След амебы
Елена Клещенко
«Химия и жизнь» №4, 2009
Безмозглый, безглазый, бесцветный шар, полностью покрытый грязью» — такими словами описал Михаил Матц из университета Техаса (о других его работах см. «Химию и жизнь», 2008, № 12) существо, найденное им с коллегами на морском дне неподалеку от Багамских островов. Чем так замечателен этот шар, точнее, шарик диаметром около 3 см, что о нем пишут и электронные СМИ, и авторитетные научные журналы?
Во время погружений на глубину 750–780 м неподалеку от острова Литл-Сан-Сальвадор на дне были обнаружены шарообразные объекты, каждый величиной с крупную виноградину. Дно было покрыто следами, которые, судя по всему, оставляли именно загадочные шарики. Причем они перемещались не только вниз по склону, но и вверх, и не под действием течения. Судя по разнообразным направлениям и поворотам траекторий, существа двигались активно. Вдобавок след имел непростую форму: канавка, окруженная валиками грунта, с еще одним валиком посередине, особенно отчетливым на конце следа, ближайшем к шарику.
При ближайшем рассмотрении бродячие морские «виноградины» оказались раковинными амебами (корненожками) Gromia sphaerica. Этот вид ранее был обнаружен в Аравийском море. В это трудно поверить, но каждый шарик — одна клетка. Под оболочкой у нее тонкий зеленоватый слой протоплазмы, покрытый оболочкой, а под ним большой водяной пузырь. С помощью такого «надувательства» и одноклеточное может стать макроскопическим объектом. Багамская громия не во всем похожа на арабскую: она не строго шарообразная, а слегка вытянутая и, кроме того, она действительно снаружи покрыта илом. (Чтобы рассмотреть пойманное животное, его сначала пришлось отмыть.) Арабская громия снаружи чистенькая, зато содержит гранулы осадка во внутреннем «пузыре», который у багамской родственницы заполнен чистой водой. Возможно, все это связано с одним, самым главным отличием: арабская амеба ведет сидячий образ жизни, тогда как багамская гуляет по дну.
Чтобы точнее определить систематическое положение багамской громии, исследователи секвенировали РНК малой субъединицы рибосомы. (Рибосомные РНК часто используются для построения филогенетических деревьев. Рибосомы — машины белкового синтеза — есть в каждой клетке, молекулы РНК, входящие в их состав, хорошо изучены, так что, сравнивая их нуклеотидные последовательности, можно делать выводы о родстве между организмами.) Перед выделением рибосомной РНК шарик сначала «схлопнули», чтобы избежать разведения клеточного материала. Новое существо действительно оказалось ближайшим родичем арабской громии.
Но еще интереснее самой амебы — ее следы. Дело в том, что они чрезвычайно напоминают следы докембрийского периода, возраст древнейших из которых — 1,8 млрд. лет. Напомним, что в кембрии около 540 млн. лет назад произошел «взрыв видообразования». Палеонтологическая летопись показывает, что именно тогда — и очень быстро, по палеонтологическим меркам — возникает как бы из ниоткуда множество разнообразных форм многоклеточных организмов.
 Как замечают палеонтологи Стефан Бенгтсон и Биргер Расмуссен в статье для «Сайенс», посвященной багамской громии, следы и останки животных превращаются в окаменелости в совершенно разных условиях, поэтому обычно в руки ученых попадает либо след, либо животное, но не то и другое вместе.
Как замечают палеонтологи Стефан Бенгтсон и Биргер Расмуссен в статье для «Сайенс», посвященной багамской громии, следы и останки животных превращаются в окаменелости в совершенно разных условиях, поэтому обычно в руки ученых попадает либо след, либо животное, но не то и другое вместе.Так вот, следы багамской громии удивительно напоминают докембрийские отпечатки. При этом животное, во-первых, одноклеточное, во-вторых, шарообразное. Как же громия перемещается и почему оставляет странный след с валиком на дне?
Предполагаемый способ передвижения громии лучше всего описывает выражение «катится колбасой»: длинная ось «виноградины» ориентирована перпендикулярно следу, и вокруг этой оси она вращается. Движется она, скорее всего, с помощью тонких выростов цитоплазмы (филоподий). А средний валик, возможно, образуется из «переработанного» грунта: амеба извлекает из него питательные вещества и затем сбрасывает позади себя.
Практически нет сомнений в том, что двухколейные следы оставляет именно амеба, коль скоро шарики всегда находятся на конце следа. К сожалению, пока не удалось зафиксировать процесс перемещения по простой причине: движутся они очень медленно. Одна из амеб была найдена рядом с кораллом, растущим на скорлупке морского ежа (см. фото). Судя по размерам и положению коралла, грунт оставался нетронутым в течение нескольких лет, значит, возраст следа может составлять недели, если не месяцы.
К сожалению, пока не удалось зафиксировать процесс перемещения по простой причине: движутся они очень медленно. Одна из амеб была найдена рядом с кораллом, растущим на скорлупке морского ежа (см. фото). Судя по размерам и положению коралла, грунт оставался нетронутым в течение нескольких лет, значит, возраст следа может составлять недели, если не месяцы.
Так или иначе, находка расширила наши представления о том, каким мог быть мир докембрия. Возможно, таинственные предки многоклеточных были похожи на громию. А вдруг современные громии — «живые ископаемые», прямые потомки докембрийских животных, не слишком изменившиеся с тех давних времен? Это было бы еще интереснее.
Что еще можно почитать о багамской Gromia sphaerica и ее следах:
- Stefan Bengtson, Birger Rasmussen. New and ancient trace makers // Science, 2009, January 16, v. 323, p. 346–347.
- Mikhail V. Matz, Tamara M. Frank, N. Justin Marshall, Edith A. Widder, Sönke Johnsen. Giant deep-sea protist produces bilaterian-like traces // Current biology, 2008, v.
 18, p. 1849.
18, p. 1849.
Хомни, или Захват Земли новым сверхорганизмом
- Гея Винс
- BBC Future
Подпишитесь на нашу рассылку ”Контекст”: она поможет вам разобраться в событиях.
Автор фото, Getty
Эволюционирует ли наш биологический вид до какого-то нового уровня? Корреспондент BBC Future выдвигает провокационный тезис: человечество преображается, и последствия этого процесса для жизни на нашей планете уже очевидны.
В древнегреческой мифологии у богини Земли Геи было девять сыновей-титанов, которые попытались захватить власть не только над Землей, но и над всей Вселенной. Знакомьтесь — еще один титан. Это новое существо. Оно появилось на свет лишь несколько десятилетий назад, но уже так же сильно влияет на жизнь планеты, как и фитопланктон или леса, регулирующие температуру, погоду и состав воздуха, которым мы дышим.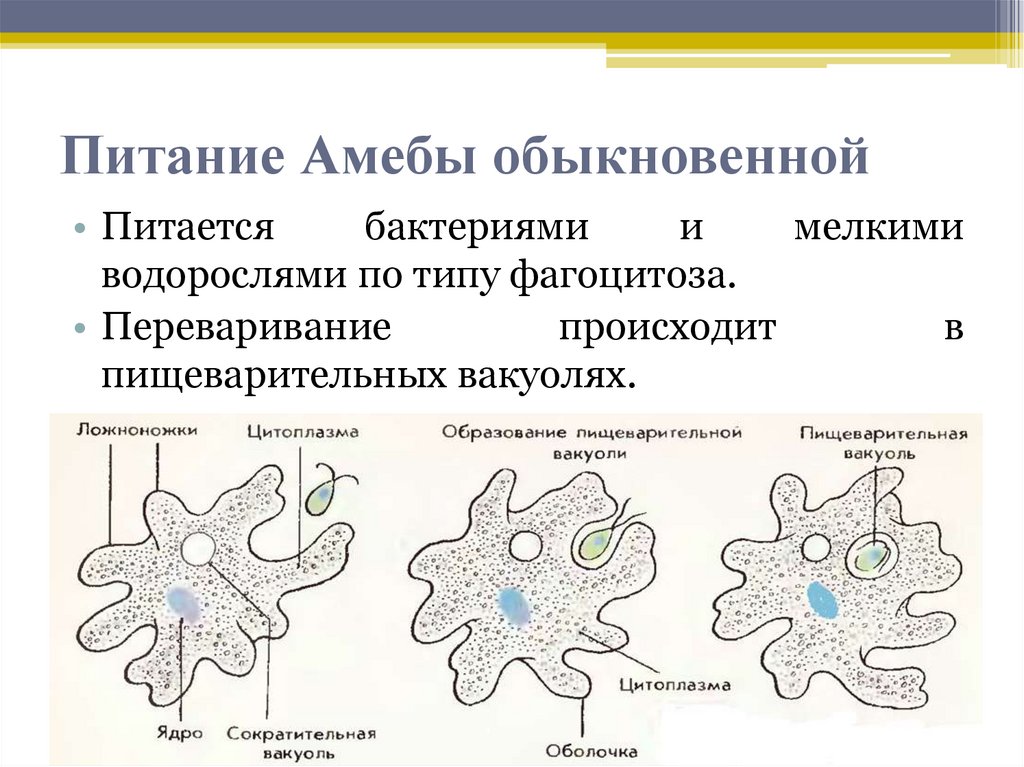
Это новое существо — мы или, точнее, то, во что превращается человечество. Весь наш биологический вид, Homosapiens, эволюционирует до уровня некоего сверхорганизма; назовем его Homoomnis, или «хомни».
Мы уже являемся основной силой, формирующей нашу планету. Говорят, что в результате наших действий началась новая геологическая эпоха — антропоцен, или “эпоха человека”. Хомни — продукт этой эпохи, продукт индустриализации, роста населения, глобализации и революции в сфере коммуникационных технологий; и этот продукт обладает огромной силой.
Хомни может воздействовать на биосферу и имеет свои потребности: сегодня в каждый момент времени он использует 18 тераватт (триллионов ватт) электроэнергии, тратит 9 000 миллиардов кубометров воды в год, занимает 40% общей площади суши под сельскохозяйственные угодья и расходует множество других природных ресурсов и полезных ископаемых.
Только время покажет, станет ли хомни ответственным пользователем нашей планеты или чудовищем, уничтожающим жизнь, а с ней и самого себя. Но некоторые признаки заметны уже сейчас, и в этой статье мы рассмотрим, кто такой хомни и как он влияет на наш биологический род, на планету и на остальных обитателей Земли.
Но некоторые признаки заметны уже сейчас, и в этой статье мы рассмотрим, кто такой хомни и как он влияет на наш биологический род, на планету и на остальных обитателей Земли.
Сила толпы
Чтобы разобраться в природе хомни, рассмотрим сначала один из простейших и древнейших одноклеточных организмов — амебу, которую называют слизевиком. Она появилась в процессе эволюции около 600 миллионов лет назад и расселилась в почве по всему миру, от Антарктики до Арктики. Большую часть своего жизненного цикла она ведет обычную жизнь простой амебы. Но иногда эти отдельные клетки собираются тысячами, образуя единый организм, который может передвигаться ползком, пульсировать, отращивать щупальца и даже преодолевать препятствия.
Автор фото, Thinkstock
Подпись к фото,Человечество — плесень на поверхности Земли?
Пропустить Подкаст и продолжить чтение.
Подкаст
Что это было?
Мы быстро, просто и понятно объясняем, что случилось, почему это важно и что будет дальше.
эпизоды
Конец истории Подкаст
Ученые назвали слизевиков моделью социума, наблюдая за тем, как слаженно действуют отдельные амебы, стремясь к общей цели, во имя которой они даже иногда жертвуют собой. К примеру, если в почве становится мало пищи, амебы соединяются, образуя щупальце, которое выползает на свет. Как только щупальце достигает поверхности, часть амеб формируют над землей стебелек, создавая из своих тел твердую целлюлозную оболочку и погибая при этом. Затем остальные амебы перемещаются по стебельку наверх и ждут где-нибудь в капле, пока мимо не пройдет какое-нибудь животное, с помощью которого можно добраться до новых земель. И все это делает простейший организм.
Человеческий мозг чем-то похож на такого слизевика. Отдельные клетки мозга — нейроны — нельзя назвать мыслящими или разумными, но когда все 86 миллиардов нейронов функционируют как единый организм, человеческий мозг превращается в нечто гораздо большее, чем просто совокупность его составных элементов. Он способен мыслить и находить нестандартные способы обработки информации.
Он способен мыслить и находить нестандартные способы обработки информации.
Мы до сих пор не понимаем, где в этой системе зарождаются и развиваются мысли, черты характера или особенности поведения, и как происходит организация нейронов для стимулирования подобных процессов, но каким-то образом из самых обычных материалов формируется сознание.
Интеллект, творческий потенциал и коммуникабельность хомни можно сравнить с диалоговой системой, состоящей из взаимосвязанных мозгов всех людей, в том числе живших ранее и оставивших культурное и интеллектуальное наследие, а также искусственных «мозгов» наших технических изобретений, таких как компьютерные программы и «Википедия». Хомни — это глобальная сеть цивилизаций, через которую уже передается поток знаний, направленный на защиту человечества. Как скворцы всей стаей неожиданно меняют направление полета, так и хомни вдруг начинает себя вести непредсказуемо.
Но это лишь одна сторона медали.
«Искусственный человек»
Не так давно я была в Белизе и, сидя в джунглях, наблюдала за работой колонии огненных муравьев. Каждый из них по отдельности — грозное создание длиной в несколько сантиметров, которое сильно кусается. Но вместе они образуют, по выражению американского энтомолога Э. О. Уилсона, «сверхорганизм» из тысяч слаженно действующих муравьев, которые систематично пожирают все на своем пути, уничтожая деревья и посевы, сгрызая изоляцию с проводов и другого электронного оборудования.
Каждый из них по отдельности — грозное создание длиной в несколько сантиметров, которое сильно кусается. Но вместе они образуют, по выражению американского энтомолога Э. О. Уилсона, «сверхорганизм» из тысяч слаженно действующих муравьев, которые систематично пожирают все на своем пути, уничтожая деревья и посевы, сгрызая изоляцию с проводов и другого электронного оборудования.
В некоторых местах муравьи даже «крышуют» тлю и жуков, которые взамен носят им нектар. Кроме того, сверхорганизм нашел способ выживания в условиях тропических ливней — когда начинается потоп, муравьи цепляются друг за друга, образуя «спасательный плот».
Автор фото, Getty
Подпись к фото,Муравьи тоже умеют «крышевать» чужой бизнес
Истощив все имеющиеся ресурсы, муравьиная община перебирается на новое место. Муравьи за 50 с лишним миллионов лет до людей наладили полноценное хозяйство: они выращивают и доят «скот» (тлю), строят замысловатые жилища, держат рабов и ведут масштабные бои с применением психологического оружия.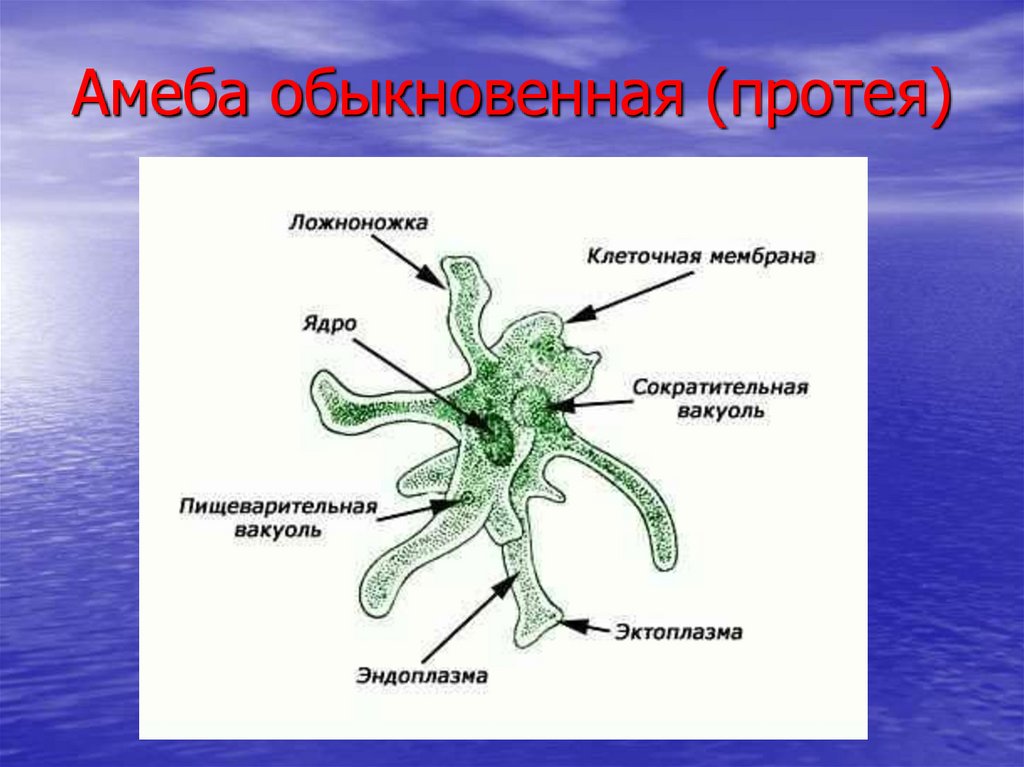 Сравнивать их с людьми можно до бесконечности.
Сравнивать их с людьми можно до бесконечности.
Применить понятие сверхорганизма к людям несколько сложнее — по отдельности мы более самостоятельны, чем клетки и даже муравьи. Однако за прошедшие века этот механизм представал в самых разных образах.
Древние греки полагали, что все мы являемся клетками высшего существа. Некоторые уподобляли задачи, исполняемые каждым из нас в обществе, — такие как выработка энергии или добыча пищи — функциям отдельных органов тела.
Английский философ Томас Гоббс, живший в XVII веке, писал, что общество образует «искусственного человека», который функционирует (через цивилизацию) таким образом, чтобы обеспечить собственное выживание.
Вступая в эпоху антропоцена, мы наблюдаем практическое воплощение этих представлений с беспрецедентным размахом. Отдельные люди или общества на местном или региональном уровне могут влиять на ландшафт, водоемы или разнообразные формы жизни, но наш сверхвид действует в планетарных масштабах.
Автор фото, AP
Подпись к фото,Ледники тают не просто так. Готов ли новый сверхорганизм взять на себя ответственность за планету?
Хомни уже контролирует три четверти запасов пресной воды на Земле, занимает более трех четвертей поверхности суши, свободной ото льда, и оказывает воздействие на атмосферу, различные формы жизни, а также на химические и биологические свойства океана.
Хомни даже уже начали засорять космос телескопами, спутниками и прочим искусственным мусором. Воздействие хомни на окружающую среду подобно наступлению аргентинских муравьев в глобальном масштабе.
В современном обществе отдельные люди все меньше и меньше способны функционировать независимо друг от друга, для нас все делает сверхорганизм: питает, одевает, снабжает энергией множество наших приборов, информирует и лечит нас и даже помогает нам размножаться за счет суррогатного материнства и ЭКО. В ближайшие десятилетия доступ к интернету, вероятно, уже будет обеспечен во всех уголках земного шара, мы еще больше интегрируемся в сетевое общество, а те, кто останется за пределами нового сверхорганизма, окажутся в культурной и технологической изоляции.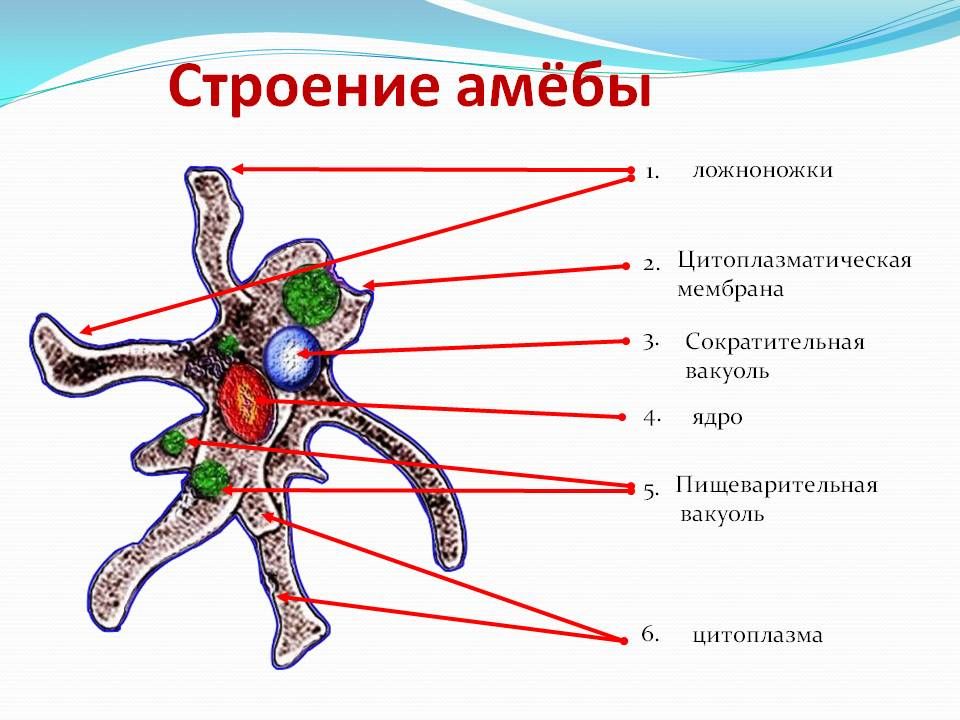
Кроме того, в ближайшие десятилетия доступ к электроэнергии, санитарно-гигиеническим средствам и антибиотикам станет почти всеобщим, за счет чего человек в эпоху антропоцена в среднем будет жить вдвое дольше и иметь базовые представления о научных, геополитических, культурных и социальных факторах, движущих миром. Итогом станет настолько новый образ жизни, что это будет практически равносильно появлению нового подвида. Да, антропоцен и хомни являются продуктом деятельности современного человека, но теперь уже хомни создает нового человека.
Чудовищные проблемы
И здесь возникает интересный парадокс. Возможно, люди в процессе эволюции прошли этап естественного отбора, пережив менее сильных конкурентов, но, как говорит австралийский палеонтолог Тим Фланнери, в результате сформировался не «мир, живущий по законам джунглей», а дружное сообщество.
Фланнери считает, что в данный момент формируется взаимозависимое глобальное общество, живущее на основе общих убеждений, — некая «цивилизация идей», благодаря которой на Земле воцарится справедливость и экологическое благоденствие.
Автор фото, Getty
Подпись к фото,Оптимисты считают, что человечество рано или поздно превратит Землю в цветущий сад…
Таков оптимистичный прогноз развития хомни, основанный на том, что большинство людей стремятся ладить друг с другом и возделывать окружающую среду. Прав ли Фланнери в своих альтруистичных суждениях о человечестве, и если да, то в какой степени, — по-прежнему большой вопрос.
Отдельные люди могут до некоторого уровня оказывать влияние на хомни, примерно как отдельный нейрон подает сигнал, рождающий мысль, результатом которой становится картина, песня или изобретение чего-нибудь вроде iPod. Или как человек пишет одно сообщение в «Твиттере», которое потом превращается в мем и начинает видоизменяться и расходиться по интернету. Считается, что для достижения “переломного момента” требуется не так много: достаточно, чтобы новое убеждение поддержали 10% людей, и оно распространится на большинство населения.
Но хотя отдельные личности, может быть, и будут способны оказывать на хомни определенное влияние, сейчас совершенно не ясно, что и как надо делать, чтобы выжить в эпоху антропоцена.
Воздействие хомни уже заметно по изменениям планетарного масштаба, подобных которым не было уже несколько миллионов лет и которые влияют на людей и на наши взаимоотношения в природном мире. Мы меняем климат, увеличивая количество выбросов СО2 в атмосферу, мы сокращаем биологическое разнообразие Земли, вызывая опасения ученых о потенциальном шестом за всю историю планеты массовом уничтожении отдельных биологических видов, мы “благоустраиваем” сушу, возводя на ней гигантские города из бетона, стали и стекла.
Если отдельный человек принимает решение о сокращении потребления пресной воды или о более ответственном отношении к окружающей среде, это почти никак не влияет на состояние рек или температуру воздуха во всем мире.
Хомни — это, по сути, всего лишь понятие, которое я придумала, чтобы показать: наш сверхорганизм обладает качествами, превосходящими качества просто скопления людей. В других примерах из животного мира, например, в пчелиных ульях или муравейниках, смерть матки означает гибель всей колонии. Однако у хомни нет отдельной матки, поэтому этот организм так же силен, как целый лес. Для того чтобы уничтожить хомни, оставив лишь горстку людей, потребовалась бы катастрофа планетарного масштаба: эпидемия, колоссальное изменение экологической обстановки, падение астероида или ядерная война.
Однако у хомни нет отдельной матки, поэтому этот организм так же силен, как целый лес. Для того чтобы уничтожить хомни, оставив лишь горстку людей, потребовалась бы катастрофа планетарного масштаба: эпидемия, колоссальное изменение экологической обстановки, падение астероида или ядерная война.
Однако нашим первым побуждением всегда будет укреплять хомни, а не уничтожать его — мы созданы для сотрудничества в обществе; большинство последних открытий, изобретений и прочих достижений является результатом труда целых коллективов.
Секрет улучшения условий жизни в антропоцене состоит в признании власти человека, но одновременно и в поддержании в нем человечности: стремления к налаживанию отношений с соседями и к охране окружающей среды. Помогут ли в этом крупные международные соглашения? Возможно.
Я считаю, что в будущем, и это подтверждают снимки с множества мощных спутников и дистанционно управляемых камер, которые позволяют отслеживать отдельные деревья в джунглях или определять степень таяния ледяной корки, мы начнем понимать хомни и более точечно контролировать его действия.
Может быть, после этого мы станем использовать потенциал хомни для восстановления планеты и заботы о ней. В противном случае последствия могут быть чудовищными.
Прочитать оригинал этой статьи на английском языке можно на сайте BBC Future.
Взгляд на образ жизни амебы Willaertia magna во время выращивания в биореакторе с использованием транскриптомики и протеомики М.В., Бурки Ф. и соавт. Изменения в классификации, номенклатуре и разнообразии эукариот. Дж. Эукариот. микробиол. 2018: jeu.12691. doi: 10.1111/jeu.12691. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2. Bertelli C., Greub G. Боковые обмены генами формируют геномы микроорганизмов, устойчивых к амебам. Передний. Клетка. Заразить. микробиол. 2012; 2 doi: 10.3389/fcimb.2012.00110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3. Панек Т., Чепичка И. Разнообразие Heterolobosea. Жене. Дайверы. Микроорг. 2012 г.: 10.5772/35333. [CrossRef] [Google Scholar]
[CrossRef] [Google Scholar]
4. de Jonckheere J.F., Dive D.G., Pussard M., Vickerman K. Willaertia Magna gen. ноябрь, сп. ноябрь (Vahlkampfiidae), теплолюбивая амеба, встречающаяся в различных средах обитания. [(по состоянию на 12 августа 2018 г.)]; Доступно в Интернете: https://eurekamag.com/research/001/281/001281223.php
5. Robinson B.S., Christy P.E., De Jonckheere J.F. Временная стадия жгутиконосца (мастигота) у вахлкампфиидной амебы Willaertia magna и ее возможное эволюционное значение. Биосистемы. 1989; 23: 75–86. doi: 10.1016/0303-2647(89)
-5. [PubMed] [CrossRef] [Google Scholar]
6. Клархольм М. Выпас простейших бактерий в почве — влияние и важность. микроб. Экол. 1981; 7: 343–350. doi: 10.1007/BF02341429. [PubMed] [CrossRef] [Google Scholar]
7. Родригес-Сарагоса С. Экология свободноживущих амеб. крит. Преподобный Микробиолог. 1994;20:225–241. doi: 10.3109/10408419409114556. [PubMed] [CrossRef] [Google Scholar]
8. Греуб Г., Рауль Д. Микроорганизмы, устойчивые к свободноживущим амебам. клин. микробиол. 2004; 17:413–433. doi: 10.1128/CMR.17.2.413-433.2004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
клин. микробиол. 2004; 17:413–433. doi: 10.1128/CMR.17.2.413-433.2004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Rowbotham T.J. Предварительный отчет о патогенности Legionella pneumophila для пресноводных и почвенных амеб. Дж. Клин. Патол. 1980; 33: 1179–1183. doi: 10.1136/jcp.33.12.1179. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Роуботэм Т.Дж. Выделение Legionella pneumophila из клинических образцов через амеб и взаимодействие тех и других изолятов с амебами. Дж. Клин. Патол. 1983; 36: 978–986. doi: 10.1136/jcp.36.9.978. [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Стаут Дж.Э., Ю.В.Л. Легионеллез. Н. англ. Дж. Мед. 1997; 337: 682–687. doi: 10.1056/NEJM199709043371006. [PubMed] [CrossRef] [Google Scholar]
12. Барбари Дж. М., Филдс Б. С., Фили Дж. К., Горман Г. В., Мартин В. Т. Выделение простейших из воды, связанное со вспышкой легионеллеза, и демонстрация внутриклеточного размножения Legionella pneumophila. заявл. Окружающая среда. микробиол. 1986;51:422–424. doi: 10.1128/AEM.51.2.422-424.1986. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Scheikl U., Sommer R., Kirschner A., Rameder A., Schrammel B., Zweimüller I., Wesner W., Hinker M. , Валочник Дж. Свободноживущие амебы (FLA), встречающиеся вместе с легионеллами в промышленных водах. Евро. Дж. Протистол. 2014; 50:422–429. doi: 10.1016/j.ejop.2014.04.002. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
14. Dupuy M., Binet M., Bouteleux C., Herbelin P., Soreau S., Héchard Y. Допустимость свежевыделенных экологических штаммов амеб для роста Legionella pneumophila. ФЭМС микробиол. лат. 2016;363:fnw022. дои: 10.1093/женский/fnw022. [PubMed] [CrossRef] [Google Scholar]
15. Килвингтон С., Стивенс С., Эберт Ф., Мишель Р., Бичинг Дж. Р. Сравнительное исследование Willaertia magna (свободноживущая амеба) из разных географических районов с использованием целых полиморфизм длин рестрикционных фрагментов рРНК и малых субъединиц рРНК. Дж. Протозол. Рез. 1995; 5: 97–107. [Google Scholar]
16. Линдер Дж.В.-К. Ewert Свободноживущие амебы, защищающие легионеллу в воде: верхушка айсберга? Сканд. Дж. Заразить. Дис. 1999; 31: 383–385. дои: 10.1080/00365549950163833. [PubMed] [CrossRef] [Google Scholar]
17. Cirillo J.D., Falkow S., Tompkins L.S. Рост Legionella pneumophila в Acanthamoeba castellanii усиливает инвазию. Заразить. Иммун. 1994;62:3254–3261. doi: 10.1128/IAI.62.8.3254-3261.1994. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Делафонт В., Родье М.-Х., Мезоннев Э., Като Э. Vermamoeba vermiformis: интересующая свободноживущая амеба. микроб. Экол. 2018;76:991–1001. doi: 10.1007/s00248-018-1199-8. [PubMed] [CrossRef] [Академия Google]
19. Dey R., Bodennec J., Mameri M.O., Pernin P. Свободноживущие пресноводные амебы различаются по своей восприимчивости к патогенной бактерии Legionella pneumophila . ФЭМС микробиол. лат. 2009; 290:10–17. doi: 10.1111/j.1574-6968. 2008.01387.x. [PubMed] [CrossRef] [Google Scholar]
20. Hasni I., Jarry A., Quelard B., Carlino A., Eberst J.-B., Abbe O., Demanèche S. Внутриклеточное поведение трех Legionella pneumophila Штаммы из трех штаммов амеб, включая Willaertia magna C2c Maky. Возбудители. 2020;9:105. doi: 10.3390/pathogens
05. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]21. Amoeba|Biocide by Nature. [(по состоянию на 7 января 2020 г.)]; Доступно в Интернете: http://www.amoeba-biocide.com/fr
22. Нефф Р.Дж. Очистка, аксеническое культивирование и описание почвенной амебы, Acanthamoeba sp. Дж. Протозол. 1957; 4: 176–182. doi: 10.1111/j.1550-7408.1957.tb02505.x. [CrossRef] [Google Scholar]
23. Weekers P.H.H., Vogels G.D. Аксеническое культивирование свободноживущих амеб, Acanthamoeba castellanii и Hartmannella vermiformis в хемостате. Дж. Микробиол. Методы. 1994;19:13–18. doi: 10.1016/0167-7012(94)
-3. [CrossRef] [Google Scholar] 24. Beshay U. , Friehs K., Azzam A.-E.-M., Flaschel E. Анализ поведения Dictyostelium discoideum в иммобилизованном состоянии с помощью непрерывного культивирования. Биопроцесс. Биосист. англ. 2003; 26: 117–122. doi: 10.1007/s00449-003-0339-8. [PubMed] [CrossRef] [Google Scholar]
25. Mimouni V., Ulmann L., Pasquet V., Mathieu M., Picot L., Bougaran G., Cadoret J.-P., Morant-Manceau A. , Schoefs B. Потенциал микроводорослей для производства биологически активных молекул, представляющих фармацевтический интерес. Курс. фарм. Биотехнолог. 2012;13:2733–2750. дои: 10.2174/138920112804724828. [PubMed] [CrossRef] [Google Scholar]
26. Newman J.D., Marshall J., Chang M., Nowroozi F., Paradise E., Pitera D., Newman K.L., Keasling J.D. Высокоуровневое производство аморфы -4,11-диен в биореакторе с двухфазным разделением метаболически сконструированной кишечной палочки. Биотехнолог. биоинж. 2006; 95: 684–691. doi: 10.1002/bit.21017. [PubMed] [CrossRef] [Google Scholar]
27. Папаспириди Л. -М., Алигианнис Н., Кристакопулос П., Скальтсунис А.-Л., Фокиалакис Н. Производство биологически активных метаболитов, представляющих фармацевтический и нутрицевтический интерес, путем погружения в воду. ферментация Pleurotus ostreatus в биореакторе периодического действия с мешалкой. Procedia Food Sci. 2011; 1:1746–1752. doi: 10.1016/j.profoo.2011.09.257. [CrossRef] [Google Scholar]
28. Хасни И., Челха Н., Баптист Э., Мамери М.Р., Лачуер Дж., Плассон Ф., Колсон П., Скола Б.Л. Изучение потенциальной патогенности Willaertia magna путем исследования переноса генов патогенности бактерий в ее геном. науч. Отчет 2019; 9: 1–12. doi: 10.1038/s41598-019-54580-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Ван З., Герштейн М., Снайдер М. RNA-Seq: революционный инструмент для транскриптомики. Нац. Преподобный Жене. 2009 г.;10:57–63. doi: 10.1038/nrg2484. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Ломсадзе А., Бернс П.Д., Бородовский М. Интеграция картированных считываний RNA-Seq в автоматическое обучение алгоритма поиска эукариотических генов. Нуклеиновые Кислоты Res. 2014;42:e119. doi: 10.1093/nar/gku557. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Vogel C., Marcotte E.M. Взгляд на регуляцию содержания белка на основе протеомного и транскриптомного анализов. Нац. Преподобный Жене. 2012;13:227–232. doi: 10.1038/nrg3185. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Фриц-Лейлин Л.К., Прочник С.Е., Джинджер М.Л., Дакс Дж.Б., Карпентер М.Л., Филд М.С., Куо А., Паредес А., Чепмен Дж., Фам Дж. и др. Геном Naegleria gruberi проливает свет на многогранность ранних эукариот. Клетка. 2010; 140:631–642. doi: 10.1016/j.cell.2010.01.032. [PubMed] [CrossRef] [Google Scholar]
33. Opperdoes F.R., De Jonckheere J.F., Tielens A.G.M. Метаболизм Naegleria gruberi. Междунар. Дж. Паразитол. 2011;41:915–924. doi: 10.1016/j.ijpara.2011.04.004. [PubMed] [CrossRef] [Академия Google]
34. Alsam S., Sissons J., Dudley R., Khan N.A. Механизмы, связанные с фагоцитозом Acanthamoeba castellanii (T4). Паразитол. Рез. 2005; 96: 402–409. doi: 10.1007/s00436-005-1401-z. [PubMed] [CrossRef] [Google Scholar]
35. Кларк М., Лохан А.Дж., Лю Б., Лагкувардос И., Рой С., Зафар Н., Бертелли К., Шильде К., Кианианмомени А., Бюрглин Т.Р. и др. Геном Acanthamoeba castellanii подчеркивает обширный латеральный перенос генов и раннюю эволюцию передачи сигналов тирозинкиназы. Геном биол. 2013;14:R11. doi: 10.1186/gb-2013-14-2-r11. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Хан Н.А. Акантамеба: Биология и растущее значение для здоровья человека. ФЭМС микробиол. 2006; 30: 564–595. doi: 10.1111/j.1574-6976.2006.00023.x. [PubMed] [CrossRef] [Google Scholar]
37. De Jonckheere J. Использование аксенической среды для дифференциации патогенных и непатогенных изолятов Naegleria fowleri. заявл. Окружающая среда. микробиол. 1977; 33: 751–757. doi: 10.1128/AEM. 33.4.751-757.1977. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Bioréacteur Pilot GPC. [(по состоянию на 21 марта 2020 г.)]; Доступно в Интернете: http://www.gpcbio.com/bioracteurpilot.html
39. Bolger A.M., Lohse M., Usadel B. Trimmomatic: гибкий триммер для данных последовательностей Illumina. Биоинформатика. 2014;30:2114–2120. doi: 10.1093/биоинформатика/btu170. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
40. Aherfi S., Andreani J., Baptiste E., Oumessoum A., Dornas F.P., Andrade A.C., Chabriere E., Abrahao J., Levasseur А., Рауль Д. и др. Большой открытый пангеном и малый основной геном для гигантских пандоравирусов. Передний. микробиол. 2018;9 doi: 10.3389/fmicb.2018.01486. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41. Ким Д., Лангмид Б., Зальцберг С.Л. HISAT: быстродействующий выравниватель с низкими требованиями к памяти. Нац. Методы. 2015;12:357–360. doi: 10.1038/nmeth.3317. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Андерс С., Пил П.Т., Хубер В. HTSeq — платформа Python для работы с высокопроизводительными данными секвенирования. Биоинформатика. 2015; 31: 166–169. doi: 10.1093/биоинформатика/btu638. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
43. Huerta-Cepas J., Forslund K., Coelho L.P., Szklarczyk D., Jensen L.J., von Mering C., Bork P. Fast Genome- Широкая функциональная аннотация с помощью назначения ортологии от eggNOG-Mapper. Мол. биол. Эвол. 2017;34:2115–2122. дои: 10.1093/молбев/msx148. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
44. Татусов Р.Л., Гальперин М.Ю., Натале Д.А., Кунин Е.В. База данных COG: инструмент для анализа функций и эволюции белков в масштабе генома. Нуклеиновые Кислоты Res. 2000; 28:33–36. doi: 10.1093/нар/28.1.33. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Канехиса М., Гото С. КЭГГ: Киотская энциклопедия генов и геномов. Нуклеиновые Кислоты Res. 2000; 28: 27–30. doi: 10.1093/нар/28.1.27. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Marchler-Bauer A., Derbyshire M.K., Gonzales N.R., Lu S., Chitsaz F., Geer L.Y., Geer R.C., He J., Gwadz M., Hurwitz D.I., et al. CDD: сохраненная база данных доменов NCBI. Нуклеиновые Кислоты Res. 2015;43:D222–D226. doi: 10.1093/nar/gku1221. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Хантер С., Апвайлер Р., Эттвуд Т.К., Байрох А., Бейтман А., Биннс Д., Борк П., Дас У., Догерти Л., Дюкен Л. и др. InterPro: Интегративная база данных сигнатур белков. Нуклеиновые Кислоты Res. 2009 г.;37:D211–D215. doi: 10.1093/nar/gkn785. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Бахманн Б.О., Равель Дж. Глава 8. Методы in silico прогнозирования путей биосинтеза микробных поликетидов и нерибосомных пептидов на основе данных о последовательности ДНК. Методы Энзимол. 2009; 458:181–217. doi: 10.1016/S0076-6879(09)04808-3. [PubMed] [CrossRef] [Google Scholar]
49. Ziemert N., Podell S., Penn K., Badger J.H., Allen E., Jensen P.R. Поиск доменов натуральных продуктов NaPDoS: основанный на филогенезе биоинформационный инструмент для классификации вторичных разнообразие генов метаболитов. ПЛОС ОДИН. 2012;7:e34064. doi: 10.1371/journal.pone.0034064. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Альмагро Арментерос Дж. Дж., Сальваторе М., Эмануэльссон О., Винтер О., фон Хейне Г., Элофссон А., Нильсен Х. Обнаружение сигналов последовательности в целевых пептидах с использованием глубокого обучения. Жизнь наук. Альянс. 2019; 2 doi: 10.26508/lsa.201
9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Prokisch H., Andreoli C., Ahting U., Heiss K., Ruepp A., Scharfe C., Meitinger T. MitoP2: The mitochondrial база данных протеома — теперь включает данные о мышах. Нуклеиновые Кислоты Res. 2006; 34:D705–D711. дои: 10.1093/нар/gkj127. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Эдгар Р.К. MUSCLE: множественное выравнивание последовательностей с высокой точностью и высокой пропускной способностью. Нуклеиновые Кислоты Res. 2004; 32: 1792–1797. doi: 10.1093/nar/gkh440. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Кумар С., Стечер Г., Тамура К. MEGA7: Молекулярно-эволюционный генетический анализ, версия 7.0 для больших наборов данных. Мол. биол. Эвол. 2016; 33:1870–1874. doi: 10.1093/molbev/msw054. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
54. Летуник И., Борк П. Интерактивное древо жизни (iTOL) v3: онлайн-инструмент для отображения и аннотации филогенетических и других деревьев. Нуклеиновые Кислоты Res. 2016;44:W242–W245. doi: 10.1093/nar/gkw290. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. He L., Diedrich J., Chu Y.-Y., Yates J.R. Извлечение точной информации о прекурсоре для тандемных масс-спектров с помощью RawConverter. Анальный. хим. 2015;87:11361–11367. doi: 10.1021/acs.analchem.5b02721. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
56. Ma B., Zhang K., Hendrie C., Liang C., Li M., Doherty-Kirby A., Lajoie G. PEAKS: мощное программное обеспечение для секвенирования пептидов de novo методом тандемной масс-спектрометрии. Быстрое общение. Масс-спектр. РКМ. 2003; 17: 2337–2342. doi: 10.1002/rcm.1196. [PubMed] [CrossRef] [Google Scholar]
57. McCafferty J., Reid R., Spencer M., Hamp T., Fodor A. Peak Studio: инструмент для визуализации и анализа файлов анализа фрагментов. Окружающая среда. микробиол. Отчет 2012; 4: 556–561. дои: 10.1111/j.1758-2229.2012.00368.х. [PubMed] [CrossRef] [Google Scholar]
58. Хан Н.А. Акантамеба: биология и патогенез. 2-е изд. Кэйстер Академик Пресс; Норфолк, Великобритания: 2018. [Google Scholar]
59. Отомо Т., Отомо К., Томчик Д.Р., Мачиус М., Розен М.К. Структурные основы опосредованной Rho GTPase активации формина mDia1. Мол. Клетка. 2005; 18: 273–281. doi: 10.1016/j.molcel.2005.04.002. [PubMed] [CrossRef] [Google Scholar]
60. Hug L.A., Stechmann A., Roger A.J. Филогенетическое распределение и история белков, участвующих в анаэробном метаболизме пирувата у эукариот. Мол. биол. Эвол. 2010;27:311–324. дои: 10.1093/молбев/msp237. [PubMed] [CrossRef] [Google Scholar]
61. Эльсбах П. Бактерицидный/увеличивающий проницаемость белок (BPI) в антибактериальной защите хозяина. Дж. Лейкок. биол. 1998; 64:14–18. doi: 10.1002/jlb.64.1.14. [PubMed] [CrossRef] [Google Scholar]
62. Cosson P., Lima W.C. Внутриклеточное уничтожение бактерий: является ли Dictyostelium модельным макрофагом или пришельцем?: Внутриклеточное уничтожение бактерий в Dictyostelium . Клетка. микробиол. 2014; 16:816–823. дои: 10.1111/см.12291. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. Winterbourn C.C., Kettle A.J. Окислительно-восстановительные реакции и уничтожение микробов в нейтрофильной фагосоме. Антиоксид. Окислительно-восстановительный сигнал. 2012; 18: 642–660. doi: 10.1089/ars.2012.4827. [PubMed] [CrossRef] [Google Scholar]
64. Hong Y., Kang J.-M., Joo S.-Y., Song S.-M., Lê HG, Thái T.L., Lee J., Goo Ю.-К., Чунг Д.-И., Сон В.-М. и др. Молекулярные и биохимические свойства цистеиновой протеазы Acanthamoeba castellanii. Корейский Дж. Параситол. 2018;56:409–418. doi: 10.3347/kjp.2018.56.5.409. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
65. Турк В., Стока В., Васильева О., Ренко М., Сун Т., Турк Б., Турк Д. Катепсины цистеина: из структура, функции и регулирование к новым границам. Биохим. Биофиз. Акта. 2012; 1824: 68–88. doi: 10.1016/j.bbapap.2011.10.002. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
66. Миянага А. Структура и функция ферментов биосинтеза поликетидов: различные стратегии производства структурно разнообразных поликетидов. Бионауч. Биотехнолог. Биохим. 2017;81:2227–2236. дои: 10.1080/09168451.2017.1391687. [PubMed] [CrossRef] [Google Scholar]
67. Робинсон Дж.А. Комплексы поликетидсинтазы: их структура и роль в биосинтезе антибиотиков. Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 1991; 332: 107–114. doi: 10.1098/rstb.1991.0038. [PubMed] [CrossRef] [Google Scholar]
68. Сингх Д.Р. БИОДЕГРАДАЦИЯ КСЕНОБИОТИКОВ – ПУТЬ ДЕТОКСИКАЦИИ ОКРУЖАЮЩЕЙ СРЕДЫ. Междунар. Дж. Дев. Рез. 2017;7:14082–14087. [Google Scholar]
69. Миранда Э.Р., Нам Э.А., Куспа А., Шаульский Г. Транспортер ABC, AbcB3, опосредует экспорт цАМФ в развитии D. discoideum. Дев. биол. 2015;397: 203–211. doi: 10.1016/j.ydbio.2014.11.006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
70. Eichinger L., Pachebat J.A., Glöckner G., Rajandream M.-A., Sucgang R., Berriman M., Song J., Olsen Р., Шафрански К., Сюй К. и др. Геном социальной амебы Dictyostelium discoideum. Природа. 2005; 435:43–57. doi: 10.1038/nature03481. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
71. Brown S., Clastre M., Courdavault V., O’Connor S.E. Производство de novo растительного алкалоида стриктозидина в дрожжах. проц. Натл. акад. науч. США. 2015;112:3205–3210. doi: 10.1073/pnas.1423555112. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
72. Zysset-Burri D.C., Müller N., Beuret C., Heller M., Schürch N., Gottstein B., Wittwer M. Полногеномная идентификация факторов патогенности свободноживущей амебы Naegleria fowleri. Геномика BMC. 2014; 15 doi: 10.1186/1471-2164-15-496. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Liechti N., Schürch N., Bruggmann R., Wittwer M. Геном Naegleria lovaniensis, основа сравнительного подхода к выявлению факторов патогенности патогенной для человека амебы N. fowleri. Геномика BMC. 2018;19doi: 10.1186/s12864-018-4994-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
74. Хасни И., Андреани Дж., Колсон П., Ла Скола Б. Описание вирулентных факторов и горизонтального переноса генов амебы, связанной с кератитом Acanthamoeba Triangularis по анализу генома. Возбудители. 2020;9:217. doi: 10.3390/pathogens17. [Статья PMC бесплатно] [PubMed] [CrossRef] [Google Scholar]
75. Челха Н., Хасни И., Луазани А.С., Левассер А., Ла Скола Б., Колсон П. Vermamoeba vermiformis CDC-19черновая последовательность генома показывает значительный оборот генов, в том числе с кандидатами на типы радиации и гигантскими вирусами. науч. Отчет 2020; 10 doi: 10.1038/s41598-020-62836-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
76. Детеринг Х., Эбишер Т., Дабровски П.В., Радонич А., Ницше А., Ренар Б.Ю., Кидерлен А.Ф. Первый проект последовательности генома Balamuthia mandrillaris возбудитель амебного энцефалита. Объявление генома. 2015; 3 doi: 10.1128/genomeA.01013-15. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
77. Greninger A.L., Messacar K., Dunnebacke T., Naccache S.N., Federman S., Bouquet J., Mirsky D., Nomura Y., Yagi S., Glaser C., et al. Клиническая метагеномная идентификация энцефалита Balamuthia mandrillaris и сборка проекта генома: продолжающийся случай секвенирования эталонного генома. Геном Мед. 2015; 7 doi: 10.1186/s13073-015-0235-2. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
78. Fan M., Sun X., Xu N., Liao Z., Li Y., Wang J., Fan Y., Cui D. , Ли П., Мяо З. Интеграция анализа глубокого транскриптома и протеома салициловой кислоты, регулирующей высокотемпературный стресс у Ulva prolifera. науч. Отчет 2017; 7: 1–19.. doi: 10.1038/s41598-017-11449-w. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Lee M.V., Topper S.E., Hubler S.L., Hose J., Wenger C.D., Coon J.J., Gasch A.P. Динамическая модель изменений протеома раскрывает новые роли изменение транскрипции у дрожжей. Мол. Сист. биол. 2011;7:514. doi: 10.1038/msb.2011.48. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
80. де Соуза Абреу Р., Пенальва Л.О., Маркотт Э.М., Фогель С. Глобальные признаки уровней экспрессии белков и мРНК. Мол. Биосист. 2009 г.;5:1512–1526. doi: 10.1039/b908315d. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
81. Флетчер Д.А., Маллинс Р.Д. Клеточная механика и цитоскелет. Природа. 2010; 463:485–492. doi: 10.1038/nature08908. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
82. Текле Йонас И., Уильямс Джессика Р. Цитоскелетная архитектура и ее эволюционное значение у амебоидных эукариот и их способ передвижения. Р. Соц. Открытая наука. 2016;3:160283. doi: 10.1098/rsos.160283. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
83. Atteia A., van Lis R., Gelius-Dietrich G., Adrait A., Garin J., Joyard J., Rolland N., Martin W. Пируватформиат-лиаза и новый путь эукариотического синтеза АТФ в митохондриях хламидомонады. Дж. Биол. хим. 2006; 281:9909–9918. doi: 10.1074/jbc.M507862200. [PubMed] [CrossRef] [Google Scholar]
84. Barberà M.J., Ruiz-Trillo I., Tufts J.Y.A., Bery A., Silberman J.D., Roger A.J. Sawyeria marylandensis (Heterolobosea) имеет гидрогеносому с новыми метаболическими свойствами. Эукариот. Клетка. 2010;9: 1913–1924. doi: 10.1128/EC.00122-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
85. Цаусис А.Д., Нивлтова Э., Шутак Р., Хрди И., Тачези Дж. Производство немитохондриального водорода в Naegleria gruberi. Геном биол. Эвол. 2014; 6: 792–799. doi: 10.1093/gbe/evu065. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
86. Сиддики Р., Хан Н.А. Война микробных миров: кто выигрывает от взаимодействия между акантамёбами и бактериями? Эксп. Паразитол. 2012; 130:311–313. doi: 10.1016/j.exppara.2012.01.021. [PubMed] [CrossRef] [Академия Google]
87. Дене Э., Купа-Гуталанд Б., Назарет С., Пеландакис М., Фавр-Бонте С. Разнообразие свободноживущих амеб в почвах и связанных с ними условно-патогенных бактерий человека. Паразитол. Рез. 2017;116:3151–3162. doi: 10.1007/s00436-017-5632-6. [PubMed] [CrossRef] [Google Scholar]
88. Akya A., Pointon A., Thomas C. Механизм, участвующий в фагоцитозе и уничтожении Listeria monocytogenes полифагой Acanthamoeba. Паразитол. Рез. 2009;105:1375–1383. doi: 10.1007/s00436-009-1565-z. [PubMed] [CrossRef] [Академия Google]
89. Тишков В.И., Савин С.С., Ясная А.С. Белковая инженерия пенициллинацилазы. Acta Nat. 2010;2:47–61. doi: 10.32607/20758251-2010-2-3-47-61. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
90. Iqbal J., Siddiqui R., Khan N. A. Acanthamoeba и бактерии производят противомикробные препараты, нацеленные на своих аналогов. Паразит. Векторы. 2014;7:56. дои: 10.1186/1756-3305-7-56. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
91. Zucko J., Skunca N., Curk T., Zupan B., Long PF, Cullum J., Kessin R.H., Hranueli D. Поликетидсинтаза гены и потенциал натуральных продуктов Dictyostelium discoideum. Биоинформа. Оксф. англ. 2007; 23: 2543–2549. doi: 10.1093/биоинформатика/btm381. [PubMed] [CrossRef] [Google Scholar]
92. Rozgaj R. [Микробная деградация ксенобиотиков в окружающей среде] Arh. высокий Рада Токсикол. 1994; 45: 189–198. [PubMed] [Google Scholar]
93. Janssen DB, Dinkla IJT, Poelarends GJ, Terpstra P. Бактериальная деградация ксенобиотических соединений: эволюция и распределение активности новых ферментов. Окружающая среда. микробиол. 2005; 7: 1868–1882. doi: 10.1111/j.1462-2920.2005.00966.x. [PubMed] [CrossRef] [Академия Google]
94. Ли В.В., Ли Дж.Дж. Моделирование и анализ данных секвенирования РНК: обзор со статистической точки зрения. Квант. биол. Пекин, Китай. 2018;6:195–209. doi: 10.1007/s40484-018-0144-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
95. Манга П., Клингеман Д.М., Лу Т.-Ю.С., Мельхорн Т.Л., Пеллетье Д.А., Хаузер Л.Дж., Уилсон С.М., Браун С.Д. Репликаты, числа считывания и другие важные соображения по планированию эксперимента для микробной РНК-секвенции, идентифицированной с использованием наборов данных Bacillus thuringiensis. Передний. микробиол. 2016;7 doi: 10.3389/fmicb.2016.00794. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Schurch NJ, Schofield P., Gierliński M., Cole C., Sherstnev A., Singh V., Wrobel N., Gharbi K., Симпсон Г.Г., Оуэн-Хьюз Т. и др. Сколько биологических повторов необходимо в эксперименте RNA-seq и какой инструмент дифференциальной экспрессии следует использовать? РНК. 2016; 22: 839–851. doi: 10.1261/РНК.053959.115. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
97. Rapaport F., Khanin R., Liang Y., Pirun M., Krek A. , Zumbo P., Mason C.E., Socci N.D., Betel D. Комплексная оценка методов дифференциального анализа экспрессии генов для данных секвенирования РНК. Геном биол. 2013;14:3158. doi: 10.1186/ru-2013-14-9-р95. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
98. Liu Y., Zhou J., White K.P. Исследования дифференциальной экспрессии РНК-секвенций: больше последовательности или больше репликации? Биоинформатика. 2014;30:301–304. doi: 10.1093/биоинформатика/btt688. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Проверка здравомыслия: «Интервьюирование» амебы относительно цели жизни
Как я упоминал в предыдущих блогах, однажды я занимался юмором/ обозреватель образа жизни Dayton Daily News. Главным редактором, подписавшим контракт с этим концертом, был Джефф Брюс, который сейчас пишет детектив, серьезно посвященный, ну, обозревателю (чья колонка сильно отличается от моей). . Джефф также недавно брал у меня интервью для своего книжного клуба; вы можете посмотреть это интервью здесь.
Разговор с Джеффом навеял приятные воспоминания о моей колонке «Проверка здравомыслия», которая выходила каждый понедельник в разделе «Жизнь» Dayton Daily News в течение десяти с половиной лет.
Размышляя об этом и о наших нынешних трудных временах, я вспомнил еще одно интервью, которое я дал для своей колонки. Я взял интервью — ну, я вообразил, что взял интервью — у амебы. Нам с амебой было так много о чем поговорить, что наши обсуждения превратились в интервью, состоящее из двух частей.
Вот обе колонки для вашего удовольствия и, возможно, для вдохновения.
Интервью с амебой
Недавно я наткнулся на новостной ролик о научном эксперименте с участием амеб.
Любители амеб, не волнуйтесь; этот научный эксперимент не включал испытания лекарств или подводки для глаз на амебах, или введение им болезнетворных штаммов, ну, других амеб, или лишение амебы пищи или сна амебы.
На самом деле ученые создали идеальную среду для амебы. Итак, что случилось с этими амёбами, плавающими в идеальной среде без стресса?
Они умерли.
Просто опустились на дно своих идеальных маленьких чашек Петри с 5 звездами и тут же перевернулись.
Я точно не знаю, как переворачивается капля, меняющая форму, но результаты этого удивительно жестокого эксперимента впечатляют; на самом деле настолько захватывающим, что я хотел бы взять интервью у одной из этих амеб, когда она опускалась на дно, ее маленькие ложноножки безнадежно размахивали.
Я: Извините, мистер — или это мисс Амеба —
Амеба (вздыхает): Не надо формальностей. Зови меня просто Амеба.
Я: О, хорошо. Так что, Амеба, я не понимаю. Почему ты и все твои приятели-амебы просто сдаешься и умираешь? У вас было все, что вы могли бы хотеть или в чем нуждаться!
Амеба: Звучит здорово, не так ли? Это то, о чем мы все думали, когда записывались на этот концерт. И да, у нас было все это. Идеальный солевой раствор. Не бойтесь столкнуться с хищным планктоном. Бактериальный буфет на любой вкус. Даже круглосуточный доступ к нашим любимым кабельным каналам. Все от CNN до BFN.
Я: CNN? БФН?
Амеба: Знаешь, новости Центрального ядра. Бинарная сеть деления. Хотя тот последний был поздно ночью, только для взрослых.
Я: Верно. Ну, если ваша жизнь была такой идеальной, что же случилось, что вы все начали умирать?
Амеба: Вот что случилось! Жизнь стала слишком идеальной. Отсутствие борьбы означает отсутствие проблем. Конечно, кому не нужно время от времени расслабляться, плавать и потягивать стакан Каберне из водорослей? Но через некоторое время… скучно.
Я: Итак, вы все умираете со скуки?
Амеба: Думаю, правильнее будет сказать, что мы умираем, потому что у нас отняли цель.
Я: Ваша цель состояла в том, чтобы избегать планктона, есть бактерии и разделиться с самим собой!
Амеба: Эй, не бей! Таким образом, мы, амебы, не очень хорошо разбираемся в промышленности или творчестве. У нас все еще есть основная цель. Когда эта цель была достигнута вместо нас вместо на нас, затем наша цель закончилась, и поэтому мы просто сдались.
Я: Хммм. Знаете, я вижу корреляцию между амебой и людьми. Людям тоже нужна цель.
Амеба: Угу. Нравится мучить амёб, создавая для них идеальные миры?
Я: Ты что, саркастичная разновидность амебы?
Амеба: На самом деле, я Pelomyxa palustris.
Я: Ой. Как бы то ни было, человеческая цель принимает различные формы — соревнование, творчество, поиск знаний, помощь другим. И есть множество способов, которыми эта цель проявляется.
Амеба: Включает ли это создание воображаемых разговоров с амебами?
Я: Эй, я многому научился из нашей небольшой беседы! Почему, если даже амебам нужно бросить вызов, чтобы достичь цели, тогда эта потребность должна быть частью всей жизни… подожди, куда ты идешь?
Амеба: Этот разговор был достаточно сложным, чтобы вдохновить меня. Забудьте о погружении на дно. Я возвращаюсь наверх и требую, чтобы бактериальный буфет был убран!
Я: Вы тоже избавляетесь от CNN и BFN?
Амеба: Не будь смешным.
Продолжение интервью с амебой
Всего два месяца назад я написал колонку о научном эксперименте с участием амеб, в котором амебы впали в уныние и безжизненно погрузились на дно чашек Петри, когда все испытали стресс. были удалены из их pH-сбалансированной среды.
Воображаемый разговор со свободной от стресса, но больной амебой (да, это именно то, что представляют себе писатели) показал, что сталкиваться с трудностями и преодолевать их необходимо, чтобы сделать жизнь достойной жизни.
Что ж, амебы снова в новостях.
Вы можете прочитать все подробности на сайте livecience.com, но основная история заключается в том, что в трудные времена d ictyostelium discoideum амебы, которые обычно предпочитают быть суровыми индивидуалистами, решают подружиться с другими dictyostelium discoideums . (Или это discoideii ? Во всяком случае…)
Когда условия жизни становятся особенно трудными (например, начинает заканчиваться бактериальный буфет, начинает размножаться планктон, питающийся амёбами, и т. д.), эти амебы объединяются, чтобы создать «сообщество». индивидуумов», чтобы стать многоклеточным организмом. Затем амебы либо становятся спорами, которые могут выживать, размножаться и передавать генетическую информацию будущим амёбам… либо около 20% из них становятся стеблями, которые поднимают споры над землей, чтобы они могли затем рассеяться к более благоприятным условиям жизни. условия.
Недостаток возвышающегося стебля? Амебы с этой обязанностью должны сначала умереть, прежде чем превратиться в стебли.
Это забавный научный факт, который, с одной стороны, может показаться довольно мрачной аналогией важности сообщества в эти трудные времена.
Но давайте изменим эту смерть-превращением-в-поднимающие-стебли, чтобы она стала чем-то более, ну, возвышающим, но все же жертвенным.
Было бы легкомысленно просто предположить, например, что можно подумать о том, чтобы немного меньше есть вне дома, чтобы пожертвовать на продовольственную кладовую, поскольку многие люди уже сокращают такие предметы роскоши, а также предметы первой необходимости, чтобы взять с собой заботиться о себе и своих близких.
На самом деле, давать другим в столь пугающие времена кажется почти нелогичным. Заманчиво отступить, уцепиться за то, что есть, даже спрятаться.
Но я думаю, что dictyostelium discoideums правы. В трудные времена, как никогда, важно объединиться. Чтобы помочь друг другу.
Конечно, эта помощь может принимать форму пожертвования товаров, услуг или финансов.
Но помогать друг другу можно и в форме улыбки. Еще немного терпения. Еще немного прослушивания. Разговор, которого обычно не бывает.
Более того, помощь другим не обязательно должна быть чем-то заметным. Это может быть молитва. Корректировка отношения. Пытаюсь увидеть людей… даже незнакомцев… особенно незнакомцев… в более добром свете.
Тот водитель, который подрезал вас сегодня утром? Ладно, может быть, он придурок. А может, он спешит на собеседование при приеме на работу или на помощь близкому человеку.
Та женщина в бакалейной лавке, которая встала в очередь за 15 или меньше с 15 или больше? Ладно, может быть, она придурок. Или, может быть, она настолько увлеклась проблемой, что честно не увидела знак 15 или меньше.
Рискуя перейти от аналогии с амебой к каламбуру с амебой, я закончу комментарием — я уверен, что вы поняли, кхм, дрейф — и вопросом: почему амебы внезапно стали новостями -достоин, что может быть более подходящим материалом для создания аналогий?
Я был высшей формой жизни на этой планете четыре с лишним десятилетия и никогда не думал об амебах.