АмГПГУНовая страница
В вашем браузере отключен JavaScript, поэтому некоторые возможности сайта будут недоступны.
Как включить JavaScript ?
Результаты 1 — 10 из 183
Начало | Пред. | 1 2 3 4 5 | След. | Конец | Все
| Изменено | Курс | Школа | Класс | ФИО | Обратный контакт | |||
|---|---|---|---|---|---|---|---|---|
| 03.11.2022 12:13:48 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ16 с УИОП | 10А | Куберская Полина Степановна | 79242251090 | |||
02. 11.2022 11.202219:17:56 | ИПиП Тренинг «Как преодолеть страх перед ОГЭ и ЕГЭ» | гимназия №1 | 11 | Какаджанов Артур Илмуратович | 89141617834 | |||
| 01.11.2022 12:18:54 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ 37 | 9В | Молодцева Алиса Андреевна | +79141968033 | |||
| 01.11.2022 10:30:00 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ №42 | 8А | Глушакова Алина Евгеньевна | 89143169842 | |||
01. 11.2022 11.202209:21:26 | ИПиП Профессиональная проба «Педагог дошкольного образования и изобразительного искусства» | МОУ СОШ с УИОП №16 | 10Б | Сосина Виталина Витальевна | 89147792000 | |||
| 01.11.2022 00:15:42 | ИПиП Тренинг «Как преодолеть страх перед ОГЭ и ЕГЭ» | инженерная школа | 9а | Иманрммти мамин иоиит | 89095586522 | |||
| 31.10.2022 23:59:06 | ФФиМК Лингвистическая игра «Тайны фразеологии» | МОУ СОШ С УИОП 16 | 10А | Куберская Полина Степановна | 79242251090 | |||
31. 10.2022 10.202210:19:05 | ИПиП Мастер-класс по выполнению задания # 39 посменной части ЕГЭ по английскому языку | МБОУ СОШ 2 с.п. «Село Хурба» | 11 | Шульга Варвара Викторовна | 89638280255 | |||
| 30.10.2022 20:26:54 | ИПиП Мастер-класс по выполнению задания # 39 посменной части ЕГЭ по английскому языку | МОУ «Инженерная школа» | 11 А | Болдырев Семен | 89990876239 | |||
| 30.10.2022 20:24:22 | ИПиП Мастер-класс по выполнению задания # 39 посменной части ЕГЭ по английскому языку | МОУ «Инженерная школа» | 11 А | Рыжкина Александра Викторовна | 89141633647 | |||
| Всего: 183 | ||||||||
Результаты 1 — 10 из 183
Начало | Пред. | 1 2 3 4 5 | След. | Конец | Все
| 1 2 3 4 5 | След. | Конец | Все
определение прокариотов и эукариотов, особенности строения органов движения и их функции
Биология
12.11.21
11 мин.
Клетки классифицируются на прокариоты и эукариоты. В первую группу входят вирусы, зеленые водоросли, у которых имеется цитоплазма с молекулой ДНК или РНК. Эукариотические организмы содержат ядро с нуклеопротеидами. Особенности строения и функций движения органоидов (постоянные структуры растительных и животных клеток) изучаются на уроках биологии в 8 классе.
Оглавление:
- Бактерии и археи
- Ложноножки и миофибриллы
- Инфузория и амеба
- Зеленая эвглена
Клетки классифицируются на прокариоты и эукариоты. В первую группу входят вирусы, зеленые водоросли, у которых имеется цитоплазма с молекулой ДНК или РНК. Эукариотические организмы содержат ядро с нуклеопротеидами.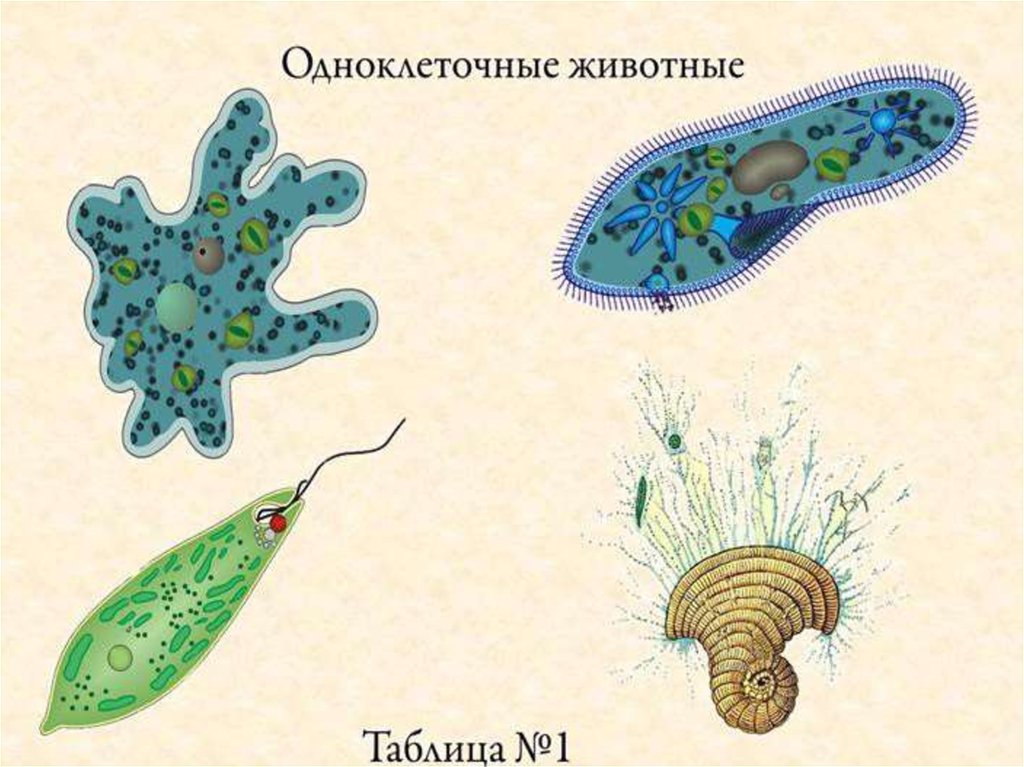 Особенности строения и функций движения органоидов (постоянные структуры растительных и животных клеток) изучаются на уроках биологии в 8 классе.
Особенности строения и функций движения органоидов (постоянные структуры растительных и животных клеток) изучаются на уроках биологии в 8 классе.
Основные понятия
Доядерные, или прокариоты, — одноклеточные организмы, у которых отсутствует ядро и мембрана. Прокариоты не способны дифференцироваться в многоклеточные, так как они не развиваются. Эукариоты — живые организмы с одной или несколькими клетками. К органам перемещения одноклеточных относятся жгутики, реснички, ложноножки, миофибриллы.
К многоклеточным относятся человек, животные, растения. На бронхиальном эпителии расположены реснички, защищающие дыхательные пути от инородных тел. К особенностям строения сперматозоидов можно отнести наличие жгутиков.
Под одним из методов проявления жизнедеятельности подразумевается такое движение в биологии, которое позволяет организму активно взаимодействовать со средой, включая:
- перемещаться в пространстве;
- захватывать пищу.

Принципы классификации
Органы движения в растительных и животных многоклеточных организмов являются органеллами, сформированными из белков и фосфолипидов.
Таблица с перечислением особенностей органоидов движения у простейших и многоклеточных
| Название органа | Структура | Функции | Вид организма |
| Жгутики | Наросты на мембране | Перемещение | Жгутиковая инфузория |
| Реснички | Тоненькие выросты на клетке | Вывод пыли | Ресничная инфузория |
| Ложноножки | Цитоплазма | Питание и движение | Амеба обыкновенная |
| Миофибриллы | Тоненькие нити | Сокращение мышц | Человек |
В человеческом организме могут бывать жгутики и реснички. Они очищают легкие, защищая эпителий, обеспечивая размножение.
Бактерии и археи
Жгутик является органоидом движения эукариотов. Он обеспечивает передвижение организмов в жидкости. Жгутики могут находиться в зооспорах, протистах, половых клетках. Они представлены в виде маленьких наростов, окруженных пленкой. Жгутики имеют цитоскелет, где происходит гидролиз АТФ.
Можно перечислить основные функции жгутиков:
- контакт с субстратами;
- иммунная защита.
Кроме общих функций, по строению реснички и жгутики относятся к комплексным структурным элементам. Жгутик — организм, какой состоит из 9 пар микротрубочек с мостиками. Дополнительно может существовать переходная область эксонемы. В центре находятся ось и центриоли.
Около 80% бактерий состоят из жгутиков, размещенных на противоположных их полюсах.
Отличительная характеристика жгутиков-бактерий — размещение в оболочках. Для вращения затрачивается энергия, получаемая при гидролизе АТФ.
Процесс перемещения осуществляется по часовой стрелке.
Кратко о строении бактериального жгутика:
- Базальное тело.
- Муреиновый чехол, окруженный оболочкой.
В отличие от бактерий, жгутики архей представлены в виде 7−13 разных генов. Для формирования структуры используются единые опероны. Если рассмотреть рисунок с ресничками, можно сделать вывод, что они являются тонкими органеллами в виде волосков. Реснички выполняют функции рецепторов. Они не двигаются.
Их поверхность покрыта мембраной. Реснички состоят из микротрубочек с белковыми структурами. В центре размещено базальное тело с центриолями. В процессе движения реснички ударяются. Направление и характер ударов зависят от количества входящих в состав ионов кальция.
Ложноножки и миофибриллы
К органеллам относятся псевдоподии, представленные в виде цитоплазматических выростов, у которых нет оболочки. Подобные органеллы называются ложноножками. Они всасывают питательные компоненты, обеспечивают перемещение организмов со скоростью до 0,2 мм/мин. В процессе движения органеллы закрепляются в субстрате, захватывая пищу, формируя пищеварительную вакуоль.
Они всасывают питательные компоненты, обеспечивают перемещение организмов со скоростью до 0,2 мм/мин. В процессе движения органеллы закрепляются в субстрате, захватывая пищу, формируя пищеварительную вакуоль.
Под воздействием нейронов органоиды уменьшаются. При сокращении выделяется энергия, необходимая для определения силы мышцы. При выполнении упражнений число миофибрилл увеличивается. Процесс называется гипертрофией. Если органоида большая, она делится на несколько маленьких. Травма способствует уменьшению числа миофибрилл и получению меньше энергии.
Инфузория и амеба
Инфузория туфелька — одноклеточный организм, который передвигается с помощью ресничек.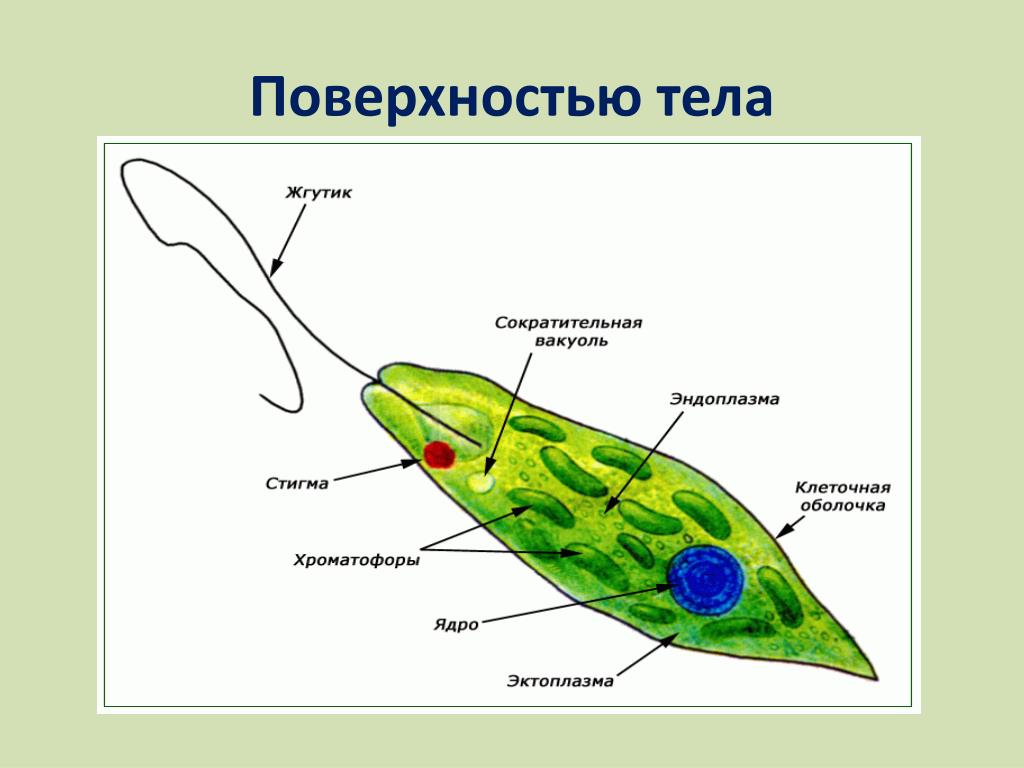 Они выполняют волнообразные движения. Движение осуществляется тупой частью вперед и только в одной плоскости. Они совершают удар после выпрямления. В выгнутом положении происходит возвратный удар. Между атаками наблюдается небольшая задержка.
Они выполняют волнообразные движения. Движение осуществляется тупой частью вперед и только в одной плоскости. Они совершают удар после выпрямления. В выгнутом положении происходит возвратный удар. Между атаками наблюдается небольшая задержка.
Скорость перемещения туфельки — 2,5 мм/с. Ее направление меняется при перегибе тела. При появлении на пути преграды происходит столкновение. Инфузория двигается в противоположную сторону. Для всех ресничек туфельки характерно одно строение со жгутиками зеленой эвглены. Вокруг основания ресничек образуется базальное зерно. Они участвуют в движении тела.
Для некоторых туфелек характерно соединение ресничек, что позволяет им двигаться с большой скоростью. Движение высокоорганизованных простейших осуществляется за счет сокращений. Мионемы (волокна) помогают организмам набрать большую скорость.
Размер простейшей амебы достигает до 0,5 мм. По форме тело полиподиальное, что объясняется наличием нескольких псевдоподий (выросты, у которых предусмотрена внутренняя циркуляция цитоплазмы).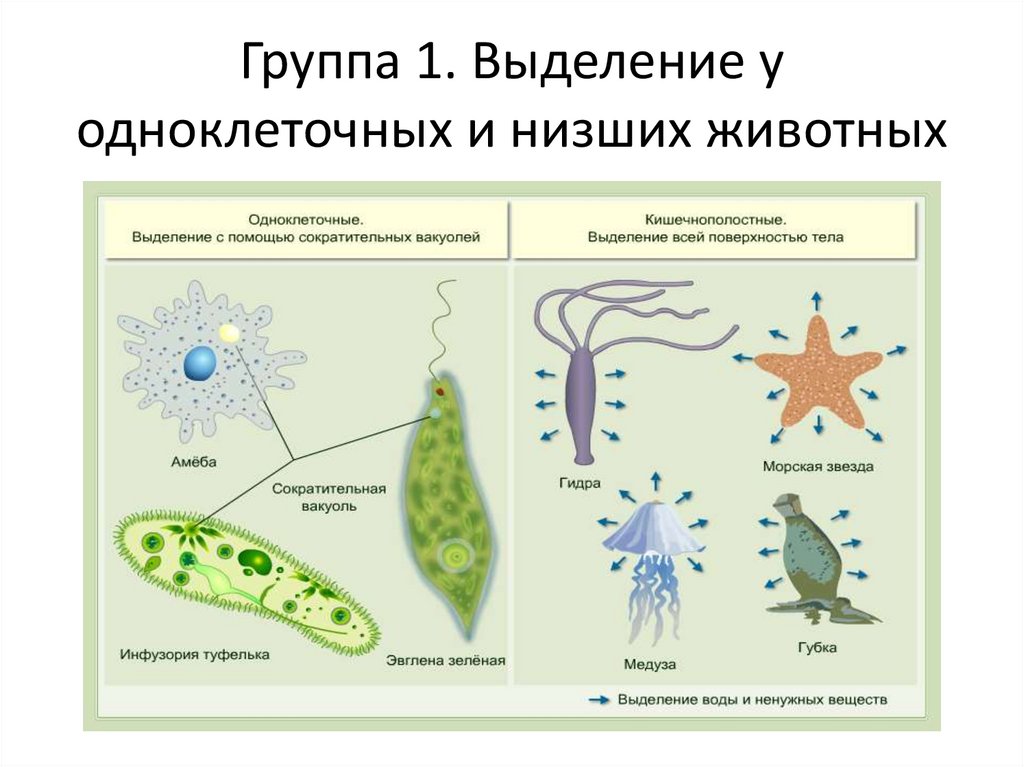 При движении они направляются в различные стороны. Скорость перемещения достигает 0,2 мм/мин.
При движении они направляются в различные стороны. Скорость перемещения достигает 0,2 мм/мин.
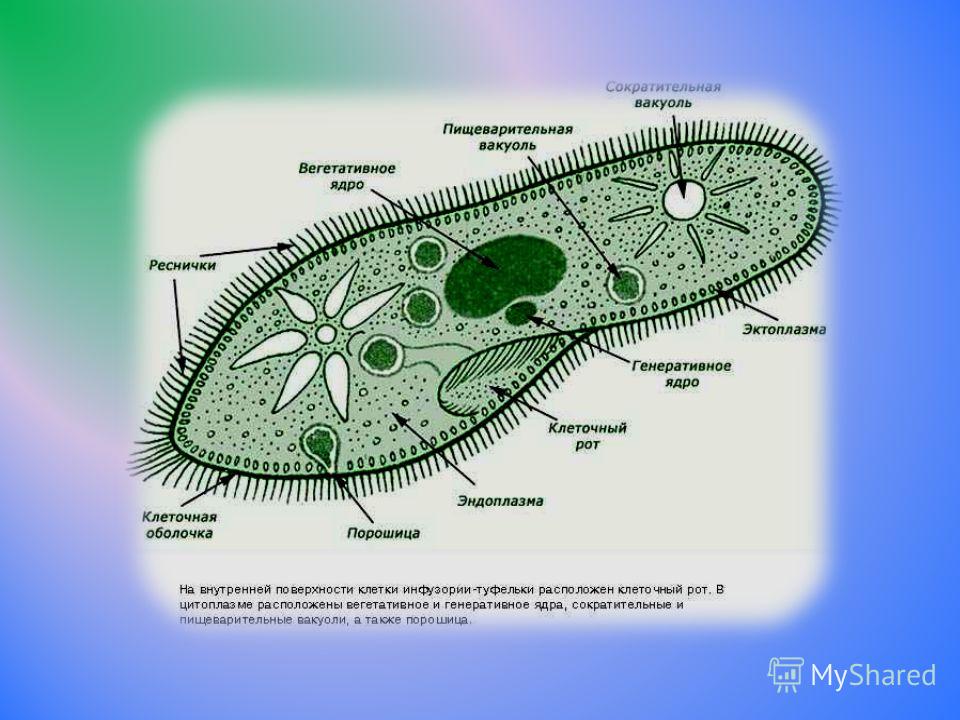
Зеленая эвглена
Простейшее относится к классу жгутиковые. Тело имеет веретенообразную форму с жгутиками, число которых достигает 10. Для каждого вида характерен определенный механизм движения. Большинство жгутиковых вращаются в виде конуса. Перемещение считается эффективным, если угол вершины конуса равен 45 °C.
Они могут совершать 40 оборотов за секунду. Вращательное движение может сопровождаться волнообразным покачиванием. Подобный характер перемещения характерен для одножгутиковых. Многожгутиковые имеют волосинки, расположенные в одной плоскости.
У зеленой эвглены жгутики покрыты оболочкой. Она является продолжением внешнего слоя (пелликула). Внутри жгутика существует цитоплазма. Если нити находятся по периферии, они отвечают за движение. Реснички и жгутики обеспечивают передвижение в пространстве, доставку питательных веществ к мембранам с последующим их проникновением в организм.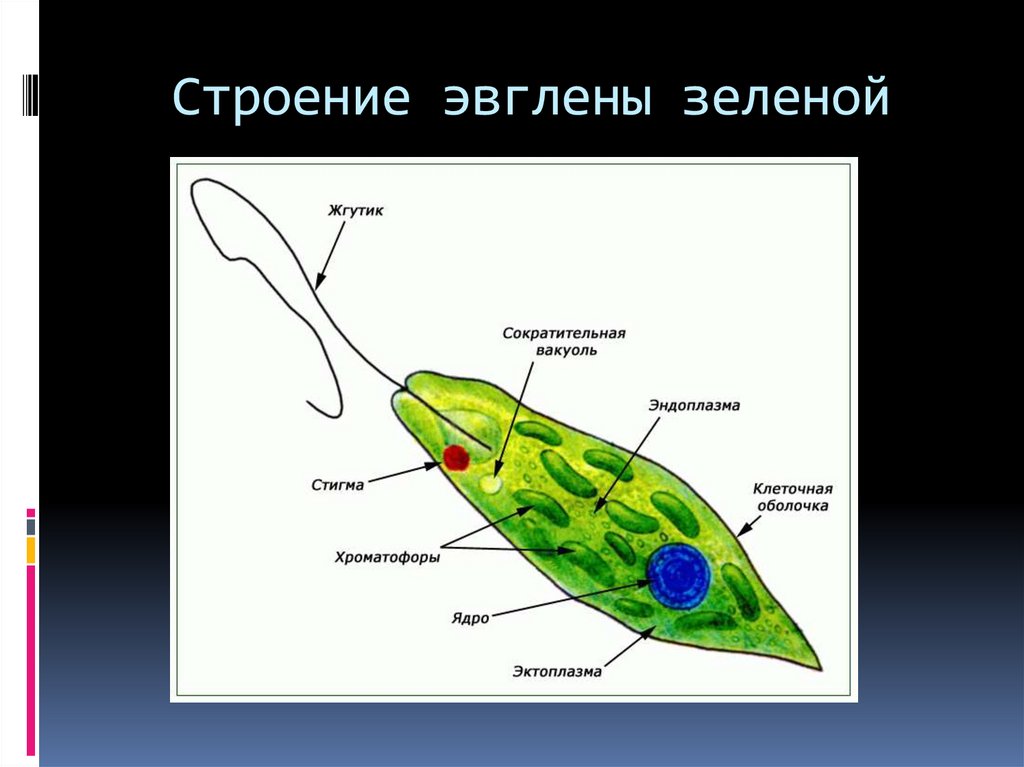
Определение и примеры одноклеточных — Биологический онлайн-словарь
Одноклеточные
прил.
[ˌjuːnɪˈsɛljʊlə]
Определение: Имеющие или состоящие только из одной клетки
Содержание
Одноклеточные организмы – это организмы, состоящие только из одной клетки, которые выполняют все жизненно важные функции, включая метаболизм, выделение и размножение. Одноклеточные организмы могут быть как прокариотами, так и эукариотами. Примерами одноклеточных организмов являются бактерии, археи, одноклеточные грибы и одноклеточные протисты. Хотя одноклеточные организмы не видны невооруженным глазом, они играют незаменимую роль в окружающей среде, промышленности и медицине. Некоторые из них также могут быть инфекционными или патогенными для человека, животных и растений.
Одноклеточный Определение
Что такое одноклеточный организм? В отличие от многоклеточных организмов одноклеточные организмы — или одноклеточные организмы — это группы различных живых организмов, состоящие только из одной клетки.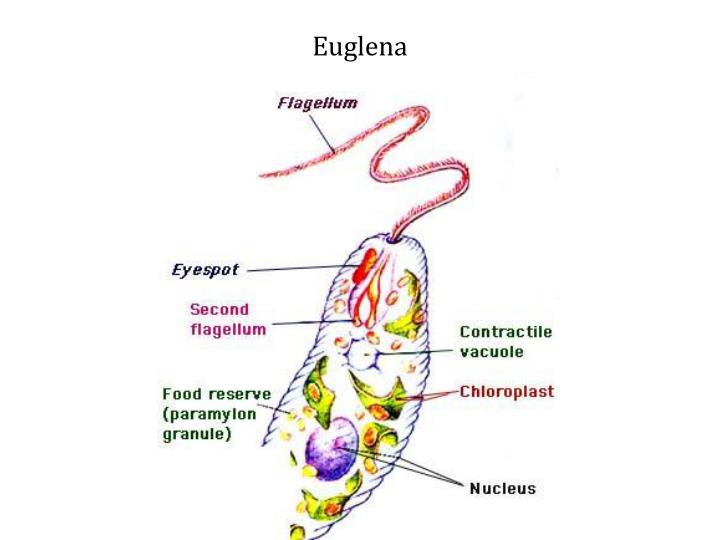 И эта клетка выполняет все жизненно важные функции, такие как гомеостаз, обмен веществ и размножение. Более того, отдельная клетка должна иметь возможность получать и использовать энергию, избавляться от отходов и транспортировать материалы. Напротив, многоклеточные организмы состоят из нескольких клеток, и эти клетки играют определенную роль и могут функционировать вместе как единое целое (ткань).
И эта клетка выполняет все жизненно важные функции, такие как гомеостаз, обмен веществ и размножение. Более того, отдельная клетка должна иметь возможность получать и использовать энергию, избавляться от отходов и транспортировать материалы. Напротив, многоклеточные организмы состоят из нескольких клеток, и эти клетки играют определенную роль и могут функционировать вместе как единое целое (ткань).
Клетка одноклеточного организма имеет протоплазму, содержащую различные белки, липиды, углеводы и нуклеиновые кислоты. Протоплазма окружена клеточной мембраной, которая отделяет внутренние компоненты клетки от внешней среды. Однако любая клетка должна иметь возможность взаимодействовать с внешней средой, чтобы получать молекулы извне и выводить наружу отходы.
Бактерии одноклеточные? Да! На самом деле одноклеточными являются не только бактерии, но и археи. И бактерии, и археи являются прокариотическими организмами. Однако одноклеточность свойственна не только прокариотам. Некоторые эукариоты также живут поодиночке. Примерами одноклеточных эукариот являются одноклеточные водоросли, одноклеточные грибы и простейшие.
Некоторые эукариоты также живут поодиночке. Примерами одноклеточных эукариот являются одноклеточные водоросли, одноклеточные грибы и простейшие.
Большинство живых существ, состоящих только из одной клетки, микроскопичны и не видны невооруженным глазом. Одноклеточные организмы изобилуют в природе. Даже в экстремальных местообитаниях встречаются одноклеточные организмы. Некоторые археи, например, могут выживать в экстремальных условиях, поэтому их называют экстремофилами . Обычно они устойчивы к экстремальным условиям, таким как температура или pH.
Рисунок 1: Что означает одноклеточный? Вот пример одноклеточного организма. Клетка способна жить отдельно или независимо от других клеток. Кредит: ck12.org Одноклеточные (биологическое определение): (1) Имеющие или состоящие только из одной клетки. (2) Относящийся к организму, все функции которого выполняются внутри одной клетки. Этимология: Латинское «ūni», «ūnus», что означает «один» + «cellularis», «cellul(a)», что означает живая клетка. Синоним: одноклеточный. Сравните: многоклеточный.
Синоним: одноклеточный. Сравните: многоклеточный.
Значение одноклеточных организмов
Одноклеточные организмы могут жить независимо, но они играют важную роль в экосистеме. Кроме того, они также используются в медицине, например, при разработке антибиотиков.
Экологические функции одноклеточных организмов
Несколько штаммов эубактерий полезны для окружающей среды, поскольку они разлагают разлагающееся и мертвое органическое вещество и фиксируют свои соединения в почве. Некоторые штаммы помогают бороться с загрязнением, например штаммы Pseudomonas , которые способны разлагать разливы нефти в почве и океанах. Другие штаммы бактерий могут разлагать тяжелые металлы в сточных водах. Эубактерии имеют промышленные преимущества. Они используются в производстве сыра, ликера и других пищеварительных продуктов. Эубактерии являются основным источником производства антибиотиков, поэтому они важны с медицинской точки зрения. Цианобактерии — это тип эубактерий, обладающих способностью осуществлять фотосинтез и способствующих выработке кислорода, поддерживающего кислородный баланс в атмосфере.
Цианобактерии — это тип эубактерий, обладающих способностью осуществлять фотосинтез и способствующих выработке кислорода, поддерживающего кислородный баланс в атмосфере.
Медицинское значение
Некоторые археи используются в фармацевтической промышленности для производства антибиотиков, которые по своей структуре и способу действия отличаются от бактериальных антибиотиков. Такие антибиотики могут лечить пациентов по разным механизмам, следовательно, они могут решить проблему устойчивости бактерий к антибиотикам.
Типы одноклеточных организмов
Одноклеточные организмы подразделяются на две основные группы: прокариоты, и эукариоты. Они находятся повсюду вокруг нас и даже внутри нашего тела. Но из-за крошечного размера клеток они не видны невооруженным глазом, если только не используется специальное оборудование, такое как микроскопы. Тем не менее, они могут образовывать колонии, что хорошо видно невооруженным глазом.
A. Прокариоты
Являются ли прокариоты одноклеточными? Прокариоты составляют подавляющее большинство одноклеточных организмов. Что такое прокариотическая клетка? Прокариот по определению означает отсутствие специализированных связанных с мембраной органелл, таких как ядро и митохондрии. Их ДНК свободна в цитоплазме в области, известной как нуклеоид .
Что такое прокариотическая клетка? Прокариот по определению означает отсутствие специализированных связанных с мембраной органелл, таких как ядро и митохондрии. Их ДНК свободна в цитоплазме в области, известной как нуклеоид .
Что такое прокариоты? Прокариоты — высокоадаптивные клетки к окружающим условиям. Прокариоты могут использовать органические и неорганические материалы для метаболизма. Примерами прокариотических клеток являются архей и эубактерий. Одним из способов отличить бактерии от архей является их клеточная стенка. Те бактерии, у которых есть клеточные стенки, имеют пептидогликан в своих клеточных стенках, тогда как клеточная стенка архей имеет псевдопептидогликана 9.0015 .
Рисунок 2: Схематическая диаграмма клеточной структуры прокариотического одноклеточного организма.1. Археи
Археи являются одним из древних представителей прокариот. Они размножаются почкованием, фрагментацией или бинарным делением.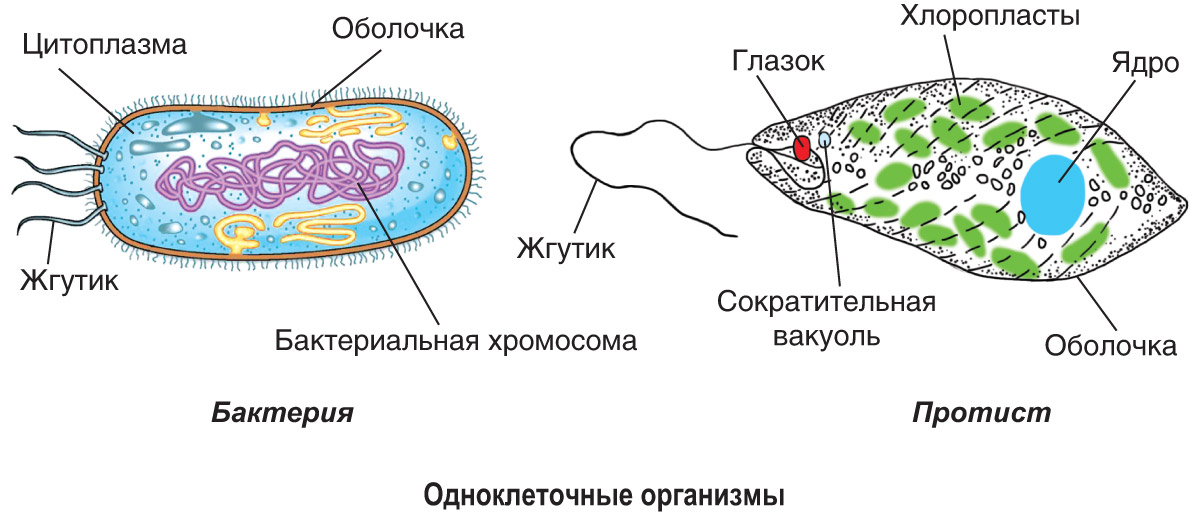 Археи обычно живут в экстремальных условиях, в то время как другие живут в нормальных условиях, таких как океаны, почвы и определенные части человеческого тела. Однако они обычно непатогенны для человека. Метаногены — это археи, способные производить газообразный метан. Они производят метан, восстанавливая углекислый газ до водорода и производя энергию в виде аденозинтрифосфата.
Археи обычно живут в экстремальных условиях, в то время как другие живут в нормальных условиях, таких как океаны, почвы и определенные части человеческого тела. Однако они обычно непатогенны для человека. Метаногены — это археи, способные производить газообразный метан. Они производят метан, восстанавливая углекислый газ до водорода и производя энергию в виде аденозинтрифосфата.
Бактерии одноклеточные? Домен eubacteria широко известен как бактерии; это одноклеточные организмы, которые живут почти повсюду вокруг нас. Являются ли бактерии прокариотами? Бактерии — наиболее распространенные прокариотические одноклеточные организмы. По своему строению они делятся на грамположительные и грамотрицательные бактерии. Бактерии живут? Да, бактерии — это живые организмы, которые обычно размножаются путем бинарного деления. Некоторые эубактерии обладают способностью образовывать споры, чтобы защитить свой генетический материал от повреждения окружающей средой, такой как радиация или химические вещества. Некоторые бактерии патогенны для человека и других живых существ. Другие, тем не менее, полезны и присутствуют в организме в виде нормальная микробиота , которая может либо защищать организм от вторжения микроорганизмов, либо вырабатывать витамины, которые помогают различным клеткам организма выполнять свои нормальные функции.
Некоторые бактерии патогенны для человека и других живых существ. Другие, тем не менее, полезны и присутствуют в организме в виде нормальная микробиота , которая может либо защищать организм от вторжения микроорганизмов, либо вырабатывать витамины, которые помогают различным клеткам организма выполнять свои нормальные функции.
B. Эукариоты
Могут ли эукариоты быть одноклеточными? Эукариоты могут быть одноклеточными или многоклеточными организмами. Однако одноклеточные эукариоты имеют общие отличительные черты многоклеточных эукариот на клеточном уровне. В частности, все эукариоты содержат связанные с мембраной органеллы, такие как ядро и митохондрии. Эукариоты размножаются половым путем или бесполым путем. При половом размножении увеличивается генетическое разнообразие, в отличие от бесполого размножения, при котором обычно образуется клон родителя. Тем не менее бесполое размножение способствует быстрому распространению организма.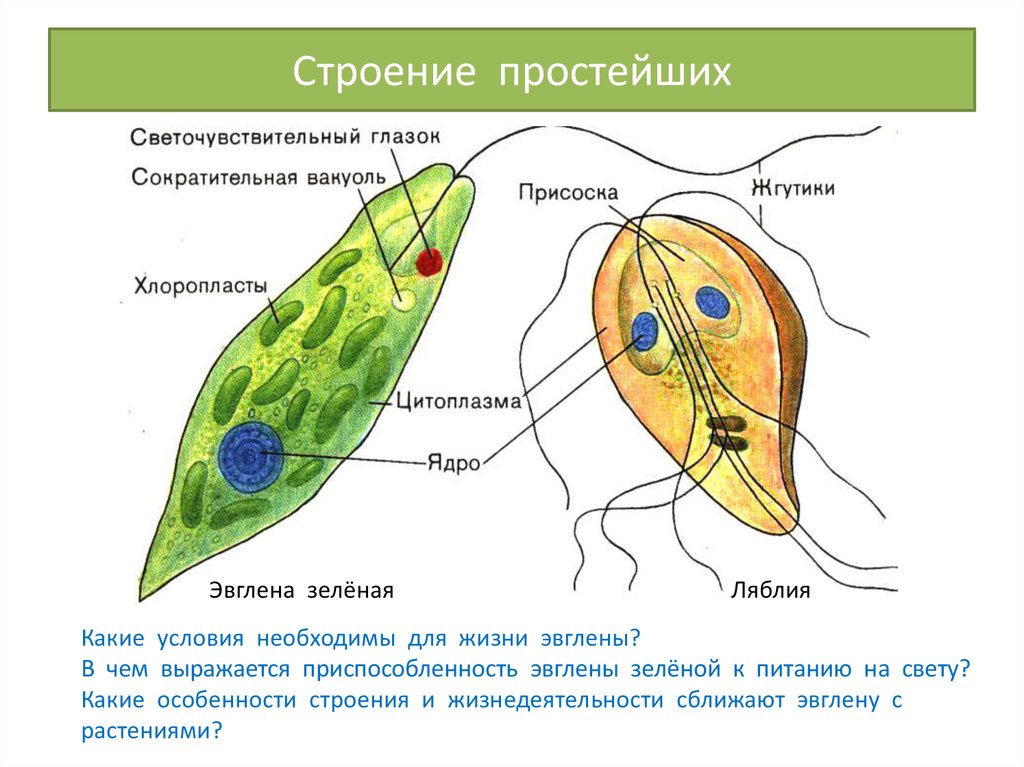 Примеры эукариотических одноклеточных организмов включают одноклеточные протисты и грибы.
Примеры эукариотических одноклеточных организмов включают одноклеточные протисты и грибы.
1. Протисты
Простейшие, водоросли, слизевики и водяные плесени относятся к королевству Protista. Есть ли у протистов ядро? Они эукариоты? Протисты имеют ядро, содержащее их генетический материал, поэтому они эукариоты.
- Простейшие: простейших — это одноклеточные эукариоты, обитающие в почве или воде. Они различаются по форме, размеру, способу питания и клеточной структуре. Простейшие могут быть обнаружены в организме как часть нормальной флоры или как источник болезней. Однако лишь немногие паразиты могут вызывать заболевания. Паразитические патогены, оказывающие большое влияние на здоровье, представлены споровиками Plasmodium spp. , которые вызывают малярию, одну из ведущих причин смертности среди африканских детей. Малярия передается человеку через инфицированных комаров. Они вводят споровики в кровь хозяина, вызывая малярию.
 Другие простейшие автотрофны, способны к фотосинтезу. Они свободно живут. Простейшие, являющиеся патогенными, имеют довольно сложный жизненный цикл, переходя от одного хозяина к другому. Простейшие обычно размножаются бесполым путем путем шизогонии, почкования или деления, тогда как лишь немногие простейшие, такие как Paramecium размножаются половым путем.
Другие простейшие автотрофны, способны к фотосинтезу. Они свободно живут. Простейшие, являющиеся патогенными, имеют довольно сложный жизненный цикл, переходя от одного хозяина к другому. Простейшие обычно размножаются бесполым путем путем шизогонии, почкования или деления, тогда как лишь немногие простейшие, такие как Paramecium размножаются половым путем.
Капсула вырабатывается некоторыми простейшими в определенных условиях для защиты организма. Это форма цисты простейших, и это стадия жизненного цикла, на которой организм может выжить в неблагоприятных условиях, включая истощение питательных веществ, воды и кислорода, или когда организм подвергается воздействию сильной жары или токсичных химических веществ. Киста также может защищать простейших за пределами хозяина, поскольку некоторые паразитические простейшие выделяются из организма одного хозяина и заражают другого.
- Водоросли: Многие водоросли являются одноклеточными.
 Обычно они встречаются на скалах, почве или в прибрежных водах. Самый крупный одноклеточный организм – одна из водных водорослей. Водоросли — фотоавтотрофы, подобные растениям. Однако из-за различий в строении растений и водорослей последняя группа не рассматривается как растения.
Обычно они встречаются на скалах, почве или в прибрежных водах. Самый крупный одноклеточный организм – одна из водных водорослей. Водоросли — фотоавтотрофы, подобные растениям. Однако из-за различий в строении растений и водорослей последняя группа не рассматривается как растения. - Плесневые грибы: Водяные и слизевики представляют собой одноклеточные организмы, питающиеся органическим разлагающимся веществом. Водяные плесени (оомицеты) нитевидные; они напоминают грибы. Однако, в отличие от грибов, имеющих хитиновые клеточные стенки, водяные плесени (как и слизевики) имеют целлюлозные клеточные стенки. Слизевики также являются одноклеточными, хотя при стрессе они способны превращаться в кажущиеся многоклеточными организмы.
2. Грибы
Грибы
Грибы одноклеточные или многоклеточные? Большинство грибов существуют как многоклеточные организмы, и лишь немногие из них одноклеточные. Дрожжи представляют собой одноклеточную форму грибов; они широко распространены и обычно находятся на покрытии листьев и плодов. Дрожжи, такие как Candida albicans , могут прикрепляться к эпителиальным клеткам человека и вызывать заболевание (кандидоз). Другие виды дрожжей также могут вызывать серьезные нечеловеческие заболевания. Обычно дрожжи размножаются либо почкованием, либо делением. Они могут расти и выживать в аэробных или анаэробных условиях. Когда кислорода много, они аэробно метаболизируют углеводы в воду и углекислый газ. В случаях истощения кислорода дрожжи выполняют анаэробную ферментацию углеводов с образованием этанола и углекислого газа. Эта форма ферментации используется в различных отраслях промышленности, таких как выпечка, производство вина и пивоварение.
Есть ли у грибов ядро? Поскольку грибы являются эукариотами, в их клетках есть ядро, которое несет их генетический материал.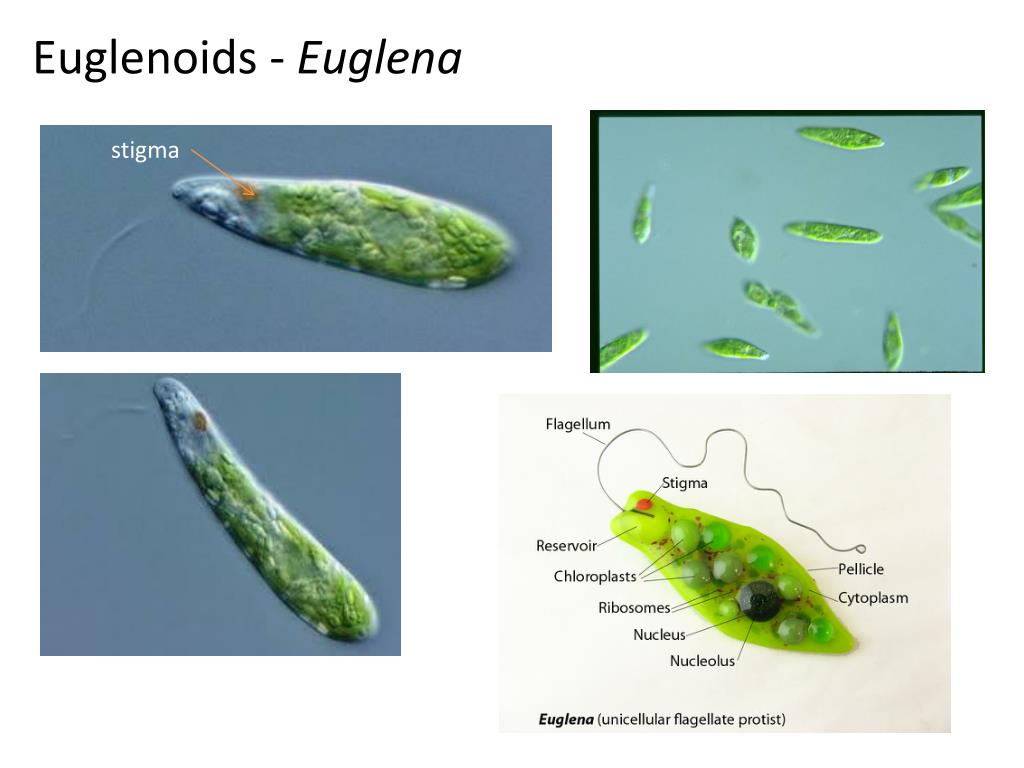 Грибы имеют важное экономическое значение, поскольку они используются во многих отраслях промышленности, они также несут ответственность за разложение мертвых веществ, поэтому они считаются важным компонентом пищевой цепи. Большинство растений зависят от грибов, которые помогают им поглощать воду и минералы из почвы корнями. Люди используют грибы, такие как грибы, в пищевой и фармацевтической промышленности.
Грибы имеют важное экономическое значение, поскольку они используются во многих отраслях промышленности, они также несут ответственность за разложение мертвых веществ, поэтому они считаются важным компонентом пищевой цепи. Большинство растений зависят от грибов, которые помогают им поглощать воду и минералы из почвы корнями. Люди используют грибы, такие как грибы, в пищевой и фармацевтической промышленности.
Эволюционная история одноклеточных организмов
Все клетки имеют некоторые общие черты, такие как хранение генетического материала в форме нуклеиновой кислоты, использование аденозинтрифосфата для получения энергии и будучи окружен клеточной мембраной. Эти сходства обусловлены эволюцией организмов и наличием общих предков. С другой стороны, различия между организмами являются результатом приспособления к окружающей среде посредством естественного отбора.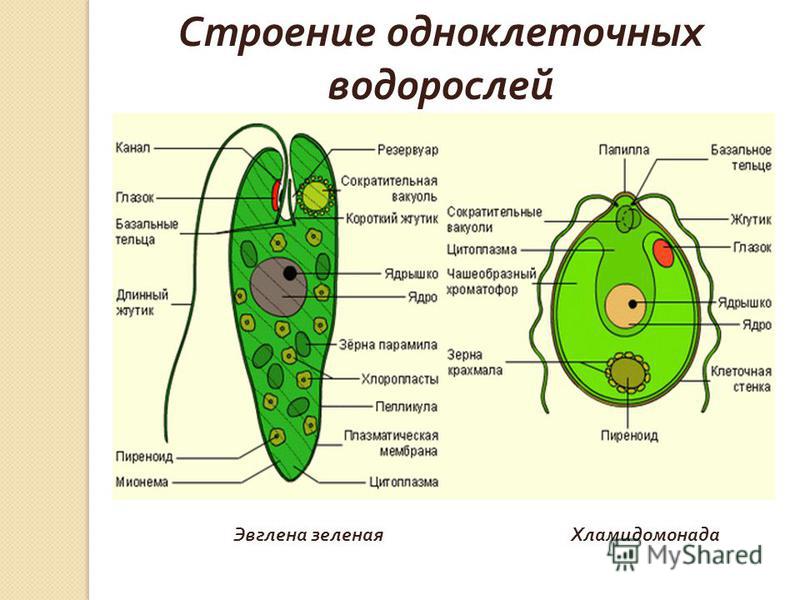
Существует три основных типа организмов, разделенных на три домена: (1) эукариоты (состоящие из эукариот) и два типа прокариот: (2) бактерии и (3) археи. Они были классифицированы в соответствии со структурой рибосом в каждом типе клеток.
Одноклеточные организмы возникли более 3 миллиардов лет назад, когда произошел горизонтальный перенос генов между разными видами, что в конечном итоге привело к образованию трех разных типов клеток. Эта теория была предложена после анализа геномов трех клеток и получения результатов, показывающих, что они имеют общий генетический материал.
Самая старая окаменелость, найденная на поверхности Земли, принадлежала одноклеточному прокариоту. Он образовался около 3,5 миллиардов лет назад. После этого совсем недавно, около 2,5 миллиардов лет назад, начали появляться эукариоты. Эндосимбиотическая теория показывает, что эукариоты произошли от прокариотических клеток, живших внутри эукариот. Например, сходство между митохондриальной рибосомой, которая является компонентом эукариотической клетки, и рибосомой бактериальной клетки свидетельствует о том, что эукариоты произошли от этих эндосимбиотических отношений между двумя прокариотами. Наука таксономия проясняет эволюционные отношения между различными видами.
Наука таксономия проясняет эволюционные отношения между различными видами.
Примеры одноклеточных организмов
Одноклеточные организмы необходимы для жизни и благополучия всех других существ на Земле. Они могут производить полезные вещества, разлагать мертвую материю и защищать других существ от некоторых инфекций. В этом разделе описаны некоторые одноклеточные примеры.
Фитопланктон
Фитопланктоны, т. е. водные одноклеточные протисты, являются примерами эукариотических клеток, обитающих либо в пресной, либо в соленой воде. Зеленые водоросли и диатомеи относятся к фитопланктону. Они осуществляют фотосинтез для получения энергии. Фитопланктон цветет, когда окружающие условия благоприятны. Эти цветы производят большое количество кислорода в земной атмосфере.
Amoeba
Amoeba является одним из примеров эукариотических клеток. Амеба вид. живут во влажной среде, такой как влажная почва, разлагающаяся растительность или внутри человеческого тела. Амебы используют свои щупальца для охоты на более мелкие бактерии, которыми они питаются. Эти щупальца называются ложноножками, они используются при движении, касании и охоте на добычу.
Амеба вид. живут во влажной среде, такой как влажная почва, разлагающаяся растительность или внутри человеческого тела. Амебы используют свои щупальца для охоты на более мелкие бактерии, которыми они питаются. Эти щупальца называются ложноножками, они используются при движении, касании и охоте на добычу.
Nitrosomonas и Nitrobacter
Nitrosomonas и Nitrobacter являются примерами прокариот. Эти бактерии могут использовать любой источник углерода, например различные источники энергии или углекислый газ, для создания сложных химических веществ, содержащих азот. Затем в результате окисления этих азотистых соединений нитрозомонас образуется нитрит. После этого нитраты образуются в результате окисления нитритов видами Nitrobacter. Продукт этого процесса используется в сельском хозяйстве.
Эти бактерии могут использовать любой источник углерода, например различные источники энергии или углекислый газ, для создания сложных химических веществ, содержащих азот. Затем в результате окисления этих азотистых соединений нитрозомонас образуется нитрит. После этого нитраты образуются в результате окисления нитритов видами Nitrobacter. Продукт этого процесса используется в сельском хозяйстве.
Эвглена
Еще один уникальный одноклеточный организм — Эвглена .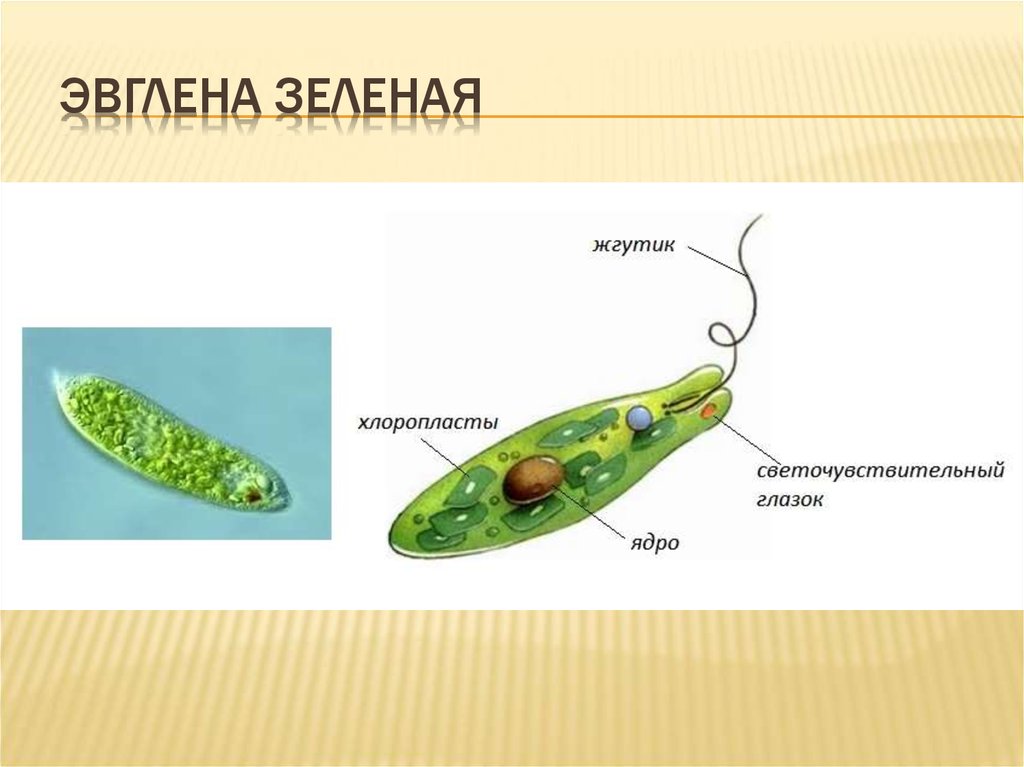 Этот вид может либо производить свою собственную пищу, либо получать ее из внешнего источника. Таким образом, эвглены рассматриваются как «гибриды растений и животных» одноклеточные организмы. Они в основном зеленые из-за питания зелеными водорослями при плохом освещении, которого недостаточно для осуществления фотосинтеза организмом. С другой стороны, если света достаточно, он будет производить кислород во время фотосинтеза.
Этот вид может либо производить свою собственную пищу, либо получать ее из внешнего источника. Таким образом, эвглены рассматриваются как «гибриды растений и животных» одноклеточные организмы. Они в основном зеленые из-за питания зелеными водорослями при плохом освещении, которого недостаточно для осуществления фотосинтеза организмом. С другой стороны, если света достаточно, он будет производить кислород во время фотосинтеза.
Попробуйте ответить на приведенный ниже тест, чтобы проверить, что вы уже узнали об одноклеточных.
Викторина
Выберите лучший ответ.
1. Что такое одноклеточное?
Организм с несколькими клетками
Организм, состоящий из одной клетки
Организм с клетками, действующими как единое целое
2. Что из следующего верно?
Все одноклеточные организмы являются прокариотами.
Все одноклеточные организмы одноклеточные.
Все одноклеточные организмы являются бактериями.
3. Что из перечисленного является примером одноклеточного организма?
Red blood cell
Muscles
Pathogen
4. Unicellular organisms lacking nucleus
Bacteria
Amoeba
Protists
5. Unicellular autotrophs
Amoebae
Oomycetes
Algae
Отправьте результаты (необязательно)
Ваше имя
На адрес электронной почты
Далее
Происхождение, потери и приобретения хлоропластов
Эйткен, А. и Р.Ю. Станьер. 1979. Характеристика пептидогликана цианелл Cyanophora paradoxa. J. Gen. Microbiol. 112:219–223.
и Р.Ю. Станьер. 1979. Характеристика пептидогликана цианелл Cyanophora paradoxa. J. Gen. Microbiol. 112:219–223.
Перекрёстная ссылка КАС Google ученый
Аллен, Дж.Ф., Н.Г. Холмс, К. В. Маллино и К. Э. Сандерс. 1986. Подробнее о стэкинге тилакоидных мембран. Тренды Биохим. науч. 11:320.
Перекрёстная ссылка КАС Google ученый
Бхаттачарья Д., Х.Дж. Элвуд, Л.Дж. Гофф и М.Л. Согин. 1990. Филогения Gracilaria lemaneiformis (Rhodophyta) на основе анализа последовательности области, кодирующей малую субъединицу рибосомной РНК. J. Phycol. 26:181–186.
Перекрёстная ссылка КАС Google ученый
Блобель Г. 1980. Топогенез внутриклеточных белков. Проц. Натл. акад. науч. США 77: 1496–1500.
Перекрёстная ссылка пабмед КАС Google ученый
Brandes, D. 1965. Наблюдения за очевидным способом образования чистых лизосом. Дж. Ультраструктур. Рез. 12:63–80.
1965. Наблюдения за очевидным способом образования чистых лизосом. Дж. Ультраструктур. Рез. 12:63–80.
Перекрёстная ссылка пабмед КАС Google ученый
Кавалье-Смит, Т. 1978. Эволюционное происхождение и филогения микротрубочек, митотических веретен и жгутиков эукариот. Биосистемы 10:93–114.
Перекрёстная ссылка пабмед КАС Google ученый
Кавалье-Смит, Т. 1981а. Эукариотические царства: семь или девять? Биосистемы 14:461–481.
Перекрёстная ссылка КАС Google ученый
Кавальер-Смит, Т. 1981b. Происхождение и ранняя эволюция эукариотической клетки. п. 33–84. В: Molecular and Cellular Aspects of Microbial Evolution , MJ Carlile, JF Collins, and B.E.B. Мозли (ред.). Симпозиум Общества общей микробиологии 32. Издательство Кембриджского университета.
Google ученый
Кавалье-Смит, Т. 1982. Происхождение пластид. БиоЛ Дж. Линн. соц. 17: 289–306.
Перекрёстная ссылка Google ученый
Кавалье-Смит, Т. 1983. Эндосимбиотическое происхождение митохондриальной оболочки. п. 265–279. В: Endocytobiology II ,W. Швеммлер и Х.Е.А. Шенк (ред.). де Грюйтер, Берлин.
Google ученый
Кавальер-Смит, Т. 1986. Царство Хромиста: происхождение и систематика. стр. 309–347. В: Прогресс в психологических исследованиях , том 4. Ф.Э. Раунд и Д.Дж. Чепмен (ред.). Биопресс Лтд., Бристоль.
Google ученый
Кавалье-Смит, Т. 1987а. Glaucophyceae и происхождение растений. Эволюция. Тенденции Растения 2:75–78.
Google ученый
Кавалье-Смит, Т. 1987b. Бактериальная сегрегация ДНК: ее двигатели и позиционный контроль. Дж. Теор. биол. 127:361–372.
1987b. Бактериальная сегрегация ДНК: ее двигатели и позиционный контроль. Дж. Теор. биол. 127:361–372.
Перекрёстная ссылка КАС Google ученый
Кавалер-Смит, Т. 1987c. Происхождение грибов и псевдогрибов. Симп. Брит. Микол. соц. 13:339–353. В: Эволюционная биология грибов , A.D.M. Райнер, К.М. Брейзер и Д. Мур (ред.). Издательство Кембриджского университета.
Google ученый
Кавалер-Смит, Т. 1987d. Одновременное симбиотическое происхождение митохондрий, хлоропластов и микротел. год. Академик Нью-Йорка науч. 503:55–71.
Перекрёстная ссылка КАС Google ученый
Кавалер-Смит, Т. 1987e. Происхождение клеток эукариот и архебактерий. год. Академик Нью-Йорка науч. 503:17–54.
Перекрёстная ссылка КАС Google ученый
Кавалер-Смит, Т. 1989. Королевство Хромиста. п. 379–405. In: Хромофитные водоросли: проблемы и перспективы , JC Green, BSC Ледбитер и У.К. Дайвер (ред.). Издательство Оксфордского университета.
1989. Королевство Хромиста. п. 379–405. In: Хромофитные водоросли: проблемы и перспективы , JC Green, BSC Ледбитер и У.К. Дайвер (ред.). Издательство Оксфордского университета.
Google ученый
Кавалье-Смит, Т. 1990а. Мегаэволюция микроорганизмов: объединение живых и ископаемых свидетельств. Ред. Micropal. 33:145–154.
Google ученый
Кавальер-Смит, Т. 1990b. Симбиотическое происхождение пероксисом. п. 515–521. В: Эндоцитобиология IV , П. Нардон, В. Джанинацци-Пирсон, А.М. Гренье, Л. Маргулис и Д.К. Смит (ред.). Национальный институт агрономических исследований, Париж.
Google ученый
Кавальер-Смит, Т. 1991а. Эволюция прокариотических и эукариотических клеток. п. 217–272. В: Основы медицинской клеточной биологии , Vol. I , Г.Э. Биттар (ред.). Дж.А.И. Press, Гринвич, Коннектикут.
Press, Гринвич, Коннектикут.
Google ученый
Кавалер-Смит, Т. 1991b. Эволюция клеток. п. 271–304. В: Эволюция жизни. Материалы конференции Конгресса Института перспективных исследований в Киото , С. Осава и Т. Хондзё (ред.). Спрингер-Верлаг, Токио.
Google ученый
Кавалер-Смит, Т. 1991c. Филогения интронов: новая гипотеза. Тенденции Жене. 7: 145–148.
Перекрёстная ссылка КАС Google ученый
Кавальер-Смит, Т. 1991d. Диверсификация клеток у гетеротрофных жгутиконосцев. п. 113–131. В: Биология свободноживущих гетеротрофных жгутиконосцев , Д.Дж. Паттерсон и Дж. Ларсен (ред.). Кларендон Пресс, Оксфорд.
Google ученый
Кавалер-Смит, Т. 1991e. Архамебы: предки эукариот? Биосистемы 24:25–38.
Перекрёстная ссылка Google ученый
Кавальер-Смит, Т. 1992а. Царство Protozoa и его 16 типов. J. Протозоол. , В печати. Кавалье-Смит, Т. 1992b. Происхождение хлоропластов и царство Plantae. Доп. Бот. Рез. , В печати.
1992а. Царство Protozoa и его 16 типов. J. Протозоол. , В печати. Кавалье-Смит, Т. 1992b. Происхождение хлоропластов и царство Plantae. Доп. Бот. Рез. , В печати.
Google ученый
Кавалье-Смит, Т. и Дж.Дж. Ли. 1985. Простейшие как хозяева эндосимбиозов и превращение симбионтов в органеллы. J. Протозоол. 32: 376–379.
Google ученый
Dodge, JD 1985. Повторное изучение взаимоотношений между одноклеточным хозяином и эукариотическим эндосимбионтом со специальной ссылкой на Glenodinium foliaceum Dinophyceae. п. 1015–1026. В: Endocytobiology II , H.E.A. Шенк и В. Швеммлер (ред.). де Грюйтер, Берлин.
Google ученый
Дус, Р. и Дж. Джоярд. 1981. Происходит ли пластидная оболочка из эндоплазматического ретикулума? Тренды Биохим. науч. 6: 237–239.
Перекрёстная ссылка КАС Google ученый
Дорн, А.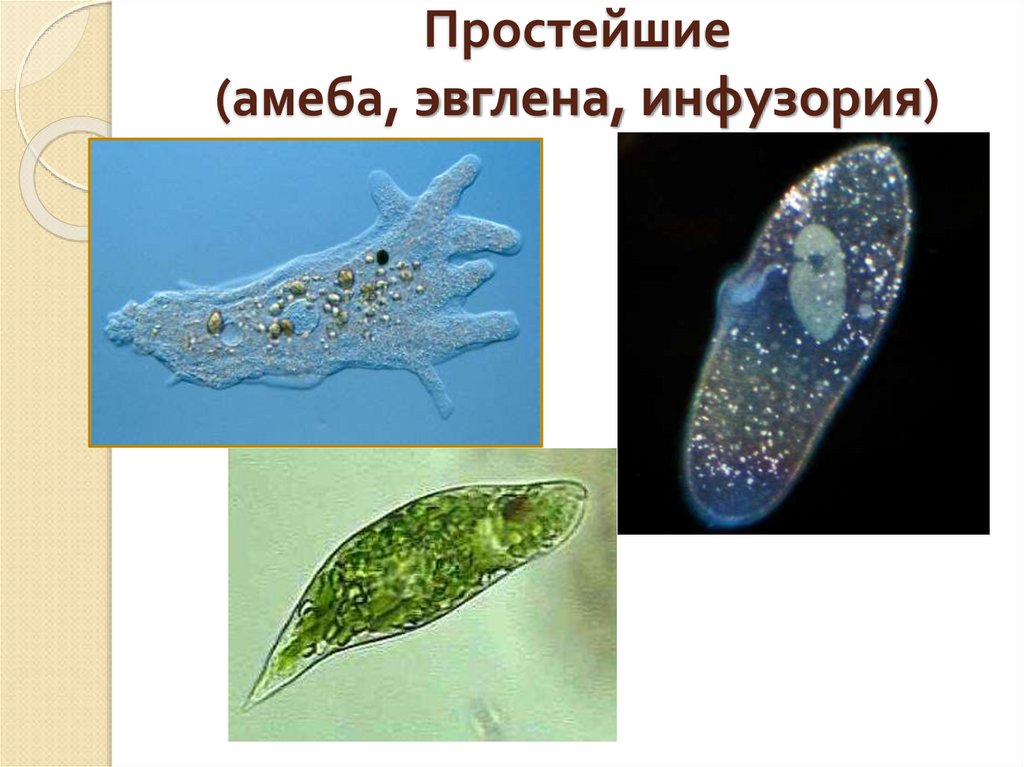 -Дж., Дж. Джоярд, М.А. Блок и Р. Дус. 1985. Локализация фосфатидихолина во внешней оболочке мембраны хлоропластов шпината. J. Cell Biol. 100:1690–1697.
-Дж., Дж. Джоярд, М.А. Блок и Р. Дус. 1985. Локализация фосфатидихолина во внешней оболочке мембраны хлоропластов шпината. J. Cell Biol. 100:1690–1697.
Перекрёстная ссылка пабмед КАС Google ученый
Дорн, А.-Дж., Дж. Джоярд и Р. Дус. 1990. Действительно ли тилакоиды содержат фосфатидилхолин? Проц. Натл. акад. науч. США 87:71–74.
Перекрёстная ссылка пабмед КАС Google ученый
Догерти, Э.К. и М.Б. Аллен. 1960. Является ли пигментация ключом к филогении протистов? п. 129–144. В: Comparative Biochemistry of Photoreactive Systems , E.C. Dougherty and M.B. Аллен (ред.). Академик Пресс, Нью-Йорк.
Google ученый
Дуглас, С.Э., Калифорния Мерфи, Д.Ф. Спенсер и М. В. Грей. 1991. Водоросли криптомонады представляют собой эволюционные химеры двух филогенетически различных одноклеточных эукариот. Природа 350:148–151.
Природа 350:148–151.
Перекрёстная ссылка пабмед КАС Google ученый
Эшбах, С., С. Дж. Б. Хофманн, У.-Г. Майер, П. Ситте и П. Хансманн. 1991. Эукариотический геном размером 660 т.п.н.: электрофоретический кариотип нуклеоморфа и ядра клетки криптомонадной водоросли, Пиреномонады салина. Нуклеиновые Кислоты Res. 19: 1779–1781.
Перекрёстная ссылка пабмед КАС Google ученый
Эврард, Дж.Л., М. Кунц и Дж.Х. Weil. 1990. Нуклеотидная последовательность пяти генов рибосомных белков цианелл Cyanophora paradoxa: значение, касающееся филогенетических отношений между цианеллами и хлоропластами. Дж. Мол. Эвол. 30:16–25.
Перекрёстная ссылка пабмед КАС Google ученый
Фармер, Массачусетс и Р.Э. Тример. 1991. Таксономическое переописание эвгленоидных жгутиконосцев на основе ультраструктурных признаков. J. Phycol. 27:20.
J. Phycol. 27:20.
Google ученый
Галло, Ж.-М. и Дж. Шревел. 1985. Гомологии между белками паражгутиковых стержней трипаносом и эвгленоидов, выявленные с помощью моноклональных антител. евро. Дж. Клеточная биология. 36:163–168.
ПабМед КАС Google ученый
Gibbs, S.P. 1978. Хлоропласты Euglena , возможно, произошли от симбиотических зеленых водорослей. Кан. Дж. Бот. 56:2883–2889.
Перекрёстная ссылка Google ученый
Гиббс, С.П. 1981. Эндоплазматический ретикулум хлоропласта, структура, функция и эволюционное значение. Междунар. Преподобный Цитол. 72:49–99.
Перекрёстная ссылка Google ученый
Gillott, MA and S.P. Gibbs. 1980. Нуклеоморф криптомонады: ее ультраструктура и эволюционное значение. J. Phycol. 16: 558–568.
J. Phycol. 16: 558–568.
Перекрёстная ссылка Google ученый
Серый, M.W. и W.F. Дулитл. 1982. Доказана ли эндосимбионтная гипотеза? Микробиолог. Откр. 46:1–42.
ПабМед КАС Google ученый
Гринвуд, AD 1974. Cryptophyta в отношении филогении и фотосинтеза. 8-й Международный конгресс по электронной микроскопии, Канберра.
Google ученый
Гроссман, А., А. Манодори и Д. Снайдер. 1990. Светособирающие белки диатомовых водорослей — их связь со связывающими хлорофилл и белками высших растений и их способ транспорта в пластиды. Мол. Генерал Жене. 224:91–100.
Перекрёстная ссылка пабмед КАС Google ученый
Хансманн, П. и С. Эшбах. 1990. Выделение и предварительная характеристика ядра и нуклеоморфа криптомонады Pyrenomonas salina.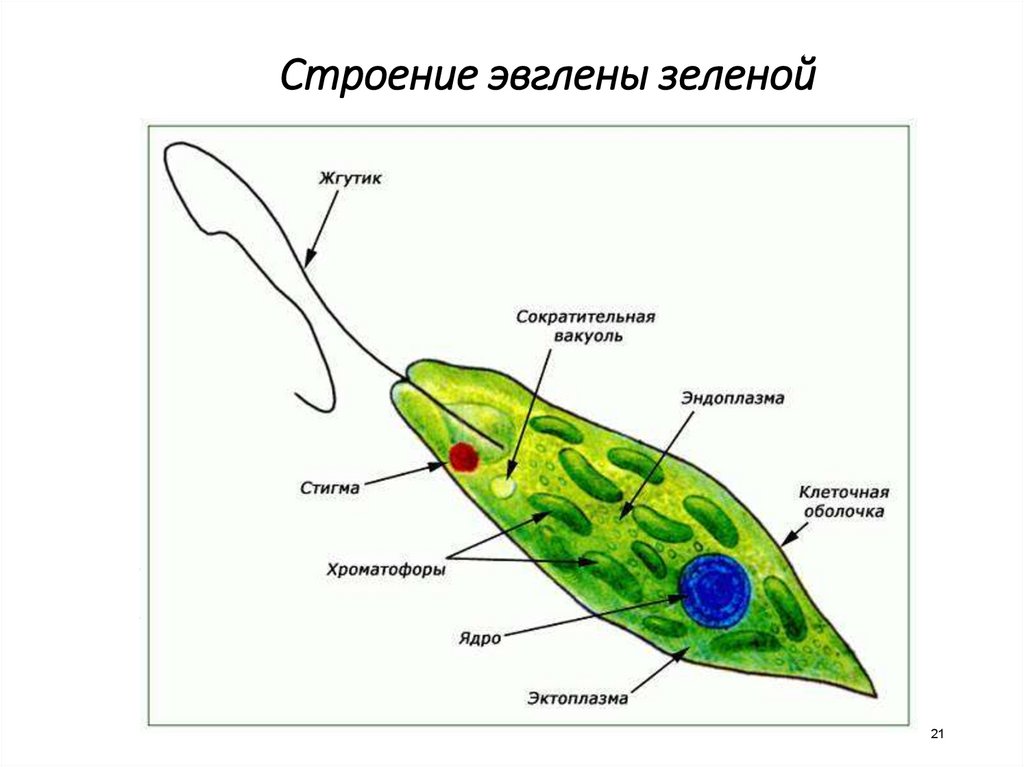 Евро. Дж. Клеточная биология. 52: 373–378.
Евро. Дж. Клеточная биология. 52: 373–378.
ПабМед КАС Google ученый
Хендрикс, Л., Р. Де Баре, Ю. Ван де Пер, Дж. Нифс, А. Горис и Р. Де Вахтер. 1991. Эволюционное положение родофита Porphyra umbilicalis и базидиомицета Leucosporidium scottii среди других эукариот, выведенное из полных последовательностей малой рибосомной субъединицы РНК. Дж. Мол. Эвол. 32:167–177.
Перекрёстная ссылка пабмед КАС Google ученый
Хердман, М. и Р.Ю. Станьер. 1977. Цианелла: хлоропласт или эндосимбиотический прокариот? FEMS Письмо. 1:7–12.
Перекрёстная ссылка КАС Google ученый
Хибберд, Д.Дж. и Р.Э. Норрис. 1984. Цитология и ультраструктура Chlorarachnion reptans (Chlorarachniophyta divisio nova , Chlorarachniophyceae classis nova).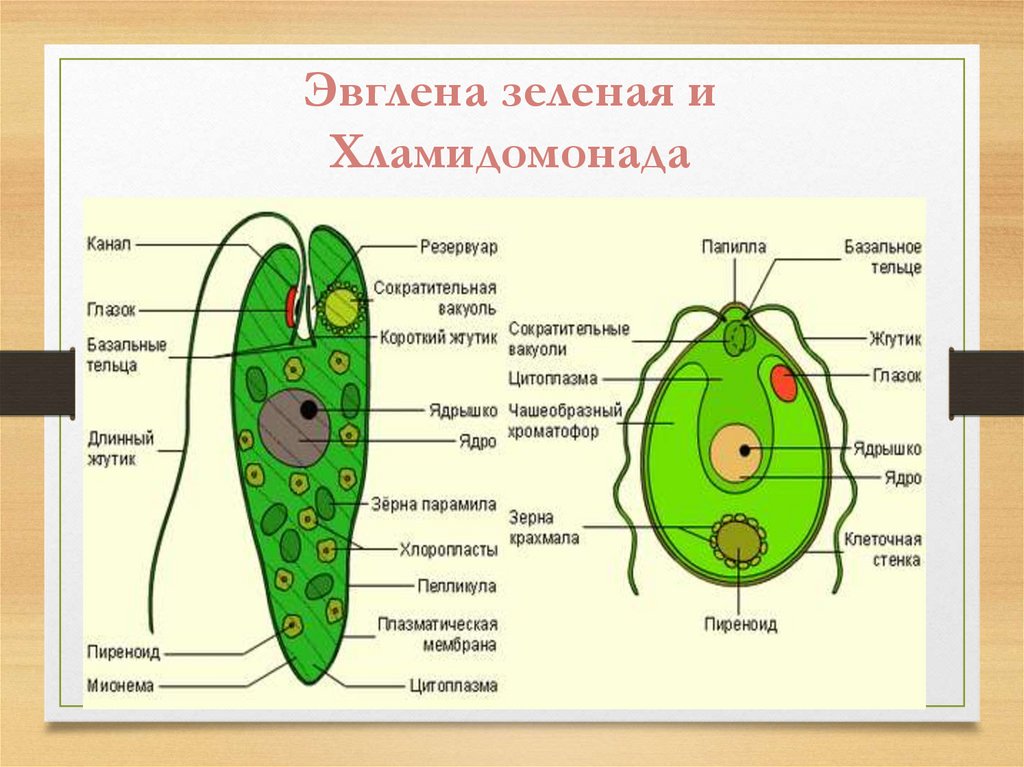 Дж. Фикол. 20:310–330.
Дж. Фикол. 20:310–330.
Перекрёстная ссылка Google ученый
Хёфельд И., Дж. Оттен и М. Мелконян. 1988. Сократительные жгутики эукариот: задействован центрин. Протоплазма 147:16–24.
Перекрёстная ссылка Google ученый
Джеффри, К. 1971. Таллофиты и царства — критика. Кью. Бык. 25: 291–299.
Перекрёстная ссылка Google ученый
Джоярд Дж., М.А. Блок, А.-Дж. Дорн и Р. Дус. 1987. Сравнение оболочек пластид высших растений и водорослей и наружных мембран цианобактерий (сине-зеленых водорослей). п. 123–133. В: Развитие водорослей (молекулярные и клеточные аспекты). В. Визнер, Д.Г. Робинсон и Р.К. Старр (ред.). Springer-Verlag, Берлин.
Google ученый
Kies, L. 1988. Влияние пенициллина на морфологию и ультраструктуру Cyanophora, Gloeochaete и Glaucocystis (Glaucocystophyceae) и их цианелл. Междунар. Дж. Эндоцитоб. Сотовый рез. 5: 361–372.
Междунар. Дж. Эндоцитоб. Сотовый рез. 5: 361–372.
Google ученый
Кивич П.А. и П.Л. Уолн. 1984. Оценка возможной филогенетической связи между Euglenophyta и Kinetoplastida. Происхождение Жизнь 13:269–288.
Перекрёстная ссылка Google ученый
Кляйн, Р. и А. Кронквист. 1967. Рассмотрение эволюционного и таксономического значения некоторых биохимических, микроморфологических и физиологических признаков таллофитов. Q. Rev. Biol. 42:105–296.
ПабМед КАС Google ученый
Нолл, А. Х. 1989. Эволюция и исчезновение в морской среде; некоторые ограничения, налагаемые фитопланктоном. Фил. Транс. Рой. Соц Лонд. Б, 325:229–290.
Перекрёстная ссылка Google ученый
Лахти, Си Джей и П. Джей Джонсон. 1991. Trichomonas vaginalis гидрогеносомные белки синтезируются на свободных полирибосомах и могут подвергаться процессингу при созревании. Мол. Биохим. Паразитол. 46:307–310.
Trichomonas vaginalis гидрогеносомные белки синтезируются на свободных полирибосомах и могут подвергаться процессингу при созревании. Мол. Биохим. Паразитол. 46:307–310.
Перекрёстная ссылка пабмед КАС Google ученый
Ламберт Д.Х., Д.А. Брайант, В.Л. Стируолт, Дж. М. Даббс, С.Э. Стивенс и Р. Д. Портер. 1985. Карта генов для генома Cyanophora paradoxa cyanelle. J. Бактериол. 164: 659–664.
ПабМед КАС Google ученый
Ларуш Л., К. Тремблей, К. Симар и Г. Белленев. 1991. Характеристика кДНК, кодирующей связанный с PSII хлорофилл alb , связывающий белок (Cab) из Chlamydomonas moewusii , не относящийся ни к типу I, ни к типу II. Курс. Жене. 19: 285–288.
Перекрёстная ссылка пабмед КАС Google ученый
Лаутерборн, Р. 1895. Protozoenstudien. II. Paulinella chromatophora нов. ген., нояб. spec., ein beschalter Rhizopode des Süsswassers mit blaugrünen chromatophoren-artigen Einschlüssen. З. Висс. Зоол. 59: 537–544.
1895. Protozoenstudien. II. Paulinella chromatophora нов. ген., нояб. spec., ein beschalter Rhizopode des Süsswassers mit blaugrünen chromatophoren-artigen Einschlüssen. З. Висс. Зоол. 59: 537–544.
Google ученый
Лефорт-Тран, М. 1981. Трехслойная организация хлоропластной оболочки Euglena (значение и функции). Бер. Д. Ч. Бот. Гэс. 94: 463–476.
Google ученый
Левин Р.А. и Л. Ченг. 1989. Прохлорон: микробная загадка. Чепмен Холл, Нью-Йорк.
Перекрёстная ссылка Google ученый
Ли, К.-В. и быть. Вулканы. 1987. Четыре новых апохлоротических диатомовых водоросли. руб. Фикол. Дж. 22: 375–382.
Перекрёстная ссылка Google ученый
Лонсдейл, Д.М., Т.П. Ходж, С.Дж. Хоу и Д.Б. Стерн. 1983. Митохондриальная ДНК кукурузы содержит последовательность, гомологичную гену большой субъединицы рибулозо-1,5-бисфосфаткарбоксилазы ДНК хлоропластов. Сотовый 34:1007–1014.
1983. Митохондриальная ДНК кукурузы содержит последовательность, гомологичную гену большой субъединицы рибулозо-1,5-бисфосфаткарбоксилазы ДНК хлоропластов. Сотовый 34:1007–1014.
Перекрёстная ссылка пабмед КАС Google ученый
Людвиг, М. и С.П. Гиббс. 1985. ДНК присутствует в нуклеоморфе криптомонад: еще одно свидетельство того, что хлоропласт произошел от эукариотического эндосимбионта. Протоплазма 127:9–20.
Перекрёстная ссылка Google ученый
Манодори А. и А.Р. Гроссман. 1990. Гомология последовательностей светособирающих полипептидов растений и диатомей Phaeodactylum tricornutum. Том. III, с. 541–544. В: Текущие исследования в области фотосинтеза , М. Балчевски (ред.). Клювер, Дордрехт.
Google ученый
Маргулис, Л. 1970. Происхождение эукариотических клеток. Издательство Йельского университета, Нью-Хейвен.
Издательство Йельского университета, Нью-Хейвен.
Google ученый
Маргулис, Л. 1981. Симбиоз в эволюции клеток. Фриман, Сан-Франциско.
Google ученый
Маргулис, Л. и Р. Обар. 1985. Heliobacterium и происхождение хризопластов. Биосистемы 17:317–325.
Перекрёстная ссылка пабмед КАС Google ученый
Мацубаяси Т., Т. Вакасуги, К. Шинозаки, К. Ямагути-Шинозаки, Н. Заита, Т. Хидака, Б.Ю. Мэн, К. Охто, М. Танака, А. Като, Т. Маруяма, и М. Сугиура. 1987. Активно экспрессируются шесть хлоропластных генов (ndhA-F) , гомологичных митохондриальным генам человека, кодирующим компоненты НАДН-дегидрогенозы дыхательной цепи: определение сайтов сплайсинга в пре-мРНК ndhA и ndhB . Мол. Генерал Жене. 210:385–393.
Перекрёстная ссылка пабмед КАС Google ученый
Макфадден, Г. И. 1990. Доказательства того, что хлоропласты криптомонад произошли от фотосинтезирующих эукариотических эндосимбионтов. J. Cell Sci. 95:303–308.
И. 1990. Доказательства того, что хлоропласты криптомонад произошли от фотосинтезирующих эукариотических эндосимбионтов. J. Cell Sci. 95:303–308.
КАС Google ученый
Мерешковский, К. 1905. Über Natur и Ursprung der Chromatophoren im Pflanzenreiche. биол. центральный б. 25: 593–604.
Google ученый
Мерещковский, К. 1910. Теория плазменного искусства и основы симбиогенеза, einer neuen Lehre von der Entstehung der Organismen. биол. центральный б. 30:278–303, 321–347, 353–367.
Google ученый
Михаловски, С. Б., Б. Пфанцагль, В. Лёффельхардт и Х. Дж. Бонерт. 1990. Оперон гена рибосомного белка cyanelle S10 spc из Cyanophora paradoxa. Мол. Генерал Жене. 224:222–231.
Перекрёстная ссылка пабмед КАС Google ученый
Миньо, Дж.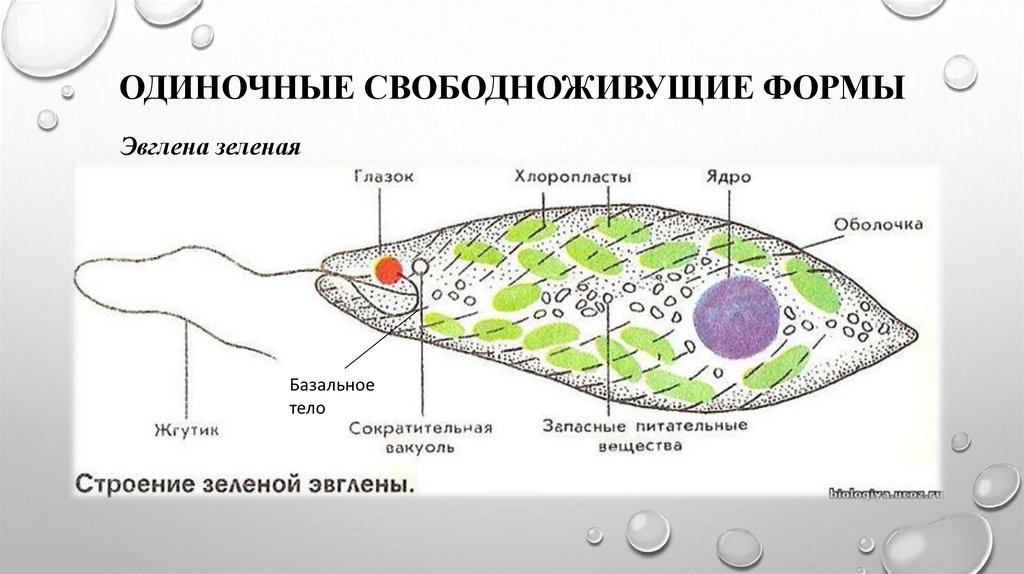 П. и Э. Брюгероль. 1975. Ультраструктурный этюд жгутиковых фаготрофов Colponema loxodes Stein. Protistologica 11:429–444.
П. и Э. Брюгероль. 1975. Ультраструктурный этюд жгутиковых фаготрофов Colponema loxodes Stein. Protistologica 11:429–444.
Google ученый
Моструп, Ø. 1978. О филогенетической достоверности жгутикового аппарата зеленых водорослей и других растений, содержащих хлорофилл а и b . Биосистемы 10:117–144.
Перекрёстная ссылка пабмед КАС Google ученый
Нойманн-Спалларт, К., Дж. Якович, М. Краус, М. Брандтнер, Х. Дж. Бонерт и В. Лёффельхардт. 1991. rps10 , не зарегистрированный для пластидных ДНК, расположен в геноме цианеллы Cyanophora paradoxa и котранскрибируется с генами оперона str . Курс. Жене. 19:313–315.
Перекрёстная ссылка пабмед КАС Google ученый
Окли, Б.Р. и Ф.Дж.Р. Тейлор. 1978. Доказательства нового типа эндосимбиотической организации в популяции инфузории Mesodinium rubrum из Британской Колумбии. Биосистемы 10:361–369.
Биосистемы 10:361–369.
Перекрёстная ссылка пабмед КАС Google ученый
Осафунэ Т., Дж.А. Шифф и Г. Хасе. 1990. Иммунолокализация апопротеина LHCPII в Golgi Euglena. р. 735–738. В: Текущие исследования в области фотосинтеза , Том. III , М. Бальчевский (ред.). Клювер, Дордрехт.
Google ученый
Pascher, A. 1929. Über die Natur der blaugrünen Chromatophoren des Rhizopoden Paulinella chromatophora. Зоол. Анц. 81:189–194.
Google ученый
Перассо, Р., А. Бароин, Л. Х. Ку, Дж. П. Бачеллери и А. Адаутт. 1989. Происхождение водорослей. Природа 339:142–144.
Перекрёстная ссылка пабмед КАС Google ученый
Пичерский Э., Дж. М. Логсдон, Дж. М. МакГрат и Р.А. Стасис. 1991. Фрагменты пластидной ДНК в ядерном геноме томата: распространенность, хромосомное расположение и возможный механизм интеграции. Мол. Генерал Жене. 225:453–458.
Перекрёстная ссылка пабмед КАС Google ученый
Раган, Массачусетс, 1989. Биохимические пути и филогения эукариот. п. 145–160. В: Иерархия жизни , Б. Фернхольм, К. Бремер и Х. Йорнвалл (ред.). Эльзевир.
Google ученый
Рэйвен, П.Х. 1970. Множественное происхождение пластид и митохондрий. Наука 169:641–646.
Перекрёстная ссылка пабмед КАС Google ученый
Remillard, S.P., E.Y. Лай, Ю.Ю. Леви, К. Фултон. 1990. Дифференциальная экспрессия гена кальциневрина В во время дифференцировки Naegleria . J. Cell Biol. 111:355а.
Google ученый
Рикин А. и С.Д. Шварцбах. 1988. Чрезвычайно большие и медленно процессируемые предшественники Euglena , собирающие свет, связывающие хлорофилл a/b белков фотосистемы II. Проц. Натл. акад. науч. США 85:5117–5121.
Перекрёстная ссылка пабмед КАС Google ученый
Рис, Х. 1961. Ультраструктура и молекулярная организация генетических систем. Кан. Ж. Жене. Цитол. 3:95–120.
ПабМед КАС Google ученый
Рис, Х. и В. Плант. 1962. Ультраструктура ДНК-содержащих участков хлоропласта Chlamydomonas. Дж. Клеточная биология. 13:383–391.
Перекрёстная ссылка пабмед КАС Google ученый
Шенк, Х.Э.А., К. Поралла, Т. Хартнер, Р. Даймель и Д. Тиль. 1985. Липиды в Cyanophora paradoxa I. Необычные структуры жирных кислот Cyanocyta korschikoffiana. Междунар. Дж. Эндоцит. Сотовый рез. 2: 233–238.
Google ученый
Schnepf, E. 1964. Zur Feinstruktur von Geosiphon pyriforme: Ein Versuch zur Deutung Cytoplasmatischer Membranen und Kompartimente. Архив. для микробиологии 49:112–131.
Перекрёстная ссылка Google ученый
Schnepf, E. 1969. Leucoplasten bei Nitschia alba. Эстерр. Бот. 116: 65–69.
Перекрёстная ссылка Google ученый
Шнепф, Э. и Г. Дайхгрэбер. 1984. Мизоцитоз, разновидность эндоцитоза с последствиями для разделения при эндосимбиозе. Наблюдения в Паульсенелла (Dinophyta). Натурвисс. 71: 218–219.
Перекрёстная ссылка Google ученый
Шнепф, Э. и М. Эльбрахтер. 1988. Cryptophycean-подобный хлоропласт, связанный с двойной мембраной, у динофлагеллят, Dinophysis Ehrenb.: эволюционные, филогенетические и токсикологические последствия. Бот. 90 марта 290 101: 196–203.
Google ученый
Шивджи М.С. 1991. Организация генома хлоропластов красной водоросли Porphyra yezoensis. Курс. Жене. 19:49–54.
Перекрёстная ссылка КАС Google ученый
Симейстер Г., К. Буххольц и В. Хахтель. 1990. Гены пластидного фактора удлинения Tu и рибосомного белка S7, а также шесть генов тРНК на ДНК размером 73 т.п.н. из Astasia longa , которая напоминает хлоропластную ДНК Euglena. Мол. Генерал Жене. 220:425–432.
Перекрёстная ссылка пабмед КАС Google ученый
Станьер, Р.Ю. 1970. Некоторые аспекты биологии клеток и их возможное эволюционное значение. п. 1–38. В: Организация и контроль в прокариотических клетках. соц. Генерал микробиол. Симп. 20, л.с. Чарльз и B.C.J.G. Рыцарь (ред.). Издательство Кембриджского университета.
Google ученый
Стюарт, К.Д. и К.Р. Маттокс. 1975. Сравнительная цитология, эволюция и классификация зеленых водорослей с некоторыми соображениями о происхождении других организмов с хлорофиллами а и b. Бот. Откр. 41:104–135.
Перекрёстная ссылка Google ученый
Стивенсон, Дж.К., Р.Г. Драгер, Д.В. Коппертино, Д.А. Кристофер, К.П. Дженкинс, Г. Йепиз-Пласенсия и Р. Б. Халлик. 1991. Межцистронные интроны III группы в полицистронных рибосомных белковых оперонах хлоропластов. Мол. Генерал Жене. 222:183–192.
Google ученый
Тейлор, Ф.Дж.Р. 1978. Проблемы разработки явной гипотетической филогении низших эукариот. BioSysters 10:67–89.
Перекрёстная ссылка КАС Google ученый
Тессье, Л.-Х., М. Келлер, Р.Л. Чан, Р. Фурнье, Дж.Х. Вейль и П. Имбо. 1991. Короткие лидерные последовательности могут быть перенесены с малых РНК на незрелые мРНК путем транс-сплайсинга в Эвглена. EMBO J. 10: 2621–2625.
ПабМед КАС Google ученый
Томсен, Х. А. и К. Оутс. 1978. Balaniger balticus gen. и др. ноябрь (Prymnesiophyceae) из прибрежных вод Дании. J. Mar. BioL Assoc. Великобритания 58: 773–779.
Перекрёстная ссылка Google ученый
Томас Р.Н. и Э. Р. Кокс. 1973. Наблюдения за симбиозом .Peridinium balticum и его внутриклеточная водоросль. I. Ультраструктура. J. Phycol. 9: 304–323.
Google ученый
Тример, Р.Э. и М. А. Фармер. 1991. Ультраструктурное сравнение митотического аппарата, пищевого аппарата, жгутикового аппарата и цитоскелета у эвгленоидов и кинетопластид. Протоплазма 164:91–104.
Перекрёстная ссылка Google ученый
Тернер, С., Т. Бургер-Вирсма, С.Дж. Джованнони, К.Р. Мур и Н.Р. Шаг. 1989. Отношение прохлорофита Prochlorothrix hollandica к зеленым хлоропластам. Природа 337:380–382.
Перекрёстная ссылка пабмед КАС Google ученый
Урбах Э., Д.Л. Робертсон и С.В. Чисхолм. 1992. Множественное эволюционное происхождение прохлорофитов в цианобактериальной радиации. Природа 355:267–270.
Перекрёстная ссылка пабмед КАС Google ученый
ван Вален, Л. М. 1986. Проводоросли и значение адаптивных фаций. Эволюция. Теория 8:29.
Google ученый
Уотли, Дж.М., П. Джон и Ф.Р. Уотли. 1979. От внеклеточного к внутриклеточному: создание митохондрий и хлоропластов. проц. Рой. соц. Лонд. Б. 204: 165–187.
Перекрёстная ссылка КАС Google ученый
Уилкокс, Л.В. 1989. Многослойные структуры у двух динофлагеллят, Katodinium campylops и Woloszynskia paschal. Дж. Фикол. 25:785–789.