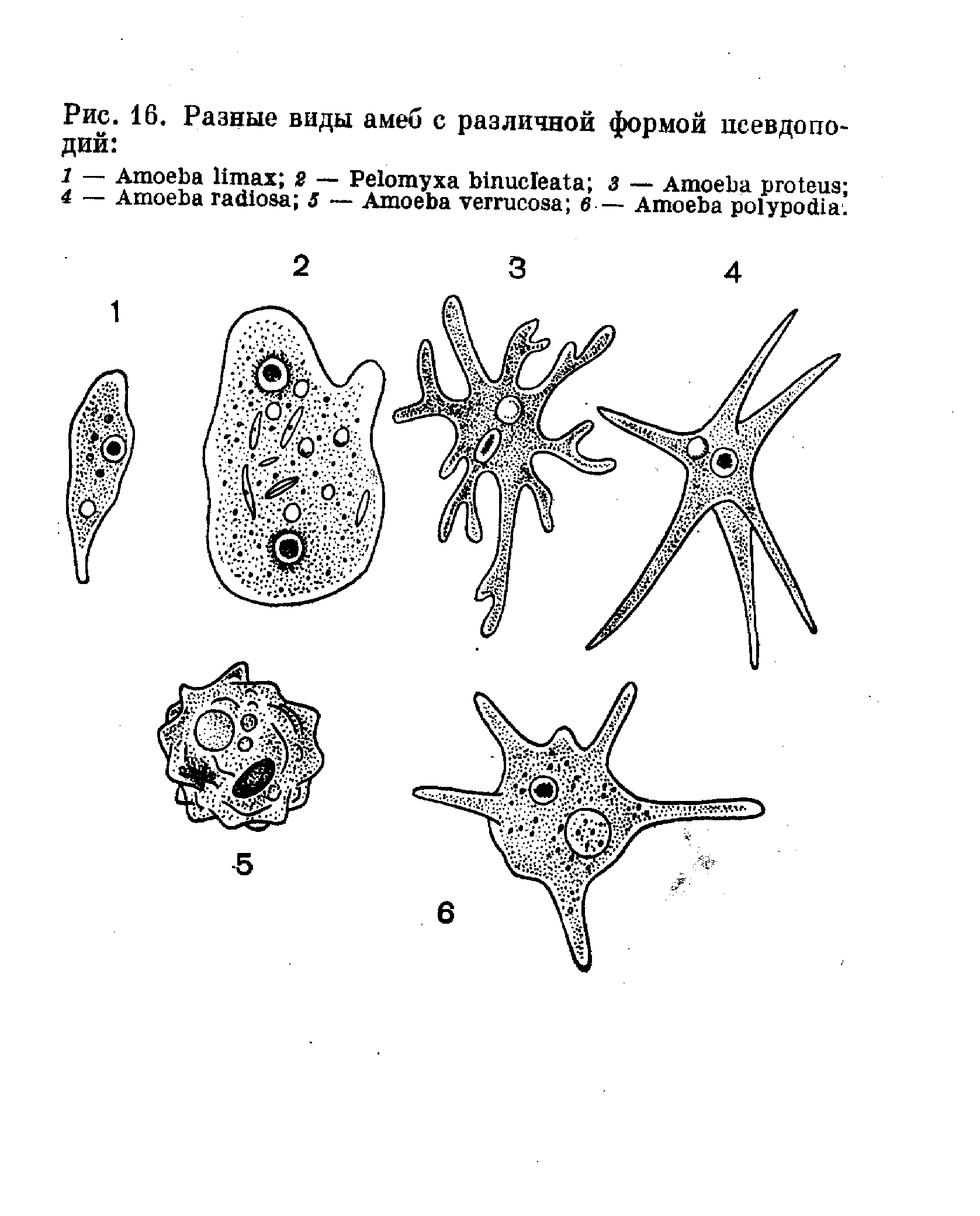
Агрегация у D. discoideum может происходить между амебами, которые генетически различны, и поэтому эволюционная теория предсказывает отбор мошенников — генотипов, которые получают выгоду от стебля, но не вносят свой вклад в его производство [12–15].Действительно, исследования природных изолятов показали, что генетически различные штаммы D. discoideum могут образовывать химеры в лаборатории, которые могут различаться по своему расположению к предспоровым и предстебельным областям слизняка [8]. Генетические скрининги для изучения мошеннического поведения у лабораторного штамма также выявили многочисленные гены, которые при нарушении приводят к чрезмерному представлению этого мутанта в спорах [16].
Продемонстрированная повсеместность и легкость социального мошенничества у D. discoideum ставит загадку: что поддерживает жертв в природе? Одна из возможностей состоит в том, что мошенники имеют более низкую приспособленность, чем кооператоры, когда не являются химерами.Если это так, то преимущество приспособленности, полученное мошенниками, может быть уменьшено или устранено механизмами, которые приводят к разделению мошенников и кооператоров на отдельные плодовые тела [14,17,18]. Есть два объяснения тому, как может произойти это разделение. Одна из возможностей заключается в том, что мошенники и жертвы редко взаимодействуют между собой, поскольку структура популяции пассивно ведет к формированию преимущественно клональных плодовых тел. Другая возможность заключается в том, что штаммы отделяются друг от друга до или во время многоклеточного развития, что является формой дискриминации по родству.Дискриминация родственников отличается от распознавания родственников тем, что последний термин относится к когнитивным процессам, тогда как дискриминация родственников описывает наблюдаемые модели поведения [19–22]. Доказательства кин-дискриминации предоставлены исследованием другого вида (D. purpureum), которое показало, что клетки отделялись от неидентичных клеток во время многоклеточного развития, хотя мошенничества не наблюдалось [23]. Однако у D. discoideum признаки дискриминации носят лишь косвенный характер: генетически отличные клоны обнаруживаются в непосредственной близости в почве [24], но в плодовых телах часто доминирует один клон, по крайней мере, на богатом субстрате оленьих фекалий, где обнаружено большинство дикорастущих плодовых тел [17].
Есть два объяснения тому, как может произойти это разделение. Одна из возможностей заключается в том, что мошенники и жертвы редко взаимодействуют между собой, поскольку структура популяции пассивно ведет к формированию преимущественно клональных плодовых тел. Другая возможность заключается в том, что штаммы отделяются друг от друга до или во время многоклеточного развития, что является формой дискриминации по родству.Дискриминация родственников отличается от распознавания родственников тем, что последний термин относится к когнитивным процессам, тогда как дискриминация родственников описывает наблюдаемые модели поведения [19–22]. Доказательства кин-дискриминации предоставлены исследованием другого вида (D. purpureum), которое показало, что клетки отделялись от неидентичных клеток во время многоклеточного развития, хотя мошенничества не наблюдалось [23]. Однако у D. discoideum признаки дискриминации носят лишь косвенный характер: генетически отличные клоны обнаруживаются в непосредственной близости в почве [24], но в плодовых телах часто доминирует один клон, по крайней мере, на богатом субстрате оленьих фекалий, где обнаружено большинство дикорастущих плодовых тел [17].
D. discoideum представляет собой генетически управляемую модельную систему, поэтому понимание того, способна ли она обнаруживать и ограничивать кооперацию в соответствии с генетической дистанцией, является важным шагом на пути к выявлению генетической основы лежащих в основе механизмов. Например, исследования мутантов csaA D. discoideum, у которых отсутствует молекула межклеточной адгезии gp80, показали, что клеток csaA – имеют тенденцию теряться из химерных агрегатов с клетками дикого типа на природных субстратах. предполагая, что различия в клеточной адгезии между штаммами могут способствовать различению [25,26].Однако о сегрегации между дикими изолятами D. discoideum не сообщалось, что делает неясным актуальность этого вывода.
Мы исследовали несколько паттернов дискриминации у D. discoideum. Во-первых, мы напрямую проверили, способны ли генетически разные изоляты отделяться друг от друга во время многоклеточного развития. Во-вторых, мы определили, в какой степени генетическая дистанция между штаммами влияет на степень этого исключения. Наконец, мы изучили фенотипическую основу сегрегации среди различных смесей в свете различных возможных объяснений сортировки, основанных на предыдущей работе [27,28].
Наконец, мы изучили фенотипическую основу сегрегации среди различных смесей в свете различных возможных объяснений сортировки, основанных на предыдущей работе [27,28].
Результаты
Чтобы изучить взаимосвязь между генетическим сходством штаммов и степенью сегрегации, которую они проявляют во время формирования плодовых тел, мы выполнили попарные смеси эталонного штамма и группы природных изолятов (таблица S1). Для оценки генетических дистанций между штаммами мы генотипировали их по 12 полиморфным микросателлитным локусам, рассредоточенным по всему геному. Мы рассчитали стандартизированное евклидово расстояние между штаммами на основе размеров их микросателлитных аллелей и использовали его в качестве оценки генетической дивергенции и, таким образом, в качестве показателя вероятности того, что штаммы имеют общие аллели (таблица S2).Генетическая дистанция, таким образом, похожа на родство в том смысле, что обе меры являются оценками идентичности по происхождению, хотя формально они различаются, поскольку последнее выражается относительно частот аллелей в эталонной популяции [29,30].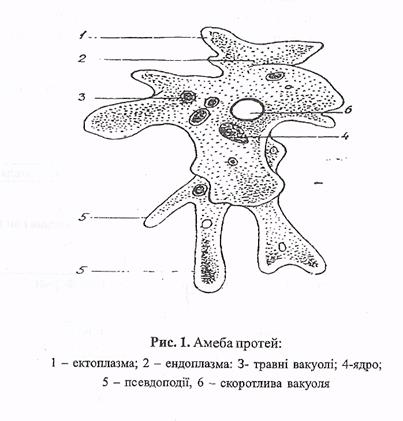 Что еще более важно, поскольку генетическое расстояние учитывает не только аллельную идентичность, но и расстояние между аллелями, оно обеспечивает большее разрешение, чем меры родства, основанные на общих аллелях для дивергентных штаммов, взятых из разных географических мест.
Что еще более важно, поскольку генетическое расстояние учитывает не только аллельную идентичность, но и расстояние между аллелями, оно обеспечивает большее разрешение, чем меры родства, основанные на общих аллелях для дивергентных штаммов, взятых из разных географических мест.
Сначала мы смешали лабораторный штамм AX4-GFP (меченый путем трансформации геном зеленого флуоресцентного белка) с каждым из 14 природных изолятов, штаммом, из которого он был получен (натуральный изолят NC4), и немеченым AX4 (контроль).Для каждой смеси мы объединяли меченые и немеченые амебы в равных пропорциях, наносили смесь на влажные нитроцеллюлозные фильтры и давали им возможность агрегировать и формировать плодовые тела. Из каждой смеси отбирали по десять плодовых тел и определяли соотношение флуоресцентных и нефлуоресцентных спор в каждом из них. Мы использовали среднюю дисперсию этой доли по плодовым телам, основанную как минимум на трех независимых во времени повторах, в качестве оценки степени сегрегации для данной пары штаммов.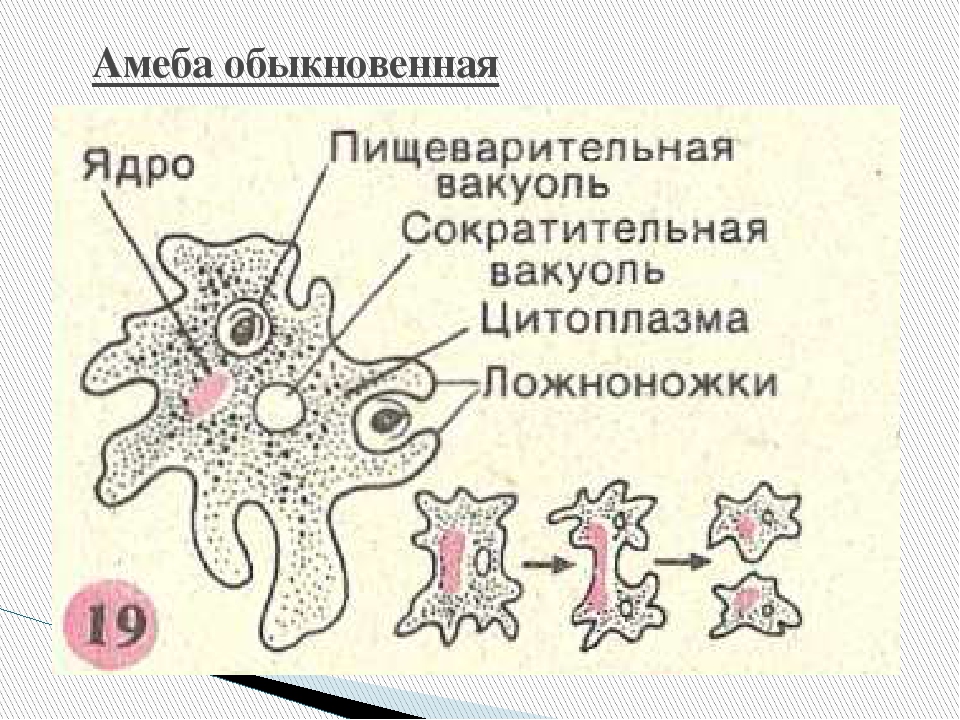
Эксперимент по смешиванию может иметь несколько результатов (рис. 1). В отсутствие какой-либо дискриминации все плодовые тела должны демонстрировать одинаковые пропорции двух клонов, что приводит к низкой дисперсии в этом показателе и отсутствию различий между смесями изолятов с разным генетическим расстоянием (рис. 1А). При исключительном различении себя-не-я люди должны сотрудничать и формировать плодовые тела с генетически идентичными клетками, но отделяться от всех других штаммов, что приводит к строго бинарной реакции (рис. 1В).В качестве альтернативы, если степень дискриминации зависит от генетического сходства между штаммами, мы ожидаем увидеть градуированную связь между генетическим расстоянием и степенью сортировки (рис. 1C).
Рисунок 1. Гипотетические закономерности дискриминации
(левая панель) Отклонение отдельных плодовых тел от среднего значения популяции. Каждый символ (+) представляет отдельное плодовое тело, а смеси нанесены на график в порядке увеличения генетического расстояния. (Правая панель) Изменчивость плодовых тел в зависимости от генетического расстояния.Незакрашенные кружки представляют собой контрольную смесь между генетически идентичными мечеными и немечеными клетками. Полные кружки представляют собой смеси между генетически различными клетками. Рассматриваем три гипотезы:
(Правая панель) Изменчивость плодовых тел в зависимости от генетического расстояния.Незакрашенные кружки представляют собой контрольную смесь между генетически идентичными мечеными и немечеными клетками. Полные кружки представляют собой смеси между генетически различными клетками. Рассматриваем три гипотезы:
(A) Нет дискриминации. На левой панели показано, что каждое плодовое тело содержит одинаковые пропорции двух клонов. Правая панель показывает, что результирующая дисперсия среди плодовых тел невелика, и нет никакой разницы между самосмесями (незаштрихованные кружки) и несамосмесями (закрашенные кружки).
(B) Исключительная дискриминация себя-не-я.Левая панель показывает, что меченый штамм хорошо сочетается с генетически идентичными клетками, но плохо с другими клонами. Правая панель показывает, что существует разница между дисперсией собственного микширования и чужих смесей, но нет разницы между чужими смесями.
(C) Дискриминация по генетическому сходству. На левой панели показано, что смеси генетически идентичных клеток дают хорошо смешанные плодовые тела, но по мере увеличения генетического расстояния между клонами наблюдается сегрегация на отдельные плодовые тела.Правая панель показывает увеличение дисперсии пропорционально генетической дистанции между штаммами.
На левой панели показано, что смеси генетически идентичных клеток дают хорошо смешанные плодовые тела, но по мере увеличения генетического расстояния между клонами наблюдается сегрегация на отдельные плодовые тела.Правая панель показывает увеличение дисперсии пропорционально генетической дистанции между штаммами.
https://doi.org/10.1371/journal.pbio.0060287.g001
Результаты экспериментов по смешиванию подтверждают третью модель (рис. 2). Когда AX4-GFP смешивали либо с немеченым AX4 (контроль), либо с родительским диким изолятом NC4 (ранг генетического расстояния 1 и 2 соответственно), пропорция GFP-позитивных спор была одинаковой между разными плодовыми телами (рис.2А), а дисперсия была низкой (рис. 2В), что указывает на низкую сортировку. Однако смеси AX4-GFP с изолятами с увеличивающимся генетическим расстоянием приводили к большей сегрегации, что отражалось в более высокой дисперсии, а смеси наиболее генетически отдаленных штаммов приводили к образованию плодовых тел двух классов, что указывает на то, что штаммы отделились друг от друга (рис. 2). Мы наблюдали очень значимую корреляцию между генетической дистанцией и дисперсией (коэффициент корреляции Пирсона: r = 0.773, n = 16, двусторонний p <0,0001), что указывает на усиление сегрегации пропорционально генетической дистанции между штаммами. Поскольку генетические расстояния были ненормально распределены, мы также провели непараметрическую корреляцию, которая также была очень значимой (ранговая корреляция Спирмена: ρ = 0,631, n = 16, двусторонняя p = 0,009). Наконец, несмотря на ограниченное разрешение для различения более отдаленных родственных штаммов, анализы, в которых генетическая дистанция оценивалась на основе количества общих аллелей, а не различий в размерах аллелей, дали столь же значимые результаты (ранговая корреляция Спирмена: ρ = 0.798, n = 16, p = 0,0002).
2). Мы наблюдали очень значимую корреляцию между генетической дистанцией и дисперсией (коэффициент корреляции Пирсона: r = 0.773, n = 16, двусторонний p <0,0001), что указывает на усиление сегрегации пропорционально генетической дистанции между штаммами. Поскольку генетические расстояния были ненормально распределены, мы также провели непараметрическую корреляцию, которая также была очень значимой (ранговая корреляция Спирмена: ρ = 0,631, n = 16, двусторонняя p = 0,009). Наконец, несмотря на ограниченное разрешение для различения более отдаленных родственных штаммов, анализы, в которых генетическая дистанция оценивалась на основе количества общих аллелей, а не различий в размерах аллелей, дали столь же значимые результаты (ранговая корреляция Спирмена: ρ = 0.798, n = 16, p = 0,0002).
Рисунок 2. Увеличение сегрегации в зависимости от генетического расстояния в смешанных плодовых телах
Референтные клетки (AX4-GFP) смешивали в равных пропорциях с тестовыми клетками с разным генетическим расстоянием, и смеси давали возможность сформировать плодовые тела.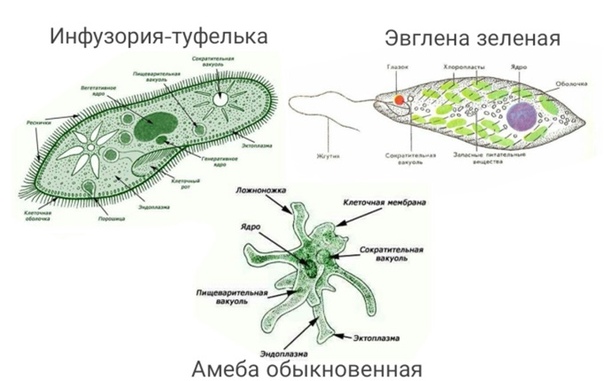 Количество GFP-позитивных и негативных спор определяли в десяти индивидуальных плодовых телах для каждой из трех или четырех независимых смесей.
Количество GFP-позитивных и негативных спор определяли в десяти индивидуальных плодовых телах для каждой из трех или четырех независимых смесей.
(A) Объединенные данные из повторных смесей, показывающие долю GFP-позитивных спор в каждом плодовом теле (+), центрированные вокруг среднего значения и нанесенные на график в зависимости от рангового генетического расстояния от эталонного штамма AX4.
(B) Средняя дисперсия доли GFP-позитивных спор для каждого из 16 штаммов, построенная, как указано выше, на основе трех или четырех независимых экспериментов по смешиванию для каждой пары штаммов. Корреляция между дисперсией и генетической дистанцией была положительной и статистически значимой (корреляция Спирмена ρ = 0,631, n = 16, p = 0,009), что указывает на большую сегрегацию с увеличением генетической дистанции. Данные для штаммов QS33 и QS32 нанесены на график отдельно (ранговые генетические расстояния 11 и 12 соответственно), но им присвоены одинаковые ранги для целей расчета коэффициента ранговой корреляции Спирмена.
https://doi.org/10.1371/journal.pbio.0060287.g002
Чтобы проверить общность результатов, мы повторили наши эксперименты с другой комбинацией штаммов. Мы выбрали два природных изолята (QS32 и QS33), которые плохо смешивались с AX4-GFP, но были тесно связаны друг с другом (идентичны по всем исследованным нами микросателлитным локусам), и третий штамм (QS38), который в равной степени отличался от обоих ( Таблица S1). Если степень дискриминации можно предсказать на основе генетического сходства, то генетически сходные штаммы QS32 и QS33 должны хорошо смешиваться друг с другом и отделяться от генетически отдаленного штамма QS38.Чтобы проверить это предсказание, мы пометили штамм QS32 жизненно важным флуоресцентным красителем и разработали его в попарных смесях с двумя другими штаммами и с немечеными клетками QS32 в качестве контроля. Мы наблюдали небольшую сегрегацию в контрольной смеси и в смеси генетически сходных штаммов QS32 и QS33 (рис. 3). Напротив, смешивание QS32 с генетически отдаленным штаммом QS38 привело к получению плодовых тел с более изменчивым соотношением меченых спор, что указывает на более сильную сегрегацию.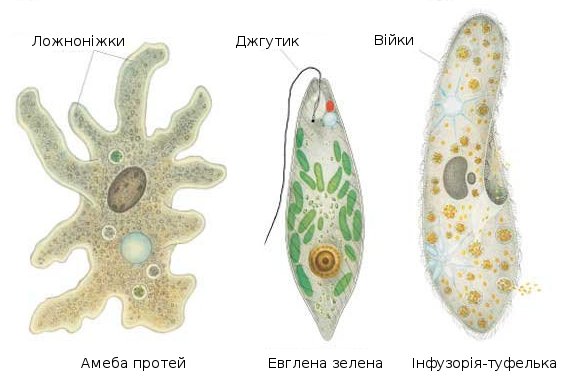 Эти результаты согласуются с исследованиями, проведенными с меченым лабораторным штаммом, предполагая, что свойство генетически связанной сегрегации является транзитивным [31] и устойчивым к выбору штамма.
Эти результаты согласуются с исследованиями, проведенными с меченым лабораторным штаммом, предполагая, что свойство генетически связанной сегрегации является транзитивным [31] и устойчивым к выбору штамма.
Рисунок 3. Свойство сегрегации является транзитивным и устойчивым к выбору штамма различное географическое положение) и QS38 (идентичные по одному генетическому маркеру). Смешанным клеткам давали возможность сформировать плодовые тела, и количество флуоресцентных и нефлуоресцентных спор определяли по меньшей мере в десяти плодовых телах.Доля флуоресцентных спор в каждом плодовом теле (+) представлена как функция рангового генетического расстояния от эталонного штамма QS32.
https://doi.org/10.1371/journal.pbio.0060287.g003
Сегрегация может быть результатом дифференциальной агрегации или постагрегативной сегрегации. Чтобы различать эти возможности, мы трансформировали один из природных изолятов вектором экспрессии GFP (QS44-GFP) и смешали его с генетически отличающимся штаммом, помеченным вектором экспрессии DsRed (AX4-DsRed, рисунок 4A).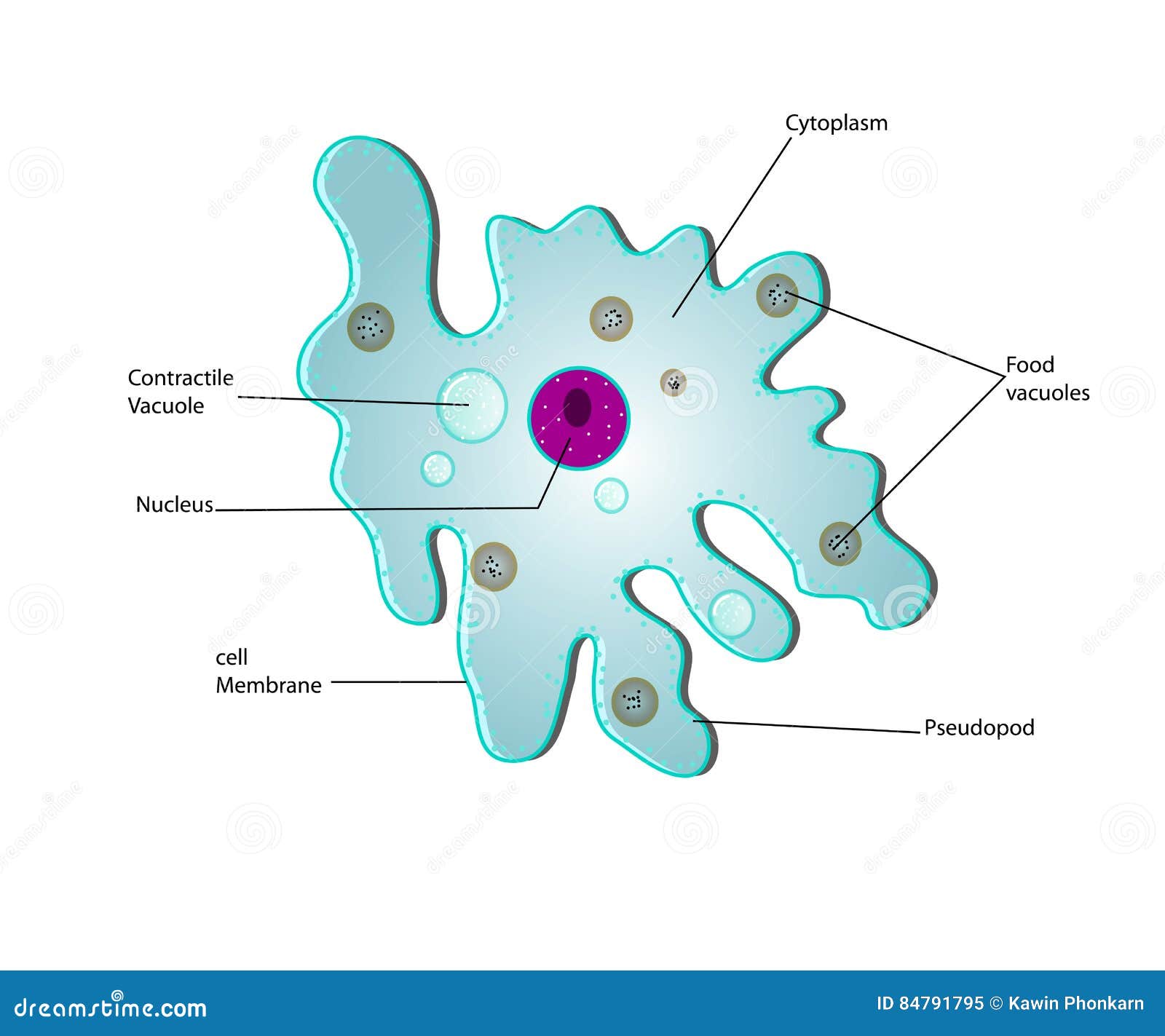 В качестве контроля мы также смешали клетки AX4-GFP с клетками AX4-DsRed (рис. 4B). Как в экспериментальной, так и в контрольной смесях меченые и немеченые клетки были хорошо перемешаны при первоначальном посеве на агар, чтобы вызвать развитие (рис. 4А и 4В, 0 ч). По мере развития обе смеси инициировали агрегацию, и все клетки двигались к одним и тем же центрам агрегации независимо от их генетического сходства (рис. 4А и 4В, 9 ч). Однако кластеры дифференциально меченых клеток становились все более очевидными в смесях генетически разнородных штаммов (рис. 4А, 9 ч), тогда как генетически идентичные клетки оставались смешанными (рис. 4В, 9 ч).Сегрегация генетически несходных штаммов продолжалась на протяжении всей стадии агрегации, после чего происходило частичное разделение меченых и немеченых клеток на разные агрегаты, а также сегрегация внутри агрегатов (рис. 4А, 13 ч). Таким образом, генетически разные штаммы разделяются, но делают это несовершенно. Контрольные смеси не показали сегрегации на этом этапе или в любое более позднее время (рис. 4В, 13 ч и неопубликованные данные). Мы наблюдали подобное расщепление в смесях AX4 с генетически различными изолятами QS32 и QS38 (неопубликованные данные).Постагрегативная природа сегрегации предполагает, что сортировка не является результатом различий во времени развития или использования разных хемоаттрактантов, которые, как известно, уменьшают межвидовой химеризм [27].
В качестве контроля мы также смешали клетки AX4-GFP с клетками AX4-DsRed (рис. 4B). Как в экспериментальной, так и в контрольной смесях меченые и немеченые клетки были хорошо перемешаны при первоначальном посеве на агар, чтобы вызвать развитие (рис. 4А и 4В, 0 ч). По мере развития обе смеси инициировали агрегацию, и все клетки двигались к одним и тем же центрам агрегации независимо от их генетического сходства (рис. 4А и 4В, 9 ч). Однако кластеры дифференциально меченых клеток становились все более очевидными в смесях генетически разнородных штаммов (рис. 4А, 9 ч), тогда как генетически идентичные клетки оставались смешанными (рис. 4В, 9 ч).Сегрегация генетически несходных штаммов продолжалась на протяжении всей стадии агрегации, после чего происходило частичное разделение меченых и немеченых клеток на разные агрегаты, а также сегрегация внутри агрегатов (рис. 4А, 13 ч). Таким образом, генетически разные штаммы разделяются, но делают это несовершенно. Контрольные смеси не показали сегрегации на этом этапе или в любое более позднее время (рис. 4В, 13 ч и неопубликованные данные). Мы наблюдали подобное расщепление в смесях AX4 с генетически различными изолятами QS32 и QS38 (неопубликованные данные).Постагрегативная природа сегрегации предполагает, что сортировка не является результатом различий во времени развития или использования разных хемоаттрактантов, которые, как известно, уменьшают межвидовой химеризм [27].
Рисунок 4. Сортировка штаммов во время многоклеточного развития
Клетки, экспрессирующие либо GFP, либо DsRed, смешивали в равных пропорциях и оставляли для развития на чашках с агаром. Снимки были сделаны в указанные моменты времени развития, и показано объединенное изображение двух флуорофоров.
(A) Смесь генетически несходных штаммов AX4-DsRed и QS44-GFP демонстрирует усиление сегрегации со временем.
(B) Смесь генетически идентичных штаммов AX4-DsRed и AX4-GFP не показывает сегрегации.
https://doi.org/10.1371/journal.pbio.0060287.g004
Обсуждение
Наши результаты показывают, что социальные амебы различают генетически сходные и несходные клетки постепенным образом, больше смешиваясь с первыми и отделяясь от вторых в процессе многоклеточного развития.Сходство паттернов сегрегации среди ряда различных изолятов предполагает общий основной механизм и согласуется с различиями, которые могут возникать из-за дифференциальной клеточной адгезии [28]. Например, межклеточная адгезия необходима для потока клеток, а пространственное разделение предстебельных и предспоровых клеток, которое также происходит в холмике после агрегации, объясняется различиями в относительной адгезивности этих разных типов клеток [26,32]. –35].
Мы предлагаем два эволюционных объяснения этих паттернов дискриминации, которые не исключают друг друга [31,36,37].Одна возможность состоит в том, что генетический дрейф или адаптация к различным компонентам окружающей среды вызывает генетическую дивергенцию в локусах, которые вызывают дискриминацию, что в конечном итоге приводит к уменьшенной способности формировать химерные плодовые тела, процесс, аналогичный аллопатрическому видообразованию [38]. В качестве альтернативы дивергенция может возникнуть в результате отбора, чтобы избежать издержек химеризма, включая мошенничество. Предыдущие исследования D. discoideum показали, что мошенники широко распространены в природе [8,39] и что его геном содержит многочисленные гены, которые при мутациях придают мошенническое поведение [16].Знание распределения генетической изменчивости в природе может помочь определить, как часто чужеземные особи встречаются друг с другом, и важность отбора в управлении паттернами дискриминации, которые мы здесь показываем [31]. Тем не менее, наши данные дают возможное объяснение высокого уровня клональности плодовых тел в природных популяциях [17], а также сосуществования мошенников и жертв в непосредственной близости друг от друга в природе [24].
Хотя мы наблюдали сильную корреляцию между генетическим расстоянием и степенью сегрегации, большинство плодовых тел сохранили некоторую репрезентативность обоих клонов, что указывает на некоторую способность коагрегировать и образовывать многоклеточные структуры, которые охватывают почти весь ареал вида.Более того, продемонстрированная способность этих штаммов к сегрегации, но их неспособность сделать это полностью, предполагает, что важный компонент выяснения роли отбора в способности к различению будет включать количественную оценку не только затрат на формирование химер, таких как мошенничество, но и преимущества, включая увеличение совокупного размера [31,40]. Наши результаты также отличаются от большинства других примеров дискриминационного поведения у микробов, таких как производство токсинов у бактерий [41] и вегетативная несовместимость у грибов [42].В этих случаях дискриминационное поведение демонстрирует сильное распознавание себя-не-я, а фенотипы дискриминации, которые часто включают гибель клеток, строго бинарны.
Интересно, что социальная несовместимость встречается и у почвенной бактерии Myxococcus xanthus, которая также образует многоклеточные плодоносящие структуры в ответ на голодание. В этой системе наблюдается сильный антагонизм между географически разными изолятами, при этом смешение вызывает снижение спорообразования или даже вымирание популяции [43].Напротив, мы наблюдаем мало признаков антагонизма, но увеличивающееся избегание общительности у генетически несходных штаммов, поведение, которое должно ограничить преимущества приспособленности, обеспечиваемые обманом, и помочь объяснить сохранение альтруизма у этого вида. В более общем плане различия между M. xanthus и D. discoideum в их реакции на чужеродных особей иллюстрируют, что разные микробы, несмотря на большое сходство черт истории их социальной жизни, могут находить разные решения проблемы обеспечения сотрудничества [44,45].
Материалы и методы
Штаммы и условия культивирования.
В смесях с AX4-GFP мы выращивали все штаммы-конкуренты, включая AX4, на чашках с SM-агаром (на литр: 10 г глюкозы, 10 г Bacto Peptone (Oxoid), 1 г дрожжевого экстракта (Oxoid), 1 г MgSO 4 , 1,9 г KH 2 PO 4 , 0,6 г K 2 HPO 4 , 20 г агара) в сочетании с Klebsiella pneumoniae при комнатной температуре. Мы выращивали эталонный штамм AX4-GFP аксиально в среде HL5 с добавлением 5 мкг/мл G418 при встряхивании при 22 °C для поддержания экспрессии GFP [46].
Эксперименты по смешиванию.
Мы собирали каждый штамм во время средней экспоненциальной фазы роста, дважды промывали клетки холодным буфером KK2 (14,0 мМ K 2 HPO 4 и 3,4 мМ K 2 HPO 4 , pH = 6,4), и ресуспендировали их при плотности 6 × 10 7 клеток/мл в буфере КК2. Для каждой смеси мы объединяли два штамма в равных пропорциях и наносили аликвоту 1,5 × 10 7 клеток на нитроцеллюлозный фильтр плотностью 3.5 × 10 6 кл/см 2 . В качестве контроля мы также высевали каждый штамм по отдельности при той же плотности клеток, что и смеси. Мы поместили фильтры в чашки Петри на одну фильтровальную подушку (Pall), которая была пропитана 1,5 мл буфера PDF (20,1 мМ KCl, 5,3 мМ MgCl 2 · 6H 2 O, 9,2 мМ K 2 HPO 4 , 13,2 мМ KH 2 PO 4 , 0,5 г/л сульфата стрептомицина, pH = 6,4), поместили их во влажную камеру и инкубировали при 22 °C в темноте для развития.После формирования плодовых тел (около 24 часов) мы случайным образом отбирали не менее десяти отдельных плодовых тел из каждого фильтра смеси. Мы ресуспендировали споры из каждого плодового тела в 10 мкл детергента для устранения амеб и подсчитывали споры с использованием фазово-контрастной и флуоресцентной микроскопии для определения доли GFP-позитивных спор. Мы насчитали около 400 спор на каждое плодовое тело.
Окрашивание клеточным трекером.
Мы выращивали каждый дикий изолят до средней экспоненциальной фазы в сочетании с K.pneumoniae на чашках с SM-агаром, дважды промывали клетки холодным буфером КК2 и ресуспендировали при плотности 1 × 10 7 клеток/мл. Мы окрашивали клетки с помощью CellTracker Green CMFDA (Molecular Probes) в соответствии с рекомендованным производителем протоколом со следующими модификациями. Мы добавили реагент для отслеживания клеток в концентрации 50 мкМ, инкубировали клетки в течение 30 минут, дважды промыли их холодным буфером KK2 и инкубировали еще 30 минут в буфере KK2, чтобы позволить клеткам вывести избыток красителя.После окрашивания клетки ресуспендировали с плотностью 6 × 10 7 клеток/мл, смешивали в равных пропорциях с немечеными клетками и наносили смесь на фильтры, как описано выше. После формирования плодовых тел мы собрали споры из отдельных плодовых тел в детергенте и проанализировали долю флуоресцентных спор на проточном цитометре BD LSRII.
Микросателлитное генотипирование.
Мы генотипировали штаммы по 12 микросателлитным локусам, которые были разбросаны по всему геному.Эти микросателлитные локусы были созданы на основе последовательности AX4 (наш очаговый штамм) и описаны ранее [47]. Для выделения геномной ДНК споры собирали с 5–10 плодовых тел и инкубировали их в смеси 150 мкл 5% Bio-Rad Chelex-10 и 10 мкл 20 мг/мл протеиназы К в течение 4 ч при 56 °С. затем 30 мин при 98°С. Каждый микросателлитный локус амплифицировали с помощью ПЦР с использованием флуоресцентно меченных праймеров (таблица S2). Полученный продукт анализировали на секвенаторе ABI 3100 и программах GeneScan 3.7 и GENOTYPER использовали для определения размера фрагмента. Чтобы оценить генетические расстояния, мы рассчитали стандартизированное евклидово расстояние между штаммами, используя размер продукта ПЦР в качестве количественной переменной (таблица S1). Родство и генетическое сходство, основанные на многолокусных, многовариантных методах кластеризации, включающих евклидовы расстояния между гаплотипами, были описаны ранее [48-51]. Мы использовали стандартизированную евклидову метрику расстояния, которая масштабирует каждое локус по его дисперсии, так что они вносят одинаковый вклад [52].Кластеризация, основанная на размере аллеля, а не на количестве общих аллелей, обеспечивает лучшее разрешение генетических различий, особенно для более отдаленных родственников. Это аналог пошаговой мутационной модели (SMM). Было показано, что SMM работает хорошо [53–56], особенно когда частота микросателлитных мутаций низка и нет сильного направленного смещения в изменении длины аллеля, оба из которых были показаны для этих локусов (описаны в [47]) . Были повторены анализы, где расстояния рассчитывались на основе наличия общих аллелей, где аллели считались идентичными, если расчетный размер был в пределах 3 п.н. от аллеля для AX4.
Штаммовая конструкция.
Мы инокулировали споры природного изолята QS44 из плодовых тел в чашки Петри, содержащие среду HL5 с добавлением 10% эмбриональной бычьей сыворотки, и выращивали полученные клетки в глубинной культуре до тех пор, пока они полностью не покрывали поверхность. Мы трансформировали клетки вектором экспрессии, содержащим репортер S65T-GFP, управляемый промотором act15. Условия трансформации были изменены по сравнению с протоколами, описанными ранее [57]. Мы собирали клетки после двукратной промывки в ледяном буфере KK2 и ресуспендировали их в концентрации 1 × 10 8 клеток/мл в ледяном буфере H-50.Мы смешали 100 мкл клеточной суспензии с 5–15 мкг плазмиды в кювете для электропорации с зазором 0,1 см и провели электропорацию смеси при 0,95 кВ и 25 мФ три раза с интервалом примерно в 5 с. Трансформанты выращивали в погруженной культуре с HL5, дополненной 8 мкг/мл G418, и выбранные клоны тестировали на экспрессию GFP с помощью флуоресцентной микроскопии. Мы проверили генетический фон с помощью микросателлитного генотипирования.
Статистические методы.
Пропорции были преобразованы из квадратного корня арксинуса, чтобы гарантировать, что дисперсия была статистически независимой от среднего значения, и, таким образом, учесть различия между смесями в общей доле флуоресцентных спор [58].Преобразование не влияет на статистическую значимость результатов. Генетические расстояния были ненормально распределены, поэтому мы также провели непараметрическую (ранг Спирмена) корреляцию. Все парные смешивания с эталонным штаммом AX4-GFP проводили не менее трех раз и использовали для расчета средней дисперсии. Три смеси (NC4, QS43 и QS45) были воспроизведены четыре раза. В таких случаях мы рассчитывали среднюю дисперсию по всем четырем повторам, хотя включение четвертого повтора не влияло на статистическую значимость наших результатов.
Благодарности
Мы благодарим Дж. Ландольта за предоставление нескольких штаммов и Т. Купера, К. Фостера, О. Гилберта и нескольких анонимных рецензентов за полезные обсуждения, советы и комментарии к более ранним версиям этой рукописи
Вклад авторов
ЕАО и МК провели экспериментальную работу. EAO, MK, GS, DCQ и JES задумали исследование, обсудили результаты и написали рукопись.
Примечание добавлено в доказательство
Аналогичные результаты были получены в отношении Dictyostelium purpureum Mehdiabadi et al.[59].
Каталожные номера
- 1. Гамильтон В. Д. (1964) Генетическая эволюция социального поведения. я, II. J Theor Biol 7: 1–52.
- 2. Вест С.А., Гриффин А.С., Гарднер А., Диггл С.П. (2006) Теория социальной эволюции микроорганизмов. Nat Rev Microbiol 4: 597–607.
- 3. Crespi BJ (2001) Эволюция социального поведения у микроорганизмов. Тенденции Ecol Evol 16: 178–183.
- 4. Уэст С.А., Диггл С.П., Баклинг А., Гарднер А., Гриффин А.С. (2007) Социальная жизнь микробов.Annu Rev Ecol Evol Syst 38: 53–77.
- 5. Акерманн М., Чао Л. (2004) Эволюция сотрудничества: два за одного. Курр Биол 14: R73–R74.
- 6. Рейни П.Б., Рейни К. (2003)Эволюция сотрудничества и конфликтов в экспериментальных бактериальных популяциях. Природа 425: 72–74.
- 7. Суонсон А.Р., Ваделл Э.М., Кавендер Дж.К. (1999)Глобальное распространение диктиостелид в лесных почвах. Дж. Биогеогр 26: 133–148.
- 8. Strassmann JE, Zhu Y, Queller DC (2000)Альтруизм и социальное мошенничество у социальной амебы Dictyostelium discoideum.Природа 408: 965–967.
- 9. Kessin RH (2001) Dicytostelium — Эволюция, клеточная биология и развитие многоклеточности. Кембридж (Соединенное Королевство): Издательство Кембриджского университета.
- 10. Шаульский Г., Кессин Р. Х. (2007) Холодная война социальных амеб. Курр Биол 17: R684–R692.
- 11. Гадагкар Р., Боннер Дж. Т. (1994) Социальные насекомые и социальные амебы. J Biosci 19: 219–245.
- 12. Мейнард Смит Дж., Сатмари Э. (1995) Основные переходы в эволюции.Оксфорд: WH Freeman.
- 13. Гросберг Р.К., Стратманн Р.Р. (1998)Одноклеточный, двухклеточный, эритроцитный, синеклеточный: сохранение одноклеточной стадии в многоклеточных историях жизни. Тенденции Ecol Evol 13: 112–116.
- 14. Buss LW (1982) Паразитизм соматических клеток и эволюция совместимости соматических тканей. Proc Natl Acad Sci U S A 79: 5337–5341.
- 15. Басс Л.В. (1987) Эволюция индивидуальности. Принстон: Издательство Принстонского университета.
- 16. Санторелли Л.А., Томпсон С.Р.Л., Вильегас Э., Светц Дж., Динь С. и соавт. (2008) Факультативные мутанты-обманщики раскрывают генетическую сложность сотрудничества у социальных амеб. Природа 451: 1107–1110.
- 17. Гилберт О.М., Фостер К.Р., Мехдиабади Н.Дж., Штрассманн Дж.Е., Квеллер Д.К. (2007) Высокое родство поддерживает многоклеточное сотрудничество в социальной амебе, контролируя мутантов-мошенников. Proc Natl Acad Sci USA 104: 8913–8917.
- 18. Ennis HL, Dao DN, Pukatzki SU, Kessin RH (2000) Dictyostelium amoebae, лишенные белка F-box, образуют споры, а не стебель в химере с диким типом.Proc Natl Acad Sci U S A 97: 3292–3297.
- 19. Holmes WG (2004) Ранняя история гамильтонианских исследований распознавания родства. Энн Зул Фенн 41: 691–711.
- 20. Матео Дж. М. (2004) Системы распознавания и биологическая организация: компонент восприятия социального признания. Энн Зул Фенн 41: 729–745.
- 21. Танг-Мартинес З. (2001) Механизмы распознавания родства и эволюция распознавания родства у позвоночных: критическая переоценка.Поведенческий процесс 53: 21–40.
- 22. Бекофф М., Байерс Дж. А. (1986) Что означает «родственное признание». Этология 72: 342–345.
- 23. Мехдиабади Н.Дж., Джек С.Н., Талли Фарнхэм Т., Платт Т.Г., Калла С.Е. и др. (2006) Родственные предпочтения в социальном микробе. Природа 442: 881–882.
- 24. Fortunato A, Strassmann JE, Santorelli L, Queller DC (2003)Совместное появление в природе различных клонов социальной амебы Dicytostelium discoideum. Мол Экол 12: 1031–1038.
- 25. Queller DC, Ponte E, Bozzaro S, Strassmann JE (2003)Эффекты зеленой бороды с одним геном у социальной амебы Dictyostelium discoideum. Наука 299: 105–106.
- 26. Ponte E, Bracco E, Faix J, Bozzaro S (1998)Обнаружение тонких фенотипов: случай молекулы клеточной адгезии csA в Dicytostelium . Proc Natl Acad Sci U S A 95: 9360–9365.
- 27. Рапер К.Б., Том С. (1941)Межвидовые смеси у Dicytosteliaceae.Ам Дж. Бот 28: 69–78.
- 28. Sternfeld J (1979) Доказательства дифференциальной клеточной адгезии как механизма сортировки различных видов слизевиков. J Embryol Exp Morphol 53: 163–178.
- 29. Queller DC (1994) Генетическое родство в вязких популяциях. Эвол Экол 8: 70–73.
- 30. Hardy OJ (2003)Оценка попарного родства между людьми и характеристика процессов изоляции на расстоянии с использованием доминантных генетических маркеров.Мол Экол 12: 1577–1588.
- 31. Гросберг Р.К. (1988) Эволюция специфичности аллоузнавания у клональных беспозвоночных. Q Rev Biol 63: 377–412.
- 32. Коутс Дж. К., Харвуд А. Дж. (2001) Межклеточная адгезия и передача сигнала во время развития Dicytostelium . J Cell Sci 114: 4349–4358.
- 33. Гомер Р.Х. (2001) Неправильный размер. Nat Rev Mol Cell Biol 2: 48–54.
- 34. Вонг Э., Ян С., Ван Дж., Фуллер Д., Лумис В.Ф. и др.(2002) Нарушение гена, кодирующего молекулу клеточной адгезии DdCAD-1, приводит к аберрантной сортировке клеток и распределению типов клеток во время развития Dictyostelium . Развитие 129: 3839–3850.
- 35. Weijer CJ (2004) Морфогенез Dictyostelium . Curr Opin Genet Dev 14: 392–398.
- 36. Crozier R (1986) Генетические способности к распознаванию клонов у морских беспозвоночных должны поддерживаться путем отбора чего-то еще. Эволюция 40: 1100–1101.
- 37. Крозье Р. (1987)Генетические аспекты распознавания родственников: концепции, модели и синтез. В: Fletcher DJC, компакт-диск Michener, редакторы. Распознавание родства у животных. Чичестер: Джон Вили и сыновья.
- 38. Койн Дж. А., Орр Х. А. (2004) Видообразование. Сандерленд (Массачусетс): Sinauer Associates.
- 39. Fortunato A, Queller DC, Strassmann JE (2003)Линейная иерархия доминирования среди клонов химер социальной амебы Dictyostelium discoideum.J Evol Biol 16: 438–445.
- 40. Фостер К.Р., Фортунато А., Страссманн Дж. Э., Квеллер Д. К. (2002) Затраты и выгоды от того, чтобы быть химерой. Proc R Soc Lond B 269: 2357–2362.
- 41. Райли М.А., Верц Дж.Е. (2002) Бактериоцины: эволюция, экология и применение. Annu Rev Microbiol 56: 117–137.
- 42. Гласс Н.Л., Джейкобсон Д.Дж., Шиу ПКТ (2000)Генетика слияния гиф и вегетативной несовместимости у нитчатых аскомицетов. Annu Rev Genet 34: 165–186.
- 43. Fiegna F, Velicer GJ (2005)Эксплуатационный и иерархический антагонизм в кооперативной бактерии. PLoS Biol 3 (11): 1980–1987.
- 44. Трависано М., Велисер Г.Дж. (2004) Стратегии борьбы с микробными мошенниками. Тенденции микробиол 12: 72–78.
- 45. Velicer GJ (2003) Социальная борьба в микробном мире. Trends Microbiol 11: 330–337.
- 46. Суссман М. (1987) Культивирование и синхронный морфогенез Dictyostelium в контролируемых экспериментальных условиях.Методы Cell Biol 28: 9–29.
- 47. McConnell R, Middlemist S, Scala C, Strassmann JE, Queller DC (2007)Необычно низкая частота мутаций микросателлитов в Dicytostelium discoideum, организме с необычно обильными микросателлитами. Генетика 177: 1499–1507.
- 48. Блуин М.С., Парсонс М., Лакайль В., Лотц С. (1996) Использование микросателлитных локусов для классификации людей по родству. Мол Экол 5: 393–401.
- 49. Blouin MS (2003) Основанные на ДНК методы реконструкции родословной и анализа родства в природных популяциях.Тенденции Ecol Evol 18: 503–511.
- 50. Smouse PE, Peakall R (1999)Пространственный автокорреляционный анализ индивидуальной мультиаллельной и мультилокусной генетической структуры. Наследственность 82: 561–573.
- 51. Excoffier L, Smouse PE, Quattro JM (1992) Анализ молекулярной дисперсии, выведенный из метрических расстояний между гаплотипами ДНК: приложения к данным об ограничении митохондриальной ДНК человека. Генетика 131: 479–491.
- 52. Лоу А., Харрис С., Эштон П. (2004)Экологическая генетика: дизайн, анализ и применение.Оксфорд: Издательство Блэквелл.
- 53. Вальдес А.М., Слаткин М., Фреймер Н.Б. (1993)Частоты аллелей в микросателлитных локусах: пересмотр модели ступенчатой мутации. Генетика 133: 737–749.
- 54. Shriver MD, Jin L, Chakraborty R, Boerwinkle E (1993) Распределение частот аллелей VNTR в модели пошаговой мутации: подход компьютерного моделирования. Генетика 134: 983–993.
- 55. Ди Риенцо А., Петерсон А.С., Гарза Дж.С., Вальдес А.М., Слаткин М. и др.(1994) Мутационные процессы локусов повторов простой последовательности в человеческих популяциях. Proc Natl Acad Sci U S A 91: 3166–3170.
- 56. Takezaki N, Nei M (1996) Генетические расстояния и реконструкция филогенетических деревьев по микросателлитной ДНК. Генетика 144: 389–399.
- 57. Панг К.М., Лайнс М.А., Кнехт Д.А. (1999) Переменные, контролирующие уровень экспрессии экзогенных генов у Dictyostelium . Плазмида 41: 187–197.
- 58. Zar JH (1999) Биостатистический анализ.Река Аппер-Сэдл (Нью-Джерси): Прентис-холл.
- 59. Mehdiabadi NJ, Kronforst MR, Queller DC, Strassmann JE (2008)Филогения, репродуктивная изоляция и родственное распознавание социальной амебы Dictyostelium purpureum. Эволюция. В прессе.