Косослой — это… Что такое Косослой?
Косослой Измерение косослойности Тангентальный наклон волоконКососло́й — порок строения древесины, выражающийся в отклонении направления древесных волокон от продольной оси сортимента — древесного ствола, пилопродукции или шпона.
Естественный косослой
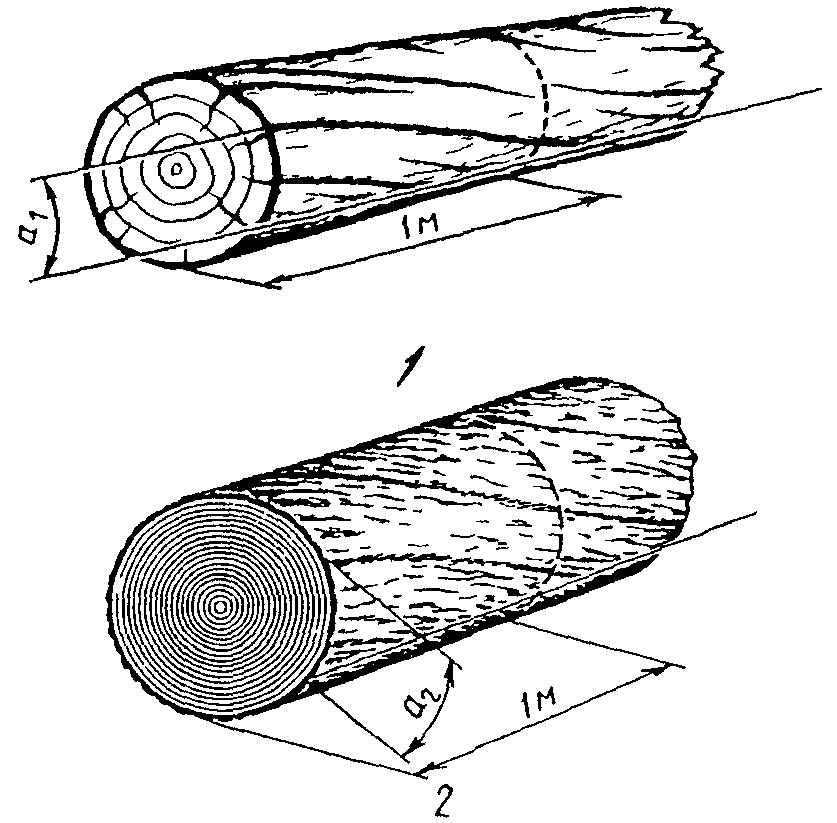
Естественный косослой, или тангентальный (тангенциальный) наклон волокон выражается в расположении волокон по спирали вокруг оси дерева. Опытный человек может определить косослойное дерево ещё на корню, покрытое корой. Определяется по трещинам на поверхности коры или, у окорённого дерева, по трещинам на стволе. У разных видов и пород идёт либо по часовой стрелке, либо против часовой. Данный порок настолько широко распространён у многих древесных пород, что может считаться скорее нормой, чем отклонением от неё. Проявляется почти у всех видов деревьев, особенно часто у сосны, ели, лиственницы и граба. Не ограничивается стволом, распространяется и на сучья и ветки; увеличивается от сердцевины к периферии и по высоте ствола.
Дерево, поражённое косослоем, называется косослойным деревом. Существуют целые косослойные леса. Есть мнение, что они появляются вследствие передачи этого порока через семена, на что указывает повальное его распространение в некоторых местностях, например, косослой очень распространён в Туруханском крае, по Енисею, где, как указывает ЭСБЕ, волокна делают почти целый оборот вокруг ствола на 1 фут (30,48 см).
Измеряется в процентах: отклонение волокон в 1 см на 1 м длины составляет 1 % косослойности древесины. Особенно значительно снижает прочность древесины на растяжение и изгиб косослойность больше 5 %. При измерении первый метр от комлевого торца в расчёт не принимают. Косослой увеличивает прочность древесины при раскалывании, порождает повышенную продольную усушку и коробление.
Искусственный косослой
Радиальный наклон волокон, или искусственный косослой (неофиц. ) или перерезание годовых слоёв (неофиц.) — отклонение направления годовых слоёв от продольной оси пиломатериала, шпона или деталей. Образуется при пилении лесоматериалов со сбежистостью, закомелистостью или кривизной. Может образоваться и при неправильной распиловке прямослойной древесины. Древесина с таким пороком плохо воспринимает поперечную нагрузку, не подходит для гнутья. Измеряется так же, как и естественный косослой. Затрудняет механическую обработку (строжку и тёску).
) или перерезание годовых слоёв (неофиц.) — отклонение направления годовых слоёв от продольной оси пиломатериала, шпона или деталей. Образуется при пилении лесоматериалов со сбежистостью, закомелистостью или кривизной. Может образоваться и при неправильной распиловке прямослойной древесины. Древесина с таким пороком плохо воспринимает поперечную нагрузку, не подходит для гнутья. Измеряется так же, как и естественный косослой. Затрудняет механическую обработку (строжку и тёску).
См. также
Литература
- Лесная энциклопедия: В 2-х т./Главный редактор Воробьев Г. И.; редколлегия: Анучин Н. А., Атрохин В. Г., Виноградов В. Н. и др. — М.: Советская энциклопедия, 1985 г. — 563 с., ил.
- Пороки древесины. Альбом/Миллер В. В., Вакин А. Т.. — М., Л.: Катологиздат НКТП СССР, 1938. — 171 с.
- Вакин А. Т., Полубояринов О. И., Соловьёв В. А.. Альбом пороков древесины. — М.: Лесная промышленность, 1969.
- Вакин А. Т., Полубояринов О. И.
 , Соловьёв В. А.. Альбом пороков древесины. — М.: Лесная промышленность, 1980.
, Соловьёв В. А.. Альбом пороков древесины. — М.: Лесная промышленность, 1980.
Ссылки
Пороки древесины и их виды: сучки дефекты трещины
При заготовки древесины различают различные пороки, дефекты, повреждения.
Это — ненормальное развитие некоторых частей ствола, различного вида сучки, трещины, повреждения дереворазрушающими грибами, насекомыми и др.
Многие пороки древесины появляются во время роста дерева, но выявить их в растущем дереве не всегда воможно; большей частью их обнаруживают уже после того, как дерево срублено.
Пороками древесины называют недостатки, свойственные лесным материалам и обусловленные отклонениями от нормального строения, повреждениями (механическими и микологическими) растущего дерева и срубленной древесины.
Некоторые пороки могут возникнуть в процессе заготовки, транспортирования и хранения материала.
Пороки в зависимости от характера, размеров и расположения снижают сорт древесины или же делают ее совершенно непригодной для производственных целей. Некоторые пороки (волнистость древесных волокон и др.) оказываются полезными, улучшая текстуру древесины для столярных изделий.
Некоторые пороки (волнистость древесных волокон и др.) оказываются полезными, улучшая текстуру древесины для столярных изделий.
Наименование пороков и их подробная характеристика установлены ГОСТ.
Наиболее часто встречающихся пороки:
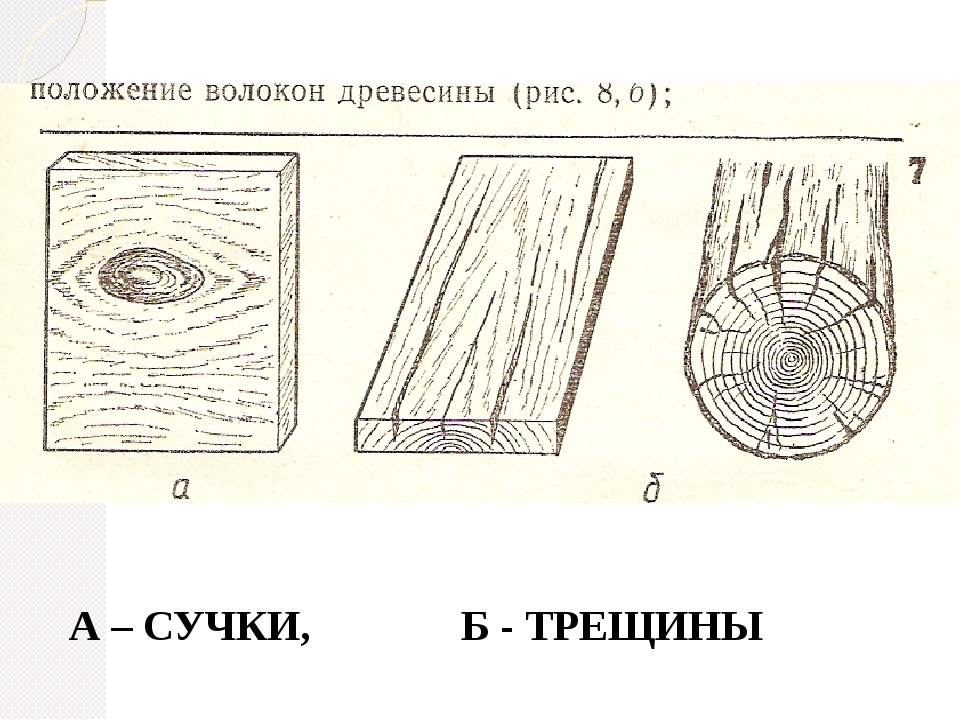
- сучки,
- трещины,
- неправильность строения,
- гнили, цветные окраски.
Разновидности трещин встречающихся в древесине. Влияние трещин на строительные материалы
Трещины, наружные и внутренние, встречаются на растущем дереве, а также на заготовленном лесоматериале и изделиях из древесины.
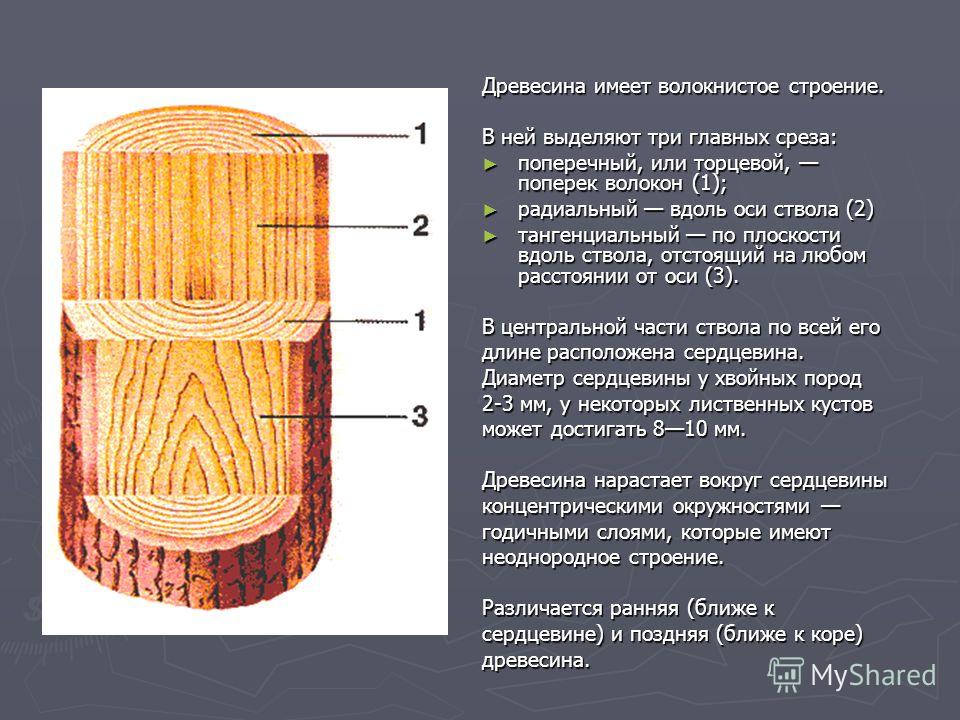
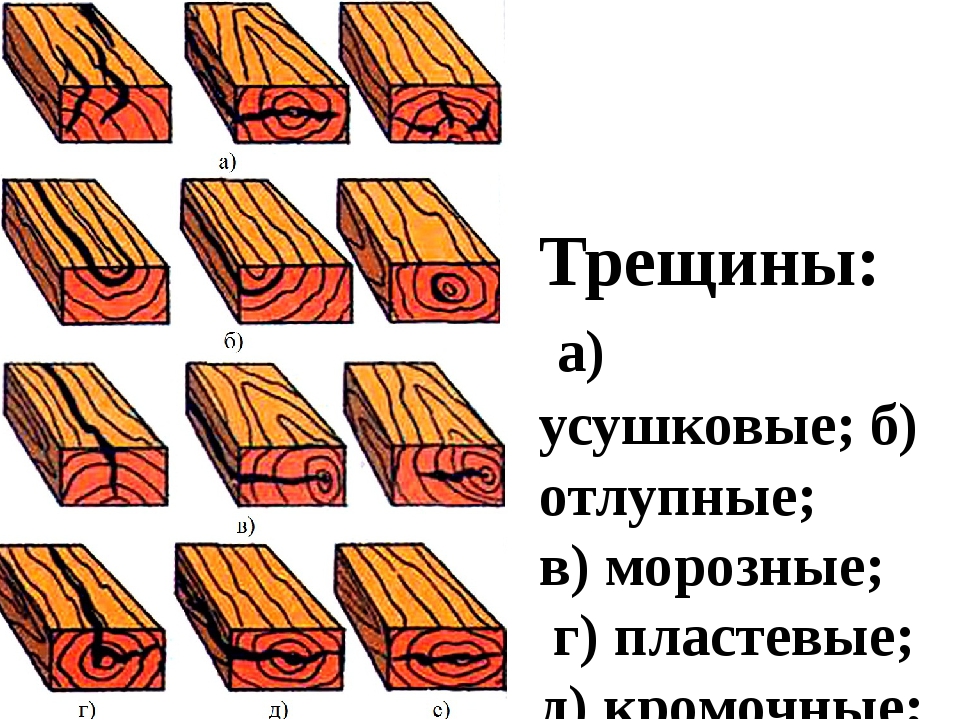
Влияние трещин на качество материала зависит от их размера, количества, вида, а также и положения по длине, ширине и толщине лесоматериала. Трещины задерживают влагу, попадающую извне, а это в свою очередь повышает влажность самой древесины и создает условия для развития в ней грибов. Метик (рис. 1) одна или несколько широких внутренних продольных трещин, постепенно суживающихся от центра к периферии ствола.
Рис. 1. Метики: 1 — простой; 2 — лучистый
Простой метик
Различают простой метик (рис. 1,), когда трещина проходит по диаметру ствола, и лучистый (рис. 1, 2), когда две или несколько трещин расположены по радиусам, под углом друг к другу. Метик называется согласным, если трещина идет по стволу, оставаясь в одной плоскости, и несогласным, если трещина идет винтообразно. Метик влияет на качество материала при распиловке бревен.
Несогласный метик
Несогласный метик сильно снижает выход полноценного материала, так как количество доброкачественных досок, выпиленных из такого бревна, будет небольшим. При сушке материалов метик увеличивается в размерах.
Рис. 2. Отлуп: 1 — в круглом лесе; 2 — в доске.
Отлуп
Отлуп — внутренняя трещина, идущая по годичному слою (рис. 2) и распространяющаяся на некоторую длину ствола.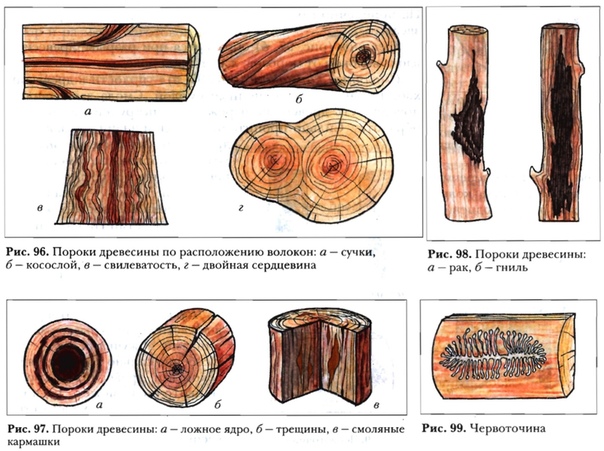 На торцах она имеет вид дугообразных трещин, не заполненных смолой. Различают отлуп частичный, когда трещина занимает часть годичного слоя, и кольцевой, если она распространяется по всему годичному слою.
На торцах она имеет вид дугообразных трещин, не заполненных смолой. Различают отлуп частичный, когда трещина занимает часть годичного слоя, и кольцевой, если она распространяется по всему годичному слою.
В пиломатериалах эти трещины представляют собой луночки на торцах, а на боковых поверхностях — продольные трещины или желобчатые углубления.
Рис. 3. Трещины: 1 — торцовая в доске; 2 — на боковой поверхности в доске с сердцевиной, выходящей на широкую грань
Наружные трещины
Наружные трещины, появляющиеся при высыханий на бревнах и различных лесных материалах, называются трещинами усушки; различают торцовые и боковые трещины (рис. 3): Трещины в круглом лесе, брусьях и досках появляются прежде всего на торцах, так как интенсивность испарения влаги через единицу торцовой поверхности (вследствие большой пористости торца)в несколько раз больше, чем при высыхании боковой поверхности.
Неодинаковая по величине усушка, а также неравномерность высыхания древесины вследствие медленного продвижения воды от более влажных участков к менее влажным (особенно в поперечном направлении) — причина появления внутренних напряжений в древесине. Эти напряжения вызывают растрескивание лесоматериалов при высыхании. У подсохшего снаружи отрезка древесины волокна наружной зоны под влиянием усушки стремятся сократить свои размеры, в то время как во внутренних слоях влажность еще велика (больше 30%) и усушки нет. В результате наружные волокна древесины оказываются под действием растягивающих усилий, а внутренние под действием сжимающих.
Эти напряжения вызывают растрескивание лесоматериалов при высыхании. У подсохшего снаружи отрезка древесины волокна наружной зоны под влиянием усушки стремятся сократить свои размеры, в то время как во внутренних слоях влажность еще велика (больше 30%) и усушки нет. В результате наружные волокна древесины оказываются под действием растягивающих усилий, а внутренние под действием сжимающих.
Трещины усушки
Трещины усушки обычно имеют радиальное направление, поскольку древесина разрывается по сердцевинным лучам, так как связь между продольными волокнами и сердцевинными лучами наиболее слабая.
Наиболее подвержены растрескиванию дуб и бук — породы, имеющие крупные сердцевинные лучи.
Максимальной ширины наружные трещины достигают на поверхности лесоматериалов и постепенно суживаются к центру. Пиломатериалы растрескиваются меньше, чем круглый лес. Размер и характер трещин зависят от породы дерева, быстроты сушки и т. п.
Торцовые трещины
Торцовые трещины увеличивают количество отходов, поэтому необходимо предотвращать их появление.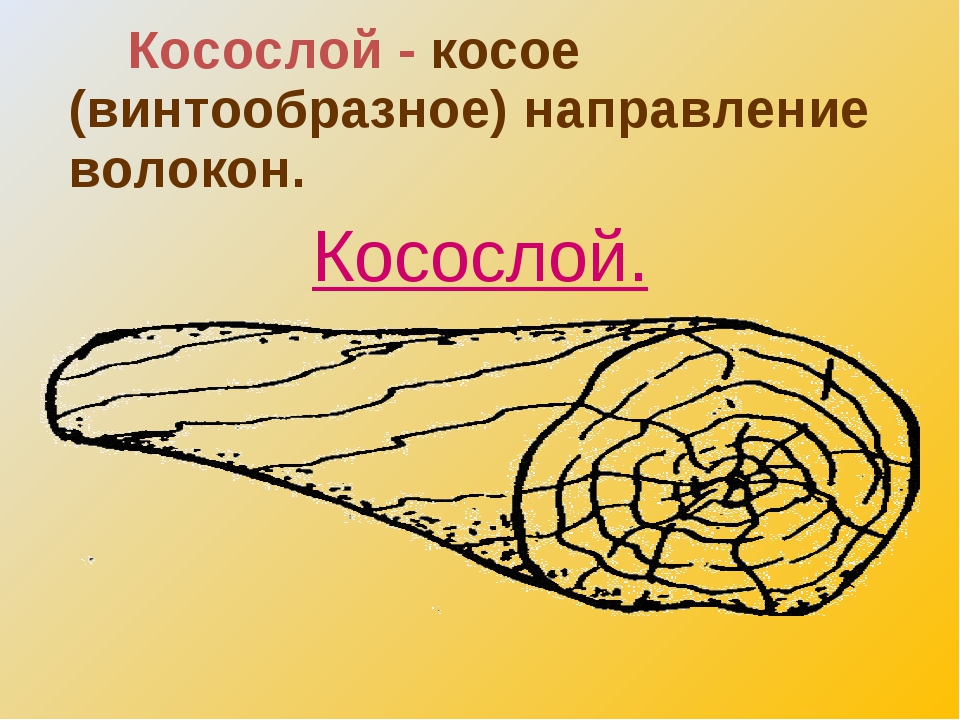 При хранении лесоматериалов в штабелях торцы затеняют тонкими досками (наторцовниками), вертикально прикрепленными к штабелю, покрывают краской или составами, которые препятствуют быстрому испарению влаги из древесины
При хранении лесоматериалов в штабелях торцы затеняют тонкими досками (наторцовниками), вертикально прикрепленными к штабелю, покрывают краской или составами, которые препятствуют быстрому испарению влаги из древесины
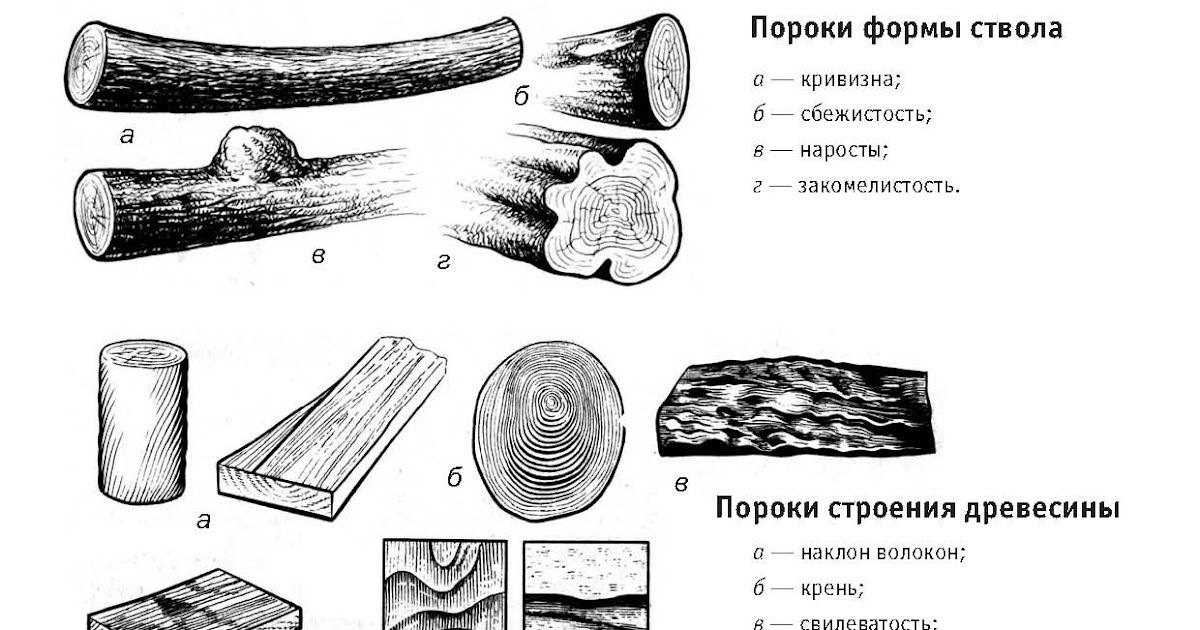
Пороки строения древесины, ствола
К группе пороков, образовавшихся вследствие неправильности строения древесины, относят: косослой, наклон годичных слоев, свилеватость, завиток и крень.
Косослой
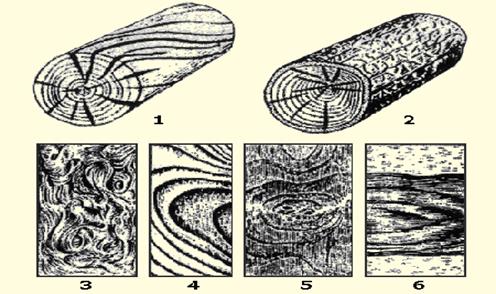
Косослоем называется винтообразное направление волокон древесины от комля к вершине. Небольшие отклонения волокон от прямолинейного направления наблюдаются почти у каждого дерева, но пороком считается лишь такое отклонение, когда наклон волокон по отношению к ребру доски или бруса составляет более 5°. Часто волокна имеют винтообразное направление по всей толщине ствола, но иногда косое расположение имеют только наружные слои дерева, а внутренние в центральной части ствола сохраняют прямое направление.
Рис. 1. Косослой: 1— косослойное бревно; 2 — косослойная доска
Косослой определяют по косым трещинам на боковой поверхности подсушенного материала, а в сыром материале по полоскам смолы или синевы, идущим вдоль волокон (рис.
Косослойная древесина легко растрескивается и имеет пониженную прочность. Снижение прочности (особенно на растяжение и изгиб) при косослое является результатом того, что действующее вдоль длины доски или бруса усилие направлено под углом к волокнам и стремится разъединить их сдвинуть или оторвать.
Влияние косослоя на механические свойства древесины начинает заметно сказываться, когда величина его превышает 5°. При больших углах наклона волокон или годичных слоев наблюдается резкое снижение предела прочности.
Крень
Крень — утолщение поздней части годичных слоев с резко повышенной твердостью на одной стороне торца (рис. 1). Встречается на торце в виде темноокрашенных участков и на боковых поверхностях пиломатериалов в виде сплошной полосы темноокрашенной древесины.
Крень почти всегда сопровождается внецентренным расположением сердцевины и вызывает значительное коробление пиломатериалов не только в поперечном, но и в продольном направления.
Прорость
Прорость — омертвение части древесины или коры в результате повреждений поверхности ствола. Прорость бывает открытой и закрытой. В последнем случае омертвевшая древесина, иногда даже поверх коры, обрастает сомкнутыми слоями живой древесины. Открытая прорость наблюдается на боковой поверхности бревна или доски в виде более или менее широкой продольной ложбины (рис. 2).
Рис. 2. Прорость: 1—открытая; 2 — закрытая; 3 — следы прорости в доске.
Засмолок
Засмолок, встречающийся на древесине хвойных пород, характеризуется тем, что участок древесины, прилегающий к месту повреждения ствола дерева, обильно пропитывается смолой. В пиломатериалах засмоленные участки значительно темнее окружающей здоровой древесины. На качество древесины засмолок из-за своих небольших размеров особого влияния не оказывает, но затрудняет обработку (строжку).
Сбежистость
Сбежистость — ненормальное уменьшение толщины круглого леса или ширины необрезной доски на всем протяжении от комля к вершине. Дерево считается сбежистым, если диаметр его изменяется на каждый метр более чем на 2 см. Сбежистость увеличивает количество отходов при распиловке круглого леса и влечет за собой перерезание годичных слоев в пиломатериалах.
Кривизна
Кривизна (искривление ствола) служит одной из причин перерезания годичных слоев в пиломатериалах. Кривизна в зависимости от ее размеров и вида может понижать сорт деловой древесины, в результате чего иногда приходится переводить ее в категорию дровяной.
Наклон годичных слоев, завиток, свилеватость древесины
Особенности строения древесины задают методику обработки и изготовлению строительных материалов.
Наклон годичных слоев получается в пиломатериалах в тех случаях, когда бревно распиливают под некоторым углом к продольному направлению, и чаще всего при распиловке параллельно сердцевине закомелистых, сбежистых и кривых бревен.
Косослойная древесина обычно раскалывается по радиусу годичных слоев, а при наклоне по годичным слоям (рис. 6).
Рис. 6. Раскалываемость древесины: 1— косослойной; 2 — с наклоном годичных слоев
Завиток, называемый также присучковым косослоем, представляет собой местное искривление годичных слоев из-за наличия сучков. На поверхностях пиломатериалов завиток имеет вид частично перерезанных и изогнутых годичных слоев (рис. 7).
Если завиток расположен на нижней кромке (в зоне наибольшего изгибающего момента), он особенно сильно влияет на прочность древесины при изгибе. В этом случае действующее на волокна растягивающее усилие, разъединяющее их, особенно опасно, так как сопротивление древесины отрыву волокон незначительно. Завиток почти не снижает прочности древесины, если он расположен на сжатой кромке или в мало-напряженных участках.
Рис.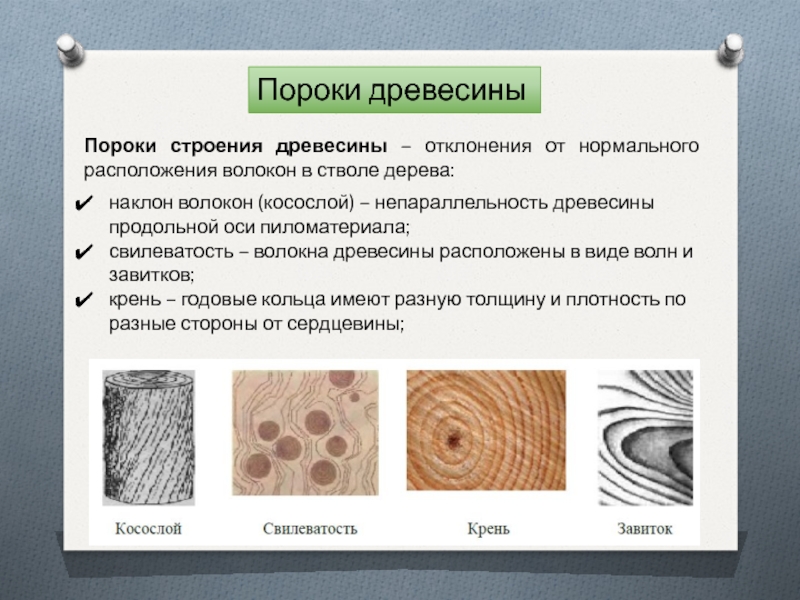 7. Присучковой косослой (завиток) в доске, выпиленной вблизи сучка.
7. Присучковой косослой (завиток) в доске, выпиленной вблизи сучка.
Свилеватость
Свилеватость выражается в резко волнистом или путаном расположении древесных волокон. Свилеватость обычно наблюдается в нижней (комлевой) части ствола. Сопротивляемость древесины изгибу в этом случае понижается, а сопротивление сжатию и раскалыванию повышается. Свилеватая древесина трудно поддается обработке — строганию, раскалыванию. Свилеватую древесину клена, ореха, ясеня, карельской березы и других пород применяют в виде фанеры и в столярно-отделочных работах, в которых она ценится за свою красивую текстуру.
|
|
|
|
Отклонение от нормы в строении ствола дерева, внешнего вида и формы,
а также нарушение физического состояния и повреждение древесины, снижающие
ее качество и возможность применения, называют пороками древесины. |
|
|
Кривизна |
Искривление ствола вдоль оси дерева. Она может быть простой (ствол имеет один изгиб) и сложный (ствол имеет несколько изгибов в разном направлении). |
|
Закомелистость |
Закомелистость
характеризуется сильным утолщением или увеличением диаметра комля по отношению
к стволу дерева. |
|
Ройки |
Выявлены продольными углублениями в комлевой части ствола. Поперечный распил торца бревна выглядит звездообразным с волнистыми расположением годичных колец.. При распиле на доски большую часть ствола выбраковывают в отходы, поскольку такие доски сильно коробятся и имеют пониженную прочность. |
|
Наросты |
Местное
резкое утолщение ствола различной формы и размеров, в большинстве случаев
встречаются на лиственных породах – березе, клене, ольхе, дубе и др.
наросты бывают двух видов – наплывы и капы. |
|
Наплывы: |
Это внутреннее заболевание дерева, сопровождающееся наростами с гладкой поверхностью, чаще бывают на комлевой части дерева. |
|
Капы: |
Выражены более рельефной поверхностью; при очистке от коры рельеф выглядит в виде капель. |
|
|
Древесина наростов очень плохо поддается обработке, но зато отличается красивой свилеватой текстурой. |
|
|
Широко
используется при изготовлении художественных изделий, при облицовке мебели
шпоном. |
|
Сучки |
Сучки имеются во всех древесных породах, это наиболее неизбежный порок
древесины. |
|
Косослой |
Косослой
представляет собой ярко выраженное косое (винтообразное) расположение
волокон в древесине относительно продольной оси ствола. |
|
Свилеватость |
Свилеватость в древесине выражена волнистым и перепутанным расположением волокон, чаще всего встречаются на лиственных породах в комлевой части ствола. Свилеватость повышает прочность древесины, красоту ее фактуры, но затрудняет обработку, резьбе поддается с большим трудом. Очень большой свилеватостью отличается карельская береза, за что так высоко и ценится. |
|
Крень |
Крень часто образуется в древесине искривленных и наклонно стоящих стволов. Крень нарушает однородность строения древесины, понижает прочность, способствует сильному продольному короблению досок и брусьев. |
|
Двойная сердцевина |
Двойная
сердцевина ярко выражена при поперечном распиле ствола в месте раздвоения. |
|
Внутренняя заболонь |
Внутренняя заболонь – группа годичных колец-слоев, расположенных в ядровой древесине, имеющая окраску, свойства и строение заболони. На торце ствола она ярко выражена в виде одного или нескольких колец разной ширины, более светлых, чем ядро древесины. Такой порок наблюдается в стволах лиственных пород, особенно у дуба и ясеня. |
|
Ложное ядро |
Ложное
ядро представляет собой темную окраску разных оттенков внутренней части
ствола, возникающую в березе, ольхе, клене и др. |
|
Прорость |
Проростью называют полностью или частично сухую древесину или кору. Проросль возникает в результате наружных повреждений дерева и значительно снижает прочность древесины. Она может быть закрытой и открытой. |
|
Трещины |
Трещины в стволах деревьев возникают в результате разрывов древесины вдоль волокон. Они образуются от усушки, морозобоя, отлупа -–внутренней трещины между соседними годовыми слоями. Трещина может быть и метиковой – образованной внутри ствола по радиусу или диаметру. |
|
Смоляной кармашек |
Смоляной
кармашек – небольшая полость между слоями в древесине хвойных пород,
заполненная смолой. |
|
Рак |
Рак – это рана на поверхности ствола дерева, возникшая в результате заражения паразитическим грибом и бактериями. На месте заражения древесина не нарастает, а с противоположной стороны ствола ввиду усиленного прироста образуется характерное вздутие (опухоль). |
|
Червоточина |
Разрушение
древесины личинками и жуками, поедающими лубяной слой дерева
свежесрубленных неокоренных лесоматериалов. При очистке древесины от коры
хорошо видны следы деятельности насекомых в виде ходов и бороздок различной
формы и конфигурации. Червоточина может быть поверхностной, когда жуки
проделывают ходы в коре и лубяном слое; глубокой и неглубокой, когда ходы
жуков-древесников проникают на несколько сантиметров в глубину или даже
пронизывают древесину насквозь. |
|
Гнили |
Гнили возникают в сухостойной, валежной и срубленной древесине под воздействием дереворазрушающих грибов. В начальной стадии гниения появляется ненормальная для древесины окраска, меняется механическая прочность, и постепенно дерево разрушается, превращаясь в труху. |
|
Внутренняя трухлявая гниль |
Развивается
в сердцевинной части ствола растущего дерева. Инфекция в виде бактерий и
грибов проникает в ствол и корни через сломанные сучки и другие раны и разрушает
дерево. Внутренняя трухлявая гниль настолько поражает древесину, что она
превращается в порошок или выдирается пучками волокон. Трухлявая гниль
встречается в древесине всех пород. |

 При изготовлении досок из этой части ствола неизбежны
большие отходы. Полученный материал при распиловке — невысокого качества,
так как появляется большое количество перерезанных волокон.
При изготовлении досок из этой части ствола неизбежны
большие отходы. Полученный материал при распиловке — невысокого качества,
так как появляется большое количество перерезанных волокон.

 Косослой вызывает
коробление (скручивание) досок.
Косослой вызывает
коробление (скручивание) досок.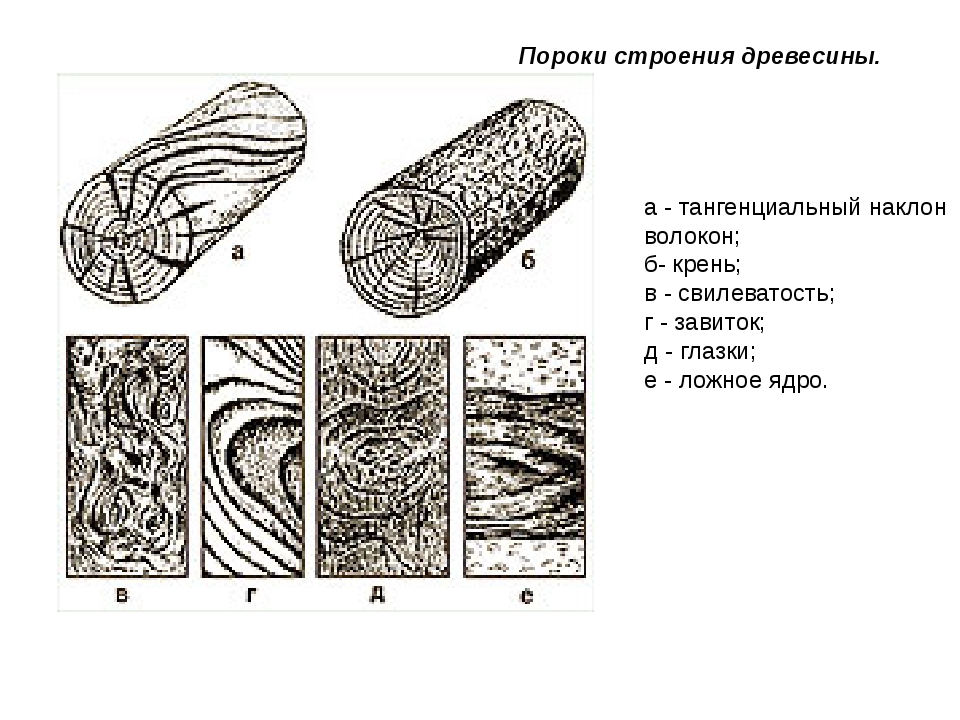 Торец дерева в этом месте обычно имеет овальную форму. Часто между двумя
сердцевинами бывает закрытая прорость (заросшая кора). Необычная форма
распила вместе с текстурным рисунком могут дать интересный декоративный
эффект.
Торец дерева в этом месте обычно имеет овальную форму. Часто между двумя
сердцевинами бывает закрытая прорость (заросшая кора). Необычная форма
распила вместе с текстурным рисунком могут дать интересный декоративный
эффект. Ложное ядро бывает
круглой, эксцентричной, звездчатой или лопастной формы. От заболони л.я.
отличается более темной окраской.
Ложное ядро бывает
круглой, эксцентричной, звездчатой или лопастной формы. От заболони л.я.
отличается более темной окраской.

Урок по технологии в 5-м классе по теме «Древесина. Обработка древесины»
Цели:
- Образовательные – систематизировать и обобщить изученный материал по указанной теме (свойства древесины, недостатки, пороки, виды инструментов, породы древесин).
- Воспитательные – содействовать воспитанию у учащихся аккуратности, ответственности, неравнодушия к результатам своего труда, практической значимости изготовляемого изделия.
- Развивающие – развитие творческой активности учащихся.
Ход урока
1. Урок начинается с развивающей игры:
На доске записано несколько слов в произвольном порядке, из которых надо составить предложение.
Человек, материал, который, древности, в глубокой, распространенный, обрабатывать, научился.
У учащихся должно получиться следующее предложение:
Распространенный материал, который человек научился обрабатывать в глубокой древности.
Ребята, как вы думаете, что это за материал?
Древесина.
2. Учащимся сообщается тема урока: Древесина. Обработка древесины.
Давайте вспомним все, что мы изучали по данной теме.
1). Ребята, оглядитесь вокруг и назовите мне как можно больше предметов, сделанных из древесины. Перечисляют.
2). Определите на глаз длину карандаша, длину указки (в см).
Учитель проверяет с помощью линейки, у кого из ребят получился наиболее точный ответ.
3). Фронтальный опрос.
— Как называется профессия рабочего, занятого ручной обработкой древесины? (Столяр).
— Какие виды инструментов вы знаете? Приведите пример.
(Режущие: ножовка, пила, дрель, рубанок; измерительные: линейка, угольник, циркуль, транспортир).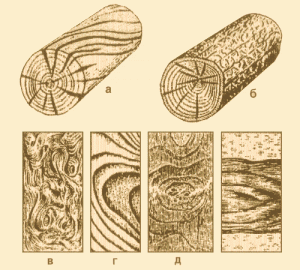
— Какие древесные породы вам известны? Приведите пример.
(Лиственные: береза, осина, дуб; хвойные: ель, сосна, пихта).
— Перечислите свойства древесины.
(Высокая прочность, хорошо обрабатывается инструментами, деревянные детали легко склеиваются, соединяются гвоздями и шурупами, деревянные детали имеют красивый внешний вид).
— Есть ли у древесины недостатки? Если есть, то какие?
(Портится от сырости, коробится при высыхании, легко горит).
3. Проверка домашнего задания.
Повторим пороки древесины, задавая друг другу вопросы. Вопросы учащиеся готовили друг другу дома. Задавая вопросы, они должны не только правильно их задать, но и внимательно выслушать ответ.
Вопросы могут быть следующего содержания:
- Отклонения древесины от нормального состояния, внешнего вида и формы, а также повреждения древесины (пороки).
- Винтообразное расположение волокон в древесине (косослой).

- Разрыв древесины вдоль волокон от морозов, жары, при сушке (трещина).
- Повреждение древесины в виде отверстий, прогрызаемых личинками и жучками (червоточины).
- Части ветвей, заключенные в древесине ствола (сучки).
- Поражение древесины паразитирующими грибами (гниль).
Самостоятельная работа с тестами.
1. Технология это:
1. наука, посвященная изучению природы;
2. наука, посвященная изучению общества;
3. наука о преобразовании материалов, энергии и информации;
4. наука о строение материи.
2. Изготовление детали из древесины начинается:
1. с разметки заготовки;
2. с выбора заготовки;
3. с обработки заготовки;
4. с зачистки заготовки.
3. Твердой лиственной породой является:
1. липа;
2. дуб;
3. береза;
4. осина.
осина.
4. Мягкой лиственной породой является:
1. бук;
2. клен;
3. береза;
4. дуб.
5. Для чернового строгания древесины используют:
1. рубанок деревянный;
2. фуганок;
3. шерхебель;
4. рубанок ручной электрический.
6. Сведения о процессе изготовления изделий приведены на:
1. чертеже изделия;
2. техническом рисунке;
3. на сборочном чертеже;
4. на технологической карте.
7. Ровные и гладкие поверхности из древесины получают при помощи:
1. лучковой пилы;
2. ножовки;
3. рубанка;
4. двуручной пилы.
8. Процесс создания мозаики по дереву – это:
1. выпиливание;
2. полирование;
3. инкрустация;
4. резьба.
9. Видом художественной обработки древесины является:
1. сверление
2. пиление
пиление
3. строгание
4. выжигание
5. Далее учащиеся демонстрируют свои работы, сделанные из древесины (разделочные доски) и рассказывают о последовательности их выполнения.
Каждый учащийся должен уметь назвать последовательность изготовления разделочной доски:
- Выбор заготовки, строгание.
- Разметка заготовки по шаблону.
- Выпиливание по разметке, сверление отверстия.
- Шлифовка.
- На одной стороне доски делается рисунок, покрывается лаком.
Дети отвечают на вопрос:
Где используется данное изделие?
В быту, для разделки различных продуктов.
6. Подведение итогов, самоанализ свой работы на уроке самими учащимися, выставление оценок.
7. Домашнее задание.
Составить кроссворд по теме: «Древесина. Обработка древесины».
#самоизоляция — источник.
 .. — Экология Камчатки
.. — Экология КамчаткиКак зимуют растения?🌿❄️
Наконец-то у нас выдалась снежная зима. И сейчас, когда травянистые растения и многие кустарники скрылись под снегом, самое время поговорить о том, как они зимуют.
Датский ботаник К. Раункиер по способу перезимовки разделил все растения на следующие типы:
1) фанерофиты, 2) хамефиты, 3) гемикриптофиты, 4) геофиты (криптофиты) и 5) терофиты.
☘️1 Фанерофиты — это особая группа, в которую входят лишенные защиты растения, перезимовывающие почки которых значительно возвышаются над поверхностью почвы и снежным покровом. Это все наши деревянистые растения: деревья и кустарники, большинство которых сбрасывает свои листья, хотя некоторые, например, хвойные, зимуют в зеленом виде. В умеренном климате фанерофитов по сравнению с другими типами очень мало. Они составляют всего лишь около 8% всей нашей флоры.
☘️2. Хамефиты или, иначе, карликовые растения — это обычные обитатели северных территорий. Хамефиты имеют зимующие почки, расположенные невысоко над поверхностью почвы.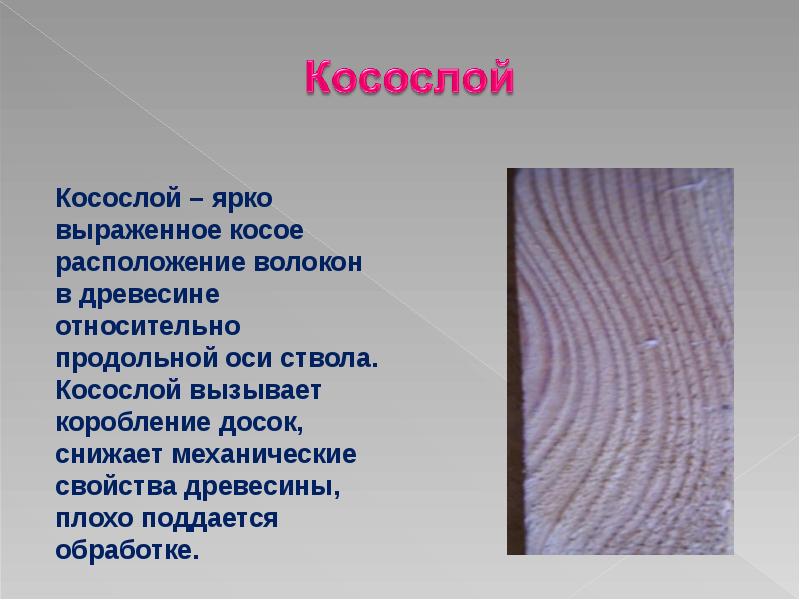 В чем же преимущество такого приспособления? После схода снежного покрова поверхностный слой почвы и поверхность самих растений начинает сильно прогреваться солнечными лучами, а почва даже на незначительной глубине еще остается мерзлой и задерживает развитие растений. Поэтому приподнятые над поверхностью почвы побеги с почками получают некоторое дополнительное количество тепла и начинают раньше развиваться. Ведь вегетативный сезон на севере так короток! А карликовый рост дает возможность растениям прятаться под защиту снежного покрова.
В чем же преимущество такого приспособления? После схода снежного покрова поверхностный слой почвы и поверхность самих растений начинает сильно прогреваться солнечными лучами, а почва даже на незначительной глубине еще остается мерзлой и задерживает развитие растений. Поэтому приподнятые над поверхностью почвы побеги с почками получают некоторое дополнительное количество тепла и начинают раньше развиваться. Ведь вегетативный сезон на севере так короток! А карликовый рост дает возможность растениям прятаться под защиту снежного покрова.
☘️3. Гемикриптофиты (в переводе означает «наполовину скрытые растения») это растения, зимующие почки которых располагаются обычно близко в поверхности или даже выступают над уровнем почвы. В наших лесах, особенно на лугах, они широко распространены. Среди гемикриптофитов есть виды с отмирающей на зиму наземной частью, а есть и с зимующей. Многие из них имеют прикорневые листовые розетки. К числу таких растений относится около половины всей нашей флоры.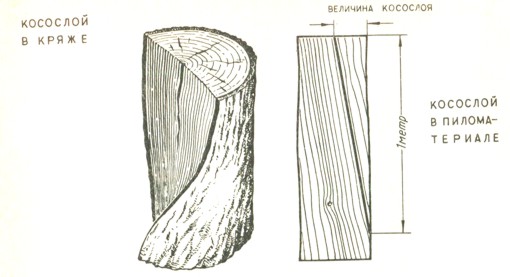
☘️4. Геофиты (криптофиты) — в переводе — «земляные растения». Это многолетние растения, почки возобновления которых находятся на подземных органах, скрытых в почве или под водой. Для этого у них есть подземные органы – клубни, корневища. Растения этого типа очень обычны в наших лесах и на лугах. Все эти растения относятся к числу летнезеленых, т.е. у них надземные побеги сохраняются лишь в течение летнего сезона и на зиму отмирают.
☘️5. Терофиты неблагоприятный период года переживают на стадии семян, наземные и подземные органы у них отмирают. То есть это однолетние растения. В большей степени они распространены в степях и пустынях, там, где нет снежного покрова. Но и у нас есть такие однолетники, например, в семействе капустовых и маковых.
Фото Ирины Щегловой
|
Пороки древесины |
Нормы ограничения пороков в пиломатериалах для сортов |
||||
|
отборного |
1-го |
2-го |
3-го |
4-го |
|
|
2. |
|||||
|
2.1. Пластевые и кромочные, в том числе выходящие на торец |
Допускаются длиной в долях длины пиломатериала, не более: |
Допускаются при условии сохранения целостности пиломатериала |
|||
|
Неглубокие |
Неглубокие и глубокие |
||||
|
1/6 |
1/4 |
1/3 |
1/2 |
||
|
Глубокие |
|||||
|
1/10 |
1/6 |
||||
|
2.2. Пластевые сквозные, в том числе выходящие на торец |
Допускаются длиной в мм, не более: |
Допускаются общей длиной в долях длины пиломатериала, не более: |
|||
|
100 |
150 |
200 |
|||
|
1/6 |
1/4 |
||||
|
2. |
Не допускаются |
Допускаются на одном торце длиной в долях ширины пиломатериала,не более: |
Допускаются при условии сохранения целостности пиломатериала |
||
|
1/4 |
1/3 |
1/2 |
|||
|
Примечание. Допускаемые размеры трещин установлены для пиломатериала с влажностью древесины не более 22%, при большей влажности эти размеры трещин уменьшаются вдвое. |
|||||
|
3. Пороки строения древесины |
|||||
|
3.1. Наклон волокон |
Допускается не более 5% |
Допускается |
|||
|
3.2. Крень |
Не допускаются |
Допускается не более 20% площади пласти пиломатериала |
Допускается |
||
|
3. |
Односторонние на любом однометровом участке длины в количестве 1 шт. длиной не более 50 мм |
Допускаются на любом однометровом участке длины пиломатериала в шт., не более: |
Допускаются |
||
|
2 |
4 |
||||
|
3.4. Сердцевина и двойная сердцевина |
Не допускается |
Допускается без отлупных и радиальных трещин только в пиломатериалах толщиной 40 мм и более |
Допускается |
||
|
3.5. Прорость |
Не допускается |
Допускается односторонняя шириной в долях соответствующей стороны пиломатериала, не более: |
Допускается |
||
|
1/10 |
1/5 |
1/4 |
|||
|
и длиной в долях длины пиломатериала, не более: |
|||||
|
1/20 |
1/10 |
1/10 |
|||
|
3. |
Не допускается |
Допускается протяжением в долях длины пиломатериала до: |
Допускается |
||
|
1/5 |
1/3 |
||||
|
но не более 1 м. |
|||||
|
4. Грибные поражения |
|||||
|
4.1. Грибные ядровые пятна (полосы) |
Не допускаются |
Допускаются общей площадью в % от площади пиломатериала, не более: |
Допускаются |
||
|
10 |
20 |
||||
|
4.2. Заболонные грибные окраски и плесень |
Не допускаются |
Допускаются поверхностные в виде пятен и полос. |
Допускаются |
||
|
10 |
20 |
50 |
|||
|
4.3. Гнили |
Не допускаются |
Не допускаются |
Допускается только пестрая ситовая ядровая гниль в виде пятен и полос-общей площадью не. более 10% площади пиломатериала |
||
|
5. Биологические повреждения |
|||||
|
5.1. Червоточина |
Допускается неглубокая на обзольных частях пиломатериала |
Допускается на любом однометровом участке длины пиломатериала в шт., не более: |
|||
|
2 |
3 |
6 |
|||
|
6. |
|||||
|
6.1. Инородные включения (проволока, гвозди, металлические осколки и др.) |
Не допускаются |
||||
|
6.2. Обзол (в обрезных пиломатериалах) |
Острый не допускается |
Допускается ту поп и острый при условии, что пластb пропилены не менее, чем на 1/2ширины, а кромки не менее, чем на 3/4 длины пиломатериала. |
|||
|
Тупой допускается на .пластях и кромках размером в долях ширины соответствующих сторон .пиломатериала без ограничения по длине, не более: |
|||||
|
1/6 |
1/6 |
1/6 |
1/3 |
||
|
Допускается на отдельных участках кромок размером в долях ширины кромки, не более: |
|||||
|
1/3 |
1/3 |
1/3 |
2/3 |
||
|
и протяженностью в долях длины пиломатериала, не более: |
|||||
|
1/6 |
1/6 |
1/6 |
1/4 |
||
|
Примечания: · Кора на обзолах экспортных пиломатериалов не допускается. · Обрезные пиломатериалы, соответствующие по всем показателям требованиям определенного сорта, но с обзолом, превышающим установленную норму для этого сорта, допускается переводить в необрезные с сохранением сортности. |
|||||
|
6.3. Скос пропила |
В пиломатериалах один торец (в экспортных пиломатериалах – оба торца) должен быть опилен перпендикулярно к продольной оси пиломатериала. Отклонение от перпендикулярности торца к пласти и кромке допускается до 5% ширины и толщины пиломатериала соответственно. |
||||
|
6.4. Взрывы, накол |
Допускаются в пределах отклонении от номинальных размеров, установленных в ГОСТ 24454—80 |
Допускаются глубиной в 3 мм, не более |
Допускаются |
||
|
Примечание. Наколы нормируются только в экспортных пиломатериалах. |
|||||
|
7. Покоробленности |
|||||
|
7.1. Покоробленность продольная по пласти и кромке, крыловатость |
Допускается стрела прогиба в долях длины пиломатериала в %, не более: |
Допускаются |
|||
|
0,2 |
0,2 |
0,2 |
0,4 |
||
|
Примечание. В необрезных пиломатериалах продольная покоробленность по кромке не нормируется. |
|||||
|
7.2. Покоробленность поперечная |
Допускается стрела прогиба в долях ширины пиломатериала в %, не более: |
Допускаются |
|||
|
1 |
1 |
1 |
2 |
||
|
Примечания: · Нормы покоробленности установлены для пиломатериалов с влажностью не более 22%. · Пороки древесины, не упомянутые в настоящем стандарте, допускаются. |
|||||
 Трещины
Трещины 3. Торцовые (кроме трещин усушки)
3. Торцовые (кроме трещин усушки) 3. Кармашки
3. Кармашки 6. Рак
6. Рак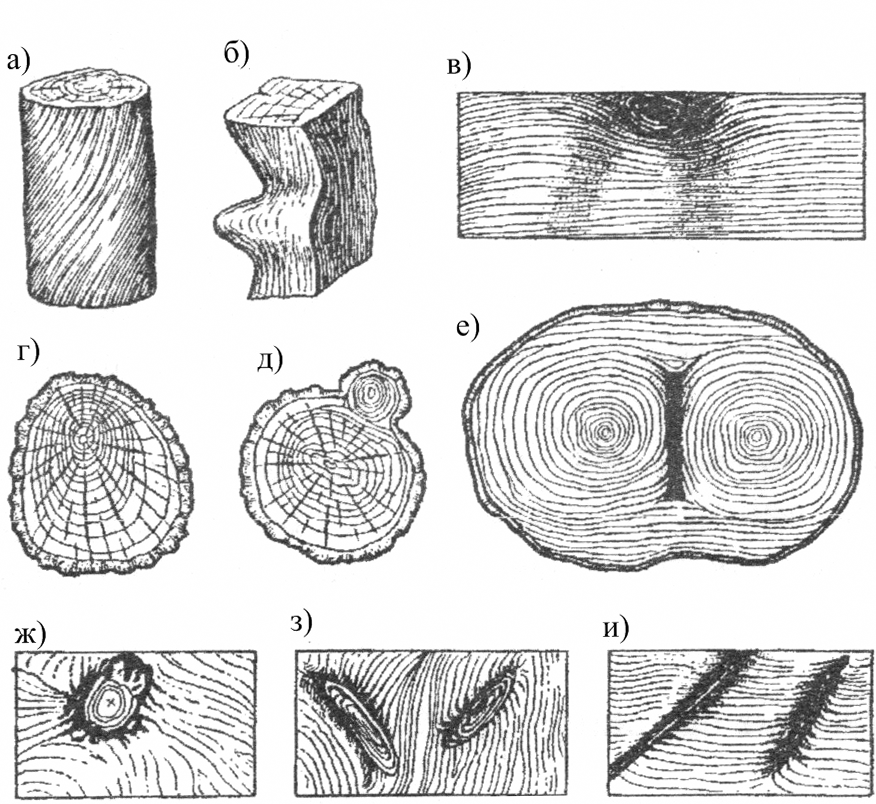 Инородные включения, механические повреждения, и пороки обработки
Инородные включения, механические повреждения, и пороки обработки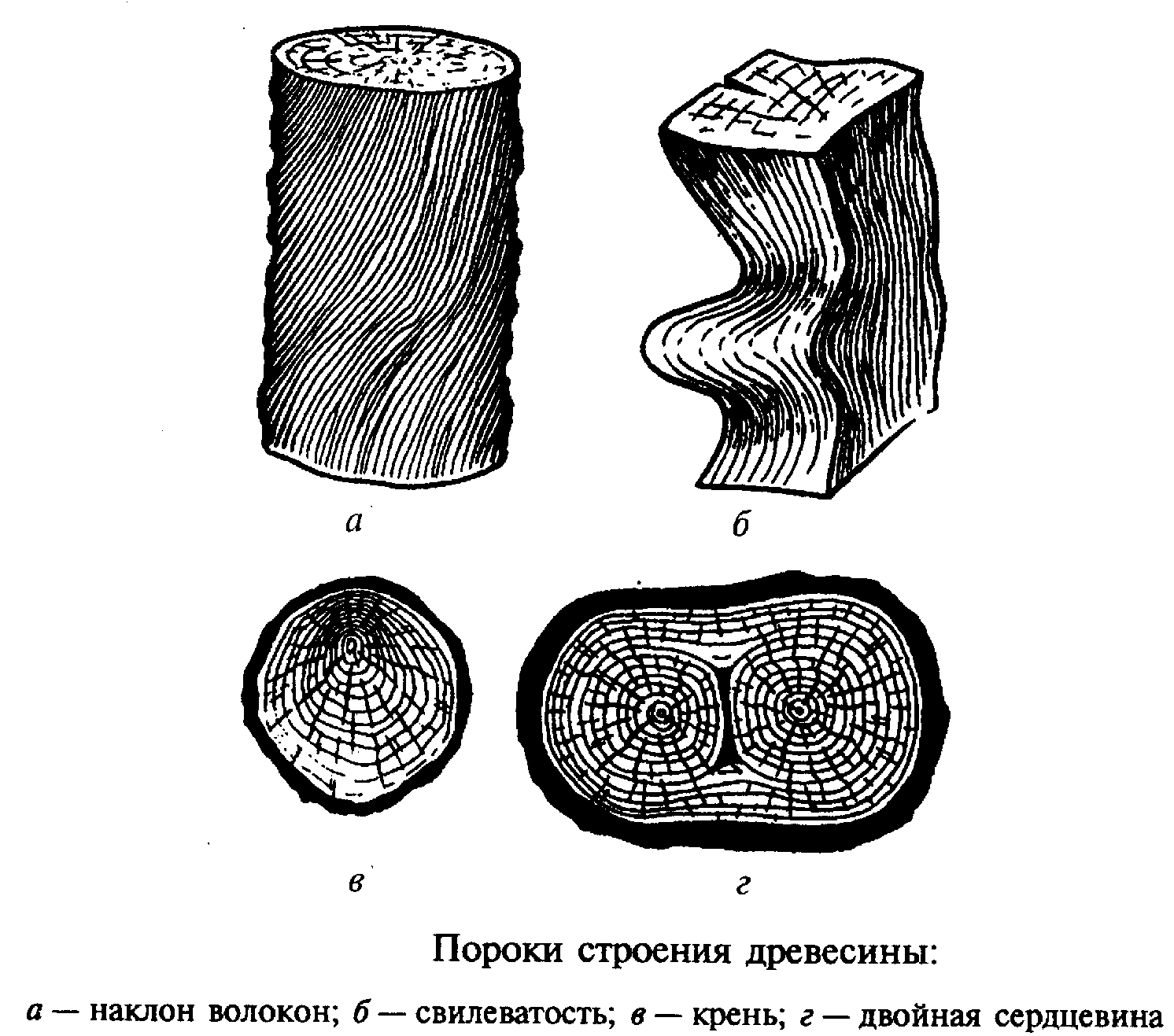
Пороки древесины
Навигация:
Главная → Все категории → Строительное материаловедение
Пороками древесины называют изъяны, возникающие на стадиях роста или хранения дерева, что непосредственно снижает качество и ограничивает использование лесоматериалов в строительных конструкциях. Их принято подразделять на ряд специфических групп: сучки, трещины, пороки формы ствола, пороки строения древесины, химические окраски, грибные поражения, повреждения насекомыми, инородные включения, деформации. Пороки растущего дерева обычно переходят в срубленную древесину. В ней возникают и собственные пороки — грибные поражения, химические окраски и Др. Полная характеристика пороков и их влияния на качество древесины приведены в ГОСТ 2140—81.
Сучки — живые и отмершие, закрытые и заросшие, здоровые и загнившие представляют собой части (основания) ветвей, заключенные в древесине (рис. 7.13). Они нарушают однородность строения древесины, затрудняют механическую обработку пиломатериалов, нарушают закономерность распределения внутренних напряжений, уменьшают рабочее сечение пиломатериала и, следовательно, прочность древесины, ее сортность. Нередко прочность снижается на 50% и более, а в тонких досках и брусьях — еще выше.
7.13). Они нарушают однородность строения древесины, затрудняют механическую обработку пиломатериалов, нарушают закономерность распределения внутренних напряжений, уменьшают рабочее сечение пиломатериала и, следовательно, прочность древесины, ее сортность. Нередко прочность снижается на 50% и более, а в тонких досках и брусьях — еще выше.
Рис. 7.13. Сучки различных видов: а — здоровый; б — роговой; в — выпадающий
Для несущих деревянных конструкций допускается применять древесину без сучков или со здоровыми сросшимися сучками, количество и размеры которых ограничены для каждого сорта материала. Сучки — основные сортоопределяющие факторы у древесины. Особенно вредны выпадающие, рыхлые и табачные несросшиеся сучки.
Ненормальные окраски и гнили возникают под влиянием различных причин в период роста или хранения ствола и имеют различную степень вредности. Среди этой разновидности пороков наиболее выделяются: ложное ядро — встречается чаще всего у бука, березы и других заболонных и спелодревесных пород, иногда располагается эксцентрично. Оно отличается от настоящего ядра древесины по времени образования, форме и размерам. Ложное ядро не оказывает значительного негативного влияния, если отсутствуют признаки загнивания, но может нарушить общую фактуру изделия. Пятнистость (радиальная и тангенциальная) наблюдается на торце в виде пятен шириной не больше годичного слоя, вытянутых на радиальном срезе незначительно в виде узких полос, а на тангенциальном — в виде широких полос, вытянутых вдоль ствола, иногда на несколько метров. Этот порок ухудшает внешний вид пиломатериалов, не влияя на их механические свойства.
Оно отличается от настоящего ядра древесины по времени образования, форме и размерам. Ложное ядро не оказывает значительного негативного влияния, если отсутствуют признаки загнивания, но может нарушить общую фактуру изделия. Пятнистость (радиальная и тангенциальная) наблюдается на торце в виде пятен шириной не больше годичного слоя, вытянутых на радиальном срезе незначительно в виде узких полос, а на тангенциальном — в виде широких полос, вытянутых вдоль ствола, иногда на несколько метров. Этот порок ухудшает внешний вид пиломатериалов, не влияя на их механические свойства.
Ложное ядро и пятнистость характерны для растущего дерева.
Однако изменение окраски нередко наблюдается и у срубленной древесины при ее хранении. В частности, к ним относятся химические и грибные окраски.
К химическим окраскам относятся желтизна, оранжевая окраска, продубина, чернильные пятна, дубильные потеки. Окраска проникает на глубину 1—5 мм, но мало влияет на физико-механические свойства древесины, ухудшая только внешний вид пиломатериалов.
Грибные окраски (гнили) поражают отмирающую и мертвую древесину. На отмирающей при медленном подсыхании образуется прелость вследствие поражения складскими грибами: плесень, цветной налет глубиной 1—2 мм, цветная заболонь, синева, коричневая или желтая окраска (в виде лимонно-желтых пятен и полос в отличие от химической окраски). Эти окраски развиваются за счет содержимого реагента.
Более опасными являются изменения окраски древесины, связанные с формированием гнили у растущего дерева и мертвой древесины.
Гнили у растущего дерева — пестрая, белая и бурая — образуются под воздействием грибов. Белая, пестрая (коррозийные) и бурая (деструктивная) гнили развиваются постепенно с появлением пятен и полос до появления мягкой гнили, имеющей трухлявово-локнистую структуру и легко ломающейся пальцами. Гнилая древесина почти полностью теряет свои позитивные механические свойства, сильно понижается плотность; она непригодна к использованию в строительстве.
Гнили мертвой древесины являются одним из самых опасных пороков. Они могут развиваться после вторичного увлажнения в лежалой древесине и открытых деревянных сооружениях. Особенно распространена бурая деструктивная гниль, вызываемая грибами заборным, столбовым, шпальным й др. Древесина становится бурой, появляются поперечные и продольные трещины; древесина быстро приобретает трухлявость и распадается на куски.
Они могут развиваться после вторичного увлажнения в лежалой древесине и открытых деревянных сооружениях. Особенно распространена бурая деструктивная гниль, вызываемая грибами заборным, столбовым, шпальным й др. Древесина становится бурой, появляются поперечные и продольные трещины; древесина быстро приобретает трухлявость и распадается на куски.
В закрытых, плохо проветриваемых помещениях с высокой влажностью и умеренной температурой развивается бурая деструктивная гниль под воздействием так называемых домово-шахтных грибов, настоящего домового гриба (грибница ватообразная с каплями желтоватой жидкости), пленочного домового гриба (плодовое тело пленчатое распростертое), белого домового гриба (плодовые тела белые, мягкие, распростертые с большими угловатыми порами), шахтного или пластинчатого домового гриба (развивается в подземных сооружениях).
Эти разновидности грибов делают древесину непригодной для применения, опасной для окружающих древесных материалов.
Водослой — участок древесины, сильно пропитанный водой, расположенный на месте ядра, спелой древесины, ложного ядра или радиальной пятнистости. Как правило, водослой связан с очагами гнили.
Как правило, водослой связан с очагами гнили.
Водослойная древесина отличается повышенными усушкой и разбуханием; после высыхания образуются трещины.
Грибные ядровые пятна и полосы — измененная окраска в зоне ядра, вызываемая грибами в растущем дереве, наблюдается на торцах и продольном разрезе. Этот порок называют также темнина или краснина. Ухудшая внешний вид пиломатериалов, на механические свойства почти не оказывает заметного влияния.
Значительное влияние на снижение механической прочности древесины оказывают пороки, именуемые трещинами и представляющие собой разрывы древесины вдоль волокон. Они характерны как для растущего дерева, так и для мертвой древесины, хотя их вид и размеры различные.
Трещины продольные — метиковые, простые и сложные. Эти трещины проходят через сердцевину ствола, находясь обычно в пределах зоны ядра или спелой древесины. Они идут по длине ствола от комля до зоны живых сучков, расширяясь от периферийной части ствола к сердцевине.
Простые метиковые трещины расположены по одному диаметру и идут по длине ствола в одной плоскости; сложные — по одному диаметру, но по спирали или по нескольким диаметрам (тогда по длине ствола идут по нескольким плоскостям).
Отлупные трещины (неполные и кольцевые) — внутренние трещины, проходящие в зоне ядра или спелой древесины между годичными слоями. Отлуп как порок особенно часто встречается у дуба, осины, пихты, тополя, ели. Как и метиковые, отлупные трещины образуются при раскачивании дерева ветром и при резкой смене температуры.
Метики и отлуп, возникнув в растущем дереве, увеличиваются в своих размерах в срубленном дереве при его высыхании.
Морозные трещины — открытые радиальные трещины в растущем дереве, видимые на поверхности ствола (чаще в комлевой части дерева) и распространяющиеся до сердцевины. Возникают зимой при резком охлаждении стволов. Они поражают толстые стволы клена, бука, дуба, ясеня, а иногда осину и липу.
Рис. 7.14. Метиковые трещины в бревне.
7.14. Метиковые трещины в бревне.
Кривизна ствола — искривление по длине ствола; бывает простая кривизна —- характеризуется одним изгибом по длине ствола и сложная — несколькими изгибами.
Закомелистостъ — резкое увеличение диаметра комлевой части (в 1—2 раза и более) по сравнению с диаметром сортамента, измеренным на расстоянии 1 м от комлевого торца (частый случай сбе-жистости). Этот порок также уменьшает выход пиломатериалов и шпона.
Пороки строения древесины. Наклон волокон (косослой) — волокна располагаются не параллельно оси ствола, а винтообразно, по спирали. Косослой оказывает отрицательное влияние на качество древесины, ее физико-механические свойства. Наклон волокон более 5% значительно снижает предел прочности древесины при растяжении вдоль волокон и поперечном изгибе. С изменением влажности древесина с наклоном волокон склонна к значительному короблению и скручиванию.
Крень бывает сплошная и местная. Характерна для комлевой части наклонных и искривленных стволов хвойных пород. Это резкое утолщение поздней зоны годичных слоев при эксцентричном расположении сердцевины.
Это резкое утолщение поздней зоны годичных слоев при эксцентричном расположении сердцевины.
Плотность древесины сильно развитой крени на 15—40% выше нормальной (кремнина) мешает механической обработке древесины.
Свилеватость (волнистая и путаная) — неправильность в строении древесины, резковолнистое или беспорядочное расположение древесных волокон. Встречается преимущественно у лиственных пород в нижней комлевой части ствола (у осины, клена, ясеня, березы, тополя и др.). Свилеватость снижает прочность древесины при растяжении, изгибе, сжатии, но увеличивает прочность при скалывании. Свилеватую древесину используют в качестве отделочного строительного материала.
Повреждения насекомыми (червоточина). Червоточиной называют повреждения древесины в виде поверхностных бороздок, внутренних ходов (каналов) и отверстий, проделанных насекомыми, а чаще всего их личинками (жуками-короедами, жуками-усачами, долгоносиками, домовыми жуками-точильщиками, некоторыми разновидностями муравьев, бабочками (древоточцами и стеклянницами), термитами и др. ).
).
Степень пораженности древесины этим пороком определяют количеством отверстий на 1 м лесоматериала, а в фанере — количеством отверстий на один лист.
Кроме того, следует отметить дефекты, которые возникают в основном при неправильных режимах обработки древесины.
Трещины усушки образуются в срубленной древесине и пиломатериалах при несоблюдении установленного режима сушки. Различают трещины торцовые, пластевые, кромочные, сквозные. Трещины усушки имеют меньшую толщину, чем метиковые и морозные, но также снижают прочность и сортность древесины.
Покоробленность относится к деформациям, возникающим при распиловке или сушке древесины; она затрудняет или даже полностью исключает использование пиломатериалов.
Механические повреждения при обработке древесины — отщеп, скол, вырыв и другие также затрудняют использование ее по назначению. Сюда же относятся раны, например, огневые, паразитарные — механические, сухостойкость, пожарная подсушина и др.
Похожие статьи:
Строительные термины и определения
Навигация:
Главная → Все категории → Строительное материаловедение
Статьи по теме:
Главная → Справочник → Статьи → Блог → Форум
Произошла ошибка при установке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Сходства и различия в композитных приложениях
Работа над композитами из целлюлозного волокна обычно строго разделена на две отдельные области исследований в зависимости от происхождения волокна, то есть из древесины и из однолетних растений, представляющих две разные отрасли лесного и сельского хозяйства. соответственно. В данной статье проводится параллельная оценка древесных волокон и растительных волокон, чтобы выявить их сходства и различия в отношении их использования в качестве армирующих материалов в композитах, а также для обеспечения взаимной передачи знаний и технологий между двумя областями исследований.В статье дается введение в морфологию, химию и ультраструктуру волокон, моделирование механических свойств волокон, волоконные заготовки, используемые для производства композитов, типичные механические свойства композитов, моделирование механических свойств.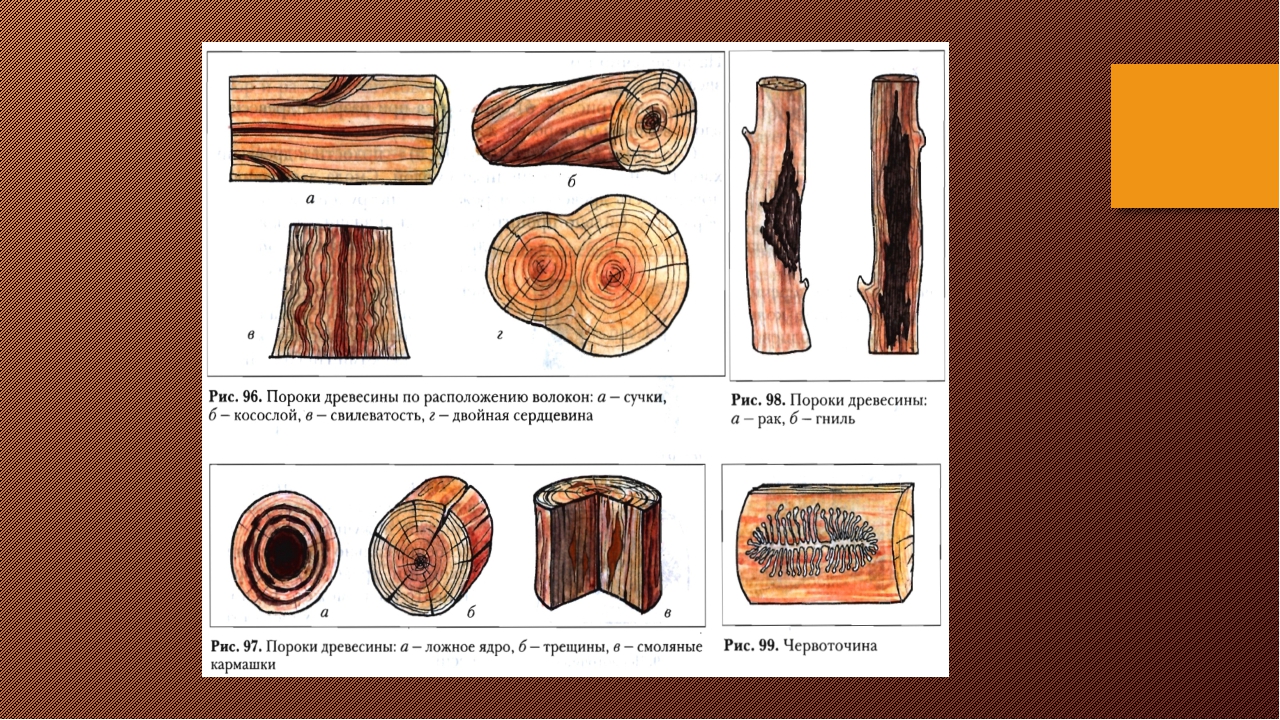 с упором на композиты, имеющие случайную ориентацию волокон и значительную пористость, и, наконец, чувствительность композитов к влаге. Характеристики композитов из древесных и растительных волокон сравниваются с синтетическими стеклянными и углеродными волокнами, обычно используемыми для композитов, и обсуждаются преимущества и недостатки различных волокон.
с упором на композиты, имеющие случайную ориентацию волокон и значительную пористость, и, наконец, чувствительность композитов к влаге. Характеристики композитов из древесных и растительных волокон сравниваются с синтетическими стеклянными и углеродными волокнами, обычно используемыми для композитов, и обсуждаются преимущества и недостатки различных волокон.
1. Введение
Композиты на основе целлюлозных волокон древесины и растений представляют собой относительно новый и многообещающий класс композиционных материалов [1–4]. Они безвредны для окружающей среды и обладают хорошими техническими характеристиками. Для нескольких несущих нагрузок, где обычно используются композиты из стекловолокна или углеродного волокна, композиты из целлюлозного волокна могут быть достойной альтернативой. Это особенно актуально для приложений, где экологические преимущества (возобновляемость, биоразлагаемость) играют важную роль, а высокие механические свойства не являются основной мотивацией. За последнее десятилетие было собрано огромное количество научной литературы по целлюлозным волокнам для применения в композитах (например, см. Недавние обзоры [5–8]), хотя публикации, как правило, делятся на две отдельные области в зависимости от происхождения волокон. , то есть из древесины или однолетних растений. Причина этого разделения, возможно, заключается в том, что производители сырья ищут новые рынки для своих волокон (тяга к технологиям), а конечным потребителям (тяга рынка) еще предстоит использовать потенциал целлюлозных волокон, независимо от их происхождения. волокна.Производители сырья в данном случае, то есть лесное хозяйство для производства древесных волокон и сельское хозяйство для производства растительных волокон, разработали свои специальные технологии в цепочке создания стоимости для производства волокон в зависимости от традиционного использования волокон. Для производства древесного волокна были построены целлюлозные заводы, которые производят сырье для производства бумаги и картона.
За последнее десятилетие было собрано огромное количество научной литературы по целлюлозным волокнам для применения в композитах (например, см. Недавние обзоры [5–8]), хотя публикации, как правило, делятся на две отдельные области в зависимости от происхождения волокон. , то есть из древесины или однолетних растений. Причина этого разделения, возможно, заключается в том, что производители сырья ищут новые рынки для своих волокон (тяга к технологиям), а конечным потребителям (тяга рынка) еще предстоит использовать потенциал целлюлозных волокон, независимо от их происхождения. волокна.Производители сырья в данном случае, то есть лесное хозяйство для производства древесных волокон и сельское хозяйство для производства растительных волокон, разработали свои специальные технологии в цепочке создания стоимости для производства волокон в зависимости от традиционного использования волокон. Для производства древесного волокна были построены целлюлозные заводы, которые производят сырье для производства бумаги и картона. Что касается растительных волокон, текстильные технологии совершенствуются для производства пряжи и тканей. Ввиду того, что область исследований в области целлюлозных волокон смещается в сторону достижения технических требований конечных пользователей, и довольно независимого существования сообществ исследователей древесных и растительных волокон, соответственно, эта статья была написана, чтобы пролить дополнительный свет на сходства и различия этих двух типов целлюлозных волокон (древесного и растительного происхождения) с точки зрения промышленного использования для производства композитов из целлюлозных волокон для структурного применения.Характеристики волокон также будут сравниваться с характеристиками основных современных конкурентов, то есть композитов, изготовленных из синтетических волокон, таких как стеклянные и углеродные волокна. Сходства и различия различных волокон для композитов будут обсуждаться в свете будущих возможностей в инженерных приложениях. Приближая дисциплины науки о древесине и растительном волокне, можно было бы надеяться на взаимную передачу знаний, поскольку эти две области исследований развивались довольно независимо и, таким образом, достигли разных уровней понимания в отношении различных аспектов, таких как методы описания.
Что касается растительных волокон, текстильные технологии совершенствуются для производства пряжи и тканей. Ввиду того, что область исследований в области целлюлозных волокон смещается в сторону достижения технических требований конечных пользователей, и довольно независимого существования сообществ исследователей древесных и растительных волокон, соответственно, эта статья была написана, чтобы пролить дополнительный свет на сходства и различия этих двух типов целлюлозных волокон (древесного и растительного происхождения) с точки зрения промышленного использования для производства композитов из целлюлозных волокон для структурного применения.Характеристики волокон также будут сравниваться с характеристиками основных современных конкурентов, то есть композитов, изготовленных из синтетических волокон, таких как стеклянные и углеродные волокна. Сходства и различия различных волокон для композитов будут обсуждаться в свете будущих возможностей в инженерных приложениях. Приближая дисциплины науки о древесине и растительном волокне, можно было бы надеяться на взаимную передачу знаний, поскольку эти две области исследований развивались довольно независимо и, таким образом, достигли разных уровней понимания в отношении различных аспектов, таких как методы описания. , обработка волокна, обработка преформ волокна и производство композитов.
, обработка волокна, обработка преформ волокна и производство композитов.
Статья представляет собой введение в (i) морфологию, химию и ультраструктуру волокон, (ii) моделирование механических свойств волокон, (iii) заготовки волокна, доступные для производства композитов, (iv) типичные механические свойства композитов, (v) моделирование механических свойств композитов с особым акцентом на композитах, имеющих случайную ориентацию волокон и значительную пористость, и (vi) чувствительность композитов к влаге.Кроме того, приводятся примеры применения новых композитов, а затем рассматриваются будущие перспективы использования древесных и растительных волокон для производства композитов из целлюлозных нановолокон. Наконец, дается краткое описание различий и вытекающих из этого преимуществ двух типов целлюлозных волокон, а именно древесных и растительных волокон, а также различия и вытекающие из них преимущества целлюлозных волокон по сравнению с синтетическими волокнами.
2.
 Морфология, химия и ультраструктура волокон
Морфология, химия и ультраструктура волокон Plantae — одно из пяти царств живых организмов, которое включает зеленые растения, то есть мхи, папоротники, голосеменные растения (например, мхи, папоротники, голосеменные).g., древесина хвойных пород) и покрытосеменных (например, лиственных и однолетних растений). Клетки зеленых растений окружены жесткой клеточной стенкой , , и это основная характеристика, отличающая их от клеток животных. В некоторых типах клеток клеточные стенки увеличены, чтобы иметь превосходные механические свойства, которые обеспечивают требуемые структурные характеристики растений. Размеры этих так называемых волокон различаются у разных растений, но их общая форма чаще всего имеет удлиненную длину в диапазоне 1–50 мм и диаметр в диапазоне 15–30 мкм м.С точки зрения композитного армирования волокна удобно сгруппировать по длине. (I) Короткие волокна (1–5 мм), обычно происходящие из пород древесины (например, ели, сосны, березы, эвкалипта) и обычно используемые для изготовления композитов с плоскими изотропными свойствами, то есть композитов с неспецифической (случайной) ориентацией волокон. (ii) Длинные волокна (5–50 мм), обычно получаемые из однолетних видов растений (например, лен, конопля, джут) и обычно используемые для изготовления композитов с анизотропными свойствами, то есть композитов с определенной ориентацией волокон.У живых зеленых растений, когда волокна полностью развиты, их внутриклеточные органеллы начинают дегенерировать, в результате чего волокна имеют пустую центральную полость, так называемый просвет. В древесных волокнах площадь просвета составляет 20–70% площади поперечного сечения волокна [9]. Напротив, однолетние растительные волокна, такие как конопля и лен, имеют относительно меньшую площадь просвета в диапазоне 0–5% [3, 10].
(ii) Длинные волокна (5–50 мм), обычно получаемые из однолетних видов растений (например, лен, конопля, джут) и обычно используемые для изготовления композитов с анизотропными свойствами, то есть композитов с определенной ориентацией волокон.У живых зеленых растений, когда волокна полностью развиты, их внутриклеточные органеллы начинают дегенерировать, в результате чего волокна имеют пустую центральную полость, так называемый просвет. В древесных волокнах площадь просвета составляет 20–70% площади поперечного сечения волокна [9]. Напротив, однолетние растительные волокна, такие как конопля и лен, имеют относительно меньшую площадь просвета в диапазоне 0–5% [3, 10].
Основным химическим компонентом клеточной стенки является целлюлоза, которая представляет собой неразветвленный полисахаридный полимер, состоящий из звеньев глюкозы.Для древесных волокон средняя длина целлюлозной цепи составляет 5 мкм, м, что соответствует степени полимеризации (т. Е. Единиц глюкозы) 10 000 [9]. Эта молекулярная линейность делает целлюлозу сильно анизотропной с теоретической жесткостью и прочностью около 130 и 15 ГПа, соответственно, в направлении цепи [11]. Цепи целлюлозы расположены параллельно, образуя пучки, которые обозначены как микрофибриллы . В некоторых областях микрофибрилл молекулы глюкозы целлюлозных цепей расположены в виде высокоупорядоченной кристаллической структуры.Двумя другими основными химическими составляющими клеточной стенки являются гемицеллюлоза и лигнин. Гемицеллюлоза — это гетерогенная группа полисахаридов, характеризующаяся тем, что они короткие и разветвленные. Лигнин — это сильно разветвленный полимер, состоящий из фенилпропановых звеньев, организованных в сложную трехмерную структуру. В дополнение к организации трех химических компонентов, структурная сложность клеточной стенки увеличивается за счет организации ряда слоев, различающихся углом между микрофибриллами целлюлозы и продольной осью волокна.Угол наклона микрофибрилл целлюлозы в различных слоях, помимо относительной толщины слоев, определяет общие механические характеристики волокон.
Эта молекулярная линейность делает целлюлозу сильно анизотропной с теоретической жесткостью и прочностью около 130 и 15 ГПа, соответственно, в направлении цепи [11]. Цепи целлюлозы расположены параллельно, образуя пучки, которые обозначены как микрофибриллы . В некоторых областях микрофибрилл молекулы глюкозы целлюлозных цепей расположены в виде высокоупорядоченной кристаллической структуры.Двумя другими основными химическими составляющими клеточной стенки являются гемицеллюлоза и лигнин. Гемицеллюлоза — это гетерогенная группа полисахаридов, характеризующаяся тем, что они короткие и разветвленные. Лигнин — это сильно разветвленный полимер, состоящий из фенилпропановых звеньев, организованных в сложную трехмерную структуру. В дополнение к организации трех химических компонентов, структурная сложность клеточной стенки увеличивается за счет организации ряда слоев, различающихся углом между микрофибриллами целлюлозы и продольной осью волокна.Угол наклона микрофибрилл целлюлозы в различных слоях, помимо относительной толщины слоев, определяет общие механические характеристики волокон. Таким образом, в целом клеточная стенка древесины и растительных волокон по существу организована как композитный ламинат с рядом пластинок с по-разному ориентированными, жесткими и прочными микрофибриллами полукристаллической целлюлозы, встроенными в матрицу из гемицеллюлозы и лигнина.
Таким образом, в целом клеточная стенка древесины и растительных волокон по существу организована как композитный ламинат с рядом пластинок с по-разному ориентированными, жесткими и прочными микрофибриллами полукристаллической целлюлозы, встроенными в матрицу из гемицеллюлозы и лигнина.
В отличие от целлюлозных волокон синтетические волокна, которые традиционно используются для армирования композитов, таких как стеклянные и углеродные волокна, являются монолитными и имеют гораздо более простую ультраструктуру.Стекловолокно в основном состоит из молекул оксида кремния, организованных в аморфную конфигурацию. Углеродные волокна состоят из атомов углерода в графитовых слоях, которые организованы в пакетную турбостратную конфигурацию.
В таблице 1 приведены ключевые числа химического состава и ультраструктуры целлюлозных волокон. Содержание целлюлозы в необработанных волокнах находится в диапазоне от 40 до 50% по весу для древесных волокон и в диапазоне от 60 до 70% по весу для растительных волокон. Соответственно, содержание гемицеллюлозы и лигнина в древесных волокнах выше, и это особенно верно для лигнина, содержание которого в древесных волокнах составляет около 30 мас.% По сравнению с только примерно 5 мас.% В растительных волокнах.Химический состав древесных и растительных волокон явно отличается друг от друга. Кроме того, древесные волокна демонстрируют более низкую степень кристалличности целлюлозы, чем растительные волокна, с типичными значениями в диапазонах 55–70 и 90–95% соответственно. Угол микрофибрилл в древесных волокнах варьируется в диапазоне 3–50 ° в зависимости от типа и расположения волокон в древесине (например, поздняя и ранняя древесина) [12], тогда как угол микрофибрилл в растительных волокнах более постоянен в диапазон 6–10 ° [13].
Соответственно, содержание гемицеллюлозы и лигнина в древесных волокнах выше, и это особенно верно для лигнина, содержание которого в древесных волокнах составляет около 30 мас.% По сравнению с только примерно 5 мас.% В растительных волокнах.Химический состав древесных и растительных волокон явно отличается друг от друга. Кроме того, древесные волокна демонстрируют более низкую степень кристалличности целлюлозы, чем растительные волокна, с типичными значениями в диапазонах 55–70 и 90–95% соответственно. Угол микрофибрилл в древесных волокнах варьируется в диапазоне 3–50 ° в зависимости от типа и расположения волокон в древесине (например, поздняя и ранняя древесина) [12], тогда как угол микрофибрилл в растительных волокнах более постоянен в диапазон 6–10 ° [13].
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 %)
%) Влияние обработки по химическому составу волокон приведен в таблице 1. Как правило, как для древесных, так и для растительных волокон содержание целлюлозы увеличивается после обработки за счет удаления нецеллюлозных остатков волокон (например, пектинов и восков). Эффект технологической обработки наиболее отчетливо виден для сильно обработанных текстильных волокон конопли в исследовании Madsen et al. [14], где содержание целлюлозы достигло 91% мас. / Мас.
Как правило, как для древесных, так и для растительных волокон содержание целлюлозы увеличивается после обработки за счет удаления нецеллюлозных остатков волокон (например, пектинов и восков). Эффект технологической обработки наиболее отчетливо виден для сильно обработанных текстильных волокон конопли в исследовании Madsen et al. [14], где содержание целлюлозы достигло 91% мас. / Мас.
Влияние условий выращивания и обработки на химический состав и ультраструктуру целлюлозных волокон обычно приводит к волокнам с более разнообразными свойствами, чем у синтетических волокон.Это часто считается одним из основных недостатков использования целлюлозных волокон для армирования композитов. Однако считается, что это беспокойство вызвано общей неопределенностью в отношении причины изменчивости свойств и отсутствием системы классификации качества целлюлозных волокон, например, аналогичной системе, существующей для классификации твердых материалов. древесина. Следует также отметить, что изменчивость механических свойств волокон может положительно влиять на чувствительность к надрезам и трещиностойкость композитов [15, 16].
3. Моделирование механических свойств волокон
Микромеханические модели могут быть полезны для понимания того, как химический состав и ультраструктура целлюлозных волокон влияют на их механические свойства. С геометрической точки зрения клеточная стенка в целлюлозных волокнах может быть аппроксимирована слоями концентрических цилиндрических оболочек. На рисунке 1 показана такая идеализированная геометрия волокна. Резюме того, как ультраструктурные особенности клеточной стенки влияют на механические свойства волокон, были составлены Neagu et al.[17], Салмен и Бургерт [18] и Салмен [19].
В принципе, наиболее важными механическими свойствами волокон при использовании в композитах являются жесткость и прочность в осевом направлении, то есть в направлении длины волокна. Предполагается, что именно в этом направлении волокна несут нагрузку при использовании в композитах. В случае целлюлозных волокон ключевые ультраструктурные особенности, которые влияют на осевые механические свойства волокон, следующие: (i) Размер люмена . Только стенка ячейки несет нагрузку, то есть механические свойства волокна пропорциональны площади поперечного сечения стенки ячейки. Чем больше относительный размер просвета, тем ниже жесткость и прочность волокон. (Ii) Содержание целлюлозы. В исследовании Thygesen et al. [20] было обнаружено, что увеличение содержания целлюлозы в волокнах хорошо коррелирует с увеличением их жесткости и прочности. Кроме того, известно, что кристалличность целлюлозы и соотношение размеров кристаллитов влияют на жесткость клеточной стенки в направлении микрофибрилл (например,г., [21]). (iii) Угол из микрофибриллы . С помощью классической теории ламината (вращение ортотропной пластины в плоскости) можно продемонстрировать, что эффективные упругие свойства волокон в осевом направлении зависят от локальной жесткости в направлении микрофибрилл, умноженной на cos 4 , где — угол микрофибриллы. Таким образом, жесткость (и прочность) волокна очень чувствительна к углу наклона микроволокон, даже если механические свойства в направлении микроволокон постоянны.
Только стенка ячейки несет нагрузку, то есть механические свойства волокна пропорциональны площади поперечного сечения стенки ячейки. Чем больше относительный размер просвета, тем ниже жесткость и прочность волокон. (Ii) Содержание целлюлозы. В исследовании Thygesen et al. [20] было обнаружено, что увеличение содержания целлюлозы в волокнах хорошо коррелирует с увеличением их жесткости и прочности. Кроме того, известно, что кристалличность целлюлозы и соотношение размеров кристаллитов влияют на жесткость клеточной стенки в направлении микрофибрилл (например,г., [21]). (iii) Угол из микрофибриллы . С помощью классической теории ламината (вращение ортотропной пластины в плоскости) можно продемонстрировать, что эффективные упругие свойства волокон в осевом направлении зависят от локальной жесткости в направлении микрофибрилл, умноженной на cos 4 , где — угол микрофибриллы. Таким образом, жесткость (и прочность) волокна очень чувствительна к углу наклона микроволокон, даже если механические свойства в направлении микроволокон постоянны. Эта тенденция также отражается в более точных и подробных микромеханических моделях (например, в исследовании Hofstetter et al. [22]). Малый угол микрофибрилл растительных волокон делает их сильно анизотропными (что также относится к синтетическим углеродным волокнам, но не к стекловолокнам), и это приводит к относительно низким поперечным механическим свойствам.
Эта тенденция также отражается в более точных и подробных микромеханических моделях (например, в исследовании Hofstetter et al. [22]). Малый угол микрофибрилл растительных волокон делает их сильно анизотропными (что также относится к синтетическим углеродным волокнам, но не к стекловолокнам), и это приводит к относительно низким поперечным механическим свойствам.
Из трех вышеупомянутых ультраструктурных особенностей наиболее важным фактором, который необходимо учитывать при моделировании механических свойств волокон, является, вероятно, угол микрофибрилл, поскольку просветы волокон могут либо сжаться (как для ранней древесины в волокнах, подвергнутых химической пульверизации). или заполнены смолой с низкой вязкостью во время производства композитов, а содержание целлюлозы является внутренним свойством, которое примерно постоянно для растительных волокон и постоянное, хотя и более низкое, для древесных волокон (см.Таблица 1). Зависимость жесткости волокна от угла микрофибриллы является хорошо известным эффектом (например, [23]) и может быть описана классической теорией ламината [24].
4. Преформы из волокон
Типы преформ из целлюлозных волокон, которые будут использоваться для изготовления композитов, в принципе идентичны преформам для синтетических волокон, хотя необходимо учитывать некоторые особенности. Ниже приведены подробные сведения о преформах из дерева и растительных волокон.
4.1. Преформы из древесного волокна
Древесные волокна доступны по низкой цене в виде целлюлозных волокон (рис. 2 (а)). Из них делают бумажные листы или картон для упаковки. Одним из способов изготовления композитов на основе древесных волокон является использование таких волокнистых матов (рис. 2 (b)), которые можно пропитать, например, с использованием метода литья с переносом смолы (например, [46]). Вязкая термореактивная смола пропитывает замкнутый древесноволокнистый мат с помощью сосуда под давлением, прикрепленного к входному отверстию формы, а иногда также с помощью вакуумного всасывания на выходе.Эта технология производства подходит только для смол с низкой вязкостью, обычно термореактивных. Термопласты обычно имеют высокую вязкость в расплавленном состоянии, и формование с переносом смолы не подходит, поскольку время пропитки было бы слишком большим или требуемое высокое давление могло бы вызвать сильную деформацию волокнистого мата. Вместо этого можно рекомендовать смешанную технологию. Используя бумагоделательные машины, например, так называемые французские или финские листоформовочные машины в лабораторных масштабах, можно производить маты, состоящие из смешанных волокон древесной массы и термопластических волокон (например.г., [46]). Термопластические волокна предпочтительно должны иметь такие же размеры, что и волокна пульпы, чтобы иметь примерно такие же гидродинамические свойства во время процесса формирования, что способствует эффективному смешиванию. Термопластические волокна можно формовать до диаметров около 30 мкм и м и нарезать до длины примерно 3 мм (аналогично размерам волокон целлюлозы). Когда мат из смешанных волокон высохнет, его можно поместить в горячий пресс, и можно будет формовать композитные компоненты.
Термопласты обычно имеют высокую вязкость в расплавленном состоянии, и формование с переносом смолы не подходит, поскольку время пропитки было бы слишком большим или требуемое высокое давление могло бы вызвать сильную деформацию волокнистого мата. Вместо этого можно рекомендовать смешанную технологию. Используя бумагоделательные машины, например, так называемые французские или финские листоформовочные машины в лабораторных масштабах, можно производить маты, состоящие из смешанных волокон древесной массы и термопластических волокон (например.г., [46]). Термопластические волокна предпочтительно должны иметь такие же размеры, что и волокна пульпы, чтобы иметь примерно такие же гидродинамические свойства во время процесса формирования, что способствует эффективному смешиванию. Термопластические волокна можно формовать до диаметров около 30 мкм и м и нарезать до длины примерно 3 мм (аналогично размерам волокон целлюлозы). Когда мат из смешанных волокон высохнет, его можно поместить в горячий пресс, и можно будет формовать композитные компоненты. Этот метод не ограничивается только плоскими пластинами для испытаний материалов, но также может изготавливаться сложные детали с двойной кривизной [47].
Этот метод не ограничивается только плоскими пластинами для испытаний материалов, но также может изготавливаться сложные детали с двойной кривизной [47].
Бумажная промышленность включает в себя огромную инфраструктуру для производства древесноволокнистых матов. Предполагаемые объемы таких волокнистых преформ для применения в композитах чрезвычайно малы по сравнению с объемами производства обычной бумаги и картона. Тем не менее, есть возможность использовать накопленный опыт и использовать небольшие бумажные фабрики для производства композитных преформ.В лабораторном масштабе используются два основных метода имитации процесса производства бумаги. Наиболее распространены листы, полученные путем динамического формования листов и обычные листы. При динамическом формовании листа струя волокнистой суспензии направляется к вращающемуся проволочному барабану [48]. Волокна осаждаются на проволоке, а вода проходит через проволоку. В зависимости от разницы скоростей струи и проволоки волокна будут ориентироваться в продольном направлении (по окружности вращающегося барабана). Затем может быть получен анизотропный лист с характеристиками, аналогичными тем, которые производятся на бумажных фабриках, где волокна предпочтительно ориентированы в продольном направлении по сравнению с поперечным направлением. В листах для рук волокна смешаны с водой в большом контейнере. Вода резко выпускается на дно емкости, и волокна откладываются на плоской проволоке внизу. В этом случае волокна преимущественно беспорядочно ориентированы в плоскости. Таким образом, основное различие между листами, которые формируются с использованием динамического формирователя листов, и ручных листов состоит в том, что первые, как правило, анизотропны в плоскости, а вторые изотропны в плоскости.
Затем может быть получен анизотропный лист с характеристиками, аналогичными тем, которые производятся на бумажных фабриках, где волокна предпочтительно ориентированы в продольном направлении по сравнению с поперечным направлением. В листах для рук волокна смешаны с водой в большом контейнере. Вода резко выпускается на дно емкости, и волокна откладываются на плоской проволоке внизу. В этом случае волокна преимущественно беспорядочно ориентированы в плоскости. Таким образом, основное различие между листами, которые формируются с использованием динамического формирователя листов, и ручных листов состоит в том, что первые, как правило, анизотропны в плоскости, а вторые изотропны в плоскости.
4.2. Преформы из растительного волокна
Типы преформ из растительного волокна, доступных для композитов, показаны на Рисунке 3. Ниже приводится описание их обработки и характеристики.
После того, как волокна были извлечены из растений с помощью процесса вымачивания с последующим рядом механических процессов, волокна могут быть преобразованы в нетканые маты с помощью методов воздушной укладки и иглопробивки [49] . Ориентация волокон в нетканых матах номинально в плоскости случайна, но они могут показывать предпочтительную ориентацию волокон в машинном направлении [50].В качестве альтернативы, волокна могут быть преобразованы в непрерывную пряжу пряжи с использованием различных методов прядения, таких как кольцевое прядение, роторное прядение, оберточное прядение и воздушно-струйное прядение [51]. Кольцевое прядение — самый распространенный метод. Во время прядения непрерывный пучок почти параллельных волокон (так называемая лента) скручивается так, что волокна принимают спиралевидную конфигурацию. Влияние угла закручивания волокна на механические свойства композитов рассматривалось в нескольких исследованиях [52–54].Кроме того, площадь поперечного сечения пряжи (которая косвенно определяется ее линейной плотностью, выраженной в единицах г / 1000 м) и степень уплотнения пряжи являются другими важными характеристиками пряжи, которые, однако, получили ограниченное внимание в перспективе. композитной арматуры [14].
Ориентация волокон в нетканых матах номинально в плоскости случайна, но они могут показывать предпочтительную ориентацию волокон в машинном направлении [50].В качестве альтернативы, волокна могут быть преобразованы в непрерывную пряжу пряжи с использованием различных методов прядения, таких как кольцевое прядение, роторное прядение, оберточное прядение и воздушно-струйное прядение [51]. Кольцевое прядение — самый распространенный метод. Во время прядения непрерывный пучок почти параллельных волокон (так называемая лента) скручивается так, что волокна принимают спиралевидную конфигурацию. Влияние угла закручивания волокна на механические свойства композитов рассматривалось в нескольких исследованиях [52–54].Кроме того, площадь поперечного сечения пряжи (которая косвенно определяется ее линейной плотностью, выраженной в единицах г / 1000 м) и степень уплотнения пряжи являются другими важными характеристиками пряжи, которые, однако, получили ограниченное внимание в перспективе. композитной арматуры [14]. Можно предположить, что степень уплотнения пряжи коррелирует с проницаемостью пряжи для пропитки матрицы во время изготовления композитов. Необходимы дополнительные исследования для улучшения понимания корреляции между различными структурными характеристиками пряжи из растительных волокон и механическими характеристиками пряжи в композитах.
Можно предположить, что степень уплотнения пряжи коррелирует с проницаемостью пряжи для пропитки матрицы во время изготовления композитов. Необходимы дополнительные исследования для улучшения понимания корреляции между различными структурными характеристиками пряжи из растительных волокон и механическими характеристиками пряжи в композитах.
Преформы пряжи из растительных волокон могут быть использованы непосредственно для производства композитов путем намотки из смеси нитей вместе с термопластичной филаментной нитью с последующим прессованием [41], или пряжу можно использовать для изготовления преформ тканых и не гофрированных материалов. . Ткани изготавливаются с различными узорами плетения, такими как полотняное, саржевое и атласное переплетение, в которых пряжа по-разному переплетена в двух основных, ортогональных и плоских направлениях. Пряжа в двух направлениях может иметь разную линейную плотность, и их можно размещать на разном расстоянии друг от друга.Ткани предлагают возможность иметь плоскую конфигурацию пряжи в двух измерениях, разработанную для соответствия профилю нагрузки данного композитного приложения. Ткани из льняных, джутовых и хлопковых волокон широко доступны, но чаще всего они предназначены для текстильных приложений, а не для композитных материалов. Ткани без обжима состоят из пряжи, которая не скрепляется путем вплетения друг в друга, а вместо этого сшивается тонкими и гибкими нитями (обычно из термопластичного полиэстера).Это означает, что пряжа полностью растянута; то есть у них нет обжима, так как они не должны переходить друг под друга. Отдельные слои параллельных нитей, удерживаемые вместе поперечно направленными сшивающими нитями, обозначают одноосные не гофрированные ткани. Такие одноосные слои укладываются и сшиваются вместе, образуя двухосные или многоосные не гофрированные ткани с определенной плоской ориентацией пряжи, например, ± 45 °, 0 ° / 90 ° и 0 ° / + 45 ° / -45 ° / 90 °. . В последнее время ряд европейских компаний приступили к производству не гофрированных тканей из льняных волокон.Таким образом, впервые в продаже появились ткани из растительных волокон, специально предназначенные для композитов.
Ткани из льняных, джутовых и хлопковых волокон широко доступны, но чаще всего они предназначены для текстильных приложений, а не для композитных материалов. Ткани без обжима состоят из пряжи, которая не скрепляется путем вплетения друг в друга, а вместо этого сшивается тонкими и гибкими нитями (обычно из термопластичного полиэстера).Это означает, что пряжа полностью растянута; то есть у них нет обжима, так как они не должны переходить друг под друга. Отдельные слои параллельных нитей, удерживаемые вместе поперечно направленными сшивающими нитями, обозначают одноосные не гофрированные ткани. Такие одноосные слои укладываются и сшиваются вместе, образуя двухосные или многоосные не гофрированные ткани с определенной плоской ориентацией пряжи, например, ± 45 °, 0 ° / 90 ° и 0 ° / + 45 ° / -45 ° / 90 °. . В последнее время ряд европейских компаний приступили к производству не гофрированных тканей из льняных волокон.Таким образом, впервые в продаже появились ткани из растительных волокон, специально предназначенные для композитов.
5. Механические свойства композитов
Механические свойства композитов из древесных и растительных волокон подробно охарактеризованы и проанализированы. Однако были охарактеризованы в основном свойства при растяжении, а также свойства при изгибе и в некоторой степени также свойства при ударе, поскольку их относительно легко измерить, и они обычно используются для оценки различных материалов в процессе разработки материалов.Другие более сложные механические свойства, такие как усталость [55–58] и ползучесть [59], изучены в меньшей степени.
В Таблице 2 представлены типичные значения свойств при растяжении (жесткости и прочности) композитов из древесного и растительного волокна, а также значения для композитов из стекловолокна и углеродного волокна. Замечательно высокая жесткость и прочность на 26 ГПа и 247 МПа соответственно для фенолформальдегидных композитов, пропитанных крафт-бумагой [34], до сих пор не были достигнуты для древесноволокнистых композитов.Эти материалы были разработаны во время Второй мировой войны для использования в обшивке крыльев самолетов. Помимо этих крайних результатов Кокса и Пеппера [34], можно заметить, что композиты из целлюлозных волокон (как древесных, так и растительных волокон) с номинальной случайной ориентацией волокон в плоскости, изготовленные с использованием преформ из рыхлых волокон, бумаги и нетканые маты, обладают умеренными растягивающими свойствами с жесткостью в диапазоне 4–8 ГПа и прочностью в диапазоне 30–60 МПа. Что касается композитов из стекловолокна, с аналогичной случайной ориентацией волокон в плоскости, демонстрирующих жесткость в диапазоне 5–7 ГПа и прочность в диапазоне 80–100 МПа, композиты из целлюлозного волокна в целом демонстрируют сравнимую жесткость и несколько меньшую прочность. .Хорошо известно, что можно использовать различные химические подходы для управления межфазным соединением с целью повышения прочности композитов из целлюлозных волокон. Ацетилирование — это один из видов обработки поверхности, который можно использовать для уменьшения полярности волокон, делая их более совместимыми с (обычно) неполярной матрицей [60].
Помимо этих крайних результатов Кокса и Пеппера [34], можно заметить, что композиты из целлюлозных волокон (как древесных, так и растительных волокон) с номинальной случайной ориентацией волокон в плоскости, изготовленные с использованием преформ из рыхлых волокон, бумаги и нетканые маты, обладают умеренными растягивающими свойствами с жесткостью в диапазоне 4–8 ГПа и прочностью в диапазоне 30–60 МПа. Что касается композитов из стекловолокна, с аналогичной случайной ориентацией волокон в плоскости, демонстрирующих жесткость в диапазоне 5–7 ГПа и прочность в диапазоне 80–100 МПа, композиты из целлюлозного волокна в целом демонстрируют сравнимую жесткость и несколько меньшую прочность. .Хорошо известно, что можно использовать различные химические подходы для управления межфазным соединением с целью повышения прочности композитов из целлюлозных волокон. Ацетилирование — это один из видов обработки поверхности, который можно использовать для уменьшения полярности волокон, делая их более совместимыми с (обычно) неполярной матрицей [60]. Также связующие агенты, такие как малеиновый ангидрид, могут использоваться для образования ковалентных связей между волокнами и матрицей [61]. В исследовании Clemons [30] (Таблица 2) было показано, что прочность композитов древесное волокно / ПП увеличилась с 28 до 52 МПа при использовании малеинового ангидрида в качестве связующего агента, тогда как жесткость не показала никаких изменений.Аналогичное значительное увеличение прочности с 40 до 60 МПа было обнаружено при исследовании композитов джутовое волокно / полипропилен Андерсеном и Плакеттом [37].
Также связующие агенты, такие как малеиновый ангидрид, могут использоваться для образования ковалентных связей между волокнами и матрицей [61]. В исследовании Clemons [30] (Таблица 2) было показано, что прочность композитов древесное волокно / ПП увеличилась с 28 до 52 МПа при использовании малеинового ангидрида в качестве связующего агента, тогда как жесткость не показала никаких изменений.Аналогичное значительное увеличение прочности с 40 до 60 МПа было обнаружено при исследовании композитов джутовое волокно / полипропилен Андерсеном и Плакеттом [37].
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Литье под давлением; 2 нить-намотка.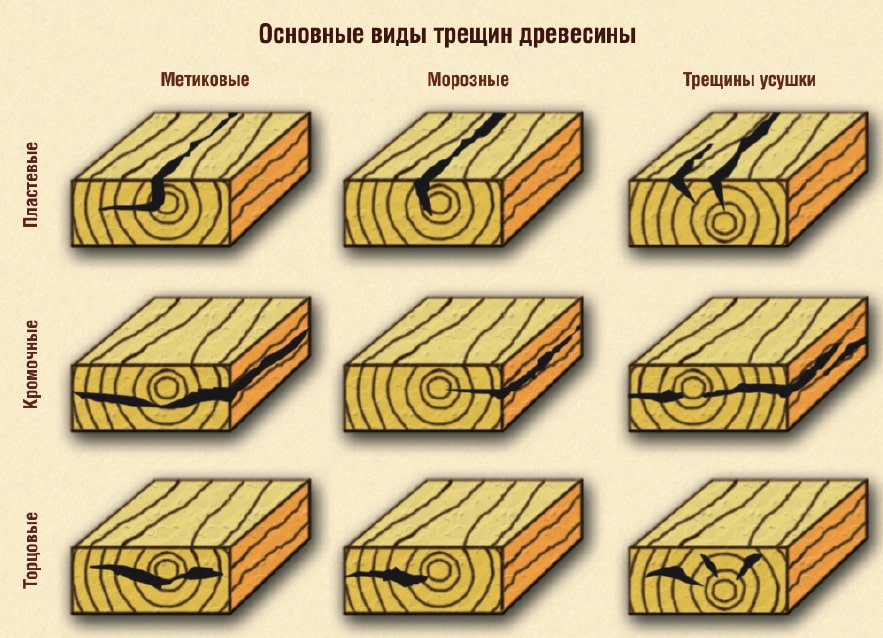 Машинное направление; b поперечное направление; c высокомодульные волокна; d высокопрочные волокна. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
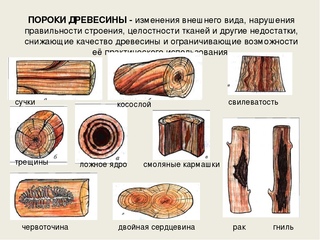 2
2 При использовании преформ с номинально однонаправленными волокнами, таких как пряжа и не гофрированные ткани, свойства при растяжении композитов заметно повышаются при жесткости в диапазоне 20–32 ГПа и прочности в диапазоне 130–340 МПа. (Таблица 2). Кроме того, по сравнению с композитами со случайной ориентацией волокон в плоскости, объемное содержание волокна в однонаправленных композитах в целом выше (до 50–55%) [62].Это связано с лучшей упаковочной способностью выровненных волокон по сравнению с произвольно ориентированными волокнами. Однако по сравнению с синтетическими волокнами сборки из целлюлозных волокон обычно имеют более низкую упаковочную способность [63], что означает, что максимальное объемное содержание волокна обычно ниже в композитах из целлюлозных волокон. Это часть объяснения более низкой жесткости однонаправленных композитов из целлюлозного волокна по сравнению с композитами из стекловолокна со значениями примерно 30 и 45 ГПа соответственно. Жесткость целлюлозных волокон в композитах оценивается в диапазоне 20–90 ГПа [41], что для целлюлозных волокон наилучшего качества сопоставимо со стеклянными волокнами с жесткостью в диапазоне 70–87 ГПа [45]. Что касается прочности, однонаправленные композиты из целлюлозного волокна показывают радикально более низкие значения — около 300 МПа по сравнению с около 1000 МПа для композитов из стекловолокна. Объяснение низкой прочности однонаправленных композитов из целлюлозного волокна в настоящее время неизвестно, однако ожидается, что дефекты волокна, которые вносятся в волокна во время их обработки, играют большую роль [64].
Жесткость целлюлозных волокон в композитах оценивается в диапазоне 20–90 ГПа [41], что для целлюлозных волокон наилучшего качества сопоставимо со стеклянными волокнами с жесткостью в диапазоне 70–87 ГПа [45]. Что касается прочности, однонаправленные композиты из целлюлозного волокна показывают радикально более низкие значения — около 300 МПа по сравнению с около 1000 МПа для композитов из стекловолокна. Объяснение низкой прочности однонаправленных композитов из целлюлозного волокна в настоящее время неизвестно, однако ожидается, что дефекты волокна, которые вносятся в волокна во время их обработки, играют большую роль [64].
Свойства при растяжении в таблице 2 хорошо иллюстрируют текущее состояние композитов из целлюлозного волокна, где жесткость приемлема и сопоставима с композитами из стекловолокна, но прочность требует улучшения. Из-за низкой плотности целлюлозных волокон особые механические свойства композитов из целлюлозного волокна особенно конкурентоспособны по сравнению с композитами из стекловолокна. Кроме того, если эти конкретные свойства нормализовать относительно стоимости, композиты из целлюлозного волокна также хорошо сравниваются с композитами из углеродного волокна.Другими словами, для приложений большого объема, где вес является проблемой, например, при упаковке и транспортировке, композиты из целлюлозного волокна, вероятно, будут основными конкурирующими материалами.
Кроме того, если эти конкретные свойства нормализовать относительно стоимости, композиты из целлюлозного волокна также хорошо сравниваются с композитами из углеродного волокна.Другими словами, для приложений большого объема, где вес является проблемой, например, при упаковке и транспортировке, композиты из целлюлозного волокна, вероятно, будут основными конкурирующими материалами.
6. Моделирование механических свойств композитов
Для композитных материалов количественная связь между микроструктурой и механическими свойствами обычно называется микромеханикой, и это было предметом обширных исследований для высокоэффективных композитов. Микромеханические модели, разработанные для этих материалов, обычно применимы также и для композитов из целлюлозных волокон с некоторыми модификациями, учитывающими особенности целлюлозных волокон.Безусловно, наибольшее внимание привлекла связь между микроструктурой и упругими свойствами. Жесткость является одним из важнейших параметров конструкции, и она также поддается моделированию, поскольку жесткость представляет собой среднее общее свойство, в отличие от прочности, которая обычно контролируется локально самыми большими дефектами в материалах.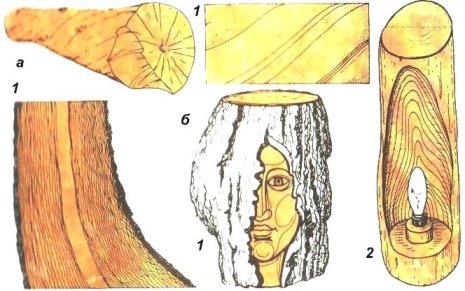
6.1. Композиты со случайной ориентацией волокон
Композиты со случайным распределением ориентации волокон в плоскости, которое обычно имеет место для композитов из древесных волокон, можно рассматривать как стопку однонаправленных слоев, где относительная толщина каждого слоя определяется по волокну. распределение ориентации.Это известно как аналогия ламината, где классическая механика ламината может использоваться для соотнесения упругих свойств гипотетического однонаправленного слоя с характеристиками композитной пластины. Аналогия с ламинатом схематически проиллюстрирована на рисунке 4. Предполагается, что распределение ориентации волокон является симметричным, что обычно имеет место для матов из древесного волокна, изготовленных с помощью традиционных методов влажного формования [65]. Это означает, что материалы глобально ортотропны, а глобальная матрица жесткости может быть описана пятью упругими константами, а именно продольным и поперечным модулями Юнга, большим и второстепенным коэффициентами Пуассона и модулем сдвига. Компоненты в глобальной матрице жесткости могут быть определены с помощью стандартизированных макроскопических испытаний, а распределение ориентации волокон может быть найдено путем анализа изображений сканированных участков волокнистого мата [66].
Компоненты в глобальной матрице жесткости могут быть определены с помощью стандартизированных макроскопических испытаний, а распределение ориентации волокон может быть найдено путем анализа изображений сканированных участков волокнистого мата [66].
Волокна хвойной целлюлозы имеют коэффициент формы около 100 [67]. С механической точки зрения эти волокна можно рассматривать как непрерывные, то есть имеющие бесконечную длину, поскольку неэффективные длины вблизи концов волокна относительно малы, как это может быть рассчитано с помощью теорий сдвига и запаздывания [68, 69].Вклад жесткости древесных волокон в однонаправленные слои в аналогии ламината затем может быть описан простыми механическими моделями, такими как модель правила смесей для продольных упругих свойств и модель Халпина-Цая для поперечных упругих свойств и свойств сдвига ( например, [70]). Что касается внеосевых свойств, модель концентрического цилиндра Хашина является более точной [71]. Последняя модель использовалась Neagu et al. [72] для обратного расчета жесткости древесных волокон на основе измеренной жесткости композитов и, таким образом, ранжирования различных химических обработок древесных волокон с точки зрения их эффективности армирования в композитах.Пример показан на рисунке 5, где показано влияние отбеливания и остаточного лигнина (характеризуемого числом Каппа) на определяющую жесткость волокна. Это служит иллюстрацией того, как микромеханический подход может быть использован для определения оптимального уровня отбеливания независимо от содержания волокна и ориентации волокон в композитах. Эти последние два параметра могут быть трудно воспроизводимыми при производстве композитов.
[72] для обратного расчета жесткости древесных волокон на основе измеренной жесткости композитов и, таким образом, ранжирования различных химических обработок древесных волокон с точки зрения их эффективности армирования в композитах.Пример показан на рисунке 5, где показано влияние отбеливания и остаточного лигнина (характеризуемого числом Каппа) на определяющую жесткость волокна. Это служит иллюстрацией того, как микромеханический подход может быть использован для определения оптимального уровня отбеливания независимо от содержания волокна и ориентации волокон в композитах. Эти последние два параметра могут быть трудно воспроизводимыми при производстве композитов.
6.2. Композиты с незначительным содержанием пористости
В композитах с целлюлозным волокном пористость обычно вносит заметный вклад в общий объем композита с содержанием пористости до 10% [62].Напротив, в композитах из стекла и углеродного волокна накоплен значительный объем знаний о снижении содержания пористости ниже 1% [73]. В целом в композитах из целлюлозного волокна пористостью, как правило, нельзя пренебрегать, и ее следует учитывать при оценке характеристик композитов.
В целом в композитах из целлюлозного волокна пористостью, как правило, нельзя пренебрегать, и ее следует учитывать при оценке характеристик композитов.
На рисунке 6 показаны примеры трех типов пористости, которые обычно можно найти в композитах из целлюлозного волокна: пористость просвета волокна, пористость на границе раздела и пористость пропитки. В исследовании Madsen et al.[62], содержание пористости коррелирует с содержанием волокна и матрицы, и представлена модель численной корреляции между массовым и объемным содержанием компонентов композита. Входными параметрами являются (i) плотность волокон и матрицы, которую можно измерить методами пикнометрии и плавучести, (ii) ряд эмпирических констант пористости, которые можно измерить по изображениям композитных микроструктур, и (iii) максимально достижимый объемная доля волокна, которую можно определить по характеристикам уплотнения волоконной сборки.Модель предсказывает объемные доли волокон, матрицы и пористости как функцию массовой доли волокна. Модель применима к композитам в целом, но особенно актуальна для композитов с относительно высокой пористостью, что обычно имеет место для композитов из целлюлозного волокна.
Модель применима к композитам в целом, но особенно актуальна для композитов с относительно высокой пористостью, что обычно имеет место для композитов из целлюлозного волокна.
На рис. 7 (а) показаны экспериментальные данные и модельные прогнозы объемного состава серии однонаправленных композитов льняное волокно / термопластичная матрица с переменными весовыми долями волокон.Объемные доли волокон и пористость увеличиваются в зависимости от массовой доли волокна до определенного значения, при котором после того, как объемная доля волокна остается постоянной, пористость начинает увеличиваться более резко. Массовая доля переходного волокна составляет 0,61. Таким образом, данные композиты должны изготавливаться с массовой долей волокна 0,61, чтобы иметь наилучшее возможное сочетание высокой объемной доли волокна и низкой пористости, и, как будет показано далее, это приводит к композитам с максимально достижимой жесткостью при растяжении.
Прогнозы объемного состава композитов могут быть интегрированы с микромеханическими моделями. Это было сделано в исследовании Madsen et al. [74], применяя правило модели смесей для жесткости композитов. Использовалась модифицированная версия модели, в которой был учтен эффект пористости, определяющий концентрацию напряжений в композитах. На рисунке 7 (b) показаны экспериментальные данные и модельные прогнозы жесткости однонаправленных композитов из льняного волокна и термопластичной матрицы.Жесткость монотонно увеличивается в зависимости от массовой доли волокна до определенного значения, после которого она начинает уменьшаться. Весовая доля переходного волокна составляет 0,61 (т.е. значение, определенное при моделировании объемного состава композитов), и здесь композиты показывают максимально достижимую жесткость около 35 ГПа. Он демонстрирует, что модели могут использоваться в качестве руководства для проектирования композитов с существенной пористостью, таких как композиты из целлюлозного волокна, для получения оптимального объемного состава, ведущего к оптимальным механическим характеристикам.
Это было сделано в исследовании Madsen et al. [74], применяя правило модели смесей для жесткости композитов. Использовалась модифицированная версия модели, в которой был учтен эффект пористости, определяющий концентрацию напряжений в композитах. На рисунке 7 (b) показаны экспериментальные данные и модельные прогнозы жесткости однонаправленных композитов из льняного волокна и термопластичной матрицы.Жесткость монотонно увеличивается в зависимости от массовой доли волокна до определенного значения, после которого она начинает уменьшаться. Весовая доля переходного волокна составляет 0,61 (т.е. значение, определенное при моделировании объемного состава композитов), и здесь композиты показывают максимально достижимую жесткость около 35 ГПа. Он демонстрирует, что модели могут использоваться в качестве руководства для проектирования композитов с существенной пористостью, таких как композиты из целлюлозного волокна, для получения оптимального объемного состава, ведущего к оптимальным механическим характеристикам.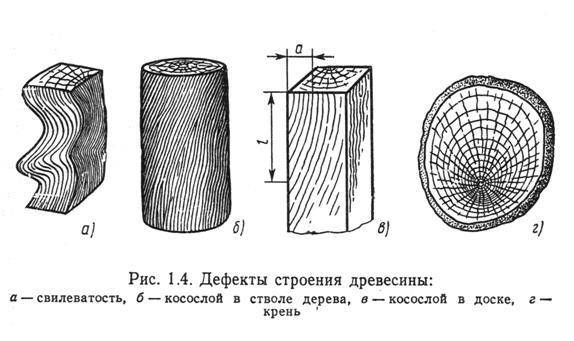
7. Чувствительность композитов к влаге
По сравнению с композитами с обычными волокнами, ахиллесова пята композитов из целлюлозного волокна — это их способность впитывать влагу, что приводит к набуханию, нестабильности размеров и потенциальному ухудшению механических свойств. Гидрофильность волокон обусловлена обилием доступных гидроксильных групп в гемицеллюлозе, в аморфной целлюлозе и на поверхности кристаллитов целлюлозы. Для конструкционных материалов чувствительность к влаге обычно считается недостатком, и ее следует по возможности снизить.Для композитов из целлюлозных волокон это может быть сделано путем сшивания полимеров клеточной стенки в волокнах [76], использования жесткой и гидрофобной матрицы [77] и использования влагонепроницаемого покрытия [78].
То, как набухание волокон влияет на стабильность размеров композитов, сложно из-за нерегулярной микроструктуры сборки волокон. Одним из способов изолировать гигрорасширение волокон и количественно оценить его вклад в гигрорасширение композитов является использование микромеханических моделей. Они аналогичны моделям, главным образом разработанным для теплового расширения и остаточных напряжений в композитах с керамической матрицей. Тепловое расширение и гигрорасширение регулируются одними и теми же физическими уравнениями, в которых термическая и гигральная деформации определяются температурой и влажностью соответственно. Микромеханические модели гигрорасширения композитов включают также параметры упругих свойств волокон и составляющих матрицы. В исследовании Neagu et al. [48], измерения скручивания полос композитов из древесного волокна и матов из древесного волокна были использованы для определения коэффициента поперечного гигрорасширения древесных волокон, равного примерно 0.10 деформаций на относительную влажность. Это согласуется с немногочисленными имеющимися в литературе данными о гигрорасширении целлюлозных волокон [79].
Они аналогичны моделям, главным образом разработанным для теплового расширения и остаточных напряжений в композитах с керамической матрицей. Тепловое расширение и гигрорасширение регулируются одними и теми же физическими уравнениями, в которых термическая и гигральная деформации определяются температурой и влажностью соответственно. Микромеханические модели гигрорасширения композитов включают также параметры упругих свойств волокон и составляющих матрицы. В исследовании Neagu et al. [48], измерения скручивания полос композитов из древесного волокна и матов из древесного волокна были использованы для определения коэффициента поперечного гигрорасширения древесных волокон, равного примерно 0.10 деформаций на относительную влажность. Это согласуется с немногочисленными имеющимися в литературе данными о гигрорасширении целлюлозных волокон [79].
В исследовании Madsen et al. [80] при гигрорасширении однонаправленных композитов пеньковое волокно / термопластичная матрица, изменения размеров, как было обнаружено, хорошо коррелировали с объемной долей волокна.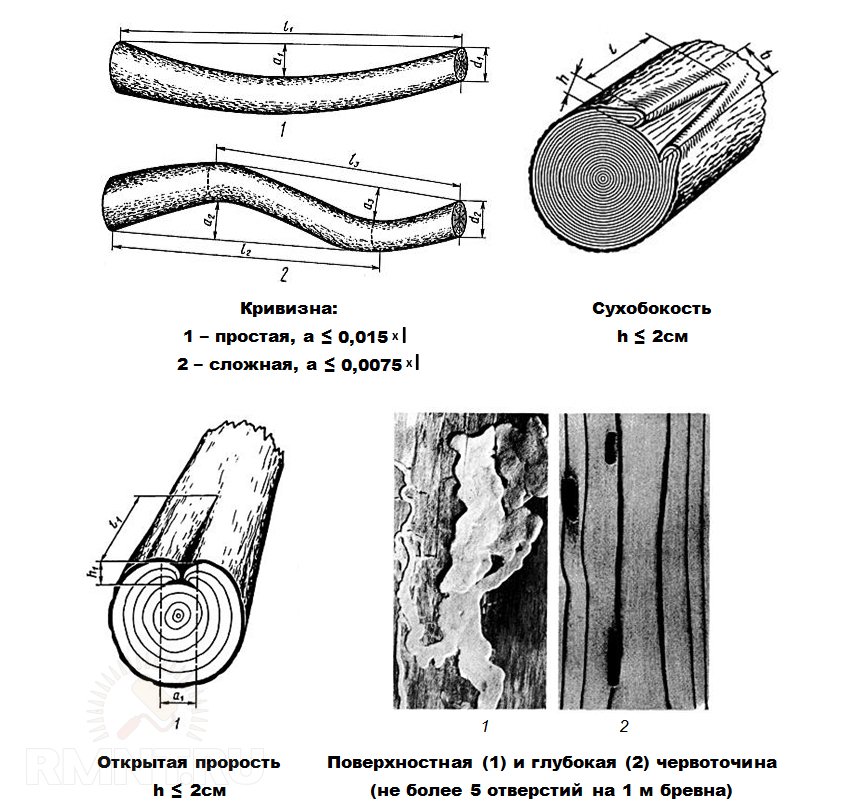 Выбранные результаты показаны на фиг. 8. При использовании эталонной влажности 65% относительной влажности было обнаружено, что поперечное гигрорасширение составило 0.9% и 3,3% при относительной влажности 85 и 100% соответственно для композитов с максимальной объемной долей волокна 0,43. Гигрорасширение в продольном направлении оказалось низким и слегка отрицательным, что предположительно связано с релаксацией остаточных напряжений растяжения в матрице, вызванной влагой. На рисунке видно, что точки экспериментальных данных хорошо моделируются линиями микромеханической модели.
Выбранные результаты показаны на фиг. 8. При использовании эталонной влажности 65% относительной влажности было обнаружено, что поперечное гигрорасширение составило 0.9% и 3,3% при относительной влажности 85 и 100% соответственно для композитов с максимальной объемной долей волокна 0,43. Гигрорасширение в продольном направлении оказалось низким и слегка отрицательным, что предположительно связано с релаксацией остаточных напряжений растяжения в матрице, вызванной влагой. На рисунке видно, что точки экспериментальных данных хорошо моделируются линиями микромеханической модели.
Стеклянные и углеродные волокна не впитывают влагу, хотя стеклянные волокна чувствительны к коррозии под воздействием окружающей среды в присутствии влаги и растягивающего напряжения [81].Чувствительность к влаге у древесных волокон выше, чем у растительных волокон, поскольку первые содержат большее относительное количество гемицеллюлозы, которая является наиболее гидрофильным полимером в клеточной стенке.
8. Применение композитов
В Европе композиты из целлюлозного волокна в основном используются в автомобильной промышленности. Применяемые волокнистые преформы представляют собой рыхлые волокна, используемые для технологий литья под давлением, и нетканые маты, используемые для методов компрессионного формования. Из-за номинальной случайной ориентации волокон в этих композитах они обладают только умеренными механическими свойствами (см. Таблицу 2), но это, тем не менее, делает их хорошо подходящими для использования в неструктурных компонентах, таких как дверные накладки, подкладки багажника и полки для пакетов.Низкие цены на рыхлые волокна и нетканые маты из целлюлозных волокон по сравнению с их синтетическими аналогами создают сильную мотивацию для использования этих двух преформ в автомобильной промышленности. За пределами Европы более распространено использование неструктурных компонентов на основе целлюлозных волокон, и древесные волокна являются гораздо более предпочтительным типом волокон.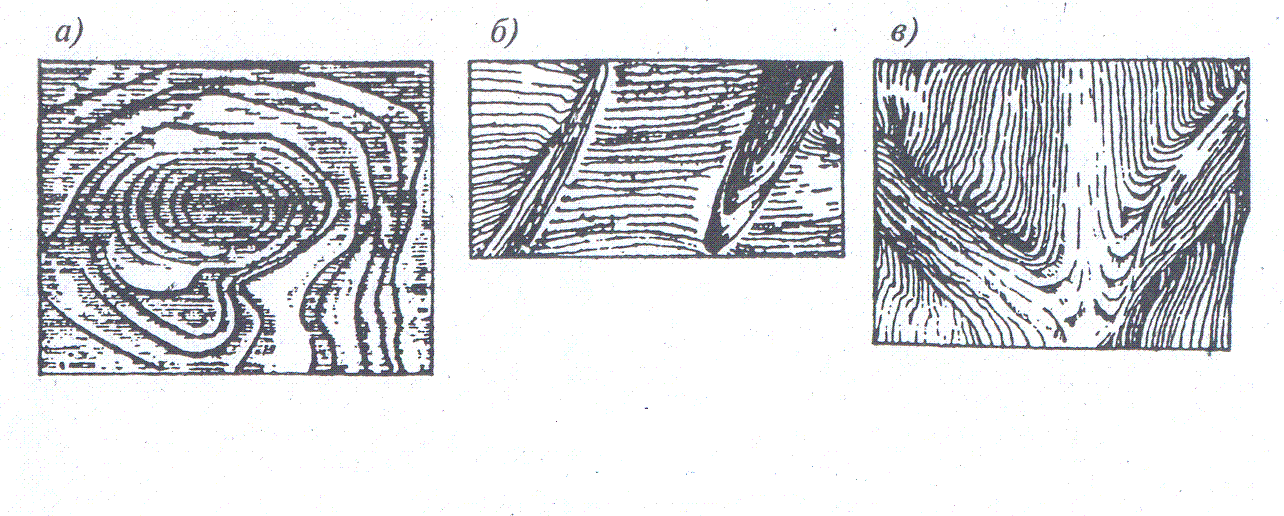 В Северной Америке основными приложениями являются строительные компоненты, такие как настилы, оконные профили и полы.
В Северной Америке основными приложениями являются строительные компоненты, такие как настилы, оконные профили и полы.
Недавно, в контексте исследовательских и опытно-конструкторских проектов, был проведен ряд демонстраций, демонстрирующих хороший потенциал композитов из целлюлозного волокна в новых областях применения (см. Рисунок 9).(i) Скульптура «», представленная в Музее современного искусства Луизианы, Дания, специальная выставка «Зеленая архитектура для будущего», победитель премии JEC за инновации 2010 года, совместное предприятие 20 компаний, координируемое 3XN Architects, Дания. (ii) Обод колеса , проект 7-й рамочной программы ЕС, NATEX (2008–2012). (iii) Стул для детей, разработанный научно-исследовательским институтом Innventia AB совместно с целлюлозно-бумажной промышленностью и архитекторами, представленный на Миланской мебельной ярмарке. 2009 г.(iv) Панели двойной кривизны , спроектированные М. Ларсеном и К.Р. Nielsen, представленный на «Klimaforum09 / Ideas at work» в связи с COP15, Копенгаген, 2009 г. , и на выставке JEC 2010. (v) Малая лопасть ротора для использования в ветряной машине, проект 7-й рамочной программы ЕС , ВУДИ (2009–2012). (vi) Выставочные стенды в Шведском морском историческом музее в Стокгольме, Швеция, проект 7-й рамочной программы ЕС, WOODY (2009–2012).
, и на выставке JEC 2010. (v) Малая лопасть ротора для использования в ветряной машине, проект 7-й рамочной программы ЕС , ВУДИ (2009–2012). (vi) Выставочные стенды в Шведском морском историческом музее в Стокгольме, Швеция, проект 7-й рамочной программы ЕС, WOODY (2009–2012).
9.Перспективы на будущее: композиты из нановолокна
В последние годы значительное внимание было направлено на композиты, изготовленные из нановолокон целлюлозы (например, [82]). Как уже было описано, клеточная стенка древесных и растительных волокон структурирована как композит с микрофибриллами целлюлозы, встроенными в матрицу из гемицеллюлозы и лигнина. Микрофибриллы целлюлозы имеют поперечные размеры в диапазоне 10–100 нм и аксиальные размеры в диапазоне микрометров, и поэтому они подходят в качестве армирования в композитах из нановолокон.Идея состоит в том, чтобы добиться значительных улучшений технических свойств с добавлением нановолокон по сравнению с волокнами микрометрового диапазона. Это можно объяснить высокой удельной поверхностью наноразмерных волокон, которая влияет на свойства окружающей матрицы. Успех композитов из нановолокон особенно очевиден, если учитывать лишь незначительное добавление волокон и при этом сохраняется дисперсность нановолокон.
Это можно объяснить высокой удельной поверхностью наноразмерных волокон, которая влияет на свойства окружающей матрицы. Успех композитов из нановолокон особенно очевиден, если учитывать лишь незначительное добавление волокон и при этом сохраняется дисперсность нановолокон.
Композиты на основе углеродных нанотрубок в течение относительно долгого времени были очень многообещающими, но пока не получили широкого применения [83].Разница между нановолокнами целлюлозы и углеродными нанотрубками заключается в способности нановолокон целлюлозы связываться друг с другом посредством водородных связей, тогда как поверхность углеродных нанотрубок химически инертна. Нановолокна целлюлозы могут образовывать очень прочную сетку и, кроме того, хорошо связываться с материалами полимерной матрицы с полярными группами. Это, однако, также приводит к трудностям обработки, поскольку нановолокна целлюлозы имеют тенденцию к агрегированию и после влажной обработки требуется много времени для высыхания. Таким образом, технологичность и производительность являются взаимодополняющими и противоположными поведениями.Функциональные гидроксильные группы волокон можно модифицировать для улучшения диспергирования и обрабатываемости, хотя обычно это сопровождается повышенными затратами. Основные проблемы для композитов из целлюлозных нановолокон, вероятно, состоят в том, чтобы научиться производить объемные композитные компоненты с сохранением тонкости и дисперсности нановолокон. Сырьем являются сами целлюлозные волокна из древесной массы или растительных волокон, поэтому затраты на сырье пренебрежимо малы по сравнению с затратами на производство.
Таким образом, технологичность и производительность являются взаимодополняющими и противоположными поведениями.Функциональные гидроксильные группы волокон можно модифицировать для улучшения диспергирования и обрабатываемости, хотя обычно это сопровождается повышенными затратами. Основные проблемы для композитов из целлюлозных нановолокон, вероятно, состоят в том, чтобы научиться производить объемные композитные компоненты с сохранением тонкости и дисперсности нановолокон. Сырьем являются сами целлюлозные волокна из древесной массы или растительных волокон, поэтому затраты на сырье пренебрежимо малы по сравнению с затратами на производство.
10. Общее сравнение волокон
Как показано в разделах выше, древесные и растительные волокна в некоторых отношениях схожи, а в других — различаются. Таким образом, в зависимости от предполагаемого применения один конкретный тип волокна подходит больше, чем другой. Далее приводится общее сравнение, чтобы выделить некоторые преимущества древесины по сравнению с растительными волокнами и наоборот. Точно так же целлюлозные волокна сравнивают с их синтетическими аналогами, стеклянными и углеродными волокнами.
Точно так же целлюлозные волокна сравнивают с их синтетическими аналогами, стеклянными и углеродными волокнами.
Преимущества древесных волокон по сравнению с растительными волокнами заключаются в следующем.(i) Низкая стоимость, доступная на целлюлозных фабриках. (ii) Относительно короткие волокна означают лучшую технологичность. (iii) Развитая инфраструктура, доступная на целлюлозно-бумажных комбинатах, для производства больших объемов по низкой цене. (iv) Преформы можно изготавливать из бумаги -технологии производства. (v) Могут быть достигнуты достаточно однородные партии целлюлозы. (vi) Не конкурирует с выращиванием продовольственных культур.
Преимущества растительных волокон по сравнению с древесными волокнами заключаются в следующем. (I) Высокая продуктивность и выход. (Ii) Высокое содержание целлюлозы, высокая степень кристалличности целлюлозы, низкий угол наклона микрофибрилл, небольшой просвет означают отличные механические свойства волокон. .(iii) Относительно длинные волокна означают возможность контролировать ориентацию волокон и их укладку. (iv) Текстильные технологии могут использоваться для производства пряжи, тканых материалов и не гофрированных тканей.
(iv) Текстильные технологии могут использоваться для производства пряжи, тканых материалов и не гофрированных тканей.
Несмотря на различия, древесные и растительные волокна имеют больше общего, чем то, что их отличает друг от друга. Можно упомянуть некоторые преимущества целлюлозных волокон по сравнению со стеклянными и углеродными волокнами. (I) возобновляемый. (Ii) биоразлагаемый. (Iii) свет, то есть композиты обладают хорошими специфическими свойствами, которые важны для автомобилей и упаковки.(iv) Недорогое сырье.
Основными недостатками целлюлозных волокон по сравнению со стеклянными и углеродными волокнами являются следующие: (i) Умеренные механические свойства. (Ii) Чувствительность к влаге, приводящая к нестабильности размеров и потенциальному ухудшению механических свойств. (Iii) Не полностью разработаны технологии изготовления композитов.
Приведенные выше списки ни в коем случае не являются исчерпывающими, они служат только для демонстрации некоторых свойств древесины и растительных волокон в контексте применения композитов.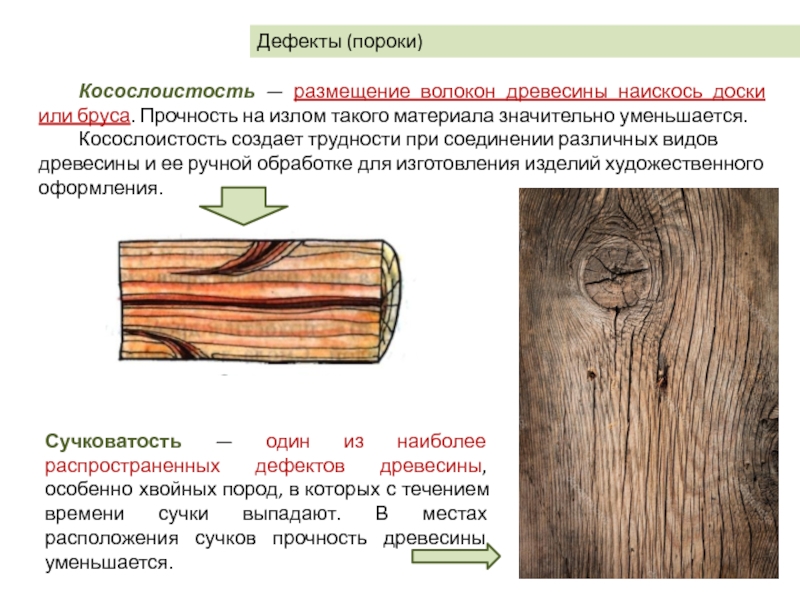 В дальнейшем развитии композитов из целлюлозных волокон важную роль играют как достоинства, так и недостатки. Конкретные преимущества определяют, какие области применения актуальны. Например, сочетание низкой стоимости, возобновляемости и способности к биологическому разложению делает композиты из целлюлозного волокна подходящими материалами для применения в упаковке. Недостатки ограничивают возможности их применения. Исследования того, как устранить эти недостатки, могут расширить распространение целлюлозных волокон как экологически чистой альтернативы синтетическим волокнам.Если целлюлозные волокна можно обработать, чтобы лучше сохранить присущую им высокую жесткость и прочность, и их можно модифицировать, чтобы они стали менее гидрофильными, они также могут стать потенциальными армирующими волокнами в современных конструкционных наружных применениях, например, в лопастях ротора для ветряных турбин и в нагрузках. транспортировка компонентов в транспортных средствах.
В дальнейшем развитии композитов из целлюлозных волокон важную роль играют как достоинства, так и недостатки. Конкретные преимущества определяют, какие области применения актуальны. Например, сочетание низкой стоимости, возобновляемости и способности к биологическому разложению делает композиты из целлюлозного волокна подходящими материалами для применения в упаковке. Недостатки ограничивают возможности их применения. Исследования того, как устранить эти недостатки, могут расширить распространение целлюлозных волокон как экологически чистой альтернативы синтетическим волокнам.Если целлюлозные волокна можно обработать, чтобы лучше сохранить присущую им высокую жесткость и прочность, и их можно модифицировать, чтобы они стали менее гидрофильными, они также могут стать потенциальными армирующими волокнами в современных конструкционных наружных применениях, например, в лопастях ротора для ветряных турбин и в нагрузках. транспортировка компонентов в транспортных средствах.
Понимание развития древесных волокон и волокон при росте деревьев
Ян Кирби изучает торцевую текстуру куска пиломатериала с 10-кратной лупой, текстура древесины показана справа. Поскольку древесина в большинстве мастерских существует в основном в виде пиломатериалов, приобретаемых с заданными размерами и строганием, легко забыть, что каждая часть изначально была получена из какой-то части живого дерева. Возможно, еще менее очевидно то, что качества, которые в первую очередь побудили вашу покупку, — цвет, зернистость, блеск, вес, твердость — открыты для объяснения, которое начинается на клеточном уровне. В первой статье серии, состоящей из двух частей, я расскажу о некоторых деталях анатомии древесины и ее функциях, которые можно узнать только при изучении под мощным микроскопом, и свяжу их с тем, что можно увидеть невооруженным глазом и ручным объективом с 10-кратным увеличением.
Поскольку древесина в большинстве мастерских существует в основном в виде пиломатериалов, приобретаемых с заданными размерами и строганием, легко забыть, что каждая часть изначально была получена из какой-то части живого дерева. Возможно, еще менее очевидно то, что качества, которые в первую очередь побудили вашу покупку, — цвет, зернистость, блеск, вес, твердость — открыты для объяснения, которое начинается на клеточном уровне. В первой статье серии, состоящей из двух частей, я расскажу о некоторых деталях анатомии древесины и ее функциях, которые можно узнать только при изучении под мощным микроскопом, и свяжу их с тем, что можно увидеть невооруженным глазом и ручным объективом с 10-кратным увеличением.Растущее дерево: корни, ствол, листья
Дерево состоит из трех частей, каждая из которых выполняет определенную функцию. Корни закрепляют дерево в земле и впитывают грунтовые воды, содержащие минеральные соли из почвы. Ствол переносит этот раствор, называемый соком, от корней к листьям; в нем хранится еда; он удерживает слои живых клеток, необходимые для роста дерева; и придает жесткость кроне — меньшим ветвям и веточкам, на которых растут листья. Листья поглощают углекислый газ из воздуха, выделяют кислород и путем фотосинтеза обогащают сок сахарами, которые передаются по внутренней коре и используются для стимулирования роста.
Листья поглощают углекислый газ из воздуха, выделяют кислород и путем фотосинтеза обогащают сок сахарами, которые передаются по внутренней коре и используются для стимулирования роста.Древесина
Эта диаграмма показывает основы того, как древесина и где древесина образуется в дереве с самого начала процесса. Прямо под корой находится микроскопически тонкий слой живых клеток, называемый камбием, который покрывает дерево от земли до кроны. Клетки камбия растут и делятся. Половина ячеек составляет либо древесину, либо кору; другая половина остается в камбии, чтобы снова расти и делиться. Новые клетки внутри камбия становятся одним из древесных элементов.Клетки снаружи становятся корой, которая делится на два слоя. Внутренняя кора несет богатый сахаром сок с листьев, чтобы питать камбий и корни. Внешняя кора защищает хрупкий камбий от вторжения насекомых, грибов, животных, а также от сильной жары и холода.Переменный рост
Несмотря на горячие надежды плотников на обратное, деревья не существуют для того, чтобы предоставить нам пригодный для использования прямозернистый плоский материал — и многие из его характеристик роста свидетельствуют об этом противоречии. Например, из-за множества факторов, включая почву, погоду и близость других деревьев, деревья не растут с одинаковой скоростью. Некоторые деревья растут по спирали. Вы можете наблюдать это в трещинах на поверхности деревянной опоры. Скорость накрутки варьируется. Многие большие тропические деревья будут двигаться по спирали в одном направлении, скажем, шесть периодов роста, а затем еще несколько периодов роста в противоположном направлении. Такое ламинирование ствола, несомненно, должно повысить его устойчивость к нагрузкам, но оно также создает проблемы для плотника при обработке преобразованной доски.Если по какой-то причине срубленное дерево показывает асимметричный рост — сердце находится ближе к одному краю, а не к центру — его обычно выбрасывают. Изготовленные из него доски склонны к внезапному разрушению, а при высыхании он деформируется и становится непригодным для использования.
Например, из-за множества факторов, включая почву, погоду и близость других деревьев, деревья не растут с одинаковой скоростью. Некоторые деревья растут по спирали. Вы можете наблюдать это в трещинах на поверхности деревянной опоры. Скорость накрутки варьируется. Многие большие тропические деревья будут двигаться по спирали в одном направлении, скажем, шесть периодов роста, а затем еще несколько периодов роста в противоположном направлении. Такое ламинирование ствола, несомненно, должно повысить его устойчивость к нагрузкам, но оно также создает проблемы для плотника при обработке преобразованной доски.Если по какой-то причине срубленное дерево показывает асимметричный рост — сердце находится ближе к одному краю, а не к центру — его обычно выбрасывают. Изготовленные из него доски склонны к внезапному разрушению, а при высыхании он деформируется и становится непригодным для использования.Заболонь и сердцевина
После пяти-десяти лет роста древесина в центре всех деревьев претерпевает сложные химические изменения. У большинства деревьев переход от заболони к сердцевине очевиден из-за изменения цвета. У некоторых бледный цвет сердцевины трудно отличить от бледного цвета древесины хвойных пород.Химические вещества, вызывающие изменения, трудно идентифицировать, и все вместе они известны как экстрактивные вещества. Тилозы встречаются во время этой переходной фазы, хотя и не у всех видов. Они выглядят как блестящая тонкая пленка, которая закупоривает сосуды. Происходит то, что тонкая мембрана, которая когда-то была частью стенки сосуда, разрушается через крошечные отверстия, называемые ямками, в полость сосуда из-за разницы давления в ткани.
У большинства деревьев переход от заболони к сердцевине очевиден из-за изменения цвета. У некоторых бледный цвет сердцевины трудно отличить от бледного цвета древесины хвойных пород.Химические вещества, вызывающие изменения, трудно идентифицировать, и все вместе они известны как экстрактивные вещества. Тилозы встречаются во время этой переходной фазы, хотя и не у всех видов. Они выглядят как блестящая тонкая пленка, которая закупоривает сосуды. Происходит то, что тонкая мембрана, которая когда-то была частью стенки сосуда, разрушается через крошечные отверстия, называемые ямками, в полость сосуда из-за разницы давления в ткани.Кольца роста
Сосуды многих видов деревьев, произрастающих в регионах с умеренным климатом, где есть четко выраженные периоды роста и покоя, образуют годичное кольцо, хорошо заметное на поперечном срезе.Мы называем эти деревья кольцевыми пористыми. Некоторые породы умеренного пояса, такие как береза, тополь и платан, а также большинство тропических лиственных пород, например красное дерево, имеют диффузную пористость. То есть сосуды появляются случайным образом на протяжении всего периода роста.
То есть сосуды появляются случайным образом на протяжении всего периода роста.Зерно, текстура и рисунок
Некоторая путаница связана с тем, что подразумевается под зерном, текстурой и рисунком, когда они используются для описания деревянной поверхности. Хорошим ориентиром является то, что зернистость относится к древесным волокнам относительно длины дерева на гранях и краях куска древесины.Текстура — это относительный размер и вариации элементов. Рисунок относится к рисунку на доске, вызванному расположением различных элементов и характером текстуры.Хвойные и лиственные породы
Деревья обычно делятся на хвойные (игольчатые) и лиственные (широколиственные). Считается, что древесина хвойных пород на нашей планете предшествовала лиственным, потому что их структура проще и примитивнее. В отличие от древесины твердых пород, осмотр древесины мягких пород с помощью 10-кратной линзы не позволяет выявить гораздо больше, чем можно увидеть при общем осмотре.Однако более внимательное рассмотрение показывает важные сходства: оба они состоят в основном из целлюлозы, и оба проходят фазу роста от заболони до сердцевины.
Элементы из мягкой древесины
Камбий — это микроскопически тонкий слой живых клеток, который покрывает дерево от земли до кроны. Клетки камбия растут и делятся. Половина новых ячеек состоит из дерева или коры; другая половина остается в камбии. Новые клетки внутри камбия становятся одним из древесных элементов (волокна, паренхима, лучи и т. Д.).Внешние клетки становятся корой Клетки камбия, которые делятся и образуют древесную ткань, являются так называемыми инициалами, потому что они инициируют образование специализированных клеток-предшественников после деления клеток. Есть два типа инициалов: веретенообразные инициалы и лучевые инициалы. В типичной мягкой древесине веретенообразная начальная буква образует только один тип элемента, называемый трахеидой. Трахеиды бывают разных типов и составляют большую часть древесины хвойных пород. В основном они в 80-100 раз длиннее своего диаметра.Они также сильно различаются по толщине стенок в зависимости от того, когда они откладываются в течение вегетационного периода. Таким образом они выполняют задачи по транспортировке и поддержке. Инициалы луча в мягкой древесине сложны, потому что они могут функционировать, чтобы хранить и обеспечивать пищу в течение некоторого времени в течение вегетационного периода после того, как другие элементы завершили свой рост. Большинство лучей имеют ширину всего одну клетку и не видны при всестороннем осмотре.
Таким образом они выполняют задачи по транспортировке и поддержке. Инициалы луча в мягкой древесине сложны, потому что они могут функционировать, чтобы хранить и обеспечивать пищу в течение некоторого времени в течение вегетационного периода после того, как другие элементы завершили свой рост. Большинство лучей имеют ширину всего одну клетку и не видны при всестороннем осмотре.Элементы из твердых пород дерева
В древесине лиственных пород веретенообразная начальная буква образует три различных элемента, называемых сосудами, волокном и паренхимой.Начальный луч отвечает только за один элемент — лучи. Каждый элемент играет определенную роль в дереве, и каждый достаточно четко определен, чтобы его можно было идентифицировать с помощью ручной линзы 10x при просмотре на поперечном сечении. Сосуды проводят воду и питательные вещества от корней к листьям. Это длинные трубчатые элементы с тонкими стенками и большими полостями. Хотя каждое судно короткое, они соединены последовательно по вертикали и могут простираться на некоторое расстояние вверх по дереву. Волокно — это механический или опорный элемент древесины.Он закладывается в конце вегетационного периода. Вы можете описать этот элемент как обратную конструкцию судна. Он короткий, с заостренным концом, толстой стенкой и небольшой полостью, которую не видно с помощью 10-кратного объектива. Паренхима — это, по сути, запасная ткань. Он откладывается вертикально у разных пород разными способами в течение вегетационного периода дерева. Лучи — самый сложный из четырех элементов. В некоторых случаях они продолжают функционировать в течение вегетационного периода, чтобы обеспечивать или хранить пищу.Их размер, форма и количество сильно различаются в зависимости от вида. У некоторых видов, таких как тополь и ива, лучи имеют ширину всего одну клетку и поэтому не видны. У большинства видов хорошо видны лучи. Лучи в дубе — это относительно огромные конструкции, сотни ячеек в высоту и десятки в ширину. На поперечном срезе лучи расходятся наружу, как спицы на колесе. Когда вы разбиваете бревно, оно обычно раскалывается по серии лучей.
Волокно — это механический или опорный элемент древесины.Он закладывается в конце вегетационного периода. Вы можете описать этот элемент как обратную конструкцию судна. Он короткий, с заостренным концом, толстой стенкой и небольшой полостью, которую не видно с помощью 10-кратного объектива. Паренхима — это, по сути, запасная ткань. Он откладывается вертикально у разных пород разными способами в течение вегетационного периода дерева. Лучи — самый сложный из четырех элементов. В некоторых случаях они продолжают функционировать в течение вегетационного периода, чтобы обеспечивать или хранить пищу.Их размер, форма и количество сильно различаются в зависимости от вида. У некоторых видов, таких как тополь и ива, лучи имеют ширину всего одну клетку и поэтому не видны. У большинства видов хорошо видны лучи. Лучи в дубе — это относительно огромные конструкции, сотни ячеек в высоту и десятки в ширину. На поперечном срезе лучи расходятся наружу, как спицы на колесе. Когда вы разбиваете бревно, оно обычно раскалывается по серии лучей. Затем вы можете увидеть их на разделенной стороне бревна на переднем фасаде или в виде пластины.
Затем вы можете увидеть их на разделенной стороне бревна на переднем фасаде или в виде пластины.Читаем до конца
Здесь вы можете увидеть различия в паренхиме, волокнах, сосудах и лучах у трех разных типов древесины. Поскольку четыре элемента, обнаруженные в древесине лиственных пород — сосуды, волокна, паренхима, лучи — уникально представлены в каждом виде, способность анализировать эти различия является ключом к возможности идентифицировать тип или породу любой древесины. В качестве введения в идентификацию древесины я выбрал три хорошо известных и широко используемых породы: красный дуб, белый дуб и африканское красное дерево.■ Сосуды: это крупные сосуды ранней древесины, образующие концентрические кольца. Это тонкостенные транспортные элементы с большими полостями, необходимые в начале весенне-летнего периода роста. Обычно они резко становятся меньше, хотя на обоих дубах их продолжают делать и использовать в течение всего вегетационного периода. Основное различие между дубами состоит в том, что сосуды из красного дуба не содержат каких-либо включений, тогда как сосуды из белого дуба забиты тилозами. (Эффект от этого мы увидим, когда посмотрим на использование в Части 2.) Сосуды из красного дерева меньше и одинакового размера. Некоторые делятся на пары, а некоторые на тройки. Белый налет в некоторых сосудах — обычная особенность и, вероятно, какая-то форма отложения десен. ■ Паренхима: У обоих дубов паренхима — это ткань, которая окружает крупные сосуды. Белый дуб легко увидеть из-за его белого цвета и типичных пламенеющих форм; у красного дуба он имеет оттенок коричневого и полностью окружает более мелкие сосуды, которые намного легче увидеть, чем в белом дубе.Паренхима у красного дерева очень редкая, заметна только по светлым краям некоторых сосудов. ■ Волокно: Волокно на каждой фотографии представляет собой темный фоновый материал. При таком малом увеличении невозможно увидеть элементы с толстыми стенками и небольшими полостями как отдельные предметы.
Основное различие между дубами состоит в том, что сосуды из красного дуба не содержат каких-либо включений, тогда как сосуды из белого дуба забиты тилозами. (Эффект от этого мы увидим, когда посмотрим на использование в Части 2.) Сосуды из красного дерева меньше и одинакового размера. Некоторые делятся на пары, а некоторые на тройки. Белый налет в некоторых сосудах — обычная особенность и, вероятно, какая-то форма отложения десен. ■ Паренхима: У обоих дубов паренхима — это ткань, которая окружает крупные сосуды. Белый дуб легко увидеть из-за его белого цвета и типичных пламенеющих форм; у красного дуба он имеет оттенок коричневого и полностью окружает более мелкие сосуды, которые намного легче увидеть, чем в белом дубе.Паренхима у красного дерева очень редкая, заметна только по светлым краям некоторых сосудов. ■ Волокно: Волокно на каждой фотографии представляет собой темный фоновый материал. При таком малом увеличении невозможно увидеть элементы с толстыми стенками и небольшими полостями как отдельные предметы. В обоих дубах вы увидите очень тонкие светлые линии, проходящие горизонтально через волокно. Это паренхиматозный материал. В красном дереве волокно — это красная основа, которая не является ни лучами, ни сосудами. ■ Лучи: на каждой фотографии лучи представляют собой линии, идущие более или менее вертикально.Они обильны у белого дуба, меньше у красного дуба. У обоих дубов между более крупными очень мелкие лучи. У красного дерева лучи примерно одинаковой толщины. Их разделяют немногим более двух диаметров сосудов, и они огибают сосуды.
В обоих дубах вы увидите очень тонкие светлые линии, проходящие горизонтально через волокно. Это паренхиматозный материал. В красном дереве волокно — это красная основа, которая не является ни лучами, ни сосудами. ■ Лучи: на каждой фотографии лучи представляют собой линии, идущие более или менее вертикально.Они обильны у белого дуба, меньше у красного дуба. У обоих дубов между более крупными очень мелкие лучи. У красного дерева лучи примерно одинаковой толщины. Их разделяют немногим более двух диаметров сосудов, и они огибают сосуды.Поведение отдельных древесных волокон на излом определяется геометрическими ограничениями: исследования ESEM на месте трех типов волокон
Сравнение измерений отдельных волокон внутри и снаружи камеры ESEM показало снижение прочности на разрыв для волокон, которые были испытаны в на месте.Данные измерений in situ и ex situ показаны в таблице 1.
Таблица 1 Максимальные нагрузки ( F max ) волокон, испытанных ex situ и in situ Все три группы волокон (ранняя древесина, переходная древесина и поздняя древесина) показали более низкие максимальные нагрузки при тестировании под микроскопом. Различия в температуре и предварительной обработке волокон (никогда не сушились / повторно увлажнялись) не полностью объясняют различное поведение внутри и снаружи.Следовательно, уменьшение может быть результатом радиационного поражения. Хорошо известно, что радиационное повреждение может быть серьезной проблемой в режиме ESEM, особенно когда водяной пар используется в качестве газа для визуализации. Радиационное повреждение вызывается присутствием воды, потому что высокоэнергетические первичные электроны создают радикалы во время процессов ионизации (Jenkins and Donald 1997; Kitching and Donald 1998). Предполагается, что эти радикалы могут привести к серьезному повреждению образцов. Подробнее о радиационном повреждении воды и влажных проб см. E.г. Китчинг и Дональд (1998) или Ройалл и др. (2001). Для минимизации излучения поддерживали низкое ускоряющее напряжение (8 кВ) и короткое время воздействия пучка. Однако, хотя в зонах, чувствительных к лучу, таких как ямы, повреждений не наблюдалось (Kifetew and Sandberg 2000), различия в максимальных нагрузках внутри и снаружи камеры указывали на радиационное повреждение.
Различия в температуре и предварительной обработке волокон (никогда не сушились / повторно увлажнялись) не полностью объясняют различное поведение внутри и снаружи.Следовательно, уменьшение может быть результатом радиационного поражения. Хорошо известно, что радиационное повреждение может быть серьезной проблемой в режиме ESEM, особенно когда водяной пар используется в качестве газа для визуализации. Радиационное повреждение вызывается присутствием воды, потому что высокоэнергетические первичные электроны создают радикалы во время процессов ионизации (Jenkins and Donald 1997; Kitching and Donald 1998). Предполагается, что эти радикалы могут привести к серьезному повреждению образцов. Подробнее о радиационном повреждении воды и влажных проб см. E.г. Китчинг и Дональд (1998) или Ройалл и др. (2001). Для минимизации излучения поддерживали низкое ускоряющее напряжение (8 кВ) и короткое время воздействия пучка. Однако, хотя в зонах, чувствительных к лучу, таких как ямы, повреждений не наблюдалось (Kifetew and Sandberg 2000), различия в максимальных нагрузках внутри и снаружи камеры указывали на радиационное повреждение. Более тщательное изучение данных показало, что тонкостенные волокна из ранней древесины пострадали больше, чем другие типы волокон, что подтверждает первое предположение, поскольку глубина проникновения электронного луча должна быть одинаковой для всех трех типов волокон.Это означает, что доля радиационных повреждений выше для тонких, чем для толстостенных образцов.
Более тщательное изучение данных показало, что тонкостенные волокна из ранней древесины пострадали больше, чем другие типы волокон, что подтверждает первое предположение, поскольку глубина проникновения электронного луча должна быть одинаковой для всех трех типов волокон.Это означает, что доля радиационных повреждений выше для тонких, чем для толстостенных образцов.
На рис. 2 показан типичный образец деформации и разрушения одного волокна из ранней древесины при растяжении и соответствующая кривая нагрузка-смещение.
Рис. 2Характерная картина деформации волокна из ранней древесины и соответствующая кривая сила-смещение; бар 50 мкм; a – f на схеме относятся к соответствующим микрофотографиям, которые представлены ниже; стрелки in b указывают на начальное складывание / разрушение клеточной стенки
В начальной части кривой наклон был менее крутым из-за дополнительной деформации из-за регулировки узла точечных отверстий лиственных рамок, несущих волокно.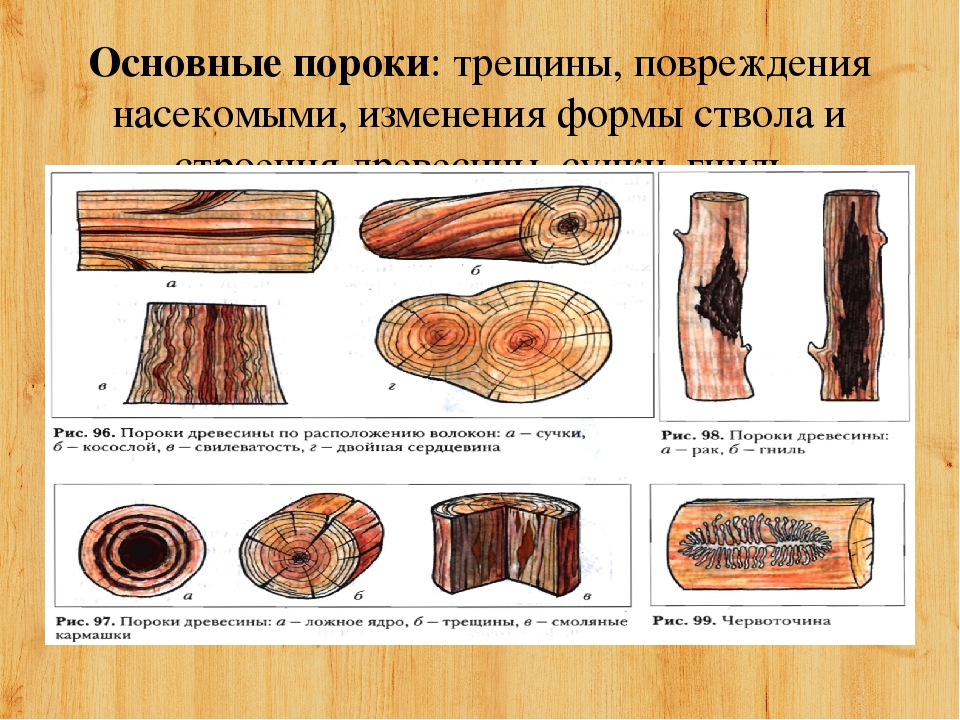 Падение нагрузки на кривой «сила-перемещение» было вызвано остановкой двигателя, необходимого для получения изображения; уменьшение силы было связано с релаксацией напряжения во время паузы. На микрофотографии 2а показан участок ненагруженного волокна. Видимая деформация началась со складывания / разрушения клеточной стенки при довольно низких уровнях нагрузки и деформации, показанных стрелками на рис. 2b. Дальнейшее растяжение привело к образованию трещин в районе этих складок. Регистрация одновременного усилия показала частичное падение нагрузки при появлении первых трещин.Для полного отделения волокна требовалось дополнительное напряжение. Удивительно, но образование трещин не обязательно происходило в непосредственной близости от карьеров или полей карьера.
Падение нагрузки на кривой «сила-перемещение» было вызвано остановкой двигателя, необходимого для получения изображения; уменьшение силы было связано с релаксацией напряжения во время паузы. На микрофотографии 2а показан участок ненагруженного волокна. Видимая деформация началась со складывания / разрушения клеточной стенки при довольно низких уровнях нагрузки и деформации, показанных стрелками на рис. 2b. Дальнейшее растяжение привело к образованию трещин в районе этих складок. Регистрация одновременного усилия показала частичное падение нагрузки при появлении первых трещин.Для полного отделения волокна требовалось дополнительное напряжение. Удивительно, но образование трещин не обязательно происходило в непосредственной близости от карьеров или полей карьера.
Для волокон переходной древесины наблюдались два типичных механизма разрушения. Что касается первой картины разрушения, волокна были разделены на две части трещиной, непосредственно пересекающей всю ячейку, как показано на рис. 3. При ~ 60% предельной нагрузки не наблюдалось явной картины деформации (3b). Даже незадолго до смертельного перелома зарождения трещины не было видно.
3. При ~ 60% предельной нагрузки не наблюдалось явной картины деформации (3b). Даже незадолго до смертельного перелома зарождения трещины не было видно.
Картина деформации переходного древесного волокна и соответствующая кривая силы-смещения, бар 50 мкм
Второй типичный образец деформации волокон переходной древесины показан на рис. 4. Видимая деформация началась со складывания ячейки, которое преимущественно происходило вблизи окаймленных ямок [верхняя часть волокна на микрофотографии (4b)] и ямок. поля [нижняя часть волокна на микрофотографии (4b)].
Фиг.4Устойчивость переходного древесного волокна при растяжении и соответствующая кривая силы-смещения, бар 50 мкм
Подобно волокнам из ранней древесины, частичное падение нагрузки наблюдалось, когда первые трещины появлялись в среде карьера (4c) и волокно начало разрушаться. Для полного разделения волокна требовалось дополнительное натяжение (4г). На микрофотографии 4д показан путь разрушения трещины. Трещина распространялась через ямочное поле частично параллельно продольной оси волокна.В отличие от волокон ранней древесины, складывание стенок ячеек и последующее разрушение начинались при значительно более высоких уровнях нагрузки и начинались вблизи структурных слабых мест, в данном случае ямочного поля.
Для полного разделения волокна требовалось дополнительное натяжение (4г). На микрофотографии 4д показан путь разрушения трещины. Трещина распространялась через ямочное поле частично параллельно продольной оси волокна.В отличие от волокон ранней древесины, складывание стенок ячеек и последующее разрушение начинались при значительно более высоких уровнях нагрузки и начинались вблизи структурных слабых мест, в данном случае ямочного поля.
Обычная картина излома волокон поздней древесины (рис. 5) была аналогична той, что показана на рис. 3 (переходное древесное волокно), с трещиной, разделяющей целую клетку в поперечном направлении без предварительной видимой деформации.
Рис. 5Характерная картина деформации волокна поздней древесины и соответствующая кривая сила-смещение, бар 50 мкм
В дополнение к описанному механизму разрушения для волокон, содержащих большее количество ямок или ямок, наблюдалось складывание ячейки при высоких уровнях нагрузки. В некоторых случаях наблюдали небольшое перекручивание ячеек поздней древесины во время деформации, хотя крутильное движение ограничивалось фиксацией концов волокон клеем. Предположительно, структурно слабые места вдоль волокна могут способствовать описанному поведению кручения.
В некоторых случаях наблюдали небольшое перекручивание ячеек поздней древесины во время деформации, хотя крутильное движение ограничивалось фиксацией концов волокон клеем. Предположительно, структурно слабые места вдоль волокна могут способствовать описанному поведению кручения.
Предел прочности на разрыв трех типов волокон, измеренный на месте в камере ESEM, нельзя сравнивать напрямую, потому что радиационное повреждение, вероятно, повлияло на предельные силы. Однако испытания ex situ ясно показали, что тонкие волокна ранней древесины были менее прочными, чем толстостенные волокна (Таблица 1).Аналогичным образом Mott et al. (2002) обнаружили более низкую прочность на разрыв для волокон ранней древесины, когда они тестировали делигнифицированные волокна, которые сгибаются и изгибаются еще до деформации при растяжении из-за потери поперечной стабильности, вызванной удалением лигнина. Они объяснили более низкую прочность на разрыв ранней древесины более высоким углом микрофибриллы и питтингом. Также был обнаружен немного более высокий угол микрофибриллы целлюлозы в ранней древесине по сравнению с переходной древесиной и поздней древесиной (Таблица 1), но из-за наблюдаемых структур трещин в наших данных было принято предположение, что различия между ранней древесиной, переходной древесиной и поздней древесиной не могут быть полностью объяснены. небольшими различиями в угле и питтинге микрофибрилл целлюлозы.Исследования деформации в ESEM показали, что коробление при растяжении имеет место в тонкостенных волокнах и что вероятность коробления уменьшается с уменьшением размера ячеек и увеличением толщины стенок ячеек. Сравнивая характер разрушения трех типов волокон, можно сказать, что трещины в тонкостенных ячейках в основном были вызваны складыванием ячеек и их последующим разрушением, что в литературе называется «коробление при растяжении» (Jeronimidis 1980; Pagano and Halpin 1968; Page et al. 1971). . С увеличением толщины стенок ячеек и уменьшением размера ячеек видимые структурные слабые места, такие как окаймленные ямы или поля ямок, становились все более актуальными с точки зрения зарождения трещин.
Также был обнаружен немного более высокий угол микрофибриллы целлюлозы в ранней древесине по сравнению с переходной древесиной и поздней древесиной (Таблица 1), но из-за наблюдаемых структур трещин в наших данных было принято предположение, что различия между ранней древесиной, переходной древесиной и поздней древесиной не могут быть полностью объяснены. небольшими различиями в угле и питтинге микрофибрилл целлюлозы.Исследования деформации в ESEM показали, что коробление при растяжении имеет место в тонкостенных волокнах и что вероятность коробления уменьшается с уменьшением размера ячеек и увеличением толщины стенок ячеек. Сравнивая характер разрушения трех типов волокон, можно сказать, что трещины в тонкостенных ячейках в основном были вызваны складыванием ячеек и их последующим разрушением, что в литературе называется «коробление при растяжении» (Jeronimidis 1980; Pagano and Halpin 1968; Page et al. 1971). . С увеличением толщины стенок ячеек и уменьшением размера ячеек видимые структурные слабые места, такие как окаймленные ямы или поля ямок, становились все более актуальными с точки зрения зарождения трещин. Следовательно, более низкая прочность на разрыв тонкостенных волокон из ранней древесины может быть объяснена короблением при растяжении.
Следовательно, более низкая прочность на разрыв тонкостенных волокон из ранней древесины может быть объяснена короблением при растяжении.
Пагано и Халпин (1968) изучали механизмы потери устойчивости при растяжении с помощью тонкостенных анизотропных цилиндров, состоящих из нейлоновых волокон, заключенных в резиновую матрицу под разными углами спирали. Испытания на растяжение с ограничением вращения образцов привели к короблению цилиндров. Аналогичным образом, потеря устойчивости при растяжении наблюдалась при испытаниях отдельных волокон (Пейдж и др., 1971).Пейдж и Эль-Хоссейни (1983) описали коробление при растяжении тонкостенных волокон целлюлозы и объяснили сопротивление толстостенных волокон короблению критическим напряжением потери устойчивости, которое зависит от реакции волокна на напряжения изгиба в клеточной стенке.
Жесткие целлюлозные фибриллы внутри деревянной клеточной стенки имеют тенденцию выравниваться в направлении нагрузки во время растяжения (Gordon and Jeronimidis 1980, Keckes et al. 2003). Возникающие в результате напряжения сдвига заставляют тонкостенную трубку изгибаться под действием растягивающей нагрузки.Однако коробление при растяжении не представляет проблемы для искусственно изготовленных тонких цилиндрических элементов, армированных волокном, поскольку эти трубки обычно состоят из плетеных волокон или волокон, намотанных под углами + Θ и −Θ к продольной оси, что приводит к менее анизотропной системе. (например, Харт и Флек, 2000; Соден и др., 1993). Что касается растяжения древесных тканей, изгибу отдельного волокна внутри ткани препятствуют соседние клетки из-за их в основном прямоугольной формы и z-спиральной ориентации микрофибрилл целлюлозы, что приводит к встречно вращающимся фибриллам целлюлозы двух соседних клеточных стенок, склеенных вместе. сложными средними ламелями, (например,г. Букер 1996; Lichtenegger et al. 1999).
2003). Возникающие в результате напряжения сдвига заставляют тонкостенную трубку изгибаться под действием растягивающей нагрузки.Однако коробление при растяжении не представляет проблемы для искусственно изготовленных тонких цилиндрических элементов, армированных волокном, поскольку эти трубки обычно состоят из плетеных волокон или волокон, намотанных под углами + Θ и −Θ к продольной оси, что приводит к менее анизотропной системе. (например, Харт и Флек, 2000; Соден и др., 1993). Что касается растяжения древесных тканей, изгибу отдельного волокна внутри ткани препятствуют соседние клетки из-за их в основном прямоугольной формы и z-спиральной ориентации микрофибрилл целлюлозы, что приводит к встречно вращающимся фибриллам целлюлозы двух соседних клеточных стенок, склеенных вместе. сложными средними ламелями, (например,г. Букер 1996; Lichtenegger et al. 1999).
Доказательства ближнего спирального порядка в 30-нм хроматиновых волокнах ядер эритроцитов
Abstract
Сворачивание хроматина у эукариот укладывается в геном в ограниченный объем ядра клетки. Формирование структур хроматина более высокого порядка снижает доступность ДНК, тем самым внося свой вклад в контроль основных функций генома, таких как транскрипция, репликация и репарация ДНК. Считается, что 30-нм волокно является первым иерархическим уровнем сворачивания хроматина, но расположение нуклеосом в компактном 30-нм волокне ранее было неизвестно.Мы использовали криоэлектронную томографию срезов стекловидного тела для определения структуры компактного нативного 30-нм волокна ядер эритроцитов птиц. Преобладающая геометрия 30-нм волокна, выявленная при усреднении субтомограмм, представляет собой левостороннюю двухзаходную спираль с примерно 6,5 нуклеосом на 11 нм, в которой нуклеосомы расположены лицом к лицу, но смещены от своих сверхспиральных осей на осевое перемещение примерно 3,4 нм и азимутальное вращение примерно 54 °. Нуклеосомы образуют шахматный узор при наблюдении в направлении, перпендикулярном оси волокна, но не пересекаются между собой.Упаковка нуклеосом внутри волокон показывает большие межнуклеосомные расстояния от центра к центру, чем предполагалось ранее, что исключает возможность взаимодействий между ядрами и объясняет, как факторы транскрипции и регуляции могут получать доступ к нуклеосомам.
Формирование структур хроматина более высокого порядка снижает доступность ДНК, тем самым внося свой вклад в контроль основных функций генома, таких как транскрипция, репликация и репарация ДНК. Считается, что 30-нм волокно является первым иерархическим уровнем сворачивания хроматина, но расположение нуклеосом в компактном 30-нм волокне ранее было неизвестно.Мы использовали криоэлектронную томографию срезов стекловидного тела для определения структуры компактного нативного 30-нм волокна ядер эритроцитов птиц. Преобладающая геометрия 30-нм волокна, выявленная при усреднении субтомограмм, представляет собой левостороннюю двухзаходную спираль с примерно 6,5 нуклеосом на 11 нм, в которой нуклеосомы расположены лицом к лицу, но смещены от своих сверхспиральных осей на осевое перемещение примерно 3,4 нм и азимутальное вращение примерно 54 °. Нуклеосомы образуют шахматный узор при наблюдении в направлении, перпендикулярном оси волокна, но не пересекаются между собой.Упаковка нуклеосом внутри волокон показывает большие межнуклеосомные расстояния от центра к центру, чем предполагалось ранее, что исключает возможность взаимодействий между ядрами и объясняет, как факторы транскрипции и регуляции могут получать доступ к нуклеосомам.
Основной повторяющейся единицей хроматина является нуклеосома, структура которой известна с атомным разрешением (1). In vitro в присутствии линкерных гистонов и умеренной концентрации одновалентных катионов массивы нуклеосом принимают неправильную зигзагообразную топологию (2, 3).Дальнейшее увеличение количества одновалентной соли и / или присутствие двухвалентных катионов приводит к более компактным структурам, называемым 30-нм хроматиновыми волокнами, в которых предполагается, что нуклеосомы спирально упакованы с плотностью в диапазоне 6-11 нуклеосом на оборот. (4, 5). Биологическая значимость этого уплотнения подтверждается прямой видимостью 30-нм волокон in situ с помощью электронной микроскопии (ЭМ) в некоторых типах клеток эволюционно далеких видов (6-8). Несмотря на это фундаментальное знание, разрешение электронных микрофотографий было недостаточным для визуализации организации отдельных нуклеосом в компактном 30-нм волокне.Таким образом, расположение нуклеосом внутри волокна и, соответственно, интерпретация основных функций хроматина была основана на косвенных методах, что привело к различным моделям, которые до сих пор оспаривались (9).
Были предложены два основных типа моделей: первая модель представляет собой однозарядную спираль, в которой последовательные нуклеосомы расположены в виде соленоида (4, 10), а вторая модель представляет собой скрученную двухстартовую спираль, образованную зигзагом. лента нуклеосом (11). Самые последние свидетельства в пользу модели одного старта получены в результате измерений двумерных (2D) электронных микроскопических изображений массивов нуклеосом, восстановленных in vitro, которые предполагают встречно-гребенчатый соленоид с линкерной ДНК, изогнутой внутри волокна (12).Двухстартовая модель была недавно подтверждена кристаллографическим исследованием тетрануклеосомы (13), а также экспериментами по перекрестному связыванию (14).
Криоэлектронная томография обладает уникальной способностью визуализировать молекулярные сборки в неизмененном замороженном гидратированном состоянии в условиях, близких к естественным. При обычной электронной микроскопии пластиковых срезов образец химически фиксируют, обезвоживают, заливают смолой и окрашивают. Эти стадии, особенно процесс дегидратации, приводят к структурной перестройке, которая особенно заметна для полиэлектролитов, таких как молекулы ДНК (15).Другой недостаток традиционной микроскопии заключается в том, что просматривается окружающее пятно, а не сама биологическая структура. В криоэлектронной томографии образцы замораживают под высоким давлением, а сохраненный замороженно-гидратированный образец затем разрезают при криотемпературах без дегидратации. Поскольку образцы неокрашены, интенсивности, наблюдаемые на изображениях застеклованных образцов, представляют собой истинные плотности макромолекул. Здесь мы используем криоэлектронную томографию срезов стекловидного тела для прямой визуализации расположения нуклеосом в уплотненных 30-нм волокнах зрелых ядер эритроцитов курицы без использования фиксаторов, дегидратации или каких-либо сшивающих агентов, используемых в обычной электронной микроскопии и других методах.
Эти стадии, особенно процесс дегидратации, приводят к структурной перестройке, которая особенно заметна для полиэлектролитов, таких как молекулы ДНК (15).Другой недостаток традиционной микроскопии заключается в том, что просматривается окружающее пятно, а не сама биологическая структура. В криоэлектронной томографии образцы замораживают под высоким давлением, а сохраненный замороженно-гидратированный образец затем разрезают при криотемпературах без дегидратации. Поскольку образцы неокрашены, интенсивности, наблюдаемые на изображениях застеклованных образцов, представляют собой истинные плотности макромолекул. Здесь мы используем криоэлектронную томографию срезов стекловидного тела для прямой визуализации расположения нуклеосом в уплотненных 30-нм волокнах зрелых ядер эритроцитов курицы без использования фиксаторов, дегидратации или каких-либо сшивающих агентов, используемых в обычной электронной микроскопии и других методах.
Результаты
Крио-ЭМ ядер куриных эритроцитов.

Зрелые ядра эритроцитов птиц, как хорошо известно, содержат 30-нм хроматиновые волокна при выделении с ионной силой, близкой к физиологической, например в буфере, содержащем 2 мМ MgCl 2 и 75 мМ одновалентных катионов (16). Мы изолировали ядра куриных эритроцитов в таком буфере и витрифицировали их в буфере, содержащем 20 мМ NaCl с 20% декстраном 40 кДа. Осмотический эффект этого конкретного декстрана поддерживает уплотнение волокна, эквивалентное ионной силе 2 мМ MgCl 2 (17) (рис.S1). Ядра были неповрежденными и сохраняли свою форму за счет процедур изоляции и витрификации. Ядра куриных эритроцитов полезны для анализа данных, потому что ( и ) они содержат небольшое количество негистонового хромосомного белка и, следовательно, не содержат других белковых комплексов, которые могли бы повлиять на обработку изображения (18), и ( ii ) межволоконное расстояние составляет достаточно большой, чтобы волокна можно было извлекать отдельно. Хроматин эритроцитов курицы содержит полный набор линкерного гистона h2 (0. 4 h2 на нуклеосому) и H5 (0,9 на нуклеосому) (19) и является представителем транскрипционно неактивного хроматина (20). В этом исследовании мы количественно анализируем срезы стекловидного тела этих ядер, которые показывают отчетливые 30-нм волокна как в 2D-проекциях, так и в трехмерных (3D) реконструкциях (рис. 1 A — F ).
4 h2 на нуклеосому) и H5 (0,9 на нуклеосому) (19) и является представителем транскрипционно неактивного хроматина (20). В этом исследовании мы количественно анализируем срезы стекловидного тела этих ядер, которые показывают отчетливые 30-нм волокна как в 2D-проекциях, так и в трехмерных (3D) реконструкциях (рис. 1 A — F ).
Крио-ЭМ изображения хроматина в куриных эритроцитах. ( A ) Двухмерное обзорное изображение криогенного среза ядра куриного эритроцита толщиной примерно 50 нм.Поперечные сечения волокон (виды сверху) имеют более высокий контраст по сравнению с продольными сечениями волокон и, таким образом, намного более очевидны, но видны все возможные ориентации. Некоторые продольно разрезанные волокна выделены красными прямоугольниками; некоторые волокна в поперечном сечении обозначены красными кружками. Длина бокса составляет 200 нм, а ширина — 50 нм. Вставка: 1D осредненный по вращению спектр мощности ядра показывает сильный пик в диапазоне 30–40 нм. ( B ) Срез толщиной 10 нм, полученный при трехмерной реконструкции ядра эритроцита курицы.Ядро содержит компактные, но отчетливые и хорошо разделенные 30-нм волокна, в которых видны отдельные нуклеосомы. Продольное волокно и поперечное сечение волокна обозначены красным прямоугольником и красным кружком соответственно. ( C и D ) Поперечные сечения отдельных волокон; срезы имеют толщину 1,2 нм. ( E и F ) Продольные срезы отдельных волокон; срезы имеют толщину 1,2 нм. Размер и форма позиций с высокой плотностью хорошо соответствуют отдельным нуклеосомам.
( B ) Срез толщиной 10 нм, полученный при трехмерной реконструкции ядра эритроцита курицы.Ядро содержит компактные, но отчетливые и хорошо разделенные 30-нм волокна, в которых видны отдельные нуклеосомы. Продольное волокно и поперечное сечение волокна обозначены красным прямоугольником и красным кружком соответственно. ( C и D ) Поперечные сечения отдельных волокон; срезы имеют толщину 1,2 нм. ( E и F ) Продольные срезы отдельных волокон; срезы имеют толщину 1,2 нм. Размер и форма позиций с высокой плотностью хорошо соответствуют отдельным нуклеосомам.
Усреднение субтомограмм нативных 30-нм волокон из куриных эритроцитов.
Усреднение субтомограмм включает в себя выравнивание множества похожих трехмерных изображений (так называемых субтомограмм) с эталонным шаблоном, который итеративно улучшает отношение сигнал / шум данных до сходимости (21). Несмотря на то, что отличное качество реконструкции позволяет нам непосредственно визуализировать нуклеосомы в реконструкции (Рис. 1 C — F ), процедура усреднения выполняется для улучшения отношения сигнал / шум и усреднения. недостающий клин.Мы обнаружили волокна в томографических реконструкциях путем взаимной корреляции с фильтрованным бинарным цилиндром, диаметром 30 нм и длиной 60 нм, который вращался и перемещался в пространстве (22).
1 C — F ), процедура усреднения выполняется для улучшения отношения сигнал / шум и усреднения. недостающий клин.Мы обнаружили волокна в томографических реконструкциях путем взаимной корреляции с фильтрованным бинарным цилиндром, диаметром 30 нм и длиной 60 нм, который вращался и перемещался в пространстве (22).
Длина цилиндра шаблона была выбрана равной 60 нм, чтобы избежать выбора видов сверху в качестве видов сбоку. Поскольку цилиндр не содержит какой-либо структурно значимой информации, любые особенности с высоким разрешением, возникающие после выравнивания, должны исходить из самих данных, что позволяет полностью избежать смещения шаблона.Таким образом, мы автоматически извлекли приблизительно 1000 субтомограмм 30-нм волокон и применили процедуру выравнивания субтомограмм и усреднения (подробный протокол см. В SI Methods ). Это включало выравнивание каждой извлеченной 3D-субтомограммы с начальным бинарным цилиндром, где отсутствующий клин учитывался и компенсировался как в алгоритмах выравнивания, так и в алгоритмах усреднения (21), пока структура не достигла сходимости. Первоначально мы классифицировали субтомограммы, чтобы удалить ложные срабатывания, а также волокна с неправильной ориентацией оси волокна (21, 23).Среднее значение субтомограммы с полной длиной 60 нм имело низкое разрешение из-за структурной неоднородности волокон на большом расстоянии, но при маскировке в центральной области примерно 40 нм вдоль оси волокна разрешение улучшалось. Это указывает на то, что спиральный порядок в волокне явно краткосрочный (не более 3 круговоротов нуклеосом). Субтомограммы были повторно выровнены с использованием этой маски с угловым ограничением, которое предотвращало поворот волокон более чем на 30 ° от их первоначально рассчитанной оси волокна.
Первоначально мы классифицировали субтомограммы, чтобы удалить ложные срабатывания, а также волокна с неправильной ориентацией оси волокна (21, 23).Среднее значение субтомограммы с полной длиной 60 нм имело низкое разрешение из-за структурной неоднородности волокон на большом расстоянии, но при маскировке в центральной области примерно 40 нм вдоль оси волокна разрешение улучшалось. Это указывает на то, что спиральный порядок в волокне явно краткосрочный (не более 3 круговоротов нуклеосом). Субтомограммы были повторно выровнены с использованием этой маски с угловым ограничением, которое предотвращало поворот волокон более чем на 30 ° от их первоначально рассчитанной оси волокна.
Результирующая структура на средней субтомограмме представляет собой волокно диаметром примерно 32 нм и длиной примерно 60 нм (рис. S2), содержащее дискретные эллиптические плотности, размер которых соответствует отдельным нуклеосомам. Пространственное положение и ориентация нуклеосом можно четко распознать (рис.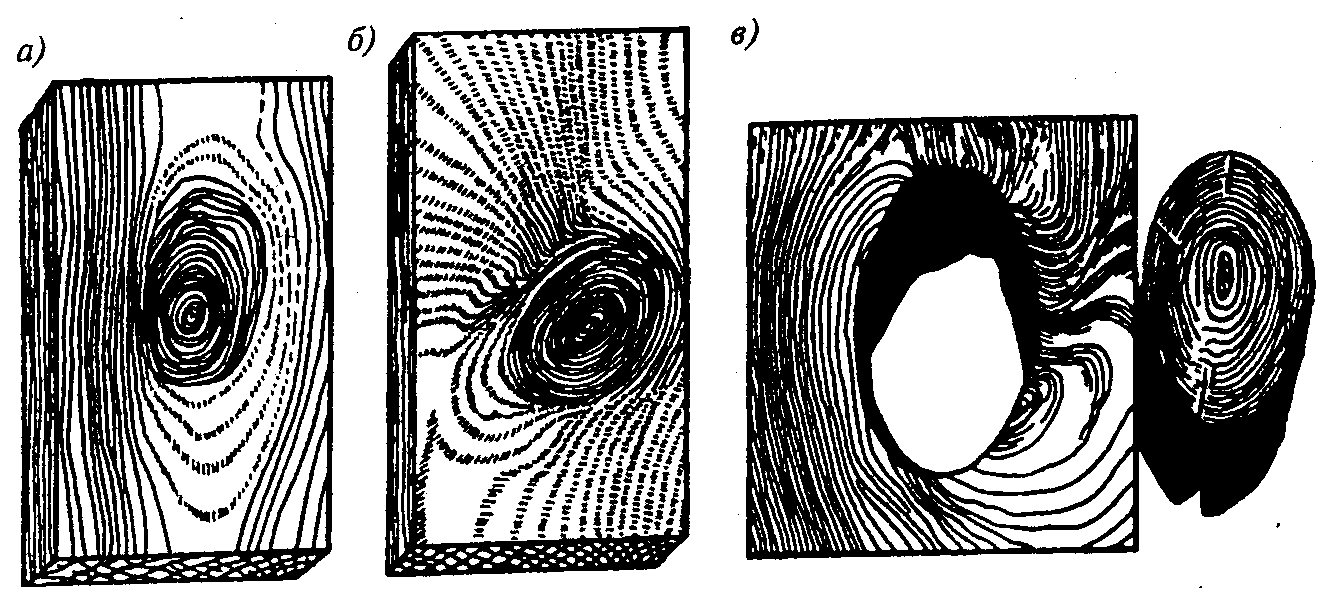 2). Поперечный разрез средней субтомограммы показывает шесть радиально организованных нуклеосом, расположенных на равном расстоянии друг от друга в плоскости (рис. 2 E и F ).На 11 нм приходится 6,5 нуклеосом, в результате чего шаг спирали составляет примерно 22,8 нм. Нуклеосомы демонстрируют шахматный узор, если смотреть на волокна сбоку (рис. 2 C и 5 A ), что указывает на то, что нуклеосомы расположены выше и между нуклеосомами предыдущего круговорота, без пересечения. График распределения углов Эйлера (рис. S3) показывает, что существует полный охват углов, что приводит к изотропной функции рассеяния точки. Поскольку ориентации волокон распределены равномерно и недостающий клин был компенсирован, среднее значение субтомограммы не содержит какой-либо недостающей информации в пространстве Фурье.Разрешение средней субтомограммы, оцененной корреляцией Фурье-оболочки при отсечении 0,5, составляет 4,3 нм (рис. S4).
2). Поперечный разрез средней субтомограммы показывает шесть радиально организованных нуклеосом, расположенных на равном расстоянии друг от друга в плоскости (рис. 2 E и F ).На 11 нм приходится 6,5 нуклеосом, в результате чего шаг спирали составляет примерно 22,8 нм. Нуклеосомы демонстрируют шахматный узор, если смотреть на волокна сбоку (рис. 2 C и 5 A ), что указывает на то, что нуклеосомы расположены выше и между нуклеосомами предыдущего круговорота, без пересечения. График распределения углов Эйлера (рис. S3) показывает, что существует полный охват углов, что приводит к изотропной функции рассеяния точки. Поскольку ориентации волокон распределены равномерно и недостающий клин был компенсирован, среднее значение субтомограммы не содержит какой-либо недостающей информации в пространстве Фурье.Разрешение средней субтомограммы, оцененной корреляцией Фурье-оболочки при отсечении 0,5, составляет 4,3 нм (рис. S4).
Средняя субтомограмма волокон эритроцитов курицы. ( A ) Изображение расположения и взаимного расположения срезов и изоповерхности, визуализированное в B — H . ( B ) Расчетное изображение проекции, параллельное оси волокна, через среднее значение субтомограммы. Видны шесть радиально ориентированных эллиптических плотностей, окружающих центральную плотность.Размер одной из радиально расположенных плотностей соответствует размерам отдельной нуклеосомы. ( C ) Компьютерное проекционное изображение, рассчитанное перпендикулярно оси волокна через периферию волокна, показывает узор нуклеосом в шахматном порядке. ( D ) Продольный срез толщиной 0,6 нм, показывающий соседние нуклеосомы вне суперпиральной оси, присутствующие в волокне. ( E и F ) Два среза толщиной 0,6 нм на расстоянии 10 нм друг от друга, показывающие радиально расположенные нуклеосомы.( G ) Изоповерхность средней субтомограммы показана в перспективе. Визуализируются два круговорота волокна. Указаны ось волокна и расстояние между нуклеосомами.
( A ) Изображение расположения и взаимного расположения срезов и изоповерхности, визуализированное в B — H . ( B ) Расчетное изображение проекции, параллельное оси волокна, через среднее значение субтомограммы. Видны шесть радиально ориентированных эллиптических плотностей, окружающих центральную плотность.Размер одной из радиально расположенных плотностей соответствует размерам отдельной нуклеосомы. ( C ) Компьютерное проекционное изображение, рассчитанное перпендикулярно оси волокна через периферию волокна, показывает узор нуклеосом в шахматном порядке. ( D ) Продольный срез толщиной 0,6 нм, показывающий соседние нуклеосомы вне суперпиральной оси, присутствующие в волокне. ( E и F ) Два среза толщиной 0,6 нм на расстоянии 10 нм друг от друга, показывающие радиально расположенные нуклеосомы.( G ) Изоповерхность средней субтомограммы показана в перспективе. Визуализируются два круговорота волокна. Указаны ось волокна и расстояние между нуклеосомами. ( H ) Изоповерхность среднего значения субтомограммы, очерченная на двух пороговых уровнях, чтобы указать центр положений нуклеосом на верхнем пороге и появление 30-нм волокна на нижнем пороге. Изоповерхность на верхнем пороге показывает различные плотности в эллиптической форме, изображая центр положений отдельных нуклеосом.Изоповерхность на нижнем пороге была выбрана для точного представления полной нуклеосомы и показана как прозрачная, псевдоцветная в соответствии с радиусом цилиндра. Красным обозначены позиции на внешнем радиусе волокна, а зелено-синие позиции — ближе к центру волокна. Положения нуклеосом хорошо определены.
( H ) Изоповерхность среднего значения субтомограммы, очерченная на двух пороговых уровнях, чтобы указать центр положений нуклеосом на верхнем пороге и появление 30-нм волокна на нижнем пороге. Изоповерхность на верхнем пороге показывает различные плотности в эллиптической форме, изображая центр положений отдельных нуклеосом.Изоповерхность на нижнем пороге была выбрана для точного представления полной нуклеосомы и показана как прозрачная, псевдоцветная в соответствии с радиусом цилиндра. Красным обозначены позиции на внешнем радиусе волокна, а зелено-синие позиции — ближе к центру волокна. Положения нуклеосом хорошо определены.
Сопоставление нуклеосом и идеализированное волокно после наложения симметрии. ( A и B ) Два класса из процедуры классификации мононуклеосом, отображающие узор в виде шахматной доски, который полностью совместим с видами сбоку среднего значения субтомограммы, если смотреть в направлении, перпендикулярном оси волокна.Классы имеют немного отличающуюся организацию нуклеосом, но оба показывают нуклеосомы, которые сопоставлены лицом к лицу, но смещены от своих суперспиральных осей.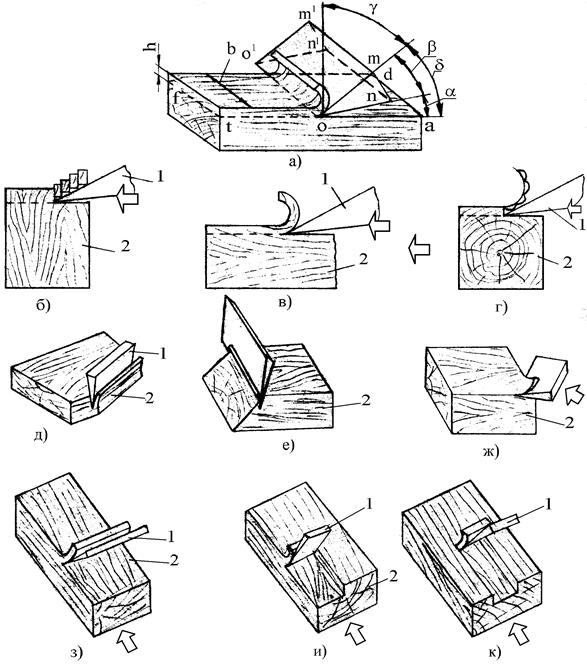 Сверхспиральная ось параллельна оси x. ( C ) Репрезентативный класс из процедуры классификации мононуклеосом, изображающий прямое наложение нуклеосом по оси, которое отличается от того, что видно на средней субтомограмме, что может быть связано с локальными искажениями спиральной структуры.( D ) Изоповерхность идеализированного волокна после симметризации на основе определенных параметров спирали. ( E ) Индивидуальные нуклеосомы помещаются в идеализированное волокно (визуализируется на прозрачной бело-голубой изоповерхности). Ось волокна и суперспиральная ось нуклеосомы указаны стрелками.
Сверхспиральная ось параллельна оси x. ( C ) Репрезентативный класс из процедуры классификации мононуклеосом, изображающий прямое наложение нуклеосом по оси, которое отличается от того, что видно на средней субтомограмме, что может быть связано с локальными искажениями спиральной структуры.( D ) Изоповерхность идеализированного волокна после симметризации на основе определенных параметров спирали. ( E ) Индивидуальные нуклеосомы помещаются в идеализированное волокно (визуализируется на прозрачной бело-голубой изоповерхности). Ось волокна и суперспиральная ось нуклеосомы указаны стрелками.
Определение параметров симметрии.
Мы не вводили симметрию во время итеративной процедуры выравнивания, таким образом, предположение о спиральной симметрии было полностью исключено.Когда автокорреляционная функция среднего значения субтомограммы построена в цилиндрических координатах, она показывает замечательную периодичность на радиусе 13 нм (рис. 3 A ). В цилиндрических координатах пики автокорреляции можно рассматривать как точки решетки спиральной сети, которая может быть проиндексирована векторами направления (Рис. 3 B ). Вектор направления на самый высокий пик корреляции (HD) и вектор второго самого высокого пика корреляции (SD) показаны на фиг. 3 B .HD показывает направление основной спирали (обозначено розовой стрелкой), по которому можно рассчитать расстояние повторения нуклеосом. SD указывает расстояние между спиралями спирали (второй круговорот обозначен синей стрелкой) (рис. 3 B ). Примечательно, что автокорреляционная функция показывает отдельные точки решетки вплоть до третьего порядка, захватывая все нуклеосомы в пределах двух круговоротов, показывая, что среднее значение субтомограммы имеет примерно 6,5-кратную симметрию (рис. 3 C ). И первая, и вторая спирали пересекают горизонтальную линию (экватор), что свидетельствует о том, что двухзаходная спираль — это наименьшее возможное начало.
В цилиндрических координатах пики автокорреляции можно рассматривать как точки решетки спиральной сети, которая может быть проиндексирована векторами направления (Рис. 3 B ). Вектор направления на самый высокий пик корреляции (HD) и вектор второго самого высокого пика корреляции (SD) показаны на фиг. 3 B .HD показывает направление основной спирали (обозначено розовой стрелкой), по которому можно рассчитать расстояние повторения нуклеосом. SD указывает расстояние между спиралями спирали (второй круговорот обозначен синей стрелкой) (рис. 3 B ). Примечательно, что автокорреляционная функция показывает отдельные точки решетки вплоть до третьего порядка, захватывая все нуклеосомы в пределах двух круговоротов, показывая, что среднее значение субтомограммы имеет примерно 6,5-кратную симметрию (рис. 3 C ). И первая, и вторая спирали пересекают горизонтальную линию (экватор), что свидетельствует о том, что двухзаходная спираль — это наименьшее возможное начало.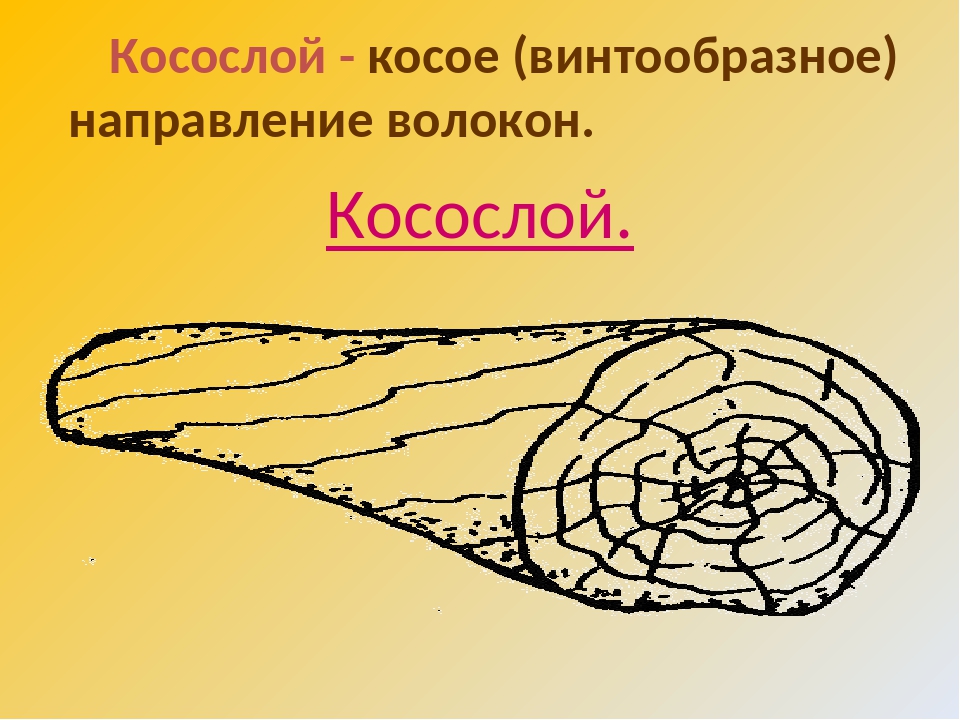
Волокно показывает двухстартовую организацию. ( A ) Автокорреляционная функция среднего значения субтомограммы в цилиндрических координатах на радиусе 13 нм. Красные сферы, обозначающие положения локальных максимумов, демонстрируют замечательную периодичность и используются для определения узлов решетки. ( B ) Идеализированная решетка, показанная серыми сферами, была рассчитана на основе положений локальных максимумов. В идеализированной решетке первая спираль обозначена розовой стрелкой, а вторая спираль указана синей стрелкой.Позиции нуклеосом, видимые в реальных данных, индексируются небольшими числами. Обе спирали пересекают экватор в точках, обозначенных розовым и синим кружками соответственно. Это показывает, что волокно имеет четкую двухзаходную геометрию. ( C ) График автокорреляционной функции в положении основной спирали показывает, что среднее значение субтомограммы имеет примерно 6,5-кратную симметрию с поворотом на субъединицу примерно 54 °. ( D ) Изображение идеализированной решетки в декартовом пространстве в B , облегчение индексации нуклеосом на решетке.Нумерация нуклеосом идентична нумерации B. Двухзаходная спираль видна в декартовых координатах. ( E ) Вид сверху положений нуклеосом, размещенных в положениях корреляционных пиков, показанных в B , с линкерной ДНК, смоделированной серым цветом, которая, как предполагается, пересекает ось волокна. Указаны азимутальное вращение и межцентровое расстояние между нуклеосомами. ( F ) Вид сбоку изображения в E (связанный поворотом на 90 °) в стереоскопическом виде с косыми глазами.
( D ) Изображение идеализированной решетки в декартовом пространстве в B , облегчение индексации нуклеосом на решетке.Нумерация нуклеосом идентична нумерации B. Двухзаходная спираль видна в декартовых координатах. ( E ) Вид сверху положений нуклеосом, размещенных в положениях корреляционных пиков, показанных в B , с линкерной ДНК, смоделированной серым цветом, которая, как предполагается, пересекает ось волокна. Указаны азимутальное вращение и межцентровое расстояние между нуклеосомами. ( F ) Вид сбоку изображения в E (связанный поворотом на 90 °) в стереоскопическом виде с косыми глазами.
Путем вывода параметров симметрии мы таким образом определили, что среднее значение субтомограммы представляет собой левостороннюю спираль с аксиальным переносом (Δ z ) на субъединицу 3,4 нм (± 0,48 нм) и азимутальным вращением (Δ ϕ ). ) на субъединицу -54 ° (± 0,4 °), что связывает каждую нуклеосому с ее соседом с межцентровым расстоянием в пределах круговорота 10,3 нм (± 0,12 нм) (рис. 3 и Movie S1). Это дает примерно 6,7 нуклеосом на оборот (рис. 3 E и F ) или примерно 6.5 нуклеосом на 11 нм. Это число указывает на то, что волокна полностью компактны, потому что скорость уплотнения 6-7 нуклеосом на 11 нм виток соответствует точке насыщения уплотнения в изолированных волокнах хроматина эритроцитов курицы, согласно измерениям, предоставленным в предыдущих исследованиях (5, 24). Мы проверили воспроизводимость окончательного среднего значения субтомограммы, используя три отдельных набора данных из разных томограмм с независимыми параметрами инициализации, и результаты были очень схожими.Среди обработанных наборов данных полученные субтомограммы показали примерно 6,5-кратную структуру с Δ z на субъединицу примерно 3,6 нм (± 0,6 нм) и азимутальным вращением (Δ ϕ ) на субъединицу примерно -54 ° (± 3 °) (выравнивание серии наклона показано в таблице S1).
3 и Movie S1). Это дает примерно 6,7 нуклеосом на оборот (рис. 3 E и F ) или примерно 6.5 нуклеосом на 11 нм. Это число указывает на то, что волокна полностью компактны, потому что скорость уплотнения 6-7 нуклеосом на 11 нм виток соответствует точке насыщения уплотнения в изолированных волокнах хроматина эритроцитов курицы, согласно измерениям, предоставленным в предыдущих исследованиях (5, 24). Мы проверили воспроизводимость окончательного среднего значения субтомограммы, используя три отдельных набора данных из разных томограмм с независимыми параметрами инициализации, и результаты были очень схожими.Среди обработанных наборов данных полученные субтомограммы показали примерно 6,5-кратную структуру с Δ z на субъединицу примерно 3,6 нм (± 0,6 нм) и азимутальным вращением (Δ ϕ ) на субъединицу примерно -54 ° (± 3 °) (выравнивание серии наклона показано в таблице S1).
Классификация: вариативность и гибкость волокон.
Наше среднее значение описывает наиболее статистически значимую симметрию, которая характерна для набора данных, но после классификации субтомограмм (21) мы видим, что существует некоторый уровень вариации и гибкости внутри волокон. Чтобы идентифицировать структурную изменчивость, мы классифицировали окончательный набор выровненных частиц с помощью анализа главных компонентов (PCA) с использованием масок длиной 30 нм. Эта процедура классификации показала, что существуют различия в симметриях извлеченных волокон. Поскольку мы игнорировали вариации, меньшие нашего предела разрешения, мы могли убедиться, что идентифицировали истинные структурные вариации, потому что плотности, которые мы классифицировали, были порядка размеров нуклеосомы. Большинство средних значений класса (65%) показывают 6 нуклеосом на поперечное сечение, но другие показывают 5 (11%) или 7 (16%), демонстрируя уровень вариабельности, который может быть отнесен на счет незначительных искажений или изгибов в отдельных волокнах. (Инжир.4). Однако мы никогда не получаем классы с более чем 7 нуклеосом на оборот, что противоречит чрезвычайно высокому коэффициенту упаковки 11 нуклеосом на 11 нм, полученному при измерениях реконструированных массивов in vitro (12).
Чтобы идентифицировать структурную изменчивость, мы классифицировали окончательный набор выровненных частиц с помощью анализа главных компонентов (PCA) с использованием масок длиной 30 нм. Эта процедура классификации показала, что существуют различия в симметриях извлеченных волокон. Поскольку мы игнорировали вариации, меньшие нашего предела разрешения, мы могли убедиться, что идентифицировали истинные структурные вариации, потому что плотности, которые мы классифицировали, были порядка размеров нуклеосомы. Большинство средних значений класса (65%) показывают 6 нуклеосом на поперечное сечение, но другие показывают 5 (11%) или 7 (16%), демонстрируя уровень вариабельности, который может быть отнесен на счет незначительных искажений или изгибов в отдельных волокнах. (Инжир.4). Однако мы никогда не получаем классы с более чем 7 нуклеосом на оборот, что противоречит чрезвычайно высокому коэффициенту упаковки 11 нуклеосом на 11 нм, полученному при измерениях реконструированных массивов in vitro (12). Очевидно, что в природе структурная изменчивость может существовать в пределах заданного набора параметров. Это, конечно, может быть другим случаем для восстановленных волокон, где требуется структурная однородность, даже если это не может быть истинным отражением структуры в природе.
Очевидно, что в природе структурная изменчивость может существовать в пределах заданного набора параметров. Это, конечно, может быть другим случаем для восстановленных волокон, где требуется структурная однородность, даже если это не может быть истинным отражением структуры в природе.
Структурная изменчивость в наборе данных. Показано характеристическое подмножество результирующих средних значений класса субтомограммы из классификации PCA; срезы имеют толщину 1,2 нм. Большинство средних значений класса показывают 6 нуклеосом на поперечное сечение ( B ), но другие показывают 5 ( A ) или 7 нуклеосом на поперечное сечение ( C ), что демонстрирует уровень вариабельности с нашими данными. Нуклеосомы имеют разную плотность, потому что они расчетно разделены на разную высоту.( D ) Круговая диаграмма, показывающая процентное содержание частиц, составляющих каждый класс.
Характеристики собственного 30-нм волокна.
Автокорреляционная функция в цилиндрических координатах однозначно показывает, что волокно имеет двухзаходную геометрию, образованную двумя противоположными спиралями, что согласуется с двухзаходной моделью (рис. 3 B и D ). Симметрия не может быть объяснена спиралью с одним началом, потому что подъем на нуклеосому исключает возможность завершения числа нуклеосом на 11 нм в пределах определенного шага.Для выполнения этого требования потребуется гораздо меньшая аксиальная трансляция (Δ z ) на субъединицу (<1,7 нм). Подгоняя нуклеосомы к плотности с использованием алгоритма, учитывающего параметры симметрии, можно определить, что двухзаходная геометрия волокна образована двумя противоположными спиралями, где вторая спираль начинается под углом 203 ° по отношению к первой. .
3 B и D ). Симметрия не может быть объяснена спиралью с одним началом, потому что подъем на нуклеосому исключает возможность завершения числа нуклеосом на 11 нм в пределах определенного шага.Для выполнения этого требования потребуется гораздо меньшая аксиальная трансляция (Δ z ) на субъединицу (<1,7 нм). Подгоняя нуклеосомы к плотности с использованием алгоритма, учитывающего параметры симметрии, можно определить, что двухзаходная геометрия волокна образована двумя противоположными спиралями, где вторая спираль начинается под углом 203 ° по отношению к первой. .
Из-за разрешения средней субтомограммы мы не можем однозначно определить связь ДНК между нуклеосомами.Однако мы постоянно видим плотности, пересекающие ось волокна на средней субтомограмме и даже на отдельных субтомограммах при благоприятных углах проекции (Рис. 1 C и D ). Расчетные проекции вниз по оси волокна показывают плотность в полости волокна, которая не может быть объяснена доменами h2 / H5 или гистонового хвоста из-за их небольшого размера и расположения (рис. 2 B ) ( SI Methods ). Поскольку ранее не было сообщений о том, что такая плотность визуализируется в реальных данных, мы предполагаем, что это является дополнительным свидетельством того, что ДНК действительно пересекает волокно.
2 B ) ( SI Methods ). Поскольку ранее не было сообщений о том, что такая плотность визуализируется в реальных данных, мы предполагаем, что это является дополнительным свидетельством того, что ДНК действительно пересекает волокно.
Затем мы состыковали нуклеосомы в индивидуально сегментированные плотности нуклеосом средней субтомограммы с помощью методов, основанных на взаимной корреляции (22). Хотя точность ограничена разрешением, стыковка однозначна (рис. S5). Подгонка нуклеосомы в пределах плотности одиночной нуклеосомы показывает, что угол между суперспиральной осью нуклеосомы и осью волокна составляет примерно 90 ° с отклонением примерно 20 ° (Movie S2). Этот угол согласуется с предыдущими значениями, определенными из измерений фотохимического дихроизма для хроматина эритроцитов курицы (10), и разумно согласуется с углом между сверхспиральными осями стопок нуклеосом в идеализированной модели волокна, полученной на основе рентгеновской структуры тетрануклеосом.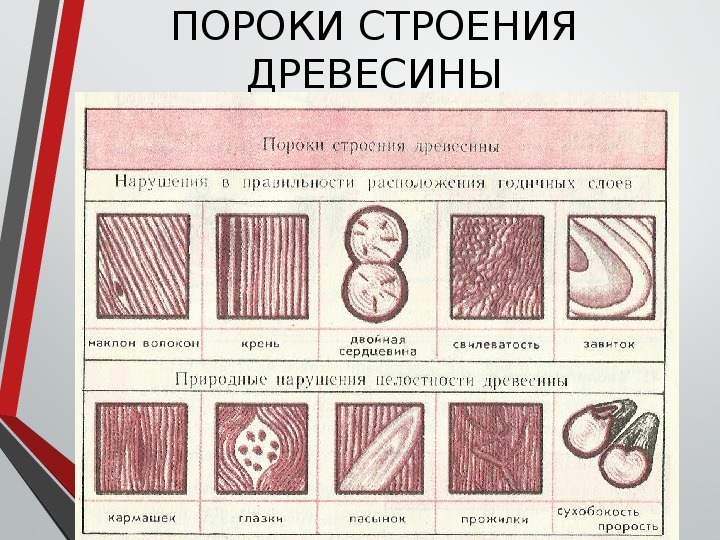 где угол был увеличен до 70 °, чтобы уменьшить стерическую интерференцию между тетрануклеосомами (13).Нуклеосомы в нашем волокне в основном расположены лицом к лицу, но смещены относительно своих сверхспиральных осей. Мы предполагаем, что нуклеосомы пронумерованы в соответствии с линейным массивом, зависящим от связности ДНК, где нуклеосома n2 присоединена к n1 и n3 . В волокне нуклеосомы расположены так, что нуклеосома n8 асимметрично расположена над нуклеосомами n1 и n6 , без пересечения. В то время как нуклеосомы n6 и n8 находятся в одном круговороте, нуклеосома n1 находится в последующем круговороте (рис.3 D и F и Movie S2).
где угол был увеличен до 70 °, чтобы уменьшить стерическую интерференцию между тетрануклеосомами (13).Нуклеосомы в нашем волокне в основном расположены лицом к лицу, но смещены относительно своих сверхспиральных осей. Мы предполагаем, что нуклеосомы пронумерованы в соответствии с линейным массивом, зависящим от связности ДНК, где нуклеосома n2 присоединена к n1 и n3 . В волокне нуклеосомы расположены так, что нуклеосома n8 асимметрично расположена над нуклеосомами n1 и n6 , без пересечения. В то время как нуклеосомы n6 и n8 находятся в одном круговороте, нуклеосома n1 находится в последующем круговороте (рис.3 D и F и Movie S2).
Дифференциальное прямое совмещение.
Чтобы визуализировать локальные области порядка, которые, возможно, не были захвачены процессом усреднения цилиндрической субтомограммы, мы использовали дополнительный метод для определения локального расположения нуклеосом, окружающих другую рассматриваемую нуклеосому.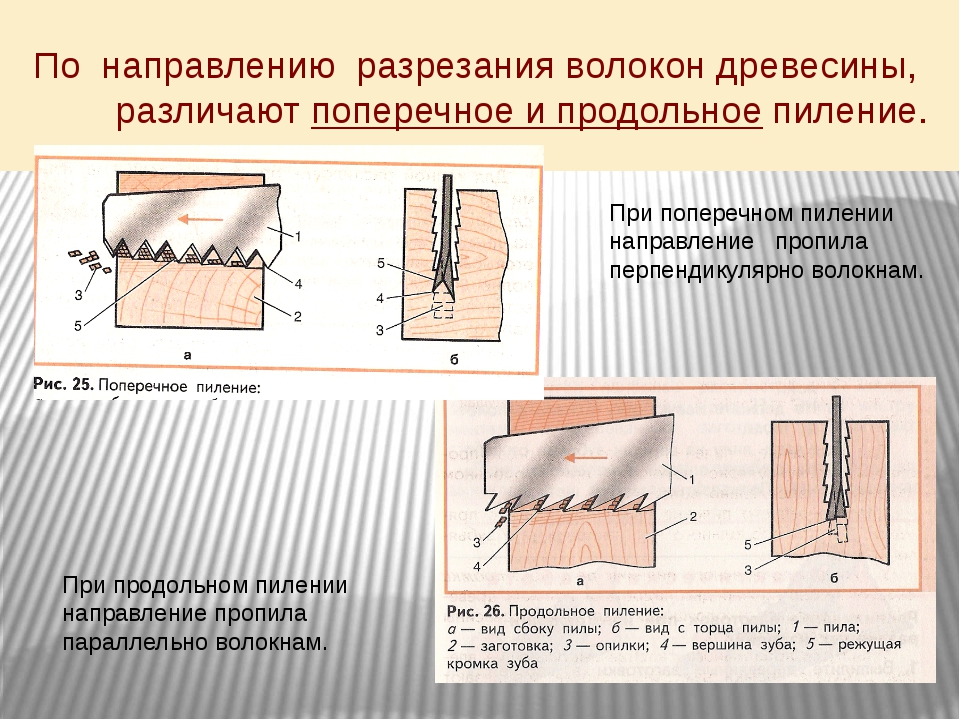 Таким образом, мы автоматически извлекали субтомограммы вокруг плотных позиций с приблизительным объемом одиночной нуклеосомы и выровняли их с шаблоном мононуклеосомы (25) ( SI Methods ).Большинство результирующих классов (рис. 5 A — C ) демонстрируют упорядоченную организацию нуклеосом, расположены лицом к лицу и демонстрируют узор в виде шахматной доски, который полностью совместим с видами сбоку среднего значения субтомограммы, если смотреть на направление, перпендикулярное оси волокна. Идеализированная симметризованная модель нашего волокна, рассчитанная на основе спиральных параметров, полученных из среднего значения субтомограммы, отчетливо демонстрирует эту геометрию (рис. 5 D и E ; подробности процедуры симметризации представлены в SI Text ).Внутри классов нуклеосомы имеют склонность ориентироваться лицом к лицу со смещением от их суперспиральных осей (Рис. 5 A и B ). Однако в некоторых случаях классы также обнаруживают нуклеосомы, расположенные рядом с их суперспиральными осями, выровненными, что отличается от того, что видно на средней субтомограмме (Рис.
Таким образом, мы автоматически извлекали субтомограммы вокруг плотных позиций с приблизительным объемом одиночной нуклеосомы и выровняли их с шаблоном мононуклеосомы (25) ( SI Methods ).Большинство результирующих классов (рис. 5 A — C ) демонстрируют упорядоченную организацию нуклеосом, расположены лицом к лицу и демонстрируют узор в виде шахматной доски, который полностью совместим с видами сбоку среднего значения субтомограммы, если смотреть на направление, перпендикулярное оси волокна. Идеализированная симметризованная модель нашего волокна, рассчитанная на основе спиральных параметров, полученных из среднего значения субтомограммы, отчетливо демонстрирует эту геометрию (рис. 5 D и E ; подробности процедуры симметризации представлены в SI Text ).Внутри классов нуклеосомы имеют склонность ориентироваться лицом к лицу со смещением от их суперспиральных осей (Рис. 5 A и B ). Однако в некоторых случаях классы также обнаруживают нуклеосомы, расположенные рядом с их суперспиральными осями, выровненными, что отличается от того, что видно на средней субтомограмме (Рис. 5 C ), и может быть отнесено к локальным искажениям спиральной структуры. Таким образом, мы обнаружили, что нуклеосомы могут выравниваться лицом к лицу с относительным сдвигом в сопоставлении примерно на 4 нм.Эти вариации в расположении нуклеосом лицом к лицу могут отражать структурную изменчивость в локальном масштабе, возникающую в результате изгиба, скручивания или интеркаляции волокон.
5 C ), и может быть отнесено к локальным искажениям спиральной структуры. Таким образом, мы обнаружили, что нуклеосомы могут выравниваться лицом к лицу с относительным сдвигом в сопоставлении примерно на 4 нм.Эти вариации в расположении нуклеосом лицом к лицу могут отражать структурную изменчивость в локальном масштабе, возникающую в результате изгиба, скручивания или интеркаляции волокон.
В большинстве кристаллических структур межцентровое расстояние между двумя одинаково ориентированными нуклеосомами составляет примерно 5,7 нм, что оставляет небольшое расстояние примерно 4 Å между гранями октомера (1). Однако нуклеосомы в нашем волокне связаны с межцентровым расстоянием в круговороте примерно 10.3 нм (рис. 3 E и рис. S6). Это значение зависит от расстояния от оси волокна, потому что соседние нуклеосомы расположены под углом друг к другу (Movie S3). Таким образом, из-за геометрии нашего волокна нуклеосомная ДНК в центре волокна почти соприкасается (расстояние примерно 2 нм), но нуклеосомная ДНК на периферии волокна очень доступна, оставляя межнуклеосомный зазор размером примерно 10 нм (рис. S6).
S6).
Обсуждение
Используя криоэлектронную томографию, мы показали, что наиболее преобладающей формой хроматина в ядрах куриных эритроцитов является 30-нм волокно, расположенное в виде двухзаходной спирали примерно с 6.7 нуклеосом на ход. Эта геометрия подобна структуре тетрануклеосомы (13) и модели двухстартового кросс-линкера, полученной на основе данных, основанных на исследованиях хроматина с использованием восстановленных волокон хроматина (14). По нашим данным, случаи электронной плотности, пересекающей ось волокна, и наличие центральной плотности в полости волокна обеспечивают прямую поддержку организации сшивающего агента. Таким образом, переход от нерегулярной зигзагообразной ленты, наблюдаемой при низком содержании соли, к компактному 30-нм волокну может происходить без серьезных топологических изменений, тем самым сохраняя двухзаходную геометрию.Однако важное различие между этими уровнями уплотнения состоит в том, что, хотя нерегулярное зигзагообразное волокно демонстрирует мало или совсем не показывает взаимодействия нуклеосом лицом к лицу (2), мотив стэкинга лицом к лицу представляет собой существенную особенность наблюдаемого волокна с длиной волны 30 нм. непосредственно в ядрах.
непосредственно в ядрах.
Структурное расположение нуклеосом более высокого порядка также выявляется с помощью классификации субтомограмм, что приводит к шахматному рисунку, который отражает виды сбоку 30-нм волокна (Рис. 5 A — C ).Лицом к лицу соседние нуклеосомы могут происходить при выровненных суперспиральных осях или со сдвигом положения суперспиральных осей по отношению друг к другу. Спиральный подъем на нуклеосому в среднем на нашей субтомограмме аналогичен подъему нуклеосом в идеализированной модели с перекрестными линкерами (13). Однако большее азимутальное вращение в нашем волокне приводит к более однородному распределению нуклеосом по сравнению с идеализированной моделью кросс-линкера (13), где уложенные друг на друга нуклеосомы образуют два плотных круговорота, которые широко разделены.В этом аспекте наша модель ближе к модели соленоида (12), в которой круговороты расположены более равномерно, но без пересечения нуклеосом. Мы предполагаем, что однородная, почти гексагональная упаковка нуклеосом (рис. 3 B ) является более энергетически выгодным решением для минимизации отталкивания от некомпенсированных отрицательных зарядов нуклеосомной ДНК.
3 B ) является более энергетически выгодным решением для минимизации отталкивания от некомпенсированных отрицательных зарядов нуклеосомной ДНК.
Лицом к лицу связывание нуклеосомных взаимодействий ранее было продемонстрировано в различных кристаллических структурах (1, 13, 25).Однако мы обнаружили, что нуклеосомы in situ взаимодействуют на расстоянии от центра до центра примерно 10,3 нм, оставляя большое расстояние не менее 3 нм между двумя ядрами октомера (рис. 3 E и рис. S6), что намного больше, чем указано в кристаллических структурах. Это расстояние связывания исключает взаимодействия ядро-ядро (13, 26), но допускает взаимодействия, опосредованные N-концевыми хвостами гистонов во взаимодействии «хвост-ядро». Большое межнуклеосомное расстояние предполагает высокую доступность грани и хвостов октамера для функциональных взаимодействий в компактном хроматине in situ, что согласуется с высокой динамической проницаемостью, наблюдаемой методами световой микроскопии (27). Довольно открытая структура указывает на то, что механизмы кроме механического барьера доступности ДНК, создаваемого плотной упаковкой нуклеосом, участвуют в подавлении транскрипции.
Довольно открытая структура указывает на то, что механизмы кроме механического барьера доступности ДНК, создаваемого плотной упаковкой нуклеосом, участвуют в подавлении транскрипции.
В отличие от искусственных массивов с определенной длиной нуклеосомного повтора и однородным составом гистонов, нативный хроматин имеет внутренние локальные вариации в длине повтора нуклеосом, а также модификации и варианты гистонов ядра и линкера. Мы демонстрируем, что, несмотря на эту изменчивость, нативный хроматин демонстрирует замечательные особенности структурной регулярности в компактных 30-нм волокнах.Наше исследование обеспечивает трехмерное изображение организации нативных волокон хроматина в компактном состоянии. Он также обеспечивает структурную интерпретацию предыдущих исследований контроля активности генов с использованием системы дифференцировки куриного эритроида (28). Тип модификаций гистонов, а также отсутствие HP1 в транскрипционно молчаливом хроматине куриных эритроцитов указывают на то, что он отличается от конститутивного гетерохроматина (29), и поэтому мы предполагаем, что его 30-нм волоконная организация может быть более репрезентативной для факультативного гетерохроматина.Эта гипотеза согласуется с недавними исследованиями Kizilyaprak et al. (6) на фоторецепторной клетке палочек мыши, где участки гетерохроматина, обогащенные факультативным гетерохроматином, демонстрируют организацию волокон с длиной волны 30 нм; тогда как кластерный конститутивный хроматин появляется беспорядочно, что согласуется с концепцией расплава хроматина (17). Другие доказательства представлены исследованиями политенных хромосом у Cheronomus tentans , где прекращение транскрипции приводило к расширенным «вздутым» волокнам хроматина, перестраивающимся в 30-нм волокна (7).
Экстраполяция характеристик 30-нм волокна куриного эритроцита на факультативный гетерохроматин in situ дает структурные ключи к реактивации репрессированных генов. Межнуклеосомные промежутки, как видно из нашего 30-нм волокна, могут обеспечивать начальную «пору» для первых факторов транскрипции, таких как FoxA, которые взаимодействуют с гистонами h4 и h5 (30), чтобы получить доступ к их сайтам связывания в компактном хроматине. Действительно, полностью неактивные геномы куриных эритроцитов также могут быть реактивированы после помещения в активную цитоплазматическую среду, такую как гетерокарионы (31).Поскольку межнуклеосомный зазор на периферии нашего волокна составляет приблизительно 10 нм, большие молекулы, такие как фактор хроматина RCC1, могут связываться непосредственно с поверхностью октамера (32) без необходимости разуплотнения волокна. Дальнейший криоэлектронный томографический анализ, применяемый к этим и другим системам, может выявить важную информацию о расположении волокон хроматина в различных функциональных состояниях.
Методы
Ядра куриных эритроцитов выделяли из суспензии куриных эритроцитов (Charles River Laboratories) по существу, как описано ранее (8, 16).Ядра замораживали под высоким давлением, а затем выполняли криосрезы с использованием алмазного ножа под углом 35 ° с номинальной подачей резания, установленной на 80–100 нм. Серии наклона собирали на микроскопе Tecnai G2 Polara (FEI), записывали на ПЗС-камеру 2k × 2k пикселей при уровне расфокусировки от -8 мкм до -6,5 мкм, с размером пикселя на уровне образца 0,6 нм. Полная доза электронов составляла приблизительно 90 e — / Å 2 на серию наклона. Серии наклона были совмещены с использованием программного обеспечения для выравнивания без реперных точек Alignator (33).Субтомограммы волокон были извлечены, итеративно выровнены и усреднены с использованием алгоритмов, учитывающих отсутствующий клин, до тех пор, пока окончательная структура не достигнет сходимости (рис. S7). Вся обработка выполнялась с использованием субтомограмм с размером вокселя 1,2 нм и выполнялась с помощью пользовательских скриптов, написанных в MATLAB (The Mathworks), которые доступны по адресу http://www.biophys.uni-frankfurt.de/frangakis. Для объемной визуализации и визуализации изоповерхностей используется программный пакет Amira (Mercury Computer Systems, http: // www.amiravis.com) (34) и пакет Chimera (35). Подробная информация о протоколах, подготовке образцов, методах электронной микроскопии и методах обработки изображений доступна в SI Text .
Благодарности
Мы хотели бы поблагодарить Хелен Сайбил, Кристофа Мюллера и Тревора Сьюэлла за внимательное чтение рукописи; кластер Франкфуртера для высокомолекулярных комплексов; и Европейский исследовательский совет за стартовый грант компании A.S.F.
Сноски
Авторы: A.С.Ф. и M.E. разработали исследования; M.P.S., M.E. и A.S.F. проведенное исследование; M.P.S., M.E. и A.S.F. проанализированные данные; и M.P.S., M.E., and A.S.F. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Размещение данных: Карта электронной плотности 30-нм волокна депонирована в базе данных макромолекулярных структур EBI под номерами доступа. EMD-1781 и EMD-1782.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1108268108/-/DCSupplemental.
Доступен бесплатно в режиме онлайн через опцию открытого доступа PNAS.
5.1: Крахмал и целлюлоза — Химия LibreTexts
Полисахариды — это самые распространенные в природе углеводы, которые выполняют различные функции, такие как хранение энергии или как компоненты стенок растительных клеток. Полисахариды — это очень большие полимеры, состоящие из десятков и тысяч моносахаридов, соединенных гликозидными связями.Три наиболее распространенных полисахарида — это крахмал, гликоген и целлюлоза. Эти три упоминаются как гомополимеры , потому что каждый дает только один тип моносахарида (глюкозы) после полного гидролиза. Гетерополимеры могут содержать сахарные кислоты, аминосахара или неуглеводные вещества в дополнение к моносахаридам. Гетерополимеры широко распространены в природе (камеди, пектины и другие вещества), но не будут обсуждаться далее в этом учебнике. Полисахариды представляют собой невосстанавливающие углеводы, не имеют сладкого вкуса и не подвергаются мутаротации.
Крахмал
Крахмал является наиболее важным источником углеводов в рационе человека и составляет более 50% потребляемых нами углеводов. Он встречается в растениях в виде гранул, и их особенно много в семенах (особенно в зернах злаков) и клубнях, где они служат формой хранения углеводов. Распад крахмала до глюкозы питает растение в периоды пониженной фотосинтетической активности. Мы часто думаем о картофеле как о «крахмалистой» пище, однако другие растения содержат гораздо больший процент крахмала (картофель 15%, пшеница 55%, кукуруза 65% и рис 75%).Технический крахмал представляет собой белый порошок.
Крахмал представляет собой смесь двух полимеров: амилозы и амилопектина. Природные крахмалы состоят примерно на 10–30% из амилазы и на 70–90% из амилопектина. Амилоза — это линейный полисахарид, полностью состоящий из единиц D-глюкозы, соединенных α-1,4-гликозидными связями, которые мы видели в мальтозе (часть (а) на рисунке 5.1.1). Экспериментальные данные показывают, что амилоза не является прямой цепью глюкозных единиц, а вместо этого свернута, как пружина, с шестью мономерами глюкозы на виток (часть (b) на рисунке 5.1.1). При таком свертывании амилоза в ядре имеет достаточно места для размещения молекулы йода. Характерный сине-фиолетовый цвет, который появляется при обработке крахмала йодом, обусловлен образованием амилозо-йодного комплекса. Этот цветовой тест достаточно чувствителен, чтобы обнаруживать даже мельчайшие количества крахмала в растворе.
Рисунок 5.1.1: Амилоза. (а) Амилоза представляет собой линейную цепь из единиц α-D-глюкозы, соединенных вместе α-1,4-гликозидными связями.(б) Из-за водородных связей амилоза приобретает спиральную структуру, содержащую шесть единиц глюкозы на оборот.
Амилопектин представляет собой полисахарид с разветвленной цепью, состоящий из единиц глюкозы, связанных главным образом α-1,4-гликозидными связями, но иногда с α-1,6-гликозидными связями, которые ответственны за разветвление. Молекула амилопектина может содержать многие тысячи единиц глюкозы с точками ветвления, встречающимися примерно через каждые 25–30 единиц (рис. 5.1.2). Спиральная структура амилопектина нарушается из-за разветвления цепи, поэтому вместо темно-сине-фиолетовой окраски амилозы с йодом амилопектин дает менее интенсивный красновато-коричневый цвет.
Рисунок 5.1.2 : Представление разветвления амилопектина и гликогена. И амилопектин, и гликоген содержат точки ветвления, которые связаны через α-1,6-связи. Эти точки ветвления чаще встречаются в гликогене.
Декстрины представляют собой полисахариды глюкозы промежуточного размера. Блеск и жесткость, придаемые одежде крахмалом, обусловлены присутствием декстринов, образующихся при глажке одежды. Из-за их характерной липкости при намокании декстрины используются в качестве клея на марках, конвертах и этикетках; как связующие вещества для удерживания пилюль и таблеток вместе; и как пасты.Декстрины перевариваются легче, чем крахмал, и поэтому широко используются при коммерческом приготовлении детского питания.
Полный гидролиз крахмала дает последовательные стадии глюкозы:
крахмал → декстрины → мальтоза → глюкоза
В организме человека несколько ферментов, известных под общим названием амилазы, последовательно расщепляют крахмал до пригодных для использования единиц глюкозы.
Гликоген
Гликоген — это углевод, являющийся энергетическим резервом животных.Практически все клетки млекопитающих содержат некоторые запасенные углеводы в форме гликогена, но особенно много его в печени (4–8% от веса ткани) и в клетках скелетных мышц (0,5–1,0%). Как и крахмал в растениях, гликоген находится в виде гранул в клетках печени и мышц. При голодании животные потребляют эти запасы гликогена в течение первого дня без еды для получения глюкозы, необходимой для поддержания метаболического баланса.
Примечание
Около 70% общего гликогена в организме хранится в мышечных клетках.Хотя процентное содержание гликогена (по весу) выше в печени, гораздо большая масса скелетных мышц хранит большее общее количество гликогена.
Гликоген структурно очень похож на амилопектин, хотя гликоген более разветвлен (8–12 единиц глюкозы между ветвями), а ветви короче. При обработке йодом гликоген дает красновато-коричневый цвет. Гликоген может быть расщеплен на его субъединицы D-глюкозы путем кислотного гидролиза или с помощью тех же ферментов, которые катализируют расщепление крахмала.У животных фермент фосфорилаза катализирует распад гликогена до фосфатных эфиров глюкозы.
Целлюлоза
Целлюлоза, волокнистый углевод, содержащийся во всех растениях, является структурным компонентом стенок растительных клеток. Поскольку земля покрыта растительностью, целлюлоза является самым распространенным из всех углеводов, на ее долю приходится более 50% всего углерода, содержащегося в царстве растений. Волокна хлопка и фильтровальная бумага почти полностью состоят из целлюлозы (около 95%), древесина составляет около 50% целлюлозы, а сухой вес листьев составляет около 10–20% целлюлозы.Наибольшее применение целлюлоза используется в производстве бумаги и бумажных изделий. Хотя использование нецеллюлозных синтетических волокон растет, вискоза (из целлюлозы) и хлопок по-прежнему составляют более 70% текстильного производства.
Подобно амилозе, целлюлоза представляет собой линейный полимер глюкозы. Однако он отличается тем, что единицы глюкозы соединены β-1,4-гликозидными связями, образуя более протяженную структуру, чем амилоза (часть (a) на рисунке 5.1.3). Эта крайняя линейность позволяет образовывать много водородных связей между группами ОН в соседних цепях, заставляя их плотно упаковываться в волокна (часть (b) на Рисунке 5.1.3). В результате целлюлоза слабо взаимодействует с водой или любым другим растворителем. Например, хлопок и дерево полностью нерастворимы в воде и обладают значительной механической прочностью. Поскольку целлюлоза не имеет спиральной структуры, она не связывается с йодом с образованием окрашенного продукта.
Рисунок 5.1.3 : Целлюлоза. (а) В структуре целлюлозы имеется обширная водородная связь. (b) На этой электронной микрофотографии клеточной стенки водоросли стенка состоит из последовательных слоев целлюлозных волокон, расположенных параллельно.
Целлюлоза дает D-глюкозу после полного кислотного гидролиза, но люди не могут метаболизировать целлюлозу как источник глюкозы. В наших пищеварительных соках отсутствуют ферменты, которые могут гидролизовать β-гликозидные связи, содержащиеся в целлюлозе, поэтому, хотя мы можем есть картофель, мы не можем есть траву. Однако некоторые микроорганизмы могут переваривать целлюлозу, потому что они вырабатывают фермент целлюлазу, который катализирует гидролиз целлюлозы. Присутствие этих микроорганизмов в пищеварительном тракте травоядных животных (таких как коровы, лошади и овцы) позволяет этим животным разлагать целлюлозу из растительного материала до глюкозы для получения энергии.Термиты также содержат микроорганизмы, выделяющие целлюлазу, и поэтому могут питаться древесной пищей. Этот пример еще раз демонстрирует крайнюю стереоспецифичность биохимических процессов.
Идентификация древесины и угля в Южном штате Мэриленд
Основы анатомии древесины и угляСогласно The Wood Handbook:
Волокнистость древесины сильно влияет на ее качество. использовал.В частности, древесина состоит в основном из пустотелых, удлиненных, веретенообразные ячейки, расположенные параллельно каждой другой вдоль ствола дерева. Когда пиломатериалы и другие изделия вырезаны из дерева, характеристики этих волокнистых ячейки и их расположение влияют на такие свойства, как прочность и усадка, а также структура текстуры древесины (USDA: 2007: 2-1).
Лиственные и хвойные породы
Деревья делятся на два широких класса, обычно называемые «твердые породы» и «мягкие породы».» Однако некоторые хвойные породы на самом деле тверже, чем некоторые из них. древесина твердых и некоторых твердых пород мягче, чем древесина мягких пород. Например, из таких хвойных пород, как длиннолистная сосна и дугласская ель, получается древесина, обычно тверже, чем липа и осина. Ботанически мягкие породы — голосеменные или хвойные, которые являются растениями с голые семена, то есть они не заключены в завязь цветок. Обычно хвойные породы представляют собой шишковидные растения с игольчатыми или чешуйки, как вечнозеленые листья.Некоторые хвойные деревья, например лиственницы и лысый кипарисовик, однако, теряют иголки осенью и зима. Примеры хвойных пород: сосны, ели, секвойи и можжевельники. Нажмите здесь для просмотра ключа Softwood . С ботанической точки зрения лиственные породы представляют собой покрытосеменные, у которых семена заключены в завязь цветка. Обычно они растения с широкими листьями, которые, за редким исключением в умеренный регион, теряют листья осенью или зимой.Большинство импортируемых тропических пород древесины — это твердые породы. Нажмите здесь, чтобы просмотреть Ключ из твердой древесины .
Нажмите для просмотра увеличенного изображения .
| Филогения 5 лесов: Сосна белая восточная ( Pinus strobus ), кипарисовик лысый ( Taxodium distichum ), Тополь тюльпан ( Liriodendron tulipifer ), Папайя ( Asimina) triloba ), вишня ( Prunus serotina ) и Хурма ( Diospyros virginiana ). |
Общие конструктивные особенности
Самолеты секции
Дерево — трехмерный материал, клетки тканей, ориентированные в трех разных плоскостях зрения (уголь) или плоскости сечения (дерево). Самолет, пересекающий ствол или ветвь, перпендикулярная волокну, называется поперечной или поперечное сечение (XS). Самолет, идущий в продольном направлении от центр ствола или ветви до коры называется лучевой раздел (R).Плоскость, перпендикулярная радиальной плоскости и является касательной к годовым кольцам, называется касательной раздел (T).
Нажмите
для просмотра увеличенного изображения .
| Схема показаны плоскости сечения, коры, годичных колец, заболони и сердцевина. |
Приросты (кольца)
Для всех кольцевых и полукольцевых твердых пород древесины и некоторые хвойные породы в округе Калверт, разницы достаточно между ранней древесиной одного года и предыдущей поздней древесиной до производят годовые кольца роста, видимые невооруженным глазом.Для по большей части возраст дерева можно рассчитать, посчитав годичные кольца либо из культи, либо из других поперечных сечений багажника. Внутренняя часть кольца роста сформировалась первой в период вегетации называется ранней древесиной, а внешняя часть формируется позже в вегетационный период, поздняя древесина. Earlywood характеризуется клетками с относительно большими полостями и тонкими стенками, а клетки поздней древесины имеют меньшие полости и более толстые стенки.Переход от ранней древесины к поздней может быть заметной или постепенной, в зависимости от от таксона и условий произрастания при его формировании.
Щелкните каждое изображение, чтобы увеличить изображение .
| | | |||
| Схематическое изображение ствола дерева, срезанного к центру, обнажающего 6 кожухов роста или годичных колец (внизу).В верхней части каждого ножна показана высота дерева в конце каждого вегетационного периода (справа). От: Фулже, А. 1969. Демонстрация свойств древесины в классе. USDA / FPL. (Общественное достояние) | Конечное зерно кипариса лысого ( Taxodium distichum ) древесный уголь показывает несколько годичных колец. | Конечное зерно древесного угля Pawpaw (Asimina triloba) с несколькими кольцами роста. |
Эрливуд и латвуд
В течение вегетационного периода дерево будет производить клетки в начале сезона (ранняя древесина) и клетки к конец сезона (поздняя древесина).Во время стресса (засуха, дефолиация, пожара) некоторые деревья дают так мало ранней древесины, что годичное кольцо исчезает в прошлогодней древесине.
| Нажмите
здесь для просмотра увеличенного изображения . | Нажмите
здесь для просмотра увеличенного изображения . | |
| Поперечное сечение
группы Yellow Pine ( Pinus sp .), показывая 2 частичный рост кольца | Поперечное сечение
компании Trumpet Vine ( Campsis) radicans ), показывающий рост граница кольца, ранняя древесина (вверху) и поздняя древесина (внизу) предыдущего кольцо роста. |
Заболонь и сердцевина
Заболонь расположена между корой и сердцевиной древесины. Заболонь содержит как живые, так и мертвые клетки и функции. в первую очередь в хранении продуктов и механическом транспорте воды или сока.Слой заболони может быть разной по толщине и толщине. количество содержащихся в нем годичных колец. Заболонь обычно колеблется от 1-1 / 2 до 2 дюймов в радиальной толщине. Как правило, чем больше У сильно растущих деревьев слои заболони шире. Много секунд Деревья товарного размера состоят в основном из заболони.
Как правило, сердцевина состоит из неактивных клетки, которые не проводят воду и не хранят пищу. Часто эти экстрактивные вещества затемняют сердцевину древесины и придают таким породам, как черный орех и вишня — их характерный цвет.Экстракты сердцевины древесины у некоторых видов, таких как саранча, красный кедр и красное дерево сделать древесину устойчивой к грибкам или насекомым. Заболонь Однако не все виды устойчивы к гниению. У некоторых видов таких как зола, гикори и некоторые дубы, поры (сосуды забиваться в большей или меньшей степени известными врастаниями как тилосы. Сердцевина имеет поры, плотно закупоренные тилозами, как и белый дуб, подходит для плотной бондаря, так как это предотвращает прохождение жидкости через поры.Тайлосы тоже делают пропитку с жидкими консервантами сложно.
Нажмите на изображение
для увеличения изображения .
| Вид зерна красной шелковицы ( Morus rubra ) с торца, виден коры. |
|
Анатомические особенности
Деревянные клетки
Ячейки древесины составляют структурные элементы древесины и состоят из разных размеров и форм.Сухие деревянные клетки могут быть пустым или частично заполненным отложениями, например камедью и смолой. Сосуды могут быть закупорены тилозами, соседними клетками паренхимы. тот шар в сосуд, закрывая его. Основная деревянная клетка типы, которые функционируют в транспортном и опорном исполнении (имеют удлиненную и заостренные на концах) — это волокна и трахеиды. Кроме того По сравнению с волокнами, древесина твердых пород имеет ячейки относительно большого диаметра, которые функционируют при транспортировке жидкостей, сосудов (или пор).В транспортную функцию также выполняют трахеиды хвойных пород. и лиственных пород. И лиственные, и хвойные породы имеют ячейки, которые сгруппированы в ткани, ориентированные горизонтально в радиальном направлении от сердцевины к коре. Эти клетки проводят жидкости радиально. и называются лучами. Лучше всего увидеть лучи на концах волокон. (XS) или четвертичные (R) поверхности. Древесина также имеет другие клетки, известные как продольная или осевая паренхима, которая функционирует в основном для хранение продуктов питания, экстрактивных веществ и / или кристаллов.Все эти ячейки типы и их расположение в тканях, трехмерно, составляет основу микроскопического описания и идентификации леса.
Вовлеченные клетки (древесина мягких и твердых пород
Хвойные породы
| Щелкните здесь, чтобы просмотреть увеличенное изображение. |
| Схема, показывающая типы клеток мягкой древесины и камбиальные инициалы, из которых они были получены.Камбиальные инициалы показаны примерно в тангенциальном аспекте. На трахеидах показаны два типа расположения ямок. Изменено из Джейн, Ф.В. 1970. Структура дерева. |
Осевые / вертикальные трахеиды
Осевые трахеиды являются основным типом клеток мягкой древесины, составляющая более 90% клеток в древесине. Их радиальный диаметр меняется в зависимости от темпов роста, порядок, сложившийся во время вегетационный период (трахеиды ранней древесины больше, чем трахеиды поздней древесины) и условия окружающей среды.В некоторых хвойных породах, таких как Лысый Кипарис. ( Taxodium distichum ) и Секвойя ( Sequoia sempervirens ), первые сформированные трахеиды всегда достаточно широки для развития межтрахеидных питтинг, который имеет более одного ряда ямок и является диагностическим ценность.
| Радиальный срез кипариса лысого (Taxodium distichum) с двояковыпуклой (шириной 2) окаймленными ямками (стрелки) в осевых трахеидах. |
Спирали
У некоторых пород древесины есть спиральные утолщения на стенах. их трахеид, и они могут быть диагностически значимыми. Однако археологическая или деградировавшая древесина часто имеет спиралевидный чеки в клеточных стенках, выглядящие как спиральные утолщения.
Лучи трахеиды
Лучи трахеиды похожи на маленькие осевые трахеиды, ориентированные радиально.Как и в осевых трахеидах, они имеют ямки с круглой окантовкой. (CBP), только меньше. Лучи трахеиды могут иметь гладкие стенки как White Pine Group или стены имеют заостренные выступы (зубчатые) внутри них, как в группах желтых и красных сосен. Главное отличие между елью и лиственницей — форма поперечного сечения CBP в лучевых трахеидах.
Нажимайте каждый для просмотра увеличенного изображения .
| Радиальный разрез группы белых сосен ( Pinus sp. ), показывающая 1 ряд лучевых трахеид и 5 рядов клеток лучевой паренхимы внизу. | Радиальный разрез сосны желтой
Группа Pinus sp.) С 3 рядами лучевых трахеид. |
Осевая / вертикальная Паренхима
Осевая паренхима у хвойных пород обычно диффузно присутствует в поздней части годичного кольца.Обычно они содержат цветное содержимое и их легко увидеть. при малых увеличениях. В тангенциальном виде стены, разделяющие соседние клетки будут либо гладкими, либо узловатыми.
Луч Паренхима
Паренхима ската у хвойных пород имеет либо гладкую торцевые стенки или торцевые стенки с отчетливыми утолщениями на них (узловые торцевые стенки).
Нажимайте каждый для просмотра увеличенного изображения .
| Радиальное сечение
из Hemlock ( Tsuga spp. ), показывающий клеток паренхимы луча с узловатыми концевыми стенками. | Радиальный сечение кипариса лысого ( Taxodium distichum ) показаны клетки паренхимы луча с гладким концом стены. |
Эпителиальные клетки смоляных каналов
Во многих хвойных и некоторых лиственных породах клетки расположены в осевом или радиальном направлении в каналах или каналах, которые производят смолы, дубильные вещества и другие вторичные соединения, используемые для защиты против патогенов, таких как насекомые и грибки.В древесине хвойных пород смола каналы хорошо видны как на X, так и на T. Может иметь толстый клеточные стенки (эпителий) как у ели ( Picea sp. ) и лиственницы ( Larix sp. ) или тонкие стенки, как у сосны ( Pinus spp. ).
Нажимайте каждый для просмотра увеличенного изображения .
| Поперечное сечение
сосны ( Pinus sp.) с одинарным полимерным воздуховодом с тонкостенными эпителиальными клетками. | Поперечное сечение
Ель ( Picea sp. ) с 3 смоляными трубами с толстыми стенками. эпителиальные клетки. |
| Тангенциальная участок сосны ( Pinus sp.), показывающий сингл смоляной проток с тонкостенными эпителиальными клетками. Счастливчик участок руки, сохранивший эпителий. | Тангенциальный
срез сосны ( Pinus sp. ), на котором виден единственный полимерный проток с тонкостенными эпителиальными клетками . Нормальный участки руки с вырванным эпителием. |
| Радиальный
секция болиголова ( Tsuga spp.), показывающее клеток паренхимы луча с узловатыми торцевыми стенками. | Радиальный сечение кипариса лысого ( Taxodium distichum ) показаны клетки паренхимы луча с гладким концом стены. |
Тангенциальное сечение
Ель ( Picea sp.) | тангенциальный
срез ели ( Picea sp. ) , показывающий единственный смоляной проток с толстостенными эпителиальными клетками. (Поляризованный свет). |
Межполевые ямы (не ячейки)
Когда лучи пересекаются с осевыми трахеидами, клетки прилегающие друг к другу имеют межклеточные связи называется кросс-полевые ямы.Они там, где осевая система и луч системы крест и являются важными диагностическими признаками.
Нажмите на изображении, чтобы увеличить изображение .
| Схематические изображения язвы на радиальных стенках клеток паренхимы луча хвойных пород. [x — ямки в разрезе по оси x; y — ямы в разрезе разрез по оси y; поперечный обзор — ямы, как видно в виде поверхности в поперечном поле] Модифицировано из Jane, F.W. 1970. |
Структура древесины
Лиственных пород
Типы ячеек
Древесина лиственных пород отличается от древесины мягких пород тем, что (обычно) сосуды и волокна в дополнение к паренхиме и трахеидам. Некоторые примитивные породы древесины не имеют сосудов. Более примитивный трахеида является хорошим проводником жидкости и хорошей опорой элемент.Волокна превратились в плохие проводники, но отличные поддерживающие элементы, в то время как сосуды развиваются для отличной проводимости но плохая поддержка. Паренхима может содержать цветные соединения или кристаллы или быть сегментированными и выровненными по горизонтали в ткани. Элементы сосуда соединяются вертикально (по волокну) в сосуды, имеющие перфорационные пластины различного типа. Все четыре типы клеток превратились в разные размеры, формы и орнаменты в отдельные таксоны и составляют основу всех точных микроскопических идентификации.
Нажмите на изображение, чтобы увеличить изображение .
| Схема с указанием типов ячеек твердой древесины и камбиальных инициалов, которые произвел их. Несколько видов перфорации показаны в сосудах.Изменено от Джейн, F.W. 1970. Структура древесины. | Эволюция волокон и члены сосудов из трахеид (разделение труда / функции). |
Тайлос
В некоторых лиственных породах при формировании сердцевины баллон вазицентрической паренхимы внутрь соседних сосудов элементы. Тилосы эффективно блокируют неиспользуемые сосуды, поскольку пути распространения патогенов, таких как насекомые и грибки.Присутствие тилозов в белом дубе позволяет использовать его как «плотную бондону» не пропускающие жидкости (вино, виски, пиво, вода и т. д.), в отличие от красного дуба, который используется как «слабина бондаря» для галантереи. Тайлозы могут казаться маленькими или большими, с толстыми или тонкие стенки и при правильном использовании является хорошим диагностическим признаком.
Нажмите для просмотра увеличенного изображения .
| ||
| Радиальный
секция американского черного ореха ( Juglans nigra ) с тилозами. | Радиальный
срез черной саранчи ( Robinia pseudoacacia ) показывая тилосы. |
Перфорационные пластины
Сосуды отличаются от трахеид наличием перфорации на их концах, чтобы обеспечить эффективную транспортировку жидкости. Эти отверстия могут быть полностью открытыми (простая перфорация). или может иметь поперечные перекладины (ступеньки) (скалярная перфорация). Полосы могут быть немногочисленными и толстыми или многочисленными и тонкими и могут быть диагностическими. ценность.
Нажмите на изображении для увеличения изображения .
| Радиальный срез воскового мирта ( Myrica cerifera ), показывая 2 сосуда, 1 с простым перфорация и еще одна с чешуйчатой перфорацией. |
Межсосудные (IV) карьеры
Соединения между соседними элементами сосуда называются межсосудистыми (IV) ямами.Их легко увидеть в тангенциальном вид, кроме случаев, когда распределение сосудов одиночное. Они различаются размером с пределов большинства оптических микроскопов (2µ) до линейной точечной коррозии, которая почти охватывает сосуд (до 50 мкм).
| РАЗМЕР ЯМКИ | ПРИМЕР |
| Минуты (2 — 4µ) | Береза |
| Маленький (5-7µ) | Горная лавра |
| Средний (8 — 10µ) | Клен |
| Большой (> 10µ), включает линейный | Орех |
Щелкните каждое изображение для просмотра увеличенное изображение .
| Тангенциальная
разрез березы ( Betula sp. ) , показывающий мельчайшие ямки между сосудами (2 — 4µ). | Тангенциальный
разрез клена ( Acer sp. ), показывающий ям между сосудами среднего размера (5 — 10µ). |
| Тангенциальное сечение ореха ( Juglans
sp. ) с изображением больших ям между сосудами (стрелки) 10-16µ. |
IV. Расположение
Расположение IV ямок может быть напротив каждой. другие, чередующиеся по горизонтали или состоящие из длинных отверстий, охватывающий член сосуда (линейный или скалярный).
| Тип | Пример |
| напротив | Тополь тюльпан |
| Альтернативный | Клен |
| Линейная / Скалярная | Сладкая жевательная резинка |
Нажмите
каждое изображение для увеличения изображения .
| Тангенциальная секция тюльпанового тополя
( Liriodendron tulipifera ) с противоположными ямками между сосудами (стрелки). | Тангенциальное сечение клена ( Acer sp. ) стрелками показаны альтернативные ямы между сосудами). |
| Тангенциальное сечение Red Gum ( Liquidambar styraciflua ) с линейным межсосудистым пространством ямки наконечников стрел). |
Ray-Vessel (RV) Точечная коррозия
Соединения между элементами емкости и прилегающими клетки лучевой паренхимы называются ямками лучевых сосудов (ПС). Это может быть такой же, как IV питтинг, или намного больше (саликоид) до линейного.
Нажмите каждое изображение для увеличения изображения .
| Радиальный разрез Вяза ( Ulmus
sp.) с ямками скатных сосудов. |
Спирали
Некоторые породы дерева имеют спиральные утолщения на стенах. членов их судов. Некоторые таксоны имеют ограниченные спирали. к наконечникам элементов сосуда. Некоторые спирали могут быть тонкими и равномерно расположенными, не проходит через межсосудистую (IV) ямку, как у Maple ( Acer sp. ). Остальные спирали более толстые и грубо уложенные и действительно проходят через внутривенную ямку, как Cherry ( Prunus sp.), Липа ( Tilia sp. ) и Холли ( Ilex sp. ). Тем не мение, археологическая или деградированная древесина часто проверяется по спирали стенки клеток выглядят как спиральные утолщения.
Нажмите на каждом изображении для просмотра увеличенного изображения.
| Тонкие спирали (клен) | грубый Спирали (вишня и липа) | |
| Тангенциальное сечение Maple (Acer sp.) показаны прекрасные спирали. | Тангенциальные срезы черешни ( Prunus serotina ) с крупными спиралями. |
Трахеиды
Хотя хвойные породы состоят в основном из трахеид, древесина твердых пород может иметь как сосудистые трахеиды (например, древесина мягких пород), так и вазицентрические трахеиды, окружающие сосуды в таких лесах, как Дуб.
Нажмите на изображении для увеличения изображения .
| Тангенциальная секция дуба ( Quercus sp. ), показывающий вазицентрические трахеиды (наконечник стрелки) с ямы с круглой окантовкой. |
Волокна
Основным типом опорной ячейки в древесине твердых пород является волокно либриформ. Внутри этих ячеек могут быть перегородки. (перегородка) или нет (без перегородки).Также есть группа ячеек (волокна трахеиды), которые по структуре занимают промежуточное положение между волокнами и трахеиды. Эти клетки также можно разделить на септатные. или без перегородки.
Нажмите каждое изображение для просмотра увеличенное изображение.
| Поперечный разрез группы дуба
( Quercus sp.), с умеренно толстостенными волокнами. | Поперечный разрез черной саранчи ( Robinia pseudoacacia ), демонстрирующий очень толстые волокна с стенками. |
Аксиальная паренхима
Пряди паренхимы, ориентированные вертикально, или с зерном внутри древесины называются осевой паренхимой. Они сгруппированы в две большие категории; апотрахеальная паренхима и паратрахеальная паренхима.Апотрахеальный означает «без сосуды »и представляет собой осевую паренхиму, не соприкасающуюся сосуды в поперечном сечении. Типы апотрахеальной паренхимы включают: диффузные, сетчатые или ломаные и полосчатые. Паратрахеальный означает «вокруг сосудов» и является осевой паренхимой. который полностью или частично окружает пересекающиеся суда раздел. Типы паратрахеальной паренхимы включают вазицентрическую («Окружающее судно»), плоское (крылатое) и сливное (сросшиеся крылья).
Щелкните по каждому
для просмотра увеличенного изображения .
Диффузный сетчатый
Полосовой
Схема различных типов апотрахеальной
паренхима лиственных пород, если смотреть в разрезе.
Сосуды представлены кружками, лучи — вертикальными линиями и
паренхима красными участками.
Щелкните по каждому
для просмотра увеличенного изображения .
Васицентрик Алиформ
Конфлюэнт
Схема различных типов паратрахеального отдела
паренхима лиственных пород, если смотреть в разрезе.
Сосуды представлены кружками, лучи — вертикальными линиями и
паренхима красными участками.
Лучи
Лучи состоят из клеток паренхимы лиственных пород а также паренхимы и трахеид лучей хвойных пород. В лиственных породах Лучи классифицируются по ширине ячеек и неоднородности. Лучи могут иметь ширину в одну ячейку в тангенциальном (однорядном) виде или в несколько ячеек широкие (многорядные). Они могут быть только одного типа ячейки (квадратная или уплощенный), называемый гомоклеточным, или он может иметь оба клетки в теле луча и от одного до многих рядов квадратов или прямостоячие клетки по периферии (гетероклеточные).
Щелкните по каждому
для просмотра увеличенного изображения .
Однорядные лучи Многорядные лучи
Гомоклеточные лучи Гетероклеточные лучи
Различные типы лучей в твердых породах древесины в тангенциальном, радиальном и поперечные сечения.
Щелкните по каждому для просмотра увеличенного изображения .
| Радиальный разрез Maple ( Acer sp. )
показаны гомоклеточных лучей, состоящих исключительно из проходящих клетки. | Радиальный разрез
Тюльпан Тополь Liriodenron tulipifera ) показан гетероклеточный луч , состоящий из пролегающих клеток (верхняя половина рисунка) и стойки (наконечники стрел). |
| Радиальный
срез ивы ( Salix spp. |