Саркомастигофоры | Саркодовые | Амеба протей (обыкновенная), амеба дизентерийная, радиолярия | |
Жгутиковые | Эвглена зеленая, вольвокс, трипаносома африканская, лейшмания, трихомонада, лямблия печеночная | ||
Споровики | Кокцидиеобразные | Малярийный плазмодий | |
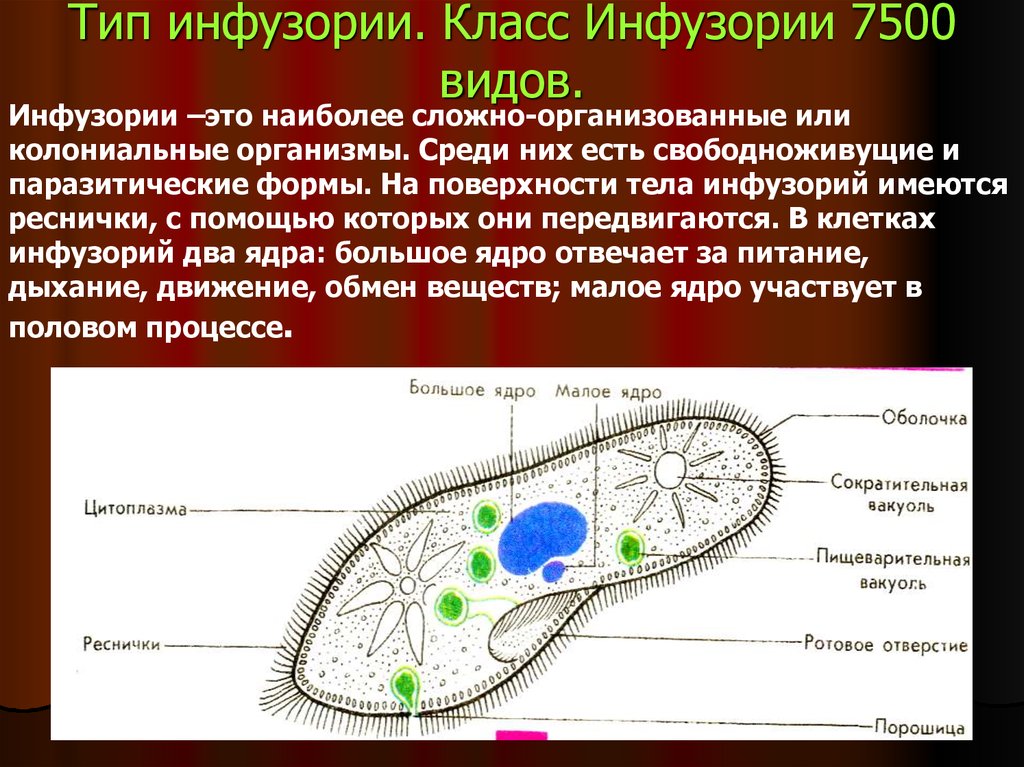
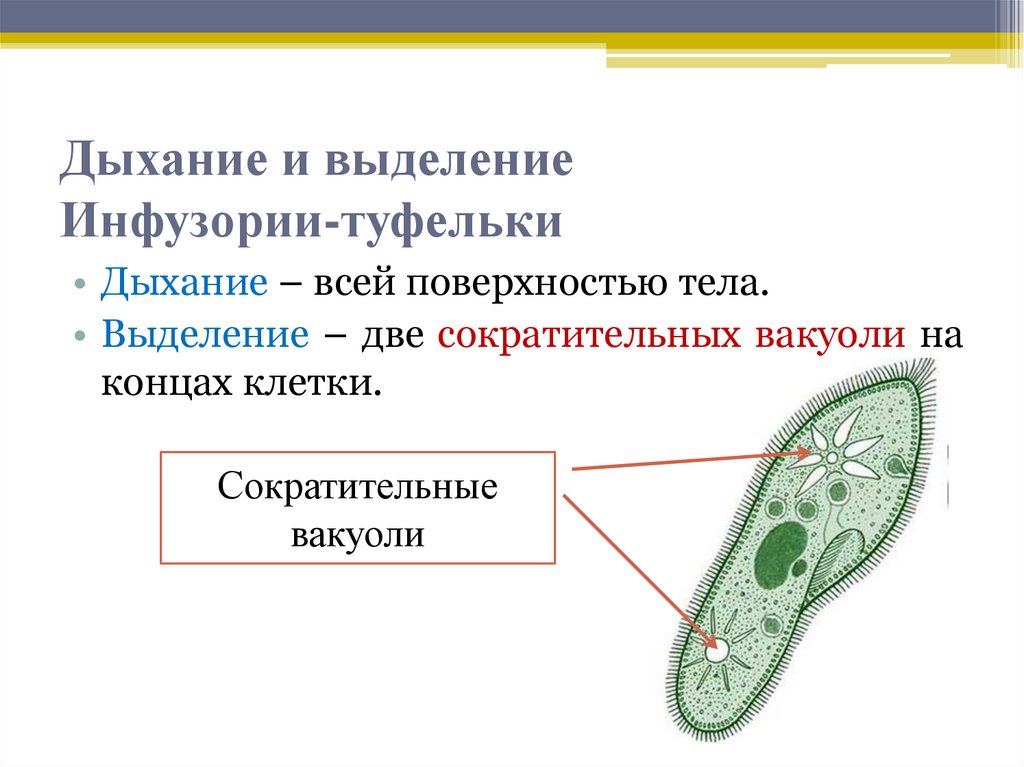
Инфузории | Ресничные | Инфузоря-балантидий, инфузория-туфелька, инфузория-трубач | |
Трихофриоз |
 — М.: АСТ: Астрель; Владимир; ВКТ, 2009 2.Биология: Животные: учеб. для 7-8 кл. общеобразоват. Учреждений. — 7-е изд. — М.: Просвещение, 2000. 3.Биология: учеб.-справ.пособие / А.Г.Лебедев. М.: АСТ: Астрель. 2009. 4.Биология. Полный курс общеобразовательной средней школы: учебное пособие для школьников и абитуриентов / М.А.Валовая, Н.А.Соколова, А.А. Каменский. — М.: Экзамен, 2002. 5.Биология для поступающих в вузы. Интенсивный курс / Г.Л.Билич, В.А.Крыжановский. — М.: Издательство Оникс, 2006.
— М.: АСТ: Астрель; Владимир; ВКТ, 2009 2.Биология: Животные: учеб. для 7-8 кл. общеобразоват. Учреждений. — 7-е изд. — М.: Просвещение, 2000. 3.Биология: учеб.-справ.пособие / А.Г.Лебедев. М.: АСТ: Астрель. 2009. 4.Биология. Полный курс общеобразовательной средней школы: учебное пособие для школьников и абитуриентов / М.А.Валовая, Н.А.Соколова, А.А. Каменский. — М.: Экзамен, 2002. 5.Биология для поступающих в вузы. Интенсивный курс / Г.Л.Билич, В.А.Крыжановский. — М.: Издательство Оникс, 2006. 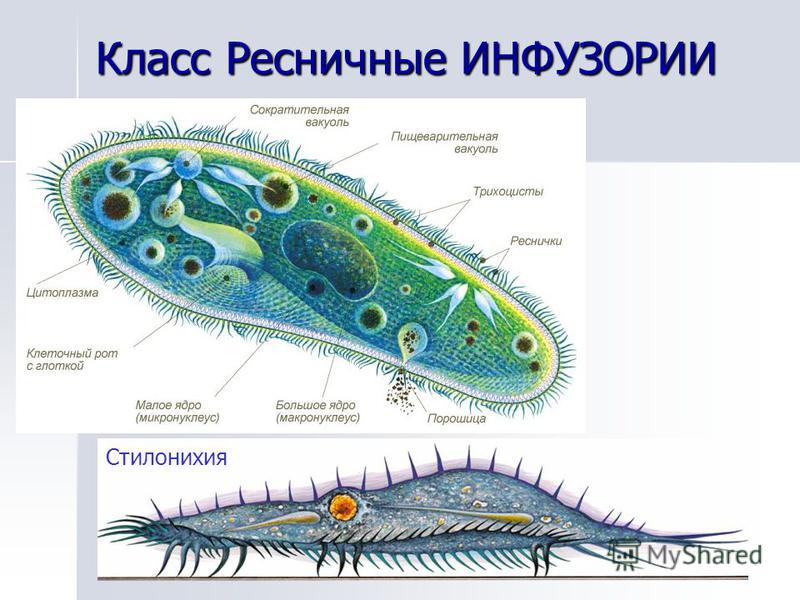
Признаки простейших одноклеточных | Амеба обыкновенная (Класс Корненожки) | Эвглена зеленая (класс Жгутиконосцы) | Инфузория ту-фелька (класс Ин-фузории) |
Строение | Состоит из цитоплазмы, ядра, сократительной вакуоли, ложноножки, пищеварительной вакуоли (см. рис) | Состоит из оболочки, ядра, жгутика, глазок, сократительной вакуоли, питательных веществ, хлоропласты (см. рис.) | Состоит из мембраны, малого и большого ядра, сократительной и пищеварительной вакуоли, рот, порошица, реснички (см. |
Движение | «Перетекание» с по-мощью ложноножек | Передвижение с помощью жгутика | Передвижение с помощью рес-ничек |
Кормом могут быть бактерии, микроскопические водоросли. Амёба захватывает пищу, вытягивая ложноножки на любом участке тела. Они обволакивают добычу и вместе с небольшим количеством воды погружают её в цитоплазму. Так образуется пищеварительная вакуоль — фагоцитоз, за-хват капель жидкости — пиноцитоз. Из пищеварительной вакуоли растворимые продукты пищеварения поступают в цитоплазму, а непереваренные остатки выводятся из организма в любой части клетки. | Автотрофное (фото-синтез) или гетеро-трофное (фагоцитоз и пиноцитоз) | Питаются различными микроорганизмами, преимущественно бактериями. Движением ресничек, расположенных вдоль ротового углубления, загоняют в него добычу. Вместе с водой она попадает в клеточный рот, затем в глотку. | |
Размноже-ние | Амеба размножается делением. При этом ядро делится надвое. Образовавшиеся новые ядра расходятся в стороны, и между ними появляется поперечная перетяжка, разделяющая амёбу на две дочерние клетки, которые живут самостоятельно. Через некоторое время молодые амёбы также начинают делиться. Для размножения благоприятна температура воды около +20 °С. | Размножение организмов данного вида эвглен бесполое — делением клетки пополам, в отличие от инфузории-туфельки, для которой характерен еще и половой процесс. | Инфузории размножаются бесполым путём — поперечным делением, как амёбы. Первым делится надвое малое ядро, затем большое. Одновременно появляется поперечная перетяжка. Она со временем разделяет инфузорию на две молодые (дочерние) клетки. Они растут и при хорошем питании и оптимальной температуре уже на следующие сутки становятся взрослыми и снова могут делиться. Для инфузорий характерен и половой процесс в форме конъюгации (слия-ние двух клеток и обмен генети-ческой информа-цией) |


_______________
Источник информации: Биология в таблицах и схемах./ Издание 2е, — СПб.: 2004.
Класс Жгутиковые
Строение . У жгутиковых имеются жгутики, служащие органоидами движения и способствующие захвату пищи. Их может быть один, два или множество. Движением жгутика в окружающей воде вызывается водоворот, благодаря которому мелкие взвешенные в воде частички увлекаются к основанию жгутика, где имеется небольшое отверстие — клеточный рот, ведущий в глубокий канал-глотку.
Генетический аппарат у большинства жгутиковых представлен одним ядром, но существуют также двуядерные (например, лямблии) и многоядерные (например, опалина) виды.

Цитоплазма четко делится на тонкий наружный слой — прозрачную эктоплазму и глубже лежащую эндоплазму.
Способ питания. По способу питания жгутиковые делятся на три группы. Автотрофные организмы как исключение в царстве животных синтезируют органические вещества (углеводы) из углекислого газа и воды при помощи хлорофилла и энергии солнечного излучения. Хлорофилл находится в хроматофорах, сходных по организации с пластидами растений. У многих жгутиконосцев с растительным типом питания имеются особые аппараты, воспринимающие световые раздражения, — стигмы.
Осморегуляторная и отчасти выделительная функции выполняются у жгутиковых, как у саркодовых, сократительными вакуолями, которые имеются у свободноживущих пресноводных форм.

Размножение. У жгутиковых отмечается половое и бесполое размножение. Обычная форма бесполого размножения — продольное деление.
Среда обитания. Жгутиковые широко распространены в пресных водоемах, особенно небольших и загрязненных органическими остатками, а также в морях. Многие виды паразитируют у различных животных и человека и тем самым приносят большой вред (трипоносомы, паразиты кишечника и др.).
Общими чертами организации простейших являются следующие:
Большинство простейших-одноклеточные, реже колониальные организмы. Их одноклеточное тело обладает функциями целостного организма, которые выполняются органеллами общего назначения (ядро, эндоплазматическая сеть, комплекс Гольджи, лизосомы, митохондрии, рибосомы и др.) и специального (пищеварительные и сократительные вакуоли, жгутики, реснички и др.). Согласованно функционируя, они обеспечивают отдельной клетке возможность существования в качестве самостоятельного организма.
Покровы
простейших представлены либо
только плазматической
мембраной, либо
еще и плотной, довольно гибкой и
эластичной оболочкой — пелликулой, придающей
им относительное постоянство формы
тела.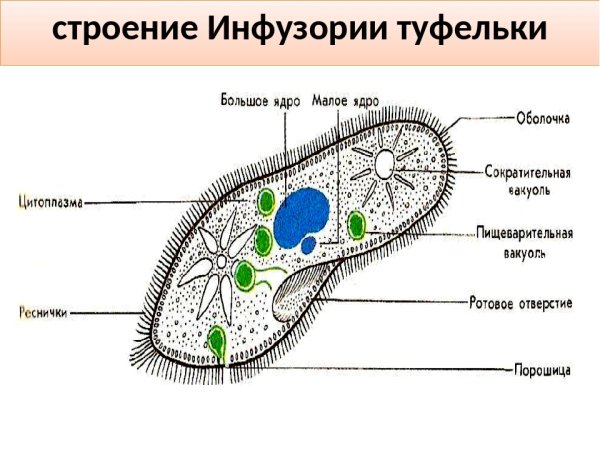
Органоиды движения большинства видов — ложноножки, жгутики или многочисленные короткие реснички.
У пресноводных одноклеточных имеется 1 -2 сократительные вакуоли, основная функция которых состоит в поддержании постоянства осмотического давления, осуществляемого за
Раздражимость у простейших проявляется в форме таксисов.
Большинство
простейших обладает способностью
переносить неблагоприятные условия в
состоянии покоящейся стадии —цисты. При
этом клетка округляется, втягивает или
отбрасывает органоиды движения и
покрывается плотной защитной оболочкой.
Стадия цисты дает возможность простейшему
не только переживать в неактивном
состоянии неблагоприятные условия, но
и расселяться.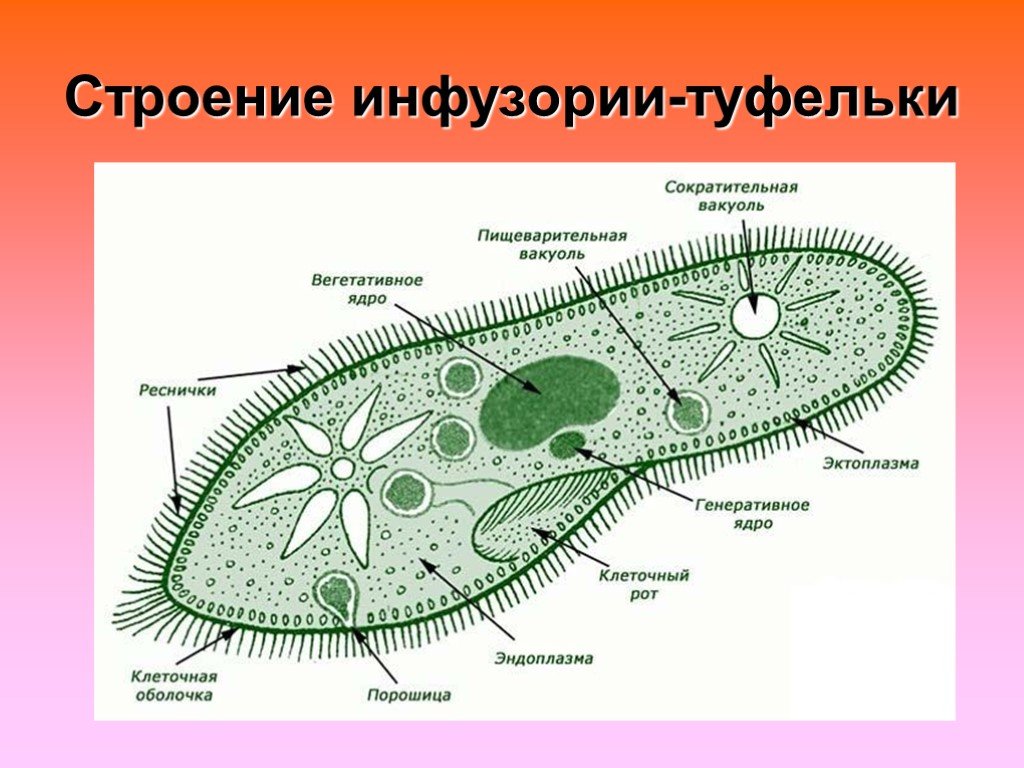
Простейшие подразделяются на классы: Корненожки, Жгутиковые, Инфузории, Споровики.
Эволюция одноклеточных, растительного и животного мира
Эволюция одноклеточных организмов
До
1950-х годов не удавалось обнаружить следы
докембрийской жизни на уровне одноклеточных
организмов, поскольку микроскопические
останки этих существ невозможно выявить
обычными методами палеонтологии. Важную
роль в их обнаружении сыграло открытие,
сделанное в начале XX в. Ч. Уолкотом. В
докембрийских отложениях на западе
Северной Америки он нашел слоистые
известняковые образования в виде
столбов, названные позднее строматолитами.
В 1954 г. было установлено, что строматолиты
формации Ганфлинт (Канада) образованы
останками бактерий и сине-зеленых
водорослей. У берегов Австралии обнаружены
и живые строматолиты, состоящие из этих
же организмов и очень сходные с ископаемыми
докембрийскими строматолитами.
Самые ранние из бактерий (прокариоты) существовали уже около 3,5 млрд. лет назад. К настоящему времени сохранились два семейства бактерий: древние, или археобактерии (галофильные, метановые, термофильные), и эубактерии (все остальные). Таким образом, единственными живыми существами на Земле в течение 3 млрд. лет были примитивные микроорганизмы. Возможно, они представляли собой одноклеточные существа, сходные с современными бактериями, например клостридиями, живущими на основе брожения и использования, богатых энергией органических соединений, возникающих абиогенно под действием электрических разрядов и ультрафиолетовых лучей. Следовательно, в эту эпоху живые существа были потребителями органических веществ, а не их производителями.
Гигантский
шаг на пути эволюции жизни был связан
с возникновением основных биохимических
процессов обмена — фотосинтеза и дыхания
и с образованием клеточной организации,
содержащей ядерный аппарат (эукариоты). Эти «изобретения», сделанные еще на
ранних стадиях биологической эволюции,
в основных чертах сохранились у
современных организмов. Методами
молекулярной биологии установлено
поразительное единообразие биохимических
основ жизни при огромном различии
организмов по другим признакам. Белки
почти всех живых существ состоят из 20
аминокислот. Нуклеиновые кислоты,
кодирующие белки, монтируются из четырех
нуклеотидов. Биосинтез белка осуществляется
по единообразной схеме, местом их синтеза
являются рибосомы, в нем участвуют и-РНК
и т-РНК. Подавляющая часть организмов
использует энергию окисления, дыхания
и гликолиза, которая запасается в АТФ.
Эти «изобретения», сделанные еще на
ранних стадиях биологической эволюции,
в основных чертах сохранились у
современных организмов. Методами
молекулярной биологии установлено
поразительное единообразие биохимических
основ жизни при огромном различии
организмов по другим признакам. Белки
почти всех живых существ состоят из 20
аминокислот. Нуклеиновые кислоты,
кодирующие белки, монтируются из четырех
нуклеотидов. Биосинтез белка осуществляется
по единообразной схеме, местом их синтеза
являются рибосомы, в нем участвуют и-РНК
и т-РНК. Подавляющая часть организмов
использует энергию окисления, дыхания
и гликолиза, которая запасается в АТФ.
Рассмотрим
подробнее особенности эволюции на
клеточном уровне организации жизни.
Наибольшее различие существует не между
растениями, грибами и животными, а между
организмами, обладающими ядром (эукариоты)
и не имеющими его (прокариоты). Последние
представлены низшими организмами —
бактериями и сине-зелеными водорослями
(цианобактерии, или цианеи), все остальные
организмы — эукариоты, которые сходны
между собой по внутриклеточной
организации, генетике, биохимии и
метаболизму.
Различие между прокариотами и эукариотами заключается еще и в том, что первые могут жить как в бескислородной (облигатные анаэробы), так и в среде с разным содержанием кислорода (факультативные анаэробы и аэробы), в то время как для эукариотов, за немногим исключением, обязателен кислород. Все эти различия имели существенное значение для понимания ранних стадий биологической эволюции.
Сравнение прокариот и эукариот по потребности в кислороде приводит к заключению, что прокариоты возникли в период, когда содержание кислорода в среде изменилось. Ко времени же появления эукариот концентрация кислорода была высокой и относительно постоянной.
Первые
фотосинтезирующие организмы появились
около 3 млрд. лет назад. Это были анаэробные
бактерии, предшественники современных
фотосинтезирующих бактерий. Предполагается,
что именно они образовали самые древние
среди известных строматолитов. Обеднение
среды азотистыми органическими
соединениями вызывало появление живых
существ, способных использовать
атмосферный азот. Такими организмами,
способными существовать в среде,
полностью лишенной органических
углеродистых и азотистых соединений,
являются фотосинтезирующие азотфиксирующие
сине-зеленые водоросли. Эти организмы
осуществляли аэробный фотосинтез. Они
устойчивы к продуцируемому ими кислороду
и могут использовать его для собственного
метаболизма. Поскольку сине-зеленые
водоросли возникли в период, когда
концентрация кислорода в атмосфере
колебалась, вполне допустимо, что они
— промежуточные организмы между
анаэробами и аэробами.
Такими организмами,
способными существовать в среде,
полностью лишенной органических
углеродистых и азотистых соединений,
являются фотосинтезирующие азотфиксирующие
сине-зеленые водоросли. Эти организмы
осуществляли аэробный фотосинтез. Они
устойчивы к продуцируемому ими кислороду
и могут использовать его для собственного
метаболизма. Поскольку сине-зеленые
водоросли возникли в период, когда
концентрация кислорода в атмосфере
колебалась, вполне допустимо, что они
— промежуточные организмы между
анаэробами и аэробами.
С уверенностью предполагается, что фотосинтез, в котором источником атомов водорода для восстановления углекислого газа является сероводород (такой фотосинтез осуществляют современные зеленые и пурпурные серные бактерии), предшествовал более сложному двустадийному фотосинтезу, при котором атомы водорода извлекаются из молекул воды. Второй тип фотосинтеза характерен для цианей и зеленых растений.
Фотосинтезирующая
деятельность первичных одноклеточных
имела три последствия, оказавшие решающее
влияние на всю дальнейшую эволюцию
живого.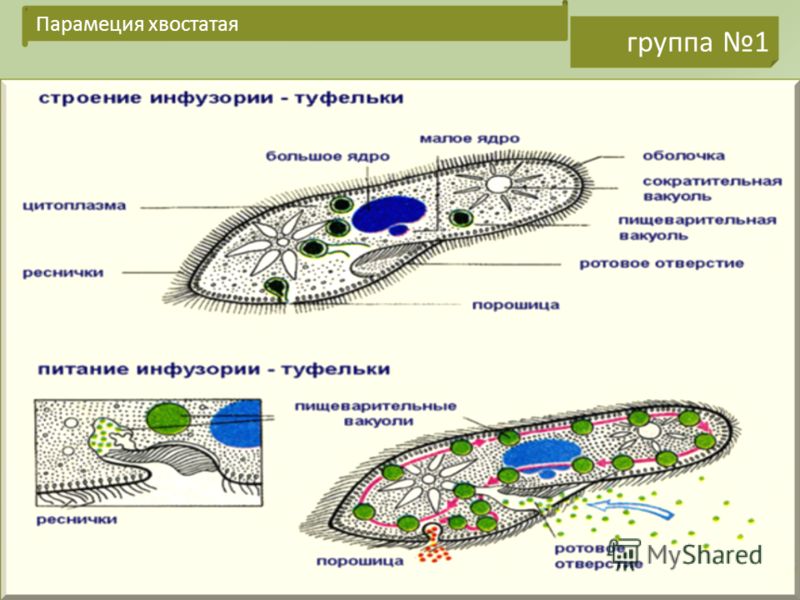 Во-первых, фотосинтез освободил
организмы от конкуренции за природные
запасы абиогенных органических
соединений, количество которых в среде
значительно сократилось. Развившееся
посредством фотосинтеза автотрофное
питание и запасание питательных готовых
веществ в растительных тканях создали
затем условия для появления громадного
разнообразия автотрофных и гетеротрофных
организмов. Во-вторых, фотосинтез
обеспечивал насыщение атмосферы
достаточным количеством кислорода для
возникновения и развития организмов,
энергетический обмен которых основан
на процессах дыхания. В-третьих, в
результате фотосинтеза в верхней части
атмосферы образовался озоновый экран,
защищающий земную жизнь от губительного
ультрафиолетового излучения космоса,
Во-первых, фотосинтез освободил
организмы от конкуренции за природные
запасы абиогенных органических
соединений, количество которых в среде
значительно сократилось. Развившееся
посредством фотосинтеза автотрофное
питание и запасание питательных готовых
веществ в растительных тканях создали
затем условия для появления громадного
разнообразия автотрофных и гетеротрофных
организмов. Во-вторых, фотосинтез
обеспечивал насыщение атмосферы
достаточным количеством кислорода для
возникновения и развития организмов,
энергетический обмен которых основан
на процессах дыхания. В-третьих, в
результате фотосинтеза в верхней части
атмосферы образовался озоновый экран,
защищающий земную жизнь от губительного
ультрафиолетового излучения космоса,
Еще
одно существенное отличие прокариот и
эукариот заключается в том, что у вторых
центральным механизмом обмена является
дыхание, у большинства же прокариот
энергетический обмен осуществляется
в процессах брожения. Сравнение
метаболизма прокариот и эукариот
приводит к выводу об эволюционной связи
между ними. Вероятно, анаэробное брожение
возникло на более ранних стадиях
эволюции. После появления в атмосфере
достаточного количества свободного
кислорода аэробный метаболизм оказался
намного выгоднее, так как при окислении
углеводов в 18 раз увеличивается выход
биологически полезной энергии в сравнении
с брожением. Таким образом, к анаэробному
метаболизму присоединился аэробный
способ извлечения энергии одноклеточными
организмами.
Вероятно, анаэробное брожение
возникло на более ранних стадиях
эволюции. После появления в атмосфере
достаточного количества свободного
кислорода аэробный метаболизм оказался
намного выгоднее, так как при окислении
углеводов в 18 раз увеличивается выход
биологически полезной энергии в сравнении
с брожением. Таким образом, к анаэробному
метаболизму присоединился аэробный
способ извлечения энергии одноклеточными
организмами.
Когда же появились эукариотические клетки? На этот вопрос нет точного ответа, но значительное количество данных об ископаемых эукариотах позволяет сказать, что их возраст составляет около 1,5 млрд. лет. Относительно того, каким образом возникли эукариоты, существуют две гипотезы.
Одна
из них (аутогенная гипотеза) предполагает,
что эукариотическая клетка возникла
путем дифференциации исходной
прокариотической клетки. Вначале
развился мембранный комплекс: образовалась
наружная клеточная мембрана с впячиваниями
внутрь клетки, из которой сформировались
отдельные структуры, давшие начало
клеточным органоидам. От какой именно
группы прокариот возникли эукариоты,
сказать невозможно.
От какой именно
группы прокариот возникли эукариоты,
сказать невозможно.
Другую
гипотезу (симбиотическую) предложила
недавно американский ученый Маргулис.
В ее обоснование она положила новые
открытия, в частности обнаружение у
пластид и митохондрий внеядерной ДНК
и способности этих органелл к
самостоятельному делению. Л. Маргулис
предполагает, что эукарио-тическая
клетка возникла вследствие нескольких
актов симбиогенеза. Вначале произошло
объединение крупной амебовидной
прокариотной клетки с мелкими аэробными
бактериями, которые превратились в
митохондрии. Затем эта симбиотическая
прокариотная клетка включила в себя
спирохетоподобные бактерии, из которых
сформировались кинетосомы, центросомы
и жгутики. После обособления ядра в
цитоплазме (признак эукариот) клетка с
этим набором органелл оказалась исходной
для образования царств грибов и животных.
Объединение прокариотной клетки с
цианеями привело к образованию пластидной
клетки, что дало начало формированию
царства растений. Гипотеза Маргулис
разделяется не всеми и подвергается
критике.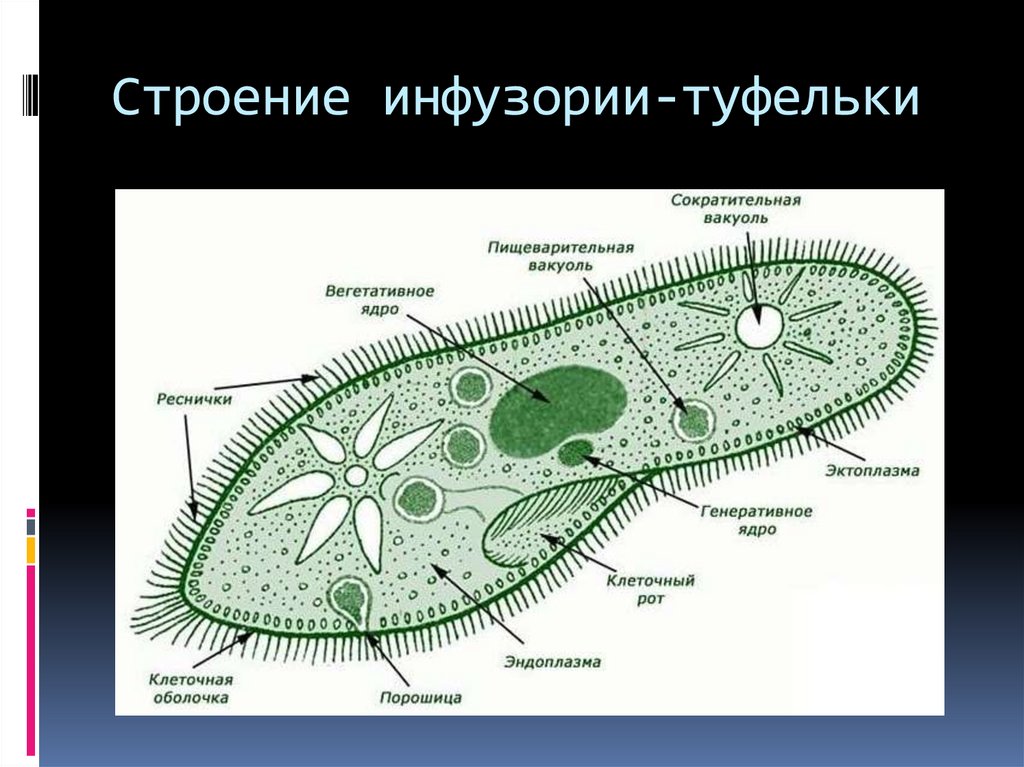 Большинство авторов придерживается
аутогенной гипотезы, более соответствующей
дарвиновским принципам монофилии,
дифференциации и усложнения организации
в ходе прогрессивной эволюции.
Большинство авторов придерживается
аутогенной гипотезы, более соответствующей
дарвиновским принципам монофилии,
дифференциации и усложнения организации
в ходе прогрессивной эволюции.
В эволюции одноклеточной организации выделяются промежуточные ступени, связанные с усложнением строения организма, совершенствованием генетического аппарата и способов размножения.
Самая примитивная стадия — агамная прокариотная — представлена цианеями и бактериями. Морфология этих организмов наиболее проста в сравнении с другими одноклеточными (простейшими). Однако уже на этой стадии появляется дифференциация на цитоплазму, ядерные элементы, базальные зерна, цитоплазматическую мембрану. У бактерий известен обмен генетическим материалом посредством конъюгации. Большое разнообразие видов бактерий, способность существовать в самых разных условиях среды свидетельствуют о высокой адаптивности их организации.
Следующая
стадия — агамная эукариотная —
характеризуется дальнейшей дифференциацией
внутреннего строения с формированием
высокоспециализированных органоидов
(мембраны, ядро, цитоплазма, рибосомы,
митохондрии и др.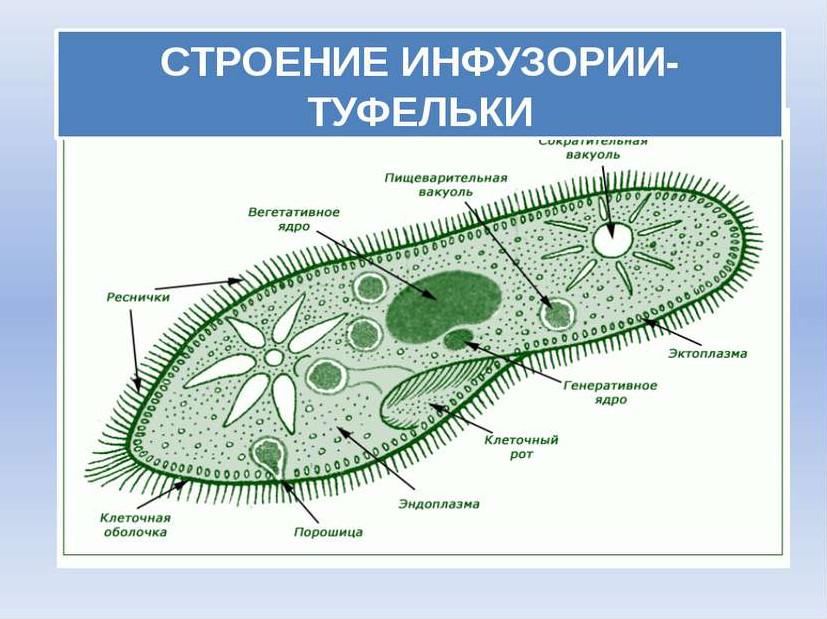 ). Особо существенной
здесь была эволюция ядерного аппарата
— образование настоящих хромосом в
сравнении с прокариотами, у которых
наследственное вещество диффузно
распределено по всей клетке. Эта стадия
характерна для простейших, прогрессивная
эволюция которых шла по пути увеличения
числа одинаковых органоидов (полимеризация),
увеличения числа хромосом в ядре
(полиплоидизация), появления генеративных
и вегетативных ядер — макронуклеуса и
микронуклеуса (ядерный дуализм). Среди
одноклеточных эукариотных организмов
имеется много видов с агамным размножением
(голые амебы, раковинные корненожки,
жгутиконосцы).
). Особо существенной
здесь была эволюция ядерного аппарата
— образование настоящих хромосом в
сравнении с прокариотами, у которых
наследственное вещество диффузно
распределено по всей клетке. Эта стадия
характерна для простейших, прогрессивная
эволюция которых шла по пути увеличения
числа одинаковых органоидов (полимеризация),
увеличения числа хромосом в ядре
(полиплоидизация), появления генеративных
и вегетативных ядер — макронуклеуса и
микронуклеуса (ядерный дуализм). Среди
одноклеточных эукариотных организмов
имеется много видов с агамным размножением
(голые амебы, раковинные корненожки,
жгутиконосцы).
Прогрессивным
явлением в филогенезе простейших было
возникновение у них полового размножения
(гамогонии), которое отличается от
обычной конъюгации. У простейших имеется
мейоз с двумя делениями и кроссинговером
на уровне хроматид, и образуются гаметы
с гаплоидным набором хромосом. У некоторых
жгутиковых гаметы почти неотличимы от
бесполых особей и нет еще разделения
на мужские и женские гаметы, т.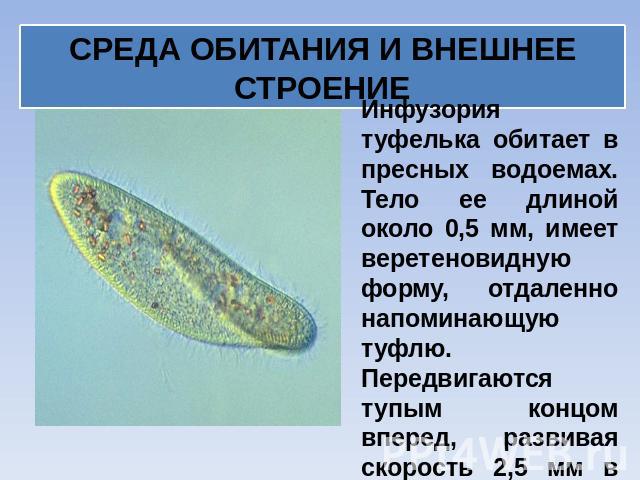 е.
наблюдается изогамия. Постепенно в ходе
прогрессивной эволюции происходит
переход от изогамии к анизогамии, или
разделению генеративных клеток на
женские и мужские, и к анизогамной
копуляции. При слиянии гамет образуется
диплоидная зигота. Следовательно, у
простейших наметился переход от агамной
эукариотной стадии к зиготной — начальной
стадии ксеногамии (размножение путем
перекрестного оплодотворения). Последующее
развитие уже многоклеточных организмов
шло по пути совершенствования способов
ксеногамного размножения.
е.
наблюдается изогамия. Постепенно в ходе
прогрессивной эволюции происходит
переход от изогамии к анизогамии, или
разделению генеративных клеток на
женские и мужские, и к анизогамной
копуляции. При слиянии гамет образуется
диплоидная зигота. Следовательно, у
простейших наметился переход от агамной
эукариотной стадии к зиготной — начальной
стадии ксеногамии (размножение путем
перекрестного оплодотворения). Последующее
развитие уже многоклеточных организмов
шло по пути совершенствования способов
ксеногамного размножения.
Животные, состоящие из единственной клетки, располагающей ядром, называются одноклеточными организмами.
В них сочетаются характерные особенности клетки и независимого организма.
Одноклеточные животные
Животные подцарства Одноклеточных или Простейших обитают в жидких средах. Внешние формы их разнообразны — от аморфных особей, не имеющих определенных очертаний, до представителей со сложными геометрическими формами.
Насчитывается около 40 тысяч видов одноклеточных животных.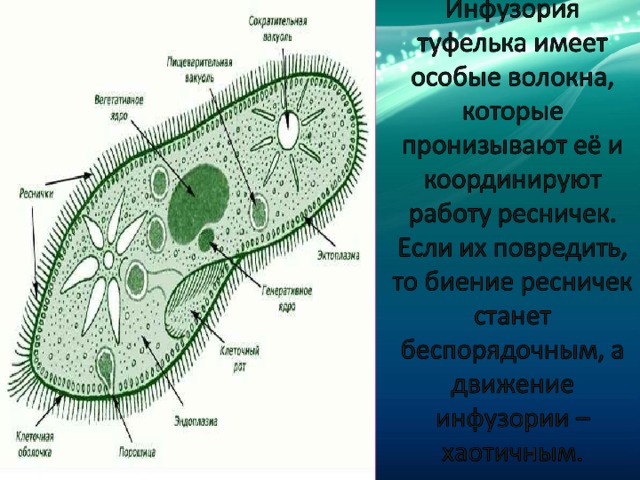 К наиболее известным относятся:
К наиболее известным относятся:
- амеба;
- зеленая эвглена;
- инфузория-туфелька.
Амеба
Принадлежит классу корненожки и отличается непостоянной формой.
Она состоит из оболочки, цитоплазмы, сократительной вакуоли и ядра.
Усвоение питательных веществ осуществляется с помощью пищеварительной вакуоли, а кормом служат другие простейшие, такие как водоросли и . Для респирации амебе необходим кислород, растворенный в воде и проникающий через поверхность тела.
Зеленая эвглена
Обладает вытянутой веерообразной формой. Питается за счет превращения углекислого газа и воды в кислород и продукты питания благодаря световой энергии, а также готовыми органическими веществами при отсутствии света.
Относится к классу жгутиковые.
Инфузория-туфелька
Класс инфузории, своими очертаниями напоминает туфельку.
Пищей служат бактерии.
Одноклеточные грибы
Грибы отнесены к низшим бесхлорофилльным эукариотам.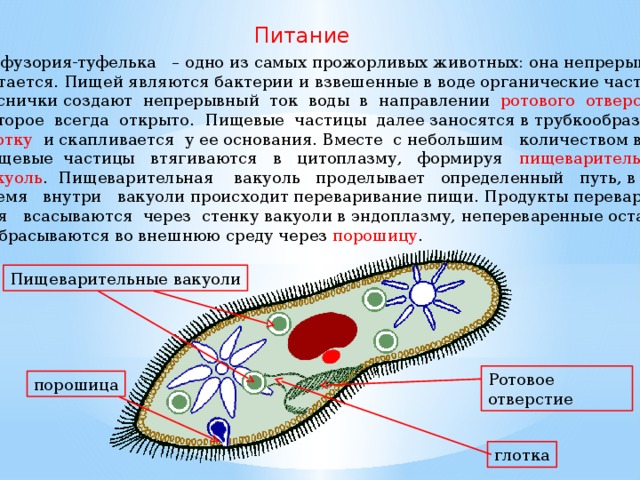 Они отличаются наружным пищеварением и содержанием хитина в клеточной стенке. Тело образует грибницу, состоящую из гифов.
Они отличаются наружным пищеварением и содержанием хитина в клеточной стенке. Тело образует грибницу, состоящую из гифов.
Одноклеточные грибы систематизированы в 4 основных классах:
- дейтеромицеты;
- хитридиомицеты;
- зигомицеты;
- аскомицеты.
Ярким примером аскомицетов служат дрожжи, широко распространенные в природе. Скорость их роста и размножения велика благодаря особенному строению. Дрожжи состоят из одиночной клетки округлой формы, размножающейся почкованием.
Одноклеточные растения
Типичным представителем низших одноклеточных растений, часто встречающихся в природе, являются водоросли:
- хламидомонада;
- хлорелла;
- спирогира;
- хлорококк;
- вольвокс.
Хламидомонада отличается от всех водорослей подвижностью и наличием светочувствительного глазка, определяющего места наибольшего скопления солнечной энергии для фотосинтеза .
Многочисленные хлоропласты заменены одним большим хроматофором. Роль насосов, откачивающих излишки жидкости, выполняют сократительные вакуоли. Передвижение осуществляется при помощи двух жгутиков.
Роль насосов, откачивающих излишки жидкости, выполняют сократительные вакуоли. Передвижение осуществляется при помощи двух жгутиков.
Зеленые водоросли хлореллы, в отличие от хламидомонады, обладают типичными растительными клетками. Плотная оболочка защищает мембрану, а в цитоплазме расположено ядро и хроматофор. Функции хроматофора сходны с ролью хлоропласт наземных растений.
С хлореллой схожа водоросль шарообразной формы хлорококк. Местом ее обитания служит не только вода, но и суша, стволы деревьев, растущих во влажной среде.
Кто открыл одноклеточные организмы
Честь открытия микроорганизмов принадлежит голландскому ученому А. Левенгуку.
В 1675 году он разглядел их в микроскоп собственного изготовления. За мельчайшими существами закрепилось название инфузория, а с 1820 года их стали называть простейшими животными.
Зоологами Келлекером и Зибольдом в 1845 году одноклеточные были отнесены к особому типу животного царства и разделены на две группы:
- корненожки;
- инфузории.
Как выглядит клетка одноклеточного животного
Строение одноклеточных организмов возможно изучить лишь с помощью микроскопа. Тело простейших существ состоит из единственной клетки, выполняющей роль независимого организма.
В состав клетки входят:
- цитоплазма;
- органоиды;
- ядро.
Со временем, в результате приспособления к окружающей среде, у отдельных видов одноклеточных появились специальные органоиды движения, выделения и питания.
Кто такие простейшие
Современная биология относит простейших к парафилетической группе животноподобных протистов. Наличие в клетке ядра, в отличие от бактерий, включает их в список эукариотов.
Клеточные структуры разнятся с клетками многоклеточных. В живой системе простейших присутствуют пищеварительные и сократительные вакуоли, у некоторых наблюдаются схожие с ротовой полостью и анальным отверстием органеллы.
Классы простейших
В современной классификации по признакам отсутствует отдельный ранг и значение одноклеточных.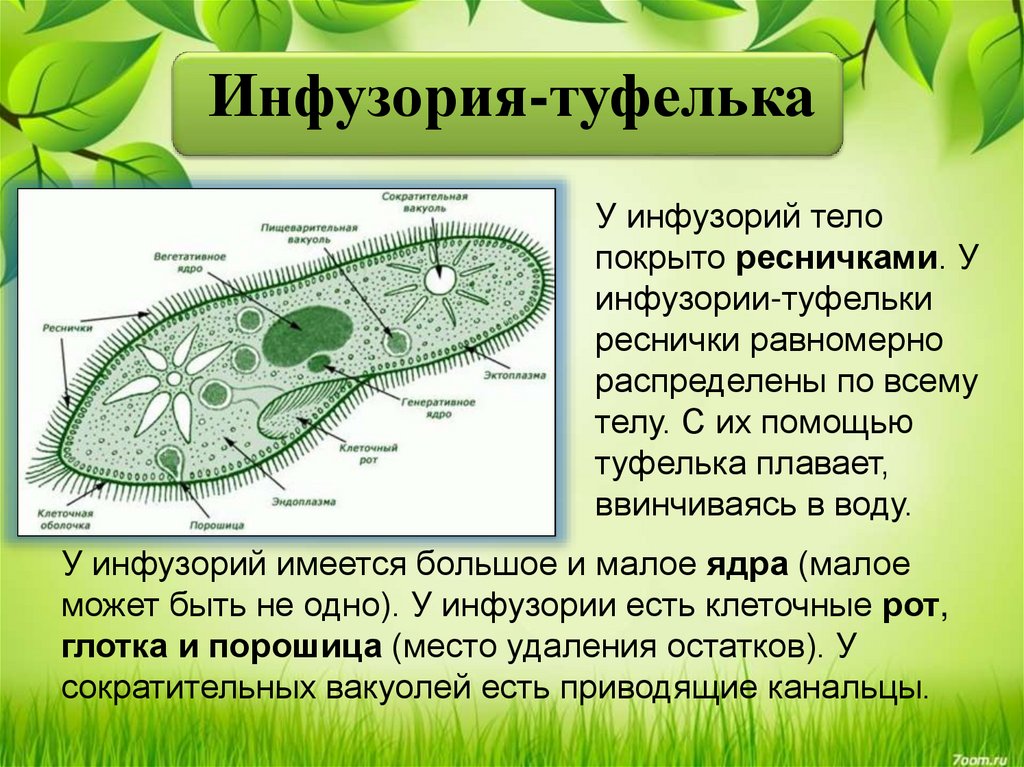
Лабиринтула
Их принято подразделять на следующие типы:
- саркомастигофоры;
- апикомплексы;
- миксоспоридии;
- инфузории;
- лабиринтулы;
- асцестоспородии.
Устаревшей классификацией считается деление простейших на жгутиковых, саркодовых, ресничных и споровиков.
В каких средах обитают одноклеточные
Средой обитания простейших одноклеточных служит любая влажная среда. Амеба обыкновенная, эвглена зеленая и инфузория-туфелька являются типичными обитателями загрязненных пресных водных источников.
Наука долгое время относила опалин к инфузориям, благодаря внешнему сходству жгутиков с ресничками и наличию двух ядер. В результате тщательных исследований родство было опровергнуто. Половое размножение опалин происходит в результате копуляции, ядра одинаковые, а ресничный аппарат отсутствует.
Заключение
Биологическую систему невозможно представить без одноклеточных организмов, являющихся источником питания других животных.
Простейшие организмы способствуют образованию горных пород, служат показателями загрязненности водоемов, участвуют в круговороте углерода . Широкое применение микроорганизмы нашли в биотехнологиях.
Большая энциклопедия школьника
Большая энциклопедия школьникауникальное издание, содержащее весь свод знаний, необходимый ученикам младших классов. Для детей, собирающихся в 1-й класс, она послужит незаменимым помощником для подготовки к школе. В этой энциклопедии ребенок сможет найти любую интересующую его информацию, в понятном и простом для него изложении. Вы подбираете слова и определения для простых вещей, которые надо объяснить ребенку? Сомневаетесь в формулировках? Просто возьмите «Большую энциклопедию школьника» и найдите нужный ответ вместе с малышом!
Математика в стихах
Развитие речи
Азбука в картинках
Игры на развитие внимания
Как правильно выбрать школу
Ваш ребенок левша
Как готовить домашнее задание
Контрольные и экзамены
Большая энциклопедия школьника — это твой надёжный путеводитель в мире знаний. Она проведёт сквозь извилистые лабиринты наук и раскроет завесу великих тайн Вселенной.
С ней ты поднимешься высоко к звёздам и опустишься на дно самых глубоких морей,
ты научишься видеть мельчайшие организмы и осязать огромные пространства Земли.
Отправившись в это увлекательное путешествие, ты значительно расширишь свой кругозор и поднимешься
на новую ступень развития. Отныне никакие вопросы учителей не смогут поставить тебя в тупик,
ты сможешь найти выход из любой ситуации. Мир знаний зовёт тебя. В добрый путь!
Она проведёт сквозь извилистые лабиринты наук и раскроет завесу великих тайн Вселенной.
С ней ты поднимешься высоко к звёздам и опустишься на дно самых глубоких морей,
ты научишься видеть мельчайшие организмы и осязать огромные пространства Земли.
Отправившись в это увлекательное путешествие, ты значительно расширишь свой кругозор и поднимешься
на новую ступень развития. Отныне никакие вопросы учителей не смогут поставить тебя в тупик,
ты сможешь найти выход из любой ситуации. Мир знаний зовёт тебя. В добрый путь!
| Ребенок не хочет учить буквы Ребенок не хочет учить буквы
— Понимаете, ведь надо что-то делать! — с тревогой говорила мне полная, хорошо одетая дама, едва умещающаяся на стуле. Ее ноги в аккуратных лодочках были плотно сжаты (юбка до середины колена казалась слегка коротковатой для такой монументальной фигуры), руки сложены на коленях. | Past continuous passive Страдательный залог образуется с помощью вспомогательного глагола ‘to be’. Страдательный залог глагола ‘to repair’ в группе ‘continuous’ : To be repaired = Быть исправленным. The road is being repaired = Дорогу чинят. The road is not being repaired = Дорогу не чинят. Is the road being repaired? = Чинят ли дорогу? The road was being repaired = Дорогу чинили. The road was not being repaired = Дорогу не чинили. Was the road being repaired? = Чинили ли дорогу? Страдательный … |
| Определение формулы органического вещества по его молярной массе Задание: Определить формулу углеводорода, если его молярная масса равна 78 г. | У У
ЗВУК (У).
1) Удобная буква!
Удобно в ней то,
Что можно на букву
Повесить пальто.
У – сучок,
В любом лесу
Ты увидишь букву У.
2) ФОНЕТИЧЕСКАЯ ЗАРЯДКА.
— Как воет волк! ( у – у – у )
3) ЗАДАНИЯ.
а) Подними руку, если услышишь звук (у):
паук, цветок, лужа, диван, стол, стул, голуби, курица. |
 — Ей же на тот год в школу, все ее сверстники уже читают, а она даже буквы …
— Ей же на тот год в школу, все ее сверстники уже читают, а она даже буквы …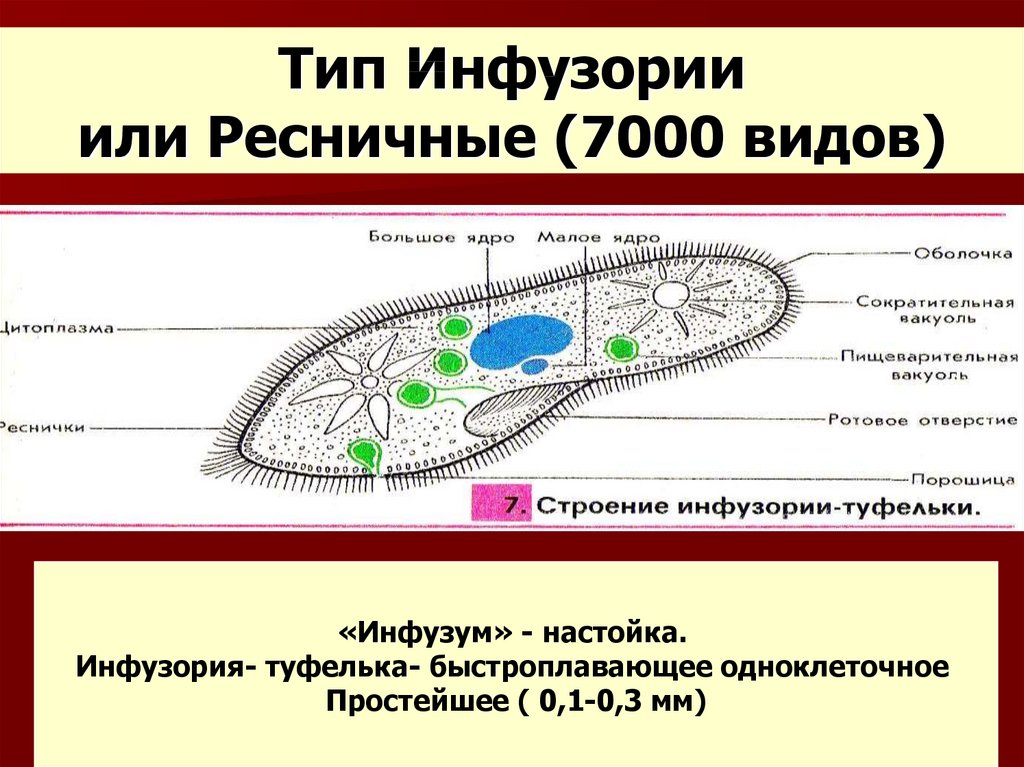 № п/п
Последовательность действий
Выполнение действий
1.
Записать общую формулу углеводорода.
Общая формула углеводорода СхНу
2.
Найти молярную массу углеводорода в общем виде.
М(СхНу)=12х +у
3.
Приравнять найденное в общем виде значение молярной массы к данному в …
№ п/п
Последовательность действий
Выполнение действий
1.
Записать общую формулу углеводорода.
Общая формула углеводорода СхНу
2.
Найти молярную массу углеводорода в общем виде.
М(СхНу)=12х +у
3.
Приравнять найденное в общем виде значение молярной массы к данному в …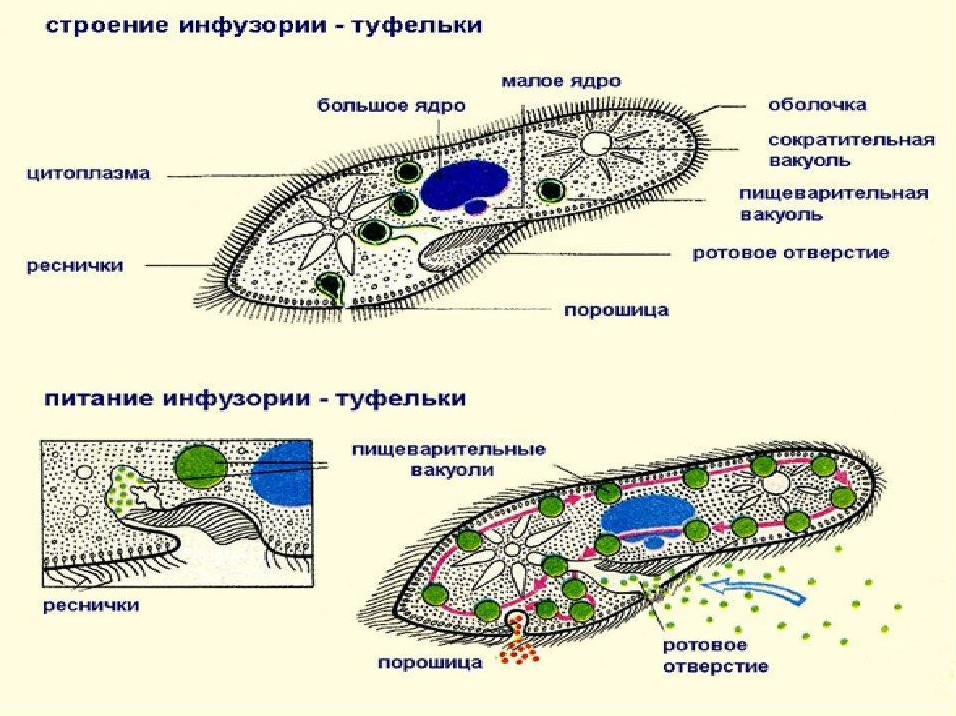 б) Где стоит (у)?
Зубы, утка, наука, кенгуру …
б) Где стоит (у)?
Зубы, утка, наука, кенгуру …Морфологическая и молекулярная филогения дилептидных и трахелиидных инфузорий: Разрешение на основании класса Litostomatea (Ciliophora, Rhynchostomatia)
Бергер Х. Монография Urostyloidea (Ciliophora, Hypotricha) Monogr. биол. 2006; 85:1–1304. [Google Scholar]
Claparède É., Lachmann J. Исследования инфузорий и корненожек. Мем. Инст. физ. женев. 1859; 6: 261–482. [Google Scholar]
Корлисс Дж.О. 2-е изд. Пергамон Пресс; Оксфорд, Нью-Йорк, Торонто, Сидней, Париж, Франкфурт: 1979. Ресничные простейшие. Характеристика, классификация и справочник по литературе. [Google Scholar]
Драгеско Дж. Захват и проглатывание продуктов из инфузорий. Бык. биол. о. бельг. 1962; 96: 123–167. [Google Scholar]
Dragesco J. Обзор жанра Dileptus , Dujardin 1871 (Ciliata Holotricha) (систематическая, цитологическая, биологическая) Bull. биол. о. бельг. 1963; 97: 103–145. [Google Scholar]
бельг. 1963; 97: 103–145. [Google Scholar]
Dragesco J., Métain C. La Capitol des proies chez Dileptus gigas (Cilié Holotrice) Bull. соц. Зоол. о. 1948; 73: 62–65. [Google Scholar]
Дюжарден Ф. Энциклопедическая библиотека Роре; Париж: 1841 г. Histoire naturelle des zoophytes. Infusoires, comprenant la physiologie et la classification de ces animaux et la maniére de les etudier a l’aide du микроскоп. [Google Scholar]
Эренберг К.Г. Верлаг фон Леопольд Фосс; Лейпциг: 1838 г. Die Infusionsthierchen als vollkommene Organismen. Ein Blick in das tiefere organische Leben der Natur. [Академия Google]
Элвуд Х.Дж., Олсен Г.Дж., Согин М.Л. Последовательности гена малой субъединицы рибосомной РНК инфузории гипотрихоза Oxytrichanova и Stylonychia pustulata . Мол. биол. Эвол. 1985; 2: 399–410. [PubMed] [Google Scholar]
Фойсснер В. Infraciliatur, Silberliniensystem und Biometrie einiger neuer und wenig bekannter terrestrischer, limnischer und Mariner Ciliaten (Protozoa: Ciliophora) aus den Klassen Kinetofragminophora, Colpodea und Polyhymenophora.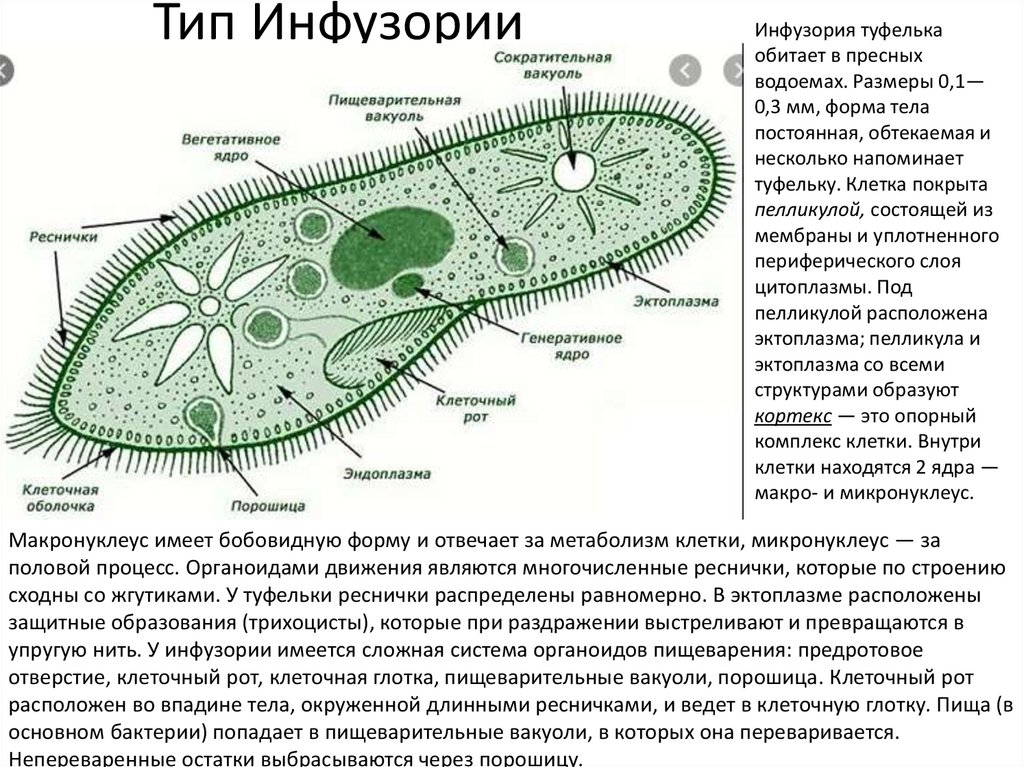 Стапфия (Линц) 1984;12:1–165. [Google Scholar]
Стапфия (Линц) 1984;12:1–165. [Google Scholar]
Фойснер В. Основные методы световой и сканирующей электронной микроскопии для таксономических исследований реснитчатых простейших. Евро. Дж. Протистол. 1991; 27: 313–330. [PubMed] [Google Scholar]
Фойснер В. Фаунистические и таксономические исследования инфузорий (Protozoa, Ciliophora) из чистых рек Баварии (Германия) с описаниями новых видов и экологическими заметками. Лимнологика. 1997; 27: 179–238. [Google Scholar]
Фойсснер В. Обновленный сборник мировых почвенных инфузорий (Protozoa, Ciliophora) с экологическими примечаниями, новыми записями и описаниями новых видов. Евро. Дж. Протистол. 1998;34:195–235. [Google Scholar]
Foissner W. Два новых наземных спатидиевых (Protozoa, Ciliophora) из тропической Африки: Arcuospathidium vlassaki и Arcuospathidium bulli . биол. Плодородный. Почвы. 2000; 30: 469–477. [Google Scholar]
Фойснер В. Два замечательных почвенных спатидиевых (Ciliophora: Haptorida), Arcuospathidium pachyoplites sp.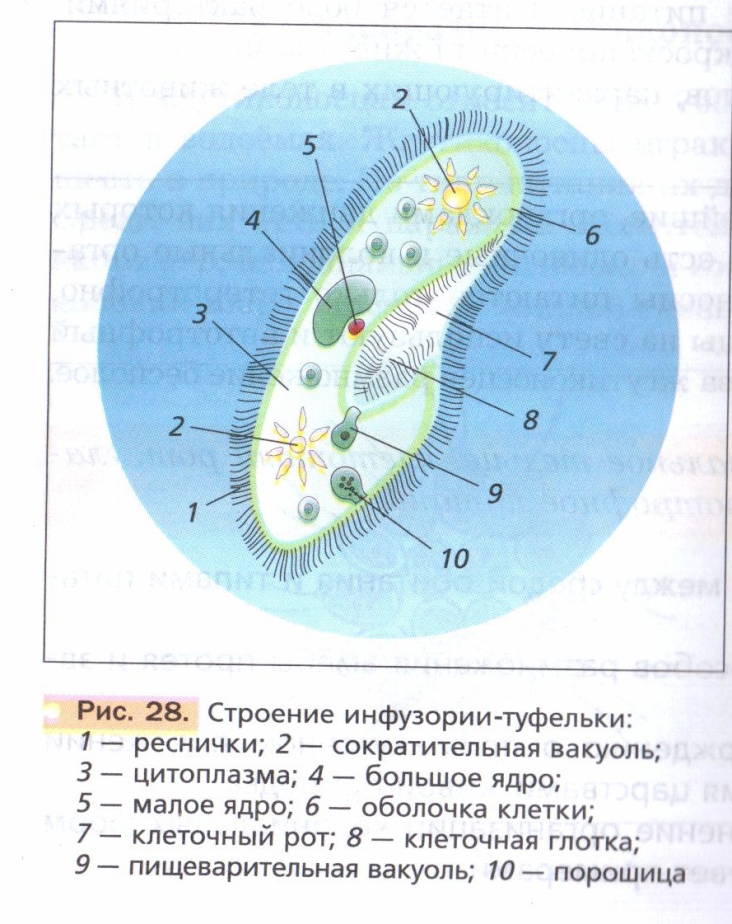 н. и Spathidium faurefremieti ном. н. Акта Протозол. 2003; 42: 145–159. [Академия Google]
н. и Spathidium faurefremieti ном. н. Акта Протозол. 2003; 42: 145–159. [Академия Google]
Фойснер В., Фойснер И. Оральные монокинетиды у свободноживущих гапторовых инфузорий Enchelydium polynucleatum (Ciliophora, Enchelyidae): ультраструктурные данные и филогенетические последствия. Дж. Протозол. 1985; 32: 712–722. [Google Scholar]
Foissner W., Foissner I. Тонкое строение Fuscheria terricola Berger et al., 1983 и предлагаемая новая классификация подкласса Haptoria Corliss, 1974 (Ciliophora, Litostomatea) Arch. Протистенк. 1988; 135: 213–235. [Академия Google]
Фойсснер В., Бергер Х., Блаттерер Х., Кохманн Ф. Таксономическая и экологическая ревизия Ciliaten des Saprobiensystems – Band IV: Gymnostomatea, Loxodes . Суктория. Informationsberichte де Байер. Landesamtes für Wasserwirtschaft. 1995;1(95):1–540. [Google Scholar]
Фойсснер В., Бергер Х., Шаумбург Дж. Идентификация и экология пресноводных планктонных инфузорий.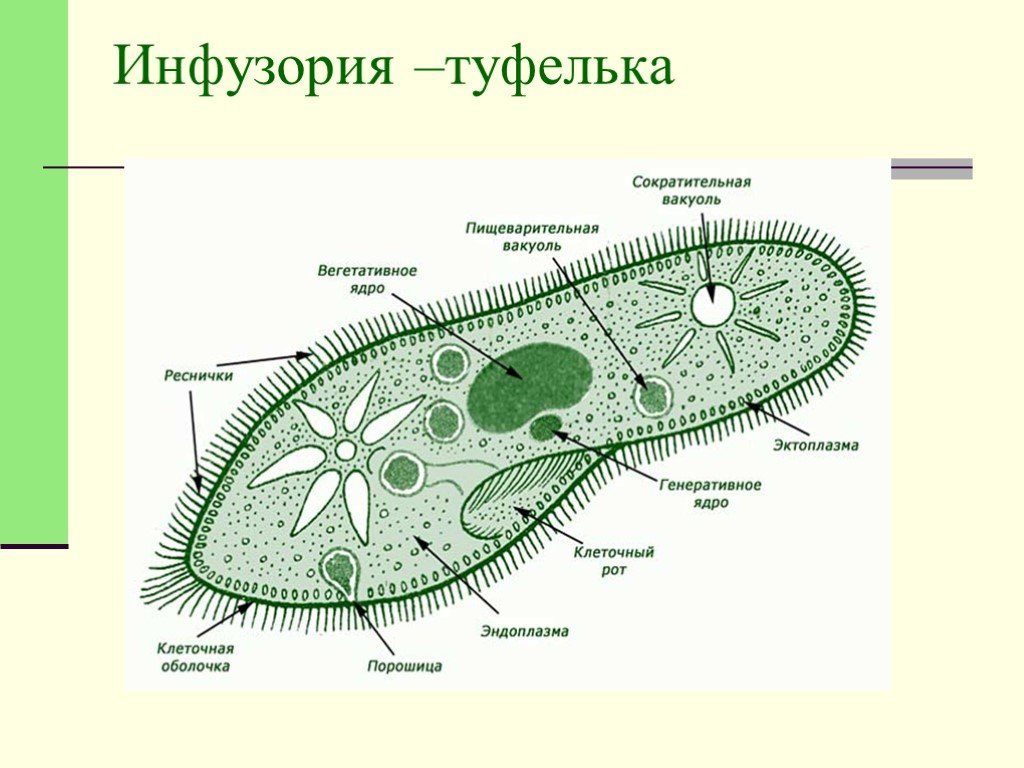 Informationsberichte де Байер. Landesamtes für Wasserwirtschaft. 1999;3(99):1–793. [Академия Google]
Informationsberichte де Байер. Landesamtes für Wasserwirtschaft. 1999;3(99):1–793. [Академия Google]
Фойсснер В., Агата С., Бергер Х. Почвенные инфузории (Protozoa, Ciliophora) из Намибии (Юго-Западная Африка) с акцентом на двух контрастирующих средах: регионе Этоша и пустыне Намиб. Денисия. 2002; 5:1–1459. [Google Scholar]
Габилондо Р., Фойсснер В. Четыре новых фушериевых почвенных инфузории (Ciliophora: Haptorida) из четырех биогеографических регионов. Акта Протозол. 2009; 48:1–24. [Google Scholar]
Gao S., Song W., Ma H., Clamp J.C., Yi Z., Al-Rasheid K.A.S., Chen Z., Lin X. Филогения шести родов подкласса Haptoria (Ciliophora, Litostomatea) выведено из последовательностей гена, кодирующего малую субъединицу рибосомной РНК. Дж. Эукариот. микробиол. 2008; 55: 562–566. [PubMed] [Академия Google]
Голиньская К. Макронуклеус в Dileptus cygnus и его изменения в делении. Акта Протозол. 1965; 3: 143–151. [Google Scholar]
Голиньская К. Сравнительные исследования морфологии Dileptus anatinus sp. н. (Holotricha, Gymnostomata) Acta Protozool. 1971; 8: 367–378. [Google Scholar]
н. (Holotricha, Gymnostomata) Acta Protozool. 1971; 8: 367–378. [Google Scholar]
Golińska K. Влияние пуромицина на процессы регенерации у Dileptus anatinus , Golinska 1971. Acta Protozool. 1974; 12: 289–306. [Академия Google]
Golińska K. Курс ремоделирования in situ поврежденного ротового аппарата у Dileptus (Ciliata, Gymnostomata) Acta Protozool. 1978; 17: 47–67. [Google Scholar]
Голиньская К. Регуляция цилиарного рисунка у Dileptus (Ciliata). I. Чувствительные реснички и их превращение в локомоторные реснички. Дж. Эмбриол. Эксп. Превращаться. 1982; 68: 99–114. [PubMed] [Google Scholar]
Golińska K. Регуляция цилиарного рисунка у Dileptus (Ciliata). II. Формирование коркового домена сенсорных ресничек из домена локомоторных ресничек. Дж. Клеточные науки. 1983;62:459–475. [PubMed] [Google Scholar]
Golińska K. Комплексы кортикальных органелл, их структура, формирование и влияние на форму клетки у инфузории, Dileptus . Протоплазма. 1991; 162: 160–174. [Google Scholar]
Протоплазма. 1991; 162: 160–174. [Google Scholar]
Голиньская К. Формирование и ориентация элементов скелета при развитии ротовой области у инфузории, Dileptus . Акта Протозол. 1995; 34: 101–113. [Google Scholar]
Голиньская К., Афонькин С.Ю. Подготовительные изменения и развитие конъюгационного перехода у инфузории, Дилептус . Протоплазма. 1993; 173: 144–157. [Google Scholar]
Grain J., Golińska K. Структура и ультраструктура Dileptus cygnus Claparède et Lachmann, 1859, Cilié Holotrice Gymnostome. Протистология. 1969; 5: 269–291. [Google Scholar]
Холл Т.А. BioEdit: удобный редактор выравнивания биологических последовательностей и программа анализа для Windows 95/98/NT. Симптом нуклеиновых кислот. сер. 1999;41:95–98. [Google Scholar]
Хейс М.Л. Цитологическое исследование на Diлептус anser . Транс. Являюсь. микроск. соц. 1938; 57: 11–25. [Google Scholar]
Hennig W. University of Illinois Press; Урбана, Чикаго, Лондон: 1966. Филогенетическая систематика. [Google Scholar]
Филогенетическая систематика. [Google Scholar]
Хёрандль Э. Парафилетические и монофилетические таксоны — эволюционные и кладистические классификации. Таксон. 2006; 55: 564–570. [Google Scholar]
Хёрандл Э., Стюесси Т.Ф. Парафилетические группы как естественные единицы биологической классификации. Таксон. 2010;59:1641–1653. [Академия Google]
Международная комиссия по зоологической номенклатуре (ICZN) 4-е изд. Международный фонд зоологической номенклатуры; Лондон: 1999. Международный кодекс зоологической номенклатуры. [Google Scholar]
Янковский А.В. Новые роды классов Gymnostomatea и Ciliostomea [Новые роды классов Gymnostomatea и Ciliostomea] Mater. IV. Конф. уч. Секц. зоол. (Кишинев) 1967:36. (на русском языке) [Google Scholar]
Янковский А.В. Конспект новой системы типа Ciliophora // Труды зоол. Инст. (Ленинград) 1980;94:103–121. (in Russian) [Google Scholar]
Джонс Э.Э., мл. Инцистментация, эксцистментация и ядерный цикл у инфузории Dileptus anser . Дж. Элиша Митчелл Науч. соц. 1951; 67: 205–217. [Google Scholar]
Дж. Элиша Митчелл Науч. соц. 1951; 67: 205–217. [Google Scholar]
Kahl A. Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 2. Holotricha außer den im 1. Teil behandelten Prostomata. Tierwelt Dtl. 1931; 21: 181–398. [Google Scholar]
Лейпе Д.Д., Бернхард Д., Шлегель М., Согин М.Л. Эволюция генов 16S-подобных рибосомных РНК у таксонов цилиофор Litostomatea и Phyllopharyngea. Евро. Дж. Протистол. 1994;30:354–361. [Google Scholar]
Липскомб Д.Л. Парсимония, гомология и анализ мультистатических признаков. кладистический. 1992; 8: 45–65. [Google Scholar]
Людвиг В., Странк О., Вестрам Р., Рихтер Л., Мейер Х., Ядхукумар, Бюхнер А., Лай Т., Степпи С., Джобб Г., Фёрстер В., Бретске И. ., Гербер С., Гинхарт А.В., Гросс О., Груманн С., Герман С., Йост Р., Кениг А., Лисс Т., Люссманн Р., Мэй М., Нонхофф Б., Райхель Б., Штрелов Р., Стаматакис А., Штукманн Н., Вилбиг А., Ленке М., Людвиг Т., Боде А., Шлейфер К.-Х. ARB: программная среда для данных последовательности. Нуклеиновые Кислоты Res. 2004; 32: 1363–1371. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Нуклеиновые Кислоты Res. 2004; 32: 1363–1371. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Линн Д.Х. 3-е изд. Спрингер; Дордрехт: 2008. Ресничные простейшие. Характеристика, классификация и справочник по литературе. [Google Scholar]
Линн Д.Х., Смолл Э.Б. Phylum Ciliophora Doflein, 1901. В: Ли Дж. Дж., Лидейл Г. Ф., Брэдбери П., редакторы. 2-е изд. об. 1. Аллен Пресс; Лоуренс, Канзас: 2002. стр. 371–656. (Иллюстрированный справочник по простейшим). [Google Scholar]
Медлин Л.К., Элвуд Х.Дж., Стикель С., Согин М.Л. Характеристика ферментативно амплифицированных эукариотических 16S-подобных рРНК-кодирующих областей. Ген. 1988;71:491–499. [PubMed] [Google Scholar]
Мюллер К. PRAP-вычисления поддержки Бремера для больших наборов данных. Мол. Филогенет. Эвол. 2004; 31: 780–782. [PubMed] [Google Scholar]
Мюллер, О.Ф., 1773. Vermium terrestrium et fluviatilium, seu animalium infusoriorum, helminthicorum et testaceorum, non marinorum, succincta historia.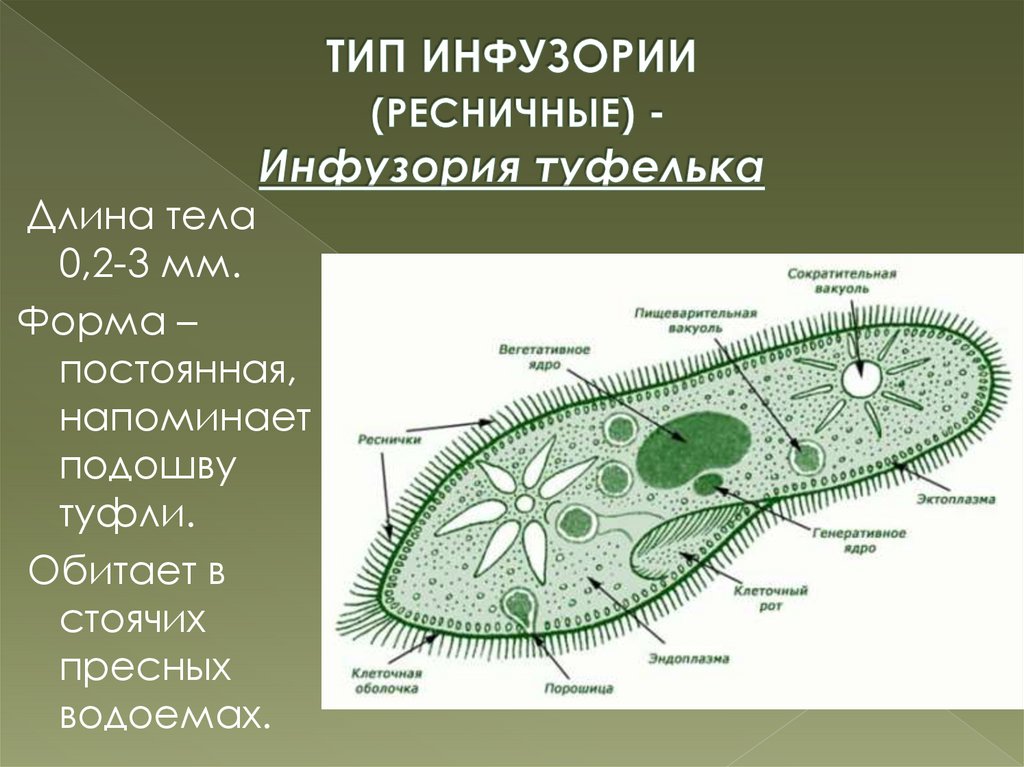 Heineck & Faber, Havniae & Lipsiae.
Heineck & Faber, Havniae & Lipsiae.
Пан Х., Гао Ф., Ли Дж., Линь С., Аль-Фаррадж С.А., Аль-Рашеид К.А.С. Морфология и филогения двух новых плевростоматидных инфузорий, Epiphyllum shenzhenense н. сп. и Loxophyllum spirellum n. сп. (Protozoa, Ciliophora) из мангровых водно-болотных угодий, Южный Китай. Дж. Эукариот. микробиол. 2010;57:421–428. [PubMed] [Google Scholar]
Penard E. Georg & Cie; Женева: 1922. Этюды сюр-ле-инфузуаров д’дьюс. [Google Scholar]
Посада Д., Крэндалл К.А. MODELTEST: тестирование модели замещения ДНК. Биоинформатика. 1998; 14:817–818. [PubMed] [Google Scholar]
Райков И.Б. Ядра инфузорий. В: Hausmann K., Bradbury BC, редакторы. Инфузории: клетки как организмы. Фишер Верлаг; Штутгарт, Йена, Нью-Йорк: 1996. стр. 221–242. [Google Scholar]
Ронквист Ф., Хюльзенбек Дж. П. MrBayes 3: Байесовский филогенетический вывод в смешанных моделях. Биоинформатика. 2003; 19: 1572–1574. [PubMed] [Google Scholar]
Schewiakoff W. Организация и систематика инфузорий Aspirotricha ( Holotricha auctorum ) [Организация и систематика инфузорий Aspirotricha ( Holotricha auctorum )] Зап. имп. акад. Наук (8e Série) 1896; 4: 1–395. (in Russian) [Google Scholar]
Организация и систематика инфузорий Aspirotricha ( Holotricha auctorum ) [Организация и систематика инфузорий Aspirotricha ( Holotricha auctorum )] Зап. имп. акад. Наук (8e Série) 1896; 4: 1–395. (in Russian) [Google Scholar]
Schmidt S.L., Foissner W., Schlegel M., Bernhard D. Молекулярная филогения Heterotrichea (Ciliophora, Postciliodesmatophora) на основе последовательностей генов малых субъединиц рРНК. Дж. Эукариот. микробиол. 2007; 54: 358–363. [PubMed] [Академия Google]
Шенборн В. А. Цимсен; Wittenberg Lutherstadt: 1966. Beschalte Amöben (Testacea) [Google Scholar]
Schrank F. von P. Bd. 3. Штейн; Landshut bei Krüll: 1803 г. (Fauna Boica. Durchgedachte Geschichte der in Baiern einheimischen und zahmen Thiere). [Google Scholar]
Шимано С., Санбе М., Касахара Ю. Связь между наблюдениями под световым микроскопом и молекулярным анализом с помощью ПЦР одиночных клеток для инфузорий. Микробы Окружающая среда. 2008; 23: 356–359. [PubMed] [Академия Google]
Song W.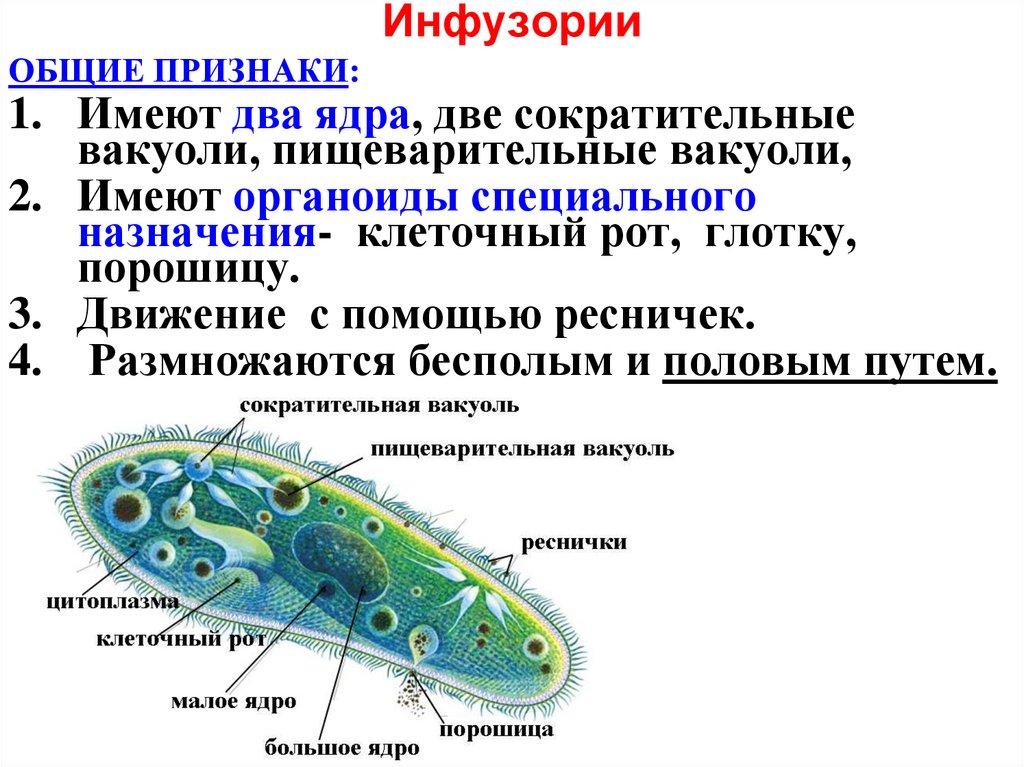 , Wilbert N. Taxonomische Untersuchungen an Aufwuchsciliaten (Protozoa, Ciliophora) im Poppelsdorfer Weiher, Bonn. Лаутерборния. 1989; 3: 2–221. [Google Scholar]
, Wilbert N. Taxonomische Untersuchungen an Aufwuchsciliaten (Protozoa, Ciliophora) im Poppelsdorfer Weiher, Bonn. Лаутерборния. 1989; 3: 2–221. [Google Scholar]
Stamatakis A., Hoover P., Rougemont J. Алгоритм быстрой загрузки для веб-серверов RAxML. Сист. биол. 2008; 75: 758–771. [PubMed] [Google Scholar]
Strüder-Kypke M.C., Lynn D.H. Анализ последовательности гена малой субъединицы рРНК подтверждает парафилию олиготриховых инфузорий sensu lato и поддерживает монофилию подклассов Oligotrichia и Choreotrichia (Ciliophora, Spirotrichea) J. Zool . (Лондон) 2003; 260:87–97. [Google Scholar]
Strüder-Kypke M.C., Wright A.-D.G., Foissner W., Chatzinotas A., Lynn D.H. Молекулярная филогения литостомовых инфузорий (Ciliophora, Litostomatea) с акцентом на свободноживущие роды гапториан. Протист. 2006; 157: 261–278. [PubMed] [Google Scholar]
Стрюдер-Кипке М.С., Корнилова О.А., Линн Д.Х. Филогения трихостомных инфузорий (Ciliophora, Litostomatea) эндосимбиотических у якутской лошади ( Equus caballus ) Eur.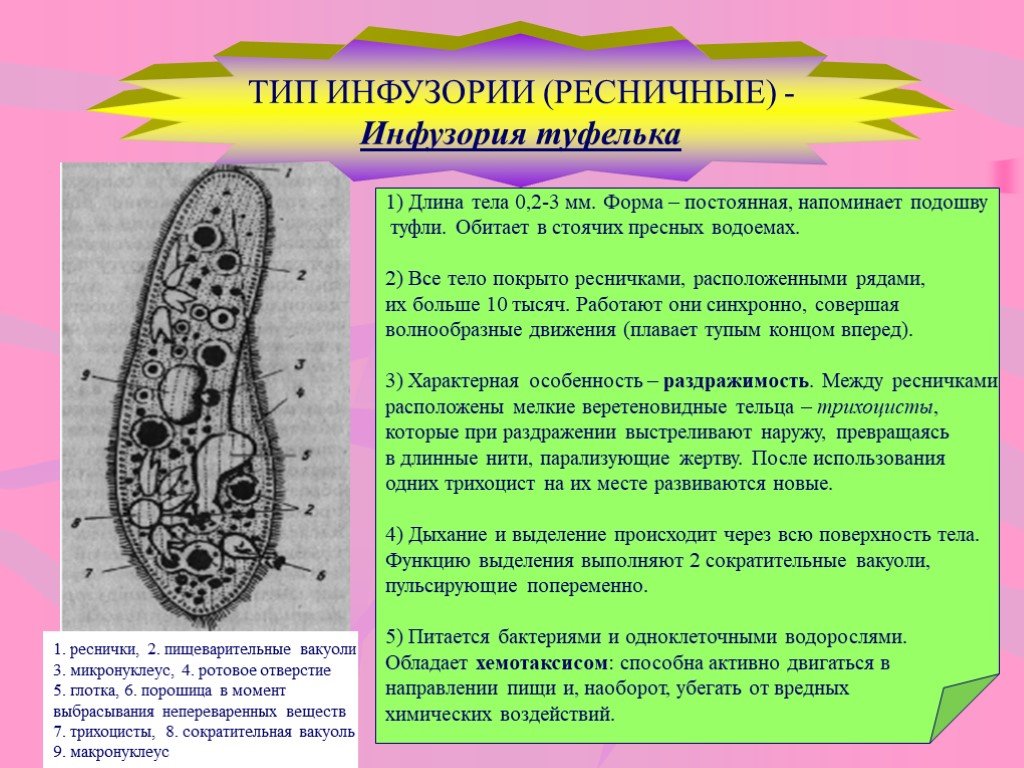 Дж. Протистол. 2007;43:319–328. [PubMed] [Академия Google]
Дж. Протистол. 2007;43:319–328. [PubMed] [Академия Google]
Своффорд Д.Л. Синауэр Ассошиэйтс, Инк.; Сандерленд, Массачусетс: 2003. PAUP*. Филогенетический анализ с использованием экономии (* и другие методы). Версия 4. [Google Scholar]
Thamm M., Schmidt S.L., Bernhard D. Взгляд на филогению рода Stentor (Heterotrichea, Ciliophora) с особым акцентом на эволюцию макронуклеуса на основе данных рДНК SSU. Акта Протозол. 2010;49:149–157. [Google Scholar]
Вячный П., Фойсснер В. Морфология, конъюгация и постконъюгационная реорганизация Dileptus tirjakovae н. сп. (Ciliophora, Haptoria) J. Eukaryot. микробиол. 2008; 55: 436–447. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Vďačný P., Foissner W. Описание четырех новых почвенных дилептидов (Ciliophora, Haptoria) с примечаниями об адаптации к почвенной среде. Акта Протозол. 2008; 47: 211–230. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Vďačný P., Foissner W. Онтогенез Dileptus terrenus и Pseudomonilicaryon brachyproboscis (Ciliophora, Haptoria) J. Eukaryot. микробиол. 2009; 56: 232–243. [PMC free article] [PubMed] [Google Scholar]
Eukaryot. микробиол. 2009; 56: 232–243. [PMC free article] [PubMed] [Google Scholar]
Vďačný P., Orsi W., Foissner W. Молекулярные и морфологические доказательства родственных связей классов Armophorea и Litostomatea (Ciliophora, Intramacronucleata, Lamellicorticata infraphyl. nov. ), с учетом базальных гапторид литостоматов. Евро. Дж. Протистол. 2010; 46: 298–309. [PubMed] [Google Scholar]
Вячный П., Бурланд В.А., Орси В., Эпштейн С.С., Фойснер В. Филогения и классификация Litostomatea (Protista, Ciliophora) с акцентом на свободноживущие таксоны и ген 18S рРНК . Мол. Филогенет. Эвол. 2011;59: 510–522. [PubMed] [Google Scholar]
Винникова Н.В. Конъюгация у Dileptus anser (O.F.M.) (Gymnostomatida, Tracheliidae) Acta Protozool. 1974; 12: 275–287. (на русском языке с английским переводом названия и английской аннотацией) [Google Scholar]
Винникова Н.В. Тонкие структурные изменения макронуклеусов Dileptus anser О.Ф.М. при конъюгации. Акта Протозол. 1974; 13: 97–106. (на русском языке с английским переводом названия и аннотацией на английском языке) [Google Scholar]
1974; 13: 97–106. (на русском языке с английским переводом названия и аннотацией на английском языке) [Google Scholar]
Винникова Н.В. Конъюгация у инфузории Dileptus anser . I. Ультраструктура микроядер при митозе и мейозе. Протистология. 1976; 12:7–24. [Google Scholar]
Visscher J.P. Пищевые реакции у инфузорий, Dileptus gigas , с особым упором на функцию трихоцист. биол. Бык. Мар биол. лаборатория (Вудс-Хоул) 1923; 45: 113–143. [Google Scholar]
Visscher J.P. Конъюгация у реснитчатых простейших, Dileptus gigas , со специальной ссылкой на ядерные явления. Дж. Морфол. 1927; 44: 383–415. [Google Scholar]
Wenrich D.H. Наблюдения за некоторыми пресноводными инфузориями (Protozoa) II. Парадилептус , н. ген. Транс. Являюсь. микроск. соц. 1929; 48: 352–365. [Google Scholar]
Wiens J.J. Кодирование морфологических вариаций внутри видов и высших таксонов для филогенетического анализа. В: Wiens JJ, редактор. Филогенетический анализ морфологических данных. Пресса Смитсоновского института; Вашингтон, Лондон: 2000. стр. 115–145. [Академия Google]
Пресса Смитсоновского института; Вашингтон, Лондон: 2000. стр. 115–145. [Академия Google]
Wright A.-D.G., Lynn D.H. Монофилия трихостомных инфузорий (Phylum Ciliophora: Class Litostomatea) протестирована с использованием новых последовательностей 18S рРНК из вестибулиферид, Isotricha кишечные и Dasytricha ruminantium , а также 1000sutum11, 90 haptorian . Евро. Дж. Протистол. 1997; 33: 305–315. [Google Scholar]
Райт А.-Д.Г., Линн Д.Х. Филогенетический анализ инфузорий рубца семейства Ophryoscolecidae на основе последовательностей 18S рибосомной РНК с новыми последовательностями из Диплодиний , Эудиплодиний и Офриосколекс . Можно. Дж. Зул. 1997; 75: 963–970. [Google Scholar]
Райт А.-Д.Г., Дехорити Б.А., Линн Д.Х. Филогения инфузорий рубца Entodinium , Epidinium и Polyplastron (Litostomatea: Entodiniomorphida), выведенная из последовательности малой субъединицы рибосомальной РНК.