Петли карточные, рояльные, секретерные
- Интернет магазин СуперСам
- ПЕТЛИ
- Петли карточные, рояльные, секретерные
По названию (А — Я)По названию (Я — А)По возрастанию ценыПо убыванию ценыПо коду товара (А — Я)По коду товара (Я — А)
15255075100
GTV Петля 270 клип-он для ДСП 18мм, ZP-KT0270-BE
Артикул: ZP-KT0270-BE
248.40р.
248. 40р.
40р.
SETE Петля барная, скрытого монтажа, сталь, H-BC
Артикул: H-BC
180.00р.
180.00р.
Ламберная петля
32.00р.
32.
Петля для стола 180 град. никель W-2014
98.00р.
98.00р.
Петля для стола 180 град. хром SHX-0129
120.00р.
120. 00р.
00р.
Петля карточная 140*34 б/п
16.50р.
16.50р.
Петля карточная 22*24 цинк
13.00р.
13. 00р.
00р.
Петля карточная 30*40 цинк
23.00р.
23.00р.
Петля карточная 50*40 цинк
25.00р.
25. 00р.
00р.
Петля рояльная L500 цинк
70.00р.
70.00р.
Петля секретерная d35 х h22, никель (к-кт- 2шт)
Артикул: M500
315.00р.
315. 00р.
00р.
Рояльная петля (цинк) L=1000мм
140.00р.
140.00р.
Рояльная петля (цинк) L=1750мм
336.00р.
336. 00р.
00р.
Рояльная петля (цинк) L=700мм
160.00р.
160.00р.
Рояльная петля латунь L=1750мм
170.00р.
170.
Петля секретерная — P7915 | кухонные аксессуары и фурнитура
- Lb12921 Мебельная секретерная петля для дверей, открывающихся вниз с помощью лифта.
- Размер: диаметр чашки 35 мм, глубина 12 мм.
- Материал: металл.
- Покрытие: гальваническое никель.
- Петля врезная для фасадов толщиной от 16 до 22 мм.
- Кликните на картинку — Вы увидите чертеж мебельной фурнитуры.
Производитель: LogAtask
P7915 Петля секретерная Отзывы
Ваш отзыв может быть первым.
Написать отзыв
По Москве:
Срок доставки:
2-3 рабочих дня.
Доставка до подъезда:
Свыше 3300 р. — БЕСПЛАТНО.
Менее 3300 р. — 330 р.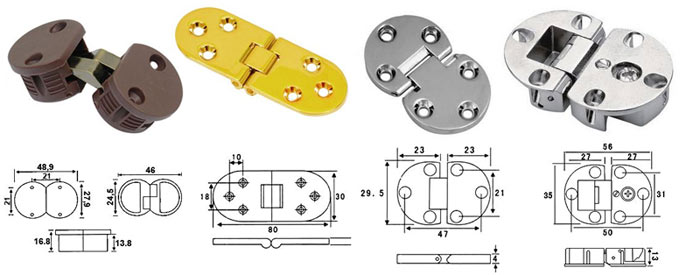
За МКАД — 20 р./км (в одну сторону).
Часы доставки оговариваются индивидуально.
Если вес товара превышает 30 кг,
то стоимость доставки может повыситься.
В случае отказа, клиент должен оплатить транспортные расходы.
По России::
Предоплата 100%. Отправка транспортной компанией (ТК).
Свыше 3300 р. — доставка до ТК БЕСПЛАТНО. Оплата до Вашего города ТК по получению.
Менее 3300 р. — доставка до ТК 330 р. Оплата до Вашего города ТК по получению.
Оплату за товар можно произвести переводом через банк или при помощи электронных денег.
Срок и стоимость доставки зависит от удаленности региона и объема груза.
Схема доставки по регионам России:
Вы оформляете заказ на сайте интернет-магазина. Доставка продукции транспортной компанией.
Стоимость доставки оплачиваете транспортной компании при получении груза.
Посылкой товар отправляем в редких случаях, если позволяет объем, например, почта не принимает посылку, когда размер одной и сторон превышает 80 сантиметров.
Отправка через день, после оплаты.
Примечание.
А города московской области, находящиеся чуть дальше:
Королёв, Одинцово, Щёлково, Пушкино, Краснознаменск, Ивантеевка, Зеленоград,
Дубна, Дмитров, Дедовск, Лобня будет небольшая доплата, при такой незначительной цене на аксессуары для кухни и мебельную фурнитуру.
По регионам доставлены заказы в города: Альметьевск, Астрахань, Брянск, Братск,
Волгоград, Владимир, Грозный, Дальний Восток, Ейск, Калининград.
Ленинградскую область, Липецк, Минск, Мурманск, Новгород Великий и Новгород Нижний,
Омск, Петрозаводск, Сахалин и многие другие.
Важно:
Если комплектующие мебельной системы вы покупаете в разных магазинах.
Где совмещение деталей вызывает у нас сомнение.
В этом случае мы не выполняем такие заказы или просим предоплату.
Кроме того, любая компания имеет право отказать в выполнении, по различным причинам.
Это могут быть какие то не выполнимые условия по наличию товара, по времени и расстоянию доставки,
ОТКАЗ ОТ ОПЛАТЫ ИЛИ ОБЫЧНОЕ ХАМСТВО.
МЫ ОСТАВЛЯЕМ ЗА СОБОЙ ПРАВО ОТКАЗЫВАТЬ В ОБСЛУЖИВАНИИ БЕЗ ОБЪЯСНЕНИЯ ПРИЧИН!
Также мебельные ручки, петли и направляющие — предоплата. Если вы не уверены и ехать на склад вам далеко, то заказывайте одну ручку, если устраивает довезем остальное на следующий день.
Вы можете оплатить заказ любым удобным Вам способом: карта, наличные, перечислением. На некоторые виды мебельной фурнитуры возможна предоплата.
Небольшие дисульфидные петли в пептидных гормонах опосредуют самоагрегацию и сортировку секреторных гранул
. 2022 27 января; 5 (5): e202101279.
doi: 10.26508/lsa.202101279.
Печать 2022 май.
Дженнифер Рек 1 , Николь Бере 1 , Эрхан Демирджи 1 , Кристина Прешанотто-Башонг 1 , Мартин Списс 2
Принадлежности
- 1 Биоцентр, Базельский университет, Базель, Швейцария.
- 2 Биоцентр, Базельский университет, Базель, Швейцария [email protected].
- PMID: 35086936
- PMCID: PMC8807871
- DOI:
10.
 26508/лса.202101279
26508/лса.202101279
Бесплатная статья ЧВК
Дженнифер Рек и др. Альянс наук о жизни. .
Бесплатная статья ЧВК
. 2022 27 января; 5 (5): e202101279.
doi: 10.26508/lsa.202101279. Печать 2022 май.
Авторы
Дженнифер Рек 1 , Николь Бере 1 , Эрхан Демирджи 1 , Кристина Прешанотто-Башонг 1 , Мартин Списс 2
Принадлежности
- 1 Биоцентр, Базельский университет, Базель, Швейцария.

- 2 Биоцентр, Базельский университет, Базель, Швейцария [email protected].
- PMID: 35086936
- PMCID: PMC8807871
- DOI: 10.26508/лса.202101279
Абстрактный
В отличие от конститутивно секретируемых белков, пептидные гормоны хранятся в плотно упакованных секреторных гранулах до регулируемого высвобождения при стимуляции. Секреторные гранулы образуются в TGN путем самоагрегации прогормонов в виде функциональных амилоидов. Было показано, что нонапептидный гормон вазопрессин, образующий небольшую дисульфидную петлю, отвечает за образование гранул своего предшественника в TGN, а также за токсическую фибриллярную агрегацию развернутых мутантов в ER. Некоторые другие предшественники гормонов также содержат подобные небольшие дисульфидные петли, что указывает на их функцию в качестве общего устройства, обеспечивающего агрегацию для сортировки гранул. Чтобы проверить эту гипотезу, мы изучили способность малых дисульфидных петель различных предшественников гормонов опосредовать агрегацию в ER и TGN. Они действительно индуцировали агрегацию ER в клетках Neuro-2a и COS-1. Слитые с конститутивно секретируемым репортерным белком, они также способствуют сортировке в секреторные гранулы, усиливают стимулированную секрецию и повышают нерастворимость луброла в клетках AtT20. Эти результаты подтверждают гипотезу, что маленькие дисульфидные петли действуют как новые сигналы для сортировки в секреторные гранулы путем самоагрегации.
Некоторые другие предшественники гормонов также содержат подобные небольшие дисульфидные петли, что указывает на их функцию в качестве общего устройства, обеспечивающего агрегацию для сортировки гранул. Чтобы проверить эту гипотезу, мы изучили способность малых дисульфидных петель различных предшественников гормонов опосредовать агрегацию в ER и TGN. Они действительно индуцировали агрегацию ER в клетках Neuro-2a и COS-1. Слитые с конститутивно секретируемым репортерным белком, они также способствуют сортировке в секреторные гранулы, усиливают стимулированную секрецию и повышают нерастворимость луброла в клетках AtT20. Эти результаты подтверждают гипотезу, что маленькие дисульфидные петли действуют как новые сигналы для сортировки в секреторные гранулы путем самоагрегации.
© 2022 Рек и др.
Заявление о конфликте интересов
 gov/pub-one»> Авторы заявляют об отсутствии конфликта интересов.
gov/pub-one»> Авторы заявляют об отсутствии конфликта интересов.
Цифры
Рисунок 1.. Отчет о циклах CC для ER…
Рисунок 1. Репортер цикла CC для агрегации ER.
Схематическое изображение пре-провазопрессина, который состоит из…
Рисунок 1. Репортер цикла CC для агрегации ER. Схематическое изображение пре-провазопрессина, состоящего из сигнальной последовательности, CCv (нонапептид вазопрессин с дисульфидной связью красного цвета), нейрофизина II (NPII) и гликопептида (gp; сайт гликозилирования обозначен ромбом) и усеченные конструкции NP∆. Для перечисленных конструкций последовательность CCv была заменена последовательностью Pro1, последовательностями СС-петли амилина (ССа), кальцитонина (ССс), ренина (ССr) или N-концевой СС-петли пролактина (CCpN). С-концевая СС-петля пролактина (CCpC) была вставлена на С-конце между последовательностью NPII и гексагистидином (His 9).0119 6 ) тег. Шкала показывает количество аминокислот в провазопрессине.
С-концевая СС-петля пролактина (CCpC) была вставлена на С-конце между последовательностью NPII и гексагистидином (His 9).0119 6 ) тег. Шкала показывает количество аминокислот в провазопрессине.
Рисунок 2. Агрегация ER конструкций CC-NP∆…
Рисунок 2. Агрегация ER конструкций CC-NP∆ в клетках COS-1 и Neuro-2a.
(А, В) COS-1…
Рисунок 2. Агрегация ER конструкций CC-NP∆ в клетках COS-1 и Neuro-2a. (A, B) Клетки COS-1 (A) и Neuro-2a (B) временно трансфицировали конструкциями NP∆, перечисленными на рис. 1, окрашивали антителами против His 6 и визуализировали с помощью иммунофлуоресцентной микроскопии. . На вставках показаны увеличения указанных областей. Шкала баров, 10 мкм. (C, D) Количественно определяли количество клеток с агрегатами (т. е. демонстрирующих >3 явных накоплений) от 100 до 150 экспрессирующих клеток для каждой конструкции в каждом из трех независимых экспериментов для Pro1 и всех конструкций CC-NP∆, и отдельно для Pro1 и NP∆, окрашенных экспериментально. Статистическую значимость рассчитывали с использованием непарных т тест. n = 3, нс, не значимо; * P ≤ 0,05; ** Р ≤ 0,01; *** Р ≤ 0,001; **** Р ≤ 0,0001.
(C, D) Количественно определяли количество клеток с агрегатами (т. е. демонстрирующих >3 явных накоплений) от 100 до 150 экспрессирующих клеток для каждой конструкции в каждом из трех независимых экспериментов для Pro1 и всех конструкций CC-NP∆, и отдельно для Pro1 и NP∆, окрашенных экспериментально. Статистическую значимость рассчитывали с использованием непарных т тест. n = 3, нс, не значимо; * P ≤ 0,05; ** Р ≤ 0,01; *** Р ≤ 0,001; **** Р ≤ 0,0001.
Рисунок S1.. Конструкции CC-NP∆ совместно локализуются с…
Рисунок S1.. Конструкции CC-NP∆ колокализуются с маркером ER PDI.
(A, B) Иммунофлуоресцентные микрофотографии…
Рисунок S1.. Конструкции CC-NP∆ колокализуются с маркером ER PDI.
(A, B) Иммунофлуоресцентные микрофотографии клеток COS-1 (A) или Neuro-2a (B), котрансфицированных конструкциями NP∆, перечисленными на рис. 1, и PDI с меченым myc. Клетки окрашивали анти-His 6 и анти-myc антителами для конфокальной микроскопии. Шкала баров, 10 мкм.
Рисунок 3. Ультраструктура агрегатов петель CC.
Рисунок 3. Ультраструктура агрегатов петель CC.
Клетки Neuro-2a были ложно трансфицированы или трансфицированы…
Рисунок 3. Ультраструктура агрегатов петель CC. Клетки Neuro-2a имитировали или трансфицировали указанными конструкциями слияния NP∆, окрашивали иммунозолотом с использованием антител против His 6 и вторичных антител, конъюгированных с золотом размером 10 нм, и визуализировали с помощью электронной микроскопии. Показаны репрезентативные меченые золотом агрегаты конструкций CC-NP∆. Единственными плотными структурами, обнаруженными в ложно- и Pro1-NP∆-трансфицированных клетках, были секреторные гранулы размером около 70 нм, лишенные иммунозолота (белые стрелки). На вставках показаны двукратные увеличения соответствующих областей в рамках. Масштабная линейка, 500 нм.
Показаны репрезентативные меченые золотом агрегаты конструкций CC-NP∆. Единственными плотными структурами, обнаруженными в ложно- и Pro1-NP∆-трансфицированных клетках, были секреторные гранулы размером около 70 нм, лишенные иммунозолота (белые стрелки). На вставках показаны двукратные увеличения соответствующих областей в рамках. Масштабная линейка, 500 нм.
Рисунок S2.. Репортерные конструкции для анализа гранул…
Рисунок S2. Репортерные конструкции для анализа сортировки гранул в стабильных клеточных линиях AtT20.
(А) Схема…
Рисунок S2. Репортерные конструкции для анализа сортировки гранул в стабильных клеточных линиях AtT20. (A) Схематическое изображение родительского репортерного белка A1Pimyc, состоящего из сигнальной последовательности, зрелой последовательности A1Pi, His 6 и метки myc.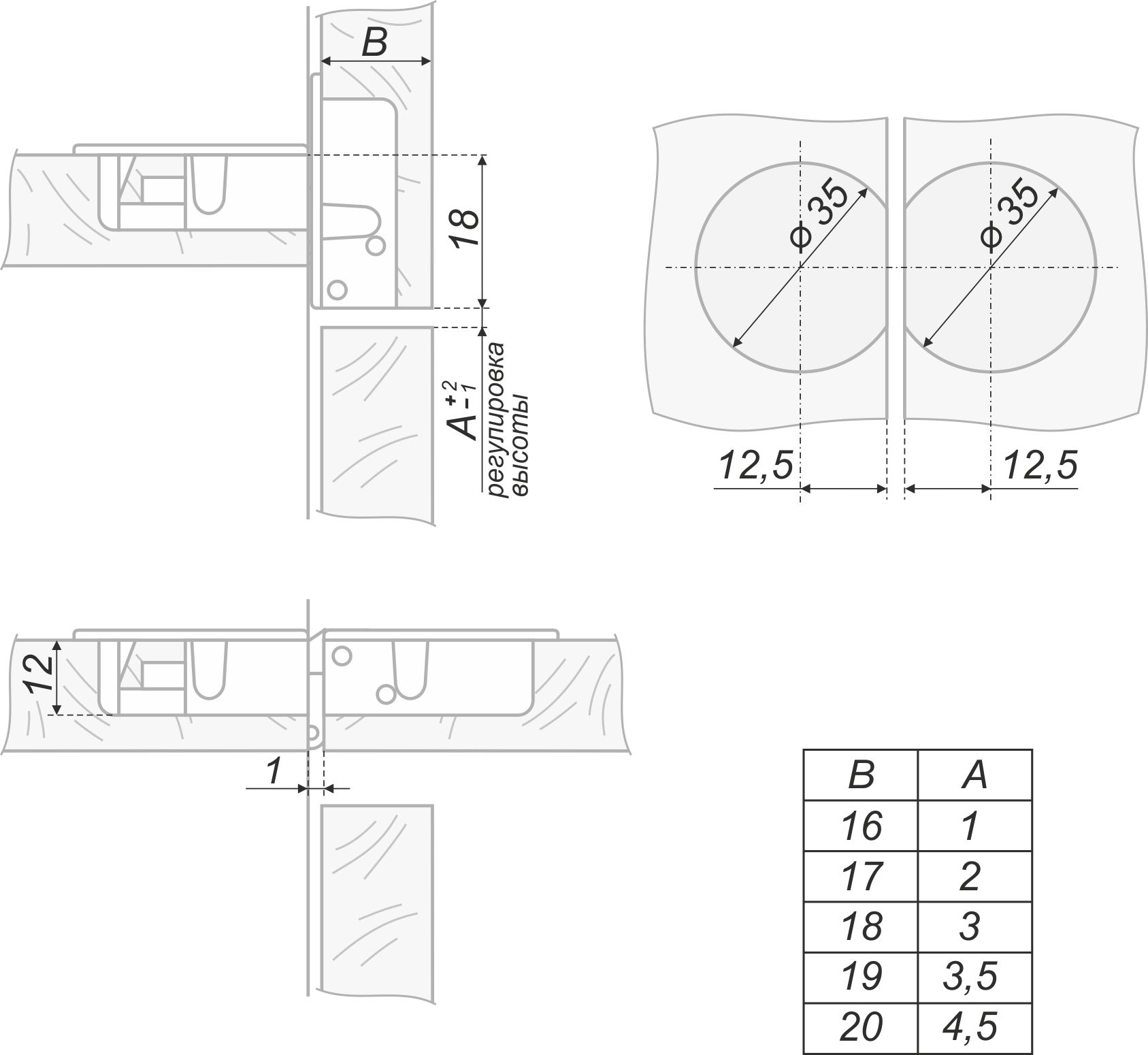 Последовательности петли CC были вставлены между сигналом и A1Pi для получения конструкций CC-A1Pimyc. Для конструкций 2xCC-A1Pimyc вторую копию той же последовательности петли СС вставляли на С-конце A1Pi. (B) Были созданы клеточных линий AtT20, стабильно экспрессирующих A1Pimyc, CC-A1Pimyc или 2xCC-A1Pimyc. Для идентификации клональных линий со сходными уровнями экспрессии их метили [ 35 S]метионин в течение 30 мин. Конструкции подвергали иммунопреципитации и анализировали с помощью электрофореза в SDS-геле и авторадиографии. Показаны наиболее подходящие линии, использованные в этом исследовании, проанализированные вместе с контрольной клеточной линией, экспрессирующей A1Pimyc (указанные номера клонов). Две полосы, соответствующие высокоманнозным и сложным гликозилированным формам производных A1Pi, обозначены закрашенными стрелками. Уровни экспрессии, нормализованные к неспецифическому белку ∼130 кДа (незаштрихованная стрелка) в качестве контроля загрузки и к A1Pimyc, указаны ниже каждой дорожки.
Последовательности петли CC были вставлены между сигналом и A1Pi для получения конструкций CC-A1Pimyc. Для конструкций 2xCC-A1Pimyc вторую копию той же последовательности петли СС вставляли на С-конце A1Pi. (B) Были созданы клеточных линий AtT20, стабильно экспрессирующих A1Pimyc, CC-A1Pimyc или 2xCC-A1Pimyc. Для идентификации клональных линий со сходными уровнями экспрессии их метили [ 35 S]метионин в течение 30 мин. Конструкции подвергали иммунопреципитации и анализировали с помощью электрофореза в SDS-геле и авторадиографии. Показаны наиболее подходящие линии, использованные в этом исследовании, проанализированные вместе с контрольной клеточной линией, экспрессирующей A1Pimyc (указанные номера клонов). Две полосы, соответствующие высокоманнозным и сложным гликозилированным формам производных A1Pi, обозначены закрашенными стрелками. Уровни экспрессии, нормализованные к неспецифическому белку ∼130 кДа (незаштрихованная стрелка) в качестве контроля загрузки и к A1Pimyc, указаны ниже каждой дорожки. Каждая панель соответствует одному гелю с одинаковой экспозицией, при этом черные линии обозначают удаленные дорожки. (B, C) Уровни экспрессии PPc-A1Pimyc определяли по сравнению с уровнями A1Pimyc путем метаболического мечения, как на панели (B).
Каждая панель соответствует одному гелю с одинаковой экспозицией, при этом черные линии обозначают удаленные дорожки. (B, C) Уровни экспрессии PPc-A1Pimyc определяли по сравнению с уровнями A1Pimyc путем метаболического мечения, как на панели (B).
Рисунок 4. Цистеины петель СС…
Рисунок 4. Цистеины петель СС окисляются в секретируемых репортерных слитых белках.
Петля CC…
Рисунок 4. Цистеины петель СС окисляются в секретируемых репортерных слитых белках. белков слияния петель CC с A1Pimyc иммунопреципитировали из среды продуцирующих клеточных линий AtT20, восстановленных с помощью TCEP (+красный) или без (-), перед алкилированием свободных тиолов цистеина хлорацетамидом. Белки расщепляли трипсином (CCv-, CCa- и CCc-NP∆) или Lys-C (CCr- и CCpN-NP∆) и анализировали с помощью масс-спектрометрии ожидаемые массы полученных пептидов СС-петли с цистеины либо присутствуют в виде дисульфидов, либо в алкилированной форме. Сигналы (площади пиков основных ионных частиц) нормировали по интенсивности пептида LQHLENELTHDIITK внутри A1Pi и представляли в условных единицах (усл. ед.). Пептидные последовательности, показанные слева от каждого графика: остатки петли CC выделены зеленым цветом с красными цистеинами, либо с красной дисульфидной связью, либо с двумя голубыми треугольниками, указывающими на алкилирование, N-концевая последовательность A1Pi выделена синим цветом, а линкерная последовательность выделена черным цветом. . Закрашенные кружки и квадраты обозначают сигналы без и с редукцией соответственно. Алкилированные пептиды обнаруживались только после восстановления. Дисульфидные формы пептидов выявлялись без восстановления и восстанавливались при обработке восстанавливающим агентом (часто малоэффективным). Пептиды для CCpN-NP∆ (↓LPICPGGAARCQVTTGEDPQGDAAQK↓, связанные дисульфидной связью или алкилированные) не были обнаружены. Для каждой конструкции показаны результаты двух или трех независимых экспериментов.
Сигналы (площади пиков основных ионных частиц) нормировали по интенсивности пептида LQHLENELTHDIITK внутри A1Pi и представляли в условных единицах (усл. ед.). Пептидные последовательности, показанные слева от каждого графика: остатки петли CC выделены зеленым цветом с красными цистеинами, либо с красной дисульфидной связью, либо с двумя голубыми треугольниками, указывающими на алкилирование, N-концевая последовательность A1Pi выделена синим цветом, а линкерная последовательность выделена черным цветом. . Закрашенные кружки и квадраты обозначают сигналы без и с редукцией соответственно. Алкилированные пептиды обнаруживались только после восстановления. Дисульфидные формы пептидов выявлялись без восстановления и восстанавливались при обработке восстанавливающим агентом (часто малоэффективным). Пептиды для CCpN-NP∆ (↓LPICPGGAARCQVTTGEDPQGDAAQK↓, связанные дисульфидной связью или алкилированные) не были обнаружены. Для каждой конструкции показаны результаты двух или трех независимых экспериментов.
Рисунок 5. Циклы CC способствуют сортировке A1Pi…
Рисунок 5. Петли CC способствуют сортировке A1Pi в секреторные гранулы.
(A, B) Линии клеток, экспрессирующие…
Рисунок 5. Петли CC способствуют сортировке A1Pi в секреторные гранулы. (A, B) Клеточные линии, экспрессирующие A1Pimyc с EBFP или без него, или слитые белки A1Pimyc с одной (A) или двумя последовательностями петли CC (B) окрашивали на myc и эндогенный CgA. Стрелки указывают на секреторные гранулы в кончиках клеток в контрольных клеточных линиях. Шкала баров, 10 мкм. (C) Для количественного анализа клеточные линии, экспрессирующие A1Pimyc или указанные петлевые конструкции CC, выращивали в смеси с клетками A1Pimyc+EBFP, окрашивали и визуализировали, как в (A) и (B). Гранулы на кончиках, определяемые окрашиванием CgA, количественно оценивали по сигналу myc и нормализовали по сигналу A1Pimyc в клетках A1Pimyc+EBFP на том же покровном стекле. 20 изображений с двумя-восьми клетками (всего ~100 клеток) каждой из них анализировали на покровное стекло в трех независимых экспериментах. Статистическую значимость определяли с использованием непарных т тест. нс, несущественный; * P ≤ 0,05; ** Р ≤ 0,01; *** Р ≤ 0,001.
Гранулы на кончиках, определяемые окрашиванием CgA, количественно оценивали по сигналу myc и нормализовали по сигналу A1Pimyc в клетках A1Pimyc+EBFP на том же покровном стекле. 20 изображений с двумя-восьми клетками (всего ~100 клеток) каждой из них анализировали на покровное стекло в трех независимых экспериментах. Статистическую значимость определяли с использованием непарных т тест. нс, несущественный; * P ≤ 0,05; ** Р ≤ 0,01; *** Р ≤ 0,001.
Рисунок 6. Петли CC опосредуют стимулированную секрецию…
Рисунок 6. Петли CC опосредуют стимулированную секрецию через функциональные секреторные гранулы.
(A) Родительские клетки AtT20…
Рисунок 6. Петли CC опосредуют стимулированную секрецию через функциональные секреторные гранулы.
(A) Родительские клетки AtT20 выращивали в двух отдельных лунках, одну из которых инкубировали в течение 1 ч с секреционной средой, не содержащей двухвалентных катионов (-), а другую со стимулирующей средой, содержащей BaCl 2 (+), обе содержащие нанотела VHH-mCherry (NB) в качестве контроля загрузки. Собранные среды подвергали количественному иммуноблот-анализу в ближней инфракрасной области на предмет эндогенно экспрессированного CgA. Полосы зеленого цвета (закрашенные стрелки) представляют полноразмерные формы CgA и протеолитически процессированные формы, продуцируемые в секреторных гранулах, красная полоса — нанотело (незаштрихованные стрелки). Цифры под дорожками представляют количество клеток (в 10 6 на лунку) для нормализации. (B) Стимулированную секрецию определяли путем количественного определения интенсивностей всех форм CgA, нормализованных к сигналу нанотела и количеству клеток, и выражали как отношение стимулированных к нестимулированным значениям. Показаны средние значения и стандартное отклонение трех независимых экспериментов, а также указаны индивидуальные значения. (C) Эксперименты со стимулированной секрецией, как на панели (A), проводили с клеточными линиями, экспрессирующими указанные конструкции A1Pimyc, в течение 30 мин. Полосы зеленого и красного цвета представляют конструкции A1Pimyc и нанотело соответственно. (D) Стимулированную секрецию количественно определяли, как в (B). Показаны среднее значение и стандартное отклонение трех независимых экспериментов, при этом отдельные значения окрашены в соответствии с экспериментом. Статистическую значимость рассчитывали с использованием непарного теста t по сравнению с контролем A1Pimyc. нс, несущественный; * P ≤ 0,05; ** Р ≤ 0,01.
Показаны средние значения и стандартное отклонение трех независимых экспериментов, а также указаны индивидуальные значения. (C) Эксперименты со стимулированной секрецией, как на панели (A), проводили с клеточными линиями, экспрессирующими указанные конструкции A1Pimyc, в течение 30 мин. Полосы зеленого и красного цвета представляют конструкции A1Pimyc и нанотело соответственно. (D) Стимулированную секрецию количественно определяли, как в (B). Показаны среднее значение и стандартное отклонение трех независимых экспериментов, при этом отдельные значения окрашены в соответствии с экспериментом. Статистическую значимость рассчитывали с использованием непарного теста t по сравнению с контролем A1Pimyc. нс, несущественный; * P ≤ 0,05; ** Р ≤ 0,01.
Рисунок 7. Нерастворимость ПОМС и CC-A1Pimyc…
Рисунок 7. Нерастворимость конструкций POMC и CC-A1Pimyc в Lubrol.
Нерастворимость конструкций POMC и CC-A1Pimyc в Lubrol.
(A) Клетки AtT20 инкубировали…
Рисунок 7. Нерастворимость конструкций POMC и CC-A1Pimyc в Lubrol. (A) Клетки AtT20 инкубировали в течение 1 ч с 1,5% раствором луброла при 4°C. Постядерный супернатант центрифугировали при 50000 g в течение 1 часа. Равные фракции супернатанта (S) и осадка (P) анализировали иммуноблоттингом на наличие эндогенных РОМС. В то время как полноразмерные формы ПОМС в основном обнаруживались в супернатанте, большая часть обработанных NPP осаждается. (B) Количественно определяли процент нерастворимых в луброле NPP. Показаны среднее значение и стандартное отклонение четырех независимых экспериментов. (C) Экстракцию Lubrol проводили, как в (A), с использованием антител против myc. Чтобы лучше отделить гликозилированный ER с высоким содержанием маннозы (hm) от сложных гликозилированных форм Гольджи/пост-Гольджи (c) конструкций A1Pimyc на геле, образцы инкубировали с эндогликозидазой H для дегликозилирования форм с высоким содержанием маннозы. (D) Количественно определяли процент нерастворимых в луброле комплексных гликозилированных форм белков. Среднее значение и стандартное отклонение соответствуют четырем независимым экспериментам, за исключением 2xCCv с тремя экспериментами, где отдельные значения окрашены экспериментом. Статистическую значимость рассчитывали с использованием непарных т тест. нс, несущественный; * P ≤ 0,05; ** Р ≤ 0,01.
(D) Количественно определяли процент нерастворимых в луброле комплексных гликозилированных форм белков. Среднее значение и стандартное отклонение соответствуют четырем независимым экспериментам, за исключением 2xCCv с тремя экспериментами, где отдельные значения окрашены экспериментом. Статистическую значимость рассчитывали с использованием непарных т тест. нс, несущественный; * P ≤ 0,05; ** Р ≤ 0,01.
Рисунок 8. Цистеины необходимы для…
Рисунок 8. Цистеины необходимы для сортировки CCc-A1Pimyc в секреторные гранулы.
(А)…
Рисунок 8. Цистеины необходимы для сортировки CCc-A1Pimyc в секреторные гранулы. (A) Цистеины CCc мутировали в пролины в PPc-A1Pimyc. (B) Клетки PPc-A1Pimyc с такими же уровнями экспрессии, как у A1Pimyc в контрольных клетках (показаны на рис. S2C), были окрашены и визуализированы, как на рис. 5. Масштабная линейка, 10 мкм. (C) Для количественного анализа клетки, экспрессирующие PPc-A1Pimyc, выращивали вместе с клетками A1Pimyc+EBFP на одном покровном стекле, окрашивали и визуализировали, а сигнал myc в гранулах на кончиках клеток определяли количественно и нормализовали к сигналу A1Pimyc в A1Pimyc Клетки +EBFP на том же покровном стекле. 20 изображений с двумя-восьми клетками (всего ~100 клеток) каждой из них анализировали на покровное стекло в трех независимых экспериментах. Для сравнения результаты для A1Pimyc и CCc-A1Pimyc на рис. 5C показаны серым цветом. Статистическую значимость определяли с использованием непарных т тест. * P ≤ 0,05.
(B) Клетки PPc-A1Pimyc с такими же уровнями экспрессии, как у A1Pimyc в контрольных клетках (показаны на рис. S2C), были окрашены и визуализированы, как на рис. 5. Масштабная линейка, 10 мкм. (C) Для количественного анализа клетки, экспрессирующие PPc-A1Pimyc, выращивали вместе с клетками A1Pimyc+EBFP на одном покровном стекле, окрашивали и визуализировали, а сигнал myc в гранулах на кончиках клеток определяли количественно и нормализовали к сигналу A1Pimyc в A1Pimyc Клетки +EBFP на том же покровном стекле. 20 изображений с двумя-восьми клетками (всего ~100 клеток) каждой из них анализировали на покровное стекло в трех независимых экспериментах. Для сравнения результаты для A1Pimyc и CCc-A1Pimyc на рис. 5C показаны серым цветом. Статистическую значимость определяли с использованием непарных т тест. * P ≤ 0,05.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Амилоидоподобная агрегация провазопрессина при несахарном диабете и сортировка секреторных гранул.

Бере Н., Хаслер Ф., Прешьянотто-Башонг С., Бирк Дж., Рутисхаузер Дж., Списс М. Берет Н. и соавт. БМС Биол. 2017 26 января; 15 (1): 5. дои: 10.1186/с12915-017-0347-9. БМС Биол. 2017. PMID: 28122547 Бесплатная статья ЧВК.
Механизм сортировки секреторных белков и образования секреторных гранул в нейроэндокринных клетках.
Чанат Э. Чанат Э. C R Seances Soc Biol Fil. 1993;187(6):697-725. C R Seances Soc Biol Fil. 1993. PMID: 7834494 Французский.
Связанная дисульфидом петля хромогранина В обеспечивает связывание с мембраной и направляет сортировку от транс-сети Гольджи к секреторным гранулам.
Гломбик М.М., Кремер А., Салм Т.
 , Хаттнер В.Б., Гердес Х.Х.
Гломбик М.М. и соавт.
EMBO J. 15 февраля 1999 г .; 18 (4): 1059–70. doi: 10.1093/emboj/18.4.1059.
ЭМБО Дж. 1999.
PMID: 10022847
Бесплатная статья ЧВК.
, Хаттнер В.Б., Гердес Х.Х.
Гломбик М.М. и соавт.
EMBO J. 15 февраля 1999 г .; 18 (4): 1059–70. doi: 10.1093/emboj/18.4.1059.
ЭМБО Дж. 1999.
PMID: 10022847
Бесплатная статья ЧВК.Концентрация гормонов в секреторных гранулах: уровни контроля.
Дэнни PS. Дэннис П.С. Мол Селл Эндокринол. 2001 г. 25 мая; 177 (1-2): 87-93. doi: 10.1016/s0303-7207(01)00437-3. Мол Селл Эндокринол. 2001. PMID: 11377824 Обзор.
Механизм сортировки пептидных гормонов и механизм биогенеза секреторных гранул секретогранином III, белком, связывающим холестерин, в эндокринных клетках.
Такеучи Т., Хосака М. Такеучи Т. и др. Curr Diabetes Rev.
 2008 Feb;4(1):31-8. дои: 10.2174/157339908783502406.
Curr Diabetes Rev. 2008.
PMID: 18220693
Обзор.
2008 Feb;4(1):31-8. дои: 10.2174/157339908783502406.
Curr Diabetes Rev. 2008.
PMID: 18220693
Обзор.
Посмотреть все похожие статьи
Рекомендации
- Арне Э., Глаттер Т., Вигано С., Шуберт К.В., Нигг Э.А., Шмидт А. (2016)Оценка и повышение точности количественного определения в экспериментах по количественному определению белка на основе изобарических масс-меток. J Proteome Res 15: 2537–2547. 10.1021/acs.jproteome.6b00066 — DOI — пабмед
- Ануп А.
 , Ранганатан С., Дас Б.Д., Джха Н.Н., Пратихар С., Гош С., Сахай С., Кумар С., Дас С., Комбрабайл М. и др. (2014) Выяснение роли дисульфидной связи в образовании амилоида и обратимости фибрилл соматостатина-14: отношение к его хранению и секреции. J Biol Chem 289: 16884–16903. 10.1074/jbcM114548354
—
DOI
—
ЧВК
—
пабмед
, Ранганатан С., Дас Б.Д., Джха Н.Н., Пратихар С., Гош С., Сахай С., Кумар С., Дас С., Комбрабайл М. и др. (2014) Выяснение роли дисульфидной связи в образовании амилоида и обратимости фибрилл соматостатина-14: отношение к его хранению и секреции. J Biol Chem 289: 16884–16903. 10.1074/jbcM114548354
—
DOI
—
ЧВК
—
пабмед
- Ануп А.
- Арван П., Касл Д. (1998)Сортировка и хранение во время биогенеза секреторных гранул: взгляд назад и взгляд вперед.
 Биохим J 332: 593–610. 10.1042/bj3320593
—
DOI
—
ЧВК
—
пабмед
Биохим J 332: 593–610. 10.1042/bj3320593
—
DOI
—
ЧВК
—
пабмед
- Арван П., Касл Д. (1998)Сортировка и хранение во время биогенеза секреторных гранул: взгляд назад и взгляд вперед.
- Beuret N, Hasler F, Prescianotto-Baschong C, Birk J, Rutishauser J, Spiess M (2017)Амилоидоподобная агрегация провазопрессина при несахарном диабете и сортировка секреторных гранул. БМС Биол 15: 5. 10.1186/s12915-017-0347-9 — DOI — ЧВК — пабмед
- Beuret N, Stettler H, Renold A, Rutishauser J, Spiess M (2004) Экспрессия регулируемых секреторных белков достаточна для образования гранулоподобных структур в конститутивно секретирующих клетках.
 J Biol Chem 279: 20242–20249. 10.1074/jbcM310613200
—
DOI
—
пабмед
J Biol Chem 279: 20242–20249. 10.1074/jbcM310613200
—
DOI
—
пабмед
- Beuret N, Stettler H, Renold A, Rutishauser J, Spiess M (2004) Экспрессия регулируемых секреторных белков достаточна для образования гранулоподобных структур в конститутивно секретирующих клетках.
Типы публикаций
термины MeSH
- 9 0031
- Скачать PDF
- Полный текст
Поделиться
Твиттер Фейсбук Электронная почта LinkedIn
- Процитировать это
- Разрешения
вещества
слепых кишечных петель у собак: влияние на секрецию гейденгайновского кармана | ДЖАМА Хирургия
Слепые кишечные петли у собак: влияние на секрецию гейденгайновского кармана | ДЖАМА Хирургия | Сеть ДЖАМА [Перейти к навигации]Эта проблема
Статья
Декабрь 1963 г.
ДЖОН Х. ЛАНДОР, MD ; В. КЕРМИТ БЕЙКЕР, MD ; ДЕННИС Ф. ХАЙТ, BA
Принадлежности автора
КОЛУМБИЯ, Миссури
Отделение хирургии Медицинской школы Университета Миссури.
Арка Сур. 1963;87(6):1048-1052. doi:10.1001/archsurg.1963.01310180164027
Полный текст
Абстрактный
Недавние обзоры 1,2 привлекли внимание к синдрому слепой петли как к причине дефицита витамина B 12 (цианокобаламина) и, реже, стеатореи. Этот синдром может возникать всякий раз, когда в тонкой кишке возникает застой либо в результате хирургического создания сегмента кишки, который не опорожняется должным образом, либо в результате патологических процессов, таких как воспалительные стриктуры или множественные дивертикулы. Клинические проявления этого состояния обычно можно контролировать, по крайней мере временно, назначением антибиотиков широкого спектра действия, таких как хлорамфеникол и антибиотики группы тетрациклинов, и считается, что чрезмерное размножение бактерий в плохо опорожняемом сегменте кишечника является причиной расстроенная физиология.