Краснокнижный вид | ООПТ России
Главная
Краснокнижный вид
Классификация: Основные группы > Algae and other protists (Водоросли и другие простейшие) > Ciliophora (Инфузории) > Oligohymenophorea > Peniculida > Stokesiidae > Stokesia
| Таксон | Stokesia vernalis |
Правовой статус
| Документ | Приложение | Дата | Номер таксона | Указан как | Статус | Категория | Дополнительно |
|---|---|---|---|---|---|---|---|
| Об утверждении перечней видов беспозвоночных животных, высших растений, грибов, лишайников и водорослей, занесенных в Красную книгу Нижегородской области | 1999-03-15 | 1 | добавлен | Д |
Описания
| Издание |
(2003) Красная книга Нижегородской области. Том 1. Животные. Нижний Новгород 380 Том 1. Животные. Нижний Новгород 380 |
| Таксон указан как | Stokesia vernalis |
| Категория | Д: Неопределенный вид, для которого нет достаточных данных, чтобы конкретизировать его статус. Один из двух видов рода. |
| Морфологическое описание |
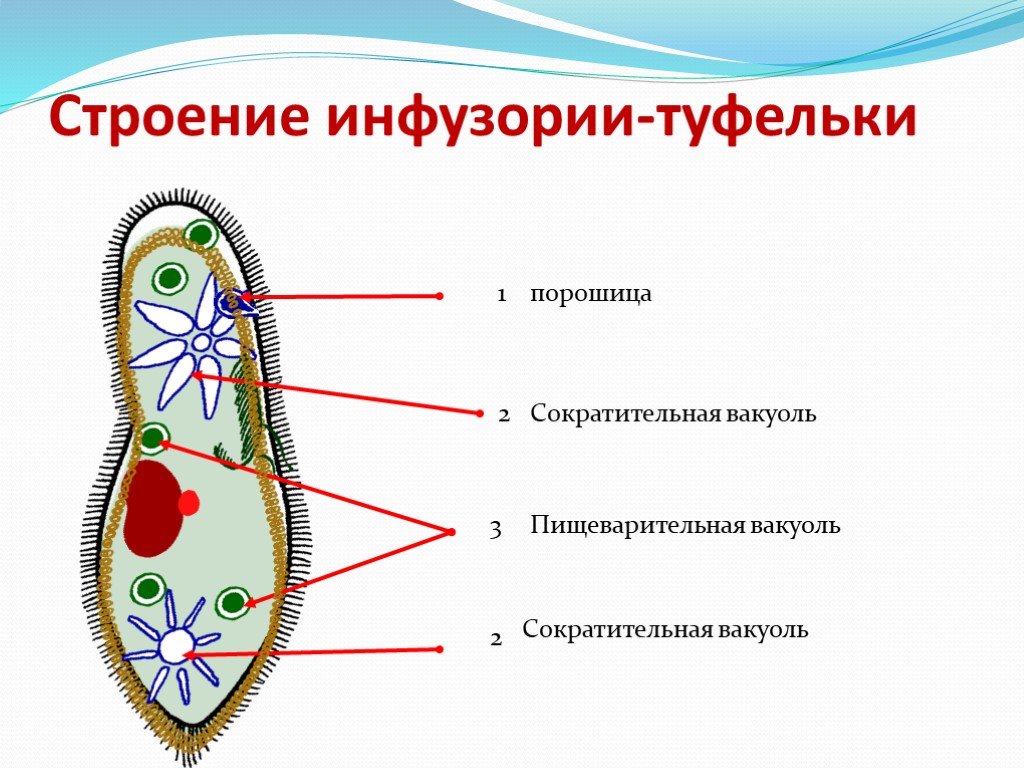
Достаточно крупная инфузория, имеющая форму асимметричного конуса. Размеры стокезии варьируют, наиболее крупными оказываются весенние формы: диаметр клетки 175–200 мкм, длина (высота конуса) 100–150 мкм; летние формы несколько мельче – 120 и 80 мкм, соответственно. Клеточный рот расположен на плоской брюшной стороне, соответствующей основанию конуса, в виде изогнутой щели и способен округляться во время приема пищи. Величина рта составляет 1/3–1/2 длины клетки. Ресничный предротовой аппарат представлен мерцательной мембраной, тремя рядами мембранелл со свободными несклеенными ресничками и погружен внутрь тела. Ресничный покров клетки хорошо развит. Стокезия двигается плавно, покачиваясь относительно вершины конуса. Макронуклеус овальный, расположен в центре клетки, сократительная вакуоль находится около вершины конуса. Инфузория имеет мощный слой трихоцист. Ресничный покров клетки хорошо развит. Стокезия двигается плавно, покачиваясь относительно вершины конуса. Макронуклеус овальный, расположен в центре клетки, сократительная вакуоль находится около вершины конуса. Инфузория имеет мощный слой трихоцист. |
| Распространение | Известна в водоемах Западной Европы, Северной Америки. Описана в составе планктонных сообществ крупных холодноводных озер европейской части России и Сибири (Онежское, Ладожское, Отрадное, Байкал), а также верхневолжских водохранилищ и р. Волги. В центральной части Европейской России зарегистрирована в планктоне оз. Светлояр, Пустынских озер Нижегородской области, оз. Открытого и оз. Сосновского Чувашии. Более южное распространение ограничивается указанием стокезии в составе придонного комплекса инфузорий в оз. Кагул (Молдавия). |
| Образ жизни |
Места обитания. В составе планктонного сообщества пелагиали слабо проточных озер, водохранилищ, в реках как результат сноса планктона из расположенных выше по течению водоемов. Особенности биологии. Характеризуется как холодолюбивый стенотермный вид с оптимумом температуры в пределах 0–10°С. Питается бактериями и водорослями (Stephanodiscus hantzchii Grun.), часто характеризуется как активный фитофаг, по экспериментальным данным способна поедать детрит и поглощать растворенное органическое вещество. В свою очередь служит пищей для коловраток рода Asplanchna, хищных инфузорий, например, Bursaria truncatella O. F. M.. ll. В весеннем планктоне встречается в смешанных популяциях вместе с Amphileptus trachelioidesZach. Предпочитает чистые насыщенные кислородом воды (90–100% насыщения) с низким содержанием органических веществ (13–18 мг O 2/л) при рН 7,5–8,2 и является индикатором олиго βезосапробной зон водоемов. Летние более мелкие формы содержат зоохлорелл и отличаются более высоким темпом роста и, соответственно, небольшим временем генерации: 12–23 часов. Особенности биологии. Характеризуется как холодолюбивый стенотермный вид с оптимумом температуры в пределах 0–10°С. Питается бактериями и водорослями (Stephanodiscus hantzchii Grun.), часто характеризуется как активный фитофаг, по экспериментальным данным способна поедать детрит и поглощать растворенное органическое вещество. В свою очередь служит пищей для коловраток рода Asplanchna, хищных инфузорий, например, Bursaria truncatella O. F. M.. ll. В весеннем планктоне встречается в смешанных популяциях вместе с Amphileptus trachelioidesZach. Предпочитает чистые насыщенные кислородом воды (90–100% насыщения) с низким содержанием органических веществ (13–18 мг O 2/л) при рН 7,5–8,2 и является индикатором олиго βезосапробной зон водоемов. Летние более мелкие формы содержат зоохлорелл и отличаются более высоким темпом роста и, соответственно, небольшим временем генерации: 12–23 часов. |
| Численность |
Сведения малочисленны, тенденции изменения оценить трудно. В водоемах Нижегородской области и Чувашии является малочисленным видом инфузорного планктона: средние показатели численности составляют примерно 0,4 млн. экз./м3. Наибольшее количество инфузорий (1,2 млн. экз./м В водоемах Нижегородской области и Чувашии является малочисленным видом инфузорного планктона: средние показатели численности составляют примерно 0,4 млн. экз./м3. Наибольшее количество инфузорий (1,2 млн. экз./м |
| Лимитирующие факторы |
Особенности биологии вида, связанные с повышенной требовательностью к условиям обитания (содержание кислорода, РОВ, скорость течения и пр. ). Эвтрофикация водоемов и нарушение гидрологического режима в результате деятельности человека. ). Эвтрофикация водоемов и нарушение гидрологического режима в результате деятельности человека. |
| Охранные меры | Принятые меры охраны. Местообитания охраняются в составе 2 ПП: «Озеро Светлояр» и «Пустынские озера». Необходимые меры охраны. Реализация мер охраны олиготрофных озер, обеспечивающих сохранение гидрологического режима данных водоемов и запрещающих загрязнение воды, что способствует сохранению данного комплекса инфузорий. |
| Ссылки | 1. Арсланова, 1983. 2. Арсланова, 1989. 3. Гаевская, 1949. 4. Лазарева, 1980. 5. Мамаева, 1979. 6. Чорик, 1967. 7. Шень Юнь Фень, 1960. 8. Noland, 1959. 9. Orr, 1954. 10. Pace, 1982. |
| Составители | Т. П. Станковска |
Инопланетянин с планеты Земля / Хабр
Spirostomum — это род свободноживущих инфузорий, относящихся к классу Heterotrichea. Встретить их можно, как в солёной, так и пресной воде. Несмотря на то, что они одноклеточные, представители некоторых из них могут вырастать в длину до 4 мм (0, 16 дюйма). Только посмотрите на этих удивительных симпатяг в сравнении с рачком Дафнией — многоклеточным организмом! К слову, данных животных, я выловил в стоячей воде сливного канала в Белоруссии, своей любимой и родной стране.
Встретить их можно, как в солёной, так и пресной воде. Несмотря на то, что они одноклеточные, представители некоторых из них могут вырастать в длину до 4 мм (0, 16 дюйма). Только посмотрите на этих удивительных симпатяг в сравнении с рачком Дафнией — многоклеточным организмом! К слову, данных животных, я выловил в стоячей воде сливного канала в Белоруссии, своей любимой и родной стране.
Описание и характеристика рода.
Клетка инфузории имеет червеобразную форму. В поперечном сечении клетка в основном овальной, или кругло-подобной формы. Хвостовые концы могут быть уплощены. Поэтому среди инфузорий, виды этого рода узнаются сразу, особенно, если рядом плавают другие причудливые представители этого типа. В моём случае, помимо наших червеобразных лапочек, плавала инфузория, похожая на музыкальный инструмент. Настолько она причудлива. Эта была инфузория трубач, которую вы увидите в моём видео ниже.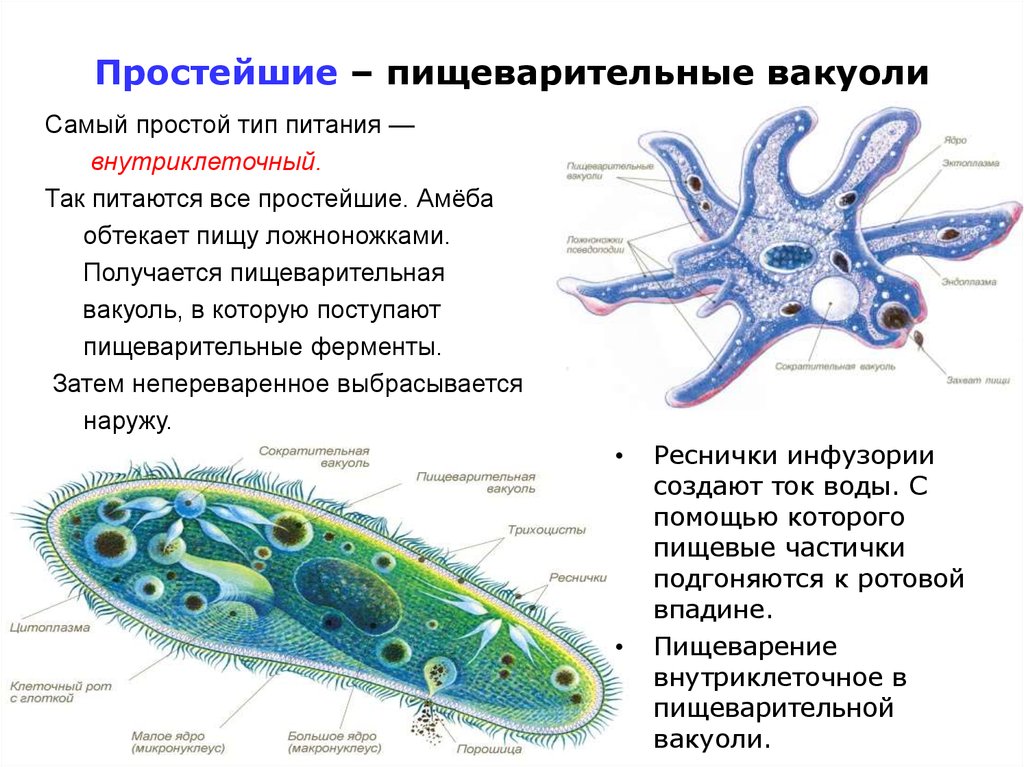 Внешне она напоминает музыкальную трубу, поросшую странными волосами (ресничками).
Внешне она напоминает музыкальную трубу, поросшую странными волосами (ресничками).
Но речь сейчас не о трубачах, а особенностях строения инфузории Спиростомиум, доминирующей в том сливном канале. И, пожалуй, самая интересная особенность их заключается в том, что их пищеварительная, а точнее экскреторная вакуоль может достигать настолько больших размеров, что способна занимать весь «хвостовой» конец клетки. Экскреторная вакуоль инфузории представляет собой гигантскую гранулу, которая, порой сливаясь с цитоплазматической мембраной, изливает вредные продукты обмена во внешнюю среду. Чтобы не умереть от своих собственных избытков вредных продуктов обмена, а также от недостатка пищи, наши инфузории должны уметь перемещаться.
Пример мембранеллДля этого у наших миниатюрных друзей есть короткие реснички на «теле» клетки, которые расположены продольными рядами вдоль «тела». Питаются инфузории с помощью мембранелл, структур, образующих область возле рта, называемой адрональной зоной. Сам рот находится на перистоме — возвышенной поверхности тела. Так вот, мембранеллы инфузорий этого рода очень интересны. Они окаймляют перистом и могут занимать от 1/4 до 2/3 длины клетки в зависимости от вида.
Сам рот находится на перистоме — возвышенной поверхности тела. Так вот, мембранеллы инфузорий этого рода очень интересны. Они окаймляют перистом и могут занимать от 1/4 до 2/3 длины клетки в зависимости от вида.
Чем же ещё интересны эти инфузории?
ЭндосимбиозИнтересны эти инфузории и тем, что встречаются виды, имеющие в своих клетках эндосимбионтов. Так, например, S. semivirescens в микроскоп кажется ярко-зеленым из-за того, что он упакован эндосимбиотическими водорослями рода Chlorella.
РазмножениеСогласно симбиогенетической теории пластиды, как и митохондрии, произошли в результате «захвата» древней цианобактерии предшественником эукариотической «хозяйской» клетки. Вполне возможно, что учёные опирались в построении этой теории и на подобные организмы, которые заключают в себя одноклеточные водоросли
Стоит упомянуть и размножение. Спиростомум размножается путём простого бинарного деления, которое в некоторых случаях может следовать сразу за конъюгацией.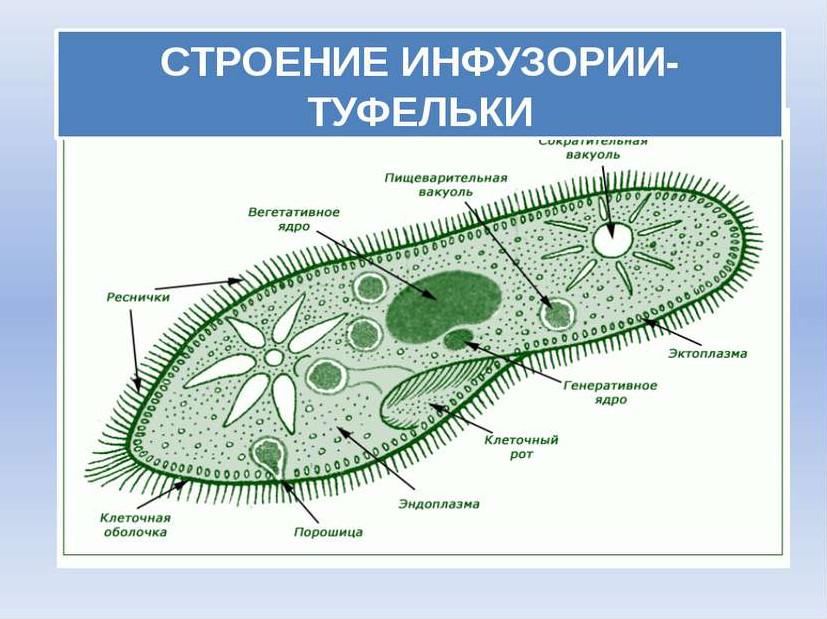 Конъюгацией у инфузорий называют обмен половыми ядрами (микронуклеусами) с последующим их попарным слиянием в синкарион. Синкарион — ядро дробления, образующееся в результате слияния половых ядер. Впоследствии синкарион делится с образованием новых половых и вегетативных ядер, которые затем перемещаются из взрослого клона в дочернего. Отличие от конъюгации, например, водорослей, заключается в том, что инфузории не образуют гамет, которые потом образуют зиготу при слиянии. Кроме того, конъюгация инфузорий не сопровождается привычным размножением, то есть увеличением числа клеток, поэтому конъюгация у инфузорий является типичным примером полового процесса без привычного размножения. Водоросли при конъюгации после образования зиготы образуют зигоспору, которая перезимовывает и весной делится мейозом, давая 4 гаплоидные споры. Даже при хологамии, аналоге конъюгации у одноклеточных, образуется зигота, которая даёт 4 гаплоидные споры, а инфузория против таких выкрутасов!
Конъюгацией у инфузорий называют обмен половыми ядрами (микронуклеусами) с последующим их попарным слиянием в синкарион. Синкарион — ядро дробления, образующееся в результате слияния половых ядер. Впоследствии синкарион делится с образованием новых половых и вегетативных ядер, которые затем перемещаются из взрослого клона в дочернего. Отличие от конъюгации, например, водорослей, заключается в том, что инфузории не образуют гамет, которые потом образуют зиготу при слиянии. Кроме того, конъюгация инфузорий не сопровождается привычным размножением, то есть увеличением числа клеток, поэтому конъюгация у инфузорий является типичным примером полового процесса без привычного размножения. Водоросли при конъюгации после образования зиготы образуют зигоспору, которая перезимовывает и весной делится мейозом, давая 4 гаплоидные споры. Даже при хологамии, аналоге конъюгации у одноклеточных, образуется зигота, которая даёт 4 гаплоидные споры, а инфузория против таких выкрутасов!
Забавно наблюдать (видео выше), как это животное способно резко сокращать свои размеры и сжиматься, будто маленькая пружина. Такое ощущение, что на это уходит буквально всего нескольких миллисекунд! Это не стоп кадр, или монтаж, это реально она так сокращается. Помимо сжатия, эти инфузории способны увеличивать свои размеры в результате большого стресса, например, в высыхающей капле воды. Этот момент мне также удалось запечатлеть.
Такое ощущение, что на это уходит буквально всего нескольких миллисекунд! Это не стоп кадр, или монтаж, это реально она так сокращается. Помимо сжатия, эти инфузории способны увеличивать свои размеры в результате большого стресса, например, в высыхающей капле воды. Этот момент мне также удалось запечатлеть.
Есть ли какие-нибудь интересные научные исследования связанные с этими инфузориями?
Как я говорил выше, резкое сокращение размеров «тела», очень интересный момент и не только для меня. Ученые из Технологического института Джорджии сделали замечательное открытие. Биолог Саад Бхамла, получив грант на изучение Spirostomum ambiguum, обнаружил невероятную способность этой инфузории с огромной скоростью сокращать своё «тело». В течение доли секунды Spirostomum ambiguum сокращает свое «тело», уменьшая его длину более чем на 60%. Это проходит с ускорением в 200 м/с² и нагрузкой равной 14g. Прямо самый быстрый организм на планете, олимпийский чемпион микромира! Это достаточно интересно, поскольку человеческому глазу не под силу заметить постепенное сокращение инфузории.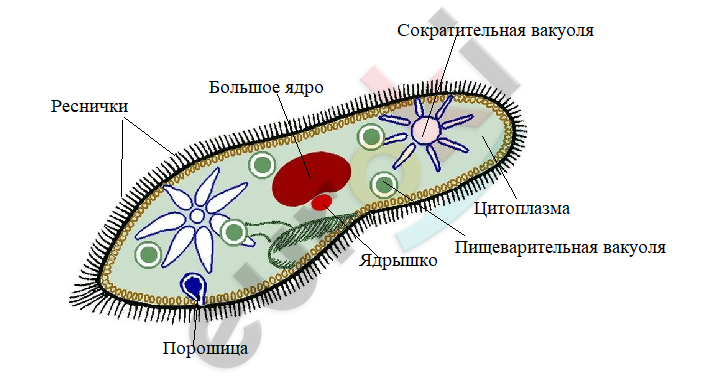
Этот механизм обеспечивает быстрое реагирование на внешние стимулы, что является ключом к выживанию. Плавающие организмы генерируют обильные потоки, которые сохраняются в водной среде и Спиростомум в ходе эволюции «научился» быстро отвечать на такой внешний сигнал. Биофизики назвали данное открытие «гидродинамическими триггерными волнами», которые распространяются — подобно цепной реакции — в сотни раз быстрее, чем скорость плавания. Исследование показало, что сокращение одной клетки (передатчик) генерирует дальние вихревые потоки, которые, в свою очередь, могут запускать соседние клетки (приемники). Результаты позволяют предположить, что такая сигнализация может способствовать организации совместно живущих сообществ на больших расстояниях и влиять на долгосрочное поведение через экспрессию генов. Так же при сокращениях выделяются токсины, синхронизированные выбросы которых, могут способствовать отпугиванию крупных хищников или обездвиживанию крупной добычи. Биологи предполагают, что схожий механизм регулирования поведения через гидродинамические триггерные волны есть и у других протистов.
Инфузория Спиростомум как индикатор загрязнения водной среды.
Представители типа протисты всё больше привлекают внимание исследователей, занимающихся поиском надежных индикаторов загрязнения водной среды. Имея достаточно большие размеры (позволяют рассматривать организмы под малым увеличением), Spirostomum ambiguum может стать идеальным кандидатом на роль организма для биоиндикации воздействия на окружающую среду химических токсикантов, γ-излучения и электромагнитного поля. Так польскими учёными из Медицинского университета Варшавы был разработан Спиротокс (Spirotox)- краткосрочный тест на острую токсичность с применением Spirostomum ambiguum (Grzegorz Nałecz-Jawecki, 2004). Впервые он был представлен на 6-м Международном симпозиуме по оценке токсичности в Берлине в 1993 году.
В течение 10 лет была проведена оценка чувствительности S. ambiguum ко многим классам токсикантов. Было установлено, что спиротокс очень чувствителен к тяжелым металлам, фунгицидам и фармацевтическим препаратам, используемым для лечения заболеваний нервной системы человека. С другой стороны, он был в целом менее чувствителен к простым органическим веществам, чем стандартные биопробы. Спиротокс также может использоваться для анализа «цветения» цианобактерий.
С другой стороны, он был в целом менее чувствителен к простым органическим веществам, чем стандартные биопробы. Спиротокс также может использоваться для анализа «цветения» цианобактерий.
Аналогичные исследования проводили и российские учёные в МГУ и Обнинском институте. Результаты этих двух исследований показывают перспективность использования Спиростомума в качестве незаменимой модели не только для выявления негативных факторов среды их обитания, но и для объяснения механизмов таких воздействий на одноклеточный и/или многоклеточный организм.
Вот такие они, инфузории, пойманные в моей родной Белоруссии!
Примечаниепольский микроскоп PZO MB30Для съёмки первых двух видео использовался мой личный польский микроскоп PZO MB30, который оснащён двумя родными объективами 5Х и 40Х и двумя от советского микроскопа на 10Х и 90Х. Окуляры родные на 10Х. В качестве камеры использовался Xiaomi Redmi Note 4
Eschenbach Typ 3455Для съёмки последующих двух видео использовался немецкий микроскоп Eschenbach Typ 3455 в родной комплектации. В качестве камеры выступал смартфон POCO X3 NFS. Дополнительной обработки видео не подвергалось.
В качестве камеры выступал смартфон POCO X3 NFS. Дополнительной обработки видео не подвергалось.
Материал брался стандартной ёмкостью на 120 мл в мелком, стоячем водоёме. Всем начинающим снимать подобные видео рекомендую начинать по этой методике. В дальнейшем можно переходить к более сложным методикам сбора материала.
P.S. Статья написана биологом Евгением Будько и опубликована в научном сообществе Фанерозой.
Источникиhttps://aspidisca.livejournal.com/37589.html
https://en.wikipedia.org/wiki/Spirostomum
https://pikabu.ru/story/bolshe_negepard_nazvano_samoe_byistroe_vmire_sushchestvo_6086766
https://zen.yandex.ru/media/zhivotniymir/samoe-bystroe-suscestvo-v-mire-600b261f1924cc0331468060
https://zen.yandex.ru/media/id/5b4c7dc488b68f00a9861d0d/naideno-samoe-bystroe-suscestvo-na-planete-5b6f7dc2955bc500a8b9995c
https://www.nature. com/articles/s41586-019-1387-9
com/articles/s41586-019-1387-9
https://cyberleninka.ru/article/n/infuzoriya-spirostoma-kak-indikator-zagryazneniya-vodnoy-sredy
https://pubmed.ncbi.nlm.nih.gov/15269908/
https://translate.yandex.ru/translate?lang=en-ru&url=https%3A%2F%2Fen.wikipedia.org%2Fwiki%2FMembrinella&view=c
https://ru.wikipedia.org/wiki/Хологамия
https://ru.wikipedia.org/wiki/Конъюгация_у_инфузорий
https://ru.wikipedia.org/wiki/Конъюгация_у_водорослей
Содержимое пищевых вакуолей у инфузории Balantidium jocularum (Balantididae), симбионта в кишечнике рыбы-хирурга Naso tonganus (Acanthuridae)
. 2006 г., июль-август; 53(4):269-74.
doi: 10.1111/j.1550-7408.2006.00101.x.
Джей Норман Грим 1
принадлежность
- 1 Факультет биологических наук, а/я 5640, Университет Северной Аризоны, Флагстафф, Аризона 86011, США.
 [email protected]
[email protected]
- PMID: 16872294
- DOI: 10.1111/j.1550-7408.2006.00101.x
Дж. Норман Грим. Дж Эукариот микробиол. 2006 июль-август.
. 2006 г., июль-август; 53(4):269-74.
doi: 10.1111/j.1550-7408.2006.00101.x.
Автор
Дж Норман Грим 1
принадлежность
- 1 Факультет биологических наук, ящик 5640, Университет Северной Аризоны, Флагстафф, Аризона 86011, США.
 [email protected]
[email protected]
- PMID: 16872294
- DOI: 10.1111/j.1550-7408.2006.00101.x
Абстрактный
В течение последних 16 лет инфузория Balantidium jocularum была собрана из кишечника многих особей рыбы-хозяина, Naso tonganus, собранной на Большом Барьерном рифе недалеко от острова Лизард в Австралии. Инфузории для данного исследования потребления пищи были выделены в 1988, 1989, 2003 и 2005. Девятнадцать образцов B. jocularum исследовали в просвечивающем электронном микроскопе для определения содержимого как пищевых вакуолей, так и предполагаемой разряжающей вакуоли цитопрокта. Пищевые вакуоли содержали палочковидные бактерии, туго закрученные спирилловидные бактерии и один или несколько жгутиконосцев эвгленид.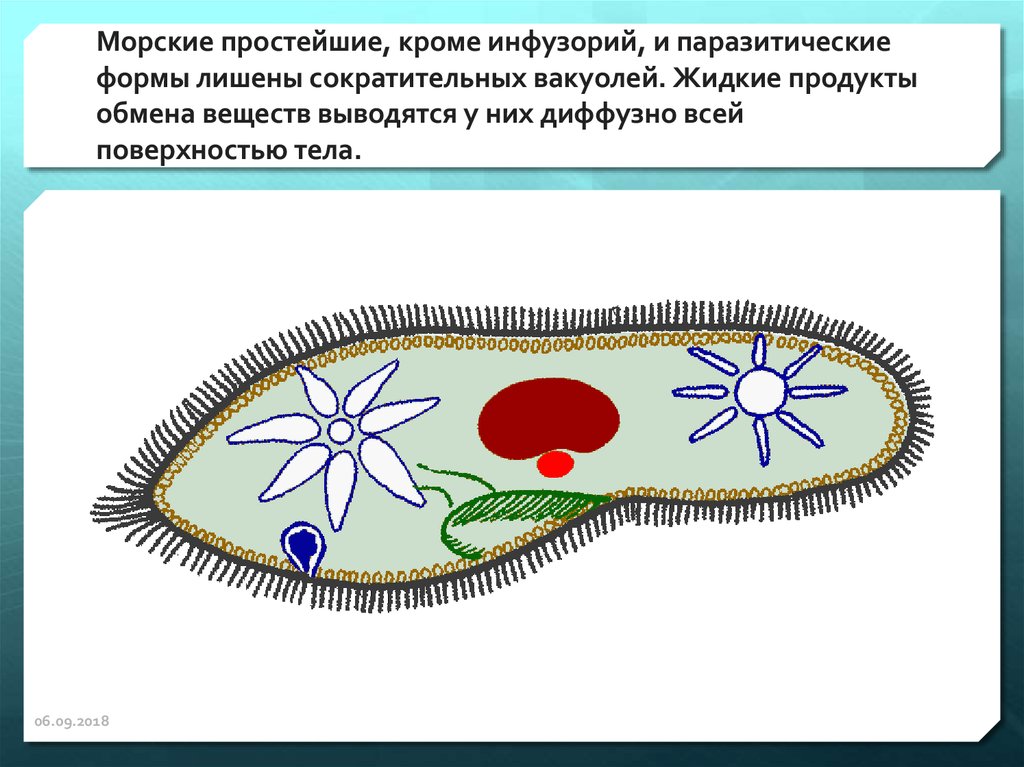 В некоторых балантидиях несколько иной формы, чем типовые виды В. jocularum, в препаратах протаргола под световым микроскопом наблюдалась крупная бактерия Epulopiscium fishelsoni. Некоторые предполагаемые фаголизосомы содержали спирилловидные бактерии, которые были явно интактными, а другие содержали частично переваренные жгутиконосцы. Пища в одной разгружаемой вакуоли цитопрокта состояла из нормально выглядящих спириллиформных бактерий, некоторых других бактерий и без жгутиконосцев. Результаты свидетельствуют о неизбирательном приеме пищи и избирательном пищеварении; следовательно, несколько неэффективная обработка пищевых продуктов.
В некоторых балантидиях несколько иной формы, чем типовые виды В. jocularum, в препаратах протаргола под световым микроскопом наблюдалась крупная бактерия Epulopiscium fishelsoni. Некоторые предполагаемые фаголизосомы содержали спирилловидные бактерии, которые были явно интактными, а другие содержали частично переваренные жгутиконосцы. Пища в одной разгружаемой вакуоли цитопрокта состояла из нормально выглядящих спириллиформных бактерий, некоторых других бактерий и без жгутиконосцев. Результаты свидетельствуют о неизбирательном приеме пищи и избирательном пищеварении; следовательно, несколько неэффективная обработка пищевых продуктов.
Похожие статьи
Новые наблюдения за инфузориями рода Vestibulongum (Pycnotrichidae): вестибулярная ультраструктура, макроядерные эндосимбиотические бактерии, биогеография и доказательства специфичности хозяина.
Грим Дж.

Новое описание рода и типового вида инфузорий Balantidium entozoon.
Грим Дж. Н., Буонанно Ф. Грим Дж. Н. и соавт. Евр Дж Протистол. 2009 г., август; 45 (3): 174–82. doi: 10.1016/j.ejop.2008.10.001. Epub 2009 28 февраля. Евр Дж Протистол. 2009. PMID: 19251405
Рециркуляция мембран в цитопрокте Tetrahymena.
Аллен Р.Д., Вольф Р.В. Аллен Р.Д. и соавт. Дж. Клеточные науки. 1979 февраль; 35: 217-27. doi: 10.1242/jcs.35.1.217. Дж. Клеточные науки.
 1979.
PMID: 106060
1979.
PMID: 106060Новые виды Balantidium и Paracichlidotherus (Ciliophora), населяющие кишечник четырех видов рыб-хирургов с островов Тувалу, Тихий океан.
Грим Дж. Н., Клементс К. Д., Байфилд Т. Грим Дж. Н. и соавт. Дж Эукариот микробиол. 2002 март-апрель;49(2):146-53. doi: 10.1111/j.1550-7408.2002.tb00359.x. Дж Эукариот микробиол. 2002. PMID: 12043962
Оценка безопасности и питательной ценности ГМ-растений и производных пищевых продуктов и кормов: роль испытаний кормления животных.
Рабочая группа EFSA по ГМО по испытаниям кормления животных. Рабочая группа EFSA по ГМО по испытаниям кормления животных. Пищевая химическая токсикол. 2008 март; 46 Приложение 1: S2-70.
 doi: 10.1016/j.fct.2008.02.008. Epub 2008, 13 февраля.
Пищевая химическая токсикол. 2008.
PMID: 18328408
Обзор.
doi: 10.1016/j.fct.2008.02.008. Epub 2008, 13 февраля.
Пищевая химическая токсикол. 2008.
PMID: 18328408
Обзор.
Посмотреть все похожие статьи
Цитируется
Проблемы, с которыми сталкиваются высокополиплоидные бактерии с ограничениями наследования ДНК.
Ангерт ER. Гнев ER. Геном Биол Эвол. 2021 8 июн;13(6):evab037. doi: 10.1093/gbe/evab037. Геном Биол Эвол. 2021. PMID: 33677487 Бесплатная статья ЧВК. Обзор.
Переизоляция и переописание Balantidium duodeni Stein, 1867 (Litostomatea, Trichostomatia).
Чистякова Л.В., Костыгов А.Ю., Корнилова О.А., Юрченко В. Чистякова Л.В. и соавт. Паразитол рез.
 2014 ноябрь;113(11):4207-15. doi: 10.1007/s00436-014-4096-1. Epub 2014 4 сентября.
Паразитол рез. 2014.
PMID: 25185665
2014 ноябрь;113(11):4207-15. doi: 10.1007/s00436-014-4096-1. Epub 2014 4 сентября.
Паразитол рез. 2014.
PMID: 25185665Экстремальная полиплоидия у крупной бактерии.
Менделл Дж. Э., Клементс К. Д., Чоат Дж. Х., Ангерт Э. Р. Mendell JE, et al. Proc Natl Acad Sci U S A. 6 мая 2008 г .; 105 (18): 6730-4. doi: 10.1073/pnas.0707522105. Epub 2008 29 апреля. Proc Natl Acad Sci U S A. 2008. PMID: 18445653 Бесплатная статья ЧВК.
термины MeSH
Физиология сократительных вакуолей | Журнал экспериментальной биологии
Skip Nav Destination
ИССЛЕДОВАТЕЛЬСКАЯ СТАТЬЯ|
01 января 1939 г.
Дж. А. Китчинг
Информация об авторе и статье
Полученный: 19 июн 1938
Онлайновый ISSN: 1477-9145
Печатный ISSN: 0022-0949
Copyright © 1939 Компания Биологов ООО
1939
J Exp Biol 9 0157 (1939) 16 (1): 34–37.
https://doi.org/10.1242/jeb.16.1.34
История статьи
Получено:
19 июня 1938 г.
Связанный контент
Опубликована сопутствующая статья: Физиология сократительных вакуолей: I. Осмотические отношения
Опубликована сопутствующая статья: Физиология сократительных вакуолей: II. Контроль объема тела у морской перитрихи
Опубликована сопутствующая статья:
Физиология сократительных вакуолей: III. Водный баланс пресноводной перитричи
Водный баланс пресноводной перитричи
Это компаньон для: Физиология сократительных вакуолей: V. Влияние кратковременных колебаний температуры на пресноводную перитриховую инфузорию
Это компаньон для: Физиология сократительных вакуолей: VI. Температура и осмотический стресс
Это компаньон для: Физиология сократительных вакуолей: VII. Осмотические отношения у сукториана с особым упором на механизм контроля вакуолярного выброса
Это компаньон для: Физиология сократительных вакуолей: VIII. Водные отношения сукторианца Подофря во время кормления
Опубликована сопутствующая статья:
Физиология сократительных вакуолей: X. Влияние высокого гидростатического давления на сократительные вакуоли сукториана
Влияние высокого гидростатического давления на сократительные вакуоли сукториана
Опубликована сопутствующая статья: Физиология сократительных вакуолей: IX. Влияние внезапных изменений температуры на сократительную вакуоль A Suctorian; с обсуждением механизма сокращения
Опубликована сопутствующая статья: Физиология сократительных вакуолей: XI. Влияние тяжелой воды на водный баланс сукториана.
- Разделенный экран
- Взгляды
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- Нажмите здесь, чтобы открыть pdf в другом окне PDF для
- Значок версии статьи Версии
- Версия записи
01 января 1939 г.
- Версия записи
01 января 1939 г.
- Делиться
- Твиттер
- MailTo
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
Citation
Дж. А. Китчинг; Физиология сократительных вакуолей: IV. Заметка об источниках эвакуированной воды и о функции сократительных вакуолей у морских простейших. J Exp Biol 1 января 1939 г. ; 16 (1): 34–37. doi: https://doi.org/10.1242/jeb.16.1.34
; 16 (1): 34–37. doi: https://doi.org/10.1242/jeb.16.1.34
Скачать файл цитаты:
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- КонецПримечание
- РефВоркс
- Бибтекс
Расширенный поиск
У перитриховых инфузорий может образовываться много пищевых вакуолей без видимого твердого содержимого. Вода в этих вакуолях переходит в общую цитоплазму.
У пресноводных Peritricha скорость поглощения жидкости пищевыми вакуолями обычно составляет от 8 до 20% скорости выделения жидкости сократительной вакуолью. Предполагается, что большая часть эвакуированной воды поступает в животное путем осмоса через общую поверхность тела.
У морских Peritricha скорость поглощения жидкости пищевыми вакуолями примерно уравновешивает скорость выхода сократительной вакуоли.
 Удаление воды, поступающей в пищевые вакуоли, считается основной функцией сократительной вакуоли морских инфузорий.
Удаление воды, поступающей в пищевые вакуоли, считается основной функцией сократительной вакуоли морских инфузорий.
Для Paramecium caudatum было показано, что скорость поступления воды в пищевые вакуоли значительно меньше скорости выхода воды из сократительной вакуоли (Eisenberg, 19).25). Китчинг (1938 b ) пришел к выводу, что у большинства пресноводных простейших основная часть воды, эвакуируемой сократительной вакуолью, поступает через поверхность тела; но что в некоторых формах, при которых большое количество жидкости проглатывается за один «глоток» или за очень короткое время, это может обеспечить большую часть выхода сократительной вакуоли. Представлялось желательным получить оценку скорости поглощения воды пищевыми вакуолями для сравнения со скоростью выхода сократительной вакуоли простейших, отличных от 9.0302 Парамеций . Соответственно, это вещество было исследовано у пресноводных и морских перитриховых инфузорий. Представлялось возможным, что у морских простейших, у которых скорость поступления воды через поверхность тела, вероятно, очень мала, основная часть воды, элиминируемая сократительной вакуолью, могла поступать в пищевые вакуоли. Это предположение, по-видимому, предлагало разумное функциональное объяснение существования сократительных вакуолей у многих морских инфузорий.
Представлялось возможным, что у морских простейших, у которых скорость поступления воды через поверхность тела, вероятно, очень мала, основная часть воды, элиминируемая сократительной вакуолью, могла поступать в пищевые вакуоли. Это предположение, по-видимому, предлагало разумное функциональное объяснение существования сократительных вакуолей у многих морских инфузорий.
Пресноводные Peritricha были получены из ног изопод Asellus из пруда на территории университета в Бристоле. Солоноватая вода Peritricha была получена на зеленой водоросли Cladophora , которая росла на корабле, постоянно пришвартованном в доке на реке Эйвон в Бристоле. Морские перитрихи были получены по почте из Плимута, где они заселили Cladophora в аквариуме. Фактические виды, использованные в каждом эксперименте, указаны в Таблице I.
Таблица I
Вид Большой
Во время наблюдений организмы поливали уже описанным способом (Китчинг, 1934), за исключением того, что температура не контролировалась. Комнатная температура находилась между 14 и 20°С, но ни в одном эксперименте не менялась более чем на полградуса.
Комнатная температура находилась между 14 и 20°С, но ни в одном эксперименте не менялась более чем на полградуса.
Измерения диаметра вакуолей производили с помощью окуляра винтового микрометра Zeiss. В случае с пищевыми вакуолями, вероятно, была допущена значительная экспериментальная ошибка, поскольку вакуоли не были сферическими при первом проглатывании. Их измеряли, как только они приобретали шарообразную форму (в течение 12–1 мин.), но, возможно, они претерпели некоторую усадку до этого времени в случае пресноводных перитрих, так что рассчитанные значения скорости поглощения жидкости в пищевые вакуоли, возможно, слишком низкие.
У Pentricha, орошаемого морской водой или фильтрованной водой из пруда (в зависимости от того, что подходит), пищевые вакуоли поступали через регулярные промежутки времени, обычно около 1-3 минут. Почти все они были лишены видимого твердого содержимого, хотя в них могли присутствовать бактерии. Окруженные цитоплазмой тела, эти вакуоли сокращались, медленно или быстро, пока не становились невидимыми. Сморщивание происходило, когда они находились на некотором расстоянии от пищевода или поверхности тела, так что следует предположить, что содержащаяся в них вода переходила в цитоплазму. (Фактическая скорость прохождения жидкости через вакуолярную мембрану будет рассмотрена в следующей статье.)
Сморщивание происходило, когда они находились на некотором расстоянии от пищевода или поверхности тела, так что следует предположить, что содержащаяся в них вода переходила в цитоплазму. (Фактическая скорость прохождения жидкости через вакуолярную мембрану будет рассмотрена в следующей статье.)
«Пустые» пищевые вакуоли при сморщивании исчезали из поля зрения, обратного выброса содержимого пищевых вакуолей в глотку не наблюдалось. Гринвуд (1894) показал, что таким образом избавляются от неперевариваемых твердых веществ вместе с некоторым количеством воды, но я не мог обнаружить соответствующего выброса воды в случае «пустых» пищевых вакуолей. Действительно образуется много пустых пищевых вакуолей, из которых вся вода переходит в цитоплазму.
Различные пресноводные, солоноватые и морские воды Peritricha орошались фильтрованной прудовой водой, фильтрованной солоноватой водой — соленостью, эквивалентной 5% морской воды — из реки Эйвон в Бристоле и Плимута «вне» морской водой соответственно. Наблюдения проводились за частотой и диаметром сократительной вакуоли и пищевых вакуолей. Результаты сведены в Таблицу I.
Наблюдения проводились за частотой и диаметром сократительной вакуоли и пищевых вакуолей. Результаты сведены в Таблицу I.
У пресноводных Peritricha скорость поступления жидкости в пищевые вакуоли в большинстве случаев составляет от 8 до 20 % скорости выхода из сократительной вакуоли. С другой стороны, у морских Peritricha скорости поглощения и выхода были примерно одинаковыми, и сомнительно, чтобы наблюдаемые различия были существенными ввиду довольно большой ошибки эксперимента (см. выше).
У пресноводных Peritricha скорость всасывания жидкости из пищевых вакуолей составляет лишь малую часть скорости выведения сократительной вакуоли. В пресной воде Zoothamnium sp.? использованный в предыдущей работе (Kitching, 1938 a ) меньше одной десятой. При таких формах следует предполагать, что большая часть эвакуированной воды поступила через общую поверхность тела путем осмоса и что сократительная вакуоль функционирует как осморегуляторный механизм. Но у морских Peritricha скорости поглощения пищевыми вакуолями и выброса сократительной вакуоли примерно уравновешиваются (в довольно широких пределах погрешности эксперимента), и можно заключить, что основной функцией сократительной вакуоли является устранение избытка вода, поступающая с пищевыми вакуолями. Скорость удаления воды вместе с неперевариваемым материалом из старых пищевых вакуолей ничтожно мала.
Но у морских Peritricha скорости поглощения пищевыми вакуолями и выброса сократительной вакуоли примерно уравновешиваются (в довольно широких пределах погрешности эксперимента), и можно заключить, что основной функцией сократительной вакуоли является устранение избытка вода, поступающая с пищевыми вакуолями. Скорость удаления воды вместе с неперевариваемым материалом из старых пищевых вакуолей ничтожно мала.
Сразу же возникает вопрос, почему морские корненожки не нуждаются в сократительных вакуолях? Возможный ключ к этой проблеме лежит в том факте, что Rhizopoda обычно формируют пищевые вакуоли только тогда, когда пища доступна; они обычно не берут «пустые» или почти «пустые». По данным Mast & Hahnert (1935), Amoeba иногда эвакуирует из старых пищевых вакуолей неперевариваемые остатки вместе с некоторым количеством воды. Возможно, таким образом поддерживается баланс между поглощением и выходом у морских Rhizopoda без необходимости наличия сократительной вакуоли.
В случае морских простейших ясно, что значительное количество солей должно поглощаться пищевыми вакуолями, и неизвестно, эвакуируются ли они сократительной вакуолью или как они удаляются. Возможная эвакуация эндогенной воды сократительной вакуолью (предположенная Камада, 1935) уже обсуждалась (Китчинг, 1936). Количество такой воды должно быть чрезвычайно малым и не может обеспечить более мельчайшей части продукции. Остается возможность проникновения небольшого количества воды через поверхность тела за счет осмотического давления клеточных белков. Однако, по-видимому, у морских Peritricha основной функцией сократительной вакуоли является удаление воды, приносимой пищевыми вакуолями.
Айзенберг
,
E.
(
1925
).
Арх. Biol., Paris,
35
,
441
.
Гринвуд
,
М.
(
1894
).
Филос. Транс. Б
,
185
,
355
.
Камада
,
Т.
(
1935
).
J. Факт. науч. Токийский университет
.
4
,
49
.
Комплектация
,
J. A.
(
1934
).
Дж. эксп. Биол
.
11
,
364
.
https://doi.org/10.1242/jeb.11.4.364
Китчинг
,
Дж. А.
(
1936
).
Природа, Лондон,
138
,
287
.
https://doi.org/10.1038/138287a0
Китчинг
,
J.A.
(
1938a
).
Дж. эксп. Биол
.
15
,
143
.
https://doi.org/10.1242/jeb.15.1.143
Комплектация
,
J. A.
(
1938b
).