Тест по биологии Тип инфузории 7 класс
Тест по биологии Тип инфузории 7 класс с ответами. Тест включает 2 варианта, в каждом по 7 заданий.
Вариант 1
A1. Какую функцию выполняет большое ядро инфузории?
1) обмена веществ
2) двигательную
3) половую
4) дыхательную
А2. В каком слое инфузории расположены опорные волоконца?
1) в эндоплазме
2) в цитоплазме
3) в эктоплазме
4) в оболочке
А3. Где происходит переваривание пищи у инфузории?
1) в вакуоли
2) в клеточном рте
3) в желудочке
4) в ядре
А4. В каком направлении происходит деление клетки инфузории?
1) поперек тела
2) вдоль продольной оси тела
3) по любой оси тела
4) по диагонали
В1. С помощью какого процесса происходит обновление и перераспределение генетического материала у инфузорий?
В2. Как питаются паразитические инфузории?
C1.
Вариант 2
A1. Какую функцию выполняет малое ядро инфузории?
1) обмена веществ
2) двигательную
3) размножения
4) дыхательную
А2. Какие особые образования инфузорий могут превращаться в длинную упругую нить?
1) соединительный мостик
2) опорные волоконца
3) трихоцисты
4) сократительная вакуоль
А3. Как находит свою добычу инфузория-туфелька?
1) чувствует запах веществ
2) чувствует наличие химических веществ
3) определяет на ощупь
4) с помощью длинной упругой нити
А4. С помощью какого органоида подвижное ядро переходит из одной инфузории в другую при половом процессе?
1) соединительного мостика
2) цитоплазматического мостика
3) цисты
4) клеточного рта
В1.
Какой способ размножения характерен для инфузорий?В2. Какие инфузории ведут неподвижный образ жизни?
C1.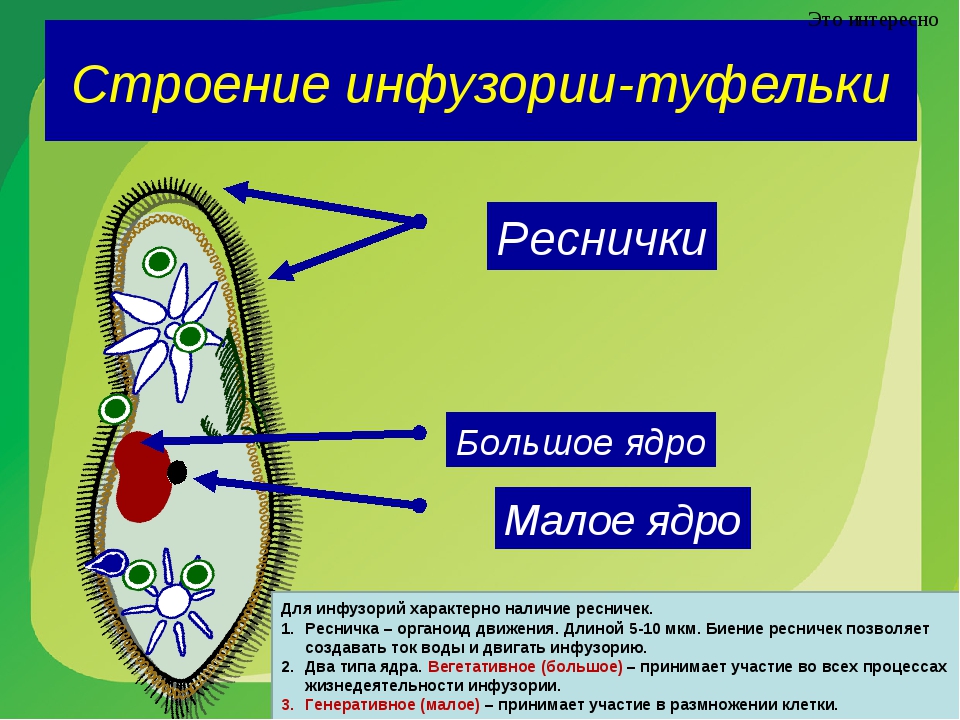 Опишите особенности дыхания инфузории.
Опишите особенности дыхания инфузории.
Ответы на тест по биологии Тип инфузории 7 класс
Вариант 1
А1-1
А2-3
А3-1
А4-1
В1. Конъюгация
В2. Всей поверхностью тела
С1. Особенности передвижения инфузории:
1) передвигается тупым концом вперед,
2) находится в постоянном волнообразном движении,
3) скорость передвижения 2,5 мм в секунду,
4) в время движения поворачивается вокруг оси своего тела,
5) туфелька резко взмахивает ресничками — ножками и плавно возвращая их на место, работает словно веслами в лодке и таких взмахов 30 раз в секунду,
6) направление движения меняется за счет изгибаний тела,
7) ресничек 10 000 — 15 000.
Вариант 2
А1-3
А2-3
А3-2
А4-1
В1. Бесполый, половой
В2. Сосущие, сувойки
С1. Особенности дыхания инфузории:
1) осуществляется всей поверхностью тела организма,
2) способна существовать за счет гликолиза при низкой концентрации воды.
Тип Инфузории. Процесс питания и выделения
Стр. 42
Вспомните
1.Каковы способы размножения одноклеточных животных и растений?
Деление, почкование, спорообразование, вегетативное размножение.
2.Какую функцию выполняет в клетке ядро?
Ядро — регулирует процессы, протекающие в клетке, обеспечивает передачу наследственных признаков дочерним клеткам при размножении.
Стр. 46
Вопросы после параграфа
1.Докажите на конкретных примерах, что инфузории имеют более сложное строение, чем саркодовые и жгутиконосцы.
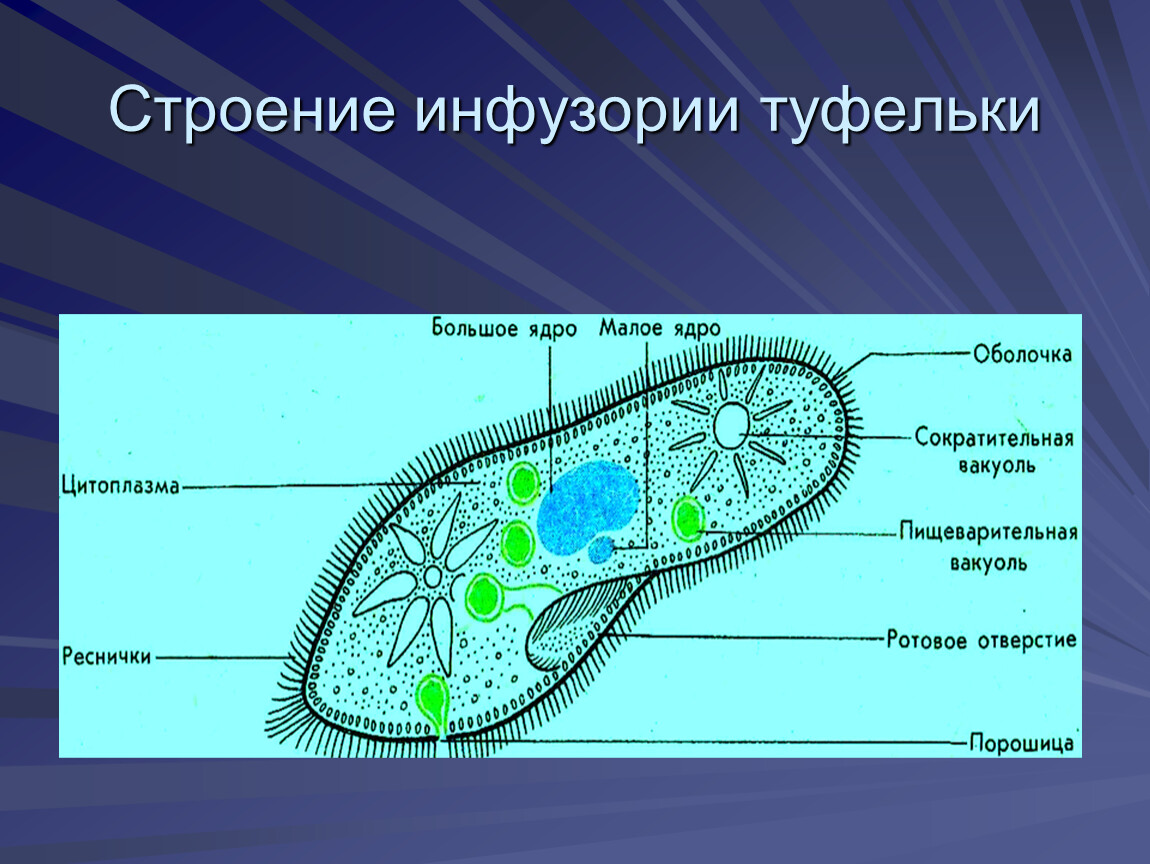
В клетке инфузории два ядра. Одно большое ядро отвечает за питание, дыхание, движение, обмен веществ, а малое ядро участвует в половом процессе.
2.Установите связь между усложнением строения инфузории — туфельки и процессами питания и выделения.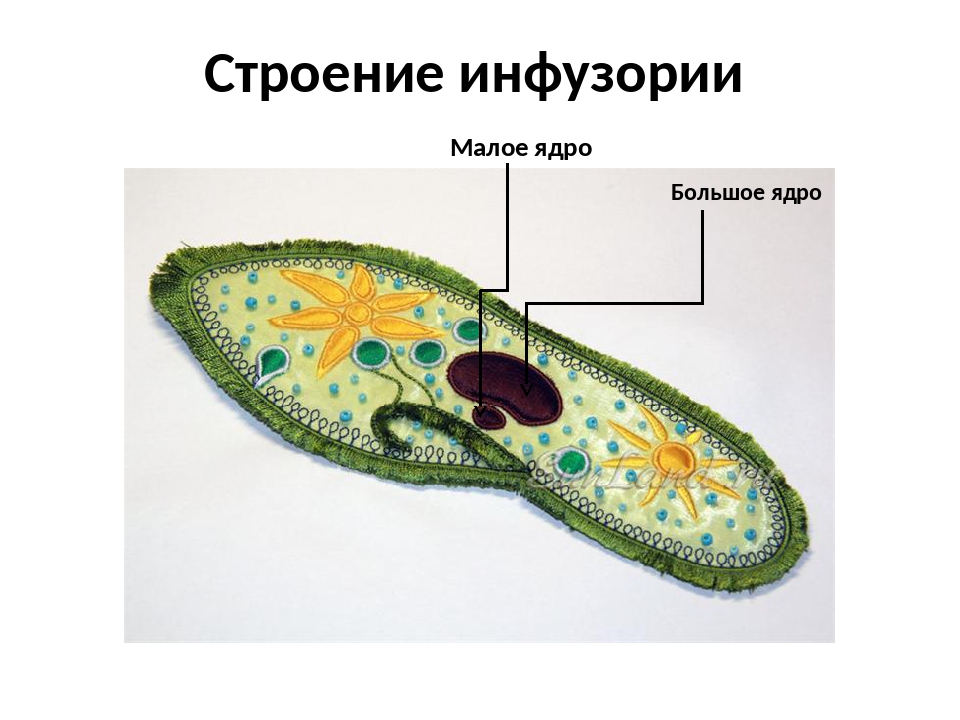
На теле инфузории имеется углубление — клеточный рот, который переходит в клеточную глотку. Около рта располагаются более толстые и длинные реснички. Они загоняют в глотку вместе с потоком воды бактерий — основную пищу туфельки. На дне глотки пища попадает в пищеварительные вакуоли, которые перемещаются в теле инфузории током цитоплазмы. В вакуолях пища переваривается, питательные вещества поступают из вакуолей в цитоплазму и используются для жизнедеятельности. Непереваренные остатки выбрасываются наружу в заднем конце тела через особую структуру — порошицу. В организме инфузории-туфельки находятся две сократительные вакуоли, которые располагаются у переднего и заднего концов тела. Они сокращаются поочерёдно два-четыре раза в минуту. Каждая вакуоль состоит из центрального резервуара и направленных к нему каналов. Жидкость сначала заполняет каналы, потом попадает в центральный резервуар и оттуда изгоняется наружу. Через сократительные вакуоли из тела инфузории выводится излишек воды вместе с растворёнными в ней вредными продуктами жизнедеятельности.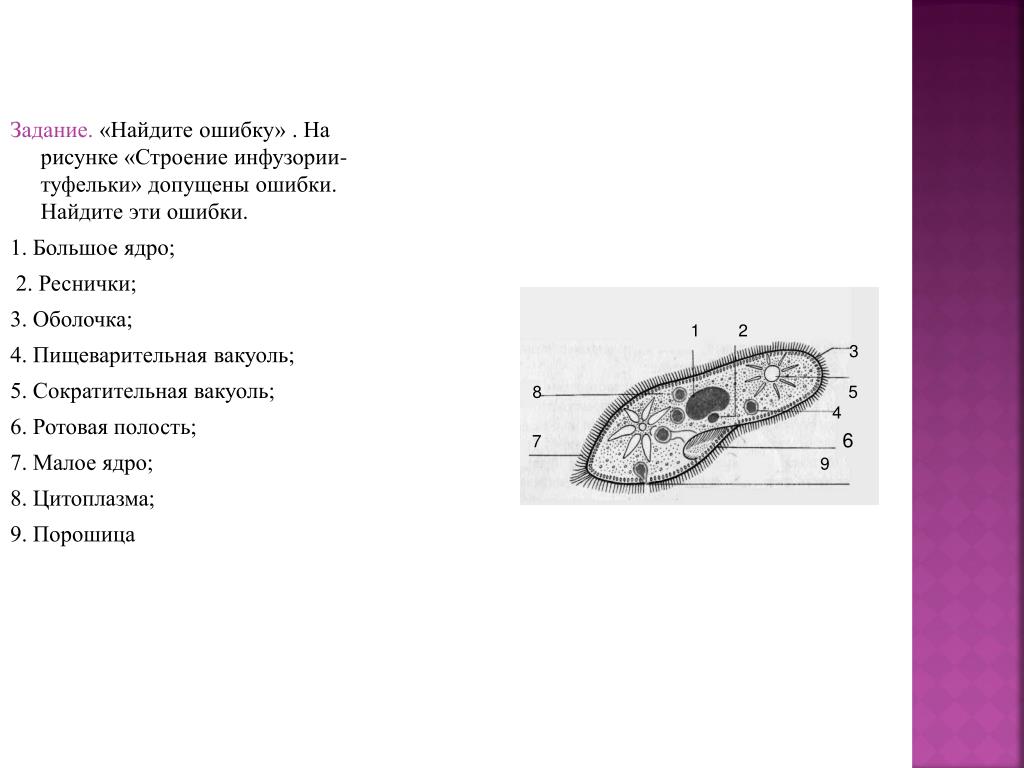
3.Охарактеризуйте особенности процесса размножения инфузории — туфельки.
Инфузории — туфельки обычно размножаются бесполым путём — делением надвое. В отличие от жгутиконосцев, инфузории делятся поперёк тела. Ядра делятся на две части, и в каждой новой инфузории, оказывается по одному большому, и по одному малому ядру. Каждая из двух дочерних инфузорий получает часть органоидов (например, сократи, тельные вакуоли), а другие образуются заново.
При половом процессе увеличения числа особей не происходит. Две инфузории соприкасаются друг с другом, их оболочки в этом месте растворяются, и образуется мостик из цитоплазмы. Большие ядра исчезают, а малые дважды делятся, и в каждой инфузории образуется по четыре малых ядра. Три из них разрушаются, а четвёртое снова делится, так что в каждой инфузории остаётся по два малых ядра. Затем инфузории обмениваются ядрами: по цитоплазматическому мостику одно из ядер от каждой особи переходит в другую особь и там сливается с оставшимся ядром.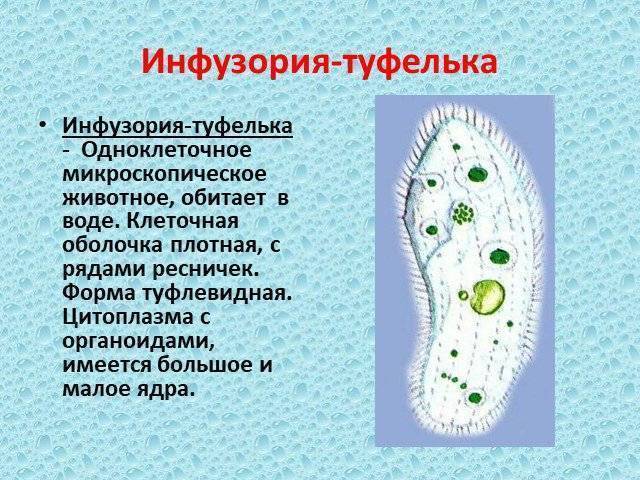 Такой половой процесс называют конъюгацией.
Такой половой процесс называют конъюгацией.
4.Объясните, почему половой процесс не является половым размножением. В чем его биологическое значение?
При половом процессе между клетками происходит обмен наследственным (генетическим) материалом, так что в каждом из участников процесса он обновляется. Это увеличивает жизнестойкость организмов.
Тест по теме «Простейшие», 7 класс
Тема «Простейшие»
вариант № 1
1. В неблагоприятных условиях амеба обыкновенная выделяет вокруг себя плотную
защитную оболочку:
1) цитоплазматическую мембрану 2) ложноножки 3) цисту 4) сократительную
вакуоль
2. Эвглена зеленая относится к типу:
1) жгутиконосцы 2) саркодовые 3) инфузории 4) споровики
3. Органоидами передвижения инфузории-туфельки являются:
1) ложноножки 2) реснички 3) жгутик 4) циста
4. Выберите простейшее, которое может питаться как растение
Выберите простейшее, которое может питаться как растение
1) инфузория-туфелька 2) радиолярии 3) обыкновенная амеба 4) зеленая эвглена
5. У инфузории-туфельки сократительная вакуоль служит для:
1) передвижения 2) выделения вредных веществ 3) питания
4) дыхания.
6. Какой газ выделяют при дыхании простейшие
1) кислород 2) азот 3) углекислый газ 4) угарный газ
7. Малярийный паразит обитает в организме человека в
1) лимфе 2) клетках крови 3) эпителиальной ткани 4) тканевой
жидкости
8. В половом процессе инфузорий основную роль играет
1) малое ядро 2) большое ядро 3) оба ядра 4) цитоплазма
9. Какую функцию в теле инфузории-туфельки выполняют органоиды, обозначенные на
рисунке цифрой 1?
1) выделения из организма вредных веществ 2) передвижения
3) ориентации в среде обитания 4) защиты от механических воздействий среды
10. Какой признак позволяет отнести обыкновенную амёбу к подцарству Простейшие?
1) мелкие размеры 2) обитание в водной среде 3) способность к передвижению
4) одноклеточное строение
11. Сократительная вакуоль присутствует у
Сократительная вакуоль присутствует у
1) пресноводных простейших 2)морских простейших 3) паразитических
простейших 4) всех простейших
12. Установите соответствие между признаками и организмами: к каждой позиции, данной
в первом столбце, подберите соответствующую позицию из второго столбца.
Признаки Организмы
А) наличие в клетке двух ядер 1) Инфузория-туфелька
Б) передвижение с помощью ресничек 2) Амеба обыкновенная
В) осуществление захвата пищи с помощью ложноножками
Г) непостоянная форма тела
Д) размножение только бесполым способом
13. Назовите органоиды эвглены зеленой, обозначенные цифрами 2,3, 4 и их функции.
Тема «Простейшие»
вариант № 2
1. Простейшие в состоянии цисты
1) образуют половые клетки 2) размножаются бесполым путём
3) способны к автотрофному питанию 4) переносят неблагоприятные условия
2. Какую функцию выполняет органоид у зелёной эвглены, обозначенный на рисунке
Какую функцию выполняет органоид у зелёной эвглены, обозначенный на рисунке
вопросительным знаком?
1) обеспечивает реакции на свет 2) контролирует обмен веществ
3) осуществляет автотрофное питание 4) выделяет продукты обмена
3. Жидкие продукты жизнедеятельности у инфузории-туфельки выводятся через
1) порошицу 2) клеточный рот 3) пищеварительные вакуоли
4) сократительные вакуоли
4. Эвглена зеленая отличается от инфузории-туфельки наличием
5. Какой газ выделяют простейшие при дыхании
1) кислород 2) углекислый газ 4) угарный газ 4) азот
6. Что нового появляется у инфузории в отличие от амебы и эвглены?
1) ядро 2) сократительная вакуоль 3) порошица 4) пищеварительная вакуоль
7. Непереваренные остатки пищи у инфузории туфельки выводятся через
Непереваренные остатки пищи у инфузории туфельки выводятся через
1) клеточный рот 2) порошицу 3) сократительную вакуоль
4) пищеварительную вакуоль
8. Укажите функцию органоида обозначенного на схеме строения амёбы цифрой 5?
1) поглощение кислорода и удаления углекислого газа 2) регуляции
обмена веществ 3) удаление остатков непереваренной пищи
4) удаления избытка воды и растворённых веществ
9. Какие простейшие образуют колонии
1) амёба 2) вольвокс 3) лямблии 4) инфузория-туфелька
10. Две сократительные вакуоли имеются у
1) эвглены зелёной 2) амёбы 3) радиолярии
4) инфузории-туфельки
11. Инфузория-туфелька передвигается с помощью
1) ресничек 2) жгутиков 3) ложноножек
12. Установите соответствие между признаками и организмами: к каждой позиции, данной
в первом столбце, подберите соответствующую позицию из второго столбца.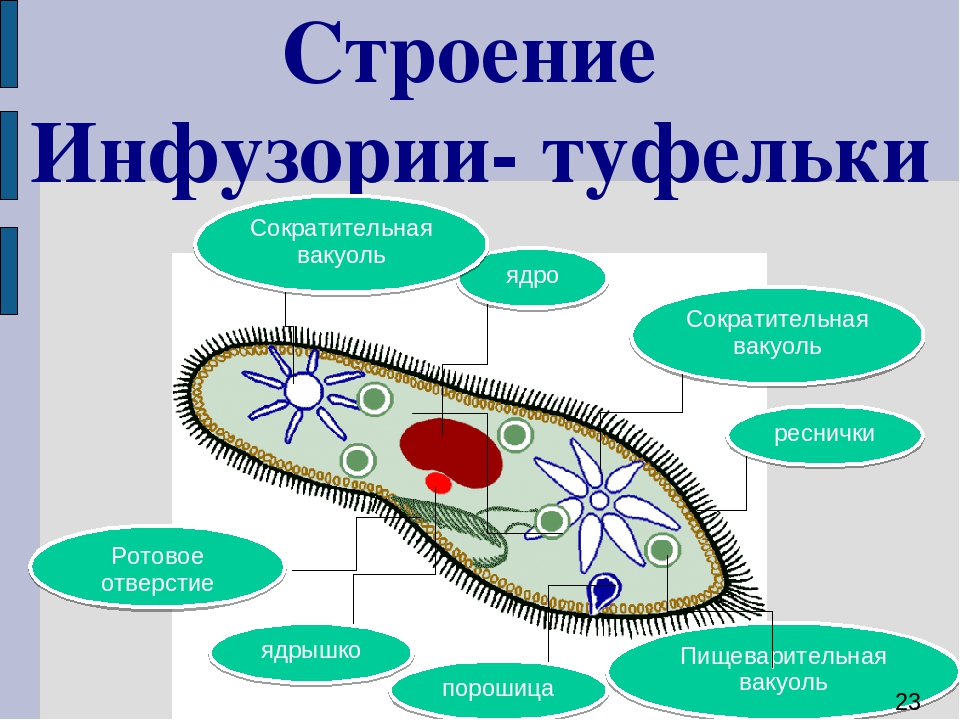
Признаки Организмы
А) наличие в клетке 1 ядра 1) Инфузория-туфелька
Б) передвижение с помощью ресничек 2) Амеба обыкновенная
В) осуществление захвата пищи с помощью ложноножками
Г) непостоянная форма тела
Д) размножение только бесполым и половым способом
13. Назовите органоиды эвглены зеленой, обозначенные цифрами 1,3, 6 и их функции.
ГЛОССАРИЙ КУРСА ЗООЛОГИЯ БЕСПОЗВОНОЧНЫХ — Бирг В.С. Зоология беспозвоночных
ГЛОССАРИЙ КУРСА «ЗООЛОГИЯ БЕСПОЗВОНОЧНЫХ»
(минимальный набор терминов по курсу)
Аборальный орган — орган равновесия и регуляции движения у гребневиков.
Аборальный полюс — полюс тела, противоположный оральному.
Автогамия — разновидность полового процесса у простейших, при котором сливаются два половых ядра, образовавшиеся из одного ядра клетки.
Автотрофный тип питания — тип питания, при котором живые организмы самостоятельно синтезируют органические вещества.
Адолескарии — инцистированные церкарии тех видов трематод, у которых инцистирование происходит во внешней среде.
Акрон — смотри простомиум.
Аксоподии — разновидность псевдоподий, характеризующаяся наличием опорных скелетных образований, расположенных по оси псевдоподии.
Аксостиль — органоид части жгутиконосцев, имеющий вид изогнутого комплекса микротрубочек, имеет опорное значение.
Алиментарный способ заражения — заражение хозяина путем проглатывания инвазионной стадии паразита, локализованной в теле промежуточного хозяина.
Амбулакральная система — особая вододвигательная система у иглокожих, образуется как производное одной из целомических закладок.
Амебоциты — амебоидные клетки в теле губок, выполняют функции захвата пищевых частиц (у части губок), переваривания, транспорта питательных веществ, расположены в мезохилле.
Амфибластула — один из типов организации личинок губок, в этом случае анимальное (верхнее) полушарие состоит из мелких жгутиконосных клеток, а вегетативное (нижнее) из крупных безжгутиковых клеток.
Анаморфоз — тип постэмбрионального развития членистоногих, при котором отрождаются личинки с неполным числом сегментов и их число увеличивается с каждой линькой.
Анизогамия — разновидность копуляции, при которой гаметы нерезко различаются морфологически и физиологически.
Антимеры — одинаковые части тела животного, на которые оно разделяется при проведении плоскости симметрии.
Археоциты — недифференцированные клетки мезохилла губок, способны образовывать клетки других типов, в частности, из них формируются половые клетки и геммулы.
Аскон — наиболее простой тип организации губок, у губок такой организации парагастральная полость выстлана хоаноцитами, поверхность тела — пинакоцитами.
Базальная мембрана — бесклеточный слой, подстилающий эпителий.
Бентос — совокупность организмов, обитающих на дне водоема.
Билатеральная симметрия — такой тип симметрии, при котором через тело животного можно провести только одну плоскость симметрии.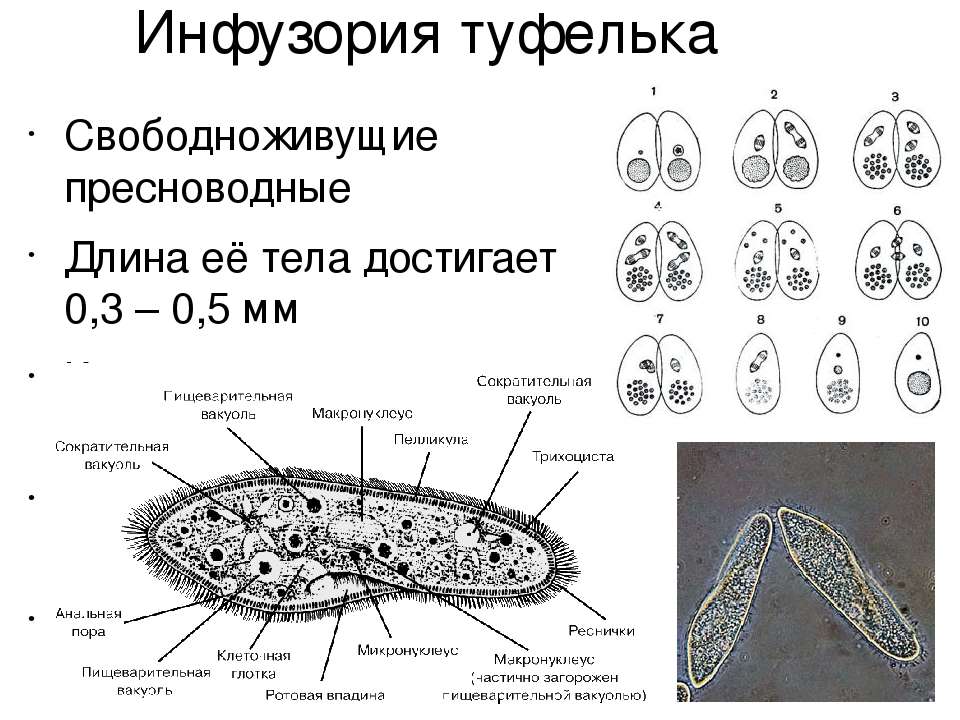
Биофильтрация — способ питания, заключающийся в улавливании из воды, протекающей через специальные структуры животного (у губок — система пор и каналов), пищевых частиц.
Бластея — гипотетический предок многоклеточных, шаровидный бластулообразный организм.
Бластомеры — клетки, возникающие из зиготы в ходе ее дробления, образуют стенку бластулы.
Бластопор — первичный рот, отверстие, через которое первичная кишка сообщается с внешней средой.
Бластостиль — один из типов особей колонии гидроидных полипов, рта не имеет, не питается. Функция — образование медуз.
Бластоцель — полость бластулы. Бластула — стадия развития многоклеточных, возникающая из зиготы путем ее дробления, состоит из бластомеров, имеет форму полого шара.
Велигер — личинка, характерная для большинства моллюсков, видоизмененная трохофора.
Вентральный – брюшной.
Вращательная симметрия — такой тип симметрии, при котором одинаковые части тела животного закономерно повторяются либо вокруг какой-либо точки (центра симметрии), либо оси симметрии, либо плоскости (плоскости симметрии).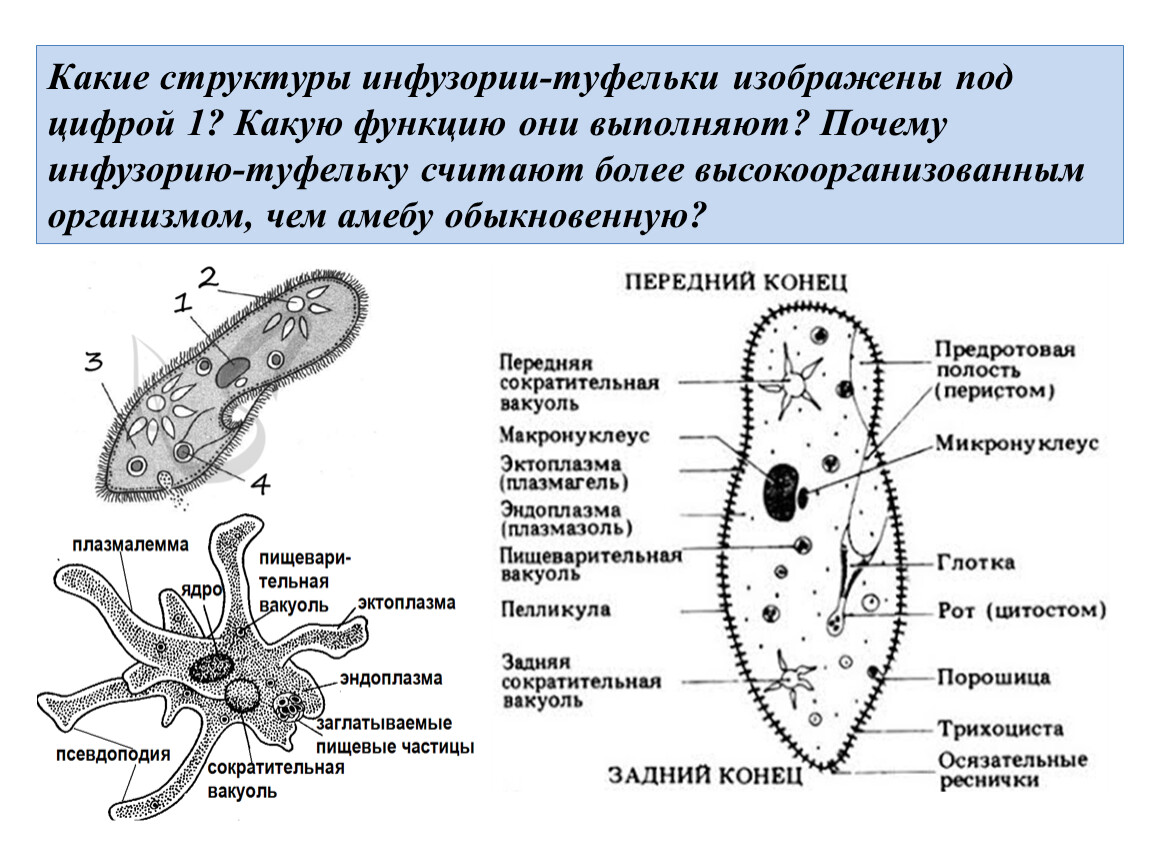
Гамета — половая клетка с гаплоидным набором хромосом.
Гаметическая редукция — разновидность ядерного цикла, при котором мейоз непосредственно предшествует формированию гамет.
Гаметоциста — сферическое образование, одетое плотной оболочкой, внутри которой содержатся гаметы. Характерно для грегарин. Образуется после завершения гамогонии.
Гамонт — стадия жизненного цикла простейших, из которой формируется одна или несколько гамет.
Гамогония — формирование гамет.
Гамонтоциста — сферическое образование, одетое плотной оболочкой, внутри которой содержатся гамонты. Характерно для грегарин. Образуется после приобретения двумя гамонтами полусферической формы. После формирования гамет получает наименование гаметоцисты.
Ганглии — нервные узлы, скопления нервных клеток.
Гастральная полость — полость первичной кишки, полость гаструлы, кишечная полость кишечнополостных.
Гастроваскулярная система — совокупность разветвлений полости первичной кишки кишечнополостных.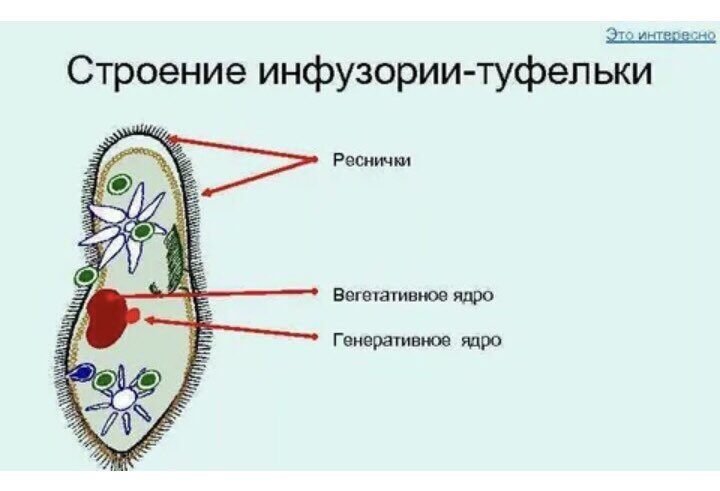
Гастрея — в соответствии со взглядами Э.Геккеля — гипотетический предок многоклеточных, двухслойное животное, имеющее наружный (эктодерма) и внутренний (энтодерма) слои клеток, первичную кишку и бластопор, возникает из бластеи путем впячивания половины клеток во внутрь.
Гаструла — двухслойная стадия развития многоклеточных, возникает из бластулы путем гаструляции, происходящей в той или иной форме. Имеет наружный (эктодерма) и внутренний (энтодерма) слои клеток, первичную кишку и бластопор.
Гаструляция — образование двухслойной гаструлы из бластулы, чаще всего происходит путем инвагинации или иммиграции, реже — деламинацией и эпиболией.
Геммула — внутренняя почка губок, имеет вид шаровидного скопления археоцитов, одетого оболочкой.
Гетерогония — жизненный цикл с правильным чередованием двуполого и гермафродитного размножения.
Гетерономная метамерия — тип метамерного строения тела, в этом случае сегменты разных участков тела организованы различно.
Гетеротрофный тип питания — питание организма готовыми органическими веществами.
Гидрант — один из типов особей колонии гидроидных полипов, характеризуется типичной полипоидной организацией, активно питается.
Гиподерма — разновидность кожного покрова, характеризуется синцитиальным строением, на ее поверхности, как правило, находится кутикула.
Главная ось — прямая линия, соединяющая оральный и аборальный полюса
Глохидий — личинка некоторых пресноводных двустворчатых моллюсков, паразитирующая на коже рыб.
Голозойный тип питания — разновидность гетеротрофного типа питания, при котором организм заглатывает сравнительно крупные, оформленные частицы пищи.
Гомономная метамерия — тип метамерного строения тела, в этом случае сегменты разных участков тела организованы одинаково (или почти одинаково).
Дейтомерит — задний отдел тела грегарин.
Дейтоцеребрум — второй отдел головного мозга членистоногих, иннервирует антеннулы.
Деламинация — способ гаструляции за счет деления клеток в плоскости, параллельной поверхности бластулы.
Детерминативное дробление — тип дробления, при котором на очень ранних стадиях развития определена дальнейшая судьба бластомеров.
Диморфизм — наличие внутри вида двух морфологически отличающихся типов особей (например, полип и медуза у кишечнополостных с метагенезом). Чаще всего представлен половой диморфизм.
Диссепименты — перегородки между следующими друг за другом сегментами тела членистых животных, образованы двойным целомическим эпителием, возникают вследствие того, что в каждом сегменте развивается своя пара целомических зачатков.
Дорзальный — спинной.
Желточник — орган женской половой системы плоских червей, производящий желточные клетки (содержат запас питательных веществ для зародыша). По происхождению является видоизмененным яичником.
Жизненный цикл — морфогенез животного между двумя одноименными стадиями его развития (например, от зиготы до зиготы).
Зигота — результат слияния гамет, обладает диплоидным набором хромосом.
Зиготическая редукция — разновидность ядерного цикла, при котором первое деление зиготы является редукционным.
Изогамия — разновидность копуляции, при которой обе участвующие гамета равноценны морфологически и физиологически.
Имаго — половозрелая стадия насекомых.
Иммиграция — способ гаструляции за счет перемещения части клеток из стенки бластулы в ее полость; может совершаться со всех полюсов бластулы (мультиполярная иммиграция) или преимущественно с одного полюса (униполярная иммиграция).
Инвагинация — способ гаструляции за счет впячивания одной половины бластулы в другую.
Инвертированные глаза — органы зрения, у которых световоспринимающая часть светочувствительной клетки обращена от света.
Инокулятивный путь заражения — способ проникновения инвазионной стадии паразитического животного в тело позвоночного хозяина со слюной кровососа — переносчика при укусе.
Интерстициал — животное, обитающее между частицами субстрата.
Интерстициальные клетки — недифференцированные клетки кишечнополостных, способны образовывать клетки других типов.
Интроверт — способная вворачиваться и выворачиваться передняя часть тела.
Кинетопласт — органоид жгутиконосцев отряда Kinetoplastida, представляет собой относительно обособленную часть их гигантской митохондрии, в кинетопласте сосредоточена митохондриальная ДНК.
Кинетосома — базальная часть жгутика (реснички).
Кинобласт — наружный слой жгутиковых клеток фагоцителлы. Эпителизован. Выполняет функции движения, подгона пищи, восприятия раздражений.
Клоака — задняя часть кишечника в том случае, если туда открываются протоки половых и/или выделительных органов.
Книдоциль — чувствительный вырост стрекательной клетки.
Колленциты — звездчатые клетки, выполняющие опорную функцию, расположены в мезохилле.
Коловращательный аппарат — совокупность определенным образом расположенных ресничек переднего конца тела коловраток, выполняет функцию движения и подгона пищи.
Коксальные железы — выделительные органы свойственные паукообразным. Являются видоизмененными целомодуктами.
Коксальные органы — выворачивающиеся тонкостенные пузыри у основания ног некоторых членистоногих.
Комиссуры — поперечные стволы нервной системы, в нервной системе узлового типа соединяют одноименные ганглии.
Коннективы — продольные стволы нервной системы, в нервной системе узлового типа соединяют разноименные ганглии.
Коноид — органоид Apicomplexa, представляющий собой скопление фибриллярных элементов в форме конуса, обеспечивает проникновение паразита в клетку хозяина.
Контаминативный путь заражения — способ проникновения инвазионной стадии паразитического животного в тело позвоночного хозяина, при котором паразит активно проникает через покровы или слизистую. В этом случае кровосос-переносчик обеспечивает возможность контакта паразита с телом позвоночного.
Конъюгация — форма полового процесса, при которой клетки временно соединяются и обмениваются частями ядерного аппарата, слияния цитоплазмы при этом не происходит.
Копуляция — форма полового процесса, при которой происходит слияние гамет.
Корацидий — личиночная стадия жизненного цикла лентецов, обитает в водной среде.
Ктенидий — тип органов дыхания моллюсков, имеющий вид лопасти с двоякоперистыми жаберными лепестками.
Кутикула — неклеточная наружная часть покрова некоторых беспозвоночных, секретируется эпителием (или гиподермой).
Ларвальные сегменты — личиночные сегменты тела членистых животных. Возникают при превращении трохофоры в метатрохофору.
Лауреров канал — орган женской половой системы трематод, начинается от оотипа, открывается наружу, служит для вывода наружу излишков желточных клеток.
Лейкон — наиболее сложный тип организации губок, у губок такой организации мезохилл утолщен, в его толще образуются камеры, выстланные хоаноцитами, эти камеры соединяются с внешней средой и парагастральной полостью каналами. Парагастральная полость выстлана пинакоцитами.
Личинка — неполовозрелая стадия развития животного, заметно отличающаяся от взрослого по морфологии и экологии.
Лобоподии — разновидность ложноножек, характеризуется небольшой длиной, на конце закруглены.
Ложная кутикула — внутриклеточное защитное образование эпидермиса (или гиподермы).
Мадрепоровая пластинка — скелетная пластинка, пронизанная порами, с которой начинается амбулакральная система иглокожих.
Макрогамета — в случае анизогамии и оогамии — крупная гамета.
Макромеры — в случае неравномерного дробления — крупные бластомеры.
Макронуклеус — крупное ядро инфузорий, содержащее полиплоидное количество хромосом, регулирует обмен веществ.
Максиллы — нижние челюсти членистоногих, являются видоизмененными конечностями.
Мальпигиевы сосуды — тип органов выделения, свойственный паукообразным и трахейнодышащим. Представляют собой выросты кишечника, причем у трахейнодышащих они эктодермальны, а у паукообразных — энтодермальны.
Мандибулы — верхние челюсти членистоногих, являются видоизмененными конечностями.
Мантия — складка кожи, полностью или частично покрывающая тело животного.
Марита — гермафродитная стадия жизненного цикла трематод (раздельнопола только у кровяных двуусток), паразитирующая в позвоночном хозяине.
Медуза — одна из двух форм тела кишечнополостных. Пелагическая форма, оральный полюс обращен вниз, тело сплющено в плоскости, перпендикулярной главной оси, тело дисковидное или зонтиковидное, щупальца расположены по краю тела.
Мезентерий — двухслойная продольная перегородка, возникающая у вторичнополостных животных за счет того, что целом закладывается парно, справа и слева от кишечника. Стенки целомических мешков соприкасаются над и под кишечником. За счет этого образования кишечник подвешен к стенке тела.
Мезенхима — совокупность диффузно расположенных клеток между эктодермой и энтодермой.
Мезодерма — третий зародышевый листок, формируется между эктодермой и энтодермой.
Мезоглея — неклеточный слой между эктодермой и энтодермой у кишечнополостных (может содержать клетки, которые проникают в мезоглею за счет выселения из эктодермы или энтодермы).
Мезохилл — совокупность элементов, лежащих у губок между слоем пинакоцитов и слоем хоаноцитов. Содержит студенистый белковый матрикс и многообразные клеточные элементы. В базовых учебниках применяется термин «мезоглея», однако мезохилл губок не равнозначен мезоглее кишечнополостных.
Мерозоит — стадия жизненного цикла споровиков, возникающая в результате шизогонии.
Метамеры — одинаковые части тела животного, повторяющиеся вдоль оси тела.
Метаморфоз — изменение строения тела во время превращения личинки во взрослое животное.
Метанефридии — отдельный каналец метанефридиальной выделительной системы, На внутреннем конце открывается воронкой в целом. Имеет эктодермальное происхождение.
Метатрохофора — личиночная стадия кольчатых червей, формируется из трохофоры за счет образования ларвальных сегментов.
Метацеркарии — инцистированные церкарии тех видов трематод, у которых инцистирование происходит в организме второго промежуточного хозяина.
Метагенез — жизненный цикл с правильным чередованием полового и бесполого размножения.
Микрогамета — в случае анизогамии и оогамии — мелкая гамета.
Микромеры — в случае неравномерного дробления — мелкие бластомеры.
Микронуклеус — малое ядро инфузорий, содержащее диплоидное число хромосом, ответственно за передачу наследственной информации при половом процессе.
Миксоцель — особый тип полости тела (смешанная полость тела), свойственная членистоногим. В ходе ее формирования объединяются первичная и вторичная полости тела..
Миоциты — клетки, способные сокращаться, содержат актин-миозиновый комплекс.
Мирацидии — личинки трематод, свободно плавающие, покрытые ресничками, после проникновения в первого промежуточного хозяина превращаются в спороцисту.
Монотомия — способ деления клеток простейших на две одинаковые дочерние клетки, при котором после деления происходит рост дочерних клеток до достижения размеров материнской клетки.
Моноэнергидная клетка — клетка, имеющая одно ядро с гаплоидным или диплоидным набором хромосом.
Науплиус — личинка ракообразных, по строению соответствует метатрохофоре кольчатых червей.
Наяда — водная нимфа у насекомых, имеющая специальные приспособления к обитанию в воде.
Нейросекреторная клетка — нервная клетка, способная выделять гормоны.
Нейстон — животные, обитающие на поверхности воды.
Нектон — животные, обитающие в толще воды и способные плыть против течения.
Неотения — способность животных размножаться на личиночной стадии.
Нефромиксии — орган некоторых полихет, образовавшийся в результате срастания целомодукта и протонефридия (метанефридия).
Нимфа — ювенильная стадия развития клещей и насекомых, мало отличающаяся от взрослой особи (за исключением размеров и развития некоторых систем органов).
Окончательный хозяин — хозяин, в котором паразит проходит стадию полового размножения.
Омматидий — простой глазок, входящий в состав сложного глаза членистоногих.
Олигомерный — имеющий малое число сегментов.
Онкосфера — личиночная стадия жизненного цикла цепней, обитает в организме промежуточного хозяина.
Оогамия — форма полового процесса, при которой гаметы резко различны морфологически и физиологически.
Оокинета — зигота, характеризующаяся способностью к движению, характерна для Haemosporidia.
Оотип — орган женской половой системы плоских червей, в котором происходит образование сложного яйца. Ооциста — зигота, одетая плотной оболочкой.
Оральный полюс — полюс, на котором возникает бластопор.
Ортогон — один из типов организации нервной системы. Включает несколько продольных нервных стволов, соединенных кольцевыми комиссурами.
Ось симметрии — прямая линия, проходящая через тело животного так, что одинаковые части (антимеры) в правильном порядке расположены вокруг этой линии, иначе говоря, при вращении вокруг оси симметрии на определенный угол тело животного совмещается сама с собой.
Оскулум — устье, отверстие, через которое вода из парагастральной полости губок поступает во внешнюю среду.
Осфрадии — органы химического чувства моллюсков, расположенные в мантийной полости.
Палинтомия — способ деления клеток простейших на две одинаковые дочерние клетки, при котором после деления не происходит роста клеток, так что с каждым последующим делением размер дочерних клеток уменьшается вдвое.
Парагастральная полость — пространство в теле губки, куда поступает вода из пор или каналов и откуда эвакуируется наружу через оскулум. Иногда называется атриальной полостью.
Параподия — парные метамерные боковые выросты тела полихет, несут локомоторную функцию. Могут выполнять также дыхательную функцию.
Паренхима — ткань из аморфно расположенных клеток, расположена между эктодермой и энтодермой, выполняет различные функции (транспортную, опорную, формирования половых продуктов, запасающую).
Паренхимула — один из типов организации личинок губок, у личинок этого типа жгутиконосные клетки расположены на поверхности, а амебоидные — внутри. Возникает их целобластулы путем иммиграции.
Партеногенез — разновидность полового размножения, в этом случае новая особь развивается из яйцеклетки без оплодотворения.
Педицеллярии — подвижные скелетные образования иглокожих, имеющие форму щипцов (пинцетов), очищающие кожу от инородных частиц.
Пелликула — комплекс мембраны простейшего и лежащих под ней субмембранных образований.
Пенис — копулятивный орган, не способный выворачиваться в ходе осуществления полового акта.
Первичная полость тела — полость тела, не имеющая собственных стенок и непосредственно граничащая с органами. Возникает за счет расхождения или редукции клеток паренхимы.
Перикардий — околосердечная сумка, полость ее представляет собой участок целома, стенка образована целомическим эпителием.
Перистомиум — сложный сегмент кольчатых червей, возникающий путем срастания нескольких сегментов передней части тела. Возникновение перистомиума расценивают как начальный этап цефализации. Несет на себе рот животного.
Пероральный путь заражения — способ проникновения инвазионной стадии паразитического животного в тело в тело хозяина путем проглатывания (per os — через рот). Реализуется фекально-оральным способом или алиментарным способом.
Пигидиум — анальная лопасть кольчатых червей, является неметамерной частью тела. У членистоногих гомологичная часть тела называется тельсон.
Пилидий — тип личинки, свойственный немертинам.
Пинакоциты — покровные клеточные элементы тела губок.
Пиноцитоз — поглощение клеткой жидкостей за счет впячивания клеточной мембраны.
Плазмодий — многоядерная амебоидная клетка.
Плакула — в соответствии с взглядами О. Бючли — гипотетический предок многоклеточных, имеющий вид пластинчатой колонии жгутиконосцев.
Планктон — организмы, обитающие в толще воды и не способные плыть против течения.
Планула — личинка кишечнополостных. Характерен наружный ресничный покров, небольшая гастральная полость, отсутствие рта, передне — задняя полярность.
Плейстон — полупогруженные в воду плавающие организмы.
Плероцеркоид — личиночная стадия жизненного цикла лентецов, обитает в организме второго промежуточного хозяина.
Плоскость симметрии — плоскость, которая разделяет тело на две зеркально-равные части, расположенные относительно друг друга как предмет и его зеркальное отражение.
Погруженный эпителий — тип организации эпителия, в этом случае апикальные части клеток образуют над базальной мембраной синцитий, а базальные части клеток расположены под базальной мембраной и сохраняют индивидуальность.
Полимерный — имеющий большое число сегментов.
Полиморфизм — наличие внутри вида более двух морфологически отличающихся типов особей (примером являются Siphonophora).
Полип — одна из двух форм тела кишечнополостных, сидячее (редко — слабо подвижное) животное цилиндрической формы, к субстрату обращен аборальный полюс, характерно наличие венчика щупалец вокруг рта.
Полиэнергидная клетка — клетка, имеющая два или более ядер (с гаплоидным или диплоидным набором хромосом).
Пороциты — клетки тела губок, образующие поры.
Постларвальные сегменты — совокупность сегментов тела членистых животных, возникающих после и позади ларвальных сегментов за счет поочередного образования в зоне роста.
Поступательная симметрия — такой вид симметрии, при котором одинаковые части тела животного (метамеры) повторяются вдоль тела животного.
Почкование — способ деления клеток простейших, при котором дочерние клетки резко различаются по размерам.
Проглоттид — членик ленточных червей.
Промежуточная редукция — разновидность ядерного цикла, при котором редукционное деление находится между двумя вегетативными стадиями.
Промежуточный хозяин — хозяин, в котором не происходит полового размножения паразита (возможно прохождение бесполого размножения).
Протоподит — основание конечности членистоногих.
Простомиум — головная лопасть кольчатых червей, является неметамерной частью тела. У членистоногих гомологичная часть тела называется акрон.
Протомерит — второй отдел тела грегарины.
Протонефридий — отдельный каналец протонефридиальной выделительной системы, на внутреннем конце несет терминальную клетку с мерцательным пламенем. Имеет эктодермальное происхождение.
Протоцеребрум — передний отдел головного мозга членистоногих, иннервирует акрон и глаза.
Процеркоид — личиночная стадия жизненного цикла лентецов, обитает в первом промежуточном хозяине.
Рабдиты — защитные структуры эпителия плоских червей.
Радиальная симметрия — разновидность вращательной симметрии, при которой одинаковые части тела животного закономерно повторяются вокруг оси симметрии, у многоклеточных ось симметрии гетерополярна.
Радула — терка, пластинка с набором зубчиков в глотке большинства моллюсков.
Редия — стадия жизненного цикла трематод, обитающая в первом промежуточном хозяине, возникает в результате размножения спороцист, размножается партеногенетически.
Ризоподии — разновидность ложноножек, характеризуются разветвленностью и способностью образовывать анастомозы.
Ропалии — сложные комплексы органов чувств, характерные для сцифоидных медуз.
Роптрии — парные органоиды Apicomplexa, имеющие вид мешковидных структур, лежащих позади коноида. Заполнены ферментами, выделяющимися наружу в ходе проникновения в клетку хозяина.
Сенсиллы — чувствительные клетки беспозвоночных, имеет чувствующий отросток и отросток, идущий к центральной нервной системе.
Септы — складки энтодермы (между двумя слоями энтодермы находится мезоглея) в гастральной полости некоторых кишечнополостных полипоидной организации.
Сизигий — результат соединения двух гамонтов, эта стадия характерна для грегарин.
Сикон — один из типов организации тела губок. У губок такого типа имеются карманы, выстланные хоаноцитами, которые каналами связаны с наружной стенкой тела, а широким основанием открываются в парагастральную полость. Парагастральная полость выстлана пинакоцитами.
Симметрия — (в зоологическим смысле) закономерная повторяемость одинаковых частей тела животного в пространстве.
Синкарион — ядро, возникающее при слиянии половых ядер инфузорий.
Синцитий — многоклеточная структура с частично или полностью отсутствующими границами между клетками, часто представляет собой многоядерную цитоплазматическую массу.
Сифоноглифы — ротовой желобок, находится на одном или обоих краях щелевидно сплющенной глотки коралловых полипов.
Склеробласты — клеточные элементы мезохилла губок, формирующие скелетные образования.
Сколекс — «головка» — начальная часть тела ленточных червей, несущая органы прикрепления.
Сложное яйцо — яйцо, включающее — кроме яйцеклетки — желточные клетки, несущие запас питательного материала, характерно для плоских червей.
Спикулы — скелетные образования губок в виде одноосных или многоосных игл.
Спиральное дробление — тип дробления, при котором направление веретена деления закономерно изменяется, вследствие этого бластомеры ложатся не точно друг над другом, а чередуясь.
Спонгин — рогоподобное эластичное вещество, выделяемое особыми клетками мезохилла губок, сеть волокон спонгина выполняет скелетную функцию.
Спорогония — процесс формирования спорозоитов из зиготы, включает редукционное деление и митоз.
Спорозоит — стадия жизненного цикла споровиков, возникающая из зиготы в результате редукционного деления и последующего митоза. Служит для заражения следующей особи хозяина.
Спороциста — стадия жизненного цикла трематод, обитающая в первом промежуточном хозяине, возникает как преобразование мирацидия, размножается партеногенетически.
Сократительная вакуоль — органоид простейших, имеющий вид пузырька, осуществляющего регулярную пульсацию. Главная функция — осморегуляция, также выполняет функцию выделения жидких продуктов обмена веществ.
Стробиляция — своеобразный процесс почкования сцифистом, приводящий к отделению эфир.
Стробила — у ленточных червей — совокупность всех члеников тела. У сцифоидных медуз — совокупность особей возникающих в ходе стробиляции, до тех пор, пока они соединены вместе.
Сцифистома — полипоидная стадия развития сцифоидных медуз.
Тагма — отдел тела (у членистоногих).
Тегумент — погруженный эпителий без ресничек.
Телобласты — клетки, из которых образуется все мезодерма трохофорных животных.
Тельсон — смотри пигидиум.
Тельце Мелиса — орган половой системы плоских червей, имеет вид совокупности одноклеточных желез, окружающих оотип, выделяет жидкость, заполняющую оотип и матку.
Тифлозоль — складка стенки кишечника, вдающаяся в полость кишки. Характерна для олигохет.
Трансмиссивное заболевание — возбудители которого передаются через животных-переносчиков, являющихся кровососами.
Трахеи — органы дыхания трахейнодышащих и некоторых паукообразных. Имеют вид трубок, начинающихся на поверхности тела и заканчивающихся в полости тела или в органах. Строго говоря, выполняют роль транспорта газов, газообмен происходит только в тканях.
Тритоцеребрум — задний отдел головного мозга членистоногих. Относится по происхождению к первому сегменту тела, иннервирует антенны.
Трихоцисты — защитные образования инфузорий, имеют форму короткой палочки, располагаются в кортексе, при раздражении выстреливают наружу, превращаясь в длинную нить .
Трофосома — орган погонофор (Vestimentifera), в котором обитают симбиотические хемосинтезирующие бактерии, вероятно, является видоизмененным кишечником, утратившим сообщение с внешней средой.
Трохофора — тип организации личинки многих кольчатых червей и моллюсков.
Фагоцитоз — поглощение клеткой сравнительно крупных оформленных частиц за счет впячивания мембраны.
Фагоцителла — в соответствии с воззрениями И.И. Мечникова — гипотетический предок многоклеточных, возникающий из шаровидной колонии жгутиконосцев за счет перемещения (иммиграции) отдельных клеток бластулы в бластоцель, причем это перемещение обусловлено захватом пищевой частицы и ее фагоцитозом.
Фагоцитобласт — внутренний пласт безжгутиковых клеток фагоцителлы. Характерно аморфное расположение. Выполняет функции пищеварения и распределения питательных веществ.
Фекально-оральный способ заражения — способ заражения, при котором инвазионная стадия (как правило, снабженная защитными оболочками), попадает во внешнюю среду из кишечника одного хозяина и случайно проглатывается другим хозяином с загрязненной пищей или водой.
Филоподии — разновидность ложноножек, характеризуются нитевидной формой, способны образовывать анастомозы. От ризоподий отличаются отсутствием микротрубочек.
Финна — пузыревидная стадия в жизненном цикле ленточных червей.
Хиастоневрия — перекрест плевровисцеральных стволов нервной системы, характерен для части брюхоногих моллюсков.
Хлорагогенные клетки — несколько измененные клетки внутреннего целомического слоя, составляющие наружную стенку средней кишки у олигохет. Участвуют в процессах выделения.
Хоаноциты — клеточные элементы тела губок, имеющие жгутик и цитоплазматический воротничок, выполняют вододвигательную функцию и (у многих губок) функцию захвата пищевых частиц.
Целлюляризация — гипотетический способ формирования многоклеточного строения из полиэнергидного одноклеточного за счет обособления вокруг каждого ядра участка цитоплазмы.
Целобластула — один из типов организации личинок губок, целобластула состоит из одинаковых жгутиконосных клеток.
Целом — вторичная полость тела. Отличительный признак — наличие собственных стенок, мезодермальной выстилки. Закладывается в форме парных мешковидных мезодермальных мешочков в каждом сегменте тела.
Целомодукт — парный канал, одним концом отирающийся в целом, другим — наружу. Имеет мезодермальное происхождение. Первичная функция — выведение половых продуктов, часто выполняет также функцию органа выделения.
Ценур — разновидность финны, способной образовывать (путем бесполого размножения) несколько сколексов.
Центр симметрии — воображаемая точка внутри тела животного. Любая проведенная через эту точку прямая по обе стороны от нее и на равных расстояниях встречает одинаковые (соответственные) точки тела.
Церкарии — личиночные стадии жизненного цикла трематод, возникают в результате партеногенетического размножения редий (или дочерних спороцист), выходят во внешнюю среду, выполняют поисково-расселительную функцию.
Церкомер — хвостовой придаток с тремя парами эмбриональных крючьев у цистицеркоида.
Цикломорфоз — закономерные сезонные изменения морфологического строения разных поколений, не связанные с типом размножения. Характерен для дафний, коловраток.
Циррус — копулятивный орган плоских червей, выворачивающийся в ходе осуществления полового акта.
Циста — стадия жизненного цикла, характеризующаяся наличием защитной оболочки.
Цистицерк — наиболее распространенный тип финны, имеет вид пузырька, в полость которого ввернут сколекс.
Цистицеркоид — разновидность личинки ленточных червей, имеет пузыревидную переднюю часть (в нее ввернут сколекс) и хвостовой придаток с тремя парами эмбриональных крючьев.
Шизогония — способ деления клеток простейших, при котором сначала несколько раз делится ядро, а затем вокруг каждого из ядер обособляется участок цитоплазмы.
Шизонт — стадия жизненного цикла споровиков, размножающаяся шизогонией.
Экзоподит наружная ветвь конечности у членистоногих.
Эктодерма — наружный слой клеток гаструлы, а также наружный слой клеток кишечнополостных и гребневиков.
Энергида — часть цитоплазмы клетки, обмен веществ которой регулируется одним ядром.
Эндон — скопление нервных клеток, лежащее под органом равновесия. В ходе эволюции выделяется из общего диффузного нервного сплетения. Часто принимает на себя функции общего координирующего центра, в этом случае именуется церебральным ганглием или эндонным мозгом.
Эндоподит — внутренняя ветвь конечности у членистоногих.
Энтодерма — внутренний слой клеток гаструлы, а также внутренний слой клеток кишечнополостных и гребневиков.
Эпиболия — способ гаструляции, заключающийся в обрастании подвижными, бедными желтком клетками клеток, богатых желтком.
Эпимерит — передний отдел тела грегарины, обычно несет органоиды прикрепления.
Эпиподит — нечленистый отросток протоподита конечности членистоногих, выполняющий функции дыхания. Может быть разветвленным.
Эфира — личинка сцифоидных медуз.
Эхинококк — наиболее сложный тип строения пузыревидной стадии жизненного цикла ленточных червей. Отличается крупными размерами и формированием внучатых поколений.
Ювенильный — неполовозрелый.
Ядерный дуализм — наличие в клетке ядер двух типов: макронуклеус (ы) — полиплоиден, управляет вегетативными функциями, микронуклеус (ы) — диплоиден, ответственен за половой процесс.
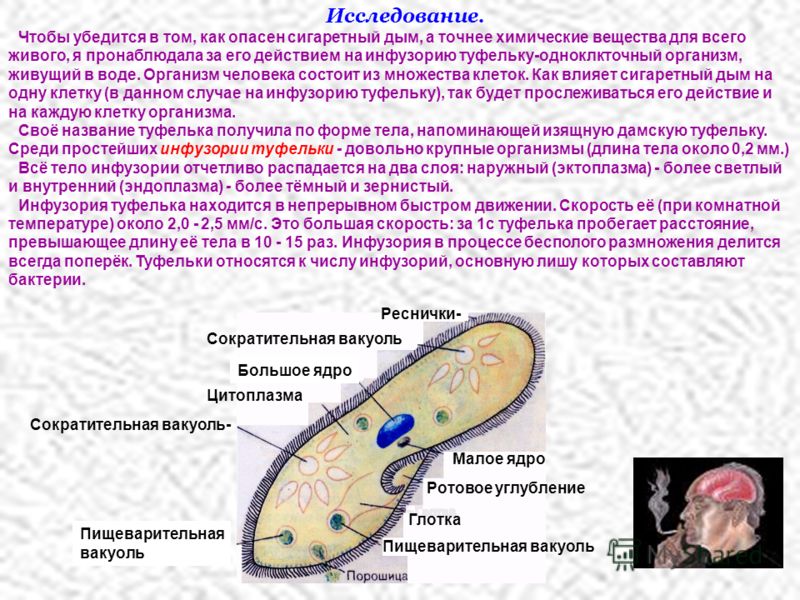
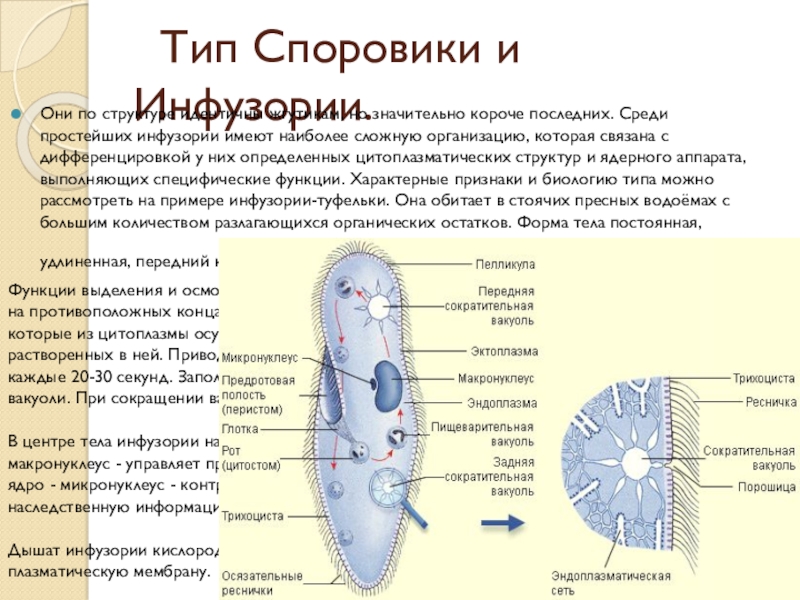
Пелликула инфузорий
Определение 1
Пелликула инфузорий – это оболочка простейших одноклеточных организмов, которая имеет весьма плотную структуру и позволяет им поддерживать форму и также защищает от негативного воздействия окружающей среды.
Особенности пелликулы инфузорий
Инфузории имеют весьма разнообразную форму. Они могут быть:
- овальными;
- продолговатыми.
Размеры инфузорий варьируются в пределах от тридцати до сорока микрометров. Это наиболее сложно устроенные простейшие, которые имеют весьма оригинально устроенные покровы. Цитоплазма их всегда имеет два слоя:
- наружный – эктоплазма или эластичная пелликула;
- внутренний – эндоплазма.
Снаружи пелликула часто имеет скульптурированную структуру с закономерно расположенными утолщениями. Утолщения инфузории туфельки в области пелликулы имеют форму правильных шестиугольников, напоминающих пчелиные соты. Для пелликулы такая структура весьма оправдана, поскольку существенно повышает прочность данного образования.
Снаружи тело инфузории покрывается ресничками, которые начинаются от пелликулы и ее базальных телец. Количество ресничек весьма большое и может составлять до 10 тысяч. Согласно внутренней структуре реснички состоят из двух фибрилл и девяти фибрилл, доходящих до периферии.
Они продолжаются в кинетосоме и приобретают тройную структуру. Если реснички располагаются не рядами, а равномерно, то это является примитивным признаком инфузорий. Локомоторный аппарат инфузорий специализируется в двух направлениях:
- реснички концентрируются на определенных участках тела;
- реснички слипаются и сохраняют собственную индивидуальность в составе крупных рабочих комплексов.
Когда реснички соединяются в один или несколько рядов, то образуется мерцательная перепонка, которая находится на пелликуле. Все эти структуры получили название мембранеллы. Если соединяются реснички, расположенные на пелликуле в виде кисточки, то образуется цирри.
Самый сложный аппарат ресничек находится у инфузорий в области ротового отверстия. Существуют такие инфузории, которые имеют сократительные волоконца или мионемы, они способны к резкому сокращению. Это существенно увеличивает возможность движения инфузорий, несмотря на плотность пелликулы.
В пелликуле инфузорий присутствуют специализированные образования, которые называются трихоцисты.
Определение 2
Трихоцисты – это маленькие защитные веретенообразные образования которые расположены в цитоплазме инфузории перпендикулярно поверхности ее тела. При действии каких-либо внешних раздражителей они выбрасываются наружу и принимают форму длинных нитей с остриями на концах.
Помимо того в пелликуле инфузорий присутствуют своеобразные структуры в виде блях и шестиугольников, подобные трихоцистам. Их нити тоже способны скручиваться и раскручиваться. У некоторых инфузорий пелликула может иметь стекловидную структуру, и она может располагаться в виде складок.
Другие инфузории имеют нежную панцеревидную пелликулу. Перистом в виде узкой полоски от переднего полюса до середины тела. Край перистома (первичного отдела клеточного рта) окружен двумя мембранами.
Другие особенности организма инфузорий
Наиболее типичное строение имеет пелликула инфузории – туфельки. Она называется видом простейшим из класса ресничных. Ее название произошло от формы тела.
Эти живые организмы являются обитателями пресных водоемов любого типа, особенно много инфузорий находится в местах со стоячей водой. Среди простейших организмов инфузория имеет относительно крупные размеры, которые достигают половины миллиметра.
Под наружной мембраной у инфузории туфельки располагается пелликула. У этих особей она представляет собой плотный цитоплазматический слой с цистернами – альвеолами и микротрубочками, и другими элементами цитоскелета.
Поскольку пелликула имеет эластичную структуру, инфузория может менять направление своего тела при столкновении с препятствием. Причем этот процесс происходит моментально.
Два ядра инфузории-туфельки имеют разное строение и выполняют различные функции. Малое ядро диплоидное, имеет округлую форму; большое ядро полиплоидное, имеет бобовидную форму. Малое ядро отвечает за половое размножение, а большое ядро руководит синтезом всех белков клетки инфузории-туфельки.
Питание инфузории также обусловлено строением пелликулы. Они могут проглотить взвешенные в воде частички, которые имеют для животного особенную пищевую ценность. Клеточный рот инфузории продолжается в клеточную глотку, возле которого находится комплекс специализированных ресничек, собирающихся в сложные комплексы. Они производят волнообразные движения, и пища попадает в глотку с потоком воды.
У основания глотки может быть сформирована достаточно крупная пищеварительная вакуоль, ускоряющая всасывание питательных веществ. Они возвращаются в цитоплазму и используются для нужд организма. Все ненужные вещества выводятся в окружающую среду через порошицу.
Порошица – это участок тела инфузории с недоразвитой пелликулой.
В структуре вакуоли выделяют резервуар и канальцы. Вода из цитоплазмы попадает в резервуар и по канальцам она поступает в резервуар, выталкиваюсь наружу через пору. Главная функция сократительных вакуолей называется осморегуляцией. Сократительные вакуоли позволяют удалить те остатки азотистого обмена, которые имеют сложную структуру и не могут пройти через пелликулу.
Дыхание инфузории также связано с переходом газов через пелликулу, которая несмотря на свою плотность дает газам возможность диффундировать через всю поверхность тела. А при пониженной концентрации кислорода в воде инфузория живет за счет гликолиза.
Тест: Одноклеточные животные
ФИО автора материала – Павлова Оксана Анатольевна
Место работы (название образовательного учреждения) – МБОУ «СОШ №22» г. Череповец
Должность автора – учитель биологии
Тема 1. Тип Простейшие
Внимательно прочитайте задание. Продумайте ответ и запишите его в тетради для проверочных работ.
Вариант №1
I. Напишите определения следующих понятий: жгутики, порошица, органоиды.
II. Выберите правильный ответ:
Изучение зеленой эвглены позволяет сделать вывод о родстве растений и животных, так как она:
а) имеет светочувствительный глазок
б) дышит всей поверхностью тела
в) питается как растение и как животное
г) при дыхании поглощает кислород
2. Цистой называется…
а) состояние одноклеточных организмов, при котором образуется плотная оболочка
б) плотная оболочка клетки
в) название простейшего
г) название заболевания, вызванного простейшими
3. Какие из перечисленных простейших не являются паразитами?
а) малярийный плазмодий
б) дизентерийная амеба
в) вольвокс
г) лямблия
4. Строение какого животного опровергает утверждение, что все простейшие – это
одноклеточные животные?
а) инфузории-туфельки в) фораминиферы
б) лямблии г) вольвокса
5. Поглощение кислорода у амёбы обыкновенной осуществляется…
а) пищеварительной вакуолью в) стигмой
б) сократительной вакуольюг) всей поверхностью тела
6. Какую функцию выполняет малое ядро у инфузории-туфельки?
а) дыханияв) пищеварения
б) участвует в половом процессег) регуляция обменных процессов
7. Цисты у одноклеточных выполняют функцию…
а) фотосинтезав) защиты и распространения
б) размноженияг) движения
III. Напишите название животного, изображенного на рисунке. Зарисуйте в тетрадь и подпишите органоиды.
IV. Найдите соответствие между названиями простейших и их особенностями:
Название простейшего | Характеристика |
А. Инфузория-туфелька | 1. Паразит |
Б. Дизентерийная амеба | 2. Колония |
В. Вольвокс | 3. Одиночный организм |
4. Передвижение при помощи ложноножек | |
5. Передвижение при помощи ресничек | |
6. Передвижение при помощи жгутиков | |
7. Два разных по величине ядра | |
8. Наличие хлорофилла |
Тема 1. Тип Простейшие
Внимательно прочитайте задание. Продумайте ответ и запишите его в тетради для проверочных работ.
Вариант №2
I. Напишите определения следующих понятий: циста, ложноножки, сократительная вакуоль.
II. Выберите правильный ответ:
Только паразитический образ жизни ведут следующие простейшие…
а) саркодовые (корненожки) в) споровики
б) инфузории г) жгутиконосцы
2. Малярийный плазмодий распространяется:
а) самостоятельно
б) при помощи комара-анофелеса
в) при помощи мухи це-це
г) при помощи крупного рогатого скота
3. Какие простейшие животные могут питаться как растения и как животные?
а) саркодовые (корненожки) в) инфузории
б) жгутиконосцыг) споровики
4. В организме какого простейшего находятся 2 ядра: большое и малое?
а) амеба дизентерийнаяв) амеба обыкновенная
б) эвглена зеленаяг) инфузория-туфелька
Органоидами движения у амёбы обыкновенной являются…
а) жгутикив) трихоцисты
б) ресничкиг) ложноножки
6. Какую функцию выполняет большое ядро у инфузории-туфельки?
а) дыханияв) пищеварения
б) участвует в половом процессег) регуляция обменных процессов
Светочувствительный глазок есть у…
а) малярийного плазмодияв) амёбы
б) эвглены зелёнойг) лямблии
III. Напишите название животного, изображенного на рисунке. Зарисуйте в тетрадь и подпишите органоиды.
IV. Найдите соответствие между понятиями и характеристиками:
Понятие | Характеристика |
А. Хозяин | 1. Животное, которое переносит паразита от одного животного или человека к другому |
Б. Переносчик паразита | 2. Организм, на котором обитает другое животное, приносящее вред первому |
В. Паразит | 3. Животное, которое обитает в организме другого животного или человека и приносит ему вред |
4. Малярийный комар | |
5. Малярийный плазмодий | |
6. Дизентерийная амеба | |
7. Человек |
ПРОВЕРКА И ОЦЕНКА
Коды ответов
Тема 1. Тип Простейшие
Вариант №1
I.
Жгутики – органоиды движения жгутиконосцев.
Порошица – отверстие в мембране, через которое происходит выброс непереваренных частиц из клетки.
Органоиды – постоянные структуры клетки, которые выполняют определенные функции.
II.
III. Инфузория-туфелька:
1 – реснички
2 – большое ядро
3 – пищеварительная вакуоль
4 – сократительная вакуоль
5 – рот
6 – порошица
IV.
Вариант №2
I.
Циста – состояние одноклеточных организмов, при котором образуется плотная оболочка.
Ложноножки – временные выросты цитоплазмы для передвижения корненожек.
Сократительная вакуоль – органоиды, выполняющие функцию выделения в организме простейших.
II.
III. Амёба протей:
1 – ложноножки
2 – сократительная вакуоль
3 – пищеварительная вакуоль
4 – ядро
5 – цитоплазма
IV.
Адрес публикации: https://www.prodlenka.org/metodicheskie-razrabotki/137594-test-odnokletochnye-zhivotnye
Какую функцию выполняет малое ядро инфузории?
Изменчивость климата(влажность, жара, холод, и тп)природных зон(болотистая местность, пустыня, тундра, джунгли и тп), рельефов, количество добычи / растений.
1) Крахмал является Субстратом в данном опыте. А ферментом является слюна,точнее фермент содержащийся в ней — Амилаза.
2) Нет,не могла.Так как в этом опыте, на накрахмаленном бинте рисуют букву слюной. А фермент Амилаза,входящий в состав слюны,расщепляет крахмал до дисахарида мальтозы. Поэтому буква получится бесцветной.
3) Нет,слюна не будет расщеплять крахмал,если ее прокипятить. Так как при нагревании ее более 50°, фермент Амилаза теряет активность,следовательно, крахмал не гидролизуется(не расщепляется).
Погладь меня. Мне не нравится что ты снимаешь ( фотографируешь ) меня.
Тебе нужно посмотреть раздел в учебнике (если он у тебя есть!)) ) про скелет, у у рептилий добавились шейные позвонки, стал более развит крестец (таз), само слово пресмыкающиеся, они же рептилии, идет от того, что они пресмыкаются к земле (крокодил). Дыхательная система. Легкие пресмыкающихся сохраняют мешкообразное строение, но их внутренняя структура много сложнее, чем у земноводных. Внутренние стенки легочных мешков имеют складчатое ячеистое строение, напоминающее пчелиные соты, что значительно увеличивает дыхательную поверхность.
Выделительная система. У рептилий туловищная почка заменяется тазовой, или вторичной, почкой (метанефрос) . Располагаются почки в тазовой области с брюшной стороны клоаки и по бокам от нее. Почки продолговатой формы, покрыты брюшиной только с нижней стороны. От почек отходят тончайшие мочеточники, которые открываются на спинной стороне клоаки.
В отличие от амфибий мозжечок пресмыкающихся крупный, что отвечает большей сложности и интенсивности их движений.
Головной мозг делится на пять отделов: передний, промежуточный, средний, мозжечок и продолговатый .
Усложнение движений и развитие общений между животными у пресмыкающихся связано с прогрессивным развитием органов чувств. Также сердечная система подвергается эволюции-кровь, идущая к мозгу становится более артериальной (насыщенной кислородом), у лягушек в мозг идет смешанная кровь. Самыми развитыми тут оказались крокодилы, у них возникает практически полноценная перегородка между предсердиями, как у человека….
1)кисть ( смородина,капуста,черемуха,солодка)
2)колос ( подорожник, любка,ятрышник)
3) щиток (яблоня, груша, боярышник)
4)зонтик (лук, женьшень, примула)
5) головка ( клевер)
6) корзинка (одуванчик, василек, ромашка)
СЛОЖНЫЕ:
1) метелка ( виноград, сирень,просо, овес, рис)
2) сложный колос ( пшеница, рожь, ячмень)
3) сережка ( береза, ива,тополь, осина)
4) сложный зонтик (морковь, трынь трава, тмин)
5) сложный щиток (калина, рябина)
Микронуклеус зародышевой линии — обзор
РНК-опосредованное развитие инфузорий
P. tetraurelia — одноклеточный эукариот, который содержит два функционально различных ядра, а именно микроядра зародышевой линии и соматические макронуклеары. Диплоидные микроядра зародышевой линии, которые претерпевают мейоз, транскрипционно неактивны во время вегетативного роста, тогда как высокополиплоидные соматические макронуклеусы (800 n) отвечают за экспрессию генов на протяжении всего жизненного цикла, но теряются после оплодотворения.Очень своеобразен процесс спаривания P. tetraurelia . Действительно, сразу после мейоза три из четырех гаплоидных ядер дегенерируют. У каждого партнера по конъюгации оставшееся ядро затем дублируется посредством митотического деления. Эта дупликация позволяет осуществлять взаимный обмен гаплоидными ядрами между спаривающимися парамециями. После кариогамии образующиеся зиготы представляют собой диплоидные микронуклеусы и расплывающиеся макронуклеары. Следовательно, необходимо создать новые зиготические макронуклеусы.Это достигается после двух делений микроядер путем массивной эндорепликации и обширных перестроек двух из четырех ядер, лежащих на задней стороне клетки. Хромосомы сильно фрагментированы на более короткие молекулы, закрытые добавлением de novo теломер [56], но наиболее яркой особенностью этих перестроек является точное удаление десятков тысяч однокопийных коротких некодирующих внутренних последовательностей (IES) [57]. , 58], что делает макронуклеары вытесненной версией микроядер.Как можно выполнить такое изумительное усилие редактирования?
Первые намеки на эпигенетическое соединение, участвующее в этом процессе перестройки всего генома, были получены в экспериментах по трансформации P. tetraurelia [59–61]. Когда последовательность IES интегрируется в вегетативные макронуклеары, вырезание соответствующих IES в новых макронуклеарах полового потомства специфически ингибируется [62]. Удержание IES, которое делает его присутствующим во всех макромолекулярных копиях, затем наследуется по материнской линии (цитоплазматически) в следующих половых поколениях.Это явно напоминало механизм, основанный на эпигенетической гомологии. Позже, у T. thermophila было показано, что онтогенетические перестройки зависят от гена TWI1 , который кодирует белок, гомологичный Piwi-подобным белкам [63], от гена DCL1 , кодирующего Dicer-подобный белок. [64], а на Ema1p — предполагаемая РНК-геликаза [65]. В P. tetraurelia идентификация Nowa1 и Nowa2, двух РНК-связывающих белков, необходимых для удаления IES из развивающихся макронуклеаров [66], также указала на то, что перекрестные помехи между ядрами, работающие во время реаранжировки генома, связаны с РНКи. путь.Таким образом, в отличие от канонического пути РНКи, эта вторая зависимая от гомологии система молчания ограничивается половым развитием, именно тогда, когда происходят перестройки ДНК зародышевой линии. Он производит специфический класс 25-нуклеотидной миРНК, называемый «сканирующими РНК» (scnRNAs) [67,68]. Было показано, что микроинъекция 25-нуклеотидного синтетического РНК-дуплекса, имитирующего структуру scnRNA, действительно способствует удалению гомологичных IES в развивающихся зиготических макронуклеусах [69]. Более того, в Paramecium небелковые транскрипты (ncRNAs), продуцируемые из соматического материнского макронуклеуса (лишенного IESs), необходимы для удаления IES в развивающемся зиготическом макронуклеусе [69].На основе этого набора данных была предложена целая модель «сканирования генома» [70,71] (рис. 13.1C). Согласно этой модели, программа эпигенетического развития, приводящая к массивному, но точному удалению ДНК, будет основана на геномном вычитании между индуцирующими делецию scnRNA и защитными некодирующими транскриптами.
Изучение полового развития у P. tetraurelia и T. thermophila многое внесло в эпигенетические области, особенно благодаря новым интригующим представлениям о том, насколько разнообразными могут быть механизмы, зависимые от гомологии.Рекрутирование аппарата PTGS у инфузорий, чтобы помочь сформировать новый соматический геном, свободный от эгоистичных элементов ДНК, напоминает роли, приписываемые PTGS в защите геномов нитчатых грибов и в определении территорий геномных гетерохроматина S. pombe . У других протистов, таких как T. brucei , было показано, что транспозоны реактивируются у мутантов с дефицитом PTGS, подтверждая роль PTGS в защите генома от экспрессии и, возможно, экспансии мусорной ДНК [72].
Ядро и цитоплазма | Анатомия и физиология
Цели обучения
- Опишите структуру и функцию ядра
- Объясните организацию ДНК в ядре
- Опишите структуру и функцию основных клеточных органелл
Теперь, когда вы узнали, что плазматическая мембрана окружает все клетки, вы можете погрузиться в прототип клетки человека, чтобы узнать о ее внутренних компонентах и их функциях.Клетки животных содержат три основных участка: плазматическую мембрану, ядро и цитоплазму. Ядро — это центральная органелла клетки, которая содержит ДНК клетки (рис. 3.6). Цитоплазма состоит из двух частей: цитозоля и органелл. Цитозоль , желеобразное вещество внутри клетки, обеспечивает жидкую среду, необходимую для биохимических реакций. Органелла («маленький орган») — это один из нескольких различных типов мембранных тел в клетке, каждое из которых выполняет уникальную функцию.Подобно тому, как различные органы тела работают вместе в гармонии, выполняя все функции человека, множество различных клеточных органелл работают вместе, чтобы поддерживать здоровье клетки и выполнять все ее важные функции.
Рисунок 3.6. Прототипная клетка человека Хотя это изображение не указывает на какую-либо конкретную человеческую клетку, это прототип клетки, содержащей первичные органеллы и внутренние структуры. ЯдроЯдро — самая большая и самая заметная из органелл клетки (Рисунок 3.7). Ядро обычно считается центром управления клеткой, потому что оно хранит все генетические инструкции для производства белков. Интересно, что некоторые клетки тела, например мышечные, содержат более одного ядра, которое называется многоядерным. Другие клетки, такие как эритроциты (эритроциты) млекопитающих, вообще не содержат ядер. По мере созревания эритроциты выбрасывают свои ядра, освобождая место для большого количества молекул гемоглобина, которые переносят кислород по всему телу. Без ядер продолжительность жизни эритроцитов коротка, поэтому организм должен постоянно производить новые.
Рисунок 3.7. Ядро Ядро — это центр управления клеткой. Ядро живых клеток содержит генетический материал, который определяет всю структуру и функции этой клетки.Внутри ядра находится план, который диктует все, что клетка будет делать, и все продукты, которые она будет производить. Эта информация хранится в ДНК. Ядро отправляет «команды» клетке через молекулярные мессенджеры, которые транслируют информацию из ДНК.Каждая клетка вашего тела (за исключением половых клеток) содержит полный набор вашей ДНК. Когда клетка делится, ДНК должна быть продублирована, чтобы каждая новая клетка получала полный набор ДНК. В следующем разделе мы исследуем структуру ядра и его содержимое, а также процесс репликации ДНК.
Организация ядра и его ДНК
Как и большинство других клеточных органелл, ядро окружено мембраной, называемой ядерной оболочкой .Это мембранное покрытие состоит из двух смежных липидных бислоев с тонким жидким пространством между ними. Эти два бислоя охватывают ядерные поры. Ядерная пора — это крошечный проход для прохождения белков, РНК и растворенных веществ между ядром и цитоплазмой. Внутри ядерной оболочки находится гелеобразная нуклеоплазма с растворенными веществами, которые включают строительные блоки нуклеиновых кислот. Также может быть темная масса, часто видимая под простым световым микроскопом, называемая ядрышком (множественное число = ядрышки).Ядрышко — это область ядра, которая отвечает за производство РНК, необходимой для построения рибосом. После синтеза вновь образованные субъединицы рибосомы покидают ядро клетки через ядерные поры. Генетические инструкции, которые используются для построения и поддержания организма, упорядочены в цепях ДНК. Внутри ядра расположены нити , хроматин , состоящий из ДНК и связанных белков (рис. 3.8). Хроматин — это волокнистая форма ДНК, которая позволяет эффективно упаковывать ДНК в ядре, сохраняя при этом структуру, позволяющую синтезировать белки на ранних стадиях.Вдоль нитей хроматина ДНК обернута вокруг набора из белков гистона . Когда клетка находится в процессе деления, хроматин конденсируется в хромосомы, так что ДНК можно безопасно транспортировать к «дочерним клеткам». Хромосома состоит из ДНК и белков; это конденсированная форма хроматина. Подсчитано, что у человека почти 22 000 генов распределены по 46 хромосомам.
Рисунок 3.8. Макроструктура ДНК Нити ДНК обернуты вокруг поддерживающих гистонов.Эти белки все больше связываются и конденсируются в хроматин, который плотно упаковывается в хромосомы, когда клетка готова к делению.Органеллы эндомембранной системы
Набор из трех основных органелл вместе формирует внутри клетки систему, называемую эндомембранной системой. Эти органеллы работают вместе для выполнения различных клеточных задач, включая задачу производства, упаковки и экспорта определенных клеточных продуктов. Органеллы эндомембранной системы включают эндоплазматический ретикулум, аппарат Гольджи и везикулы.
Эндоплазматическая сеть
Эндоплазматический ретикулум (ER) представляет собой систему каналов, которые являются продолжением ядерной мембраны (или «оболочки»), покрывающей ядро, и состоящей из того же материала липидного бислоя. ER можно рассматривать как серию извилистых магистралей, похожих на водные каналы Венеции. ER обеспечивает проходы по большей части клетки, которые функционируют при транспортировке, синтезе и хранении материалов.Обмоточная структура ER приводит к большой площади мембранной поверхности, которая поддерживает его многие функции (рис. 3.9).
Рисунок 3.9. Эндоплазматическая сеть (ER) (a) ER представляет собой извилистую сеть тонких мембранных мешочков, находящихся в тесной связи с ядром клетки. Гладкая и шероховатая эндоплазматическая сеть очень различаются по внешнему виду и функциям (источник: ткань мыши). (b) Rough ER усеяна многочисленными рибосомами, которые являются участками синтеза белка (источник: ткань мыши).EM × 110000. (c) Smooth ER синтезирует фосфолипиды, стероидные гормоны, регулирует концентрацию клеточного Ca ++ , метаболизирует некоторые углеводы и расщепляет определенные токсины (источник: ткань мыши). EM × 110 510. (Микрофотографии предоставлены Медицинской школой Риджентс Мичиганского университета © 2012)Эндоплазматический ретикулум может существовать в двух формах: грубый ER и гладкий ER. Эти два типа ER выполняют очень разные функции и могут быть найдены в очень разных количествах в зависимости от типа клетки.Грубый ER (RER) называется так, потому что его мембрана усеяна встроенными гранулами — органеллами, называемыми рибосомами, что придает RER неровный вид.
A рибосома — органелла, которая служит местом синтеза белка. Его можно обнаружить свободно плавающим в цитоплазме или прикрепленным к ER. Он состоит из двух субъединиц рибосомной РНК, которые оборачиваются вокруг мРНК, чтобы запустить процесс трансляции, стадию синтеза белка. Синтез белка состоит из двух стадий: транскрипции и трансляции.Транскрипция происходит внутри ядра и представляет собой фазу синтеза белков, в которой мРНК копируется из ДНК. МРНК покидает ядро через ядерные поры и переходит к рибосоме. Затем рибосома «считывает» или интерпретирует инструкции внутри мРНК и использует РНК переноса (тРНК) для связывания аминокислот в соответствующем порядке с образованием белка ( рис. 3.10 ). Как правило, белок синтезируется внутри рибосомы и высвобождается внутри канала грубого ER, где к нему могут быть добавлены сахара (посредством процесса, называемого гликозилированием), прежде чем он будет транспортирован внутри везикулы на следующий этап процесса упаковки и транспортировки. : аппарат Гольджи.
Рисунок 3.10. От ДНК к белку: транскрипция через трансляцию Транскрипция в ядре клетки производит молекулу мРНК, которая модифицируется и затем отправляется в цитоплазму для трансляции. Транскрипт расшифровывается в белок с помощью молекул рибосомы и тРНК.Smooth ER (SER) не содержит этих рибосом. Одна из основных функций гладкого ЭПР — синтез липидов. Гладкий ER синтезирует фосфолипиды, основной компонент биологических мембран, а также стероидные гормоны.По этой причине клетки, вырабатывающие большое количество таких гормонов, например, из женских яичников и мужских семенников, содержат большое количество гладкого ЭПР. В дополнение к синтезу липидов гладкий ЭПР также изолирует (то есть накапливает) и регулирует концентрацию ионов кальция в мышечной и нервной ткани. Гладкий ER дополнительно метаболизирует некоторые углеводы и выполняет роль детоксикации в печени, расщепляя определенные токсины. В отличие от гладкого ER, основная работа грубого ER — это синтез и модификация белков, предназначенных для клеточной мембраны или для экспорта из клетки.Для этого синтеза белка многие рибосомы прикрепляются к ER (придавая ему вид грубого ER).
Аппарат Гольджи
Аппарат Гольджи отвечает за сортировку, модификацию и отгрузку продуктов, поступающих из неотложной неотложной помощи, во многом как почтовое отделение. Аппарат Гольджи выглядит как сложенные стопкой плоские диски, почти как стопки блинов странной формы. Как и ER, эти диски являются перепончатыми. У аппарата Гольджи есть две разные стороны, каждая из которых играет свою роль.Одна сторона аппарата принимает продукты в виде пузырьков. Эти продукты сортируются в аппарате, а затем выпускаются с противоположной стороны после переупаковки в новые пузырьки. Если продукт должен быть экспортирован из клетки, везикула мигрирует на поверхность клетки и сливается с клеточной мембраной, и груз секретируется (рис. 3.11).
Рисунок 3.11. Аппарат Гольджи (a) Аппарат Гольджи управляет продуктами грубого ER, а также производит новые органеллы, называемые лизосомами.Белки и другие продукты ER отправляются в аппарат Гольджи, который организует, модифицирует, упаковывает и маркирует их. Некоторые из этих продуктов транспортируются в другие области клетки, а некоторые выводятся из клетки посредством экзоцитоза. Ферментативные белки упаковываются как новые лизосомы (или упаковываются и отправляются для слияния с существующими лизосомами). (б) Электронная микрофотография аппарата Гольджи.Лизосомы
Некоторые из белковых продуктов, упакованных Гольджи, содержат пищеварительные ферменты, которые должны оставаться внутри клетки для использования при расщеплении определенных материалов.Везикулы, содержащие ферменты, высвобождаемые Гольджи, могут образовывать новые лизосомы или сливаться с существующими лизосомами. Лизосома — это органелла, содержащая ферменты, которые расщепляют и переваривают ненужные клеточные компоненты, такие как поврежденная органелла. (Лизосома похожа на разрушительную бригаду, которая сносит старые и ненадежные здания по соседству.) Аутофагия («самопоедание») — это процесс переваривания клеткой собственных структур. Лизосомы также важны для расщепления инородного материала.Например, когда определенные клетки иммунной защиты (лейкоциты) фагоцитируют бактерии, бактериальная клетка транспортируется в лизосому и переваривается находящимися внутри ферментами. Как можно догадаться, такие клетки фагоцитарной защиты содержат большое количество лизосом. При определенных обстоятельствах лизосомы выполняют более грандиозную и ужасную функцию. В случае поврежденных или нездоровых клеток лизосомы могут открыться и высвободить свои пищеварительные ферменты в цитоплазму клетки, убивая клетку.Этот механизм «самоуничтожения» называется автолиз и контролирует процесс гибели клеток (механизм, называемый «апоптоз»).
Посмотрите это видео, чтобы узнать об эндомембранной системе, которая включает грубую и гладкую ER и тело Гольджи, а также лизосомы и везикулы. Какова основная роль эндомембранной системы?
Органеллы для производства энергии и детоксикации
Помимо функций, выполняемых эндомембранной системой, клетка выполняет множество других важных функций.Подобно тому, как вы должны потреблять питательные вещества, чтобы обеспечить себя энергией, каждая из ваших клеток должна принимать питательные вещества, некоторые из которых превращаются в химическую энергию, которая может использоваться для поддержания биохимических реакций. Еще одна важная функция клетки — детоксикация. Люди поглощают всевозможные токсины из окружающей среды, а также производят вредные химические вещества в качестве побочных продуктов клеточных процессов. Клетки печени, называемые гепатоцитами, выводят многие из этих токсинов.
Митохондрии
A митохондрия (множественное число = митохондрии) представляет собой мембранную бобовидную органеллу, которая является «преобразователем энергии» клетки.Митохондрии состоят из внешней двухслойной липидной мембраны, а также дополнительной внутренней двухслойной липидной мембраны (рис. 3.12). Внутренняя мембрана сильно сложена в извилистые структуры с большой площадью поверхности, называемые кристами. Именно вдоль этой внутренней мембраны ряд белков, ферментов и других молекул выполняет биохимические реакции клеточного дыхания. Эти реакции преобразуют энергию, хранящуюся в молекулах питательных веществ (таких как глюкоза), в аденозинтрифосфат (АТФ), который обеспечивает клетку полезной клеточной энергией.Клетки постоянно используют АТФ, поэтому митохондрии постоянно работают. Молекулы кислорода необходимы во время клеточного дыхания, поэтому вы должны постоянно вдыхать их. Одной из систем организма, которая использует огромное количество АТФ, является мышечная система, потому что АТФ требуется для поддержания мышечного сокращения. В результате мышечные клетки заполнены митохондриями. Нервным клеткам также требуется большое количество АТФ для работы своих натриево-калиевых насосов. Следовательно, отдельный нейрон будет загружен более чем тысячей митохондрий.С другой стороны, костная клетка, которая не так метаболически активна, может иметь всего пару сотен митохондрий.
Рисунок 3.12. Митохондрия Митохондрии — это фабрики преобразования энергии клетки. (а) Митохондрия состоит из двух отдельных двухслойных липидных мембран. Вдоль внутренней мембраны расположены различные молекулы, которые вместе производят АТФ, главную энергетическую валюту клетки. (б) Электронная микрофотография митохондрий. EM × 236000.(Микрофотография предоставлена Медицинской школой Риджентс Мичиганского университета © 2012)Пероксисомы
Как и лизосомы, пероксисома представляет собой мембраносвязанную клеточную органеллу, которая в основном содержит ферменты (рис. 3.13). Пероксисомы выполняют несколько различных функций, включая метаболизм липидов и химическую детоксикацию. В отличие от пищеварительных ферментов, содержащихся в лизосомах, ферменты в пероксисомах служат для передачи атомов водорода от различных молекул к кислороду, производя перекись водорода (H 2 O 2 ).Таким образом, пероксисомы нейтрализуют яды, такие как алкоголь. Чтобы понять важность пероксисом, необходимо понять концепцию активных форм кислорода.
Рисунок 3.13. Пероксисома Пероксисомы — это мембранные органеллы, которые содержат множество ферментов для детоксикации вредных веществ и метаболизма липидов.Активные формы кислорода (АФК) , такие как пероксиды и свободные радикалы, являются высокореактивными продуктами многих нормальных клеточных процессов, включая митохондриальные реакции, которые производят АТФ и метаболизм кислорода.Примеры ROS включают гидроксильный радикал ОН, H 2 O 2 и супероксид (O 2 —). Некоторые АФК важны для определенных клеточных функций, таких как клеточные сигнальные процессы и иммунные ответы против чужеродных веществ. Свободные радикалы реактивны, потому что они содержат свободные неспаренные электроны; они могут легко окислять другие молекулы по всей клетке, вызывая клеточное повреждение и даже смерть клетки. Считается, что свободные радикалы играют роль во многих деструктивных процессах в организме, от рака до ишемической болезни сердца.С другой стороны, пероксисомы контролируют реакции, нейтрализующие свободные радикалы. Пероксисомы производят большие количества токсичного H 2 O 2 в процессе, но пероксисомы содержат ферменты, которые превращают H 2 O 2 в воду и кислород. Эти побочные продукты безопасно попадают в цитоплазму. Подобно миниатюрным установкам для очистки сточных вод, пероксисомы нейтрализуют вредные токсины, чтобы они не наносили вред клеткам. Печень — это орган, который в первую очередь отвечает за детоксикацию крови перед ее распространением по телу, а клетки печени содержат исключительно большое количество пероксисом.Защитные механизмы, такие как детоксикация внутри пероксисомы и некоторых клеточных антиоксидантов, служат для нейтрализации многих из этих молекул. Некоторые витамины и другие вещества, содержащиеся в основном во фруктах и овощах, обладают антиоксидантными свойствами. Антиоксиданты действуют, окисляясь сами, останавливая каскады деструктивных реакций, инициируемых свободными радикалами. Однако иногда АФК накапливаются за пределами возможностей такой защиты. Окислительный стресс — это термин, используемый для описания повреждения клеточных компонентов, вызванного ROS.Из-за своих характерных неспаренных электронов АФК могут запускать цепные реакции, в которых они удаляют электроны из других молекул, которые затем становятся окисленными и реакционноспособными, и делают то же самое с другими молекулами, вызывая цепную реакцию. АФК могут вызвать необратимое повреждение клеточных липидов, белков, углеводов и нуклеиновых кислот. Поврежденная ДНК может привести к генетическим мутациям и даже к раку. Мутация — это изменение нуклеотидной последовательности в гене в ДНК клетки, потенциально изменяющее белок, кодируемый этим геном.Другие заболевания, которые, как считается, вызываются или обостряются ROS, включают болезнь Альцгеймера, сердечно-сосудистые заболевания, диабет, болезнь Паркинсона, артрит, болезнь Хантингтона и шизофрению, среди многих других. Примечательно, что эти заболевания во многом связаны с возрастом. Многие ученые считают, что окислительный стресс является одним из основных факторов старения.
Цитоскелет
Так же, как костный скелет структурно поддерживает человеческое тело, цитоскелет помогает клеткам сохранять свою структурную целостность.Цитоскелет представляет собой группу волокнистых белков, которые обеспечивают структурную поддержку клеток, но это только одна из функций цитоскелета. Компоненты цитоскелета также имеют решающее значение для подвижности клеток, воспроизводства клеток и транспортировки веществ внутри клетки. Цитоскелет образует сложную нитевидную сеть по всей клетке, состоящую из трех различных видов волокон на основе белков: микрофиламентов, промежуточных волокон и микротрубочек (рис.14). Самой толстой из трех является микротрубочка , структурная нить, состоящая из субъединиц белка, называемого тубулином. Микротрубочки поддерживают форму и структуру клеток, помогают сопротивляться сжатию клетки и играют роль в расположении органелл внутри клетки. Микротрубочки также составляют два типа клеточных придатков, важных для движения: реснички и жгутики. Реснички обнаружены на многих клетках тела, включая эпителиальные клетки, выстилающие дыхательные пути дыхательной системы.Реснички движутся ритмично; они постоянно бьются, перемещая отходы, такие как пыль, слизь и бактерии, вверх по дыхательным путям, от легких к рту. Удары ресничек клеток в женских фаллопиевых трубах перемещают яйцеклетки из яичника в матку. Флагеллум (множественное число = жгутики) — это придаток больше реснички и специализированный для передвижения клеток. Единственная жгутиковая клетка у человека — это сперматозоид, который должен продвигаться к женским яйцеклеткам.
Рисунок 3.14. Три компонента цитоскелета Цитоскелет состоит из (а) микротрубочек, (б) микрофиламентов и (в) промежуточных филаментов. Цитоскелет играет важную роль в поддержании формы и структуры клеток, стимулировании клеточного движения и содействии делению клеток.Очень важная функция микротрубочек — устанавливать пути (наподобие железнодорожных путей), по которым генетический материал может тянуться (процесс, требующий АТФ) во время деления клетки, так что каждая новая дочерняя клетка получает соответствующий набор хромосом.Две короткие идентичные структуры микротрубочек, называемые центриолями, находятся рядом с ядром клеток. Центриоль может служить точкой клеточного происхождения для микротрубочек, выходящих наружу в виде ресничек или жгутиков, или может способствовать разделению ДНК во время деления клетки путем формирования митотического веретена (волокна веретена).
Структура парамециевой клетки
Поделиться — это забота!
В этом разделе мы рассмотрим клеточную структуру парамеция .Если не упомянуто, мы ссылаемся на Paramecium caudatum как на типичный пример клетки парамеция .
У нас также есть 4 серии сообщений в блоге о парамеции:
Часть I. Биологическая классификация парамеций — название, история и эволюция
Часть II. Структура ячейки Paramecium
Часть III. Размножение парамеций, физиология и поведение
Часть IV.Естественное обитание и выращивание парамеция — найдите парамеций для своего микроскопического проекта
Анатомия парамеция
[На этом рисунке] Помеченная диаграмма, показывающая анатомию клетки Paramecium.
Каждая структура / органелла и ее функция будут объяснены в этой статье.
Парамеций носит мягкую броню, называемую пленкой
.Тело клетки парамеции окружено жесткой, но эластичной мембраной, называемой пленкой.Пелликула состоит из тонкого студенистого вещества, вырабатываемого клеткой. Слой пленки придает парамеции определенную форму и хорошо защищает ее клеточное содержимое. Пленка также эластична по своей природе, что позволяет парамеции немного изменять свою форму.
Кожа парамециума покрыта множеством крошечных волосков, называемых ресничками
Пелликулу покрывают множество крошечных волосков, называемых ресничками, (единственная ресничка). Реснички — это выступы, распределенные по всей клетке.Одна клетка P. caudatum может иметь 5000–6000 ресничек.
Реснички — очень крошечные структуры — примерно 0,25 мкм в диаметре и до 20 мкм в длину. Мы можем видеть только общее движение ресничек на клетках, но трудно увидеть их четко под обычным световым микроскопом.
[В этом видео] Перемещение ресничек Paramecium под микроскопом.
На клетках парамеций присутствуют реснички двух типов: реснички полости рта и реснички тела. Реснички полости рта находятся на поверхности бороздки полости рта. Они помогают собирать пищевые материалы. Реснички тела находятся на поверхности тела и облегчают его передвижение. Они действуют как микроскопические весла, перемещая организм в одном направлении.
Реснички тела расположены продольными рядами (по оси от головы к хвосту) с довольно равномерной длиной по всей клетке. Есть также несколько более длинных ресничек, присутствующих на заднем конце клетки (совершенно очевидно у P. caudatum ).Они образуют хвостового пучка ресничек (отсюда и название caudatum ).
[На этом рисунке] Различные типы ресничек на клетке парамеция — оральные реснички, реснички тела и хвостовой пучок.
Реснички
под микроскопомУченые потратили много времени и усилий на изучение ресничек парамеции. Почему?
Это связано с тем, что реснички не являются исключительными для микроорганизмов, таких как парамеции или инфузории. Фактически, у нас также есть реснички на наших клетках.Например, подвижные реснички находятся на респираторном эпителии, выстилающем дыхательные пути, где они очищают наши легкие, вымывая слизь и грязь.
Продвинутая микроскопия является мощным инструментом в исследованиях клеточной биологии такого рода. Например, сканирующий электронный микроскоп (СЭМ) позволяет нам увидеть морфологию, ориентацию и плотность ресничек парамеция. С помощью просвечивающего электронного микроскопа (ПЭМ) мы можем увидеть ультраструктуру ресничек в поперечном срезе. С помощью иммунофлуоресцентного окрашивания на основе антител ученые могут даже увидеть, какие виды белков вносят вклад в структуру, движение и рост ресничек.
[На этом рисунке] Ученые использовали современные микроскопы, чтобы ответить на свои вопросы о ресничках парамеция. Слева: СЭМ показывает морфологию ресничек (Фото: Джудит Л. Ван Хаутен). В центре: ПЭМ дает нам детальное изображение поперечного сечения ресничек (Фото: Ричард Аллен). Справа: флуоресцентный микроскоп показывает нам, как реснички закрепляются на поверхности клетки.
Строение пленки и ресничек
С помощью современной микроскопии ученые теперь в деталях знают, как реснички растут и движутся.Как вы можете видеть на иллюстрации ниже, слой пленки не гладкий. Вместо этого есть много бугорков (называемых альвеолами) с углублениями на пленке. Ресничка выходит через центральное отверстие каждого углубления с якорем на базальном теле.
[На этом рисунке] Детальная структура ресничек и пленки.
Ученые также обнаружили, что находится внутри каждого волоска ресничек. Ресничка состоит из пучков микротрубочек. Микротрубочки — это белковые волокна внутри клеток, выполняющие множество функций.Микротрубочки могут служить межклеточной магистралью для транспортировки молекул и органелл. Во время деления клетки волокна микротрубочек, выступающие из двух центросом, разделяют хромосомы в новые ядра.
[На этом рисунке] Организация реснички.
Каждая ресничка содержит девять пар микротрубочек, образующих внешнюю часть кольца, и две центральные микротрубочки. Эта структура известна как аксонема. Микротрубочки удерживаются вместе за счет сшивания белков.Моторные белки, называемые динеином, проходят через каждое спаренное волокно микротрубочек.
Фотография предоставлена LadyofHats на вики.
Моторные белки (динеин) используют АТФ в качестве энергии для ползания по микротрубочкам. Когда белки динеина с одной стороны движутся вверх, а с другой — вниз, ресничка изгибается. Повторение циклов сгибания-расслабления заставляет реснички действовать как весла, качаясь взад и вперед, создавая движение.
Посмотрите, как реснички делают волну
Если реснички одинаково машут взад и вперед, клетки никуда не денутся.Ход вперед и назад должен быть различных фаз, , чтобы создать значимую движущую силу.
Ученые использовали микроскоп с высокоскоростной видеокамерой, чтобы запечатлеть, как бьются реснички, заставляя двигаться все тело парамеция. Они выглядят довольно умно!
[В этом видео] Движение ресничек демонстрирует красивую метахрональную волнообразную координацию, когда между соседними ресничками поддерживается постоянная разность фаз.
Анализируя кадр за кадром на высокоскоростной видеозаписи, ученые обнаружили, что парамеций плавает так же, как мы плывем при переднем ползании.
Эффективный ход (вперед) : Во время эффективного удара ресничка поднимается вверх (чтобы привлечь больше воды) и ударяется о воду, таким образом перемещая тело вперед и отправляя воду назад.
Восстановительный (обратный) ход : Во время восстановительного хода ресничка возвращается в исходное положение своим обратным движением.Ресничка имеет тенденцию изгибаться и оставаться ближе к поверхности клетки, чтобы минимизировать сопротивление.
[На этом рисунке] Образец штрихов ресничек парамеция.
Движение ресничек можно разделить на Эффективный (вперед) и Восстановительный (назад) ход. Два вида гребков поочередно повторяются, чтобы продвигать тело парамеции, когда мы плывем в стиле ползания вперед.
В отличие от нас, у которых только два плеча, парамеция клетка имеет тысячи ресничек. Чтобы плавать эффективно, все реснички не двигаются одновременно.Реснички группируются в два типа согласованных ритмов.
Ритм синхронный — Реснички поперечного ряда движутся одновременно.
Метахронный ритм — Реснички продольного ряда бьются друг за другом. Это создает метахрональные волны, проходящие от переднего конца к заднему.
[На этом рисунке] Реснички движутся в метахронном ритме, создавая движущую силу.
Еще одно интересное поведение — это способ побега парамеций.Если парамеций наталкивается на препятствие, биение ресничек прекращается и меняется на противоположное. Это заставляет парамеций плыть назад, чтобы держаться подальше от препятствий или хищников.
Как быстро может двигаться парамеций?
Вы можете задаться вопросом, с какой скоростью может двигаться парамеций? Они двигаются быстрее олимпийских золотых медалистов!
Большинство инфузорий, таких как парамеции, прекрасно умеют плавать. Почему реснички? Когда ваш рост меньше миллиметра, вода похожа на липкий сироп.Плавать, как рыба, было бы не очень эффективно! Если вы хотите быстро плавать и уметь маневрировать, реснички — лучший выбор.
Реснички парамеции движутся, как множество крошечных весел, продвигая организм по воде со скоростью « в четыре раза больше длины его тела в секунду ». Для P. caudatum , который имеет длину 300 микрометров (мкм), он может плавать со скоростью 1200 мкм в секунду (что равно 0,0027 мили в час). Если Майкл Фелпс (6 футов 4 дюйма или 1,93 м) будет плавать как парамеций, он будет плавать со скоростью 7.72 метра в секунду и преодоление дистанции 100 метров за 12,95 секунды. Это в четыре раза быстрее мирового рекорда по плаванию!
[На этом рисунке] Три способа передвижения простейших.
Реснички — скоординированно бить плавать. Псевдоножки — ползать по поверхности, изменяя форму клетки. Жгутик — плывет, вращаясь как пропеллер.
Фотография предоставлена Lumen.
Как питается парамеций?
Паз для полости рта
Парамеции питаются другими микроорганизмами, такими как бактерии, дрожжи или водоросли.Они питаются через систему, которая работает так же, как наш рот-пищевод-желудок. На вентро-боковой стороне тела парамеции имеется большое косое неглубокое углубление, называемое оральной бороздкой . Эта ротовая бороздка придает животному асимметричный вид.
Ротовая канавка служит для входа пищевых материалов в камеру. Поверхность ротовой бороздки покрывают реснички ротовой полости. Эти оральные реснички бьются, создавая входящий поток воды и принося пищу в оральную бороздку.
[На этом рисунке] Детальный вид системы питания парамеций.
Вы можете следить за красными стрелками, чтобы отслеживать путь пищи, проходящей через систему кормления парамеций. Сначала частицы пищи собираются в бороздку полости рта за счет движения ресничек полости рта. Пищевые материалы перемещаются из цитостома в цитофаринкс, а затем в пищевые вакуоли путем фагоцитоза. Пищеварительные ферменты внутри пищевых вакуолей расщепляют пищу на небольшие молекулы питательных веществ. После того, как питательные вещества абсорбируются клеткой в цитоплазму, неперевариваемый мусор выводится из анальной поры.
Цитостом, цитофаринкс и пищевая вакуоль
Конец ротовой бороздки соединяется с воронкообразной структурой, называемой цитостом или устьем клеток . Реснички полости рта также покрывают просвет цитостома, доставляя частицу пищи вниз в воронку цитостома, которая простирается в цитофаринкс.
Цитофаринкс представляет собой трубчатую структуру (как и наш пищевод), которая ведет к пищевым вакуолям. Пищевые вакуоли образуются почкованием из заднего конца цитофаринкса.Пищевые вакуоли функционируют так же, как наш желудок, и содержат переваривающие ферменты, расщепляющие пищевые материалы на молекулы питательных веществ.
Процесс питания путем поглощения через пищевые вакуоли, называемый фагоцитоз , обычен для одноклеточных микроорганизмов, таких как амеба. Однако только у представителей семейств инфузорий и эвглен развивается особая система цитостом-цитофаринкс.
Парамеция какает?
Анальные поры
Ответ — да.У парамеций есть свой способ выведения. После того, как питательные вещества из переваренной пищи всасываются в цитоплазму, в пищевых вакуолях все еще остается неперевариваемый мусор. Отходы будут выбрасываться из структуры, называемой анальной порой или cytoproct .
Различные одноклеточные эукариоты имеют анальную пору. Анальная пора парамеция — это область пленки, не покрытая гребнями и ресничками. Тонкая пленка позволяет вакуолям сливаться с поверхностью клетки и опорожняться.
Специализированная «Кожа» тела клетки парамеция
Эктоплазма и эндоплазма
В отличие от многоклеточных организмов, у которых есть слой выделенных клеток кожи в качестве защитного барьера, у одноклеточных парамеций развивается «клеточная кожа» для защиты.
Как мы упоминали выше, самый внешний слой — это мягкая оболочка из пелликулы и ресничек. С пленкой связан узкий периферический слой специализированной твердой цитоплазмы, называемый эктоплазмой .Под эктоплазмой находится более жидкий тип цитоплазмы: эндоплазма . Эта область содержит большинство клеточных компонентов и органелл.
[На этом рисунке] Подробное изображение структуры кожи парамеции.
На этом изображении с высоким разрешением клетки парамеция вы видите два слоя цитоплазмы: эктоплазму и эндоплазму. Трихоцисты — это защитные органеллы, встроенные в слой эктоплазмы. (Изображение изменено из работы Майкла Плевки)
По сравнению с остальной цитоплазмой (эндоплазмой), эктоплазма образует тонкий, плотный и прозрачный внешний слой, содержащий трихоцисты и фибриллярные структуры.Корни ресничек также закрепляются в слое эктоплазмы. Пелликула и эктоплазма вместе служат защитной кожей для парамеций.
Трихоцист
Trichocyst (trick-o-sists) — это небольшая веретенообразная органелла, расположенная в эктоплазме с крохотной порой, открытой на поверхности пленки. Трихоцисты располагаются перпендикулярно эктоплазме. Трихоцисты заполнены плотной преломляющей жидкостью, содержащей набухшие вещества. Когда клетки получают механические, химические или электрические раздражители, трихоцисты выделяют свое содержимое и становятся длинными тонкими колючими шипами.После их разряда из кинетосом образуются новые.
Точная функция трихоцист не совсем ясна, хотя популярная теория гласит, что они важны для защиты от хищников. Трихоцисты также могут способствовать адгезии клеток и поддерживать тело клетки парамеции.
[На этом рисунке] Трихоцисты Paramecium.
Трихоцисты — это веретенообразные органеллы, которые могут выделять жалящие нити в качестве защиты от хищников. Слева: ПЭМ-изображение, показывающее трихоцисту, встроенную в эктоплазму.При получении внешних стимулов ядро трихоцисты проглатывает и выталкивает иглу из оболочки. (Изображение: Bannister, J. Cell Sci. 11: 899-929, 1972). Справа: сильно увеличенное фазово-контрастное изображение, показывающее, что парамеций запускает свои шипастые трихоцисты для защиты. (Изображение: Walter Dawn, Encyclopædia Britannica)
Что находится внутри тела клетки парамеции?
Цитоплазма и органеллы
Подобно нормальной эукариотической клетке, внутри слоя пелликулярной оболочки парамеция находится желеобразное вещество, называемое цитоплазмой .Цитоплазма включает цитозоль и все органеллы . Цитозоль внутри клетки похож на сгущенный суп. Это сложная смесь всех видов веществ, растворенных в воде. Вы можете найти небольшие молекулы, такие как ионы (натрий, калий или кальцин), аминокислоты, нуклеотиды (основные единицы ДНК), липиды, сахара и большие макромолекулы, такие как белки и РНК.
Парамеций имеет все обычные органеллы эукариотических клеток (ссылка на клеточную биологию), включая митохондрии (электростанции клетки), эндоплазматический ретикулум и рибосомы (где синтезируется белок), аппарат Гольджи (почтовое отделение внутри клеток), лизосомы ( хранение пищеварительных ферментов), пероксисомы (химическая лаборатория внутри клеток).
В отличие от растительных клеток, парамеций не имеет хлоропластов.
В отличие от обычных эукариотических клеток, парамеций имеет два ядра , большое и маленькое. Парамеций также состоит из двух типов вакуолей: сократительной вакуоли и пищевой вакуоли , которых нет в клетках человека.
Paramecium работает на двухъядерном процессоре — Macronucleus и Micronucleus
Самая необычная особенность парамеций — их ядра.У них есть два типа ядер, которые различаются по форме, содержанию и функциям.
[На этом рисунке] Макронуклеус (МА) и Микронуклеус (МИ) в клетке P. putrinum . Белые и черные стрелки указывают на симбиотические бактерии внутри цитоплазмы.
Фотография предоставлена: MDPI
Микронуклеус
Ядра двух типов: , микроядра, и макронуклеусы. Микроядро диплоидно; то есть он содержит две копии каждой хромосомы парамеция (ядро человека также диплоидно).Микроядро содержит всю ДНК (называемую геномом), которая присутствует в организме. Эта ДНК передается от одного поколения к другому при воспроизводстве.
Макронуклеус
С другой стороны, макронуклеус содержит подмножество ДНК из микронуклеуса. Эти фрагменты ДНК копируются из микроядра в макронуклеус, поскольку они несут гены, которые часто необходимы клетке парамеции. Гены в макронуклеусе активно транскриптируются в мРНК, а затем транслируются в белки.Макронуклеус полиплоидный или содержит несколько копий каждой хромосомы, иногда до 800 копий.
Функция микронуклеуса и макронуклеуса
Другими словами, функция микроядра заключается в поддержании генетической стабильности и обеспечении передачи нужных генов следующему поколению. Его также называют зародышевой линией или генеративным ядром. Макронуклеус играет роль в нерепродуктивных функциях клетки, включая экспрессию генов, необходимых для повседневного функционирования клетки.Макронуклеус еще называют вегетативным ядром.
Если использовать компьютер в качестве метафоры, микронуклеус — это жесткий диск, на котором хранится полная копия программы клетки. Макронуклеус действует как оперативная память (RAM), в которой хранятся рабочие данные и машинные коды. Компьютер загружает с жесткого диска в оперативную память только те программы, которые используются в данный момент. В клетке парамеции более активные гены (что означает, что клетке нужно больше этих белков, кодируемых этими генами) могут иметь больше копий в макронуклеусе.
Другая причина иметь два разных ядра заключается в том, что это механизм, с помощью которого парамеции и другие инфузории могут предотвращать генетических вторжений (то есть фрагментов ДНК, которые проникают в геном, например, ДНК вируса).
При наличии двух ядер, если фрагмент ДНК находится в микроядре, но не в макронуклеусе, он будет удален во время следующего раунда деления клетки. Другими словами, если что-то чужеродное попадет в микроядерный геном, то при создании следующего макронуклеуса оно будет удалено и не будет включено в экспрессированную версию [транскрибируемую] геном.Этот механизм функционирует как примитивная иммунная система ДНК; то есть изучение генома и попытки не допустить вторжения элементов.
Морфологически макронуклеус имеет форму почки или эллипсоид. Микроядро находится рядом с макронуклеусом. Это небольшая и компактная конструкция сферической формы. У всех видов парамеций есть одно макронуклеус. Однако количество микроядер может варьироваться в зависимости от вида. Например, у P. aurelia два микроядра, а у P. multimicronucleatum — несколько.
Два вида вакуолей, жизненно важных для парамеции
Название «вакуоль» описывает эти органеллы, которые кажутся прозрачными и имеют тенденцию быть заполненными жидкостью. Вакуоли берут на себя определенные функции в клетке парамеции. Парамеций имеет два типа вакуолей: сократительные вакуоли и пищевые вакуоли .
Сократительные вакуоли действуют как водяные помпы для осморегуляции
Одна клетка парамеции имеет две звездообразные сократительные вакуоли , расположенные на каждом конце тела.Они заполнены жидкостью и находятся в фиксированных положениях между эндоплазмой и эктоплазмой. Сократительные вакуоли ответственны за осморегуляцию , или вывод избыточной воды из клетки.
Как бороться с «осмосом» — это универсальная задача для всех живых существ. Это особенно важно для одноклеточных микроорганизмов, таких как парамеций.
Как известно, каждая клетка окружена клеточной мембраной. В этой мембране есть крошечные отверстия, через которые проходят маленькие молекулы (например, вода), но не большие (например, соль).Из-за этой природы клеточная мембрана частично проницаема. Осмос — это перемещение молекул воды из области с высокой концентрацией воды (разбавленный раствор) в область с низкой концентрацией воды (концентрированный раствор) через частично проницаемую мембрану.
[На этом рисунке] Клетки животных (например, эритроциты на этом графике) чувствительны к давлению осмоса. Когда наши клетки находятся в среде « Isotonic » (например, наша кровь), входящие и исходящие молекулы воды равны, и клетки безопасны.Если окружающая среда становится « Hypotonic », что означает меньше растворенных веществ (минералов), чем изотоническая, вода будет перемещаться в клетки для достижения баланса. Клетки будут набухать и даже лопаться (лизироваться), если из клетки не удалить лишнюю воду. С другой стороны, « Hypertonic » связано с большим количеством растворенных веществ в окружающей среде и может вызвать сокращение клеток.
Сократительные вакуоли регулируют количество воды внутри клетки. В пресной воде, которая является гипотонической средой для парамеций, вода поступает в клетку путем осмоса.Сократительные вакуоли вытесняют воду из клетки, сокращаясь и предотвращая поглощение клеткой слишком большого количества воды или даже разрыв.
Как работают сократительные вакуоли?
Каждая сократительная вакуоль соединена с несколькими радикальными каналами (которые образуют ее звездообразную форму). Избыточная вода отводится от всего тела парамеции и через эти каналы подается в сократительные вакуоли. Накопление воды приводит к увеличению размеров вакуоли. Как только резервуар достигает порога сдерживания, сократительная вакуоль сжимается, выпуская избыток воды через поры на пленке.
Две сократительные вакуоли работают независимо. Задняя сократительная вакуоль находится близко к цитофаринксу и, следовательно, сокращается быстрее из-за прохождения большего количества воды. Когда сократительная вакуоль схлопывается, она может периодически исчезать и поэтому называется временными органами.
[На этом рисунке] Изображение Paramecium с высоким разрешением, показывающее две звездообразные сократительные вакуоли и макронуклеус. Это изображение было сделано Рохелио Морено из Панама-Сити, Панама, с использованием дифференциального интерференционного контраста (ДИК) при 40-кратном увеличении.Это изображение получило 4-е место на конкурсе микрофотографии Nikon в 2013 году.
Пылесосы для пищевых продуктов
Когда клетка парамеция собирает пищу через ротовую бороздку и проходит через цитостом в направлении дна цитофаринкса, эти пищевые материалы инкапсулируются в пищевых вакуолях . Пищевые вакуоли затем сливаются с органеллами, называемыми лизосомами , ферменты которых расщепляют молекулы пищи и проводят пищеварение.
Пищевые вакуоли неконтрактильные и имеют примерно сферическую форму.Они служат изолированным отсеком, позволяющим ферментам расщеплять только частицы пищи, но не другие органеллы. Размеры пищевых вакуолей варьируются в зависимости от количества пищи и прогресса пищеварения. Неперевариваемый мусор будет выброшен из отверстия на пленке, которое называется анальной порой или cytoproct .
Похожие сообщения
Биологическая классификация парамеций — название, история и эволюция
Размножение, физиология и поведение Paramecium
Поделиться — это забота!
Органеллы парамеция и их функции — Наука в домашних условиях для детей
Что такое парамеций?
Парамеций — это одноклеточный (одноклеточный) эукариотический организм, обычно встречающийся в стоячей воде.Хотя очень маленький, иногда большой парамеций можно увидеть как крошечные пятнышки, мечущиеся в пробе воды. Парамеций может быть около 0,5 мм в длину.
Откройте для себя
С помощью метрической линейки начертите линию длиной 0,5 мм. Это дает вам представление о том, насколько малы парамеции.
Что такое эукариотика?
Эукариоты — это организмы с одной или несколькими клетками, которые содержат связанное с мембраной ядро или ядра. Эукариотические клетки содержат другие органеллы (органы внутри клетки).Представленный на диаграмме парамеций не включает все органеллы, содержащиеся в эукариотической клетке. Это простое изображение парамеция, которая, как и все эукариоты, имеет другие органеллы, включая митохондрии и эндоплазматический ретикулум. Многоклеточные эукариоты включают животных, растения и грибы. Одноклеточные эукариоты включают парамеций.
Содержимое парамеции связано с клеточной мембраной e , которая покрыта пленкой , — жесткой, но эластичной мембраной.Пленка придает парамеции определенную форму, но она достаточно гибкая, чтобы допускать небольшие изменения формы.
Ниже приведены части и функции клеток, которые поддерживают жизнь одноклеточного парамеция. Цифры в скобках относятся к схеме парамеции.
Дыхание
Кислород входит, а углекислый газ выходит через клеточную мембрану. Энергия вырабатывается в митохонриях (не показаны) посредством реакции дыхания , представленной этим уравнением:
Еда + Кислород дает углекислый газ + вода + Энергия
Эндоплазматический ретикулум (не показан) хранит пищу.
Передвижение
Покрывает пленку (внешнее покрытие клетки) множество крошечных волосоподобных структур, называемых ресничками (1) . Биение ресничек очень похоже на движение лодочных весел: они толкают парамеций так, что ее передний (передний) конец перемещается вперед. У Paramecium нет глаз, поэтому, если они ударяются о что-то твердое, они отступают, меняют направление и снова движутся вперед, как показано на схеме.
Проглатывание
Проглатывание попадает в организм с пищей.На стороне парамеции примерно посередине находится оральная бороздка (2) . По мере продвижения парамеций вода с пищей, включая бактерии и водоросли, попадает в ротовую канавку. На заднем конце ротовой бороздки находится глотка, в которой собирается пища. По мере того, как больше пищи собирает концы баллончиков пищевода, они в конечном итоге отламываются в виде пищевой вакуоли (3) .
Пищеварение
Пищеварение — это расщепление пищи на мелкие частицы, которые могут быть использованы клеткой.Пищевая вакуоль (3), отрывается от пищевода на заднем конце и движется через цитоплазму (5), , которая представляет собой гелеобразный материал внутри клетки. По мере движения пищевой вакуоли фермент из цитоплазмы попадает в вакуоль и переваривает скопившуюся пищу, включая бактерии, водоросли и / или дрожжи. После переваривания питательные вещества перемещаются через мембрану вакуоли в цитоплазму. По мере продолжения этого процесса вакуоль уменьшается в размерах. Когда сморщенная вакуоль завершает свой путь, вакуоль и оставшаяся в ней непереваренная пища выводится через анальную пору (4) .
Экскреторная
Наряду с анальной порой (4) , , которая удаляет твердые отходы, парамеций имеет сократительных вакуолей (6) , которые выводят воду. Сократительные вакуоли расположены у поверхности и на обоих концах клетки. Отработанный газ, диоксид углерода, проходит через клеточную мембрану в воду вне клетки.
Репродукция
Paramecium размножаются бесполым путем бинарного деления, при котором полностью выросший организм делится на две отдельные дочерние клетки; Микроядро (8) содержит генетический материал, необходимый для воспроизводства.
Центр управленияm acronucleus (7) контролирует все функции выживания организмов.
Применение в реальном мире: карьераИзучение микроорганизмов является частью многих направлений биологии, медицины и здравоохранения. Постоянные исследования небесных тел расширяют потребность в понимании влияния микрогравитации на микроорганизмы. Куда бы люди ни пошли, они несут с собой микроорганизмы. НАСА сейчас изучает микроорганизмы, взятые с Международной космической станции (МКС) . Как пребывание в космосе меняет микробы на МКС. Нет предела карьере, которая продолжит развиваться благодаря космической программе НАСА, такой как A стромикробиология, микробный инженер, генная инженерия и т. Д. …………………………
Для получения дополнительной информации о микробах и окружающей их среде см.
Дженис ВанКлив «Экология для каждого ребенка».Связанные
Макроядерная последовательность генома инфузорий Tetrahymena thermophila, модельный эукариот
Сборка генома и общая структура хромосомы
Секвенирование и сборка.
Используя методы физического выделения, МАК очищали из культуры T. thermophila штамма SB210 и использовали для создания множества библиотек секвенирования разного размера (таблица S1). Создание больших (более 10 т.п.н.) библиотек вставок не увенчалось успехом — обычная проблема при работе с геномами, богатыми AT. Приблизительно 1,2 миллиона парных концевых последовательностей были сгенерированы из библиотек и собраны с использованием Celera Assembler [37]. В начальной сборке митохондриальный геном (мтДНК; который присутствовал из-за некоторого загрязнения препарата MAC митохондриями) и сильно амплифицированная хромосома рДНК не собирались хорошо по сравнению с опубликованными последовательностями этих молекул [38,39].Вероятно, это было связано с тем, что контиги от этих молекул имели более высокую глубину покрытия, чем контиги от других хромосом, что заставило Celera Assembler рассматривать их как повторяющуюся ДНК. Таким образом, мы разделили чтение последовательностей на три блока (мтДНК, рДНК и основная ДНК MAC) и сгенерировали сборки для каждого блока отдельно. Это привело к умеренному улучшению, и, таким образом, три отдельные сборки использовались для всех последующих анализов. Подробная информация о последовательности и сборке представлена в таблицах 1 и S2.
Основная сборка MAC содержит 1 971 каркас (контиги, которые были связаны в более крупные части с помощью информации о парных парах) с общим предполагаемым размером 104,1 Мб. Возможно, наиболее важно, используя комбинацию вычислительной и экспериментальной идентификации теломер, мы обнаружили, что многие концы каркаса соответствуют концам хромосом. Сто двадцать пять каркасов, составляющих 44% длины собранного генома, закрыты теломерами на обоих концах и, таким образом, вероятно, представляют собой полные MAC-хромосомы.Сто двадцать дополнительных каркасов, охватывающих еще 31% генома, закрыты теломерами на одном конце (Таблицы 1 и S3).
Точность и полнота сборки.
В целом, все анализы показывают высокую точность сборок MAC. Например, все 75 локусов MAC, которые находятся в разных генетических группах совокупного ассортимента (и, следовательно, должны находиться на разных хромосомах [40]), отображаются на разных каркасах, и все пары локусов, которые коассортируются (и, следовательно, должны находиться на одной хромосоме) либо отображаются на один и тот же скаффолд, либо на два не полностью закрытых скаффолда, чей совокупный размер меньше, чем у соответствующей MAC хромосомы (Table S4).Для 24 полностью собранных хромосомных каркасов, для которых мы знаем соответствующий физический размер хромосомы, существует очень сильная корреляция между физическим размером и длиной сборки. Кроме того, нет случаев, когда каркас значительно длиннее физического размера соответствующей хромосомы (рис. 3А). Наконец, все 96 последовательностей MIC, которые, как известно, соседствуют с сайтами Cbs [24,41,42], которые соответствуют каркасу MAC, сделали это только на конце каркаса.
Рисунок 3.Размеры скаффолда
(A) Размеры скаффолда по сравнению с размером хромосомы MAC. Голубые ромбы представляют собой каркасы, покрытые теломерами с обоих концов. Красные квадраты и зеленые треугольники представляют собой незавершенные каркасы, покрытые теломерами с одного или обоих концов, соответственно.
(B) Распределение размеров каркасов, покрытых теломерами с обоих концов.
https://doi.org/10.1371/journal.pbio.0040286.g003
Общая точность сборок указывает на то, что многие из потенциальных трудностей, обсуждаемых во Введении, не были значительными.Например, мы видим мало доказательств полиморфизма среди прочтений, что, вероятно, является отражением использования инбредной линии и процесса фенотипического ассортимента, который приводит к полногеномным гомозиготным линиям MAC [43]. Кроме того, поиск известных последовательностей, специфичных для MIC, показывает, что степень загрязнения MIC очень низка (например, соединения Cbs имеют 0,044-кратное покрытие, что примерно в 200 раз меньше, чем объемные хромосомы MAC) и ограничиваются небольшими контигами (большинство меньше чем 5 кб). Равномерная глубина покрытия контигов и точность сборок также предполагают, что хромосомы присутствуют в примерно одинаковом количестве копий и что только ограниченные количества повторяющейся ДНК присутствуют в MAC, оба из которых дополнительно обсуждаются ниже.
Общая длина каркаса намного меньше предсказанного размера генома от 180 до 200 мегабайт [14]. Учитывая точность сборок, большое количество хромосом, частично или полностью закрытых, и тот факт, что все (более 200) известные последовательности ДНК MAC обнаружены в сборках, мы заключаем, что сборки представляют собой очень большие (более 95 %) фракция генома. Таким образом, мы пришли к выводу, что предыдущие оценки размера генома были неточными (что неудивительно, учитывая, что они были сделаны почти 30 лет назад) и что размер генома близок к 105 мегабайтам.Однако возможно, что некоторые хромосомы или области были недостаточно представлены в наших библиотеках из-за смещения очистки или клонирования, и, таким образом, нельзя сделать вывод об отсутствии какого-либо конкретного гена или функции просто из-за их отсутствия в наших текущих сборках.
Оценка количества хромосом MAC.
Общее количество хромосом MAC неизвестно. Теломерное покрытие каркасов позволяет нам установить минимальную границу для этого числа на уровне 185 (125 плюс половина 120). Один из способов оценки действительного числа — это анализ прочтений, не содержащих теломеры рДНК; 3328 таких считываний могут быть связаны в общей сложности с 370 концами каркаса.Это соответствует приблизительно 9-кратному охвату (3,328 / 370), что существенно не отличается от общего покрытия MAC-хромосом, равного 9,08, указывая на то, что нет значительной недопредставленности прочтений теломер (Таблицы 1 и S3). Таким образом, поскольку всего имеется 4058 таких считываний (остальные не могут быть связаны), мы оцениваем, что существует приблизительно 451 конец теломер (4058/9), и, таким образом, имеется примерно 225 хромосом (451/2). Независимая оценка фактического числа хромосом может быть сделана, если предположить, что распределение размеров полностью закрытых хромосом (см. Рисунок 3B) является репрезентативным для генома в целом.Поскольку эти 125 закрытых хромосом составляют 43,5% от общей длины сборки, это может предсказать всего 287 хромосом (125 / 0,435). Это, вероятно, будет завышенной оценкой, поскольку более крупные хромосомы статистически менее вероятно, что они будут в полностью собранном наборе. Действительно, средний размер полностью собранных хромосом составляет 359 т.п.н., тогда как оценки среднего размера MAC-хромосом, полученные с помощью гель-электрофореза в импульсном поле, значительно выше [29,41]. Таким образом, мы заключаем, что существует от 185 до 287 хромосом, скорее всего, где-то около 225.
Отсутствие многих стандартных глобальных признаков эукариотических хромосом.
Отметим, что мы искали, но не смогли найти многие из того, что считается стандартными глобальными характеристиками эукариотических хромосом. Напр., Мы не смогли найти последовательности или структурные особенности, общие для нескольких хромосом, которые могли бы рассматриваться как кандидаты на центромерные регионы. Это согласуется с экспериментальными исследованиями [44]. Кроме того, хотя у многих эукариот определенные гены и повторяющиеся элементы группируются около теломер [45–51], мы не можем обнаружить здесь какой-либо такой кластеризации.Это не потому, что эти функции не отличаются друг от друга; например, содержание GC (Рисунок S1) и плотность генов (Рисунок S2) сильно различаются. Вместо этого, отсутствие сходной глобальной структуры между MAC-хромосомами, вероятно, связано с отсутствием процессов, которые помогают генерировать ключевые особенности нормальных эукариотических хромосом (например, митоз и мейоз, которые у T. thermophila ограничены MIC).
Количество копий хромосомы MAC одинаково.
Высокое качество и полнота сборок предполагают, что вариация числа копий по крайней мере среди большинства MAC-хромосом относительно невелика, поскольку в противном случае ассемблер рассматривал бы контиги из чрезмерно представленных хромосом как повторяющуюся ДНК.Такое равномерное количество копий согласуется с генетическими экспериментальными данными для шести хромосом [31], но его универсальность для всех хромосом неизвестна. Мы поняли, что относительное количество копий хромосом можно оценить по глубине охвата наших сборок (при условии, что клонирование и секвенирование были относительно случайными). Когда все каркасы исследованы, глубина покрытия становится удивительно однородной (рис. 4). Уменьшение однородности и покрытия, наблюдаемое по мере уменьшения размера каркаса, вероятно, является отражением как случайного низкого покрытия некоторых регионов, так и того, что некоторые из небольших каркасов являются контаминантами МПК.Когда в анализ включены только каркасы, покрытые теломерами на обоих концах, наблюдаемое покрытие последовательностей становится еще более однородным (красные ромбы на рисунке 4). Хотя мы не можем исключить, что некоторые меньшие, не полностью собранные хромосомы поддерживаются с разным числом копий, наблюдаемая однородность указывает на то, что репликация и / или сегрегация большей части или всех основных хромосом MAC находится под скоординированным регулированием.
Общие характеристики предсказанных генов, кодирующих белки, и некодирующих РНК
Прогнозы генов, кодирующих белок.
Мы идентифицировали 27 424 предполагаемых гена, кодирующих белок в геноме (таблица 2), большое количество для одноклеточных видов. Эти генные модели были протестированы путем сопоставления тегов экспрессируемых последовательностей (EST) со сборками генома с использованием PASA [52]. Мы отмечаем, что большинство этих EST были созданы после того, как модели были построены (Таблица S5). Из 9122 идентифицированных кластеров EST большинство либо не конфликтует с генными моделями (49,5%), либо относительно небольшие (17,7% имеют пропущенный экзон и 9,8% предполагают, что модели необходимо объединить или разделить).Только 408 (4,4%) кластеров являются межгенными по сравнению с генными моделями. Хотя они могут представлять собой пропущенные гены или участки генов, они также могут быть некодирующими РНК (нкРНК) или загрязнением геномной ДНК библиотек кДНК. Кроме того, предсказанные интроны и интроны, полученные из EST, весьма схожи по распределению размеров, за исключением коротких и длинных крайних значений (Рисунок S3), содержания GC (16,3% против 16,7%) и сайтов сплайсинга [только небольшое количество (85) из У интронов на основе EST есть исключения из соединений 5′-GT… AG-3 ‘, предполагаемых моделью — это могут быть просто ошибки секвенирования].Эти анализы показывают, что генные модели относительно надежны и их должно быть более чем достаточно для того, чтобы делать общие прогнозы относительно кодирующего потенциала этого вида.
Две другие линии доказательств предполагают, что предсказанное число генов не завышено. Во-первых, большое количество предсказанных генов совпадают с известными или предсказанными генами других видов (14 916 имеют совпадение BLASTP с E-значением лучше, чем 10 -10 ), а во-вторых, экспериментальные исследования сложности мРНК предсказывают транскрипцию не менее 25 000 генов со средним размером 1 200 п.н. [53].Отметим также, что последовательность самой большой MAC-хромосомы другой инфузории, Paramecium tetraurelia, , указывает на высокую плотность кодирования, а экстраполяция на полный геном предсказывает, по крайней мере, 30 000 генов, кодирующих белок [54].
нкРНК и использование всех 64 кодонов для кодирования аминокислот.
нкРНК, обнаруженные в геноме, перечислены в таблице S6. Обращаем внимание на несколько новых открытий. Из 174 предполагаемых генов 5S рРНК (таблица S6A) 19 не соответствуют ни одному из четырех ранее описанных генов T.thermophila [55,56]. Эти 19 отличаются друг от друга простыми заменами нуклеотидов в 34 положениях, а также различными вставками, делециями и усечениями и могут представлять собой псевдогены. Кроме того, присутствуют две формы мяРНК U2 (Таблица S6C), которые мы назвали U2 (четыре гена) и U2var (пять генов). Семейства функциональных генов РНК экспрессируются повсеместно в течение жизненного цикла T. thermophila, а также в стрессовых условиях (репрезентативные данные показаны на рисунке S4).Самый большой класс — это тРНК с 700 идентифицированными (таблицы S6B и S6D), число, согласующееся с оценками, основанными на гибридизации [57].
Одной из наиболее необычных особенностей T. thermophila и некоторых других инфузорий является использование альтернативного генетического кода, в котором канонические стоп-кодоны UAG и UAA кодируют глутамин [58]. Важность и возраст этого альтернативного кода отражены в геноме наличием 39 тРНК для этих кодонов. Примечательно, что анализ генома также выявил присутствие тРНК, которая, как предполагается, декодирует оставшийся стоп-кодон, UGA.Многочисленные доказательства указывают на то, что это действующая тРНК селеноцистеина (Sec), так называемой 21-й аминокислоты. У тех эукариотических видов, которые используют Sec, большинство кодонов UGA все еще вызывают терминацию трансляции, в то время как те мРНК, которые кодируют Sec-содержащие пептиды, имеют характерный мотив последовательности стебель-петля в 3′-области UTR, который управляет включением Sec [59,60]. Предполагаемая тРНК-Sec T. thermophila была идентифицирована путем анализа последовательности генома и показала, что она транскрибируется и ацилируется [61], и мы обнаружили, что она экспрессируется и заряжается и что ее заряд может находиться под отдельным регуляторным контролем со стороны других тРНК. (Рисунок S4A).Кроме того, мы идентифицировали шесть генов T. thermophila с кодонами UGA в рамке считывания, которые выравниваются (после редактирования генных моделей) с известными кодонами Sec их гомологов из других видов эукариот и которые имеют консенсус «стебель-петля» и, таким образом, вероятно, будут кодируют селенопротеины. Таким образом, мы заключаем, что UGA почти наверняка транслируется в Sec, что сделало бы T. thermophila первым известным организмом, использующим все 64 триплетных кодона для определения включения аминокислот.
Эволюция генома
Ошибка использования кодонов и аминокислот.
Хотя T. thermophila может использовать все 64 кодона, но не все они одинаково. Наиболее важным аспектом использования кодонов у этого вида является то, что богатые AT кодоны, как правило, используются чаще, чем другие [62,63]. Таким образом, хотя смещение AT в геноме наиболее сильно в некодирующих областях, где считается, что отбор ослаблен, он наблюдается даже в кодирующих областях. Фактически, притяжение AT настолько велико в кодирующих областях, что аминокислотный состав белков смещен в сторону кодонов с высоким содержанием AT, как это видно у других видов с экстремальным смещением AT (например.г., [64]). Хотя общее использование кодонов смещено по сравнению с кодонами, богатыми GC, на уровне каждого гена существует значительная вариация в степени смещения. Мы определили два доминантных паттерна этой вариации от одного гена к другому. Основная закономерность состоит в том, что для большинства генов используемые кодоны являются просто отражением общего содержания AT в гене (рис. 5). Различия между генами обусловлены вариациями содержания AT в пределах генома (см. Рисунок 5A), хотя мы не смогли выявить механизм, лежащий в основе этой вариации (например,g., нет кластеризации генов с высоким или низким AT возле теломер). Однако существует менее распространенный образец вариации от одного к другому, что очень важно. Существует подмножество генов (показано красным), которые используют общий предпочтительный набор кодонов, который отличается от такового для среднего гена, и кодоны в этом наборе не сильно коррелируют с содержанием AT генов. Хотя о существовании такого предпочтительного набора кодонов для этого вида сообщалось [62,63], анализ генома позволяет более точно определить набор и гены, которые его используют.Всего, используя относительно консервативное отсечение (рис. 5В), мы идентифицировали 232 таких гена.
Рисунок 5. Использование кодонов
(A) Эффективное количество кодонов (ENc; мера общего смещения кодонов) для каждой предсказанной ORF отображается в зависимости от GC3 (доля кодонов, которые являются синонимичными в позиции третьего кодона, которые имеют либо гуанин или цитозин в этом положении). Верхний предел ожидаемого смещения, основанного только на GC3, представлен черной кривой; большинство ОРС T. thermophila группируются под кривой [красные точки, как на (B)].
(B) Анализ главных компонентов относительного использования синонимичных кодонов у T. thermophila. 232 гена в хвосте распределения в форме запятой (те, которые используют наиболее предвзятые кодоны) окрашены в красный цвет.
(C) Анализ главных компонентов относительного использования синонимичных кодонов в P. falciparum.
https://doi.org/10.1371/journal.pbio.0040286.g005
Считается, что использование геном предпочтительных кодонов обеспечивает более эффективную или точную трансляцию [65].Это, по-видимому, имеет место здесь, поскольку из предсказанных генов с использованием предпочтительного подмножества многие, вероятно, имеют функции домашнего хозяйства, и, хотя они составляют только 0,85% всех предсказанных генов, 12,5% всех EST сопоставляются с ними (Таблица S7). . Хотя некоторые из них не имеют совпадений EST и теоретически могут представлять ошибочно предсказанные гены, кажется маловероятным, что ложные гены будут использовать предпочтительный набор кодонов. Таким образом, мы прогнозируем, что эти гены-выбросы либо сильно экспрессируются (по крайней мере, в некоторых из условий, с которыми обычно сталкивается организм), либо выполняют некоторую критическую функцию, требующую точной трансляции.
Считается, что различия в использовании кодонов между генами оказывают лишь небольшое влияние на приспособленность. Чтобы естественный отбор эффективно воздействовал на различия в использовании кодонов и, таким образом, создавал предпочтительное подмножество, факторы, усиливающие генетический дрейф (например, небольшие размеры популяции, инбридинг), должны быть слабее, чем силы отбора [66]. Таким образом, хотя использование кодонов, вероятно, находится под давлением отбора у всех видов, не все могут развить предпочтительные наборы кодонов. Например, хотя у него такое же смещение AT с T.thermophila, , в апикомлексе Plasmodium falciparum не было обнаружено предпочтительного набора (рис. 5C), что, возможно, отражает его паразитический образ жизни и ограниченный эффективный размер популяции. Присутствие предпочтительной подгруппы у T. thermophila, вероятно, является отражением большого эффективного размера популяции из-за его свободно живущего, воспроизводящего половым путем образа жизни (см. [66,67] для дополнительного обсуждения большого размера популяции этого вида).
Гены, происходящие из пластид, не могут быть идентифицированы.
Один из вопросов, представляющих особый интерес, на который геном T. thermophila может пролить свет, касается времени происхождения пластид, обнаруженных у apicomplexans и dinoflagellates, других членов альвеолят [68,69]. Хотя пластиды в этих линиях различаются (например, пластиды apicomplexans, известные как апикопласты, даже не участвуют в фотосинтезе), оба, как полагают, имеют происхождение из красных водорослей [70]. Это привело к предположению, что пластиды в этих линиях являются результатом единственного эндосимбиотического события между предком apicomplexans и динофлагеллят и красной водорослью, при этом ядро водорослей потеряно, а пластида водорослей сохраняется.Ключевой вопрос заключается в том, произошел ли этот вторичный эндосимбиоз до или после отделения инфузорий от двух других ветвей. Возможность того, что это произошло до расщепления инфузорий, известна как хромальвеолатная гипотеза [71].
Для того, чтобы гипотеза хромальвеолатов была верной, потеря пластид должна была произойти у инфузорий, скорее всего, в основании дерева инфузорий, поскольку современные инфузории, как известно, не содержат пластиды. Если бы у предка инфузорий когда-то была пластида, возможно, что некоторые гены, полученные из пластид, были бы перенесены в ядерный геном (как это произошло во многих линиях, включая апикомплексы и динофлагелляты [72]), и, кроме того, некоторые такие гены будут все еще можно найти у Т.термофила. Чтобы проверить эту возможность, мы построили филогенетические деревья всех генов в геноме и провели поиск генов с паттерном ветвления, соответствующим пластидному происхождению (см. Материалы и методы). Для T. thermophila, w e не видит никакого сигнала для генов пластидного происхождения, который превышает шум, наблюдаемый в таких автоматизированных филогенетических анализах.
Несколько линий свидетельств предполагают, что это не общий недостаток филогенетического подхода, использованного здесь. Например, мы использовали тот же подход для идентификации и каталогизации пластидных генов в других линиях, включая растение Arabidopsis thaliana и apicomplexan P.falciparum. Кроме того, такой подход был использован для обнаружения прошлых эндосимбиозов у других эукариотических клонов [73]. Наконец, используя тот же подход, мы идентифицировали 91 вероятно происходящий из митохондрий ген (Таблица S8) в ядерном геноме T. thermophila. Это важно, потому что гены, происходящие из митохондрий, обычно труднее идентифицировать, чем гены, происходящие из пластид [74], отчасти потому, что пластидный симбиоз появился позже [75].
Тем не менее, поскольку возможно, что наш филогеномный скрининг мог пропустить некоторые гены, происходящие из пластид, мы также провели целенаправленный поиск генов, которые, как можно было ожидать, будут сохранены, используя апикопласт в качестве модели.Апикопласты участвуют в биосинтезе жирных кислот, изопреноидов и гема. Пути биосинтеза жирных кислот и изопреноидов представляют особый интерес, потому что пути, происходящие из пластид, отличаются от аналогичных путей в цитоплазме эукариот [76]. В случае биосинтеза изопреноидов гены белков в каноническом эукариотическом цитозольном мевалонатном пути присутствуют, как и ожидалось на основании экспериментальных исследований [77–79], но не было обнаружено никаких ферментов, участвующих в пути DOXP, происходящем из пластид.Что касается биосинтеза жирных кислот, хотя T. thermophila не требует экзогенного поступления жирных кислот для роста, не удалось найти никаких доказательств полной версии пути типа I (обычно цитозольного). Хотя присутствуют по крайней мере некоторые гены пути типа II, их недостаточно для синтеза жирных кислот de novo и, по-видимому, более вероятно, что они происходят из митохондрии, чем из пластиды.
На основании общего и целевого поиска мы пришли к выводу, что в настоящее время нет доказательств наличия пластидных или предковых пластидных генов у T.термофила. Это не исключает возможности того, что другие инфузории имеют ферменты, происходящие из пластид или даже пластиды, но в настоящее время нет доказательств, подтверждающих это, несмотря на обширные ультраструктурные наблюдения [80,81]. Если у инфузорий отсутствуют все доказательства пластиды, это может означать либо то, что предполагаемое раннее происхождение хромальвеолатной пластиды неверно, либо то, что предок T. thermophila (и, возможно, все инфузории) полностью потерял пластиду и все определяемые гены, происходящие из пластид. .Последняя возможность имеет прецедент, так как некоторые апикомплексы, такие как Cryptosporidia, потеряли свои апикопласты и имеют немного, если таковые имеются, производных от пластид генов в своих ядерных геномах [82,83]. Было высказано предположение, что эта потеря является результатом рационализации метаболизма в ответ на паразитарный образ жизни. Решение вопроса о том, присутствовала ли пластида у предка инфузорий, будет важно для нашего понимания эволюции пластид и их биохимических взаимоотношений с эукариотическими хозяевами.
ВырезаниеIES нацелено на чужеродную ДНК, а не на повторяющуюся ДНК
как таковую.Как обсуждалось во Введении, существует множество параллелей между процессом вырезания IES и другими явлениями подавления повторяющихся элементов, такими как RIP и образование гетерохроматина. Несмотря на эти параллели, процессы значительно различаются по своим механизмам действия и, следовательно, вероятно, имеют разные краткосрочные и долгосрочные эволюционные последствия. Например, у видов с RIP вся повторяющаяся ДНК становится мишенью для мутационной инактивации, что привело к резкому подавлению эволюционной диверсификации за счет дупликации генов [84,85].Процесс вырезания IES приводит к исключению определенных последовательностей ДНК MIC из транскрипционно активного MAC. Экспериментальное введение чужеродных трансгенов в МИК показало, что с увеличением числа копий МИК повышается эффективность удаления трансгена [86]. Следовательно, можно предсказать такое же подавление дупликации генов, что и при RIP. Однако, вместо того, чтобы нацеливаться на повторяющуюся ДНК как таковую, было предложено, чтобы эксцизия IES специфически нацелена на чужеродную ДНК, которая вторглась в MIC зародышевой линии, но не представлена в MAC [35,87,88].Дублирование гена MIC и функциональная диверсификация все еще возможны в этом сценарии, если при каждом событии конъюгации копии гена не расходятся в последовательности, достаточной для того, чтобы их можно было распознать как чужеродные и исключить из MAC; так как секс является частым явлением в естественных популяциях T. thermophila [89], это должно быть так. Поэтому мы стремились использовать данные последовательности генома как для проверки гипотезы о чужеродной ДНК, так и для изучения последствий процесса эксцизии IES для эволюции T.геном thermophila.
Анализ генома позволяет выявить несколько линий доказательств, которые убедительно подтверждают гипотезу о чужеродной ДНК. Во-первых, в МАК присутствует небольшое, но, тем не менее, значительное количество повторяющейся ДНК. Это лучше всего видно при анализе каркасов, которые соответствуют полным хромосомам MAC, которые вряд ли содержат контаминацию MIC IES. Эти каркасы содержат диспергированные повторы, составляющие 2,3% всей ДНК. Это означает, что некоторая повторяющаяся ДНК обходит процесс вырезания IES.Вторая линия доказательств прибывает из исследования небольших контигов и синглтонов (несобранных последовательностей) в данных сборки. Известные специфические для MIC элементы, такие как транспозоны REP и Tlr 1 [90,91], обнаруживаются только в этих небольших контигах, которые, таким образом, явно обогащены специфичной для MIC ДНК (а также для повторяющейся ДНК; см. Рисунок S5). Фактически, небольшие контиги содержат гомологи необычайно широкого диапазона кладов мобильных элементов (TE) для одноклеточного эукариота [92,93], включая многие ранее не описанные в Tetrahymena (Таблица S9).Мы не находим подходящих совпадений с TE ни в одном из крупных контигов. Таким образом, транспозоны в целом, по-видимому, очень эффективно отфильтровываются процессом вырезания IES. Тандемные и диспергированные повторы в MAC, по-видимому, соответствуют неинвазивной ДНК (например, генам 5S рРНК). Взятый вместе, тот факт, что мобильные (и, вероятно, инвазивные) элементы ДНК не попадают в MAC, в сочетании с тем фактом, что как тандемные, так и диспергированные неинвазивные повторы избегают процесса вырезания, указывает на сильную поддержку гипотезы чужеродной ДНК.
У организмов с RIP, поскольку вся дуплицированная ДНК является мишенью [94], диверсификация генов путем дупликации подавляется. Например, доля всех генов Neurospora crassa, обнаруженных в паралогичных семьях, составляет всего 19%, значение, которое находится ниже общей линии корреляции между этой долей и общим числом генов [84]. Кроме того, очень немногие пары генов имеют более 80% идентичности аминокислотной последовательности [84]. В соответствии с гипотезой чужеродной ДНК, мы не видим таких признаков подавления диверсификации семейства генов у T.термофила. В геноме обнаружено большое количество паралоговых генов (1970 семейств генов, включая 10 851 предсказанный белок) (Таблица 3). Доля генов таких семейств у T. thermophila (39%) намного выше, чем у N. crassa. Хотя эта доля не так высока, как можно было бы предсказать из наблюдаемой корреляции между общим числом генов и фракцией, обнаруженной в паралогичных семьях [84], доля пар генов, имеющих более 80% аминокислотной идентичности, намного выше, чем в N .crassa и аналогично тому, что обнаружено у других секвенированных эукариот.
Поскольку возможно, что некоторые из 1970 семейств генов могли возникнуть в результате дупликаций, произошедших до начала процесса вырезания IES, более полезно изучить недавние дупликации . Мы искали такие дупликации множеством способов, включая идентификацию генов, дублированных в линии T. thermophila относительно других линий, для которых доступны геномы (Таблица S10), и путем поиска пар паралогов с очень похожими последовательностями.Оба этих класса широко распространены у T. thermophila, , что дополнительно указывает на то, что иссечение IES не оказывает значительного влияния на расширение семейств генов «нативных» генов. Таким образом, инфузорийная система нацеливания на вторгающуюся ДНК имеет значительно другие последствия, чем RIP.
Большое количество генов у
T. thermophila.Расширение семейств генов помогает объяснить высокое количество генов у T. thermophila, , которое выше, чем у других протистов, и даже превосходит количество некоторых многоклеточных (Таблица 4).События дублирования, по-видимому, распределены по эволюционному времени, причем некоторые из них были древними, а некоторые — совсем недавними. Мы искали, но не нашли доказательств ни полногеномной, ни сегментарной дупликации. Мы действительно находим огромное количество тандемно дублированных генов. Всего было обнаружено 1603 тандемных кластера от двух до 15 генов, включая 4276 генов; 67% этих кластеров представляют собой простые пары генов, а 96% содержат пять или меньше генов. Таким образом, многие гены-паралоги T.thermophila — это результат отдельных небольших событий дублирования.
Большое количество генов у T. thermophila по сравнению с некоторыми другими одноклеточными эукариотами не просто отражение расширения семейства генов. Например, когда недавние экспансии генов объединяются в наборы ортологов, мы обнаруживаем, что у людей и T. thermophila больше ортологов друг с другом (2280), чем у людей и дрожжей S. cerevisiae (2097) или T. thermophila и P. .falciparum (1,325) (рис. 6), несмотря на родственные связи между животными и грибами, с одной стороны, и инфузориями и апикомплексами, с другой.Мы отмечаем, что это не означает, что люди и T. thermophila в целом более похожи друг на друга, чем на виды в сестринских типах. Например, люди и S. cerevisiae действительно разделяют некоторые процессы, которые развились у общего предка грибов и животных. Кроме того, для ортологов, обнаруженных у всех эукариот, гены человека и S. cerevisiae более похожи по последовательности друг на друга, чем на гены T. thermophila. Большее количество ортологов, общих между людьми и T. thermophila, является отражением как потери генов в других эукариотических линиях, так и сохранения T.термофила. В соответствии с этим выводом, существует 874 человеческих гена с ортологами у T. thermophila, но не у S. cerevisiae, 58 из которых соответствуют локусам, связанным с заболеваниями человека (Таблица S12). Таким образом, анализ генома показывает множество случаев, когда T. thermophila может продолжать дополнять экспериментальные исследования дрожжей как модельной системы для клеточной биологии эукариот (и человека) [13].
Рисунок 6. Ортологи, общие для T. thermophila и отдельных эукариотических геномов
Диаграмма Венна, показывающая ортологи, общие для человека, дрожжей S.cerevisiae, апикомплексан P. falciparum, и T. thermophila. Дупликации генов, специфичных для клонов, в каждом из организмов были идентифицированы и рассматривались как один единственный ген (или суперортолог). Затем попарные взаимные наилучшие совпадения по BLASTP были идентифицированы как предполагаемые ортологи.
https://doi.org/10.1371/journal.pbio.0040286.g006
Дупликация гена как индикатор важных биологических процессов
Одним из мотивов получения последовательности генома организма является продвижение изучения уже исследуемых процессов.Многие исследователи, в том числе те, кто никогда раньше не работал с этим видом, воспользовались общедоступными данными, чтобы достичь этой цели (например, [24,95–103]). Вместо того, чтобы сосредоточить наш биоинформатический анализ на этих хорошо изученных процессах, мы решили искать доказательства в предсказанном протеоме процессов, имеющих особое значение для организма. Наш подход был относительно простым — мы искали избыточное количество (по сравнению с другими эукариотами) в списках семейств паралогичных генов или расширений семейств генов, специфичных по клонам, связанных с различными процессами.Такой подход был выбран по нескольким причинам. Во-первых, поиск различий в больших семействах генов не так подвержен ошибкам аннотации, как поиск, сфокусированный на отдельных генах. Кроме того, большие семейства генов явно вносят вклад в большое количество генов, присутствующих в T. thermophila по сравнению с другими одноклеточными эукариотами. Мы отмечаем, что многие из доступных геномов одноклеточных эукариот принадлежат паразитам, которые были выбраны для секвенирования в основном из-за их медицинской значимости, и что они не являются репрезентативными (например,г., многие имеют довольно маленькие геномы). Наиболее важно то, что наличие больших семейств генов и недавние дупликации генов, вероятно, указывают на функциональное разнообразие, недавние эволюционные инновации и давление отбора, оказываемое на этот организм.
Наш анализ семейств паралогичных генов и, в частности, недавно дублированных членов таких семейств показывает важность процессов, связанных с восприятием изменений окружающей среды и реагированием на них. Мы выделяем здесь пять таких процессов: трансдукция сигнала, мембранный транспорт, протеолитическое расщепление, конструирование и изменение формы и движения клеток и мембранный перенос.Все эти процессы имеют решающее значение для свободноживущего гетеротрофного образа жизни этого организма. В следующих разделах мы обсудим, что анализ генома показывает об этих процессах у T. thermophila, уделяя особое внимание расширению генов, связанных с этими функциями, по сравнению с другими видами.
Сигнальная трансдукция и расширения семейств киназ.
Множество генов, предположительно играющих роль в передаче сигналов, были идентифицированы в наших скринингах паралоговых генов.Из них мы решили провести углубленный анализ киназ, потому что они представляют собой такое разнообразное семейство белков и потому, что было обнаружено, что они играют важную роль в сенсорных и регуляторных процессах на всем древе жизни. Всего в геноме было идентифицировано 1069 предсказанных протеинкиназ (таблицы 5 и S11A). Это соответствует примерно 3,8% предсказанного протеома, фракции значительно больше, чем примерно 2,3% у грибов, Drosophila, и позвоночных [104].Среди них были обнаружены представители 54 известных семейств и подсемейств киназ [105]. Некоторые семейства, обнаруженные у большого разнообразия эукариот [106], не были обнаружены. Сюда входят киназа контрольной точки CHK1 / RAD53, связанная с киназой PI3 киназа TRRAP, две циклин-зависимые киназы (CDK7 и CDK8, которые могут быть функционально заменены родственным расширенным семейством CDC2) и два плохо консервативных класса (Bub1 и Haspin). которые могли быть пропущены поисками гомологии последовательностей. Несмотря на сообщения о наличии фосфотирозина в T.thermophila [107], явных членов группы тирозинкиназ идентифицировать не удалось. Однако геном кодирует некоторые белки, которые могут быть альтернативными тирозинкиназами, включая множественные киназы двойной специфичности (например, Wee1, Ste7, TTK и Dyrk), а также пять членов родственной группы TKL, которые могут опосредовать фосфорилирование тирозина в слизи. плесень Dictyostelium discoideum [106]. Двенадцать классов киназ обнаружены у T. thermophila и человека, но не у дрожжей, и, таким образом, являются очевидными примерами сохранения предковых функций эукариот, обсуждаемых выше.Некоторые из генов этих классов вовлечены в этиологию заболеваний человека (Dyrk1A, DNAPK, SGK1, RSK2, Wnk1 и Wnk4) [108].
Ключевой особенностью кинома T. thermophila является расширение нескольких классов киназ по сравнению с другими секвенированными организмами (Таблица 5). Последствия некоторых из этих расширений можно предсказать, основываясь на известных функциях членов семьи. Напр., Семейства митотических киназ Aurora, CDC2 и PLK все существенно расширились, возможно, отражая дополнительную сложность передачи сигналов, требуемую двумя ядрами, которые одновременно участвуют в очень разных процессах в одной и той же цитоплазме клетки.Также расширены множественные киназы, которые взаимодействуют с сетью микротрубочек [109,110] [напр., Nima-related kinases (NRKs) и семейство ULK], возможно, отражая диверсификацию цитоскелетных систем (обсуждается более подробно ниже). Из семейств киназ с известными функциями наиболее поразительным является присутствие 83 гистидиновых протеинкиназ (HPK), которые обычно участвуют в передаче сигналов из внешней среды [111]. HPK обнаруживаются преимущественно в двухкомпонентных регуляторных системах бактерий, архей, простейших и растений и отсутствуют у многоклеточных животных.Большинство HPK T. thermophila имеют домены-приемники субстрата, и многие из них, как предполагается, являются трансмембранными рецепторами.
Полное значение киномного разнообразия T. thermophila трудно предсказать, поскольку большая часть диверсификации произошла в классах, функции которых плохо изучены. Например, во многих известных семейств киназ белки T. thermophila сильно различаются по последовательности как по отношению к таковым у других видов, так и друг к другу (например.g., см. рисунок S6). Возможности диверсификации T. thermophila, возможно, лучше всего видны в том факте, что 630 (приблизительно 60%) киназ не могут быть отнесены к какому-либо известному семейству или подсемейству [105]. Всего в этом геноме идентифицировано 37 новых классов киназ и сотни уникальных белков. Присутствие стольких новых киназ и расширений во многих известных классах киназ одновременно является показателем универсальности эукариотического домена протеинкиназы, наблюдаемой в других клонах [112], и наводит на мысль о большой разработке функций, специфичных для ресничек.
Диверсификация мембранных транспортных систем.
Многие из наиболее широко распространенных семейств генов T. thermophila кодируют белки, которые, как предполагается, участвуют в мембранном транспорте. Мембранные транспортеры играют решающую роль в реагировании на изменения в окружающей среде и использовании доступных ресурсов. Поэтому мы провели более тщательный анализ предполагаемых переносчиков у этого вида. В целом T. thermophila обладает надежной и разнообразной коллекцией предсказанных систем мембранного транспорта (Таблицы 6 и S11B).Сравнение с другими эукариотами [113] обнаруживает некоторые интересные различия с точки зрения обоих классов переносчиков и предполагаемых перемещаемых субстратов. Например, T. thermophila имеет больше представителей в каждом из четырех основных семейств, чем человек. Кроме того, он кодирует гораздо большее количество переносчиков в суперсемействе ABC, потенциал-управляемых ионных каналов (VIC) и АТФаз P-типа, чем любой другой секвенированный вид эукариот (Таблица 6), включая других свободноживущих протистов, диатомовых водорослей. Thalassiosira pseudonana, и слизевик D.дискоидеум. Что касается субстратов, был идентифицирован чрезвычайно обширный набор переносчиков, вероятно, специфичных для неорганических катионов (таблица 6). Большинство из них являются транспортерами канального типа и АТФазами P-типа, транспортирующими катионы. Интересно, что, несмотря на очевидную массовую амплификацию переносчиков катионов, T. thermophila имеет очень ограниченный набор переносчиков неорганических анионов: были идентифицированы только по одному члену для сульфатного, фосфатного, арсенита и хромат-иона, а прогнозируемые анионные каналы отсутствуют.Причина разницы в амплификации переносчиков катионов и анионов неясна.
Как и в случае с киназами, некоторые из наиболее интересных свойств выявляются при изучении специфичных для клонов дупликаций переносчиков. Недавние кластеры включают белки канала K + (285 членов), транспортеры ABC (152 члена), транспортирующие катионы АТФазы (59 членов), белки бета субъединицы канала K + (22 члена), оксалат: формиатные антипортеры (24 члена). члены), транспортеры сахара (22 члена) и АТФазы, транспортирующие фосфолипиды (20 членов).Экспансия белков канала K + , которые являются переносчиками VIC-типа, была особенно значительной и продолжалась.
Всего было предсказано 308 селективных каналов VIC-типа K + , что намного больше, чем у любых других секвенированных видов, и более чем в три раза больше, чем у людей (89). Мультигенное семейство калиевых ионных каналов также было идентифицировано у P. tetraurelia [114] и, таким образом, может быть общей характеристикой некоторых инфузорий. Некоторые данные свидетельствуют о том, что это расширение у инфузорий может быть адаптивным.Во-первых, каналы K + контролируют пассивное проникновение K + через мембрану, что важно для подвижности ресничек [115]. Во-вторых, новая аденилилциклаза с предполагаемым N-концевым ионным каналом K + регулирует образование универсального вторичного мессенджера цАМФ в инфузориях и апикомплексах [116,117], который может помочь в ответе на внезапные изменения ионного окружения. T. thermophila кодирует шесть гомологов этой аденилатциклазы / транспортера K + , тогда как паразитические apicomplexans P.falciparum и Cryptosporidium parvum кодируют только по одному.
Присутствующие надежные транспортные системы, вероятно, являются отражением поведенческой и физиологической универсальности T. thermophila как свободноживущего одноклеточного организма и его воздействия на широкий спектр различных субстратов в естественной среде. Изучение конкретных типов расширений позволяет предположить, что функции, связанные с переносом K + и других катионов, значительно разнообразились.Таким образом, такие функции могут играть роль во многих уникальных аспектах биологии этого вида и инфузорий в целом.
Протеолитический процессинг.
T. thermophila — прожорливый хищник, поэтому можно ожидать, что он обладает широким разнообразием протеолитических ферментов. Анализ предсказанных белков в T. thermophila показывает некоторые противоречивые результаты, относящиеся к этой идее. С одной стороны, многие из крупнейших кластеров дупликаций, специфичных для клонов, относятся к протеазам (например,, папаин, лейшманолизин). С другой стороны, общее количество идентифицированных протеаз (480) относительно невелико с точки зрения доли протеома (1,7%) по сравнению с другими модельными организмами, которые были секвенированы и аннотированы [118–120]. Конфликт, скорее всего, является отражением разнообразия физиологических процессов, в которых функционируют протеазы [121]. Таким образом, мы рассмотрели подклассификацию типов протеаз, присутствующих более подробно.
Используя номенклатуру протеаз Merops, основанную на внутренних эволюционных и структурных отношениях [119], T.Протеазы thermophila были разделены на пять каталитических классов и 40 семейств. Это: 43 аспарагиновые протеазы, принадлежащие двум семействам, 211 цистеиновые протеазы, принадлежащие 11 семействам, 139 металлопротеаз, принадлежащие 14 семействам, 73 сериновые протеазы, принадлежащие 12 семействам, и 14 треониновых протеаз, принадлежащих семейству T1 (таблицы 7 и S11C). Некоторые уникальные особенности T. thermophila можно увидеть при сравнении с P. falciparum, который является наиболее близкородственным секвенированным видом, для которого опубликован подробный анализ его протеаз [122].В обоих геномах присутствует 21 семейство протеаз. Например, высококонсервативные треониновые протеазы и семейства убиквитинкарбоксил-концевых гидролаз (C12 и C19) отражают решающую роль АТФ-зависимой системы убиквитин-протеасома, которая участвует в контроле клеточного цикла и стрессовой реакции [123] . У T. thermophila присутствует девятнадцать семейств протеаз, но не P. falciparum. Один из них включает лейшманолизин (M8), первоначально идентифицированный у кинетопластидного паразита Leishmania major и предположительно участвующий в процессинге поверхностных белков [124–126].Это семейство значительно расширено (до 48 членов, включая 15 в тандемном массиве) у T. thermophila и предполагает, что здесь могут быть важны поверхностные белковые процессы, хотя функции лейшманолизин-связанных протеаз у некинетопластидных эукариот остаются неясными. Семейства карбоксипептидазы A (M14) и карбоксипептидазы Y (S10) расширены до 28 и 25 членов, соответственно, у T. thermophila, , что может отражать многочисленные и разнообразные функции. Только четыре семейства протеаз присутствуют у P.falciparum не обнаружены у T. thermophila. Среди них — метакаспаза (C14), наследственный тип каспазы, который характерен для апоптоза или апоптозоподобных путей передачи сигнала [127].
Самыми большими кластерами расширенных протеаз у T. thermophila являются все цистеиновые протеазы, которые составляют 44% от общего протеазного комплемента. Два наиболее известных семейства из этого класса — это семейство папаина (C1), которое является наиболее многочисленным и сложным семейством, состоящим из 114 членов, и семейство убиквитинкарбоксил-концевой гидролазы 2 (UCh3, C19), состоящее из 47 членов.Возможно, что биохимическая активность паралогов внутри этих семейств сохраняется, но они используются в разных частях клетки (или вне клетки) или на разных стадиях развития T. thermophila.
Компоненты и регуляторы цитоскелета.
Инфузории имеют очень сложную архитектуру цитоскелета [128] с сильно поляризованными типами клеток, которые собирают 18 типов микротрубочковых органелл в определенных местах вдоль переднезадней и дорсовентральной оси.Поэтому мы стремились определить, отражено ли это разнообразие в геноме. Как и в случае с анализом протеазы, описанным выше, первоначальное сравнение количества конкретных типов цитоскелетных и связанных с микротрубочками белков было несколько неоднозначным (числа для людей и T. thermophila показаны в таблицах 8 и S11D). Напр., Хотя двигатели кинезина и динеина, а также киназы, ассоциированные с микротрубочками, по-видимому, расширяются, структурные компоненты ресничек и участники пути внутрижладжкового транспорта не расширяются.Кроме того, некоторые типы белков цитоскелета, по-видимому, отсутствуют у T. thermophila; , они включают белки промежуточных филаментов (включая ядерные ламины), как уже предполагалось в биохимических исследованиях [129], некоторые белки, связанные с микротрубочками (MAP2, MAP4 и Tau, для которых не были обнаружены неживотные гомологи эукариот) и некоторые актин-связывающие белки. (например, α-актинин). Чтобы лучше понять, какую роль гены, участвующие в функциях микротрубочек и цитоскелета, могли играть в диверсификации этого вида, мы сосредоточили анализ на некоторых генах с очевидным расширением: тубулинах, динеинах и регуляторных белках.
Тубулины.
Тубулины являются ключевыми структурными компонентами микротрубочек, и у эукариот они встречаются во многих формах [130]. В геноме T. thermophila филогенетический анализ гомологов тубулина (рис. 7) выявляет присутствие одного или двух генов, каждый из основных подсемейств альфа (α), бета (β) и гамма (γ) (как сообщалось ранее [ 131–133]) и по одному в каждой из дельта (δ), эпсилон (ε) и эта (η), которые обнаруживаются у организмов, которые обладают центриолями / базальными тельцами [134–136].Кроме того, T. thermophila кодирует неканонические гомологи тубулина, которые можно разделить на две категории. К первой категории относятся гены, наиболее похожие на канонические α- или β-тубулины. Эти девять генов (три α-подобных и шесть β-подобных) лишены характерных мотивов для посттрансляционных модификаций хвостового домена (полиглутамилирование и полиглицилирование), которые существенны для функции их канонических аналогов [137–139]. Три β-подобных гена ( BLT1, / TTHERM_01104960, TTHERM_01104970 и TTHERM_01104980) образуют тандемный кластер с межгенными интервалами менее 2 т.п.н.Мы предполагаем, что эти гены функционируют, возможно, избыточно, в формировании или функционировании некоторых из многих высокоспециализированных систем микротрубочек клеток T. thermophila. Экспериментальный анализ BLT1 , β-подобного тубулина, показал, что его продукт локализуется в небольшом субнаборе микротрубочек и не включается в растущие аксонемы ресничек (К. Кларк и М. Горовский, неопубликованные данные). Генетическая делеция этого гена или α-подобного гена TTHERM_00647130 не дала явного фенотипа (R.Се и М.А. Горовский, неопубликованные данные).
Рисунок 7. Разнообразие гена тубулина у T. thermophila
На рисунке показано дерево соединения соседей, построенное на основе выравнивания clustalX. Сокращения видов: Hs, H. sapiens; Dm, D. melaogaster; Sc, S. cerevisiae; Tt, T. thermophila; Pt, P. tetraurelia; Cr, C. reinhardtii; Tb, T. brucei; Ec, E. coli; Xl, Xenopus laevis. Прокариотический ортолог тубулина, Escherichia coli FtsZ, был использован в качестве внешней группы.
https://doi.org/10.1371/journal.pbio.0040286.g007
Вторая категория неканонических гомологов тубулина состоит из трех новых белков (TTHERM_00550910, TTHERM_01001250 и TTHERM_01001260), которые попадают в кладу с тубулином P. tetraurelia iota. . Два из них (TTHERM_01001250 и TTHERM_01001260) тесно связаны друг с другом (рис. 7) и тесно связаны в геноме и, таким образом, вероятно, возникли в результате недавней тандемной дупликации.Функции этих генов неизвестны, но поскольку они пока уникальны для инфузорий, они могут отвечать за функции микротрубочек, специфичные для этого типа.
Динеины.
Динеины, которые были впервые обнаружены в Tetrahymena [140], представляют собой молекулярные моторы, которые перемещаются по трекам микротрубочек, процесс, критический для многих активностей T. thermophila, включая биение ресничек, кариокинез, деление MAC, корковую организацию и фагоцитоз. Многие из этих действий имеют решающее значение для восприятия изменений в окружающей среде и реагирования на них.Каждый динеиновый комплекс состоит из одной, двух или трех тяжелых цепей (содержащих двигательную активность) и конкретных комбинаций более мелких субъединиц, включая промежуточные, легкие промежуточные и легкие цепи, которые регулируют двигательную активность и связывание динеина с его молекулярным грузом. [141–143]. У организмов с ресничками или жгутиками существует множество изоформ динеинов, включая динеины аксонемного внешнего плеча, аксонемные динеины внутреннего плеча и неаксонемные или «цитоплазматические» динеины. Каждый из них специализируется на своем внутриклеточном расположении и выполняемых клеточных задачах [144].
Всего мы идентифицировали 21 легкую цепь, пять промежуточных цепей, две легкие промежуточные цепи и 25 тяжелых цепей (Таблица S13). Экспрессию каждого гена, а также структур экзон / интрон большинства подтверждали с помощью ОТ-ПЦР и, при необходимости, секвенирования продукта ОТ-ПЦР. По большей части, семейства субъединиц динеина T. thermophila похожи на семейства других модельных организмов; однако есть некоторые интересные отличия. Легкие цепи LC3A и 3B T. thermophila наиболее сходны с LC3 и LC5 зеленой водоросли Chlamydomonas reinhardtii [145].Эти белки принадлежат к большему семейству белков, связанных с тиоредоксином, и без биохимических данных, идентифицирующих один или оба белка как часть динеинового комплекса, может быть преждевременно маркировать их как компоненты динеина. Легкая цепь LC4 принадлежит к семейству белков, связанных с кальмодулином, и может регулировать кальций-зависимое обращение ресничек. T. thermophila экспрессирует два гена LC4, возможно, обеспечивая альтернативные или дополнительные способы контроля подвижности ресничек по сравнению с видами, которые экспрессируют только один.В других системах LC8 ассоциирован с несколькими разными динеиновыми и нединеиновыми комплексами, а T. thermophila экспрессирует один канонический LC8, а также пять дивергентных LC8-подобных генов с неизвестными функциями.
Возможно, наиболее интересным открытием является то, что T. thermophila экспрессирует 25 тяжелых цепей динеина. К ним относятся 14 ранее описанных генов DYH [146,147] и 11 новых, все из которых, по-видимому, являются аксонемными. Сложность семейства DYH может представлять механизм, с помощью которого организм может точно настраивать активность ресничек, производить специализированные реснички (например,g., оральные и задние реснички) и / или быстро генерируют большое количество новых ресничек. Наряду с этим произошла экспансия и других моторных белков. Например, имеется 78 кинезинов, больше, чем в любом другом секвенированном организме ([101] и Таблица 8). Кроме того, хотя миозинов меньше, чем у людей (13 против 22), 12 из 13 генов T. thermophila составляют единственный новый класс миозинов, не обнаруженный у других организмов [102,148].
Регуляция микротрубочек и процессов, связанных с микротрубочками.
Среди расширенных генов T. thermophila есть множество, участвующих в регуляции микротрубочек или связанных с микротрубочками процессов. Одним из примеров является тубулин-тирозин-лигаза-подобные доменные белки, несколько членов которых были идентифицированы как ферменты, ответственные за полиглутамилирование α- или β-тубулина [149]. T. thermophila кодирует 50 белков, подобных тирозинлигазе тубулина, по сравнению с 14 белками у человека. Другим примером является семейство протеинкиназ NRK, которое, как упоминалось выше, претерпело большое распространение в T.термофила. NRKs часто обнаруживаются связанными с микротрубочковыми органеллами [150], такими как центриоли, базальные тельца и жгутики, и играют множество ролей, включая регуляцию созревания центросом [151] и иссечение жгутиков [152]. Мы идентифицировали 39 NRK в T. thermophila, что примерно в три раза превышает количество таких локусов у человека. Филогенетический и функциональный анализ подтвердил, что эта диверсификация адаптировала членов этого семейства к различным субклеточным локализациям и цитоскелетным ролям [103].Таким образом, такие генные экспансии могут позволить изоформам белка с различной направленностью регулировать функцию одного и того же типа органелл в разных местах или создавать разные свойства одних и тех же структурных строительных материалов (например, микротрубочек), которые используются в качестве каркасов для построения различных типов органелл. .
Секреторные пути и мембранный трафик.
Помимо обычных органелл, T. thermophila поддерживает несколько более специализированных мембраносвязанных компартментов, включая альвеолы (общие с другими альвеолами), сократительную вакуоль (обнаруженную у многих протистов) и отдельные, функционально отличные макронуклеары и микроядра [128].Он также имеет несколько путей интернализации плазматической мембраны, а также конститутивного и регулируемого экзоцитоза [128, 153]. Сортировка и транспортировка компонентов мембраны являются критическими функциями для всех этих действий. Анализ генома выявляет гомологи многих ключевых белков, которые, как известно из других эукариот, участвуют в образовании и слиянии везикул, включая все основные классы белков оболочки (Таблица S14). Одно интересное открытие, сделанное анализом генома, заключается в том, что T. thermophila кодирует восемь белков, связанных с динамином, больше, чем большинство других секвенированных одноклеточных эукариот, и два из них, Drp1p и Drp2p, развили новую функцию в эндоцитозе [96] (A.Рахаман и А. П. Туркевиц, неопубликованные данные). Более того, филогенетический анализ показал, что привлечение динамина к роли в эндоцитозе происходит независимо путем конвергентной эволюции в клонах животных и инфузорий [96].
Диверсификация мембранного транспорта более очевидна в отношении Rab белков, которые представляют собой небольшие мономерные GTPases, которые регулируют процессы слияния и деления мембран. T. thermophila, с 69 Rab (Таблица S15), имеет большее количество линий, чем у людей (у которых их 60), чем у многих одноклеточных видов, таких как Saccharomyces cerevisiae, у которого 11 [154] и Trypanosoma brucei , , в котором 16 [155].На основании локализации и функциональных исследований, включая сравнение дрожжей и людей [156], Rab были разделены на восемь групп [157]. Филогенетический анализ (Рисунок S7) показывает, что T. thermophila кодирует представителей всех групп, кроме IV и VII, которые участвуют в позднем эндоцитозе и транспорте Гольджи, соответственно. Для группы VII это, по-видимому, отражает потерю, специфичную для клонов, поскольку геномы T. brucei и Entamoeba histolytica имеют несколько гомологов в этой группе.Два Rabs T. thermophila оказались гомологичными Rab28 и Rab32, которые не были отнесены ни к одной из этих групп; Ранее считалось, что Rab32 ограничен клонами млекопитающих. Группы II и V Rab, участвующие в эндоцитозе, особенно велики у T. thermophila и включают несколько гомологов Rab2, Rab4 и Rab11 в группе II. Это может отражать сложность поддержания по крайней мере двух основных путей интернализации мембраны. Кроме того, 29 Rab у T. thermophila не могут объединяться ни с одной из групп Rab, более широко встречающихся среди эукариот.В этой группе 20 кластеров в три клады, обозначенные Tetrahymena клады I, II и III на рисунке S7, которые могут представлять собой излучения, специфичные для инфузорий. Остальные девять очень расходятся и могут представлять очень древние события дублирования и / или изменения, связанные с наймом для новой функции. Поскольку однозначное выравнивание среди таких расходящихся Rabs затруднено, их отношения станут более ясными по мере секвенирования дополнительных родственных геномов.
Недавно большое количество Rab было обнаружено у множества амебоидных протистов, включая D.discoideum, E. histolytica [158] и парабазалид Trichomonas vaginalis [159]. Было высказано предположение, что диверсификация этих видов связана с их амебоидным образом жизни [159]. Однако наличие значительной диверсификации у T. thermophila предполагает, что различный образ жизни простейших может сопровождаться их собственным брендом значительной диверсификации Rab.
Выводы и планы на будущее
При секвенировании и сборке генома MAC T. thermophila ожидалось много серьезных проблем, которые обычно не наблюдаются в проектах генома эукариот.В целом, однако, сборки удивительно точны и представляют собой превосходный охват генома. Это, вероятно, во многом связано с низким уровнем повторяющейся ДНК, одной из особенностей генома MAC, которая изначально побудила нас выбрать его для секвенирования. Данные о последовательности в наших текущих собраниях, безусловно, достаточно полны для детального анализа предсказанной биологии этого вида, как мы сообщали здесь и другие показали. Кроме того, последовательность генома уже используется во многих функциональных геномных исследованиях с использованием имеющихся мощных экспериментальных инструментов.В этом отношении будет очень полезно провести сравнительный анализ с последовательностями генома других инфузорий, таких как P. tetraurelia и Oxytricha trifallax, , которые находятся в стадии разработки.
Одна из наших основных целей — получить полную последовательность генома MAC, и еще предстоит решить некоторые проблемы. Поскольку нам не удалось получить качественные данные о последовательностях из больших клонов-вставок, любая область генома MAC, содержащая значительные количества повторяющейся ДНК, не могла бы собраться должным образом.Чтобы преодолеть эту ловушку, мы теперь используем HAPPY mapping [162] в качестве альтернативного подхода к получению такой связывающей информации. Также известно, что по крайней мере концы по крайней мере двух MAC-хромосом, присутствующих сразу после конъюгации, исчезают во время последующего вегетативного роста, что может указывать на то, что эти хромосомы неспособны к долгосрочному поддержанию [41]. Как и ожидалось, мы не находим в нашей базе данных последовательности, соответствующие этим концам. Таким образом, потребуются альтернативные методы для получения последовательностей этих областей и любых других, потерянных во время раннего вегетативного роста.Несмотря на эти проблемы, все данные свидетельствуют о том, что можно будет закрыть весь геном MAC.
Конечно, один только геном MAC не дает нам полной картины генома T. thermophila. Секвенирование генома MIC будет сложнее из-за большего количества повторяющейся ДНК. Однако мы сможем использовать геном MAC в качестве основы, и, таким образом, в некотором смысле секвенирование MIC будет эквивалентно закрытию генома, а не независимому проекту.Мы уже начали в этой области с определения последовательности, смежной с соединениями MIC Cbs, и сопоставления их со сборками MAC, а также наоборот — с использованием последовательностей, смежных с теломерами MAC, для извлечения областей, фланкирующих MIC Cbs [24,41].
Наличие последовательности MIC и сопоставление MIC с MAC будет полезно для понимания многих аспектов биологии T. thermophila, которые мы не можем изучать с помощью MAC. К ним относятся функция центромеры, особенности теломер MIC и степень, в которой MAC и MIC у T.thermophila и другие инфузории являются эквивалентом соматических и половых клеток. Возможно, наиболее важным является то, что наличие обоих геномов позволит провести подробный анализ процесса перестройки ДНК в масштабе всего генома. Только имея обе последовательности генома, мы можем полностью понять биологию этого удивительного вида.
Структура генома зародышевой линии Tetrahymena thermophila и связь с массивно реаранжированным соматическим геномом
[…] В рукописи есть некоторые оставшиеся проблемы, которые необходимо решить, прежде чем она будет признана принятой, как указано ниже.В частности, все три рецензента и рецензент считают, что работа будет иметь наибольшее влияние, если она будет более читаемой для широкой аудитории eLife. Текст мог бы дать читателям немного больше рекомендаций относительно общих выводов их выводов, как подробно описано ниже. Кроме того, рукописи было бы полезно иметь сводную цифру, иллюстрирующую основные результаты: повторяющиеся богатые центромерные области, консервативные позиции CBS и не поддерживаемые хромосомы.
1) Большая часть информации из подразделов «Секвенирование и сборка генома MIC» и «Построение и анализ суперсборок MIC по длине хромосомы» может быть изложена в краткой форме в тексте, а представленные детали могут быть поместите в Методы или в дополнительный материал. Это поможет читать статью более гладко.
Как было предложено рецензентом, подробный материал в этом разделе был перемещен в раздел «Материалы и методы» и на новый рисунок 1 — приложение к рисунку 1.Мы также переместили детали секвенирования и сборки генома в «Материалы и методы», а сводку статистики сборки (бывшая таблица 1) — в дополнительный файл 1A.
2) Зашифрованная ДНК, предназначенная для Mac, описанная в подразделе «ДНК, предназначенная для зашифрованных макронуклеарных хромосом», представляет интерес, и авторы описывают механизмы как отличные от того, что есть в гипотрихах. Было бы полезно иметь цифру, чтобы следовать тому, что обнаружили авторы. Они заявляют, что это будет рассмотрено в другой рукописи, но если они собираются сообщить об этом хотя бы в некоторой степени здесь, цифра и немного больше деталей о том, что отличается, очень помогут.
Мы поняли, что сравнение с дескремблированием генов у Spirotrichs может быть преждевременным. Мы удалили отдельный заголовок этого раздела и переместили упрощенный абзац, без ссылки на дескремблирование, под обсуждение поломки хромосомы. Как следствие, мы считаем, что больше нет необходимости добавлять еще одну цифру.
3) Первые шесть абзацев подраздела «MIC Centromeres» можно сокращать и сокращать. Весь раздел особенно длинный и умозрительный, а местами повторяющийся.Понятие мейотического драйва вставлено в середину описания структуры и сравнения с другими организмами. Можно ли отредактировать раздел, чтобы подтянуть прозу, а может, и сократить?
Значительно сокращен раздел центромер MIC. Некоторая подробная информация была перенесена на новый рисунок 2 — дополнение к рисунку 1 и в дополнительный файл 1E.
4) Обсуждение запрограммированного удаления последовательностей Cen в шестом абзаце подраздела «Центромеры MIC» должно иметь цифру, позволяющую читателю проследить за структурами, о которых идет речь.
Обсуждение запрограммированного удаления последовательностей Cen было сокращено и упрощено; в результате мы не считаем, что для этого требуется дополнительная цифра.
5) В седьмом абзаце подраздела «МИК Центромеры» обсуждаются не поддерживаемые хромосомы, которые обсуждаются позже. Это не относится к разделу «MIC Centromeres», где оно находится в настоящее время.
Обсуждение необслуживаемых хромосом перенесено в более поздний, более подходящий раздел рукописи.
6) В девятом абзаце подраздела «Центромеры MIC» обсуждение транскриптов из микрофонных последовательностей на самом деле представляет собой краткое изложение литературы и гипотезу и должно проводиться в отдельном обсуждении, а не в середине раздела, посвященного центромерам. .
В ответ на этот пункт и пункт 14 эта спекулятивная гипотеза была опровергнута.
7) Обсуждение Cbs в подразделах «Семейство последовательностей хромосомных разрывов (Cbs)», «Набор из 225 Cbs в геноме MIC» и «Вырождение мотива Cbs среди функциональных Cbs» должно быть значительно сжатым.Консенсусная последовательность уже была известна, поэтому авторам следует тратить на это меньше времени и просто начинать новую информацию, которую предлагают дополнительные последовательности.
В соответствии с рекомендациями, этот раздел был значительно сокращен.
8) Аналогичным образом обсуждение сохранения CBS у разных видов в подразделах «Сохранение участков разрыва хромосом у видов Tetrahymena» и «Дублирование областей Cbs на эволюционной шкале времени» должно быть значительно сокращено и объединено с идентификацией Сайты CSB обсуждались выше.
По предложению рецензента мы значительно сократили разделы, переместив детали в Материалы и методы и дополнительные файлы. Однако мы считаем, что объединение идентификации и характеристики всех 225 Cbs в T. thermophila с обсуждением сохранения Cbs у разных видов было бы неуместным. Одним из основных результатов предыдущего анализа является полная картина диапазона изменчивости последовательности Cbs; небольшая выборка Cbs, идентифицированных у других видов, попадает в этот диапазон, как мы указываем в этом разделе.Однако наиболее интересным выводом из последнего анализа является сохранение местоположений Cbs, что приводит к сохранению размера и содержания генов MAC-хромосом. Этот вывод полностью независим от анализа сохранения последовательности Cbs и был бы невозможен без межвидового сравнения в этом разделе.
9) Подраздел «Не поддерживаемые хромосомы MAC» можно объединить с обсуждением фенотипических наборов в десятом абзаце подраздела «Центромеры MIC», и этот раздел следует сократить.
В целях сокращения рукописи обсуждение фенотипического ассортимента, на которое ссылается рецензент, было перенесено в легенду Дополнительного файла 1E. В любом случае мы считаем, что обсуждение необслуживаемых хромосом стоит само по себе. Как и предполагалось, он был сокращен.
10) Резюме и заключение не потребовались бы, если бы авторы заявили результаты, а затем написали краткое обсуждение.
Чтобы избежать повторения результатов (и удлинения рукописи), которое было бы необходимо, если бы был включен отдельный раздел для обсуждения, мы включили краткие обсуждения, которые вытекают непосредственно из результатов по всей рукописи.Мы значительно сократили и переориентировали исходный раздел «Резюме и выводы», который теперь называется «Выводы и будущие направления».
11) На рисунке 2 кажется, что было бы полезно, если бы авторы обозначили область, которая, по их мнению, содержит центромеру каждой хромосомы.
Запрошенное изменение внесено в рисунок.
12) В четвертом абзаце подраздела «Дублирование областей Cbs на эволюционной шкале времени» требуется точка после «взаимосвязи».
Исправление сделано.
13) В четвертом абзаце подраздела «Дублирование областей Cbs на эволюционной шкале времени». Идея о том, что CBS вводится инвазией транспозонов, приводящей к использованию этой последовательности в качестве сайта фрагментации хромосомы, является убедительной, но способ ее описания не является обязательным. Это не было бы «связано» с инвазией транспозонов, это было бы результатом инвазии и последующего одомашнивания фермента разрушения.Это один из примеров, связанных с моим общим замечанием о том, что рукопись может быть написана более кратко и ясно.
«Связанный с» был изменен на «возникший из», как это было предложено.
14) Описание возможной роли RNAi и Dicer-like 1 в функции центромеры должно быть уменьшено. В дополнение к Mochizuki, K et al. (2004) в статье процитирована роль Dcl1 в митозе и мейозе, во втором исследовании Malone, CD et al. (2005) исследовали фенотипы потери функции Dcl1 и не обнаружили доказательств какой-либо роли Dcl1 в сегрегации хромосом.Различия между результатами двух исследований никогда не согласовывались, поэтому использование только одного исследования в качестве вспомогательной информации для Обсуждения может ввести в заблуждение менее информированного читателя.
Это исправление было сделано. См. Пункт 6 выше.
15) Эта рукопись включает данные из двух других рукописей. А именно, аннотация мобильного элемента (TE) (Kapusta et al.) И отображение делеций (Cassidy-Hanley et al.), Данные, используемые для хромосомного ландшафта (рисунок 2) и для определения центромерных областей 5 хромосом зародышевой линии (на рисунке 2 и таблица 2).Документ TE, кажется, «отправлен», тогда как отображение удаления находится «в стадии подготовки». Я предлагаю включить в эту рукопись схему удаления. Поскольку эта рукопись несколько многословна и спекулятивна, будет нетрудно сжать текст (сократить рукопись), чтобы освободить место для данных удаления, которые на самом деле уже присутствуют, за исключением, возможно, дополнительной таблицы и / или рисунка, и параграф в Материалы и методы. Это не должно препятствовать более подробной второй статье по этому вопросу.Что касается аннотации к транспонируемым элементам, я надеюсь, что ее можно будет опубликовать вместе с этой рукописью. Было бы даже лучше (но, вероятно, невозможно) представить хотя бы основные результаты в настоящей рукописи, чтобы получить исчерпывающий обзор хромосом зародышевой линии Tetrahymena, включая не только CBS, но и аннотации TE.
В этом пункте сделаны два предложения. Публикация нашего анализа последовательностей Tetrahymena IES рядом с текущей рукописью, как было предложено рецензентом, была нашей первоначальной целью.Редакция eLife решила не полностью рецензировать рукопись IES в том виде, в каком она была первоначально представлена, но предложила вариант переноса «ключевых результатов» в первую рукопись. Первоначально мы решили продвинуться вперед с полным представлением первой рукописи, но после прочтения обзоров и консультаций с рецензирующим редактором Кэтлин Коллинз мы решили воспользоваться первоначальным советом и перенести ключевые результаты анализа IES во всеобъемлющий (но существенно сжатый и тщательно реорганизованная) рукопись о структуре и реорганизации генома.Второе предложение заключалось в том, чтобы включить в эту рукопись данные о картировании хромосомных делеций. Исследование делеционного картирования было многолетним усилием под руководством доктора Донны Кэссиди-Хэнли и с участием многих студентов-исследователей. Лишь небольшая часть идентифицированных делеций была информативной относительно местоположения центромер. Мы обеспокоены тем, что полная публикация методов и частичных результатов этих усилий может поставить под угрозу способность доктора Кэссиди-Хэнли и его коллег опубликовать полную историю в высококачественном журнале и получить то внимание и освещение, которых заслуживает эта история.Мы не считаем, что отсутствие этих подробных результатов умаляет нашу рукопись. Однако мы включили новый рисунок 2 — добавление к рисунку 1, показывающий, как набор делеций использовался для разграничения центромеры области хромосомы 5. Остальные центромеры были разграничены аналогичным образом. Мы считаем, что эта цифра проясняет, как был проведен анализ делеции, без ущерба для будущей публикации всего исследования.
16) Подраздел «ДНК, предназначенная для зашифрованных макронуклеарных хромосом» является слабым.Слово «скремблированный» вводит в заблуждение, поскольку предлагаемый механизм (HR между повторами) отличается от того, что Oxytricha использует материнскую направляющую РНК для дескремблирования MDS. Сравнение с соединением V (D) J вводит в заблуждение, поскольку этот процесс включает путь негомологичного соединения концов (NHEJ), а не HR. Наконец, это основано на ДНК MAC от одного карионида с последующим выделением одного полностью отобранного вегетативного клона, не так ли? Как можно подтвердить такой редкий процесс, не глядя на ДНК по независимо преобразованным и сортированным линиям?
См. Наш ответ на пункт №2.
17) Как идентифицируются IES? Они просто оперативно определены как области в сборке MIC, которые удаляются в сборке MAC? Есть ли другие критерии? В пятом абзаце подраздела «Центромеры MIC» упоминается, что «мы охарактеризовали тысячи IES, специфичных для MIC». Что такое характеристика, просто идентификация на основе выравнивания последовательностей? Или это ссылка на данные отдельной рукописи TE? В «Материалы и методы» ничего не нашел.
Теперь эта информация включена. Смотрите наш ответ на пункт 15.
18) Чтобы избежать путаницы, я думаю, следует уточнить, что IES в Paramecium — это многочисленные короткие, уникальные копии некодирующих последовательностей (во многих случаях остатки транспозонов), в то время как IES в Tetrahymena являются (или включают) переносимые элементы и другие повторяющиеся последовательности (как четко указано в подразделе «Центромеры MIC»). Это следует прояснить во втором абзаце Введения (и почему 40 000 вместо опубликованных 45 000 IES Paramecium?).
Предлагаемые исправления внесены.
19) Поскольку теломерные повторы MIC и MAC различаются, и известно точное количество хромосом MIC и MAC, может ли численность теломерных повторов (и количество Cbs?) В считывании позволить оценить контаминацию MAC (подраздел «Центромеры MIC») , последний абзац)? Если теломерные повторы MIC и ранее охарактеризованные субтеломерные области могут быть идентифицированы в парах считывания (как в библиотеках коротких вставок, так и в библиотеках прыжков), могут ли они быть сопоставлены с каркасами MIC (подраздел «Построение и анализ суперсборок MIC с длиной хромосомы» ”, Последний абзац)? При наличии всего 10 концов хромосом MIC (но 169-кратного охвата) это может быть сложно.
Мы ценим предложение рецензента оценить степень загрязнения MAC путем сравнения повторов теломер MAC и MIC и попытаться идентифицировать и связать теломеры MIC с другими каркасами. Мы искали теломерные повторы MIC в сборках и чтениях, но, к сожалению, нашли мало примеров, и они малоинформативны. Мы добавили текст в соответствующий раздел (Материалы и методы в ответ на пункт №1), разъясняющий этот вопрос. В любом случае, мы не думаем, что оценка загрязнения ПДК может значительно улучшить или изменить наши выводы.Мы четко признаем, что существует некоторое загрязнение, которое неизбежно с учетом экспериментальных ограничений, но, как указано в первом абзаце материалов и методов: «Путем микроскопического подсчета очищенных ядер (с учетом относительной ядерной плоидности) мы оцениваем загрязнение макронуклеарная геномная ДНК составляла менее 2% ». Мы уверены, что это загрязнение не оказало существенного влияния на сборку генома MIC каким-либо образом, что могло бы повлиять на выводы этой статьи. Следовательно, текст, идентифицированный рецензентом, был изменен, чтобы объяснить плохую сборку теломерных областей как результат их повторяющейся природы, как это наблюдается в сборках генома многих других организмов.
20) В первом предложении подраздела «Секвенирование и сборка генома MIC», «очень богатый AT»: можете ли вы указать значения для сборок MAC и MIC (77,7% и 76,3%) или среднее (~ 77%) в скобках?
Эти значения (сборка MAC 77,7% и сборка MIC 77,9%) были добавлены в соответствующий раздел материалов и методов.
21) В последнем предложении подраздела «Секвенирование и сборка генома MIC» допущена опечатка («как будет описано»).Что еще более важно, это предложение не совсем оправдано, так как только небольшая часть последовательностей, ограниченных зародышевой линией Paramecium, была аннотирована и опубликована на сегодняшний день. Я не понимаю, как можно провести глобальное сравнение, к тому же оно сделано в отдельном документе TE. То, что эта организация отличается, было известно давно из молекулярных исследований перестройки генома.
Данный текст удален.
22) В абзаце шестом подразделе «MIC Centromeres».Предположение о том, что хромосомы Paramecium MAC являются плечами хромосом MIC, было сильно спекулятивным, когда оно было опубликовано в 2008 году, и основывалось на косвенных доказательствах, а именно на обратной корреляции между содержанием G + C и длиной хромосомы MAC. Было высказано предположение, что эта обратная корреляция может быть объяснена смещенной конверсией генов, что указывает на то, что хромосомы MAC могут быть пропорциональны по размеру плечам хромосом MIC (поскольку хромосомы MIC подвергаются мейотической рекомбинации). Совсем недавно была опубликована работа по центромерам Paramecium, так что было бы разумно удалить спекулятивное утверждение и сослаться только на работу по центромерам.Более того, я думаю, что еще слишком рано говорить о фундаментальных различиях в архитектуре хромосом зародышевой линии, по крайней мере, в том, что касается центромер, поскольку может оказаться, что центромеры теряются в ходе обычного процесса элиминации IES каждого организма.
Мы изменили текст, чтобы удалить спекулятивное утверждение относительно Paramecium центромер, а также утверждение о «фундаментальном различии» между центромерами двух видов и включить ссылку на недавно опубликованное исследование, как было предложено рецензентом.
23) Рисунок 3. Логотип последовательности. В логотипе Cbs не указана частота каждой базы, как указано в легенде. Логотип представлен в виде битов информации. Однако это было неправильно рассчитано. Никакие A или T не могут нести 2 бита информации в геноме, который превышает 75% A + T. Логотип следует рассчитывать с учетом частоты каждой основы в геноме, а не предполагать равные частоты. Это заставит C выделяться намного больше. При использовании seqlogo доступен просмотр «базовой частоты», но результат не такой приятный.
Благодарим рецензента за обнаружение ошибки в легенде рисунка; это было исправлено, чтобы идентифицировать ось Y как биты, то есть как единицы сохранения, а не единицы частоты нуклеотидов. Что касается предложения поднять базовый уровень до уровня общегеномной сохранности, что предлагает рецензент, в этом предложении есть определенная ценность. Но общегеномные частоты (и связанные с ними биты сохранения) включают частоты кодирующих последовательностей, которые имеют содержание G + C выше среднего.Сохранение истинного уровня сохранения некодирующей Cbs-смежной последовательности обеспечивает значимый внутренний контроль. Мы предпочитаем сохранять представление измерений сохранения без каких-либо предположений, что, например, не дает искаженного представления о разнице в сохранении между T в положениях Cbs 1 и 15.
24) В четвертом абзаце подраздела «Сохранение участков хромосомных разрывов у видов Tetrahymena». Я предполагаю, что термин «центрические хромосомы» относится к хромосомам MAC, которые происходят из центральных областей хромосом MIC.По тексту было бы легче следовать, если бы они были названы центрическими MAC-хромосомами. В конце того же абзаца «развязка» правильная или должна быть «развязка»?
После пересмотра термин «центрические хромосомы» больше не появляется в основном тексте, но он был заменен на «центрические хромосомы MAC» в легенде к Таблице 1. В легенде к дополнительному файлу 1E мы также теперь ссылаемся на « Центрические хромосомы MAC, происходящие из центромерных областей хромосом MIC «.Предлагаемое изменение с «разъединения» на «разъединение» было сделано. Это предложение теперь появляется в дополнительном файле 2C.
25) Ссылка Lhuillier-Akakpo et al., 2015 является неполной.
Ссылка исправлена.
https://doi.org/10.7554/eLife.19090.045Cell | Encyclopedia.com
Клетка — это мельчайший живой компонент организмов и основная единица жизни. В многоклеточных живых существах совокупность клеток, которые работают вместе для выполнения аналогичных функций, называется тканью ; различные ткани, выполняющие согласованные функции, образуют органы; и органы, которые работают вместе для выполнения общих процессов, образуют системы организма.Пищеварительная система человека , например, состоит из различных органов, включая желудок, поджелудочную железу и кишечник. Ткань, выстилающая кишечник, называется эпителиальной тканью. Эпителиальная ткань, в свою очередь, состоит из особых клеток, называемых эпителиальными клетками. В тонком кишечнике эти эпителиальные клетки специализируются на их абсорбционной функции: каждая эпителиальная клетка покрыта тысячами небольших выступов, называемых микроворсинками. Многочисленные микроворсинки значительно увеличивают площадь поверхности тонкой кишки, через которую питательных веществ могут всасываться в кровоток.
Типы клеток
Многоклеточные организмы содержат огромное количество высокоспециализированных клеток. Растения содержат клетки корня, клеток листа, клеток и стволовых клеток, . У людей есть клетки кожи, нервные клетки и половые клетки. Каждый вид ячейки структурирован для выполнения узкоспециализированной функции. Часто изучение структуры клетки позволяет многое узнать о ее функциях в организме . Например, как мы уже видели, эпителиальные клетки тонкой кишки специализируются на абсорбции из-за многочисленных микроворсинок, которые заполняют их поверхность.Нервные клетки или нейроны — это еще один вид специализированных клеток, форма которых отражает функцию. Нервные клетки состоят из тела клетки и длинных отростков, называемых аксонами, которые проводят нервные импульсы. Дендриты — это более короткие отростки, принимающие нервные импульсы.
Сенсорные клетки — клетки, которые обнаруживают сенсорную информацию из внешней среды и передают эту информацию в мозг — часто имеют необычные формы и структуры, которые способствуют их функции. Клетки-палочки в сетчатке глаза , например, не похожи ни на одну другую клетку в человеческом теле.Эти клетки имеют форму стержня и имеют светочувствительную область, содержащую множество мембранных дисков. В каждый диск встроен специальный светочувствительный пигмент, который улавливает световых лучей . Когда пигмент получает свет из внешней среды, нервные клетки глаза срабатывают, чтобы посылать нервный импульс в мозг . Таким образом, люди могут обнаруживать свет.
Клетки, однако, также могут существовать как одноклеточные организмы. Например, организмы, называемые протистами, являются одноклеточными организмами.Примеры простейших включают микроскопический организм под названием Paramecium и одноклеточную водоросль под названием Chlamydomonas.
Прокариоты и эукариоты
В живых существах различают два типа клеток. Прокариоты (буквально «перед ядром») — это клетки, не имеющие отдельного ядра. Большинство прокариотических организмов одноклеточные, например бактерий и водорослей . С другой стороны, эукариотические (буквально «истинное ядро») организмы имеют отчетливое ядро и высокоорганизованную внутреннюю структуру.Внутри эукариот присутствуют отчетливые органеллы, небольшие структуры, каждая из которых выполняет определенный набор функций. Эти органеллы связаны мембранами. Прокариоты, помимо отсутствия ядра, также лишены этих мембраносвязанных органелл.
Размер и количество клеток
По оценкам, тело взрослого человека содержит около 60 триллионов клеток. Большинство этих клеток, за некоторыми исключениями, настолько малы, что для их наблюдения необходим микроскоп . Небольшой размер клеток выполняет определенную функцию в функционировании организма.Если бы клетки были больше, многие процессы, выполняемые клетками, не могли бы происходить эффективно. Чтобы представить себе эту концепцию, подумайте об эпителиальных клетках кишечника, которые обсуждались ранее. Что, если бы эпителий кишечника состоял из одной большой клетки, а не из тысяч маленьких клеток? Большая ячейка имеет большой объем или содержимое. Площадь поверхности или мембраны этой большой клетки является местом, через которое питательные вещества попадают в тонкий кишечник для доставки в кровоток.Поскольку объем этой большой ячейки настолько велик, площадь поверхности для сравнения относительно мала. Следовательно, большие ячейки имеют отношение площади поверхности к объему . Только определенное количество питательных веществ может пройти через ограниченную площадь мембраны этой большой клетки. При небольшом соотношении площади поверхности к объему количество веществ, попадающих в ячейку и выходящих из нее, строго ограничено.
Однако, если эпителий кишечника разделен на тысячи более мелких клеток, объем остается прежним, но площадь поверхности — количество клеточных мембран — значительно увеличивается.Через клетки кишечного эпителия может проходить гораздо больше питательных веществ. Поэтому маленькие клетки имеют большое отношение площади поверхности к объему. Большое отношение площади поверхности к объему мелких клеток делает перенос веществ в клетки и из них чрезвычайно эффективным.
Другая причина небольшого размера клеток заключается в том, что управлять клеточными процессами легче в маленькой клетке, чем в большой. Клетки — это динамичные живые существа. Клетки переносят вещества из одного места в другое, воспроизводятся и вырабатывают различные ферменты и химические вещества для экспорта во внеклеточную среду.Все эти действия выполняются под руководством ядра, центра управления клеткой. Если бы ядру пришлось контролировать большую клетку, то это направление могло бы нарушиться. Вещества, транспортируемые из одного места в другое, должны будут преодолевать большие расстояния, чтобы добраться до места назначения; воспроизводство большой клетки было бы чрезвычайно сложным делом; и продукция на экспорт будет производиться не так эффективно. Ячейки меньшего размера из-за их более управляемого размера контролируются гораздо эффективнее, чем ячейки большего размера.
Структура и функции клеток
Основная структура всех клеток, будь то прокариот, и эукариот, одинакова. Все клетки имеют плазменную мембрану , через которую вещества проходят внутрь и из клетки. За исключением нескольких незначительных отличий, плазматические мембраны у прокариот и эукариот одинаковы. Внутренняя часть обоих типов клеток называется цитоплазмой. В цитоплазму эукариот встроены клеточные органеллы; цитоплазма прокариот не содержит органелл.Наконец, оба типа клеток содержат небольшие структуры, называемые рибосомами, , которые участвуют в синтезе белка. Состоящие из двух белковых субъединиц рибосомы не ограничены мембранами; следовательно, они не считаются органеллами. У эукариот рибосомы либо связаны с органеллами, эндоплазматическим ретикулумом, либо существуют как «свободные» рибосомы в цитоплазме. Прокариоты содержат только свободные рибосомы.
Строение прокариот
Примером типичного прокариота является бактериальная клетка.Бактериальные клетки могут иметь форму стержней, сфер или штопоров. Все прокариоты ограничены плазматической мембраной. Над этой плазматической мембраной находится клеточная стенка, а у некоторых бактерий капсула, состоящая из желеобразного материала, покрывает клеточную стенку. Многие бактерии, вызывающие болезни у животных, имеют капсулы. Капсула обеспечивает дополнительный уровень защиты для бактерий, и часто патогенные бактерии с капсулами вызывают гораздо более серьезное заболевание , чем бактерии без капсул.
В цитоплазме прокариот находится нуклеоид — область, в которой находится генетический материал (ДНК).Этот нуклеоид не является настоящим ядром, потому что он не ограничен мембраной. Также в цитоплазме находятся многочисленные рибосомы. Эти рибосомы не прикреплены к какой-либо структуре и поэтому называются «свободными» рибосомами.
К клеточной стенке некоторых бактерий прикреплены жгутики , хлыстоподобные структуры, обеспечивающие движение. У некоторых бактерий также есть пили, которые представляют собой короткие выступы в виде пальцев, которые помогают бактериям прикрепляться к тканям. Бактерии не могут вызвать заболевание, если они не могут прикрепиться к тканям.Бактерии, вызывающие пневмонию, , например, прикрепляются к тканям легкого. Бактериальные пили значительно облегчают это прикрепление к тканям, и поэтому, как и капсулы, бактерии с пилями часто более вирулентны, чем бактерии без них.
Структура эукариот
Органеллы, обнаруженные у эукариот, включают мембранную систему, состоящую из плазматической мембраны, эндоплазматического ретикулума, тельца Гольджи и везикул; ядро; цитоскелет; и митохондрии. Кроме того, клеток растений имеют особые органеллы, которых нет в клетках животных.Эти органеллы представляют собой хлоропласты, клеточную стенку и вакуоли.
Мембранная система
Мембранная система клетки выполняет множество важных функций. Эта система контролирует вход и выход веществ в ячейку и из нее, а также обеспечивает производство и упаковку веществ. внутри клетки. Мембранная система клетки состоит из плазматической мембраны, в которой заключено содержимое клетки; эндоплазматический ретикулум, который производит липиды и белков ; тело Гольджи, которое упаковывает вещества, производимые внутри клетки; и различные везикулы, выполняющие разные функции.
Плазматическая мембрана
Плазматическая мембрана клетки часто описывается как «избирательно проницаемая»; то есть плазматическая мембрана устроена так, что только определенные вещества могут пересекать ее границы. Плазматическая мембрана состоит из двух слоев молекул, называемых фосфолипидами. Каждая молекула фосфолипида состоит из фосфатной «головы» и двух цепочек жирных кислот, которые свисают с головы.
Ориентация этих двух участков молекулы фосфолипида имеет решающее значение для функции плазматической мембраны.Фосфатная область является гидрофильной (буквально «водолюбивой») и привлекает воды . Область жирных кислот является гидрофобной (буквально «ненавидящей воду») и отталкивает воду. В фосфолипидном бислое плазматической мембраны слои фосфолипидов расположены так, что две фосфатные гидрофильные области обращены наружу, к водянистой внеклеточной среде, и внутрь, к клеточной цитоплазме, которая также содержит воду. Две части цепей из гидрофобных жирных кислот обращены друг к другу, образуя водонепроницаемую защиту.Таким образом, плазматическая мембрана является водонепроницаемой и притягивающей воду. Он функционирует как граница между содержимым клетки и внешней клеточной средой, но также позволяет переносить воду и другие вещества через свои границы.
В плазматические мембраны эукариот встроены различные белки. Эти белки выполняют несколько различных функций в клетке. Некоторые белки представляют собой насосы или каналы для импорта и экспорта веществ. Другие белки, называемые антигенами, служат маркерами идентификации клетки.Другие белки помогают клетке формировать связи с другими клетками. Поскольку эти мембранные белки часто выступают из клеточной мембраны во внеклеточную среду, они также имеют гидрофобные и гидрофильные области. Части белков, которые встроены в плазматическую мембрану, являются гидрофобными, а части белков, которые выходят наружу во внеклеточную среду, являются гидрофильными.
Ученые, изучающие плазматические мембраны, используют термин «жидко-мозаичная модель» для описания структуры плазматических мембран.«Мозаичная» часть модели описывает способ встраивания белков в плазматическую мембрану. «Жидкая» часть модели объясняет жидкую природу плазматических мембран. Вместо того, чтобы фиксироваться в одном месте внутри плазматической мембраны, эксперименты показали, что фосфолипиды демонстрируют некоторое движение внутри плазматических мембран, иногда перемещаясь вбок, иногда (хотя и редко) перескакивая с одного фосфолипидного слоя на другой. Мембранные белки также перемещаются внутри плазматической мембраны, хотя и медленнее, чем фосфолипиды.
Эндоплазматический ретикулум
Эндоплазматический ретикулум (что означает «внутри цитоплазмы» и «сеть») состоит из уплощенных листов, мешочков и трубок мембраны, которые покрывают все пространство цитоплазмы эукариотической клетки. Эта внутренняя мембранная система является продолжением двойной мембраны, окружающей ядро клетки. Следовательно, закодированные инструкции, которые ядро отправляет для синтеза белков, поступают непосредственно в эндоплазматический ретикулум. Внутри клетки эндоплазматический ретикулум синтезирует липиды и белки.Белки, которые синтезирует эндоплазматический ретикулум, такие как ферменты, экспортируются из клетки для выполнения различных функций в организме. Белки, которые вырабатываются в клетке для использования ею, например, в качестве каналов в плазматической мембране, производятся свободными рибосомами, которые усеивают цитоплазму.
В эукариотической клетке обнаружены два типа эндоплазматического ретикулума. Грубая эндоплазматическая сеть усеяна рибосомами на внешней стороне. Эти рибосомы являются участками синтеза белка.Как только белок синтезируется на рибосоме, он оказывается заключенным в везикулу, небольшой мембранно-связанный «пузырек». Везикула перемещается в другую органеллу, тело Гольджи. Внутри тела Гольджи белки везикулы дополнительно модифицируются, прежде чем они будут экспортированы из клетки. Клетки, которые специализируются на секреции белка, содержат большое количество грубого эндоплазматического ретикулума. Например, клетки поджелудочной железы, которые продуцируют белок инсулин , имеют обильный грубый эндоплазматический ретикулум.Плазматические клетки, белые клетки крови , которые секретируют иммунные белки, называемые антителами, настолько переполнены грубым эндоплазматическим ретикулумом, что трудно различить другие органеллы в цитоплазме.
Другой тип эндоплазматической сети — гладкая эндоплазматическая сеть. Гладкая эндоплазматическая сеть не имеет рибосом и является участком липидного обмена . Здесь макромолекулы, содержащие липиды, разбиваются на составные части. Кроме того, гладкая эндоплазматическая сеть участвует в синтезе липидсодержащих макромолекул.Гладкая эндоплазматическая сеть встречается в клетках не так часто, как грубая эндоплазматическая сеть. Большое количество гладкой эндоплазматической сети обнаруживается в клетках, которые специализируются на липидном обмене. Например, клетки печени удаляют алкоголя, и лекарства из кровотока. Клетки печени имеют впечатляющую сеть гладкой эндоплазматической сети. Точно так же клетки яичников и семенников, которые производят липидсодержащие гормонов, эстроген и тестостерон, содержат большое количество гладкой эндоплазматической сети.
Тело Гольджи
Тело Гольджи, названное в честь своего первооткрывателя, итальянского ученого XIX века Камилло Гольджи, является одной из органелл самой необычной формы. Тело Гольджи, чем-то напоминающее стопку блинов, состоит из уложенных друг на друга, ограниченных мембранами, сплющенных мешочков. Тело Гольджи окружают многочисленные маленькие пузырьки, связанные с мембраной. Тело Гольджи и его везикулы выполняют функцию сортировки, модификации и упаковки макромолекул, которые секретируются клеткой или используются внутри клетки для различных функций.
Тело Гольджи можно сравнить с отделом отправки и получения в большой компании. Каждое тело Гольджи в клетке имеет лицо цис , что аналогично принимающему отделу отдела. Здесь тело Гольджи получает макромолекулы, синтезированные в эндоплазматическом ретикулуме, заключенном в пузырьки. Лицо trans тела Гольджи аналогично отделению доставки отдела и является местом, откуда модифицированные и упакованные макромолекулы доставляются к месту назначения.
Внутри тела Гольджи к макромолекулам добавляются различные химические группы, чтобы гарантировать, что они достигнут своего предназначения. Таким образом, тело Гольджи прикрепляет «адрес» к каждой полученной макромолекуле. Например, клетки слизистой оболочки кишечника, называемые бокаловидными клетками, выделяют слизь. Белковый компонент слизистой оболочки, называемый муцином, модифицируется в теле Гольджи путем добавления углеводных групп. Из тельца Гольджи модифицированный муцин упаковывается в пузырьки.Везикула, содержащая слизистый груз, сливается с плазматической мембраной бокаловидной клетки и выходит во внеклеточную среду.
Везикулы
Везикулы представляют собой небольшие мембранные сферы, которые содержат различные макромолекулы. Некоторые везикулы, как мы видели, используются для транспортировки макромолекул из эндоплазматического ретикулума в тело Гольджи и из тела Гольджи в различные места назначения. Везикулы особого типа выполняют и другие функции. Лизосомы — это везикулы, которые содержат ферменты, участвующие в клеточном пищеварении.Некоторые протисты, например, поглощают другие клетки для еды. В процессе, называемом фагоцитозом, протист окружает частицу пищи и поглощает ее везикулами. Эта пища, содержащая везикулы, транспортируется в цитоплазме протиста до тех пор, пока не соприкоснется с лизосомой. Пищевой пузырек и лизосома сливаются, и ферменты внутри лизосомы высвобождаются в пищевой пузырек. Ферменты разбивают пищу на более мелкие части для использования простейшим.
Лизосомы, однако, обнаруживаются во всех типах клеток.Во всех клетках лизосомы переваривают старые изношенные органеллы. Они также играют роль в самоуничтожении старых клеток. Хотя ученые не понимают триггерный механизм этого самоуничтожения, клетки, которые не функционируют должным образом из-за старости, по-видимому, самоперевариваются с помощью лизосом. Смерть клеток также является компонентом нормальных процессов развития . Например, у человеческого зародыша руки и ноги похожи на паутину. По мере развития клетки, составляющие эти сети, медленно самоуничтожаются, освобождая пальцы.
Пероксисомы, как следует из их названия, содержат перекись водорода . Пероксисомы участвуют в окислении многих материалов, включая жиры. При окислении к молекуле добавляется кислорода . При добавлении кислорода к жирам образуется перекись водорода . Любой, кто лечил порез перекисью водорода, знает, что это вещество смертельно опасно для клеток. Следовательно, окисление жиров происходит внутри мембран пероксисом, так что вредное химическое вещество не просачивается в цитоплазму клетки.
Ядро
Ядро — это центр управления клеткой. Под микроскопом ядро выглядит как темное пятно с более темной областью, называемой ядрышком, в центре которой находится. Ядрышко — это место, где производятся субъединицы рибосом. Ядро окружает двойная мембрана, называемая ядерной оболочкой. Ядерная оболочка испещрена крошечными отверстиями, называемыми ядерными порами.
Ядро управляет всей клеточной деятельностью, контролируя синтез белков.Ядро содержит закодированные инструкции по синтезу белков в спиральной молекуле, называемой дезоксирибонуклеиновой кислотой (ДНК) . ДНК клетки упакована в ядре в структурной форме, называемой хроматином . Хроматин состоит из ДНК, плотно намотанной вокруг сферических белков, называемых гистонами. Когда клетка готовится к делению, ДНК раскручивается от гистонов и принимает форму хромосом, X-образные структуры, видимые внутри ядра до деления клетки .Хроматиновая упаковка ДНК позволяет всей ДНК клетки помещаться в объединенное пространство ядра. Если бы ДНК не была упакована в хроматин, она бы вылилась в пространство, примерно в 100 раз превышающее размер самой клетки.
Первый этап синтеза белка начинается в ядре. Внутри ядра ДНК транслируется в молекулу, называемую рибонуклеиновой кислотой-мессенджером (мРНК). мРНК затем покидает ядро через ядерные поры. Попадая в цитоплазму, мРНК прикрепляется к рибосомам (либо связанным с эндоплазматическим ретикулумом, либо свободным в цитоплазме) и инициирует синтез белка.Белки, предназначенные для экспорта из клетки, действуют как ферменты, которые участвуют во всех химических реакциях организма . Поскольку ферменты необходимы для всех химических процессов организма — от клеточного дыхания до пищеварения, — направление синтеза этих ферментов, по сути, контролирует всю деятельность организма. Следовательно, ядро, которое содержит инструкции по синтезу этих белков, направляет всю клеточную деятельность и, следовательно, все процессы в организме.
Цитоскелет
Цитоскелет — это «скелетный» каркас клетки.Однако вместо кости скелет клетки состоит из трех видов белковых нитей, которые образуют сети. Эти сети придают клетке форму и обеспечивают движение клеток. Три типа цитоскелетных волокон — это микротрубочки, актиновые филаменты и промежуточные филаменты.
Микротрубочки имеют диаметр 25 нанометров и состоят из белковых субъединиц, называемых тубулином. Каждая микротрубочка состоит из одиннадцати пар субъединиц тубулина, расположенных в виде кольца. В клетках животных микротрубочки возникают из области клетки, называемой центром организации микротрубочек (MTOC), расположенной рядом с ядром.Из этого центра микротрубочки расходятся веером через клетку, образуя сеть «треков», по которым перемещаются различные органеллы внутри клетки. Микротрубочки также образуют небольшие парные структуры, называемые центриолями внутри клеток животных. Эти структуры не считаются органеллами, потому что они не ограничены мембранами. Ученые когда-то думали, что центриоли образуют микротрубочки, которые разделяют клетку во время деления клетки; теперь известно, что каждая центриоль с парой раздвигается во время деления клетки и указывает план, по которому делится клетка.
Некоторые эукариотические клетки перемещаются с помощью микротрубочек, прикрепленных к внешней части плазматической мембраны. Эти микротрубочки называются жгутиками и ресничками. Жгутики и реснички имеют одинаковую структуру: кольцо из девяти триплетов тубулина, расположенных вокруг двух субъединиц тубулина. Разница между жгутиками и ресничками заключается в их движении и количестве. Жгутики прикреплены к клетке с помощью аппарата, похожего на «кривошип», который позволяет жгутикам вращаться. Обычно жгутиковые клетки имеют только один или два жгутика.Реснички же, с другой стороны, не прикрепляются с помощью «кривошипа» и колеблются взад и вперед, обеспечивая движение. Ресничные клетки обычно имеют сотни таких выступов, покрывающих их поверхность. Например, протист Paramecium перемещается с помощью одного жгутика, а протист Didinium покрыт многочисленными ресничками. Ресничные клетки также выполняют важные функции в организме человека. Дыхательные пути человека и других животных выстланы ресничными клетками, которые выносят мусор и бактерии вверх, из легких в горло.Там мусор либо кашляет из горла, либо попадает в пищеварительный тракт, где пищеварительные ферменты уничтожают вредные бактерии.
Нити актина имеют диаметр 8 нанометров и состоят из двух нитей белкового актина, намотанных друг на друга. Нити актина особенно заметны в мышечных клетках, где они обеспечивают сокращение мышечной ткани.
Промежуточные волокна имеют диаметр 10 нанометров и состоят из волокнистых белков. Из-за своей относительной силы они функционируют в основном для закрепления органелл в цитоплазме.
Митохондрии
Митохондрии — это электростанции клеток. Каждая митохондрия в форме сосиски покрыта внешней мембраной; внутренняя мембрана митохондрии сложена на отсеки, называемые кристами (что означает «ящик»). Матрица или внутреннее пространство, созданное кристами, содержит ферменты, необходимые для многих химических реакций, которые в конечном итоге превращают молекулы пищи в энергии .
Клетки содержат от сотен до тысяч митохондрий.Интересным аспектом митохондрий является то, что они содержат собственные последовательности ДНК, хотя и не в том количестве, которое содержится в ядре. Наличие этой отдельной ДНК, наряду с сходством митохондрий с одноклеточными прокариотами, привело к теории эволюции эукариот , названной эндосимбиотической теорией. Эта теория постулирует, что митохондрии когда-то были отдельными прокариотами, которые были поглощены другими прокариотами. Вместо того, чтобы перевариваться, митохондриальные прокариоты оставались внутри поглощающей клетки и выполняли свои функции высвобождения энергии.На протяжении миллионов лет эти симбиотические отношения способствовали эволюции эукариотической клетки.
Органеллы растений
Клетки растений имеют несколько органелл, которых нет в клетках животных. Это пластиды, вакуоли и клеточная стенка.
Пластиды
Пластиды — это органеллы везикулярного типа, которые у растений выполняют множество функций. Амилопасты хранят крахмал, а хромопласты хранят молекулы пигмента, которые придают некоторым растениям яркий оранжевый и желтый цвет.
Хлоропласты — это пластиды, которые осуществляют фотосинтез , процесс, в котором вода и углекислый газ превращаются в сахара.Внутри хлоропластов находится сложная мембранная система. Тилакоиды рассекают хлоропласты пополам, и к этим платформам прикреплены стопки перепончатых мешочков, называемых грана. В каждой грануле содержатся ферменты, необходимые для фотосинтеза. Мембранная система хлоропластов омывается жидкостью, называемой стромой, которая также содержит ферменты.
Подобно митохондриям, хлоропласты напоминают некоторых древних одноклеточных прокариот, а также содержат собственные последовательности ДНК. Считается, что их происхождение в эукариотах возникло в результате эндосимбиотических отношений между фотосинтетическим одноклеточным прокариотом, который был поглощен и остался внутри другой прокариотической клетки.
Вакуоли
Вакуоли растений — это большие пузырьки, связанные одной мембраной. Во многих клетках растений они занимают около 90% клеточного пространства. Они выполняют множество функций в клетке, включая хранение органических соединений, продуктов жизнедеятельности, пигментов и ядовитых соединений, а также функции пищеварения.
Клеточная стенка
Все клетки растений имеют клеточную стенку, которая покрывает плазматическую мембрану. Клеточная стенка растений состоит из жесткого углеводного вещества, называемого , целлюлоза , расположенного в матрице или сети других углеводов.Стенка клетки обеспечивает дополнительный уровень защиты между содержимым клетки и внешней средой. Например, хрустящее яблоко объясняется наличием этих клеточных стенок.
См. Также Клеточное дыхание; Хлоропласт; Хромосома; Фермент; Эукариоты; Жгутики; Ген; Мейоз; Митоз; Нейрон; Ядро, клеточное; Орган; Рибонуклеиновая кислота (РНК); Ткань.
Ресурсы
книги
Барритт, Грег Дж. Общение в клетках животных. Oxford: Oxford University Press, 1992.
Bittar, F. Edward, ed. Химия живой клетки. Гринвич, Коннектикут: JAI Press, 1992.
Брей, Деннис. Перемещения клеток. Нью-Йорк: Гарланд Пресс, 1992.
Кэрролл, Марк. Органеллы. Нью-Йорк: Гилфорд Пресс, 1989. Поверхность клетки. Plainview, NY: Cold Spring Harbor Laboratory Press, 1992.
периодические издания
Мэддокс, Джон. «Почему микротрубочки растут и сжимаются.» Nature 362 (18 марта 1993 г.): 201.
Панте, Нелли и Уели Эби.» Комплекс ядерных пор «. The Journal of Cell Biology 122 (сентябрь 1993 г.): 5-6.
Скотт, Дж. Д. и Т. Поусон. «Cell Communication: The Inside Story.» Scientific American 282 (июнь 2000 г.): 54-61.
Шей, Джерри В. и Вудринг Э. Райт ». Hayflick, Его предел и клеточное старение ». Nature Reviews / Molecular Cell Biology (1 октября 2000 г.): 72-76.
Kathleen Scogna
КЛЮЧЕВЫЕ ТЕРМИНЫ
- Актиновая нить
—Тип цитоскелетной нити, обладающей сократительными свойствами.
- Амилопласт
—Пластида растительной клетки, которая хранит крахмал.
- Центриоль
— парные структуры, состоящие из микротрубочек; в клетках животных управляет плоскостью деления клеток.
- Хлоропласт
—Зеленая органелла у высших растений и водорослей, в которых происходит фотосинтез.
- Хромопласт
—Пластида растительной клетки, содержащая желтый и оранжевый пигменты.
- Реснички
—Короткие выступы, состоящие из микротрубочек, которые покрывают поверхность некоторых клеток и обеспечивают движение.
- Cisface
— Сторона (или «лицо») тела Гольджи, которая принимает везикулы, содержащие макромолекулы.
- Crista
—пл., cristae, складки внутренней мембраны митохондрии.
- Цитоплазма
— Вся протоплазма в живой клетке, расположенная вне ядра, в отличие от нуклеоплазмы , , которая является протоплазмой в ядре.
- Цитоскелет
—Сеть различных белковых нитей, прикрепленных к клеточной мембране и различным органеллам, которая составляет основу формы и движения клеток.
- Дезоксирибонуклеиновая кислота (ДНК)
— Генетический материал клетки, содержащий закодированные инструкции по синтезу белков
- Эндоплазматический ретикулум
— Сеть мембран, которая распространяется по всей клеточной сети; участвует в синтезе белков и липидном обмене.
- Эндосимбиотическая теория
— теория, которая предполагает, что митохондрии, хлоропласты и другие эукариотические органеллы первоначально возникли внутри клеток в результате симбиоза между одноклеточным прокариотом и другим прокариотом.
- Эукариотическая клетка
— клетка, генетический материал которой находится на хромосомах внутри ядра, заключенного в оболочку. У эукариотических клеток также есть органеллы, которые выполняют определенные метаболические задачи и поддерживаются цитоскелетом, который проходит через цитоплазму, придавая клетке форму и форму.
- Flagellum
— нитевидный придаток определенных клеток, таких как сперматозоиды, который контролирует их передвижение.
- Модель жидкой мозаики
— Модель, описывающая природу плазматической мембраны; «мозаичная» часть описывает белки, встроенные в плазматическую мембрану, а «жидкая» часть описывает текучесть плазматической мембраны.
- Тело Гольджи
— Органелла, которая производит, сортирует и транспортирует макромолекулы внутри клетки.
- Granum
— мешки внутри хлоропласта, содержащие фотосинтетические ферменты.
- Гидрофильный
— «Водолюбивый»; описывает фосфатную часть фосфолипида.
- Гидрофобный
— «Ненависть к воде»; описывает жирнокислотную часть фосфолипида.
- Промежуточная нить
—Тип цитоскелетной нити, которая закрепляет органеллы.
- Лизосома
— везикула, содержащая пищеварительные ферменты.
- Матрица
— Внутреннее пространство митохондрии, образованное кристами.
- Микротрубочка
— тип цитоскелетной нити; компонент центриолей, жгутиков и ресничек.
- Митохондрия
— Электростанция клетки; содержит ферменты, необходимые для превращения пищи в энергию.
- Ядерная оболочка
— Двойная мембрана, окружающая ядро.
- Ядерная пора
—Крошечные отверстия, пронизывающие ядерную оболочку.
- Нуклеоид
— Область в прокариоте, где расположена ДНК клетки.
- Ядрышко
— Более темная область внутри ядрышка, где образуются рибосомные субъединицы.
- Ядро
—центр управления клеткой; содержит ДНК.
- Органелла
— мембраносвязанный клеточный «орган», который выполняет определенный набор функций внутри эукариотической клетки.
- Пероксисома
— везикула, которая окисляет жиры и другие вещества и накапливает перекись водорода.
- Фосфолипид
— молекула, состоящая из фосфатной головки и двух цепей жирных кислот, свисающих с головы; компонент плазматической мембраны.
- Фосфолипидный бислой
— Двойной слой фосфолипидов, составляющих плазматическую мембрану.
- Фотосинтез
—В растениях процесс, при котором углекислый газ и вода превращаются в сахара.
- Пили
—Короткие выступы, которые помогают бактериям прикрепляться к тканям.
- Плазменная мембрана
—Мембрана клетки.
- Пластид
— везикулярная органелла, обнаруженная в клетках растений.
- Прокариот
—Клетка без истинного ядра.
- Протист
— одноклеточный эукариотический организм.
- Рибонуклеиновая кислота
-РНК; молекула, транслируемая с ДНК в ядре, которая направляет синтез белка в цитоплазме; это также генетический материал многих вирусов.
- Рибосома
— Белок, состоящий из двух субъединиц, который участвует в синтезе белка.
- Строма
—Материал, которым омывается внутренняя часть хлоропластов в растительных клетках.
- Отношение площади поверхности к объему
—Зависимость между площадью поверхности, обеспечиваемой плазматической мембраной, и объемом содержимого клетки.
- Тилакоид
— мембранная структура, которая делит пополам внутреннюю часть хлоропласта.
- Transface
— Сторона (или «лицо») тела Гольджи, которое высвобождает наполненные макромолекулами везикулы для транспортировки.
- Тубулин
—белок, содержащий микротрубочки.
- Вакуоль
—Мембранно-замкнутая структура внутри клеток, в которых хранятся пигменты, вода, питательные вещества и отходы.
- Везикула
— мембраносвязанная сфера, которая содержит в клетках различные вещества.