1. Кто открыл мир простейших животных? А. Левенгук Б. Аристотель В. Ламарк Г. Линней 2. Выберите правильные суждения:
A. Простейшие животные состоят только из одной клетки Б. В колониях простейших имеются отличные от других специализированные клетки B. Все простейшие питаются только готовыми органическими веществами Г. Неблагоприятные условия простейшие переносят, превращаясь в цисту 3. Простейшие могут обитать: А. В воде Б. В почве В. В живых организмах Г. Во всех перечисленных средах 4. Количество видов простейших, известное к настоящему времени: А. Менее 1 тыс. Б. Около 7 тыс. В. Около 70 тыс. Г. Около 700 тыс. 5. Корненожки передвигаются с помощью: А. Парных ножек Б. Ложноножек В. Ресничек Г. Корней 6. Ложноножки представляют собой: А. Скелетные образования Б. Выпячивания цитоплазмы В. Многоклеточные отростки Г. Жгутики 7. Наружный или внутренний минеральный скелет имеют: А. Только корненожки Б. Только радиолярии В. Корненожки и радиолярии Г. Все простейшие 8. Минеральный скелет простейших не обеспечивает: А. Поддержание формы тела Б. Увеличение поверхности тела В. Защиту организма Г. Запасание питательных веществ 9. В период неблагоприятных условий простейшие: А. Усиленно питаются Б. Интенсивно размножаются В. Превращаются в цисту Г. Ведут обычный образ жизни 10. Все виды жгутиконосцев перемещаются с помощью: А. Одного подвижного жгутика Б. Двух подвижных жгутиков В. Разного количества жгутиков Г. Множества ресничек 11. Все виды жгутиконосцев питаются: А. Как растения, т. е. только фотосинтезом с помощью хлорофилла Б. Как животные, т. е. только готовым органическим веществом В. На свету – как растения, а в темноте – как животные Г. Разным видам жгутиконосцев свойственны разные способы питания: растительного, животного или смешанного 12. К кишечным паразитам животных и человека относятся: A. Инфузория туфелька Б. Трихомонада, лямблия и дезинтерийная амеба B. Трипаносома, лейшмания и малярийный плазмодий Г. Все паразитические простейшие 13. К паразитам крови животных и человека относятся: A.
Поддержание формы тела Б. Увеличение поверхности тела В. Защиту организма Г. Запасание питательных веществ 9. В период неблагоприятных условий простейшие: А. Усиленно питаются Б. Интенсивно размножаются В. Превращаются в цисту Г. Ведут обычный образ жизни 10. Все виды жгутиконосцев перемещаются с помощью: А. Одного подвижного жгутика Б. Двух подвижных жгутиков В. Разного количества жгутиков Г. Множества ресничек 11. Все виды жгутиконосцев питаются: А. Как растения, т. е. только фотосинтезом с помощью хлорофилла Б. Как животные, т. е. только готовым органическим веществом В. На свету – как растения, а в темноте – как животные Г. Разным видам жгутиконосцев свойственны разные способы питания: растительного, животного или смешанного 12. К кишечным паразитам животных и человека относятся: A. Инфузория туфелька Б. Трихомонада, лямблия и дезинтерийная амеба B. Трипаносома, лейшмания и малярийный плазмодий Г. Все паразитические простейшие 13. К паразитам крови животных и человека относятся: A. Инфузория туфелька Б. Трихомонада, лямблия и дезинтерийная амеба B. Трипаносома, лейшмания и малярийный плазмодий Г. Все паразитические простейшие 14. Органическое вещество в водоеме производит: А. Пресноводная гидра Б. Инфузория туфелька В. Эвглена зеленая Г. Амеба 15. Какая из перечисленных ниже болезней не вызывается паразитическими простейшими? А. Малярия Б. Грипп В. Дизентерия Г. Сонная болезнь 16. Какие из простейших имеют непостоянную форму тела? А. Амебы Б. Радиолярии В. Инфузории Г. Все простейшие
Инфузория туфелька Б. Трихомонада, лямблия и дезинтерийная амеба B. Трипаносома, лейшмания и малярийный плазмодий Г. Все паразитические простейшие 14. Органическое вещество в водоеме производит: А. Пресноводная гидра Б. Инфузория туфелька В. Эвглена зеленая Г. Амеба 15. Какая из перечисленных ниже болезней не вызывается паразитическими простейшими? А. Малярия Б. Грипп В. Дизентерия Г. Сонная болезнь 16. Какие из простейших имеют непостоянную форму тела? А. Амебы Б. Радиолярии В. Инфузории Г. Все простейшие
Тип простейшие – Зоология – Kaz-Ekzams.ru
admin 22.07.2010
Зоология
простейшие
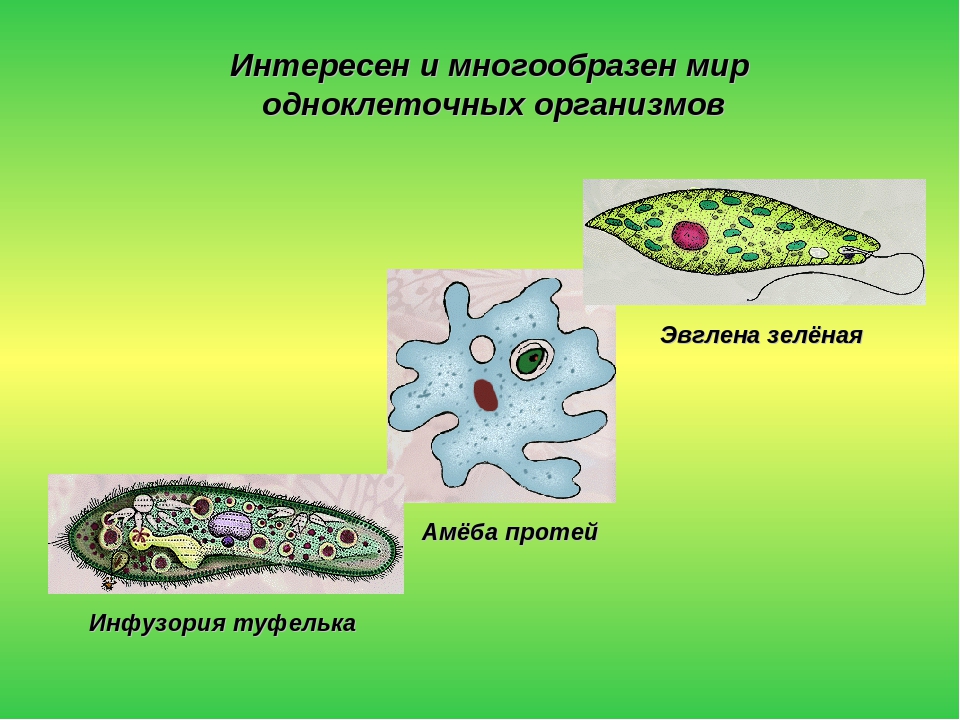
К простейшим относят животных, тело которых состоит из одной клетки. Клетка простейшего — это целостный организм, которому присущи основные физиологические процессы: дыхание, пищеварение, выделение, раздражимость. Таковы амебы, инфузории и др. Некоторые виды простейших образуют колонии (вольвокс). Простейшие обитают в пресных водоемах, морях и почве.
Простейшие обитают в пресных водоемах, морях и почве.
Некоторые из них паразитируют на других организмах. Подавляющее большинство простейших обладает способностью к инцистированию, т. е. образованию при наступлении неблагоприятных условий (понижение температуры, высыхание водоема) покоящейся стадии — цисты, покрытой плотной оболочкой. Циста напоминает споры грибов или других бесцветковых растений. Образование цисты способствует не только выживанию в неблагоприятных условиях, но и распространению простейших. При благоприятных условиях такое животное покидает оболочку цисты, начинает питаться и размножаться.
В типе простейших насчитывают около 25 000 видов, относящихся к нескольким классам.
КЛАСС КОРНЕНОЖКИ
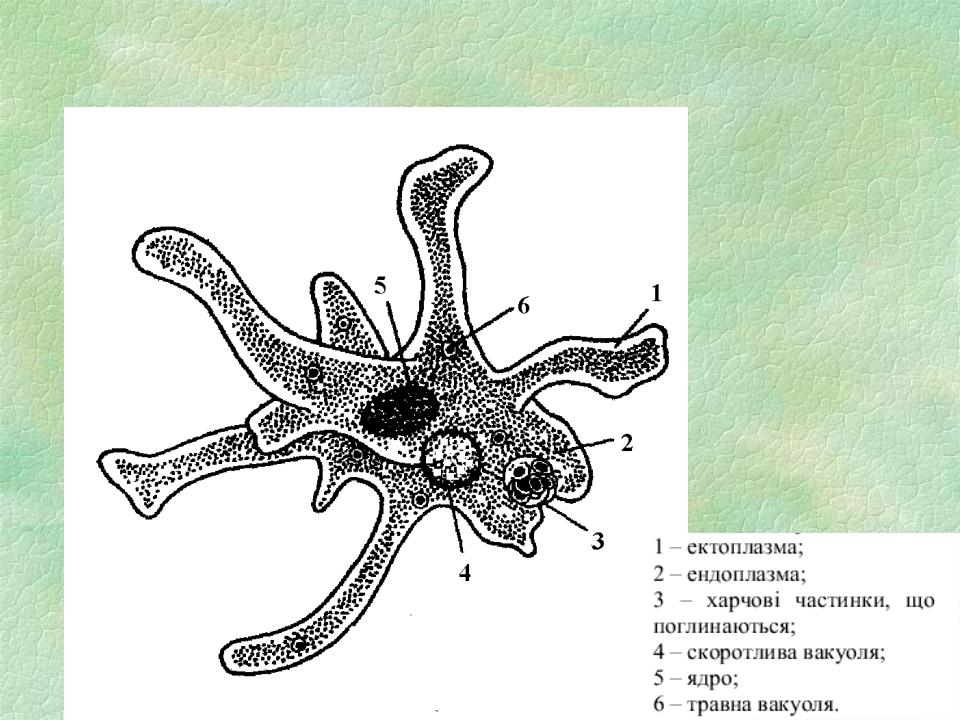
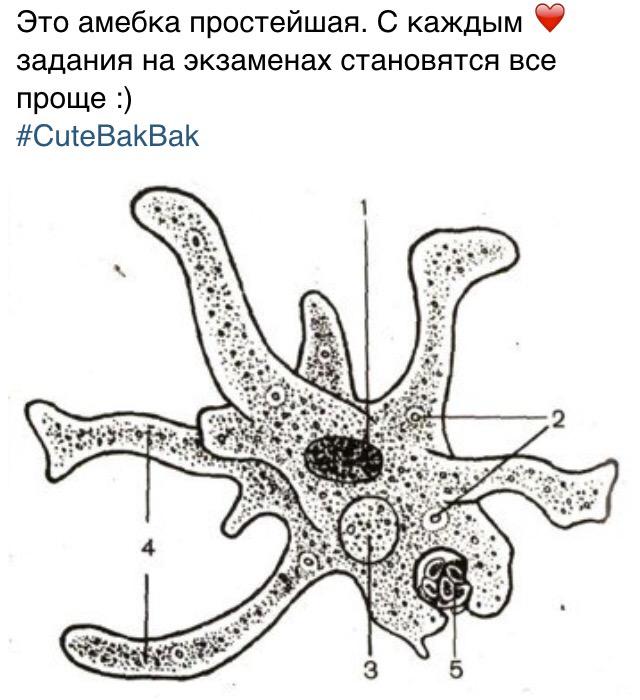
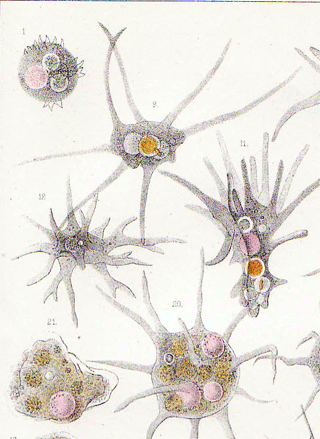
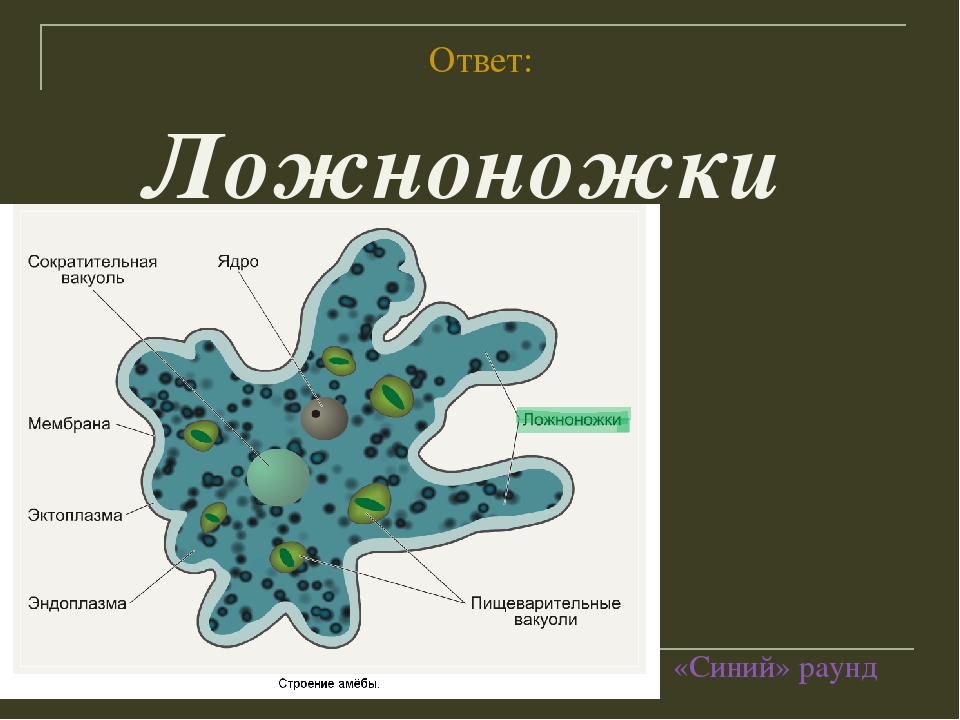
Класс корненожек объединяет наиболее примитивных одноклеточных животных, тело которых лишено плотной оболочки, а потому не имеет постоянной формы (19). Для них характерно образование ложноножек, которые представляют собой временно образующиеся выросты цитоплазмы, способствующие передвижению и захвату пищи. Ложноножки похожи на небольшие корни растения (отсюда и название класса).
Ложноножки похожи на небольшие корни растения (отсюда и название класса).
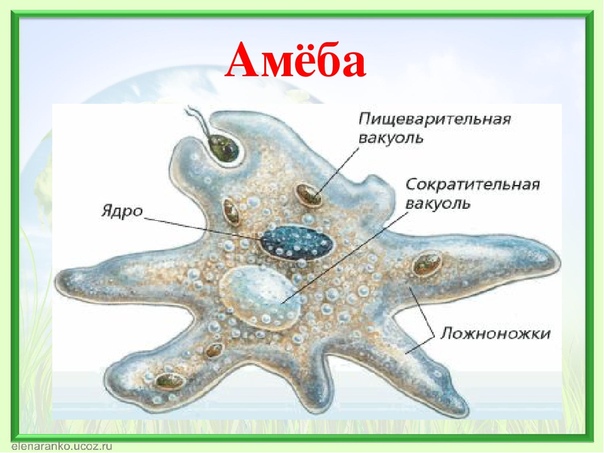
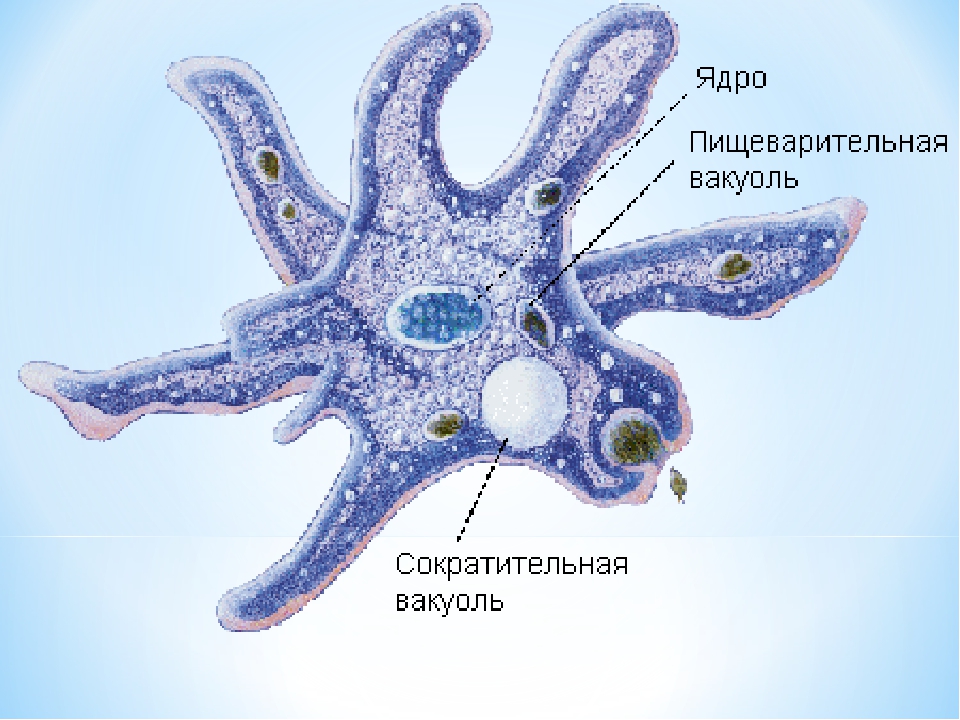
Один из представителей класса — амеба обыкновенная — состоит из комочка полужидкой, тягучей цитоплазмы, внутри которой заключено круглое ядро. Цитоплазма находится в постоянном движении. Это способствует передвижению амебы, так как часть цитоплазмы, перетекая, образует вырост — ложноножку. Одновременно ложноножки с противоположной стороны укорачиваются и амеба «перетекает» на новое место.
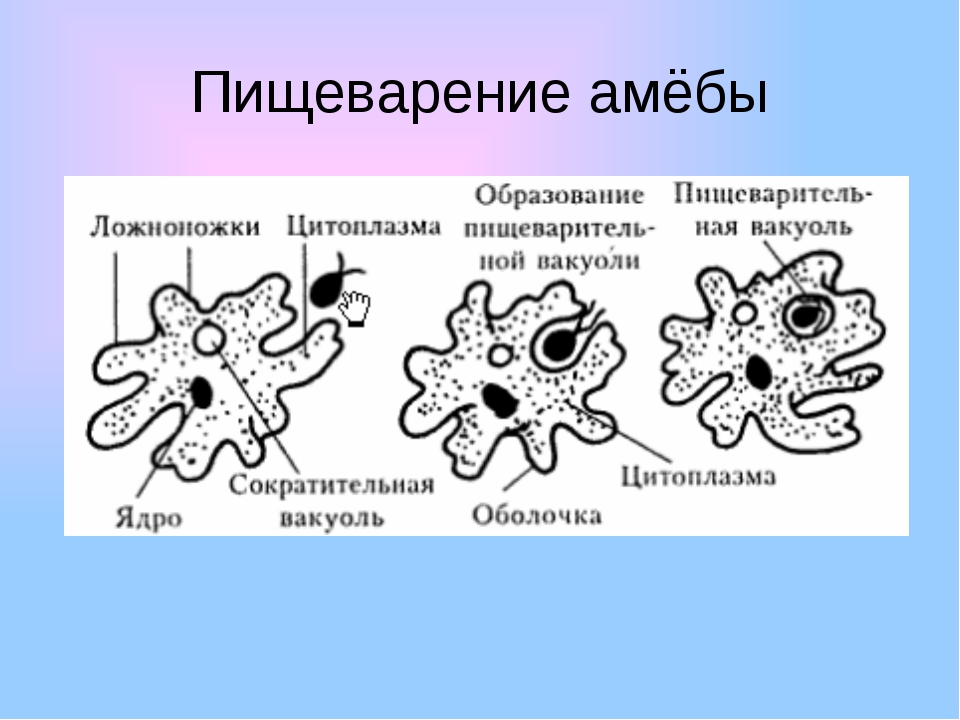
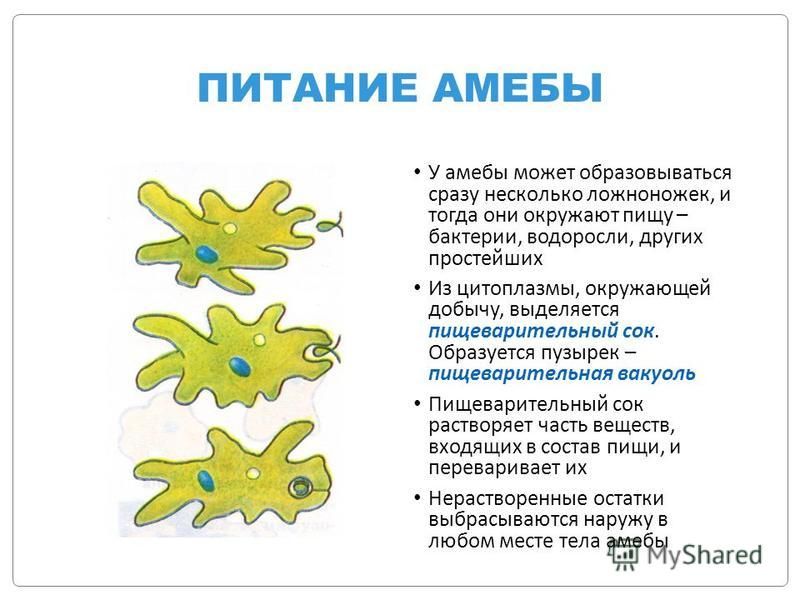
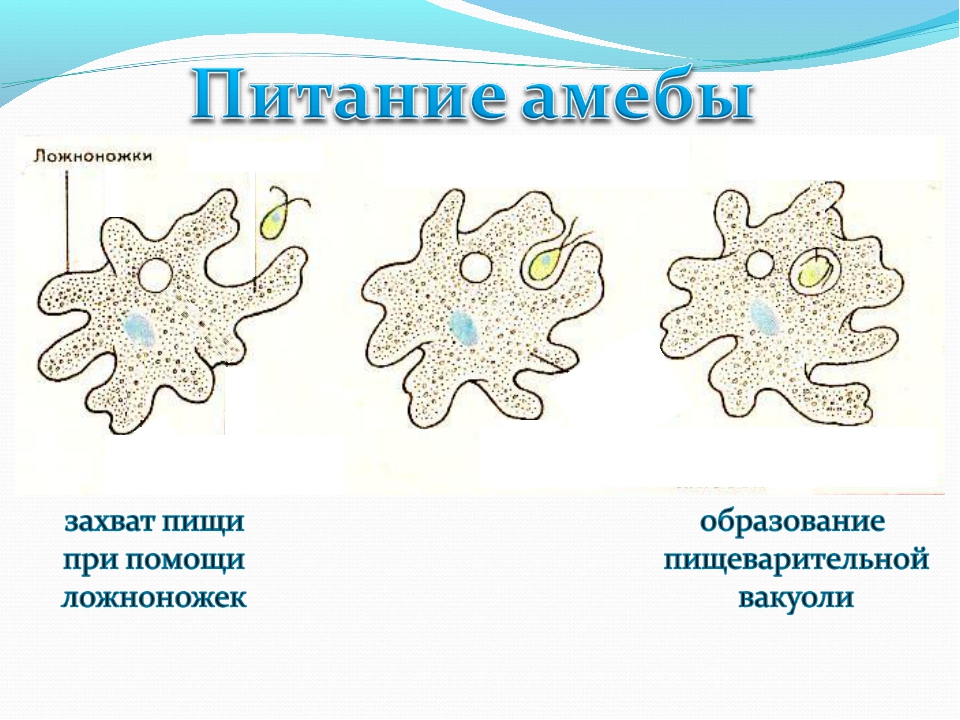
Несмотря на простоту своего строения, амеба обнаруживает все основные свойства, присущие животному организму. При резких толчках, при прибавлении к воде некоторых веществ, например кристалликов поваренной соли, амеба втягивает все свои ложноножки и принимает шарообразную форму, следовательно, амеба обладает раздражимостью. Амебы питаются, обволакивая бактерии, водоросли своими ложноножками. Погруженная в цитоплазму добыча переваривается под действием выделяемого пищеварительного сока (внутриклеточное пищеварение). Пузырьки с пищевыми включениями называют пищеварительными вакуолями 19.
Пузырьки с пищевыми включениями называют пищеварительными вакуолями 19.
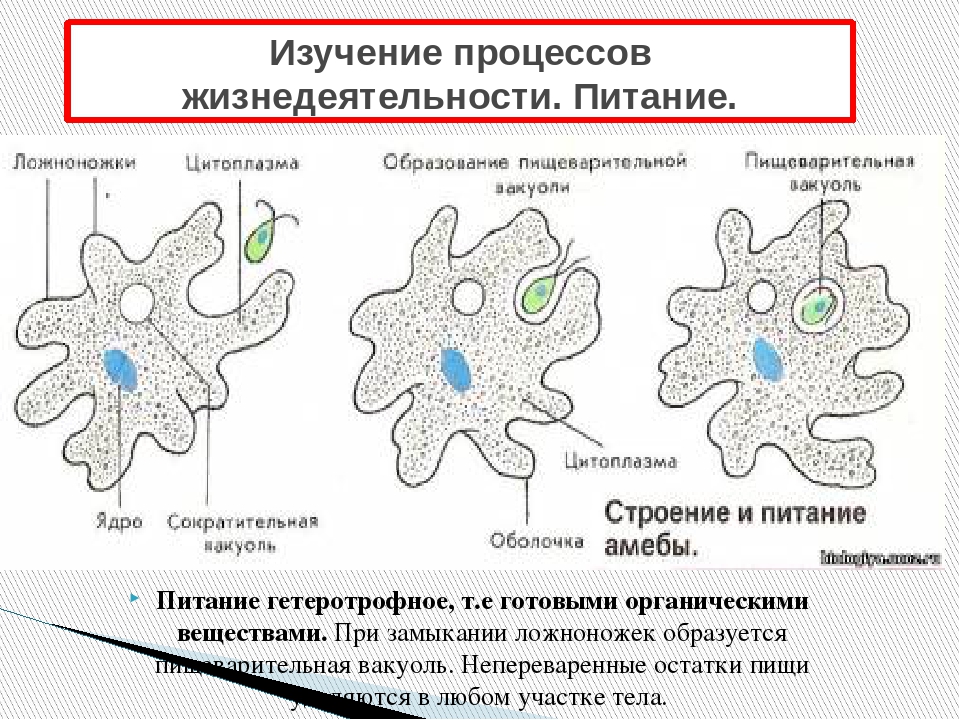
Постоянно поступающая в цитоплазму вода содержит кислород. При участии кислорода происходит расщепление сложных органических веществ, поступивших с пищей, на более простые. При этом выделяется энергия, необходимая для движения и пищеварения, а растворенный в воде углекислый газ и вредные для амебы жидкие продукты распада удаляются наружу через пульсирующий в цитоплазме пузырек — сократительную вакуоль. Происходит обмен веществ, в результате чего тело амебы растет. Размножается амеба делением. Ядро делится надвое, обе половинки его расходятся в стороны, между ними на теле амебы образуется перетяжка, а затем из одной материнской клетки образуются две самостоятельные, дочерние амебы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Просмотров: 1 374
Тематическое и поурочное планирование по биологии. 7 класс 7 класс
http://www.modernlib.ru/books/zaharova_natalya/tematicheskoe_i_pourochnoe_planirovanie_po_biologii_7_klass/read/ К учебнику В.Б. Захарова, Н.И. Сонина «Биология. Многообразие живых организмов. 7 класс» (М.: Дрофа)Методическое пособие Рекомендовано Российской Академией образования
ОТ АВТОРАПредлагаемое пособие предназначено учителю, работающему с учебником В.Б. Захарова и Н.И. Сонина «Биология. Многообразие живых организмов» (7 класс). Оно поможет в планировании уроков, определении их структуры и содержания. Опираясь на собственный опыт, а также учитывая возможности школы (для проведения лабораторных работ, экскурсий, просмотра видеофильмов), учитель может следовать рекомендациям пособия и применять их частично или целиком. Особенно, в отношении внутриурочного распределения времени на разные формы работы (проверка домашних заданий, изложение нового материала и его закрепление).  1 При отсутствии достаточной лабораторной базы соответствующие работы могут быть заменены уроками с более полным изложением разнообразия, образа жизни и значения наиболее известных групп живых организмов (например, покрытосеменных растений, насекомых, амфибий, рептилий, птиц, млекопитающих). 1 При отсутствии достаточной лабораторной базы соответствующие работы могут быть заменены уроками с более полным изложением разнообразия, образа жизни и значения наиболее известных групп живых организмов (например, покрытосеменных растений, насекомых, амфибий, рептилий, птиц, млекопитающих).
Каждую тему предлагается завершать проверочной работой с применением тестов, которые все шире используются в качестве единых государственных экзаменов, а также контрольных вопросов и заданий. В пособие включены некоторые сведения, дополняющие основные положения, раскрываемые в основном тексте учебника. Они приводятся как дополнения или пояснения, а также в виде развернутых ответов на некоторые вопросы, позволяющие несколько расширить представления учащихся об изучаемых биологических объектах. Разумеется, такие вопросы целесообразно задавать классу в качестве методического приема при закреплении изучаемого материала или в случаях, когда учитель уже изложил на предыдущем уроке новые сведения. Выполнение тестовых заданий в проверочных работах оценивается баллами (например, по 10-бальной системе), которые перед выставлением итоговой оценки в журнал и дневники переводятся в принятую школами 4-бальную шкалу. Пособие адресовано учителям и репетиторам.
Достарыңызбен бөлісу: |
 Методический прием авторов учебника, когда в заключительных заданиях требуется определить принадлежность изображаемых организмов к тем или иным систематическим категориям, потребовал подстраховки для учителя в виде соответствующих ответов, приводимых в пособии. Более того, необходимыми сведениями сопровождаются и те рисунки в учебнике, которые также полезно снабдить некоторыми комментариями. Это относится, в первую очередь, к названиям изображаемых организмов, о чем нередко учащиеся спрашивают учителя на уроках.
Методический прием авторов учебника, когда в заключительных заданиях требуется определить принадлежность изображаемых организмов к тем или иным систематическим категориям, потребовал подстраховки для учителя в виде соответствующих ответов, приводимых в пособии. Более того, необходимыми сведениями сопровождаются и те рисунки в учебнике, которые также полезно снабдить некоторыми комментариями. Это относится, в первую очередь, к названиям изображаемых организмов, о чем нередко учащиеся спрашивают учителя на уроках.
Рецензия на фильм Жена астронавта от Misha.naumoff2012 13.01.2018
Долгое время обозрение этого фильма стояло в планах, но на некоторое время выпал из них. В свободное от праздников время захотелось посмотреть чего-нибудь душещипательного с претензией на мурашки по коже. Но как оказалось кино получилось слабоватым, местами посредственным, но на удивление смотрибельным. DVD-издание этого фильма выходило в далёком 2008-ом году, когда фильму было десять лет. Поэтому будут небольшие отсылки в дальнейшем обзоре. Далее впечатления от просмотра и мысли по этому поводу.
Сюжет рассказывает нам о двух героях астронавтах, побывавших в космосе, где чинили космический спутник, но оказалось что НАСА потеряло связь с ними на две минуты и что же происходило в эти две минуты станет основным вопросом фобии главной героини. В пику популярности актёра Джонни Деппа стоило ожидать появления на физических носителях фильмов ранних, менее ярких но с заветным именем звезды в титрах. На этот раз свадебными генералами на своих же свадьбах стали пират-брадобрей Депп в компании с мастерицей перевоплощения Шарлиз Терон. Фильму без малого двадцать лет, но смотрится он астрономически, без каламбуров и преувеличений.
В свободное от праздников время захотелось посмотреть чего-нибудь душещипательного с претензией на мурашки по коже. Но как оказалось кино получилось слабоватым, местами посредственным, но на удивление смотрибельным. DVD-издание этого фильма выходило в далёком 2008-ом году, когда фильму было десять лет. Поэтому будут небольшие отсылки в дальнейшем обзоре. Далее впечатления от просмотра и мысли по этому поводу.
Сюжет рассказывает нам о двух героях астронавтах, побывавших в космосе, где чинили космический спутник, но оказалось что НАСА потеряло связь с ними на две минуты и что же происходило в эти две минуты станет основным вопросом фобии главной героини. В пику популярности актёра Джонни Деппа стоило ожидать появления на физических носителях фильмов ранних, менее ярких но с заветным именем звезды в титрах. На этот раз свадебными генералами на своих же свадьбах стали пират-брадобрей Депп в компании с мастерицей перевоплощения Шарлиз Терон. Фильму без малого двадцать лет, но смотрится он астрономически, без каламбуров и преувеличений.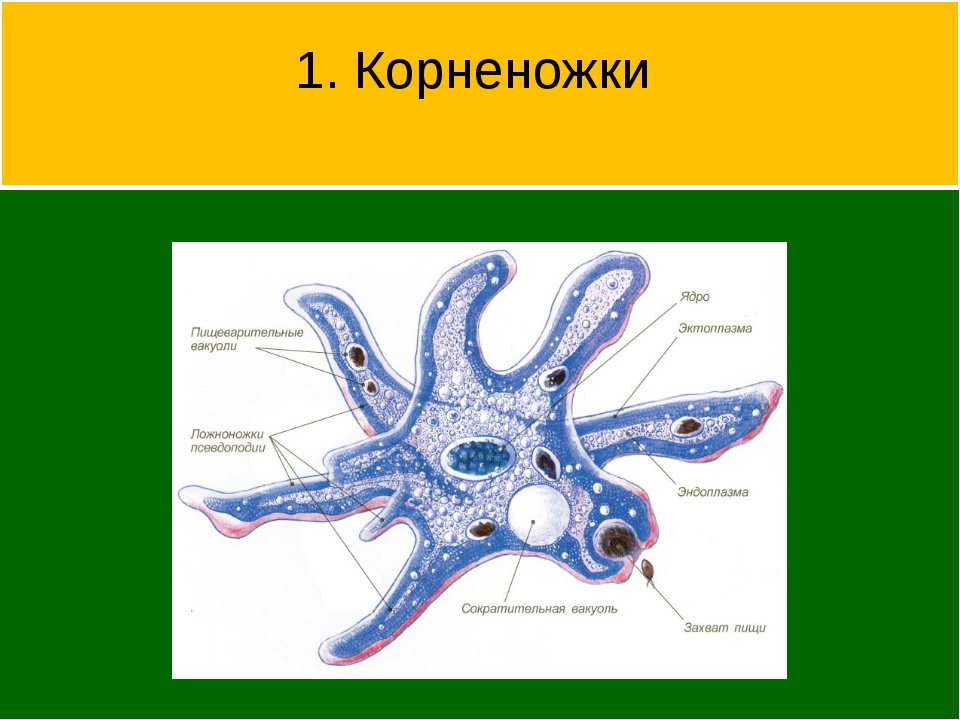

Амёбы / Зоология для учителя
По сложности своего строения (в пределах одноклеточной организации) инфузории представляют собой высший класс среди других простейших, и, казалось бы, не с них, а с форм действительно наиболее простого строения следовало бы начать ознакомление с типом простейших, как это и делается во всех вузовских учебниках.
Однако по соображениям методического порядка автору этой книги представляется более целесообразным начать изучение простейших именно с инфузорий: живой материал по ним более доступен для непосредственных наблюдений, на нем легче приобрести навыки изучения «мира в капле воды» и вместе с тем получить и общее ориентировочное представление о внешности и размерах существ из этого мира, скрытого от наших глаз в повседневной жизни.
И уже на этой основе от монографического изучения инфузории-туфельки удобнее перейти к амёбе — животному организму, действительно наиболее простому по своему строению (правда, далеко не все методисты-биологи разделяют такую точку зрения, считая, что и в школе изучение животного мира должно начинаться с амёбы).
По сравнению с инфузориями тело амёбы имеет гораздо более простое строение (рис. 35). Оно состоит из комочка полужидкой, тягучей протоплазмы, внутри которой заключено одно круглое ядро. Протоплазма на поверхности не выделяет оболочки, и потому амёба не имеет постоянной формы. То на одном, то на другом месте её тела вытягиваются выросты протоплазмы — так называемые ложноножки. Постепенно в вытянувшуюся ложноножку переходит все больше и больше протоплазмы, а ложноножки с противоположной стороны сокращаются; в результате амёба медленно переползает с места на место.
«Поймать» амёбу можно следующим способом. На поверхность культуры, в которой могут быть амёбы, осторожно кладут покровное стёклышко (благодаря поверхностному натяжению жидкости и своему лёгкому весу оно не потонет).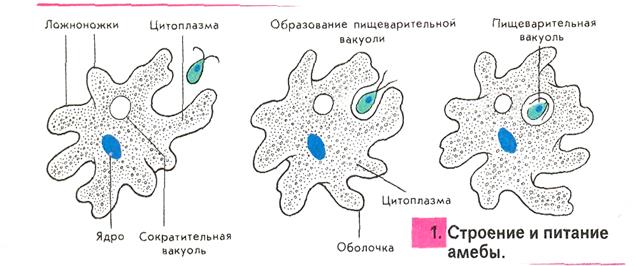 Через сутки покровное стёклышко снимают и переносят на предметное стекло, покрыв им капельку воды, взятой с поверхностного слоя той же культуры.
Через сутки покровное стёклышко снимают и переносят на предметное стекло, покрыв им капельку воды, взятой с поверхностного слоя той же культуры.
Рассматривают препарат под микроскопом, не отвлекаясь наблюдениями над инфузориями и другими существами. Поиски амёб требуют большого внимания: вначале потревоженные амёбы имеют вид округлых комочков и только некоторое время спустя начинают менять свою форму и выпускают ложноножки. Название «амёбы» относится не к одному зоологическому виду, а к целому отряду, заключающему в себе ряд отдельных родов и видов, различающихся между собой и по размерам, и по характеру очертаний ложноножек, и по образу жизни.
Несмотря на простоту своего строения, амёбы обнаруживают все основные свойства, присущие животному организму. При резких толчках, при прибавлении к воде некоторых веществ амёба втягивает все свои ложноножки и принимает шарообразную форму; следовательно, амёба обладает раздражимостью: она отвечает определённым образом на изменения внешних условий, на механические и химические раздражения.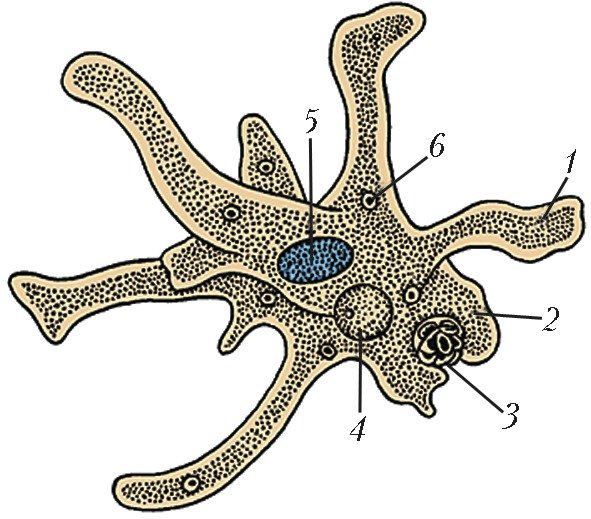 Амёба, как уже было сказано, передвигается при помощи своих ложноножек.
Амёба, как уже было сказано, передвигается при помощи своих ложноножек.
Далее, амёбы питаются: прикасаясь к какой-нибудь бактерии или мелкой водоросли, она обволакивает её своими ложноножками: погруженная в протоплазму добыча переваривается, её питательный материал усваивается амёбой, то есть переходит в состав её протоплазмы, а непереваренные остатки выбрасываются вон. Внутри тела появляется светлый водянистый пузырёк, который вскоре исчезает, выбрасывая жидкость наружу, а затем образуется снова; это сократительная вакуоля, через которую происходит выделение излишней воды и продуктов распада протоплазмы.
В кипячёной воде, лишённой кислорода, амёба погибает; следовательно, кислород необходим ей для дыхания.
В результате совершающегося обмена веществ тело амёбы растёт. Достигнув своей предельной величины, амёба размножается путём деления: делится ядро, обе половинки его расходятся в стороны, между ними на теле образуется перетяжка, и, наконец, из одного материнского организма образуются две самостоятельные, дочерние амёбы.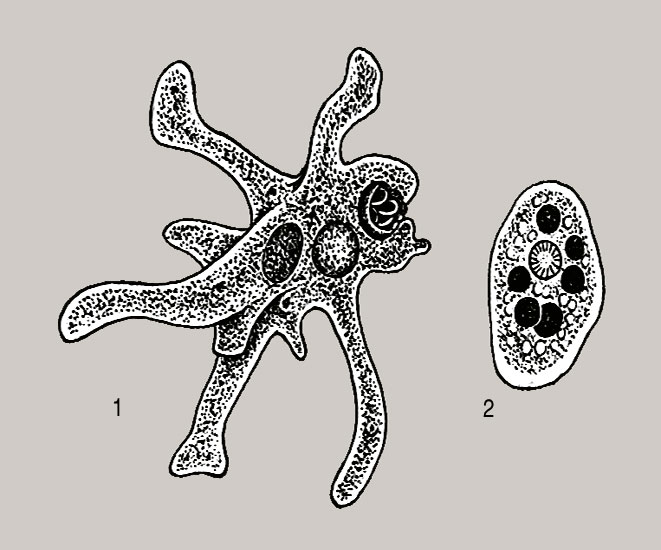
При наступлении неблагоприятных условий амёба выделяет на поверхности тела цисту и впадает в состояние скрытой жизни. В таком виде амёба напоминает спору грибов или других бесцветковых растений.
Тренировочный вариант по русскому языку №6 ЕГЭ
ПОДГОТОВКА К ЕГЭ по русскому языку
ТРЕНИРОВОЧНЫЙ ВАРИАНТ №6 — 2021 год.
Прочитайте текст.
(1)У амёбы протей тело представляет собой одну клетку до 0,25 мм длиной. (2) Клетка перемещается, так как ее форма постоянно меняется. (3)Во время движения на передней стороне клетки возникают выросты, в которые постепенно перетекает цитоплазма из центральной и задней части клетки. 4)(…) выросты называются ложноножками. (5)Когда на пути амёбы возникает скопление бактерий или остатки других организмов, ложноножки замыкают их в себе и начинается процесс пищеварения у амёбы протей.
Задание 1
Укажите варианты
ответов, в которых верно передана ГЛАВНАЯ информация, содержащаяся в тексте. Запишите номера этих предложений.
Запишите номера этих предложений.
1) Во время движения на передней стороне клетки возникают выросты, в которые постепенно перетекает цитоплазма из центральной и задней части клетки, они называются ложноножками.
2)Тело амебы протей – всего одна клетка, которая, изменяя форму, передвигается, а благодаря выростам-ложноножкам, (куда перетекает цитоплазма), у нее происходит процесс пищеварения в местах скопления бактерий.
3)Изменяя свою форму, одноклеточная амеба протей перемещается, а на передней части клетки возникают выросты-ложноножки, которые организуют процесс пищеварения в местах скопления бактерий.
4)Благодаря выростам, у амебы происходит процесс пищеварения.
5)Тело амебы представляет собой одну клетку до 0,25 мм длиной, которая перемещается и питается.
Задание 2.
Самостоятельно
подберите указательное местоимение, которое должно стоять на месте пропуска в
четвертом (4) предложении текста. Запишите это местоимение.
Запишите это местоимение.
Задание 3
Прочитайте фрагмент словарной статьи, в которой приводятся значения слова ФОРМА. Определите значение, в котором это слово употреблено в втором (2) предложении текста. Выпишите цифру, соответствующую этому значению в приведённом фрагменте словарной статьи.
ФО́РМА, -ы, ж.
1. Внешние очертания, наружный вид предмета. Листья перистой формы. [Сталь], кипящую, зачерпнули из печи железной ложкой, и, застыв, сталь сохранила форму ложки.
2. мн. ч. (фо́рмы, форм). Очертания человеческого тела, фигуры. На креслах сидел молодой человек лет 28. Все формы его выражали атлетическую силу тела.
3. Внешнее выражение чего-л., обусловленное определенным содержанием, сущностью; тип, устройство, способ организации чего-л. Форма государственного правления. Формы классовой борьбы.
Задание 4
В одном из приведённых
ниже слов допущена ошибка в постановке ударения: НЕВЕРНО выделена буква,
обозначающая ударный гласный звук. Выпишите это слово.
Выпишите это слово.
недУг
придАв
обогнАла
назвалА
кОгтя
Задание 5
В одном из приведённых ниже предложений НЕВЕРНО употреблено выделенное слово. Исправьте лексическую ошибку, подобрав к выделенному слову пароним. Запишите подобранное слово.
В образе героя писатель ПРЕДСТАВИЛ поместное дворянство, ведущее разгульный образ жизни.
В академии были выставлены портреты выдающихся отечественных учёных, прошедших достойный жизненный путь и являющих собой ГОРДОСТЬ мировой науки.
Подруги знали о стремлении Даши добиться спортивных побед, поэтому преподнесли ей на день рождения ГОДОВАЛЫЙ абонемент в бассейн.
Легендарный американский боксёр по-прежнему считается одним из самых ТЕХНИЧНЫХ тяжеловесов.
В условиях мирового кризиса следует повышать ЭФФЕКТИВНОСТЬ производства.
Задание 6
Отредактируйте
предложение: исправьте лексическую ошибку, исключив лишнее слово.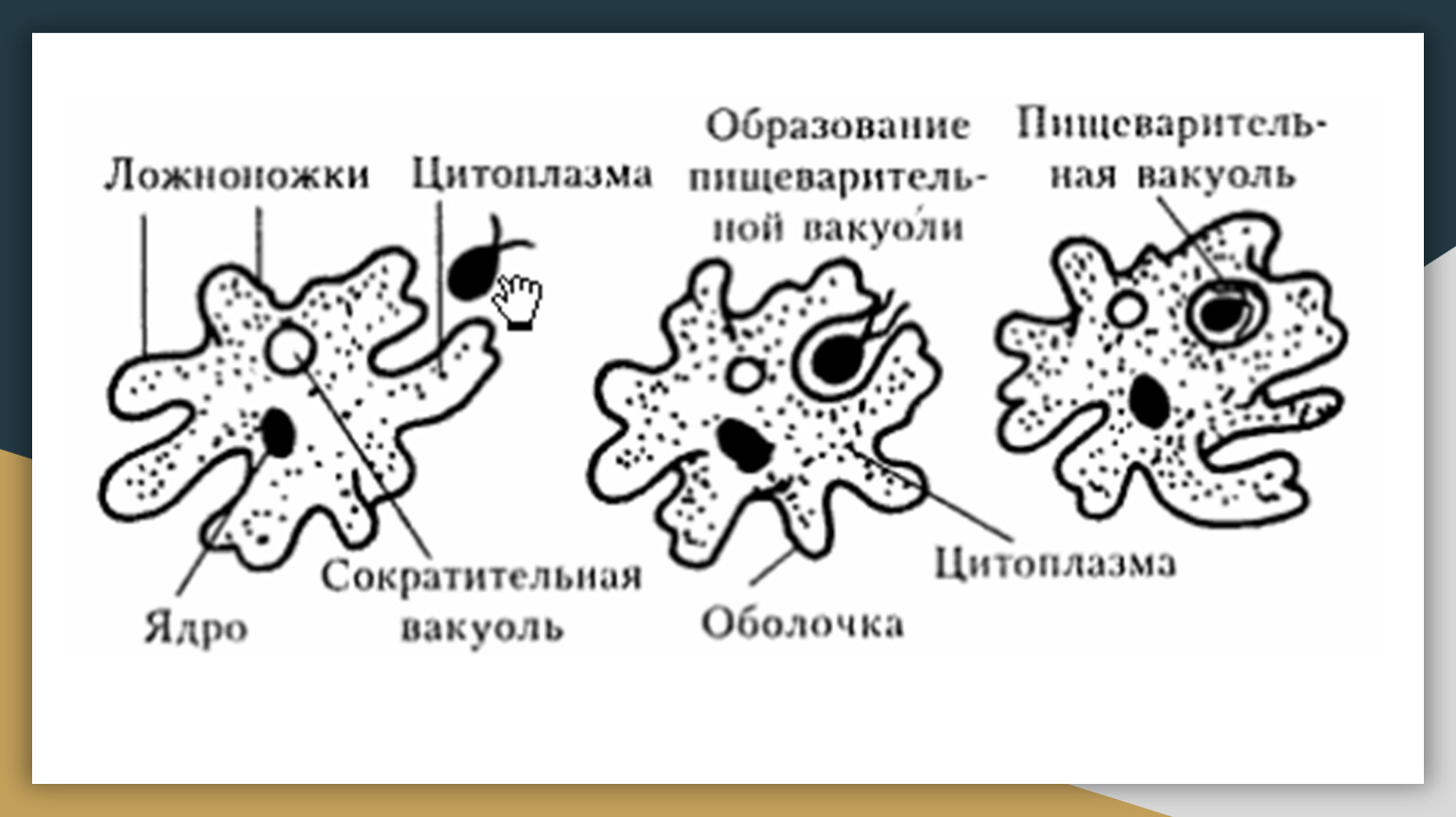 Выпишите это слово.
Выпишите это слово.
Первая игра стартовавшего в Дании чемпионата мира по плаванию не принесла никаких неожиданных сюрпризов.
Задание 7
В одном из выделенных ниже слов допущена ошибка в образовании формы слова. Исправьте ошибку и запишите слово правильно.
нет ПОЛОТЕНЕЦ
мягких КРЕСЕЛ
ШЕСТИСТАМИ жителями
без САПОГ
ТРЕМСТАМ пятидесяти рублям
Задание 8
Установите соответствие между грамматическими ошибками и предложениями, в которых они допущены: к каждой позиции первого столбца подберите соответствующую позицию из второго столбца.
ГРАММАТИЧЕСКИЕ ОШИБКИ |
| |
А) нарушение связи между подлежащим и сказуемым Б) нарушение в построении предложения с несогласованным приложением В) нарушение в построении предложения с деепричастным оборотом Г) неправильное употребление падежной формы имени существительного с предлогом Д) нарушение в построении предложения с причастным оборотом
ПРЕДЛОЖЕНИЯ 1) Город Сочи за последние годы сильно изменился. 2) Охотник пошёл напрямик и попал в общество крестьянских мальчишек, которые в поле стерегли коней. 3) По приезду домой молодой офицер хотел заняться делами, но отец был не в состоянии дать ему нужных объяснений. 4) С юных лет Мясоедов уже входил в круг художников, боровшимся с рутиной академизма. 5) Возвращаясь недавно в родные места, мне вспомнились детские страхи от впервые увиденной грозы. 6) В замечательном рассказе И. С. Тургенева «Бежином луге» повествуется об интересном случае на охоте, произошедшем с самим пистелем. 7)Обречённый судьбой на постоянную праздность, я не делал решительно ничего. 8) В просторной комнате с итальянским окном в течение двенадцати лет Н. М. Карамзин работал над «Историей государства Российского». 9) Многие из тех, кто посетил выставку, принял участие в журналистском опросе. |
|

Задание 9
Укажите варианты
ответов, в которых во всех словах одного ряда содержится безударная
проверяемая гласная корня. Запишите номера ответов.
Запишите номера ответов.
1) наполнитель, увековечить, восхвалять
2) возрастной, невразумительно, творец
3) оснащение, озаряться, располагающийся
4) формализм, победить, подражание
5) издавать, отгадать, примерять (сапоги)
Задание 10
Укажите варианты ответов, в которых во всех словах одного ряда пропущена одна и та же буква. Запишите номера ответов.
1) пр..ковылять, пр..морский, пр..быть (на станцию)
2) о..цепленный (вагон), о..гадать, на..кусить
3) ра..сеять, в..пахивать, бе..голосый
4) без..нтересный, меж..гровой, с..мпровизировать
5) беспр..будный, нед..варить, п..завчера
Задание 11
Укажите варианты ответов, в которых в обоих словах одного ряда пропущена одна и та же буква. Запишите номера ответов.
1) алюмини..вый, овлад..вать
2) (завернуть) налев.., завед..вать
3) выстра..вающий, чист..нький
4) танц..вать,
син.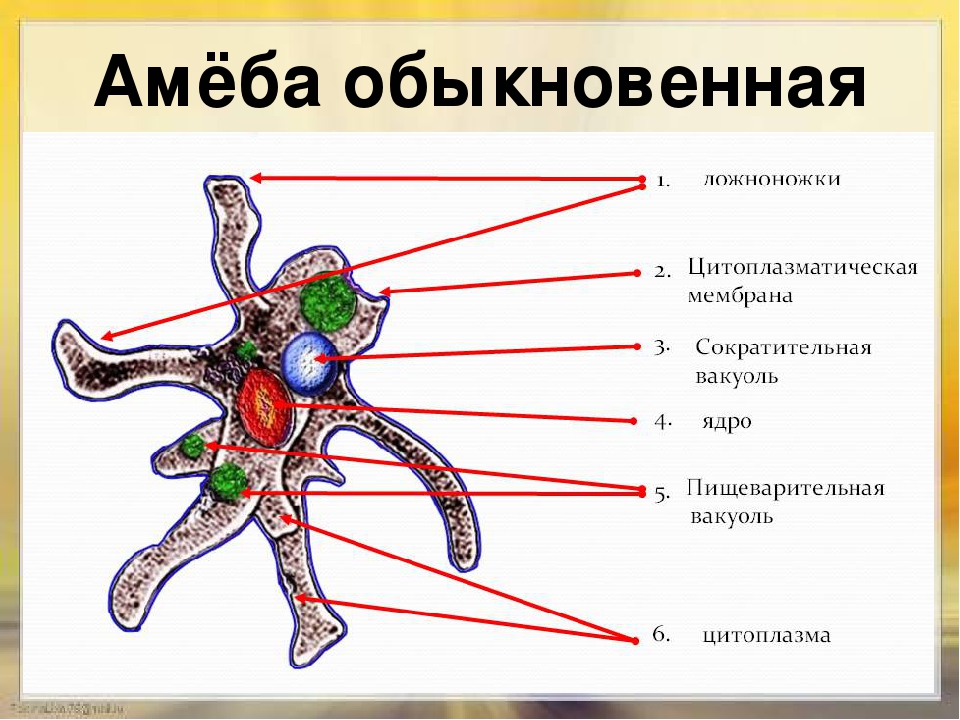 .ватый
.ватый
5) въедл..вый, цел..вой
Задание 12
Укажите варианты ответов, в которых в обоих словах одного ряда пропущена одна и та же буква. Запишите номера ответов.
1) мелька..шь, закле..вший
2) жмур..щийся (от солнца), (друзья) увид..тся
3) прилад..шь, скле..нный
4) подстав..шь, независ..мый
5) онеме..шь, поджида..мый
Задание 13
Определите предложение, в котором НЕ со словом пишется СЛИТНО. Раскройте скобки и выпишите это слово.
(НЕ)ЗАМЕТНАЯ окраска стала ярче, и древние письмена теперь видны отчётливо.
(НЕ)ВЕРЯЩИЙ в чудеса маг хитро улыбался.
Мы (НЕ)ДОЛЖНЫ легкомысленно относиться к огромному богатству, которым владеем в наших городах.
Недолгое знакомство нисколько (НЕ)МЕШАЛО нам разговаривать по-дружески.
Ни разу ещё мерцание звёзд (НЕ)КАЗАЛОСЬ таким золотым и чистым.
Задание 14
Определите предложение,
в котором оба выделенных слова пишутся СЛИТНО. Раскройте скобки и выпишите эти
два слова.
Раскройте скобки и выпишите эти
два слова.
Думы о (ЗА)ГРАНИЦЕ, о необходимости покупать чемоданы, билеты, выбирать маршрут ввели Илью Обломова, (В)ОТЛИЧИЕ от его друга Андрея Штольца, в состояние, близкое к паническому.
(НА)КОНЕЦ солнце стало сильно пригревать, большие радужные капли начали падать с крыш, покрытых снегом, (КАК)БУДТО потекли весенние слёзы.
(ПО)НАЧАЛУ первого романа трудно было судить о таланте автора, однако (ЗА)ТЕМ читатели оценили его книги по достоинству..
(В)СЛЕД за тем главные ворота завода распахнулись, и (ОТ)ТУДА выкатилось семь самосвалов.
Беда не произойдёт, если, имея (В)ВИДУ климатические изменения, (ВО)ВРЕМЯ принять меры.
Задание 15
Укажите цифру(-ы), на месте которой(-ых) пишется НН.
Писатели позднего классицизма были убежде(1)ы, что искусство предназначе(2)о для глуби(3)ого постижения человека и мироздания.
Задание 16
Расставьте знаки
препинания. Укажите два предложения, в которых нужно поставить ОДНУ запятую.
Запишите номера этих предложений.
Укажите два предложения, в которых нужно поставить ОДНУ запятую.
Запишите номера этих предложений.
1) И по богатству художественных приёмов и по идейной значимости и по разнообразию воссозданных социальных типов «Сказки» М. Е. Салтыкова-Щедрина в полной мере могут считаться одной из вершин творчества писателя.
2) Очень многие акварели того времени не имели авторов или приписывались тем или иным художникам предположительно.
3) У А.Вознесенского в стихах о любви звучат либо настроения трагизма либо ликующие ноты счастья.
4) Для защиты истины капитану не раз приходилось скрывать свои настоящие взгляды и тогда он вынужден был играть роль убеждённого сторонника академика.
5)Это печальная и трагическая участь человека бездуховного.
Задание 17
Расставьте знаки препинания:укажите цифру(-ы), на месте которой(-ых) в предложении должна(-ы) стоять запятая(-ые).
Вечером облака (1)
отражённые в воде (2) легко можно спутать с розовеющими (3) островами (4)
похожими на материки.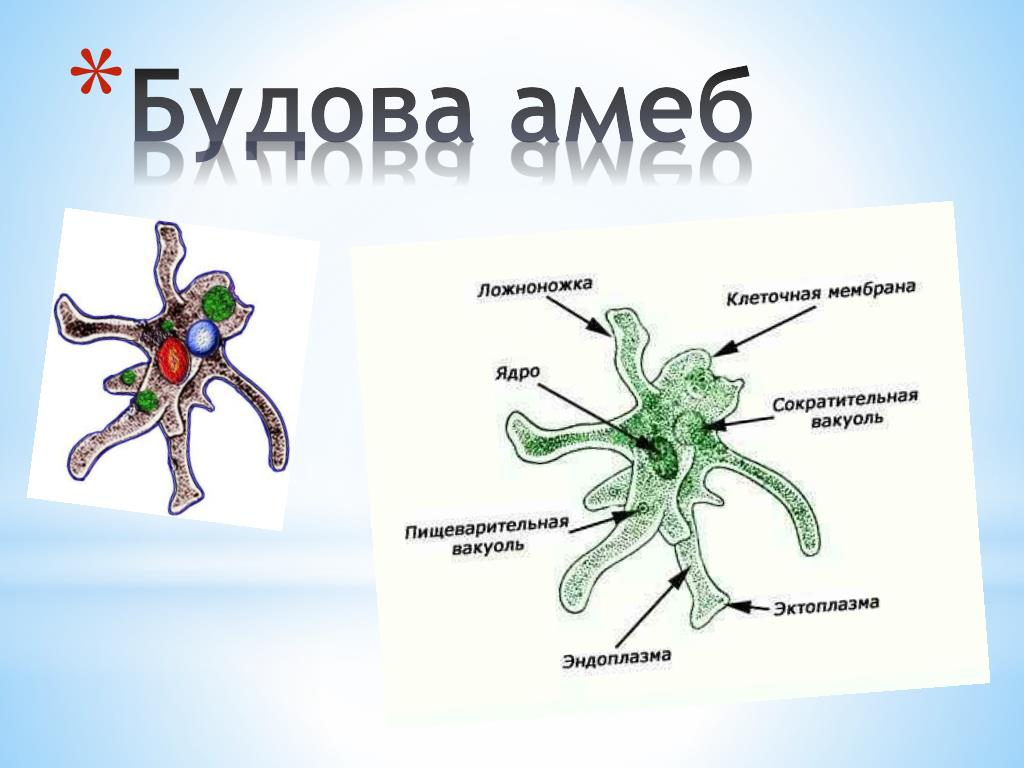
Задание 18
Расставьте знаки препинания: укажите все цифры, на месте которых в предложениях должны стоять запятые.
Третьяковская галерея (1) как известно (2) названа в честь Павла Михайловича Третьякова — купца, употребившего своё богатство на пользу общества. Увлечённый живописью, он положил начало (3) поистине (4) замечательному собранию картин.
Задание 19
Расставьте знаки препинания: укажите цифру(-ы), на месте которой(-ых) в предложении должна(-ы) стоять запятая(-ые).
Загадочное «огненное кольцо» (1) очертания (2) которого идут вдоль группы сейсмических поясов (3) представляет интерес для различных специалистов, занимающихся изучением Тихого океана.
Задание 20
Расставьте знаки препинания: укажите цифру(-ы), на месте которой(-ых) в предложении должна(-ы) стоять запятая(-ые).
Долго читать было
невозможно (1) и (2) когда не спалось (3) я слушал окружающие ночные звуки и
пытался представить (4) что скрывается за ними.
Задание 21
Найдите предложения, в которых тире ставится в соответствии с одним и тем же правилом пунктуации. Запишите номера этих предложений.
(1)У всех есть место,
которое нам бесконечно дорого, — родная земля. (2)Алтайский край является
одним из самых красивых мест земного шара. (3)Тысячи бурных рек и спокойных
прозрачных озёр, необъятный простор степей и цветущие ковры горных лугов,
светлые берёзовые рощи и таинственные урочища со следами диких животных —
всё это мило и дорого каждому, кто здесь бывал. (4)Этот край — территория
невероятных сокровищ, таящихся в глухих недрах. (5)Но Алтайский край знаменит
не только первозданной природой, главное его богатство — люди. (6)Люди спокойные
и смелые, уважающие друг друга, любящие жизнь и свою родную землю, уверенно
смотрящие в будущее. (7)Чтобы познакомиться с Алтайским краем, можно почитать о
нём книги, посмотреть фильмы, но, чтобы постичь его душой, а затем полюбить
навсегда, надо обязательно приехать на эту землю. (8)«Добро пожаловать в
удивительный Алтайский край!» — призывают к увлекательному путешествию
путеводители по туристическим маршрутам России.
(8)«Добро пожаловать в
удивительный Алтайский край!» — призывают к увлекательному путешествию
путеводители по туристическим маршрутам России.
Прочитайте текст и выполните задания 22 — 26.
(1) Это было на фронте. (2) Кормили плохо, вечно хотелось есть. (3) Иногда пищу давали раз в сутки, и то вечером. (4) Ах, как хотелось есть! (5) И вот в один из таких дней, когда уже приближались сумерки, а во рту ещё маковой росинки не было, мы, человек восемь бойцов, сидели на невысоком травянистом берегу тихонькой речушки и чуть не скулили.
(6) Вдруг видим: без гимнастёрки, что-то держа в руках, к нам бежит ещё один наш товарищ.(7) Подбежал. (8) Лицо сияющее. (9) Свёрток – это гимнастёрка, а в неё что-то завёрнуто.(10) Смотрите! – победителем восклицает Борис. (11) Разворачивает гимнастерку, и в ней живая дикая утка.
(12) — Вижу: сидит, притаилась за
кустиком! (13) Я рубаху снял и хоп! (14) Есть еда! (15) Зажарим? (16) Утка была
некрупная, молодая.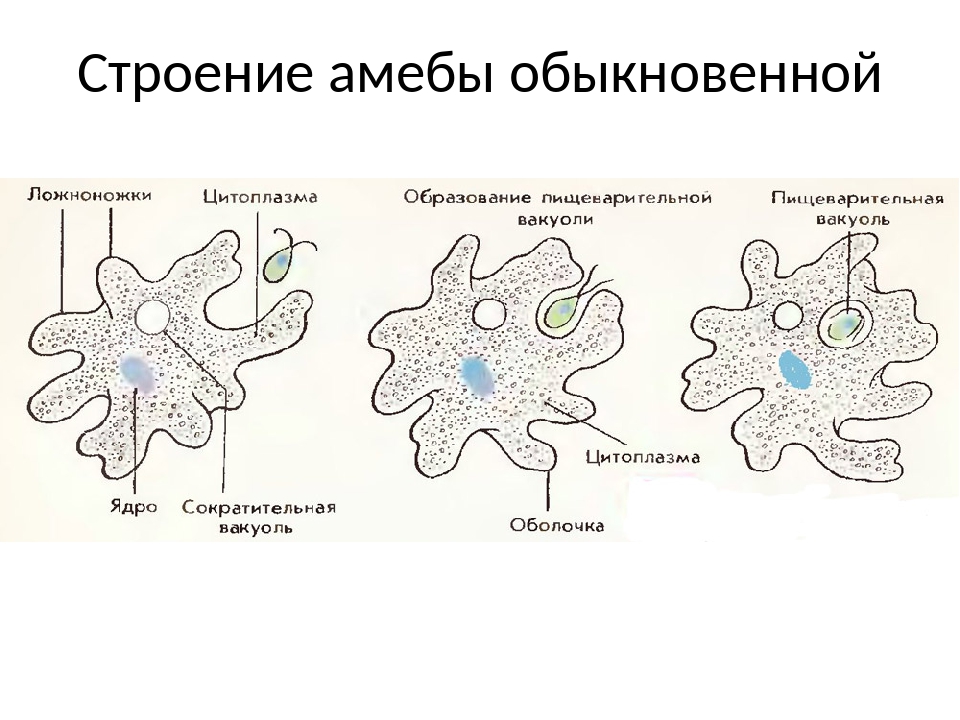 (17) Поворачивая голову по сторонам, она смотрела на нас
изумлёнными бусинками глаз. (18) Она даже не была напугана, для этого она была
ещё просто молода. (19) Она не могла понять, что это за странные милые существа
её окружают и смотрят на неё с таким восхищением. (20) Она не вырывалась, не
крякала, не вытягивала натужно шею, чтобы выскользнуть из державших её рук.
(21) Она грациозно и с любопытством озиралась. (22)Красавица уточка! (23)А мы –
грубые, пропылённые, нечисто выбритые, голодные. (24)Все залюбовались
красавицей.
(17) Поворачивая голову по сторонам, она смотрела на нас
изумлёнными бусинками глаз. (18) Она даже не была напугана, для этого она была
ещё просто молода. (19) Она не могла понять, что это за странные милые существа
её окружают и смотрят на неё с таким восхищением. (20) Она не вырывалась, не
крякала, не вытягивала натужно шею, чтобы выскользнуть из державших её рук.
(21) Она грациозно и с любопытством озиралась. (22)Красавица уточка! (23)А мы –
грубые, пропылённые, нечисто выбритые, голодные. (24)Все залюбовались
красавицей.
(25) И произошло чудо, как в доброй
сказке. (26) Кто-то просто произнёс: «(27) Отпустим!» (28)Было брошено
несколько логических реплик, вроде: «(29) Всё равно толку не добьёмся, нас
восемь человек, а она такая маленькая. (30) Ещё возиться! – (31) Подождём,
приедет же этот зараза повар со своей походной кухней-таратайкой! (32)Боря,
неси её обратно.»(33) И, уже ничем не покрывая, Борис бережно понёс утку
обратно. (34)Вернувшись, сказал:
– (35) Я её в воду пустил.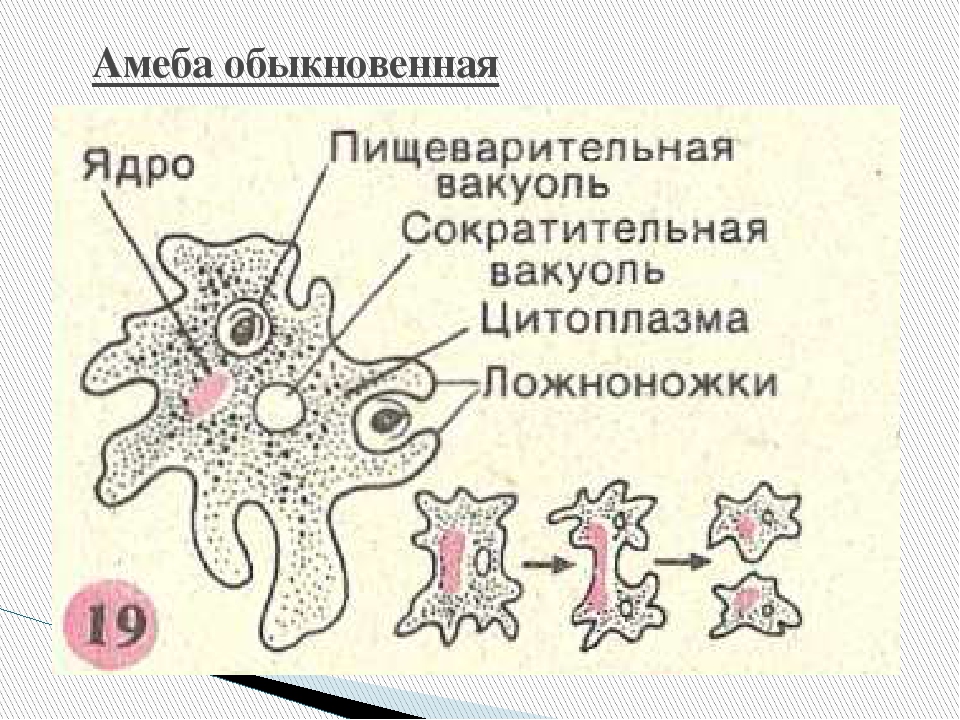 (36) Нырнула. (37) А где вынырнула, не видел. (38)
Ждал-ждал, чтоб посмотреть, но не увидел. (39) Уже темнеет.
(36) Нырнула. (37) А где вынырнула, не видел. (38)
Ждал-ждал, чтоб посмотреть, но не увидел. (39) Уже темнеет.
(40) Когда меня заматывает жизнь, когда начинаешь клясть всё и всех, теряешь веру в людей, мне хочется изо всех сил крикнуть, как однажды я услыхал вопль одного очень известного человека: «Я не хочу быть с людьми, я хочу быть с собаками!» – вот в эти минуты неверия и отчаяния я вспоминаю дикую утку и думаю: нет-нет, в людей можно верить. (41) Они способны к состраданию и великодушию.
(42) Мне могут сказать: (43) «Ну да, это были вы, интеллигенты, артисты, от вас всего можно ожидать». (44) Нет, на войне всё перемешалось и превратилось в одно целое – единое и неделимое.(45) Во всяком случае, там, где служил я. (46) Были в нашей группе и два вора, только что выпущенных из тюрьмы. (47) Один с гордостью красочно рассказывал, как ему удалось украсть подъёмный кран. (48) Видимо, был талантлив. (49) Но и он сказал: «Отпустить!»
(В.Розов).
Задание 22
Какие из высказываний
соответствуют содержанию текста? Укажите номера ответов.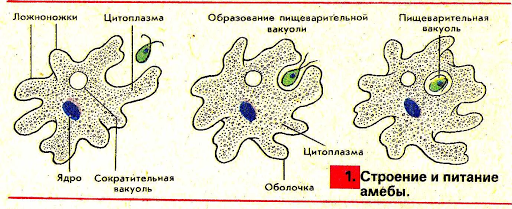
1) Один вор с гордостью предлагал украсть подъемный кран.
2) И произошло чудо: кто-то просто произнес: «Отпустим!»
3) Голодные бойцы любовались красавицей уткой.
4) На фронте рассказчика кормили плохо: вечно хотелось есть.
5) Борис бережно понес утку повару из походной кухни.
Задание 23
Какие из перечисленных утверждений являются верными? Укажите номера ответов.
1) Предложения 6,7 содержат повествование.
2) В предложении 40 содержится рассуждение.
3) Предложения 44, 45 содержат повествование.
4) В предложении 16 представлен ответ на вопрос, сформулированный в предложении 15.
5) В предложениях 1-3 представлено рассуждение.
Задание 24
Из предложений 12 – 14 выпишите разговорное слово.
Задание 25
Среди предложений 1-5
найдите такое(-ие), которое(-ые) связано(-ы) с предыдущим с помощью сочинительного
союза и указательной частицы. Напишите номер(-а) этого(-их)
предложения(-ий).
Напишите номер(-а) этого(-их)
предложения(-ий).
Задание 26
«В. Розов — мастер повествования. Троп (А)_____ («милые существа» в предложении 19) помогает писателю передать удивление дикой красавицы. Сложность чувств голодных бойцов к уточке подчёркивают синтаксический приём — (Б)_____ ( в предложениях 20, 23) и лексическое средство — (В)_____ (предложение 5)», а также синтаксическое средство — (Г)_____(в предложениях 12 -14).
Список терминов:
Тип простейшие. Классификация, образ жизни, особенности строения, размножения и развития учебное пособие по биологии
Тип простейшие. Классификация, образ жизни, особенности строения, размножения и развития 1% одноклеточным относятся животные, тело которых морфологически
источнику раздражения, другие – отрицательную, т.е. движение от источника раздражения. Эти направленные движения свободноживущих организмов, не имеющих нервных аппаратов, называются таксисами. Большинство простейших может медленно или быстро передвигаться. Способы передвижения в разных классах этого типа различны. Наиболее распространены способы передвижения с помощью ложноножек, или псевдоподий, жгутиков и ресничек. Ложноножки – это временные выпячивания цитоплазмы, форма и длина которых различны и характерны для каждого вида или группы видов. Выпуская ложноножки, животное медленно ползет, при этом форма его тела все время меняется. Жгутики – это длинные, тонкие выросты наружного слоя цитоплазмы, которые описывают при своем движении конус. В каждом жгутике внутри имеется 11 продольных волоконец (фибрилл), из которых 2 центральных и 9 периферических. Внутренние волоконца выполняют опорную функцию, а наружные – двигательную. Жгутики начинаются от особых образований – кинетосом. Процессы обмена веществ, происходящие в кинетосомах, являются источником энергии, необходимой для движения жгутиков. Реснички по строению очень сходны со жгутиками, но в отличие от последних короткие и работают наподобие весел; число их обычно велико.
Большинство простейших может медленно или быстро передвигаться. Способы передвижения в разных классах этого типа различны. Наиболее распространены способы передвижения с помощью ложноножек, или псевдоподий, жгутиков и ресничек. Ложноножки – это временные выпячивания цитоплазмы, форма и длина которых различны и характерны для каждого вида или группы видов. Выпуская ложноножки, животное медленно ползет, при этом форма его тела все время меняется. Жгутики – это длинные, тонкие выросты наружного слоя цитоплазмы, которые описывают при своем движении конус. В каждом жгутике внутри имеется 11 продольных волоконец (фибрилл), из которых 2 центральных и 9 периферических. Внутренние волоконца выполняют опорную функцию, а наружные – двигательную. Жгутики начинаются от особых образований – кинетосом. Процессы обмена веществ, происходящие в кинетосомах, являются источником энергии, необходимой для движения жгутиков. Реснички по строению очень сходны со жгутиками, но в отличие от последних короткие и работают наподобие весел; число их обычно велико. Известны и другие способы передвижения простейших: в результате сокращения особых волоконец (мионем), путем выделения жидкости из задней части тела (реактивный способ движения) и др. Продукты диссимиляции выделяются у простейших через всю поверхность тела, а у многих и через сократительные, или пульсирующие, вакуоли. Эти органоиды представляют собой светлые, прозрачные пузырьки, периодически наполняющиеся жидкостью до определенного предела, затем спадающиеся при выделении ее наружу. Главная функция сократительных вакуолей – удаление избыточной воды из клетки. Простейшие гетеротрофы. Только в классе жгутиковых, кроме многочисленных гетеротрофных видов, имеется значительное количество автотрофов. Они всасывают из окружающей их воды простые неорганические соединения (углекислый газ и соли, растворенные в воде), которые в пластидах, содержащих хлорофилл, превращаются в сложные органические вещества. У свободноживущих гетеротрофных простейших пища (мелкие организмы, остатки растений и животных) попадает в клетку вместе с водой и в цитоплазме образуются пищеварительные вакуоли, в которых сложные нерастворимые органические вещества разлагаются на более простые растворимые органические вещества.
Известны и другие способы передвижения простейших: в результате сокращения особых волоконец (мионем), путем выделения жидкости из задней части тела (реактивный способ движения) и др. Продукты диссимиляции выделяются у простейших через всю поверхность тела, а у многих и через сократительные, или пульсирующие, вакуоли. Эти органоиды представляют собой светлые, прозрачные пузырьки, периодически наполняющиеся жидкостью до определенного предела, затем спадающиеся при выделении ее наружу. Главная функция сократительных вакуолей – удаление избыточной воды из клетки. Простейшие гетеротрофы. Только в классе жгутиковых, кроме многочисленных гетеротрофных видов, имеется значительное количество автотрофов. Они всасывают из окружающей их воды простые неорганические соединения (углекислый газ и соли, растворенные в воде), которые в пластидах, содержащих хлорофилл, превращаются в сложные органические вещества. У свободноживущих гетеротрофных простейших пища (мелкие организмы, остатки растений и животных) попадает в клетку вместе с водой и в цитоплазме образуются пищеварительные вакуоли, в которых сложные нерастворимые органические вещества разлагаются на более простые растворимые органические вещества. Переваренные вещества диффундируют из вакуолей во все части клетки, где происходит их ассимиляция, непереварившиеся остатки пищи удаляются наружу, вакуоли после окончания процессов пищеварения исчезают. Большинство паразитических простейших, обитающих в тех частях тела хозяев, где имеются растворимые органические вещества, всасывают эти вещества всей поверхностью тела. У простейших наиболее распространено бесполое размножение, которое может проявляться в виде простого деления, почкования и множественного деления. Деление обычно наблюдается у свободноживущих форм: организм делится на две приблизительно равные клетки. Сначала происходит деление ядра, а потом – цитоплазмы. Реже встречается у простейших бесполое размножение в виде почкования. У большинства паразитических простейших, а также у ряда свободноживущих форм распространено множественное деление, или шизогония, при котором материнская особь распадается на большое количество дочерних. Во всех классах простейших наблюдается половое размножение.
Переваренные вещества диффундируют из вакуолей во все части клетки, где происходит их ассимиляция, непереварившиеся остатки пищи удаляются наружу, вакуоли после окончания процессов пищеварения исчезают. Большинство паразитических простейших, обитающих в тех частях тела хозяев, где имеются растворимые органические вещества, всасывают эти вещества всей поверхностью тела. У простейших наиболее распространено бесполое размножение, которое может проявляться в виде простого деления, почкования и множественного деления. Деление обычно наблюдается у свободноживущих форм: организм делится на две приблизительно равные клетки. Сначала происходит деление ядра, а потом – цитоплазмы. Реже встречается у простейших бесполое размножение в виде почкования. У большинства паразитических простейших, а также у ряда свободноживущих форм распространено множественное деление, или шизогония, при котором материнская особь распадается на большое количество дочерних. Во всех классах простейших наблюдается половое размножение. В большинстве случаев оно осуществляется путем слияния (копуляции) микрогаметы с макрогаметой. У инфузорий половой процесс осуществляется в виде конъюгации. У простейших, возникших в результате разных способов размножения, строение в какой-то степени отличается от родительских форм и не все органоиды имеются в должном количестве (например, из двух сократительных вакуолей может остаться одна). Восстановление обычного строения и недостающих органоидов совершается в процессе быстрого онтогенетического развития. Простейшие обладают широким всесветным распространением. Множество их живет в море. Некоторые входят в состав бентоса на различных глубинах – от литорали до абиссали (фораминиферы, инфузории, жгутиконосцы). Многочисленные виды радиолярий, жгутиконосцев и инфузорий являются компонентами морского планктона. Многие простейшие (жгутиконосцев, инфузории, корненожки) входят в состав пресноводного бентоса и планктона. Существуют некоторые виды, живущие во влажной почве. Наконец, широкое распространение среди всех классов простейших получил паразитизм.
В большинстве случаев оно осуществляется путем слияния (копуляции) микрогаметы с макрогаметой. У инфузорий половой процесс осуществляется в виде конъюгации. У простейших, возникших в результате разных способов размножения, строение в какой-то степени отличается от родительских форм и не все органоиды имеются в должном количестве (например, из двух сократительных вакуолей может остаться одна). Восстановление обычного строения и недостающих органоидов совершается в процессе быстрого онтогенетического развития. Простейшие обладают широким всесветным распространением. Множество их живет в море. Некоторые входят в состав бентоса на различных глубинах – от литорали до абиссали (фораминиферы, инфузории, жгутиконосцы). Многочисленные виды радиолярий, жгутиконосцев и инфузорий являются компонентами морского планктона. Многие простейшие (жгутиконосцев, инфузории, корненожки) входят в состав пресноводного бентоса и планктона. Существуют некоторые виды, живущие во влажной почве. Наконец, широкое распространение среди всех классов простейших получил паразитизм. Многие виды паразитических простейших вызывают тяжелые заболевания человека, домашних и промысловых животных. Некоторые виды паразитируют в растениях. Таким образом, простейшие имеют важное практическое значение для медицины, ветеринарии, сельского хозяйства. Принято разделение типа на шесть классов: 1. Класс жгутиковые; 2. Класс ложноножковые, или саркодовые; 3. Класс споровики; 4. Класс слизистые споровики; 5. Класс микроспоридии; 6. Класс инфузории. Класс жгутиковые Эти простейшие передвигаются с помощью жгутиков. Оболочка тела плотная, что придает определенную форму их телу и облегчает быстрое передвижение. Однако, у некоторых жгутиковых, оболочка очень тонкая и они передвигаются медленно, переливаясь всем телом во временные выросты тела – ложноножки, или псевдоподии. Эти формы наряду с ложноножками имеют и жгутики. Многие представители имеют наружные скелеты из клетчатки или других веществ. У форм, постоянно находящихся в толще Класс споровики Все представители этого класса – паразиты беспозвоночных и позвоночных животных.
Многие виды паразитических простейших вызывают тяжелые заболевания человека, домашних и промысловых животных. Некоторые виды паразитируют в растениях. Таким образом, простейшие имеют важное практическое значение для медицины, ветеринарии, сельского хозяйства. Принято разделение типа на шесть классов: 1. Класс жгутиковые; 2. Класс ложноножковые, или саркодовые; 3. Класс споровики; 4. Класс слизистые споровики; 5. Класс микроспоридии; 6. Класс инфузории. Класс жгутиковые Эти простейшие передвигаются с помощью жгутиков. Оболочка тела плотная, что придает определенную форму их телу и облегчает быстрое передвижение. Однако, у некоторых жгутиковых, оболочка очень тонкая и они передвигаются медленно, переливаясь всем телом во временные выросты тела – ложноножки, или псевдоподии. Эти формы наряду с ложноножками имеют и жгутики. Многие представители имеют наружные скелеты из клетчатки или других веществ. У форм, постоянно находящихся в толще Класс споровики Все представители этого класса – паразиты беспозвоночных и позвоночных животных. Они живут в пищеварительном тракте, в полостях тела, в кровеносной системе и в других органах хозяев. Многие выходят из организма хозяина в виде стадий, окруженных толстыми оболочками, часто называемых спорами. Взрослые споровики обычно малоподвижны или двигаются очень медленно (при помощи сократимых волоконец – мионем, или выделяя через мельчайшие поры на заднем конце тела густую жидкость). У ряда споровиков оболочки довольно тонкие. Гаметы у многих видов имеют жгутики. Сократительных вакуолей нет. Питаются споровики, всасывая растворенные органические и другие вещества всей поверхностью тела. Они разрушают ткани хозяев и отравляют их продуктами своего обмена веществ. Бесполое размножение часто чередуется с половым. Развитие споровиков у многих видов сопровождается сменой хозяев. Класс делится на два подкласса: кокцидиобразные и грегарины. Первый подкласс делится на: Отряд кокцидии. Они обитают большей частью в клетках пищеварительных органов. Многие из них паразитируют в организме сельскохозяйственных животных (кур, кроликов, коз, овец, крупного рогатого скота и др.
Они живут в пищеварительном тракте, в полостях тела, в кровеносной системе и в других органах хозяев. Многие выходят из организма хозяина в виде стадий, окруженных толстыми оболочками, часто называемых спорами. Взрослые споровики обычно малоподвижны или двигаются очень медленно (при помощи сократимых волоконец – мионем, или выделяя через мельчайшие поры на заднем конце тела густую жидкость). У ряда споровиков оболочки довольно тонкие. Гаметы у многих видов имеют жгутики. Сократительных вакуолей нет. Питаются споровики, всасывая растворенные органические и другие вещества всей поверхностью тела. Они разрушают ткани хозяев и отравляют их продуктами своего обмена веществ. Бесполое размножение часто чередуется с половым. Развитие споровиков у многих видов сопровождается сменой хозяев. Класс делится на два подкласса: кокцидиобразные и грегарины. Первый подкласс делится на: Отряд кокцидии. Они обитают большей частью в клетках пищеварительных органов. Многие из них паразитируют в организме сельскохозяйственных животных (кур, кроликов, коз, овец, крупного рогатого скота и др. ). У одного и того же хозяина могут обитать разные виды этих паразитов. Болезни, вызываемые кокцидиями, называются кокцидиозами. Заражение кокцидиями происходит в результате попадания вместе с пищей и водой в пищеварительный тракт хозяина инцистированных стадий паразитов – ооцист. В каждом ооцисте имеются спороцисты, в каждой спороцисте – узенькие подвижные клетки – спорозоиты. В кишечнике под действием пищеварительных соков оболочки ооцист и спороцист разрушаются и освободившиеся спорозоиты внедряются в клетки кишечника, печени, поджелудочной железы и превращаются в трофозоитов, которые питаются содержимым клеток хозяина и быстро растут. По окончании периода роста трофозоиты превращаются в шизонтов, которые делятся на большое количество узких, подвижных мерозоитов. Кроме присущих всем клеткам органоидов, в спорозоитах и мерозоитах обнаружены коноид – широкое кольцо в передней части клетки, служащее для опоры при внедрении их в клетки органов хозяина; роптрии – мешочки, содержащие вещество, облегчающее упомянутое внедрение; микронемы – волоконца, связанные с роптриями.
). У одного и того же хозяина могут обитать разные виды этих паразитов. Болезни, вызываемые кокцидиями, называются кокцидиозами. Заражение кокцидиями происходит в результате попадания вместе с пищей и водой в пищеварительный тракт хозяина инцистированных стадий паразитов – ооцист. В каждом ооцисте имеются спороцисты, в каждой спороцисте – узенькие подвижные клетки – спорозоиты. В кишечнике под действием пищеварительных соков оболочки ооцист и спороцист разрушаются и освободившиеся спорозоиты внедряются в клетки кишечника, печени, поджелудочной железы и превращаются в трофозоитов, которые питаются содержимым клеток хозяина и быстро растут. По окончании периода роста трофозоиты превращаются в шизонтов, которые делятся на большое количество узких, подвижных мерозоитов. Кроме присущих всем клеткам органоидов, в спорозоитах и мерозоитах обнаружены коноид – широкое кольцо в передней части клетки, служащее для опоры при внедрении их в клетки органов хозяина; роптрии – мешочки, содержащие вещество, облегчающее упомянутое внедрение; микронемы – волоконца, связанные с роптриями.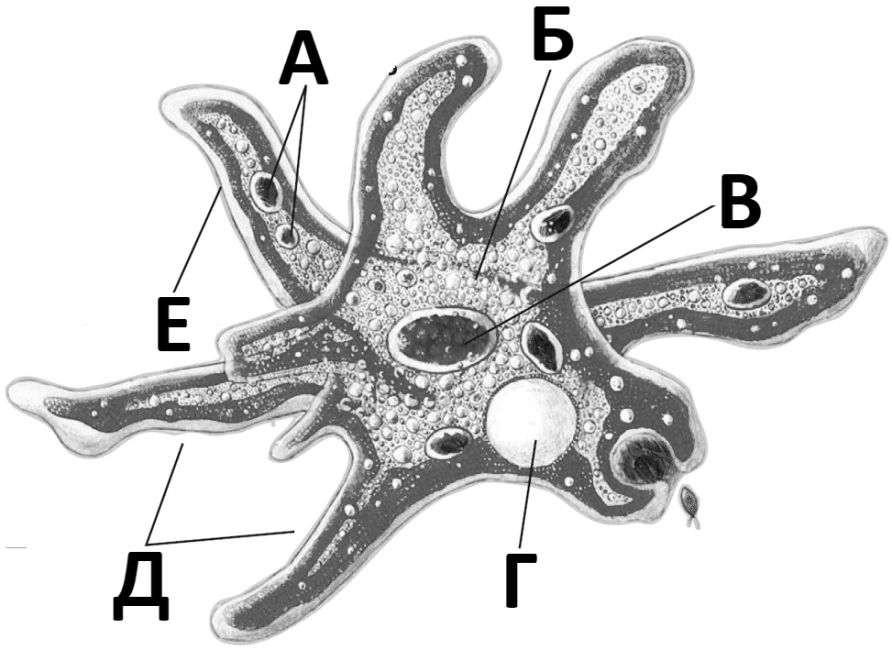 В конце передней трети описываемых клеток имеется пора. Мерозоиты выходят из разрушенных клеток органов хозяина и проникают в неповрежденные клетки. Затем из мерозоитов образуются особые клетки – гаметоциты, которые превращаются в зрелые половые клетки: женские – макрогаметы, мужские – микрогаметы. Микрогаметы и макрогаметы попарно сливаются и образуют зиготы, которые выделяют оболочки и превращаются в ооцисты. Ооцисты вместе с калом выходят наружу. Содержимое при благоприятных условиях делится на четыре споробласта. Каждый споробласт выделяет оболочку – спороцисту и делится у большинства видов на два спорозоита. Ооциста со спорозоитами способна заразить хозяев. Отряд кровяные споровики, или гемоспоридии. В отличие от кокцидий у них половое размножение происходит в одних хозяевах (различные виды комаров), а бесполое – в других (птицы, млекопитающие, человек). Гемоспоридии могут вызывать малярию. Самки комара, зараженные малярийными паразитам, питаясь кровью промежуточных хозяев, могут внести в кровь последних множество спорозоитов.
В конце передней трети описываемых клеток имеется пора. Мерозоиты выходят из разрушенных клеток органов хозяина и проникают в неповрежденные клетки. Затем из мерозоитов образуются особые клетки – гаметоциты, которые превращаются в зрелые половые клетки: женские – макрогаметы, мужские – микрогаметы. Микрогаметы и макрогаметы попарно сливаются и образуют зиготы, которые выделяют оболочки и превращаются в ооцисты. Ооцисты вместе с калом выходят наружу. Содержимое при благоприятных условиях делится на четыре споробласта. Каждый споробласт выделяет оболочку – спороцисту и делится у большинства видов на два спорозоита. Ооциста со спорозоитами способна заразить хозяев. Отряд кровяные споровики, или гемоспоридии. В отличие от кокцидий у них половое размножение происходит в одних хозяевах (различные виды комаров), а бесполое – в других (птицы, млекопитающие, человек). Гемоспоридии могут вызывать малярию. Самки комара, зараженные малярийными паразитам, питаясь кровью промежуточных хозяев, могут внести в кровь последних множество спорозоитов.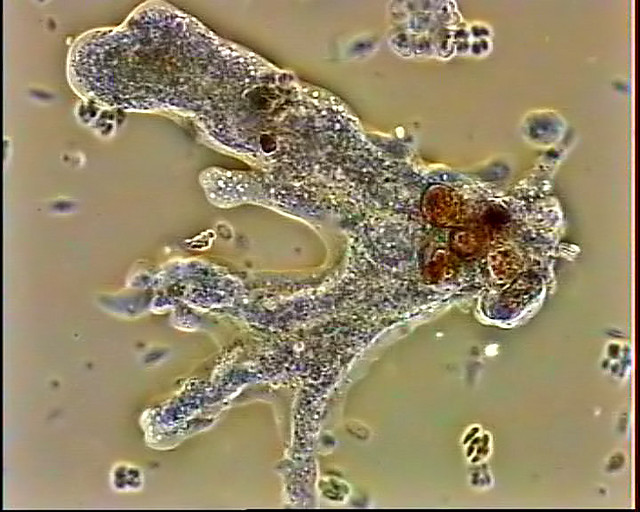 Спорозоиты проникают в клетки печени и клетки эндотелия сосудов и там проходят стадии трофозоитов и шизонтов, завершающиеся делением последних на множество мерозоитов, которые внедряются в эритроциты. Мерозоиты в эритроцитах проходят стадии трофозоита и шизонта. Шизонты затем делятся на новые мерозоиты. По окончании деления паразитов, зараженные эритроциты разрушаются, мерозоиты выходят в плазму и вновь внедряются в эритроциты. Характерно для малярийных плазмодиев образование из гемоглобина эритроцитов особого черного вещества – меланина. При разрушении эритроцитов в кровяное русло попадают, кроме меланина, ядовитые продукты обмена веществ споровиков. Освободившиеся мерозоиты проникают в новые эритроциты. На определенной стадии развития мерозоиты, проникая в эритроциты, больше уже не делятся, а превращаются в микро- и макрогаметоциты. Дальнейшее развитие гаметоцитов может происходить только в кишечнике самки комара, куда они попадают при сосании ею крови промежуточного хозяина. В кишечнике комара гаметоциты превращаются в зрелые половые клетки – микро- и макрогаметы.
Спорозоиты проникают в клетки печени и клетки эндотелия сосудов и там проходят стадии трофозоитов и шизонтов, завершающиеся делением последних на множество мерозоитов, которые внедряются в эритроциты. Мерозоиты в эритроцитах проходят стадии трофозоита и шизонта. Шизонты затем делятся на новые мерозоиты. По окончании деления паразитов, зараженные эритроциты разрушаются, мерозоиты выходят в плазму и вновь внедряются в эритроциты. Характерно для малярийных плазмодиев образование из гемоглобина эритроцитов особого черного вещества – меланина. При разрушении эритроцитов в кровяное русло попадают, кроме меланина, ядовитые продукты обмена веществ споровиков. Освободившиеся мерозоиты проникают в новые эритроциты. На определенной стадии развития мерозоиты, проникая в эритроциты, больше уже не делятся, а превращаются в микро- и макрогаметоциты. Дальнейшее развитие гаметоцитов может происходить только в кишечнике самки комара, куда они попадают при сосании ею крови промежуточного хозяина. В кишечнике комара гаметоциты превращаются в зрелые половые клетки – микро- и макрогаметы.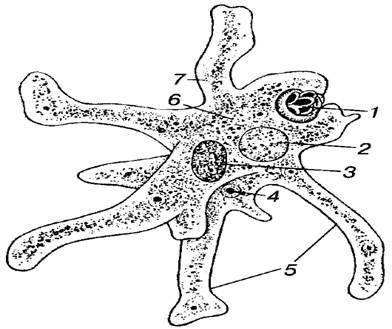 Узенькие, подвижные микрогаметы оплодотворяют округлые, малоподвижные макрогаметы, и образуются зиготы. Зиготы у гемоспоридий подвижные, они называются оокинетами. Последние, внедрившись в стенки кишечника, сильно вырастают, покрываются эластичной капсулой и превращаются в ооцисты. В ооцисте под защитой ее оболочки происходит деление на огромное количество спорозоитов. В конце концов, ооциста лопается, спорозоиты попадают в кровь комара и заносятся в его слюнные железы. Отряд пироплазмиды. Эти простейшие одну часть своего жизненного цикла проходят в организмах млекопитающих, а другую – в организмах кровососущих пастбищных клещей. Отряд мясные споровики. Они часто встречаются в виде довольно больших мешочкообразных цист главным образом в мышцах млекопитающих и птиц. Подкласс грегарины. Эти простейшие характеризуются своеобразной формой полового процесса, при котором происходит объединение двух особей (гамонтов) в сизигий, выделяющий общую оболочку. Под оболочкой и происходит формирование гамет (гаметогония) и их копуляция.
Узенькие, подвижные микрогаметы оплодотворяют округлые, малоподвижные макрогаметы, и образуются зиготы. Зиготы у гемоспоридий подвижные, они называются оокинетами. Последние, внедрившись в стенки кишечника, сильно вырастают, покрываются эластичной капсулой и превращаются в ооцисты. В ооцисте под защитой ее оболочки происходит деление на огромное количество спорозоитов. В конце концов, ооциста лопается, спорозоиты попадают в кровь комара и заносятся в его слюнные железы. Отряд пироплазмиды. Эти простейшие одну часть своего жизненного цикла проходят в организмах млекопитающих, а другую – в организмах кровососущих пастбищных клещей. Отряд мясные споровики. Они часто встречаются в виде довольно больших мешочкообразных цист главным образом в мышцах млекопитающих и птиц. Подкласс грегарины. Эти простейшие характеризуются своеобразной формой полового процесса, при котором происходит объединение двух особей (гамонтов) в сизигий, выделяющий общую оболочку. Под оболочкой и происходит формирование гамет (гаметогония) и их копуляция.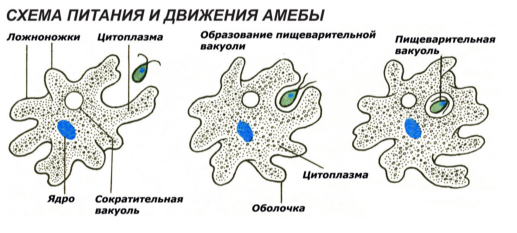 Все грегарины – паразиты различных групп беспозвоночных животных. Большую часть жизненного цикла – внеклеточные паразиты кишечника или полости тела, реже – половых желез. Передний конец тела большинства грегарин образует органоид прикрепления к стенкам кишечника – эпимерит. Снаружи тело одето
Все грегарины – паразиты различных групп беспозвоночных животных. Большую часть жизненного цикла – внеклеточные паразиты кишечника или полости тела, реже – половых желез. Передний конец тела большинства грегарин образует органоид прикрепления к стенкам кишечника – эпимерит. Снаружи тело одето
Определение, функция, движение и примеры
Определение, функция, движение и примеры
Определение: Что такое псевдоподы?
Также известные как псевдоподии (существительное в единственном числе: псевдоподии), псевдоподии представляют собой временные расширения цитоплазмы (также называемые ложными стопами), используемые для передвижения и ощущения. Их можно найти у всех саркодинов, а также у ряда жгутиковых простейших, которые существуют либо как паразиты, либо как свободноживущие организмы.
У высших животных ложноножки можно наблюдать в ряде лейкоцитов (фагоцитарных клеток), которые используют эту структуру для улавливания и уничтожения вторгшихся микробов. В зависимости от типа клетки различают четыре основных типа, которые не только различаются по внешнему виду (и общей морфологии), но и выполняют разные функции.
В зависимости от типа клетки различают четыре основных типа, которые не только различаются по внешнему виду (и общей морфологии), но и выполняют разные функции.
Например, у некоторых организмов псевдоподии содержат микротрубочки, которые вносят значительный вклад в движение клеток.
Существует четыре типа ложноножек, в том числе:
- Lobopodia
- Axopodia
- Filopodia
- Reticulopodia/Rhizopoda
- Транспортировка кремния, полученного от некоторых жертв
РетикулоподиумТакже называемые в некоторых книгах ризоподиями (или внеталамической цитоплазмой), ретикулоподии представляют собой нитевидные псевдоподии, которые разветвляются и сливаются, образуя чрезвычайно динамичную сеть.Как и в случае с аксоподиями, ретикулоподии также состоят из канальцев и цитоплазмы.
Их можно найти в ряде организмов, включая амебы Endomyxa и некоторые фораминиферы (древняя группа простейших). У этих организмов ретикулоподии участвуют в питании и передвижении.
Как и аксоподии, ретикулоподии также состоят из микротрубочек и цитоплазмы. Здесь микротрубочки, из которых состоят псевдоподии, состоят из уникального типа тубулина, известного как бета-тубулин типа 2.
 Этот тубулин образует спиральные филаменты (HF), которые являются основой микротрубочек, обнаруживаемых в фораминиферовых ретикулоподиях.
Этот тубулин образует спиральные филаменты (HF), которые являются основой микротрубочек, обнаруживаемых в фораминиферовых ретикулоподиях.У фораминифер и других организмов ретукулоподии выпячиваются через одну или несколько пор (апертуальные отверстия). Первоначально эти псевдоподии могут быть тонкими и заостренными (по внешнему виду похожи на филоподии).
По мере увеличения количества цитоплазмы в структуре псевдоподиальный ствол, известный как ножка, становится толще и разветвляется, образуя новые псевдоподии.В то время как эти псевдоподии растут и анастомозируют (связываются вместе), они образуют сеть, напоминающую паутинные нити.
Некоторые из основных характеристик ретикулоподии включают в себя:
· 9004
· , отличные от других в том, что они сильно разветвляются и формируют анастомозные сети
· в зависимости от организма, ретикулоподия могут простираться несколько сантиметров. из тела клетки организма
· Может быстро вытягиваться и втягиваться со скоростью около 20 мкм/с
· процесс, известный как ответ на бусинки
· Внутри псевдоподий транспорт частиц двунаправленный.
 Это означает, что цитоплазма течет по длине ложноножек к телу клетки и от него
Это означает, что цитоплазма течет по длине ложноножек к телу клетки и от него
ФункцииКак и аксоподии, ретикулоподии играют важную роль в питании и передвижении. Однако их основная функция заключается в сборе пищи и кормлении. Во время кормления организм распространяет ложноножки (выглядящие как неправильная паутина), которые очищают их непосредственную поверхность и собирают доступный пищевой материал для приема внутрь.
Добычей могут быть такие одноклеточные организмы, как бактерии, которые попадают в паутину и попадают в пищевые вакуоли для пищеварения.Помимо питания, ретикулоподии также используются для передвижения. Однако это не является их основной функцией.
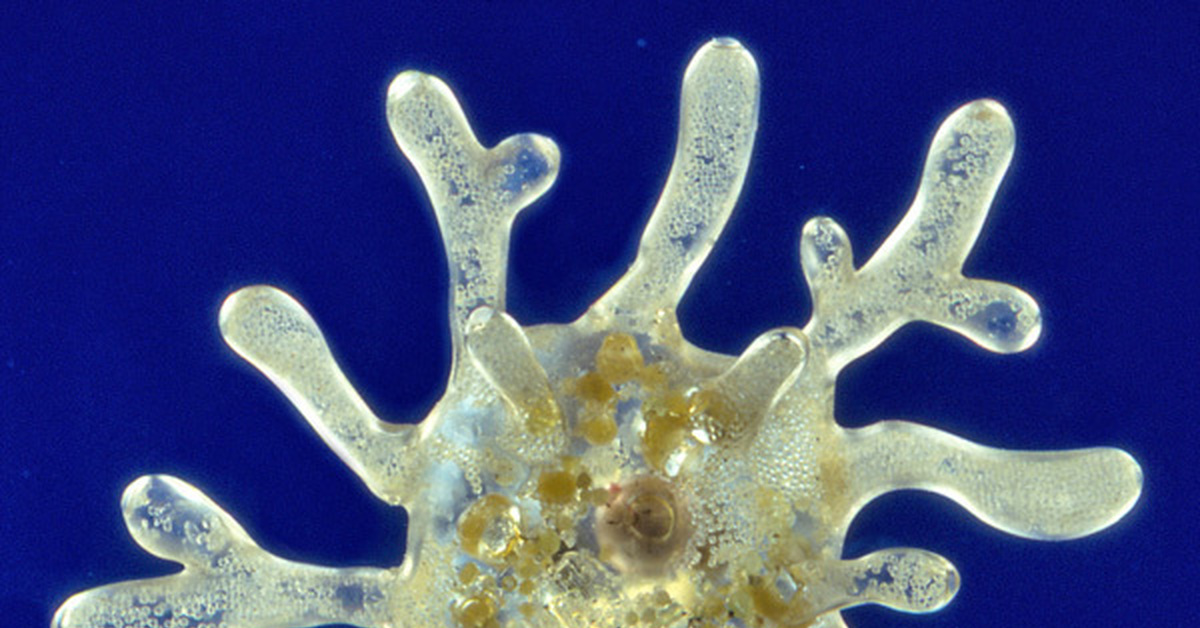
ЛобоподияLopodium является наиболее распространенным типом, встречающимся в таких организмах, как Amoeba proteus. Для лобопод характерны пальцевидные трубчатые псевдоподии, состоящие из экто- и эндоплазмы. Однако было также показано, что они содержат актин и миозин (микрофиламенты), которые способствуют общему движению.
В отличие от других ложноножек микротрубочки лобоподий развиты слабо.У многих амеб лобоподии в первую очередь участвуют в передвижении.
Формирование и передвижениеУ таких организмов на формирование лобоподий влияют химические сигналы в окружающей среде. В присутствии пищевого вещества химические сигналы влияют на направление движения амебы. Здесь молекулы (из пищевого материала) связываются с рецепторами, расположенными на клеточной мембране организма, что стимулирует образование филаментов за счет агрегации глобулярного актина.
При добавлении глобулярного актина структура (филамент) продолжает удлиняться, что, в свою очередь, вызывает выпячивание мембраны (это действие приводит к образованию псевдоподий). Выступающие лобоподы по мере своего расширения заполняются цитоплазмой. В случае исчезновения молекул глобулярный актин распадается, что останавливает дальнейшее удлинение ложноножек.
Если молекулы сохраняются, миозин, который действует как двигательные белки, взаимодействует с актином, толкая тело клетки в направлении ложноножки.
* Миозиновая активность (в качестве двигателей) требует энергии (АТФ).
* Также было показано, что вязкость цитоплазмы изменяется при ее втекании и вытекании из ложноножек.
Во время кормления лобоподии также окружают пищевой материал и поглощают его внутри пузырька, где на него воздействуют различные ферменты. Затем продукты жизнедеятельности выводятся через вакуоли, открывающиеся в окружающую среду.
Вернуться на главную страницу Protozoa
Вернуться в цитоплазму
Вернуться из Pseudopods на главную страницу MicroscopeMaster
СсылкиЧансон Ян и Татьяна Свиткина.(2011). Инициация филоподий: сосредоточьтесь на комплексе Arp2/3 и форминах. нкби.
Hou Y et al. (2013). Молекулярные доказательства неофункционализации β-тубулина у Retaria (фораминиферы и радиолярии). NBCI.
Кара Роджерс. (2011). Грибы, водоросли и протисты.
П.
 Де Вевер, П. Думитрика, Дж. П. Коле, К. Нигрини и М. Каридройт. (2001). Радиолярии в осадочной летописи.
Де Вевер, П. Думитрика, Дж. П. Коле, К. Нигрини и М. Каридройт. (2001). Радиолярии в осадочной летописи.Сэмюэл С. Баузер и Джеффри Л. Трэвис. (2000). Методы структурных исследований ретикулоподий, «мягкой части» жизненно важных фораминифер.jstor.org.
Стефани Л. Гуптон и Фрэнк Б. Гертлер. (2007). Филоподия: Пальцы, которые ходят. ResearchGate.
9Ссылки
https://royalsocietypublishing.org/doi/10.1098/rsos.160283
https://www.researchgate.net/publication/44576108_understinganding_eukaryoTy_chemotaxis_a_pseudopod-centred_view
Псевдоподии — обзор | ScienceDirect Topics
Может ли цитоскелет выполнять вычисления?
Цитоскелет, особенно микротрубочки и актиновые филаменты, необходимы для ядерных процессов, происходящих во время митотических и мейотических делений.Они также играют решающую роль в определении положения органелл и во внутриклеточном транспорте везикул и других надмолекулярных элементов; они определяют форму клетки и динамизм формы клетки.
 Все эти функции, выполняемые микротрубочками и актиновыми филаментами, являются обязательными, а не случайными процессами.
Все эти функции, выполняемые микротрубочками и актиновыми филаментами, являются обязательными, а не случайными процессами.В строго экспериментальных условиях поведение амебы (образование псевдоподий, направление ее движений, начало репродуктивной деятельности и многие физиологические процессы) предсказуемо.Поведение амебы призвано адаптировать ее к экспериментальным вызовам внешней среды. Но преднамеренные изменения, вызывающие адаптацию, подразумевают, что система предсказывает* адаптивный результат, просчитывает пути и предпринимает шаги для его достижения. Но если «предсказание — это вычисление» (Pfaffmann and Zauner, 2001), то где выполняются вычисления для определения поведения амебы?
Отец современной неврологии, испанский ученый Сантьяго Рамон-и-Кахаль, еще столетие назад высказал идею о том, что цитоскелет участвует в проведении нервного импульса, а еще в 1951 году отец-основатель нейрофизиологии Чарльз Скотт Шеррингтон, казалось бы, предвосхитил современная тенденция рассматривать цитоскелет как потенциальную «нервную систему» клетки у одноклеточных:
Нерва нет и в помине.
 Но клеточный каркас, цитоскелет, может служить.
Но клеточный каркас, цитоскелет, может служить.Sherrington (1951)
Действительно, нейроны используют цитоскелет для увеличения своей потенциальной вычислительной мощности до 10 16 битовых состояний/с (Rasmussen et al., 1990).
Экспериментально подтвержденный механизм вычисления в микротрубочках цитоскелета отсутствует. Однако предполагается, что цитоскелет обрабатывает информацию у одноклеточных и нервных клеток. Известно, например, что обучение связано с повышенной продукцией тубулинов (строительных блоков микротрубочек) в мозге цыплят и крысят и более высокой активностью микротрубочек (Hameroff, 1998; Tuszynski et al., 1998). Считается, что в нейронах микротрубочки могут использоваться для обработки электрической информации и когнитивной обработки (Priel et al., 2006). Имеются достаточные доказательства, чтобы твердо утверждать, что центриоли, MTOCs одноклеточных, являются органеллами, в которых происходит такое вычисление. В этом контексте важно помнить, что правильное размещение органелл внутри клетки также определяется MTOC.
 Действительно, неправильное расположение центриолей вызывает неспособность Chlamydomonas ’ двигаться в направлении источника света.
Действительно, неправильное расположение центриолей вызывает неспособность Chlamydomonas ’ двигаться в направлении источника света.С точки зрения ученых-компьютерщиков эти одноклеточные представляют собой биологические микромашины, а цитоскелет является их центральной системой. Отвечая на внутренние и внешние сигналы, цитоскелет реорганизуется, чтобы генерировать ответы, которые адаптируют клетку к изменившейся внешней или внутренней среде (Glade, 2008).
В модели Glade (2008) элементарными единицами биологической микромашины (цитоскелета) являются отдельные волокна (микротрубочки, актиновые филаменты и промежуточные филаменты), мономеры которых (молекулы тубулина, актина и др.) могут действовать как молекулярные биты информации. Отдельные микротрубочки связаны белками, ассоциированными с микротрубочками (MAPs) и другими белками, что делает возможной связь между волокнами сети цитоскелета. В процессе своего роста и деполимеризации микротрубочки оставляют химические следы, которые позволяют обмениваться информацией с соседними микротрубочками.
 В результате динамический скелет способен выполнять функции примитивного мозга и действовать «как автономная система, чувствительная к внешним раздражителям, придающая клеткам очень сложное поведение.(Поляна, 2008). По-видимому, он инструктирует клетку адаптивно реагировать на внешние и внутренние вызовы.
В результате динамический скелет способен выполнять функции примитивного мозга и действовать «как автономная система, чувствительная к внешним раздражителям, придающая клеткам очень сложное поведение.(Поляна, 2008). По-видимому, он инструктирует клетку адаптивно реагировать на внешние и внутренние вызовы.Функции и структура микротрубочек тесно связаны с MAP. Аксоны и дендриты нервных клеток очень богаты микротрубочками. Рост микротрубочек является результатом полимеризации димеров тубулина (молекул α- и β-тубулина), на которую влияет биохимическое окружение и особенно MAP, связанные с микротрубочками.
В модели регуляции сетей микротрубочек в клетке MAP передают цитоплазматические сигналы микротрубочкам, которые также получают сигналы от всей сети волокнистых элементов цитоскелета.MAP, в свою очередь, получают результат обработки цитоплазматической информации от микротрубочек.
Рост микротрубочек зависит от способности сети микротрубочек связываться с MAP. Когда обработка сигналов в сети хорошая, MAP связываются с микротрубочками.
 Повышенная активность связывания в сети повышает стабильность сети и сигнализирует о том, что никаких изменений не требуется. Наоборот, снижение устойчивости сети служит сигналом к изменению сетевой структуры.Адаптивная самостабилизация (рис. 1.24) основана на механизме обратной связи по структуре сети микротрубочек и распределению МАР в структуре. Система отображает некоторое элементарное обучение и подразумевает своего рода память. Адаптивная обработка информации нуждается в экспериментальной проверке, но авторы приходят к выводу, что «тонкая структура клетки является естественной средой для адаптивной обработки информации» (Pfaffmann and Conrad, 2000).
Повышенная активность связывания в сети повышает стабильность сети и сигнализирует о том, что никаких изменений не требуется. Наоборот, снижение устойчивости сети служит сигналом к изменению сетевой структуры.Адаптивная самостабилизация (рис. 1.24) основана на механизме обратной связи по структуре сети микротрубочек и распределению МАР в структуре. Система отображает некоторое элементарное обучение и подразумевает своего рода память. Адаптивная обработка информации нуждается в экспериментальной проверке, но авторы приходят к выводу, что «тонкая структура клетки является естественной средой для адаптивной обработки информации» (Pfaffmann and Conrad, 2000).Рисунок 1.24. Циклический поток управления.Модули динамики роста и обработки сигналов имеют общее представление сети микротрубочек. Модуль адаптивной самостабилизации связывает динамику роста с эффективностью обработки сигналов (Pfaffmann and Conrad, 2000).
В дополнение к своей роли в определении формы и движений клетки и расположения органелл внутри этой клетки, распространение везикул и стимулов по всей клетке и свидетельства процессов клеточного деления показывают, что в клетках млекопитающих цитоскелет участвуют в процессах экспрессии генов (Berfield et al.
 , 1997; Пак и др., 1990). Предполагается, что в нейронах, по крайней мере, именно цитоскелет сигнализирует генам о потребности в продукции белка (Georgiev, Quantum Mind Theories; http://www.quantum-mind.co.uk/danko-georgiev- c169.html). В клетках млекопитающих цитоскелет находится в контакте как с внеклеточным матриксом (ECM), так и с ядерным матриксом, который действует как преобразователь внеклеточных сигналов к ядру клетки. Промежуточные филаменты (ПФ) тесно связаны со структурными элементами ядерного матрикса и являются составной частью ядерного скелета (Tolstonog et al., 2002).
, 1997; Пак и др., 1990). Предполагается, что в нейронах, по крайней мере, именно цитоскелет сигнализирует генам о потребности в продукции белка (Georgiev, Quantum Mind Theories; http://www.quantum-mind.co.uk/danko-georgiev- c169.html). В клетках млекопитающих цитоскелет находится в контакте как с внеклеточным матриксом (ECM), так и с ядерным матриксом, который действует как преобразователь внеклеточных сигналов к ядру клетки. Промежуточные филаменты (ПФ) тесно связаны со структурными элементами ядерного матрикса и являются составной частью ядерного скелета (Tolstonog et al., 2002).В модели регуляции сетей микротрубочек в клетке MAP передают цитоплазматические сигналы микротрубочкам, которые получают сигналы от всей сети волокнистых элементов цитоскелета. MAP, в свою очередь, получают результат обработки цитоплазматической информации от микротрубочек.
Некоторые молекулы ВКМ связаны с клеточными микрофиламентами через актин-ассоциированные белки, образуя интегрированную систему с внутриклеточными элементами цитоскелета (Ingber, Folkman, 1989).
В ответ на фотостимуляцию ближним инфракрасным светом некоторые одноклеточные вытягивают псевдоподии в сторону источника света. Утверждается, что удлинение не может быть выполнено за счет продукции клеткой распространяемых химических сигналов, которые без разбора воздействовали бы на псевдоподии по всей клетке. Следовательно, микротрубочки могут передавать сигнал на специфические псевдоподии. Действительно, замечено, что после введения антимикротубулярных веществ клетка теряет эту способность к адаптации, вытягивая правые псевдоподии, хотя еще может двигаться (http://www.basic.northwestern.edu/g-buehler/nerves.htm). Вот интерпретация этого явления исследователем:
После получения световых импульсов центросома дестабилизирует радиальный ряд микротрубочек, идущих к клеточной коре, которая, в свою очередь, впоследствии вытягивает специальные псевдоподии к источникам света. Поэтому представляется, что наблюдаемая дестабилизация – это сигнал, который распространяется по микротрубочкам, как по «нервам».
http://www.basic.northwestern.edu/g-buehler/nerves.htm
Основываясь на экспериментальных данных, Гюнтер Альбрехт-Бюлер пришел к выводу, что центросома представляет собой органеллу, в которой происходит интеграция световых сигналов и откуда берут начало сигналы микротрубочек для формирования псевдоподий (Albrecht-Buehler , 1998).
Спорное и далеко не подтвержденное экспериментально объяснение системы управления было разработано физиком Роджером Пенроузом и анестезиологом Стюартом Хамероффом. Протофиламенты состоят из молекул тубулина в виде тубулиновых димеров мономеров α- и β-тубулина.Димеры тубулина могут быть основными вычислительными единицами МТ (Penrose, 2003), которые действуют как клеточные автоматы для передачи сигналов (Hameroff and Penrose, 1996). MTs распространяют и обрабатывают сигналы, и эти распространяющиеся сигналы, по-видимому, имеют отношение к способу, которым микротрубочки транспортируют различные молекулы вместе с собой, и к различным взаимосвязям между соседними микротрубочками в форме MAP (Penrose, 2003).

В другой модели Георгиев предполагает, что диполи молекул воды внутри нейронов распадаются на кванты дипольных волн, которые передают информацию в воде, воздействуя на тубулиновые «хвосты» микротрубочек, которые передают информацию по микротрубочкам.«Предполагается, что столкновения волн, образованных тубулиновыми хвостами, действуют как вычислительные ворота для контроля процессов цитоскелета» (Георгиев, 2011), включая определение мест прикрепления MAP в микротрубочках, которые, в свою очередь, передают результаты выполненных вычислений. тубулиновыми хвостами.
Псевдоподы: определение и функции — видео и стенограмма урока
Стрелки на этом изображении указывают на ложноножки. Функция псевдоподов
Псевдоподы на самом деле являются продолжением цитоплазмы или густой жидкости, которая находится внутри таких организмов, как амебы.Организм может изменять форму ложноножки, заставляя ее двигаться, появляться и исчезать.
Псевдоножки используются для движения и в качестве инструмента для захвата добычи. Чтобы двигаться с помощью псевдоподий, организм отталкивает цитоплазму к одному концу клетки, что образует выступ или псевдоподию за пределами клетки. Этот выступ удерживает существо на месте, а остальная часть клетки может следовать за ним, тем самым продвигая организм вперед. Для питания организмы вытягивают свои ложноножки, поглощая свою добычу, а затем переваривая ее с помощью ферментов.
Стрелки на этом изображении указывают на ложноножки, которые поглощают добычу синего цвета. Типы псевдоподов
Псевдоподы бывают разных форм и находятся на многих различных организмах, в первую очередь простейших , которые являются одноклеточными тварями, которые должны потреблять пищу, в отличие от водорослей, которые могут производить свою собственную посредством фотосинтеза.
 Есть ложноножки, похожие на короткие пальцы.Есть ложноножки длинные и тонкие, а есть даже ложноножки, которые перекрещиваются друг с другом, образуя разветвленную сеть.
Есть ложноножки, похожие на короткие пальцы.Есть ложноножки длинные и тонкие, а есть даже ложноножки, которые перекрещиваются друг с другом, образуя разветвленную сеть.Euglypha — это вид простейших с длинными тонкими ложноножками, называемыми филоподиями, которые можно увидеть здесь. Радиолярии, еще один тип простейших, имеют длинные толстые ложноножки, называемые аксоподиями. Они состоят из микротрубочек и могут перекрываться. Это поедающая мозг амеба, простейшее существо с короткими, похожими на пальцы ложноножками, называемыми лобоподиями.Это самый распространенный тип псевдоподий. Краткий обзор урока
Псевдоподы , или ложные ноги, встречаются у многих простейших, от мозгоядных амеб до радиолярий.
 Простейшие — это одноклеточные существа, которые должны потреблять пищу. Хотя ложноножки бывают самых разных форм и форм, у простейших они выполняют одну и ту же функцию: движение и захват добычи.
Простейшие — это одноклеточные существа, которые должны потреблять пищу. Хотя ложноножки бывают самых разных форм и форм, у простейших они выполняют одну и ту же функцию: движение и захват добычи.Например, амеба способна изменять форму и двигаться, выталкивая цитоплазму наружу, создавая псевдоподии, которые действуют как якорь, тянущий ее вперед.Они также используют ложноножки, чтобы окружать и захватывать добычу.
Итак, когда вы расслабляетесь, наблюдая закат, а поедающая мозг амеба пробирается к ткани вашего мозга, вы можете найти утешение в том, что ложноножки — это адаптация, которая дала простейшим возможность перемещаться в новые места и питаться захватывающие новые продукты, такие как ваш мозг.
Псевдоподы: ключевые слова
Условия Определения Амеба разнообразная группа крошечных одноклеточных организмов, использующих ложноножки по крайней мере в течение части своего жизненного цикла Псевдоподы фальш-ножки; выступы, которые могут появляться и исчезать из тела организма, используемые в движении и для захвата добычи Цитоплазма густая жидкость внутри таких организмов, как амеба, которая может изменять форму ложноножки, заставляя ее двигаться, появляться и исчезать Простейшие одноклеточных тварей, которые должны потреблять пищу Результаты обучения
После просмотра этого урока вы сможете:
- Давать определение ложноногам и простейшим
- Опишите, как ложноножки используются для передвижения и охоты
- Вспомнить разные типы ложноножек
Pseudopod — wikidoc
Главный редактор: C.
 Майкл Гибсон, MS, MD [1]
Майкл Гибсон, MS, MD [1]Пожалуйста, займите эту страницу и подайте заявку на пост главного редактора этой темы: Может быть один или несколько главных редакторов. Вы также можете подать заявку на должность младшего главного редактора одной из подтем ниже. Пожалуйста, напишите нам [2], чтобы указать, что вы заинтересованы в работе либо в качестве главного редактора всей темы, либо в качестве помощника главного редактора для подтемы. Обязательно прикрепите свое резюме и/или биографический очерк.
Обзор
Псевдоподии или псевдоподии (ложные ноги) являются временными отростками эукариотических клеток.Клетки, обладающие этой способностью, обычно называют амебовидными.
Формирование
Во-первых, клеточная поверхность имеет мембранный отросток, называемый ламеллиподием . Происходит полимеризация актина и формирование нитей на переднем крае, которые впоследствии сливаются друг с другом, образуя сети.
 Предполагается, что полимеризация актина лежит в основе силы, толкающей клетку вперед.
Предполагается, что полимеризация актина лежит в основе силы, толкающей клетку вперед.Функция
Механическая роль
Псевдоподы — один из трех способов передвижения одноклеточных организмов (вместе со жгутиками и ресничками).
Морфология
Псевдоподы можно разделить на несколько разновидностей в зависимости от их внешнего вида:
- Lobopodia имеют выпуклую, короткую и тупую форму, очень типичную для Amoebozoa.
- Филлоподии более тонкие и нитевидные с заостренными концами, состоящие в основном из эктоплазмы. Эти образования поддерживаются микрофиламентами.
- Ретикулоподии , также известные как ретикулярные ложноножки , представляют собой сложные образования, в которых отдельные ложноножки сливаются вместе и образуют неправильные сети.
- Аксоподии представляют собой тонкие псевдоподии, содержащие сложные массивы микротрубочек и окруженные цитоплазмой.
 Аксоподии отвечают за фагоцитоз, быстро втягиваясь в ответ на физические контакты. Они наблюдаются у радиолярий и гелиозоа. Это, очевидно, вызывает напряжение в спирали, поскольку после того, как сенсорное действие произошло, она затем умирает.
Аксоподии отвечают за фагоцитоз, быстро втягиваясь в ответ на физические контакты. Они наблюдаются у радиолярий и гелиозоа. Это, очевидно, вызывает напряжение в спирали, поскольку после того, как сенсорное действие произошло, она затем умирает.
Шаблон:SIB
Шаблон: Исходники WikiDoc
Псевдоножки эндотелиальных клеток и ангиогенез опухолей молочной железы | Cancer Cell International
В общей сложности 10 опухолей размером более 35 мм 3 были зафиксированы, залиты парафином и сделаны средние срезы толщиной 8 мкм для окрашивания и гистологического исследования.Гистологическое исследование средних срезов опухолей, окрашенных PAS, было проведено для оценки характера васкуляризации опухоли. Опухоли объемом >35 мм 3 демонстрировали соединительнотканную капсулу с кровеносными сосудами (рис. 1А). При большем увеличении в области коры в пределах примерно 100 мкм от капсулы было обнаружено несколько кровеносных сосудов, в то время как в области более 100 мкм от капсулы были обнаружены значительные признаки кровеносных сосудов и капилляров с множеством эндотелиальных псевдоножек, отходящих от капилляров (рис.
Рисунок 1 1В и С). Общее направление ложноножек параллельно поверхности капсулы опухоли. Иммуногистохимическая локализация CD-31, используемого в качестве специфического маркера эндотелия кровеносных сосудов, продемонстрировала положительную реакцию ложноножек (рис. 1Г). Области некроза опухоли наблюдались ниже подкорковой области (рис. 1E и F). Иммуногистохимическая локализация индуцируемого гипоксией фактора 1-α (HIF) показывает, что подкорковая область опухоли содержит HIF-положительные клетки, в то время как капсула опухоли, кора и некротические области опухоли не демонстрируют признаков HIF.Таким образом, области, которые оказались HIF-позитивными, были обогащены эндотелиальными псевдоножками.
1В и С). Общее направление ложноножек параллельно поверхности капсулы опухоли. Иммуногистохимическая локализация CD-31, используемого в качестве специфического маркера эндотелия кровеносных сосудов, продемонстрировала положительную реакцию ложноножек (рис. 1Г). Области некроза опухоли наблюдались ниже подкорковой области (рис. 1E и F). Иммуногистохимическая локализация индуцируемого гипоксией фактора 1-α (HIF) показывает, что подкорковая область опухоли содержит HIF-положительные клетки, в то время как капсула опухоли, кора и некротические области опухоли не демонстрируют признаков HIF.Таким образом, области, которые оказались HIF-позитивными, были обогащены эндотелиальными псевдоножками.Микрофотографии, иллюстрирующие характер васкуляризации опухоли. A. В капсуле опухоли (слева) на этом срезе, окрашенном PAS, видны кровеносные сосуды. В коре под капсулой нет кровеносных сосудов и мало эндотелиальных ложноножек, в то время как в подкорковой области вверху справа ложноножек больше.
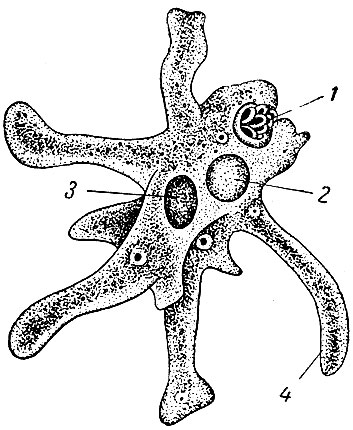 Б, при большем увеличении в подкорковой области опухоли обнаруживается кровеносный капилляр с множественными эндотелиальными псевдоножками, выступающими под прямым углом в опухолевую массу.C. Видно, что эндотелиальные псевдоножки разветвляются и иногда имеют вакуоль/просвет (стрелка). D. Эндотелиальные псевдоножки реагируют иммуногистохимически положительно на специфический маркер эндотелиальных клеток CD-31 с использованием метода комплекса авидин-биотин-пероксидазы. E, область жизнеспособных клеток видна под капсулой опухоли (слева), а область некроза видна справа. F, увеличенная подкорковая область от E. В E положительная область HIF-α между жизнеспособной и некротической тканью окрашена в коричневый цвет.
Б, при большем увеличении в подкорковой области опухоли обнаруживается кровеносный капилляр с множественными эндотелиальными псевдоножками, выступающими под прямым углом в опухолевую массу.C. Видно, что эндотелиальные псевдоножки разветвляются и иногда имеют вакуоль/просвет (стрелка). D. Эндотелиальные псевдоножки реагируют иммуногистохимически положительно на специфический маркер эндотелиальных клеток CD-31 с использованием метода комплекса авидин-биотин-пероксидазы. E, область жизнеспособных клеток видна под капсулой опухоли (слева), а область некроза видна справа. F, увеличенная подкорковая область от E. В E положительная область HIF-α между жизнеспособной и некротической тканью окрашена в коричневый цвет.Счет пересечения глазной сетки использовался для количественной оценки васкуляризации опухоли в окрашенных PAS гистологических срезах толщиной 8 мкм.Количество пересечений глазной сетки оценивали по: 1.) кровеносным сосудам и капиллярам, 2.) эндотелиальным псевдоподиям и 3.
Рисунок 2 ) областям без признаков этих структур. Было показано, что этот метод является удобной мерой объемной плотности, занимаемой распознаваемыми структурами. Результаты оценки кровеносных сосудов и эндотелиальных ложноножек в подкорковых областях опухолей представлены на рис. 2. Как показано на рис. 2, объемная плотность ложноножек в корковом слое опухоли была значительно меньше, чем в подкорковом. .
) областям без признаков этих структур. Было показано, что этот метод является удобной мерой объемной плотности, занимаемой распознаваемыми структурами. Результаты оценки кровеносных сосудов и эндотелиальных ложноножек в подкорковых областях опухолей представлены на рис. 2. Как показано на рис. 2, объемная плотность ложноножек в корковом слое опухоли была значительно меньше, чем в подкорковом. .Количественное определение различий объемной плотности псевдоножек эндотелиальных клеток в коре опухоли в пределах 100–150 мкм от хорошо васкуляризированной капсулы опухоли и в подкорковой области на расстоянии более 150 мкм под капсулой опухоли . Среднее ± SEM объемной плотности ложноножек 10 опухолей в двух разных областях опухоли указывает на значимое (p <0,001) трехкратное более высокое значение объемной плотности ложноножек в подкорковой области по сравнению с корковой областью опухоли.
Можно ли распространить наблюдение PAS-положительных эндотелиальных клеток на образцы хирургической биопсии рака молочной железы человека? Чтобы ответить на этот вопрос, были исследованы образцы от 15 пациентов с раком молочной железы, полученные из Центра медицинских наук Техасского университета в отделении патологии Сан-Антонио.
 Из этих 15 пациентов у девяти была диагностирована внутрипротоковая карцинома с ограничением протоков, или DCIC. Хотя в пяти из них было обнаружено многослойность раковых клеток в протоках, ни в одном из них не было обнаружено признаков эндотелиальных псевдоножек в этой популяции раковых клеток.Семь образцов биопсии были диагностированы как инвазивная протоковая карцинома, или «ИДС». Шесть из семи образцов IDS показали наличие эндотелиальных псевдоножек, но в меньшей степени, чем в опухоли молочной железы человека MDA MB231, использованной в текущем исследовании. Два образца опухоли IDS имели видимые участки некроза. В области опухоли, прилегающей к этим областям некроза, было больше ложноножек, чем в других частях опухоли. Таким образом, PAS-положительные псевдоподии эндотелиальных клеток обычно присутствовали в образцах IDS человека.
Из этих 15 пациентов у девяти была диагностирована внутрипротоковая карцинома с ограничением протоков, или DCIC. Хотя в пяти из них было обнаружено многослойность раковых клеток в протоках, ни в одном из них не было обнаружено признаков эндотелиальных псевдоножек в этой популяции раковых клеток.Семь образцов биопсии были диагностированы как инвазивная протоковая карцинома, или «ИДС». Шесть из семи образцов IDS показали наличие эндотелиальных псевдоножек, но в меньшей степени, чем в опухоли молочной железы человека MDA MB231, использованной в текущем исследовании. Два образца опухоли IDS имели видимые участки некроза. В области опухоли, прилегающей к этим областям некроза, было больше ложноножек, чем в других частях опухоли. Таким образом, PAS-положительные псевдоподии эндотелиальных клеток обычно присутствовали в образцах IDS человека.Две новые морфологии псевдоподов, обнаруженные в линии гемопоэтических клеток-предшественников KG1a человека и в первичных CD34+ клетках человека | Кровь
Было обнаружено, что у примитивной человеческой гемопоэтической миелоидной клеточной линии-предшественника, KG1a, характеризующейся высокой экспрессией поверхностного антигена CD34, вытягиваются длинные тонкие псевдоподии.
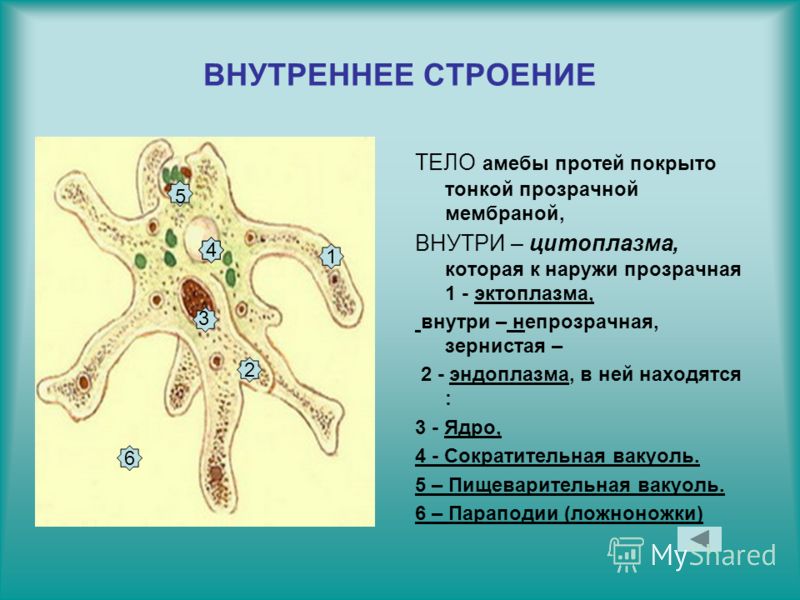 После удлинения эти псевдоподии могут принимать одну из двух недавно описанных морфологий: тенуподии или магноподии.Тенуподии очень тонкие и образуют линейные сегменты. Они прилипают к субстрату, могут многократно раздваиваться и часто соединяют мембраны клеток на расстоянии более 300 мкм друг от друга. Магнуподии намного толще, и было замечено, что они простираются более чем на 330 мкм от клетки. Магнуподы гибкие и могут демонстрировать быстрое динамическое движение, вытягиваясь или втягиваясь за несколько секунд. Во время ретракции расширенный материал часто скапливается в выпуклость, расположенную на стручке. Обе морфологии могут прикрепляться к субстратам, покрытым фибронектином, коллагеном IV и ламинином, а также к пластику.Антигены CD34 и CD44 также присутствуют на поверхности этих ножек. Первичные человеческие клетки CD34 + из печени плода, пуповинной крови, костного мозга взрослых и мобилизованной периферической крови также расширяют эти ножки. Морфология, которую демонстрируют эти ложноножки, позволяет предположить, что они могут играть как сенсорную, так и механическую роль во время миграции клеток и самонаведения после трансплантации костного мозга.
После удлинения эти псевдоподии могут принимать одну из двух недавно описанных морфологий: тенуподии или магноподии.Тенуподии очень тонкие и образуют линейные сегменты. Они прилипают к субстрату, могут многократно раздваиваться и часто соединяют мембраны клеток на расстоянии более 300 мкм друг от друга. Магнуподии намного толще, и было замечено, что они простираются более чем на 330 мкм от клетки. Магнуподы гибкие и могут демонстрировать быстрое динамическое движение, вытягиваясь или втягиваясь за несколько секунд. Во время ретракции расширенный материал часто скапливается в выпуклость, расположенную на стручке. Обе морфологии могут прикрепляться к субстратам, покрытым фибронектином, коллагеном IV и ламинином, а также к пластику.Антигены CD34 и CD44 также присутствуют на поверхности этих ножек. Первичные человеческие клетки CD34 + из печени плода, пуповинной крови, костного мозга взрослых и мобилизованной периферической крови также расширяют эти ножки. Морфология, которую демонстрируют эти ложноножки, позволяет предположить, что они могут играть как сенсорную, так и механическую роль во время миграции клеток и самонаведения после трансплантации костного мозга.
M ЛЮБЫЕ ТИПЫ КЛЕТОК демонстрируют динамическое формирование и втягивание поверхностных расширений по мере их миграции.Эти псевдоподии могут играть сенсорную или механическую роль и обычно классифицируются по их морфологии. Некоторыми примерами являются выпуклые лобоподии, используемые паразитической амебой Entamoeba histolytica , и широкие плоские ламеллоподии, продуцируемые амебой Acanthamoeba castellani .1 Клетки млекопитающих также имеют ложноножки разновидности lamellipodia вместе с сопутствующими микрошипами, похожими на тонкие волоски. отростки диаметром от 0,1 до 0,2 мкм и длиной до 20 мкм.Известно, что Т-лимфоциты образуют уроподы, которые опосредуют межклеточное взаимодействие, 2 в то время как нейриты образуют более длинные микрошипы, известные как филоподии, которые имеют решающее значение для процесса наведения конуса роста.3-6 Филоподии, расширенные нейритами, могут иметь длину до 50 мкм.
 7Тромбоциты также расширяют активные мембранные процессы, при которых динамическое взъерошивание мембраны сопровождается развертыванием микрошипов.8
7Тромбоциты также расширяют активные мембранные процессы, при которых динамическое взъерошивание мембраны сопровождается развертыванием микрошипов.8Клеточная линия острого миелогенного лейкоза, KG1a, была впервые выделена в 1980 году.9 Заметной фенотипической аномалией этих клеток является их неспособность дифференцироваться в функционально зрелые клетки, в результате чего они остаются на ранней или примитивной стадии развития. незрелые гемопоэтические клетки.11 Результатом стало первое антитело против маркера клеточной поверхности CD34, который с тех пор стал маркером выбора при отборе клеток-предшественников для гемопоэтических исследований, а также протоколов обогащения для терапии стволовыми клетками.12
Миграционные и хоуминговые характеристики гемопоэтических стволовых клеток (ГСК) представляют значительный интерес в настоящее время, поскольку эти процессы играют важную роль в приживлении гемопоэтических трансплантатов и восстановлении после трансплантации костного мозга.
 Трансплантация клеток, характеризующихся поверхностным антигеном CD34, вызывает восстановление кроветворения у пациентов, перенесших миелоабляцию. Самонаведение этих клеток в костный мозг представляет собой сложный и плохо изученный процесс, который, вероятно, включает хемокины для навигации 13, а также адгезивные взаимодействия 14-17 для направления их в соответствующие ниши в костном мозге.Известно, что из многих адгезивных взаимодействий, вовлеченных в этот процесс, антиген CD44 связывается с фибронектином, распространенным компонентом внеклеточного матрикса.18 В дополнение к формированию адгезивных прикреплений миграция также требует деформации клеток, расширения цитоплазматических проекций и создания сократительных сил. при движении.19,20 Хотя в принципе клетки CD34 + также должны подвергаться тем же физическим процессам, которые необходимы для движения, мало что известно о том, как они на самом деле выполняют эту задачу.Открытие мобилизации HSCs в кровообращение 21, позволяющее собрать популяцию приживляющихся клеток с помощью лейкофореза, дополнительно иллюстрирует важность понимания миграционных характеристик популяций примитивных гемопоэтических клеток.
Трансплантация клеток, характеризующихся поверхностным антигеном CD34, вызывает восстановление кроветворения у пациентов, перенесших миелоабляцию. Самонаведение этих клеток в костный мозг представляет собой сложный и плохо изученный процесс, который, вероятно, включает хемокины для навигации 13, а также адгезивные взаимодействия 14-17 для направления их в соответствующие ниши в костном мозге.Известно, что из многих адгезивных взаимодействий, вовлеченных в этот процесс, антиген CD44 связывается с фибронектином, распространенным компонентом внеклеточного матрикса.18 В дополнение к формированию адгезивных прикреплений миграция также требует деформации клеток, расширения цитоплазматических проекций и создания сократительных сил. при движении.19,20 Хотя в принципе клетки CD34 + также должны подвергаться тем же физическим процессам, которые необходимы для движения, мало что известно о том, как они на самом деле выполняют эту задачу.Открытие мобилизации HSCs в кровообращение 21, позволяющее собрать популяцию приживляющихся клеток с помощью лейкофореза, дополнительно иллюстрирует важность понимания миграционных характеристик популяций примитивных гемопоэтических клеток.
Мы исследовали миграционные характеристики клеток KG1a в культуре с помощью покадровой флуоресцентной микроскопии. Здесь мы сообщаем о характеристике двух новых морфологий тонких (около 1 мкм или меньше) псевдоподий, тенуподий и магнуподий, проявляемых этими клетками.Тенуподии (от латинского «тонкий» или «тонкий») очень тонкие и состоят из линейных сегментов. Они прилипают к субстрату, могут многократно раздваиваться и часто соединяют мембраны клеток на расстоянии более 300 мкм друг от друга. Магнуподии (от латинского означает большой или длинный) немного толще и, как было замечено, простираются более чем на 330 мкм от клетки. Magnupods являются гибкими и могут демонстрировать быстрое динамическое движение, раскрываясь или убираясь за несколько секунд. Когда они втягиваются, расширенный материал часто скапливается в луковицу, расположенную на стручке.Обе морфологии могут прикрепляться к субстратам, покрытым фибронектином, коллагеном IV и ламинином, а также к пластику, обработанному тканевой культурой.
 Мы наблюдали развертывание этих ножек во время миграции клеток in vitro, предполагая, что они выполняют как сенсорные, так и механические функции. Эти наблюдения могут иметь отношение к роли ножек в приживлении после трансплантации костного мозга.
Мы наблюдали развертывание этих ножек во время миграции клеток in vitro, предполагая, что они выполняют как сенсорные, так и механические функции. Эти наблюдения могут иметь отношение к роли ножек в приживлении после трансплантации костного мозга.Линия гемопоэтических клеток человека KG1a была получена от ATCC (Rockville, MD).Клетки получали криоконсервированными, а затем быстро оттаивали и суспендировали в модифицированной по Искову среде Дульбекко (IMDM; GIBCO-BRL, Гранд-Айленд, Нью-Йорк), содержащей 4 ммоль/л L-глутамина, 1,5 г/л бикарбоната натрия и 20% фетальной бычьей сыворотки. Линия содержалась в инкубаторе при 37°C, влажности 95% и 5% CO 2 . Пассирование проводили каждые 3-4 дня (согласно рекомендациям ATCC). Йодид пропидия (Boehringer Mannheim, Indianapolis, IN) использовали для проверки жизнеспособности клеток в соответствии с рекомендациями производителя.
человека CD34 + клеток были получены из нескольких источников.
Препараты Клетки CD34 + из мобилизованной периферической крови (mPB) были получены от доноров, которые были мобилизованы комбинацией цитокинов гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ) и гранулоцитарного колониестимулирующего фактора (Г-КСФ). Периферическую кровь больных подвергали лейкафорезу и выделяли мононуклеары.Обогащение CD34 проводили с помощью колонки для иммуномагнитного разделения (AmCell, Sunnyvale, CA). Обычно достигалась приблизительно 85% чистота клеток CD34 + . Затем клетки криоконсервировали в жидком азоте. После быстрого оттаивания при 37°C их суспендировали в среде Meylocult H5100 (Stem Cell Technologies Inc., Ванкувер, Британская Колумбия, Канада), содержащей 12,5% фетальной телячьей сыворотки и 12,5% лошадиной сыворотки (без дополнительных гемопоэтических факторов роста), и оставляли для восстановления. за 6-12 часов до окрашивания.
Клетки CD34 + из мобилизованной периферической крови (mPB) были получены от доноров, которые были мобилизованы комбинацией цитокинов гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ) и гранулоцитарного колониестимулирующего фактора (Г-КСФ). Периферическую кровь больных подвергали лейкафорезу и выделяли мононуклеары.Обогащение CD34 проводили с помощью колонки для иммуномагнитного разделения (AmCell, Sunnyvale, CA). Обычно достигалась приблизительно 85% чистота клеток CD34 + . Затем клетки криоконсервировали в жидком азоте. После быстрого оттаивания при 37°C их суспендировали в среде Meylocult H5100 (Stem Cell Technologies Inc., Ванкувер, Британская Колумбия, Канада), содержащей 12,5% фетальной телячьей сыворотки и 12,5% лошадиной сыворотки (без дополнительных гемопоэтических факторов роста), и оставляли для восстановления. за 6-12 часов до окрашивания.Purer CD34 + из печени плода (FL), пуповинной крови (UCB) и костного мозга взрослого человека (aBM) получали с помощью проточной цитометрической сортировки (FACStar Plus; Becton Dickinson, San Jose, CA).
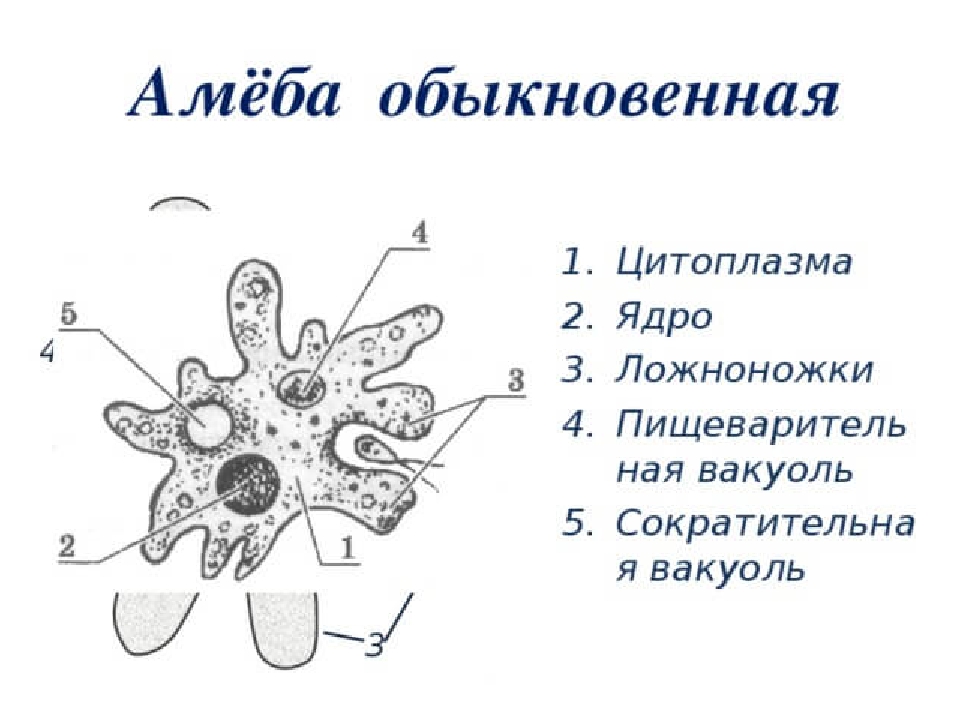 Используемое антитело к CD34, конъюгированное с флуоресцеинизотиоцианатом (FITC), представляло собой анти-HCPA-2 (клон 8G12; Becton Dickinson). Эти клетки суспендируют в той же среде Meylocult и используют в свежем виде сразу после сортировки.Количество жизнеспособных клеток было увеличено путем сортировки мертвых клеток на основе их характеристик светорассеяния.
Используемое антитело к CD34, конъюгированное с флуоресцеинизотиоцианатом (FITC), представляло собой анти-HCPA-2 (клон 8G12; Becton Dickinson). Эти клетки суспендируют в той же среде Meylocult и используют в свежем виде сразу после сортировки.Количество жизнеспособных клеток было увеличено путем сортировки мертвых клеток на основе их характеристик светорассеяния.Для визуализации клетки окрашивали флуоресцентным мембранным красителем РХ36 (Zynaxis, Malvern, PA), который возбуждает при длине волны 551 нм и излучает при 567 нм. Процедура окрашивания включает алифатические репортерные молекулы в клеточную мембрану, поэтому обнаруживаемая флуоресценция зависит от количества мембраны в любом заданном месте.22,23 Клетки окрашивали в соответствии с рекомендациями производителя. Вкратце, около 1 млн клеток суспендировали в 1 мл разбавителя С. ПХ36 разводили в разбавителе С до соотношения 8 мкл красителя на 1 мл разбавителя С.
 Разбавленный краситель объединяли с клетками (конечная концентрация 4 мкмоль/л). L) в течение 6 минут, после чего для остановки реакции окрашивания добавляли равный объем фосфатно-солевого буфера Дульбекко (PBS; GIBCO-BRL), содержащего 1% лепталба-7 (Intergen, Purchase, NY). Через 1 минуту добавляли равный объем свежей среды.Затем клетки центрифугировали при 400 g в течение 10 минут и еще два раза промывали в PBS или свежей среде.
Разбавленный краситель объединяли с клетками (конечная концентрация 4 мкмоль/л). L) в течение 6 минут, после чего для остановки реакции окрашивания добавляли равный объем фосфатно-солевого буфера Дульбекко (PBS; GIBCO-BRL), содержащего 1% лепталба-7 (Intergen, Purchase, NY). Через 1 минуту добавляли равный объем свежей среды.Затем клетки центрифугировали при 400 g в течение 10 минут и еще два раза промывали в PBS или свежей среде.Иммуноокрашивание на поверхностные антигены проводили по следующей методике. Клетки KG1a промывали в PBS (GIBCO-BRL), а затем ресуспендировали в изотоническом растворе сахарозы с концентрацией 295 мОсм, чтобы избежать образования кристаллов соли при высыхании среды. Пятьдесят микролитров клеточной суспензии сушили на предметном стекле.Сушку выполняли для сохранения хрупких структур стручков, поскольку обычные протоколы фиксации приводили к повреждению и отсутствию стручков.
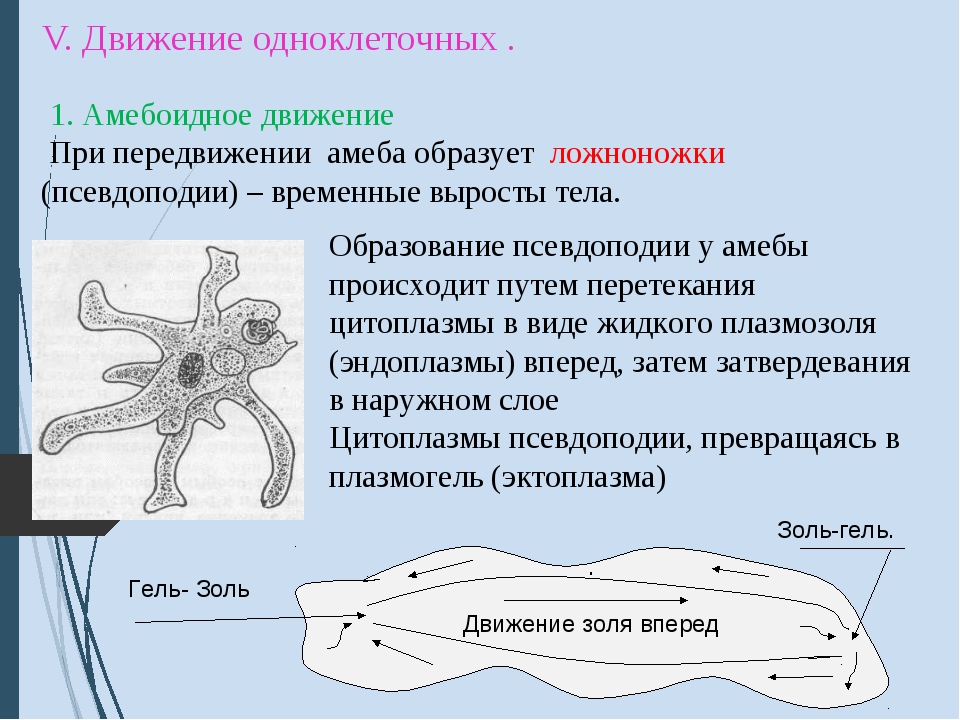 Затем на высушенные клетки наносили 50 мкл разведения 1:100 либо CD34 (конъюгированного с фикоэритрином анти-HCPA-2; Becton Dickinson), либо CD44 (конъюгированного с фикоэритрином; Pharmingen, San Diego, CA) в изотоническом растворе сахарозы. и инкубировали при 4°C в течение ночи перед визуализацией на следующий день. В качестве отрицательного контроля в той же процедуре использовали антитела изотипического контроля, конъюгированные с фикоэритрином (мышиный IgG2 b для CD44 и мышиный IgG 1 для CD34; Pharmingen).
Затем на высушенные клетки наносили 50 мкл разведения 1:100 либо CD34 (конъюгированного с фикоэритрином анти-HCPA-2; Becton Dickinson), либо CD44 (конъюгированного с фикоэритрином; Pharmingen, San Diego, CA) в изотоническом растворе сахарозы. и инкубировали при 4°C в течение ночи перед визуализацией на следующий день. В качестве отрицательного контроля в той же процедуре использовали антитела изотипического контроля, конъюгированные с фикоэритрином (мышиный IgG2 b для CD44 и мышиный IgG 1 для CD34; Pharmingen).Клетки визуализировали с помощью инвертированного флуоресцентного микроскопа Diaphot 300 (Nikon Inc, Мелвилл, штат Нью-Йорк) с объективами 20×, 40× и 100×. Изображения ПХ36 были получены с использованием набора флуоресцентных фильтров 41003 (Chroma Technologies, Brattleboro, VT). Оцифрованные изображения получали с помощью охлаждаемой ПЗС-камеры (Photometrics Inc, Tucson, AZ) и сохраняли на рабочей станции SGI O 2 (Silicon Graphics, Mountain View, CA).
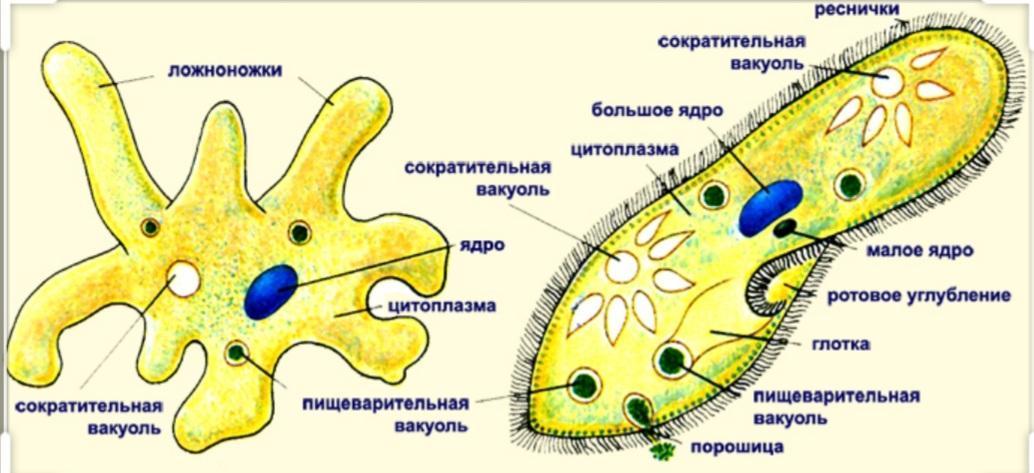 Все функции сбора и обработки контролировались программным обеспечением Isee (Inovision Corp, Дарем, Северная Каролина), которое обеспечивало полную автоматизацию системы.
Все функции сбора и обработки контролировались программным обеспечением Isee (Inovision Corp, Дарем, Северная Каролина), которое обеспечивало полную автоматизацию системы.Клетки содержались при постоянной температуре 37°C в инкубаторе, окружающем микроскоп. Инкубатор состоял из большой, герметичной, изготовленной на заказ коробки из плексигласа, обогреваемой вентилятором теплого воздуха, управляемым регулятором температуры PID (Cole-Parmer, Vernon Hills, IL).Контроль pH примерно до 7,4 был достигнут путем промывки коробки из плексигласа CO 2 . Контроллер постоянно обнаруживал CO 2 в коробке и поддерживал его концентрацию на уровне 5%. Испарение среды было снижено приблизительно до 2% в день путем заполнения соседних лунок водой и обертывания многолуночных планшетов парафильмом (American National Can, Гринвич, Коннектикут).
Клетки высевали с различной плотностью на различные подложки в пластиковых чашках для культур тканей, 96-луночных планшетах (Costar, Cambridge, MA), предметных и покровных стеклах микроскопа.
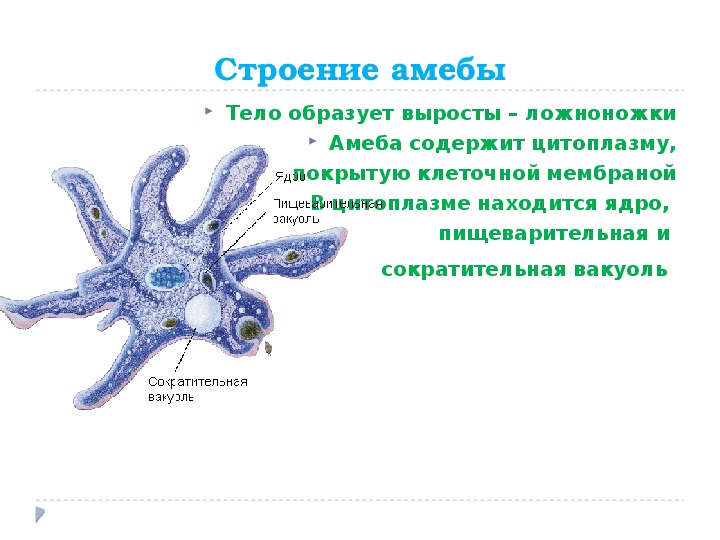 Другими протестированными поверхностями были покровные стекла, покрытые человеческим фибронектином, пластиковые чашки диаметром 35 мм и 96-луночные планшеты, покрытые мышиным коллагеном IV, мышиным ламинином и человеческим фибронектином (Becton Dickinson, Bedford, MA).
Другими протестированными поверхностями были покровные стекла, покрытые человеческим фибронектином, пластиковые чашки диаметром 35 мм и 96-луночные планшеты, покрытые мышиным коллагеном IV, мышиным ламинином и человеческим фибронектином (Becton Dickinson, Bedford, MA).Другие эксперименты включали манипулирование ячейками после того, как они развернули свои стручки. Систему микроманипуляторов (Eppendorf, Гамбург, Германия) использовали для размещения стандартной капиллярной трубки диаметром 1,1 мм (внешний диаметр) в лунке ячейки.Трубку изгибали под углом 90° примерно на 18 мм от конца (чтобы она могла поместиться в глубокие лунки небольшого диаметра) путем нагревания ее горелкой Бунзена. Этот процесс запрещал использование наконечников микропипеток, потому что высокая температура вызывала плавление тонкого стеклянного наконечника. С помощью шприца среду осторожно втягивали в пипетку, а затем выталкивали, заставляя ножки испытывать механические усилия из-за движения жидкости.

Когда клетки KG1a высевают и инкубируют в течение примерно 1 часа при 37°C, они начинают развертывать длинные тонкие стручки.Клетка KG1a, показанная на рис. 1, развернула магнупод, который простирался примерно на 94 мкм от клетки к соседней клетке. Magnupods имеют тенденцию плавать в среде и, если есть какое-либо движение жидкости, будут распространяться вниз по течению, в то время как клетка остается прикрепленной к субстрату. Это явление наиболее очевидно, когда каплю клеточной суспензии объемом 10 мкл помещают на предметное стекло. Когда среда испаряется, жидкость движется к краю капли из-за капиллярного потока, вызывающего радиальный ток.24 Клетки у края развертывают магноподы, направленные радиально наружу.
Рис. 1.
Клетка KG1a, проецирующая магнопод длиной примерно 94 мкм в сторону соседней клетки.
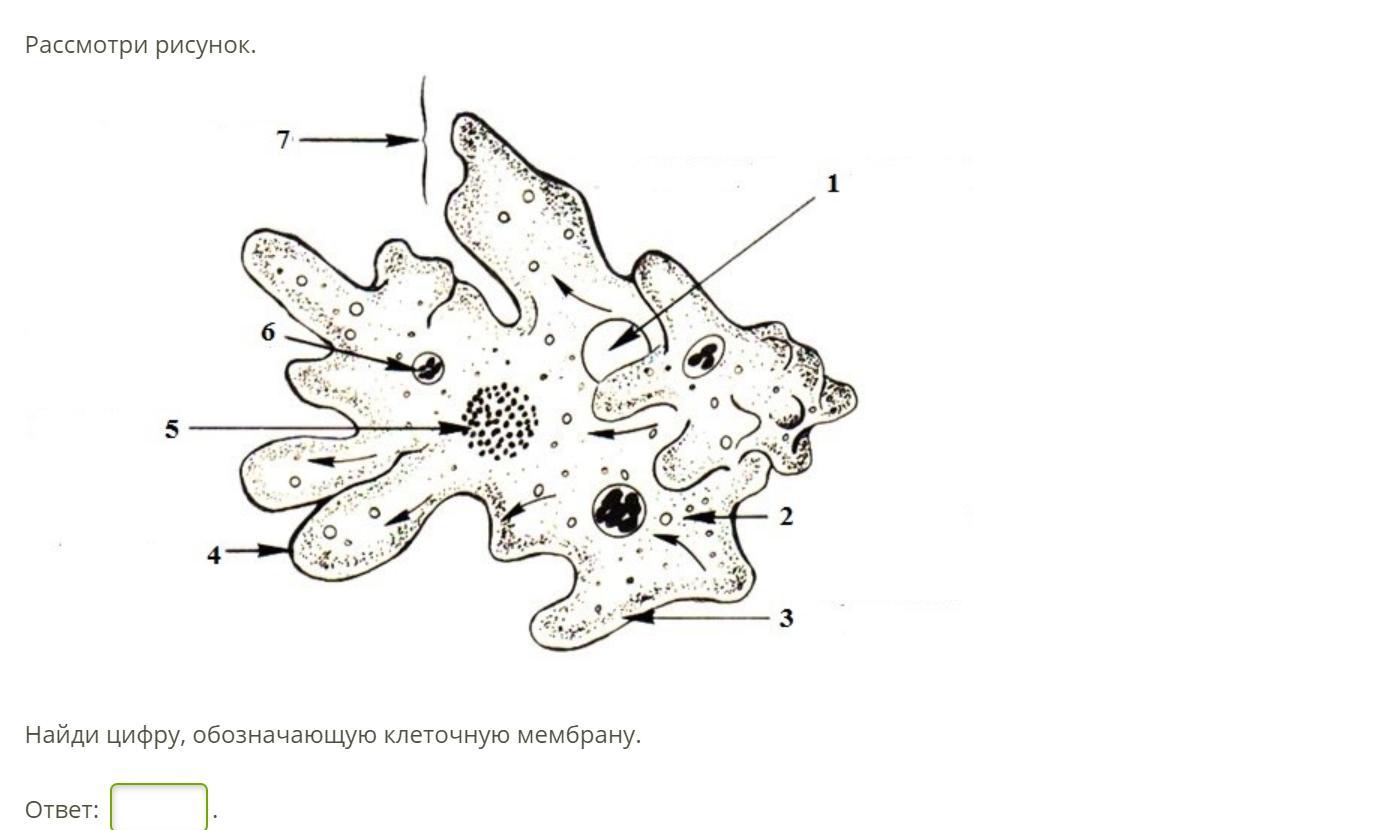 Поверхность покрывали фибронектином и использовали объектив 20×. Масштабная линейка имеет длину 50 мкм. Контрастность изображения повышена, чтобы были видны подиумы. На рисунок были вставлены неулучшенные изображения ячеек, чтобы реальный размер ячеек был более очевиден.
Поверхность покрывали фибронектином и использовали объектив 20×. Масштабная линейка имеет длину 50 мкм. Контрастность изображения повышена, чтобы были видны подиумы. На рисунок были вставлены неулучшенные изображения ячеек, чтобы реальный размер ячеек был более очевиден.Рис.1.
Клетка KG1a, проецирующая крупноногтевую пластинку длиной примерно 94 мкм в сторону соседней клетки. Поверхность покрывали фибронектином и использовали объектив 20×. Масштабная линейка имеет длину 50 мкм. Контрастность изображения повышена, чтобы были видны подиумы. На рисунок были вставлены неулучшенные изображения ячеек, чтобы реальный размер ячеек был более очевиден.
Magnupods способны быстро втягиваться, как показано на временной последовательности рис. 2a–d.Более короткие крупноножки (<20 мкм) могут втягиваться в тело клетки за считанные секунды, в то время как более длинные ножки объединяют мембрану в луковицы, которые формируются по длине стручка.
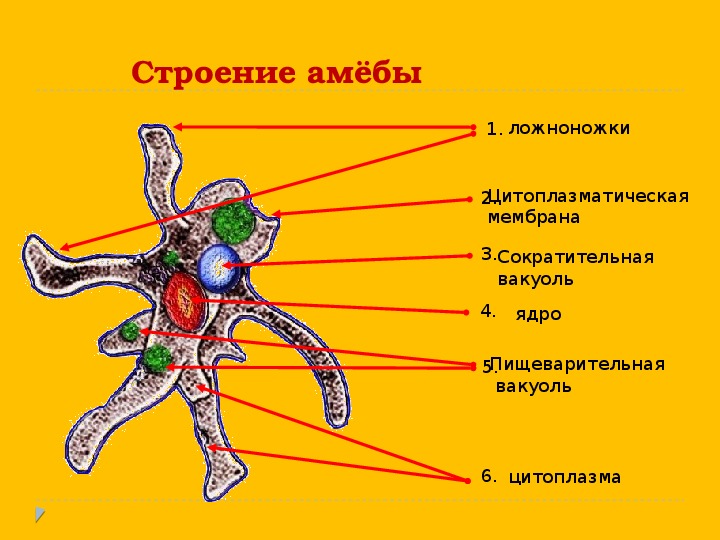 Было замечено, что ячейка, показанная на рис. 2, втягивает свой мегапод со скоростью 0,4 мкм/с. Выдвижение и втягивание магнуподов не является непрерывным процессом. После того, как магнупод выдвинулся, нередко он остается развернутым в течение нескольких часов. Стимул, ответственный за вытягивание или втягивание крупноногих позвоночников, в настоящее время неизвестен.Другими интересными особенностями морфологии большеноногих являются то, что они могут распространяться на большие расстояния от клетки, как видно на рис. 3, где показана общая длина большеножек более 330 мкм. Кончик магнупода не в фокусе, что указывает на то, что он свободно парит над поверхностью. Однако магнуподы также способны прикрепляться к субстрату. Их можно определить, применяя механические усилия с помощью микропипетки. Прикрепление определяли с помощью микропипетки, чтобы вызвать поток жидкости вокруг клетки.Ячейка и стручок деформируются и могут уплыть, за исключением случаев, когда они прикреплены к субстрату (данные не показаны).
Было замечено, что ячейка, показанная на рис. 2, втягивает свой мегапод со скоростью 0,4 мкм/с. Выдвижение и втягивание магнуподов не является непрерывным процессом. После того, как магнупод выдвинулся, нередко он остается развернутым в течение нескольких часов. Стимул, ответственный за вытягивание или втягивание крупноногих позвоночников, в настоящее время неизвестен.Другими интересными особенностями морфологии большеноногих являются то, что они могут распространяться на большие расстояния от клетки, как видно на рис. 3, где показана общая длина большеножек более 330 мкм. Кончик магнупода не в фокусе, что указывает на то, что он свободно парит над поверхностью. Однако магнуподы также способны прикрепляться к субстрату. Их можно определить, применяя механические усилия с помощью микропипетки. Прикрепление определяли с помощью микропипетки, чтобы вызвать поток жидкости вокруг клетки.Ячейка и стручок деформируются и могут уплыть, за исключением случаев, когда они прикреплены к субстрату (данные не показаны). Однако эти прикрепления не являются непрерывными вдоль магнупода, что свидетельствует о неравномерном распределении комплексов адгезии.
Однако эти прикрепления не являются непрерывными вдоль магнупода, что свидетельствует о неравномерном распределении комплексов адгезии.Рис. 2.
Ячейка KG1a, втягивающая магнитопод. Четыре изображения были сделаны в различные промежутки времени. Изображение (а) было сделано при t = 0, (б) при t = 14 секунд, (в) при t = 75 секунд и (г) при t = 150 секунд.Соответствующие длины магнупода составляют 77, 58, 42 и 14 мкм соответственно, что дает скорость втягивания примерно 0,4 мкм/с. Масштабная линейка имеет длину 50 мкм.
Рис. 2.
Ячейка KG1a, втягивающая магнупод. Четыре изображения были сделаны в различные промежутки времени. Изображение (а) было сделано при t = 0, (б) при t = 14 секунд, (в) при t = 75 секунд и (г) при t = 150 секунд. Соответствующие длины магнупода составляют 77, 58, 42 и 14 мкм соответственно, что дает скорость втягивания примерно 0.4 мкм/с. Масштабная линейка имеет длину 50 мкм.
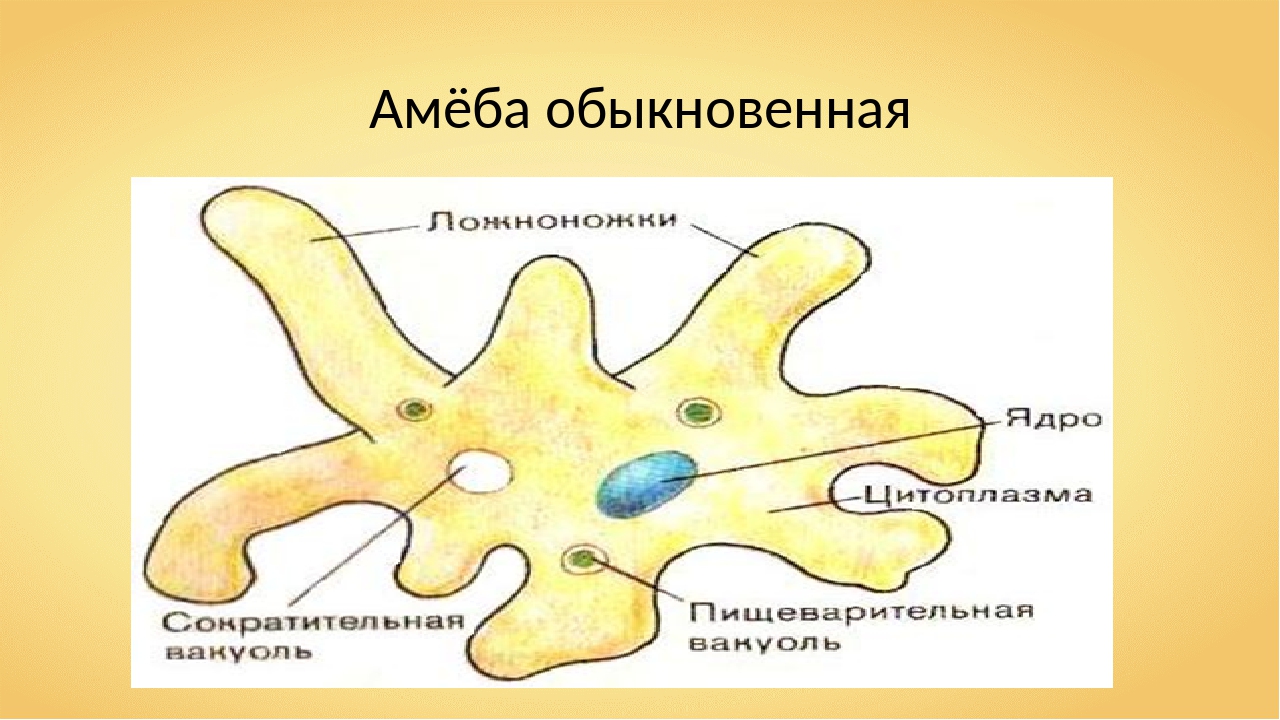
Рис. 3.
Две клетки KG1a с очень длинными магноподиями. Общая длина ножек, расширенных клеткой слева, составляет более 330 мкм. Кончик одного магнопода не в фокусе, что указывает на то, что он парит над плоскостью фокуса. Масштабная линейка на этом изображении имеет длину 100 мкм. Повышение контрастности этого изображения, чтобы ножки стали видны, также привело к тому, что фрагменты мембраны и обломки стали выглядеть как яркие объекты.
Рис. 3.
Две клетки KG1a с чрезвычайно длинными крупноногими ножками. Общая длина ножек, расширенных клеткой слева, составляет более 330 мкм. Кончик одного магнопода не в фокусе, что указывает на то, что он парит над плоскостью фокуса. Масштабная линейка на этом изображении имеет длину 100 мкм. Повышение контрастности этого изображения, чтобы ножки стали видны, также привело к тому, что фрагменты мембраны и обломки стали выглядеть как яркие объекты.
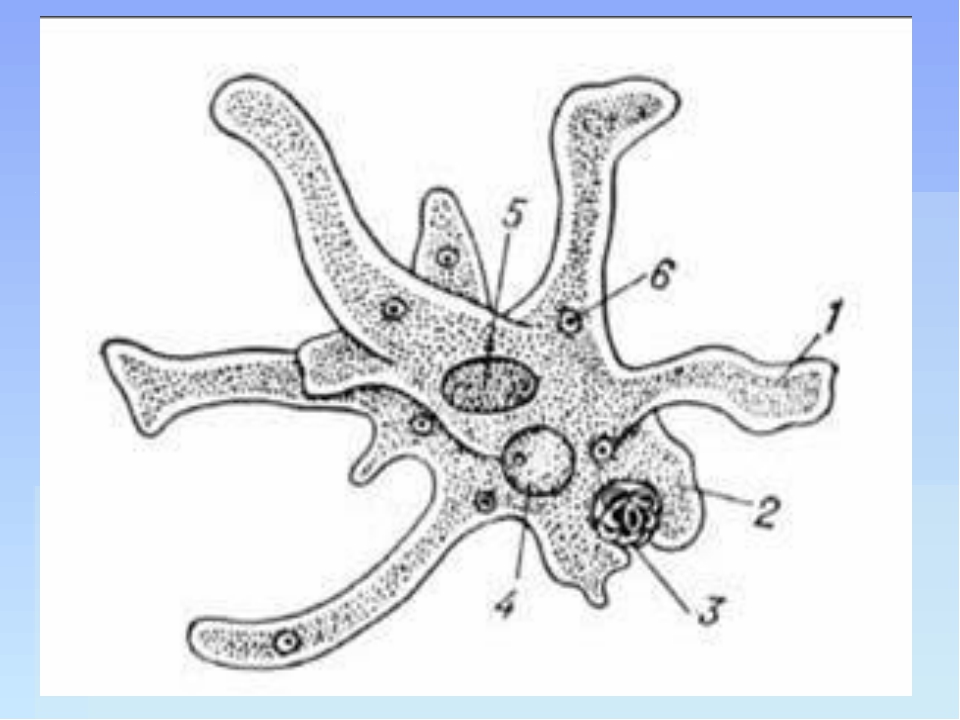
Tenupodia являются второй новой морфологией стручка, обнаруженной на клетках KG1a.Тенуподии морфологически существенно отличаются от магнуподий и ведут себя иначе. Пример образования тенуподов показан на рис. 4. На рис. 4а показана клетка KG1a, которая прорастает тениподами на субстрате, покрытом ламинином (вверху вторая клетка KG1a, проецирующая магнупод). Tenupod выглядит как линейные сегменты, которые развертываются из общей точки происхождения на клеточной поверхности. Они отходят от тела клетки и могут разветвляться или поворачиваться в определенных точках. Один из тенуподов, столкнувшись с 6.Пластиковый шарик диаметром 4 мкм перемещается вокруг него, раздваиваясь. Это бифуркационное поведение также видно на рис. 4b, на котором скопление тениподов отходит от одной клетки. Тенуподии более распространены на поверхностях, покрытых фибронектином, ламинином и коллагеном IV, чем на стекле или пластике, что свидетельствует о том, что взаимодействие с поверхностью играет важную роль в определении морфологии ножек.
 Наконец, было замечено, что тенуподы соединяют мембраны двух разных клеток, расположенных на расстоянии более 300 мкм друг от друга.Типичный пример такого поведения показан на рис. 5, на котором показаны тенуподы, простирающиеся линейным образом непосредственно к соседней ячейке. В противоположность этому, несколько тенуподов из клетки на рис. 4а развертывались по более окольному маршруту, когда они сходились в области в правом верхнем углу клетки, меняя направление, по-видимому, неслучайным образом, как будто какой-то фактор действовал, чтобы смещать направление их расширение. Взятые вместе, эти наблюдения предполагают, что тенуподии чувствуют сигналы окружающей среды и реагируют на них.Подобные способности позволили бы тенуподам нацеливаться на отдаленный источник, секретирующий растворимый фактор, или следовать по следу молекул адгезии, связанных с внеклеточным матриксом.
Наконец, было замечено, что тенуподы соединяют мембраны двух разных клеток, расположенных на расстоянии более 300 мкм друг от друга.Типичный пример такого поведения показан на рис. 5, на котором показаны тенуподы, простирающиеся линейным образом непосредственно к соседней ячейке. В противоположность этому, несколько тенуподов из клетки на рис. 4а развертывались по более окольному маршруту, когда они сходились в области в правом верхнем углу клетки, меняя направление, по-видимому, неслучайным образом, как будто какой-то фактор действовал, чтобы смещать направление их расширение. Взятые вместе, эти наблюдения предполагают, что тенуподии чувствуют сигналы окружающей среды и реагируют на них.Подобные способности позволили бы тенуподам нацеливаться на отдаленный источник, секретирующий растворимый фактор, или следовать по следу молекул адгезии, связанных с внеклеточным матриксом.Рис. 4.
(а) Клетки KG1a, демонстрирующие обе морфологии ножек.
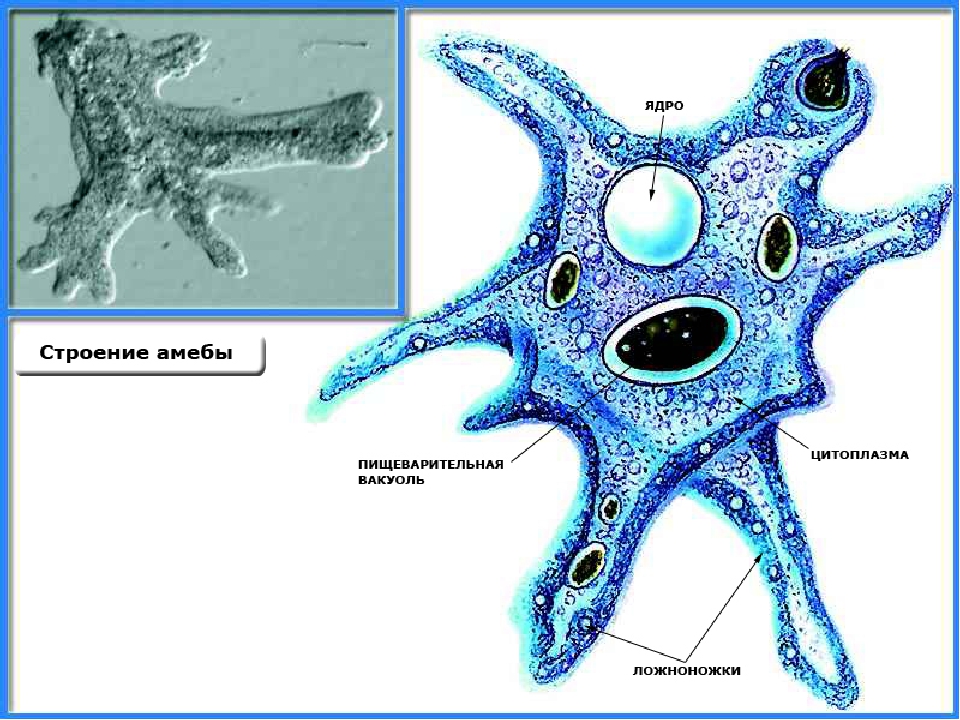 Клетка в верхнем левом углу имеет расширенный магнуподий длиной 114 мкм, тогда как в другой клетке развернуты множественные тенуподии. Один тенупод вытянулся в направлении 6.Пластиковый шарик диаметром 4 мкм перед тем, как раздвоиться и отправиться вправо от изображения, где он, по-видимому, обнаружил притягивающий его фактор. Масштабная линейка имеет длину 100 мкм. (b) На этом врезном изображении показана клетка KG1a с сильно разветвленной сетью тенуподов, которая простирается к соседней клетке KG1a, которая мигрировала через ламеллоподии. Масштабная линейка имеет длину 50 мкм.
Клетка в верхнем левом углу имеет расширенный магнуподий длиной 114 мкм, тогда как в другой клетке развернуты множественные тенуподии. Один тенупод вытянулся в направлении 6.Пластиковый шарик диаметром 4 мкм перед тем, как раздвоиться и отправиться вправо от изображения, где он, по-видимому, обнаружил притягивающий его фактор. Масштабная линейка имеет длину 100 мкм. (b) На этом врезном изображении показана клетка KG1a с сильно разветвленной сетью тенуподов, которая простирается к соседней клетке KG1a, которая мигрировала через ламеллоподии. Масштабная линейка имеет длину 50 мкм.Рис. 4.
(а) Клетки KG1a, демонстрирующие обе морфологии ножек. Клетка в верхнем левом углу имеет расширенный магнуподий длиной 114 мкм, тогда как в другой клетке развернуты множественные тенуподии.Один тенупод протянулся в направлении пластиковой бусинки размером 6,4 мкм, прежде чем раздвоиться и направиться вправо от изображения, где он, по-видимому, обнаружил притягивающий его фактор. Масштабная линейка имеет длину 100 мкм.
 (b) На этом врезном изображении показана клетка KG1a с сильно разветвленной сетью тенуподов, которая простирается к соседней клетке KG1a, которая мигрировала через ламеллоподии. Масштабная линейка имеет длину 50 мкм.
(b) На этом врезном изображении показана клетка KG1a с сильно разветвленной сетью тенуподов, которая простирается к соседней клетке KG1a, которая мигрировала через ламеллоподии. Масштабная линейка имеет длину 50 мкм.Рис.5.
Тенуподы могут простираться к другим клеткам и, по-видимому, соединять мембраны. На этом изображении тенуподии соединяют мембраны трех соседних клеток. Длина самого длинного тенупода составляет 310 мкм. Масштабная линейка имеет длину 100 мкм.
Рис. 5.
Тенуподы могут простираться к другим клеткам и, по-видимому, соединять мембраны. На этом изображении тенуподии соединяют мембраны трех соседних клеток. Длина самого длинного тенупода составляет 310 мкм. Масштабная линейка имеет длину 100 мкм.
Первичные человеческие клетки CD34 + также демонстрируют такое же поведение при формировании ножек, как и клетки KG1a.
 Изображения на рис. 6a–c показывают клетки CD34 + из FL, UCB и aBM, демонстрирующие как большие, так и тенуподии. Аналогичным образом ведут себя и мобилизованные клетки периферической крови CD34 + (данные не представлены). В более чем 40 экспериментах с первичными клетками мы наблюдали образование ножек более чем в 75% экспериментов. Формирование Magnupod и tenupod первичными клетками не было таким последовательным, как у клеточной линии KG1a, возможно, из-за изменчивости донора25 или чувствительности к обработке клеток, подготовке и условиям культивирования.Интересно, что эти морфологии ножек, по-видимому, являются общими для всех источников первичных клеток CD34 + , которые мы тестировали до сих пор.
Изображения на рис. 6a–c показывают клетки CD34 + из FL, UCB и aBM, демонстрирующие как большие, так и тенуподии. Аналогичным образом ведут себя и мобилизованные клетки периферической крови CD34 + (данные не представлены). В более чем 40 экспериментах с первичными клетками мы наблюдали образование ножек более чем в 75% экспериментов. Формирование Magnupod и tenupod первичными клетками не было таким последовательным, как у клеточной линии KG1a, возможно, из-за изменчивости донора25 или чувствительности к обработке клеток, подготовке и условиям культивирования.Интересно, что эти морфологии ножек, по-видимому, являются общими для всех источников первичных клеток CD34 + , которые мы тестировали до сих пор.Рис. 6.
Первичные клетки CD34 + человека также обнаруживают большие и тенуподии. ( а ) Клетка CD34 + из источника пуповинной крови, демонстрирующая магнупод.
 ( б ) Клетка CD34 + печени плода человека с очень длинным тенуподом. (c) ячейка aBM CD34 + с развертыванием длинного магнупода.Масштабные линейки на всех изображениях имеют длину 30 мкм.
( б ) Клетка CD34 + печени плода человека с очень длинным тенуподом. (c) ячейка aBM CD34 + с развертыванием длинного магнупода.Масштабные линейки на всех изображениях имеют длину 30 мкм.Рис. 6.
Первичные человеческие клетки CD34 + также обнаруживают большие и тенуподии. ( а ) Клетка CD34 + из источника пуповинной крови, демонстрирующая магнупод. ( б ) Клетка CD34 + печени плода человека с очень длинным тенуподом. (c) ячейка aBM CD34 + с развертыванием длинного магнупода. Масштабные линейки на всех изображениях имеют длину 30 мкм.
Поверхностные антигены CD34 и CD44 также связаны с образованием ножек в клетках KG1a, как показано на рис. 7a и b.Великолепный подиум на рис. 7а ярко окрашен на антиген CD34, тогда как окрашивание на CD44, известную молекулу поверхностной адгезии, также показало его присутствие на поверхности ножек на рис.
 7b. Отрицательные контроли всегда были тусклыми, и не наблюдалось обнаруживаемого неспецифического связывания. Тот факт, что эти подии были обнаружены при окрашивании антителами, а также на неокрашенных клетках при большом увеличении, указывает на то, что они не являются артефактом окрашивания мембраны красителем ПХ36. На момент наблюдения примерно 95% клеток KG1a были жизнеспособны.Первичные клетки не проверяли на жизнеспособность. Клетки KG1a, которые были убиты нагреванием до 65°C в течение 15 минут, не развернули никаких ножек, показывая, что, как и ожидалось, их расширение является активным процессом.
7b. Отрицательные контроли всегда были тусклыми, и не наблюдалось обнаруживаемого неспецифического связывания. Тот факт, что эти подии были обнаружены при окрашивании антителами, а также на неокрашенных клетках при большом увеличении, указывает на то, что они не являются артефактом окрашивания мембраны красителем ПХ36. На момент наблюдения примерно 95% клеток KG1a были жизнеспособны.Первичные клетки не проверяли на жизнеспособность. Клетки KG1a, которые были убиты нагреванием до 65°C в течение 15 минут, не развернули никаких ножек, показывая, что, как и ожидалось, их расширение является активным процессом.Рис. 7.
(а) Клетка KG1a, показывающая магнупод, окрашенную антителом против поверхностного маркера CD34. Масштабная линейка имеет длину 50 мкм. ( б ) Аналогичная клетка KG1a, окрашенная на наличие антигена CD44 на ее поверхности.Масштабная линейка имеет длину 100 мкм. Обратите внимание, что окрашивание антителами тел клеток на этих флуоресцентных изображениях не такое яркое, как окрашивание ПКХ36, что приводит к более равномерному усилению изображений.
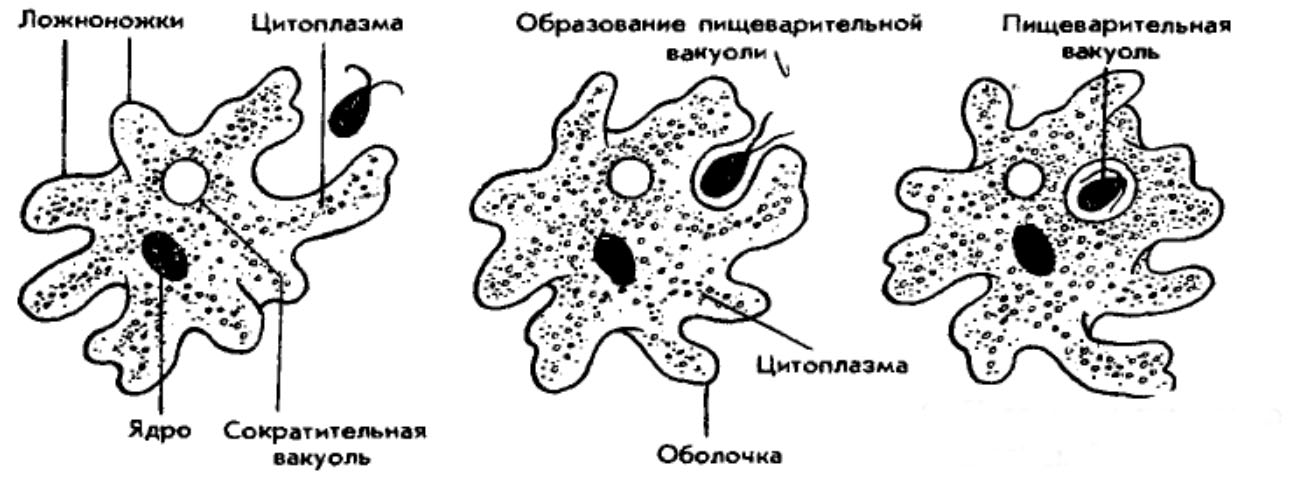
Рис. 7.
(а) Клетка KG1a, показывающая магнупод, окрашенную антителом против поверхностного маркера CD34. Масштабная линейка имеет длину 50 мкм. ( б ) Аналогичная клетка KG1a, окрашенная на наличие антигена CD44 на ее поверхности. Масштабная линейка имеет длину 100 мкм. Обратите внимание, что окрашивание антителами тел клеток на этих флуоресцентных изображениях не такое яркое, как окрашивание ПКХ36, что приводит к более равномерному усилению изображений.
Многие типы клеток демонстрируют динамическое расширение поверхности, когда они мигрируют или изменяют форму. Такие удлинения способны к динамическому формированию и ретракции с типичными скоростями для микрошипов и ламеллиподий порядка 0,1 мкм/сек. Считается, что в нейритах филоподии играют роль в развитии удлинения аксонов, способствуя сборке микротрубочек.26 Эти филоподии простираются от ламеллоподиальной области, действуя как радиальные сенсоры, и имеют решающее значение для навигации по конусам роста.
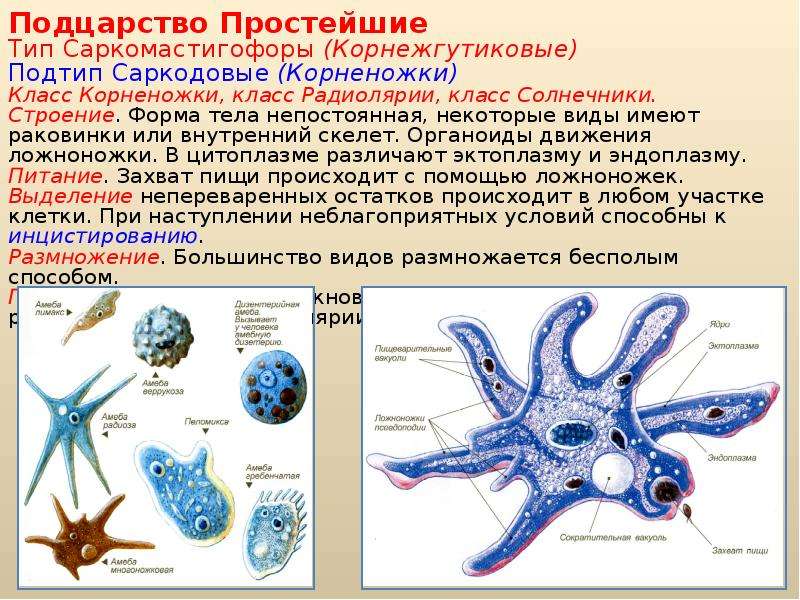 3-6 Было также обнаружено, что они несут рецепторы для определенных молекул клеточной адгезии. 27 Зрелые клетки крови также могут развертывать цитоплазматические отростки. Недавно на Т-лимфоцитах были обнаружены структуры, называемые уроподами, которые формируются во время взаимодействия лимфоцитов с эндотелиальными клетками. Было обнаружено, что Т-клетки используют уроподы для контакта и прямого общения с другими Т-клетками. Развитию уропод способствовали физиологические факторы, такие как хемокины. Таким образом, псевдоподии могут проявлять различные динамические и морфологические свойства, выполняя при этом спектр функций во многих различных типах клеток, связанных с миграцией и коммуникацией.
3-6 Было также обнаружено, что они несут рецепторы для определенных молекул клеточной адгезии. 27 Зрелые клетки крови также могут развертывать цитоплазматические отростки. Недавно на Т-лимфоцитах были обнаружены структуры, называемые уроподами, которые формируются во время взаимодействия лимфоцитов с эндотелиальными клетками. Было обнаружено, что Т-клетки используют уроподы для контакта и прямого общения с другими Т-клетками. Развитию уропод способствовали физиологические факторы, такие как хемокины. Таким образом, псевдоподии могут проявлять различные динамические и морфологические свойства, выполняя при этом спектр функций во многих различных типах клеток, связанных с миграцией и коммуникацией.Мы обнаружили, что клетки примитивной кроветворной миелоидной линии клеток-предшественников человека, KG1a, способны расширять две принципиально новые морфологии псевдоподий. Эти ножки (1) проявляют динамическое поведение растяжения и ретракции, (2) могут прикрепляться к субстрату, по которому они мигрируют, (3) по-видимому, управляются стимулами окружающей среды и (4) имеют антигены для CD34 и антител к CD44 на своей поверхности.
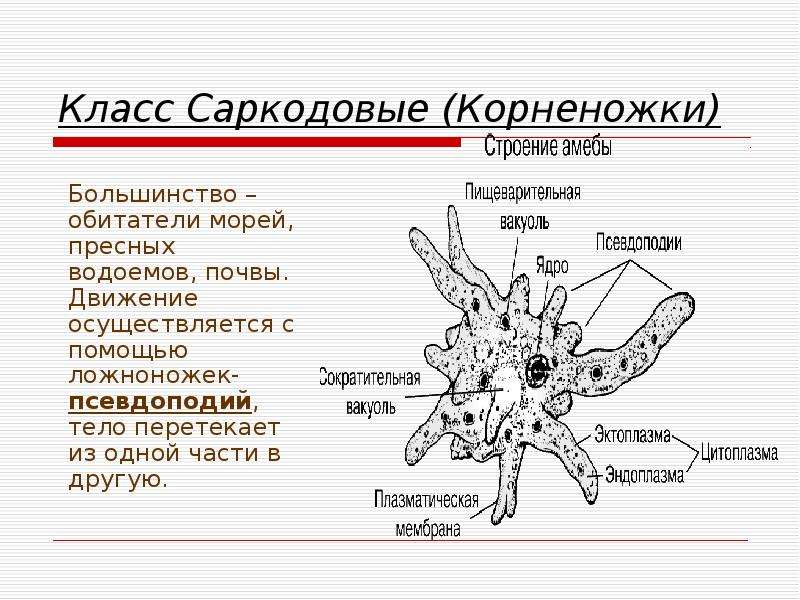 .Однако, магнуподии и тенуподии представляют собой совершенно разные морфологии, демонстрируемые клетками KG1a, и поэтому могут выполнять специализированные миграционные функции, которые уникальны для примитивных гемопоэтических клеток, такие как самонаведение.
.Однако, магнуподии и тенуподии представляют собой совершенно разные морфологии, демонстрируемые клетками KG1a, и поэтому могут выполнять специализированные миграционные функции, которые уникальны для примитивных гемопоэтических клеток, такие как самонаведение.Magnupods имеют морфологическое сходство с длинными (50 мкм) и тонкими (0,2 мкм) филоподиями, обнаруженными на нейритах.28,29 Однако они отличаются тем, что расширение Magnupod происходит на гораздо большие расстояния (>300 мкм, или примерно 30 диаметров клеток). от основного тела клетки, и они демонстрируют быстрое динамическое поведение, втягиваясь более чем в восемь раз быстрее, чем нейритные филоподии.Magnupods могут оставаться вытянутыми в течение нескольких часов, тогда как нейритные филоподии непрерывно вытягиваются и втягиваются. Было предположено, что переключение между удлинением и ретракцией нейритных филоподий отражает сдвиг в скорости полимеризации и деполимеризации актина.
 29 Это различие в динамических характеристиках может указывать на структурные, а также функциональные различия между филоподиями и магноподиями. В то время как филоподии, по-видимому, действуют в первую очередь как сенсорные придатки нейритов, крупноногие могут играть более механическую роль в миграции, подобно ламеллоподиям.Magnupods могут прилипать к субстрату в определенных точках, и недавно сообщалось, что миграционные свойства клеток KG1a усиливаются, когда они ползают по субстрату, покрытому фибронектином.30 Можно предположить, что развернутый Magnupod свободно дрейфует до тех пор, пока его поверхность не прилипнет комплексы сталкиваются с подходящими сайтами связывания на субстрате. Затем ретракция будет тянуть тело клетки к месту адгезии. Кроме того, втягиванию также могут препятствовать многочисленные точечные крепления, в результате чего материал ножек скапливается в луковицы по длине магнупода.Такое динамическое поведение делает морфологию крупноногих моллюсков уникально отличными от филоподий.
29 Это различие в динамических характеристиках может указывать на структурные, а также функциональные различия между филоподиями и магноподиями. В то время как филоподии, по-видимому, действуют в первую очередь как сенсорные придатки нейритов, крупноногие могут играть более механическую роль в миграции, подобно ламеллоподиям.Magnupods могут прилипать к субстрату в определенных точках, и недавно сообщалось, что миграционные свойства клеток KG1a усиливаются, когда они ползают по субстрату, покрытому фибронектином.30 Можно предположить, что развернутый Magnupod свободно дрейфует до тех пор, пока его поверхность не прилипнет комплексы сталкиваются с подходящими сайтами связывания на субстрате. Затем ретракция будет тянуть тело клетки к месту адгезии. Кроме того, втягиванию также могут препятствовать многочисленные точечные крепления, в результате чего материал ножек скапливается в луковицы по длине магнупода.Такое динамическое поведение делает морфологию крупноногих моллюсков уникально отличными от филоподий.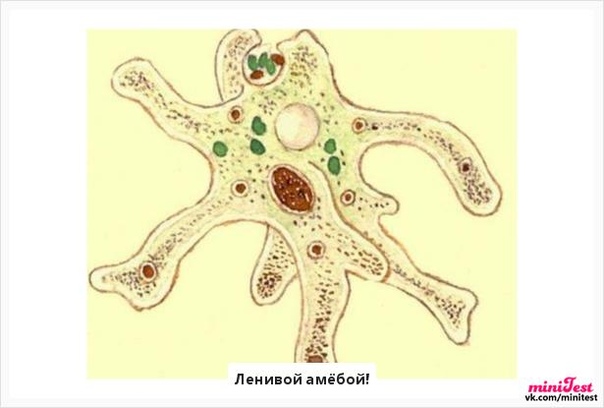
Tenupodia, с другой стороны, может выполнять роль более сенсорного типа. Они тоньше и, вероятно, не обладают структурными характеристиками, необходимыми для перетаскивания клетки. Своеобразной особенностью этой морфологии является то, что они простираются совершенно линейными сегментами и часто в виде линии непосредственно к другой клетке. Если тенупод перестанет расти, он, скорее всего, раздвоится или повернется вокруг фиксированной точки и продолжит свое линейное распространение, пока не встретит свою цель.Этот тип наведения может быть опосредован диффундирующими факторами, высвобождаемыми клеточными мишенями.31,32 Подсчитано, что домен, в котором отдельная клетка может эффективно общаться с помощью растворимого сигнала, имеет размер около 250 мкм и что связь внутри этого домена происходит от 10 до 30 минут.33 Эти теоретически полученные оценки хорошо коррелируют со временем и расстояниями, которые преодолевают тенуподы, чтобы добраться до соседней клетки или соединиться с ней.
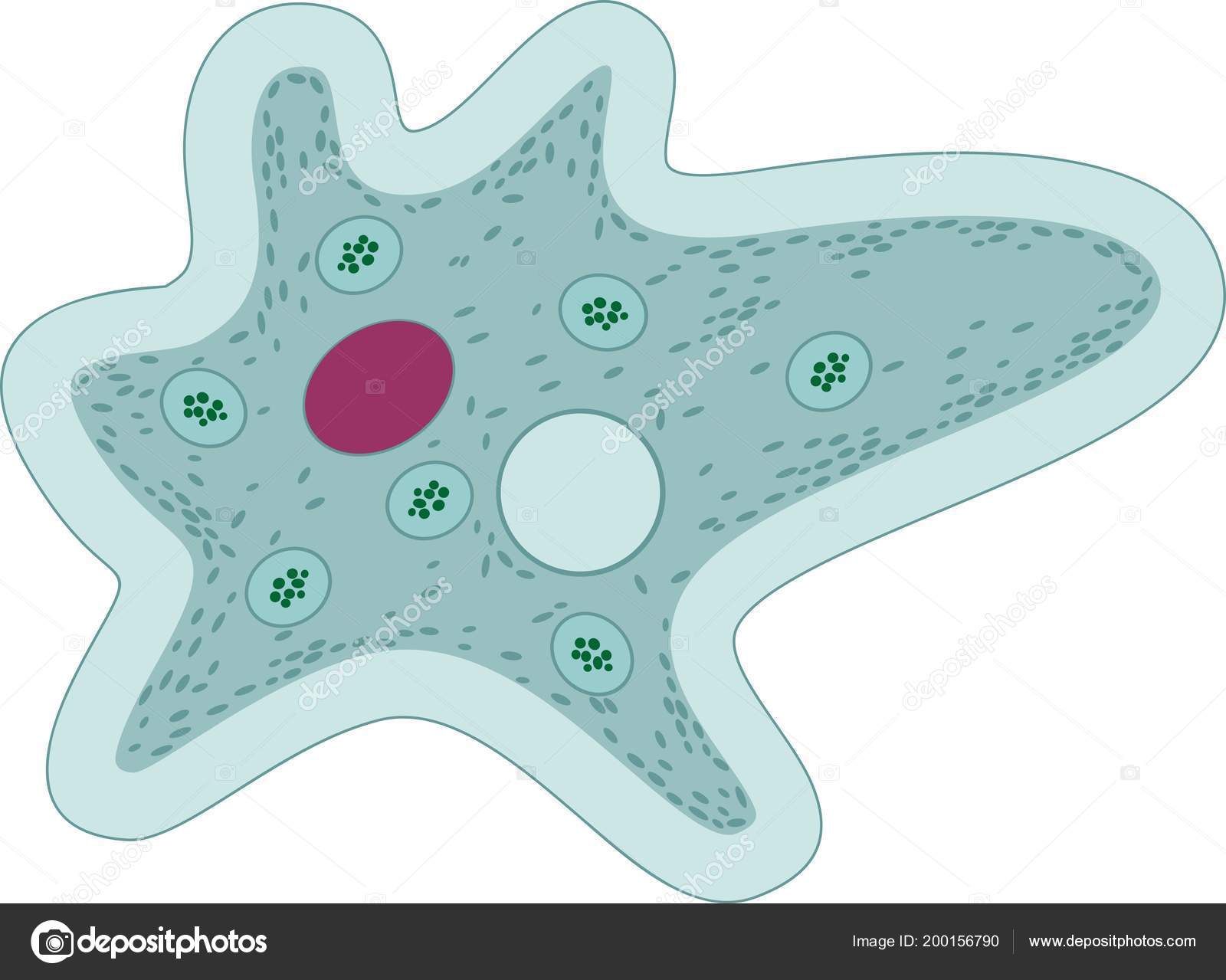 Уроподы также проявляют сходные сенсорные свойства в том смысле, что они, как было замечено, контактируют, захватывают и рекрутируют дополнительные Т-клетки во время лимфоцитарно-эндотелиального взаимодействия.2 Хотя для этого типа поведения был создан прецедент, уроподии кажутся более широкими и простираются всего на несколько диаметров клетки от клетки, что далеко не на расстояние растяжения тенуподов. Ультраструктурный анализ антигенпредставляющих дендритных клеток показывает, что эти клетки также развертывают отростки, которые намного длиннее тела клетки. с клеткой.Это высокое отношение площади поверхности к объему может также сделать тенупод очень чувствительным к сигналам окружающей среды. 5 Тенуподы также более распространены, когда клетки культивируются на поверхностях, покрытых компонентами внеклеточного матрикса, такими как фибронектин, ламинин или коллаген IV. Это указывает на то, что адгезия и контактная ориентация играют роль в их формировании и морфологии. Будь то через растворимые сигналы, поверхностную адгезию или и то, и другое, взаимодействия с окружающей средой явно влияют на морфологию тенуподов, предполагая, что выполняемая ими функция носит сенсорный характер.
Уроподы также проявляют сходные сенсорные свойства в том смысле, что они, как было замечено, контактируют, захватывают и рекрутируют дополнительные Т-клетки во время лимфоцитарно-эндотелиального взаимодействия.2 Хотя для этого типа поведения был создан прецедент, уроподии кажутся более широкими и простираются всего на несколько диаметров клетки от клетки, что далеко не на расстояние растяжения тенуподов. Ультраструктурный анализ антигенпредставляющих дендритных клеток показывает, что эти клетки также развертывают отростки, которые намного длиннее тела клетки. с клеткой.Это высокое отношение площади поверхности к объему может также сделать тенупод очень чувствительным к сигналам окружающей среды. 5 Тенуподы также более распространены, когда клетки культивируются на поверхностях, покрытых компонентами внеклеточного матрикса, такими как фибронектин, ламинин или коллаген IV. Это указывает на то, что адгезия и контактная ориентация играют роль в их формировании и морфологии. Будь то через растворимые сигналы, поверхностную адгезию или и то, и другое, взаимодействия с окружающей средой явно влияют на морфологию тенуподов, предполагая, что выполняемая ими функция носит сенсорный характер.
Предполагается, что антиген CD44 играет важную роль в хоминге и гемопоэзе35,36 благодаря его способности связываться с различными компонентами внеклеточного матрикса, включая фибронектин и коллаген.18,37 Недавно было высказано предположение, что антиген CD34 также участвует в адгезионных взаимодействиях через непрямой механизм, сигнализируя об изменениях профиля поверхности других молекул адгезии.38 Кроме того, существуют внутриклеточные хранилища белка CD34, которые могут перемещаться на плазматическую мембрану в ответ на внеклеточные сигналы. 39 Данные о составе, такие как эти, указывают на то, что может существовать взаимосвязь между ключевыми поверхностными маркерами, адгезивными белками, связанными с самонаведением, и морфологией псевдоподий. .
Возможно, наиболее интересным открытием является то, что эти ножки обнаруживаются на первичных гемопоэтических клетках человека.
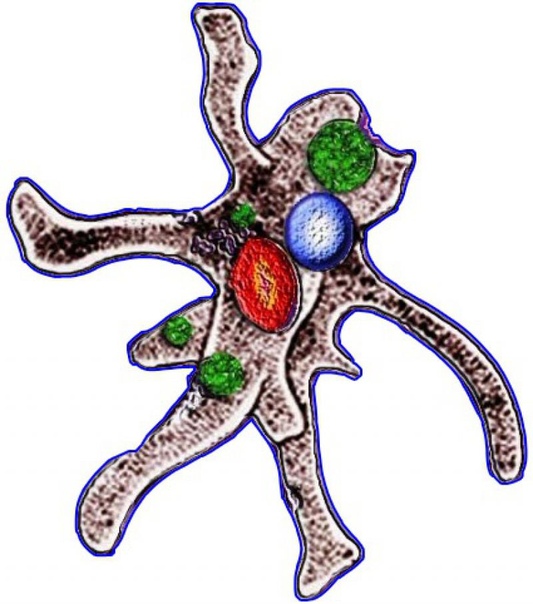 В дополнение к клеткам CD34 + KG1a, первичные человеческие клетки CD34 + из FL, UCB, aBM и mPB, которые мы тестировали, а также клетки печени плода мыши, все демонстрируют эти новые морфологии псевдоподов. Следовательно, эти новые морфологии не являются особым свойством клеточной линии KG1a. Следует также отметить, что клетки KG1a и первичные клетки человека CD34 + удлиняют отростки ламеллоподиального типа во время миграции. При отображении в режиме замедленной съемки эти клетки также демонстрируют классическое поведение ползания, как описано в литературе.1,19,40 Из клеток с покрытием только от 1% до 10% фактически развертывают магнуподы или тенуподы в любой момент времени.
В дополнение к клеткам CD34 + KG1a, первичные человеческие клетки CD34 + из FL, UCB, aBM и mPB, которые мы тестировали, а также клетки печени плода мыши, все демонстрируют эти новые морфологии псевдоподов. Следовательно, эти новые морфологии не являются особым свойством клеточной линии KG1a. Следует также отметить, что клетки KG1a и первичные клетки человека CD34 + удлиняют отростки ламеллоподиального типа во время миграции. При отображении в режиме замедленной съемки эти клетки также демонстрируют классическое поведение ползания, как описано в литературе.1,19,40 Из клеток с покрытием только от 1% до 10% фактически развертывают магнуподы или тенуподы в любой момент времени.Учитывая, что (1) известные типы псевдоподий выполняют специфические функции коммуникации и миграции, (2) примитивные гемопоэтические клетки расширяют длинные тонкие псевдоножки и (3) важной деятельностью примитивных гемопоэтических клеток являются коммуникация и миграция, возможно, что морфология и физиологическая активность магноподий и тенуподий связана с функцией, которую они выполняют во время самонаведения клетки после трансплантации.
 Эти данные могут существенно повлиять на наши представления о том, как мигрируют примитивные гемопоэтические клетки, и о роли структур цитоскелета в клинически важном процессе приживления стволовых клеток.
Эти данные могут существенно повлиять на наши представления о том, как мигрируют примитивные гемопоэтические клетки, и о роли структур цитоскелета в клинически важном процессе приживления стволовых клеток.Морфология Mayorella
Морфология MayorellaМир простейших, коловраток, нематод и олигохет
Mayorella Schaeffer, 1926 (код ID; 660, 3687, 3719, 7601) Подкласс Gymnamoebia: Отряд Amoebida: Семейство Paramoebidae (ref.Я БЫ; 7601)
Семейство Mayorellidae Schaeffer, 1926 (ссылка ID; 3719) [исх. Я БЫ; 3719]
Семейство Mayorellidae было создано Шеффером в 1926 г. для корненожек среднего и крупного размера с коническими или сужающимися ложноножками, которые не совершают прямого движения. Он включил в это семейство свой новый род Mayorella . Основной характеристикой этого рода являются многочисленные мелкие конические ложноножки, непрерывно формирующиеся вдоль переднего края клетки. Эти ложноножки всегда находятся в движении, находясь либо в состоянии растяжения, либо втягивания.(код ID; 3719) [исх. Я БЫ; 7601]
Эти ложноножки всегда находятся в движении, находясь либо в состоянии растяжения, либо втягивания.(код ID; 3719) [исх. Я БЫ; 7601]
Дактилоидные, тупые, гиалиновые субпсевдоподии одинаковой длины, не вильчатые, обычно немногочисленные, обычно образуются из гиалиновой лопасти; длина формы локомотива больше ширины; у видов, тонкая структура которых известна, покрытие сложной, ладьевидной чешуей, часто, если не всегда, двух размеров и узоров; большая и сложная область Гольджи, прилегающая к ядру, место синтеза чешуи; нет парасом; плавающие формы часто с отчетливой сферической центральной массой и тонкими расходящимися псевдоподиями.Как пресноводные, так и морские виды. (код ID; 7601)- Mayorella augusta Schaeffer, 1926 (арт. ID; 3687)
- Mayorella bigemma Schaeffer, 1918 (ref. ID; 660, 3719) указан автор и год? (код ID; 7586)
- Mayorella bulla Schaeffer, 1926 (ссылочный номер; 660, 3687, 7601)
- Mayorella cantabrigiensis Страница, 1983 г.
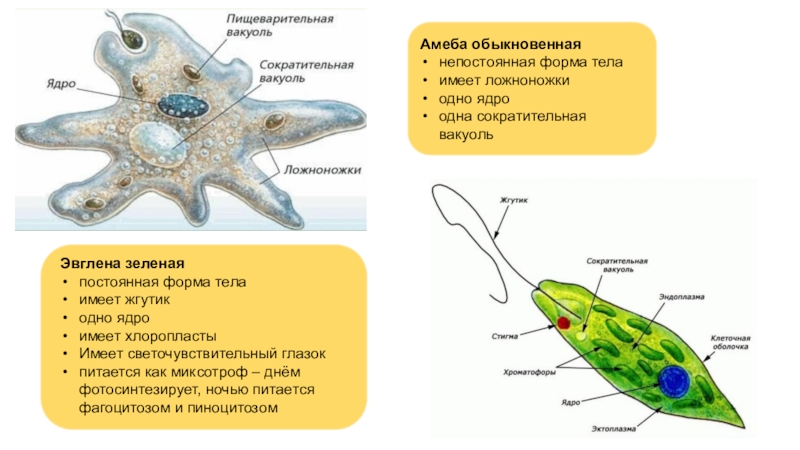 (ссылочный номер; 660 оригиналов)
(ссылочный номер; 660 оригиналов) - Mayorella clavabellans Bovee, 1970 (исх.Я БЫ; 660)
- Mayorella conipes Schaeffer, 1926 (арт. ID; 3687)
- Mayorella crystallus Schaeffer, 1926 (кат. ID; 3687)
- Mayorella gemmifera Schaeffer, 1926 (кат. ID; 3687)
- Mayorella godesae Арена, 1953 г. (номер по каталогу: 660)
- Mayorella oblonga Bovee, 1970 (код ID; 660)
- Mayorellapalestinensis Рейх, 1933 г.
См.; Acanthamoebapalestinensis (ссыл.Я БЫ; 3767) - Mayorella penardi Страница, 1972 г. (код ID; 660, 2039)
Син; Amoeba spumosa Penard, 1902 (номер ID; 660, 2039) - Mayorella riparia Страница, 1972 (ссылка ID; 2039) указан автор и год? (код ID; 7601)
- Mayorella spumosa (Gruber) (код ID; 3497)
- Mayorella stella (Schaeffer, 1926) Пейдж, 1981 (исх.
 Я БЫ; 7601, переописанная бумага), страница, 1981 (ссылка ID; 660)
Я БЫ; 7601, переописанная бумага), страница, 1981 (ссылка ID; 660) - Mayorella vespertilio Penard, 1902 (код ID; 660, 3719)
Син; Amoeba vespertilio Penard, 1902 г. (код ID; 3719) - Mayorella vespertilioides Страница, 1983 г. (ссылочный номер; 660 оригиналов)
- Mayorella viridis Leidy, 1874 (ссылка ID; 660, 3719)
Син; Amoeba viridis Leidy, 1874 (ref.Я БЫ; 3719)
Mayorella bigemma Schaeffer, 1918 (ref. ID; 660, 3719) указан автор и год? (код ID; 7586)Описания
Mayorella bigemma зарегистрирована как содержащая зоохлореллы только у трех особей. (код ID; 3719)
У особей этого вида может быть несколько сократительных вакуолей. Они могут выбрасывать свое содержимое в любую точку клеточной поверхности, за исключением псевдоподий. Вакуоль выглядит округлой непосредственно перед изгнанием.При изгнании вакуолярного содержимого сократительная вакуоль замещается слоем гиалиновой цитоплазмы. Когда все вакуолярное содержимое изгнано, гиалиновая область сохраняется как отдельная область (систолическая масса) в течение некоторого времени. В конце концов систолическая масса рассеивается за счет циклотической активности цитоплазмы. Ранние стадии наполнения трудно увидеть, но более поздние стадии диастолы включают, по крайней мере, слияние сопутствующих пузырьков. (код ID; 7586)
Когда все вакуолярное содержимое изгнано, гиалиновая область сохраняется как отдельная область (систолическая масса) в течение некоторого времени. В конце концов систолическая масса рассеивается за счет циклотической активности цитоплазмы. Ранние стадии наполнения трудно увидеть, но более поздние стадии диастолы включают, по крайней мере, слияние сопутствующих пузырьков. (код ID; 7586)Исследуемый материал
Mayorella bigemma была выделена из озера Чу, Англия (ссылка National Grid ST 596635) и идентифицирована Пейджем (1976).(код ID; 7586)
Бык Mayorella Schaeffer, 1926 (кат. ID; 660, 3687, 7601)Диагностика
Длина локомотивной формы 35-165 мкм, в среднем около 80 мкм; отношение длины к ширине около 2,0; задняя часть обычно не сильно сужена; часто один или несколько спинных гребней несколько неправильной формы; плавающая форма со сферической центральной массой и тонкими, лучистыми, часто несколько извилистыми псевдоподиями, длина которых может достигать четырех или пяти диаметров центральной клеточной массы; ядро 8,5-16,5 мкм, часто эллипсоидное, сильно деформируемое, с нерегулярными эндосомальными фрагментами, обычно рыхло слипшимися в центральной области ядра или в форме кольца; цитоплазматические кристаллы неизвестны; обычно две или несколько сократительных вакуолей; более крупные поверхностные чешуйки длиной примерно 720 нм; с выступающим рогом на каждом конце и решетчатой корзиной с открытым дном, подвешенной между концами и поддерживаемой с каждой стороны тремя вертикальными стойками из слегка вогнутого твердого основания; более мелкие чешуйки длиной примерно 150 нм, с простой лодковидной формой и мелкоячеистыми рабочими сторонами; киста не известна. (код ID; 7601)
(код ID; 7601)Наблюдаемая среда обитания
Пресная вода: США, Великобритания. (код ID; 7601)
Mayorella cantabrigiensis Страница, 1983 г. (ссылка ID; 660 оригинальная бумага)Диагностика
Локомотивная форма часто значительно шире кпереди; обычно с дактилоидными субпсевдоподиями, которые иногда встречаются отдельными парами; часто с морулированным уроидом. Ядро с центральным ядрышком; диаметр ядра 8,4-13,0 мкм, в среднем 9,4-11,1 мкм, примерно в 1,5-1,6 раза больше диаметра ядрышка.Сократительная вакуоль заменяется после опорожнения временным гиалоплазматическим участком. Цитоплазматические включения парные, каждая пара в своем пузырьке и одна пара кристаллическая. Плавающая форма округлая, иногда с короткими выступами, но не лучистая. Кисты не известны. Кутикула обычно имеет толщину 200-230 нм, иногда до 280 нм и немного больше. (код ID; 660)Тип местности
Фрешуотер, Англия. (код ID; 660)Тип слайдов
Голотип (1982:7:13:1) и паратип (1982:7:13:2), оба получены из штамма 311, в Британском музее (естественная история). (код ID; 660)
(код ID; 660)Измерения
Длина локомотивной формы 55-180 мкм, средняя 93-115 мкм; среднее отношение длины к ширине 2,4-3,3. (код ID; 660)
Mayorella spumosa (Gruber) (код ID; 3497)Описания
Форма среднего размера обычно имеет треугольные псевдоподии, короткие уроиды и ряд сократительных вакуолей, одна из которых часто крупная. Обычно видны отчетливые плазматические грануляции, а слабые дорсальные гребни редко наблюдаются у развитых образцов.(код ID; 3497)Комментарии
Обыкновенная амеба пресных вод также часто встречается в морской воде низкой солености, встречаясь в течение всего года в солоноватой и морской воде с соленостью 15,21—31,87 o/oo в Хиросимском заливе. Экземпляры, исследованные в море, обычно небольшие: длина большинства из них достигает 40–60 мкм. (код ID; 3497)Измерения
Длина без псевдоподий 40-120 мкм. (код ID; 3497)
Mayorella stella (Schaeffer, 1926) Page, 1981 (ссылка ID; 7601 переописанная бумага), Page, 1981 (ref.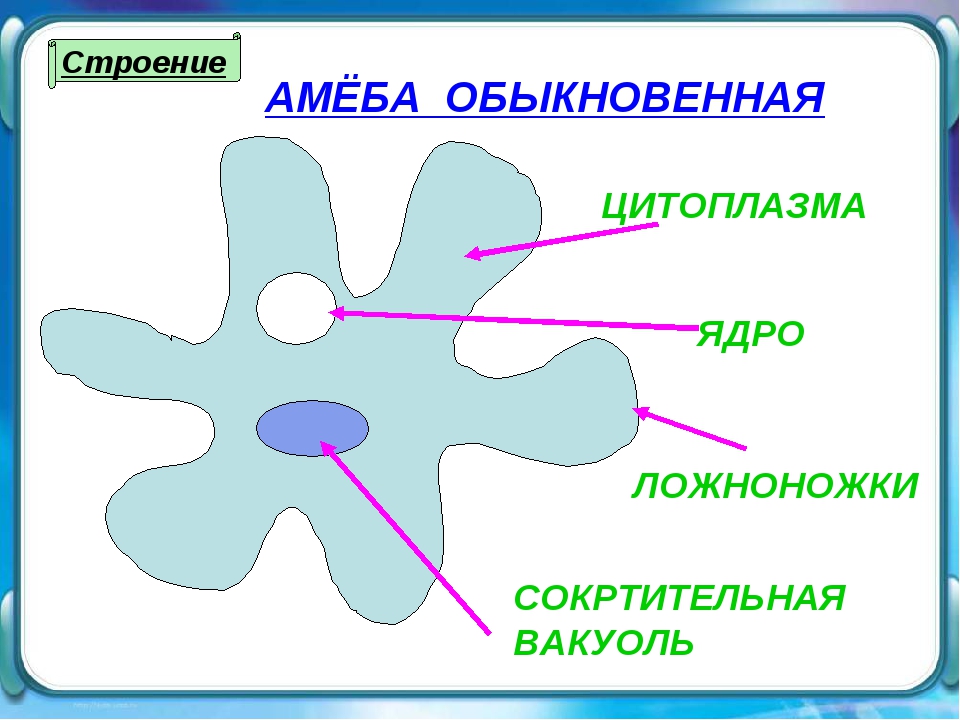 Я БЫ; 660)
Я БЫ; 660)Повторная диагностика
Длина формы локомотива сильно различается у разных штаммов, примерно от 20 до 120 мкм, значит от 35 до 65 мкм; среднее отношение длины к ширине около 3,0 или чуть меньше, но нередки удлиненные формы с отношением длины к ширине более 4,0; большое разнообразие форм; удлиненные формы иногда с продольным дорсальным гребнем; плавающая форма с более или менее сферической центральной массой и расходящимися, обычно изогнутыми псевдоподиями, длина которых может достигать или превышать двойной диаметр центральной массы; ядро часто удлиненное, часто с подобием двухъядерного состояния, когда ядро лопастное или скрученное; эндосомальный материал в длинном, лопастном, часто скрученном теле, возможно, иногда фрагментирован, не пристеночный в ядре; длина ядра 4-13 мкм, значит 7-9 мкм; цитоплазматические кристаллы неизвестны; обычно две сократительные вакуоли; более крупные чешуи длиной около 550 нм, с решетчатой корзинкой, подвешенной между двумя концами, дно которой параллельно вогнутому грунтовому дну чешуи, к которой корзинка прикреплена сбоку; более мелкие чешуйки длиной около 150 нм, с простой лодкообразной формой и мелкоячеистыми сторонами; киста не известна.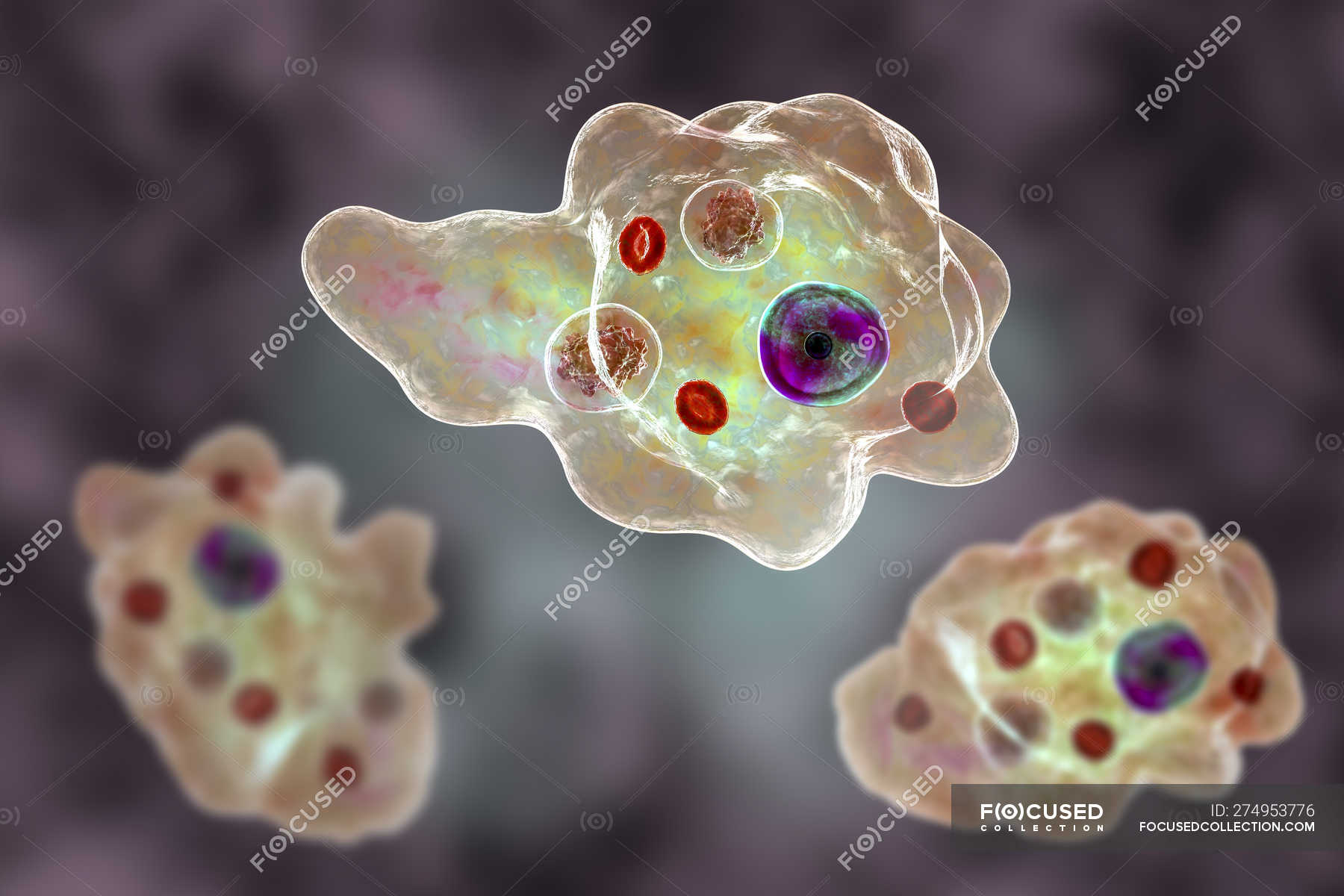 Отличается от Mayorella riparia прежде всего формой ядра и ядрышка. (код ID; 7601)
Отличается от Mayorella riparia прежде всего формой ядра и ядрышка. (код ID; 7601)Наблюдаемые местообитания
Пресная вода, США: эстуарий, Шотландия: эстуарий выдерживает соленость только до 3 0/00. (код ID; 7601)Тип слайдов
Неотип (1981:7:10:3) и паранеотип (1981:7:10:4), оба получены из штамма 272, в Британском музее (естественная история). (код ID; 7601)
Mayorella vespertilioides Страница, 1983 г. (ссылка ID; 660 оригинальная бумага)Диагностика
Локомотивная форма иногда лишена дактилоидных субпсевдоподий; часто с морулированным уроидом.Ядро с центральным ядрышком; диаметр ядра 5,6-10,0 мкм, в среднем 6,5-8,3 мкм, примерно в 1,8 раза больше диаметра ядрышка. Сократительная вакуоль заменяется после опорожнения временным гиалоплазматическим участком. Часто с парными цитоплазматическими включениями, каждая пара в своем пузырьке и одна пара кристаллическая. Плавающая форма с небольшими бугристыми выступами или с лучистыми гиалиновыми псевдоподиями, сужающимися к широкому основанию, но без правильной сферической центральной массы.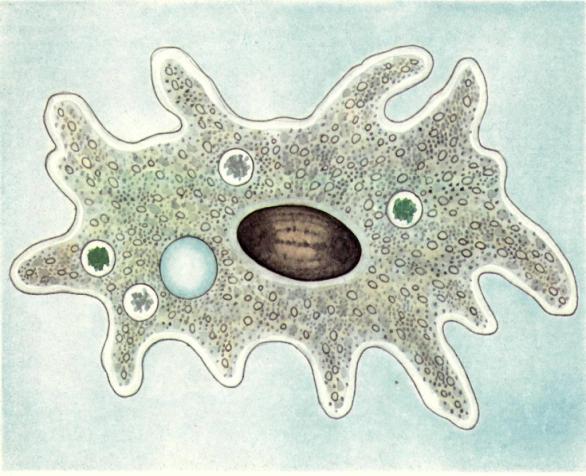 Кисты не известны. Кутикула толщиной 180-200 нм; Тельца Гольджи иногда парные.(код ID; 660)
Кисты не известны. Кутикула толщиной 180-200 нм; Тельца Гольджи иногда парные.(код ID; 660)Тип местности
Фрешуотер, Англия. (код ID; 660)Тип слайдов
Голотип (1982:7:13:3) и паратип (1982:7:13:4), оба получены из штамма 314, в Британском музее (естественная история). (код ID; 660)Измерения
Длина локомотивной формы 35-90 мкм, средняя 57-65 мкм; среднее отношение длины к ширине 2,3-2,8. (код ID; 660)
Mayorella viridis Leidy, 1874 (ссылка ID; 660, 3719)Синоним
Amoeba viridis Leidy, 1874 (ref.Я БЫ; 3719)Описания
- Световая микроскопия: Исследование препарата висячей капли с помощью световой микроскопии показало, что форма клетки обычно удлиненная, причем длина всегда больше ширины. Длина 100 клеток, измеренная от мочевой оболочки до основания псевдоподий, составила в среднем 111 мкм с диапазоном от 61 мкм до 55,5 мкм. Уроид часто был округлым. Тупые, гиалиновые, конические субпсевдоподии образуются из передней гиалиновой области клетки.
 При движении амебы передние субпсевдоподии быстро захватываются активным потоком цитоплазмы, и очень быстро образуются новые.В активном состоянии по бокам клетки также образуются конические субпсевдоподии. Субпсевдоподии не контролируют направление движения амебы, которое в наиболее активном состоянии составляло 77 мкм в минуту. Гранулярная эндоплазма, содержащая зоохлореллы, быстро стекает к переднему концу клетки. Активного заглатывания не наблюдалось, наблюдалась только одна пищевая вакуоль на организм, содержимое которой оказалось водорослевым. Однако большинству клеток, по-видимому, не хватало и пищевой вакуоли.Когда она присутствует, пищевая вакуоль имеет сферическую форму и приблизительно 11 мкм в диаметре, увеличиваясь до 16 мкм, прежде чем быстро выбросить, по-видимому, лишь частично переваренные остатки пищи. Ядро приблизительно центральное, сферическое, с центральным шаровидным ядрышком. Средний диаметр ядра 12,4 мкм, ядрышка 7,6 мкм. В культуральной среде M. viridis наблюдалось наличие многочисленных сократительных вакуолей, которые перед систолой достигают максимального диаметра 12 мкм.
При движении амебы передние субпсевдоподии быстро захватываются активным потоком цитоплазмы, и очень быстро образуются новые.В активном состоянии по бокам клетки также образуются конические субпсевдоподии. Субпсевдоподии не контролируют направление движения амебы, которое в наиболее активном состоянии составляло 77 мкм в минуту. Гранулярная эндоплазма, содержащая зоохлореллы, быстро стекает к переднему концу клетки. Активного заглатывания не наблюдалось, наблюдалась только одна пищевая вакуоль на организм, содержимое которой оказалось водорослевым. Однако большинству клеток, по-видимому, не хватало и пищевой вакуоли.Когда она присутствует, пищевая вакуоль имеет сферическую форму и приблизительно 11 мкм в диаметре, увеличиваясь до 16 мкм, прежде чем быстро выбросить, по-видимому, лишь частично переваренные остатки пищи. Ядро приблизительно центральное, сферическое, с центральным шаровидным ядрышком. Средний диаметр ядра 12,4 мкм, ядрышка 7,6 мкм. В культуральной среде M. viridis наблюдалось наличие многочисленных сократительных вакуолей, которые перед систолой достигают максимального диаметра 12 мкм. Период от начала диастолы до систолы составляет примерно 90 сек.Их легко заметить, если оставить клетки под покровным стеклом на несколько минут. Другие вакуоли диаметром до 18,5 мкм также присутствуют по всей эндоплазме. Кристаллов не обнаружено. Делящихся клеток не наблюдалось. (код ID; 3719)
Период от начала диастолы до систолы составляет примерно 90 сек.Их легко заметить, если оставить клетки под покровным стеклом на несколько минут. Другие вакуоли диаметром до 18,5 мкм также присутствуют по всей эндоплазме. Кристаллов не обнаружено. Делящихся клеток не наблюдалось. (код ID; 3719) - Электронная микроскопия: Предварительные исследования тонкой структуры M. viridis были проведены мисс С.М. Блейки и ее результаты, поскольку они касались поверхностного слоя, были изложены в статье, прочитанной в 1977 г. в Британской секции Общества протозоологов (Page & Blakey 1977).Клеточная оболочка снаружи плазматической мембраны имеет толщину 0,2 мкм и состоит из трех слоев. Прилежащий к мембране слой состоит из фибрилл, идущих параллельно плазматической мембране, и имеет толщину 94 нм. Следующий слой имеет толщину 73 нм и состоит из полос, перпендикулярных плазматической мембране. Внешний слой аналогичен, но имеет ширину всего 33 нм. Зазор различной толщины отделяет плазматическую мембрану от клеточной оболочки.
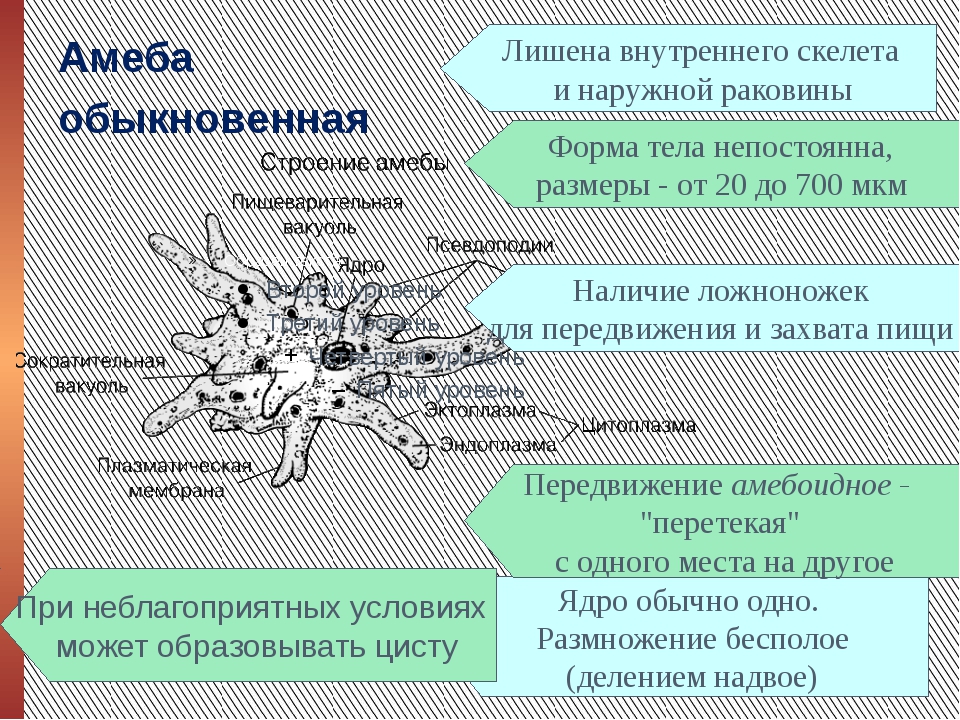 Ядро содержит крупное ядрышко неправильного очертания, обычно состоящее из двух неоднородных участков.Ядро окружают три мембраны, а везикулы, присутствующие в цитоплазме, концентрируются вокруг поверхности ядра. Активное образование везикул, по-видимому, происходит из наружной мембраны. Расстояние между тремя мембранами составляет 50 мкм. Внешняя мембрана, окружающая ядро, может представлять собой гладкий эндоплазматический ретикулум, поскольку он кажется более складчатым и менее правильным, чем две внутренние мембраны. Ядерные поры имеют диаметр 30 нм и расположены с интервалом до 1,3 мкм. Митохондрии, содержащие анастомозирующие кристы, встречаются по всей клетке, но, по-видимому, сосредоточены в области зоохлорелл.Они могут быть довольно удлиненными, достигая длины 1,6 мкм. Эндозойные зоохлореллы находятся в связанных с мембраной везикулах и обычно имеют размер 3×2 мкм. Иногда наблюдалось, что везикулы соединяются, а зоохлореллы находятся в непосредственном контакте друг с другом (возможно, после деления клеток?).
Ядро содержит крупное ядрышко неправильного очертания, обычно состоящее из двух неоднородных участков.Ядро окружают три мембраны, а везикулы, присутствующие в цитоплазме, концентрируются вокруг поверхности ядра. Активное образование везикул, по-видимому, происходит из наружной мембраны. Расстояние между тремя мембранами составляет 50 мкм. Внешняя мембрана, окружающая ядро, может представлять собой гладкий эндоплазматический ретикулум, поскольку он кажется более складчатым и менее правильным, чем две внутренние мембраны. Ядерные поры имеют диаметр 30 нм и расположены с интервалом до 1,3 мкм. Митохондрии, содержащие анастомозирующие кристы, встречаются по всей клетке, но, по-видимому, сосредоточены в области зоохлорелл.Они могут быть довольно удлиненными, достигая длины 1,6 мкм. Эндозойные зоохлореллы находятся в связанных с мембраной везикулах и обычно имеют размер 3×2 мкм. Иногда наблюдалось, что везикулы соединяются, а зоохлореллы находятся в непосредственном контакте друг с другом (возможно, после деления клеток?).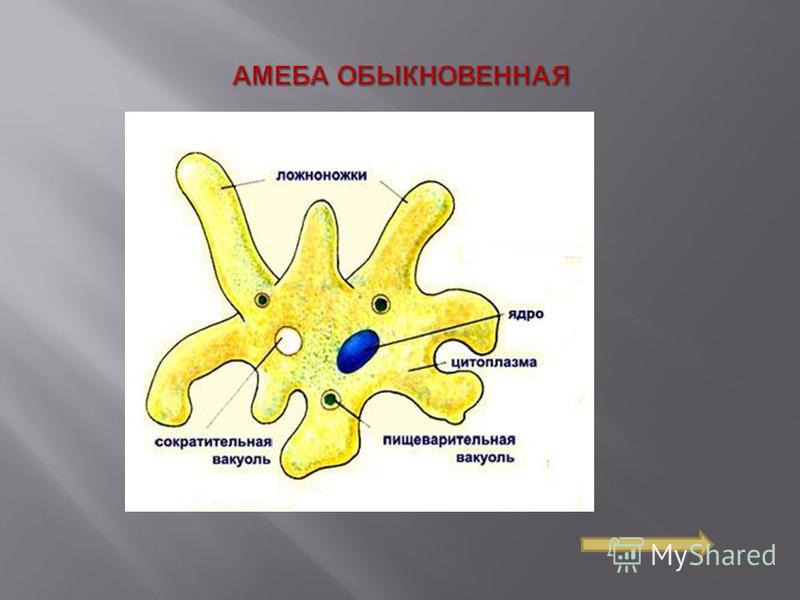 Водоросли имеют типично хлорофитовое строение с пластидой, состоящей из пучка из 3-х тилакоидных мембран. Пиреноид диаметром около 1 мкм окружен преобладающей крахмальной оболочкой размером около 4.4 мкм в поперечнике. Крахмальная оболочка инвагинирована двумя пластинками тилакоидов, одна из которых пересекает пиреноид. Диктиосомы были видны по всей цитоплазме и были либо сильно вогнутыми, либо менее вогнутыми. Расстояние по стопке сильно вогнутой диктиосомы составляет 0,3 мкм. В полученном срезе точное количество мешочков определить не удалось. Расстояние поперек менее вогнутой диктилосомы составляет 0,7 мкм, присутствуют 7 мешочков. Рибосомы разбросаны по всей цитоплазме и связаны с шероховатой эндоплазматической сетью.Плотные тельца также обнаруживаются по всей цитоплазме. (код ID; 3719)
Водоросли имеют типично хлорофитовое строение с пластидой, состоящей из пучка из 3-х тилакоидных мембран. Пиреноид диаметром около 1 мкм окружен преобладающей крахмальной оболочкой размером около 4.4 мкм в поперечнике. Крахмальная оболочка инвагинирована двумя пластинками тилакоидов, одна из которых пересекает пиреноид. Диктиосомы были видны по всей цитоплазме и были либо сильно вогнутыми, либо менее вогнутыми. Расстояние по стопке сильно вогнутой диктиосомы составляет 0,3 мкм. В полученном срезе точное количество мешочков определить не удалось. Расстояние поперек менее вогнутой диктилосомы составляет 0,7 мкм, присутствуют 7 мешочков. Рибосомы разбросаны по всей цитоплазме и связаны с шероховатой эндоплазматической сетью.Плотные тельца также обнаруживаются по всей цитоплазме. (код ID; 3719)
Комментарии
Предварительные исследования структуры поверхности M. viridis показали, что он структурно и, по-видимому, химически отличается от обычного гликокаликса (Page & Blakey, 1977).
2
1
1
Типы псевдоподий
Филоподия
Филоподии представляют собой тонкие структуры на основе актина, которые выполняют сенсорные и двигательные функции.Как и другие псевдоподии, филоподии представляют собой клеточные выпячивания и, таким образом, выходят за пределы клеточной поверхности. Однако по сравнению с псевдоподиями, обнаруженными в одноклеточных организмах, филоподии в основном встречаются в некоторых клетках многоклеточных организмов, где они проникают во внеклеточный матрикс и участвуют в передаче сигналов.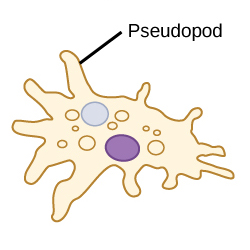
* Некоторые одноклеточные организмы, такие как представители рода Dictyostelium, используют для питания филоподии.
Филоподия Формирование
Формирование филоподий начинается с зарождения актиновых филаментов под влиянием нуклеаторов (группы белков).Хотя для объяснения инициации (индукции) филоподий были предложены две модели, процесс, по-видимому, запускается связыванием GTPase Cdc42 с важным регулятором, известным как N-WASP.
Это приводит к активации N-WASP, который, в свою очередь, связывается с профилином и Arp2/3 с образованием комплекса, который запускает формирование новой псевдоподии.
Хотя были предложены две модели инициации филоподий, это модель конвергентной элонгации и модель зарождения кончика, дальнейшие исследования показали, что они не исключают друг друга.
Согласно этим исследованиям, две модели могут фактически сосуществовать, особенно если учесть разнообразный и изменчивый характер этих структур.
Что касается строения, филоподии представляют собой тонкие цилиндрические выступы диаметром от 100 до 200 нм и длиной 10 мкм. Однако некоторые филоподии на клеточной поверхности очень короткие, едва выступающие из клеточной поверхности. Эти филоподии известны как микрошипы.
Актиновые филаменты (от 10 до 30) составляют центральное ядро структуры.Здесь нити плотно упакованы вместе параллельным образом, образуя стержень псевдоподии.
Внутри филоподий нити перекрываются и выстраиваются в единой полярности; где зазубренный конец нити ориентирован к филоподиальному концу структуры. Колючки заходят в концевой комплекс, состоящий из актин-связывающих белков и филаментов. В области основания псевдоподии филаменты впадают в актиновую сеть, расположенную под клеточной мембраной.
В то время как добавление актиновых мономеров к актиновым филаментам происходит на кончике структуры по мере ее удлинения, было показано, что филаменты постоянно возвращаются назад к основанию через ретроградный поток, который зависит от миозина. Именно скорость между этим потоком и добавлением мономера вверху влияет на общую скорость роста.
Именно скорость между этим потоком и добавлением мономера вверху влияет на общую скорость роста.
На основании исследований электронной томографии результаты показали, что этот рост происходит со скоростью около 0,2 мкм/с (которая может увеличиваться до 25 мкм в минуту) до того, как структура достигает критической длины. В этот момент филоподии либо производят новые актиновые структуры, либо начинают втягиваться.
Достигнув критической длины, некоторые филоподии связываются и сливаются с плазматической мембраной, образуя пучки актина.
* Филоподии достаточно гибкие, чтобы волноваться во внеклеточном матриксе. Тем не менее, они также достаточно прочны, что позволяет поддерживать структурную целостность, даже если длина конструкции превышает 30 мкм.
Некоторые из наиболее распространенных типов филоподий включают:
· Цитонемы. Обнаруженные в крыльях видов дрозофилы, этот тип филоподий может достигать более 800 мкм, где они участвуют в межклеточном взаимодействии
2
· Миоподии — тип филоподий, которые можно обнаружить на клеточной поверхности мышечных клеток, где они смешиваются с другими типами филоподий.
Функции В многоклеточных организмах филоподии выполняют ряд физиологических функций, включая заживление ран, передачу клеточных сигналов, а также развитие клеток. Учитывая, что филоподии простираются во внеклеточный матрикс, они способны ощущать химические вещества в своем окружении, что, в свою очередь, позволяет клетке реагировать соответствующим образом.
Здесь рецепторы внутри филоподий получают химическую информацию во внеклеточном матриксе, которая затем передается вниз в клетку (по сигнальному пути).
Во внеклеточном матриксе филоподии могут идентифицировать мишени, необходимые для адгезии, что позволяет генерировать направляющие сигналы, а также тяговые силы, которые в конечном итоге способствуют движению клетки. Благодаря этому процессу клетки способны к таким действиям, как нахождение путей аксонов и адгезия к эпителиальным клеткам, что способствует миграции клеток.
* Филоподии также способствуют застегиванию адгезии, процессу, в котором они участвуют в выравнивании и адгезии клеток, что уменьшает зазор между клетками.
Аксоподия Как и филоподии, аксоподии представляют собой длинные и тонкие отростки клеток. Однако они более жесткие (и поэтому кажутся игольчатыми), чем филоподии, которые имеют тенденцию быть более гибкими по своей природе. Их можно найти на клеточной поверхности различных организмов (например, представителей типа Antinopoda), где они участвуют в питании и передвижении.
У этих организмов аксоподий (Sin. axopodium) происходит из аксопластов (мембранная полость вблизи ядра, состоящая из микрофибриллярного материала и гранул).Здесь микрофибриллы организуются для построения стенок микротрубочек, которые затем образуют параллельные ряды, соединенные звеньями.
На основе микроскопических исследований было показано, что эти микротрубочки образуют взаимосвязанные двойные катушки (примерно 500 трубочек образуют примерно 5 витков катушки).
Микротрубочки составляют ядро аксонемы (которая является центральной частью аксоподий) и проходят по всей длине структуры.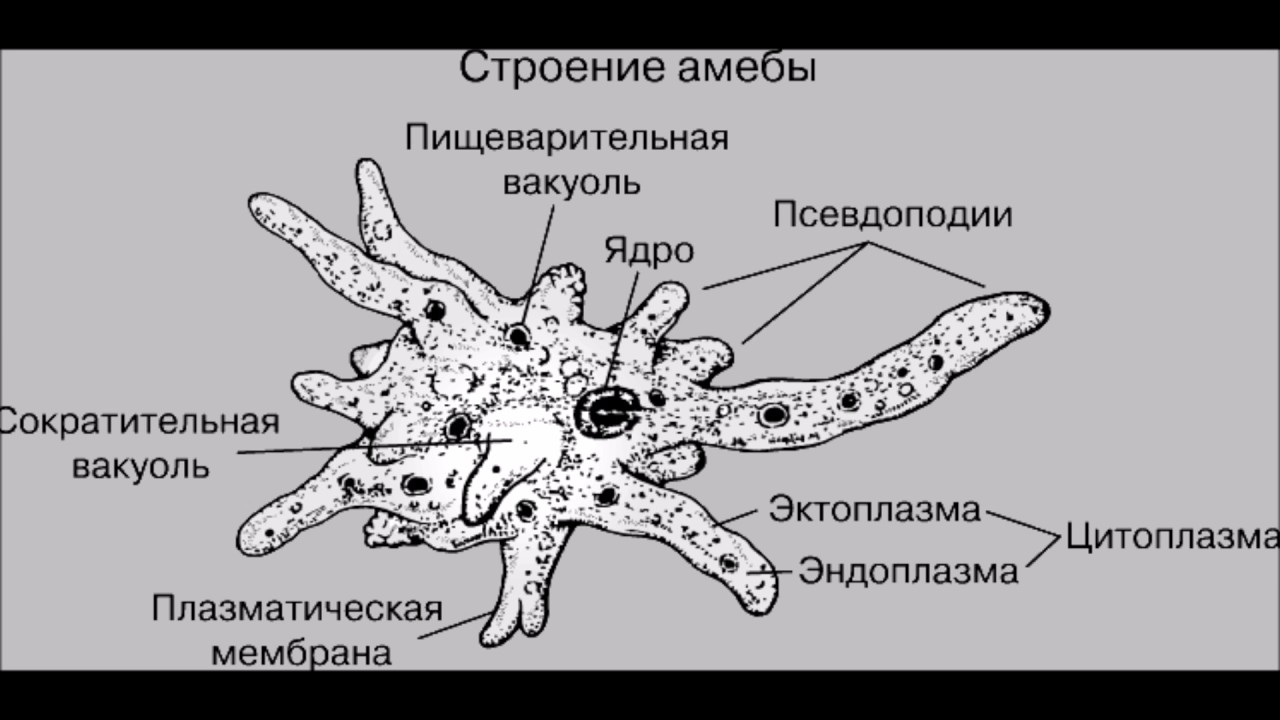 Помимо микротрубочек, структура также состоит из цитоплазмы, которая несет такие органеллы, как митохондрии, к цитосоме и от нее.
Помимо микротрубочек, структура также состоит из цитоплазмы, которая несет такие органеллы, как митохондрии, к цитосоме и от нее.
* В клетке аксоподии выходят из аксопластов через поры, расположенные на стенке капсулы. В зависимости от организма эти поры различаются по размеру и количеству. В то время как было показано, что полицистины содержат многие из этих пор, феодарианы содержат около трех из этих пор.
Показано, что укорочение аксоподий происходит во время питания. Например, было показано, что после захвата пищи быстрое сокращение происходит из-за разрушения микротрубочек.После сокращения аксоподы начинают удлиняться с нормальной скоростью, пока не достигнут нормальной длины.
Некоторые из основных характеристик AXOPODIA включают в себя:
· RegiD — как таковые, они более устойчивы к изгибам по сравнению с другими псевдоподачами
· тонкие и удлиненные — игольчатые
· во время неблагоприятных условий они могут убрать (быстрое сокращение), где нити реабсорбируются после дисперсии фибриллов в аксоплазме
· Axopodia MicroTubula имеют диаметр около 220 A
· Axopodia также потерян во время клеточного деления
· Имеют липкую поверхность, что объясняется наличием экструзом
Функции
Кормление Для таких организмов, как представители класса Actinopoda, важную роль в питании играют аксоподии. Как уже упоминалось, аксоподии имеют на своей поверхности клейкое вещество, вырабатываемое мукоцистами. Кроме того, у них также есть кинетоцисты, которые выбрасывают нитевидные структуры, которые эффективно ловят их добычу.
Как уже упоминалось, аксоподии имеют на своей поверхности клейкое вещество, вырабатываемое мукоцистами. Кроме того, у них также есть кинетоцисты, которые выбрасывают нитевидные структуры, которые эффективно ловят их добычу.
Используя эти экструсомы, эти организмы способны улавливать пищевой материал (или добычу), который затем транспортируется в тело клетки через цитоплазматический поток. В то время как более мелкая добыча может быть поймана и захвачена одним аксоподием, более крупная запутывается в нескольких аксоподиях.В некоторых случаях было показано, что несколько особей участвуют в отлове более крупной добычи.
Поддержка и передвижение Было показано, что помимо питания аксоподии также помогают удерживать простейших в положении в воде и даже способствуют передвижению. Например, благодаря контролируемому изменению длины аксопод некоторые гелиозоа оказались эффективно поперечными в водной среде. Это у других организмов достигается за счет расширения и сокращения эктоплазматических вакуолей, расположенных между аксоподиями.
Здесь организм может оставаться на месте или контролировать направление движения. Во время клеточного деления и аксоподии, и эктоплазма теряются, в результате чего организм опускается на дно.
Некоторые другие важные функции аксоподий включают:
Помимо микротрубочек, структура также состоит из цитоплазмы, которая несет такие органеллы, как митохондрии, к цитосоме и от нее. Как уже упоминалось, аксоподии имеют на своей поверхности клейкое вещество, вырабатываемое мукоцистами. Кроме того, у них также есть кинетоцисты, которые выбрасывают нитевидные структуры, которые эффективно ловят их добычу.