67. Класс Корненожки, особенности строения и важнейшие представители. Цикл развития дизентерийной амебы. Диагностика, профилактика и распространение амебиаза.
Класс корненожки – саркодовые – Sarcodina Распространение – повсеместно, но в районах с жарким климатом встречается чаще.
Наиболее примитивные простейшие, что проявляется в слабой степени дифференцировки. Цитоплазма ограничена только наружной мембраной, форма тела непостоянная.
Органоидами движения и захвата пищи служат псевдоподии. Специальное ротовое отверстие отсутствует. Поступление пищи и выделение непереваренных остатков может происходить в любом участке тела. Сократительная вакуоль одна. Выделение продуктов диссимиляции и излишков воды также может происходить в любом месте. Обычно имеется только одно ядро, хотя встречаются и многоядерные формы.
Размножение в основном бесполым путем – делением надвое.
В неблагоприятных условиях образуют цисту.
Мед значение имеет лишь отряд АМЕБ: Дизентерийная амеба(Entomoeba histolytica)
Кишечная
амеба(entomoeba
coli),
ротовая амеба(Entomoeba
gingivalis.
Жизненный цикл дизентирийной амебы. Паразитирует только у человека. В жизненном цикле встречаются следующие формы : циста,мелкая вегетативная форма(formaminuta), крупная вегетативная форма(formamagna) и тканевая.
Инвазионной является циста содержащая 4 ядра. В кишечнике человека оболочка цисты растворяется и из нее выходит четырехядерная амеба, которая быстро делится на 4 одноядерные мелкие вегетативные формы. Мелкая вегетативная форма обитает в просвет толстого кишечника, питается в основном бактериями, размножается и не вызывает заболевания. При попадании в нижние отделы кишечника она превращается в цисту, сод – ую сначала одно ядро, которое в процессе созревания делится с образованием 4ядерной цисты.
У
некоторых людей при соответствующих
условиях( охлаждение.Перегревание,
гельминтозы)малая вегетативная форма
проникает в стенки кишечника, где
инетнсивно размножается и вызывает
поражение слизистой с образованием
язв. При этом разрушаются стенки
кровеносных сосудов и возникают
кровотечения в полость кишечника.
При появлении амебных поражений кишечника мелкие вегетативные формы, находящиеся в просвете кишечника, начинают превращаться в крупную вегетативную форму. Она характеризуется крупными размерами и строением ядра: хроматин ядра образует радиальные структуры, строго в центре располагается крупная глыбка хроматина – кариосома, большая вегетативная начинает питаться эритроцитами, т.е. становится эритрофагом. Характерны тупые широкие псевдоподии и передвижение толчками.
Амебы, размножающиеся в тканях стенки кишечника, – тканевая форма, -попадая в просвет, по строению и размерам становятся сходными с крупной вегетативной формой, но не способны заглатывать эритроциты.
Внедрение амеб в слизистую стенки кишечника и ее расплавление связывают с выделением паразитами веществ, растворяющих тканевые белки.
При
лечении или нарастании защитной реакции
крупная вегетативная вновь превращается
в мелкую, которая начинает инцистироваться.
Далее или наступает выздоровление, или
заболевание переходит в хроническую
форму.
У некоторых людей мелкая форма никогда не переходит в крупную, эти люди – цистоносители, являются источником заражения окружающих. Цистоноситель подлежат обязательному лечению. Единственный источник заболевая амебиазом – чел-к. выделяющиеся с фекалиями цисты загрязняют почву и воду. Нередко фекалии используют как удобрение, так он попадают в огород и сад, где загрязняют овощи и фрукты. В кишечник попадают с немытыми овощами и фруктами, ч-з некипяченую воду, грязные руки. Механические переносчики – мухи, тараканы.
Диагностика – микроскопирование мазков фекалий. В остром периоде в мазке нах-ся крупные вегетативные формы, содержащие эритроциты. При хронической форме в мазке –цисты.
ЛП – мытье овощей и фруктов, употребление только кипяченой воды, мытье рук перед едой и после посещения туалета
ОП — борьба с загрязнением почв и воды фекалиями, уничтожение мух, сан-посвет работа, обследование на цистоносительство лиц, работающих на предприятиях общественного питания, лечение больных
Урок по теме» Тип Жгутиконосцы.
 Колониальные организмы» | План-конспект урока по биологии (7 класс) на тему:
Колониальные организмы» | План-конспект урока по биологии (7 класс) на тему:Технологическая карта урока биологии
Тема урока: Тип Жгутиконосцы. Колониальные организмы
Класс: 7
Тип урока: комбинированный
Цели урока:
обучение: сформировать знания об особенностях строения и процессах жизнедеятельности одноклеточных животных типа Жгутиконосцы и представителях колониальных организмов в связи со средой обитания
развитие: научить устанавливать причинные связи в строении животного и способом его питания на примере эвглены зеленой, развивать у учащихся умения планировать свою деятельность, работать с текстом параграфа (маркировать текст, выделять главные мысли, находить в тексте значение выделенных терминов).
воспитание: формировать у учащихся мотивацию к обучению и познавательной деятельности, учить соблюдать правила работы с микроскопами, выявлять эстетические достоинства простейших как объектов живой природы.
Ожидаемые результаты урока:
Предметные:
– называть и выделять существенные признаки строения и образа жизни животных типа Жгутиконосцы и особенности строения и образа жизни колониальных организмов
– отличать и сравнивать между собой по строению и процессам жизнедеятельности одноклеточных животных типа Корненожки и Жгутиконосцы, распознавать этих животных на рисунках и микропрепаратах
Метапредметные:
регулятивные: умение ставить цели урока, планировать ход своей деятельности по достижении результата, самоконтроль, самооценка.
коммуникативные: умение непосредственного общения в группе, умение задавать вопросы.
познавательные: умение извлекать информацию и работать по алгоритму, проводить сравнение, устанавливать и раскрывать суть причинных связей между строением организма и способом его питания, между строением и образом жизни.
Личностные:
выражать собственную убеждённость и готовность изучать объекты живой природы, выражать эстетическое отношение к живым объектам.
Используемая технология: ИКТ, технология развития критического мышления
Информационно-технологические ресурсы: учебник, рабочая тетрадь, компьютер, проектор, учебный диск, микроскопы, микропрепараты.
Этапы урока | Задачи этапа | Деятельность учителя | Деятельность учащихся | УУД |
Этап мотивации (самоопределения) к учебной деятельности | Создать благоприятный психологический настрой на работу | Организационный момент. Приветствует уч-ся, проверяет готовность к уроку, выявляет отсутствующих. Преобладает фронтальная форма работы. | Приветствуют учителя, сообщают отсутствующих. Деятельность учащихся мотивационная. Формулируют предполагаемую тему урока. Какие цели на уроке будут решаться. | Личностные УУД Уметь смотивировать себя на целенаправленную познавательную деятельность, Коммуникативное УУД. Уметь оформлять свои мысли в устной форме. Познавательные УУД. Уметь преобразовывать информацию из одной формы в другую. |
Этап актуализации и пробного учебного действия | Актуализация опорных знаний и способов действий | Входной контроль Цель: определить исходный уровень знаний на основе повторения темы: Тип Корненожки и Фораминеферы Повторим предыдущий материал. Задание (приложение №1) | Воспроизводят имеющиеся знания о строении и процессах жизнедеятельности простейших животных типа Кореножки Синтезируют имеющиеся знания. | Познавательные УУД. Уметь приводить доказательства фактов. Коммуникативное УУД. Уметь оформлять свои мысли в устной форме |
Стадия вызова – введение в проблему, актуализация знаний. | Обеспечение мотивации учения детьми, формулирование ими целей урока | Цель: В ходе урока продолжим выяснять -так ли просты простейшие? Что бы вы хотели узнать о животных типа Жгутиконосцы? На какие вопросы вам хотелось бы получить ответы? Предлагает сформулировать вопросы, спланировать работу на уроке.
| Формулируют вопросы. Почему так называется этот тип? Как дышат,питаются и размножаются Жгутиконосцы? Какое строение имеют представители типа? Что такое колония? (Все вопросы записываются на доске. Планируют свою работу на уроке | Регулятивные УУД Умение ставить цель, планировать деятельность |
Стадия осмысления – изучение новой темы | Включение учащихся в целенаправленную деятельность | Организует деятельность учащихся: В учебнике на стр. 24-25 и рисунку 9 изучите материал о строении жгутиконосцев (бодо, эвглена зелёная), сравните их. Изучите схемы, иллюстрации в учебнике. Обращает внимание на особенности питания эвглены зелёной – эвглена это растение или животное? Организует работу с микроскопами Организует работу учащихся с рабочей тетрадью (Биология: 7 класс: для учащихся общеобразовательных учреждений/С.В. Суматохин, М. : Мнемозина 2010.) задание №10 Организует чтение текста с пометками (инсерт) о особенностях строения и образе жизни колониальных организмов. Во время работы учащихся, по мере необходимости, учитель отвечает на вопросы учащихся, помогает с выполнением задания. | Работают с текстом, иллюстрациями, схемами в учебнике, рассматривают под микроскопом микропрепараты простейших, читают текст и маркируют его. | Познавательные УУД. Умение работать с информацией, смысловое чтение. Выполнение учебно-практической задачи. Коммуникативное УУД. Формируются навыки активного слушания и умение выделять главное из устного сообщения. |
Этап первичного закрепления с проговариванием во внешней речи | Обеспечение восприятия, осмысления и первичного запоминания детьми изучаемой темы: | Ответьте на вопросы в конце параграфа. Учитель предлагает учащимся изучить материал для заполнения таблицы в рабочей тетради на стр.11 задание №12 Во время работы учащихся, по мере необходимости, учитель отвечает на вопросы учащихся. | Работают с текстом, иллюстрациями, схемами в учебнике. Устанавливают соответствие между текстом и иллюстрациями. Изучают текст учебника, анализируют информацию, делают записи в тетради. | Познавательные УУД. Умение работать с информацией, смысловое чтение, умение анализировать, сравнивать. Коммуникативное УУД. Уметь оформлять свои мысли в устной форме при задавании вопросов. Умение работы в паре (общение). |
Стадия Размышление | Выявление качества и уровня усвоения знаний и способов действий, а также выявление недостатков в знаниях и способах действий, установление причин выявленных недостатков | Выходной контроль. Цель: проверить усвоение материала о особенностях строения и жизнедеятельности Жгутиконосцев Предлагает учащимся задания для самостоятельной работы. Приложение №2 | Выполняют самостоятельно тест. Проверяют по ключам ответы теста. Выставляют оценку за выполненную работу (в соответствии с критериями), затем осуществляют взаимооценивание. | Познавательные УУД. Умение работать по алгоритму, анализировать полученную информацию, подводить итоги. Регулятивные УУД Умение осуществлять контроль и коррекцию своей деятельности |
Этап включения в систему знаний и повторения | Дать качественную оценку работы класса и отдельных учащихся | Для коррекции полученных результатов в предыдущем этапе ребятам предлагаются задания базового и повышенного уровня сложности. | Выполняют предложенные задания. Осуществляют самопроверку и взаимопроверку (по предложенным ключам) | Познавательные УУД. Умение работать по алгоритму, анализировать полученную информацию, подводить итоги. Регулятивные УУД Умение осуществлять контроль и коррекцию своей деятельности Коммуникативное УУД. Умение работы в паре (общение), владение устной и письменной речью |
Информация о домашнем задании | Обеспечение понимания детьми цели, содержания и способов выполнения домашнего задания | Задаёт домашнее задание учащимся, которые выполнили задание только базового уровня, и допустили серьёзные ошибки в заданиях повышенного уровня. В соответствие с оценкой запишите домашнее задание в дневник: | Записывают в дневник домашнее задание, задают интересующие их вопросы. | |
Этап рефлексии учебной деятельности на уроке | Инициировать рефлексию детей по их собственной деятельности и взаимодействия с учителем и другими детьми в классе | Учитель организует рефлексию: Обсуждает с учащимися выставленные ими оценки и корректирует оценку учащихся. Просит продолжить предложения: Раньше не знал….. Было интересно….. Особенно запомнилось…. Предлагает составить синквейн о Простейших | Объявляют выставленную ими оценку. Задают вопросы учителю. | Личностные УУД Готовность к личностному самоопределению, самооценке на основе критерия успешной учебной деятельности.Коммуникативное УУД. Уметь оформлять свои мысли в устной форме, задать вопрос. Регулятивные УУД Умение оценивать правильность выполнения действия на уровне адекватной оценки |

 Опрос домашнего задания заканчивается составлением кластера (в центре слово Простейшие, от него слова – одна клетка, водная среда, гетеротрофы, бесполое
Опрос домашнего задания заканчивается составлением кластера (в центре слово Простейшие, от него слова – одна клетка, водная среда, гетеротрофы, бесполое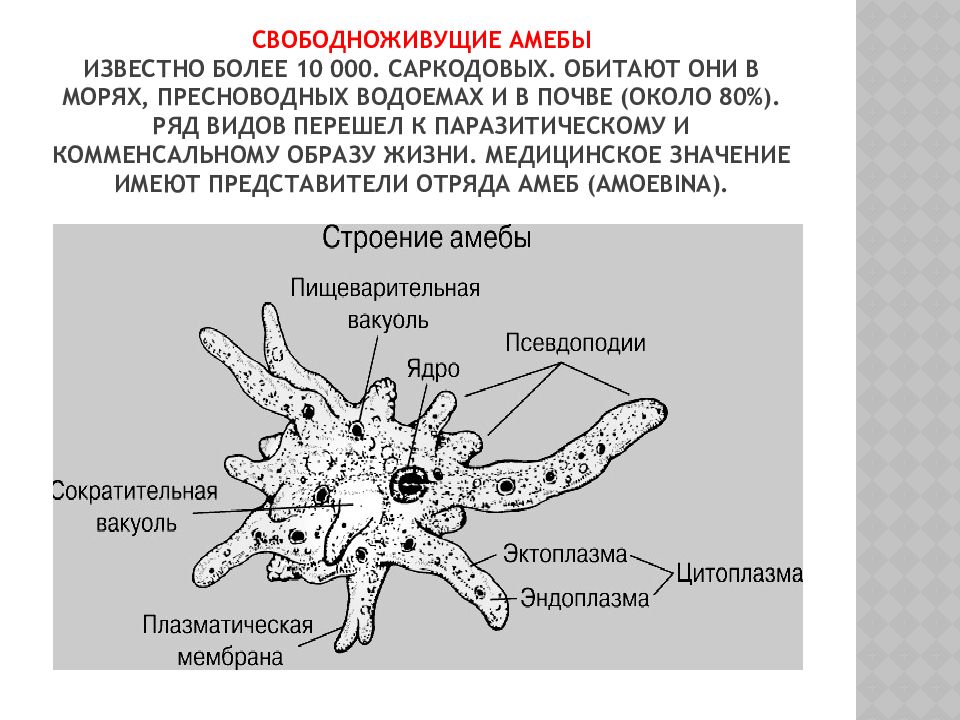 Это наши задачи)
Это наши задачи)




Приложение №1
Фронтальный опрос
- Почему подцарство изучаемых животных называется Простейшие?
- Какова их среда обитания?
- Какой способ питания характерен этим животным?
- Какие типы входят в состав подцарства?
Индивидуальный опрос
- Объясните название типа Корненожки.
- Сделать рисунок амёбы протей на доске, рассказать о её строении.

- Форма тела амёбы непостоянная. Почему жесткая оболочка совершенно не нужна и даже вредна амёбе?
- Представьте себе, что человек создал робота, передвигающегося как амёба и свободно меняющего форму, как амёба. Какие функции может выполнять этот робот?
- Как дышит амёба?
- Размножение амёбы. Рассказ – комментарий видео слайда.
- Какими преимуществами обладает амёба образующая цисту от амёбы, цисту не образующей?
- Каковы особенности строения фораминифер? Что теряет и что приобретает амёба, поселившись в ажурной раковине фораминиферы?
- Из чего построена пирамида Хеопса? (сообщение)
Приложение №2
Задание 1. Верно ли суждение:
- Тело простейшего состоит из одной клетки.
- Простейшие обитают в водной и наземно – воздушной среде.
- Амеба протей передвигается с помощью жгутиков.
- В передвижении простейших играет роль сократительная вакуоль.
- У эвглены зелёной пищеварительные вакуоли не образуются.

- Кислород в цитоплазму простейшего поступает через всю поверхность тела.
- Жгутиконосец бодо при делении образует не две, а несколько особей.
- На стадии цисты происходит расселение простейших ветром и животными.
- Из раковин погибших фораминифер образовались отложения известняка.
Задание 2. Выпишите цифры, обозначающие признаки, характерные для этих животных:
амеба обыкновенная: ______________, эвглена зеленая: ______________
Признаки: 1. Микроскопически малые животные; 2. Есть ядро; 3. Есть оболочка; 4. Пища переваривается в пищеварительных вакуолях; 5. Тело состоит из одной клетки; 6. Передвигается с помощью жгутика; 7. Есть цитоплазма; 8. Продукты распада удаляются через сократительную вакуоль; 9. Среда обитания жидкая; 10. Передвигаются с помощью ложноножек; 11. Может всасывать растворенные органические вещества; 12. На свету питается как зеленое растение, образуя органические вещества из неорганических.
Protozoa
Protozoa — одноклеточные эукариоты, принадлежащие к царству Protista.
Они существуют либо как свободноживущие организмы, либо как паразиты. Известно более 50 000 видов размером от 2 до 70 микрон (от 0,002 до 0,07 мм).
Они имеют относительно сложную внутреннюю структуру и осуществляют сложную метаболическую активность.
Органеллы простейших выполняют функции органов высших животных.
Некоторые простейшие имеют специальные структуры, которые помогают им двигаться и двигаться. Примерами таких структур являются реснички, жгутики и псевдоподии.
Общая характеристика типа Protozoa
- Царство: Protista
- Среда обитания: в основном водные, свободноживущие или паразитирующие
- Степень организации: протоплазматическая степень организации. Одна клетка выполняет все функции жизнедеятельности. Они известны как бесклеточные или неклеточные организмы, поскольку их тело состоит только из массы протоплазмы.
- Оболочка : тело простейших либо голое, либо покрыто пелликулой или хитином.

- Передвижение: органов передвижения представляют собой псевдоподии, жгутики или реснички.
- Питание: питание бывает голофитным, голозойным, сапрофитным или паразитарным.
- Пищеварение: пищеварение является внутриклеточным и происходит в пищевых вакуолях.
- Дыхание: дыхание происходит через поверхность тела.
- Осморегуляция: сократительных вакуолей помогают осморегуляции (рис. 2).
- Репродукция: бесполое размножение бинарным делением или почкованием и половое размножение сингамией или конъюгацией.
Классификация типов Protozoa
В 1985 г. Общество протозоологов опубликовало таксономическую схему, согласно которой простейшие подразделяются на четыре основных класса, показанных ниже:
- Rhizopda
- Жгутиковые/мастигофоры
- споровики
- Цилиата
Таблица, сравнивающая признаки четырех классов простейших
| Особенности | корненожки | Жгутиковые | споровики | Реснички |
| Опорно-двигательный аппарат | Псевдоподия | Жгутики | Отсутствует | Реснички |
| Среда обитания | В основном живут свободно, некоторые паразитируют | Свободноживущие или паразиты | Исключительно эндопаразиты | Свободноживущие или паразиты |
| Репродукция | Бесполое путем бинарного деления и половое путем сингамии | Половое размножение продольным делением | Бесполое размножение делением и половое размножение спорами | Бесполое размножение путем бинарного деления.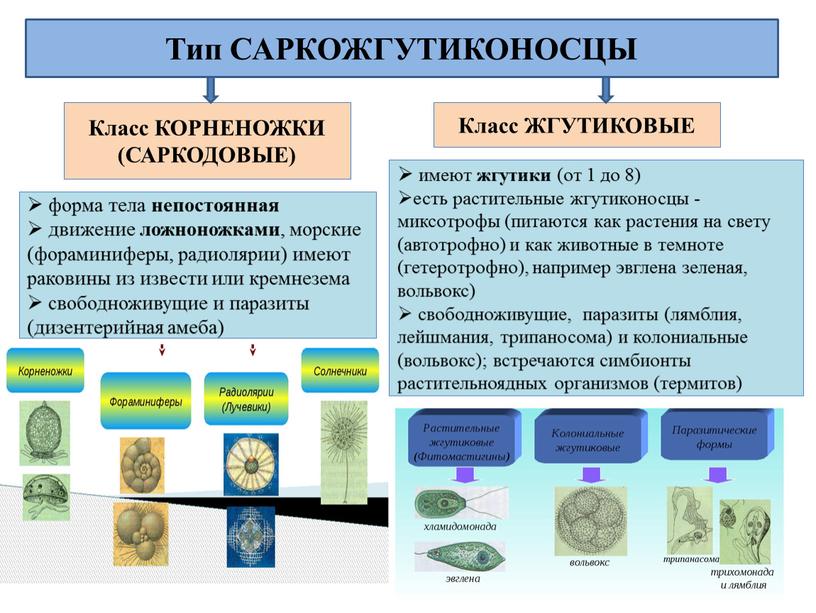 половое размножение путем конъюгации половое размножение путем конъюгации |
| Крышка корпуса | Отсутствует | Тело, покрытое целлюлозой, хитином или кремнеземом | Корпус покрыт пленкой | Корпус покрыт пленкой |
| Примеры | Амеба (рис. 2), Энтамеба | Giardia, Euglena, Trypanosoma | Плазмодий моноцистис | Paramecium (рис. 3 ), Voricella, Blantidium |
Среда обитания
Простейшие встречаются повсеместно. Они живут в самых разных средах обитания, включая пресную воду, морскую среду обитания и почву.
Их цисты можно найти даже в самых негостеприимных частях биосферы. Поскольку группа содержит много неродственных или слабородственных организмов, существуют разновидности по структуре и форме.
Большинство из них свободноживущие и питаются бактериями, водорослями или другими простейшими. Они являются важными компонентами водных и почвенных экосистем.
Они являются важными компонентами водных и почвенных экосистем.
Например, бактериоядные простейшие изобилуют на станциях очистки сточных вод с активным илом, тогда как некоторые простейшие, такие как Escherichia Coli (рис. 4), Entamoeba histolytica , Giardia lamblia имеют медицинское и экономическое значение.
Рисунок 4. Бактерия Escherichia coli ( E . coli ) грамотрицательная палочковидная бактерия, часть нормальной флоры кишечника и возбудитель диареи и воспаленияЖизненный цикл
В течение своего жизненного цикла простейшее в основном проходит через несколько стадий, различающихся по структуре и активности.
Трофозоит — это общий термин, обозначающий активную, питающуюся, размножающуюся стадию простейших. У паразитических видов трофозоиты связаны с патогенезом.
У гемофлагеллят (разновидность простейших) термины амастигота, промастигота, эпимастигота и трипомастигота представляют стадии трофозоита.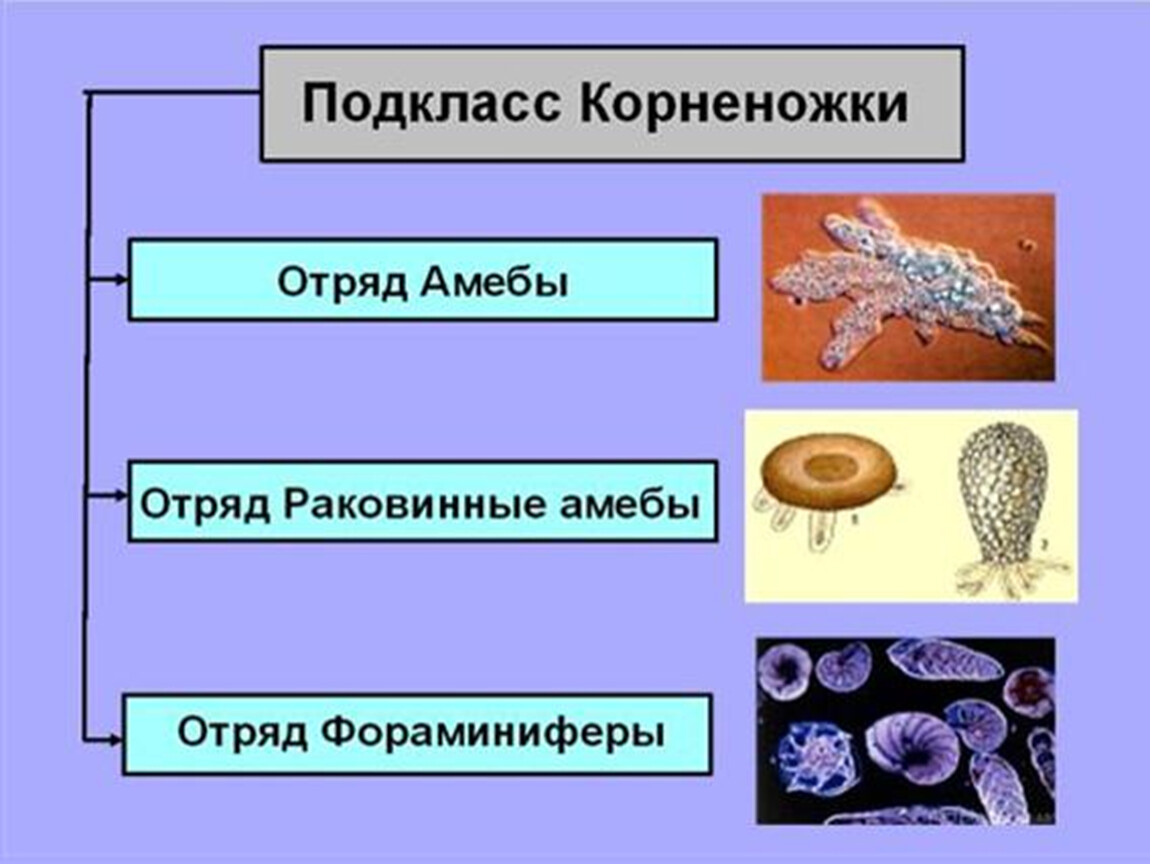
Эта стадия отличается отсутствием или наличием жгутика и положением кинетопласта, связанного со жгутиком.
Некоторые простейшие образуют цисты, содержащие одну или несколько инфекционных форм. Ооцисты — это стадии, возникающие в результате полового размножения у некоторых простейших, в то время как ооцисты Плазмодий.
Развиваются в полости тела комара-переносчика (рис. 5).
Некоторые простейшие имеют сложные жизненные циклы, требующие наличия двух разных видов хозяев, в то время как другим для завершения жизненного цикла требуется только один хозяин.
Рисунок 5. Жизненный цикл Plasmodium FalciparumПитание
Простейшие питаются по-разному: голозойным (как у животных), голофитным (как у растений), сапрофитным или паразитарным.
Амеба поглощает твердые частицы пищи или капли через рот, осуществляет пищеварение и всасывание в пищевой вакуоли и выбрасывает отходы.
У многих простейших проглоченная пища проходит через цитосомы или микропоры и затем попадает в пищевые вакуоли.
Пиноцитоз представляет собой метод проглатывания питательных веществ, при котором жидкость всасывается через небольшие временные отверстия в стенке тела организма.
Простейшие также классифицируются в зависимости от питания на три категории, перечисленные ниже:
- Автотрофы
- Гетеротрофы
- Хемогетеротрофы
Автотрофы
Автотрофы, как и некоторые жгутиковые, синтезируют углеводы из углекислого газа и воды с использованием хлорофилла в присутствии солнечного света.
Большинство фотоавтотрофных жгутиконосцев (например, Euglenida, Volvocida) склонны сочетать автотрофию с гетеротрофией. Их источником углерода являются ацетаты, простые жирные кислоты или спирты.
Будучи автотрофами на свету, эти жгутиковые превращаются в гетеротрофов в темноте.
Гетеротрофы
Большинство свободноживущих простейших являются гетеротрофами. Они зависят от широкого спектра продуктов питания. Некоторые питаются бактериями (микрофаги), тогда как другие питаются водорослями.
Плотоядные формы питаются как травоядными, так и микробоядными.
Хемогетеротрофные
В эту группу входят те, которые требуют энергии и источников органического углерода.
Размножение
Бинарное деление является наиболее распространенной формой бесполого размножения простейших. При бинарном делении органеллы организма удваиваются, а затем организм делится на два полных организма.
Множественное деление, плазмотомия и почкование — другие формы бесполого размножения.
Размножение простейших может быть бесполым, как у амеб и жгутиконосцев (заражающих человека), или как бесполым, так и половым, как у апикомплексов.
Половое размножение у Protozoa в основном осуществляется:
- сингамией (например, Chlamydomonas, Copromonas), конъюгация
- (например, Paramecium (рис. 6), Vorticella) и
- automixis (например, Actinosphaerium, Actinophrys).
- плазмогамия (например, Rhizopoda и Mycetozoa),
- регенерация и
- партаногенез (например, Actinophrys, Chlamydomonas).

Деление продольное у жгутиковых, поперечное у инфузорий, а эндодиогенез является формой бесполого деления, наблюдаемой у Toxoplasma .
Рис. 6. Конъюгация в парамециях при световой микроскопииИнициация филоподий – ПМК
1. Эллерманн В. Об обнаружении корненожек в двух случаях острого переднего полиомиелита. В: Брюс А., Брамвелл Э., Кэмпбелл М., редакторы. Обзор неврологии и психиатрии. Эдинбург: Отто Шульце Ко.; 1906. с. 353. [Google Scholar]
2. Харрисон Р.Г. Наблюдения за живым развивающимся нервным волокном. Анат Рек. 1907; 1: 116–118. [Google Scholar]
3. Gallo G, Letourneau PC. Регуляция актиновых филаментов конуса роста с помощью сигналов управления. Дж Нейробиол. 2004; 58: 92–102. [PubMed] [Google Scholar]
4. Дрис Ф., Гертлер Ф.Б. Ena/VASP: белки на кончике нервной системы. Курр Опин Нейробиол. 2008; 18:53–59. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. De Smet F, Segura I, De Bock K, Hohensinner PJ, Carmeliet P. Механизмы ветвления сосудов: филоподии на эндотелиальных концевых клетках ведут вперед. Артериосклеры Тромб Васк Биол. 2009 г.;29:639–649. [PubMed] [Google Scholar]
Механизмы ветвления сосудов: филоподии на эндотелиальных концевых клетках ведут вперед. Артериосклеры Тромб Васк Биол. 2009 г.;29:639–649. [PubMed] [Google Scholar]
6. Мачески Л.М., Ли А. Фаскин: Инвазивные филоподии, способствующие метастазированию. Коммун Интегр Биол. 2010;3:263–270. [PMC free article] [PubMed] [Google Scholar]
7. Васюхин В., Бауэр С., Инь М., Фукс Е. Направленная полимеризация актина является движущей силой межклеточной адгезии эпителия. Клетка. 2000; 100: 209–219. [PubMed] [Google Scholar]
8. Вуд В., Мартин П. Структуры в фокус-филоподии. Int J Biochem Cell Biol. 2002; 34: 726–730. [PubMed] [Академия Google]
9. Борн Дж. Н., Харрис К. М. Балансировка структуры и функции дендритных шипиков гиппокампа. Annu Rev Neurosci. 2008; 31:47–67. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Hotulainen P, Hoogenraad CC. Актин в дендритных шипиках: соединяя динамику с функцией. Джей Селл Биол. 2010; 189: 619–629. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Хейман М.Г., Шахам С. Ветки в ветви: как филоподий становится дендритом. Курр Опин Нейробиол. 2010;20:86–91. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Хейман М.Г., Шахам С. Ветки в ветви: как филоподий становится дендритом. Курр Опин Нейробиол. 2010;20:86–91. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Аль-Алван М.М., Роуден Г., Ли Т.Д., Уэст К.А. Фасцин участвует в антигенпрезентирующей активности зрелых дендритных клеток. Дж Иммунол. 2001; 166: 338–345. [PubMed] [Google Scholar]
13. Faix J, Rottner K. Создание филоподий. Curr Opin Cell Biol. 2006; 18:18–25. [PubMed] [Google Scholar]
14. Гуптон С.Л., Гертлер Ф.Б. Филоподии: пальцы, которые совершают ходьбу. наук СТКЭ. 2007; 2007:5. [PubMed] [Google Scholar]
15. Маттила П.К., Лаппалайнен П. Филоподии: молекулярная архитектура и клеточные функции. Nat Rev Mol Cell Biol. 2008;9: 446–454. [PubMed] [Google Scholar]
16. Faix J, Breitsprecher D, Stradal TE, Rottner K. Филоподия: сложные модели простых стержней. Int J Biochem Cell Biol. 2009;41:1656–1664. [PubMed] [Google Scholar]
17. Меллор Х. Роль форминов в формировании филоподий. Биохим Биофиз Акта. 2010; 1803: 191–200. [PubMed] [Google Scholar]
Биохим Биофиз Акта. 2010; 1803: 191–200. [PubMed] [Google Scholar]
18. Могильнер А., Рубинштейн Б. Физика филоподиальной протрузии. Биофиз Дж. 2005; 89: 782–795. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Могильнер А. Математика клеточной подвижности: получили ли мы ее число? Дж. Матем. Биол. 2009; 58: 105–134. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Carlsson AE, Sept D. Математическое моделирование миграции клеток. Методы Cell Biol. 2008; 84: 911–937. [PubMed] [Google Scholar]
21. Чабра Э.С., Хиггс Х.Н. Многоликость актина: сопоставление факторов сборки с клеточными структурами. Nat Cell Biol. 2007; 9: 1110–1121. [PubMed] [Google Scholar]
22. Николсон-Дикстра С.М., Хиггс Х.Н. Истощение Arp2 ингибирует листовидные выпячивания, но не линейные выпячивания фибробластов и лимфоцитов. Клеточный подвижный цитоскелет. 2008;65:904–922. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Ahmed S, Goh WI, Bu W. Домены I-BAR, IRSp53 и формирование филоподия. Цемин Селл Дев Биол. 2010;21:350–356. [PubMed] [Google Scholar]
Цемин Селл Дев Биол. 2010;21:350–356. [PubMed] [Google Scholar]
24. Goh WI, Sudhaharan T, Lim KB, Sem KP, Lau CL, Ahmed S. Взаимодействие Rif-mDia1 участвует в формировании филоподия независимо от эффекторов Cdc42 и Rac. Дж. Биол. Хим. 2011; 286:13681–13694. [PMC free article] [PubMed] [Google Scholar]
25. Коробова Ф., Свиткина Т. Молекулярная архитектура синаптического актинового цитоскелета в нейронах гиппокампа раскрывает механизм морфогенеза дендритных шипиков. Мол Биол Селл. 2010;21:165–176. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Свиткина Т.М., Буланова Е.А., Чага О.Ю., Вигневич Д.М., Кодзима С., Васильев Ю.М., и соавт. Механизм инициации филоподий путем реорганизации дендритной сети. Джей Селл Биол. 2003; 160:409–421. [PMC free article] [PubMed] [Google Scholar]
27. Коробова Ф., Свиткина Т. Комплекс Arp2/3 важен для формирования филоподий, подвижности конусов роста и нейритогенеза в нейрональных клетках. Мол Биол Селл. 2008;19:1561–1574. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Pruyne D, Evangelista M, Yang C, Bi E, Zigmond S, Bretscher A, et al. Роль форминов в сборке актина: нуклеация и ассоциация колючих концов. Наука. 2002;297: 612–615. [PubMed] [Google Scholar]
29. Romero S, Le Clainche C, Didry D, Egile C, Pantaloni D, Carlier MF. Формин является процессивным мотором, которому требуется профилин для ускорения сборки актина и связанного с ним гидролиза АТФ. Клетка. 2004; 119:419–429. [PubMed] [Google Scholar]
30. Ковар Д.Р., Поллард Т.Д. Вставка зазубренных концов актиновых филаментов в ассоциации с форминами создает силы пиконьютона. Proc Natl Acad Sci USA. 2004; 101:14725–14730. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Breitsprecher D, Kiesewetter AK, Linkner J, Urbanke C, Resch GP, Small JV, et al. Кластеризация VASP активно управляет процессивным, опосредованным доменом Wh3 удлинением актиновых филаментов. EMBO J. 2008; 27: 2943–2954. [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Hansen SD, Mullins RD. VASP представляет собой процессивную актиновую полимеразу, которой требуется мономерный актин для ассоциации с зазубренными концами. Джей Селл Биол. 2010;191:571–584. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Hansen SD, Mullins RD. VASP представляет собой процессивную актиновую полимеразу, которой требуется мономерный актин для ассоциации с зазубренными концами. Джей Селл Биол. 2010;191:571–584. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Mullins RD, Heuser JA, Pollard TD. Взаимодействие комплекса Arp2/3 с актином: нуклеация, высокоаффинное кэпирование остроконечных концов и образование сети ветвящихся филаментов. Proc Natl Acad Sci USA. 1998;95:6181–6186. [PMC free article] [PubMed] [Google Scholar]
34. Свиткина Т.М., Верховский А.Б., Маккуэйд К.М., Борисий Г.Г. Анализ актин-миозиновой системы II в эпидермальных кератоцитах рыб: механизм транслокации клеточных тел. Джей Селл Биол. 1997; 139: 397–415. [бесплатная статья PMC] [PubMed] [Google Scholar]
35. Свиткина Т.М., Борисий Г.Г. Комплекс Arp2/3 и фактор деполимеризации актина/кофилин в дендритной организации и беговой дорожке массива актиновых филаментов в ламеллоподиях. Джей Селл Биол. 1999;145:1009–1026. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Goley ED, Welch MD. Комплекс ARP2/3: актиновый зародыш достигает совершеннолетия. Nat Rev Mol Cell Biol. 2006; 7: 713–726. [PubMed] [Google Scholar]
37. Поллард Т.Д. Регуляция сборки актиновых филаментов комплексом Arp2/3 и форминами. Annu Rev Biophys Biomol Struct. 2007; 36: 451–477. [PubMed] [Google Scholar]
38. Le Clainche C, Carlier MF. Регуляция сборки актина, связанная с протрузией и адгезией при миграции клеток. Physiol Rev. 2008; 88:489–513. [PubMed] [Google Scholar]
39. Мачески Л.М., Инсолл Р.Х. Scar1 и родственный белок синдрома Вискотта-Олдрича, WASP, регулируют актиновый цитоскелет через комплекс Arp2/3. Карр Биол. 1998; 8: 1347–1356. [PubMed] [Google Scholar]
40. Innocenti M, Gerboth S, Rottner K, Lai FP, Hertzog M, Stradal TE, et al. Abi1 регулирует активность N-WASP и WAVE в различных процессах, основанных на актине. Nat Cell Biol. 2005; 7: 969–976. [PubMed] [Google Scholar]
41. Derivery E, Gautreau A. Генерация разветвленных актиновых сетей: сборка и регулирование молекулярных машин N-WASP и WAVE. Биоэссе. 2010;32:119–131. [PubMed] [Google Scholar]
Derivery E, Gautreau A. Генерация разветвленных актиновых сетей: сборка и регулирование молекулярных машин N-WASP и WAVE. Биоэссе. 2010;32:119–131. [PubMed] [Google Scholar]
42. Campellone KG, Welch MD. Гонка нуклеаторов: клеточный контроль сборки актина. Nat Rev Mol Cell Biol. 2010; 11: 237–251. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Higgs HN, Peterson KJ. Филогенетический анализ домена гомологии формина 2. Мол Биол Селл. 2005; 16:1–13. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Higgs HN. Форминовые белки: доменный подход. Тенденции биохимических наук. 2005; 30: 342–353. [PubMed] [Академия Google]
45. Гуд Б.Л., Экк М.Дж. Механизм и функция форминов в контроле сборки актина. Анну Рев Биохим. 2007; 76: 593–627. [PubMed] [Google Scholar]
46. Пол А.С., Поллард Т.Д. Обзор механизма удлинения процессивных актиновых филаментов форминами. Клеточный подвижный цитоскелет. 2009; 66: 606–617. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Yang C, Czech L, Gerboth S, Kojima S, Scita G, Svitkina T. Новые роли формина mDia2 в формировании ламеллоподий и филоподий в подвижных клетках. PLoS биол. 2007; 5:317. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Yang C, Czech L, Gerboth S, Kojima S, Scita G, Svitkina T. Новые роли формина mDia2 в формировании ламеллоподий и филоподий в подвижных клетках. PLoS биол. 2007; 5:317. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Gates J, Mahaffey JP, Rogers SL, Emerson M, Rogers EM, Sottile SL, et al. Enabled играет ключевую роль в морфогенезе эмбрионального эпителия у Drosophila. Разработка. 2007;134:2027–2039. [PubMed] [Google Scholar]
49. Falet H, Hoffmeister KM, Neujahr R, Hartwig JH. Нормальная активация комплекса Arp2/3 в тромбоцитах без WASp. Кровь. 2002; 100:2113–2122. [PubMed] [Google Scholar]
50. Вигжевич Д., Ярар Д., Уэлч М.Д., Пелокин Дж., Свиткина Т., Борисый Г.Г. Формирование филоподийных пучков in vitro из дендритной сети. Джей Селл Биол. 2003;160:951–962. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Haviv L, Brill-Karniely Y, Mahaffy R, Backouche F, Ben-Shaul A, Pollard TD, et al. Восстановление перехода от ламеллиподия к филоподию в безмембранной системе. Proc Natl Acad Sci USA. 2006; 103:4906–4911. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Proc Natl Acad Sci USA. 2006; 103:4906–4911. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Brill-Karniely Y, Ideses Y, Bernheim-Groswasser A, Ben-Shaul A. От разветвленных сетей актиновых филаментов к пучкам. Химфиз. 2009; 10: 2818–2827. [PubMed] [Академия Google]
53. Lee K, Gallop JL, Rambani K, Kirschner MW. Самосборка филоподийных структур на поддерживаемых липидных бислоях. Наука. 2010; 329:1341–1345. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Norris AD, Dyer JO, Lundquist EA. Комплекс Arp2/3, UNC-115/abLIM и UNC-34/Enabled регулируют направление аксонов и образование филоподий конусов роста у Caenorhabditis elegans . Нейронный разработчик. 2009; 4:38. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Goncalves-Pimentel C, Gombos R, Mihaly J, Sanchez-Soriano N, Prokop A. Рассечение регуляторных сетей формирования филоподий в модели конуса роста дрозофилы. ПЛОС Один. 2011;6:18340. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Квалманн Б., Келли Р.Б. Изоформы синдапина участвуют в рецептор-опосредованном эндоцитозе и организации актина. Джей Селл Биол. 2000; 148:1047–1062. [PMC free article] [PubMed] [Google Scholar]
Квалманн Б., Келли Р.Б. Изоформы синдапина участвуют в рецептор-опосредованном эндоцитозе и организации актина. Джей Селл Биол. 2000; 148:1047–1062. [PMC free article] [PubMed] [Google Scholar]
57. Бияшева А., Свиткина Т., Кунда П., Баум Б., Борисы Г. Каскадный путь образования филоподий ниже по течению от SCAR. Дж. Клеточные науки. 2004; 117: 837–848. [PubMed] [Google Scholar]
58. Johnston SA, Bramble JP, Yeung CL, Mendes PM, Machesky LM. Активность комплекса Arp2/3 на филоподиях распространяющихся клеток. BMC клеточная биология. 2008;9:65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Cohen M, Georgiou M, Stevenson NL, Miodownik M, Baum B. Динамические филоподии передают прерывистую передачу сигналов Delta-Notch для уточнения паттерна во время бокового торможения. Ячейка Дев. 2010;19:78–89. [PubMed] [Google Scholar]
60. Miki H, Sasaki T, Takai Y, Takenawa T. Индукция образования филоподия родственным WASP актин-деполимеризующим белком N-WASP. Природа. 1998; 391:93–96. [PubMed] [Google Scholar]
Природа. 1998; 391:93–96. [PubMed] [Google Scholar]
61. Cory GO, Garg R, Cramer R, Ridley AJ. Фосфорилирование тирозина 291 усиливает способность WASp стимулировать полимеризацию актина и образование филоподия. Дж. Биол. Хим. 2002; 277:45115–45121. [PubMed] [Google Scholar]
62. Bu W, Chou AM, Lim KB, Sudhaharan T, Ahmed S. Комплекс Toca-1-N-WASP связывает образование филоподий с эндоцитозом. Дж. Биол. Хим. 2009; 284:11622–11636. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Sheldon H, Andre M, Legg JA, Heal P, Herbert JM, Sainson R, et al. Активное участие Robo1 и Robo4 в формировании филоподий и подвижности эндотелиальных клеток опосредуется через WASP и другие факторы, способствующие нуклеации актина. ФАСЭБ Дж. 2009 г.;23:513–522. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Lommel S, Benesch S, Rottner K, Franz T, Wehland J, Kuhn R. Формирование пьедестала актина энтеропатогенными Escherichia coli и внутриклеточная подвижность Shigella flexneri исчезают в N-WASP-дефектных клетках. EMBO Rep. 2001; 2: 850–857. [Бесплатная статья PMC] [PubMed] [Google Scholar]
EMBO Rep. 2001; 2: 850–857. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Snapper SB, Takeshima F, Anton I, Liu CH, Thomas SM, Nguyen D, et al. Дефицит N-WASP выявляет различные пути для проекций клеточной поверхности и микробной подвижности на основе актина. Nat Cell Biol. 2001;3:897–904. [PubMed] [Google Scholar]
66. Co C, Wong DT, Gierke S, Chang V, Taunton J. Механизм прикрепления актиновой сети к движущимся мембранам: захват зазубренных концов доменами N-WASP Wh3. Клетка. 2007; 128:901–913. [Бесплатная статья PMC] [PubMed] [Google Scholar]
67. Takano K, Watanabe-Takano H, Suetsugu S, Kurita S, Tsujita K, Kimura S, et al. Небулин и N-WASP взаимодействуют, вызывая индуцированное IGF-1 образование саркомерных актиновых филаментов. Наука. 2010; 330:1536–1540. [PubMed] [Академия Google]
68. Меджиллано М.Р., Кодзима С., Эпплуайт Д.А., Гертлер Ф.Б., Свиткина Т.М., Борисий Г.Г. Ламеллиподиальный и филоподиальный режим актиновой наномашины; ключевую роль зазубренного конца нити. Клетка. 2004; 118: 363–373. [PubMed] [Google Scholar]
Клетка. 2004; 118: 363–373. [PubMed] [Google Scholar]
69. Lebrand C, Dent EW, Strasser GA, Lanier LM, Krause M, Svitkina TM, et al. Критическая роль белков Ena/VASP в формировании филоподий в нейронах и в функционировании ниже нетрина-1. Нейрон. 2004; 42:37–49. [PubMed] [Академия Google]
70. Квятковский А.В., Рубинсон Д.А., Дент Э.В., Эдвард ван Вин Дж., Лесли Дж.Д., Чжан Дж. и соавт. Ena/VASP необходим для нейритогенеза в развивающейся коре. Нейрон. 2007; 56: 441–455. [PubMed] [Google Scholar]
71. Han YH, Chung CY, Wessels D, Stephens S, Titus MA, Soll DR, et al. Потребность в члене семейства сосудорасширяющих фосфопротеинов (VASP) для клеточной адгезии, образования филоподий и хемотаксиса у Dictyostelium. Дж. Биол. Хим. 2002;17:17. [PubMed] [Академия Google]
72. Homem CC, Peifer M. Изучение роли прозрачной и активированной активности в формировании баланса между филоподиями и ламеллиподиями. Мол Биол Селл. 2009;20:5138–5155. [Бесплатная статья PMC] [PubMed] [Google Scholar]
73. Peng J, Wallar BJ, Flanders A, Swiatek PJ, Alberts AS. Нарушение родственного Diaphanous гена формина Drf1, кодирующего mDia1, показывает роль Drf3 как эффектора для Cdc42. Карр Биол. 2003; 13: 534–545. [PubMed] [Google Scholar]
Peng J, Wallar BJ, Flanders A, Swiatek PJ, Alberts AS. Нарушение родственного Diaphanous гена формина Drf1, кодирующего mDia1, показывает роль Drf3 как эффектора для Cdc42. Карр Биол. 2003; 13: 534–545. [PubMed] [Google Scholar]
74. Pellegrin S, Mellor H. GTPase Rif семейства Rho индуцирует филоподии через mDia2. Карр Биол. 2005;15:129–133. [PubMed] [Google Scholar]
75. Harris ES, Gauvin TJ, Heimsath EG, Higgs HN. Сборка филоподий формином FRL2 (FMNL3) Cytoskeleton (Hoboken) 2010;67:755–772. [Бесплатная статья PMC] [PubMed] [Google Scholar]
76. Harris ES, Rouiller I, Hanein D, Higgs HN. Механистические различия в связывающей актин активности двух форминов млекопитающих, FRL1 и mDia2. Дж. Биол. Хим. 2006; 281:14383–14392. [PubMed] [Google Scholar]
77. Lambrechts A, Kwiatkowski AV, Lanier LM, Bear JE, Vandekerckhove J, Ampe C, et al. цАМФ-зависимое фосфорилирование протеинкиназы EVL, родственника Mena/VASP, регулирует его взаимодействие с доменами актина и Sh4. Дж. Биол. Хим. 2000; 275:36143–36151. [PubMed] [Академия Google]
Дж. Биол. Хим. 2000; 275:36143–36151. [PubMed] [Академия Google]
78. Эпплуайт Д.А., Барзик М., Кодзима С., Свиткина Т.М., Гертлер Ф.Б., Борисий Г.Г. Белки Ena/VASP выполняют независимую функцию против кэппинга при формировании филоподий. Мол Биол Селл. 2007; 18: 2579–2591. [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Dent EW, Kwiatkowski AV, Mebane LM, Philippar U, Barzik M, Rubinson DA, et al. Филоподии необходимы для инициации кортикальных нейритов. Nat Cell Biol. 2007; 9: 1347–1359. [PubMed] [Google Scholar]
80. Schirenbeck A, Bretschneider T, Arasada R, Schleicher M, Faix J. Формамин dDia2, связанный с Diaphanous, необходим для формирования и поддержания филоподий. Nat Cell Biol. 2005;7:619–625. [PubMed] [Google Scholar]
81. Matusek T, Gombos R, Szecsenyi A, Sanchez-Soriano N, Czibula A, Pataki C, et al. Белки формина подсемейства DAAM играют роль во время роста аксона. Дж. Нейроски. 2008;28:13310–13319. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Block J, Stradal TE, Hanisch J, Geffers R, Kostler SA, Urban E, et al. Образование филоподий, индуцированное активным mDia2/Drf3. Дж Микроск. 2008; 231: 506–517. [PubMed] [Google Scholar]
Block J, Stradal TE, Hanisch J, Geffers R, Kostler SA, Urban E, et al. Образование филоподий, индуцированное активным mDia2/Drf3. Дж Микроск. 2008; 231: 506–517. [PubMed] [Google Scholar]
83. Steffen A, Faix J, Resch GP, Linkner J, Wehland J, Small JV, et al. Формирование филоподий в отсутствие функциональных WAVE- и Arp2/3-комплексов. Мол Биол Селл. 2006; 17: 2581–259.1. [Бесплатная статья PMC] [PubMed] [Google Scholar]
84. Pronk S, Geissler PL, Fletcher DA. Пределы устойчивости филоподия. Phys Rev Lett. 2008;100:258102. [Бесплатная статья PMC] [PubMed] [Google Scholar]
85. Beli P, Mascheroni D, Xu D, Innocenti M. WAVE и Arp2/3 совместно ингибируют образование филоподия, входя в комплекс с mDia2. Nat Cell Biol. 2008; 10: 849–857. [PubMed] [Google Scholar]
86. Sarmiento C, Wang W, Dovas A, Yamaguchi H, Sidani M, El-Sibai M, et al. Члены семейства WASP и белки формина координируют регуляцию клеточных выпячиваний в клетках карциномы. Джей Селл Биол. 2008; 180:1245–1260.