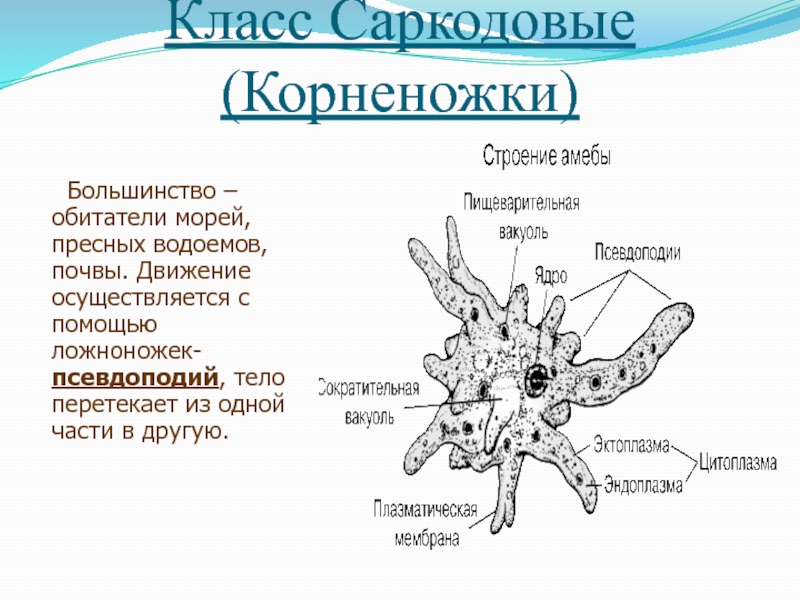
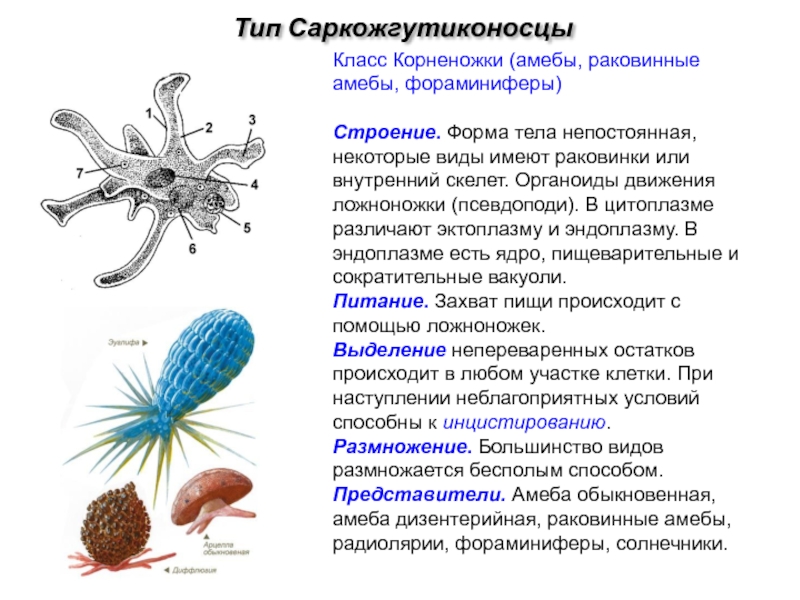
Наиболее часто встречающиеся простейшие, передающиеся с пищевыми продуктами
Согласно статистическим данным Всемирной организации здравоохранения, в мире более 4,5 млрд. человек заражены различными паразитами. В развивающихся странах, зараженность теми или иными паразитами может достигать 93-97%.
Существует огромное количество разнообразных паразитов, которые могут существовать во внутренних органах человека или населять его кожу и волосы. В том числе паразитарные заболевания могут вызывать и простейшие организмы, которые представляют группу одноклеточных или колониальных организмов. Большинство простейших — микроорганизмы, но некоторые могут достигают размеров в несколько миллиметров и хорошо видны невооружённым глазом. Настоящих многоклеточных форм среди простейших нет.
Способов попадания паразитов в организм человека довольно много, однако, одним из наиболее часто встречающихся путей заражения является заражение через пищевые продукты. Плохо промытые овощи, фрукты или зелень, недостаточно прожаренное или сваренное мясо, рыба, птица, неправильно обработанные продукты питания, могут привести к заражению простейшими.
Плохо промытые овощи, фрукты или зелень, недостаточно прожаренное или сваренное мясо, рыба, птица, неправильно обработанные продукты питания, могут привести к заражению простейшими.
Давайте же попробуем разобраться, какие простейшие, передаются с пищей и встречаются наиболее часто в нашей стране, а также как свести к минимуму риск заражения ими.
Простейшие:
Лямблиоз
|
|
Заболевание, вызывается лямблиями и поражающей кишечник человека. Лямблиоз может протекать бессимптомно, в этом случае заболевание выявляется случайно, после проведения лабораторных исследований. Клинически заболевание проявляется умеренными болями в животе, вздутием живота, тошнотой, изжогой, расстройством стула. У человека, зараженного лямблиями, могут наблюдаться повышенная утомляемость, нарушение аппетита, снижение массы тела, бледность кожных покровов, мышечные подергивания. |

Значительную роль в развитии заболевания играет состояние слизистой оболочки тонкой кишки и кислотность желудочного сока. У людей с пониженной кислотностью желудочного сока, нарушением слизистой оболочки кишечника, риск заражения лямблиозом, а также тяжесть клинических проявлений возрастают.
Балантидоз
|
|
Возбудителем балантидоза является балантидий кишечный — единственный представитель инфузорий, паразитирующий в толстом кишечнике человека. Заболевание характеризуется изъязвлениями стенки толстой кишки с признаками общей интоксикации организма, но чаще протекает с незначительными клиническими проявлениями или вовсе бессимптомно. |

Возникновению признаков заболевания способствует ряд факторов: нарушение питания, снижение активности иммунной системы, переохлаждение организма, воспалительные заболевания кишечника и другие. В местах локализации паразитов возникает воспалительно-язвенный процесс, происходит отмирание клеток кишечника с образованием язв. У больных возникает общая интоксикация, появляются боли в животе, понос, в фекалиях – слизь и кровь. Наиболее опасными осложнениями балантидиаза являются прободение язв с развитием перитонита и кишечные кровотечения.
Амебиаз
|
|
Возбудителем амебиаза является дизентерийная амеба, обитающая в кишечнике человека. Заболеваемость амебиазом довольно высока — ежегодно регистрируются несколько сотен миллионов больных во всех странах. Для нескольких десятков тысяч больных заболевание заканчивается летальным исходом. В России эндемичными районами являются Закавказье, Калмыкия, Астраханская область и районы, примыкающие к среднеазиатским республикам. |
 Заболевание характеризуется частым водянистым стулом с примесью крови и слизи, болями в животе, лихорадкой и обезвоживанием организма.
Заболевание характеризуется частым водянистым стулом с примесью крови и слизи, болями в животе, лихорадкой и обезвоживанием организма.
Если место обитания амебы в просвете толстой кишки, то паразитирование не приводит к возникновению клинических признаков заболевания. Если же дизентерийная амеба проникает в ткани слизистой оболочки толстой кишки, то образуются язвы. У больных появляются сильные боли в животе, развивается диарея (до 15 и более раз в сутки). Отличительный признак амебиаза – стул напоминает слизь и окрашен кровью («малиновое желе»). При разрушении сосудов стенки кишечника может возникнуть массивное кровотечение.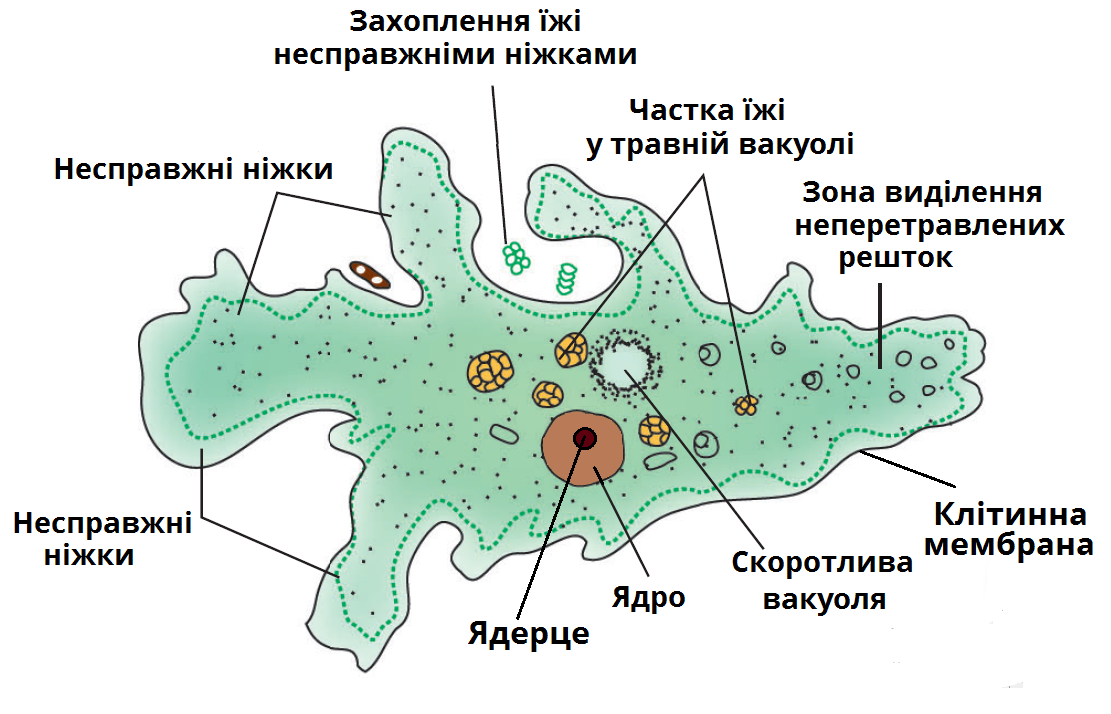
Больные амебной дизентерией подлежат обязательной госпитализации.
Токсоплазмоз
|
|
Возбудителем токсоплазмоза является токсоплазма. Заболевание характеризуется хроническим течением, поражением нервной системы, увеличением печени и селезенки, поражением скелетных мышц и миокарда. Цикл развития токсоплазмы довольно сложный: происходит не только со сменой хозяев, но и чередованием жизненных стадий паразита. Окончательные хозяева паразита – представители семейства кошачьих — американская рысь, бенгальский тигр и домашние кошки. |
Большинство случаев токсоплазмоза протекает бессимптомно. На характер проявлений заболевания существенное влияние оказывает общее состояние организма, а также активность иммунной системы.
У лиц с нарушениями иммунной системы заболевание приобретает особенно тяжелый характер. Увеличение количества очагов размножения паразита приводит к воспалительным изменениям в органах с последующим отмиранием тканей. Наиболее частое и грозное осложнение токсоплазмоза – энцефалит, летальность при котором достигает до 90%.
Профилактика паразитозов
Чтобы не допустить заражения простейшими, нужно соблюдать несколько правил, а именно:
- Соблюдать простейшие правила личной гигиены, в том числе мытье рук после улицы, перед едой, после контакта с деньгами и животными.
- Стараться избегать контакта с бродячими животными, а при наличии у вас питомца регулярно посещать ветеринарную клинику. Обязательно проводить курсы дегельминтизации животных с периодичностью, рекомендуемой ветеринаром.
- Тщательно мыть фрукты, овощи, ягоды, даже если они с Вашего приусадебного участка.

- Не пить воду и не мыть посуду и продукты питания водой, в безопасности которой Вы не уверены.
- Употреблять в пищу только продукты, которые прошли необходимую термическую обработку.
- Обеспечить защиту от насекомых на Вашей кухне. Например, засетчивать оконные проемы, накрывать готовые блюда.
- Не есть вяленую или сушеную рыбу домашнего приготовления.
- Не употреблять домашнее соленое сало.
Микроорганизмы способны вести «фермерское хозяйство», считают ученые
https://ria.ru/20110120/323961267.html
Микроорганизмы способны вести «фермерское хозяйство», считают ученые
Микроорганизмы способны вести «фермерское хозяйство», считают ученые — РИА Новости, 20.01.2011
Микроорганизмы способны вести «фермерское хозяйство», считают ученые
Ученые обнаружили, что одноклеточные амебы (простейшие из животных), питающиеся бактериями, способны культивировать их и транспортировать вместе с собой на новое место обитания, что является наиболее примитивной формой ведения фермерского хозяйства.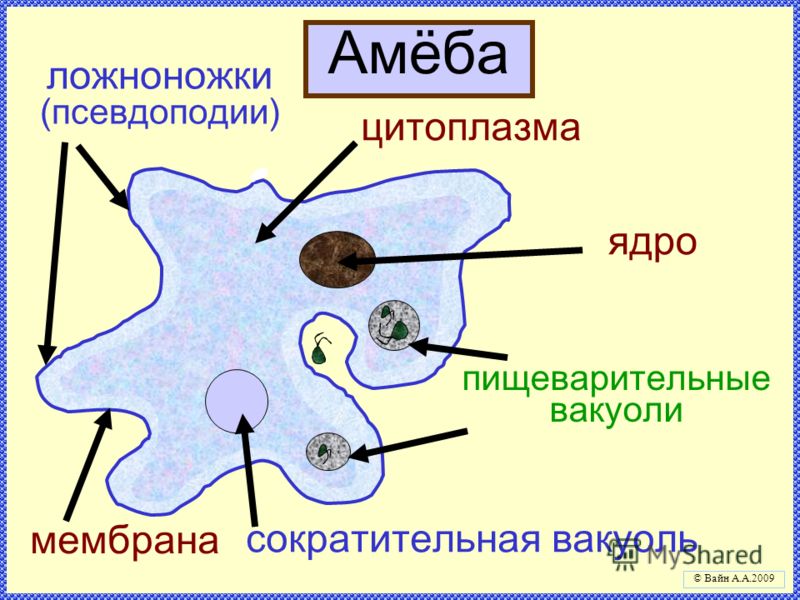
2011-01-20T11:18
2011-01-20T11:18
2011-01-20T11:20
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21.img.ria.ru/images/sharing/article/323961267.jpg?3239531611295511630
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2011
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
1
5
4. 7
7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
1
5
4.7
96
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
МОСКВА, 20 янв — РИА Новости. Ученые обнаружили, что одноклеточные амебы (простейшие из животных), питающиеся бактериями, способны культивировать их и транспортировать вместе с собой на новое место обитания, что является наиболее примитивной формой ведения фермерского хозяйства, сообщается в статье исследователей, опубликованной в журнале Nature.
Фермерство известно и у других организмов, например, муравьев, выращивающих тлю — снабжающих её кормом и защищающих и от вредителей. Однако выявленный пример фермерства (у амеб) является, по всей видимости, самой примитивной его формой.
Автор открытия, Дебра Брок (Debra Brock), завершающая к настоящему времени написание своей кандидатской диссертации в Райсовском университете в США, обнаружила, что одноклеточные организмы — амебы Dictyostelium discoideum — могут, при перемещении на новое мест обитания, брать используемые ими для питания бактерии с собой и выпускать их для разведения на новом месте.
Несмотря на то, что этот одноклеточный организм является очень популярным объектом для исследования, до сих пор никому не удавалось обнаружить такое поведение животных. Причина состоит в том, что большинство штаммов этого организма, используемого в лабораториях, получено от одного и того же штамма, широко распространяемого на коммерческой основе и выращиваемого в искусственных условиях.
Брок, работавшая в течение десяти лет в качестве лаборанта в одной из лабораторий университета, однажды столкнулась с штаммом амебы, обитающей в диких условиях, когда и обнаружила необычное поведение.
Дело в том, что когда в одном участке обитания заканчивается пища, одноклеточные амебы собираются в единый организм, примерно сантиметр длиной, называемый слизевой плесенью. Этот организм, извиваясь, передвигается на новое место обитания, и, найдя подходящее, выбрасывает наружу споры новых амеб.
Борк обнаружила, что в некоторых случаях такие организмы несут на себе бактерии. Предположив, что это патогенные микроорганизмы-паразиты, Борк попыталась их уничтожить с помощью антибиотиков, однако обнаружила, что лишенная бактерий, слизевая плесень снова набирает их, как только ей представляется такая возможность.
После попадания в благоприятные условия бактерии высвобождаются и становятся пищей для новых амеб.
При этом, если среда уже содержит какие-либо штаммы бактерий, то штаммы-фермеры размножаются на ней с большим трудом, тогда как обычные амебы чувствуют себя на новом месте вольготно. Это говорит о том, что «фермерство» для этих организмов не является всегда более выгодной стратегией выживания.
Amoeba leningradensis – Microworld
A. leningradensis c. 300 мкм – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis Page & Kalinina, 1984
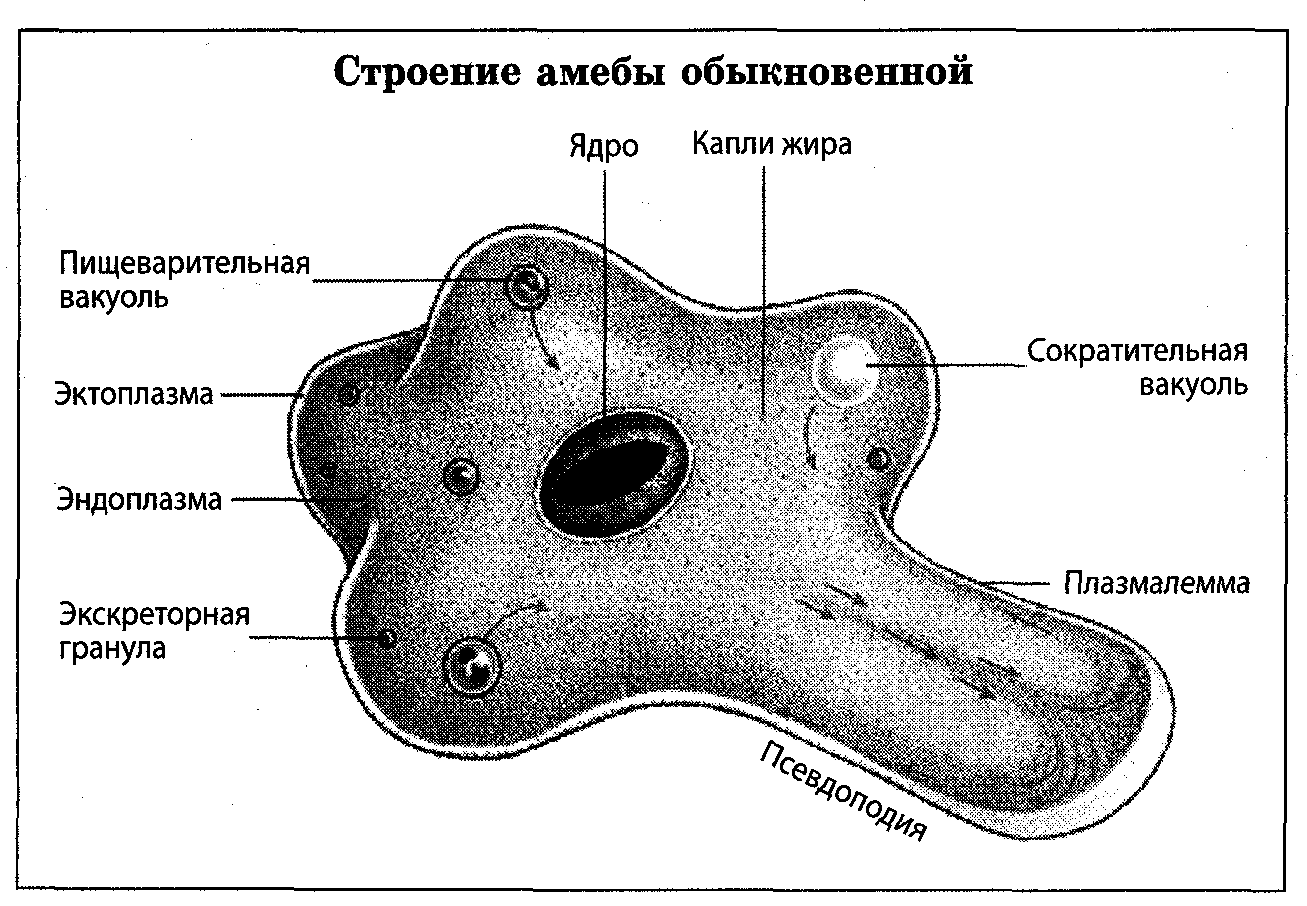
Диагноз : Локомотивная форма часто полиподиальна, длина примерно 160-550 мкм, в среднем около 360 мкм; гиалиновая шляпка менее выражена, как у A. proteus ; ядро — сфера, часто слегка сжатая, часто выглядящая яйцевидной, с наибольшим диаметром от 21 до 29мкм, в среднем 25 мкм; много цитоплазматических кристаллов, преимущественно усеченных бипирамид; уроид грубоморулированный или узловатый, иногда включает висячие остатки псевдоподий, перешедших на задний конец.
Плазматическая мембрана покрыта аморфной оболочкой толщиной 13-20 нм; ядерная оболочка с внутренней волокнистой пластинкой, дифференцированной на гексагональные призмы, пластинка обычно имеет толщину от 400 до 550 нм, внутренний диаметр каждой призмы приблизительно 165 нм, ядрышкообразный материал сосредоточен вокруг периферии ядра, но также встречается внутри: тельца Гольджи, состоящие примерно из четырех уплощенных мешочков и ассоциированные везикулы, возможно, иногда только везикулы. Ядро несовместимо при трансплантации с ядром и цитоплазмой Амеба протей.
Известная среда обитания : Пресная вода, северо-запад СССР и Флорида, США.
Примечания : Я нашел три экземпляра этого вида в образце из садового пруда в Веллингтоне, Флорида. Материал любезно предоставила мне Сандра Редекер (2014). Все образцы выглядели одинаково по своей общей форме и поведению, хотя один был вдвое длиннее (400 мкм), чем самый маленький (180 мкм). Ядро первого экземпляра имело яйцевидную форму, остальные экземпляры имели ядро шахматной формы, напоминающее протеус ядро. Ядра варьировались от 21-33 мкм в диаметре. Ядрышковый материал был рассеян по всему ядру, но более сконцентрирован на периферии. У всех экземпляров никогда не было спинных складок или гребней. Уроид был мелко сосочковым, особенность, которую я редко видел у A. proteus .
Ядро первого экземпляра имело яйцевидную форму, остальные экземпляры имели ядро шахматной формы, напоминающее протеус ядро. Ядра варьировались от 21-33 мкм в диаметре. Ядрышковый материал был рассеян по всему ядру, но более сконцентрирован на периферии. У всех экземпляров никогда не было спинных складок или гребней. Уроид был мелко сосочковым, особенность, которую я редко видел у A. proteus .
Amoeba leningradensis – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis, ядро (21 X 29 мкм) и мочевидная железа (справа) – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis ядро – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis 90 004 второй экз., c. 400 мкм в длину – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis второй экземпляр, папиллярный уроид – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis продвигающаяся псевдоподия – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis клетчатое ядро второго экземпляра, 33 мкм – Садовый пруд, Веллингтон, Флорида
Amoeba leningradensis третий экземпляр, c. 180 мкм в длину, ядро 21 мкм, шахматной формы – Садовый пруд, Веллингтон, Флорида
180 мкм в длину, ядро 21 мкм, шахматной формы – Садовый пруд, Веллингтон, Флорида
Кристаллы и липоидные тельца
Фото ниже взято из Page and Kalinina, 1984:
Обратите внимание, что на микрофотографии 5 изображено ядро A. proteus , для сравнения!
Паром Siemensma, создан 27 февраля 2019 г.; последнее изменение 31 января 2021 г.
Морфология трихамебы
Морфология трихамебыОсновное содержание
< Главная страница
Мир простейших, коловраток, нематод и олигохет
Trichamoeba Fromental, 1874 (ref. ID; 4238), Fromental, 1874 изм. Schaeffer, 1926 (ref. ID; 3687) или Fromental, 1874, исправление. Siemensma & Page, 1986 (арт. ID; 7606) Семейство Amoebidae Ehrenberg, 1838 (ссылка ID; 7606, 7615) [исх. ИДЕНТИФИКАТОР; 4238]Этот род был создан Fromentel для включения двух видов:
 Schaeffer (1926) рассмотрел таксономический статус этого рода, уточнил его и описал шесть новых видов. Два других вида были описаны Bovee (1972). Хотя Пейдж (1976) выделил четыре вида: T. villosa , T. myakka Bovee, 1972, T. osseosaccus Schaeffer, 1926 и T. cloaca Bovee, 1972, позже он оспаривал сходство между T. hirta и A. villosa и отвергли синонимы (FJ Siemensma & FC Page, unpbl.). Род Trichamoeba характеризуется наличием при локомоции уроида, состоящего из многочисленных тонких волосовидных выростов, отходящих от заднего конца, и удлиненно-булавовидной формой тела, причем передняя часть шире задней. Никаких псевдоподий, кроме субполушаровидных выступов, не образуется. Ветвящиеся псевдоподии никогда не образуются. Движение быстрое и осуществляется обычно эруптивными волнами. Известно, что лучевая стадия не встречается. Бови включены Trichamoeba в семействе Pelomyxidae Schulze, 1877, в подотряде Limacina Bovee & Jahn, 1966, вместе с двумя другими лимаксными амебами, Pelomyxa и Saccamoeba .
Schaeffer (1926) рассмотрел таксономический статус этого рода, уточнил его и описал шесть новых видов. Два других вида были описаны Bovee (1972). Хотя Пейдж (1976) выделил четыре вида: T. villosa , T. myakka Bovee, 1972, T. osseosaccus Schaeffer, 1926 и T. cloaca Bovee, 1972, позже он оспаривал сходство между T. hirta и A. villosa и отвергли синонимы (FJ Siemensma & FC Page, unpbl.). Род Trichamoeba характеризуется наличием при локомоции уроида, состоящего из многочисленных тонких волосовидных выростов, отходящих от заднего конца, и удлиненно-булавовидной формой тела, причем передняя часть шире задней. Никаких псевдоподий, кроме субполушаровидных выступов, не образуется. Ветвящиеся псевдоподии никогда не образуются. Движение быстрое и осуществляется обычно эруптивными волнами. Известно, что лучевая стадия не встречается. Бови включены Trichamoeba в семействе Pelomyxidae Schulze, 1877, в подотряде Limacina Bovee & Jahn, 1966, вместе с двумя другими лимаксными амебами, Pelomyxa и Saccamoeba .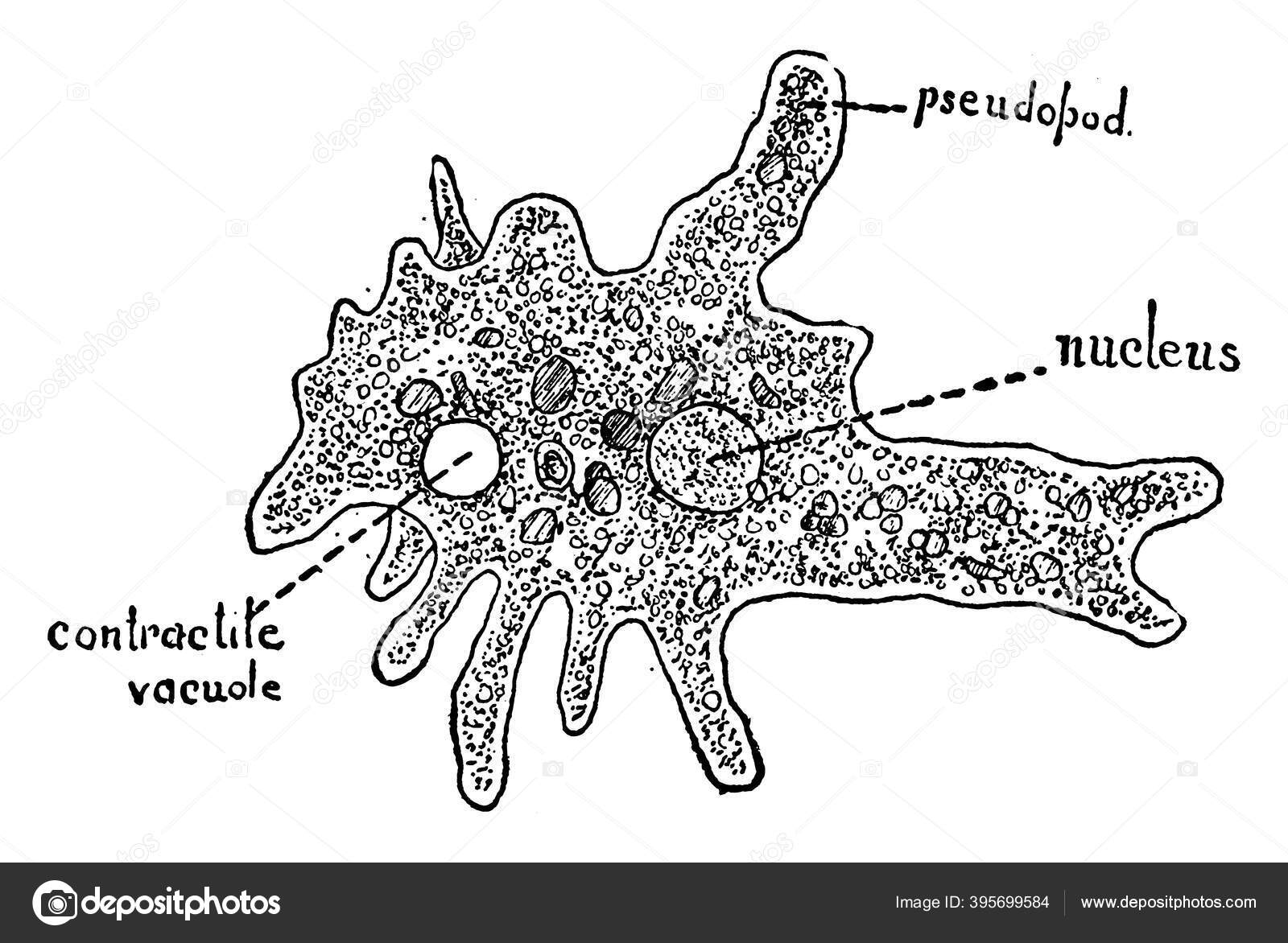 Пейдж, однако, включил Trichamoeba в семейство Amoebidae с другими полиподиальными амебами, такими как Amoeba Bory de St. Vincent, 1822, Chaos Linnaeus, 1767, и Polychaos Schaeffer, 1926, вместо этого из семейства Hartmannellidae Volkonsky, 1931, наряду с другими типично лимаксными амебами. Сообщалось о наличии как полиподиальной, так и булавовидной лимаксной формы для типового вида,
Пейдж, однако, включил Trichamoeba в семейство Amoebidae с другими полиподиальными амебами, такими как Amoeba Bory de St. Vincent, 1822, Chaos Linnaeus, 1767, и Polychaos Schaeffer, 1926, вместо этого из семейства Hartmannellidae Volkonsky, 1931, наряду с другими типично лимаксными амебами. Сообщалось о наличии как полиподиальной, так и булавовидной лимаксной формы для типового вида,  leningradensis и Trichamoeba sp. Аморфный слой на поверхности клеток T. mycophaga подтверждает его включение в род Amoebidae Trichamoeba , а не в род Hartmannellidae Saccamoeba . У последних покрытие клеточной поверхности очень тонкое и у хорошо фиксированных экземпляров имеет тонкие шестиугольные чашевидные структуры, подобные таковым у других Hartmannellidae. Другой важной характеристикой амебид является наличие внутренней волокнистой пластинки в ядрах многих родов. Сотовые пластины A. proteus , о которых сообщают Пейдж и Калинина (1984), также встречаются в A. leningradensis (1984) и P. dubium (1974). Amoeba algonquinensis (1983) не имеет сложной сотовой организации; вместо этого между нуцелолярными телами и ядерной мембраной возникают слои параллельных волокон. Наличие этих волокон в ядрах A. algonquinensis было подтверждено F.C. Паж (лиц. сообщ.). Хотя ядра большинства родов Amoebidae содержат несколько ядрышек или зернистых ядрышковых тел, P.
leningradensis и Trichamoeba sp. Аморфный слой на поверхности клеток T. mycophaga подтверждает его включение в род Amoebidae Trichamoeba , а не в род Hartmannellidae Saccamoeba . У последних покрытие клеточной поверхности очень тонкое и у хорошо фиксированных экземпляров имеет тонкие шестиугольные чашевидные структуры, подобные таковым у других Hartmannellidae. Другой важной характеристикой амебид является наличие внутренней волокнистой пластинки в ядрах многих родов. Сотовые пластины A. proteus , о которых сообщают Пейдж и Калинина (1984), также встречаются в A. leningradensis (1984) и P. dubium (1974). Amoeba algonquinensis (1983) не имеет сложной сотовой организации; вместо этого между нуцелолярными телами и ядерной мембраной возникают слои параллельных волокон. Наличие этих волокон в ядрах A. algonquinensis было подтверждено F.C. Паж (лиц. сообщ.). Хотя ядра большинства родов Amoebidae содержат несколько ядрышек или зернистых ядрышковых тел, P. fasciculatum , а также T. mycophaga имеют одно ядро с одним ядрышком. Внутри генов Trichamoeba , T. myakka и T. cloaca имеют зернистый хроматин, как и новый вид из Голландии. С другой стороны, четыре из 10 видов, включенных Schaeffer (1926) в Trichamoeba и включающих типовые виды. T. villosa (1966), имеют единую массу центрально расположенного ядрышкового материала. Сходное разнообразие в организации ядрышек отмечено у представителей рода 9.0090 Текамеба . (код ID; 4238)
[исх. ИДЕНТИФИКАТОР; 7606]
fasciculatum , а также T. mycophaga имеют одно ядро с одним ядрышком. Внутри генов Trichamoeba , T. myakka и T. cloaca имеют зернистый хроматин, как и новый вид из Голландии. С другой стороны, четыре из 10 видов, включенных Schaeffer (1926) в Trichamoeba и включающих типовые виды. T. villosa (1966), имеют единую массу центрально расположенного ядрышкового материала. Сходное разнообразие в организации ядрышек отмечено у представителей рода 9.0090 Текамеба . (код ID; 4238)
[исх. ИДЕНТИФИКАТОР; 7606] Повторная диагностика ; Одноядерный. Регулярная моноподия в передвижении, продвигающаяся за счет устойчивого потока или неэруптивного переднебокового выпячивания. Все или большинство цитоплазматических кристаллов бипирамидальны. Поверхностный слой из волокнистого материала, более или менее аморфного строения. В ядре одного из видов, исследованных электронно-микроскопически, внутренняя пластинка отсутствует. Свободная жизнь.
 (код ID; 7606)
(код ID; 7606) Примечания ; Род Trichamoeba был установлен Фроменталем (1874) для двух видов, Т. hirta и Т. radiata , без обозначения типового вида. Он определил род по двум признакам: -Nous avons cree ce жанр pour des Amibes peu difflutes, mais avec des changements assez profonds dans la forme du corps. Le tegument est orne de cils raides et non vibratiles-. Schaeffer (1926), переопределяя род, сузил его до амеб с моноподиальной формой и уроидом из многочисленных ворсинок. Однако он создал две проблемы. Во-первых, его родовое определение теперь оказалось неадекватным, поскольку оно было слишком широким. В него входят не только амебы (по определению Стр. 1976), но также и амебы, теперь признанные представителями родов Rhizamoeba (Page 1972) и Saccamoeba (Bovee 1972; Page 1974). Шеффер не делал никакого различия между одноядерными и многоядерными видами и даже предположил, что паразитический Endamoeba barreti , вероятно, принадлежал к этому роду.
 На необходимость более четкого определения этой гетерогенной группы обратили внимание Бови (1951) и Пейдж (1974). Род Saccamoeba был переопределен и отделен от Trichamoeba по Bovee (1972), в основном на том основании, что первый имеет везикулярное ядро. К сожалению, Бови не дал общего диагноза Trichamoeba независимо от определения предложенного им нового подсемейства, Trichamoebinae, члены которого, согласно этому диагнозу, имеют зернистые ядра. Это подсемейство здесь не используется, и теперь необходимо переопределить Trichamoeba . Род Rhizamoeba был выделен из Trichamoeba 9.0091 Page (1972) на основании различий в уроидальных структурах.
Вторая проблема касается типового вида Trichamoeba (стр. 1972). Schaeffer (1926) отверг один из первоначальных видов Fromental, T. radiata , из рода, поскольку он, вероятно, был Nuclearia . Другие виды Фроментела, Т. hirta и Т. lieberkuehnia Maggi, 1880, он считал синонимами вида Валлиха (1863) Amoeba villosa , и на этом основании обозначил A.
На необходимость более четкого определения этой гетерогенной группы обратили внимание Бови (1951) и Пейдж (1974). Род Saccamoeba был переопределен и отделен от Trichamoeba по Bovee (1972), в основном на том основании, что первый имеет везикулярное ядро. К сожалению, Бови не дал общего диагноза Trichamoeba независимо от определения предложенного им нового подсемейства, Trichamoebinae, члены которого, согласно этому диагнозу, имеют зернистые ядра. Это подсемейство здесь не используется, и теперь необходимо переопределить Trichamoeba . Род Rhizamoeba был выделен из Trichamoeba 9.0091 Page (1972) на основании различий в уроидальных структурах.
Вторая проблема касается типового вида Trichamoeba (стр. 1972). Schaeffer (1926) отверг один из первоначальных видов Fromental, T. radiata , из рода, поскольку он, вероятно, был Nuclearia . Другие виды Фроментела, Т. hirta и Т. lieberkuehnia Maggi, 1880, он считал синонимами вида Валлиха (1863) Amoeba villosa , и на этом основании обозначил A. villosa как типовой вид Trichamoeba . (код ID; 7606)
villosa как типовой вид Trichamoeba . (код ID; 7606) Примечания ; Обозначение типового вида Шеффером (1926) верно, поскольку он выбрал T. hirta , один из исходных видов Fromental (1874), но недействителен в его попытке синонимизации T. hirta с Amoeba villosa Wallich, 1863 , в результате чего Шеффер использовал « Trichamoeba villosa » в качестве названия типовых видов. Согласно Международному кодексу зоологической номенклатуры (Международная комиссия по зоологической номенклатуре, 1985), Amoeba villosa не может быть признан типовым видом, если он не является синонимом одного из двух видов, включенных Fromentel (статья 69). Поскольку мы не принимаем эту синонимию, мы заключаем, в соответствии с нашей интерпретацией статьи 69 и предложением Шеффера, что Шеффер (1926) действительно обозначал Т. hirta как типовой вид. Одной из причин отказа от предполагаемой синонимии является то, что длина T. hirta была, согласно увеличению цифры Фроментеля, приблизительно 125 мкм, тогда как длина 9000 A.
 villosa Wallich (1863) имел длину до 500 мкм. Кроме того, описание A. villosa и дополнительные сведения, разбросанные по трем цитируемым работам, указывают на то, что амеба Валлиха несомненно была полиподиальной. Амеба Фроментеля не была полиподиальной, и заявление Шеффера о том, что род Trichamoeba не имел «никаких ложноножек, кроме субполусферических отростков», ясно указывает на его намерение включить только моноподиальные амебы. (код ID; 7606)
villosa Wallich (1863) имел длину до 500 мкм. Кроме того, описание A. villosa и дополнительные сведения, разбросанные по трем цитируемым работам, указывают на то, что амеба Валлиха несомненно была полиподиальной. Амеба Фроментеля не была полиподиальной, и заявление Шеффера о том, что род Trichamoeba не имел «никаких ложноножек, кроме субполусферических отростков», ясно указывает на его намерение включить только моноподиальные амебы. (код ID; 7606) Типовой вид ; Trichamoeba hirta Fromental, 1874 г. (арт. ID; 7606) [исх. ИДЕНТИФИКАТОР; 7615]
Одноядерный. Регулярные моноподии в передвижении. Все или большинство цитоплазматических кристаллов бипирамидальны. Поверхностный слой нечеткого вида и ядра без заметной внутренней пластинки у одного вида, изученного под электронным микроскопом. Свободная жизнь. (код ID; 7615)
Типовой вид ; Trichamoeba hirta Fromental, 1874 г. (арт. ID; 7615)
- Trichamoeba caerulea (код ID; 2618)
- Trichamoeba clava Schaeffer, 1926 (ref.
 ID; 3687) указанный год? (код ID; 3491)
ID; 3687) указанный год? (код ID; 3491) - Trichamoeba cloaca Bovee, 1972 (арт. ID; 4238)
- Trichamoeba coerulea Schaeffer, 1926 (кат. ID; 3687)
- Trichamoeba frenzeli Лепси, 1960 г. (арт. ID; 3687 оригинальная бумага)
- Трихамеба гумия Шеффер, 1926 (ссылочный номер; 2618, 3687)
- Trichamoeba hirta Fromental, 1874 (ref. ID; 7606, 7615) указан автор и год? (код ID; 2618)
- Trichamoeba monofila Lepsi, 1960 (арт. ID; оригинальная бумага 3687)
- Trichamoeba myakka Bovee, 1972 (арт. ID; 4238)
- Trichamoeba mycophaga Chakraborty & Old, 1986 (ссылочный номер; 4238 оригинальная статья)
- Trichamoeba osseosaccus Schaeffer, 1926 (ref. ID; 3687, 4238, 7606) указан автор и год? (код ID; 2618)
- Trichamoeba pallida Schaeffer, 1926 (арт.
 ID; 2618, 3687)
ID; 2618, 3687) - Trichamoeba pavonia Lepsi, 1960 (арт. ID; оригинальная бумага 3687)
- Trichamoeba radiata (арт. ID; 2618)
- Trichamoeba schaefferi Radir, 1927 (арт. ID; 2618, 3687)
- Trichamoeba sinuosa Siemensma & Page, 1986 (ссылка ID; оригинальная бумага 7606)
- Trichamoeba sphaerarum Schaeffer, 1926 (ref. ID; 3687) указан автор и год? (код ID; 2618)
- Trichamoeba urotricha (Lepsi, 1953) (код ID; 3687)
- Trichamoeba villosa (Wallich, 1863) (ссылочный номер; 2618, 3687)
Трихамеба микофага Chakraborty & Old, 1986 (ссылка ID; оригинальная бумага 4238)
Диагностика
Трофозоиты от пальчатых до удлиненных, преимущественно полиподиальные, но могут быть моноподиальными при непрерывном передвижении, кажутся трубчатыми или булавовидными с ворсинчато-луковичным уроидом. Эктоплазматическая шапочка на псевдоподиях и на переднем крае лимаксных трофозоитов. Эндоплазма сильно зернистая с удлиненными и бипирамидальными кристаллами до 2,5 мкм длиной. Обычно одиночная сократительная вакуоль диаметром 3-15 мкм расположена в задней части клетки. Трофозоиты одноядерные со сферическими или овальными ядрами диаметром 4-10 мкм. Ядрышковый материал организован в единую нефрагментированную структуру, от овальной до удлиненной, диаметром 2,8-5,0 мкм. Размеры трофозоита: 45-136 мкм в длину и 25-9 мкм.шириной 4 мкм. Кисты округло-овальные, диаметром 21-60 мкм, стенки экто- и эндоцисты разделены аморфным материалом толщиной несколько мкм. (код ID; 4238)
Эктоплазматическая шапочка на псевдоподиях и на переднем крае лимаксных трофозоитов. Эндоплазма сильно зернистая с удлиненными и бипирамидальными кристаллами до 2,5 мкм длиной. Обычно одиночная сократительная вакуоль диаметром 3-15 мкм расположена в задней части клетки. Трофозоиты одноядерные со сферическими или овальными ядрами диаметром 4-10 мкм. Ядрышковый материал организован в единую нефрагментированную структуру, от овальной до удлиненной, диаметром 2,8-5,0 мкм. Размеры трофозоита: 45-136 мкм в длину и 25-9 мкм.шириной 4 мкм. Кисты округло-овальные, диаметром 21-60 мкм, стенки экто- и эндоцисты разделены аморфным материалом толщиной несколько мкм. (код ID; 4238)Описания
- Трофозоит: Амебы в движении часто имеют пальчато-удлиненную форму. Также часто встречается тонкая трубчатая форма, типичная для так называемой «лимаксной» формы. Трофозоиты имеют длину 45-136 мкм и ширину 25-94 мкм. Отличительной плавающей формы не наблюдается. Подвешенные клетки в водных культурах могут казаться довольно вытянутыми и разветвленными, хотя в других случаях в нарушенных культурах можно увидеть амебы неправильной округлой формы с короткими, радиально выступающими трубчатыми псевдоподиями.
 Трофозоиты плохо прикрепляются к стеклу или пластику, хотя в неподвижных культурах задняя часть многих трофозоитов может уплощаться и прикрепляться ко дну чашки и обычно распространяться во всех направлениях. Одновременно могут образовываться от одной до многих псевдоподий типа «лобоза», продвигающихся в одном или разных направлениях. В новообразованной псевдоподии присутствует гиалиновый полумесяц; однако гиалиновая эктоплазматическая шапочка может быть не видна у быстро движущихся трофозоитов, поскольку зернистая эндоплазма вскоре следует за гиалиновой эктоплазмой. Несколько псевдоподий обычно удлиняются продвигающимся трофозоитом, а количество псевдоподий постоянно меняется за счет повторяющихся подразделений и воссоединений. Только одна из них становится основной псевдоподией и определяет направление движения, но другие псевдоподии могут образовываться одновременно и взять на себя эту роль в любое время. При непрерывной локомоции трофозоиты могут иметь удлиненную, широко моноподиальную или булавовидную форму, обычно с полушаровидной задней бульбовидной (уроидной) опорой, бахромой из коротких нитей.
Трофозоиты плохо прикрепляются к стеклу или пластику, хотя в неподвижных культурах задняя часть многих трофозоитов может уплощаться и прикрепляться ко дну чашки и обычно распространяться во всех направлениях. Одновременно могут образовываться от одной до многих псевдоподий типа «лобоза», продвигающихся в одном или разных направлениях. В новообразованной псевдоподии присутствует гиалиновый полумесяц; однако гиалиновая эктоплазматическая шапочка может быть не видна у быстро движущихся трофозоитов, поскольку зернистая эндоплазма вскоре следует за гиалиновой эктоплазмой. Несколько псевдоподий обычно удлиняются продвигающимся трофозоитом, а количество псевдоподий постоянно меняется за счет повторяющихся подразделений и воссоединений. Только одна из них становится основной псевдоподией и определяет направление движения, но другие псевдоподии могут образовываться одновременно и взять на себя эту роль в любое время. При непрерывной локомоции трофозоиты могут иметь удлиненную, широко моноподиальную или булавовидную форму, обычно с полушаровидной задней бульбовидной (уроидной) опорой, бахромой из коротких нитей. Уроид имеет эктоплазматическое строение и часто соединяется с основным телом лимаксного трофозоита узким перешейком. В активных культурах скорость передвижения составляет примерно 2 мкм сек-1. Цитоплазма клеток состоит из светлой эктоплазматической зоны и плотной высокозернистой эндоплазмы, содержащей фазово-яркие кристаллы. Они либо удлиненные, менее одного мкм в длину, либо бипирамидальные и 1-2,5 мкм в длину. Количество и доля кристаллов каждого типа варьирует у разных трофозоитов. Кзади расположены сократительная вакуоль диаметром 3-15 мкм и ядро - другие цитологические детали, различимые при световом микроскопическом исследовании. Все 100 или около того исследованных трофозоитов были одноядерными. Ядра от сферических до овальных, размером 4-10 мкм в диаметре и содержат одно сферическое или овальное ядрышко диаметром 2,8-5,0 мкм. (код ID; 4238)
Уроид имеет эктоплазматическое строение и часто соединяется с основным телом лимаксного трофозоита узким перешейком. В активных культурах скорость передвижения составляет примерно 2 мкм сек-1. Цитоплазма клеток состоит из светлой эктоплазматической зоны и плотной высокозернистой эндоплазмы, содержащей фазово-яркие кристаллы. Они либо удлиненные, менее одного мкм в длину, либо бипирамидальные и 1-2,5 мкм в длину. Количество и доля кристаллов каждого типа варьирует у разных трофозоитов. Кзади расположены сократительная вакуоль диаметром 3-15 мкм и ядро - другие цитологические детали, различимые при световом микроскопическом исследовании. Все 100 или около того исследованных трофозоитов были одноядерными. Ядра от сферических до овальных, размером 4-10 мкм в диаметре и содержат одно сферическое или овальное ядрышко диаметром 2,8-5,0 мкм. (код ID; 4238) - Ультраструктура: Клетка ограничена плазматической мембраной толщиной примерно 10 нм, покрытой электронно-плотным аморфным материалом, проникающим в суспендирующую среду на глубину до 100 нм.
 Цитоплазма резко разделена на эктоплазму, совершенно лишенную органелл, и эндоплазму, переполненную органеллами. Эктоплазма наиболее широкая спереди, образуя гиалиновую шапочку. Уроиду могут соответствовать и другие участки клеточной мембраны, заброшенные в сложные инвагинации. Органеллы включают многочисленные мелкие везикулы, некоторые из которых содержат частицы пищи, бактерии и споры грибов на разных стадиях лизиса. Никаких кристаллических структур, видимых с помощью световой микроскопии, обнаружено не было. Они могли раствориться во время процедур фиксации. Пейдж и Болдок (1980) показал остатки подобных кристаллов в Polychaos fasciculatum . Бактерии свободно присутствуют в цитоплазме, что свидетельствует о симбиотических отношениях с амёбой. Митохондрии обычно имеют палочковидную или эллипсоидальную форму с везикулярными кристами. Тельца Гольджи обычны и диктиосомны по морфологии, встречаются одиночно или парно. Эндоплазматический ретикулум предустановлен по всей эндоплазме.
Цитоплазма резко разделена на эктоплазму, совершенно лишенную органелл, и эндоплазму, переполненную органеллами. Эктоплазма наиболее широкая спереди, образуя гиалиновую шапочку. Уроиду могут соответствовать и другие участки клеточной мембраны, заброшенные в сложные инвагинации. Органеллы включают многочисленные мелкие везикулы, некоторые из которых содержат частицы пищи, бактерии и споры грибов на разных стадиях лизиса. Никаких кристаллических структур, видимых с помощью световой микроскопии, обнаружено не было. Они могли раствориться во время процедур фиксации. Пейдж и Болдок (1980) показал остатки подобных кристаллов в Polychaos fasciculatum . Бактерии свободно присутствуют в цитоплазме, что свидетельствует о симбиотических отношениях с амёбой. Митохондрии обычно имеют палочковидную или эллипсоидальную форму с везикулярными кристами. Тельца Гольджи обычны и диктиосомны по морфологии, встречаются одиночно или парно. Эндоплазматический ретикулум предустановлен по всей эндоплазме. Другие присутствующие структуры представляют собой гранулы примерно 30-50 нм в диаметре. По-видимому, это продукты распада грибковых клеток. Липидные тела обычны, особенно у амеб, питающихся Fusarium macroconidia , которые содержат большие запасы липидов, иногда видны микрофибриллы. Последние различаются по встречаемости в различных срезах трофозоитов и обычно объединяются в кластеры, подобные тем, которые показал Bhowmick (1966) для Trichamoeba villosa . Ядерная мембрана различается по морфологии, иногда кажется относительно цельной, а иногда имеет вид бусинок из-за наличия большого количества пор. Нуклеоплазма наиболее плотна непосредственно внутри мембраны, с диффузной областью, простирающейся до единственного сферического ядрышка, пересекаемого очень тонкими нитями электронно-плотного материала. Было обнаружено, что скопления филаментов, собранные в пучки до пяти волокон, распределяются радиально через периферическую, более плотную нуклеоплазму. Функция этих структур неизвестна.
Другие присутствующие структуры представляют собой гранулы примерно 30-50 нм в диаметре. По-видимому, это продукты распада грибковых клеток. Липидные тела обычны, особенно у амеб, питающихся Fusarium macroconidia , которые содержат большие запасы липидов, иногда видны микрофибриллы. Последние различаются по встречаемости в различных срезах трофозоитов и обычно объединяются в кластеры, подобные тем, которые показал Bhowmick (1966) для Trichamoeba villosa . Ядерная мембрана различается по морфологии, иногда кажется относительно цельной, а иногда имеет вид бусинок из-за наличия большого количества пор. Нуклеоплазма наиболее плотна непосредственно внутри мембраны, с диффузной областью, простирающейся до единственного сферического ядрышка, пересекаемого очень тонкими нитями электронно-плотного материала. Было обнаружено, что скопления филаментов, собранные в пучки до пяти волокон, распределяются радиально через периферическую, более плотную нуклеоплазму. Функция этих структур неизвестна. На срезах препаратов амеб цист не наблюдалось. (код ID; 4238)
На срезах препаратов амеб цист не наблюдалось. (код ID; 4238) - Морфология цист: В культурах старше трех недель, хранящихся при 25°C, часть трофозоитов прекращает активную локомоцию, убирает псевдоподии и контрастирует по размеру. Эктоплазматический слой постепенно исчезает, и амеба появляется в виде плотной беспорядочной массы светоотражающих гранул и кристаллов диаметром 17-35 мкм. Хотя многие из этих клеток могут умирать, вполне вероятно, что часть из них дает начало кистам. Кисты сферические, удлиненные или овальные, размером 21-60 мкм в диаметре, могут образовываться как поодиночке, так и группами. Стенка кисты отчетливо разделена на эктоцисту и эндоцисту, разделенные желтоватым аморфным слоем толщиной несколько микрометров. Эксцизии не наблюдается. (код ID; 4238)
Экология
Было показано, что Trichamoeba mycophaga питается спорами нескольких грибов, включая Cochliobolus sativus , Cylindrocarpon didymum (Hartig) Wollenw., Fusarium oxysporum fsp. штифт (Hartig) Snyd. & Hansen, Epicoccum nigrum Link ex Link и Endothia gyrosa (Schw. Fr.) Fries. Наряду с некоторыми другими микофагами-амебами и миксобактериями эта амеба относится к очень немногим организмам, способным лизировать меланизированные пропагулы грибов в почве. Считается, что это факультативный грибной корм, также использующий бактерии в качестве пищевых организмов. (код ID; 4238)
штифт (Hartig) Snyd. & Hansen, Epicoccum nigrum Link ex Link и Endothia gyrosa (Schw. Fr.) Fries. Наряду с некоторыми другими микофагами-амебами и миксобактериями эта амеба относится к очень немногим организмам, способным лизировать меланизированные пропагулы грибов в почве. Считается, что это факультативный грибной корм, также использующий бактерии в качестве пищевых организмов. (код ID; 4238)Тип материала
Проводится авторами. Активная культура депонирована в Центре культуры водорослей и простейших Института экологии суши. (код ID; 4238)Trichamoeba osseosaccus Schaeffer, 1926 (ref. ID; 3687, 4238, 7606) указан автор и год? (код ID; 2618)
Замечания
T. osseosaccus внешне похож на T. sinuosa , но есть отличия. Во время движения амеба имеет очертания от неправильно булавовидных до овальных, с более уплощенной формой. Эта форма никогда не наблюдалась в Т. sinuosa . Bovee (1951) описывает облигатно-булавовидную форму с шириной заднего конца от половины до двух третей ширины переднего конца, тогда как Т. sinuosa более цилиндрическая форма с шириной передней части, не сильно отличающейся от ширины переднего конца. задний конец. Висячие филаменты T. osseosaccus образуются в результате прилипания эктоплазмы к субстрату; как Schaeffer, так и Bovee (1951) описывают этот сходный с коллоподием внешний вид, а рисунок уроида, сделанный Schaeffer, не оставляет сомнений в адгезивной природе филаментов. Уроид T. sinuosa никогда не имеет такой структуры, и ворсинки никогда не образуются в результате адгезии. Ядрышковый материал T. osseosaccus расположен в один слой непосредственно под ядерной мембраной, в то время как ядро T. sinuosa имеет отчетливое свободное пространство между слоем ядрышкового материала и ядрышковой мембраной. Плавающая форма с длинными псевдоподиями, наблюдаемая Bovee, никогда не наблюдалась у T. sinuosa . (код ID; 7606)
sinuosa более цилиндрическая форма с шириной передней части, не сильно отличающейся от ширины переднего конца. задний конец. Висячие филаменты T. osseosaccus образуются в результате прилипания эктоплазмы к субстрату; как Schaeffer, так и Bovee (1951) описывают этот сходный с коллоподием внешний вид, а рисунок уроида, сделанный Schaeffer, не оставляет сомнений в адгезивной природе филаментов. Уроид T. sinuosa никогда не имеет такой структуры, и ворсинки никогда не образуются в результате адгезии. Ядрышковый материал T. osseosaccus расположен в один слой непосредственно под ядерной мембраной, в то время как ядро T. sinuosa имеет отчетливое свободное пространство между слоем ядрышкового материала и ядрышковой мембраной. Плавающая форма с длинными псевдоподиями, наблюдаемая Bovee, никогда не наблюдалась у T. sinuosa . (код ID; 7606)Trichamoeba sinuosa Siemensma & Page, 1986 (ссылка ID; оригинальная статья 7606)
Диагностика
Форма локомотива обычно слегка извилистая, длиной около 200 мкм. Ядро сферическое, диаметром от 14,4 до 26,9 мкм (в среднем около 20 мкм), с периферическим слоем зернистого ядрышкового материала. Всегда присутствуют кристаллы, в основном усеченные бипирамиды. Уроид гладкий или сосочковидный, либо покрыт ворсинками. Гликокаликс толщиной около 20-25 нм над плазматической мембраной. Отсутствие различимой внутренней ядерной пластинки. Предполагаемые кисты двуядерные; эндоциста приблизительно 53 мкм в диаметре. (код ID; 7606)
Ядро сферическое, диаметром от 14,4 до 26,9 мкм (в среднем около 20 мкм), с периферическим слоем зернистого ядрышкового материала. Всегда присутствуют кристаллы, в основном усеченные бипирамиды. Уроид гладкий или сосочковидный, либо покрыт ворсинками. Гликокаликс толщиной около 20-25 нм над плазматической мембраной. Отсутствие различимой внутренней ядерной пластинки. Предполагаемые кисты двуядерные; эндоциста приблизительно 53 мкм в диаметре. (код ID; 7606)Описания
- Световая микроскопия:
- 1. Общая форма и поведение. Локомотивная форма представляла собой неразветвленный цилиндр, обычно извилистой формы, передний конец которого несколько шире заднего. Спинных гребней не наблюдалось. Существовало два способа передвижения. Первым было быстрое продвижение устойчивым потоком. Скорость составляла приблизительно 340 мкм в минуту при комнатной температуре; самая высокая наблюдаемая скорость составила 480 мкм в минуту. Второй вид локомоции был несколько медленнее.
 Амеба двигалась последовательностью неэруптивных выпуклостей, поочередно вправо и влево от переднего конца, придавая амебе сигмовидную или извилистую форму, поскольку она постоянно перенаправляла свой передний конец. Такое переднебоковое выпячивание было началом нового моноподия. Бывший передний конец оставался в виде выпуклости вдоль бока амебы, пока не достигал и не вливался в задний конец. Таким образом, эти остатки прежних передних концов можно рассматривать как небольшие изгибы вдоль моноподия. Каждый остаток потенциально может дать новый псевдоподий, что в конечном итоге приведет к новому моноподию. Формирование латеральной псевдоподии всегда совпадало со снижением скорости локомотива и обычно приводило к новому направлению движения. Короткие псевдоподии, почти полушаровидные, обычно числом две или три, появлялись, когда амеба замедляла ход и прекращала свое продвижение. Такие псевдоподии образовывались в, по-видимому, случайных точках клетки. Со временем одна псевдоподия удлинилась и превратилась в новую моноподию.
Амеба двигалась последовательностью неэруптивных выпуклостей, поочередно вправо и влево от переднего конца, придавая амебе сигмовидную или извилистую форму, поскольку она постоянно перенаправляла свой передний конец. Такое переднебоковое выпячивание было началом нового моноподия. Бывший передний конец оставался в виде выпуклости вдоль бока амебы, пока не достигал и не вливался в задний конец. Таким образом, эти остатки прежних передних концов можно рассматривать как небольшие изгибы вдоль моноподия. Каждый остаток потенциально может дать новый псевдоподий, что в конечном итоге приведет к новому моноподию. Формирование латеральной псевдоподии всегда совпадало со снижением скорости локомотива и обычно приводило к новому направлению движения. Короткие псевдоподии, почти полушаровидные, обычно числом две или три, появлялись, когда амеба замедляла ход и прекращала свое продвижение. Такие псевдоподии образовывались в, по-видимому, случайных точках клетки. Со временем одна псевдоподия удлинилась и превратилась в новую моноподию.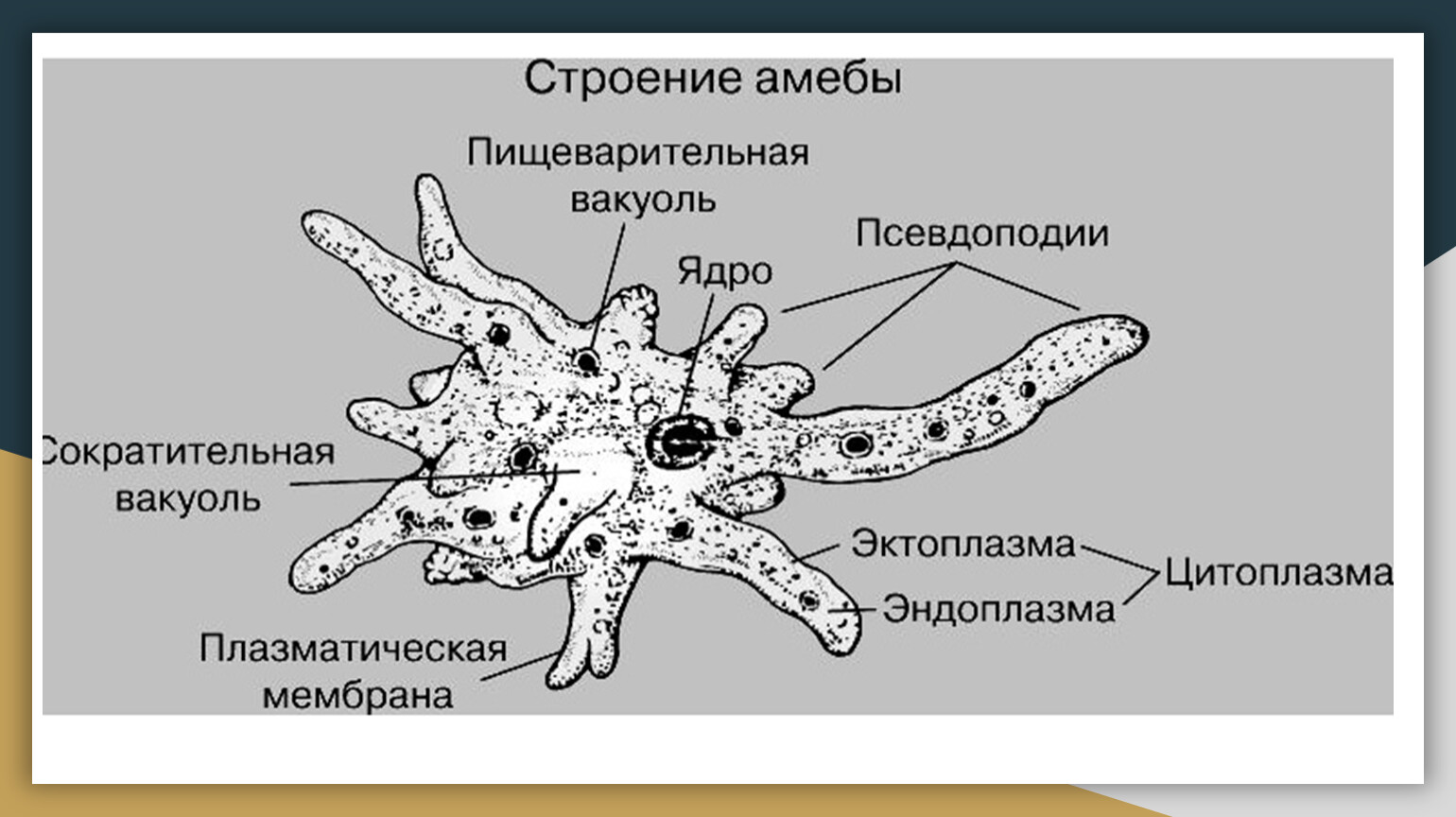 Новое направление также может быть принято путем изменения передне-задней полярности. Такой разворот мог произойти в течение нескольких секунд. Во время установившегося течения амеба меняла направление, слегка смещая передний конец в сторону, что приводило к дугообразной траектории. При продолжающейся локомоции гиалиновая шапочка отсутствовала, так как она постоянно была пронизана и заполнена гранулоплазмой. Гиалиновая шапочка появлялась при образовании нового псевдоподия, но исчезала при удлинении моноподия. Когда формировался гиалиновый колпачок, гиалоплазма никогда не сбегала назад по одной стороне клеточной массы (стр. 19).72). Плавающая форма представляла собой неправильную сферу с неравномерно выступающими в нескольких направлениях псевдоподиями, заполненными или частично заполненными гранулоплазмой. Эта плавающая форма сильно отличалась от формы Saccamoeba , которая представляла собой просто довольно неправильную сферу, часто с небольшими бугорками, но никогда с отчетливыми псевдоподиями.
Новое направление также может быть принято путем изменения передне-задней полярности. Такой разворот мог произойти в течение нескольких секунд. Во время установившегося течения амеба меняла направление, слегка смещая передний конец в сторону, что приводило к дугообразной траектории. При продолжающейся локомоции гиалиновая шапочка отсутствовала, так как она постоянно была пронизана и заполнена гранулоплазмой. Гиалиновая шапочка появлялась при образовании нового псевдоподия, но исчезала при удлинении моноподия. Когда формировался гиалиновый колпачок, гиалоплазма никогда не сбегала назад по одной стороне клеточной массы (стр. 19).72). Плавающая форма представляла собой неправильную сферу с неравномерно выступающими в нескольких направлениях псевдоподиями, заполненными или частично заполненными гранулоплазмой. Эта плавающая форма сильно отличалась от формы Saccamoeba , которая представляла собой просто довольно неправильную сферу, часто с небольшими бугорками, но никогда с отчетливыми псевдоподиями. (код ID; 7606)
(код ID; 7606) - 2. Ядро. Обычно каждая клетка содержала одно сферическое ядро, хотя иногда можно было наблюдать двуядерные особи. Предполагаемый материал ядрышка располагался мелкими кусочками, образуя регулярный слой под ядерной оболочкой. Между этим периферическим слоем зернистого материала и ядерной оболочкой всегда виднелось отчетливое, узкое, чистое пространство. Центральное однородное тело наблюдали in vivo с помощью дифференциальной интерференционно-контрастной микроскопии, а также, но очень неясно, в светлом поле. В окрашенных препаратах это тело было очень заметным. И кармин буры, и гематоксилин железа ясно показали центральную структуру. Это тело явно не является эндосомой, так как ни в одном э.м. не было обнаружено следов эндосомы. раздел. По-видимому, это оптический эффект in vivo и артефакт в окрашенных препаратах. Диаметр ядра у культивируемых амеб колебался от 16,8 мкм до 26,9 мкм.мкм (в среднем 22,3 мкм), тогда как диаметр ядра в полевой пробе амеб был меньше, 14,4-22,8 мкм (в среднем 16,4 мкм).
 Ядра измеряли у 30 амеб из каждого источника. (код ID; 7606)
Ядра измеряли у 30 амеб из каждого источника. (код ID; 7606) - 3. Уроид. Уроид не выражен. Он редко имел вид ручки. Его обычной формой было четкое пальцевидное произношение, иногда образующее острый или небольшой угол с основной клеточной массой. Очертания уроида могли меняться от момента к моменту. Уроид может быть гладким, сосочковым или покрытым мелкими ворсинками. Гладкая мочевая железа встречалась чаще, чем ворсинчатая, а спаечная мочевая железа никогда не наблюдалась. (код ID; 7606)
- 4. Кристаллы. Обычно присутствовало много бипирамидальных кристаллов размером от 2,1 до 10,8 мкм (обычно 2,4-5,2 мкм). Крупные кристаллы имели четко различимую форму с усеченными концами; мелкие кристаллы часто имели неясную форму. Все кристаллы располагались эксцентрично в мелких вакуолях, хотя часто вакуоли различимы с трудом. Диаметр такой вакуоли был примерно в два раза больше диаметра содержащегося в ней кристалла. Амебы в полевых образцах, как правило, содержат меньше кристаллов, чем культивируемые амебы.
 (код ID; 7606)
(код ID; 7606) - 5. Вакуоли. В цитоплазме часто встречались оптически пустые вакуоли. Сократительная вакуоль часто располагалась ближе к заднему концу, между ядром и мочевой оболочкой. Часто он был очень выражен, образуя полушаровидную выпуклость на очертании клеточной массы. Часто образуется путем слияния нескольких мелких вакуолей. (код ID; 7606)
- 6. Кистоподобные тела. Образование цистоподобных тел наблюдалось только в первый месяц после закладки культуры, причем их количество в каждой культуральной чашке в этот период резко снижалось. С тех пор такие органы больше никогда не образовывались. Термин «кистоподобное тело», а не «киста», используется потому, что эксцистирование клетки никогда не наблюдалось. В каждой культуральной чашке 10-20 амеб свернулись и были прочно прикреплены к субстрату. Вокруг зернистой клеточной массы формируется от одного до трех слоев. Внутренние слои были отделены от протоплазмы прозрачным пространством. Центральное тело или эндоциста имели диаметр 50,4-75,6 мкм (в среднем около 53 мкм).
 Каждая предполагаемая киста содержала два ядра, обычно сближенных друг с другом. Диаметры ядер варьировались от 12,0 мкм до 16,8 мкм. Материал ядрышка образовывал компактный почти однородный слой непосредственно под ядерной оболочкой. Цитоплазма в цистоподобных телах была сильно зернистой. (код ID; 7606)
Каждая предполагаемая киста содержала два ядра, обычно сближенных друг с другом. Диаметры ядер варьировались от 12,0 мкм до 16,8 мкм. Материал ядрышка образовывал компактный почти однородный слой непосредственно под ядерной оболочкой. Цитоплазма в цистоподобных телах была сильно зернистой. (код ID; 7606) - 7. Питание. Поглощенная пища в культурах состояла из мелких инфузорий ( Chilomonas sp., Halteria sp.), мелкой коловратки и бактерий. (код ID; 7606)
- 1. Общая форма и поведение. Локомотивная форма представляла собой неразветвленный цилиндр, обычно извилистой формы, передний конец которого несколько шире заднего. Спинных гребней не наблюдалось. Существовало два способа передвижения. Первым было быстрое продвижение устойчивым потоком. Скорость составляла приблизительно 340 мкм в минуту при комнатной температуре; самая высокая наблюдаемая скорость составила 480 мкм в минуту. Второй вид локомоции был несколько медленнее.
- Электронная микроскопия: Поверхность клеток обычно имела довольно аморфный, нечеткий вид. Однако благоприятные срезы предполагали рыхлую, но отчетливую структуру внутри гликокаликса, возможно, состоящую из тонких цилиндрических элементов, возможно, шестиугольных в поперечном сечении, с максимальным диаметром 25 или 30 нм. Лучшие разделы не давали более подробной информации, и эти элементы не были четко сформированы или достаточно дискретны, чтобы считаться гликостилями. Общая толщина гликокаликса составляла 20-25 нм над плазматической мембраной.
 Тонкая структура ядра была поразительно похожа на Hydramoeba hydroxena (Page and Robson 1983): очень неправильный контур, отсутствие какой-либо заметной внутренней волокнистой пластинки, париетальное расположение ядерного материала во многих фрагментах и полное отсутствие центрального ядрышка или кариосомы. Кусочки ядрышка были несколько меньше и многочисленнее у T. sinuosa , чем у H. hydroxena , судя по впечатлению, полученному при наблюдениях под световым микроскопом. Митохондрии в форме колбасы (т. е. удлиненные с параллельными сторонами) были довольно распространены, хотя также наблюдались многочисленные эллиптические и овальные профили. Трубчатые кристы иногда ветвятся. Система Гольджи имела форму многочисленных диктиосом, часто по нескольку в одном отделе. Такое тело обычно состояло из пяти-шести уплощенных цистерн с прилежащими пузырьками, общей шириной до 0,85 мкм. Грубый эндоплазматический ретикулум состоял из несколько уплощенных пузырьков или коротких цистерн, а не длинных трактов, как у некоторых других амебид.
Тонкая структура ядра была поразительно похожа на Hydramoeba hydroxena (Page and Robson 1983): очень неправильный контур, отсутствие какой-либо заметной внутренней волокнистой пластинки, париетальное расположение ядерного материала во многих фрагментах и полное отсутствие центрального ядрышка или кариосомы. Кусочки ядрышка были несколько меньше и многочисленнее у T. sinuosa , чем у H. hydroxena , судя по впечатлению, полученному при наблюдениях под световым микроскопом. Митохондрии в форме колбасы (т. е. удлиненные с параллельными сторонами) были довольно распространены, хотя также наблюдались многочисленные эллиптические и овальные профили. Трубчатые кристы иногда ветвятся. Система Гольджи имела форму многочисленных диктиосом, часто по нескольку в одном отделе. Такое тело обычно состояло из пяти-шести уплощенных цистерн с прилежащими пузырьками, общей шириной до 0,85 мкм. Грубый эндоплазматический ретикулум состоял из несколько уплощенных пузырьков или коротких цистерн, а не длинных трактов, как у некоторых других амебид.