Страница не найдена — Optima
- ШКАФЫ-КУПЕ
- Коллекции
- Шкафы-купе FURNITURE COLLECTIONS WARDROBE
- Шкафы-купе FURNITURE COLLECTIONS SAND-BLAST
- Шкафы-купе FURNITURE COLLECTIONS COLOR MADNESS
- Каталог изображений пескоструйной обработки по стеклу
- Цена
- Шкафы-купе заказ оптом
- Материал
- Шкафы-купе с фотопечатью
- Стиль
- Встроенные шкафы-купе
- Коллекции
- ГОСТИНЫЕ
- Коллекции
- Garmonia
- Symfonia
- Bogemia
- Мебель для гостиной
- Стенки
- Горки
- Модульные стенки
- Стиль
- Цена
- Коллекции
- КУХНИ
- Кухни Alvic
- Коллекции кухонь
- FANTASY
- SUNNY
- CUTTER
- WOOD
- Цена
- Кухни оптом
- Эконом класса
- Кухни премиум-класса
- Форма
- Маленькие кухни
- Мини кухни
- По индивидуальным размерам
- Угловые кухни
- Встроенные кухни
- Кухня для хрущевки
- Кухни 2×2 м
- Материал
- из пластика
- из массива
- из МДФ
- с эмалью
- с фотопечатью
- глянцевые
- Акриловые
- Кухни из ЛДСП
- Кухни с патиной
- Назначение
- Модульные кухни
- Стиль
- в Классическом стиле
- в стиле Прованс
- Белые кухни
- Лофт
- Модерн
- Мебель для кухни
- Каменная столешница
- Шкафы для кухни
- Барные стойки
- Фартук для кухни
- из МДФ
- Кухни на распродаже
- КАТАЛОГИ
- Фреза
- Гостиные
- GARMONIA, SYMFONIA, BOGEMIA
- Коллекции Кухни
- Серия кухни FANTASY
- Серия кухни SUNNY
- Серия кухни WOOD
- Коллекции Шкафы
- Шкафы-купе COLOR MADNESS
- Шкафы-купе SAND-BLAST
- Шкафы-купе WAREDROBE
- Каталоги изображений
- Пескоструйная обработка по стеклу
- Фотопечать по стеклу
- Фотопечать для стеновых панелей
- КОНТАКТЫ
- ШКАФЫ-КУПЕ
- Коллекции
- Шкафы-купе FURNITURE COLLECTIONS WARDROBE
- Шкафы-купе FURNITURE COLLECTIONS SAND-BLAST
- Шкафы-купе FURNITURE COLLECTIONS COLOR MADNESS
- Каталог изображений пескоструйной обработки по стеклу
- Цена
- Шкафы-купе заказ оптом
- Материал
- Шкафы-купе с фотопечатью
- Стиль
- Встроенные шкафы-купе
- Коллекции
- ГОСТИНЫЕ
- Коллекции
- Garmonia
- Symfonia
- Bogemia
- Мебель для гостиной
- Стенки
- Горки
- Модульные стенки
- Стиль
- Цена
- Коллекции
- КУХНИ
- Кухни Alvic
- Коллекции кухонь
- FANTASY
- SUNNY
- CUTTER
- WOOD
- Цена
- Кухни оптом
- Эконом класса
- Кухни премиум-класса
- Форма
- Маленькие кухни
- Мини кухни
- По индивидуальным размерам
- Угловые кухни
- Встроенные кухни
- Кухня для хрущевки
- Кухни 2×2 м
- Материал
- из пластика
- из массива
- из МДФ
- с эмалью
- с фотопечатью
- глянцевые
- Акриловые
- Кухни из ЛДСП
- Кухни с патиной
- Назначение
- Модульные кухни
- Стиль
- в Классическом стиле
- в стиле Прованс
- Белые кухни
- Лофт
- Модерн
- Мебель для кухни
- Каменная столешница
- Шкафы для кухни
- Барные стойки
- Фартук для кухни
- из МДФ
- Кухни на распродаже
- КАТАЛОГИ
- Фреза
- Гостиные
- GARMONIA, SYMFONIA, BOGEMIA
- Коллекции Кухни
- Серия кухни FANTASY
- Серия кухни SUNNY
- Серия кухни WOOD
- Коллекции Шкафы
- Шкафы-купе COLOR MADNESS
- Шкафы-купе SAND-BLAST
- Шкафы-купе WAREDROBE
- Каталоги изображений
- Пескоструйная обработка по стеклу
- Фотопечать по стеклу
- Фотопечать для стеновых панелей
- КОНТАКТЫ
- ШКАФЫ-КУПЕ
- Коллекции
- Шкафы-купе FURNITURE COLLECTIONS WARDROBE
- Шкафы-купе FURNITURE COLLECTIONS SAND-BLAST
- Шкафы-купе FURNITURE COLLECTIONS COLOR MADNESS
- Каталог изображений пескоструйной обработки по стеклу
- Цена
- Шкафы-купе заказ оптом
- Материал
- Шкафы-купе с фотопечатью
- Стиль
- Встроенные шкафы-купе
- Коллекции
- ГОСТИНЫЕ
- Коллекции
- Garmonia
- Symfonia
- Bogemia
- Мебель для гостиной
- Стенки
- Горки
- Модульные стенки
- Стиль
- Цена
- Коллекции
- КУХНИ
- Кухни Alvic
- Коллекции кухонь
- FANTASY
- SUNNY
- CUTTER
- WOOD
Цена
- Кухни оптом
- Эконом класса
- Кухни премиум-класса
Форма
- Маленькие кухни
- Мини кухни
- По индивидуальным размерам
- Угловые кухни
- Встроенные кухни
- Кухня для хрущевки
- Кухни 2×2 м
Материал
- из пластика
- из массива
- из МДФ
- с эмалью
- с фотопечатью
- глянцевые
- Акриловые
- Кухни из ЛДСП
- Кухни с патиной
- Назначение
- Модульные кухни
- Стиль
- в Классическом стиле
- в стиле Прованс
- Белые кухни
- Лофт
- Модерн
- Мебель для кухни
- Каменная столешница
- Шкафы для кухни
- Барные стойки
- Фартук для кухни
- из МДФ
- Кухни на распродаже
- КАТАЛОГИ
- Фреза
- Гостиные
- GARMONIA, SYMFONIA, BOGEMIA
- Коллекции Кухни
- Серия кухни FANTASY
- Серия кухни SUNNY
- Серия кухни WOOD
- Коллекции Шкафы
- Шкафы-купе COLOR MADNESS
- Шкафы-купе SAND-BLAST
- Шкафы-купе WAREDROBE
- Каталоги изображений
- Пескоструйная обработка по стеклу
- Фотопечать по стеклу
- Фотопечать для стеновых панелей
- КОНТАКТЫ
Запрашиваемой страницы не существует
НА ГЛАВНУЮSubaru WRX как первая тачка.
 Повышаем уровень ВЁДЕР
Повышаем уровень ВЁДЕР03.06.2020 0:19 На первый взгляд такое рвение разработчиков 5G к предоставлению услуг элементам бытовой техники кажется довольно забавным, однако Интернет вещей (Internet of Things) — это не только умный дорожный столб и разговаривающий с домохозяйкой крутой японский утюг. Интернет вещей — это еще и так называемая “умная пыль”, Smart Dust — наноструктурированные метало-кремниевые устройства непонятного назначения и от непонятных производителей. Разработка смарт-пыли ведется уже очень давно и о серьезных успехах в этом направлении пресса писала еще 20 лет назад, томясь в радостном ожидании лучезарной перспективы. Самые простые варианты смарт-пыли — это простые сенсоры и датчики, которые самопроизвольно включаются-выключаются и передают данные о своем месте пребывания. Загрузка… Однако есть разработки и посложнее. Например, еще в 2015 году IBM сообщила о создании полноценного чипа размером 7 нм. Это в 1 000 раз меньше, чем диаметр эритроцита. Следующий их чип должен был быть размером 5 нм. Правда куда его собрались вставлять IBM так и не сообщила. Наверное, в мобильные телефоны для вирусов и микробов. Кроме миниатюризации корпорации работают и в других направлениях, например — в создании чипов, которые в качестве источника энергии используют окружающие их живые клетки. Так же создаются чипы, которые заставляют те или иные типы клеток расти, заставляют те или иные клетки умирать. И все это открыто, официально, только подается как “новое прорывное лекарство”. И это только вершина официального айсберга, поскольку создаются и микроскопические устройства, работающие в группе, то есть создающие внутри организма реципиента информационную сеть, способную выполнять те или иные задачи. Причем, поскольку все эти устройства очень малы, их внедрение не требует от реципиента каких-то осознанных действий. Все почему-то ждут, что завтра представители Нового Мирового Порядка наденут на людей кандалы и потащат в очередь ставить клейма на лоб. Но этого вовсе не нужно. Микроскопические электрические устройства можно подмешивать в воду и в пищу, можно распылять с самолёта в виде химиотрасс.
 Под микроскопом энтузиастам, конечно, кое что видно: в осадках химиотрасс находят странного вида волокна, находят какие-то элементы из фольги. Но кто смотрел на глубину 5 нанометров? У людей и микроскопов таких нет. Единственная проблема для “умной пыли” — это коммуникационная связь. Моты, то есть единичные элементы смарт-пыли еще 15 лет назад вполне сносно контактировали между собой, передавая информацию на несколько метров. Проблема была передать/принять данные на расстоянии километра и более, для чего предлагались новые протоколы мобильной связи — приблизительно такие, какими сегодня хвастаются разработчики 5G. Таким образом, только спустя много лет и теперь, с появлением стандарта 5G моты могут управляться дистанционно. Например, сгруппировавшись особым образом вызвать плохое/хорошее самочувствие у своего хозяина. Или даже внушить ему какие-то мысли. Идея совершенно не фантастическая, поскольку, как мы заметили выше, совершенно официально давно ведутся разработки наноустройств, которые при попадании в кровь начинают искать те или иные клетки.
Под микроскопом энтузиастам, конечно, кое что видно: в осадках химиотрасс находят странного вида волокна, находят какие-то элементы из фольги. Но кто смотрел на глубину 5 нанометров? У людей и микроскопов таких нет. Единственная проблема для “умной пыли” — это коммуникационная связь. Моты, то есть единичные элементы смарт-пыли еще 15 лет назад вполне сносно контактировали между собой, передавая информацию на несколько метров. Проблема была передать/принять данные на расстоянии километра и более, для чего предлагались новые протоколы мобильной связи — приблизительно такие, какими сегодня хвастаются разработчики 5G. Таким образом, только спустя много лет и теперь, с появлением стандарта 5G моты могут управляться дистанционно. Например, сгруппировавшись особым образом вызвать плохое/хорошее самочувствие у своего хозяина. Или даже внушить ему какие-то мысли. Идея совершенно не фантастическая, поскольку, как мы заметили выше, совершенно официально давно ведутся разработки наноустройств, которые при попадании в кровь начинают искать те или иные клетки. А поскольку химиотрассами дышим мы все, равно как и мы все не обрабатываем большими магнитами куски поглощаемой пищи — какие-то элементы смарт-пыли наверняка есть в теле у каждого. Пока, правда, первыми осчастливили американцев, где с 1-го октября этими сожранными за десятилетия мотами корпорации будут каким-то образом управлять.
А поскольку химиотрассами дышим мы все, равно как и мы все не обрабатываем большими магнитами куски поглощаемой пищи — какие-то элементы смарт-пыли наверняка есть в теле у каждого. Пока, правда, первыми осчастливили американцев, где с 1-го октября этими сожранными за десятилетия мотами корпорации будут каким-то образом управлять.
Blum AVENTOS HF
Складной подъемник HF – идеальное решение для высоких верхних шкафов. При компактных размерах он открывается одним движением и обеспечивает полный доступ ко всему содержимому шкафа.
Механизм оборудован встроенным доводчиком Blumotion, что дает мягкую и бесшумную работу как при открывании, так и при закрывании.
Подъемные механизмы BLUM для кухни – насколько это надежно?
Как и вся фурнитура Blum, Aventos HF обладает невероятным запасом надежности. Проверка на 80000 циклов о\з позволяет говорить о 20 и более годах безупречной работы.
Разумеется, для корректной работы необходима правильная установка.
Удобный монтаж – визитная карточка BLUM. Используйте специальные шаблоны для точной присадки силового мех-ма и рычага на корпус и фасад.
Варианты установки Aventos HF
Помимо классического варианта подъемника-«гармошки» возможен также его монтаж с электрической системой открывания – SERVO-DRIVE. Приобрести соответствующие комплектующие вы можете приобрести на этой странице.
Серводрайв рекомендуется к установке на кухни без ручек для открывания фасадов «по касанию» или push-to-open.
Дизайн HF предполагает один из 3 цветов заглушек – классический светло-серый, белый шелк или глубокий серый.
Ограничитель угла открывания Aventos HF
Для работы с модулями в непосредственной близости от потолка возможна комплектация ограничителями угла открывания на 83 и 100 градусов.
Специалисты Блюм продумали все нюансы для комфорта на Вашей кухне!
Первый в своем роде курс для выпускников, посвященный правовым вопросам в аддитивном производстве — Торговая палата Columbia Montour
Благодаря недавно введенному курсу AMD 597 «Юридические вопросы в аддитивном производстве» студенты будут готовы разбираться в юридических вопросах, возникающих в аддитивном производстве, чтобы они могли стать как техническими экспертами, так и новыми лидерами мнений в отрасли. (Изображение любезно предоставлено iStock)Из кампуса Penn State World
Примечание. Сотрудники всех членов Columbia Montour Chamber, их супруги и иждивенцы имеют право на 5% скидку на обучение в кампусе PSU World.
Когда у вас есть свобода создавать что угодно, как вы защищаете свой дизайн? В тандеме, как вы гарантируете, что не нарушаете существующую интеллектуальную собственность? В индустрии аддитивного производства (AM) эти опасения усиливаются и развиваются.
«Аддитивное производство меняет дизайн продукта и то, как мы производим детали», — сказал Тимоти В.Симпсон, Пол Морроу, профессор инженерного проектирования и производства и директор программы AMD. «Это также мешает нам защищать нашу интеллектуальную собственность. Большинство инженеров не готовы думать о том, как это повлияет на то, как их компания будет предоставлять новые продукты и услуги с помощью AM ».
Учебная программа AMD, обычно называемая 3D-печатью, призвана вдохновить студентов стать как техническими экспертами, так и новыми лидерами мнений в отрасли. По словам Симпсона, благодаря новому предложенному курсу — 597 драмов РА «Правовые вопросы аддитивного производства», первому в своем роде в любой учебной программе AM, студенты получат знания, необходимые для решения юридических вопросов, возникающих в области AM.
«Аддитивное производство создает новые этические дилеммы, с которыми приходится сталкиваться компаниям», — сказал Симпсон. «Поэтому мы должны подготовить наших студентов AMD к этим вызовам, особенно когда они берут на себя руководящие роли в компаниях, стремящихся использовать AM».
По своей сути программа AMD опирается на свои междисциплинарные связи. Он объединяет дипломные работы на нескольких факультетах, включая машиностроение; промышленное и производственное машиностроение; технические науки и механика; материаловедение и инженерия; и Школа инженерного дизайна, технологий и профессиональных программ; а также в колледжах инженерии и наук о Земле и минералах.
Благодаря новому предложению курсов, впервые преподаваемых летом 2019 года профессором делового права Дэниел Р. Кахой, программа объединяет новое междисциплинарное сотрудничество с Законом штата Пенсильвания и Бизнес-колледжем Смила. Студенты, завершившие курс:
- Знать основы создания действующих юридических контрактов и уметь заключать соглашения о секретности и лицензировать.

- Понимать, как патенты и коммерческие секреты поддерживают инновации в аддитивном производстве, и уметь интерпретировать патентные притязания и документы.
- Знайте, как включить закон об ответственности за качество продукции при разработке продуктов и процессов.
- Уметь создавать охраняемые товарные знаки и понимать, как сформировать ценный бренд.
- Узнайте, как авторские права и патенты на дизайн защищают творческий контент в аддитивном производстве.
- Уметь определять обязанности по применению кибербезопасности и защиты прав на конфиденциальность.
Традиционно инженерам не нужно было хорошо разбираться в сложных юридических аспектах контрактов, соглашений о неразглашении и интеллектуальной собственности, но Симпсон объяснил, что «последствия аддитивного производства требуют от них тщательного рассмотрения интеллектуальной собственности во всех аспектах своих бизнес-моделей. .Этот курс демистифицирует юридические аспекты и то, как аддитивное производство меняет традиционные парадигмы для защиты нашей интеллектуальной собственности ».
Бренна Маккорнак, студентка, в настоящее время обучающаяся на курсе, и инженер по аддитивному производству в Cumberland Additive, согласились.
«Как работающий инженер, я считаю, что это ценная база знаний, которую нужно иметь. Я не верю, что у многих инженеров есть возможность узнать много нового о законе, особенно в своей конкретной области », — сказала она.«Те из нас, кто участвует в этом курсе, будут иметь уникальные возможности для эффективной работы в корпоративной среде или начать свой собственный бизнес, имея хорошую основу юридических знаний, касающихся аддитивного производства».
В качестве факультатива в рамках магистерской программы этот курс поможет студентам получить ценную информацию о том, как эти проблемы напрямую влияют на промышленность. Кристофер Хиггинс, партнер и соруководитель группы 3D-печати юридической фирмы Orrick, Herrington & Sutcliffe LLP, помог Кахою разработать и провести курс.Заявите о том, что принадлежит вам по праву, без каких-либо хлопот, юристы по несчастным случаям в родах в Чикаго обязательно представят ваше дело и отнесутся к нему справедливо.
Практикующий юрист, Хиггинс борется с последствиями AM в прецедентных делах об интеллектуальной собственности более пяти лет через подразделение 3D-печати и аддитивного производства, которое он основал в своей фирме.
Он объяснил, что по мере того, как отрасль продолжает развиваться, выпускники этого курса будут готовы оказать заметное влияние.
«Правовые вопросы, особенно интеллектуальная собственность, находятся в центре внимания многих компаний-производителей аддитивов», — сказал Хиггинс. «Как инженер, понимание юридических вопросов, которые могут возникнуть в аддитивном производстве, может сделать вас бесценным активом для компании. Это набор навыков, которых у большинства инженеров нет при выходе из школы, что делает этот курс уникальной возможностью для Penn State ».
За дополнительной информацией обращайтесь по электронной почте [адрес электронной почты защищен].
GE угрожает выйти на рынок топливных элементов, конкурируя с Bloom «Breaking Energy
Заявленный прорыв в технологии твердооксидных топливных элементов
Ранее на этой неделе General Electric объявила о начале предпринимательской деятельности по коммерциализации своей технологии твердооксидных топливных элементов (SOFC) для применения в стационарных установках мегаваттной мощности. Блум Энерджи, стартап топливных элементов за миллиард долларов, также работает с технологией SOFC в этом масштабе.
Блум Энерджи, стартап топливных элементов за миллиард долларов, также работает с технологией SOFC в этом масштабе.
GE заявила о недавнем «прорыве» в топливных элементах с КПД 65 процентов (при использовании с двигателем Jenbacher) и общим КПД до 95 процентов при улавливании отработанного тепла.
Джоанна Веллингтон, руководитель передовых технологий в GE Global Research и глава подразделения топливных элементов GE, заявила в сообщении: «Проблемы с затратами, связанные с этой технологией, долгое время ставили в тупик многих людей», добавив: «Мы сделали это работает, и мы заставили его работать экономично.”
Специалист по материаламGE Кристен Броснан заявляет, что использование аддитивной технологии термического напыления «для осаждения анода и электролита» делает его «легким в нанесении, [позволяя ему] выдерживать большие перепады температуры и… служить долго». Веллингтон утверждает, что это «революционный момент».
Мартин ЛаМоника сообщил о прорыве в этом году в топливных элементах SOFC от GE год назад в IEEE.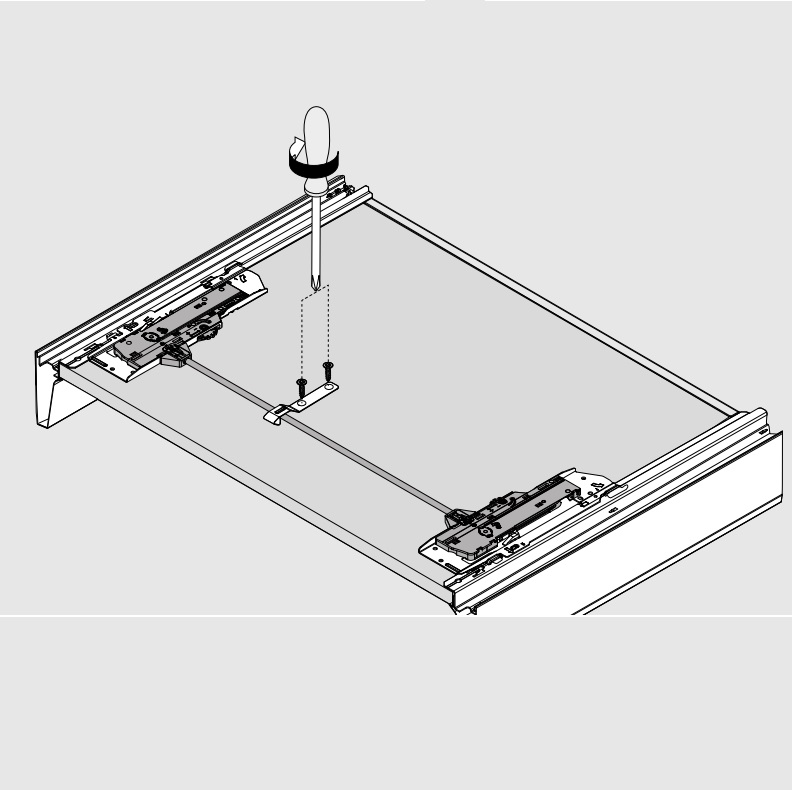 Он написал, что GE сочетает «запатентованную технологию топливных элементов со своими существующими газовыми двигателями» для замены дизельных генераторов.ЛаМоника также отметила, что технология топливных элементов GE предназначена «для работы в тандеме с двигателями GE». Он написал, что до коммерческой доступности системы еще несколько лет, и она нацелена на клиентов за пределами США, где цены на природный газ не такие низкие.
Он написал, что GE сочетает «запатентованную технологию топливных элементов со своими существующими газовыми двигателями» для замены дизельных генераторов.ЛаМоника также отметила, что технология топливных элементов GE предназначена «для работы в тандеме с двигателями GE». Он написал, что до коммерческой доступности системы еще несколько лет, и она нацелена на клиентов за пределами США, где цены на природный газ не такие низкие.
Веллингтона на посту директора GE Global Research Марк Литтл также заявил в то время о «настоящем прорыве в технологии топливных элементов».
GE работает над технологией SOFC на протяжении десятилетий. Вот слайд SOFC от GE, датируемый 2003 годом, примерно в то же время, когда Ion America сменила название на Bloom и инвестировала 1 миллиард долларов в разработку своих все еще нерентабельных топливных элементов.
GE планирует построить пилотный завод и опытно-конструкторский комплекс недалеко от Саратога-Спрингс, Нью-Йорк. По данным сайта GigaOm, в программе сейчас задействовано семнадцать человек.
По данным сайта GigaOm, в программе сейчас задействовано семнадцать человек.
Влатко Влаткович, главный технический директор подразделения GE Power Conversion, сказал в интервью Bloomberg: «Практически невозможно создать хороший топливный элемент без платины в качестве катализатора», поэтому GE пошла по пути SOFC. Но Влаткович также сказал, что до реального продукта еще «очень далеко».”
Выручка GE Conglomerate в прошлом году составила 146 миллиардов долларов. Майкл Канеллос, бывший главный редактор GTM, предположил, что это будет конгломерат, подобный Siemens или GE, который в конечном итоге выведет на рынок технологию топливных элементов, а не такой стартап, как Bloom. Другой конгломерат, UTC, который имеет обширную родословную в области топливных элементов, фактически заплатил ClearEdge Power 48 миллионов долларов за то, чтобы вывести у этой фирмы бизнес по производству топливных элементов. См. Ниже конечный результат этой транзакции.
Другие недавние новости о топливных элементах:
- Doosan из Южной Кореи, конгломерат с доходом в 21 миллиард долларов в 2013 году, только что приобрел останки обанкротившегося претендента на топливные элементы ClearEdge Power.
 По данным информационного агентства Yonhap, активы и долги проданы за 32,4 миллиона долларов, сообщило, что доход ClearEdge в 2013 году составил 68 миллионов долларов. Dow Jones сообщает, что первоначальная ставка была на уровне 48 миллионов долларов. ClearEdge привлек более 136 миллионов долларов венчурного финансирования с момента своего основания в 2006 году от Kohlberg Ventures, Applied Ventures (инвестиционное подразделение Applied Materials), Big Basin Partners и Southern California Gas Company для разработки и создания протонообменной мембраны (PEM) — топливные элементы на основе для жилых и небольших коммерческих приложений в гостиницах, многоквартирных домах и школах.В декабре 2012 года ClearEdge приобрела бизнес по производству топливных элементов технологического конгломерата United Technology Corporation (UTC) и переключила свой основной продукт PEM на технологию фосфорной кислоты UTC для своих предложений мощностью 5 и 400 кВт. Блок топливных элементов UTC мощностью 400 киловатт, работающий на природном газе, зарекомендовал себя в отрасли как один из наиболее эффективных продуктов.
По данным информационного агентства Yonhap, активы и долги проданы за 32,4 миллиона долларов, сообщило, что доход ClearEdge в 2013 году составил 68 миллионов долларов. Dow Jones сообщает, что первоначальная ставка была на уровне 48 миллионов долларов. ClearEdge привлек более 136 миллионов долларов венчурного финансирования с момента своего основания в 2006 году от Kohlberg Ventures, Applied Ventures (инвестиционное подразделение Applied Materials), Big Basin Partners и Southern California Gas Company для разработки и создания протонообменной мембраны (PEM) — топливные элементы на основе для жилых и небольших коммерческих приложений в гостиницах, многоквартирных домах и школах.В декабре 2012 года ClearEdge приобрела бизнес по производству топливных элементов технологического конгломерата United Technology Corporation (UTC) и переключила свой основной продукт PEM на технологию фосфорной кислоты UTC для своих предложений мощностью 5 и 400 кВт. Блок топливных элементов UTC мощностью 400 киловатт, работающий на природном газе, зарекомендовал себя в отрасли как один из наиболее эффективных продуктов. UTC Power была выделена из Pratt & Whitney в 1958 году и поставляла топливные элементы НАСА для космических миссий с 1966 по 2010 год.Примерно в то же время, что и апрельское банкротство, Ballard Power приобрела активы UTC, связанные с транспортными и стационарными топливными элементами, за 22 миллиона долларов.
UTC Power была выделена из Pratt & Whitney в 1958 году и поставляла топливные элементы НАСА для космических миссий с 1966 по 2010 год.Примерно в то же время, что и апрельское банкротство, Ballard Power приобрела активы UTC, связанные с транспортными и стационарными топливными элементами, за 22 миллиона долларов. - Ранее в этом месяце компания Intelligent Energy, производитель топливных элементов PEM, стала публичной и привлекла 94,1 миллиона долларов. Компания была оценена в 811 миллионов долларов, что сделало ее самой высоко оцениваемой публичной компанией по производству топливных элементов в мире. Здесь мы немного подробнее рассмотрим рынок топливных элементов.
Первоначально опубликовано на Greentech Media
Эрик Весофф, 24 июля 2014 г.
человек в пластике: неделя с 22 марта
AGILYX CORP.- Компания по переработке химикатов в Тигарде, штат Орегон, назначила Shellie Gasaway вице-президентом по развитию бизнеса.
AMCOR RIGID PLASTICS — Кармен Беккер, вице-президент и генеральный менеджер по производству специальных контейнеров производителя жесткой пластиковой упаковки в Анн-Арборе, штат Мичиган, была назначена членом совета по жидкому молоку Международной ассоциации молочных продуктов.
АМЕРИКАНСКИЙ ХИМИЧЕСКИЙ СОВЕТ — Торговая группа в Вашингтоне назначила Кимберли Уайз Уайт вице-президентом по нормативным и техническим вопросам.
ARBURG GMBH + CO. KG — Производитель термопластавтоматов в Лосбурге, Германия, назначил Паскаль Лаборде управляющим директором Arburg France и Стефаном Бюхнером управляющим директором AMKmotion GmbH + Co KG.
CHEVRON PHILLIPS CHEMICAL CO. LLC — Поставщиком смолы в Вудлендсе, штат Техас, назначен Митч Эйхелбергер, исполнительный вице-президент по полимерам и специальностям, и вице-президент по устойчивому развитию Бенджамин Мерманс.
COLORTECH — Производитель концентратов красителей и добавок в Морристауне, Теннесси. , назвал Джозефа Суанна и Джоша представителями технической службы Cisco. Рэйчел Мэтьюз и Кунтеш Десаи были назначены торговыми представителями, а Терри Брукс — старшим торговым представителем.
, назвал Джозефа Суанна и Джоша представителями технической службы Cisco. Рэйчел Мэтьюз и Кунтеш Десаи были назначены торговыми представителями, а Терри Брукс — старшим торговым представителем.
CONTINENTAL STRUCTURAL PLASTICS — Разработчик легких композитных материалов из Оберн-Хиллз, штат Мичиган, назначил Стива Руни генеральным менеджером подразделения композитов Teijin Ltd.
CURBELL PLASTICS — Дистрибьютор листов, стержней, трубок, клеев и герметиков в Орчард Парк, Н.Y. назвал региональных менеджеров по продажам Билла Раммелла и Эй Джей Лейси. Шерри Блум была назначена заместителем директора по маркетингу.
CYCLYX INTERNATIONAL LLC — Компания по переработке пластикового сырья в Портсмуте, штат Нью-Хэмпшир, назначила Рика Кокрейна старшим вице-президентом по стратегии и развитию бизнеса и главным юрисконсультом Лорен Этлингер.
DANIMER SCIENTIFIC — Разработчик и производитель биоразлагаемых материалов в Бейнбридже, штат Джорджия, назначил Брэда Роджерса вице-президентом по исследованиям и развитию технологий.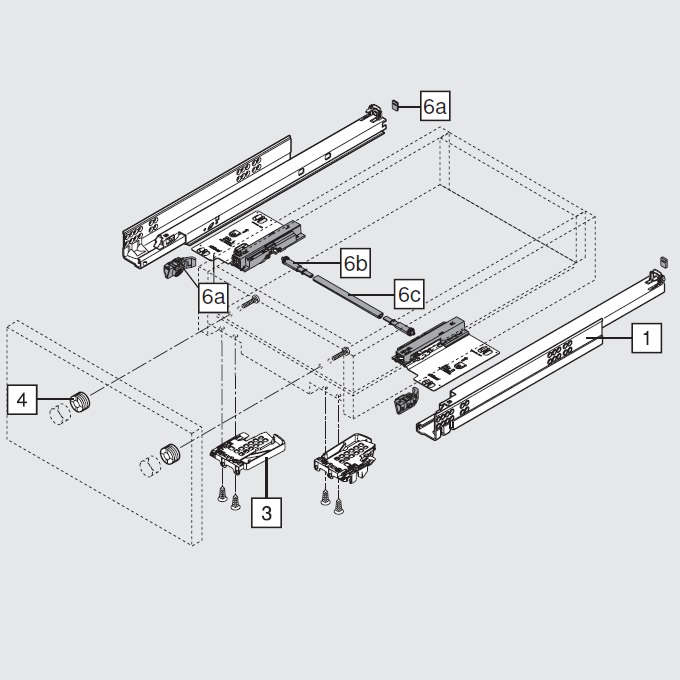
ООО «ДЭВИС-СТАНДАРТ» — Производитель экструзионных систем в Павкатаке, штат Коннектикут, назначил Эндрю Алайя вице-президентом по вторичным продажам.
DELTA TECNIC S.A. — Производитель цветных концентратов и сырья в Барселоне, Испания, назначен директором по продажам Lori Parent в США и Канаде. Родитель был включен в Plastics News ‘Women Breaking the Mold.
DIGIMARC CORP. — Производитель программного обеспечения для цифровой идентификации и обнаружения в Бивертоне, штат Орегон., назначил Тима Прайса директором по доходам и Келли Хаггерти главным директором по продуктам.
EMBALLATOR AB — Упаковочная фирма из Феникс Вэй, Англия, назвала Кристиана Олссона специалистом по полимерам для своего инновационного центра упаковочных решений в Люнгби, Швеция.
ПРОИЗВОДСТВО ЭНКОМПАССОВ — Литье под давлением в Куквилле, штат Теннеси, названо президентом Саманты Шеррелл Кросс. Крест был включен в Plastics News ‘Women Breaking the Mold.
Крест был включен в Plastics News ‘Women Breaking the Mold.
АССОЦИАЦИЯ ГИБКОЙ УПАКОВКИ — Торговая группа в Аннаполисе, Мэриленд.избрала председателем Кэти Болхоус, генерального директора Charter Next Generation, сменив Курта Бегла.
GINDUMAC GMBH — Продавец подержанной техники в Кайзерслаутерне, Германия, назвал в свой консультативный совет троих: Александр Эйслер из Weiler Werkzeugmaschinen GmbH и Kunzmann Maschinenbau GmbH; Ханс Ульрих Гольц, член исполнительного комитета KraussMaffei Technologies GmbH; и Доминик Беннер из Platform Group GmbH & Co. KG.IMG_GM.jpeg (Слева направо Эйслер, Андре, Ульрих, Беннер, Руф)
GRAHAM ENGINEERING CORP.- Производитель оборудования в Йорке, штат Пенсильвания, назначил Тодда Хадсона менеджером по продажам в юго-восточном регионе.
GRUPO TIGRE — Производитель трубопроводной арматуры в Джейнсвилле, штат Висконсин, назначил Гильерме Борнхольдт генеральным менеджером Tigre USA.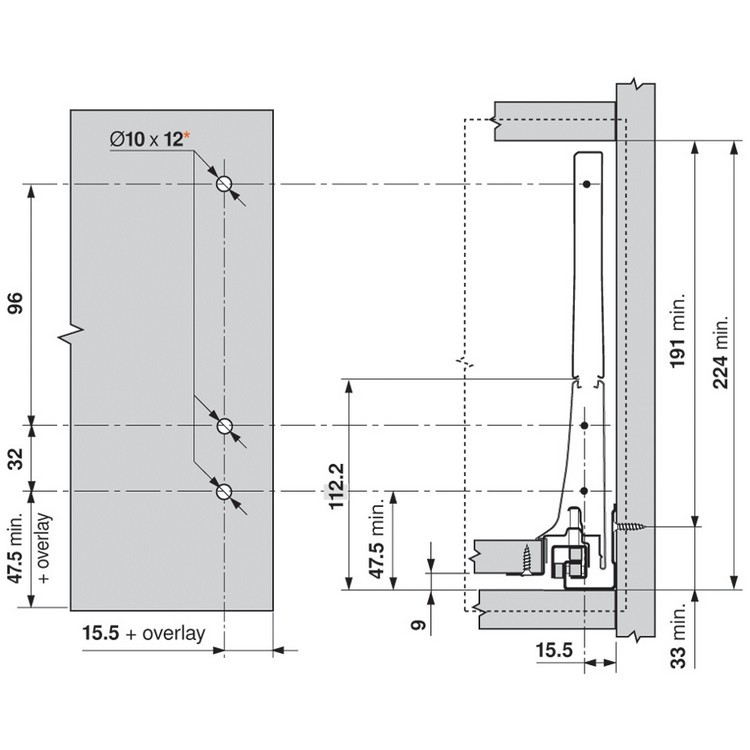
HAHN GROUP — Фирма по промышленной автоматизации и робототехнике в Райнбёллене, Германия, назначила Филиппа Унтерхальта генеральным директором и Томас Хэн генеральным директором недавно основанной RSBG Automation & Robotics Technologies GmbH.
HARWICK STANDARD DISTRIBUTION CORP.- Старший вице-президент по продажам и маркетингу Дэн Дэвис ушел на пенсию с химического поставщика в Акроне, Огайо,
.IMA SCHELLING GROUP — Поставщик оборудования для производства пластмасс в Роли, Северная Каролина, назначил Флориана Кляйншмидта региональным менеджером по продажам.
KIEFEL GMBH — Поставщик оборудования во Фрайлассинге, Германия, назван Ричардом Шмидхофером, финансовым директором.
MACK MOLDING CO. — Производитель литья под давлением в Арлингтоне, штат Вирджиния, назначил Брайана Нолана директором по персоналу своего северного подразделения.
MILLIKEN & CO.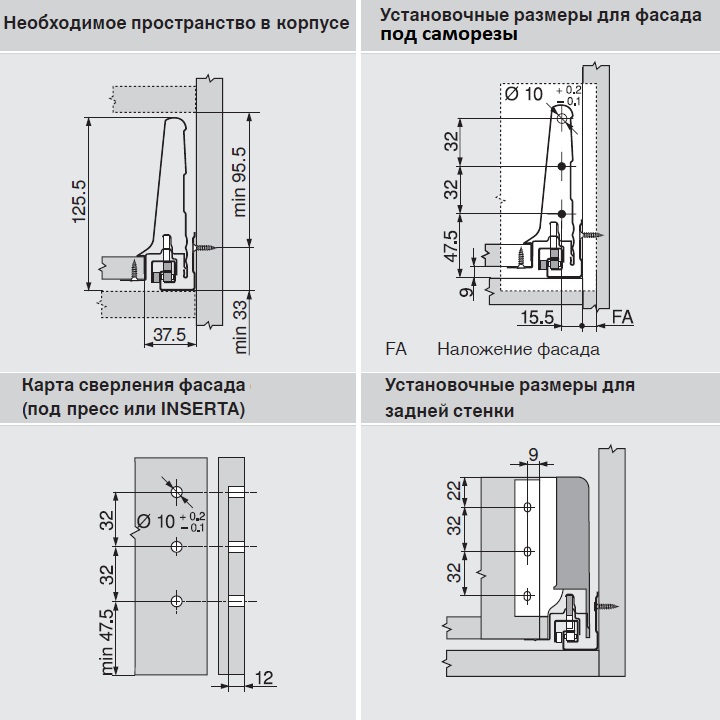 — Производитель специальных химикатов и добавок для пластмасс в Спартанбурге, Южная Каролина, назначил Синди Бойтер исполнительным вице-президентом своего химического подразделения.
— Производитель специальных химикатов и добавок для пластмасс в Спартанбурге, Южная Каролина, назначил Синди Бойтер исполнительным вице-президентом своего химического подразделения.
MYERS INDUSTRIES INC. — Иветт Дапремонт Брайт, президент Brighter Horizon Foundation, и Джеффри Крамер, генеральный директор Schweitzer-Mauduit International, были назначены в совет директоров многопрофильного производителя в Акроне, штат Огайо.
NORDSON CORP. — Производитель точного дозирующего оборудования в Вестлейке, штат Огайо, назначил Кертиса Хатчитта менеджером по развитию рынка гранулирования для компании Nordson BKG LLC, расположенной в Остине, штат Техас.
OTTO KRAHN GMBH & CO. — Специалист по изготовлению смесей в Гамбурге, Германия, назначен управляющим директором и директором по развитию Акселя Себбессе.
PRECISION CUSTOM PRODUCTS INC. — Компания по литью под давлением в ДеГрафф, штат Огайо, назначена президентом и генеральным директором Доун Билман. Билман был включен в Plastics News ‘Женщины ломают форму.
Билман был включен в Plastics News ‘Женщины ломают форму.
SACMI IMOLA S.C. — Производитель оборудования в Имоле, Италия, назначен генеральным менеджером Мауро Фенци, сменив Джулио Менголи.
SEPRO GROUP — Дистрибьютор оборудования для автоматизации в Ла-Рош-сюр-Йон, Франция, назначил Джимми Тео региональным менеджером по продажам для своего отдела продаж в Сингапуре.
SK CAPITAL PARTNERS LP — Частная инвестиционная компания в Нью-Йорке назначила Даниэля Феррари старшим директором.
ОБЩЕСТВО ИНЖЕНЕРОВ ПО ПЛАСТМАССАМ — Джон Хедрик, исполнительный вице-президент по инженерным вопросам компании Coast Systems, был назначен председателем профессионального общества в Данбери, штат Коннектикут.
ООО «СПАРТЕК» — Производитель упаковки, рулонных материалов, специальной продукции, а также визуальных и структурных листов в Мэриленд-Хайтс, Миссури, назначил Джеффа Фримена менеджером завода на своем предприятии в Макминнвилле, штат Орегон.
TANDEM GROUP INC. — Фирма производителя в Гриндейле, штат Висконсин, назвала Пэйтон Рашинг специалистом по внутренним продажам.
TEKNI-PLEX INC. — Производитель упаковки в Уэйне, штат Пенсильвания, назначил Бренду Чамулак генеральным директором и президентом с 30 июня, сменив Пола Янга, который станет председателем.
TREXEL INC. — Разработчик процесса микросотового вспенивания MuCell в Уилмингтоне, штат Массачусетс, назначил Роджера Кауфманна директором по развитию бизнеса, заменив Хартмута Траута, который вышел на пенсию.
Монументальный марафон — Wyobraska Tandem
Я являюсь членом специальной группы по планированию марафона монументов в Геринг-Скоттсблафф, Небраска.
В прошлом году, в первый год гонки, мы с Багманом проехали на наших пригородных велосипедах на одной из нескольких поездок, необходимых для сертификации трассы USA Track & Field.Я воспользовался этой возможностью, чтобы сфотографироваться и написать в блоге сообщение о курсе: «Марафон памятников и его пейзажи Западной Небраски». Я тоже сделал небольшую запись после мероприятия: «Марафон первого памятника в книгах».
Я тоже сделал небольшую запись после мероприятия: «Марафон первого памятника в книгах».
Теперь, когда мы с Багманом едем в тандеме на большие расстояния, я подумал, что нам следует проехать марафон памятников. Поскольку во время своего первого цикла тура по маршруту я выбрал наиболее интересные или иллюстративные виды, я решил, что на этот раз я буду задокументировать маршрут более объективно, делая снимки каждые полмили.
«Будет много картинок», — сказал Багман.
Ага. На расстоянии 26,2 мили два изображения на милю = 52 изображения.
Но мой первый пост по марафонской дистанции содержал 62 изображения. Какая красивая сельская местность! Это нужно задокументировать и поделиться!
Багман и я подъехали на велосипеде к Природному центру Уайлдкэт-Хиллз, немного остыли, а затем отправились на трассу. Я включил звуковые сигналы GPS и делал снимки каждые полмили, плюс-минус пройденное расстояние, пока возился с камерой.
Вот результаты наших усилий:
Миля 0: взгляд на запад / юго-запад. Парковка природного центра Wildcat Hills. Асфальтированная поверхность.
Парковка природного центра Wildcat Hills. Асфальтированная поверхность.
мили 0,5: взгляд на север / под гору. Шоссе 71. Асфальтированное покрытие.
Ой! Пропущена миля 1.0 — ветер так громко свистел на спуске, что не слышал свой GPS.
Миля 1,5: взгляд на восток. Дома в Уайлдкэт-Хиллз. Асфальтированная поверхность.
Миля 2: взгляд на восток. Асфальтированная поверхность.
- Миля 2.5: смотрит на восток / юг. Асфальтированная поверхность.
Миля 3: взгляд на запад, в долину Геринг. Асфальтированная поверхность.
- Миля 3,5: глядя на запад. Асфальтированная поверхность.
Миля 4: взгляд на запад. Асфальтированная поверхность.
- Миля 4.5: глядя на запад. Асфальтированная поверхность.
Миля 5: взгляд на запад. Асфальтированная поверхность.
Миля 5.5: взгляд на восток. Асфальтированная поверхность.
Миля 6: глядя на юг.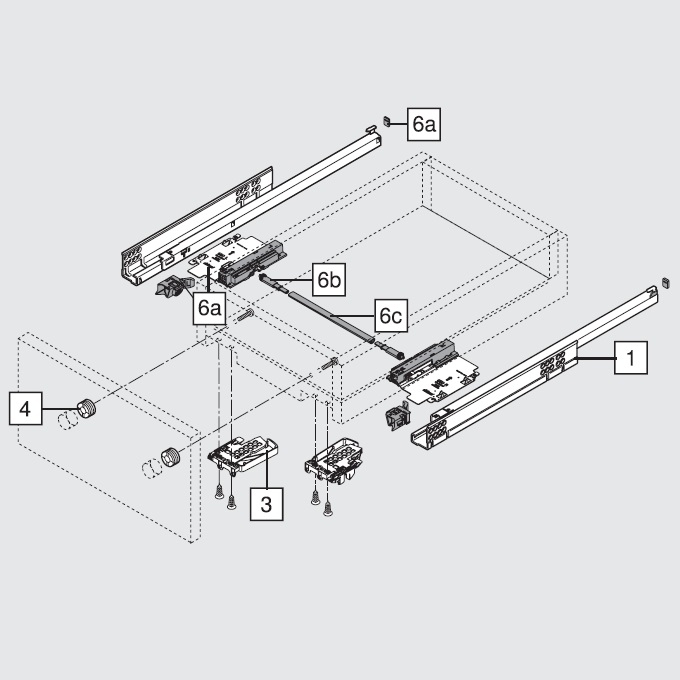 Это на Сандберг-роуд, после съезда с шоссе 71.Асфальтированная поверхность.
Это на Сандберг-роуд, после съезда с шоссе 71.Асфальтированная поверхность.
Миля 6.5: взгляд на восток. На фото сложно разглядеть, но впереди на дороге трактор. К счастью, он свернул с дороги прежде, чем мы его догнали. Асфальтированная поверхность.
Миля 7: взгляд на север. Асфальтированная поверхность.
Миля 7,5: глядя на юг. Долина Геринга (орошение) Дренаж. Асфальтированная поверхность.
Миля 8: вид на северо-запад. Поворот с Сэндберг-роуд на Локвуд-роуд. Асфальтированная поверхность.
Миля 8,5: вид на запад над кукурузным полем к национальному памятнику Скоттс-Блафф.Асфальтированная поверхность.
Миля 9: взгляд на запад. Асфальтированная поверхность.
Миля 9,5: взгляд на восток. Это могло быть то место, где на западной стороне была свалка. Редакционное решение смотреть на восток для фото. Асфальтированная поверхность.
Миля 10: вид на северо-запад, на поворот в Геринг. Асфальтированная поверхность.
Миля 10,5: глядя на север.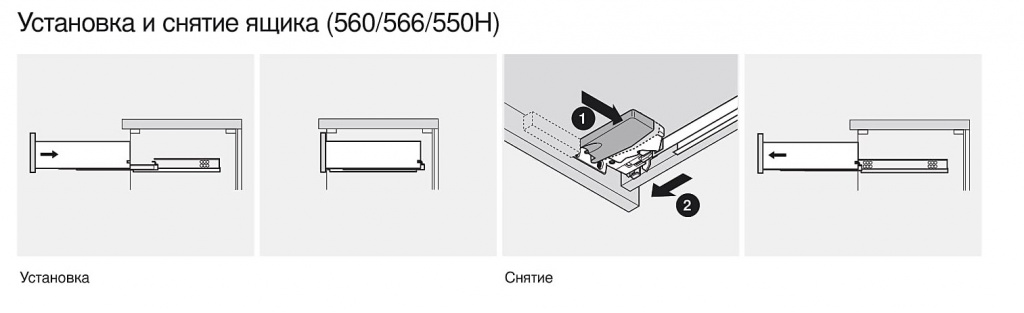 Въезд в Геринг проходит через промышленную зону. Асфальтированная поверхность.
Въезд в Геринг проходит через промышленную зону. Асфальтированная поверхность.
Миля 11: глядя на юг. Асфальтированная поверхность.
Миля 11.5: взгляд на северо-запад в сторону стадионов Орегон-Трейл-Парк. Асфальтированная поверхность.
Миля 12: вид на северо-запад на поворот жилого квартала на Файв Рокс Роуд. Асфальтированная поверхность.
Миля 12,5: глядя на восток, через плечо, на перекрестке с четырьмя остановками Five Rocks Road и M Street / Old Oregon Trail. Точка измерения находилась на перекрестке, но я был слишком занят, обращая внимание на трафик, чтобы сделать снимок. Есть несколько фотографий этой дороги конца 1800-х годов в начале развития Геринга.Раньше это была городская улица, она была частью Орегонской тропы. По этому пути прошли и наездники Пони-экспресса. Асфальтированная поверхность.
Миля 13: взгляд на юг, вниз по засаженной деревьями дороге к кладбищу Геринг. Асфальтированная поверхность.
Багман и я остановились на перерыв в музее «Наследие равнин», так что с этого момента наши отметки миль, вероятно, будут немного отклоняться из-за расстояния, пройденного на парковке музея.
Миля 13,5: вид на северо-запад, на некоторую собственность, принадлежащую Музею наследия равнин, на фоне национального памятника Скоттс-Блафф.Асфальтированная поверхность.
Миля 14: вид на северо-запад, национальный памятник Скоттс-Блафф. Асфальтированная поверхность.
Миля 14,5: глядя на север. Идеальное время, чтобы поймать первопроходческий фургон и «волов» на Орегонской тропе у национального памятника Скоттс-Блафф. Асфальтированная поверхность.
Миля 15: взгляд на север. Асфальтированная поверхность.
Миля 15,5: вид на северо-запад. Асфальтированная поверхность.
Миля 16: вид на северо-восток. Мне * нравится *, что некоторые люди здесь до сих пор разводят усачий рогатый скот. Асфальтированная поверхность.
Миля 16,5: взгляд на восток. Асфальтированная поверхность.
Миля 17: взгляд на запад. В этой группе деревьев находится очаровательный отель типа «постель и завтрак» Barn Anew. Асфальтированная поверхность.
Миля 17,5: глядя на запад. Асфальтированная поверхность.
Асфальтированная поверхность.
Миля 18: вид на юг, через поле молодых растений сахарной свеклы на Риджуэй-драйв. Перевал Митчелл находится слева от кадра (он был примерно на отметке 14,5 мили). Поверхность из гравия.
Миля 18,5: глядя на юг? юго-запад? Поверхность из гравия.
Миля 19: вид на юг от дороги оросительного канала.Маршрут от примерно 19 мили до примерно 22,5 мили проходит по грунтово-гравийной частной дороге на территории национального памятника Скоттс-Блафф, которая в основном используется для обслуживания оросительных каналов. Управлять тандемом было довольно сложно, так как поверхность варьируется от мелкого гравия до крупного гравия, утрамбованной грязи и рыхлой песчаной почвы со случайными колеями на шинах. Гравий / грязь.
Миля 19,5: вид на юг, колония луговых собачек через оросительный канал. Гравий / грязь.
Миля 20: глядя на юг.Гравий / грязь.
Миля 20,5: глядя на юг. Я думаю, что где-то в этом месте мы наткнулись на участок рыхлой почвы, и в итоге я положил руку на землю. По этому покрытию можно бегать — просто нужно быть внимательным, но местами это неприемлемо для 350-фунтового тандема с гонщиками на двух тонких дорожных шинах. Мы немного погуляли на велосипеде. Гравий / грязь.
По этому покрытию можно бегать — просто нужно быть внимательным, но местами это неприемлемо для 350-фунтового тандема с гонщиками на двух тонких дорожных шинах. Мы немного погуляли на велосипеде. Гравий / грязь.
Миля 21: взгляд на юг, на северную стену национального памятника Скоттс Блафф. Гравий / грязь.
Миля 21.5: взгляд на север в сторону бесплодных земель, когда Багман идет тандемом через еще один песчаный участок. Гравий / грязь.
Миля 22: вид на северо-восток, через пастбище, на окраину квартала передвижных домов. Гравий / грязь.
Миля 22,5: вид на юго-запад на окрестности поля для гольфа Monument Shadows. Асфальтированная поверхность.
23-я миля: взгляд с велосипедной дорожки на национальный памятник Скоттс-Блафф на запад. Асфальтированная поверхность.
Миля 23,5: вид на запад с велосипедной дорожки.Асфальтированная поверхность.
Миля 24: вид на юг с тропы на У-стрит на хранилище тюков в Геринге (перерабатывающий центр для муниципальной свалки). Асфальтированная поверхность.
Асфальтированная поверхность.
Миля 24,5: глядя на запад от Файв Рокс Роуд. Асфальтированная поверхность.
Миля 25: взгляд на юг, бульвар Медоуларк — часть зигзага по окрестностям. Асфальтированная поверхность.
Миля 25,5: вид на запад от усаженной деревьями кладбищенской дороги через бобовое поле в сторону южного обрыва национального памятника Скоттс-Блафф.Асфальтированная поверхность.
Миля 26: взгляд на запад — прямо перед жестоким поворотом ландшафта, который организаторы гонок называют «Дьявольским провалом» или «каньоном Чупакабра». Почти готово! Поверхность из гравия.
Через эти ворота и через парковку финишная черта проходит у амфитеатра Five Rocks. Поверхность из гравия.
Итак, вот и все: виды на трассу Monument Marathon с тандемного велосипеда с шагом примерно в полмили: сочетание заповедника, пастбищ, сельскохозяйственных угодий, достопримечательностей Орегонской тропы, промышленных зон и кварталов на асфальте, бетоне и грязи. / гравийные покрытия.
/ гравийные покрытия.
Это качественная сельская гонка, организованная волонтерами в пользу фонда местных общинных колледжей. Это один из четырех марафонов в Небраске: суперсельский марафон Sandhills Marathon — это небольшая гонка с ограничением регистрации, остальные — городские большие марафоны на другом конце штата в Линкольне и Омахе. Культура здесь имеет больше общего с Вайомингом, чем с остальной частью Небраски (за исключением футбольных фанатов Хаскера).
Если это похоже на ваше приключение, зарегистрируйтесь для участия в гонке на главной странице.Будем рады видеть Вас!
Чтобы узнать, что делать и что посмотреть в этом районе, посетите сайт туризма округа Скоттс-Блафф или просмотрите некоторые из заархивированных сообщений в моем другом блоге, SCB Citizen.
Вернемся на минутку к езде на велосипеде и немного поразмышляем. Чуть больше года назад я выбился из колеи, проехав 26,2 мили на велосипеде. В этом году мы проехали 47 миль, чтобы преодолеть гоночную трассу плюс расстояние до и от нашего дома, и после этого я был лишь слегка утомлен.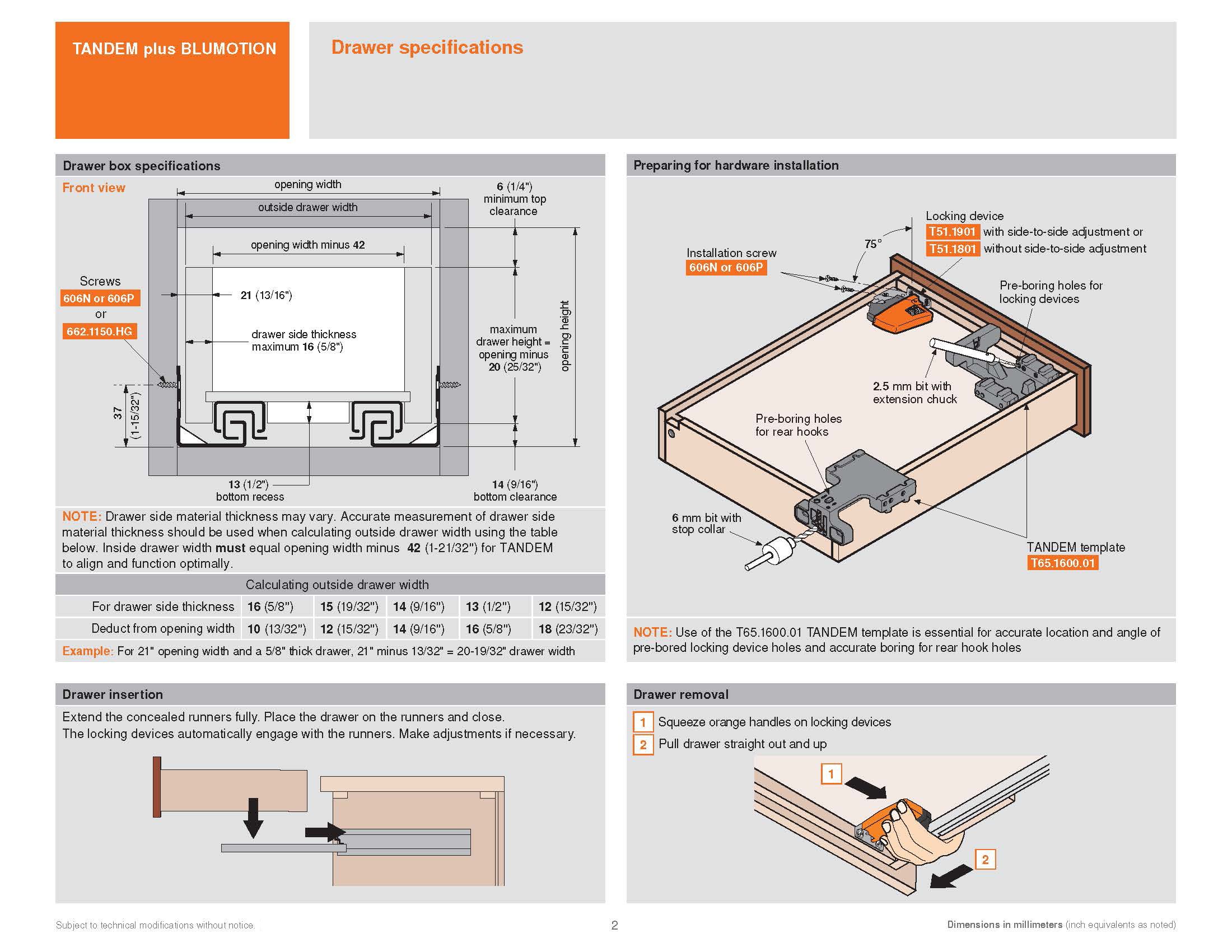 Какая разница за год!
Какая разница за год!
Авторские права 2013 Кэти Брэдшоу
Ген GIGAS гороха является гомологом T ЦВЕТОЧНОГО ЛОКУСА, необходимого для передачи трансплантата спецификации цветения, но не для реакции на фотопериод
ВВЕДЕНИЕ
У многих видов время цветения регулируется рядом факторов окружающей среды, включая продолжительность светового дня и температуры, и в последнее время много усилий было направлено на понимание молекулярных механизмов, лежащих в основе этой регуляции.Ген Arabidopsis thaliana FLOWERING LOCUS T ( FT ) занимает важное место в генетической иерархии, которая контролирует цветение и объединяет световой период, температуру, яровизацию и сигнализацию качества света. FT кодирует небольшой белок, сходный с белком фосфатидилэтаноламинсвязывающего домена млекопитающих (PEBP) (Kardailsky et al., 1999; Kobayashi et al., 1999). Arabidopsis FT экспрессируется в листьях в условиях индукции цветения, и белок FT перемещается через флоэму к верхушке побега, где он связывается с фактором транскрипции bZIP FD для активации транскрипции гена идентичности меристемы цветков APETALA1 ( AP1 ) и, возможно, других родственных генов домена MADS (Abe et al.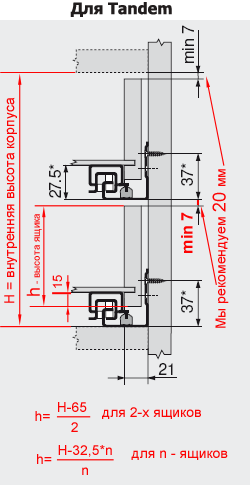 , 2005; Wigge et al., 2005; Corbesier et al., 2007; Джагер и Вигге, 2007; Mathieu et al., 2007). Было показано, что FT-подобные белки нескольких различных видов действуют аналогично FT в отношении индукции цветения, транспорта во флоэме и взаимодействия с FD-подобными белками (Lifschitz et al., 2006; Lin et al., 2007; Tamaki et al., 2007; Li and Dubcovsky, 2008), предполагая, что этот общий механизм, вероятно, широко сохраняется у цветковых растений.
, 2005; Wigge et al., 2005; Corbesier et al., 2007; Джагер и Вигге, 2007; Mathieu et al., 2007). Было показано, что FT-подобные белки нескольких различных видов действуют аналогично FT в отношении индукции цветения, транспорта во флоэме и взаимодействия с FD-подобными белками (Lifschitz et al., 2006; Lin et al., 2007; Tamaki et al., 2007; Li and Dubcovsky, 2008), предполагая, что этот общий механизм, вероятно, широко сохраняется у цветковых растений.
В Arabidopsis , FT является частью небольшого генного семейства (так называемого семейства PEBP) с пятью другими членами: TWIN SISTER OF FT ( TSF ), TERMINAL FLOWER1 ( TFL1 ). ), Arabidopsis thaliana CENTRORADIALIS, гомолог , MOTHER OF FT И TFL1 ( MFT ) и BROTHER OF FT И TFL1 ( BFT ) (Bradley et al., 1997; Mimida et al., 2001; Yoo et al., 2004, 2010; Ямагути и др., 2005). Семейства генов PEBP у других видов растений различаются по размеру: от пяти генов в томате ( Solanum lycopersicum ) до примерно 20 в рисе ( Oryza sativa ) и кукурузе ( Zea mays ) (Carmel-Goren et al. , 2003; Chardon, Damerval, 2005; Данилевская и др., 2008; Игасаки и др., 2008). Филогенетический анализ позволяет выделить три основные клады в этом семействе, соответствующие FT -подобным, TFL1 -подобным и MFT -подобным генам. FT -подобные гены способствуют цветению, тогда как TFL1 -подобные гены задерживают цветение и предотвращают превращение апикальной меристемы побега в цветочную меристему (Bradley et al., 1996; Bradley et al., 1997; Pnueli et al., 1998; Foucher et al., 2003). В Arabidopsis предполагалось, что противоположные функции FT и TFL1 происходят из-за различия одной аминокислоты в расходящейся внешней петле в четвертом экзоне (Hanzawa et al., 2005; Ahn et al., 2006) и TFL1 было предложено действовать как конкурент FT, потенциально за счет конкурентного связывания с FD (Ahn et al., 2006). Arabidopsis FT -подобная клада имеет два члена: собственно FT и близкий паралог TSF . Гены FT и TSF кодируют очень похожие белки и имеют сходный проксимальный промотор и сходные паттерны регуляции в отношении фотопериода и яровизации (Yamaguchi et al.
, 2003; Chardon, Damerval, 2005; Данилевская и др., 2008; Игасаки и др., 2008). Филогенетический анализ позволяет выделить три основные клады в этом семействе, соответствующие FT -подобным, TFL1 -подобным и MFT -подобным генам. FT -подобные гены способствуют цветению, тогда как TFL1 -подобные гены задерживают цветение и предотвращают превращение апикальной меристемы побега в цветочную меристему (Bradley et al., 1996; Bradley et al., 1997; Pnueli et al., 1998; Foucher et al., 2003). В Arabidopsis предполагалось, что противоположные функции FT и TFL1 происходят из-за различия одной аминокислоты в расходящейся внешней петле в четвертом экзоне (Hanzawa et al., 2005; Ahn et al., 2006) и TFL1 было предложено действовать как конкурент FT, потенциально за счет конкурентного связывания с FD (Ahn et al., 2006). Arabidopsis FT -подобная клада имеет два члена: собственно FT и близкий паралог TSF . Гены FT и TSF кодируют очень похожие белки и имеют сходный проксимальный промотор и сходные паттерны регуляции в отношении фотопериода и яровизации (Yamaguchi et al.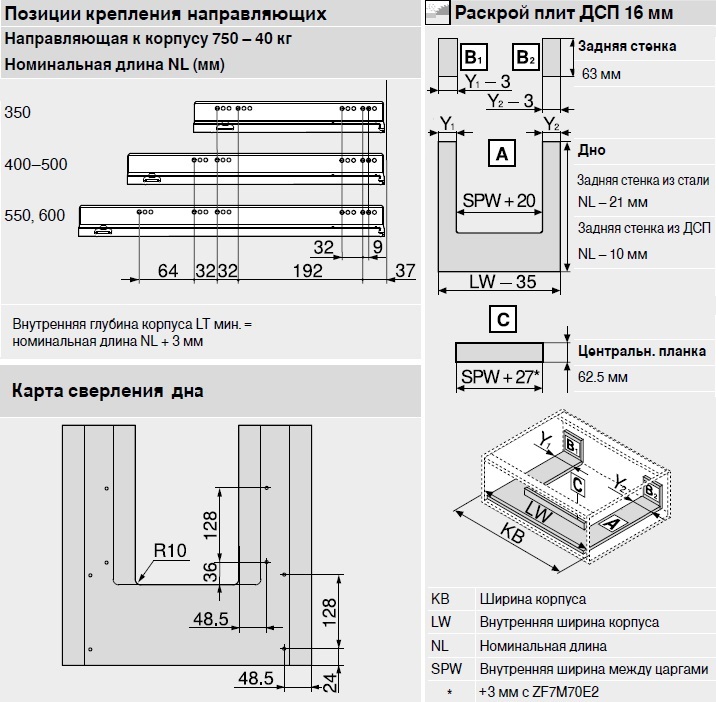 , 2005). TSF оказывает небольшое влияние на цветение добавки с FT , а белок TSF, как и FT, может действовать как мобильный индуктор цветения при экспрессии во флоэме (Jang et al., 2009). В отличие от сходства в регуляции FT и TSF , множественные FT -подобные гены у других видов демонстрируют заметные различия в регуляции (Faure et al., 2007; Kikuchi et al., 2009; Blackman et al. , 2010).
, 2005). TSF оказывает небольшое влияние на цветение добавки с FT , а белок TSF, как и FT, может действовать как мобильный индуктор цветения при экспрессии во флоэме (Jang et al., 2009). В отличие от сходства в регуляции FT и TSF , множественные FT -подобные гены у других видов демонстрируют заметные различия в регуляции (Faure et al., 2007; Kikuchi et al., 2009; Blackman et al. , 2010).
Бобовые — это большая группа растений, которая присутствует в большинстве экосистем и включает многие важные виды сельскохозяйственных культур. Большинство сельскохозяйственных культур и модельных бобовых подразделяются на две отдельные клады: так называемые галегоидные бобовые (включая горох [ Pisum sativum ], Medicago , Lotus , чечевицу [ Lens culinaris ] и нут [ Cicer arietinum ]. ]), которые в основном являются долгоживущими растениями из регионов с умеренным климатом, склонными к яровизации, и миллеттикоидными бобовыми культурами теплого сезона (включая фасоль [ Phaseolus vulgaris ], сою [ Glycine max ] и вигну [ Vigna unguiculata ]). которые происходят из более низких широт и представляют собой преимущественно растения короткого дня (Cannon et al.Мы изучаем генетический контроль цветения гороха посевного, многодневного, чувствительного к яровизации бобового растения, которое широко использовалось в ранних работах по генетическому контролю мобильных сигналов цветения (Weller et al., 1997, 2009b). . Недавно было показано, что несколько локусов гороха, контролирующих чувствительность фотопериода и сигналы мобильного цветения, являются ортологами генов Arabidopsis , участвующих в функции циркадных часов. К ним относятся LATE BLOOMER1 ( LATE1 ) и DIE NEUTRALIS ( DNE ), которые являются ортологами Arabidopsis GIGANTEA и EARLY FLOWERING4 et al., Соответственно (Hechch., 2007; Liew et al., 2009). Принимая во внимание потенциальную важность генов FT в мобильной передаче сигналов цветков, мы определили семейство генов FT у гороха, Medicago и сои и исследовали регуляцию семейства гороха FT , документируя различные паттерны экспрессии для разные участники.
которые происходят из более низких широт и представляют собой преимущественно растения короткого дня (Cannon et al.Мы изучаем генетический контроль цветения гороха посевного, многодневного, чувствительного к яровизации бобового растения, которое широко использовалось в ранних работах по генетическому контролю мобильных сигналов цветения (Weller et al., 1997, 2009b). . Недавно было показано, что несколько локусов гороха, контролирующих чувствительность фотопериода и сигналы мобильного цветения, являются ортологами генов Arabidopsis , участвующих в функции циркадных часов. К ним относятся LATE BLOOMER1 ( LATE1 ) и DIE NEUTRALIS ( DNE ), которые являются ортологами Arabidopsis GIGANTEA и EARLY FLOWERING4 et al., Соответственно (Hechch., 2007; Liew et al., 2009). Принимая во внимание потенциальную важность генов FT в мобильной передаче сигналов цветков, мы определили семейство генов FT у гороха, Medicago и сои и исследовали регуляцию семейства гороха FT , документируя различные паттерны экспрессии для разные участники. Мы также показываем, что один из этих генов соответствует ранее описанному локусу GIGAS , и исследуем роль этого гена в чувствительности к световому периоду и индукции цветения.Наши результаты предполагают, что роль семейства FT в индукции цветения потенциально более сложна у гороха, чем у Arabidopsis , включая перекрестную регуляцию транскрипции, множественные мобильные сигналы и возможную функциональную дифференциацию отдельных членов.
РЕЗУЛЬТАТЫ
Семейство генов
FT в бобовых В предыдущем обзоре генов, связанных с цветением в модельных бобовых, мы идентифицировали пять генов, подобных FT , в Medicago truncatula ( FTLa-FTLe ) и единственный ген гороха, наиболее похожий на Medicago FTLe (Hecht et al., 2005; Liew et al., 2009). Четыре дополнительных гена гороха FT были идентифицированы с помощью скрининга библиотеки и подходов ПЦР, и выделение соответствующих полноразмерных кДНК продемонстрировало, что все пять генов гороха FT экспрессируются и имеют структуру интрона / экзона, аналогичную генам FT в других виды (рис. 1А). Для двух генов нам не удалось легко выделить определенные интроны, предположительно из-за их большого размера. Чтобы получить более широкую перспективу эволюции семейства FT в составе бобовых, мы выполнили поиск BLAST генома сои в Phytozome (www.phytozome.net). Хотя геномы бобовых разделяют древнее событие дупликации всего генома, соя пережила дополнительное, более недавнее событие дупликации всего генома и в результате содержит более крупные семейства генов с легко идентифицируемыми парами гомеологов (Schmutz et al., 2010). Наши поиски выявили 10 предполагаемых полноразмерных генов FT в сое (см. Дополнительную таблицу 1 онлайн), восемь других полноразмерных генов PEPB, более похожих на Arabidopsis TFL1 , BFT или MFT (см. Таблица 1 онлайн) и несколько других очевидных псевдогенов.
1А). Для двух генов нам не удалось легко выделить определенные интроны, предположительно из-за их большого размера. Чтобы получить более широкую перспективу эволюции семейства FT в составе бобовых, мы выполнили поиск BLAST генома сои в Phytozome (www.phytozome.net). Хотя геномы бобовых разделяют древнее событие дупликации всего генома, соя пережила дополнительное, более недавнее событие дупликации всего генома и в результате содержит более крупные семейства генов с легко идентифицируемыми парами гомеологов (Schmutz et al., 2010). Наши поиски выявили 10 предполагаемых полноразмерных генов FT в сое (см. Дополнительную таблицу 1 онлайн), восемь других полноразмерных генов PEPB, более похожих на Arabidopsis TFL1 , BFT или MFT (см. Таблица 1 онлайн) и несколько других очевидных псевдогенов.
бобовых имеют три отдельные группы генов FT .
(A) Геномная организация генов гороха FT . Экзоны представлены заштрихованными прямоугольниками. Жирными пунктирными линиями обозначены интроны неизвестного размера.
Экзоны представлены заштрихованными прямоугольниками. Жирными пунктирными линиями обозначены интроны неизвестного размера.
(B) Филограмма последовательностей белка PEBP бобовых культур. Ветви со значениями начальной загрузки <60% свернуты. Анализ основан на выравнивании последовательностей, показанном на дополнительном рисунке 2 в режиме онлайн. Подробности последовательности доступны в дополнительной таблице 1 онлайн.
[Цветную версию этого рисунка см. В онлайн-статье.]
Филогенетический анализ (рисунок 1B) показывает, что гены PEBP бобовых попадают в описанные ранее клады FT , TFL1 и MFT . Рисунок 1B также показывает, что гены FT делятся на три отдельных субклада, а микросинтения вокруг генов сои и Medicago дополнительно демонстрирует сродство генов внутри каждой клады (см. Дополнительный рисунок 1 и дополнительную таблицу 2 онлайн).Эти результаты показывают, что распространение семейства FT произошло относительно рано в эволюции бобовых, и позволяют предположить, что эти три клады, вероятно, будут представлены в других культурах и модельных бобовых культурах. Чтобы обеспечить единообразие номенклатуры трех видов и рамки для наименования FT генов других бобовых, мы обозначили эти субклады как FTa , FTb и FTc и предлагаем переименовать Medicago и горох. гены соответственно (см. дополнительную таблицу 1 онлайн).
Субклад FTa представлен в Medicago и горохе двумя генами, FTa1 (ранее FTLa ) и FTa2 (ранее FTLb ), а у сои — четырьмя генами, парой гомеологов. ( FTa1 и FTa2 ) и два других связанных гена ( FTa3 и FTa4 ) без четких гомеологов. Субклад FTb также представлен двумя генами как в горохе, так и в Medicago : FTb1 (ранее FTLd в Medicago ) и FTb2 (ранее FTLe в Medicago и Medicago ). горох).В Medicago эта пара генов расположена в тандеме наверху хромосомы 7, и мы картировали ген гороха FTb1 в соответствующую область группы сцепления V. Соя также имеет четыре гена FTb в двух гомеологических блоках. на хромосомах 18 ( FTb1 и FTb2 ) и 8 ( FTb3 и FTb4 ) (см. дополнительный рисунок 1 онлайн). Субклад FTc — наименее сложный, состоящий из одного гена гороха и Medicago (ранее FTLc ) и одной пары гомеологов ( FTc1 и FTc2 ) в сое (рисунок 1).И в Medicago , и в сое эти гены расположены непосредственно рядом с генами FTa (см. Дополнительный рисунок 1 онлайн), что подтверждает их ортологическую природу и вероятное происхождение от одного и того же события дупликации.
Соя также имеет четыре гена FTb в двух гомеологических блоках. на хромосомах 18 ( FTb1 и FTb2 ) и 8 ( FTb3 и FTb4 ) (см. дополнительный рисунок 1 онлайн). Субклад FTc — наименее сложный, состоящий из одного гена гороха и Medicago (ранее FTLc ) и одной пары гомеологов ( FTc1 и FTc2 ) в сое (рисунок 1).И в Medicago , и в сое эти гены расположены непосредственно рядом с генами FTa (см. Дополнительный рисунок 1 онлайн), что подтверждает их ортологическую природу и вероятное происхождение от одного и того же события дупликации.
Предыдущие попытки определить структурные особенности, которые отличают стимулирующую цветок FT-подобную функцию от ингибирующей цветок TFL1-подобной функции, выявили две критические пары остатков: Tyr-85 / Gln-140 в FT и His-88 / Asn-144 в TFL1 (Hanzawa et al., 2005; Ahn et al., 2006). Все белки FT бобовых несут консервативный остаток Tyr, а белки FTa и FTb также содержат консервативный Gln, характерный для других белков FT (Ahn et al. , 2006). Однако белки FTc имеют общий His в этом положении и, таким образом, отличаются как от других последовательностей FT, так и от TFL1. Белки FTc также несут несколько других консервативных замен в соседней области (сегмент B в Ahn et al., 2006), которые могут иметь потенциал для изменения функции белка (см. Дополнительный рисунок 2 онлайн).
, 2006). Однако белки FTc имеют общий His в этом положении и, таким образом, отличаются как от других последовательностей FT, так и от TFL1. Белки FTc также несут несколько других консервативных замен в соседней области (сегмент B в Ahn et al., 2006), которые могут иметь потенциал для изменения функции белка (см. Дополнительный рисунок 2 онлайн).
Горох
FT Гены демонстрируют отчетливые закономерности регуляции Мы использовали эксперименты по переносу фотопериода, чтобы определить временное окно от 11 до 16 дней после посева, в течение которого растения гороха линии NGB5839 становятся необратимо приверженными к цветению в условиях длинных дней (LD). при 20 ° C (рис. 2A).Экспрессия ортолога AP1 гороха PROLIFERATING INFLORESCENCE MERISTEM ( PIM ) (Taylor et al., 2002) в апикальных зачатках (рассеченных до размера 2-3 мм) показала четкую индукцию с 21 дня в LD или 42 дня в короткие дни (SD), что соответствует появлению видимых цветочных бутонов примерно через 1 неделю. Значительная индукция ортолога гороха LFY UNIFOLIATA ( UNI ) (Hofer et al., 1997), SEPALLATA1 ( SEP1 ) (Hecht et al., 2005), и TFL1 paralog DETERMINATE ( DET / TFL1a ) (Foucher et al., 2003) также наблюдался с 21 дня в LD или 42 дня в SD (Рисунок 2B). Экспрессия ортолога гороха FD показала слабую индукцию LD на 21 день относительно уровня SD (фиг. 2B).
Значительная индукция ортолога гороха LFY UNIFOLIATA ( UNI ) (Hofer et al., 1997), SEPALLATA1 ( SEP1 ) (Hecht et al., 2005), и TFL1 paralog DETERMINATE ( DET / TFL1a ) (Foucher et al., 2003) также наблюдался с 21 дня в LD или 42 дня в SD (Рисунок 2B). Экспрессия ортолога гороха FD показала слабую индукцию LD на 21 день относительно уровня SD (фиг. 2B).
Гены гороха FT демонстрируют различные паттерны экспрессии.
(A) Перенос фотопериода определяет окно для приверженности цветению у растений дикого типа (NGB5839), выращенных в LD.Растения выращивали после посева в условиях LD и переводили в условия SD в указанные сроки. До 12 дня не было значительного эффекта от воздействия LD, а к 17-му дню не было значительного эффекта от переноса SD. Окно, в течение которого устанавливается приверженность цветку, обозначено серой штриховкой. Значения представляют собой среднее ± стандартное отклонение для n = 6
(B) и (C) Экспрессия генов идентичности соцветий (B) и FT генов (C) во время развития. Относительные уровни транскриптов определяли в рассеченных верхушках побегов или самом верхнем полностью развернутом листе во время развития в LD (16 светлых / 8 темных; белые кружки) или SD (8 светлых / 16 темных; темные кружки). Серый цвет представляет окно обязательств, определенное в (A) . Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 2 до 3 биологических повторов, каждая из которых состоит из объединенного материала от двух растений.
Относительные уровни транскриптов определяли в рассеченных верхушках побегов или самом верхнем полностью развернутом листе во время развития в LD (16 светлых / 8 темных; белые кружки) или SD (8 светлых / 16 темных; темные кружки). Серый цвет представляет окно обязательств, определенное в (A) . Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 2 до 3 биологических повторов, каждая из которых состоит из объединенного материала от двух растений.
Интересно, что гены FT обнаруживают различные паттерны регуляции в отношении фотопериода, времени и тканевой специфичности.В самом верхнем полностью увеличившемся листе проростков, выращенных методом LD, экспрессия FTb2 сильно индуцировалась к 14 дню, тогда как индукция экспрессии FTa1 и FTa2 происходила более постепенно с 14 по 35 день (рис. 2C). В соответствующей ткани проростков, выращенных на SD, FTb2 не экспрессировалось, тогда как FTa1 демонстрировало слабую индукцию к 21 дню, аналогичную LD, и FTa2 также индуцировался в той же степени, что и при LD, но с задержкой ~ 1 неделя. Транскрипт FTc не был обнаружен выше фона в ткани листа при LD или SD.
Транскрипт FTc не был обнаружен выше фона в ткани листа при LD или SD.
Мы также обнаружили значительную экспрессию генов FT в апикальных зачатках. В LD экспрессия FTa1 и FTc была явно индуцирована, а FTa2 — слабо индуцирована к 21 дню в этой ткани (фиг. 2C). В SD FTc показал самую сильную индукцию, постепенно увеличиваясь с 35-го дня до уровня LD, аналогичного дикому типу, тогда как экспрессия FTa1 и FTa2 индуцировалась более слабо с 42-го дня, как и у . PIM и другие гены идентичности соцветий (Рисунки 2B и 2C).Ни FTb1 , ни FTb2 транскрипт не экспрессировались выше фона в верхушках побегов при любом фотопериоде. Интерпретация экспрессии в этих образцах апикальных зачатков должна быть квалифицирована тем фактом, что они содержали другие ткани помимо самой апикальной меристемы побега, включая лист, черешок и сосудистые зачатки. В будущем будет интересно изучить более подробно паттерны пространственной экспрессии генов FT на верхушке побега. Тем не менее, даже при самой консервативной интерпретации, специфическая экспрессия FTc в этих образцах и отсутствие экспрессии FTb2 действительно указывают на сильную контрастирующую тканевую специфичность для экспрессии этих генов FT .
Тем не менее, даже при самой консервативной интерпретации, специфическая экспрессия FTc в этих образцах и отсутствие экспрессии FTb2 действительно указывают на сильную контрастирующую тканевую специфичность для экспрессии этих генов FT .
Гены гороха
FT способствуют цветению трансгенного Arabidopsis Затем мы проверили способность генов гороха FT дополнять мутант Arabidopsis ft-1 . На фигуре 3 показано, что репрезентативная линия 35S: Ps -FTa1 с высокой экспрессией, выращенная в LD, зацвела значительно раньше, чем нетрансформированный контроль ft-1 . FTc сверхэкспрессия привела к поразительному фенотипу раннего цветения, при котором большинство трансформантов продуцировало только две деформированные структуры стеблевого листа перед окончанием верхушки побега в одном терминальном цветке (см. Supplemental Figure 3 online).Этот фенотип сильнее, чем сообщалось для экспрессии 35S: FT у Arabidopsis , и более похож на комбинированные эффекты 35S: AP1 и 35S: FT (Kardailsky et al. , 1999). Остальные гены гороха ( FTa2 , FTb1 и FTb2 ) показали более слабую активность, при этом трансформанты зацвели раньше, чем нетрансформированный мутант ft , но не так рано, как дикий тип. Это показывает, что все пять генов гороха FT в некоторой степени обладают функцией FT , и предполагает, что разные белки FT могут отличаться по присущей им активности.
, 1999). Остальные гены гороха ( FTa2 , FTb1 и FTb2 ) показали более слабую активность, при этом трансформанты зацвели раньше, чем нетрансформированный мутант ft , но не так рано, как дикий тип. Это показывает, что все пять генов гороха FT в некоторой степени обладают функцией FT , и предполагает, что разные белки FT могут отличаться по присущей им активности.
FT проявляют разную активность у трансгенного Arabidopsis.
Комплементация мутанта Arabidopsis ft-1 генами гороха FT
(A) Типичные растения, выращенные в LD. Цветение произошло во всех линиях, но не в нетрансформированном контроле ft-1 . WT, дикий тип.
(B) Общее количество листьев в период цветения для репрезентативных линий Arabidopsis .Данные являются средними значениями ± стандартная ошибка как минимум для 10 растений.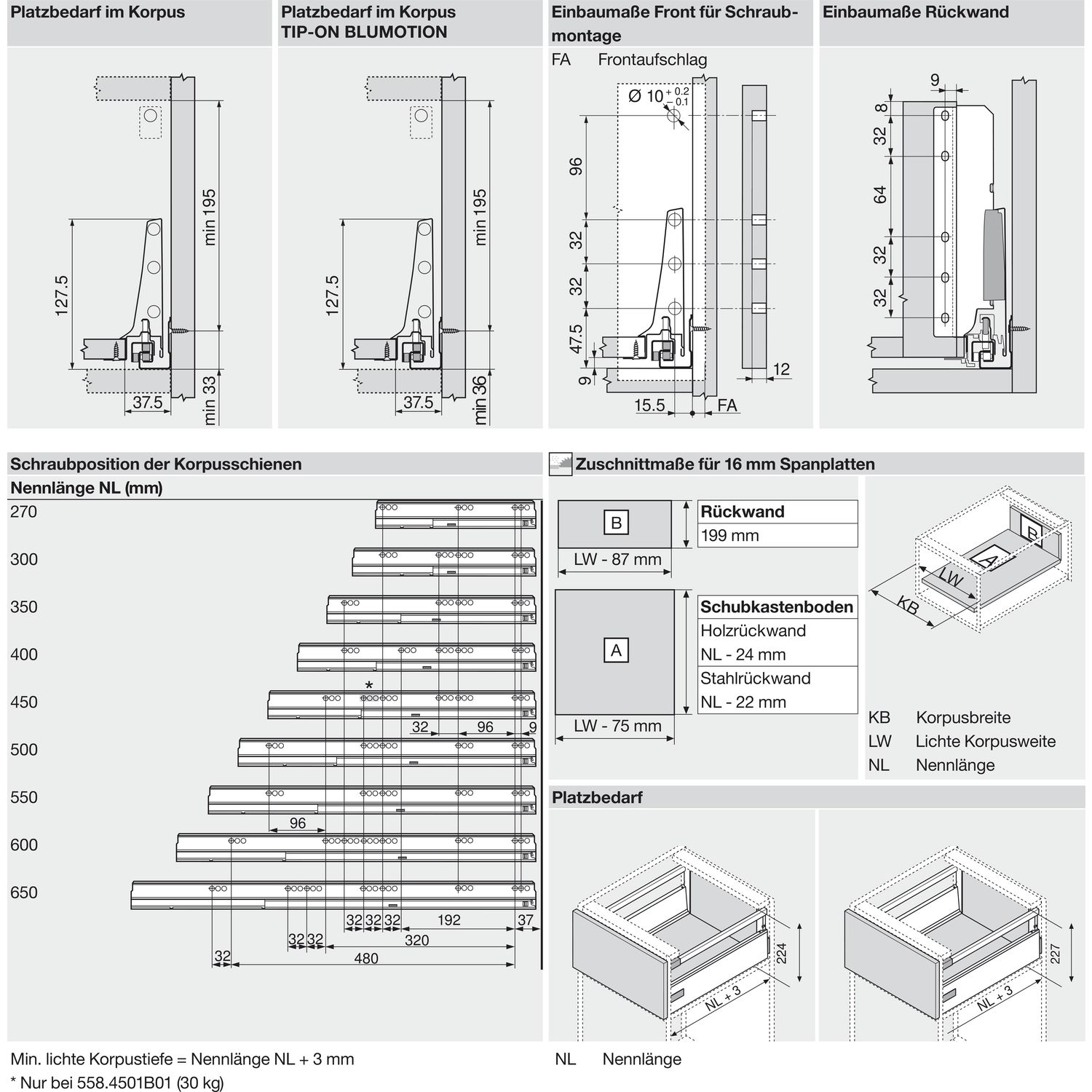
[Цветную версию этого рисунка см. В статье в Интернете.]
Аберрантная регуляция генов
FT в мутантах пути фотопериодаЗатем мы исследовали, может ли гены FT неправильно регулироваться у двух мутантов с нарушенной реакцией на фотопериод. Мутант late1 задерживает цветение и блокирует ответ LD, тогда как мутант dne способствует цветению и имеет фенотип LD при SD (Hecht et al., 2007; Liew et al., 2009).
При 16-часовом LD в шкафах для выращивания мутация late1 задержала цветение примерно на шесть узлов и индукцию PIM в рассеченных апикальных зачатках побегов на 2 недели (рис. 4A). Паттерны экспрессии для трех экспрессируемых на листьях генов FT в мутанте late1 были аналогичны тем, которые наблюдались в растениях дикого типа, выращенных на SD (рис. 2C), что согласуется с SD-подобным фенотипом выращенного LD late1. растения (Hecht et al., 2007). Индукция FTb2 в листе была полностью заблокирована late1 (фиг. 4A), что согласуется с предыдущим отчетом, показывающим более низкую экспрессию этого гена в late1 (Hecht et al., 2007). В поздних1 верхушках побегов паттерны экспрессии FTa1 , FTa2 и FTc также были аналогичны таковым в растениях дикого типа, выращенных на SD, с пониженной экспрессией FTa1 и задержкой FTc по отношению к индукции. к выращенному на LD дикому типу и без существенных различий в экспрессии FTa2 (фиг. 4A).
4A), что согласуется с предыдущим отчетом, показывающим более низкую экспрессию этого гена в late1 (Hecht et al., 2007). В поздних1 верхушках побегов паттерны экспрессии FTa1 , FTa2 и FTc также были аналогичны таковым в растениях дикого типа, выращенных на SD, с пониженной экспрессией FTa1 и задержкой FTc по отношению к индукции. к выращенному на LD дикому типу и без существенных различий в экспрессии FTa2 (фиг. 4A).
FT неправильно регулируются у мутантов с фотопериодным ответом.
(A) Экспрессия гена у дикого типа (темные кружки) и мутанта late1-2 (белые кружки) во время развития в условиях LD (16 световых / 8 темных).
(B) Экспрессия гена у дикого типа (темные кружки) и мутант dne-1 (белые кружки) во время развития в условиях SD (8 световых / 16 темных).
Относительные уровни транскриптов определяли в рассеченных верхушках побегов или в самом верхнем полностью развернутом листе. Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 2 до 3 биологических повторов, каждая из которых состоит из объединенного материала от двух растений.
Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 2 до 3 биологических повторов, каждая из которых состоит из объединенного материала от двух растений.
При 8-часовом стандартном отклонении в шкафах для выращивания мутация dne способствовала цветению примерно на восемь узлов и индукции PIM примерно на 3 недели (рис. 4B). Мутант dne также показал значительно более высокую экспрессию FTa1 , FTa2 и FTb2 в листьях и FTa1 , FTa2 и FTc в вершинах побегов.Мутант dne цветет раньше при SD, чем дикий тип при LD (Liew et al., 2009), и это также отражается в некоторых паттернах экспрессии FT . В частности, экспрессия в листьях FTa1 и FTa2 была намного сильнее в SD-выращенных dne , чем в LD-выращенных дикого типа, как и экспрессия FTa2 в апикальных зачатках. Однако экспрессия FTb2 , хотя и повышенная в dne по сравнению с диким типом при SD (фиг. 4B), не показала сильной ранней индукции (т.е.е., к 14-му дню), что типично для LD дикого типа (рис. 2С).
Однако экспрессия FTb2 , хотя и повышенная в dne по сравнению с диким типом при SD (фиг. 4B), не показала сильной ранней индукции (т.е.е., к 14-му дню), что типично для LD дикого типа (рис. 2С).
Эта неправильная регуляция генов FT в листьях мутантов late1 и dne согласуется с предыдущими выводами о том, что оба мутанта изменяют выработку передаваемых трансплантатом сигналов цветения (Hecht et al., 2007; Liew et al. ., 2009). Однако он не определяет, какой из генов FT может вносить вклад в эту передачу сигналов.
Локус
GIGAS соответствует FTa1 Ранее описанный мутант gigas ( gigas-1 ) поздно цветет при SD и цветет поздно или совсем не при LD, в зависимости от спектрального качества увеличение светового периода (Beveridge, Murfet, 1996).Второй мутантный аллель ( gigas-2 ) имеет несколько более тяжелый фенотип, неспособный зацвести ни при каких условиях LD (Reid et al. , 1996). В то время как фенотип SD мутантов gigas по существу подобен дикому типу, за исключением задержки цветения (Beveridge and Murfet, 1996), фенотип LD необычен. Несмотря на неспособность к цветению, мутанты демонстрируют ряд вегетативных изменений в ответ на LD, включая уменьшение длины междоузлий, уменьшение размера листа и разрастание пазушных почек (Beveridge and Murfet, 1996; Рисунок 5A).Этот фенотип очень отличается от фенотипа мутантов с фотопериодным ответом late1, и phyA , которые по существу фенокопируют SD-выращенные растения дикого типа (Weller et al., 2004; Hecht et al., 2007), и больше похож на фенотип других мутантов гороха, которые нарушают цветение и / или развитие соцветий, таких как нецветущие мутанты veg1 и veg2 (Reid et al., 1996; Benlloch et al., 2007). Тем не менее, нормальное цветение может быть восстановлено в гига путем прививки к дикому типу, вовлекая GIGAS в производство передаваемого трансплантатом стимула цветения (Beveridge and Murfet, 1996) и предлагая в качестве кандидатов FT -подобных генов.
, 1996). В то время как фенотип SD мутантов gigas по существу подобен дикому типу, за исключением задержки цветения (Beveridge and Murfet, 1996), фенотип LD необычен. Несмотря на неспособность к цветению, мутанты демонстрируют ряд вегетативных изменений в ответ на LD, включая уменьшение длины междоузлий, уменьшение размера листа и разрастание пазушных почек (Beveridge and Murfet, 1996; Рисунок 5A).Этот фенотип очень отличается от фенотипа мутантов с фотопериодным ответом late1, и phyA , которые по существу фенокопируют SD-выращенные растения дикого типа (Weller et al., 2004; Hecht et al., 2007), и больше похож на фенотип других мутантов гороха, которые нарушают цветение и / или развитие соцветий, таких как нецветущие мутанты veg1 и veg2 (Reid et al., 1996; Benlloch et al., 2007). Тем не менее, нормальное цветение может быть восстановлено в гига путем прививки к дикому типу, вовлекая GIGAS в производство передаваемого трансплантатом стимула цветения (Beveridge and Murfet, 1996) и предлагая в качестве кандидатов FT -подобных генов.
Геномы гороха и Medicago тесно синтеничны (Aubert et al., 2006), а расположение GIGAS примерно соответствует положению кластера FTa / FTc в Medicago (Hecht et al., 2005). Мы подтвердили консервативное расположение этих генов у гороха, а затем исследовали, может ли какой-либо из них быть нарушен у мутантов гига . Мы не обнаружили различий в кодирующих областях FTa2 и FTc в гига-1 или гигас-2 , но выявили значительные изменения в гене FTa1 у обоих мутантов по сравнению с их линиями-предшественниками cv Virtus ( гигаз-1 ) и резюме Porta ( гигаз-2 ).Мутант gigas-1 содержал замену одного нуклеотида в 3′-сайте сплайсинга интрона 2 (фиг. 5C), что, по прогнозам, приведет к пропуску экзона 3 и немедленному прекращению трансляции после Trp-88. ПЦР с праймерами FTa1 на кДНК gigas-1 привела к амплификации только ожидаемого мутантного продукта, в молекуле кДНК отсутствовал 41 п.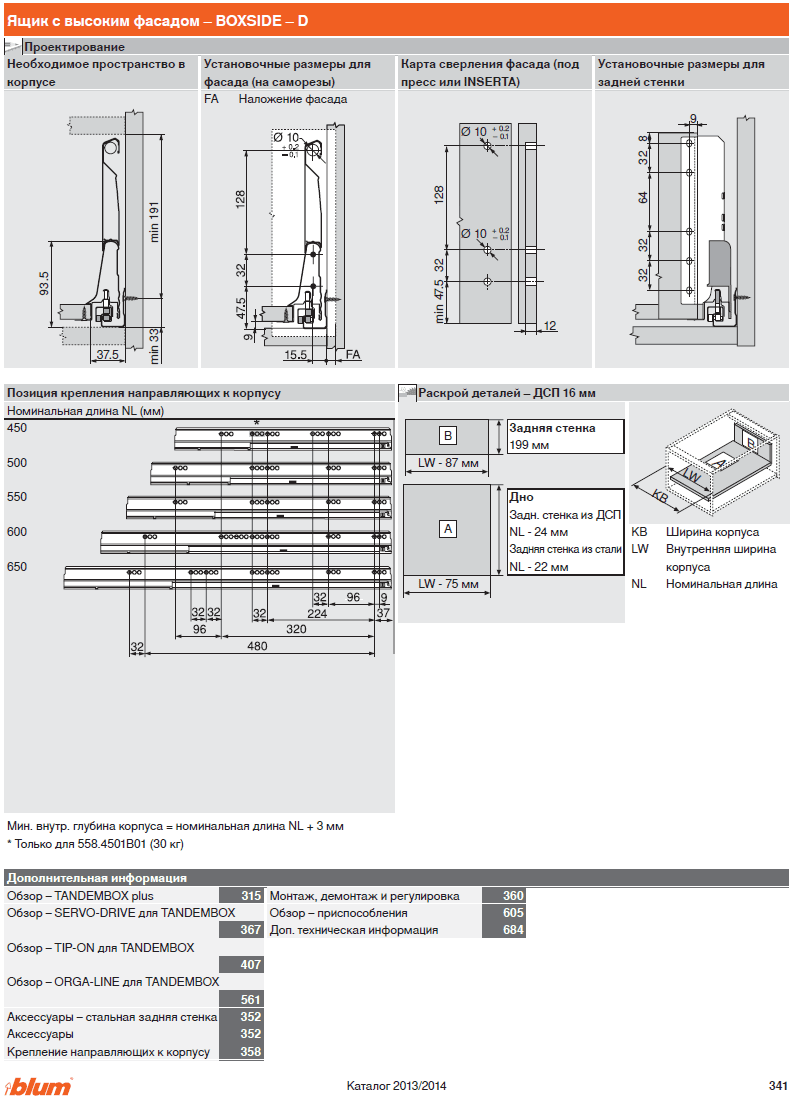 н. экзона 3, и кДНК FTa1 дикого типа не могла быть обнаружена. В случае мутанта gigas-2 неспособность амплифицировать FTa1 из геномной ДНК предполагала наличие значительной делеции или реаранжировки.Третий мутант гига ( гигас-3 ) был идентифицирован среди мутантов с поздним цветением из мутагенизированной этилметансульфонатом популяции (Hecht et al., 2007), и было обнаружено, что он несет замену C-to-T в экзоне 4 FTa1 , который преобразовал Gln-127 в стоп-кодон (рис. 5C). Последовательность FTa2 была идентична последовательности дикого типа в гига-3 , и поскольку рекомбинанты с FTc были идентифицированы во время картирования, последовательность FTc не исследовалась в гига-3 .Трансформация мутанта Arabidopsis ft-1 кДНК FTa1 из gigas-1 не смогла спасти фенотип позднего цветения (см. Дополнительный рисунок 4 онлайн), подтверждая, что функция FTa1 значительно нарушена мутация gigas-1 , как и ожидалось.
н. экзона 3, и кДНК FTa1 дикого типа не могла быть обнаружена. В случае мутанта gigas-2 неспособность амплифицировать FTa1 из геномной ДНК предполагала наличие значительной делеции или реаранжировки.Третий мутант гига ( гигас-3 ) был идентифицирован среди мутантов с поздним цветением из мутагенизированной этилметансульфонатом популяции (Hecht et al., 2007), и было обнаружено, что он несет замену C-to-T в экзоне 4 FTa1 , который преобразовал Gln-127 в стоп-кодон (рис. 5C). Последовательность FTa2 была идентична последовательности дикого типа в гига-3 , и поскольку рекомбинанты с FTc были идентифицированы во время картирования, последовательность FTc не исследовалась в гига-3 .Трансформация мутанта Arabidopsis ft-1 кДНК FTa1 из gigas-1 не смогла спасти фенотип позднего цветения (см. Дополнительный рисунок 4 онлайн), подтверждая, что функция FTa1 значительно нарушена мутация gigas-1 , как и ожидалось.
Локус GIGAS соответствует FTa1.
(A) Типичные растения двух независимых мутантов gigas и их исходных предшественников дикого типа (WT) линий Virtus (VIR) и Porta (POR).Растения выращивали в течение ~ 12 недель в условиях LD (18 световых / 6 темных) в теплице.
(B) Влияние мутации gigas-2 на цветущий узел в генетическом фоне NGB5839. Растения выращивали в фитотроне в условиях SD (8 световых / 16 темных) и LD (16 светлых / 8 темных). Значения представляют собой среднее ± стандартное отклонение для n = от 8 до 12. Нецветущий фенотип gigas LD, показанный в (A) , представлен диагональной штриховкой и стрелкой.Эти растения показаны с номинальной стоимостью, которая приблизительно соответствует узлу, в котором начался фенотип с короткими междоузлиями и сильным разветвлением.
(C) Схема гена FTa1 , показывающая природу и расположение мутаций у мутантов гига . Экзоны показаны прямоугольниками, кодирующая последовательность заштрихована серым, а нетранслируемые области — черным. В мутанте gigas-2 полностью удалено FTa1 .
Экзоны показаны прямоугольниками, кодирующая последовательность заштрихована серым, а нетранслируемые области — черным. В мутанте gigas-2 полностью удалено FTa1 .
(D) Экспрессия гена в диком типе (темные кружки) и мутант gigas-2 (белые кружки) во время развития в условиях LD (16 световых / 8 темных).Относительные уровни транскриптов определяли в рассеченных верхушках побегов или в самом верхнем полностью развернувшемся листе. Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 2 до 3 биологических повторов, каждая из которых состоит из объединенного материала от двух растений.
[Цветную версию этого рисунка см. В онлайн-статье.]
Молекулярные последствия
гига мутантов Далее мы исследовали, как потеря функции FTa1 влияет на регуляцию развития генов, связанных с идентичностью соцветия и переходом цветков. На фигуре 5D показано, что экспрессия PIM и SEP1 полностью отсутствует в гига , что согласуется с общей неспособностью мутантов гига производить цветы в условиях LD. Экспрессия UNI была намного ниже, чем у дикого типа, но все же предполагалась остаточная слабая индукция. Напротив, индукция DET и FD , хотя и относительно слабая, была сходной у дикого типа и гига (фиг.5D).Время и апекс-специфическая экспрессия FTc в растениях дикого типа при LD (рис. 2B) предполагают, что FTc может быть транскрипционной мишенью для других экспрессируемых листом генов FT , поэтому мы также исследовали, как потеря FTa1 влияет на экспрессию FTc и других генов FT . На фигуре 5D показано, что слабая индукция экспрессии FTa2 в ткани листа дикого типа полностью отсутствовала в гига , тогда как сильная индукция экспрессии FTb2 не пострадала.
На фигуре 5D показано, что экспрессия PIM и SEP1 полностью отсутствует в гига , что согласуется с общей неспособностью мутантов гига производить цветы в условиях LD. Экспрессия UNI была намного ниже, чем у дикого типа, но все же предполагалась остаточная слабая индукция. Напротив, индукция DET и FD , хотя и относительно слабая, была сходной у дикого типа и гига (фиг.5D).Время и апекс-специфическая экспрессия FTc в растениях дикого типа при LD (рис. 2B) предполагают, что FTc может быть транскрипционной мишенью для других экспрессируемых листом генов FT , поэтому мы также исследовали, как потеря FTa1 влияет на экспрессию FTc и других генов FT . На фигуре 5D показано, что слабая индукция экспрессии FTa2 в ткани листа дикого типа полностью отсутствовала в гига , тогда как сильная индукция экспрессии FTb2 не пострадала. Мутация гигабайт также предотвращала экспрессию FTa2 в апикальной ткани, тогда как FTc индуцировалась с аналогичным временем, но до более низкого уровня в гигах , чем у дикого типа (фигура 5D). Таким образом, FTa1 / GIGAS необходим для экспрессии генов идентичности соцветий PIM и SEP1 в LD, а также вносит основной вклад в индукцию UNI . Кроме того, FTa1 / GIGAS также требуется для нормальной экспрессии FTa2 в листе и верхушке и FTc в верхушке.Это предполагает, что перекрестная регуляция между генами FT может быть важной особенностью во время индукции цветения гороха.
Мутация гигабайт также предотвращала экспрессию FTa2 в апикальной ткани, тогда как FTc индуцировалась с аналогичным временем, но до более низкого уровня в гигах , чем у дикого типа (фигура 5D). Таким образом, FTa1 / GIGAS необходим для экспрессии генов идентичности соцветий PIM и SEP1 в LD, а также вносит основной вклад в индукцию UNI . Кроме того, FTa1 / GIGAS также требуется для нормальной экспрессии FTa2 в листе и верхушке и FTc в верхушке.Это предполагает, что перекрестная регуляция между генами FT может быть важной особенностью во время индукции цветения гороха.
Генетические взаимодействия
гига с другими цветущими мутантами Ввиду отчетливого цветения и молекулярных фенотипов, демонстрируемых мутантами гига и фотопериодическими ответными мутантами, мы затем исследовали их генетические взаимодействия. Удивительно, но введение мутации late1 в фон gigas восстановило цветение и вернуло общий фенотип к более сходному с фенотипом late1 (см. Дополнительный рисунок 5 онлайн).Однако мутант late1 gigas зацвел позже, чем одиночный мутант late1 по LD (рис. 6A), и разница между late1 и late1 gigas в LD была аналогична разнице между диким типом и gigas. в SD. Это указывает на то, что эффект late1 заключается просто в придании фенотипа SD мутанту gigas по LD. Это также обеспечивает дополнительную иллюстрацию того, что мутант gigas , несмотря на то, что он не цветет в LD, сохраняет сильный ответ на LD, который может быть заблокирован мутацией late1 .Двойные мутанты между гига и нечувствительным к световому периоду раннего цветения мутантом dne были легко различимы при SD по нецветущему фенотипу, очень похожему на одиночный мутант gigas , выращенный при LD (рис. 6А). Таким образом, эффект dne на gigas является обратным эффекту late1 , что придает фенотип LD мутанту gigas при выращивании в SD.
Дополнительный рисунок 5 онлайн).Однако мутант late1 gigas зацвел позже, чем одиночный мутант late1 по LD (рис. 6A), и разница между late1 и late1 gigas в LD была аналогична разнице между диким типом и gigas. в SD. Это указывает на то, что эффект late1 заключается просто в придании фенотипа SD мутанту gigas по LD. Это также обеспечивает дополнительную иллюстрацию того, что мутант gigas , несмотря на то, что он не цветет в LD, сохраняет сильный ответ на LD, который может быть заблокирован мутацией late1 .Двойные мутанты между гига и нечувствительным к световому периоду раннего цветения мутантом dne были легко различимы при SD по нецветущему фенотипу, очень похожему на одиночный мутант gigas , выращенный при LD (рис. 6А). Таким образом, эффект dne на gigas является обратным эффекту late1 , что придает фенотип LD мутанту gigas при выращивании в SD.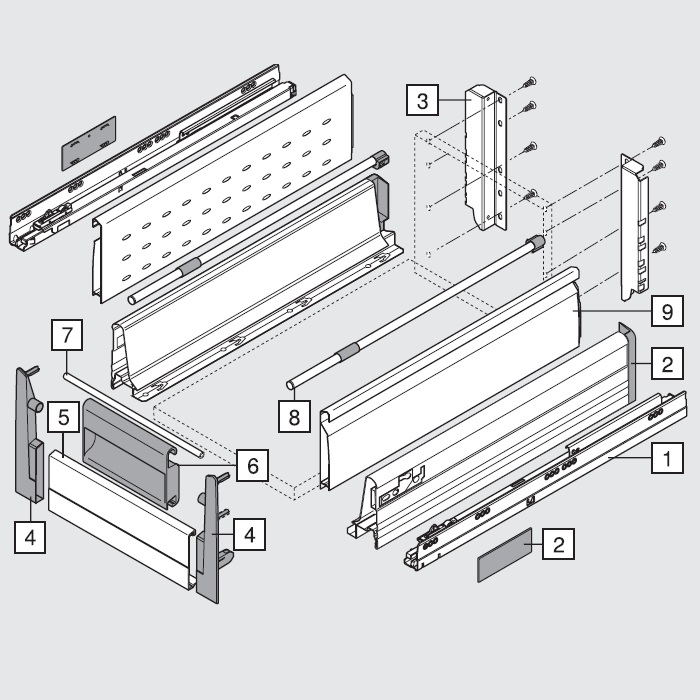 Оба эти взаимодействия можно рассматривать как существенно аддитивные в том смысле, что DNE и LATE1 оказывают свое влияние на чувствительность фотопериода независимо от GIGAS .Мы также исследовали, как взаимодействие мутаций late1 и gigas и восстановление цветения на фоне gigas к late1 может отразиться на экспрессии генов. Рисунок 6B и дополнительный рисунок 6 онлайн показывают, что позднее цветение двойного мутанта late1 gigas связано с четкой индукцией экспрессии PIM , SEP1 и UNI на верхушке, несмотря на отсутствие детектируемой экспрессии каких-либо Ген FT в ткани листа.
Оба эти взаимодействия можно рассматривать как существенно аддитивные в том смысле, что DNE и LATE1 оказывают свое влияние на чувствительность фотопериода независимо от GIGAS .Мы также исследовали, как взаимодействие мутаций late1 и gigas и восстановление цветения на фоне gigas к late1 может отразиться на экспрессии генов. Рисунок 6B и дополнительный рисунок 6 онлайн показывают, что позднее цветение двойного мутанта late1 gigas связано с четкой индукцией экспрессии PIM , SEP1 и UNI на верхушке, несмотря на отсутствие детектируемой экспрессии каких-либо Ген FT в ткани листа.
Генетические взаимодействия GIGAS.
(A) Сравнение цветковых узлов у двойных мутантов late1-2 gigas-2 , dne-1 gigas-2 и lf-22 gigas-2 у дикого типа (WT) NGB5839 , и соответствующие моногенные мутанты. Растения выращивали в фитотроне либо в режиме LD (16 световых / 8 темных) или SD (8 светлых / 16 темных). Как и на рисунке 5B, нецветущий фенотип gigas LD представлен диагональной штриховкой и стрелкой.Растения, демонстрирующие этот фенотип, показаны с номинальной стоимостью, которая приблизительно соответствует узлу, в котором начался фенотип с короткими междоузлиями и сильным разветвлением.
(B) Экспрессия гена в NGB5839 (дикий тип), gigas-2 и двойной мутант late1-2 gigas-2 во время развития в условиях LD (16 световых / 8 темных). Относительные уровни транскриптов определяли в рассеченных верхушках побегов или в самом верхнем полностью развернувшемся листе. Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 2 до 3 биологических повторов, каждая из которых состоит из объединенного материала от двух растений.Стрелки соответствуют видимому внешнему виду цветочных бутонов на рассеченных верхушках у дикого типа и двойных мутантов late1-2 gigas-2 .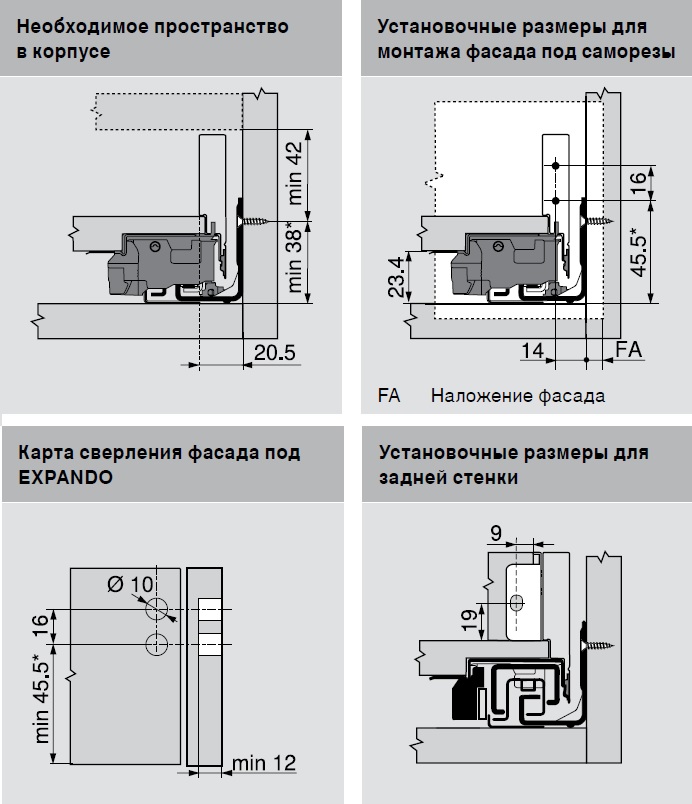
Наконец, мы исследовали генетическое взаимодействие между GIGAS / FTa1 и геном LATE FLOWERING ( LF ), паралогом Arabidopsis TFL1 (рис. 1), который задерживает цветение в фотопериод. независимым образом (Murfet, 1975; Foucher et al., 2003). Мы обнаружили, что фенотип раннего цветения предполагаемого нулевого мутанта lf ( lf-22 ) полностью эпистатичен нецветущему фенотипу gigas при LD, с двойным мутантом lf gigas , неотличимым от одиночного мутанта lf. мутант (Рисунок 6A; см. Дополнительный рисунок 5 онлайн).Это указывает на то, что GIGAS способствует цветению, отрицательно влияя на функцию LF .
Влияние
гига на передаваемые трансплантатом сигналы цветения Предыдущие эксперименты по прививке показали, что как позднецветущий фенотип SD, так и нецветущий фенотип LD мутанта gigas-1 можно исправить путем прививки к прививке дикого типа.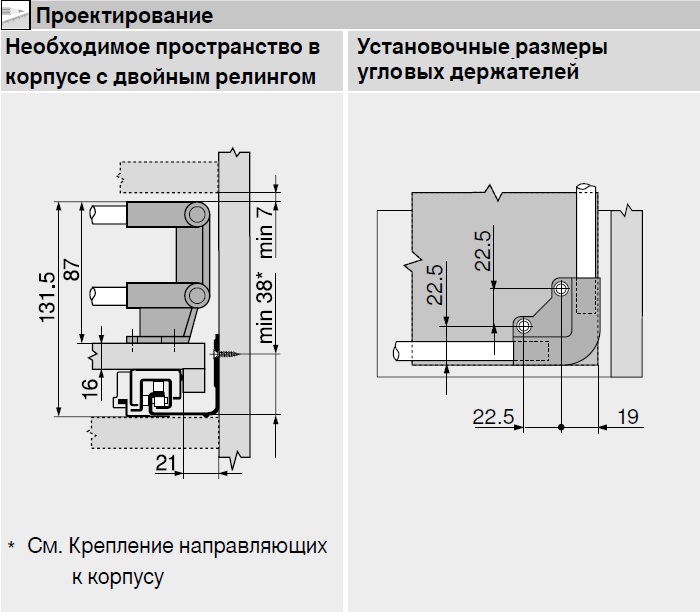 подвоя с листьями (Beveridge and Murfet, 1996). Рисунок 7A показывает, что это также верно для мутанта gigas-2 и подтверждает, что GIGAS / FTa1 необходим для производства стимула цветения, передаваемого через прививку.В предыдущих экспериментах фенотип позднего цветения мутанта late1 аналогичным образом мог быть спасен путем прививки к листовым привитым подвоям дикого типа (Hecht et al., 2007). Одна из возможных интерпретаций этих результатов заключается в том, что LATE1 действует через GIGAS / FTa1 для управления одним и тем же мобильным стимулом, которым может быть либо сам GIGAS / FTa1 , либо какой-либо другой сигнал ниже FTa1 . Такая интерпретация предсказывала бы, что запасы прививки мутанта gigas должны быть менее эффективными, чем запасы прививки дикого типа для спасения мутанта late1 .На рисунке 7B показано, что поздних1 привоев, привитых на гига подвоев, зацвели значительно позже (14,4 ± 0,1 узла), чем при прививке к подвоям дикого типа (13,2 ± 0,1 узлов, P <0,01), но все же зацвели намного раньше, чем поздних1.
подвоя с листьями (Beveridge and Murfet, 1996). Рисунок 7A показывает, что это также верно для мутанта gigas-2 и подтверждает, что GIGAS / FTa1 необходим для производства стимула цветения, передаваемого через прививку.В предыдущих экспериментах фенотип позднего цветения мутанта late1 аналогичным образом мог быть спасен путем прививки к листовым привитым подвоям дикого типа (Hecht et al., 2007). Одна из возможных интерпретаций этих результатов заключается в том, что LATE1 действует через GIGAS / FTa1 для управления одним и тем же мобильным стимулом, которым может быть либо сам GIGAS / FTa1 , либо какой-либо другой сигнал ниже FTa1 . Такая интерпретация предсказывала бы, что запасы прививки мутанта gigas должны быть менее эффективными, чем запасы прививки дикого типа для спасения мутанта late1 .На рисунке 7B показано, что поздних1 привоев, привитых на гига подвоев, зацвели значительно позже (14,4 ± 0,1 узла), чем при прививке к подвоям дикого типа (13,2 ± 0,1 узлов, P <0,01), но все же зацвели намного раньше, чем поздних1. самопривитых растения (19,7 ± 0,5 узла, P <0,01). Кроме того, поздних1 подвоев были лишь немного менее эффективны, чем дикий тип в стимулировании цветения у привоев гига (Рисунок 7B; 15,1 ± 0,2 по сравнению с 13,2 ± 0,1 узлов, P <0.01), указывая на то, что небольшое влияние late1 на экспрессию GIGAS / FTa1 в листьях (рис. 4A) оказывает лишь незначительное влияние на сигнал мобильного цветения. Эти дополнительные эффекты late1 и gigas указывают на то, что GIGAS / FTa1 действительно вносят вклад в сигнал цветения, но также предполагают роль компонента сигнала, независимого от GIGAS .
самопривитых растения (19,7 ± 0,5 узла, P <0,01). Кроме того, поздних1 подвоев были лишь немного менее эффективны, чем дикий тип в стимулировании цветения у привоев гига (Рисунок 7B; 15,1 ± 0,2 по сравнению с 13,2 ± 0,1 узлов, P <0.01), указывая на то, что небольшое влияние late1 на экспрессию GIGAS / FTa1 в листьях (рис. 4A) оказывает лишь незначительное влияние на сигнал мобильного цветения. Эти дополнительные эффекты late1 и gigas указывают на то, что GIGAS / FTa1 действительно вносят вклад в сигнал цветения, но также предполагают роль компонента сигнала, независимого от GIGAS .
GIGAS на передаваемую трансплантатом цветочную сигнализацию.
Влияние мутаций гигабайт на сигналы подвижных цветков было исследовано путем прививки 6-дневных побегов, вырезанных в эпикотиле (привитой привой), на основной стебель 3-недельных растений над самым верхним полностью развернувшимся листом (прививка). склад). Для каждой комбинации прививок показаны генотипы привоя (вверху) и подвоя (внизу), разделенные горизонтальной линией.
склад). Для каждой комбинации прививок показаны генотипы привоя (вверху) и подвоя (внизу), разделенные горизонтальной линией.
(A) Прививка к дикому типу (WT) спасает фенотип gigas-2 при LD и SD. Самостоятельно привитые гига показали фенотип, типичный для непривитых гига контрольных растений, тогда как прививка гига к дикому типу восстановила фенотип, близкий к дикому типу.
(B) Дополнительная прививка-трансмиссивная стимуляция цветения гига-2 и late1-2 в LD. Поскольку ранее наблюдали, что оба мутанта late1 и gigas влияют на стимул, передаваемый трансплантатом, их способность дополнять друг друга через соединение трансплантата была исследована в сравнении с самотрансплантатами и трансплантатами мутантов дикого типа.
(C) GIGAS — независимая от трансплантата стимуляция цветения мутантом dne-1 в SD.Важность GIGAS / FTa1 для трансмиссивного эффекта dne была исследована путем сравнения способности запасов одинарных мутантов dne и двойных мутантов dne gigas стимулировать цветение привоев дикого типа, содержащихся в SD.
(D) Экспрессия генов FT в самом верхнем расширенном листе прививки, используемой в (C) . Значения были нормализованы к уровню транскрипта гена ACTIN и представляют собой среднее ± стандартное отклонение для n = от 4 до 6 биологических повторов.
Все растения выращивали в фитотроне либо в LD (16 световых / 8 темных) или SD (8 светлых / 16 темных). Данные в (A) от до (C) являются средними ± стандартными значениями для n = от 6 до 15. Как и на рисунке 5B, нецветущий фенотип gigas-2 LD представлен в (A) от до . (C) диагональной штриховкой и стрелкой. Растения, демонстрирующие этот фенотип, показаны с номинальной стоимостью, которая приблизительно соответствует узлу, в котором начался фенотип с короткими междоузлиями и сильным разветвлением.
Мы проверили этот вывод дополнительно, изучив, как гига может повлиять на способность привитых подвоев dne стимулировать цветение через стык трансплантата в условиях SD. Мутант dne имеет повышенную экспрессию FTa1 , FTa2 и FTb2 в ткани листа (рис. 4B), и мы пришли к выводу, что устранение активности FTa1 из привитых материалов с помощью gigas-2 мутация покажет, может ли какой-либо из оставшихся экспрессируемых на листьях генов FT влиять на цветение побегов.Рисунок 7D подтверждает, что 3-недельные проростки dne gigas , выращенные на SD, не имеют транскрипта FTa1 , как ожидалось, а также не могут экспрессировать FTa2 или FTb1 , но экспрессируют значительно более высокие уровни FTb2 , чем у дикого типа. или одиночный мутант gigas . Это сопровождается повышенной способностью стимулировать цветение у привоев дикого типа в условиях SD (рис. 7C; 17,0 ± 1,2 по сравнению с 21,7 ± 0,6 узла, P <0,01), что позволяет предположить, что FTb2 связано с образованием второго подвижного сигнал цветения, не зависящий от GIGAS / FTa1 .
Мутант dne имеет повышенную экспрессию FTa1 , FTa2 и FTb2 в ткани листа (рис. 4B), и мы пришли к выводу, что устранение активности FTa1 из привитых материалов с помощью gigas-2 мутация покажет, может ли какой-либо из оставшихся экспрессируемых на листьях генов FT влиять на цветение побегов.Рисунок 7D подтверждает, что 3-недельные проростки dne gigas , выращенные на SD, не имеют транскрипта FTa1 , как ожидалось, а также не могут экспрессировать FTa2 или FTb1 , но экспрессируют значительно более высокие уровни FTb2 , чем у дикого типа. или одиночный мутант gigas . Это сопровождается повышенной способностью стимулировать цветение у привоев дикого типа в условиях SD (рис. 7C; 17,0 ± 1,2 по сравнению с 21,7 ± 0,6 узла, P <0,01), что позволяет предположить, что FTb2 связано с образованием второго подвижного сигнал цветения, не зависящий от GIGAS / FTa1 .
ОБСУЖДЕНИЕ
Становится все более очевидным, что представители семейства FT играют важную роль в переходе цветков. По сравнению с Arabidopsis , где были проведены наиболее подробные исследования, семейства FT у многих других видов значительно сложнее, а функциональный анализ гораздо менее продвинут. В этом исследовании мы охарактеризовали семейство FT в трех модельных бобовых культурах, сои, Medicago и горохе, и определили три основные ветви, которые, вероятно, являются общими для всех бобовых культур.Затем мы исследовали вклад генов гороха в контроль цветения, светочувствительность и мобильную передачу сигналов.
Экспрессия генов гороха
FT По сравнению с простым семейством Arabidopsis FT , в котором FT и TSF имеют сходные паттерны экспрессии, мы наблюдали отчетливо разные паттерны экспрессии генов гороха FT . В ткани листьев растений в LD только FTb2 явно индуцировалось в течение периода, когда растение становится физиологически приверженным к цветению (фиг. 2).Эта индукция специфична для фотопериода и вместе с очевидным отсутствием экспрессии в апикальных зачатках указывает на то, что FTb2 может играть роль или в продукции мобильного сигнала для индукции цветков в LD. Подобно FTb2 , FTa1 и FTa2 также индуцируются в листьях, но их экспрессия не регулируется по-разному в зависимости от продолжительности светового дня в течение периода до фиксации цветков в LD, что позволяет предположить, что они с меньшей вероятностью участвуют в начальной передаче сигналов для LD. цветочная индукция.Однако в апикальных зачатках FTa1 и FTc показали четкую, ускоренную LD индукцию, которая происходила после фиксации цветков с аналогичным временем индукции генов идентичности соцветий PIM , UNI и SEP1 . Это повышает вероятность того, что оба этих гена FT могут играть роль в верхушке, которая является промежуточной между мобильным белком FT и генами, определяющими идентичность соцветия, или действовать параллельно с этими генами в качестве мишеней для флоригенов.Особый интерес представляет то, что экспрессия FTc снижена как у gigas , так и у late1 мутантов (рисунки 4 и 5), а эффект late1 по крайней мере частично не зависит от GIGAS (см. Дополнительный рисунок 6). онлайн). Это представляет собой ясный пример перекрестной регуляции транскрипции среди членов семейства генов FT и дополнительно подтверждает возможность того, что FTc может интегрировать передачу сигналов дальнего действия от других генов FT .
2).Эта индукция специфична для фотопериода и вместе с очевидным отсутствием экспрессии в апикальных зачатках указывает на то, что FTb2 может играть роль или в продукции мобильного сигнала для индукции цветков в LD. Подобно FTb2 , FTa1 и FTa2 также индуцируются в листьях, но их экспрессия не регулируется по-разному в зависимости от продолжительности светового дня в течение периода до фиксации цветков в LD, что позволяет предположить, что они с меньшей вероятностью участвуют в начальной передаче сигналов для LD. цветочная индукция.Однако в апикальных зачатках FTa1 и FTc показали четкую, ускоренную LD индукцию, которая происходила после фиксации цветков с аналогичным временем индукции генов идентичности соцветий PIM , UNI и SEP1 . Это повышает вероятность того, что оба этих гена FT могут играть роль в верхушке, которая является промежуточной между мобильным белком FT и генами, определяющими идентичность соцветия, или действовать параллельно с этими генами в качестве мишеней для флоригенов.Особый интерес представляет то, что экспрессия FTc снижена как у gigas , так и у late1 мутантов (рисунки 4 и 5), а эффект late1 по крайней мере частично не зависит от GIGAS (см. Дополнительный рисунок 6). онлайн). Это представляет собой ясный пример перекрестной регуляции транскрипции среди членов семейства генов FT и дополнительно подтверждает возможность того, что FTc может интегрировать передачу сигналов дальнего действия от других генов FT .
Существование множества, по-разному регулируемых членов семейства FT , которые мы наблюдаем здесь у гороха, также было описано для нескольких других видов. Например, у ячменя ( Hordeum vulgare ) два FT -подобных гена преимущественно экспрессируются при LD, но различаются по времени развития и суточному времени, тогда как третий ген предпочтительно экспрессируется при SD (Faure et al., 2007; Kikuchi и др., 2009). У подсолнечника ( Helianthus annuus ) один из трех функциональных генов, подобных FT , специфически экспрессируется в верхушке побега, но не в листьях (Blackman et al., 2010), тогда как у кукурузы множественные гены FT экспрессируются в различных тканях, включая корни, листья, развивающиеся соцветия и семена (Данилевская и др., 2008). Следующей сложной задачей будет оценка функциональной значимости этих различий. Эксперименты по комплементации Arabidopsis показывают, что все пять белков FT гороха обладают способностью в некоторой степени функционировать подобно Arabidopsis FT. Они также предположили явные различия в активности, но для доказательства этого явно потребуется функциональный анализ в системе гороха.
GIGAS / FTa1 ФункцияИдентификация мутантов gigas-1 и gigas-2 как РНК-нулевых аллелей FTa1 позволила сделать первый шаг в понимании функций генов гороха FT . Эти мутанты ясно показывают, что FTa1 играет важную роль в контроле цветения не только при LD, но также и при SD. Исследования риса и Arabidopsis также сообщают о незначительном влиянии генов FT на цветение в неиндуктивных условиях, так что это не является неожиданностью (Yamaguchi et al., 2005; Комия и др., 2008). Однако фенотип gigas LD, который сочетает в себе неспособность зацвести с сохранением LD-ответов для других признаков, отличается от мутантов FT у других видов и показывает, что FTa1 не опосредует общий ответ на фотопериод. Большее сходство гига с идентичными мутантами по соцветию гороха, такими как veg1 и veg2 , вместо этого указывает на то, что роль GIGAS / FTa1 ограничивается индукцией цветения (Reid and Murfet, 1984; Benlloch et al., 2007). Это также согласуется с наблюдением, что экспрессия FTa1 в листьях дикого типа не индуцируется LD в окне физиологического обязательства, а происходит немного позже, параллельно с индукцией генов идентичности соцветия и собственной индукцией в верхушечных почках (рис. 2). Дополнительную поддержку обеспечивает мутант late1 , который, несмотря на отсутствие чувствительности к световому периоду, показывает лишь незначительные эффекты на экспрессию FTa1 в листьях (рис. 4A; Hecht et al., 2007) во время окна физиологической приверженности.
Помимо поддержки роли FTa1 в приобретении идентичности соцветия и исключения основной роли в реактивности фотопериода, сравнение между late1 и gigas предполагает связь между другим экспрессируемым листом геном FT , FTb2 , и реакция на фотопериод. Подобно дикому типу в SD, мутанты late1 в LD демонстрируют отсроченную индукцию FTa1 и FTc и не оказывают явного влияния на FTa2 , но не способны полностью индуцировать экспрессию FTb2 , тогда как в гига. , FTb2 индуцируется нормально (рисунки 4A и 5D).Таким образом, несколько линий доказательств идентифицируют FTb2 в качестве сильного кандидата в первичный ген FT , контролирующий фотопериодный ответ: он не экспрессируется в SD и сильно активируется во время индукции цветков в LD, и это усиление блокируется у мутанта фотопериодного ответа. и не затронуты мутантами гига .
В Arabidopsis и злаках умеренного климата гены FT также интегрируют передачу сигналов от путей яровизации (Kim et al., 2009). Мы не изучали яровизацию в этом исследовании, но предыдущий отчет показал, что яровизация могла вызвать цветение гига-1 в LD и была не менее эффективной в гига-1 , чем в диком типе для стимулирования цветения в SD ( Беверидж и Мерфет, 1996). Это указывает на то, что FTa1 не является основной мишенью для регуляции с помощью яровизации у гороха и что яровизация может противодействовать любому фактору, ответственному за вегетативный фенотип LD гига .Однако, как сообщается, яровизация гороха действует посредством различных механизмов в листьях и на верхушках (Reid and Murfet, 1975), а ответ на основе листьев в гига , по-видимому, отсутствует (Beveridge and Murfet, 1996), что позволяет предположить, что Тем не менее FTa1 может опосредовать компонент реакции яровизации. В будущем будет интересно изучить, как яровизация может регулировать гены гороха FT и более широкие молекулярные фенотипы гига и мутантов с фотопериодным ответом.
Два отчетливых сигнала подвижного цветения у гороха
Эксперименты по прививке показывают, что недостаток FTa1 в побегах может быть явно компенсирован некоторым фактором, перемещающимся из листьев или других исходных тканей (рис. 7A). Ввиду растущего числа примеров, где FT-подобные белки действуют как мобильные сигналы (Turck et al., 2008), и явная индукция FTa1 в листьях под LD, наиболее вероятное объяснение спасения трансплантата гига мутанты — это движение самого белка FTa1.Альтернативное объяснение состоит в том, что сигнал — это какая-то другая молекула, которая регулируется FTa1 . Например, поскольку выражение FTa2 в листьях зависит от GIGAS / FTa1 (рис. 5D), возможно, что FTa2 может играть эту роль.
Как и в случае гига , позднецветущий фенотип мутанта late1 также может быть спасен путем прививки к дикому типу (Hecht et al., 2007), даже если gigas и late1 мутанты имеют явно разные фенотипы и профили экспрессии FT (фиг. 4A и 5D).Простейшей интерпретацией этого могло бы быть то, что один и тот же FTa1 -зависимый сигнал является дефицитным у отпрысков обоих мутантов и может поставляться из запасов дикого типа, но несколько линий доказательств предполагают, что это не так. Во-первых, late1 мутантов показывают лишь небольшое снижение экспрессии FTa1 и имеют сходную экспрессию FTa2 по сравнению с диким типом. Во-вторых, гига привоев, привитых к поздним1 подвоям , вызывают цвести и цвести гораздо раньше, чем поздних1 прививок (Рисунок 7B; Hecht et al., 2007). Этого не было бы, если бы единственной причиной позднего цветения late1 и отсутствия цветения гига был недостаток того же мобильного сигнала. Действительно, тот факт, что среди пяти генов FT только FTa1 и FTa2 экспрессируются в ткани листа мутанта late1 , обеспечивает дополнительную поддержку идеи о том, что FTa1 отвечает за генерацию мобильного сигнала в late1 запасов, способных спасти мутанта gigas .В-третьих, значительное ускорение цветения происходит у поздних1 отпрысков при прививке в гига подвоев под LD. Тот факт, что в запасах gigas-2 отсутствует ген FTa1 , явно исключает белок FTa1 в качестве активной молекулы в этом случае, и ввиду незначительного уровня экспрессии FTa2 участие белка FTa2 также маловероятно. Кроме того, из трех экспрессируемых на листьях генов FT только FTb2 значительно экспрессируется в гига , что означает, что если спасение прививки поздних1 отпрысков гига запасов отражает FT -зависимую генерацию сигнала. в запасе уходит, то FTb2 — единственный вероятный кандидат.Этот вывод подтверждается результатами трансплантатов с двойным мутантом dne gigas при SD (рис. 7C). Как и одиночный мутант gigas , dne gigas не экспрессирует FTa1 или FTa2 , но более эффективен, чем gigas , в стимулировании цветения привоев дикого типа и экспрессирует гораздо более высокие уровни транскрипта FTb2 в листе. ткань (рис. 7D). Общей чертой обоих этих двух последних экспериментов по прививке, таким образом, является ассоциация между способностью стимулировать цветы и наличием значительной экспрессии FTb2 в листьях: в гига стада под LD и в dne gigas под SD.
В целом, эти результаты указывают на сосуществование двух различных передаваемых трансплантатом сигналов, способствующих развитию цветков у гороха, которые коррелируют с экспрессией генов FTa1 и FTb2 в ткани листа. Кроме того, разные профили экспрессии этих генов во время развития дикого типа при LD и их связь с различными фенотипами цветения у мутантов late1 и gigas предполагают гипотезу, согласно которой FTa1 и FTb2 влияют на процесс цветения через механизмы, которые, по крайней мере, частично различны.Одна правдоподобная интерпретация заключается в том, что GIGAS / FTa1 может предоставлять независимый от фотопериода мониторинг размера растения или других переменных окружающей среды и действует конкретно на индукцию цветения, тогда как FTb2 может действовать как первичный сигнал, полученный из листьев для ускорения роста растений. цветение с помощью LD, а также отвечает за другие аспекты ответа LD (Рисунок 8). Несколько недавних исследований показали, что гены FT могут играть роль, выходящую за рамки простого инициирования цветения.Например, у томатов, у которых цветение не зависит от фотопериода, гомологи FT и TFL1 взаимодействуют для регулирования роста и детерминации нескольких процессов развития (Shalit et al., 2009), а у тополя ( Populus spp) , FT и Гомологи TFL1 также играют роль в индукции покоя (Böhlenius et al., 2006; Ruonala et al., 2008; Mohamed et al., 2010). Будущее выделение мутантов FTb2 , несомненно, станет важным тестом на эту возможность у гороха и прояснит другие аспекты функции FTb2 , включая его регуляторные отношения с FTa1 и FTc и его взаимодействие с TFL1 гомологами.
Рис. 8.Модель роли и взаимодействия генов гороха FT в цветении и фотопериодической реакции.
Эта модель суммирует основные результаты исследования и основные гипотезы, основанные на них. Гены фотопериодного ответа DNE и LATE1 регулируют экспрессию двух ключевых генов FT в листьях: FTa1 и FTb2 . Мы предполагаем, что оба гена генерируют различные мобильные сигналы, влияющие на цветение на верхушке.Мобильный сигнал FTa1 специфически действует на цветение как в LD, так и в SD, потенциально за счет противодействия репрессии со стороны LF из PIM и других генов идентичности соцветий. FTa1 -зависимый сигнал также вносит вклад в индукцию FTc и потенциально в экспрессию самого FTa1 на вершине. LD индуцирует экспрессию FTb2 в листьях и генерацию второго, потенциально зависимого от FTb2 , сигнала, который влияет на цветение посредством индукции FTa1 в листьях и (прямо или косвенно) на верхушках.Независимо от FTa1 , этот второй сигнал также индуцирует FTc на верхушке, регулирует ряд других процессов, зависимых от фотопериода, и может ингибировать индукцию цветков.
[Цветную версию этого рисунка см. В онлайн-статье.]
Контроль цветения без
FTa1Цветение одиночного мутанта gigas в SD или мутанта late1 gigas в LD показывает, что у гороха, переход цветков все еще может происходить в отсутствие значительной экспрессии любого гена FT в листьях.Это предполагает наличие основного пути по умолчанию, гарантирующего, что цветение в конечном итоге произойдет независимо от продолжительности светового дня, и предполагает, что модуль FT может быть наложен на него, чтобы обеспечить средство ускорения цветения при благоприятных условиях. В Arabidopsis растения, лишенные как FT , так и TSF , все еще цветут, и известны механизмы, которые обходят FT , чтобы индуцировать экспрессию генов идентичности меристемы цветков, включая регуляцию SOC1 гиббереллином (Moon et al., 2003; Jang et al., 2009). Будет интересно изучить, действует ли аналогичный механизм и у гороха.
Эксперименты по прививке показывают положительную роль FTa1 и FTb2 в инициации цветения. Однако отсутствие цветения у мутанта gigas в LD и тот факт, что восстановление цветения с помощью мутации late1 сочетается с потерей экспрессии FTb2 , явно подразумевают более сложную регуляцию. Одна возможность может заключаться в том, что условия LD индуцируют или активируют некоторый фактор, который ингибирует образование цветков в отсутствие GIGAS / FTa1 , и одним из потенциальных кандидатов на этот фактор может быть TFL1 paralog LF (Foucher et al., 2003). Как в Arabidopsis , так и в томате белки FT и TFL1 антагонистически взаимодействуют, регулируя индукцию цветков, что позволяет предположить, что активация генов-мишеней зависит от баланса активности этих двух генов (Kardailsky et al., 1999; Kobayashi et al., 1999 ; Шалит и др., 2009). У гороха потеря функции LF приводит к чрезвычайно раннему цветению, а полный эпистаз lf на гига (Рисунок 6; см. Дополнительный рисунок 5 онлайн) согласуется с механизмом, в котором GIGAS противостоит LF -опосредованное торможение цветения.Таким образом, возникает соблазн предположить, что вегетативный фенотип гига в LD может быть результатом LD-специфического увеличения активности LF , которому обычно противодействует активность GIGAS / FTa1 у дикого типа. В Arabidopsis , TFL1 транскрипционно активируется с помощью CO (Simon et al., 1996), а в горохе как LF , так и DET экспрессируются на более высоком уровне в LD, чем в SD (рис. ), и их экспрессия задерживается ( LF ) или снижается ( DET ) в late1 по сравнению с диким типом при LD (рис. 4A).Однако тот факт, что экспрессия LF ниже в нецветущем мутанте gigas , чем в двойном мутанте late1 gigas (см. Дополнительный рисунок 6 онлайн), исключает регуляцию транскрипции LF как прямое объяснение LD -специфический вегетативный фенотип. Другие объяснения могут включать пространственно локализованную посттранскрипционную регуляцию активности LF или действие некоторых других факторов. Подробное молекулярное сравнение гигабайт с мутантами с фотопериодной реакцией может дать больше понимания этого вопроса.
В заключение, это исследование подчеркивает важную роль генов гороха FT как в регулировании времени цветения, так и в определении цветочной идентификации. Наши результаты предполагают более сложный механизм индукции цветения гороха по сравнению с Arabidopsis , который включает перекрестную регуляцию между различными генами FT с различными паттернами экспрессии и различными присущими им активностями, а также действием по крайней мере двух мобильных сигналов, из которых один — GIGAS / FTa1.Учитывая, что в настоящее время описаны расширенные семейства FT с различными паттернами регуляции у ряда видов, вполне вероятно, что такая сложность окажется относительно обычным явлением у высших растений. Наши результаты также открывают путь для углубленного функционального анализа генов FT в других бобовых культурах и должны ускорить молекулярный анализ генетической изменчивости времени цветения и чувствительности к световому периоду, которая существует у многих видов бобовых культур.
МЕТОДЫ
Растительный материал, условия роста и прививка
Происхождение мутантов gigas-1 , le-3 , dne-1 и late1-2 было описано ранее (King и Мерфет, 1985; Беверидж и Мерфет, 1996; Лестер и др., 1999; Hecht et al., 2007). Мутант gigas-2 был создан W.K. Swiecicki после мутагенеза на быстрых нейтронах сорта Porta, и мутант gigas-3 был получен из линии NGB5839 с помощью этилметансульфонатного мутагенеза (Hecht et al., 2007). Растения для всех исследований экспрессии генов (Рисунки 2B, 2C, 4, 5D и 6B), эксперименты по цветению Arabidopsis thaliana (Рисунок 3) и эксперименты по переносу фотопериода (Рисунок 2A) проводились в шкафах для выращивания при 20 ° C, тогда как цветение эксперименты по времени и прививке (рисунки 5A, 5B, 6A и 7) были проведены в фитотроне Хобарта с использованием ранее описанных питательных сред, источников света, условий фитотрона и протоколов прививки (Hecht et al., 2007). Стандартные условия SD фитотрона состояли из 8-часового фотопериода естественного света, который был продлен на 8 часов белым светом от компактных люминесцентных трубок при освещенности 10 мкмоль м -2 с -1 , чтобы получить 16-часовой световой период. LD.
Выделение гена и филогенетический анализ
Все гены гороха ( Pisum sativum ) FT , кроме FTc , были выделены из кДНК листа и геномной ДНК дикого типа (NGB5839) с использованием методов ПЦР, быстрой амплификации концов кДНК ( Набор для амплификации кДНК SMART RACE; Clontech) и обход генома (GenomeWalker Universal kit; Clontech) с использованием специфических праймеров, созданных на начальном фрагменте ДНК, полученном с помощью вырожденных праймеров (Hecht et al., 2005). Полногеномная последовательность FTc была получена путем скрининга коммерческой геномной библиотеки гороха (Clontech). Все фрагменты ПЦР были клонированы в pGEM-T easy (Promega) и секвенированы в Австралийском исследовательском центре генома. Детали грунтовки приведены в дополнительной таблице 3 онлайн. Для филогенетического дерева, показанного на рисунке 1B, аминокислотные последовательности белков PEBP бобовых были выровнены с использованием ClustalX (Thompson et al., 1997). Для филогенетического анализа в PAUP * 4 использовались дистанционные и экономичные методы.0b10 (http://paup.csit.fsu.edu/), используя выравнивание, показанное на дополнительном рисунке 2 в Интернете.
Arabidopsis КомплементацияМутация Arabidopsis ft-1 на фоне Landsberg erecta была описана ранее (Koornneef et al., 1991; Kardailsky et al., 1999; Kobayashi et al., 1999) Фрагменты полноразмерной кДНК для генов гороха FT были получены с помощью ПЦР из линии гороха дикого типа NGB5839 и дополнительных полноразмерных кДНК FTa1 из исходного мутанта gigas-1 и его предшественника сорта Virtus с использованием праймеров, перечисленных в Дополнительных материалах. Таблица 3 онлайн.Фрагменты кДНК были сначала клонированы в векторе pCR8⁄GW⁄TOPO (Invitrogen), а затем рекомбинированы в бинарный вектор pB2GW7 с использованием клонирования Gateway (Karimi et al., 2002) и подтверждены секвенированием. Трансформацию Arabidopsis проводили окунанием цветов (Bechtold et al., 1993), и фенотипы цветения нескольких независимых трансформантов на конструкцию характеризовали на протяжении нескольких поколений.
Исследования экспрессии гена
Собранная ткань состояла из обоих листочков верхнего полностью развернутого листа или апикальных почек, рассеченных до размера ~ 2 мм в ширину и 3 мм в длину.Образцы замораживали в жидком азоте и общую РНК экстрагировали с использованием системы выделения общей РНК SV (Promega). Концентрации РНК определяли спектрофотометрическим анализом с использованием NanoDrop 8000 (Thermo Scientific). Обратную транскрипцию проводили в 20 мкл с 1 мкг тотальной РНК с использованием высокоэффективной обратной транскриптазы MMLV (Epicenter) в соответствии с инструкциями производителя. RT-отрицательные (без фермента) контроли были выполнены для мониторинга загрязнения геномной ДНК. КДНК первой цепи разводили в пять раз, и 2 мкл использовали в каждой реакции ПЦР в реальном времени.Реакции ПЦР в реальном времени с использованием SYBR green chemistry (Sensimix, Quantace, Bioline) были установлены с помощью роботизированной системы обработки жидкостей CAS-1200N (Corbett Research) и проводились в течение 50 циклов в Rotor-Gene RG3000 (Corbett Research). Для каждого образца были выполнены две технические повторы и от двух до трех биологических повторов. Уровни транскрипции нескольких потенциальных референсных генов, включая EF1a (Johnson et al., 2006), UBI (Platten et al., 2005) и ACTIN (Weller et al., 2009a), были исследованы в различных сериях тканей. Было обнаружено, что из этих генов наиболее стабильно экспрессируется ACTIN , и поэтому этот ген был использован для оценки уровней транскриптов генов цветения, как описано ранее (Weller et al., 2009a). Последовательности праймеров приведены в дополнительной таблице 3 онлайн.
Дополнительные данные
Следующие материалы доступны в онлайн-версии этой статьи.
Триммер Twister T2S — стандартный 1/4 «(сухой)
Twister — это самый быстрый в мире медицинский триммер для цветов, оснащенный самой передовой в отрасли геометрией лезвий, позволяющей срезать максимально точно, не повредив урожай.
ПОЗНАКОМЬТЕСЬ С ТРИММЕРОМ TWISTER T2
Крупная обрезка цветка — отстой. Но это не обязательно. Twister T2 преображает игру, используя передовую запатентованную технологию обрезки цветочных растений, которая сделает всю работу за вас (вместо ручной обрезки). Обрезайте до 19 фунтов медицинских цветов в час с помощью одной машины.
Лучшая в отрасли производительность
Предназначен для средних и крупных медицинских операций и учреждений. Гарантированная максимальная производительность и время безотказной работы.Положительный возврат инвестиций обычно достигается менее чем за один урожай!
15-19 фунтов / час *
* сухой при стрижке в условиях, рекомендованных Twister (или влажная стрижка 77-85 фунтов)
Наши стаканы изготовлены из нержавеющей стали 304 *. Шестидюймовый диаметр уменьшает проскальзывание стенок, что обеспечивает плавный и эффективный поток продукта через стакан.
* Мы никогда не используем покрытия PFE-PTFE на основе тефлона.
Оснащенный мощным двигателем, Twister T2 способен стабильно обеспечивать максимальную производительность во время сложных сеансов обрезки большого производства.
Увеличить масштаб?
Добавить конвейеры и подключить несколько твистеров
Заезд четверки в тандеме
T2 — единственная машина для обрезки цветов на планете, предназначенная для работы в тандеме. Развивайте свой бизнес с уверенностью, добавляя при необходимости до четырех триммеров, увеличивая профессиональную производительность до 220 фунтов в час без каких-либо дополнительных затрат труда!
| Т2 | Тип | Подача фунтов / час | Корм кг / час |
| 1 | Сухая | 14-16 | 6-7 |
| 2 | Сухая | 27-29 | 12-13 |
| 3 | Сухая | 35-38 | 16-17 |
| 4 | Сухая | 44-68 | 20-30 |
| 1 | Мокрая | 35-45 | 16–21 |
| 2 | Мокрая | 75-85 | 34-39 |
| 3 | Мокрая | 125-141 | 57-64 |
| 4 | Мокрая | 141-220 | 64-100 |
Мощный, но портативный?
Проверьте T4
Нужно больше?
Добавить второй T2
ВОПРОСЫ КАЧЕСТВА.
Twister T2 был разработан с нуля, чтобы превосходить строгие требования самых требовательных медицинских учреждений на планете. Его легкая, 100% водонепроницаемая конструкция из нержавеющей стали может обеспечить высокие результаты в медицинском производстве любого размера.
Вместо того, чтобы тратить дни или недели на усердную работу, подрезая урожай вручную, теперь то же самое можно сделать с помощью T2 за считанные часы!
Легендарное качество сборки
Приобретая Twister, вы инвестируете в более чем десятилетний производственный опыт.Качество заложено в нашей ДНК. Каждый день мы стремимся создавать лучшее оборудование на планете. Как измерить этот тип качества? Успех клиентов. T2 за последние 10 лет обрезал более 10 миллионов фунтов цветов.
Сохраните профиль вашего цветка
Зачем уменьшать внешний вид вашего цветка (с помощью других сухих триммеров), если вы можете сохранить естественную красоту и качественный профиль вашего цветка нетронутыми с помощью технологии Twister’s SoftTumble ™. Как это возможно? Оставьте магическое число 11.
Легкость разборки и очистки
Разбирается за секунды, обеспечивая легкий доступ к компонентам при необходимости. Инструменты не требуются.
Нам доверяют
На Twister T2 обрабатывается большая часть цветов мира. Большинство наших крупнейших производителей используют T2 каждый день до 12-14 часов в день. Возникает вопрос: если крупнейшие предприятия на планете доверяют Twister T2, может, и вам стоит тоже? #trustthebest
Масштабируйте и снижайте затраты.
Twister T2 разработан для крупномасштабных производственных линий, которые заботятся о стабильности и контроле качества. Большинство клиентов T2 начинали с одной машины, а затем расширились и расширили свою деятельность до нескольких производственных линий, на которых одновременно работает 12-18 T2. Почему это важно? Когда вы выбираете T2, вы выбираете будущее, в котором вы можете масштабироваться.
100% водонепроницаемость.
Мы упоминали, что он водонепроницаем? Фактически, это единственный триммер на планете, который защищает вашу машину от повреждений, вызванных водой или влагой.Рассмотрим эту встроенную защиту инвестиций. Еще одна причина выбрать Twister T2.
ПОСМОТРЕТЬ T2 НА YouTube
ВЫСОКОЕ ПРОИЗВОДСТВО НИЗКОЕ ОБСЛУЖИВАНИЕ
Согласованный
Стабильная производительность означает стабильную прибыль. Добавьте конвейеры для еще большей выгоды.
Недорогое владение
Разработан и изготовлен из нержавеющей стали и анодированного алюминия. Нет необходимости постоянно менять детали из-за сколов или отслаивания краски.
Минимальное время простоя
Водонепроницаемые электрические и двигатели позволяют проводить полную промывку под высоким давлением; Полностью удаляется для полной очистки менее чем за 10 секунд без инструментов.
Прецизионная обрезка
T2 оснащен технологией QuickClick, обеспечивающей сверхбыструю и чрезвычайно точную регулировку лезвия до 0,0009 / 0,023 мм за щелчок!
Преимущества азотирования …
- Закаленная сталь
- Повышенная прочность на разрыв
- Повышенная коррозионная стойкость
- Повышенная износостойкость
- Меньше износ лезвий
- Меньше отложений на лезвиях
- Лезвия остаются острыми дольше
- Материалы разрешены для контакта с пищевыми продуктами
Коррозионно-стойкие азотированные лезвия [видео]
Сохраните свою ценную отделку
Революция в сборе и сохранении трима для Twister T2, модернизированный вариант Trim Saver ™ использует технологию радиального циклона для создания воздушного потока из вакуума, эффективно обходя крыльчатку, чтобы обеспечить лучший в своем классе трим, готовый к извлечению.
TWISTER TRIM SAVER ™
ЦИКЛОНИЧЕСКИЙ ДИЗАЙН
Впервые в отрасли, Trim Saver ™ использует технологию радиального циклона для сбора ценной обрезки, эффективно обходя крыльчатку, используемую в традиционных вакуумных системах сбора обрезков.
Вы просили, мы слушали. Лучшие триммеры на планете теперь могут работать всухую с помощью нашей новой технологии DRY Soft Tumble.
ПРОСТОТА ЭКСПЛУАТАЦИИ
Trim Saver ™ действительно работает по принципу «включай и работай». Просто подсоедините вакуумный шланг к триммеру и нажмите выключатель питания.Это так просто.
ЛУЧШАЯ ОТДЕЛКА ОТРАСЛИ
Независимо от того, используется ли ВЛАЖНЫЙ или СУХОЙ, Trim Saver ™ обеспечивает максимальную эффективность отделения обрезков, собирая более 99% вашего продукта в легко опорожняемом сборном бункере под циклоном.
ЛЕГКО ЧИСТИТЬ
Разработанный с нуля для облегчения разборки и очистки, Trim Saver ™ поставляется со съемным «контейнером для сбора обрезков», позволяющим быстро протирать в течение 2 минут между сеансами обрезки.
ОТДЕЛКА ВЛАЖНАЯ ИЛИ СУХАЯ
Обрезка с широким диапазоном влажности, от 11% до полностью увлажненного цветка.
НЕТ ОСТАНОВКИ
Наши медицинские триммеры для цветов рассчитаны на непрерывный поток и непрерывную работу.
САМАЯ НИЗКАЯ ОБРАБОТКА
Зачем переворачивать нежные цветы 20 минут, если 30 секунд будет достаточно?
ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ ПРОДУКТА
ТРИММЕР TWISTER T2
| Входное напряжение | 120 В |
| Текущий расход | 14,8А |
| Двигатель ножа | 1HP, с вентиляторным охлаждением, нержавеющая сталь |
| Тумблерный двигатель | 1 / 30HP, с эпоксидным покрытием, водонепроницаемость |
| Высота обрезки | 0.040 дюймов / 1/25 / 1 мм |
| Масса | 215 фунтов / 97,5 кг |
| Масса брутто (в сборе с листосборником) | 420 фунтов / 190 кг |
| Длина | 42,5 дюйма / 107,9 см |
| Ширина с колесами | 27,5 дюйма / 69,9 см |
| Ширина без колес | 20,5 дюйма / 52,2 см |
| Высота | 32.5 дюймов / 82 см |
| Высота без колес | 30,5 дюйма / 76 см |
| Требуется автоматический выключатель | 20А, 120В |
| Отвод | 6 дюймов / 152 мм |
TWISTER T2 ЛИСТОВОЕ КОЛЛЕКТОР
| Входное напряжение | 230 В |
| Двигатель | 2HP, одобрено UL и CSA, с вентиляторным охлаждением |
| Ампер (рабочий) | 9A |
| А (пуск) | 45A |
| Скорость двигателя | 3450 об / мин |
| Объем воздушного потока | 1700 куб. Футов в минуту |
| Статическое давление | 10.5 дюймов h30 |
| Рабочее колесо | 12 дюймов / 305 мм, сбалансированный алюминий |
| Впуск | 6 дюймов / 153 мм |
| Высота | 36 дюймов / 91,5 см |
| Масса | 123 фунта / 56 кг |
| Требуется автоматический выключатель | 15 А, 240 В |
| Разъем (штекер) | 15A 250 В (NEMA 6-15) |
| Уровень шума | 80 дБ |
| с предохранителем | Есть |
| Фильтр-мешок | 1 — 80 мкм |
| Сумка для сбора | 3–6 мил |
| Требования к генератору | 10,000 Вт |
| Калибр удлинителя | 12AWG МИНИМАЛЬНАЯ длина удлинительного шнура: 25 дюймов / 7.6 м МАКСИМУМ *** Несоблюдение этих мин. требования могут нанести ущерб |
| Запасные части | Зажимы для мешков, фильтровальные мешки, сборные мешки |
| Техническое обслуживание | Часто очищайте корпус крыльчатки. Заменить фильтр ba … |
Мы вас вернем
Более десяти лет лидируя в отрасли, мы гарантируем качество и мастерство изготовления нашего оборудования и предлагаем техническую поддержку высшего уровня, доступную 12 часов в день, 7 дней в неделю.
(1) He, J., Li, G., Chen, J., Lin, J. , Цзэн, К., Чен, Дж., Дэн, Дж. И Се, П., Длительное воздействие малых доз микроцистина вызывает у мышей неалкогольный стеатогепатит: исследование системной токсикологии. Архив токсикологии : (2016) 1-16. (2) Chen L, Chen J * , Zhang X, Xie P *. Обзор репродуктивной токсичности микроцистинов. Журнал опасных материалов . 301 (2016): 381-99. (3) Yu D, Xie P, Zeng C, Xie L, Chen J * . Эксперименты в вольерах in situ по возникновению, развитию и уменьшению черного налета и динамике связанных с ним соединений вкуса и запаха. Экологическая инженерия 87 (2016): 246-53. (4) Ван В., Шен Х, Ши П, Чен Дж, Ni L, Се П *. Экспериментальные доказательства роли гетеротрофных бактерий в формировании колоний Microcystis. Журнал прикладной психологии 28 (2016): 1111-23. (5) Hu Y, Chen J , Fan H, Xie P *, He J. Обзор нейротоксичности микроцистинов. Науки об окружающей среде и исследованиях загрязнения (2016): 1-9. (6) Цай Й, Чжан Ц., Хао Л., Чен Дж. , Се П, Чен З *. Систематическая идентификация семи генов рибосомных белков у пестрого толстолобика и их экспрессия в ответ на микроцистин-LR. Журнал токсикологических наук 41 (2016): 293-302. (7) Chen J, Xie P, Lin J, He J, Zeng C, Chen J *. Влияние микроцистина-LR на микрофлору кишечника в различных областях кишечника мышей. Журнал токсикологических наук 40 (2015): 485-494. (8) Li SC, Chen J *, Xie P, Guo XC, Fan HH, Yu DZ, Zeng C, Chen L. Роль пути детоксикации глутатиона в вызванной MCLR гепатотоксичности у крыс SD . Environment Toxi cology 30 (2015): 1470-1480. (9) Guo XC, Chen L, Chen J *, Xie P *, Li SC, He J, Li W, Fan HH, Yu DZ, Zeng C. Количественная оценка детоксикации гепатотоксичного микроцистина-LR через путь глутатиона (GSH) у крыс SD. Науки об окружающей среде и исследованиях загрязнения 22 (2015): 19273-19284. (10) Лин Дж., Чен Дж. , Хэ Дж, Чен Дж., Ян Цюй, Чжоу Дж. З., Се П *. Влияние микроцистина-LR на профиль функциональных генов бактерий и грибов в кишечнике крыс. Toxicon 96 (2015): 50-56. (11) Guo XC, Xie P *, Chen J * , Tuo X, Deng XW, Li SC, Yu DH, Zeng C. Одновременное количественное определение микроцистина-LR и его метаболитов глутатиона в печени крыс с помощью жидкостная хроматография-тандемная масс-спектрометрия. J. Chromatogr. Б. 963 (2014): 54-61. (12) Li W, Xie P, Chen J * , He J, Guo XC, Yu DZ, Chen L. Метод количественной жидкостной хроматографии и тандемной масс-спектрометрии для определения микроцистина-RR и его глутатиона и цистеина конъюгаты в плазме и желчи рыб. J. Chromatogr. Б. 963 (2014): 113-118. (13) Li W, Chen J * , Xie P, He J, Guo XC, Tuo X, Zhang W, Wu LY.Быстрое преобразование и обратимая конъюгация глутатиона, детоксикация микроцистинов у пестрого амура ( Aristichthys nobilis ). Aquat. Toxicol. 147 (2014): 18-25. (14) Chen L, Li SC, Guo XC, Xie P, Chen J *. Роль GSH в индуцированном микроцистином апоптозе в печени крыс: участие в окислительном стрессе и NF-kB. Экологическая токсикология , (2014) DOI: 10.1002 / tox.22068 (15) Chen L, Zhang X, Chen J , Zhang XZ, Fan HH, Li SC, Xie P *.NF-каппа B играет ключевую роль в пролиферации и апоптозе клеток HeLa, индуцированных микроцистином-RR. Toxicon 87 (2014): 120-130. (16) Fan HH, Cai Y, Xie P *, Xiao WH *, Chen J , Ji W, Zhao SJ. Microcystin-LR стабилизирует белок c-myc, ингибируя протеинфосфатазу 2A в клетках HEK293. Токсикология 319 (2014): 69-74. (17) Панг XH, Шен Х *, Ниу Y, Сун XX, Чен Дж, Се П *.Растворенный органический углерод и взаимосвязь с составом сообщества бактериопланктона в 3 озерных районах озера Тайху, Китай. Банка. J. Microbiol. 60 (2014): 669-680. (18) Tuo X, Chen J, Zhao SJ, Xie P *. Химический протеомный анализ потенциальных токсикологических механизмов микроцистина-RR в печени зебры ( Danio rerio ). Экологическая токсикология , (2014) DOI: 10.1002 / tox.22128 (19) Ma ZM, Niu Y, Xie P, Chen J * , Tao M, Deng XW.Соединения с неприятным запахом от разлагающихся цветов цианобактерий озера Тайху. J. Environ. Научно-Китай. 25 (2013): 495-501 (20) Zhang J, Xie P *, Tao M, Guo LG *, Chen J , Li L, Zhang XZ, Zhang L. Влияние хищничества рыб и цианобактерии на размерную структуру зоопланктона 96 субтропических озер. PLoS ONE 8 (2013 г.): e76378. (21) Чжан Д., Дэн XW, Се П *, Чен Дж. , Гуо LG.Оценка риска микроцистинов толстолобика ( Hypophthalmichthys molitrix ) из восьми эвтрофных озер в Китае. Food Chem. 140 (2013): 17-21. (22) Чжан В., Лян Г.Д., Ву ЛИ, Туо Х, Ван В.Дж., Чен Дж. , Се П *. Почему млекопитающие более восприимчивы к гепатотоксическим микроцистинам, чем рыбы: свидетельства связывания с белками плазмы и альбумина посредством равновесного диализа. Экотоксикология 22 (2013): 1012-1019. (23) Wu LY, Wang Q, Tao M, Chen J , Ma ZM, Xie P *. Предварительное исследование распределения и накопления метаболитов GSH / Cys гепатотоксических микроцистинов-RR у карпа из озера с затяжным цветением цианобактерий (озеро Тайху, Китай). B. Environ. Contam. Tox. 90 (2013): 382-386. (24) Ма ZM, Xie P *, Chen J , Niu Y, Tao M, Qi M, Zhang W, Deng XW. Microcystis цветет, влияя на концентрацию летучих органических соединений в озере Тайху. Fresen. Environ. Бык. 22 (2013): 95-102. (25) He J, Chen J * , Wu LY, Li GY, Xie P *. Метаболический ответ на пероральное воздействие макроцистина-LR у крыс по данным метабономического исследования на основе ЯМР. J. Proteom. Res. 11 (2012): 5934-5946. (26) He J, Chen J * , Xie P, Zhang DW, Li GY, Wu LY, Zhang W, Guo XC, Li S. C. Количественная оценка детоксикации гепатотоксичных микроцистинов с помощью глутатиона и цистеина пути у пестрого амура, поедающего цианобактерии. Aquat. Toxicol. 116-117 (2012): 61-68. (27) Deng XW, Xie P *, Qi M, Liang GD, Chen J * , Ma ZM, Jiang Y. Устройство для продувки и улавливания с помощью микроволнового излучения в сочетании с газовой хроматографией и масс-спектрометрией для определение пяти преобладающих запахов в отложениях, тканях рыб и клетках водорослей. J. Chromatogr. А 1219 (2012): 75-82. (28) Чжан DW, Ян Цюй, Се П, Дэн XW, Чен Дж * , Дай М.Путь детоксикации MC-LR в печени пестрого амура ( Aristichthys nobilis ): полевые и лабораторные исследования. Экотоксикология 21 (2012): 244-252. (29) Цай Й, Ли Хай, Хао Л, Ли Ги, Се П, Чен Дж, * . Идентификация гена cda у пестрого толстолобика и его экспрессия в ответ на микроцистин-LR. Ecotoxicol. Environ. Безопасность 79 (2012): 206-213. (30) Tao M, Xie P *, Chen J , Qin BQ, Zhang DW, Niu Y, Zhang M, Wang Q, Wu LY.Использование обобщенной аддитивной модели для исследования ключевых абиотических факторов, влияющих на клеточные квоты микроцистина в районах с сильным цветением озера Тайху. PLoS ONE 7 (2012): e32020. (31) Qi M, Chen J , Sun XX, Deng XW, Niu Y, Xie P *. Разработка моделей для прогнозирования преобладающих соединений вкуса и запаха в озере Тайху, Китай. PLoS ONE 7 (2012): e51976. (32) Li HY, Cai Y, Xie P *, Chen J , Hao L, Li G, Xiong Q.Идентификация и профиль экспрессии Id1 у пестрого амура в ответ на микроцистин-LR. Окружающая среда. Toxicol. Phar. 34 (2012): 324-333. (33) Ян В, Чжоу YX, Ян Дж, Ху DJ, Дин Х, Ван Дж. Х, Чен Дж *, Ли ГЙ *. Воздействие микроцистина-LR через воду изменяет уровни гормонов щитовидной железы и транскрипцию генов в оси гипоталамус-гипофиз-щитовидная железа у личинок рыбок данио. Chemosphere (2012) 87: 1301-1307. (34) Liang GD , Xie P *, Chen J * , Yu T.Сравнительные исследования зависимости DOW микроцистина-RR и -LR от pH с использованием ЖХ-МС. TheScientificWorldJournal 11 (2011): 20-26 (35) Niu Y, Shen H, Chen J , Xie P *, Yang X, Tao M, Ma ZM, Qi M. Последовательность сообществ фитопланктона формирование состава сообщества бактериопланктона в озере Тайху, Китай. Вода Рез. 45 (2011): 4169-4182. (36) Li GY, Chen J , Xie P *, Jiang Y, Wu LY, Zhang XZ.Профилирование экспрессии белка в эмбрионах рыбок данио ( Danio rerio ), подвергшихся воздействию микроцистина-LR. Протеомика 11 (2011): 2003-2018. (37) Deng XW, Liang GD, Chen J , Qi M, Xie P *. Одновременное определение восьми общих запахов в естественном водоеме с использованием автоматической продувки и ловушки в сочетании с газовой хроматографией с масс-спектрометрией. J. Chromatogr. А 1218 (2011): 3791-3798. (38) Wu LY, Xie P *, Chen J * , Zhang DW, Liang GD. Разработка и валидация метода жидкостной хроматографии и масс-спектрометрии для одновременного количественного определения микроцистина-RR и его метаболитов в печени рыб. J. Chromatogr. А 1217 (2010): 1455-1462. (39) Chen J , Xie P *, Ma ZM, Niu Y, Tao M, Deng XW, Wang Q. и абиотические параметры в заливе Гунху озера Тайху, Китай. Sci. Total Environ. 409 (2010): 314-325. (40) Zhang DW, Xie P *, Chen J * . Влияние температуры на стабильность микроцистинов в мышцах рыб и ее последствия для безопасности пищевых продуктов. Бык. Environ. Contam. Toxicol. 84 (2010): 202-207. (41) Ван Цюй, Ниу И, Се П *, Чен Дж, Ма ЗМ, Тао М, Ци М, Ву Лай, Гуо LG. Факторы, влияющие на временные и пространственные вариации микроцистинов в заливе Гонгу озера Тайху, с потенциальным риском заражения микроцистином для здоровья человека. The Scientific World Journal 10 (2010): 1795–1809. (42) Li GY, Xie P *, Li HY, Chen J , Hao L, Xiong Q. Количественное профилирование экспрессии мРНК генов суперсемейства глутатион-S-трансферазы в различных тканях толстоголового амура ( Aristichthys nobilis ). J. Biochem. Мол. Токсично. 24 (2010): 250-259. (43) Chen J, Xie P *, Li L, Xu J. Первое выявление гепатотоксических микроцистинов в сыворотке хронически подверженной воздействию человеческой популяции вместе с указанием гепатоцеллюлярного повреждения. Toxicol. Sci. 108 (2009): 81-89. (44) Чен Дж, Чжан Д.В., Се П *, Ма З.М. Одновременное определение контаминации микроцистином у различных позвоночных (рыб, черепах, уток и водоплавающих птиц) из большого эвтрофного китайского озера Тайху с токсичным цветением Microcystis . Sci. Всего Окружающая среда 925 18. 407 (2009): 3317-3322. (45) Чжан D, Xie P * , Liu Y, Chen J , Wen Z.Пространственные и временные вариации микроцистинов в гепатопанкреасе пресноводной улитки из озера Тайху. Ecotoxicol. Environ. Безопасность 72 (2009): 466-472. (46) Чжан DW , Xie P *, Chen J, Dai M, Qiu T, Liu YQ, Liang GD. Определение микроцистина-LR и его метаболитов у улитки ( Bellamya aeruginosa ), креветок ( Macrobrachium nipponensis ) и толстолобика ( Hypophthalmichthys molitrix ) из озера Тайху, Китай. Chemosphere 7 (2009): 974-981. (47) Li H, Xie P *, Zhang DW, Chen J . Первое исследование влияния микроцистина-RR на профили экспрессии генов антиоксидантных ферментов и белка теплового шока-70 у Synechocystis sp. PCC6803. Toxicon 53 (2009): 595-601. (48) Chen J , Xie P *. Накопление гепатотоксичных микроцистинов в пресноводных мидиях, личинках водных насекомых и олигохатах в большом мелководном эвтрофном озере (озеро Чаоху) в субтропическом Китае. Fresen. Environ. Бык. 17 (2008): 849-854. (49) Дай М., Се П *, Лян Г.Д., Чен Дж. , Лей ХХ. Одновременное определение микроцистина-LR и его конъюгата глутатиона в тканях рыб методом жидкостной хроматографии-тандемной масс-спектрометрии. J. Chromatogr. Б. 862 (2008): 43-50. (50) Lei HH, Xie P *, Chen J , Liang GD, Dai M, Zhang XZ. Распределение токсинов в различных тканях карася при внутрибрюшинном введении гепатотоксических микроцистинов. Окружающая среда. Toxicol. Chem. 27 (2008): 1167-1174. (51) Lei HH, Xie P *, Chen, J , Liang GD, Yu T, Jiang Y. Распределение тканей и очистка выделенных гепатотоксических цианотоксиновых микроцистинов у карася ( Carassius carassius , введенных внутрибрюшинно) в сублетальной дозе. The ScienceWorldJournal 8 (2008): 713-719. (52) Ван Кью, Се П *, Чен Дж. , Лян Г.Д.Распределение микроцистинов в различных органах (сердце, печень, кишечник, гонады, мозг, почки и легкие) крысы Wistar при внутривенной инъекции. Toxicon 52 (2008): 721-727. (53) Чен Дж, Се П *. Накопление микроцистина у пресноводных двустворчатых моллюсков из озера Тайху, Китай, и потенциальный риск для потребления человеком. Окружающая среда. Toxicol. Chem. 26 (2007): 1066-1073. (54) Чен Дж. , Се П *, Чжан Д., Лей Х.Исследования in situ закономерностей распределения и динамики микроцистинов у биоманипуляционной рыбы — пестрого амура ( Aristichthys nobilis ). Окружающая среда. Загрязнение. 147 (2007): 150-157. (55) Zhang DW, Xie P *, Liu YQ, Chen J , Liang GD. Биоаккумуляция гепатотоксичных микроцистинов в различных органах пресноводной улитки из субтропического китайского озера Тайху с плотным токсичным цветением Microcystis . Окружающая среда.Toxicol. Chem. 26 (2007): 171-176. (56) Li L, Xie P *, Chen J . Биохимические и ультраструктурные изменения печени и почек толстолобика-фитопланктона, питающегося ядовитым цветком Microcystis в озере Тайху, Китай. Toxicon 49 (2007): 1042-1053. (57) Chen J , Xie P *, Zhang D, Ke ZX, Yang H. Исследования in situ биоаккумуляции микроцистинов у фитопланкоядных толстолобиков ( Hypophthalmichthys molitrix ), разводимых в озере Тайху с плотной токсичный Microcystis цветет. Аквакультура 261 (2006): 1026-1038. (58) Чжан X, Се П *, Хао Л, Гуо NC, Гонг Ю.Г., Ху XL, Чен Дж. , Лян Г.Д. Воздействие фитопланкоядных толстолобиков на планктон и гепатотоксические микроцистины в эксперименте в вольере в эвтрофном озере Шичахай в Пекине. Аквакультура 257 (2006): 173-186. (59) Chen J , Xie P *, Guo LG, Zheng L, Ni LY. Распределение в тканях и сезонная динамика гепатотоксичных микроцистинов -LR и -RR у пресноводной улитки ( Bellamya aeruginosa ) из большого мелководного эвтрофного озера в субтропическом Китае. Окружающая среда. Загрязнение. 134 (2005): 423-430. (60) Чен Дж, Се П *. Распределение в тканях и сезонная динамика гепатотоксичных микроцистинов -LR и -RR у двух пресноводных креветок, Palaemon modestus и Macrobrachium nipponensis из большого мелководного эвтрофного озера в субтропическом Китае. Toxicon 45 (2005): 615-625. (61) Chen J , Xie P *. Сезонная динамика гепатотоксических микроцистинов в различных органах четырех пресноводных двустворчатых моллюсков из большого эвтрофного озера Тайху в субтропическом Китае и риск для потребления человеком. Environ Toxicol. 20 (2005): 572-584. (62) Li L, Xie P *, Chen J . Исследования in vivo накопления токсинов в печени и ультраструктурных изменений гепатоцитов фитопланкоядного пестрого карпа, вводимого внутрибрюшинно с экстрагированными микроцистинами. |