Предложена первая филогенетическая классификация миксомицетов • Дмитрий Леонтьев • Новости науки на «Элементах» • Систематика, Микология
Слизевики — своеобразные амебоидные протисты, освоившие наземный образ жизни и способные к образованию сложно устроенных плодовых тел. Крупнейшая и наиболее известная группа слизевиков, миксомицеты, длительное время ускользала от внимания филогенетиков. В начале 2000-х годов было установлено, что эти организмы относятся к супергруппе Amoebozoa, в пределах которой их ближайшими родственниками являются диктиостелиевые и протоспорангиевые (они же церациомиксовые) слизевики. Однако внутренняя система миксомицетов по-прежнему опиралась на морфологию плодовых тел. Первые филогенетические исследования пошатнули уверенность в адекватности традиционной классификации, но не предложили ей серьезной альтернативы. В свежем номере журнала Phytotaxa опубликована первая детальная классификация миксомицетов, основанная на молекулярно-филогенетических данных. В ней не только предложен ряд новых порядков и семейств, но и использованы иные морфологические критерии разграничения таксонов, ранее не принимавшиеся во внимание. На примере миксомицетов вновь подтверждается тезис о том, что наиболее яркие и заметные морфологические признаки являются плохими предсказателями родственных связей, поскольку имеют склонность возникать и исчезать в ходе эволюции неоднократно. Поэтому одной из важнейших задач современной систематики является поиск альтернативных морфологических (или, в более широком смысле, фенотипических) критериев для таксонов, основанных на молекулярно-филогенетических данных.
В ней не только предложен ряд новых порядков и семейств, но и использованы иные морфологические критерии разграничения таксонов, ранее не принимавшиеся во внимание. На примере миксомицетов вновь подтверждается тезис о том, что наиболее яркие и заметные морфологические признаки являются плохими предсказателями родственных связей, поскольку имеют склонность возникать и исчезать в ходе эволюции неоднократно. Поэтому одной из важнейших задач современной систематики является поиск альтернативных морфологических (или, в более широком смысле, фенотипических) критериев для таксонов, основанных на молекулярно-филогенетических данных.
Кто такие слизевики
Слизевики (англ. slime molds) — подвижные наземные одноклеточные фаготрофы, формирующие крупные спороносные структуры, плодовые тела. Макроскопические размеры и сухопутный образ жизни ставят их в один ряд с четырьмя другими группами: многоклеточными животными, зелеными растениями, настоящими грибами и суперколониальными цианобактериями (например, Nostoc pruniforme).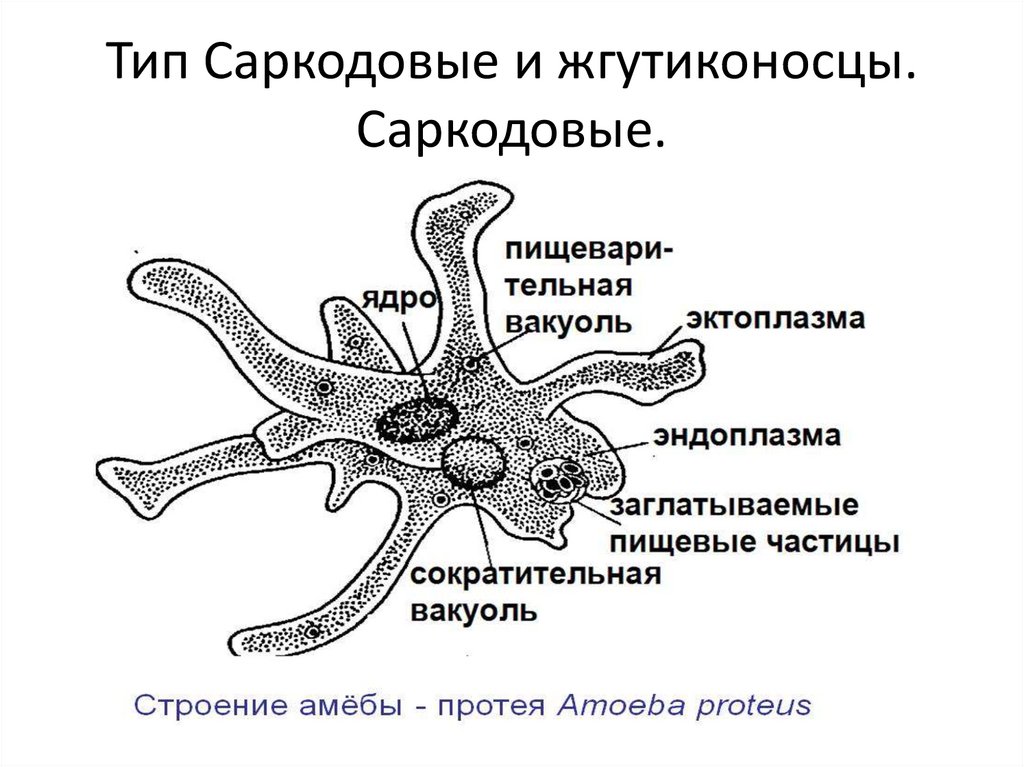 Но среди членов этого «клуба тяжеловесов» слизевики выделяются полным отсутствием истинной многоклеточности, то есть не образуют массу физиологически связанных клеток, происходящих от одной клетки-предшественницы. У них нет ни тканей, ни органов, ни даже их аналогов, как у высших грибов. Слизевик начинает жизнь в виде микроскопической амебы, которая, после ряда драматических преобразований, превращается в россыпь крупных и яркоокрашенных споровместилищ, имеющих различное, но в любом случае не тканевое строение.
Но среди членов этого «клуба тяжеловесов» слизевики выделяются полным отсутствием истинной многоклеточности, то есть не образуют массу физиологически связанных клеток, происходящих от одной клетки-предшественницы. У них нет ни тканей, ни органов, ни даже их аналогов, как у высших грибов. Слизевик начинает жизнь в виде микроскопической амебы, которая, после ряда драматических преобразований, превращается в россыпь крупных и яркоокрашенных споровместилищ, имеющих различное, но в любом случае не тканевое строение.
Ученые и обыватели многие века принимали спороносные структуры слизевиков за плодовые тела грибов. Ситуация изменилась в 1820-е годы, после того, как выдающийся миколог Элиас Магнус Фриз забыл в лесу свою шляпу-цилиндр, в которую он для сохранности положил незрелое плодовое тело слизевика. Вернувшись за шляпой вечером, Фриз обнаружил, что странный гриб… выполз на поля цилиндра, где, наконец, замер и созрел. То, что казалось плодовым телом скрытой в толще земли грибницы, было спороношением гигантского подвижного существа! Когда же Антон де Бари в 1858 году прорастил в лаборатории споры слизевика и обнаружил, что из них выползают крохотные амебы, стало ясно, что слизевики имеют не больше отношения к грибам, чем киты к рыбам.
Плодовые тела слизевиков могут формироваться двумя различными способами. Путь агрегации предполагает, что свободные клетки, напоминающие обычных амеб, собираются в тесные группы, так называемые псевдоплазмодии (рис. 2, а, б). Зрелый псевдоплазмодий ведет себя как единый многоклеточный организм: поддерживает постоянную форму, целенаправленно перемещается по субстрату, и в конце концов, в результате довольно сложного «эмбриогенеза», формирует конструкцию типа «шарик на ножке» — собственно плодовое тело. Псевдоплазмодии и плодовые тела не развиваются из одной клетки-предшественницы, как тела многоклеточных организмов, а являются временными объединениями генетически разнокачественных особей. При этом, в ходе онтогенеза плодовых тел некоторые амебы могут целенаправленно приноситься в жертву, поскольку ножка, состоящая из мертвых клеток, прочнее, чем «живая». Такая стратегия — американский биолог Джеймс Кавендер (James Cavender) в шутку назвал ее «коммунистической» — сближает ее обладателей с истинно многоклеточными организмами. Слизевики, которым свойственна агрегация и псевдоплазмодии, традиционно называют клеточными.
Слизевики, которым свойственна агрегация и псевдоплазмодии, традиционно называют клеточными.
Второй путь развития, плазмодиация, позволяет получить сложно устроенное плодовое тело из одной-единственной микроскопической амебы. Правда, для этого ей приходится стать макроскопической, разрастаясь, но не делясь. В результате образуется плазмодий (рис. 2, в, г), гигантская многоядерная амеба, которая может покрывать площадь до 1 м2, быть до 5 м в длину и весить до 20 кг (у слизевика Brefeldia maxima). Большинство плазмодиев, конечно, не столь огромны, но достаточно крупны, чтобы в Латинской Америке их употребляли в пищу. При этом плазмодий является типичной эукариотической клеткой, имеющей, правда, миллионы ядер, исключительно мощный цитоскелет и сложную систему циркуляции цитоплазмы. Достигнув определенных размеров, плазмодий переходит к формированию плодовых тел. Будучи одноклеточным, он не может пожертвовать частью клеток для построения вспомогательных структур. Поэтому все компоненты плодового тела, кроме спор, образуются из отложений полисахаридов (в основном — β-1,4-галактозаминогликана, см. гликозаминогликаны), которые накапливаются на поверхности плазмодия или внутри цистерн его эндоплазматической сети. У некоторых видов небольшое количество спороподобных клеток может остаться в ножке, но это скорее случайность, чем целенаправленная стратегия. Слизевики, которым свойственна плазмодиация, обычно называют плазмодиальными.
Поэтому все компоненты плодового тела, кроме спор, образуются из отложений полисахаридов (в основном — β-1,4-галактозаминогликана, см. гликозаминогликаны), которые накапливаются на поверхности плазмодия или внутри цистерн его эндоплазматической сети. У некоторых видов небольшое количество спороподобных клеток может остаться в ножке, но это скорее случайность, чем целенаправленная стратегия. Слизевики, которым свойственна плазмодиация, обычно называют плазмодиальными.
Клеточные слизевики бывают очень разными
Как видим, стратегии агрегации и плазмодиации имеют между собой мало общего. Поэтому биологи давно предполагали, что клеточные и плазмодиальные слизевики неродственны друг другу. В 1970-е годы выяснилось еще одно обстоятельство: стратегия клеточных слизевиков возникала в ходе эволюции неоднократно, у самых разных организмов (L. S. Olive, 1970. The Mycetozoa: a revised classification). Сегодня мы знаем, что этот экстравагантный путь развития практикуют минимум шесть неродственных групп. Среди них диктиостелиды и копромиксиды относятся к супергруппе Amoebozoa и состоят в родстве с обыкновенной амебой из школьного учебника, фонтикулиды принадлежат к Opisthokonta и считаются близкой родней настоящих грибов, а акразиды относятся к Discoba и состоят в отдаленном родстве с эвгленами и трипаносомами (см. Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2018). Пятая группа, освоившая агрегацию — это, как ни странно, инфузории (супергруппа Alveolata), правда среди них плодовые тела образует лишь один вид, Sorogena stoianovitchiae (см. S. Sharpe et al., 2015. Timing the origins of multicellular eukaryotes through phylogenomics and relaxed molecular clock analyses). Наконец, агрегация с образованием плодовых тел широко распространена у миксобактерий — прокариотов из отдела Proteobacteria. Подвижные «псевдоплазмодии» этих организмов, так называемые швармы, способны лизировать и поедать других бактерий.
Среди них диктиостелиды и копромиксиды относятся к супергруппе Amoebozoa и состоят в родстве с обыкновенной амебой из школьного учебника, фонтикулиды принадлежат к Opisthokonta и считаются близкой родней настоящих грибов, а акразиды относятся к Discoba и состоят в отдаленном родстве с эвгленами и трипаносомами (см. Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2018). Пятая группа, освоившая агрегацию — это, как ни странно, инфузории (супергруппа Alveolata), правда среди них плодовые тела образует лишь один вид, Sorogena stoianovitchiae (см. S. Sharpe et al., 2015. Timing the origins of multicellular eukaryotes through phylogenomics and relaxed molecular clock analyses). Наконец, агрегация с образованием плодовых тел широко распространена у миксобактерий — прокариотов из отдела Proteobacteria. Подвижные «псевдоплазмодии» этих организмов, так называемые швармы, способны лизировать и поедать других бактерий.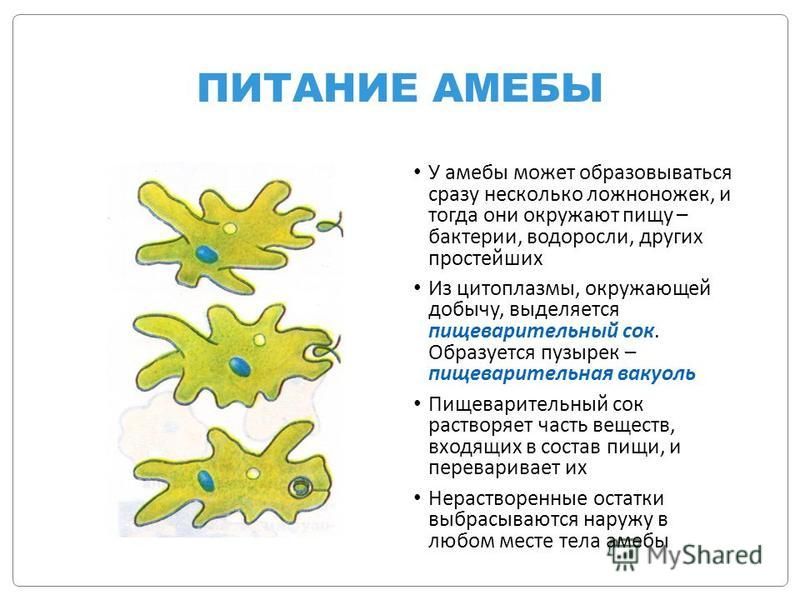 Однако миксобактерии не способны к фагоцитозу, что исключает их из перечня «классических» слизевиков.
Однако миксобактерии не способны к фагоцитозу, что исключает их из перечня «классических» слизевиков.
Из вышесказанного следует важный вывод: путь развития, характерный для клеточных слизевиков — не экзотическая эволюционная случайность, а вполне обычная и успешная стратегия, неоднократно применявшаяся самыми разными организмами. Тут уместно заметить, что мицелиальное строение независимо «изобрели» лишь две таксономические группы (настоящие и ложные грибы), а многоклеточность животного типа, судя по всему, вообще возникла на Земле всего один раз (см., например: С. Ястребов, 2016. Кембрийский взрыв). Таким образом, появление клеточных слизевиков — событие более закономерное, чем появление животных.
Плазмодиальные слизевики: три, две или шесть групп?
А что же плазмодиальные слизевики? Сколько раз в ходе эволюции возникала их жизненная стратегия? До недавнего времени биологи выделяли три группы плазмодиальных слизевиков: миксомицеты, протостелиды и плазмодиофориды. Последнюю группу можно считать слизевиками лишь с большой натяжкой: они являются внутриклеточными паразитами растений и грибов (не вполне «наземный» образ жизни), а их «плодовые тела» представляют собой комочки склеенных спор или просто аморфную споровую массу. Так что в дальнейшем мы будем считать, что истинными плазмодиальными слизевиками являются только миксомицеты и протостелиды.
Последнюю группу можно считать слизевиками лишь с большой натяжкой: они являются внутриклеточными паразитами растений и грибов (не вполне «наземный» образ жизни), а их «плодовые тела» представляют собой комочки склеенных спор или просто аморфную споровую массу. Так что в дальнейшем мы будем считать, что истинными плазмодиальными слизевиками являются только миксомицеты и протостелиды.
Протостелиды имеют очень мелкие плодовые тела, состоящие из 1–8 спор и субтильной неклеточной ножки (рис. 3). По большому счету, это просто цисты амеб, приподнятые над субстратом на стебельках (так у нее больше шансов быть подхваченной ветром). Но в отличие от обычных амеб, один плазмодий простостелид образует множество плодовых тел, от десятков до тысяч. Кроме того, часть цитоплазмы, не попавшая в споры, может быть использована для построения крупных и красивых «подставок», так называемых спороносных рожек, на поверхности которых развиваются многочисленные плодовые тела. Это можно наблюдать у обычной в лесах Евразии церациомиксы кустарничковой —
 3, в).
3, в).Миксомицеты, в отличие от протостелид — существа с очень сложной морфологией. Каждое их плодовое тело содержит от сотен до миллионов спор и включает многочисленные вспомогательные структуры (рис. 4), о которых мы подробнее расскажем ниже. Плазмодии миксомицетов также могут распадаться на зачатки мелких плодовых тел (впрочем, не столь мелких, как у протостелид), но у многих видов фаза распада сокращается или полностью выпадает, и тогда весь плазмодий превращается в одно крупное плодовое тело, от одного до десятков сантиметров в диаметре. Такое плодовое тело можно наблюдать, к примеру, у ретикулярии дождевиковой ( Reticularia lycoperdon).
Свидетельства в пользу родства миксомицетов и протостелид появились уже давно. В первой половине XX века единственную известную на тот момент протостелиду, вышеупомянутую церациомиксу, считали «экзоспоровым миксомицетом», принимая ее многочисленные плодовые тела за споры, образующиеся на поверхности одного плодового тела. В 1967–1975 годах Линдси Олайв и Кармен Стоянович из Университета Северной Каролины описали множество микроскопических протостелид, создали для них класс Protostelia (=Protosteliomycetes) и перенесли в него церациомиксу (L.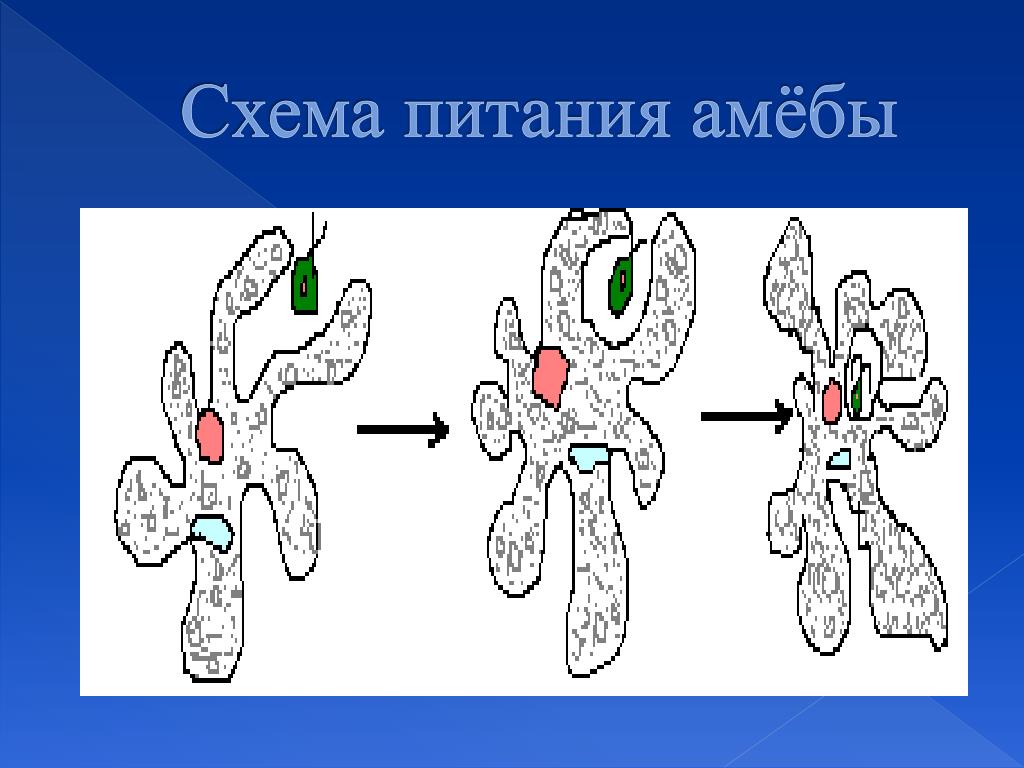 S. Olive, 1970. The Mycetozoa: a revised classification). Вместе с миксомицетами и клеточными слизевиками из класса диктиостелид, протостелиды образовали тип Eumycetozoa, «настоящие слизевики».
S. Olive, 1970. The Mycetozoa: a revised classification). Вместе с миксомицетами и клеточными слизевиками из класса диктиостелид, протостелиды образовали тип Eumycetozoa, «настоящие слизевики».
В 1990 году профессор Арканзасского университета Фредерик Шпигель (Fredrerick W. Spiegel) показал, что два рода протостелид — протоспорангиум (Protosporangium) и кластостелиум (Clastostelium), образующие отряд Protosporangiida, — демонстрируют явное сходство с миксомицетами. Само собой, возникло предположение, что миксомицеты произошли от протостелид (F. W. Spiegel, 1990. Phylum Plasmodial Slime Molds, Class Protostelida).
Все изменилось, когда в 2009 году Лора Шедвик (Lora Shadwick) с соавторами, среди которых был и Фредерик Шпигель, опубликовали результаты молекулярно-филогенетического анализа последовательностей гена рРНК малой субъединицы рибосомы 17 видов протостелид. Выяснилось, что протостелиды вообще не образуют монофилетическую группу, а представляют собой минимум шесть независимых линий, относящихся к разным, зачастую очень удаленным друг от друга ветвям супергруппы Amoebozoa (L.
К середине 2010-х класс протостелид распался, а его представители нашли свое место в системе «обычных» (неплодоносящих) амеб. Однако отряд Protosporangiida, сходство которого с миксомицетами ранее отметил Шпигель, равно как и церациомикса, неожиданно нарушили общее правило, действительно оказавшись близкими родственниками миксомицетов (S. Kang et al., 2017. Between a Pod and a Hard Test: The Deep Evolution of Amoebae). К 2019 году было твердо установлено, что состав группы Eumycetozoa (настоящих слизевиков) ограничен отрядом Protosporangiida (рода протоспорангиум и кластостелиум), церациомиксой, миксомицетами и диктиостелидами.
Классификация миксомицетов: морфология против филогенетики
Тем временем начались молекулярно-филогенетические исследования родственных связей в пределах миксомицетов, самой крупной и сложной группы слизевиков. Прогресса в этой области не наблюдалось с 1960-х годов, когда утвердилась классификация Джорджа Мартина и Константиноса Алексопулоса, делившая «эндоспоровых» (то есть настоящих) миксомицетов на пять порядков: эхиностелиевые, лицеевые, трихиевые, стемонитовые и физаровые (G. W. Martin, C. J. Alexopoulos, 1969. Myxomycetes). Отличия между порядками определялись суммой «значимых» признаков: наличием или отсутствием разрыхляющих нитей, их разветвленностью и орнаментацией, а также присутствием в плодовых телах отложений «извести» (оксалатов, карбонатов и фосфатов кальция).
Впрочем, еще в 1875 году польский миксомицетолог Юзеф Ростафинский предложил иную морфологическую классификацию, разделив миксомицеты всего на две группы: темноспоровые, имеющие темно-бурые или черные споры, и светлоспоровые (так это название обычно переводят с латыни, хотя вернее было бы назвать их «яркоспоровыми») — у них споровая масса может быть практически всех цветов радуги. В этой классификации, правда, не нашлось место роду эхиностелиум (Echinostelium), споры которого практически лишены пигментации: во времена Ростафинского этот мельчайший миксомицет был неизвестен.
Классификация Ростафинского была надолго забыта. Однако в 2005 году группа под руководством Анны Марии Фьоре-Донно (Anna Maria Fiore-Donno; сейчас она работает в Университете Кёльна) опубликовала результаты филогении двух маркерных генов (рРНК малой субъединицы рибосомы и фактора элонгации 1α) для 13 видов миксомицетов. В предложенном авторами филогенетическом дереве стало очевидно четкое разделение миксомицетов на светлоспоровых (к ним относятся порядки Лицевые и Трихиевые) и темноспоровых (Стемонитовые и Физаровые). Бесцветный же эхиностелиум на филограмме 2005 года занял сестринское положение по отношению ко всем остальным миксомицетам, то есть оказался едва ли не членом самостоятельного класса (A. M. Fiore-Donno et al., 2005. Higher-Order Phylogeny of Plasmodial Slime Molds (Myxogastria) Based on Elongation Factor 1-A and Small Subunit rRNA Gene Sequences). Однако, чем больше видов вовлекалось в анализ, тем очевиднее становилось родство эхиностелиума с темноспоровыми миксомицетами. Об этом же свидетельствовали и морфологические данные: из трех родов, в тот момент относившихся к порядку эхиностелиевых, два имеют темные, меланизированные споры, и лишь у собственно эхиностелиума они прозрачные. Кроме того, и для темноспоровых, и для эхиностелиевых характерно наличие колюмеллы — осевой структуры, расположенной в спороносной головке (споротеке) (A.
В предложенном авторами филогенетическом дереве стало очевидно четкое разделение миксомицетов на светлоспоровых (к ним относятся порядки Лицевые и Трихиевые) и темноспоровых (Стемонитовые и Физаровые). Бесцветный же эхиностелиум на филограмме 2005 года занял сестринское положение по отношению ко всем остальным миксомицетам, то есть оказался едва ли не членом самостоятельного класса (A. M. Fiore-Donno et al., 2005. Higher-Order Phylogeny of Plasmodial Slime Molds (Myxogastria) Based on Elongation Factor 1-A and Small Subunit rRNA Gene Sequences). Однако, чем больше видов вовлекалось в анализ, тем очевиднее становилось родство эхиностелиума с темноспоровыми миксомицетами. Об этом же свидетельствовали и морфологические данные: из трех родов, в тот момент относившихся к порядку эхиностелиевых, два имеют темные, меланизированные споры, и лишь у собственно эхиностелиума они прозрачные. Кроме того, и для темноспоровых, и для эхиностелиевых характерно наличие колюмеллы — осевой структуры, расположенной в спороносной головке (споротеке) (A. M. Fiore-Donno et al., 2008. Evolution of dark-spored Myxomycetes). На основании этих данных в 2013 году известный британский таксономист Томас Кавалье-Смит (Thomas Cavalier-Smith) разделил «эндоспоровые» (настоящие) миксомицеты на два надотряда: Lucisporidia (светлоспоровые) и Columellidia (колюмелловые) (T. Cavalier-Smith, 2013. Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa). Правда, использование этим автором зоологической номенклатуры поставило миксомицетологов в затруднительное положение: дело в том, что по давней традиции миксомицеты подчиняются ботанической номенклатуре, о чем прямо сказано в Международном кодексе номенклатуры водорослей, растений и грибов.
M. Fiore-Donno et al., 2008. Evolution of dark-spored Myxomycetes). На основании этих данных в 2013 году известный британский таксономист Томас Кавалье-Смит (Thomas Cavalier-Smith) разделил «эндоспоровые» (настоящие) миксомицеты на два надотряда: Lucisporidia (светлоспоровые) и Columellidia (колюмелловые) (T. Cavalier-Smith, 2013. Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa). Правда, использование этим автором зоологической номенклатуры поставило миксомицетологов в затруднительное положение: дело в том, что по давней традиции миксомицеты подчиняются ботанической номенклатуре, о чем прямо сказано в Международном кодексе номенклатуры водорослей, растений и грибов.
В начале 2010-х годов еще казалось, что традиционные порядки (отряды) миксомицетов объединяют не просто сходные, но также и родственные виды, иными словами представляют собой монофилетические группы. В этом случае, указанные порядки можно было бы сохранить без изменений, просто объединив их в предложенные Кавалье-Смитом надотряды. Однако филогенетические исследования отдельных групп миксомицетов, опубликованные в 2012–2015 годах, опровергли эти ожидания: выяснилось, что все классические порядки, а также многие семейства и рода, нуждаются в коренном пересмотре (A. M. Fiore-Donno et al., 2012. 18S rDNA Phylogeny of Lamproderma and Allied Genera (Stemonitales, Myxomycetes, Amoebozoa); A. M. Fiore-Donno et al., 2013. Two-Gene Phylogeny of Bright-Spored Myxomycetes (Slime Moulds, Superorder Lucisporidia); D. V. Leontyev et al., 2015. A critical revision of the Tubifera ferruginosa complex).
Однако филогенетические исследования отдельных групп миксомицетов, опубликованные в 2012–2015 годах, опровергли эти ожидания: выяснилось, что все классические порядки, а также многие семейства и рода, нуждаются в коренном пересмотре (A. M. Fiore-Donno et al., 2012. 18S rDNA Phylogeny of Lamproderma and Allied Genera (Stemonitales, Myxomycetes, Amoebozoa); A. M. Fiore-Donno et al., 2013. Two-Gene Phylogeny of Bright-Spored Myxomycetes (Slime Moulds, Superorder Lucisporidia); D. V. Leontyev et al., 2015. A critical revision of the Tubifera ferruginosa complex).
В 2019 году в журнале Phytotaxa вышла статья международного коллектива авторов из Украины, Германии, США и России, в которой было представлено первое обобщенное филогенетическое дерево, построенное на основании 139 полных последовательностей гена рРНК малой субъединицы рибосомы миксомицетов из всех описанных семейств и примерно 70% родов. К анализу также были привлечены результаты изучения филогении отдельных групп миксомицетов, опубликованные ранее.
В новой классификации миксомицеты (класс Myxomycetes) относятся к типу/отделу Eumycetozoa. К нему же принадлежат классы диктиостелиевых (Dictyosteliomycetes) и церациомиксовых (Ceratiomyxomycetes). Последний объединяет протостелидных родственников миксомицетов: рода церациомикса, протоспорангиум и кластостелиум (напомним, что остальные протостелиды теперь широко разбросаны по ветвям дерева Amoebozoa).
Класс Миксомицеты разделен на два подкласса, для которых авторы, уважая приоритет Кавалье-Смита, предложили ботанические версии опубликованных им названий: Lucisporomycetidae (светлоспоровые) и Columellomycetidae (колюмелловые).
Что мы узнали об эволюции миксомицетов?Подклассы светлоспоровых и колюмелловых произошли от общего предка в те времена, когда миксомицеты мало отличались от протостелид. Их плодовые тела содержали от двух (как у современного Echinostelium bisporum) до нескольких десятков (E. debaryanum) спор. Масса спор располагалась на тонкой полупрозрачной ножке, которую авторы статьи называют первичной. Ножка этого типа образуется из желеобразной массы полисахаридных молекул, выделяющейся поверхностью молодой спороносной головки.
Их плодовые тела содержали от двух (как у современного Echinostelium bisporum) до нескольких десятков (E. debaryanum) спор. Масса спор располагалась на тонкой полупрозрачной ножке, которую авторы статьи называют первичной. Ножка этого типа образуется из желеобразной массы полисахаридных молекул, выделяющейся поверхностью молодой спороносной головки.
Дальнейшие события эволюции колюмелловых миксомицетов выглядят так. В цистернах ЭПС плазмодия накапливаются полисахариды, которые, затвердевая, превращаются в нити, расположенные между спорами. Совокупность этих нитей называется капиллицием. Эту структуру ждет большое будущее: у более крупных видов капиллиций превратится в систему, обеспечивающую разрыхление и дозированное рассеивание спор.
Далее близкие родственники эхиностелиумов, представители рода барбеелла (Barbeyella) — а вернее, их далекие предки — «изобрели» меланин, кстати, весьма похожий на тот, что имеется у грибов и животных (тут уместно напомнить, что Amoebozoa состоят в отдаленном родстве с Opisthokonta, см.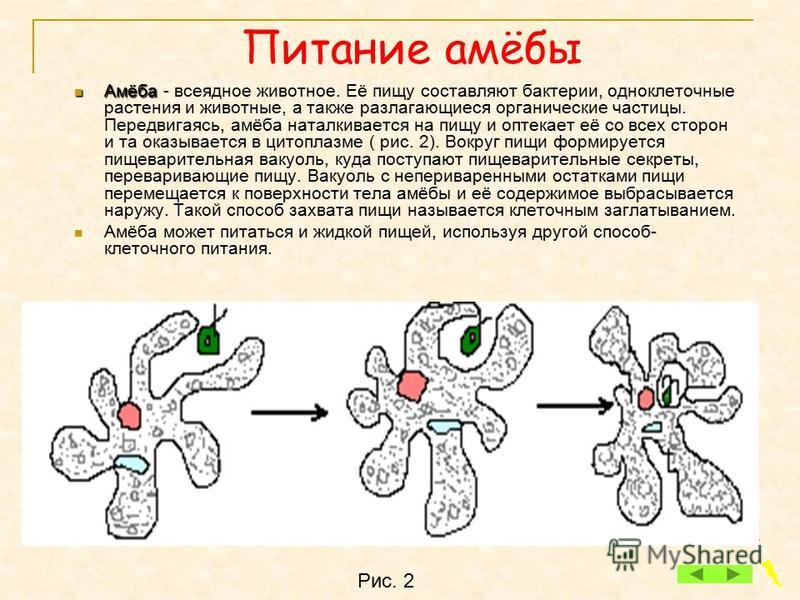 S. M. Adl et al., 2018. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes). С этого момента споры, ножки и капиллиций колюмелловых миксомицетов будут окрашены в темно-бурые или черные тона. Таких миксомицетов, следуя Ростафинскому, можно называть тёмноспоровыми.
S. M. Adl et al., 2018. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes). С этого момента споры, ножки и капиллиций колюмелловых миксомицетов будут окрашены в темно-бурые или черные тона. Таких миксомицетов, следуя Ростафинскому, можно называть тёмноспоровыми.
Еще одним изобретением предков барбееллы стал перидий — защитная белково-полисахаридная пленка, окружающая споровую массу. Появление перидия сняло ограничение на размеры спороносной головки: под защитной оболочкой даже тысячи спор могут спокойно созревать, не рассыпаясь прежде времени. Нити капиллиция у барбееллы своими концами приросли к перидию, а в центральной части головки образовали цилиндрическую ось — колюмеллу, которая внизу сомкнулась с первичной ножкой.
Следующим эволюционным шагом стало отделение от предков эхиностелиевых нового порядка — кластодермовых (таксон предложен авторами обсуждаемой статьи). В пределах этой группы случилось удивительное: колюмелла вышла за пределы головки и образовала верхнюю треть ножки. «Колюмелловая» ножка, образованная внутри вакуолей головки, оказалась намного прочнее и эластичнее первичной. Поэтому у потомков кластодермовых, порядка меридермовых, уже вся ножка является продолжением колюмеллы, а вероятным следствием этого является значительное укрупнение плодовых тел. Интересно, что представителей меридермовых длительное время относили к другой, более молодой группе (стемонитовым). Однако французские миксомицетологи-любители Марьян Мейер и Мишель Пулян обратили внимание, что у этих миксомицетов нити капиллиция сохраняют связь с перидием — этот архаичный признак имелся уже у барбееллы. На основании этого Пулян с соавторами в 2011 году описали род меридерма (Meriderma), а авторы обсуждаемой статьи возвели его, вместе с рядом родственных форм, в самостоятельное семейство и порядок.
«Колюмелловая» ножка, образованная внутри вакуолей головки, оказалась намного прочнее и эластичнее первичной. Поэтому у потомков кластодермовых, порядка меридермовых, уже вся ножка является продолжением колюмеллы, а вероятным следствием этого является значительное укрупнение плодовых тел. Интересно, что представителей меридермовых длительное время относили к другой, более молодой группе (стемонитовым). Однако французские миксомицетологи-любители Марьян Мейер и Мишель Пулян обратили внимание, что у этих миксомицетов нити капиллиция сохраняют связь с перидием — этот архаичный признак имелся уже у барбееллы. На основании этого Пулян с соавторами в 2011 году описали род меридерма (Meriderma), а авторы обсуждаемой статьи возвели его, вместе с рядом родственных форм, в самостоятельное семейство и порядок.
Дальнейшая эволюция колюмелловых связана с образованием двух крупных эволюционных ветвей — порядков стемонитовых и физаровых. Стемонитовые сделали ставку на усложнение капиллиция: у видов рода стемонитис (Stemonitis) он по уровню сложности стал напоминать архитектурные конструкции. Перидий при этом оказался практически не нужен: поверхностная сеть капиллиция надежно удерживала споры вместе до полного высыхания их поверхности. Вторая группа, физаровые, напротив, сохранили и усложнили перидий при постепенной деградации капиллиция. Случилось это так. Ранние физаровые (ими теперь считается семейство лампродермовые) обитали в почвах и плодоносили преимущественно весной, сразу после таянья снега. Для образования плодовых тел их плазмодиям требовалось выбраться на освещенную поверхность земли и, желательно, забраться на кончик листа злака или ветку кустарника. Такая повышенная подвижность потребовала активизации актин-миозинового цитоскелета, для функционирования которого необходимы ионы кальция. Такова одна из возможных причин того, что высшие физаровые (семейства дидимиевых и собственно физаровых) стали накапливать в плодовых телах огромные количества извести. Толщина известковой корки, покрывающей 5–50-сантиметровое плодовое тело фулиго гнилостного (Fuligo septica, рис.
Перидий при этом оказался практически не нужен: поверхностная сеть капиллиция надежно удерживала споры вместе до полного высыхания их поверхности. Вторая группа, физаровые, напротив, сохранили и усложнили перидий при постепенной деградации капиллиция. Случилось это так. Ранние физаровые (ими теперь считается семейство лампродермовые) обитали в почвах и плодоносили преимущественно весной, сразу после таянья снега. Для образования плодовых тел их плазмодиям требовалось выбраться на освещенную поверхность земли и, желательно, забраться на кончик листа злака или ветку кустарника. Такая повышенная подвижность потребовала активизации актин-миозинового цитоскелета, для функционирования которого необходимы ионы кальция. Такова одна из возможных причин того, что высшие физаровые (семейства дидимиевых и собственно физаровых) стали накапливать в плодовых телах огромные количества извести. Толщина известковой корки, покрывающей 5–50-сантиметровое плодовое тело фулиго гнилостного (Fuligo septica, рис. 6), может достигать нескольких миллиметров. Благодаря своей подвижности физаровые стали одними из самых распространенных и разнообразных миксомицетов. Их находят в пустынях, альпийском поясе гор и даже в Антарктиде.
6), может достигать нескольких миллиметров. Благодаря своей подвижности физаровые стали одними из самых распространенных и разнообразных миксомицетов. Их находят в пустынях, альпийском поясе гор и даже в Антарктиде.
Эволюция светлоспоровых миксомицетов началась драматически, путем потери первичной ножки при полном отсутствии колюмеллы, которая могла бы ее заменить. Новую ножку пришлось формировать из перидия, поэтому у всех светлоспоровых миксомицетов она выглядит как складчатый цилиндр, плавно переходящий в покров головки плодового тела. У наиболее ранних представителей группы, объединенных авторами обсуждаемой статьи в порядок крибрариевых, капиллиций полностью отсутствует, так что функцию рассеивания спор выполняет перфорированный перидий. У более молодого порядка ретикуляриевых (также предложенного в рассматриваемой статье) капиллиций появился, причем его источником опять-таки стал перидий: поверхность плодового тела образовала ворончатые вдавления, которые постепенно вытянулись в трубочки капиллиция. Такое строение сохранилось у альвизии Ллойд (Alwisia lloydiae), обитающей в Австралии и на Тасмании. Впрочем, многие ретикуляриевые предпочли утратить ножки и перешли на распространение спор при помощи дождевой воды и насекомых, как, например, ликогала надревная (Lycogala epidendrum).
Такое строение сохранилось у альвизии Ллойд (Alwisia lloydiae), обитающей в Австралии и на Тасмании. Впрочем, многие ретикуляриевые предпочли утратить ножки и перешли на распространение спор при помощи дождевой воды и насекомых, как, например, ликогала надревная (Lycogala epidendrum).
Вершиной эволюционного дерева светлоспоровых миксомицетов стали два порядка, лицеевые и трихиевые. Порядок лицеевые, ныне понимаемый в более узком смысле, чем раньше, объединяет формы, в основном перешедшие к плодоношению на коре деревьев. После дождя кора сохраняет влажность недолго, так что жизненный цикл миксомицета вынужденно укорачивается. Не успевая накопить значительную биомассу, лицеевые образуют очень мелкие плодовые тела. К тому же им не нужны ножки: с высоты древесного ствола споры прекрасно рассеиваются и без них. А вот миксомицеты порядка трихиевые предпочитают жить на мертвой древесине, сохраняющей влажность почти весь сезон. Здесь очень сложно добиться полного высыхания и, соответственно, рассеивания спор, поэтому капиллиций у трихиевых устроен очень сложно: он обилен, разветвлен и орнаментирован спиралями, кольцами, шипами и т. п. Благодаря этим структурам трубочки капиллиция способны пружинить и совершать самостоятельные гигроскопические движения, взрыхляя, подсушивая и понемногу рассеивая споры. Между прочим, аналогичную стратегию разрыхления споровой массы используют печеночники и хвощи.
п. Благодаря этим структурам трубочки капиллиция способны пружинить и совершать самостоятельные гигроскопические движения, взрыхляя, подсушивая и понемногу рассеивая споры. Между прочим, аналогичную стратегию разрыхления споровой массы используют печеночники и хвощи.
Анализ эволюции морфологических признаков, проведенный авторами обсуждаемой статьи, в который раз показал, что наиболее яркие и заметные морфологические признаки — такие, как наличие капиллиция, извести, ножек — являются плохими предсказателями родственных связей, поскольку имеют склонность возникать и исчезать в ходе эволюции неоднократно. Поэтому одной из важнейших задач современной систематики является поиск альтернативных морфологических (или, в более широком смысле, фенотипических) критериев для таксонов, основанных на молекулярно-филогенетических данных. Такими критериями для миксомицетов оказались крепление капиллиция к перидию, происхождение ножки и капиллиция, наличие спороподобных клеток в ножке и ряд других особенностей.
Завершая обзор новой классификации миксомицетов, хочется подчеркнуть, что филогенетическая систематика этой группы сегодня подошла к тому уровню знаний, который для животных, зеленых растений и настоящих грибов был достигнут к концу 1990-х годов. Общие контуры эволюционных событий уже проступили, но детализация родственных связей на уровне родов и видов все еще остается проблематичной. Хочется надеяться, что транскриптомный анализ и мультигенная филогения, а также критическая ревизия отдельных таксонов миксомицетов, позволят в ближайшие годы узнать намного больше об эволюции этих странных обитателей Земли.
Источник: D. V. Leontyev, M. Schnittler, S. Stephenson, Y. K. Novozhilov, O. N. Shchepin. Towards a phylogenetic classification of the Myxomycetes // Phytotaxa. 2019. V. 399. № 3. P. 209–238. DOI: 10.11646/phytotaxa.399.3.5.
Дмитрий Леонтьев
Протисты (одноклеточные эукариоты) — презентация онлайн
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
1.
 Протисты (одноклеточные эукариоты)
Протисты (одноклеточные эукариоты)2. Общая характеристика протистов
• Среда обитания – влажная почва, лужи,водоемы, организмы человека и животного.
• Размеры – от нескольких мкм до 0,2-0,6 мм.
• Наружные покровы – плазматическая мембрана,
пелликула, раковина, целлюлозная оболочка.
• Строение – ядро, цитоплазма, пищеварительные
и сократительные вакуоли.
• Органоиды
движения
–
псевдоподии
(ложноножки), жгутики и реснички.
• Дыхание – всей поверхностью тела.
3. Общая характеристика протистов
• Размножение:1) Бесполое – деление клетки на 2-е, почкование,
спорогония.
2) Половое – коньюгация и коппуляция.
• Приспособление к неблагоприятным условиям –
инцистирование – образование цисты –
стадии покоя.
• Питание:
• Автотрофное – способны к фотосинтезу.
• Гетеротрофное – готовыми органическими
веществами.
• Автогетеротрофное – смешанный тип питания.
4. Гетеротрофные протисты.
 Амеба обыкновенная.Среда
Амеба обыкновенная.Средаобитания
Придонный
ил пресных
водоемов
Размеры
0,2 – 0,5 мм
Форма тела
Постоянно
меняется
Органоиды
движения
Ложноножки
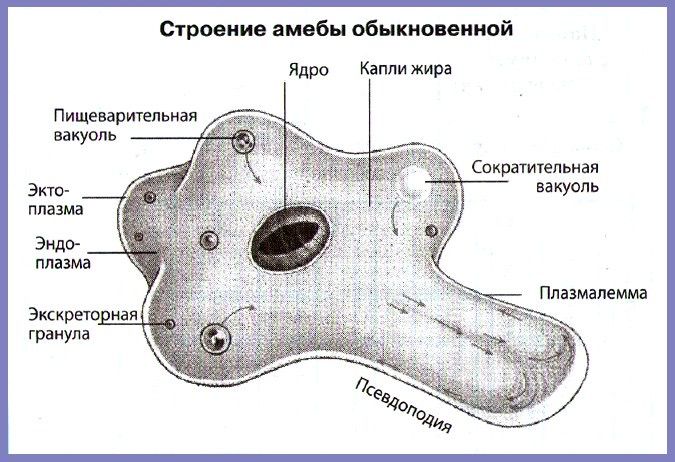
5. Строение Амебы обыкновенной
6. Питание Амебы обыкновенной
• Питаетсябактериями
и
мелкими
водорослями по типу фагоцитоза.
• Переваривание
происходит
в
пищеварительных вакуолях.
7. Дыхание и выделение
• Дыхание происходит диффузно – всейповерхностью тела.
• Выделение – при помощи сократительной
вакуоли.
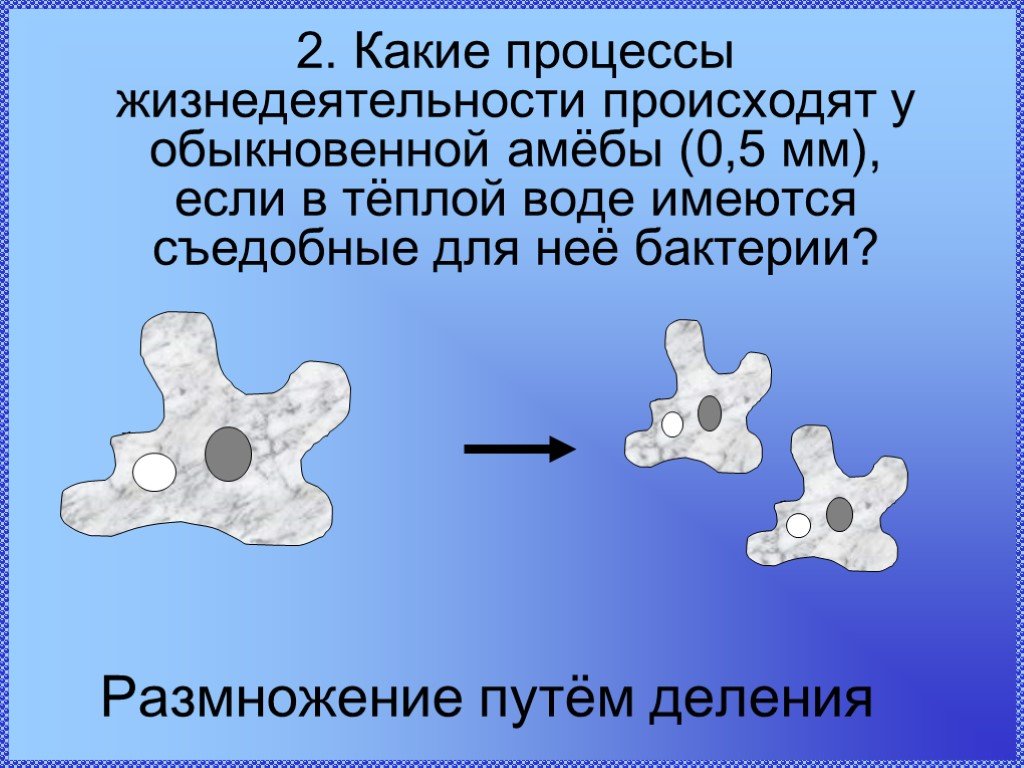
8. Размножение Амебы обыкновенной
• Размножение – бесполое – делением клеткинадвое.
• Распространяется при помощи цисты.
9. Гетеротрофные протисты. Инфузория-туфелька.
Средаобитания
Загрязненные
водоемы со
стоячей водой
Размеры
0,1 – 0,3
мм
Форма тела
Постоянная в
виде
отпечатка
стопы
человека
Органоиды
движения
Покров
Реснички
Пелликула
10.
 Строение Инфузории-туфельки
Строение Инфузории-туфельки11. Питание Инфузории-туфельки
• Пища попадает в клеточный рот и клеточнуюглотку;
• Переваривание – в пищеварительных вакуолях;
• Непереваренные остатки выводятся через
порошицу.
12. Дыхание и выделение Инфузории-туфельки
• Дыхание – всей поверхностью тела.• Выделение – две сократительных вакуоли на
концах клетки.
Сократительные
вакуоли
13. Размножение Инфузории-туфельки
• Бесполое – поперечным делением надвое.• Половое – коньюгация (объем частями
микронуклеусов, без увеличения числа
особей).
Макронуклиусы
Микронуклиусы
Бесполое размножение
Коньюгация
15. Автогетеротрофы. Эвглена зеленая
Средаобитания
Пресные
водоемы с
загрязненной
стоячей водой
Размеры
0,5 – 0,6
мм
Форма тела
Овальная,
передний
конец тупой,
задний
заостренный
Органоиды
движения
Жгутик
(вырост
цитоплазмы)
16.
 Строение Эвглены зеленой
Строение Эвглены зеленой17. Жизнедеятельность Эвглены зеленой
Питание:• На свету способна к фотосинтезу – автотроф.
• В темноте – готовыми органическими
веществами – гетеротроф.
• Дыхание – всей поверхностью тела.
• Выделение – с помощью сократительной
вакуоли.
• Размножение – продольное деление клетки
надвое.
• Приспособление к распространению – цисты.
18. Автогетеротрофы. Хламидомонада
19. Автогетеротрофы. Хлорелла
Средаобитания
Мелкие
пресные
водоемы,
влажная почва,
кора деревьев
Покров
Форма тела
Органоиды
движения
Плотная
целлюлоз
ная
оболочка
Шаровидная
Неподвижна
20. Автогетеротрофы. Хлорелла строение
21. Малярийный плазмодий
• Паразит клеток печени и эритроцитов человека,вызывает тяжелое заболевание – малярию.
• Заражение человека происходит через укус самки
малярийного
комара
(цикл
полового
размножения).

• Вегетативные формы плазмодия со слюной комара
попадают в кровь человека и заносятся в клетки
печени.
• В печени — цикл бесполого размножения
(шизогония), попадают в кровь (в эритроциты) и
разрушают эритроцит.
Симптомы:
• Высокая
температура,
поражение
печени,
селезенки, иногда головного мозга.
22. Жизненный цикл малярийного плазмодия
23. Классификация простейших
ПростейшиеКласс
Корненожки
Класс
Жгутиковые
Амеба
обыкновенная
Эвглена
зеленая
Класс
Инфузории
Инфузориятуфелька
Класс
Споровики
Малярийный
плазмодий
24. Значение протистов
В природе:• Участвуют в круговороте веществ.
• Пища для водных животных.
• Санитары в природе (очищают водоемы).
В медицине (паразиты человека):
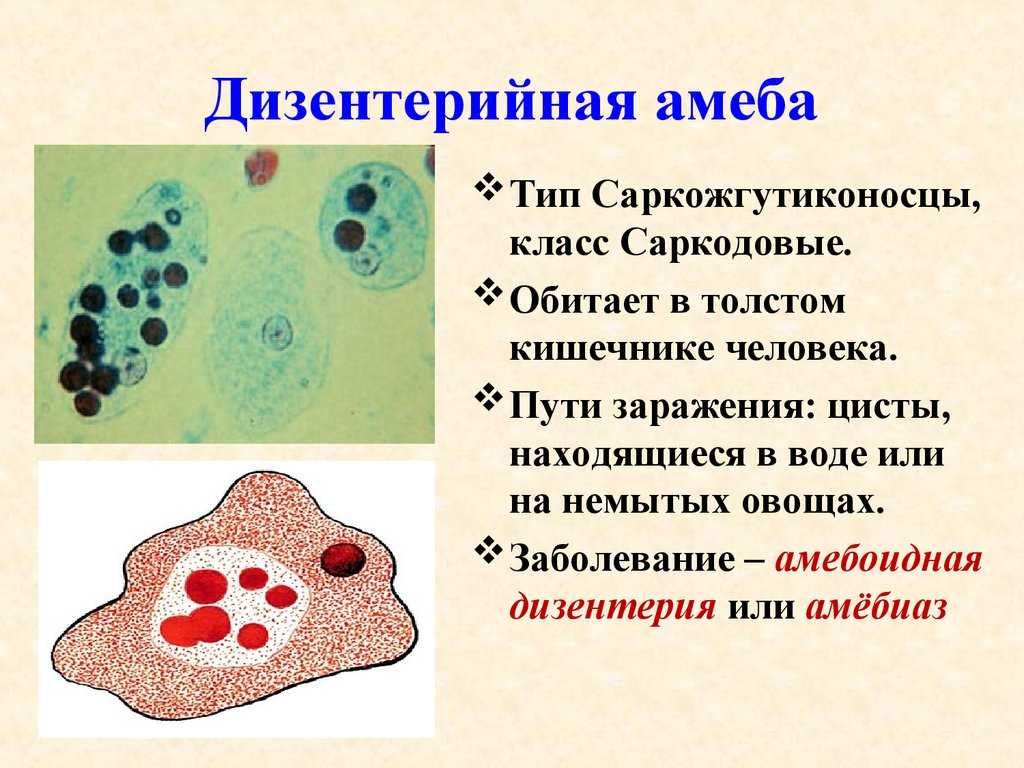
• Паразиты кишечника – дизентерийная амеба
и инфузория балантидий.
• Паразит печени и тонкого кишечника –
лямблия.
• Паразиты крови – малярийный плазмодий.
English Русский Правила
Протист — Энциклопедия Нового Света
? Протисты | ||||
|---|---|---|---|---|
| Paramecium aurelia , инфузория | ||||
| Научная классификация | ||||
| ||||
| Типичный тип | ||||
|
Протисты представляют собой гетерогенную группу живых организмов, состоящую из тех эукариот, которые не являются животными, растениями или грибами. В системах биологической классификации к ним обычно относят Царство 9.0008 Protista или Protista .
В системах биологической классификации к ним обычно относят Царство 9.0008 Protista или Protista .
Протисты более сложны, чем бактерии, поскольку у них есть клеточные ядра, но менее сложны, чем грибы, растения и животные, потому что они в основном одноклеточные, а если и многоклеточные, то у них отсутствуют высокоспециализированные ткани.
Протисты относятся к парафилетическому классу, а не к природной (монофилетической) группе, и не имеют много общего, кроме относительно простой организации. Некоторые называют это «остатками» других эукариотических царств.
Протисты являются важной частью земных экосистем и заслуживают особого внимания, поскольку составляют большую часть планктона, от которого зависит почти вся жизнь в океанах. В нескольких отношениях протисты обнаруживают удивительную гармонию и единство в творении. Небольшой фотосинтезирующий планктон использует углекислый газ и производит кислород — на самом деле большую часть кислорода в мире, — в то время как другие организмы потребляют кислород и производят углекислый газ.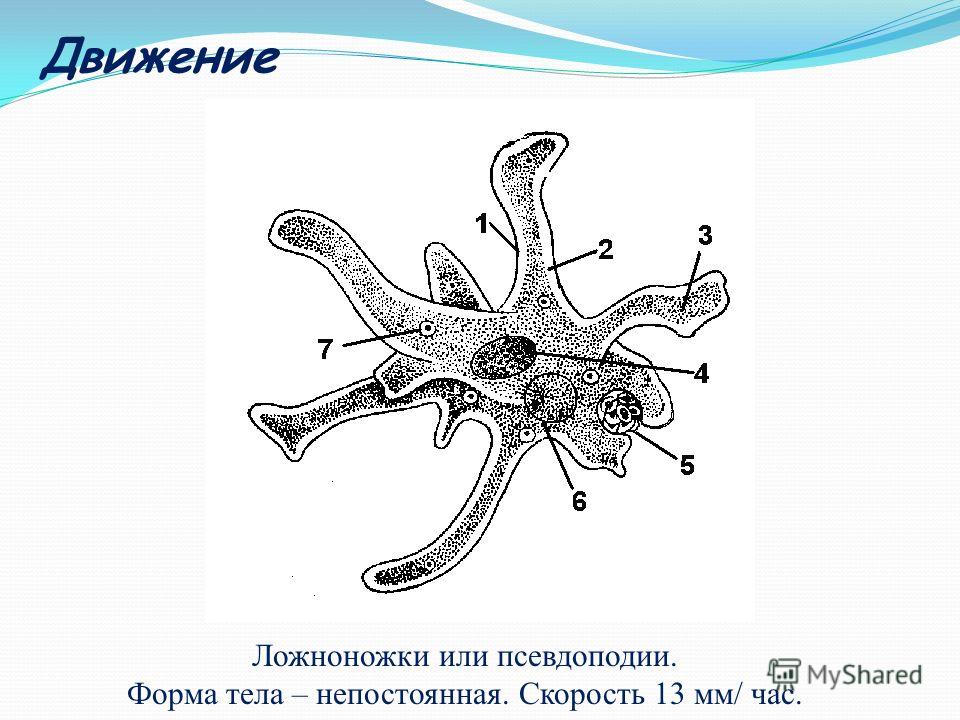 Фитоплантон улавливает солнечную энергию и служит пищей и энергией для организмов на более высоких уровнях тропиков, а чтобы замкнуть круг, они служат редуцентами при разрушении мертвых растительных и животных материалов, чтобы их можно было использовать повторно. Многие протисты, особенно водоросли, участвуют с другими организмами в симбиотических отношениях, приносящих взаимную выгоду (мутуализм).
Фитоплантон улавливает солнечную энергию и служит пищей и энергией для организмов на более высоких уровнях тропиков, а чтобы замкнуть круг, они служат редуцентами при разрушении мертвых растительных и животных материалов, чтобы их можно было использовать повторно. Многие протисты, особенно водоросли, участвуют с другими организмами в симбиотических отношениях, приносящих взаимную выгоду (мутуализм).
С другой стороны, протисты также являются причиной многих болезней, поражающих как людей, так и культурные растения, от которых мы зависим в качестве пищи. С некоторыми человеческими заболеваниями связаны уроки личной и социальной ответственности. В случае малярии (вызванной простейшими) люди имеют доступ к противомоскитным сеткам и вакцинам, а те, у кого есть средства, несут моральное обязательство помогать тем, кто не может позволить себе такие меры по спасению жизни.
Содержание
- 1 Традиционная классификация
- 1.1 Водоросли
- 1.
 2 Простейшие
2 Простейшие - 1.3 Грибовидные протисты
- 2 Филогенетические классификации
- 3 Каталожные номера
- 4 кредита
Традиционная классификация
В таксономической схеме пяти царств, разработанной Робертом Уиттакером в 1969 г., протисты входят в одно из пяти царств: Monera (прокариоты — бактерии и сине-зеленые водоросли), Protista (одноклеточные, многоклеточные и колониальные протисты), Грибы, растения и животные. Эта система остается популярной и сегодня. Точно так же протисты — это царство в таксономической схеме шести царств с добавлением архебактерий в качестве шестого царства. В трехдоменной системе Карла Вёзе протисты представляют собой царство под группировкой высшего уровня эукариот.
Когда-то бактерии также считались протистами в рамках системы трех царств: Animalia (что близко соответствует современному царству), Plantae (в которое входили не только растения, но и грибы) и Protista (все остальное). Однако большинство современных систем классификации рассматривают бактерии отдельно.
Однако большинство современных систем классификации рассматривают бактерии отдельно.
За последние 150 лет протисты были разделены на несколько групп на основе сходства с высшими царствами: растительноподобные водоросли, животноподобные простейшие и грибоподобные слизевики и водяные плесени. Эти группы часто пересекаются и были заменены филогенетическими классификациями. Однако они по-прежнему полезны как неформальные группы для описания морфологии и экологии протистов.
Водоросли
Водоросли, как и растения, собирают световую энергию посредством фотосинтеза и преобразуют ее в химическую энергию. Среди них много одноклеточных существ, которых также можно считать простейшими, например Euglena , которые приобрели хлоропласты в результате вторичного эндосимбиоза. Другие неподвижны, а некоторые (называемые морскими водорослями) действительно многоклеточные, включая представителей следующих групп:
| Хлорофиты | зеленые водоросли, относящиеся к высшим растениям | например, Ульва |
| Родофиты | красные водоросли | например, Порфира |
| Гетероконтофиты | бурые водоросли, диатомовые водоросли и т. д. д. | например, Macrocystis |
Зеленые и красные водоросли, а также небольшая группа, называемая глаукофитами, являются близкими родственниками других растений, поэтому некоторые авторы рассматривают их как Plantae, несмотря на их простую организацию. Однако большинство других видов водорослей развивались отдельно. К ним относятся гаптофиты, криптомонады, динофлагелляты, эвглениды и хлорарахниофиты, которые также считаются простейшими. Некоторые простейшие являются хозяевами эндосимбиотических водорослей, например 9.0016 Paramecium bursaria или радиолярии, которые обеспечивают их энергией, но не интегрируются в клетку.
California Kelp Forest
Водоросли встречаются во многих средах как в воде, так и на суше. Они наиболее важны, так как наряду с цианобактериями входят в состав фитопланктона Мирового океана. Фитопланктон является основным энергетическим уровнем почти всей морской жизни.
Некоторые водоросли существуют в симбиотических отношениях с другими организмами. Лишайники – это союз грибов и водорослей. В некоторых кораллах, губках и медузах обитают водоросли, которые обеспечивают их пищей, а взамен получают защиту. Водоросли также живут в волосах ленивцев в тропических лесах Южной Америки. Эти отношения дают водорослям дом, а ленивцу — зеленый цвет для маскировки.
Морские водоросли — это многоклеточные водоросли. Хотя они могут вырасти очень большими, им не хватает сложной структуры растений. Самая крупная водоросль (и самый крупный протист) — это гигантская ламинария, Macrocystis pyrifera , обитающая вдоль тихоокеанского побережья Северной и Южной Америки. Он может вырасти до 60 метров в длину (200 футов). Некоторые водоросли собирают люди для еды и других целей.
Простейшие
Простейшие в основном одноклеточные, подвижные (могут передвигаться) протисты, которые питаются путем фагоцитоза, обволакивая частицы через клеточную мембрану. Однако есть многочисленные исключения. Простейшие обычно имеют размер всего 0,01–0,5 мм, как правило, слишком малы, чтобы их можно было увидеть без микроскопа. Простейшие повсеместно распространены в водной среде и почве, обычно переживая засушливые периоды в виде цист или спор, а также включают несколько важных паразитов. По способу передвижения простейшие делятся на:
Однако есть многочисленные исключения. Простейшие обычно имеют размер всего 0,01–0,5 мм, как правило, слишком малы, чтобы их можно было увидеть без микроскопа. Простейшие повсеместно распространены в водной среде и почве, обычно переживая засушливые периоды в виде цист или спор, а также включают несколько важных паразитов. По способу передвижения простейшие делятся на:
| Амебоиды | с транзиторной псевдоподией | например, Амеба |
| Инфузории | с несколькими короткими ресничками | например, Парамеций |
| споровики | неподвижных паразитов; форма спор | например, Плазмодий |
Некоторые простейшие являются важной частью морского планктона в качестве потребителей водорослей и другого растительного фитопланктона, а также в качестве пищи для морских животных. Они также важны как редуценты как в наземной среде, так и в океане.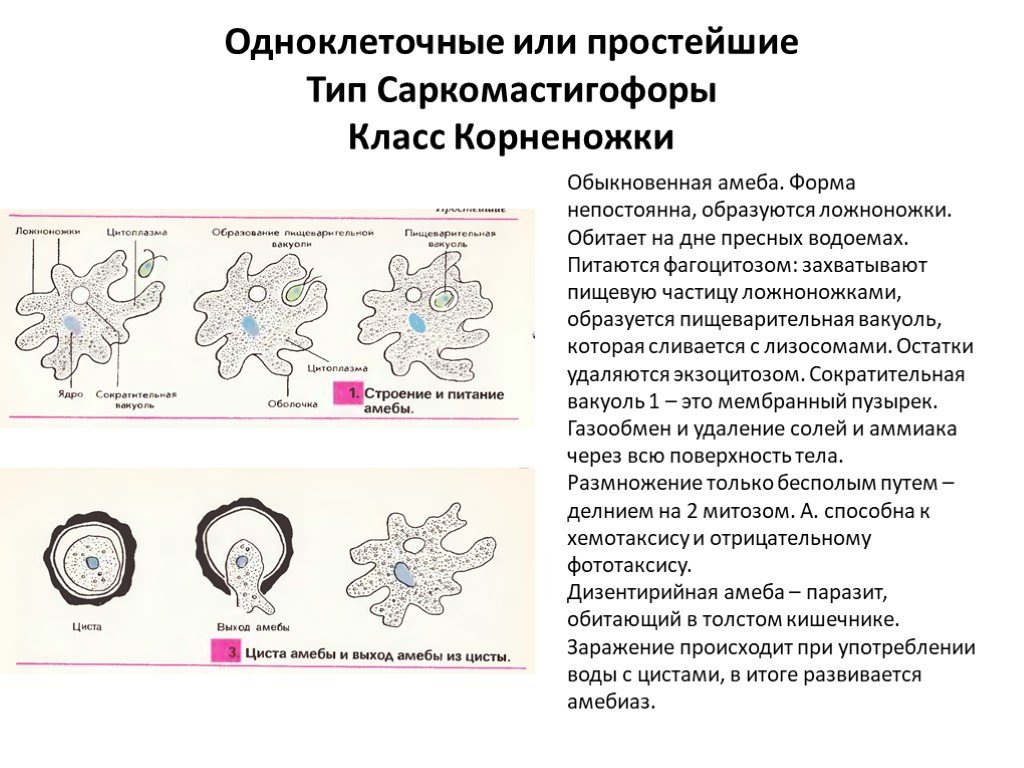 Другие являются паразитами растений и животных.
Другие являются паразитами растений и животных.
Простейшие являются причиной многих заболеваний человека. Наиболее серьезной является малярия, вызываемая четырьмя видами рода Plasmodium , простейшими, чей жизненный цикл делает их, в свою очередь, паразитами как человека, так и комаров. Малярия убивает более миллиона человек в год, большинство из них — африканские дети.[1] Другие заболевания, вызываемые простейшими, включают: африканскую сонную болезнь, амебную дизентерию, болезнь Шагаса, лямблиоз, лейшманиоз, первичный амебный менингоэнцефалит (ПАМ) и вагинит.
Грибоподобные протисты
Различные организмы с организацией на уровне протистов первоначально рассматривались как грибы, поскольку они производят спорангии, структуры, производящие и содержащие споры. К ним относятся хитриды, слизевики, водные плесени и лабиринтуломицеты.
Хитриды могут быть одноклеточными или многоклеточными. Существует около тысячи видов, большинство из которых обитает в воде или почве. Большинство — разлагатели. Некоторые из них являются паразитами и могут вызывать заболевания растений, включая кукурузу, люцерну и картофель. Один вид, Batrachochytrium dendrobatidis , по-видимому, является причиной хитридиомикоза, болезни лягушек, серьезно поражающей многие популяции диких лягушек по всему миру.
Большинство — разлагатели. Некоторые из них являются паразитами и могут вызывать заболевания растений, включая кукурузу, люцерну и картофель. Один вид, Batrachochytrium dendrobatidis , по-видимому, является причиной хитридиомикоза, болезни лягушек, серьезно поражающей многие популяции диких лягушек по всему миру.
Слизевики на газоне, США Виден след движения.
Слизевики отличаются необычным жизненным циклом. У некоторых видов отдельные одноклеточные организмы объединяются и сливаются, образуя гигантскую клетку с тысячами ядер. Это тело, называемое плазмодием, может передвигаться, питаясь бактериями, грибками и разлагающимися растительными веществами. (Это другое использование слова plasmoidium из использованного выше для рода паразитических простейших.) Слизевики встречаются во всем мире.
Водяная плесень прекрасно себя чувствует в воде и влажной почве. Считается, что они более тесно связаны с растениями, чем с грибами, поскольку имеют клеточные стенки из целлюлозы.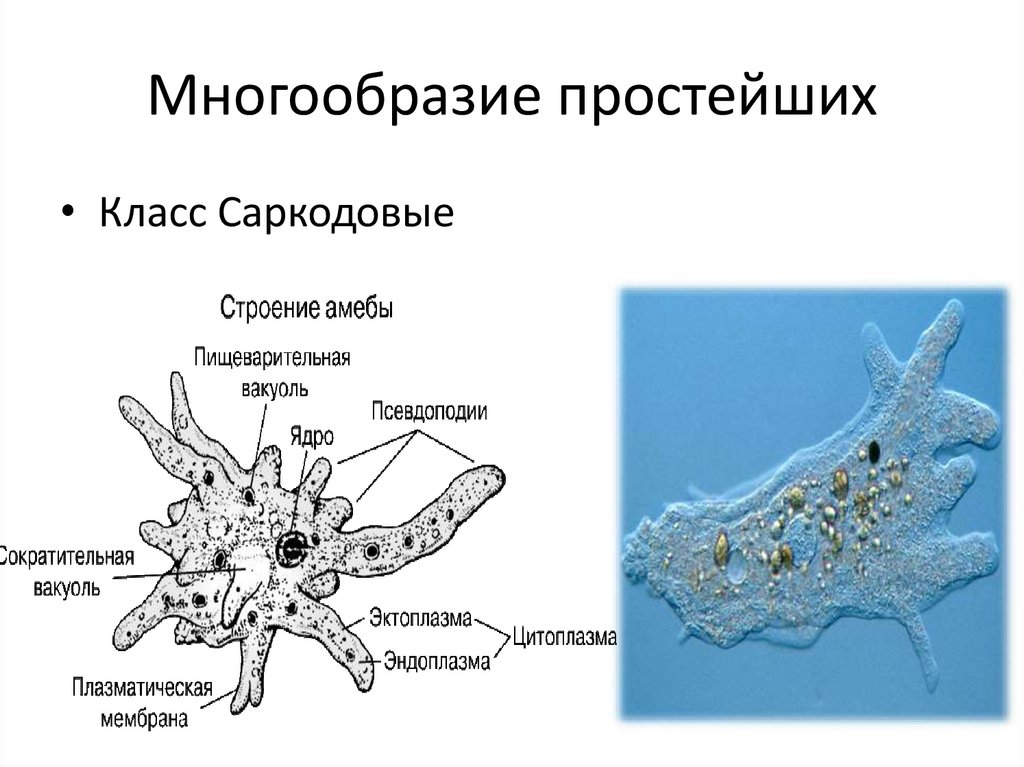 Они одноклеточные. Многие из них являются паразитами и могут вызывать заболевания у растений, грибов и животных. Один вид Phytophthora infestans вызывает фитофтороз картофеля, который привел к картофельному голоду в Ирландии.
Они одноклеточные. Многие из них являются паразитами и могут вызывать заболевания у растений, грибов и животных. Один вид Phytophthora infestans вызывает фитофтороз картофеля, который привел к картофельному голоду в Ирландии.
Лабиринтуломицеты образуют сеть трубочек или нитей, по которым скользят одноклеточные организмы для сбора пищи. Они в основном морские и разлагают мертвый растительный материал или паразитируют на растениях и водорослях или некоторых животных.
Филогенетические классификации
Классификация простейших все еще меняется. В более новых классификациях делается попытка представить монофилетические группы на основе ультраструктуры, биохимии и генетики. Монофилетическая группа состоит из общего предка и всех его потомков, тогда как таксономическая группа, которая содержит некоторых, но не всех потомков самого последнего общего предка, называется парафилетической. Поскольку протисты в целом парафилетичны, такие системы часто распадаются или покидают царство, вместо этого рассматривая группы протистов как отдельные линии эукариот. Схема по Adl и др. (2005 г.) является примером, который не беспокоится о рангах (тип, класс и т. д.).
Схема по Adl и др. (2005 г.) является примером, который не беспокоится о рангах (тип, класс и т. д.).
Ссылки
Ссылки ISBN поддерживают NWE за счет реферальных сборов
- Adl, S.M., A.G.B. Симпсон, М.А. , С. Карпов, П. Кугренс, Дж. Круг, С. Э. Лейн, Л. А. Льюис, Дж. Лодж, Д. Х. Линн, Д. Г. Манн, Р. М. Маккорт, Л. Мендоза, О. Моструп, С.Э. Мозли-Стэндридж, Т.А. Нерад, К.А. Ширер, А.В. Смирнов, Ф.В. Шпигель и М.Ф.Дж.Р. Тейлор. 2005. «Новая классификация эукариот более высокого уровня с упором на таксономию протистов». Journal of Eukaryotic Microbiology 52 (5): 399-451. [2]
- Кавалье-Смит, Т. 2003. «Филогения протистов и классификация простейших высокого уровня». Европейский журнал протистологии 39:338-348.
- Lipscomb, D. 2006. «Protozoa» (по состоянию на 15 июля 2006 г.).
- Всемирная организация здравоохранения. 2006. Global Malaria Program (по состоянию на 14 июля 2006 г.)
Credits
New World Encyclopedia авторы и редакторы переписали и дополнили Википедия статья
в соответствии со стандартами New World Encyclopedia . Эта статья соответствует условиям лицензии Creative Commons CC-by-sa 3.0 (CC-by-sa), которая может использоваться и распространяться с надлежащим указанием авторства. Упоминание должно быть выполнено в соответствии с условиями этой лицензии, которая может ссылаться как на авторов New World Encyclopedia , так и на самоотверженных добровольных участников Фонда Викимедиа. Чтобы процитировать эту статью, щелкните здесь, чтобы просмотреть список допустимых форматов цитирования. История более ранних вкладов википедистов доступна исследователям здесь:
Эта статья соответствует условиям лицензии Creative Commons CC-by-sa 3.0 (CC-by-sa), которая может использоваться и распространяться с надлежащим указанием авторства. Упоминание должно быть выполнено в соответствии с условиями этой лицензии, которая может ссылаться как на авторов New World Encyclopedia , так и на самоотверженных добровольных участников Фонда Викимедиа. Чтобы процитировать эту статью, щелкните здесь, чтобы просмотреть список допустимых форматов цитирования. История более ранних вкладов википедистов доступна исследователям здесь:
- Протист история
- Парафилия история
История этой статьи с момента ее импорта в New World Encyclopedia :
- История «Protist»
Примечание. На использование отдельных изображений, лицензированных отдельно, могут распространяться некоторые ограничения.
Классификация голых амеб (Phylum Rhizopoda)
Сумерки Sarcodina: молекулярный взгляд на полифилетическое происхождение амебоидных простейших
- J.
 Pawłowski
Pawłowski Биология
- 2008
Хотя молекулярные данные ясно указывают на то, что псевдоподиальные структуры развивались несколько раз в истории эукариот, они также показывают, что большая диверсификация происходила только у амебоидных дважды в линиях, ведущих к Amoebozoa и Rhizaria.
Pseudoparamoeba garorimi n. sp., с примечаниями о видовых различиях внутри рода
Новый морской вид голых лопастных амеб Pseudoparamoeba garorimi n. сп. (Amoebozoa, Dactylopodida), выделенные из приливно-отливных морских отложений залива Гарорим, Корея, изучались при свете и…
Phylogenetic position of the Amoeba genus Deuteramoeba (Amoebozoa, Tubulinea)
- O. Kamyshatskaya, E. Nassonova, A. Smirnov
Biology
- 2017
This work sequenced SSU rRNA gene of the CCAP 1586/ 1, представляющий типовой штамм вида Deuteramoeba mycophaga, тем самым добавив третий род к филогении этого семейства и поместив его в качестве сестринской группы к кладе Amoeba plus Chaos, тем самым подтвердив его положение, выведенное ранее из морфологических признаков.
Два новых вида рода Stenamoeba (Discosea, Longamoebia): цитоплазматический MTOC присутствует еще в одной линии амеб.
- S. Geisen, Jan Weinert, A. Kudryavtsev, A. Glotova, M. Bonkowski, A. Smirnov
Biology
European journal of protistology
- 2014
The Vampyrellid Amoebae (Vampyrellida, Rhizaria) .
- Себастьян Гесс, Андреас Зутхаус
Биология
Протисты
- 2021
Молекулярная филогения и классификация лопастных амеб.
- A. Smirnov, E. Nassonova, C. Berney, J. Fahrni, I. Bolívar, J. Pawłowski
Biology
Protist
- 2005
Amoebainte Atticle: Phylebiniation at Pedifytinia Atabiniation at Atibinia.
Используя данные о последовательности гена рибосомной РНК малой ядерной субъединицы, делается вывод, что S. pedata и опубликованный неотип S. diploidea являются родственными, но не конспецифичными.