Тест по биологии Подцарство Одноклеточные 7 класс
Тест по биологии Подцарство Одноклеточные для учащихся 7 класса. Тест включает в себя 2 варианта, каждый вариант состоит из 2 частей (часть А, часть Б). В части А — 9 заданий, в части Б — 4 задания.
1 вариант
Часть А
А1. Практически все животные питаются
1) автотрофно
2) гетеротрофно
3) в процессе фотосинтеза
4) в процессе хемосинтеза
А2. Нервная система имеется у представителей царства
1) грибов
2) бактерий
3) растений
4) животных
А3. Из одной клетки состоит тело
1) мхов
2) лишайников
3) шляпочных грибов
4) простейших животных
А4. Ложноножки простейших животных представляют собой
1) пластиды
2) ядерное вещество
3) вырост цитоплазмы
4) запас питательных веществ
А5. Органоид выделения непереваренных остатков пищи у простейших животных — это
1) ядро
2) циста
3) жгутик
4) порошица
А6. Размножение простейших животных в основном происходит путем
Размножение простейших животных в основном происходит путем
1) фагоцитоза
2) пиноцитоза
3) деления клетки
4) образования цисты
А7. Постоянная форма тела отсутствует у
1) фораминиферы
2) инфузории-туфельки
3) эвглены зеленой
4) амебы обыкновенной
А8. Только в клетках простейших животных, которые питаются автотрофно, имеется
1) хлорофилл
2) цитоплазма
3) жгутик
4) ядро
А9. Два ядра находятся в клетках животного
1) инфузории
2) эвглены
3) вольвокса
4) амебы
Часть Б
Б1. Верны ли следующие утверждения?
А. Многие простейшие животные входят в состав планктона.
Б. Фагоцитоз простейших животных связан с образованием сократительной вакуоли.
1) Верно только А
2) Верно только Б
3) Верны оба суждения
4) Неверны оба суждения
Б2. Верны ли следующие утверждения?
А. Амеба захватывает пищу ложноножками.
Б. Среди представителей простейших животных существуют многоклеточные организмы.
1) Верно только А
2) Верно только Б
3) Верны оба суждения
4) Неверны оба суждения
Б3. Выберите три верных утверждения. В клетках животных отсутствует
1) клеточная стенка
2) хлоропласт
3) цитоплазма
4) ядро
5) наружная мембрана
6) крупная вакуоль
Б4. Установите соответствие животного и его видом.
Особенность строения
А. Наличие ресничек
Б. Светочувствительный глазок
В. Ложноножки
Г. Хлоропласт
Д. Два ядра
Вид животного
1. Амеба
2. Эвглена
3. Инфузория-туфелька
2 вариант
Часть А
A1. В отличие от растений для большинства животных характерны
1) автотрофное питание
2) ограниченный рост
3) неограниченный рост
4) неподвижность
А2. Симметрия тела характерна для
1) водорослей
2) животных
3) бактерий гниения
А3. Орган передвижения инфузорий — это
Орган передвижения инфузорий — это
1) клешни
2) реснички
3) конечности
4) мускульное волокно
А4. Захватывает пищу ложноножками
1) гриб
2) эвглена
3) амеба
4) инфузория
А5. В неблагоприятных условиях простейшие животные образуют
1) цисту
2) глотку
3) порошицу
4) сократительную вакуоль
А6. Избыток воды из тела простейшего животного удаляет
1) циста
2) ложноножка
3) сократительная вакуоль
4) светочувствительный глазок
А7. Специальные органоиды питания отсутствуют у
1) амебы обыкновенной
2) инфузории-туфельки
3) эвглены зеленой
4) вольвокса
А8. Организмы, которые способны к фотосинтезу и питаются готовыми веществами, называют
1) автотрофы
2) гетеротрофы
3) миксотрофы
4) фотосинтезирующие
А9. Органом пищеварения у инфузории является
1) глотка
2) ресничка
3) малое ядро
4) сократительная вакуоль
Часть Б
Б1. Верны ли следующие утверждения?
Верны ли следующие утверждения?
А. Среди простейших животных отсутствуют паразиты.
Б. Простейшие животные способны размножаться половым путем.
1) Верно только А
2) Верно только Б
3) Верны оба суждения
4) Неверны оба суждения
Б2. Верны ли следующие утверждения?
А. Эвглена зеленая перемещается к освещенным местам.
Б. Малярийный плазмодий — простейшее паразитическое животное.
1) Верно только А
2) Верно только Б
3) Верны оба суждения
4) Неверны оба суждения
Б3. Выберите три верных утверждения. Для животных характерны особенности жизнедеятельности
1) ограниченный рост
2) неподвижность
3) активное передвижение
4) неограниченный рост
5) питание готовыми веществами
6) образование веществ на свету
Б4. Установите соответствие между процессом жизнедеятельности животного и его видом.
Процесс жизнедеятельности
А. Фагоцитоз — захват пищи ложноножками
Б.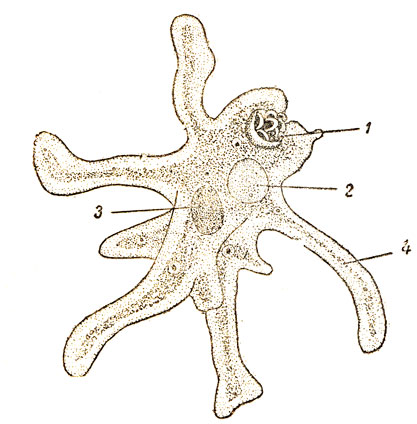 Непереваренные остатки удаляются через порошицу
Непереваренные остатки удаляются через порошицу
В. Фотосинтез
Г. Движение с помощью ресничек
Вид животного
1. Амеба
2. Эвглена
3. Инфузория-туфелька
Ответы на тест по биологии Подцарство Одноклеточные
1 вариант
А1. 2
А2. 4
А3. 4
А4. 3
А5. 4
А6. 3
А7. 4
А8. 1
А9. 1
Б1. 1
Б2. 1
Б3. 126
Б4. 32123
2 вариант
А1. 2
А2. 2
А3. 2
А4. 3
А6. 3
А7. 1
А8. 3
А9. 1
Б1. 2
Б2. 3
Б3. 135
Б4. 1323
PDF-версия
Тест Подцарство Одноклеточные
(92 Кб, pdf)
Занятие 1
Подцарство простейшие – Protozoa
Тип саркомастигофоры – Sarcomastigophora
Класс саркодовые – Sarcodina
Подкласс корненожки – Rizopoda
Отряд амебы – Amoebina
Вид амеба протей – Amoeba proteus
Класс жгутиконосцы – Mastigophora
Подкласс растительные жгутиконосцы – Phitomastigina
Виды эвглена зеленая – Euglena viridis
Вольвокс – Volvox globator
Подкласс животные жгутиконосцы – Zoomastigina
Вид трипанозома – Tripanosoma equiperdum
Отряд
амеб объединяет лишенных скелета
корненожек.
Амеба протей среди амеб наиболее крупное животное, достигающая размера 0,1-0,75 мм. Форма тела ее непостоянная. Амебы имеют вид сероватых зернистых комочков с отходящими в разных направлениях выступами цитоплазмы — псевдоподиями. Число и форма псевдоподий (ложноножек) непостоянны. Они могут появляться, а затем исчезать. Служат псевдоподии для передвижения и захватывания пищи. Цитоплазма амебы неоднородна и поэтому состоит из двух слоев. Наружный слой (эктоплазма) светлый, внутренний слой (эндоплазма) темный, зернистый. В эндоплазме находится ядро, сократительная и пищеварительная вакуоль. Сократительная или пульсирующая вакуоль имеет вид округлого светло-серого пузырька и предназначена для осморегуляции и выделения избытка воды и растворенных в ней продуктов обмена веществ.
У амебы протей пищей служат в основном мелкие простейшие и одноклеточные водоросли.
Размножаются
амебы путем деления надвое. Обитает это
простейшее в иле водоемов со стоячей,
часто загрязненной водой (с гниющими
на дне растительными остатками, листьями
и т.
— 1 —
Жгутиконосцы имеют органоиды движения — жгутики, которых может быть один, два или множество. Наряду со жгутиками у немногих видов временно или постоянно присутствует и ложноножки. Это указывает на близость между саркодовыми и жгутиконосцами, что дало основание объединить эти два класса в тип саркомастигофоры.
Эвглена
зеленная является представителем подкласса
растительных жгутиконосцев. Имеет лишь
один жгутик. Форма тела во время быстрого
поступательного движения веретеновидная,
но при замедленном движении эвглена
может укорачиваться, расширяться и даже
принимать шаровидную форму. Такая
способность к временному изменению
формы тела у простейших называется
метаболированием. Снаружи эвглена
покрыта тонкой эластической пелликулой,
выполняющей функцию оболочки. В теле
этого жгутиконосца видны ядро, хроматофоры,
окрашенные хлорофиллом в зеленый цвет,
зерна углевода парамила. У переднего
конца клеточного тала располагается
маленький красный светочувствительный
глазок или стигма.
Размножение бесполое, путем продольного деления надвое. Эвглена зеленая часто обитает в стоячих пресных водоемах, богатых легко разлагающимися органическими веществами.
Вольвокс
глобатор представляет собой шаровидную колонию
(диаметром до 2. мм). В состав колонии
входит больное количество (500-20000)
отдельных клеток, соединенных между
собой цитоплазматическими мостиками. Студенистое вещество, образующее
основную массу колонии, является
результатом ослизнения отдельных
клеток. Огромное большинство членов
колонии — вегетативные клетки,
обеспечивающие питание и движение
вольвокса. Каждая такая клетка имеет
два жгутика, ядро,
Студенистое вещество, образующее
основную массу колонии, является
результатом ослизнения отдельных
клеток. Огромное большинство членов
колонии — вегетативные клетки,
обеспечивающие питание и движение
вольвокса. Каждая такая клетка имеет
два жгутика, ядро,
— 2 —
4-глотка
5-пищеварительная вакуоль
7-сократитвльная вакуоль в приводящими каналами
8-трихоцисты
Таблица I.
Основные признаки классов простейших
Признаки | Классы | |||
жгутиковые | саркодовые | кокцидеобразные | инфузории | |
форма тела | ||||
Оболочка | ||||
Цитоплазма | ||||
Ядро | ||||
Органоиды пищеварения | ||||
Органоиды выделения | ||||
Органоиды движения | ||||
Размножение | ||||
Питание | ||||
Среда обитания | ||||
— 7 —
Инфузории
туфельки свойственны как бесполое, так
и половое размножение. Бесполое
размножение происходит путем поперечного
деления инфузории надвое. После ряда
бесполых размножений обязательно
наступает половой процесс.
Бесполое
размножение происходит путем поперечного
деления инфузории надвое. После ряда
бесполых размножений обязательно
наступает половой процесс.
Половое размножение у инфузории носит название конъюгации. Две инфузории прикладываются друг к другу, в определенных местах пелликула растворяется и образуется цитоплазматический мостик.
Макронуклеус каждой конъюгирующей вакуоли распадается, а микронуклеус делится дважды. В результате деления получается 4 ядра, из которых 3 рассасываются, а одно остается. Оставшееся ядро тоже делится и образуется два ядра. Одно из ядер остается на месте, а другое мигрирует в партнера и сливается с оставшимся в нем ядром. На этом конъюгация заканчивается и инфузории расходятся.
Инфузория туфелька относится к числу обычных обитателей мелких пресных водоемов с высоким содержанием органических веществ, живет в придонных слоях.
Для
изучения инфузории туфельки готовят
препарат из культуры этих простейших. На предметное стекло помещают небольшую
каплю культуры инфузорий и рассматривают
под микроскопом движущихся животных.
Детальное изучение строения можно
проводить на остановленных туфельках.
Остановить движение инфузории можно
следующим образом: накрыть каплю
(инфузорий) покровным стеклом, а излишек
жидкости удалить с помощью полосок
фильтровальной бумаги. Следует непременно
следить под микроскопом за процессом
удаления жидкости и прекратить его как
только туфельки остановятся. В противном
случае инфузории будут раздавлены
покровным стеклом.
На предметное стекло помещают небольшую
каплю культуры инфузорий и рассматривают
под микроскопом движущихся животных.
Детальное изучение строения можно
проводить на остановленных туфельках.
Остановить движение инфузории можно
следующим образом: накрыть каплю
(инфузорий) покровным стеклом, а излишек
жидкости удалить с помощью полосок
фильтровальной бумаги. Следует непременно
следить под микроскопом за процессом
удаления жидкости и прекратить его как
только туфельки остановятся. В противном
случае инфузории будут раздавлены
покровным стеклом.
Задание для самостоятельной работы:
Изучить эймерию магна (объектив х 90).
Нарисовать схему жизненного цикла эймерии магна.
Изучить инфузорию туфельку (объектив х 9) и зарисовать приготовив предварительно препарат.
Заполнить таблицу I. “Основные признаки классов простейших”.
Рис. 1.
Инфузория туфелька
1.
Инфузория туфелька
1-микронуклеус
2-макронуклеус
3-ротовое отверстие
— 6 —
хроматофор, стигму и две сократительные вакуоли. Встречаются и генеративные клетки (гаметы), отвечающие за размножение. Мужские (микрогаметы) и женские (макрогаметы) половые клетки сливаются, и образовавшаяся зигота дает начало новой колонии. Вольвоксы — обитатели пресных водоемов. Питание автотрофное.
Трипаносома
эквипердум. Величина ее 22-28 мкм в длину и 1,4-2,6 мкм в
ширину. Тело трипаносомы лентовидное,
заостряется к переднему и заднему концу.
Жгутик начинается близ заднего конца
от базального тельца — кинетосомы,
тянется вдоль тела и выступает на
переднем конце. Он соединен с телом
посредством ундулирующей мембраны
(волнообразной перепонки), которая
производит колебательные движения.
Размножение простое, продольное деление
надвое. Трипаносома эквипердум
паразитирует в капиллярах слизистой
оболочки половых органов непарнокопытных.
Задание для самостоятельной работы:
Изучить и зарисовать амебу протей (объектив х 40), эвглену
зеленую (объектив х 40), трипаносому эквипердум (объектив х 90).
Изучить вольвокса глобатора и раковины фораминифер
(объектив х 9).
Рис.1. Амеба протей 1-ядро 2-экто плазма 3-эндоплазма. 4-пищеварительная вакуоль 5-сократительная вакуоль 6-псевдоподии | Рис.2. Эвглена зеленая 1-ядро 2-стигма (глазок) 3-сократительная вакуоль 4-резервуар сократительной вакуоли 5-хроматофоры 6-зерна парамила 7-жгутик |
Рис. 3.
Трипаносома
3.
Трипаносома
1-ядро
2-ундулирушцая мембрана
3-жгутик
4-блефорапласт
— 3 —
Что Вы знаете о разнообразии одноклеточных животных?
Одноклеточные организмы выполняют те же функции, что многоклеточные: питаются, двигаются и размножаются. Их клетки должны быть «мастером на все руки», чтобы делать все это, что другие животных делают особые органы. Поэтому одноклеточные животные настолько непохожи на остальных, что их выделяют в отдельные Подцарство простейших.
К типу простейшие относится свыше 15000 видов животных, обитающих в морях, пресных водах, почве. Помимо свободноживущих форм, известно много паразитических, которые вызывают порой серьезные заболевания – протозоонозы.
Тело простейших состоит только из одной клетки. Форма тела простейших разнообразна. Оно может быть постоянным, иметь лучевую, двустороннюю симметрию (жгутиковые, инфузории) или вообще не иметь постоянной формы (амеба). Размеры тела простейших обычно малы – от 2-4 мк до 1,5 мм, хотя некоторые крупные особи достигают 5 мм в длину, а ископаемые раковинные корненожки имели в диаметре 3 см и более.
Размеры тела простейших обычно малы – от 2-4 мк до 1,5 мм, хотя некоторые крупные особи достигают 5 мм в длину, а ископаемые раковинные корненожки имели в диаметре 3 см и более.
Тело простейших состоит из цитоплазмы и ядра. Цитоплазма ограничена наружной цитоплазматической мембраной, в ней находятся органоиды — митохондрии, рибосомы, эндо-плазматическая сеть, аппарат Гольджи. У простейших одно или несколько ядер. Форма деления ядра – митоз. Имеется также половой процесс. Он заключается в образовании зиготы. Органоиды движения простейших – это жгутики, реснички, ложноножки; или их нет совсем. Большинство простейших, как и все прочие представители животного царства, гетеротрофные. Однако среди них имеются и автотрофные.
Особенность простейших переносить неблагоприятные условия окружающей среды – состоит в способности инцистироваться , т.е. образовывать цисту. При образовании цисты органоиды движения исчезают, объем животного уменьшается, оно приобретает округлую форму, клетка покрывается плотной оболочкой. Животное переходит в состояние покоя и при наступлении благоприятных условий возвращается к активной жизни.
Животное переходит в состояние покоя и при наступлении благоприятных условий возвращается к активной жизни.
Размножение простейших весьма разнообразно, от простого деления (бесполое размножение) до довольно сложного полового процесса – конъюгации и копуляции.
Среда обитания простейших разнообразна – это море, пресные воды, влажная почва.
Четыре основных класса простейших
1 – жгутиковые;
2 – саркодовые;
3 – споровики;
4 – инфузории.
1. Около 1000 видов, преимущественно с вытянутым овальным или грушевидном телом, составляют класс жгутиковых. Органеллы движения – жгутики, которых у различных представителей класса может быть от 1 до 8 и более. Жгутик – тонкий цитоплазматический вырост, состоящий из тончайших фибрилл. Своим основанием он прикреплен к базальному тельцу или кинетопласту . Жгутиковые движутся жгутом вперед, создавая своим движением вихревые водовороты и как бы «ввинчивая» животное в окружающую жидкую среду.
Способ питания : жгутиковых разделяют на имеющих хлорофилл и питающихся автотрофно, и на не имеющих хлорофилла и питающихся, как прочие животные, гетеротрофным способом.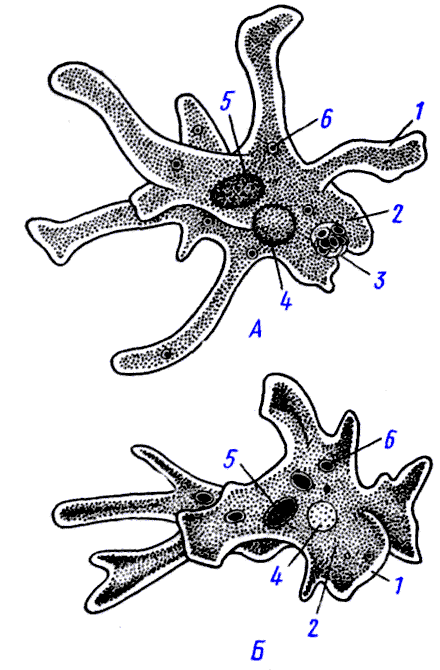 Гетеротрофы на передней стороне тела имеют особое углубление – цитостом , через который при движении жгутика пища вгоняется в пищеварительную вакуоль. Ряд форм жгутиковых питается осмотическим путем, всасывая всей поверхностью тела, растворенные органические вещества из окружающей среды.
Гетеротрофы на передней стороне тела имеют особое углубление – цитостом , через который при движении жгутика пища вгоняется в пищеварительную вакуоль. Ряд форм жгутиковых питается осмотическим путем, всасывая всей поверхностью тела, растворенные органические вещества из окружающей среды.
2. Представители класса саркодовых, или корненожек, двигаются при помощи ложноножек – псевдоподий.
Класс включает разнообразных водных одноклеточных: амеб, солнечников, лучевиков. Среди амеб, кроме форм, не имеющих скелета или раковинки, встречаются виды, имеющие домик.
Большинство саркодовых являются обитателями морей, имеются также пресноводные, живущие в почве.
Саркодовые характеризуются непостоянной формой тела. Дыхание осуществляется всей его поверхностью. Питание – гетеротрофное. Размножение – бесполое, существует также половой процесс.
3. Представители класса Споровики характеризуются тем, что в цикле своего развития образуют стадию спор .Все споровики являются паразитами человека и животных. Они паразитируют в разных тканях и клетках. Лихорадка, малокровие, желтуха – типичные признаки заболевания споровиками. Пироплазмы, бабезии относятся к отряду кровяных споровиков, поражая эритроциты млекопитающих (коров, лошадей, собак и др. домашних животных). Переносчики болезней – клещи. Кроме кровяных есть еще два отряда споровиков — кокцидии и грегарины .
Они паразитируют в разных тканях и клетках. Лихорадка, малокровие, желтуха – типичные признаки заболевания споровиками. Пироплазмы, бабезии относятся к отряду кровяных споровиков, поражая эритроциты млекопитающих (коров, лошадей, собак и др. домашних животных). Переносчики болезней – клещи. Кроме кровяных есть еще два отряда споровиков — кокцидии и грегарины .
Кокцидии паразитируют как в беспозвоночных, так и в позвоночных животных – млекопитающих, рыбах, птицах. Кокцидия токсоплазмоз вызывает болезнь человека токсоплазмоз. Им можно заразиться от любого представителя семейства кошачьих.
Грегарины паразиты только беспозвоночных, в основном – насекомых и, как правило, поселяются в кишечнике. Взрослые грегарины внешне похожи на червей. Они бывают как мелкие (10 микрон), так и весьма крупные, видимые невооруженным глазом – до 1,6 мм.
4. Представители класса Инфузорий имеют органеллы передвижения – реснички, обычно в большом числе. Так, у инфузории-туфельки число ресничек более 2000. Реснички (как и жгутики) представляют собой специальные сложно устроенные цитоплазматические выросты. Тело инфузорий покрыто оболочкой, пронизанной мельчайшими порами, через которые выходят реснички.
Реснички (как и жгутики) представляют собой специальные сложно устроенные цитоплазматические выросты. Тело инфузорий покрыто оболочкой, пронизанной мельчайшими порами, через которые выходят реснички.
В тип инфузорий объединяют наиболее высоко организованных простейших. Они – вершина достижений, совершенных эволюцией в этом Подцарстве. Инфузории ведут свободно плавающий или прикрепленный образ жизни. Обитают как в пресных, так и в соленых водах. Среди инфузорий много симбионитов и мало паразитических форм.
Достаточно среди инфузорий и паразитов беспозвоночных и позвоночных животных (включая человека). Очень много паразитических в особом отделе желудка жвачных копытных – в рубце.
У всех инфузорий не менее двух ядер. Большое ядро регулирует все жизненные процессы. Маленькое ядро играет основную роль в половом процессе.
Пища в тело животного попадает через клеточный “рот”, куда она загоняется движением ресничек; на дне глотки образуются пищеварительные вакуоли . Непереваренные остатки выводятся наружу.
Непереваренные остатки выводятся наружу.
Простейшие-строители горных пород : Почти 600 миллионов лет назад произошла «скелетная революция». Большинство живых организмов «оделось» скелетами, защитивших их от врагов. С тех пор бесчисленное поколение простейших гибли, их раковины откладывались на дне морей, километровые толщи осадков спрессовались под собственной тяжестью, превращаясь в мел и известняк. Движение земной коры поднимали осадочные породы на поверхность, сооружая из них горы. Вода вымывала минеральные вещества обратно в море, где они снова использовались для построения раковин. Так, благодаря простейшим, совершался круговорот минеральных элементов в биосфере на протяжении её истории.
Простейшие-важное звено водных экосистем : Пищевые цепи в водных экосистемах начинают с микроскопических водорослей. Вторым звеном в них обычно являются планктонные простейшие – первые потребители зелёной продукции. Затем они становятся основой питания животноядных обитателей водных экосистем _ рачков, мальков рыб и всех последующих потребителей. Когда остатки мёртвых растений и животных, опускающих на дно, их подбирают донные простейшие.
Когда остатки мёртвых растений и животных, опускающих на дно, их подбирают донные простейшие.
Немало простейших населяет и каждой миллиметр почвы, насыщенной грунтовой влагой. Вместе с другими обитателями они поддерживают плодородие почв.
Без простейших не могут существовать растительноядные животные : Ирония судьбы: растительноядные животные сами не в состоянии переваривать целлюлозу (клетчатку) — основу растительных тканей! За них это делают простейшие, заселяющей их пищевой тракт с первых дней жизни. Кишечник термита, слепая кишка зайца и желудок коровы оборудованы специальными складами для размещения этих сожителей. Хозяин усваивает лишь результат их пищеварения, а заодно и самих простейших.
Псевдоподии
Эта статья про эукариотические клетки. Для группы см. Псевдопод (полоса). Для подкаста см. Псевдопод (подкаст). О структуре в анатомии насекомых см. Proleg.
эта статья нужны дополнительные цитаты для проверка. |
 Пожалуйста помоги улучшить эту статью от добавление цитат в надежные источники. Материал, не полученный от источника, может быть оспорен и удален.
Пожалуйста помоги улучшить эту статью от добавление цитат в надежные источники. Материал, не полученный от источника, может быть оспорен и удален.Хаос Каролиненс, амебовидный лобоподии
А псевдопод или псевдоподиум (множественное число: ложноножки или псевдоподия) — временная рука-подобная проекция эукариотический клеточная мембрана которые развиваются по направлению движения. Наполненный цитоплазма псевдоподии в основном состоят из актиновые нити а также может содержать микротрубочки и промежуточные нити.[1][2] Псевдоножки используются для подвижность и проглатывание. Они часто встречаются в амебы.
Различные типы псевдоподий можно классифицировать по их отличному внешнему виду.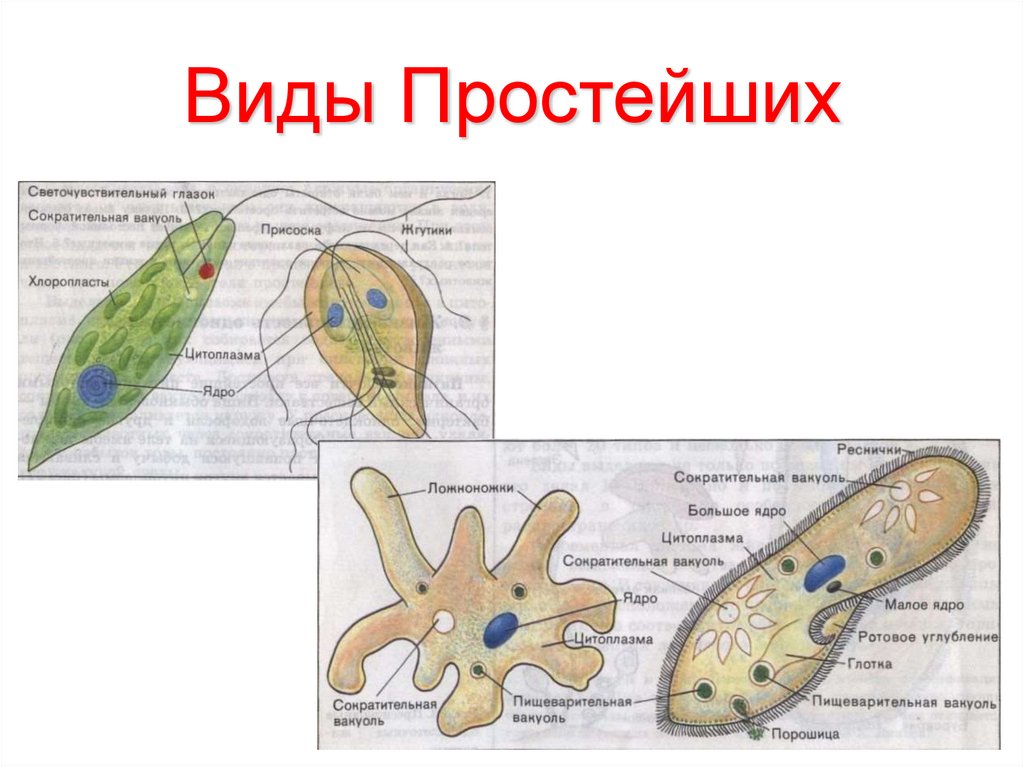 [3]Ламеллиподии широкие и тонкие. Филоподии тонкие, нитевидные и в значительной степени поддерживаются микрофиламентами. Лобоподии луковичные и амебные. Ретикулоподии представляют собой сложные структуры, несущие отдельные псевдоподии, образующие неправильные сети. Аксоподии представляют собой тип фагоцитоза с длинными тонкими ложноножками, поддерживаемыми сложными массивами микротрубочек, окруженных цитоплазмой; они быстро реагируют на физический контакт.[4]
[3]Ламеллиподии широкие и тонкие. Филоподии тонкие, нитевидные и в значительной степени поддерживаются микрофиламентами. Лобоподии луковичные и амебные. Ретикулоподии представляют собой сложные структуры, несущие отдельные псевдоподии, образующие неправильные сети. Аксоподии представляют собой тип фагоцитоза с длинными тонкими ложноножками, поддерживаемыми сложными массивами микротрубочек, окруженных цитоплазмой; они быстро реагируют на физический контакт.[4]
Однако некоторые псевдоподиальные клетки могут использовать несколько типов псевдоподий в зависимости от ситуации: большинство из них используют комбинацию ламеллиподий и филоподий для миграции.[5] (например, метастатические раковые клетки[6]). Фибробласты крайней плоти человека могут использовать миграцию на основе ламеллиподий или лобоподий в трехмерной матрице в зависимости от эластичности матрицы.[7]
Как правило, несколько псевдоподий возникают на поверхности тела (полиподиальный, например Амеба протей ), либо на поверхности тела может образоваться одиночная ложноножка (моноподиальный, например Entamoeba histolytica ).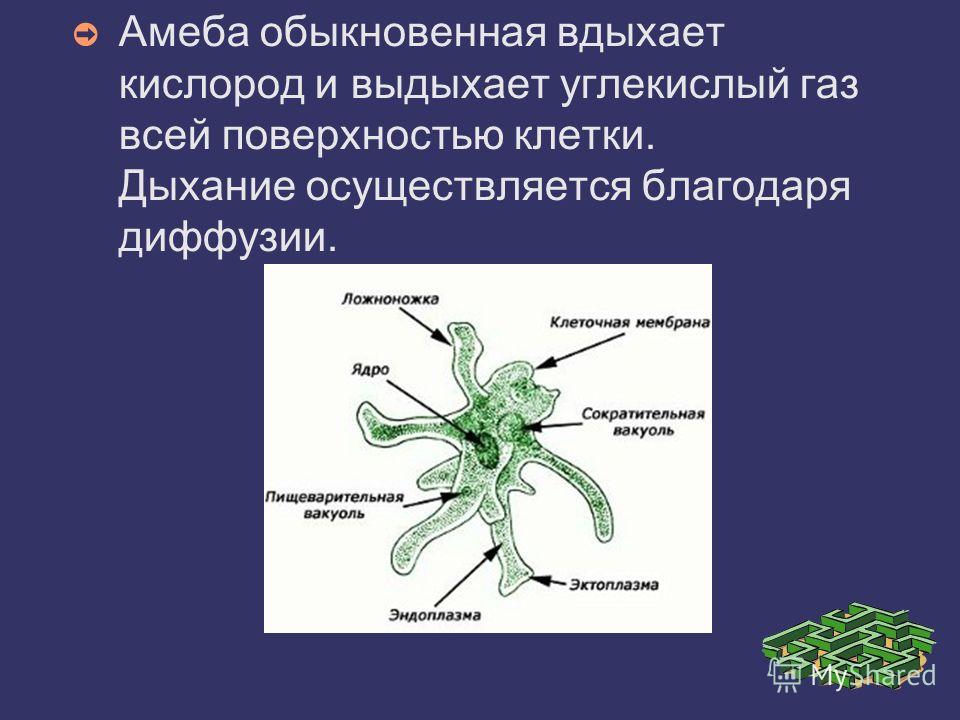 [8]
[8]
Клетки, образующие ложноножки, обычно называют амебоиды.[9]
Содержание
- 1 Формирование
- 1.1 Через внеклеточный сигнал
- 1.2 Без внеклеточного сигнала
- 2 Функции
- 3 Морфология
- 3.1 Ламеллиподии
- 3.2 Филоподии
- 3.3 Лобоподия
- 3.4 Ретикулоподии
- 3.5 Аксоподии
- 4 использованная литература
Формирование
Через внеклеточный сигнал
Чтобы двигаться к цели, ячейка использует хемотаксис. Он воспринимает внеклеточные сигнальные молекулы, хемоаттрактанты (например, цАМФ для Диктиостелиум клетки[10]), чтобы расширить псевдоподии в области мембраны, обращенной к источнику этих молекул.
Хемоаттрактанты связываются с G-белковые рецепторы, которые активируют GTPases семейства Rho (например, Cdc42, Rac) через G-белки.
Rho GTPases способны активировать WASp которые, в свою очередь, активируют Комплекс Арп2 / 3 которые служат центрами зарождения полимеризация актина. [11] Затем полимеры актина толкают мембрану по мере своего роста, образуя псевдопод. Псевдоподий может прикрепиться к поверхности через адгезионные белки (например. интегрины ), а затем тянет тело клетки вперед за счет сокращения актин-миозинового комплекса в псевдоподе.[12][13] Этот тип передвижения называется Амебоидное движение.
Rho GTPases также могут активировать фосфатидилинозитол-3-киназа (PI3K) которые набирают PIP3 к мембране на переднем крае и отсоедините PIP3-разлагающий фермент PTEN из той же области мембраны. PIP3 затем активируйте GTPases обратно через ГЭФ стимуляция. Это служит петлей обратной связи для усиления и поддержания присутствия локальной GTPase на переднем крае.[11]
В противном случае псевдоподии не могут расти на других сторонах мембраны, кроме переднего края, потому что миозиновые нити препятствуют их расширению. Эти миозиновые филаменты индуцируются циклический GMP в D. discoideum или Ро-киназа в нейтрофилы Например.[11]
Без внеклеточного сигнала
В случае отсутствия внеклеточного сигнала все движущиеся клетки перемещаются в случайных направлениях, но они могут сохранять одно и то же направление в течение некоторого времени перед поворотом. Эта функция позволяет клеткам исследовать большие области для колонизации или поиска нового внеклеточного сигнала.
В Диктиостелиум клетки, псевдоподий может образовывать либо de novo как обычно, или от существующего ложноножка, образуя Y-образный псевдоподий.
Y-образные псевдоподы используются Диктиостелиум продвигаться относительно прямо вперед, попеременно отводя левую или правую ветвь псевдоножки. В de novo псевдоподии образуются с разных сторон, чем уже существовавшие, они используются клетками для поворота.
Y-образные ложноножки встречаются чаще, чем de novo те, которые объясняют предпочтение клетки продолжать двигаться в том же направлении. Эта настойчивость модулируется PLA2 и пути передачи сигналов cGMP.[10]
Функции
Функции псевдоподий включают передвижение и прием пищи:
- Псевдоподии критически важны для обнаружения целей, которые затем могут быть поглощены; охватывающие псевдоподии называются фагоцитоз псевдоподии. Типичным примером этого типа амебоидных клеток является макрофаг.
- Они также необходимы для амебоидного передвижения. Человек мезенхимальные стволовые клетки являются хорошим примером этой функции: эти мигрирующие клетки ответственны за внутриутробное ремоделирование; например, при формировании триламинарный зародышевый диск в течение гаструляция.[14]
Морфология
Формы псевдоподий слева: полиподиальные и лобозные; моноподиальные и лобозные; нитевидный; коническая; ретикулозный; сужающиеся актиноподы; неконусные актиноподы
Псевдоподии можно разделить на несколько разновидностей по количеству выступов (моноподии и полиподии), а также по внешнему виду:
Ламеллиподии
Ламеллиподии широкие и плоские псевдоподии, используемые при передвижении. [4] Они поддерживаются микрофиламентами, которые образуются на переднем крае, создавая сетчатую внутреннюю сеть.[15]
Филоподии
Филоподии (или филозные ложноножки) тонкие и нитевидные с заостренными концами, состоящие в основном из эктоплазма. Эти образования поддерживаются микрофиламенты которые, в отличие от нитей ламеллиподий с их сетчатым актином, образуют рыхлые пучки с помощью сшивание. Это образование частично происходит из-за связывания белков, таких как фимбрины и очаровывает.[15][16]Филоподии наблюдаются в некоторых клетках животных: в части Filosa (Ризария ), в «Тестацефилозия » , в Вампиреллиды и Псевдоспориды (Ризария ) И в Nucleariida (Опистоконта ).[4]
Лобоподия
Лобоподии (или лопастные псевдоподии) луковичные, короткие и тупые.[17] Эти пальцеобразные трубчатые псевдоподии содержат как эктоплазма и эндоплазма. Их можно найти в разных типах ячеек, особенно в Лобоса и другие Амебозоа и в некоторых Heterolobosea (Экскавата ).
Лобоподии высокого давления также встречаются у людей. фибробласты путешествовать по сложной сети 3D матрица (например, млекопитающие дерма, матрица, полученная из клеток). В отличие от других псевдоподий, использующих давление, оказываемое полимеризацией актина на мембрану для расширения, фибробластные лобоподы используют ядерный поршневой механизм, заключающийся в вытягивании ядра через сократимость актомиозина, чтобы подтолкнуть цитоплазму, которая, в свою очередь, толкает мембрану, что приводит к образованию псевдоподий. Для возникновения этой миграции фибробластов на основе лобоподий требуется несприн 3, интегрины, RhoA, СКАЛА и миозин II В противном случае лобопод часто сопровождается небольшими боковыми пузыри формируются вдоль стороны клетки, вероятно, из-за высокого внутриклеточного давления во время формирования лобоподий, увеличивая частоту разрыва плазматической мембраны и коры.[18][7][19]
Ретикулоподии
Ретикулоподии (или ретикулозные ложноножки),[20] представляют собой сложные образования, в которых отдельные ложноножки сливаются и образуют неправильные сети. «Ретикулоподии». eForams. Архивировано из оригинал на 2007-07-17. Получено 2005-12-30.
Проверочная работа по теме » Простейшие»
Обновлено: 24.09.2022
Б. Для каждого типа простейших выпиши номера соответствующих утверждений.
- Тело не имеет четкой формы.
- Есть ротовое углубление.
- На переднем конце тела находится светочувствительный глазок.
- Все тело покрыто ресничками.
- Передвигаются при помощи жгутика.
- Передвигаются при помощи ложноножек.
- Клетка содержит два ядра.
- В клетке могут содержаться хлоропласты.
- Тип питания — только гетеротрофный.
- Может переходить с автотрофного на гетеротрофный тип питания.
- Способны образовывать цисту.
- Дыхание осуществляется через всю поверхность клетки.
- Являются одной из самых древних групп одноклеточных.
В. Установите, кто из представителей простейших лишний в каждом ряду. Почему?
- Амеба обыкновенная, эвглена зеленая, инфузория-туфелька.

- Амеба обыкновенная, инфузория-туфелька, лямблия.
- Амеба обыкновенная, инфузория-туфелька, инфузория-бурсария.
б) по передвижению:
- Амеба, эвглена зеленая, лямблия.
- Инфузория-туфелька, бурсария, эвглена зеленая.
- Амеба обыкновенная, инфузория-туфелька, эвглена зеленая.
По теме: методические разработки, презентации и конспекты
Проверочная работа по теме «Работа с формулами в Excel»
Проверочная работа по теме «Работа с формулами в Excel».
Проверочная работа 7 класс «Простейшие». Другие работы ищите на странице «Мои проверочные работы по биологии», в навигации
Две проверочные работы «Амеба», «Эвглена зеленая».
Методическая разработка проверочной работы по подготовке к Всероссийской проверочной работе, 11 класс
Веселова Татьяна Андреевна, учитель географии ГБОУ лицея №150 Калининского района Санкт-Петербурга.Данная методическая разработка позволит учителю подготвоить учащихся 11-х классов к итоговой ат.
Проверочная работа (самостоятельная работа, тест, контрольная работа, географический диктант) по теме «На какой Земле мы живём» (§2-4, к учебнику А. И. Алексеева, В. В. Николиной, Е. К. Липкиной «Полярная звезда», 5 класс)
Предназначение работы.Данная проверочная работа предназначена для закрепления изученного материала по теме «На какой Земле мы живём» (§2-4, к учебнику А. И. Алексеева, В. В. Николи.
Проверочная работа (самостоятельная работа, тест, контрольная работа) в формате ОГЭ/ЕГЭ по теме «Планета Земля» (§5-7, к учебнику А. И. Алексеева, В. В. Николиной, Е. К. Липкиной «Полярная звезда», 5 класс)
Предназначение работы. Данная проверочная работа предназначена для закрепления изученного материала по теме «Планета Земля» (§5-7, к учебнику А. И. Алексеева, В. В. Николиной, Е. К.
Проверочные вопросы по музыке для школьной комплексной проверочной работы.
Проверочные вопросы по музыке для школьной комплексной проверочной работы.
Паразитические простейшие
Паразитические простейшие — это одноклеточные животные, которые живут за счет других организмов, называемых хозяевами. Только в организме человека паразитирует около 30 видов простейших.
Паразитический образ жизни ведут простейшие, относящиеся к типу споровиков. Они паразитируют в организмах червей, насекомых, зверей и человека. К споровикам относят около 4 тыс. простейших.
Приспособившись к паразитическому образу жизни, споровики отличаются от других простейших. У взрослых споровиков нет органоидов движения. Пищеварительных и сократительных вакуолей у них тоже нет, так как питание осуществляется всей поверхностью клетки.
Паразиты разрушают ткани хозяина и отравляют его организм ядовитыми выделениями.
Размножение у большинства споровиков бесполое. Клетка паразита распадается на множество дочерних клеток — самостоятельных организмов.
Как человек заболевает малярией?
Для человека опасен кровяной споровик — малярийный плазмодии. Он обитает в клетках крови человека и имеет микроскопические размеры. Этот паразит распространяется малярийными комарами и вызывает опасное заболевание людей — малярию (рис. 11). Возбудителя малярии в 1880 г. описал французский врач А. Лаверан.
Рис. 11. Жизненный цикл малярийного плазмодия
Малярийных паразитов комар всасывает с кровью больного человека. В организме комара плазмодии размножаются и скапливаются в слюнных железах.
При укусе вместе со слюной комара в кровь человека проникают и малярийные плазмодии. Здесь они питаются, растут, размножаются, разрушают кровяные клетки человека и выделяют ядовитые вещества. При массовом выходе простейших из клеток в кровь у больного малярией начинается приступ лихорадки — повышение температуры, сильнейший озноб и слабость. Приступы повторяются через каждые два-три дня. Это ведет к малокровию, истощению организма.
Всемирной организацией здравоохранения (ВОЗ) разработаны и реализованы меры борьбы с малярией: обследование населения на зараженность малярийным плазмодием, лечение заболевших людей, борьба с малярийными комарами и ликвидация очагов их размножения (временных водоемов, сырых подвалов), проведение профилактических мероприятий по предупреждению распространения малярии.
Какие еще заболевания вызывают паразитические простейшие?
Наряду со споровиками паразитический образ жизни ведут некоторые представители корненожек, жгутиконосцев и инфузорий (см. рис. 11).
К паразитическим корненожкам принадлежит дизентерийная амеба. Ее в 1875 г. открыл русский ученый Ф. А. Леш. Человек заражается цистами дизентерийной амебы. Они попадают в организм с загрязненной водой или пищей. В пищеварительном тракте оболочки цист растворяются. Амеба начинает усиленно питаться, размножаться и проникает в стенки кишечника, вызывая тяжелое заболевание амебиаз.
Амебиаз называют «болезнью грязных рук». Поэтому всем, кто путешествует в странах Южной Азии и Африки, где часто встречается дизентерийная амеба, необходимо строго соблюдать правила гигиены. Это поможет уберечься от амебиаза и исключить завоз возбудителя заболевания в нашу страну.
Кишечник, печень, половые органы животных и человека поражают лямблии. Они были подробно описаны отечественным ученым Д. Ф. Лямблем. У человека чаще всего паразитирует многожгутиковая лямблия. Ее клетка похожа на разрезанную пополам грушу. Плоской стороной паразит плотно присасывается к клеткам кишечника. Человек, употребляя в пищу плохо вымытые овощи и фрукты, проглатывает цисты лямблии. Заражение лямблиями сопровождается кишечными расстройствами. Это заболевание часто бывает у детей.
Для предупреждения заболевания лямблиозом необходимо соблюдать самые простые санитарно-гигиенические правила (предохранять питьевую воду и продукты питания от загрязнения, соблюдать правила личной гигиены).
Каковы особенности одноклеточных животных?
Главная особенность простейших — микроскопические размеры и одноклеточность. При этом каждое простейшее — самостоятельный одноклеточный организм. Ему свойственны основные процессы жизнедеятельности; рост, развитие, дыхание, питание, размножение. Простейшие прошли долгий путь эволюционного развития и в настоящее время представлены разнообразием форм, приспособленных к различным условиям жизни.
Паразитические простейшие. Празиты: описание, примеры
Среди опасных организмов, поселяющихся внутри человека и наносящих вред его здоровью, значительное место занимают паразитические простейшие. Это одноклеточные животные, которые приспособились к существованию за чужой счет. Они достаточно многочисленны, всего выделяют около 30 различных видов особей, которые специализируются именно на человеке. Еще больше встречается в организмах других позвоночных и беспозвоночных животных.
Паразитические простейшие: особенности строения
Внешне и внутренне данные организмы имеют точно такое же строение, как и все остальные одноклеточные. Они имеют всего одну структурную единицу, однако она способна выполнять все жизненно важные функции, необходимые для нормальной жизнедеятельности. А именно:
- растет и развивается;
- размножается;
- питается;
- дышит;
- обладает возбудимостью и раздражимостью;
- передвигается.
Поэтому с уверенностью можно сказать, что паразитические простейшие — вполне самостоятельные, целостные живые организмы. Отличия же их от всех остальных одноклеточных в том, что они утратили пищеварительную вакуоль в связи с паразитическим образом жизни, а также сократительную. Кроме того, чаще всего лишены органоидов передвижения.
Размножаются достаточно быстро, бесполым путем. Материнская клетка просто, по достижении созревания, распадается на несколько дочерних. Они ведут самостоятельный образ жизни сразу после образования.
Как питаются паразиты в организме хозяина? Это зависит от индивидуальных особенностей каждого вида, однако в основном их пища — кровь или содержимое кишечника. Поглощают вещества они всей поверхностью тела из-за редукции вакуоли.
Классификация и представители простых паразитов
Классифицировать различных представителей рассматриваемых организмов можно следующим образом.
| Класс | Представители |
| Споровики | малярийный плазмодий, кокцидии, грегарины, пироплазмиды |
| Корненожки | дизентерийная амеба, кишечная, ротовая амеба |
| Жгутиконосцы | трихомонады, лямблии, трипаносомы, лейшмании |
| Инфузории | балантидий |
Все эти организмы — причина протозойных заболеваний именно человека. Все они — очень опасные для жизни и здоровья паразитические простейшие. Таблица включает в себя лишь самых распространенных представителей. На самом деле существуют и более редкие, их достаточно много.
Лейшмании
На сегодняшний день цифра, отражающая количество людей, пораженных этим паразитом, просто ужасна — 12 миллионов. Этот овальной формы неподвижный внутриклеточный представитель простейших имеет небольшой жгутик. Однако передвигается очень медленно и на небольшие расстояния.
Поселяясь внутри клеток хозяина, которым для лейшманий является насекомое и млекопитающее, в том числе человек, они вызывают постепенное отмирание структуры. У человека на коже, в том месте, где обитают лейшмании, формируется открытая зияющая язва, постоянно нагнивающая. Болезнь эта названа каучуковой или багдадской язвой. Лечится очень сложно и длительно, с хирургическим вмешательством.
Переносят одноклеточных этой группы москиты, поэтому очаги лейшманиоза встречаются в странах тропиков и субтропиков.
Лямблии
Класс паразитических простейших, к которым относятся лямблии — Жгутиковые. Самым опасным представителем для людей является кишечная лямблия, которая вызывает опасное заболевание — лямблиоз.
Взрослые особи паразитируют в верхнем отделе кишечника. Именно здесь они пиноцитозом поглощают все питательные вещества, которые употребляет человек. Также этот организм способен формировать цисты. В этом состоянии они могут переживать облучение ультрафиолетом и воздействие токсических веществ. Однако низкие и высокие температуры для них смертельны.
Цисты располагаются в толстой кишке, поэтому выводятся с твердыми продуктами жизнедеятельности. Заражение ими происходит через грязь, воду. Поэтому правила личной гигиены особенно важны в профилактике лямблиоза.
Чем опасен этот паразит? В процессе жизнедеятельности лямблия выделяет токсичные продукты распада, которые сильно отравляют организм изнутри.
Паразитические корненожки
К данному классу относятся разные виды амебы:
- кишечная;
- дизентерийная;
- ротовая.
Эти паразитические простейшие вызывают опасные заболевания у людей, которые в ряде случаев могут заканчиваться даже смертью.
Дизентерийная амеба — представительница жарких стран. Именно в них она — наиболее распространенный паразит. Ее особенность, как и у всех амебоидных, — отсутствие раковинных и скелетных образований. Поэтому она передвигается при помощи ложноножек. Размножается простым делением. Способна формировать цисты, также живущие в организме человека. Внутри этой структуры происходит множественное деление.
Вызывает заболевание амебиаз. Поражаются внутренние стенки кишечника, на которых амебы оставляют кровоточащие язвы. У человека наблюдается кровяной жидкий стул, боли внутри организма. Диагностировать дизентерийную амебу сложно. Поэтому она и является очень опасным представителем паразитов.
В отличие от рассмотренной формы, кишечная амеба присутствует в организме почти каждого человека и живет с ним в ладу. Не выделяет токсичных веществ и не пробуравливает стенки кишечника. Поэтому особого дискомфорта и вреда здоровью не наносит.
Ротовая амеба — обитатель зубного налета и кариесных образований. Питается бактериями, может употреблять эритроциты. Точное значение ее для человека пока не ясно.
Балантидий коли
Данный организм — представитель класса Инфузории. Достаточно крупный овальный одноклеточный организм, поселяющийся в кишечнике людей. Именно здесь он осваивается и пробуравливает стенку органа, вызывая возникновение кровоточащих, гноящихся ранок. Цисты этого простейшего легко попадают в кровяное русло. Так происходит расселение по всему организму.
Как питаются паразиты этого вида? Точно так же, как и другие представители. Всасывают питательные вещества в кишечнике человека всей поверхностью тела. Размножается бесполым делением на несколько клеток. Формирует цисты, которые также паразитируют в человеке.
Заражение цистами простейшего происходит при контакте слизистых оболочек с грязными руками, при питье сырой воды. Заболевание, вызываемое этими существами, носит название балантидиоза. Сопровождается рвотой с кровью, поносом, слабостью, сильнейшими коликами в брюшной полости.
Трихомонады
Паразитические простейшие организмы, обитающие в мочеполовой системе женщин и мужчин. Сами по себе могут вреда не наносить, однако являются местом укрытия для болезнетворных бактерий и других патогенных организмов. Способны маскироваться под клетки человека, поэтому иммунная система сама уничтожить их не способна.
Заболевание, которое вызывает непосредственно трихомонада, называется трихомониаз. Оно передается половым путем и грозит бесплодием как мужчинам, так и женщинам.
Признаки заболевания у детей
Самое страшное — это когда появляются паразиты у детей. Симптомы их возникновения следующие:
- общая слабость;
- утомляемость;
- бледность;
- головные боли;
- снижение аппетита;
- плохой сон;
- раздражительность;
- жидкий стул;
- рвота и прочие.
Самое важное — обратить на них внимание и не пускать все на самотек. Тем более что, по последним данным, на первом месте среди заболеваний стоят именно паразиты у детей. Симптомы — верный признак того, что стоит задуматься и обратиться к врачу.
Bio-Lessons
Простейшие — одноклеточные организмы, строение которых можно изучить только с помощью микроскопа. В клетке такого организма располагается цитоплазма с одним или несколькими ядрами. В настоящее время науке известно около 70 тыс. видов простейших.
Одноклеточные беспозвоночные имеют простое строение, несмотря на это, такие процессы как обмен веществ, процессы пищеварения, дыхания, размножения и движения осуществляются так же как на уровне многоклеточного организма.
Следовательно, эта самостоятельная жизнеспособная клетка — целый живой организм. Одна единственная клетка выполняет у них функции всего организма. Движутся они с помощью ложноножек, жгутиков или ресничек. Обитают в пресной и соленой воде, почве, а также ведут паразитический образ жизни в теле человека и животных. Определенные виды простейших живут колониями.
Впервые одноклеточных рассмотрел под микроскопом голландский ученый Антони ван Левенгук.
Немаловажную роль играют большая численность и широкое распространение простейших в природе. Питаются они бактериями, одноклеточными водорослями или другими простейшими. В свою очередь, сами они служат пищей для множества животных. Особое внимание им уделяется при исследовании периодов возникновения и развития живых организмов на Земле, деталей возникновения мира многоклеточных беспозвоночных.
По способам передвижения и особенностям размножения простейших делят на 7 типов. Наиболее изучены саркомастигофоры, ресничные и споровики (рис.1).
Рис.1 Классификация простейших
Многие саркодовые обитают в морях и пресных водах (амеба) (рис.2), есть и паразитирующие виды (дизентерийная амеба).
Рис.2 Строение амебы обыкновенной
Некоторые саркодовые живут в почве и моховых подушках болот и лесов. Многие имеют внутренний или наружный (в виде раковинки: арцелла, радиолярия, диффлюгия) скелет (рис.3).
Рис.3 Раковинные саркодовые
У представителей корненожек форма клетки, постоянно меняясь, образует ложноножки. Это их органеллы захвата пищи и движения (рис.4).
Рис.4 Схема питания амебы обыкновенной
Размножаются корненожки простым делением клетки пополам (рис.5)
Рис.5 Деление амебы обыкновенной
При наступлении неблагоприятных условий (высыхание водоема, низкие температуры и т.п.) простейшие образуют защитную оболочку — цисту.
Представители жгутиковых имеют постоянную форму тела и передвигаются с помощью специальных выростов — жгутиков.
Трипаносома, лямблии и трихомонада являются паразитами и вызывают заболевания как у людей так и у животных. Лейшмания вызывает лейшманиозы, поражая кожу животных и человека. Переносчиком лейшмании является москит.
К колониальным формам жгутиковых относится вольвокс, который образует колонию в виде катящегося шара.
Представители имеющие хлоропласты (эвглена зеленая), питаются за счет фотосинтеза. Красный светочувствительный глазок позволяет им находить освещенные участки водоема (рис. 6).
Рис.6 Многообразие жгутиковых
У ресничных простейших (рис. 7) клетка устроена более сложно: имеются 2 ядра, клеточный рот, глотка. Сократительные вакуоли служат для удаления избытка воды из клетки.
Рис.7 Строение инфузории-туфельки
Размножаются ресничные как простым делением клетки, так и обменом ядрами. В частности в ходе сближения простейшие обмениваются малыми (генеративными) ядрами. Данный процесс представляет собой форму полового размножения — конъюгация (рис.8).
Рис.8 Конъюгация инфузории
Органом передвижения являются реснички. (инфузория-туфелька.) Форма тела у нее постоянная. Снаружи оно покрыто плотной тонкой защитной оболочкой.
Сувойки — одиночные или колониальные формы ресничных одноклеточных, ведущие прикрепленный образ жизни (рис.
Споровики — это класс паразитических простейших (рис. 10). У них нет органов захвата пищи и передвижения. В организм хозяина они попадают при кровососании (малярийный плазмодий) или с недоброкачественной пищей. Паразитируют в клетках, тканях и органах животных и человека.
Малярию переносят комары из рода Анофелес.
Рис.10 Схема развития малярийного плазмодия
1 — укус малярийного комара
2 — проникновение плазмодия в клетки печени
3 — первая стадия деления
4 — выход плазмодия из клеток печени с дальнейшим проникновением в кровяные тельца (цикличная стадия второго деления)
5 — зрелая стадия плазмодия (вновь попадает в тело комара)
У одноклеточных беспозвоночных простое строение. Эта самостоятельная жизнеспособная клетка — целый живой организм. Они питаются одноклеточными водорослями, бактериями или другими простейшими. По способам передвижения и особенностям размножения простейших делят на 7 типов. Рассмотрены 3 из них: саркомастигофоры, ресничные и споровики.
Одноклеточных животных — простейших изучает раздел зоологии протозоология.
Тест: «Одноклеточные животные»
Тест по биологии для обучающихся 7 классов на тему: «Одноклеточные животные». Составлен в 2-х вариантах.Данный тест позволяет проверить степень усвоения материала учащимися. Ключ к тесту прилагается.
Оценить
ФИО автора материала — Павлова Оксана Анатольевна
Место работы (название образовательного учреждения) — МБОУ «СОШ №22» г. Череповец
Должность автора — учитель биологии
Тема 1. Тип Простейшие
Внимательно прочитайте задание. Продумайте ответ и запишите его в тетради для проверочных работ.
I . Напишите определения следующих понятий: жгутики, порошица, органоиды.
II. Выберите правильный ответ:
Изучение зеленой эвглены позволяет сделать вывод о родстве растений и животных, так как она:
а) имеет светочувствительный глазок
б) дышит всей поверхностью тела
в) питается как растение и как животное
г) при дыхании поглощает кислород
2. Цистой называется…
а) состояние одноклеточных организмов, при котором образуется плотная оболочка
б) плотная оболочка клетки
в) название простейшего
г) название заболевания, вызванного простейшими
3. Какие из перечисленных простейших не являются паразитами?
а) малярийный плазмодий
б) дизентерийная амеба
4. Строение какого животного опровергает утверждение, что все простейшие — это
а) инфузории-туфельки в) фораминиферы
б) лямблии г) вольвокса
5. Поглощение кислорода у амёбы обыкновенной осуществляется…
а) пищеварительной вакуолью в) стигмой
б) сократительной вакуольюг) всей поверхностью тела
6. Какую функцию выполняет малое ядро у инфузории-туфельки?
а) дыханияв) пищеварения
б) участвует в половом процессег) регуляция обменных процессов
7. Цисты у одноклеточных выполняют функцию…
а) фотосинтезав) защиты и распространения
б) размноженияг) движения
III . Напишите название животного, изображенного на рисунке. Зарисуйте в тетрадь и подпишите органоиды.
IV . Найдите соответствие между названиями простейших и их особенностями:
Название простейшего
Характеристика
Б. Дизентерийная амеба
3. Одиночный организм
4. Передвижение при помощи ложноножек
5. Передвижение при помощи ресничек
6. Передвижение при помощи жгутиков
7. Два разных по величине ядра
8. Наличие хлорофилла
Тема 1. Тип Простейшие
I . Напишите определения следующих понятий: циста, ложноножки, сократительная вакуоль.
II . Выберите правильный ответ:
Только паразитический образ жизни ведут следующие простейшие…
а) саркодовые (корненожки) в) споровики
б) инфузории г) жгутиконосцы
2 . Малярийный плазмодий распространяется:
б) при помощи комара-анофелеса
в) при помощи мухи це-це
г) при помощи крупного рогатого скота
3. Какие простейшие животные могут питаться как растения и как животные?
а) саркодовые (корненожки) в) инфузории
б) жгутиконосцыг) споровики
4 . В организме какого простейшего находятся 2 ядра: большое и малое?
а) амеба дизентерийнаяв) амеба обыкновенная
б) эвглена зеленаяг) инфузория-туфелька
Органоидами движения у амёбы обыкновенной являются…
а) жгутикив) трихоцисты
б) ресничкиг) ложноножки
6. Какую функцию выполняет большое ядро у инфузории-туфельки?
Светочувствительный глазок есть у…
а) малярийного плазмодияв) амёбы
б) эвглены зелёнойг) лямблии
IV . Найдите соответствие между понятиями и характеристиками:
1. Животное, которое переносит паразита от одного животного или человека к другому
Б. Переносчик паразита
2. Организм, на котором обитает другое животное, приносящее вред первому
3. Животное, которое обитает в организме другого животного или человека и приносит ему вред
4. Малярийный комар
5. Малярийный плазмодий
6. Дизентерийная амеба
ПРОВЕРКА И ОЦЕНКА
Жгутики — органоиды движения жгутиконосцев.
Порошица — отверстие в мембране, через которое происходит выброс непереваренных частиц из клетки.
Органоиды — постоянные структуры клетки, которые выполняют определенные функции.
2 — большое ядро
3 — пищеварительная вакуоль
4 — сократительная вакуоль
Циста — состояние одноклеточных организмов, при котором образуется плотная оболочка.
Ложноножки — временные выросты цитоплазмы для передвижения корненожек.
Сократительная вакуоль — органоиды, выполняющие функцию выделения в организме простейших.
III . Амёба протей:
2 — сократительная вакуоль
У вас недостаточно прав для добавления комментариев
Чтобы оставлять комментарии, вам необходимо авторизоваться.
Если у вас еще нет учетной записи на нашем сайте, предлагаем зарегистрироваться.
Это займет не более 5 минут.
Для скачивания материалов с сайта необходимо авторизоваться на сайте (войти под своим логином и паролем)
Если Вы не регистрировались ранее, Вы можете зарегистрироваться.
После авторизации/регистрации на сайте Вы сможете скачивать необходимый в работе материал.
Заказать рецензию на методическую разработку
можно здесь
Материал интересный и познавательный, имеете практическую значимость. Необходимо детям доступно объя. Подробнее.
С удовольствием ознакомилась Вашей разработкой. Материал понравился, очень трогательный. Думаю, что . Подробнее.
С удовольствием прочитала Ваше эссе и порадовалась, что работа приносит вам удовольствие. К сожалени. Подробнее.
Очень полезный материал. Всем нам постоянно приходится готовить материал для лекций, для выступления. Подробнее.
Оказание первой помощи в образовательных учреждениях Пройти обучение
Благодарность руководству образовательного учреждения за поддержку и развитие профессионального потенциала педагогического работника
Диплом за отличное владение и эффективное применение современных педагогических методик в условиях реализации ФГОС
- Свидетельство о регистрации средства массовой информации ЭЛ № ФС 77 — 58841 от 28 июля 2014 года выдано Федеральной службой по надзору в сфере связи, информационный технологий и массовых коммуникации (Роскомнадзор).
- Лицензия на осуществление образовательной деятельности № 4276 от 19.11.2020 года. Серия 78 ЛО № 0000171 Выдана Комитетом по образованию Правительства Санкт-Петербурга
- В соответствии с Федеральной целевой программой развития системы образования на 2011-2015 гг. и проектом концепции федеральной целевой программы развития образования на 2016-2020 гг.
Сертификат соответствия качества предоставляемых услуг рег. № 04 ЕАС1.СУ.01217 от 19.11.2019. Услуга: Дополнительное профессиональное образование.По результатам оценки оказания услуг, оценки процесса оказания услуг и проверки результатов оказываемых услуг данный документ подтверждает соответствие предоставляемых ООО «Центр Развития Педагогики» услуг всем нормативным требованиям.
Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания — обратитесь к врачу.
Читайте также:
- Помогают ли народные средства при борьбе с ОРВИ и гриппом?
- Скорая помощь при отравлении
- Прививочная кампания против гриппа и пневмонии активно проходит в Республике Алтай
- Cимптомы ВИЧ, заражение и распространение
- Рубрика здоровье: укрепляем иммунитет
Саркодовые — общая характеристика класса, строение, жизненный цикл » Kupuk.
netСаркодовые — группа одноклеточных организмов, которая ранее рассматривалась в качестве класса или подтипа. К ним относятся простейшие, не имеющие постоянной формы тела, они покрыты мембраной, состоящей из белков и липидов. Микроорганизмы не имеют оболочки, но они могут образовывать раковину или внутренний скелет.
Общее строение
Класс саркодовые представляет собой большую группу микроорганизмов, который объединяет более 11 тыс. видов, отличающихся большим разнообразием форм. Он включает простейших, которые ведут различный образ жизни.
Большую часть составляют морские обитатели, но существуют и пресноводные микроорганизмы. Некоторые простейшие могут существовать в грунте, мхе торфяных болот и лесов. Реже встречаются паразитические представители, численность которых составляет чуть больше одного процента от остальных микроорганизмов.
Тип саркодовые постоянно изменяет внешний вид, так как происходит формирование псевдоподий (ложноножек).
Если использовать электронный микроскоп, то можно увидеть тончайшую мембрану, которая имеет фибриллярное строение. Плазмалемма саркодовых представителей формирует два слоя:
- наружную плотную эктоплазму;
- внутреннюю жидкую эндоплазму.
В течение жизненного цикла оба слоя легко взаимодействуют между собой, кроме того, клеточная мембрана способна разрушаться и восстанавливаться. По-видимому, такая изменчивость слоев плазмалеммы и отсутствие плотного покрытия объясняет неопределенность конфигурации тела саркодовых.
Часть микроорганизмов обладает внутренним или наружным скелетом в виде раковин. Передвижение простейшие осуществляют с помощью псевдоподий, или ложноножек. Они представляют собой отростки разной конфигурации и предназначены также для захвата пищи.
В питание микроорганизмов входят водоросли и обыкновенные простейшие, которые перевариваются вакуолями, образующимися в цитоплазме. Это пузырьки жидкости с пищеварительными ферментами, которые выделяет плазмалемма вокруг пищевых частиц.
У пресноводных микроорганизмов имеются органы выделения — сократительные вакуоли. Саркодовые делятся на 5 отрядов:
Первые 3 отряда относятся к корненожкам, а лучевики и солнечники рассматриваются не как отряды, а как подклассы.
Отряд корненожек
Самым простым микроорганизмом, относящимся к корненожкам, считаются голые амебы. Но особенно часто встречаются обыкновенные простейшие, или протей.
Амеба обыкновенная
Обыкновенная амеба обитает в пресных водах, маленьких прудах и канавах с илистым дном, кроме того, микроорганизмы легко размножаются в условиях лаборатории. Это самый крупный представитель среди существующих простейших. Когда он передвигается, то может вырасти до 0,5 мм и его видно невооруженным глазом.
Если смотреть за амебой через микроскоп, то можно заметить несколько длинных ложноножек, напоминающих лопасти. Форма тела саркодовых постоянно изменяется, часть псевдоподий втягивается внутрь, а некоторые, наоборот, удлиняются. Ложноножки в нескольких точках закрепляются к почве, что помогает амебе передвигаться.
Если простейшее сталкивается с пищевой органикой, то оно обхватывает всю частицу и помещает ее внутрь плазмалеммы вместе с жидкостью. Так в теле образуются пузырьки с пищей, которые называют пищеварительными вакуолями, и в них происходит процесс переваривания органики.
Кроме того, в цитоплазме обыкновенного простейшего периодически появляется прозрачный пузырек. Это сократительная вакуоль, которая выполняет для микроорганизма важные жизненные задачи. Она набирает в себя воду из цитоплазмы, увеличиваясь в размерах, а затем сокращается и выбрасывает ее через отверстие наружу.
Вся эта процедура происходит в течение 5—8 минут. В теле амебы количество органических и неорганических веществ выше, чем в окружающей жидкости, поэтому вода поступает в протоплазму микроорганизма. Благодаря сократительной вакуоли, ее избыток выбрасывается наружу. Если бы этого не происходило, то амеба просто растворилась бы в воде.
Вместе с жидкостью выделяются продукты обмена веществ, то есть сократительная вакуоль принимает участие в процессе выделения. С водой в микроорганизм поступает кислород, таким образом, органоид помогает простейшему организму дышать. Как и в любой клетке, в теле амебы находится ядро, которое практически не видно, поэтому для его обнаружения специалисты применяют красители.
Размножение микроорганизма происходит единственным способом — это деление на две клетки при свободном передвижении. Оно происходит через центр ядра, которое разделяется на две половины. Периодичность этого процесса зависит от температуры обитания, питания и происходит один раз через 1—2 суток.
Паразитические микроорганизмы
Некоторые виды простейших могут вести паразитический образ жизни, поселившись в кишечнике позвоночных и беспозвоночных животных. У человека в кишечной полости обитают пять видов микроорганизмов. Четыре из них считаются безобидными, но пятый, который носит название дизентерийной амебы, может вызывать тяжелое заболевание — амебиаз.
Этот вид простейших представляет собой очень мелкие микроорганизмы (20—30 мк), которые живут в толстом кишечнике. Основной их характеристикой считается высокая подвижность. Амеба существует практически во всех уголках мира, но заболевание амебиаз встречается довольно редко, только в южных и теплых регионах.
Это объясняется тем, что наличие паразитического микроорганизма не всегда сопровождается заболеванием. В этом случае дизентерийная амеба выполняет функцию носительства, то есть она передвигается в кишечной среде, питается бактериями, но не вызывает патологических изменений. Но иногда микроорганизм активно начинает проникать в стенки кишечника.
Такое действие вызывает у человека тяжелую форму кровавого поноса. В этот период амеба меняет объект питания, начиная поедать красные кровяные клетки вместо бактерий. Если не пройти курс лечения, то амебиаз перейдет в хроническую форму, вызывая у человека сильное истощение.
Дизентерийные микроорганизмы могут жить только в организме человека, а попадая в почву или воду, они сразу погибают. Заражение осуществляется непостоянными формами существования — цистами. Когда простейшие из толстого кишечника попадают в прямую кишку, то они меняют свою форму. Они втягивают ложноножки, выкидывают пищевые частицы и становятся круглыми, а эктоплазма образует тонкое, но прочное покрытие.
Этот процесс называется инцистирование. Особенностью этого момента считается деление ядра последовательно два раза, образуя характерные четырехъядерные цисты. В таком виде они с фекалиями выводятся наружу, где продолжают существовать в течение 2—3 месяцев. Если паразитическое простейшее попадает в организм человека, то оно эксцистируется.
Тело разделяется два раза, при этом ядро не затрагивается. В итоге образуются четыре одноядерных микроорганизма, которые приступают к активной жизнедеятельности. К паразитическим микроорганизмам относят и ротовую амебу, которая обитает в полости рта человека.
Подкласс лучевики
Самый многочисленный по числу видов морской подкласс в биологической систематике саркодовых составляют лучевики, или радиолярии. Он насчитывает более 7—8 тыс. видов. Лучевики относятся к планктонным организмам, которые в основном живут в морских водах тропических и субтропических зонах.
Размеры тела радиолярий колеблются в пределах от 50 мк до 1 мм и более. Существуют некоторые объединения саркодовых, обладающих гораздо крупным телом. Значительная часть радиолярий имеет форму сферы с характерной капсулой в центре. Это органическая мембрана, расположенная вокруг цитоплазмы с ядром.
Стенки капсулы обладают множеством отверстий, через которые происходит сообщение внутрикапсулярной плазмалеммы с экстракапсулярной. Мембрана, расположенная в центре тела, считается скелетным образованием, которое защищает плазмалемму с ядром. Довольно широкую зону у радиолярий образует эктоплазма, которая обладает многочисленными включениями.
Основная их масса — это слизь, образующая мощный слой. Кроме слизи, в цитоплазме лучевиков встречаются капли жира. Все эти образования облегчают саркодовые и помогают им свободно передвигаться в морском пространстве. У некоторых радиолярий внутри цитоплазмы встречаются одноклеточные водоросли из отряда панцирных жгутиковых.
Это доказывает процесс симбиоза простейших организмов с растительным миром. Водоросли в теле простейших получают питательные вещества и углекислый газ, который образуется в результате дыхания. Подкласс лучевиков делится на четыре отряда:
Все они отличаются типичными формами скелетных образований. Недостаточно изучены процессы размножения лучевиков, хотя ученые длительное время занимались исследованием этих простейших. Проблема в том, что радиолярии долго не живут в лабораторных условиях, поэтому не удается получить полные результаты.
Скелетное образование не позволяет им делиться на две части как в случае с амебами. Скорее всего, формирование зародышей с одним ядром у них происходит как при бесполом размножении фораминифер. Этот процесс у радиолярий требует продолжения дальнейших исследований.
Псевдопод – определение, функции и примеры
Псевдопод Определение
Псевдопод происходит от греческих слов pseudes и podos , что означает «ложные» и «ноги» соответственно. Они представляют собой проекции цитоплазмы одноклеточных протистов или мембраны эукариотической клетки. Поверхность клетки, часто образованная филаментной структурой и микротрубочками, проецирует мембрану в процессе, называемом ламеллиподием, который поддерживается микрофиламентами.
Так как же передвигаются ложноножки? Затем нити на конце клетки взаимодействуют с миозином, который вызывает сокращение, приводящее к движению. Цитоплазма заполняет ламеллиподию, образуя выступы, или ложные. Эти выступы расширяются и сжимаются под действием полимеризации актина, которая толкает клетку вперед. Представьте ложноножки в виде каплевидных монстров, которых вы видите в научно-фантастических фильмах, только они микроскопические и совсем не чудовищные. Они являются частью адаптации некоторых животных, чтобы они могли эффективно перемещаться в окружающей среде.
Типы псевдоножек
Псевдоножки бывают разных форм и типов. Фактически, организмы, способные производить эти выступы, различаются в зависимости от того, к какой классификации они принадлежат, поэтому структура и форма выступов определяют его таксономические характеристики, что делает формы выступов уникальными. Чтобы лучше понять, ниже приведены различные типы форм ложноножек.
Лобоподии
Лобоподии относятся к пальцеобразным ложноногам и являются наиболее распространенным типом, наблюдаемым в природе. Они характеризуются короткими, тупыми и выпуклыми выростами, содержащими как эндоплазму, так и эктоплазму организма. Примером могут служить лопастные амебы, считающиеся самыми крупными из всех псевдоподий.
Филоподии
Отличающиеся наличием тонких и нитевидных ложноножек, филоподии способны разветвляться или анастомозировать. Они нитевидные и тонкие с заостренными концами, которые в основном состоят из эктоплазмы. Выступы поддерживаются микрофиламентами. Примеры включают Lecithium и Euglypha . Интересно, что этот тип филиподий имеет разные вариации. Например, гранулоподии похожи на филоподии, но имеют зернистую структуру, называемую экструсомами, которые больше специализируются на захвате добычи, чем на обеспечении подвижности. Другой разновидностью гранулоподий являются гранулоретикулоподии, представляющие собой нечто среднее между филиподиями и ретикулоподиями. Обычен среди видов Аллогромия, тип одноклеточных эукариот.
Ретикулоподии
Представьте, что псевдоподии используются как средство для связи с другими клетками. Ретикулоподии — это специализированные псевдоподии, которые сообщаются с другими псевдоподиями, создавая таким образом сеть, называемую ретикулумом. Также называемые ретикуло-псевдоподиями, они создают сложные выступы, в которых отдельные псевдоподии сливаются друг с другом, создавая нечто, похожее на неправильную сеть. Этот тип псевдоподия также функционирует как при приеме пищи, так и при передвижении. Примерами ретикулезных псевдоподий являются Фораминофераны .
Аксоподии
Аксоподии — это псевдоподии, поддерживаемые массивами микротрубочек. Псевдоподии покрыты цитоплазмой; таким образом, он больше всего используется для фагоцитоза или проглатывания пищевых частиц. Протисты из рода Radiolaria и Heliozoa являются примерами псевдоподиума этого типа.
На этом изображении изображена гигантская амеба из рода Chaos с тремя огромными псевдоножками лопастного типа, радиально выступающими из ее тела.
Функции псевдоподов
Псевдоподы выполняют две основные функции: (1) передвижение и (2) захват добычи или поглощение пищи. Например, амеба может ползать за счет расширения цитоплазмы и сокращения нитей. Псевдоподвыпуклость выходит наружу от края клетки, чтобы втянуть весь организм по мере его продвижения вперед.
С другой стороны, он также используется для захвата и проглатывания добычи. Они также используются для проглатывания твердых частиц, обеспечивая при этом мобильность во время охоты за едой. Это также необходимо для обнаружения добычи поблизости, что помогает таким организмам, как амеба, поглощать материю в процессе фагоцитоза. В этом процессе выступы иглы окружают пищевую частицу, создавая окруженный мембраной мешок, который отщипывается, чтобы создать пищевую вакуоль до того, как пища полностью переварится.
Примеры псевдоножек
Несколько родов царства Protista используют псевдоножки для передвижения и приема пищи. Протисты не относятся ни к растениям, ни к животным, и у них есть определенные характеристики, которые делают их достойными того, чтобы для них было зарезервировано собственное царство.
Хотя ложноножки часто классифицируются как признаки простейших, они не ограничиваются такой классификацией. Даже клетки высших животных образуют псевдоподии. Например, лейкоциты позвоночных животных используют ложноножки для поглощения инородных частиц, таких как бактерии и вирусы, в процессе, называемом фагоцитозом. Ниже приведены примеры псевдоподий.
Rhizopods
Pseudopodia является характеристикой группы простейших организмов, называемых корненожками, в царстве Protista. Они характеризуются как эукариотические клетки, мобильность которых зависит от псевдоподий. Они также используют свои псевдоподии, чтобы поглощать частицы пищи внутри вакуоли. Примеры корненожек включают Amoeba proteus, Entamoeba histolytica, Radiolarians, и Foramineferans . Эти корненожки биологически значимы. Например, скелеты Foramineferans составляют большую часть мела и известняка на планете, в то время как Entamoeba histolytica вызывает амебную дизентерию.
Лейкоциты
Лейкоциты являются важными компонентами нашей иммунной системы. Также называемые лейкоцитами, они помогают организму бороться с инфекцией, атакуя бактерии, вирусы и другие патогенные организмы, проникающие в организм. Фагоцитарные лейкоциты, такие как моноциты и нейтрофилы, образуют ложноножки, чтобы атаковать и поглощать патогены. Псевдоножки также помогают лейкоцитам ползать внутри тела подобно возбудимым системам. Это означает, что движение лейкоцитов отклоняется в сторону области, где, вероятно, будет больше лейкоцитов, атакующих конкретную инфекцию.
На этом изображении нейтрофил (желтый) атакует бактерию сибирской язвы оранжевой палочки своей псевдопией.
- Псевдоподиум – Одиночный временный отросток клетки лейкоцитов и простейших, используемый для передвижения и питания.
- Фагоцитоз – Проглатывание материалов, таких как бактерии, вирусы и разрушенные клетки, амебоидами и фагоцитами, такими как лейкоциты.
- Полимеризация актина – Это относится к росту актина, который представляет собой семейство функциональных белков, образующих микрофиламенты в эукариотических клетках, влияющих на формирование ложноножек для подвижности.
Тест
1. Что такое ложноножки?
A. Тип эукариотической клетки под царством Protista
B. Выступ, также называемый ложной ножкой цитоплазмы, как протистов, так и мембраны эукариотической клетки
C. Часть цитоплазма мелких организмов
D. Важная часть клетки, отвечающая за успешное деление клетки.
Ответ на вопрос №1
B правильно. Псевдопод относится к цитоплазматическому выступу, который присутствует как в эукариотических, так и в протистических клетках.
2. Каковы функции псевдоподий?
A. Locomotion and phagocytosis
B. Locomotion and defense
C. Phagocytosis and cell division
D. Phagocytosis and cell memory
Answer to Question #2
A is correct . Функция псевдоподий заключается в обеспечении подвижности организмов. Он также используется для фагоцитоза для поглощения пищи и других материалов.
3. Псевдоподии встречаются только у амебоидных простейших.
A. Верно
B. Неверно
Ответ на вопрос №3
Неверно. Помните, что псевдоподии встречаются не только у амебоидных простейших, но и у сложных клеточных структур, таких как лейкоциты или эритроциты.
Определение, функция, движение и примеры
Определение, функция, движение и примеры
Определение: Что такое псевдоподы?
Также известные как псевдоподии (существительное в единственном числе: псевдоподиум), псевдоподии представляют собой временные расширения цитоплазмы (также называемые ложными стопами), используемые для передвижения и ощущения. Их можно найти у всех саркодинов, а также у ряда жгутиковых простейших, которые существуют либо как паразиты, либо как свободноживущие организмы.
Сколько ног у трилобита?
Пожалуйста, включите JavaScript
Сколько ног у трилобита?
У высших животных ложноножки можно наблюдать в ряде лейкоцитов (фагоцитарных клеток), которые используют структуру для улавливания и уничтожения вторгшихся микробов. В зависимости от типа клетки различают четыре основных типа, которые не только различаются по внешнему виду (и общей морфологии), но и выполняют разные функции.
Например, у некоторых организмов псевдоподии содержат микротрубочки, которые вносят значительный вклад в движение клеток.
There are four types of pseudopods that include:
- Lobopodia
- Axopodia
- Filopodia
- Reticulopodia/Rhizopoda
Types of Pseudopodia
Filopodia
Filopodia представляют собой тонкие структуры на основе актина, которые выполняют сенсорные и двигательные функции. Как и другие псевдоподии, филоподии представляют собой клеточные выпячивания и, таким образом, выходят за пределы клеточной поверхности. Однако по сравнению с псевдоподиями, обнаруженными в одноклеточных организмах, филоподии в основном встречаются в некоторых клетках многоклеточных организмов, где они проникают во внеклеточный матрикс и участвуют в передаче сигналов.
* Некоторые одноклеточные организмы, такие как представители рода Dictyostelium, используют для питания филоподии.
Формирование филоподий
Формирование филоподий начинается с зарождения актиновых филаментов под влиянием нуклеаторов (группы белков). Хотя для объяснения инициации (индукции) филоподий были предложены две модели, процесс, по-видимому, запускается связыванием GTPase Cdc42 с важным регулятором, известным как N-WASP.
Это приводит к активации N-WASP, который, в свою очередь, связывается с Profilin и Arp2/3 с образованием комплекса, который запускает формирование новой псевдоножки.
Хотя были предложены две модели инициации филоподий, это модель конвергентной элонгации и модель нуклеации кончика, дальнейшие исследования показали, что они не исключают друг друга.
Согласно этим исследованиям, две модели могут фактически сосуществовать, особенно если учесть разнообразный и изменчивый характер этих структур.
Что касается строения, филоподии представляют собой тонкие цилиндрические выступы диаметром от 100 до 200 нм и длиной 10 мкм. Однако некоторые филоподии на клеточной поверхности очень короткие, едва выступающие из клеточной поверхности. Эти филоподии известны как микрошипы.
Актиновые филаменты (от 10 до 30) составляют центральное ядро структуры. Здесь нити плотно упакованы вместе параллельным образом, образуя стержень псевдоподии.
Внутри филоподий нити перекрываются и выстраиваются в единой полярности; где зазубренный конец нити ориентирован к филоподиальному концу структуры. Колючки заходят в концевой комплекс, состоящий из актин-связывающих белков и филаментов. В области основания псевдоподии филаменты впадают в актиновую сеть, расположенную под клеточной мембраной.
В то время как добавление актиновых мономеров к актиновым филаментам происходит на кончике структуры по мере ее удлинения, было показано, что филаменты постоянно возвращаются назад к основанию через ретроградный поток, который зависит от миозина. Именно скорость между этим потоком и добавлением мономера вверху влияет на общую скорость роста.
Основываясь на исследованиях электронной томографии, результаты показали, что этот рост происходит со скоростью около 0,2 мкм/с (которая может увеличиваться примерно до 25 мкм в минуту) до того, как структура достигает критической длины. В этот момент филоподии либо производят новые актиновые структуры, либо начинают втягиваться.
Достигнув критической длины, некоторые филоподии связываются и сливаются с плазматической мембраной, образуя пучки актина.
* Филоподии достаточно гибкие, чтобы волноваться во внеклеточном матриксе. Тем не менее, они также достаточно прочны, что позволяет поддерживать структурную целостность, даже если длина конструкции превышает 30 мкм.
Некоторые из наиболее распространенных типов филоподий включают:
· Цитонемы. Обнаруженные в крыльях видов дрозофилы, этот тип филоподий может достигать более 800 мкм, где они участвуют в межклеточном взаимодействии
· Миоподии — тип филоподий, которые можно обнаружить на клеточной поверхности мышечных клеток, где они смешиваются с другими типами филоподий.
Функции
В многоклеточных организмах филоподии выполняют ряд физиологических функций, включая заживление ран, передачу клеточных сигналов, а также развитие клеток. Учитывая, что филоподии простираются во внеклеточный матрикс, они способны ощущать химические вещества в своем окружении, что, в свою очередь, позволяет клетке реагировать соответствующим образом.
Здесь рецепторы внутри филоподий получают химическую информацию во внеклеточном матриксе, которая затем передается вниз в клетку (по сигнальному пути).
Во внеклеточном матриксе филоподии могут идентифицировать мишени, необходимые для адгезии, что позволяет генерировать направляющие сигналы, а также тяговые силы, которые в конечном итоге способствуют движению клетки. Благодаря этому процессу клетки способны к таким действиям, как нахождение путей аксонов и адгезия к эпителиальным клеткам, что способствует миграции клеток.
* Филоподии также способствуют застегиванию адгезии, процессу, в котором они участвуют в выравнивании и адгезии клеток, что уменьшает зазор между клетками.
Аксоподии
Подобно филоподиям, аксоподии представляют собой длинные и тонкие отростки клеток. Однако они более жесткие (и поэтому кажутся игольчатыми), чем филоподии, которые имеют тенденцию быть более гибкими по своей природе. Их можно найти на клеточной поверхности различных организмов (например, представителей типа Antinopoda), где они участвуют в питании и передвижении.
У этих организмов аксоподий (Sin. axopodium) происходит из аксопластов (мембранная полость вблизи ядра, состоящая из микрофибриллярного материала и гранул). Здесь микрофибриллы организуются для построения стенок микротрубочек, которые затем образуют параллельные ряды, соединенные звеньями.
На основании микроскопических исследований было показано, что эти микротрубочки образуют взаимосвязанные двойные катушки (примерно 500 трубочек образуют около 5 витков катушки).
Микротрубочки составляют ядро аксонемы (которая является центральной частью аксоподий) и проходят по всей длине структуры. Помимо микротрубочек, структура также состоит из цитоплазмы, которая несет такие органеллы, как митохондрии, к цитосоме и от нее.
* В клетке аксоподии выходят из аксопластов через поры, расположенные на стенке капсулы. В зависимости от организма эти поры различаются по размеру и количеству. В то время как было показано, что полицистины содержат многие из этих пор, феодарианы содержат около трех из этих пор.
Показано, что укорочение аксоподий происходит во время питания. Например, было показано, что после захвата пищи быстрое сокращение происходит из-за разрушения микротрубочек. После сокращения аксоподы начинают удлиняться с нормальной скоростью, пока не достигнут нормальной длины.
Некоторые из основных характеристик Axopodia включают в себя:
· жесткие — как таковые, они более устойчивы к изгибе по сравнению с другими псевдоподами
· тонкая и удлиненная — игловая лайка
40214 · . · При неблагоприятных условиях они могут втягиваться (быстро сокращаться), когда филаменты реабсорбируются после диспергирования фибрилл в аксоплазме
· Микротрубочки аксоподий имеют диаметр около 220 A
· Аксоподии также теряются во время клеточного деления
Функции
Питание
Для таких организмов, как представители класса Actinopoda, аксоподии играют важную роль в питании. Как уже упоминалось, аксоподии имеют на своей поверхности клейкое вещество, вырабатываемое мукоцистами. Кроме того, у них также есть кинетоцисты, которые выбрасывают нитевидные структуры, которые эффективно ловят их добычу.
Используя эти экструсомы, эти организмы способны улавливать пищевой материал (или добычу), который затем транспортируется в тело клетки через цитоплазматический поток. В то время как более мелкая добыча может быть поймана и захвачена одним аксоподием, более крупная запутывается в нескольких аксоподиях. В некоторых случаях было показано, что несколько особей участвуют в отлове более крупной добычи.
Поддержка и передвижение
Было показано, что помимо питания аксоподии помогают простейшим удерживать положение в воде и даже способствуют передвижению. Например, благодаря контролируемому изменению длины аксопод некоторые гелиозоа оказались эффективно поперечными в водной среде. Это у других организмов достигается за счет расширения и сокращения эктоплазматических вакуолей, расположенных между аксоподиями.
Здесь организм может оставаться на месте или контролировать направление движения. Во время клеточного деления и аксоподии, и эктоплазма теряются, в результате чего организм опускается на дно.
Некоторые из других примечательных функций Axopodia включают в себя:
- Транспортировка кремнезема, полученная из некоторой добычи
- транспортировки вакуолей
Reticulopodium
Reticulopodium
92
Reticulopodium 3
44492
. в некоторых книгах ретикулоподии представляют собой нитевидные псевдоподии, которые разветвляются и сливаются, образуя чрезвычайно динамичную сеть. Как и в случае с аксоподиями, ретикулоподии также состоят из канальцев и цитоплазмы.
Их можно найти в ряде организмов, включая амебы Endomyxa и некоторые фораминиферы (древняя группа простейших). У этих организмов ретикулоподии участвуют в питании и передвижении.
Как и аксоподии, ретикулоподии также состоят из микротрубочек и цитоплазмы. Здесь микротрубочки, из которых состоят псевдоподии, состоят из уникального типа тубулина, известного как бета-тубулин типа 2. Этот тубулин образует спиральные филаменты (HF), которые являются основой микротрубочек, обнаруживаемых в фораминиферовых ретикулоподиях.
У фораминифер и других организмов ретукулоподии выпячиваются через одну или несколько пор (апертуальные отверстия). Первоначально эти псевдоподии могут быть тонкими и заостренными (по внешнему виду похожи на филоподии).
По мере увеличения количества цитоплазмы в структуре псевдоподиальный ствол, известный как ножка, становится толще и разветвляется, образуя новые псевдоподии. В то время как эти псевдоподии растут и анастомозируют (связываются вместе), они образуют сеть, напоминающую паутинные нити.
Некоторые из основных характеристик ретикулоподий включают:
· Отличаются от других тем, что они сильно разветвлены и образуют анастомозирующие сети
· В зависимости от организма тело клетки организма
· Может быстро расширяться и втягиваться со скоростью около 20 мкм/с
· известный как бисерный ответ
· Внутри ложноножек транспорт частиц двунаправленный. Это означает, что цитоплазма течет по длине ложноножек к телу клетки и обратно
Функции
Как и аксоподии, ретикулоподии играют важную роль в питании и передвижении. Однако их основная функция заключается в сборе пищи и кормлении. Во время кормления организм распространяет ложноножки (выглядящие как неправильная паутина), которые очищают их непосредственную поверхность и собирают доступный пищевой материал для проглатывания.
Добычей могут быть такие одноклеточные организмы, как бактерии, которые попадают в паутину и попадают в пищевые вакуоли для пищеварения. Помимо питания, ретикулоподии также используются для передвижения. Однако это не является их основной функцией.
Lobopodia
Lopodium — наиболее распространенный тип, встречающийся у таких организмов, как Amoeba proteus. Для лобопод характерны пальцевидные трубчатые псевдоподии, состоящие из экто- и эндоплазмы. Однако было также показано, что они содержат актин и миозин (микрофиламенты), которые способствуют общему движению.
В отличие от других ложноножек микротрубочки лобоподий развиты слабо. У многих амеб лобоподии в первую очередь участвуют в передвижении.
Формирование и передвижение
У таких организмов на формирование лобоподий влияют химические сигналы в окружающей среде. В присутствии пищевого вещества химические сигналы влияют на направление движения амебы. Здесь молекулы (из пищевого материала) связываются с рецепторами, расположенными на клеточной мембране организма, что стимулирует образование филаментов за счет агрегации глобулярного актина.
При добавлении глобулярного актина структура (филамент) продолжает удлиняться, что, в свою очередь, вызывает выпячивание мембраны (это действие приводит к образованию псевдоподий). Выступающие лобоподы по мере своего расширения заполняются цитоплазмой. В случае исчезновения молекул глобулярный актин распадается, что останавливает дальнейшее удлинение ложноножек.
Если молекулы сохраняются, миозин, который действует как моторные белки, взаимодействует с актином, толкая тело клетки в направлении ложноножки.
* Миозиновая активность (в качестве двигателей) требует энергии (АТФ).
* Также было показано, что вязкость цитоплазмы изменяется при ее втекании и вытекании из ложноножек.
Во время кормления лобоподии также окружают пищевой материал и поглощают его внутри пузырька, где на него воздействуют различные ферменты. Затем продукты жизнедеятельности выводятся через вакуоли, открывающиеся в окружающую среду.
Вернуться на главную страницу Protozoa
Возврат в Цитоплазму
Возврат из Pseudopods в MicroscopeMaster домой
сообщить об этом объявлении
Справки
Чансон Ян и Татьяна Свиткина. (2011). Инициация филоподий: сосредоточьтесь на комплексе Arp2/3 и форминах. нкби.
Hou Y et al. (2013). Молекулярные доказательства неофункционализации β-тубулина у Retaria (фораминиферы и радиолярии). NBCI.
Кара Роджерс. (2011). Грибы, водоросли и протисты.
П. Де Вевер, П. Думитрика, Дж. П. Коле, К. Нигрини и М. Каридройт. (2001). Радиолярии в осадочной летописи.
Сэмюэл С. Баузер и Джеффри Л. Трэвис. (2000). Методы структурных исследований ретикулоподий, «мягкой части» жизненно важных фораминифер. jstor.org.
Стефани Л. Гуптон и Фрэнк Б. Гертлер. (2007). Филоподия: Пальцы, которые ходят. ResearchGate.
Ссылки
https://royalsocietypublishing.org/doi/10.1098/rsos.160283
https://www.researchgate.net/publication/44576108_Understanding_eukaryotic_chemotaxis_A_pseudopod-centred_view
Узнайте, как размещать рекламу на MicroscopeMaster!
Псевдоподоподобные базальноклеточные отростки в холецистокининовых клетках кишечника
- Список журналов
- Рукописи авторов HHS
- PMC4846361
Рез. клеточной ткани. Авторская рукопись; доступен в PMC 2016 26 апреля. 2010 август; 341(2): 289–297.
Опубликовано онлайн 2010 июня 26. DOI: 10.1007/S00441-010-0997-1
PMCID: PMC48446361
NIHMSID: NIHMS774321
PMID: 20582553
Авторская информация и лицензия Deckport.0009
- Дополнительные материалы
Холецистокинин (ХЦК) секретируется нейроэндокринными клетками, составляющими 0,1–0,5% клеток слизистой оболочки верхних отделов тонкой кишки. Используя экспрессию зеленого флуоресцентного белка (GFP), управляемого промотором CCK, у трансгенных мышей, мы применили методы иммунофлуоресценции для анализа морфологии клеток CCK. GFP и CCK совместно локализуются в нейроэндокринных клетках с небольшой аберрантной экспрессией GFP. Клетки, содержащие CCK, имеют форму колбы или веретена, и в некоторых клетках мы обнаружили дендритные отростки, подобные псевдоподиям, продемонстрированным для D-клеток кишечника, содержащих соматостатин. Большинство псевдоподий короткие, самый длинный видимый отросток проходит через три клетки. Псевдоподы обычно распространяются на соседние клетки, но некоторые переплетаются между соседними клетками. Наблюдаются также двойственные процессы. Трехмерные реконструкции предполагают, что процессы не являются однонаправленными и, таким образом, маловероятно, что они участвуют в миграции клеток CCK из крипт вверх по ворсинкам. Обильное иммуноокрашивание CCK присутствует в ложноногах, что позволяет предположить, что они высвобождают CCK на клетку-мишень. Чтобы определить тип клеток-мишеней, мы совместно окрашивали срезы антителами к хромогранину А, трилистнику-фактору-3 и сахаразе-изомальтазе. Клеточные процессы CCK почти исключительно распространяются на сахаразо-изомальтазо-положительные энтероциты. Таким образом, клетки CCK имеют клеточные отростки, возможно, вовлеченные в паракринную секрецию.
Ключевые слова: Холецистокинин, Зеленый флуоресцентный белок, I-клетки, Псевдоподы, Паракринные, Мыши, трансгенные в ответ на внутрипросветные питательные вещества (Liddle 1997). Физиологические действия CCK включают стимуляцию секреции поджелудочной железы и сокращение желчного пузыря, регуляцию опорожнения желудка и индукцию чувства насыщения (Liddle 19).94). Основываясь на исследованиях иммуноцитохимического окрашивания на световом и электронно-микроскопическом уровнях, а также с помощью просвечивающей электронной микроскопии, долгое время считалось, что питательные вещества в просвете тонкой кишки взаимодействуют с апикальной щеточной каймой клеток CCK. Это взаимодействие затем должно было бы привести к неизвестному механизму передачи цитоплазматического сигнала, что привело бы к экзоцитозу CCK из базолатеральной плазматической мембраны I-клеток и последующему распространению по всему телу либо локально путем диффузии, либо системно за счет объемного потока в системе кровообращения. Ультраструктура клеток CCK поддерживает эту концепцию, потому что эти клетки простираются от базальной мембраны эпителия слизистой оболочки до просвета кишечника и имеют апикальные микроворсинки (щеточная кайма), которые хорошо подходят для взятия проб содержимого просвета, и базально локализованную мембрану. связанные цитоплазматические гранулы, содержащие CCK, которые хорошо расположены для базолатеральной секреции путем экзоцитоза (Buchan et al. 19).78).
Недавно стала доступной новая модель для изучения клеток CCK в головном мозге; мы использовали эту модель для изучения CCK в кишечнике. Трансгенные мыши с CCK-зеленым флуоресцентным белком (GFP) были разработаны Региональным ресурсным центром мутантных мышей (MMRRC, Университет Миссури). У этих мышей повышенная экспрессия GFP управляется промотором CCK, что приводит к зеленой флуоресценции только в клетках CCK (Samuel et al. 2008). В дополнение к CCK-продуцирующим клеткам в головном мозге эндокринные CCK-клетки в тонком кишечнике этих мышей хорошо видны в флуоресцентном микроскопе. В процессе оценки распределения этих клеток по кишечнику мы заметили у многих из них необычную особенность строения, а именно наличие базальноклеточного отростка, напоминающего ложноножку, который как бы выступает на некоторое расстояние от зарождающейся клетки. Эти процессы напоминают структуры, ранее описанные в желудочно-кишечных клетках соматостатина (Larsson et al. 19).79). В случае кишечных соматостатиновых клеток ключом к пониманию функционального значения их базальных клеточных отростков является идентификация типов клеток, на которые распространяются эти отростки. Основываясь на первоначальном показе того, что отростки базальных клеток соматостатина вступают в тесный контакт с этими типами клеток-мишеней, было показано, что соматостатин регулирует в желудке секрецию гастрина G-клетками и секрецию кислоты париетальными клетками (Alumets et al. 1979). Таким образом, целью настоящего исследования было задокументировать существование базальных отростков CCK-клеток в кишечнике мышей и идентифицировать типы кишечных клеток, с которыми они контактируют, с конечной целью установления функционального значения этих структур.
Животные
Трансгенные мыши CCK-GFP были получены от MMRRC (Университет Миссури, Колумбия, Миссури, США). Они были выведены собственными силами путем скрещивания с мышами Swiss Webster дикого типа (Чарльз-Ривер, Уилмингтон, Массачусетс, США). Полученные трансгенные мыши были генотипированы с использованием протокола, предоставленного MMRRC. Уход за животными и эксперименты проводились в соответствии с установленными правилами. Животных не кормили в течение ночи со свободным доступом к воде перед забором тонкой кишки.
Иммунофлуоресценция
Мышей анестезировали смесью кетамина и ксилазина и интракардиально перфузировали охлажденным льдом свежедеполимеризованным 3,5% параформальдегидом в фосфатно-солевом буфере (PBS, 10 мМ фосфат натрия, pH 7,4, 0,9% NaCl). Проксимальные 10 см кишечника удаляли, промывали и постфиксировали в 3,5% параформальдегиде в течение 3 ч при 4°С. Ткань промывали в PBS (30 мин при 4°C), последовательно уравновешивали в холодной 10%, 20% и 30% сахарозе в 0,1M фосфате натрия, pH 7,4, и мгновенно замораживали в смеси Tissue-Tek OCT (Sakura Finetek, Торранс, Калифорния, США). Поперечные срезы (толщиной 5 мкм) собирали с помощью криостата, установленного на -19°C и помещены на предметные стекла микроскопа Super Up-Rite (Thermo Scientific, Массачусетс, США).
перечислены используемые антисыворотки, поставщики, условия окрашивания, специфичные для антител, и ссылки на их специфичность. Антисыворотка CCK была получена у кроликов против CCK 19–36, аффинно очищена и охарактеризована в нашей лаборатории. Для идентификации и локализации различных типов клеток, обнаруживаемых в эпителии слизистой оболочки тонкой кишки, использовали антисыворотки, специфичные к типу клеток: (1) хромогранин А для энтероэндокринных клеток, (2) трилистник-фактор-3 для бокаловидных клеток и (3) сахараза-изомальтаза для энтероцитов. Кроме того, антисыворотка, специфичная к рецептору CCK-1, использовалась в попытке локализовать возможные клетки-мишени CCK слизистой оболочки. Как правило, срезы фиксировали либо в 10% формалине в течение 10 мин при 4°C (ХЦК, GFP, фактор трилистника-3 и хромогранин А), либо в 50% смеси ацетона и метанола (сахараза-изомальтаза и GFP). ) в течение 10 мин при -5°С и высушивают на воздухе в течение 30 мин. Затем их промывали PBS или TBST (10 мМ TRIS, pH 7,4, 0,9).% NaCl, 0,1% Тритон Х-100) в течение 5 мин, а неспецифическое окрашивание блокировали инкубацией срезов в 10% ослиной сыворотке в течение 30 мин при комнатной температуре. Сыворотку сливали, а срезы инкубировали с первичными поликлональными антителами либо в течение ночи при 4°С, либо в течение 1 ч при комнатной температуре. После промывки PBS или TBST (3×5 мин каждая) срезы инкубировали со вторичным антителом в течение 1 ч при комнатной температуре. Предметные стекла промывали в PBS или TBST (каждый раз 3×5 мин), контрастно окрашивали 2 мкг/мл 4,6-диамидино-2-фенилиндола (DAPI; Biochemika, Германия) и помещали в реагент против выцветания Prolong Gold (Invitrogen, Carlsbad). , Калифорния, США). Отрицательные контроли включали предметные стекла, обработанные только первичной или вторичной антисывороткой.
Таблица 1
Первичные и вторичные антитела, используемые в иммунофлуоресцентном анализе ( CCK холецистокинин, GFP зеленый флуоресцентный белок, PBS фосфатно-солевой буфер, TBST XTRIS-буфер, содержащий тритон 0,10% солевой раствор)
| Первичное антитело | Вид | Источник | Буфер | Разведение | Вторичное антитело | 6 Разведение0577 | Reference | |
|---|---|---|---|---|---|---|---|---|
| CCK (aa 19–36) | Rabbit | BioSource | PBS/TBST | 1:2000 | Cy3-conjugated donkey anti-rabbit | Jackson ImmunoResearch | 1:1000 | |
| CCK (aa 19–36) | Rabbit | BioSource | PBS/TBST | 1:2000 | Dylight-488-conjugated donkey anti-rabbit | Jackson ImmunoResearch | 1:1000 | |
| CCK-1 Receptor | Rabbit | J. Walsh | TBST | 1:50 | Cy3-conjugated donkey anti-rabbit | Jackson ImmunoResearch | 1:1000 | Sternini и другие. (1999) |
| Chromogranin A | Rabbit | Abcam | TBST | 1:1000 | Cy3-conjugated donkey anti-rabbit | Jackson ImmunoResearch | 1:1000 | Duckworth and Pritchard (2009) |
| GFP | Chick | Abcam | PBS/TBST | 1:1000 | Dylight-488-conjugated donkey anti-chick | Jackson ImmunoResearch | 1:1000 | Томпсон и др. (2009) |
| GFP | Rabbit | V. Bennett | PBS/TBST | 1:1000 | Dylight-488-conjugated donkey anti-rabbit | Jackson ImmunoResearch | 1:1000 | Кижатил и др. (2009) |
| Intestinal Trefoil Factor-3 | Goat | Santa Cruz | PBS | 1:100 | Alexa-568-conjugated donkey anti-goat | Invitrogen Corporation | 1:1000 | Nishida и другие. (2009) |
| Сахароза-Изомальтаза | Коза | Санта-Крус | TBST | 1:500 | Alexa-568-конъюгированный осел | Invitrogen Corporation | 1:1000 | Gracz et al. (2010) |
Открыть в отдельном окне
Получение изображения
Изображения были получены с использованием программного обеспечения Zeiss LSM. Образцы визуализировали на инвертированном конфокальном микроскопе Zeiss с масляными объективами 40×/1,3 (Zeiss Plan NeoFluar) или 63×/1,4 (Zeiss Plan Apochrom). Отдельные оптические срезы или Z-стеки были получены путем последовательного мультитрекинга с возбуждением, установленным на 405 нм (DAPI), 488 нм (эндогенный GFP или Dylight 488) и 561 нм (Cy3 или Alexa 568) и эмиссионными фильтрами BP 420– 480, BP505–550 и LP575, с отверстиями, установленными на 1 воздушную единицу для каждого канала, и усреднением строк 8 или 16 при разрешении 1024 или 2048 пикселей. Также были получены конфокальные дифференциально-интерференционные контрастные изображения в проходящем свете.
Трехмерная визуализация
Срезы кишечной ткани (15 мкм) окрашивали на GFP и CCK, как описано выше. Иммуноокрашенные срезы исследовали с помощью конфокального микроскопа Zeiss LSM 510 с масляным объективом Plan-Apochrom 63×/1,4 и 1,2-кратным оптическим увеличением. Трехмерную многоканальную визуализацию и экспорт в виде фильмов Quicktime выполняли с использованием программного обеспечения Volocity Visualization (PerkinElmer, Уолтем, Массачусетс, США). Цифровой контраст и прозрачность всех отдельных каналов были скорректированы для оптимизации структуры всего объема отображаемой ткани. Были применены медианные фильтры (3×3) для уменьшения шума в некоторых каналах. Каналы визуализировались либо как проекции максимальной интенсивности (зеленый/GFP, красный/CCK), что позволяло видеть цвет через другие каналы, либо как флуоресценция (синий/DAPI).
Эндогенно экспрессируемый GFP, управляемый промотором CCK, заполнил цитоплазму энтероэндокринных CCK-клеток (I-клетки) в тонком кишечнике мыши (). Почти все клетки, идентифицированные иммуноокрашиванием антисывороткой к CCK, также экспрессировали GFP, указывая на то, что экспрессия GFP у трансгенных мышей была «чистой»; лишь изредка клетки GFP не окрашивались положительно на CCK (10). Интенсивность эндогенной зеленой флуоресценции, наблюдаемой в клетках CCK, была вариабельной и часто слабой. Поэтому, чтобы облегчить двойное иммунофлуоресцентное окрашивание в этих клетках, мы проверили способность антисыворотки GFP окрашивать эндогенные GFP-экспрессирующие клетки CCK в тонком кишечнике мыши. Антисыворотка против GFP, полученная как из кроличьих, так и из куриных окрашенных клеток CCK, экспрессирующих GFP (). В последующих исследованиях для окрашивания клеток CCK использовали как кроличью, так и куриную сыворотку против GFP.
Открыть в отдельном окне
Колокализация CCK (холецистокинин) и GFP (зеленый флуоресцентный белок) в клетках CCK кишечника мыши. a Эндогенно флуоресцентный GFP ( зеленый ) в клетке CCK. b Та же клетка CCK, что и в a , окрашенная кроличьей антисывороткой к GFP ( красный ), демонстрирующая, что только клетки, эндогенно экспрессирующие GFP, окрашиваются положительно на GFP в двенадцатиперстной кишке мыши. c Объединенные изображения a , б. d Эндогенно флуоресцентный GFP в клетке CCK. e Та же клетка, что и в d , окрашенная кроличьим антителом к CCK, демонстрирующая, что только клетки CCK экспрессируют GFP в двенадцатиперстной кишке мыши. f Объединенные изображения d , e. g Клетка CCK, в которой эндогенный GFP был визуализирован с помощью иммунного окрашивания сывороткой анти-GFP, выращенной у цыплят. h Та же самая клетка CCK, что и на г , окрашенная кроличьим антителом к CCK, демонстрирующая, что GFP и CCK можно окрашивать с помощью иммуноокрашивания с использованием первичных антисывороток, полученных от другого вида. i Объединенные изображения г , ч . Ориентация клеток CCK относительно просвета кишечника и основания клеток показана на и и одинакова на всех других панелях ( синий ядерное окрашивание DAPI). Бар 5 мкм
Открыть в отдельном окне
a Две желтые клетки CCK при низком увеличении, демонстрирующие как эндогенную экспрессию GFP ( зеленый ), так и положительное иммуноокрашивание на GFP кроличьей анти-GFP сывороткой ( красный ). b Два желтых клеток CCK при малом увеличении, демонстрирующих как положительное иммуноокрашивание на CCK с кроличьей сывороткой против CCK ( красный ), так и на GFP с куриной анти-GFP сывороткой ( зеленый ). Обратите внимание на отростки базальных клеток CCK ( стрелки ). Обе микрофотографии были сделаны с помощью флуоресцентной ( синий ядерное окрашивание DAPI) и дифференциально-интерференционной контрастной (DIC) оптики ( л люмен). Бар 20 мкм
клетки CCK имели колбовидную или веретенообразную форму и были более многочисленны в кишечных криптах, чем в ворсинках. Мы наблюдали псевдоподообразные базальные клеточные отростки во многих клетках CCK (4). Чтобы оценить частоту базальных клеточных процессов среди клеток CCK, мы подсчитали клетки, которые были как CCK-, так и GFP-иммунопозитивными, в шести поперечных срезах двенадцатиперстной кишки. В криптах Либеркюна в 47 из 106 таких клеток обнаруживались базальноклеточные отростки (47%). В ворсинках 73 из 112 клеток CCK имели базальноклеточные отростки (65%). Большинство базальноклеточных отростков были короткими; самый длинный визуализированный отросток (около 15 мкм) проходил через три соседние клетки. Обильное иммуноокрашивание CCK присутствовало в отростках базальных клеток, что позволяет предположить, что они могут высвобождать CCK на близлежащие клетки-мишени. В криптах базальноклеточные отростки обычно распространяются на соседнюю клетку. В ворсинках отростки иногда были изогнуты и переплетались между соседними клетками. Часто одно расширение раздваивалось на два (), и множественные отростки наблюдались в небольшом проценте клеток (25 из 75 клеток CCK с базальными отростками). Трехмерные реконструкции более толстых срезов (15 мкм) позволили предположить, что эти отростки распространяются во всех направлениях и что они, вероятно, не использовались при миграции клеток CCK из крипт вверх по продольной оси ворсинок (14). Эти реконструкции также показали, что процент клеток CCK, у которых наблюдается базолатеральный отросток, мог быть искусственно занижен, поскольку отростки могут распространяться в Z-плоскости и, таким образом, быть невидимыми при наблюдении XY на срезе толщиной 5 мкм. Эта возможность была сведена к минимуму при анализе путем тщательного фокусирования вверх и вниз по каждой CCK-иммунореактивной клетке для оценки клеточных процессов, распространяющихся во всех направлениях.
Открыть в отдельном окне
a – f Примеры клеток CCK-GFP, окрашенных куриной или кроличьей антисывороткой GFP, распространяющих базальные клеточные отростки ( стрелки ) на соседние клетки. г – i , j – l Два примера колокализации ( желтый ) ССК ( красный ) с куриным GFP ( зеленый ) в отростках клеток ССК. CCK присутствует в ложноногах ( стрелки ) вместе с GFP ( синий ядерное окрашивание DAPI). Бар 5 мкм
Открыть в отдельном окне
Трехмерная реконструкция клетки ССК, расположенной в основании ворсинки ( a ). Та же клетка (показана на всех панелях) обладает раздвоенной псевдоножкой, которая простирается в пространства, отличные от непосредственно вверх или вниз по оси крипта-ворсинка ( b – i ). GFP иммуноокрашивается куриным GFP-антителом ( зеленый ), а CCK — кроличьим CCK-антителом ( красный ). Оба флуорофора представлены в виде максимальных проекций (окрашивание ядер синим DAPI). по оси X , по оси Y и по оси Z ( зеленый , красный и синий соответственно) показаны ( внизу слева ), чтобы продемонстрировать, что каждое изображение ( 9008 b – i ) поворачивается на 45° вдоль оси Y . Ось крипты-ворсинки находится прямо напротив оси X . Прутки 10 мкм ( a ), 5 мкм ( i )
Для идентификации типа или типов клеток, на которые нацелены процессы, срезы окрашивали антителами к хромогранину А (маркер нейроэндокринных клеток), фактору трилистника-3 (бокал клеточный маркер) и сахараза-изомальтаза (маркер клеток энтероцитов). Поскольку мы никогда не наблюдали ложноножки длиной более ~ 15 мкм, мы классифицировали клетки как находящиеся в пределах 1–4 клеток или > 4 клеток от клеток CCK-GFP, чтобы оценить вероятность того, что клетки CCK передают сигналы определенному типу клеток. . Как резюмировано в , мы обнаружили, что клетки CCK-GFP иногда присутствовали рядом с другими клетками CCK, но редко рядом с другими нейроэндокринными клетками. Бокаловидные клетки чаще обнаруживались в пределах 4 клеток GFP-позитивных клеток, но это, вероятно, отражало относительное количество бокаловидных клеток по сравнению с нейроэндокринными клетками в слизистой оболочке тонкой кишки (4). Мы никогда не наблюдали, чтобы ложноножки действительно заканчивались рядом с бокаловидной клеткой (10). Напротив, клетки CCK-GFP и их базальные клеточные отростки всегда располагаются рядом с энтероцитами (4).
Открыть в отдельном окне
Двойное иммунофлуоресцентное окрашивание срезов антителами против кишечного трилистника-фактора-3 ( красный ) для идентификации бокаловидных клеток и либо CCK, либо антителами GFP кролика ( зеленый ). Базальные отростки клеток CCK не контактируют с бокаловидными клетками, положительными по фактору кишечного трилистника-3 ( L люмен, синий окрашивание ядер DAPI). Бар 10 мкм
Открыть в отдельном окне
Серия изображений Z-стека ( a – j : расстояние 0,49 мкм) части двенадцатиперстной кишки мыши, дважды окрашенной на GFP ( зеленый ) в клетках CCK и факторе трилистника кишечника-3 ( красный ) в бокаловидных клетках, чтобы продемонстрировать, что отростки базальных клеток CCK не направлены на бокаловидные клетки ( наконечников стрелок бокаловидных клеток, положительных на кишечный трилистник-фактор-3, стрелка базальный отросток CCK клеток, L просвет). Микрофотографии были сделаны с использованием как флуоресцентной, так и ДИК-оптики ( синий окрашивание ядер DAPI). Bar 20 мкм
Открыть в отдельном окне
Двойное иммунофлуоресцентное окрашивание срезов антителами к сахаразе-изомальтазе ( красный ) для идентификации энтероцитов и GFP кролика ( зеленый ) для идентификации клеток ССК. Видно, что клетки CCK и их базальные клеточные отростки тесно связаны с соседними энтероцитами. В зависимости от плоскости среза либо отсутствие окрашивания сахаразой-изомальтазой ( наконечников стрелок ) или слабое окрашивание при наличии бокаловидных клеток ( стрелка ) ( L люмен, синий ядерное окрашивание DAPI). Бар 10 мкм
Таблица 2
Близость CCK-GFP-позитивных клеток к другим типам клеток в слизистой оболочке ворсинок кишечника. Срезы кишечника окрашивали антисыворотками, направленными против хромогранина А, кишечного фактора трилистника-3 или сахаразы-изомальтазы. Близость нейроэндокринных клеток, бокаловидных клеток или энтероцитов, соответственно, к клеткам CCK была установлена путем определения того, находились ли они в пределах 1-4 клеток или >4 клеток от GFP-положительных клеток (9).0005 GFP+ ) клетки видны в том же срезе. Подсчитывали не менее 100 клеток каждого типа. Результаты выражены в процентах от общего числа подсчитанных клеток
| Антиген | Близость к клеткам GFP+ | ||
|---|---|---|---|
| Within 1–4 cells (%) | >4 cells away (%) | Total (%) | |
| Non-GFP chromogranin-positive | 0 | 100 | 100 |
| Кишечный фактор трилистника-3 | 10 | 90 | 100 |
| Sucrase-isomaltase | 100 | 0 | 100 |
| GFP (CCK cells) | 4 | 96 | 100 |
Open в отдельном окне
Было показано, что соматостатиновые клетки кишечника распространяют базальные цитоплазматические отростки на соседние типы клеток и высвобождают соматостатин из этих псевдоподообразных структур (Larsson et al. 19).79). Считается, что эти клетки высвобождают соматостатин путем экзоцитоза из базолатеральных отростков с последующей диффузией на небольшие расстояния (паракринное действие) для взаимодействия с рецепторами соматостатина на клетках-мишенях. Используя этот анатомический путь, эффективные концентрации соматостатина могут быть доставлены относительно быстро к определенным клеткам-мишеням. Хорошо известно, что в желудке этот паракринный путь соматостатина регулирует секрецию гастрина и соляной кислоты (Alumets et al. 19).79). Базальные цитоплазматические клеточные отростки этих паракринных соматостатиновых клеток заканчиваются луковицеобразными вздутиями (Alumets et al., 1979). Подобные базальные цитоплазматические клеточные процессы наблюдались в кишечных пептидных клетках YY (PYY) (Karaki et al. 2006; Lundberg et al. 1982). Более того, длинные тонкие базальные отростки, описанные на клетках подвздошной кишки крысы, окрашиваются антисывороткой против гормона желудка, гастрина (Larsson and Rehfeld, 1978). Функциональное значение этих структур на клетках PYY и гастрина неизвестно.
Считается, что некоторые действия кишечных CCK опосредованы афферентными нейронами блуждающего нерва (Dockray 2009). Таким образом, базальные цитоплазматические отростки CCK-клеток, наблюдаемые в настоящем исследовании, могут быть направлены на близлежащие окончания афферентных нейронов блуждающего нерва под базальной пластинкой эпителия слизистой оболочки кишечника. Тем не менее, несколько линий доказательств указывают на отсутствие тесного анатомического контакта между клетками CCK в эпителии слизистой оболочки тонкой кишки и кишечных нервов. Немногие клетки CCK крысы находятся в тесном анатомическом контакте (<5 мкм) с афферентными аксонами блуждающего нерва, и последние не производят терминальных специализаций вблизи клеток CCK (Berthoud and Patterson 19).96). Иммуноцитохимическая локализация рецепторов CCK-A (теперь называемых рецепторами CCK-1) в тонком кишечнике крысы выявила обильную экспрессию в телах и волокнах нейронов, но не в нервных окончаниях, иннервирующих слизистую оболочку, в которой расположены CCK-клетки (Sternini et al. 1999). Мы повторили исследование Sternini et al. на крысах. (1999) на нашей мышиной модели и, используя ту же антисыворотку к рецептору CCK, смогли подтвердить отсутствие нейронов, экспрессирующих CCK-рецептор, в слизистой оболочке двенадцатиперстной кишки (данные не показаны). Авторадиографическая локализация связывания насыщаемого радиолиганда CCK не выявила рецепторов CCK в слизистой оболочке тонкого кишечника собак (Mantyh et al. 19).94). В соответствии с этими выводами мы не обнаружили окрашивания рецептора CCK-1 в клетках слизистой оболочки кишечника (данные не показаны). Эти наблюдения предполагают, что клетки CCK не оказывают прямого нейрокринного действия на кишечные нервы, но не исключают паракринного действия.
Поскольку большинство клеток CCK, по-видимому, направляют свои базальные клеточные отростки к соседним или близлежащим энтероцитам, мы можем логически предположить, высвобождается ли CCK из этих отростков для воздействия на энтероциты. Основные функции энтероцитов — всасывание и секреция жидкости, транспорт ионов, витаминов и продуктов переваривания питательных веществ. Наше изучение литературы не выявило каких-либо опубликованных описаний эффектов CCK на эти функции энтероцитов. Однако анализ тканей взрослых мышей с помощью обратной транскрипции с помощью полимеразной цепной реакции выявил экспрессию рецептора CCK1 (также известного как рецептор CCK-A) в двенадцатиперстной кишке, тонкой кишке (предположительно тощей или подвздошной кишке) и толстой кишке. Лакурс и др. 1997). Этот анализ не идентифицировал тип кишечных клеток, экспрессирующих рецептор CCK1, поэтому этот молекулярный анализ может просто отражать хорошо известную экспрессию рецепторов CCK1 блуждающими афферентными нервами, присутствующими в стенках тонкой и толстой кишки (Sternini et al., 1999). ).
Возможна иная интерпретация функционального значения базальных отростков ХЦК в тонкой кишке. Было показано, что липидная перфузия просвета тонкой кишки крысы стимулирует повышенную секрецию ХЦК, и этот эффект блокируется предварительной обработкой препаратом Pluronic L-81, который ингибирует образование хиломикронов (Raybould et al. 19).98). Т.о., базальные клеточные процессы CCK, описанные здесь, могут функционировать для получения информации от энтероцитов, а не для передачи сигналов энтероцитам. Эта концепция также подтверждается отсутствием луковичных вздутий на концах отростков базальных клеток CCK, в отличие от наблюдаемых на концах отростков базальных соматостатиновых клеток, которые, как известно, участвуют в паракринной передаче сигналов (Alumets et al. 1979). . Интересно, что другая эндокринная клетка тонкого кишечника, которая стимулируется поглощенными липидами, клетка PYY, также демонстрирует базально-клеточные отростки (Karaki et al. 2006; Lundberg et al. 19).82). Будущие исследования должны различать эти различные возможности.
Рис. 4, ролик
Щелкните здесь для просмотра. (12M, mov)
Работа выполнена при поддержке гранта NIH DK38626.
Авторы благодарят Сэма Джонсона за его помощь в получении изображений и признательны центру световой микроскопии Университета Дьюка.
Дополнительные материалы в электронной форме Электронная версия этой статьи (doi:10.1007/s00441-010-0997-1) содержит дополнительные материалы, доступные авторизованным пользователям.
- Alumets J, Ekelund M, El Munshid HA, Hakanson R, Loren I, Sundler F. Топография клеток соматостатина в желудке крысы: возможное функциональное значение. Сотовые Ткани Res. 1979; 202: 177–188. [PubMed] [Google Scholar]
- Berthoud HR, Patterson LM. Анатомические взаимоотношения между афферентными волокнами блуждающего нерва и ХЦК-иммунореактивными энтероэндокринными клетками в слизистой оболочке тонкого кишечника крыс. Acta Anat (Базель) 1996; 156: 123–131. [PubMed] [Google Scholar]
- Buchan AM, Polak JM, Solcia E, Capella C, Hudson D, Pearse AG. Электронно-иммуногистохимические доказательства того, что I-клетка кишечника человека является источником CCK. Кишка. 1978;19:403–407. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Dockray GJ. Универсальность блуждающего нерва. Физиол Поведение. 2009; 97: 531–536. [PubMed] [Google Scholar]
- Duckworth CA, Pritchard DM. Подавление апоптоза, гиперплазия крипт и измененная дифференцировка в эпителии толстой кишки мышей с нулевыми генами. Гастроэнтерология. 2009; 136: 943–952. [PubMed] [Google Scholar]
- Gracz AD, Ramalingam S, Magness ST. Экспрессия Sox9 маркирует субпопуляцию CD24-экспрессирующих эпителиальных стволовых клеток тонкой кишки, которые образуют органоиды in vitro. Am J Gastrointest Liver Physiol. 2010;298:G590–G600. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Karaki S, Mitsui R, Hayashi H, Kato I, Sugiya H, Iwanaga T, Furness JB, Kuwahara A. Рецептор жирных кислот с короткой цепью, GPR43, экспрессируется Энтероэндокринные клетки и тучные клетки слизистой оболочки кишечника крыс. Сотовые Ткани Res. 2006; 324:353–360. [PubMed] [Google Scholar]
- Кижатил К., Бейкер С.А., Аршавский В.Ю., Беннетт В. Анкирин-Г способствует транспорту циклических нуклеотид-управляемых каналов к сенсорным клеткам фоторецепторов палочек. Наука. 2009 г.;323:1614–1617. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lacourse KA, Lay JM, Swanberg LJ, Jenkins C, Samuelson LC. Молекулярная структура гена рецептора CCK-A мыши. Biochem Biophys Res Commun. 1997; 236:630–635. [PubMed] [Google Scholar]
- Ларссон Л.И., Рефельд Дж.Ф. Распределение гастрина и клеток CCK в желудочно-кишечном тракте крыс. Доказательства существования трех различных типов клеток, сохраняющих СООН-концевую иммунореактивность гастрина. Гистохимия. 1978; 58: 23–31. [PubMed] [Академия Google]
- Ларссон Л.И., Гольтерманн Н., де Магистрис Л., Рефельд Дж.Ф., Шварц Т.В. Клеточные процессы соматостатина как пути паракринной секреции. Наука. 1979; 205: 1393–1395. [PubMed] [Google Scholar]
- Liddle RA. Холецистокинин. В: Walsh JH, Dockray GJ, редакторы. Пептиды кишечника: биохимия и физиология. ворон; Нью-Йорк: 1994. С. 175–216. [Google Scholar]
- Лиддл Р.А. Холецистокининовые клетки. Annu Rev Physiol. 1997; 59: 221–242. [PubMed] [Google Scholar]
- Лундберг Дж. М., Татемото К., Терениус Л., Хеллстром П. М., Матт В., Хёкфельт Т., Хамбергер Б. Локализация пептида YY (PYY) в эндокринных клетках желудочно-кишечного тракта и влияние на кровоток и перистальтику кишечника. Proc Natl Acad Sci USA. 1982;79:4471–4475. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Mantyh CR, Pappas TN, Vigna SR. Локализация рецепторов холецистокинина А и холецистокинина В/гастрина в верхних отделах желудочно-кишечного тракта собак. Гастроэнтерология. 1994; 107:1019–1030. [PubMed] [Google Scholar]
- Нисида К., Камизато М., Каваи Т., Масуда К., Такео К., Тешима-Кондо С., Танахаши Т., Рокутан К. Интерлейкин-18 является решающим фактором, определяющим уязвимость прямой кишки мыши к психосоциальным стресс. ФАСЭБ Дж. 2009 г.;23:1797–1805. [PubMed] [Google Scholar]
- Raybould HE, Meyer JH, Tabrizi Y, Liddle RA, Tso P. Ингибирование опорожнения желудка в ответ на кишечные липиды зависит от образования хиломикронов. Am J Physiol. 1998; 274: R1834–R1838. [PubMed] [Google Scholar]
- Сэмюэл Б.С., Шайто А., Мотоике Т., Рей Ф.Е., Бакхед Ф., Манчестер Дж.К., Хаммер Р.Е., Уильямс С.К., Кроули Дж., Янагисава М., Гордон Дж.И. Влияние кишечной микробиоты на ожирение хозяина модулируется короткоцепочечным рецептором, связанным с G-белком, Gpr41, связывающим жирные кислоты. Proc Natl Acad Sci USA. 2008; 105:16767–16772. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Стернини С., Вонг Х., Фам Т., Де Джорджио Р., Миллер Л.Дж., Кунц С.М., Рив Дж.Р., Уолш Дж.Х., Рэйбоулд Х.Е. Экспрессия рецепторов холецистокинина А в нейронах, иннервирующих желудок и кишечник крыс. Гастроэнтерология. 1999; 117:1136–1146. [PubMed] [Google Scholar]
- Томпсон Л.Х., Грилиш С., Кирк Д., Бьорклунд А. Реконструкция нигростриарного дофаминового пути в мозге взрослых мышей. Евр Джей Нейроски. 2009; 30: 625–638. [PubMed] [Академия Google]
Хемотаксис: выводы из расширяющейся псевдоподии | Journal of Cell Science
Skip Nav Destination
КОММЕНТАРИЙ|
15 сентября 2010 г.
Питер Дж. М. Ван Хаастерт
Информация об авторе и статье
Номер в сети: 1477-9137
Номер для печати: 0021-9533
© 2010.
2010
J Cell Sci (2010) 123 (18): 3031–3037.
https://doi.org/10.1242/jcs.071118
- Разделенный экран
- Просмотры
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- Делиться
- MailTo
- Твиттер
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
Цитирование
Питер Дж. М. Ван Хаастерт; Хемотаксис: выводы из расширяющейся псевдоподии. J Cell Sci 15 сентября 2010 г.; 123 (18): 3031–3037. doi: https://doi.org/10.1242/jcs.071118
Скачать файл цитаты:
- Ris (Zotero)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- РефВоркс
- Бибтекс
Расширенный поиск
Хемотаксис — один из самых увлекательных процессов в клеточной биологии. Неглубокие градиенты хемоаттрактанта направляют движение клеток, а сложная сеть сигнальных путей каким-то образом инструктирует двигательный аппарат индуцировать ложноножки в направлении этих градиентов. Захватывающие новые эксперименты подошли к хемотаксису с точки зрения расширяющейся псевдоподии. Эти недавние исследования показали, что в отсутствие внешних сигналов клетки используют эндогенные сигналы для высокоупорядоченного расширения ложноножек, которые проявляются главным образом в виде чередующихся правых и левых расщеплений. Кроме того, хемоаттрактанты активируют другие сигнальные молекулы, которые индуцируют позиционное смещение этой базальной системы, так что расширяющиеся псевдоподы ориентируются в сторону градиента. В этом комментарии я рассматриваю результаты этих недавних экспериментов, которые в совокупности дают новый взгляд на движение клеток и хемотаксис.
Ключевые слова:
Диктиостелиум, Актин, Движение клеток, хемотаксис, Pseudopod
Многие клетки имеют способ миграции, известный как амебоидное движение, которое характеризуется частыми изменениями формы клеток в результате удлинения выступов (Friedl and Wolf, 2009; Lammermann and Sixt, 2009). Выпячивания амебоидной клетки часто называют псевдоподами или ламеллиподами; эти выступы могут принимать различную форму, которые называются тонкими филоподиями или выпуклыми лобоподиями. Псевдоподы имеют решающее значение для движения клеток, потому что они определяют скорость, направление и траекторию движения клетки. Соседние клетки могут координировать расширения псевдоподий, тем самым способствуя коллективной миграции клеток в дополнение к другим процессам, таким как контактное управление (Weijer, 2009).). Важным аспектом клеточной подвижности является способность клеток реагировать на сигналы направления ориентированным движением. Градиенты диффузных химических веществ вызывают хемотаксис (Hoeller and Kay, 2007; Weiner, 2002). Другими сигналами направления, которые могут вызвать направленное движение, являются температурные градиенты (термотаксис) или электрические поля (электротаксис) (Bahat and Eisenbach, 2006; Zhao, 2009), которые здесь не рассматриваются. Эти сигналы каким-то образом модулируют направление псевдоподий, так что в среднем клетки двигаются в направлении позиционных сигналов.
Как амебоидные клетки перемещаются и ориентируются, используя химические градиенты? Многие эксперименты решают этот вопрос, используя стратегию, которая включает воздействие на клетки градиента хемоаттрактанта с последующим измерением пространственно-временной активации сигнальных молекул, которые в конечном итоге делают возможным ориентированное движение. Этот «сигнально-центрированный» подход к пониманию клеточного движения является очень мощным средством, с помощью которого могут быть идентифицированы задействованные сигнальные пути, а также фундаментальные механизмы, лежащие в основе определения градиента, нарушения симметрии (которые могут быть вовлечены в определение клеточной полярности или формирование передний край) и усиление сигнала (которое создает внутриклеточный градиент сигнальных молекул, который намного круче, чем градиент внеклеточного хемоаттрактанта) (Franca-Koh et al., 2006; Insall, 2010; King and Insall, 2009).; Мерло и Фиртель, 2003 г.; Шнайдер и Хо, 2006). Многие из этих экспериментов были проведены с Dictyostelium . Этот генетически податливый организм движется сходным образом со многими другими амебоидными клетками и проявляет хемотаксис в ответ на очень неглубокие градиенты внеклеточного хемоаттрактанта циклического АМФ (цАМФ) (Kay et al., 2008). Неглубокий градиент цАМФ индуцирует активацию рецепторов цАМФ и связанных с ними G-белков способом, который приблизительно пропорционален крутизне градиента и лишь немного сильнее на переднем крае, чем на заднем крае клетки. Напротив, небольшая GTPase Ras и многие нижележащие компоненты активируются гораздо сильнее на переднем крае, чем на заднем крае клетки (Jin et al., 2000; Zhang et al., 2008). По крайней мере четыре сигнальных пути способствуют хемотаксису Клетки Dictyostelium : путь фосфоинозитид-3-киназы (PI3K), который продуцирует фосфатидилинозитол (3,4,5)-трифосфат [PtdIns(3,4,5) P 3 ], который, в свою очередь, активирует Akt ( также известный как путь PKB; путь TorC2, который активирует PKBR1, растворимую гуанилатциклазу (рГЦ), которая активируется на переднем крае и продуцирует цГМФ; и путь с участием PLA2, механизм которого неизвестен (Kamimura et al., 2008; Veltman et al., 2008). Из-за множества сигнальных путей и их сложной регуляции петлями положительной и отрицательной обратной связи трудно установить связь между сигнальными путями и локомоционным аппаратом в Dictyostelium и других организмов, а также определить, как клетки на самом деле вытягивают ложноножки в направлении градиента.
Недавно исследователи применили дополнительный «псевдоподоцентрированный» подход, чтобы понять движение клеток. При таком подходе исследователи подробно наблюдают за тем, как клетки расширяют ложноножки, а затем пытаются интегрировать эти наблюдения с тем, что известно об установленных сигнальных путях (Andrew and Insall, 2007; Arrieumerlou and Meyer, 2005; Bosgraaf and Van Haastert, 2009).а; Инсал, 2010 г.; Ли и др., 2008 г.; Маэда и др., 2008 г.; Такаги и др., 2008). Было высказано предположение, что ложноножки являются самоорганизующимися структурами, что означает, что их организация в значительной степени контролируется изнутри (Karsenti, 2008). Хотя внешние сигналы могут запускать формирование и локализацию псевдоподии, в остальном псевдоподия следует типичному жизненному циклу (см. вставку 1). Недавнее внимание к ложноногам последовало за новаторской работой, проведенной в 1980-х годах, которая включала компьютерный анализ движения клеток, растяжения ложноножек и хемотаксиса (Potel and Mackay, 19). 79; Варнум-Финни и др., 1987; Варнум и Солл, 1984). Современная визуализация живых клеток и усовершенствованные компьютерные алгоритмы теперь позволяют проводить более точный анализ клеток и псевдоножек. В исследованиях, которые берут
Вставка 1. Три основные характеристики псевдоподий
Псевдоподы являются самоорганизующимися структурами (Эндрю и Инсалл, 2007; Босграаф и Ван Хаастерт, 2009a; Карсенти, 2008) Внешние сигналы могут инициировать формирование псевдоподии, но затем псевдоподия претерпевает ряд изменений формы независимо от внешних сигналов. Это означает, что хемоаттрактанты могут модулировать время и положение на клеточной поверхности, где будет формироваться ложноножка, но в остальном они будут иметь ограниченное влияние на другие свойства, такие как период роста или длина ложноножки.
Псевдоподии вытянуты перпендикулярно поверхности клетки (Mogilner and Oster, 1996; Van Haastert and Bosgraaf, 2009a) это положение клетки. Это означает, что во время навигации в хемотаксических градиентах псевдоподии не изгибаются в направлении градиента. Для направленного движения псевдоподии должны формироваться на стороне клетки, ближайшей к градиенту. Клетка очень неправильной формы не может должным образом хемотаксировать.
Псевдоподы образуются de novo или путем расщепления текущей псевдоподии (Andrew and Insall, 2007; Bosgraaf and van Haastert, 2009b) де ново). Расщепляющиеся ложноножки предпочтительно вытягиваются, чередуясь вправо или влево под небольшими углами, заставляя клетку двигаться по устойчивой зигзагообразной траектории. Псевдоподии de novo вытянуты в случайном направлении. Это означает, что отношение расщепляющихся псевдоподий к количеству псевдоподий de novo определяет постоянство движения клеток. С точки зрения хемотаксиса постоянство клеточного движения функционирует как память и интегратор направленной информации. Мутантные клетки с дефектами расщепления псевдоподий обладают очень плохой персистенцией, неэффективно перемещаются в окружающей среде и имеют нарушенный хемотаксис.
Подход, ориентированный на ложноножки, собираются большие наборы данных о пространственно-временных свойствах ложноножек, которые расширяются за счет клеток при отсутствии или наличии сигналов направления. Кроме того, исследования клеток, несущих мутации в генах, которые кодируют различные белки, участвующие в клеточном движении и хемотаксисе, начинают раскрывать механизмы, с помощью которых специфические сигнальные пути индуцируют образование псевдоподов в направлении градиента хемоаттрактанта. В этом комментарии я резюмирую эти недавние эксперименты и обсуждаю последствия этих коллективных открытий для клеточной биологии удлинения псевдоподий, движения клеток и хемотаксиса.
Традиционно движение клетки измеряют, отслеживая положение центроида клетки во времени, которое определяется как геометрический центр масс или периметр двумерного изображения клетки. Простые компьютерные алгоритмы могут обнаружить этот центроид и разбить траекторию на дискретные шаги и повороты (Li et al. , 2008; Soll et al., 2003). Однако, несмотря на то, что отслеживание центроидов позволило получить важные сведения о движении амеб (Li et al., 2008; Shenderov and Sheetz, 1997), он не может быть использован для количественного анализа ложноножек, поскольку лишь небольшая часть ложноножек клетки вызывает изменение направления ее движения (Bosgraaf, van Haastert, 2009b). Поэтому используются другие алгоритмы для идентификации контура клетки и определения псевдоподий на основе расширяющейся выпуклой области (Machacek and Danuser, 2006; Soll et al., 2003). Простое и привлекательное описание ложноножки — это вектор, соединяющий две точки на поверхности клетки: место начала и окончания роста выпячивания соответственно (Bosgraaf and Van Haastert, 2009).в). Измерения векторов, представляющих тысячи ложноножек, можно использовать для статистического анализа свойств ложноножек, включая частоту, размер, время роста и направление движения к хемоаттрактантам. Кроме того, анализ этих векторов с помощью алгоритмов автокорреляции может предоставить информацию о долговременных взаимодействиях псевдоподий (см. ниже), понимание которых сыграло важную роль в раскрытии стохастического порядка формирования псевдоподий в отсутствие внешних сигналов (Босграаф и ван Хаастерт). , 2009 г.б).
Наблюдая за положением на клеточной поверхности, где вытягиваются ложноножки, Эндрю и Инсолл обнаружили, что ложноножки часто образуются путем расщепления существующих ложноножек. У Dictyostelium и других организмов наблюдались две формы расщепления (Andrew, Insall, 2007; Bosgraaf, van Haastert, 2009b). При «Y-расщеплении» выпячивание без органелл разделяется на две псевдоподии, обе из которых увеличиваются в размерах, а затем обычно одна псевдоподия втягивается, а другая сохраняется. При «одностороннем» разделении новый псевдопод формируется сбоку от текущего псевдопода; новая псевдоподия некоторое время растет, останавливается, а затем сбоку от нее образуется новая псевдоподия. В поляризованных клетках односторонние расщепления преобладают над Y-расщеплениями, вероятно, потому, что образование односторонних расщеплений лучше сохраняет клеточную полярность. Помимо образования новой псевдоподии путем расщепления существующей псевдоподии, иногда псевдоподия образуется de novo, т. е. на части клеточной поверхности, которая какое-то время не участвовала в растяжении псевдоподии. Как будет показано ниже, расщепление псевдоподий формирует основу для персистентного движения и навигации клеток. На рис. 1 показана 9Клетка 0005 Dictyostelium , образующая псевдоногу de novo, за которой следуют три односторонних расщепления.
Удлиняющееся выпячивание создает противодействующие силы в плазматической мембране, и энергетически наиболее благоприятное направление псевдоподии перпендикулярно мембране (Mogilner and Oster, 1996) (рис. 2). Эта гипотеза была проверена на ложноногах, расширенных клетками Dictyostelium в буфере и клетками, перемещающимися в градиенте хемоаттрактанта. Действительно, морфометрический анализ показал, что клетки всегда вытягивают ложноножки перпендикулярно клеточной поверхности с относительно небольшим стандартным отклонением (Van Haastert and Bosgraaf, 2009). а). Градиент хемоаттрактанта не вызывает искривления этой ложноножки (статистически менее чем на 2 градуса). Чтобы двигаться в направлении градиента, клетка должна сформировать множество ложноножек на той стороне клетки, которая обращена к градиенту. В дополнение к этому позиционному смещению, эффективный хемотаксис требует, чтобы клетка имела гладкую эллипсоидную форму (Fig. 2). Это было продемонстрировано при изучении мутанта Dictyostelium , имеющего неправильную форму за счет делеции формина; хотя ложноножки возникли в передней части клетки, было обнаружено, что они простираются во многих разных направлениях, что приводит к плохому хемотаксису (Van Haastert and Bosgraaf, 2009).а). Представление о том, что псевдоподии всегда перпендикулярны кривизне клетки, подразумевает, что направление, которому следует псевдоподия, определяется положением на клеточной поверхности, где формируется псевдоподия, в сочетании с кривизной в этом положении.
Рис. 1.
Посмотреть в большом размереСкачать слайд
Клетка Dictyostelium движется в буфере. ( A ) Псевдоподы обозначены желтыми стрелками непосредственно перед формированием ложноножки, и они образуются либо de novo (0 секунд; указано пустой стрелкой), либо образуются в результате расщепления текущей ложноножки (18, 42, 66 секунд; указано) заполненными стрелками). ( B ) 15-минутный трек камеры. Темно-серая область указывает на ячейку в начале, тогда как светло-серая область представляет собой поверхность, покрытую ячейкой во время движения. Трек иллюстрирует результат статистического анализа нескольких сотен ложноножек: расщепление происходит преимущественно попеременно вправо-влево под небольшим углом, что заставляет клетку продолжать двигаться в том же направлении. И наоборот, псевдоподия de novo может расширяться в любом направлении, вызывая тем самым случайный поворот.
Рис. 1.
Увеличить Загрузить слайд
Клетка Dictyostelium движется в буфере. ( A ) Псевдоподы обозначены желтыми стрелками непосредственно перед формированием ложноножки, и они образуются либо de novo (0 секунд; указано пустой стрелкой), либо образуются в результате расщепления текущей ложноножки (18, 42, 66 секунд; указано) заполненными стрелками). ( B ) 15-минутный трек камеры. Темно-серая область указывает на ячейку в начале, тогда как светло-серая область представляет собой поверхность, покрытую ячейкой во время движения. Трек иллюстрирует результат статистического анализа нескольких сотен ложноножек: расщепление происходит преимущественно попеременно вправо-влево под небольшим углом, что заставляет клетку продолжать двигаться в том же направлении. И наоборот, псевдоподия de novo может расширяться в любом направлении, вызывая тем самым случайный поворот.
Работа, проведенная в прошлом столетии, показала, что амебоидные клетки демонстрируют так называемое коррелированное случайное блуждание: направление текущего движения клетки коррелирует с направлением ее движения в прошлом, и поэтому клетки двигаются с упорством (Гейл и Boone, 1970; Patlak, 1953; Potel and Mackay, 1979). Продолжительность этой корреляции представляет собой время персистенции, которое для многих амебоидных клеток в буфере составляет несколько минут, в течение которых вытягивается около десяти ложноножек. Статистический анализ расщепления псевдоподий и образования псевдоподий de novo у Dictyostelium показал, что угол между двумя расщепляющимися ложноножками составляет ~55 градусов, и что за расщеплением справа часто следует расщепление слева (Bosgraaf and van Haastert, 2009b). Это попеременное расщепление под относительно небольшим углом приводит к устойчивой зигзагообразной траектории. Напротив, псевдоподии de novo расширяются в любом направлении, не имеют предпочтения вправо-влево по отношению к предыдущей или следующей псевдоподии и, следовательно, вызывают случайный поворот в направлении движения клетки. Как следствие, время персистенции движения зависит от соотношения расщепления и образования псевдоподий de novo.
Исследования мутантов Dictyostelium показали, что образование псевдоподий de novo ингибируется путем цГМФ, который индуцирует миозиновые филаменты в коре тела клетки, тогда как расщеплению псевдоподов способствует передача сигналов PLA2 посредством неизвестного механизма (Bosgraaf and van Haastert, 2009б). Эти сигнальные пути используются клетками для модулирования поведения при поиске пищи (Van Haastert and Bosgraaf, 2009b). Клетки с обильным питанием имеют низкую активность PLA2 и цГМФ и образуют множество псевдоподий de novo со случайными поворотами, вызывая броуновское движение на небольшой площади. Голодание индуцирует активность PLA2 и цГМФ, тем самым сильно усиливая расщепление ложноножек, что приводит к постоянному движению клеток. Вследствие усиленного расщепления голодающие клетки посещают гораздо большую площадь в единицу времени, хотя они не расширяют больше ложноножек, чем не голодающие клетки. Похоже, что основой постоянного движения клеток является баланс чередующихся правых и левых расщеплений псевдоподий и случайных поворотов псевдоподий de novo. Помимо своей роли в поиске пищи, это стойкое расширение псевдопода используется для эффективной навигации в градиентах хемоаттрактантов.
Рис. 2.
Посмотреть в большом размереСкачать слайд
Псевдоподы вытянуты перпендикулярно поверхности клетки. ( A ) Неравные силы на мембране, окружающей выступ, который не является перпендикулярным (слева), толкают ложноножки перпендикулярно поверхности клетки (справа). ( B ) Псевдоножки, которые индуцируются на той стороне клеток дикого типа, которые обращены к градиенту (заштрихованная область на схеме справа), хорошо направлены к градиенту. ddia2 -нулевые клетки, которые имеют очень неравномерную кривизну поверхности, обнаруживают нарушенный хемотаксис, даже несмотря на то, что на той стороне клетки, которая обращена к градиенту, также образуются псевдоподии.
Рис. 2.
Посмотреть в большом размереСкачать слайд
Псевдоподы вытянуты перпендикулярно поверхности клетки. ( A ) Неравные силы на мембране, окружающей выступ, который не является перпендикулярным (слева), толкают ложноножки перпендикулярно поверхности клетки (справа). ( B ) Псевдоножки, которые индуцируются на той стороне клеток дикого типа, которые обращены к градиенту (заштрихованная область на схеме справа), хорошо направлены к градиенту. ddia2 -нулевые клетки, которые имеют очень неравномерную кривизну поверхности, обнаруживают нарушенный хемотаксис, даже несмотря на то, что на той стороне клетки, которая обращена к градиенту, также образуются псевдоподии.
Эксперименты с микропипетками, заполненными хемоаттрактантом, показали, что экспонированные клетки быстро расширяют ложноножки в направлении градиента (Gerisch and Keller, 19).81). В таких экспериментах создается впечатление, что градиент индуцирует псевдоподию, т.е. клетка считывает градиент, а затем, на основе восходящего накопления сигнальных молекул, вытягивает псевдоподию в эту сторону (Bourne, Weiner, 2002; Meili, Firtel, 2003; Вайнер, 2002). С этой точки зрения было высказано предположение, что небольшая разница концентраций хемоаттрактанта в клетке обрабатывается механизмами возбуждения-адаптации или обратной связи-инактивации для создания сильного локального внутриклеточного сигнала для образования псевдоподий (Iglesias and Devreotes, 2008; Kutscher et al. , 2004; Левин и др., 2006). Этот «градиентно-ориентированный» взгляд на хемотаксис может скрывать более общее и концептуальное понимание ориентированного движения. Для подтверждения этого положения можно использовать «мысленный эксперимент». Предположим, что клетка движется в очень мелком градиенте хемоаттрактанта, который вызывает небольшую, но значительную активацию внутриклеточных сигнальных путей в определенной области клетки. Таким образом, эндогенный пространственно-временной ритм расширения псевдоподий подвергается небольшому индуцированному градиентом пространственно-временному смещению, которое может изменить вероятность того, где и когда сформируется новый псевдопод. Мысленный эксперимент предполагает, что неглубокий градиент не индуцирует псевдоподии, но он накладывает позиционное смещение на псевдоподию, которая уже может сформироваться. С этой точки зрения на хемотаксис, направленной «псевдоподами», локальные механизмы возбуждения-адаптации не являются фундаментальными для направленного движения. На самом деле, любой механизм, который вызывает позиционное смещение формирования псевдоподий, будет индуцировать направленное движение, тогда как механизмы возбуждения-адаптации могут повышать точность, с помощью которой клетка может обнаруживать неглубокие градиенты.
Эксперименты с Dictyostelium показали, что неглубокий градиент хемоаттрактанта не влияет на частоту, скорость роста, продолжительность жизни и размер отростков ложноножек (Andrew and Insall, 2007; Bosgraaf and Van Haastert, 2009a). Таким образом, клеточное движение продолжается с небольшим градиентом в соответствии с эндогенным циклом самоорганизующихся псевдоподий. Градиент хемоаттрактанта оказывает на ложноножки три основных эффекта: избирательное втягивание, ориентированное вытягивание и подавление образования ложноножек de novo (Bosgraaf and Van Haastert, 2009).а). Селективная ретракция основана на стремлении поляризованных клеток двигаться только с одной псевдоножкой за раз. Таким образом, когда клетки иногда имеют несколько ложноножек, они обычно втягивают все ложноножки, кроме одного, чтобы сохранить полярность. В градиенте хемоаттрактанта сохраняется ложноподия, которая лучше всего ориентируется в направлении градиента (Andrew, Insall, 2007; Bosgraaf, Van Haastert, 2009a). Хотя избирательная ретракция легко наблюдается во время хемотаксиса, поляризованные клетки обычно двигаются только с одним псевдоподом, который образуется в результате одностороннего расщепления. Такие клетки хемотаксируют, потому что градиент хемоаттрактанта индуцирует небольшое смещение направления новых ложноножек в сторону градиента (Bosgraaf and Van Haastert, 2009).а). В буфере новый расщепляющийся псевдопод начинается на небольшом расстоянии от кончика текущего псевдопода, что приводит к углу ~ 55 градусов, который наблюдался между расщепляющимися псевдоподами (рис. 3). Неглубокий градиент вызывает небольшое смещение положения, в котором начинается псевдоподия, так что она срабатывает немного ближе к той стороне клетки, которая обращена к градиенту (рис. 3). Угол между последующими ложноножками не 55 градусов, а больше или меньше в зависимости от расположения хемоаттрактанта. Вследствие этого позиционного смещения в среднем больше ложноножек вытягивается в направлении градиента, чем в других направлениях.
Как обсуждалось выше, вполне вероятно, что индуцированное градиентом позиционное смещение удлинения псевдоподий является результатом комбинирования эндогенных активаторов псевдоподов (что привело бы к расщеплению под углом 55 градусов) и активаторов, индуцированных градиентом. Используя сигнальные мутанты Dictyostelium , мы продемонстрировали, что по крайней мере три пути способствуют этому позиционному смещению: путь PI3K, путь TorC2 и белок sGC (Bosgraaf and Van Haastert, 2009a). Эти белки и/или их продукты накапливаются на восходящей стороне клетки, где они увеличивают вероятность того, что в этой области начнет формироваться ложноножка (Kamimura et al., 2008; Merlot and Firtel, 2003; Parent et al., 19).98; Veltman and Van Haastert, 2006) (рис. 4). Рисунок 3 На рисунке показана клетка, сформировавшая ложноножку слева и вытянувшая новую ложноножку. ( A ) Влияние собственных сигналов на формирование ложноножек в буфере. Линии показывают измеренную вероятность того, что новый псевдопод начнется в данной позиции. Было высказано предположение, что четыре регулятора участвуют в удлинении ложноножек, которое происходит в отсутствие внешних сигналов: цГМФ индуцирует миозиновые филаменты, которые ингибируют образование ложноножек de novo в теле клетки, PLA2 стимулирует расщепление ложноножек, неизвестные регуляторы ингибируют образование новых ложноножек. псевдоподия на кончике текущей псевдоподии и неизвестные регуляторы также стимулируют попеременное расщепление вправо-влево. Черная стрелка указывает наиболее вероятное направление расширения новых псевдоподий. ( B ) Влияние внутренних и внешних сигналов на образование ложноножек во время хемотаксиса. Градиент цАМФ активирует цГМФ и PLA2, тем самым усиливая расщепление псевдоподов, и локально активирует белок PI3K, TorC2 и sGC. Вероятность того, что новый псевдопод сформируется в данном месте, зависит от относительной силы эндогенных сигналов и сигналов, индуцированных хемоаттрактантом. Красная стрелка указывает вероятное направление новой псевдоподии в буфере, тогда как черная стрелка указывает направление в градиенте; изменение направления определяется как смещение, вызванное градиентом. Адаптировано из ранее опубликованного рисунка (Bosgraaf and Van Haastert, 2009 г.).а). Рисунок 3 На рисунке показана клетка, сформировавшая ложноножку слева и вытянувшая новую ложноножку. ( A ) Влияние собственных сигналов на формирование ложноножек в буфере. Линии показывают измеренную вероятность того, что новый псевдопод начнется в данной позиции. Было высказано предположение, что четыре регулятора участвуют в удлинении ложноножек, которое происходит в отсутствие внешних сигналов: цГМФ индуцирует миозиновые филаменты, которые ингибируют образование ложноножек de novo в теле клетки, PLA2 стимулирует расщепление ложноножек, неизвестные регуляторы ингибируют образование новых ложноножек. псевдоподия на кончике текущей псевдоподии и неизвестные регуляторы также стимулируют попеременное расщепление вправо-влево. Черная стрелка указывает наиболее вероятное направление расширения новых псевдоподий. ( B ) Влияние внутренних и внешних сигналов на образование ложноножек во время хемотаксиса. Градиент цАМФ активирует цГМФ и PLA2, тем самым усиливая расщепление псевдоподов, и локально активирует белок PI3K, TorC2 и sGC. Вероятность того, что новый псевдопод сформируется в данном месте, зависит от относительной силы эндогенных сигналов и сигналов, индуцированных хемоаттрактантом. Красная стрелка указывает вероятное направление новой псевдоподии в буфере, тогда как черная стрелка указывает направление в градиенте; изменение направления определяется как смещение, вызванное градиентом. Адаптировано из ранее опубликованного рисунка (Bosgraaf and Van Haastert, 2009 г.).а).
Наконец, в дополнение к этим пространственным эффектам, градиент также вызывает подавление образования псевдоподий de novo, за счет чего увеличивается время сохранения движения. Это усиленное хемоаттрактантом расщепление псевдоподов опосредуется двумя другими сигналами посредством активации PLA2 и продукции cGMP с помощью sGC (Bosgraaf and Van Haastert, 2009a).
Расщепление псевдопод индуцирует постоянство движения: клетка имеет сильную тенденцию продолжать движение в том же направлении (Andrew and Insall, 2007; Bosgraaf and van Haastert, 2009).б; Ли и др., 2008). Как упоминалось выше, постоянство за счет расщепления псевдоподов действует как память о направлении с временной шкалой около десяти расширений псевдоподов. В градиенте память постоянства функционирует как интегратор позиционного смещения, которому подверглись предыдущие ~ 10 псевдоподий. Следовательно, смещение, которое градиент оказывает на каждую псевдоподию, может быть очень небольшим, потому что клетка становится лучше ориентированной в сторону градиента при каждом последующем расщеплении псевдоподии, пока не будет достигнуто устойчивое состояние.
Рис. 4.
Просмотреть в большом размереСкачать слайд
Интеграция подходов, ориентированных на сигнал и псевдоножек. Исследования, которые изучают и отслеживают трансдукцию градиента цАМФ, выявили участие нескольких сигнальных путей и продемонстрировали центральную роль в активации Ras. Исследования, изучающие, как формируются псевдоподии, определили способ движения клеток и роль сигнальных путей в удлинении псевдоподий. cAR, рецептор цАМФ; RasC/G, два Ras-белка, участвующих в хемотаксисе; sGCp, белок sGC, АК, арахидоновая кислота как потенциальный медиатор активации PLA2.
Рис. 4.
Просмотреть в большом размереСкачать слайд
Интеграция подходов, ориентированных на сигнал и псевдоножек. Исследования, которые изучают и отслеживают трансдукцию градиента цАМФ, выявили участие нескольких сигнальных путей и продемонстрировали центральную роль в активации Ras. Исследования, изучающие, как формируются псевдоподии, определили способ движения клеток и роль сигнальных путей в удлинении псевдоподий. cAR, рецептор цАМФ; RasC/G, два Ras-белка, участвующих в хемотаксисе; sGCp, белок sGC, АК, арахидоновая кислота как потенциальный медиатор активации PLA2.
Чувствительность клетки к очень мелким градиентам зависит от отношения сигнал/шум, т. е. способности ощущать разницу в концентрации хемоаттрактанта в клетке (сигнал) на фоне неизбежного шума, создаваемого стохастическими вариациями. занятости и активации рецепторов (Berg and Purcell, 1977; Rappel and Levine, 2008; Ueda and Shibata, 2007; Van Haastert and Postma, 2007). Шум можно значительно уменьшить, если градиент измеряется ячейкой неоднократно, а затем усредняется (Ueda, Shibata, 2007; Van Haastert, Postma, 2007). Усреднение пространственной информации во времени требует быстрых взаимодействий лиганд-рецептор в сочетании с пространственной и временной интеграцией активности рецептора. В Dictyostelium , комплекс цАМФ-рецептор имеет время жизни ~1 секунду. Медленно диффундирующие вторичные мессенджеры, такие как PtdIns(3,4,5) P 3 , интегрируют взаимодействия цАМФ-рецептора в течение ~10-секундного интервала; более длительные периоды интеграции для этих молекул невозможны, потому что диффузия вызывает потерю пространственной информации (Van Haastert and Postma, 2007). Память, которую несут десять расщеплений псевдоподов, увеличивает время интеграции до ~ 180 секунд. Поскольку шум снижается с квадратным корнем из времени интегрирования, коллективное усреднение информации о рецепторах за счет медленного распространения вторичных мессенджеров и расщепления псевдоподов увеличивает чувствительность клеток к обнаружению неглубоких градиентов примерно в 13 раз.
Эксперименты показывают, что решающим моментом для клеточного движения и хемотаксиса является положение на клеточной поверхности, где зарождается ложноножка, потому что после этого начинается самоорганизация, и ложноножки растут перпендикулярно клеточной поверхности в течение определенного времени и на определенное расстояние . Вероятно, нет принципиальных различий между движением клеток в буфере и движением в неглубоких или крутых градиентах хемоаттрактанта. Упорядоченное расширение расщепления псевдоподов и формирование псевдоподов de novo, что приводит к постоянному движению в буфере, продолжается с той же частотой в неглубоких градиентах. Низкие концентрации индуцированных градиентом сигнальных молекул недостаточны, чтобы вмешиваться во время цикла псевдоподии, но индуцируют позиционное смещение, где начинается новая псевдоподия. Крутой градиент индуцирует высокие локальные концентрации активности, индуцирующей псевдоподии, которая не только сильно влияет на положение новой псевдоподии, но также может мешать эндогенному времени удлинения псевдоподии.
Наблюдения за расширением ложноножек у Dictyostelium предполагают, что хемотаксис не является детерминированным: градиент не определяет удлинение ложноножек, время или направление. Вместо этого хемотаксис является вероятностным: клетки имеют сильный базальный цикл псевдоподий с неслучайными вероятностями времени и места, в которых происходит удлинение псевдоподий. Градиент хемоаттрактанта индуцирует смещение этих вероятностей, так что в среднем больше ложноножек инициируется на той стороне клетки, которая ближе всего к градиенту. Этот вероятностный взгляд на хемотаксис может быть верным и для других способов ориентированного движения клеток, таких как электротаксис или термотаксис. В электрических полях сигнальные молекулы, такие как PI3K и sGC, перемещаются к анодной или катодной стороне приложенного электрического поля и увеличивают вероятность образования псевдоподий, тем самым вызывая направленное движение (Sato et al., 2009).; Чжао и др., 2006).
Хотя большая часть этого обсуждения основана на результатах исследований Dictyostelium , весьма вероятно, что хемотаксис других организмов опосредуется подобным стохастическим смещением базального цикла псевдоподий (Arrieumerlou and Meyer, 2005). Расщепление псевдоподий — основа персистентного клеточного движения — наблюдается в разных типах клеток (Andrew and Insall, 2007). В клетках многих организмов хемоаттрактанты регулируют F-актин в переднем крае для образования выпячиваний и регулируют миозиновые филаменты в задней части клетки, чтобы подавить образование псевдоподов de novo и способствовать образованию втягивающихся уроподов. Однако молекулы, которые регулируют рост цитоскелета и ложноножек, могут различаться у разных организмов. В частности, роль PI3K-PtdIns(3,4,5) P 3 передача сигналов в этих процессах, вероятно, сохраняется между Dictyostelium , нейтрофилами и другими типами клеток в других организмах. myosin в Dictyostelium опосредуется Rho-ассоциированной протеинкиназой (ROCK) в клетках млекопитающих (Li et al., 2005; Schneider and Haugh, 2006; Smirnova and Segall, 2007).
Сигнальные пути, индуцированные хемоаттрактантами, часто запутываются в сложных нелинейных петлях обратной связи, вызывая нарушение симметрии, адаптацию и усиление (Iglesias and Devreotes, 2008). Такие сложные регуляторные пути, вероятно, были включены в ходе эволюции для улучшения отношения сигнал/шум, тем самым делая возможным хемотаксис в более мелких градиентах. Однако в этой стохастической модели хемотаксиса, управляемого псевдоподами, ни одна из этих петель не является существенной для определения градиента. Действительно, 9Мутантные клетки 0005 Dictyostelium , в которых ингибированы все четыре сигнальных фермента (PI3K, TorC2, PLA2 и sGC), обнаруживают хороший хемотаксис, но только в очень крутых градиентах (Veltman et al. , 2008). В мутантных клетках, в которых активен только один из четырех ферментов, хемотаксис происходит в градиентах, которые примерно в 8 раз меньше, тогда как наличие всех четырех путей обеспечивает хемотаксис в градиентах, которые примерно в 150 раз меньше (мои неопубликованные результаты). .
Предлагаю подойти к проблеме хемотаксиса с двух сторон (рис. 4). Подход, который следует за сигналом, переносимым градиентом цАМФ, исследует временную и пространственную активацию сигнальных путей. Этот подход очень эффективен для обнаружения регуляторных механизмов в восходящей части процесса сенсорной трансдукции, таких как адаптация и нарушение симметрии, которые, вероятно, происходят на уровне активации Ras (Zhang et al., 2008). Наоборот, подход, ориентированный на псевдоподии, может обеспечить более механистическое понимание того, как сигнальные пути синергируют, как они взаимодействуют с цитоскелетом и как они влияют на формирование псевдоподий. Эти методы позволят охарактеризовать базальный способ формирования псевдоподий, что будет способствовать нашему пониманию хемотаксиса.
Я благодарен Леонарду Босграафу, Арьяну Кортхольту и Инеке Кейзер-Ганнинк за обсуждения и критические комментарии к рукописи.
Эндрю
Н.
,
Инсолл
Р. Х.
(
2007
).
Хемотаксис в неглубоких градиентах опосредуется независимо от PtdIns 3-киназы за счет предвзятого выбора между случайными выступами
.
Нац. Клеточная биол.
9
,
193
—
200
.
Arrieumerlou
C.
,
Meyer
T.
(
2005
).
Модель локальной связи и параметр компаса для эукариотического хемотаксиса
.
Дев. Ячейка
8
,
215
—
227
.
Бахат
А.
,
Айзенбах
М.
(
2006
).
Термотаксис сперматозоидов
.
Мол. Клеточный эндокринол.
252
,
115
—
119
.
Berg
H.C.
,
Purcell
E. M.
(
1977
).
Физика хеморецепции
.
Биофиз. Дж.
20
,
193
—
219
.
Bosgraaf
L.
,
Van Haastert
P. J. M.
(
2009a
).
Навигация хемотаксических клеток за счет параллельной передачи сигналов к сохранению и ориентации ложноножек
.
PLoS ONE
4
,
e6842
.
Bosgraaf
L.
,
Ван Хаастерт
PJM
(
2009b
).
Упорядоченное удлинение псевдоподий амебоидной клеткой при отсутствии внешних сигналов
.
PLoS ONE
4
,
e5253
.
Bosgraaf
L.
,
Van Haastert
PJM
(
2009c
).
Quimp3, автоматизированный алгоритм отслеживания псевдоподов
.
Ячейка Adh. Мигр.
4
,
1
.
Bourne
H. R.
,
Weiner
O.
(
2002
).
Химический компас
.
Природа
419
,
21
.
Franca-Koh
J.
,
Камимура
Y.
,
Devreotes
стр.
(
2006
).
Навигационные сигнальные сети: хемотаксис у Dictyostelium discoideum
.
Курс. мнение Жене. Дев.
16
,
333
—
338
.
Friedl
P.
,
Wolf
K.
(
2009
).
Пластичность клеточной миграции: многомасштабная модель настройки
.
J. Cell Biol.
188
,
11
—
19
.
Гейл
М. Х.
,
Бун
К. В.
(
1970
).
Движение фибробластов мыши в культуре тканей
.
Биофиз. Дж.
10
,
980
—
993
.
Гериш
Г.
,
Келлер
Х.У.
(
1981
).
Хемотаксическая переориентация гранулоцитов, стимулированных микропипетками, содержащими fMet-Leu-Phe
.
J. Cell Sci.
52
,
1
—
10
.
Hoeller
O.
,
Kay
R.
(
2007
).
Хемотаксис в отсутствие градиентов PIP3
.
Курс. биол.
17
,
813
—
817
.
Иглесиас
P. A.
,
Devreotes
PN
(
2008
).
Навигация по моделям хемотаксиса
.
Курс. мнение Клеточная биол.
20
,
35
—
40
.
Insall
RH
(
2010
).
Понимание хемотаксиса эукариот: точка зрения, ориентированная на ложноножки
.
Нац. Преподобный Мол. Клеточная биол.
11
,
453
—
458
.
Джин
Т.
,
Чжан
Н.
,
Лонг
Ю.
,
Parent
C.A.
,
Devreotes
PN
(
2000
).
Локализация бетагамма-комплекса G-белка в живых клетках при хемотаксисе
.
Наука
287
,
1034
—
1036
.
Камимура
Ю.
,
Сюн
Ю.
,
Иглесиас
P. A.
,
Hoeller
O.
,
Bolourani
P.
,
Devreotes
P. N.
(
999999444444444444444444444444444444444444444444444. N.
(
).
Независимая от PIP3 активация TorC2 и PKB на переднем крае клетки опосредует хемотаксис
.
Курс. биол.
18
,
1034
—
1043
.
Карсенти
Э.
(
2008
).
Самоорганизация в клеточной биологии: краткая история
.
Нац. Преподобный Мол. Клеточная биол.
9
,
255
—
262
.
Kay
R. R.
,
Langridge
P.
,
Traynor
D.
,
Hoeller
O.
(
9O.
(
O.
(
4.0004 2008
).
Изменение направления в изучении хемотаксиса
.
Нац. Преподобный Мол. Клеточная биол.
9
,
455
—
463
.
King
J. S.
,
Insall
R. H.
(
2009
).
Хемотаксис: путь вперед с Dictyostelium
.
Ячейка трендов. биол.
19
,
523
—
530
.
Kutscher
B.
,
Devreotes
P.
,
Iglesias
P. A.
(
2004
).
Механизм локального возбуждения, глобального торможения для определения градиента: интерактивный апплет
.
Науч. СТКЭ
2004
,
пл3
.
Lammermann
T.
,
Sixt
M.
(
2009
).
Механические способы миграции «амебоидных» клеток
.
Курс. мнение Клеточная биол.
21
,
636
—
644
.
Levine
H.
,
Kessler
D. A.
,
Rappel
W. J.
(
2006
).
Направленное зондирование при эукариотическом хемотаксисе: сбалансированная модель инактивации
.
Проц. Натл. акад. науч. США
103
,
9761
—
9766
.
LI
L.
,
Norrelykke
S. F.
,
COX
E. C.
(
2008
).
Персистентное движение клеток в отсутствие внешних сигналов: стратегия поиска эукариотических клеток
.
PLoS ONE
3
,
e2093
.
Li
Z.
,
Dong
X.
,
Wang
Z.
,
Liu
W.
,
Deng
N.
,
Дин
Ю.
,
Тан
Л.
,
Хла
Т.
,Zeng
R.
,
Li
L.
, и др. . (
2005
).
Регулирование PTEN малыми GTPases Rho
.
Нац. Клетка. биол.
7
,
399
—
404
.
Махачек
М.
,
Данусер
Г.
(
2006
).
Морфодинамическое профилирование фенотипов протрузии
.
Биофиз. Дж.
90
,
1439
—
1452
.
Maeda
Y. T.
,
Inose
J.
,
Matsuo
M. Y.
,
Iwaya
S.
,
Sano
M.
(
2008
).
Упорядоченные паттерны формы и ориентации клеток при спонтанной миграции клеток
.
PLoS ONE
3
,
e3734
.
Meili
R.
,
Firtel
R. A.
(
2003
).
Два шеста и компас
.
Ячейка
114
,
153
—
156
.
Мерло
С.
,
Фиртел
Р. А.
(
2003
).
Прокладывая путь: определение направления посредством фосфатидилинозитол-3-киназы и других сигнальных путей
.
J. Cell Sci.
116
,
3471
—
3478
.
Могильнер
А.
,
Остер
Г.
(
1996
).
Физика ламеллоподиального выпячивания
.
Евро. Биофиз. Дж.
25
,
47
—
53
.
Parent
C. A.
,
Blacklock
B. J.
,
Froehlich
W. M.
,
Murphy
D. B.
,
Devreotes
P. N.
(
1998
).
Сигнальные события G-белка активируются на переднем крае хемотаксических клеток
.
Ячейка
95
,
81
—
91
.
Patlak
C.S.
(
1953
).
Случайное блуждание с инерционностью и внешним смещением
.
Бык. Мат. Биофиз.
15
,
311
—
338
.
Потел
М. Дж.
,
Маккей
С. А.
(
1979
).
Преагрегационное движение клеток Dictyostelium
.
J. Cell Sci.
36
,
281
—
309
.
Rappel
WJ
,
Levine
H.
(
2008
).
Шум рецепторов и определение направления при эукариотическом хемотаксисе
.
Физ. Преподобный Летт.
100
,
228101
.
Sato
M. J.
,
Kuwayama
H.
,
van Egmond
W. N.
,
Takayama
A. L.
,
Takagi
H.
,
Ван Хаастерт
П. Дж.
,
Янагида
Т.
,
Уэда
М.
(
2009
).
Переключение направления индуцированной электрическим сигналом миграции клеток посредством циклического гуанозинмонофосфата и передачи сигналов фосфатидилинозитолом
.
Проц. Натл. акад. науч. США
106
,
6667
—
6672
.
Schneider
I. C.
,
Haugh
J.M.
(
2006
).
Механизмы определения градиента и хемотаксиса: консервативные пути, разнообразная регуляция
.
Клеточный цикл
5
,
1130
—
1134
.
Шендеров
А. Д.
,
Шитз
М. П.
(
1997
).
Обратно коррелированные циклы скорости и поворота у амебы: колебательная модель клеточной локомоции
.
Биофиз. Дж.
72
,
2382
—
2389
.
Смирнова
Т.
,
Сегал
Дж. Э.
(
2007
).
Амебоидный хемотаксис: будущие проблемы и возможности
.
Ячейка Adh. Мигр.
1
,
165
—
170
.
Солл
D. R.
,
Wessels
D.
,
HEID
P. J.
,
VOSS
E.
(
2003
).Компьютерная реконструкция и анализ движения трехмерной ячейки
.
ScientificWorldJournal
3
,
827
—
841
.
Такаги
Ч.
,
SATO
M. J.
,
Yanagida
T.
,
UEDA
M.
(
2008
).
Функциональный анализ спонтанного движения клеток в различных физиологических условиях
.
PLoS ONE
3
,
e2648
.
Уэда
М.
,
Шибата
Т.
(
2007
).
Стохастическая обработка сигналов и трансдукция при хемотаксическом ответе эукариотических клеток
.
Биофиз. Дж.
93
,
11
—
20
.
Van Haastert
PJM
,
Postma
M.
(
2007
).
Смещенное случайное блуждание из-за стохастических флуктуаций взаимодействия хемоаттрактанта с рецептором на нижнем пределе обнаружения
.
Биофиз. Дж.
93
,
1787
—
1796
.
Van Haastert
P. J.
,
Bosgraaf
L.
(
2009a
).
Локальная кривизна клеток направляет псевдоподии к хемоаттрактантам
.
HFSP J.
3
,
282
—
286
.
Van Haastert
PJM
,
Bosgraaf
L.
(
2009b
).
Стратегия поиска пищи амебоидными клетками за счет увеличения длины прогона, вызванного голоданием
.
PLoS ONE
4
,
e6814
.
Варнум
Б.
,
Солл
Д. Р.
(
1984
).
Влияние цАМФ на подвижность одиночных клеток Dictyostelium
.
J. Cell Biol.
99
,
1151
—
1155
.
Varnum-Finney
B. J.
,
Voss
E.
,
Soll
D. R.
(
1987
).
Частота и ориентация формирования псевдоподий хемотаксиса Dictyostelium discoideum amebae в пространственном градиенте: дополнительные доказательства временного механизма
.
Селл Мотил. Цитоскелет
8
,
18
—
26
.
Veltman
D.M.
,
Van Haastert
P.J.M.
(
2006
).
Белок гуанилатциклазы и продукт цГМФ независимо контролируют переднюю и заднюю части хемотаксирующих клеток Dictyostelium
.
Мол. биол. Сотовый
17
,
3921
—
3929
.
Veltman
D. M.
,
Keizer-Gunnink
I.
,
Van Haastert
P. J. M.
(
2008
).
Четыре ключевых сигнальных пути, опосредующих хемотаксис у Dictyostelium discoideum
.
J. Cell Biol.
180
,
747
—
753
.
Weijer
CJ
(
2009
).
Коллективная миграция клеток в процессе развития
.
J. Cell Sci.
122
,
3215
—
3223
.
Weiner
НД
(
2002
).
Регуляция клеточной полярности во время эукариотического хемотаксиса: хемотаксический компас
.
Курс. мнение Клеточная биол.
14
,
196
—
202
.
Zhang
S.
,
Charest
P. G.
,
Firtel
R. A.
(
2008
).
Пространственно-временная регуляция активности Ras обеспечивает направленное зондирование
.
Курс. биол.
18
,
1587
—
1593
.
Чжао
М.
(
2009
).
Электрические поля при заживлении ран. Основной сигнал, управляющий миграцией клеток
.
Семин. Сотовый Дев. биол.
20
,
674
—
682
.
Чжао
М.
,
Сун
Б.
,
Пу
Дж.
,
Wada
T.
,
Reid
B.
,
Tai
G.
,
Wang
F.
,
Guo
A.
,
Вальчиско
П.
,
Гу
Ю.
, и др. . (
2006
).
Электрические сигналы контролируют заживление ран с помощью фосфатидилинозитол-3-ОН киназы-гамма и PTEN
.
Природа
442
,
457
—
460
.
Узнайте о псевдоподах | Chegg.com
Псевдоподы Определение
Псевдоподы определяются как небольшие выступы, которые выходят из протистов и помогают в движении и захвате добычи.
Обзор псевдоподов
Псевдоподы определяются как выросты, которые выходят из клеточной мембраны простейших и помогают организму захватывать добычу и перемещаться из одного места в другое. Он состоит из микротрубочек, которые в значительной степени поддерживают движение протистов. Ламеллиподий заполняется цитоплазмой, образуя выступы псевдоподий. Эти типы выступов сокращаются и расширяются во время полимеризации актина. Эти сокращения и расслабления толкают клетку вперед. У некоторых животных псевдоподии представляют собой приспособления, которые помогают животным эффективно перемещаться в окружающей среде.
Есть вопрос по этой теме?
What you’ll learn:
- pseudopod Definition
- Overview of Pseudopod
- Types of Pseudopods:
- Functions of Pseudopod:
- Examples of Pseudopod:
Types of Pseudopods:
Pseudopods are of different sizes , типы и формы. Различают следующие типы псевдоподий:
Лобоподии: Этот тип ложноножек имеет форму пальца и чаще всего встречается в природе.
Они делятся на луковичные, тупые и короткие отростки, которые включают как эктоплазму, так и эндоплазму. Этот тип ложноножек наблюдается у лопастных амеб, у которых ложноножки одни из самых крупных.Филоподии: Этот тип ложноножек похож на нить и имеет тонкую структуру. Он чрезвычайно тонкий и нитевидный с острыми концами, состоящими из эктоплазмы. Микрофиламенты поддерживают проекцию этого типа ложноножек. Примером организма с филоподиями является Euglypha. Филоподии состоят из различных модификаций. Например, филоподии похожи на гранулоподии. Гранулоподии представляют собой структуру с гранулами, и эти гранулы известны как экструсомы. Гранулоретикулоподия — это другое несоответствие гранулоподии, которое находится между ретикулоподией и филиподией.
Ретикулоподии: образует сеть, известную как ретикулум. Он также известен как ретикулезные псевдоподии. Он образует выступообразный комплекс, в котором псевдоподии сливаются друг с другом и образуют неправильную сетчатую структуру.
Этот тип ложноножек наблюдается у Foramineferans.Аксоподии: псевдоподии этого типа покрыты цитоплазмой; следовательно, он чаще всего используется для проглатывания пищевых частиц и процесса фагоцитоза. К организмам, имеющим этот тип псевдоподий, относятся Heliozoa и Radiolaria.
Функции псевдоподов:
Псевдоподы в основном выполняют две функции — заглатывание пищи (захват добычи) и передвижение. Например, амеба может ползти за счет сокращения нити и расширения цитоплазмы. По мере продвижения вперед ложноножки выпячиваются от края клетки, чтобы поглотить целый организм. Помимо этого, ложноножки также используются в процессе проглатывания и захвата добычи. Они также используются при проглатывании гранулированных веществ. Это необходимо для обнаружения добычи. Он поддерживает такие организмы, как амебы, и помогает им удалять антигены и токсины из организма посредством процесса, называемого фагоцитозом.
Примеры псевдоподов:
Различные типы организмов используют этот отросток или псевдопод для приема внутрь. Псевдопод также важен для осуществления процесса передвижения. Эти прогнозы можно наблюдать в следующем:
Ризоподы: Псевдоподии — это особенность простейших организмов группы, известных как корненожки, важный представитель царства протистов. Он помогает этим организмам во время передвижения. Псевдоножки помогают корненогам перемещаться из одного места в другое. Он также помогает процессу проглатывания, захватывая добычу, которая находится рядом с корненожками.
Лейкоциты: Они играют очень важную роль в обеспечении защиты организма. Это часть иммунной системы. Он также известен как лейкоциты и защищает организм от вирусов, бактерий и патогенов, которые атакуют части тела. Эти клетки имеют ложноножки в качестве выступов, которые осуществляют процесс фагоцитоза и помогают защитить организм от антигенов.
Продолжайте учиться
Что изучать дальше на основе учебной программы колледжа
Цикл лимонной кислотыСхема промежуточных филаментовЗапас питательных веществ и цитозольФункции микрофиламентаДвижение жгутикаДвижение в клеткеПроцесс окислительного фосфорилированияКератин как промежуточные филаменты
Морфологические критерии псевдоподов в поверхностной микроскопии | JAMA Dermatology
Морфологические критерии псевдоподов в поверхностной микроскопии | JAMA Дерматология | Сеть ДЖАМА [Перейти к навигации]Эта проблема
- Скачать PDF
- Полный текст
Поделиться
Твиттер Фейсбук Эл.
адрес
LinkedIn- Процитировать это
- Разрешения
Артикул
Апрель 1995 г.
Скотт В. Мензис, MBBS, PhD ; Керри А. Кротти, MBBS, BSc (Med), FRCPA ; Уильям Х. Маккарти, MBBS, MEd, FRACS
Принадлежности авторов
Сиднейское отделение меланомы, хирургическое отделение (доктор Мензис и Маккарти) и отделение патологии (доктор Кротти) Сиднейского университета (Австралия).
Арка Дерматол. 1995;131(4):436-440. doi: 10.1001 / archderm.1995.016064010
Полный текст
Абстрактный
Фон и дизайн: Было показано, что микроскопия поверхности кожи in vivo (масляная эпилюминесценция, дерматоскопия и дерматоскопия) значительно улучшает клиническую диагностику меланомы. Псевдоножки — это морфологическая особенность, видимая при поверхностной микроскопии, которая соответствует радиальному росту опухоли при меланоме. Хотя это один из наиболее специфических поверхностных микроскопических признаков инвазивной меланомы, он остается плохо определенным. Мы изучили 239пигментные поражения, 80 меланом (62 инвазивных и 18 in situ) и 159 случайно выбранных пигментных немеланом. Мы сфотографировали эти поражения in vivo с помощью иммерсионного масла и камеры Heine Dermaphot (Heine Ltd, Herrsching, Germany). Затем мы оценили поражения «слепым» способом на наличие псевдоножек на основе строго определенных морфологических критериев.
Результаты: Мы определили морфологические критерии псевдоподии. Как определено, псевдоподия сохранила 97% специфичность и 23% чувствительность для инвазивной меланомы. Не было замечено различий в средней толщине Бреслоу между меланомами с псевдоножками и без них. Ни в одной из меланом in situ не было обнаружено псевдоподий.