Биология Полости тела. Способы передвижения животных
|
Тема нашего урока «Способы передвижения животных. Полости тела» На уроке мы должны сформировать представление о полости тела животных, ее видах и значении, об эволюционном направлении в изменении типа полостей тела животных. А также познакомиться с основными типами движения животных, показать эволюционное направление в изменении способов движения.
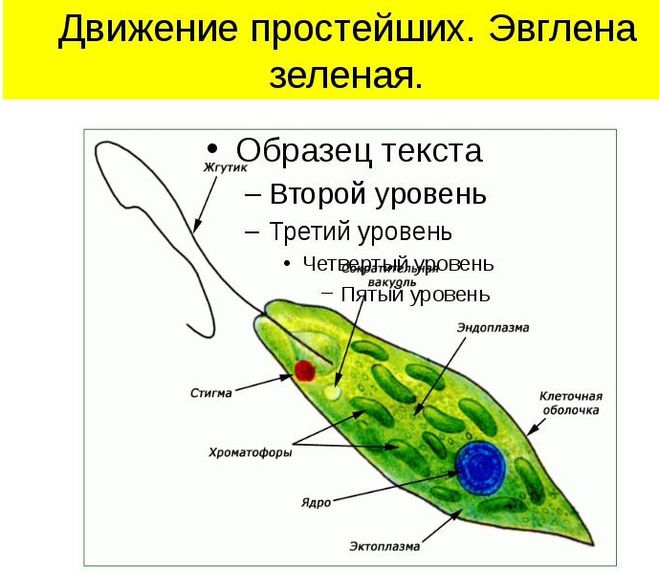
Движение — одно из основных свойств живых организмов. Несмотря на многообразие существующих активных способов передвижения, их можно разделить на 3 основных типа: Амебовидное движение. Движения при помощи жгутиков и ресничек. Движение с помощью мышц.
Давайте познакомимся с типами передвижения животных.
Амебовидное движение присуще одноклеточным организмам из класса Саркодовые. Движения при помощи жгутиков и ресничек характерно не только для жгутиконосцев и инфузорий, оно присуще некоторым многоклеточным животным и их личинкам. Строение всех жгутиков и ресничек практически одинаково. Вращаясь или взмахивая, жгутики и реснички создают движущую силу и закручивают тело вокруг собственной оси. Увеличение числа ресничек убыстряет передвижение. Такой способ движения свойствен обычно мелким беспозвоночным животным, обитающим в водной среде. Движение с помощью мышц осуществляется у многоклеточных животных.Любое движение — это очень сложная, но слаженная деятельность групп мышц в организме.
Мышцы образованы мышечной тканью. Мышечная ткань бывает гладкая и поперечно-полосатая. Главная особенность мышечной ткани — способность сокращаться. За счет сокращения мышц и осуществляется движение. У круглых червей поочередное сокращение продольных мышц вызывает характерные изгибы тела. За счет этих телодвижений червь двигается вперед. У кольчатых червей мускулатура состоит из кольцевых и продольных мышц. При переменном сокращении этих мышц тело дождевого червя может сокращаться и удлиняться.
Очень интересный способ движения – это реактивное движение.
Наибольший интерес представляет реактивный двигатель кальмара. Более сложное развитие мускулатуры в процессе эволюции приобрели хордовые животные. У рыб мускулатура развита хорошо. Мощные мускулы идут вдоль тела, по обеим сторонам позвоночника. Эти боковые мышцы не сплошные, а состоят из отдельных пластинок мышечных отрезков, или сегментов, которые идут один позади другого и разделены между собой тонкими волокнистыми прослойками. Обратите внимание, с какой скоростью плавают рыбы.
По сравнению с рыбами у земноводных только часть туловищной мускулатуры сохраняет лентовидное строение, у них развиваются специализированные мышцы.
Еще сложнее мускулатура пресмыкающихся. Короткие конечности пресмыкающихся, расположенные по бокам туловища, не поднимают тело высоко над землей, и оно волочится по земле. При всем при этом ящерицы быстро бегают. А вот движения крокодила на суше менее быстры и проворны, чем его движения в воде, где он превосходно плавает и ныряет. Его длинный и мускулистый хвост сдавлен с боков и служит хорошим рулем.
Волнообразные изгибания тела — самый распространенный способ ползания змей. Спокойно ползущая змея — удивительно красивое и завораживающее зрелище. Кажется, ничего не происходит. Движения почти незаметны. Тело вроде бы неподвижно лежит и в то же время быстро перетекает. Ощущение легкости передвижения змеи обманчиво. В ее удивительно сильном теле синхронно и размеренно работает множество мускулов, точно и плавно переносящих тело. Ползание змей может быть достаточно стремительным. Однако даже самые быстрые змеи редко развивают скорость, превышающую 8 км/ч. Рекорд скорости ползания — 16-19 км/ч, и принадлежит он черной мамбе.
Птицы приспособлены к полету. У них наиболее развиты большие грудные мышцы, опускающие крылья. Поднимают крылья подключичные мышцы, которые также хорошо развиты и расположены под большими грудными мышцами. Сильно развиты у птиц мускулы задних конечностей и шеи. мышцы у птиц. Другие способы движения птиц весьма разнообразны. Широко применяется условное подразделение птиц на древесных, наземных и водных. Это указывает на известные различия между ними в отношении не только среды обитания, но и способа движения.
Для древесных птиц, помимо полета, характерно еще лазанье по ветвям и стволам деревьев. Для наземных — хождение и бег, а для водных — плаванье и ныряние.
Лазание по ветвям деревьев представляет собой, по-видимому, изначальную форму передвижения птиц. Так должны были передвигаться непосредственные предки птиц из пресмыкающихся, так, несомненно, передвигался в основном археоптерикс, обладающий лишь слабой способностью к полету, так передвигается еще не летающий птенец гоацина, обхватывающий ветки пальцами ног и помогающий себе в лазании передними конечностями, которые снабжены относительно хорошо развитыми пальцами. Несомненно, как приспособление к такому передвижению сложилась типичная нога птицы с тремя пальцами, направленными вперед, и одним, обращенным назад, — для обхватывания веток. В процессе дальнейшей специализации выработалась так называемая лазательная нога. Она имеет мощные пальцы, из которых два обращены вперед, а два — назад, укороченные голень и цевку и способна совершать отводящие и до известной степени вращательные движения. Но все таки основной способ передвижения птиц – это полет. Предлагаем вам познакомиться с интересными фактами из жизни птиц.
Мышечная система млекопитающих достигает исключительного развития и сложности, насчитывает несколько сот мышц. Наиболее развиты мышцы конечностей и туловища, что связано с характером передвижения. За счет этого млекопитающие могут совершать сложные движения и перемешаться в пространстве. Самые быстрые животные – гепарды. Они могут бежать со скоростью 120 км в час Кенгуру- рекордсмен по прыжкам в длину. Внимание! Для понимания данной темы «Полость тела» необходимо ознакомиться с основными понятиями. Полость тела – пространство, расположенное между стенками тела и внутренними органами. Полостная жидкость – жидкость, которая находится в первичной полости тела и омывает внутренние органы. Первичная полость тела – пространство между стенкой тела и кишечником, в котором расположены внутренние органы, не имеющее собственной оболочки. Вторичная полость тела – пространство между стенкой тела и внутренними органами; ограничено собственными эпителиальными оболочками и заполнено жидкостью.
Первичная полость тела заполнена жидкостью и выполняет множество функций: поддержание формы тела, опора, транспорт питательных веществ и накопление ненужных продуктов жизнедеятельности организма. Она присутствует у круглых червей. У более развитых животных, начиная от кольчатых червей, появляется вторичная полость тела, которая является более прогрессивной. Она поделена перегородками, полостная жидкость есть только у кольчецов, у более высокоорганизованных животных отсутствует. Такая полость, в которой пищеварительная, выделительная, нервная, кровеносная системы и внутренние стенки тела не омываются полостной жидкостью и отделены от нее стенками, состоящими из одного слоя эпителиальных клеток, называется вторичной полостью тела.
У всех хордовых полость тела вторичная. В отличие от кольчатых червей вторичная полость тела хордовых не содержит полостной жидкости, и внутренние органы свободно располагаются в полости. |
Способы передвижения животных. Картинка: (На выбор) Вариант1. Хорошо было бы , если можно вставить динамическую картинку, показывающую движение животных, например бег слонов, прыжки кенгуру, плавание рыб. Вариант 2. Картинки, показывающие бегущих, прыгающих, плавающих, летающих животных.
Далее всплывающий текст и к нему картинка:
Основные типы движения: Амебовидное движение. Картинка: амеба обыкновенная, раковинные амебы, радиолярии, фораминиферы.
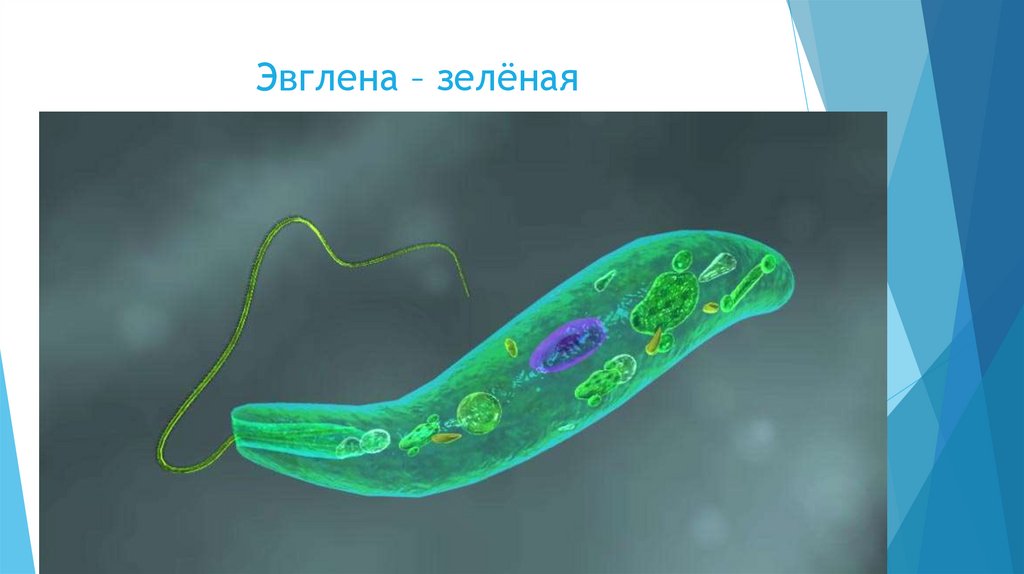
Движения при помощи жгутиков и ресничек. Картинка: эвглена зеленая инфузория сувойка лямблия лейшмания
Движение с помощью мышц. Мышечная ткань
Гладкая Поперечно-полосатая
Картинка: Рисунок гладкой и поперечно-полосатой мышечной ткани. Аскарида и острица Картинка: аскарида. острица Мышцы дождевого червя. Картинка: поперечный срез через тело дождевого червя.
Реактивное движение Картинка: осьминог, кальмар, каракатиц, медуза
Схема внутреннего строения кальмара. Картинка: Схема внутреннего строения кальмара
Мускулатура рыбы. Картинка: Схема мускулатуры рыбы. Строение мышцы. Картинка: Схема строения мышцы Схема строения плавников рыбы
Пауза 10 секунд: в это время учащиеся самостоятельно читают текст на экране. Скорость плавания рыб. Картинка: рыба-парусник рыба-парусник 95 км в час, Картинка: тунец тунец— 64 км в час Картинка: форель форель — 24 км в час Картинка: меч-рыба меч-рыба способна разгоняться до 130 км в час.
Мускулатура лягушки. Картинка: Мускулатура лягушки.
Мускулатура пресмыкающихся. Картинка: Ящерица, варан. Крокодил в воде и на суше. Картинка Крокодил в воде Крокодил на суше
Картинка: кобра, эфа, удав. Хорошо, если можно вставить динамическую картинку: волнообразно ползущая змея.
Рекордсменка ползания черная мамба. Картинка: черная мамба.
Схема мускулатура птиц.
Картинка Схема грудной мышцы птицы Диафрагма — это куполообразная мышца, отграничивающая брюшную полость от грудной полости. Картинка Схема, показывающая расположение диафрагмы. Способы передвижения птиц.
Летают ходят и бегают плавают и ныряют
Картинка: Лазание поползня, полет ласточки, бег страуса ныряние пингвина, плавание гусей и уток. птенец гоацина на ветке.
Картинка: строение задних конечностей птиц.
Пауза 10 секунд: в это время учащиеся самостоятельно читают текст на экране. Наиболее быстро летают стрижи. Скорость полета 300 км/ч. Воробьи ,синицы летают со скоростью 30—60 км/ч, аисты, цапли — 30 км/ч., кулики, голуби, 65 км/ч. утки, гуси, соколы 95 км/ч.
Картинка: гепард Самые быстрые животные – гепарды. Картинка: кенгуру Кенгуру- рекордсмен по прыжкам в длину.
Каждый термин появляется на экране согласованно с чтением текста. Полость тела
Полость тела – пространство, расположенное между стенками тела и внутренними органами. Полостная жидкость – жидкость, которая находится в первичной полости тела и омывает внутренние органы. Первичная полость тела – пространство между стенкой тела и кишечником, в котором расположены внутренние органы, не имеющее собственной оболочки. Вторичная полость тела – пространство между стенкой тела и внутренними органами; ограничено собственными эпителиальными оболочками и заполнено жидкостью.
Картинка: Схема горизонтальный срез Первичная полость Картинка: Схема горизонтальный срез Дождевого червя
Картинка: грудная и брюшная полость собаки |
 При медленном перемещении кальмар пользуется большим ромбовидным плавником, периодически изгибающимся. Для быстрого броска он использует реактивный двигатель. Мышечная ткань – мантия окружает тело моллюска со всех сторон, объем ее полости составляет почти половину объема тела кальмара. Животное засасывает воду внутрь мантийной полости, а затем резко выбрасывает струю воды через узкое сопло. Это сопло снабжено специальным клапаном, и мышцы могут его поворачивать, изменяя направление движения. Двигатель кальмара очень экономичен, он способен развивать скорость до 60 – 70 км/ч. Недаром кальмара называют “живой торпедой”.
При медленном перемещении кальмар пользуется большим ромбовидным плавником, периодически изгибающимся. Для быстрого броска он использует реактивный двигатель. Мышечная ткань – мантия окружает тело моллюска со всех сторон, объем ее полости составляет почти половину объема тела кальмара. Животное засасывает воду внутрь мантийной полости, а затем резко выбрасывает струю воды через узкое сопло. Это сопло снабжено специальным клапаном, и мышцы могут его поворачивать, изменяя направление движения. Двигатель кальмара очень экономичен, он способен развивать скорость до 60 – 70 км/ч. Недаром кальмара называют “живой торпедой”.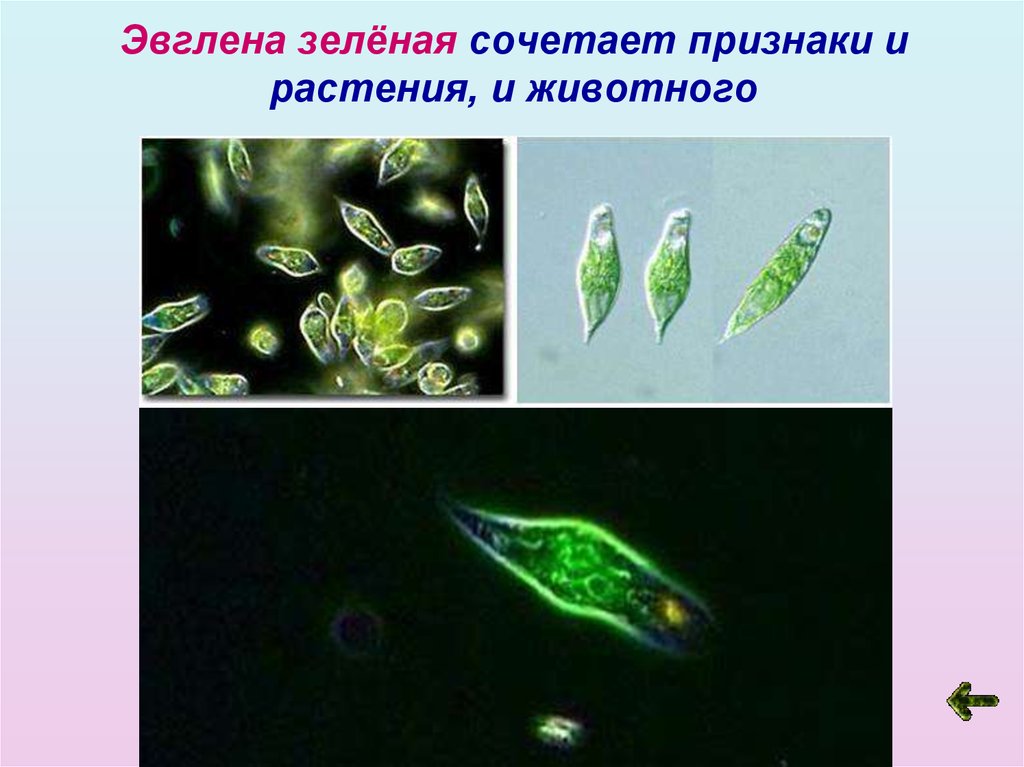 Число сегментов соответствует числу позвонков. Когда в каком-нибудь сегменте сокращаются соответствующие мышечные волокна, то они тянут позвонки в свою сторону, и позвоночник изгибается; если же сокращаются мышцы на противоположном боку, то и позвоночник изгибается в другую сторону. Таким образом, и скелет рыбы, и одевающая его мускулатура состоят из повторяющихся однородных между собой частей — позвонков и мышечных сегментов. Мышцы обеспечивают движения плавников, челюстей и жаберных крышек. В связи с плаванием наиболее развиты мышцы спины и хвоста.
Число сегментов соответствует числу позвонков. Когда в каком-нибудь сегменте сокращаются соответствующие мышечные волокна, то они тянут позвонки в свою сторону, и позвоночник изгибается; если же сокращаются мышцы на противоположном боку, то и позвоночник изгибается в другую сторону. Таким образом, и скелет рыбы, и одевающая его мускулатура состоят из повторяющихся однородных между собой частей — позвонков и мышечных сегментов. Мышцы обеспечивают движения плавников, челюстей и жаберных крышек. В связи с плаванием наиболее развиты мышцы спины и хвоста.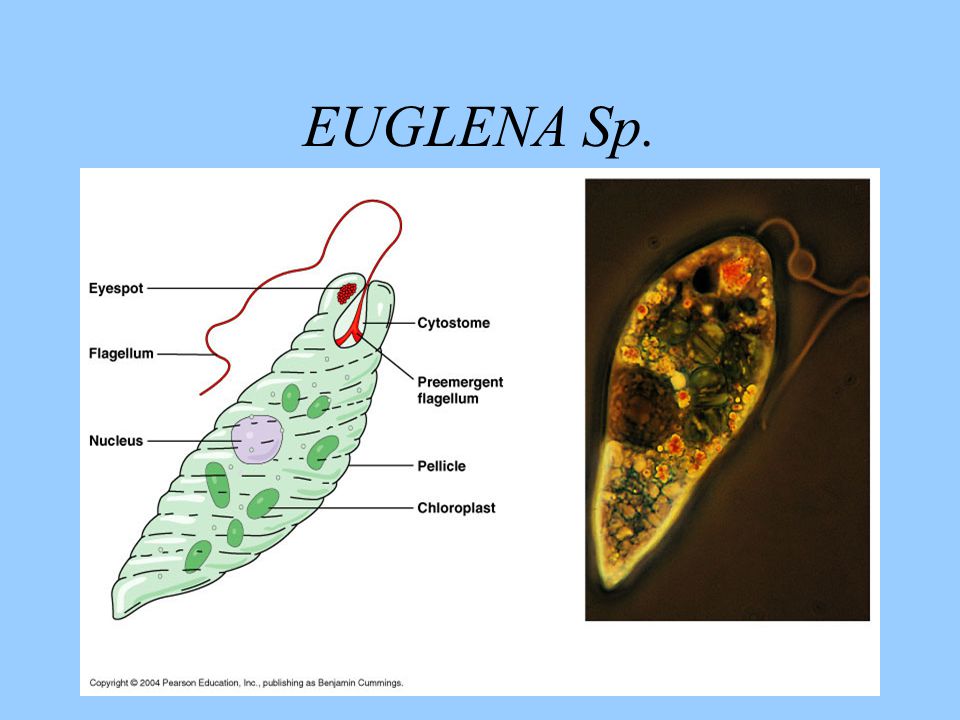 У лягушки, например, более 350 мускулов. Наиболее крупные и мощные из них связаны со свободными конечностями.
У лягушки, например, более 350 мускулов. Наиболее крупные и мощные из них связаны со свободными конечностями.



 Вторичная полость разделена собственными эпителиальными оболочками, благодаря чему тело разделяется на сегменты. Развиваются дыхательная, кровеносная и другие системы органов, то есть у организмов наблюдается дифференциация и специализация систем органов и тканей..
Вторичная полость разделена собственными эпителиальными оболочками, благодаря чему тело разделяется на сегменты. Развиваются дыхательная, кровеносная и другие системы органов, то есть у организмов наблюдается дифференциация и специализация систем органов и тканей..






Открыт хищный родственник красных водорослей • Сергей Ястребов • Новости науки на «Элементах» • Микробиология, Альгология, Систематика, Эволюция
Не дело философа определять, как далеко простирается царство растений и где начинается класс животных или где начинается жизнь.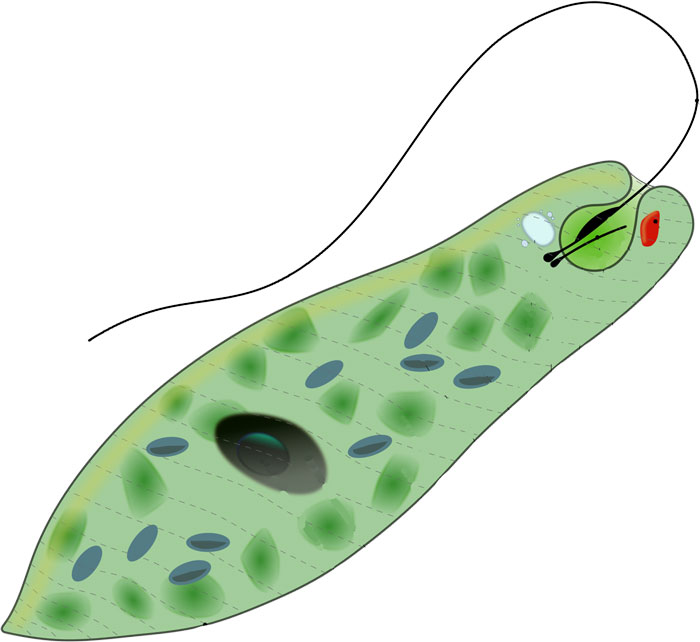 Это проблемы, относящиеся к отдельным наукам. Скорее всего, до самого конца мира мы так и не узнаем, где в точности проходят эти границы.
Это проблемы, относящиеся к отдельным наукам. Скорее всего, до самого конца мира мы так и не узнаем, где в точности проходят эти границы.
Стефан Свежавски, «Святой Фома, прочитанный заново»
Описан новый род одноклеточных организмов, которые являются ближайшими родственниками красных водорослей, но совершенно не похожи на них по образу жизни: это хищные жгутиконосцы, успевшие вторично утратить способность к фотосинтезу. По всей вероятности, это означает, что общие предки красных водорослей и зеленых растений были одноклеточными хищниками, которые даже после приобретения хлоропластов долго сохраняли смешанный тип питания (хищничество + фотосинтез). Жгутиконосец Rhodelphis — это дожившее до наших дней одноклеточное хищное растение.
30 мая 2019 года безвременно, в возрасте всего лишь 66 лет, умер известный российский биолог Александр Петрович Мыльников. Доктор наук, сотрудник Института биологии внутренних вод имени И. Д. Папанина (ИБВВ РАН), он был одним из крупнейших современных протистологов. Мыльников прославился как мастер электронной микроскопии и сделал много открытий, серьезно прояснивших наши представления об эволюции одноклеточных организмов. Установленные им факты уже давно вошли в университетские курсы. Мыльников был очень активным исследователем, признанным во всем мире (см. его страницу на Researchgate). Последняя работа с его участием вышла в журнале Nature уже посмертно. В числе ее авторов есть и другие известные протистологи, например сотрудник ИБВВ Денис Викторович Тихоненков и канадский профессор Патрик Килинг (Patrick Jonhn Keeling). Эта работа посвящена описанию удивительного одноклеточного организма, устройство которого позволяет нам узнать кое-что новое об истоках эволюции растений (рис. 1).
Мыльников прославился как мастер электронной микроскопии и сделал много открытий, серьезно прояснивших наши представления об эволюции одноклеточных организмов. Установленные им факты уже давно вошли в университетские курсы. Мыльников был очень активным исследователем, признанным во всем мире (см. его страницу на Researchgate). Последняя работа с его участием вышла в журнале Nature уже посмертно. В числе ее авторов есть и другие известные протистологи, например сотрудник ИБВВ Денис Викторович Тихоненков и канадский профессор Патрик Килинг (Patrick Jonhn Keeling). Эта работа посвящена описанию удивительного одноклеточного организма, устройство которого позволяет нам узнать кое-что новое об истоках эволюции растений (рис. 1).
Клетки в клетках
В последние два с лишним миллиарда лет облик жизни на Земле определяют в основном эукариоты, обладатели сложных клеток с ядрами. К ним относятся животные, растения, грибы и многочисленные одноклеточные существа, которых собирательно называют протистами. Отличие сложной эукариотной клетки от простой прокариотной огромно: не вдаваясь сейчас в детали, вполне можно сказать, что это принципиально разные уровни организации живой материи. Понятно, что проблема становления эукариот очень важна для эволюционной биологии.
Отличие сложной эукариотной клетки от простой прокариотной огромно: не вдаваясь сейчас в детали, вполне можно сказать, что это принципиально разные уровни организации живой материи. Понятно, что проблема становления эукариот очень важна для эволюционной биологии.
Основой для ответа на вопрос «откуда взялись эукариоты?» является симбиогенетическая теория, согласно которой эукариотная клетка возникла в результате взаимовыгодного объединения (симбиоза) двух или нескольких прокариот. Принципиально важно, что при этом происходило вселение одних клеток в другие (эндосимбиоз). Эту теорию нельзя назвать новой: она сформировалась на рубеже XIX–XX веков, после долгого периода забвения возродилась в 1960-х благодаря Линн Маргулис (Lynn Margulis), а на русском языке её современную версию впервые изложил еще в 1973 году Армен Леонович Тахтаджян (А. Л. Тахтаджян, 1973. Четыре царства органического мира (DjVu, 6 Мб)). Молекулярная биология подтверждает симбиогенетическую теорию настолько однозначно, что споры о её верности или неверности давно прекратились. Иное дело — как конкретно всё это происходило. Поток данных, добываемых современными биологами, порождает множество вопросов на эту тему, и исследователи планомерно ищут ответы на них.
Иное дело — как конкретно всё это происходило. Поток данных, добываемых современными биологами, порождает множество вопросов на эту тему, и исследователи планомерно ищут ответы на них.
У эукариот есть два типа внутриклеточных структур (органелл), которые точно имеют симбиотическое происхождение, то есть являются бывшими бактериями. Это митохондрии, обеспечивающие дыхание, и хлоропласты, обеспечивающие фотосинтез. В наши дни никто из серьезных биологов не сомневается, что эти органеллы возникли эндосимбиотически. Симбиоз с предками митохондрий начался очень давно, на заре становления эукариот. Есть все основания считать, что у общего предка всех современных эукариот митохондрии уже были (см. Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.11.2017). Сейчас на наших глазах идут оживленные дискуссии о том, в какой именно момент и по какому сценарию предок эукариот, который сам был не бактерией, а археей, вступил в симбиоз с предком митохондрий (см.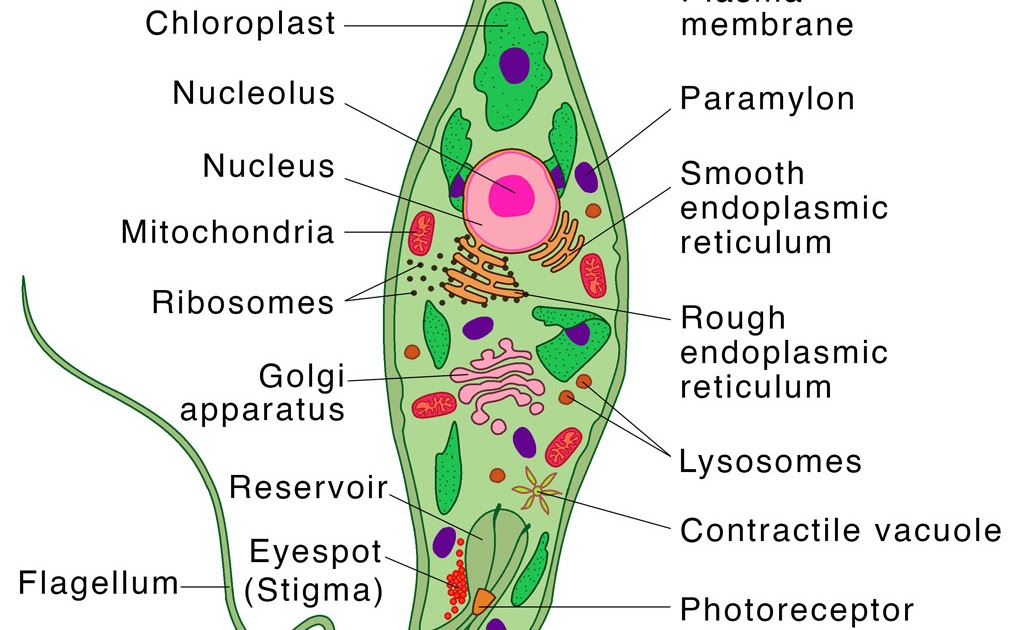 Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016). Но в любом случае это было до того, как сложились «нормальные» эукариотные клетки, знакомые нам из современной живой природы. В общем, союз с митохондриями по большому счету относится скорее к предыстории эукариот.
Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016). Но в любом случае это было до того, как сложились «нормальные» эукариотные клетки, знакомые нам из современной живой природы. В общем, союз с митохондриями по большому счету относится скорее к предыстории эукариот.
С хлоропластами дело обстоит совсем иначе. Во-первых, они есть далеко не у всех эукариот: многие современные группы (в том числе и та, к которой относится человек) не имели их никогда. Во-вторых, в отличие от приобретения митохондрий, которое, по-видимому, было уникальным и неповторимым событием, приобретение хлоропластов совершенно точно происходило много раз. Хлоропласты имеют богатейшую историю, насыщенную неожиданными явлениями (см., например: P. J. Keeling, 2013. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution; рис. 2). Тому, кто захочет разобраться в ней детально, можно посоветовать сначала взглянуть на современную версию эволюционного древа эукариот (её беглое описание можно найти, например, в статье: Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06. 02.2019). Здесь же достаточно сказать, что это древо включает в себя от 6 до 10 (в зависимости от способа подсчета) крупных ветвей, ни одна из которых — увы — не соответствует традиционным царствам животных, растений или грибов. Хорошо это или плохо, но система живой природы, знакомая нам из учебников XX века, ушла в прошлое безвозвратно. Крупные эволюционные ветви эукариот, приблизительно соответствующие по значению классическим царствам, сейчас принято называть супергруппами (supergroup).
02.2019). Здесь же достаточно сказать, что это древо включает в себя от 6 до 10 (в зависимости от способа подсчета) крупных ветвей, ни одна из которых — увы — не соответствует традиционным царствам животных, растений или грибов. Хорошо это или плохо, но система живой природы, знакомая нам из учебников XX века, ушла в прошлое безвозвратно. Крупные эволюционные ветви эукариот, приблизительно соответствующие по значению классическим царствам, сейчас принято называть супергруппами (supergroup).
Краткая история хлоропластов
Итак, предками всех хлоропластов в конечном счете являются цианобактерии, прокариотные организмы, более известные как синезеленые водоросли. Наша история началась с того, что некий жгутиконосец поглотил цианобактерию, которая превратилась в хлоропласт. Потомки этого жгутиконосца разделились на три эволюционные ветви: микроскопические глаукофитовые водоросли, красные водоросли и зеленые растения (см. рис. 3). Все вместе они образуют супергруппу Archaeplastida. Именно с возникновения эволюционного ствола архепластид началась история фотосинтезирующих эукариот. Другое название этой супергруппы — Plantae, что значит попросту «растения». Здесь и далее мы будем использовать названия «архепластиды» и «растения» как синонимы.
Именно с возникновения эволюционного ствола архепластид началась история фотосинтезирующих эукариот. Другое название этой супергруппы — Plantae, что значит попросту «растения». Здесь и далее мы будем использовать названия «архепластиды» и «растения» как синонимы.
Цианобактерии относятся к грамотрицательным бактериям, главная особенность которых состоит в уникальном строении клеточной оболочки. Любая грамотрицательная бактерия имеет две настоящие клеточные мембраны — внутреннюю и наружную. Между ними находится тонкая клеточная стенка, состоящая, как и у всех бактерий, из пептидогликана. Получается эдакий сэндвич. Когда цианобактерия становится хлоропластом, обе ее мембраны остаются на своем месте. Поэтому хлоропласты глаукофитовых водорослей, красных водорослей и зеленых растений всегда имеют две мембраны (у глаукофит между ними сохраняются и остатки пептидогликановой клеточной стенки). Такие хлоропласты называются первичными. Самые заметные их обладатели — зеленые водоросли и особенно высшие растения, создавшие растительный покров земной суши.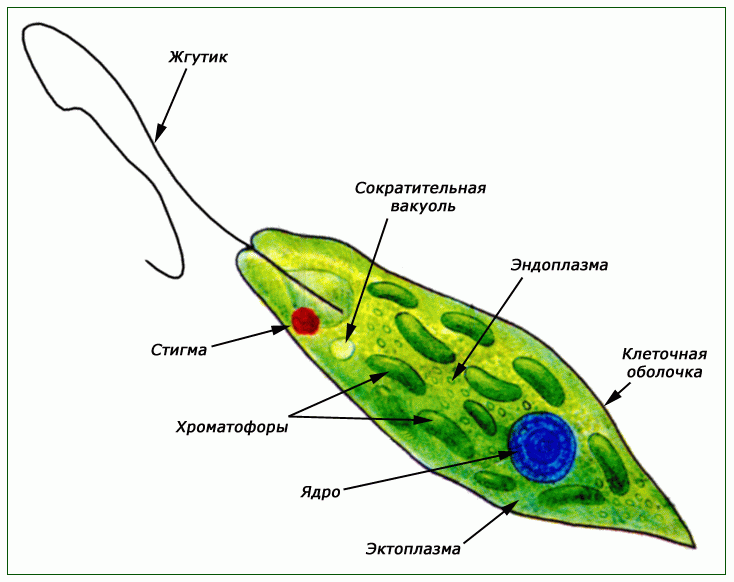
Но это — только часть разнообразия хлоропластов. Другая часть, причем с некоторой точки зрения гораздо более значительная, образуется вот как: хищный эукариот поглощает другого эукариота, внутри которого уже есть первичный хлоропласт, и заставляет этого эукариота в свою очередь работать хлоропластом. Такие хлоропласты называются вторичными. В одних случаях они возникли из зеленых водорослей, в других — из красных. Типичный вторичный хлоропласт имеет четыре мембраны: к двум мембранам цианобактерии добавляется внешняя мембрана первого хозяина (зеленой или красной водоросли) и мембрана пищеварительной вакуоли второго хозяина (хищника, захватившего эту водоросль). Как правило, обладатели вторичных хлоропластов рано или поздно теряют хищничество и обращаются сами в истинные водоросли, то есть в организмы, питающиеся только путем фотосинтеза.
Зеленые водоросли стали вторичными хлоропластами у эвгленовых жгутиконосцев (супергруппа Discoba) и у микроскопических морских хлорарахниофитовых водорослей, которые часто имеют своеобразный облик фотосинтезирующих амеб (супергруппа Rhizaria). Во времена, когда это не было известно, такой характер эволюции изрядно мешал исследователям разобраться в родственных связях эукариот. Например, ботаников постоянно сбивал с толку тот факт, что эвглена зелёная по структуре хлоропластов очень близка к зеленым водорослям, но при этом ни по каким другим признакам ничего общего с зелеными водорослями у нее нет! Разгадка состоит в том, что зеленые водоросли родственны не эвглене — она относится к совершенно другой супергруппе, — а ее хлоропластам.
Во времена, когда это не было известно, такой характер эволюции изрядно мешал исследователям разобраться в родственных связях эукариот. Например, ботаников постоянно сбивал с толку тот факт, что эвглена зелёная по структуре хлоропластов очень близка к зеленым водорослям, но при этом ни по каким другим признакам ничего общего с зелеными водорослями у нее нет! Разгадка состоит в том, что зеленые водоросли родственны не эвглене — она относится к совершенно другой супергруппе, — а ее хлоропластам.
Судьба тех вторичных хлоропластов, которые образовались из красных водорослей, была еще сложнее. В числе их обладателей — бурые, золотистые, желтозеленые и диатомовые водоросли, входящие в супергруппу Stramenopiles, а также еще три группы преимущественно одноклеточных водорослей: динофлагелляты (супергруппа Alveolata), криптофиты (супергруппа Cryptista) и гаптофиты (супергруппа Haptista; тут надо отметить, что родственные связи криптофит и гаптофит до сих пор неясны, и их отнесение к самостоятельным супергруппам может оказаться временным). Ситуация дополнительно усложняется тем, что многие обладатели вторичных хлоропластов потом их потеряли. Например, в супергруппе Alveolata есть две большие группы, потеря хлоропластов у которых подтверждена электронной микроскопией и молекулярной генетикой, причем обе они фигурируют в школьных учебниках зоологии. Это паразитические споровики, к которым относится малярийный плазмодий, и инфузории, к которым относится всем известная туфелька. Строго говоря, как минимум у споровиков вторичный хлоропласт, происходящий из красной водоросли, даже и не исчез: он сохранился в виде бесцветной структуры, называемой апикопластом, которая утратила способность к фотосинтезу, но продолжает выполнять кое-какие другие биохимические функции.
Ситуация дополнительно усложняется тем, что многие обладатели вторичных хлоропластов потом их потеряли. Например, в супергруппе Alveolata есть две большие группы, потеря хлоропластов у которых подтверждена электронной микроскопией и молекулярной генетикой, причем обе они фигурируют в школьных учебниках зоологии. Это паразитические споровики, к которым относится малярийный плазмодий, и инфузории, к которым относится всем известная туфелька. Строго говоря, как минимум у споровиков вторичный хлоропласт, происходящий из красной водоросли, даже и не исчез: он сохранился в виде бесцветной структуры, называемой апикопластом, которая утратила способность к фотосинтезу, но продолжает выполнять кое-какие другие биохимические функции.
В ходе эволюции устройство вторичных хлоропластов может упрощаться. Например, у эвглен и у панцирных водорослей динофлагеллят они имеют уже не четыре мембраны, а три (исчезает, скорее всего, та мембрана, которая соответствует внешней мембране зеленой или красной водоросли — она наименее физиологически необходима).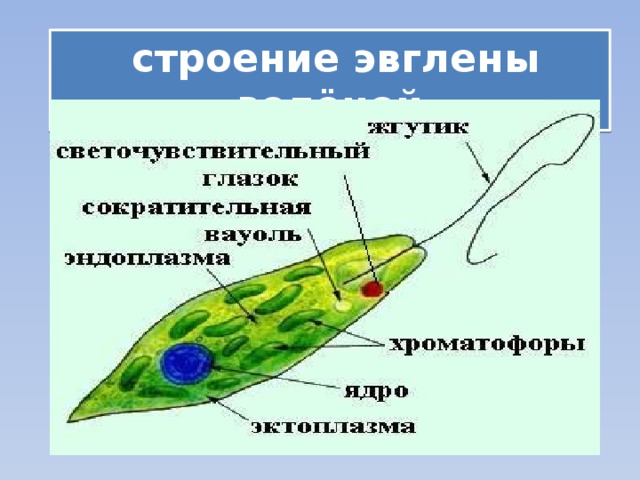 У криптомонад и у хлорарахниофит между второй и третьей мембранами хлоропласта сохраняется нуклеоморф — маленькое, но функционирующее ядро красной (у криптомонад) или зеленой (у хлорарахниофит) поглощенной водоросли. В других группах оно успело исчезнуть.
У криптомонад и у хлорарахниофит между второй и третьей мембранами хлоропласта сохраняется нуклеоморф — маленькое, но функционирующее ядро красной (у криптомонад) или зеленой (у хлорарахниофит) поглощенной водоросли. В других группах оно успело исчезнуть.
У некоторых эукариот приобретение хлоропластов происходило неоднократно: они теряли их, а потом опять приобретали. Особенно это свойственно почему-то динофлагеллятам. Такого бешеного разнообразия хлоропластов, как у динофлагеллят, нет ни в одной другой группе эукариот. Представим себе одноклеточную динофлагелляту, которая потеряла хлоропласт, вернулась к хищничеству, а потом «приняла решение» завести хлоропласт заново. Как она может это сделать? Очевидный и давно опробованный способ — поглотить другого эукариота, у которого хлоропласт уже есть. В роли этого эукариота может оказаться зеленая, криптофитовая, гаптофитовая или диатомовая водоросль (все эти варианты реально известны). Если поглощена будет зеленая водоросль, то она просто-напросто превратится в новый вторичный хлоропласт. Поглощение криптофитовой, гаптофитовой или диатомовой водоросли по результату ничем не отличается… кроме того, что их собственные хлоропласты — вторичные, образовавшиеся из красных водорослей. Поэтому, если такая водоросль, в свою очередь, становится хлоропластом, то возникает конструкция из четырех последовательно вставленных друг в друга клеток — трех эукариотных и одной прокариотной: динофлагеллята заключает в себе криптофитовую, гаптофитовую либо диатомовую водоросль, внутри которой находится некогда поглощенная красная водоросль, а уж внутри той — бывшая цианобактерия. Такие хлоропласты называются третичными.
Поглощение криптофитовой, гаптофитовой или диатомовой водоросли по результату ничем не отличается… кроме того, что их собственные хлоропласты — вторичные, образовавшиеся из красных водорослей. Поэтому, если такая водоросль, в свою очередь, становится хлоропластом, то возникает конструкция из четырех последовательно вставленных друг в друга клеток — трех эукариотных и одной прокариотной: динофлагеллята заключает в себе криптофитовую, гаптофитовую либо диатомовую водоросль, внутри которой находится некогда поглощенная красная водоросль, а уж внутри той — бывшая цианобактерия. Такие хлоропласты называются третичными.
Как мы видим, большинство случаев приобретения эукариотами хлоропластов связано с поглощением не цианобактерий (как можно было бы ожидать), а других эукариот, внутри которых поглощенные цианобактерии уже есть. Таким образом, в конечном счете все эукариоты обязаны своими хлоропластами общему предку супергруппы Archaeplastida, который некогда «приручил» захваченную цианобактерию и тем самым сделал возможными все описанные эволюционные авантюры. Известно одно-единственное исключение: пресноводная раковинная амеба Paulinella chromatophora, относящаяся к супергруппе Rhizaria, использует в качестве хлоропластов именно захваченных цианобактерий, причем таких же, какими хищные виды тех же паулинелл обычно питаются. Это уникальный пример независимого приобретения первичных хлоропластов. Но это случилось по меркам истории эукариот совсем недавно — от 90 до 140 миллионов лет назад — и никаких масштабных эволюционных последствий пока не имело (L. Delaye et al., 2016. How really ancient is Paulinella chromatophora?).
Известно одно-единственное исключение: пресноводная раковинная амеба Paulinella chromatophora, относящаяся к супергруппе Rhizaria, использует в качестве хлоропластов именно захваченных цианобактерий, причем таких же, какими хищные виды тех же паулинелл обычно питаются. Это уникальный пример независимого приобретения первичных хлоропластов. Но это случилось по меркам истории эукариот совсем недавно — от 90 до 140 миллионов лет назад — и никаких масштабных эволюционных последствий пока не имело (L. Delaye et al., 2016. How really ancient is Paulinella chromatophora?).
За этим исключением, вся живая природа Земли обязана своими хлоропластами архепластидам, то есть растениям в эволюционном, а не экологическом смысле этого слова. Вот почему так важно понять, как именно они их приобрели.
Родельфис и эволюция
О том, как появились самые первые хлоропласты, существуют разные мнения. Например, знаменитый биоинформатик Евгений Викторович Кунин с коллегами в свое время отважились допустить, что в эволюционной линии растений вообще никогда не существовало фагоцитоза, то есть свойственного хищным эукариотам механизма активного поглощения пищевых объектов (N. Yutin et al., 2009. The origins of phagocytosis and eukaryogenesis (PDF, 1 Мб)). Тогда способ приобретения первичных хлоропластов архепластид должен быть принципиально иным, чем способ приобретения всех вторичных хлоропластов (и первичных хлоропластов паулинеллы тоже), поскольку уж в этих-то случаях фагоцитоз определенно был задействован. Действительно, предполагать тут можно всякое: ранние этапы эволюции растений, прямо скажем, очень темны.
Yutin et al., 2009. The origins of phagocytosis and eukaryogenesis (PDF, 1 Мб)). Тогда способ приобретения первичных хлоропластов архепластид должен быть принципиально иным, чем способ приобретения всех вторичных хлоропластов (и первичных хлоропластов паулинеллы тоже), поскольку уж в этих-то случаях фагоцитоз определенно был задействован. Действительно, предполагать тут можно всякое: ранние этапы эволюции растений, прямо скажем, очень темны.
Новое исследование российских, канадских и немецких протистологов бросает луч света на эти таинственные события. Героем статьи стал хищный жгутиконосец Rhodelphis, один вид которого открыли в небольшом пресном озере в бассейне реки Десна, а другой — в морском песке у берега Южно-Китайского моря. По внешнему облику родельфисы — довольно заурядные бесцветные жгутиконосцы с двумя жгутиками. Однако анализ транскриптомов показал, что на эволюционном древе эукариот родельфисы занимают довольно неожиданное положение, а именно — являются сестринской группой по отношению ко всем красным водорослям. Это означает, что они входят в супергруппу Archaeplastida, и не просто входят, а располагаются на древе этой группы довольно близко к ее общему предку, хотя и не у самого корня.
Это означает, что они входят в супергруппу Archaeplastida, и не просто входят, а располагаются на древе этой группы довольно близко к ее общему предку, хотя и не у самого корня.
Между тем родельфис вовсе не похож ни на какое растение. Он — настоящий хищник. С помощью ложноножек, образующихся в задней части клетки, он запросто поглощает не только бактерий, но и других эукариот. Надо сказать, что для примитивных эукариот такое хищничество в целом довольно обычно — ни по общему облику, ни по образу жизни, ни по способу питания родельфис среди них особо не выделяется (см., например: Выделена новая супергруппа эукариот, «Элементы», 18.04.2018). Нет оснований сомневаться, что хищничество родельфиса — примитивная черта. А это, в свою очередь, означает, что предки растений все-таки были хищниками, владевшими фагоцитозом. И более того, даже внутри эволюционного ствола Archaeplastida хищничество продержалось еще довольно долго.
Интересной особенностью родельфисов, которую отметили исследователи, оказались трубчатые кристы митохондрий. Что это значит? Митохондрии образовались из альфа-протеобактерий, которые, как и цианобактерии, являются грамотрицательными. Поэтому митохондрия, точно так же как и первичный хлоропласт, имеет две мембраны — наружную и внутреннюю. Кристами называются складки внутренней мембраны, предназначенные для увеличения ее поверхности (этого требует физиологическая функция митохондрии). Самые распространенные варианты формы крист — пластинчатые и трубчатые, причем еще в 1970-х было замечено, что в крупных эволюционных ветвях этот признак очень стабилен. Предлагалось даже разделить большинство эукариот на группы Lamellicristata (с пластинчатыми кристами) и Tubulicristata (с трубчатыми кристами; см. Я. И. Старобогатов, 1986. К вопросу о числе царств эукариотных организмов (DjVu, 1 Мб)). И хотя эта идея столкновения с фактами не выдержала, форма крист митохондрий осталась признаком, важным для крупномасштабной системы. Интересно то, что — как опять же давно известно — у представителей супергруппы Archaeplastida, то есть у растений, кристы, как правило, пластинчатые.
Что это значит? Митохондрии образовались из альфа-протеобактерий, которые, как и цианобактерии, являются грамотрицательными. Поэтому митохондрия, точно так же как и первичный хлоропласт, имеет две мембраны — наружную и внутреннюю. Кристами называются складки внутренней мембраны, предназначенные для увеличения ее поверхности (этого требует физиологическая функция митохондрии). Самые распространенные варианты формы крист — пластинчатые и трубчатые, причем еще в 1970-х было замечено, что в крупных эволюционных ветвях этот признак очень стабилен. Предлагалось даже разделить большинство эукариот на группы Lamellicristata (с пластинчатыми кристами) и Tubulicristata (с трубчатыми кристами; см. Я. И. Старобогатов, 1986. К вопросу о числе царств эукариотных организмов (DjVu, 1 Мб)). И хотя эта идея столкновения с фактами не выдержала, форма крист митохондрий осталась признаком, важным для крупномасштабной системы. Интересно то, что — как опять же давно известно — у представителей супергруппы Archaeplastida, то есть у растений, кристы, как правило, пластинчатые. Трубчатые кристы родельфиса — редкий для этой супергруппы признак, который может оказаться очень древним.
Трубчатые кристы родельфиса — редкий для этой супергруппы признак, который может оказаться очень древним.
Ну а как же с хлоропластами? Они есть у глаукофитовых водорослей, которые, по общепринятой версии, являются древнейшей эволюционной ветвью растений (более древней, чем ветвь красных водорослей и родельфиса). У красных водорослей они тоже есть. Все это означает, что хлоропласты наверняка уже были у ближайшего общего предка всех современных архепластид. Но тогда родельфис тоже должен был бы их получить в наследство. Ну, и где они у него?
Электронная микроскопия не обнаружила у родельфиса хлоропластов (во всяком случае, пока). Но вот молекулярная биология, как оказалось, может к этому кое-что добавить. Хорошо известно, что белки, необходимые для работы хлоропластов, но кодируемые ядерными генами, всегда синтезируются за пределами хлоропласта и только потом транспортируются в него. При этом они обязательно распознаются специальными белковыми комплексами, которые встроены во внешнюю и внутреннюю мембраны хлоропласта (см. TIC/TOC complex). Так вот у родельфиса удалось обнаружить не менее четырех белков, входящих в эти комплексы. Нашлись у него и некоторые другие белки, связанные с хлоропластами — в их числе ферредоксин, обычно участвующий в фотосинтезе. В то же время большинство других белков фотосинтеза у родельфиса не найдено, и клетки его, как уже говорилось бесцветные, а не окрашенные, как у фотосинтезирующих растений. Вывод: у родельфиса, вероятно, всё-таки есть первичный хлоропласт, но только маленький, рудиментарный и бесцветный. От фотосинтеза эта эволюционная линия успела отказаться.
TIC/TOC complex). Так вот у родельфиса удалось обнаружить не менее четырех белков, входящих в эти комплексы. Нашлись у него и некоторые другие белки, связанные с хлоропластами — в их числе ферредоксин, обычно участвующий в фотосинтезе. В то же время большинство других белков фотосинтеза у родельфиса не найдено, и клетки его, как уже говорилось бесцветные, а не окрашенные, как у фотосинтезирующих растений. Вывод: у родельфиса, вероятно, всё-таки есть первичный хлоропласт, но только маленький, рудиментарный и бесцветный. От фотосинтеза эта эволюционная линия успела отказаться.
Подводя итог, родельфис вполне можно описать как одноклеточное хищное растение. Причем, что самое главное, его хищность — первичная, унаследованная прямо от общего предка всех современных эукариот, а не вторичная, как у растущей в наших болотах росянки или у триффидов из великого романа Уиндема.
Что касается общих предков родельфисов и красных водорослей, то они скорее всего были миксотрофами, то есть организмами со смешанным типом питания. Они совмещали фотосинтез и хищничество. Затем красные водоросли (как и зеленые растения) отказались от хищничества, а родельфис — от фотосинтеза. Ну а общий предок всех архепластид, вероятно, был обыкновенным хищным жгутиконосцем.
Таким образом, последовательность древнейшей эволюции хлоропластов теперь проясняется. Очевидно, все началось с хищного жгутиконосца, который просто захватывал цианобактерий как пищевые объекты. Потом цианобактерии были «приручены» и наступил этап миксотрофного питания, который, судя по положению родельфисов на эволюционном древе, был достаточно долгим: следуя этой логике, приходится сделать вывод, что даже общий предок красных и зеленых водорослей еще мог быть только миксотрофом. И наконец, в нескольких эволюционных ветвях миксотрофия исчезла. Так возникли растения в наиболее классическом смысле слова — организмы, относящиеся к эволюционной ветви Archaeplastida (= Plantae) и питающиеся только фотосинтезом.
Загадка красных водорослей
Уже упоминалось, что ближайшими современными родственниками родельфисов оказались красные водоросли. Эти организмы давно и хорошо знакомы человеку; многие из них удостоились народных названий, например ирландский мох (Irish moss, Chondrus crispus), в изобилии заселяющий литораль Атлантического океана. Красные водоросли живут почти исключительно в морях, хотя надо отметить, что пресноводные формы среди них все же попадаются: например, известный научно-популярный журнал «Батрахоспермум» получил свое название именно в честь пресноводной красной водоросли. Иногда красные водоросли бывают одноклеточными, но чаще они многоклеточные — нитчатые, пластинчатые или ветвящиеся. Благодаря особым фотосинтетическим пигментам они могут жить глубоко в море, там, где никаким другим водорослям света для фотосинтеза уже не хватает, уходя таким образом от конкуренции. Группа эта очень древняя: самые ранние красные водоросли, обнаруженные палеонтологами, имеют возраст 1,2 миллиарда лет (см. N. J. Butterfield, 2015. Proterozoic photosynthesis — a critical review). Причем они относятся к современному классу бангиевых водорослей, которые, таким образом, являют собой поразительный по глубине пример живых ископаемых (для сравнения — ближайшие ископаемые родичи прославленной латимерии имеют возраст всего-то 70 миллионов лет). По-видимому, это самые древние живые ископаемые среди всех многоклеточных организмов — во всяком случае, эукариотных (вездесущие колониальные цианобактерии не в счет).
У красных водорослей есть одна удивительная особенность, давно ставящая биологов в тупик. Это — полное отсутствие жгутиков. В этой статье уже не раз встречалось слово «жгутиконосцы», которое, собственно, и обозначает эукариот, имеющих жгутики. Когда-то такими были все эукариоты без исключения. Жгутики — это универсальные двигательные структуры, унаследованные современными эукариотами от их одноклеточного общего предка. Они часто сохраняются и после перехода к многоклеточности: например, у большинства многоклеточных животных, включая человека, с помощью жгутиков движутся сперматозоиды. Случаи отказа от жгутиков не очень многочисленны и связаны, как правило, с наземным образом жизни. Например, жгутики совершенно утрачены у высших грибов и у цветковых растений. Этим организмам они просто не нужны: жизненные циклы высших грибов и цветковых растений рассчитаны на полностью сухопутное существование, поэтому размножение у них давным-давно идет без участия активно плавающих половых клеток. Но вот многоклеточные животные такой высокой степени адаптации к наземности не достигли. А красные водоросли — это вообще чисто водные существа. Никаких видимых причин для отказа от жгутиков у них нет. Однако ни у одной известной красной водоросли жгутики не появляются ни на какой стадии жизненного цикла. Это очень странно, тем более что красные водоросли испытывают по этой причине явные трудности: например, из-за того, что мужские половые клетки у них неподвижны, им приходится «изобретать» специальные сложные структуры для оплодотворения, хотя со жгутиковыми половыми клетками всё было бы намного проще.
Лет тридцать назад было популярно мнение, что красные водоросли — это чрезвычайно примитивные эукариоты, у которых жгутики еще просто не успели появиться. Увы, современная филогенетика отвергает эту гипотезу напрочь. Здесь надо искать какое-то другое объяснение.
В 2015 году вышла очень интересная работа, указывающая путь к решению этой проблемы (H. Qiu et al., 2015. Evidence of ancient genome reduction in red algae (Rhodophyta)). Она посвящена геномам красных водорослей. Прежде всего, оказалось, что эти геномы по меркам эукариот маленькие — всего 5–10 тысяч генов. Причем это относится не только к одноклеточным, но и к достаточно крупным и сложным многоклеточным красным водорослям, включая тот же ирландский мох. Детальный анализ показал, что многие гены, свойственные почти всем эукариотам, у красных водорослей почему-то потеряны. В частности, у них отсутствует целый набор генов, обеспечивающих сборку жгутиков и работу центриолей (опорных структур, которые всегда располагаются в основаниях жгутиков). Красные водоросли не образуют жгутиков не потому, что им это не пригодилось бы, а потому, что они на это физически неспособны: у них нет генов, кодирующих необходимые белки. Ни жгутик, ни центриоль они сформировать не могут. Кроме того, они потеряли ряд генов, никак не связанных со жгутиками, но имеющих отношение к разным другим клеточным структурам и механизмам. Например, у красных водорослей не оказалось генов, обычно контролирующих синтез некоторых компонентов клеточных мембран (см. гликозилфосфатидилинозитол). Причем известно, что для многих самых разных эукариот — грибы, трипаносома, мышь — потеря этих генов вообще смертельна. За счет утраты множества генов у красных водорослей заметно обеднен метаболизм, то есть обмен веществ; некоторые биохимические пути у них целиком исчезли.
Авторы делают твердый вывод: все современные красные водоросли произошли от «генетически обнищавшего предка» (gene depauperate ancestor). И это «обнищание» в самом деле составляет для них проблему: показано, что для восполнения недостающих элементов метаболизма красные водоросли склонны «рекрутировать» бактериальные гены, проникающие в их клетки путем постоянно идущего в природе горизонтального переноса генов (ГПГ). А вот жгутики восстановить так легко не получается.
Возможно, что именно из-за сужения адаптивных возможностей, вызванного массовой потерей генов, красные водоросли не смогли завоевать сушу (J. Collen, 2015. Win some, lose some: genome evolution in red algae (PDF)). В конце концов, почему бы земным лесам и травам не быть красными? Однако зеленым растениям освоить сушу удалось, а красным водорослям нет. Зато красные водоросли внесли в эволюцию жизни на Земле другой важный вклад: как мы уже знаем, они «подарили» хлоропласты многим другим группам эукариот — бурым водорослям и иже с ними.
У современных морских или пресноводных красных водорослей нет ни малейших причин отказываться ни от жгутиков, ни от многих других полезных вещей. Это означает, что их эволюционный путь был на редкость непрямым. Судя по всему, в начале эволюционной ветви красных водорослей произошла какая-то генетическая катастрофа: за короткое (конечно, по эволюционным меркам) время они, по грубой оценке, потеряли около четверти генов, свойственных зеленым растениям и большинству других эукариот. Почему это произошло — пока никто не знает. Может быть, предки красных водорослей на каком-то эволюционном этапе были паразитами — ведь известно, что паразитизм часто приводит к редукции генома (см. Бубонная чума была уже 3800 лет назад, «Элементы», 15.08.2018). Но тогда совершенно непонятно, почему они сохранили фотосинтез. А может быть, потеря генов была связана с жизнью в каком-то экстремальном местообитания, но в каком — пока трудно даже представить. Интересующая нас «генная катастрофа» должна была произойти не менее полутора миллиардов лет назад (позже разные ветви красных водорослей уже разделились), так что реконструировать ее условия — дело не самое простое.
Открытие родельфиса добавляет к складывающемуся «пазлу» очень важный фрагмент. У родельфиса нет ни малейших признаков той массивной потери генов, которая произошла у красных водорослей. Все те гены, отсутствие которых специфично для красных водорослей, у него оказались на месте. Общий предок красных водорослей и родельфисов был в этом отношении совершенно «нормален». Особенно интересно, что геном родельфиса богат внутригенными некодирующими последовальностями — интронами, в то время как у красных водорослей число интронов резко уменьшено. Дело в том, что интроны сами по себе нефункциональны. Их массовая потеря обычно бывает связана просто с уменьшением общего размера генома, которое, в свою очередь, связано с уменьшением размера клеток (см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Итак, возможно, что одним из факторов постигшей красные водоросли «генной катастрофы» было предельное уменьшение абсолютного размера клеток, случившееся на стадии, когда эти водоросли еще были одноклеточными. Такие примеры в других эволюционных ветвях известны (H. Qiu, 2016. Travel light: reductive genome evolution in free-living eukaryotes (PDF, 1 Мб)). Полной разгадки тайны красных водорослей эти соображения не дают, но ключ к ней — быть может.
Источник: Ryan M. R. Gawryluk, Denis V. Tikhonenkov, Elisabeth Hehenberger, Filip Husnik, Alexander P. Mylnikov and Patrick J. Keeling. Non-photosynthetic predators are sister to red algae // Nature. 2019. Published 17 July 2019. DOI: 10.1038/s41586-019-1398-6
Сергей Ястребов
Euglena Viridis Stok Fotoğraf, Resimler ve Görseller
Görsel
- Görsel
- Fotoğraf
- İllüstrasyon
- Vektörler
- Video
52
euglena viridis stok fotoğraf ve görselini inceleyin veya daha fazla stok fotograf ve görsel keşfetmek için yeni bir arama başlatın.Телефон:
En popüler
tek hücreli organizmaların vektör illustrasyonu. амеба протей paramecium caudatum ve euglena viridis. простейшие — euglena viridis стоковые иллюстрацииTek hücreli organizmaların вектор illustrasyonu. Amoeba proteus…
euglena vektör anatomisi. eğitici illüstrasyon — euglena viridis стоковые иллюстрацииEuglena vektör anatomisi. Eğitici illüstrasyon
Tek hücreli organizmalar seti (protozoa): paramecium caudatum, amoeba proteus, chlamydomonas ve euglena viridis — euglena viridis фондовые иллюстрацииTek hücreli organizmalar seti (protozoa): Paramecium caudatum,…
иллюстрацииСтруктура эвглены
euglena — euglena viridis стоковые фотографии и изображенияEuglena
euglena зеленые простейшие, изолированные на белом фоне — euglena viridis фондовые иллюстрацииEuglena зеленые простейшие, изолированные на белом фоне
boyama sayfası. tek hücreli organizmaların seti (protozoa): paramecium caudatum, amoeba proteus ve euglena viridis — euglena viridis стоковые иллюстрации Boyama sayfası. Наборы организмов (простейшие):…
Tek hücreli organizmalar seti (protozoa): Paramecium caudatum,…
Tek hücreli organizmaların seti (protozoa): paramecium caudatum, amoeba proteus ve euglena viridis — euglena viridis фондовые иллюстрации .. биотоп пруда с микроскопическими одноклеточными организмами: простейшие (paramecium caudatum, амеба протей, хламидомонада, эвглена виридис), зеленые водоросли и бактерии под увеличительным стеклом — эвглена виридис иллюстрацииБиотоп пруда с микроскопическими одноклеточными организмами: простейшие (Par
контур эвглены зеленой простейшей, выделенной на белом фоне — эвглена зеленая иллюстрацииконтур эвглены зеленой простейшей, выделенной на белом фоне
эвглена анатомическая — эвглена виридис фондовая иллюстрация proteus, chlamydomonas, paramecium caudatum öğrenme küçük bilim adamları — euglena viridis стоковые иллюстрацииLaboratuvarda Protozoa Одноклеточная амеба Proteus, Chlamydomonas,
büyüteçli küçük karakter protozoa unicellular amoeba proteus, chlamydomonas ile büyük su bardağının i̇çine bakın — euglena viridis stock illustrations Büyüteçli Küçük Karakter Protozoa Unicellular Amoeba Proteus,. ..
Euglena viridis ile boyama sayfası
euglena viridis yapısı — euglena viridis стоковые иллюстрацииEuglena viridis yapısı
euglena viridis под увеличительным стеклом на белом фоне — euglena viridis стоковые иллюстрации0016 Euglena viridis под увеличительным стеклом, изолированным на белом… Euglena viridis honor. 1830 — Euglena viridis стоковые иллюстрацииEuglena viridis honor. 1830
tek hücreli organizmalar seti (protozoa): paramecium caudatum, amoeba proteus, chlamydomonas ve euglena viridis — euglena viridis стоковые иллюстрацииtek hücreli organizmalar seti (protozoa): Paramecium caudatum,… caudatum, amoeba proteus ve euglena viridis — euglena viridis стоковые иллюстрации
Tek hücreli organizmaların seti (protozoa): Paramecium caudatum,…
микроскопические одноклеточные организмы: простейшие (paramecium caudatum, амеба протей, хламидомонада, эвглена зеленая), зеленые водоросли (хлорелла, спирогира) и бактерии под увеличительным стеклом — euglena viridis stock иллюстрацииМикроскопические одноклеточные организмы: простейшие (Paramecium caudatum,
euglena ‘nın şeması. bu başlıklar ile euglena viridis yapısı — euglena viridis стоковые иллюстрацииEuglena ‘nın şapısı viridis.0003 эвглена ‘нин шемаси. bu başlıklar ile euglena viridis yapısı — эвглена зеленая стоковые иллюстрации
Euglena ‘nın şeması. Bu başlıklar ile Euglena viridis yapısı
euglena viridis yapısı — euglena viridis stock illustrationsEuglena viridis yapısı
eğitim oyunu: çıkartmaları şeklinde hazır bileşenlerden euglena viridis montajı — euglena viridis stock illustrationsEğitim oyunu: çıkartmaları şeklinde hazır bileşenlerden Euglena…
eğitim oyunu : çıkartmaları şeklinde hazır bileşenlerden euglena viridis montajı — euglena viridis стоковые иллюстрацииEğitim oyunu: çıkartmaları şeklinde hazır bileşenlerden Euglena…
başlıkları ile euglena viridis yapısı ile boyama sayfası — euglena viridis stock illustrationsBaşlıkları ile Euglena viridis yapısı ile boyama sayfası
black silhouette of euglena viridis under magnifying glass isolated on white background — euglena viridis стоковые иллюстрации Черный силуэт Euglena viridis под увеличительным стеклом. ..
Черный силуэт микроскопических одноклеточных организмов: простейшие (P
tek hücreli organizmaların seti (protozoa): paramecium caudatum, amoeba proteus ve euglena viridis — euglena viridis иллюстрацииTek hücreli organizmaların seti (protozoa): paramecium 9003,… биология зоология изображение: euglena viridis — euglena viridis стоковые иллюстрации
Старинная биология зоология изображение: Euglena Viridis
euglena viridis — euglena viridis стоковые иллюстрацииEuglena Viridis
euglena viridis под увеличительным стеклом на белом фоне — euglena viridis стоковые иллюстрацииEuglena viridis под увеличительным стеклом на белом фоне…
euglena viridis, vibrionidae, monas termo — euglena viridis стоковые иллюстрацииEuglena Viridis, Vibrionidae, Monas Termo
euglena viridis siyah siluet — euglena viridis стоковые иллюстрацииEuglena viridis siyah siluet
boyama sayfası. tek hücreli organizmalar seti (простейшие): paramecium caudatum, amoeba proteus, chlamydomonas ve euglena viridis — euglena viridis стоковые иллюстрацииБояма сайфаси. Tek hücreli organizmalar seti (protozoa):…
набор микроскопических одноклеточных организмов: простейшие (paramecium caudatum, амеба протей, хламидомонада, эвглена виридис) и зеленые водоросли (хлорелла, спирогира), выделенные на белом фоне — эвглена виридис иллюстрацииSet микроскопических одноклеточных организмов: простейшие (Paramecium…0016 euglena anahat vektör illustrasyon, bilim eğitimli sanat
224 euglena Стоковые фото, картинки и изображения
Protozoa amoeba proteus, paramecium caudatum, euglena viridis, unicellorganisms under the микроскоп, реалистический, иллюстрация для энциклопедии, изолированное изображение на белом фоне00PREMIUM эвглены зеленой. protozoaPREMIUM
Licmophora sp. водоросли, морские и пресноводные диатомеи под микроскопом. род донных, фотосинтетических и эпифитных диатомейPREMIUM
Эвглена изолирована на белом фоне illustrationPREMIUM
Схематическое изображение стадий деления зеленой эвглены. PREMIUM
Хламидомонада — гаплоидный одноклеточный эукариот.PREMIUM
Векторная иллюстрация эвглены. структура и описание меченого микроорганизма. Схема биологических внутренних частей рода одноклеточных жгутиковых эукариот. схема с расположением жгутика и рыльца.PREMIUM
Структура и схема эвгленыPREMIUM
Euglena, одноклеточный микроорганизм PREMIUM
Licmophora sp. водоросли, морские и пресноводные диатомеи под микроскопом. род донных, фотосинтетических и эпифитных диатомейPREMIUM
Эвглена зеленая. векторная иллюстрация микроорганизма. яркая цветная иллюстрация.PREMIUM
Перспектива структуры водорослейPREMIUM
Коллекция делений простейших в виде набора биологических контуров одноклеточных эукариотPREMIUM
Схематическое изображение эвглены грацилисPREMIUM
Эвглена — род одноклеточных жгутиковых эукариот под микроскопом для обучения.PREMIUM
Происхождение инфузорий с ресничками и способы развития, векторная иллюстрация. PREMIUM
Планктон. векторная иллюстрация с небольшим организмом фитопланктона и зоопланктона на тему экологической биологической природы дикой природы. ПРЕМИУМ
Зеленая эвглена. ПРЕМИУМ
Зеленая эвглена. ПРЕМИУМ
. бактерии в лабораторном микроскопе, структура клеток амебы или простейших, биология протистов, микробиологияПРЕМИУМ
Euglena — род одноклеточных жгутиковых эукариот. PREMIUM
Licmophora sp. водоросли, морские и пресноводные диатомеи под микроскопом. род донных, фотосинтезирующих и эпифитных диатомейPREMIUM
Licmophora sp. водоросли, морские и пресноводные диатомеи под микроскопом. род донных, фотосинтетических и эпифитных диатомейPREMIUM
Cladophora sp. водоросли. структураPREMIUM
Euglena — род одноклеточных жгутиковых эукариот.PREMIUM
Набор одноклеточных организмов в каракулях на белом фоне иллюстрацииPREMIUM
Анатомия эвгленыPREMIUM
Licmophora sp. водоросли, морские и пресноводные диатомеи под микроскопом. род бентических, фотосинтетических и эпифитных диатомовых водорослейPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для образования.PREMIUM
Микроскоп с анатомической структурой эвглены на белом фоне иллюстрацииPREMIUM
Планктон Chlamydomonas. небольшое одноклеточное зеленое животное с усиками и жгутиками. flat cartoonPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Зеленая эвглена, простейшие на черном фонеPREMIUM
3 Eugle это род одноклеточных жгутиковых эукариот под микроскопом для изучения. ПРЕМИУМ
Клетки простейших, простейших и амебных микроорганизмов, микроорганизмы-переносчики. одноклеточные клетки амебы и протистов в лабораторном микроскопе, типы простейших эукариотических организмовPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Три типа водорослей: asterionella, anabaena, euglenaPREMIUM
одноклеточные жгутиковые эукариоты под микроскопом для изучения. ПРЕМИУМ
Простейшие бактерии. самые известные и простые бактерии: euglenaPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения.PREMIUM
Эвглена — род одноклеточных жгутиковых эукариот под микроскопом для изучения.PREMIUM
Клетки амёб, простейших и одноклеточных протистов, бактерии-переносчики эвглены или организм paramecium. простейшие микроорганизмы, бактерии деления и бинарные протисты амебы хламидомонады микробиологияПРЕМИУМ
Euglena — род одноклеточных жгутиковых эукариот. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Планктон Chlamydomonas. небольшое одноклеточное зеленое животное с усиками и жгутиками. flat cartoonPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Эвглена в цвете и рисунок на белом фоне. ПРЕМИУМ
Планктон Chlamydomonas. небольшое одноклеточное зеленое животное с усиками и жгутиками. flat cartoonPREMIUM
Набор иллюстраций различных типов одноклеточных организмовPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для образования. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena дополненияPREMIUM
Эвглена изолированная векторная иллюстрация. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для обучения.
Раздражительность в примере инфографической диаграммы эвгленозоа эвглена движется к свету, но избегает прямого солнечного света стимул рецептор импульсная реакция одноклеточный организм биология наука образование вектор ПРЕМИУМ
Ученый мужского пола, работающий на ноутбуке, читающий информацию и изучающий простейшие одноклеточные paramecium caudatumPREMIUM
Амеба и эвгленаPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena эукариоты под микроскопом для изучения.ПРЕМИУМ
Эвглена — род одноклеточных жгутиковых эукариот под микроскопом для образования.ПРЕМИУМ
Эвглена — род одноклеточных жгутиковых эукариот под микроскопом для образования.ПРЕМИУМ
Планктон Chlamydomonas. маленькое одноклеточное зеленое животное с усиками и жгутиками. ПРЕМИУМ
Векторная иллюстрация набора схем одноклеточных: водоросли, амебы, эвглены. paramecium и дрожжиPREMIUM
EuglenaPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Нематоды происходят от эвглены, выделены на белом отростке, векторная иллюстрация. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Простейшие бактерии. самые известные и простые бактерии: эвглена, хламидомонада, амеба и парамецийPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Структура водорослей крупным планомPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Шаблон целевой страницы Amoeba proteus. ученые в лаборатории изучают одноклеточные простейшие, женщина с болями в животеPREMIUM
Микроводоросли. водные организмы и бактерии. зеленый фитопланктон. плоская иллюстрацияPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Концепция экстремального крупного плана бактерий-паразитов амебы. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. и каракули на белом фоне illustrationPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Крошечный персонаж с лупой заглянет внутрь огромного жидкого стакана с простейшими одноклеточными амёбами proteus, chlamydomonasPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом вид для изучения.PREMIUM
Набор одноклеточных организмов, выделенных на белом фоне иллюстрацииPREMIUM
ChlamidomonasPREMIUM
Анатомия клеток протистов с контурной диаграммой структуры микроорганизма эвгленыPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения. PREMIUM
Микроводоросли. водные организмы и бактерии. зеленый фитопланктон. плоская иллюстрацияPREMIUM
Эвглена мидримуши Эвглена персонаж иллюстрация клип артPREMIUM
Микроводоросли. водные организмы и бактерии. зеленый фитопланктон. плоская иллюстрацияPREMIUM
Euglena — род одноклеточных жгутиковых эукариот под микроскопом для изучения.