Вольвокс | PHYWE
Nach oben
Информация
- Контактное лицо
- Условия сотрудничества
- Декларация о конфиденциальности
- Вводные данные
Обслуживание
- Краткий обзор услуг
- Скачать
- Каталоги
- Вебинары и Видео
- Связаться со службой поддержки клиентов
Компания
- О нас
- Качественная политика
- Безопасность в классе
Please note
* Prices subject to VAT.
We only supply companies, institutions and educational facilities. No sales to private individuals.
Please note: To comply with EU regulation 1272/2008 CLP, PHYWE does not sell any chemicals to the general public. We only accept orders from resellers, professional users and research, study and educational institutions.
Пожалуйста, введите имя, под которым должна быть сохранена Ваша корзина.
Сохраненные корзины вы можете найти в разделе My Account.
Название корзины
АЛЬГОЛОГИЯ:О научных названиях отдела зеленые водоросли и классов собственно зеленые водоросли и требуксиевые
On the scientific names of the division Chlorophyta, and the classes Chlorophyceae, and Trebouxiophyceae
Тарасов К.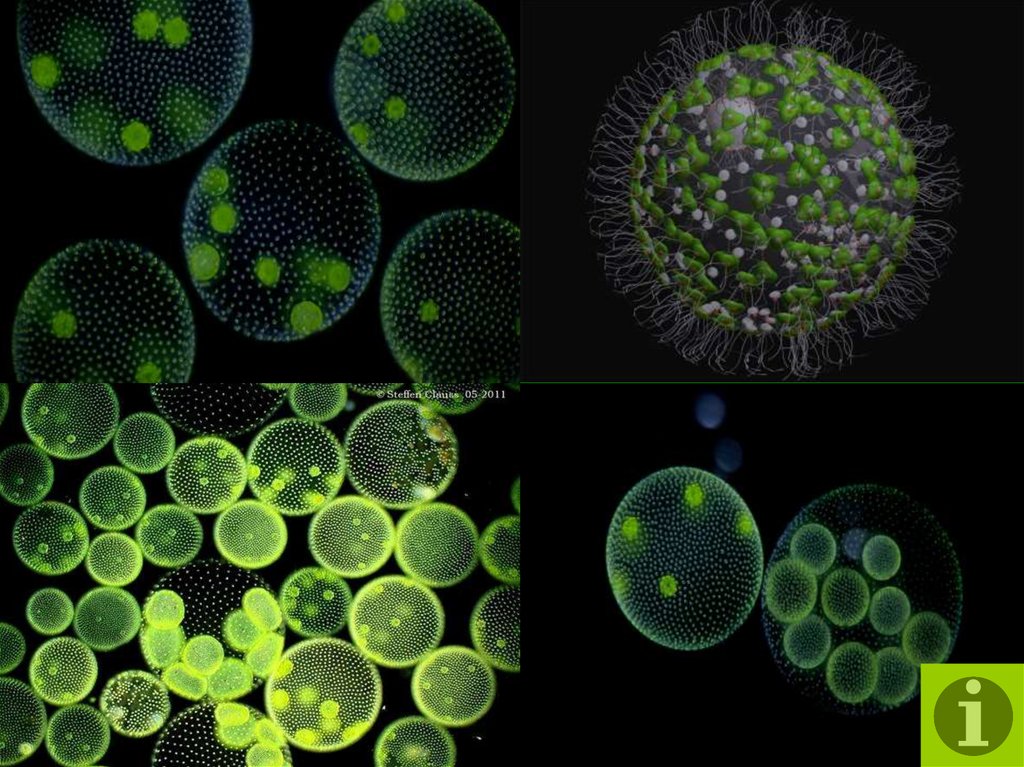 Л.
Л.
Konstantin L. Tarasov
Московский государственный университет имени М. В. Ломоносова,
биологический факультет, кафедра микологии и альгологии
УДК 582.21:57.06
Считается, что научное название Chlorophyta, обычно используемое для зелёных водорослей, явно неудачно, поскольку оно означает «зелёные растения», но все высшие растения – также зелёные растения. Аналогично название Chlorophyceae для одного из классов зелёных водорослей тоже неудачно, поскольку оно означает «зелёные водоросли», но они представляют только один класс зелёных водорослей. Для этих таксонов предлагаются названия соответственно Chlorellophyta и Volvocophyceae. Поскольку род Chlorella, от которого производится предлагаемое название отдела, относится к классу Trebouxiophyceae, название последнего может быть заменено на Chlorellophyceae.
Ключевые слова: зелёные водоросли; научные названия.
Зеленые водоросли – обширнейшая группа низших эукариот, занимающая во многих отношениях центральное положение в системе растений.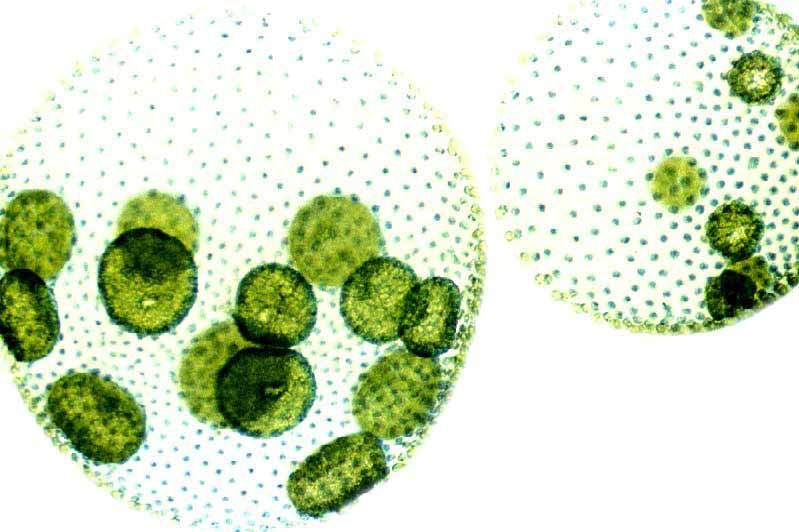 Многие ее представители играют огромную роль в различных экосистемах, особенно пресноводных. Некоторые широко используются в экспериментальных исследованиях.
Многие ее представители играют огромную роль в различных экосистемах, особенно пресноводных. Некоторые широко используются в экспериментальных исследованиях.
В разных современных системах они понимаются по-разному, в разном объеме, но в любой системе имеется отдел под названием Chlorophyta. Название это явно неудачно, так как означает «зеленые растения». Получается, что все высшие растения (а во многих современных системах и харофиты) – НЕ зеленые растения. Такое название отдела имеет смысл только в том случае, если мы понимаем его в самом широком смысле, то есть включающим и зеленые водоросли в традиционном понимании, и все высшие растения.
В то же время, многими ботаниками-систематиками, особенно академиком А.Л. Тахтаджяном, давно отстаивался тезис о желательности (или даже обязательности) образования всех научных названий таксонов рангом выше рода от родовых названий (например, отдел Magnoliophyta от рода Magnolia и т.д.).
В нашем же случае представляется особенно целесообразным образование названия отдела Chlorophyta от родового названия.
В качестве такого родового названия можно предложить Chlorella Beijerinck 1890.
Каковы могут быть аргументы в пользу такого выбора?
1. Род Chlorella относится в любой системе к отделу Chlorophyta, который и в сокращенном виде хотя бы в силу своего названия занимает центральное положение среди водорослей.
2. От этого родового названия уже образованы названия семейства и порядка — соответственно Chlorellaceae и Chlorellales.
3. Предлагаемое название похоже на предыдущее, то есть в определенном смысле не является совсем новым.
4. Суффикс «-ell-» имеет уменьшительное значение: предлагаемое название отдела как бы попутно дает понять, что зеленые водоросли – «братья меньшие» настоящих зеленых, то есть высших растений.
Соответственно я хотел бы сделать другое предложение – присвоить название Chlorellophyceae классу Trebouxiophyceae, поскольку именно в нем находятся сейчас и род Chlorella, и семейство Chlorellaсеае, и порядок Chlorellales.
Для класса же Chlorophyceae я предлагаю название Volvocophyceae – по роду
опубликовано — июнь 2012 г.
On the scientific names of the division Chlorophyta, and the classes Chlorophyceae, and Trebouxiophyceae
Konstantin L. Tarasov
The scientific name Chlorophyta usually used for green algae is supposed to be obviously inappropriate since in means “green plants”, but all higher plants are also green plants. Similarly, the name Chlorophyceae for one of chlorophytan class is inappropriate too, since in means “green algae”, but they represent only one class of green algae.
Key words: Chlorophyta; scientific names.
Об авторe
Тарасов Константин Львович — Tarasov Konstantin L.
кандидат биологических наук, доцент
ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), Биологический ф-т, каф. микологиии и альгологии
Корреспондентский адрес: Россия, 119991, Москва, Ленинские горы, д. 1, стр. 12, Биологический факультет МГУ
ССЫЛКА НА СТАТЬЮ:
Тарасов К.Л. О научных названиях отдела зеленые водоросли и классов собственно зеленые водоросли и требуксиевые // Вопросы современной альгологии. 2012. № 1 (1). URL: http://algology.ru/95
2012. № 1 (1). URL: http://algology.ru/95
Карта сайта
Вольвокс — первое многоклеточное существо?. Исчезнувший мир
Вольвокс — первое многоклеточное существо?
Зоологи и ботаники давно спорят о жгутиконосцах — растения они или животные?
Предмет их спора столь невелик, что простым глазом его не увидишь. Жгутиконосцы микроскопические существа — живые «шарики», «колбаски», «лодочки», с тонкими, жгутиковидными хвостиками, которыми они ударяют по воде и плывут.
В каждой луже миллиарды жгутиконосцев. Под микроскопом видно, что они зеленые: полным-полно у них под прозрачной «кожицей» хлорофилловых зернышек. Значит, это растения?
Решить непросто.
Тысячи хвостатых шариков, словно молекулы в тепловом движении, беспорядочно скачут в капле воды. Вот один наткнулся на бактерию. Втянул ее в миниатюрный «ротик» и… проглотил.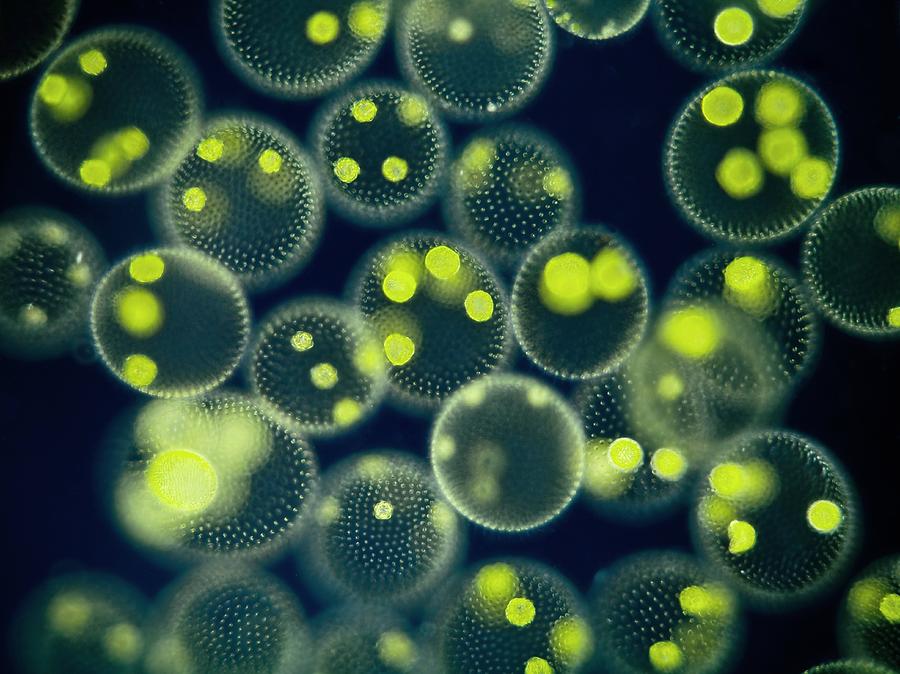 У растений нет ртов. И глаз тоже нет. А у жгутиконосцев есть «карие» глазки. Обычно это просто бурое или красноватое пятнышко, воспринимающее свет. Но иногда оно углублено в виде чаши, в которой лежит линзовидный комочек крахмала. Он прозрачный — это хрусталик первородного глаза.
У растений нет ртов. И глаз тоже нет. А у жгутиконосцев есть «карие» глазки. Обычно это просто бурое или красноватое пятнышко, воспринимающее свет. Но иногда оно углублено в виде чаши, в которой лежит линзовидный комочек крахмала. Он прозрачный — это хрусталик первородного глаза.
Так это животное?
Все зависит от погоды. В солнечный день, когда много света, жгутиконосцы — скорее растения. Занимаются фотосинтезом, из углекислого газа и воды изготовляют сахар. Им и питаются. В пасмурную погоду, когда света мало, некоторые из них переходят на другую диету: ловят бактерий и мелкие водоросли.
Поэтому и нелегко биологам решить: с кем же они наконец имеют дело? Зоологи считают, что жгутиконосцы (по крайней мере некоторые из них) — простейшие животные. А ботаники числят их в разряде низших водорослей.
Вольвокс, говорит Д. Апдайк, «интересует нас потому, что он изобрел смерть. Амебы никогда не умирают… Но вольвокс, этот подвижный, перекатывающийся шар водорослей… нечто среднее между растением и животным под микроскопом он кружится, как танцор на рождественском балу, — впервые осуществив идею сотрудничества, ввел жизнь в царство неизбежной — в отличие от случайной — смерти».
До него, до вольвокса, смерть на Земле была необязательна и, так сказать, незаконна. Все одноклеточное живое никогда не умирало естественной смертью, только насильственной. Размножаясь, одноклеточная жизнь делилась пополам. А разделившись, жила вновь в удвоенном числе. Но когда одноклеточные жгутиконосцы объединились и образовали вольвокс, все они приобрели в этом объединении разную квалификацию. Одни сохранили привилегии половых клеток — эти, размножаясь, жили вечно в своих потомках. Другие сделались клетками соматическими, то есть бесполым телом колонии, и всякий раз умирали теперь после того, как их половые сестры и братья размножались.
Так смерть стала обязательным и законным по кодексу природы финалом жизни. До этого была лишь случайность.
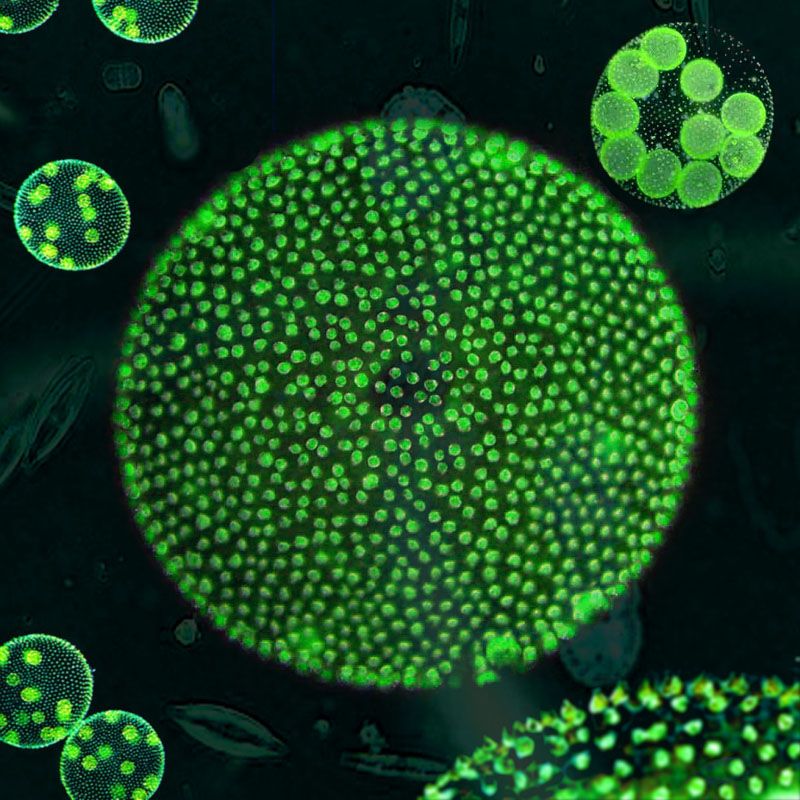
Вольвокс — подвижный живой шарик (в диаметре до 3 миллиметров). Внутри он студенистый, а снаружи весь усеян жгутиконосцами (одноклеточными водорослями с двумя хвостиками-жгутиками, колебания которых приводят вольвокс в движение). Это настоящая колония зеленых жгутиконосцев — на поверхности вольвокса их от 200 до 50 тысяч. Есть у соединившихся в единое целое жгутиконосцев примитивные глазки — стигмы. На одном полюсе шара они лучше развиты, на другом хуже. Более «глазастым» полюсом вольвокс и плывет вперед.
Это настоящая колония зеленых жгутиконосцев — на поверхности вольвокса их от 200 до 50 тысяч. Есть у соединившихся в единое целое жгутиконосцев примитивные глазки — стигмы. На одном полюсе шара они лучше развиты, на другом хуже. Более «глазастым» полюсом вольвокс и плывет вперед.
Почти все составляющие сферическую колонию клетки размножаться не способны (ни вегетативно, ни половым путем). Только около десятка самых крупных из них в нужное время плодятся, создавая дочерние колонии внутри живого шара.,
«Колонию вольвокса, может быть, правильнее рассматривать как многоклеточный организм, поскольку… не все клетки колонии равноценны. Возможно, что колониальность имела большое значение в эволюции органического мира и явилась переходным этапом к многоклеточным животным» (Ю. Полянский).
Так большинство ученых и полагает: от колониальных простейших организмов, подобных вольвоксу, и произошли многоклеточные растения и животные.
А теперь подведем итог. Как шла в пространстве и во времени биохимическая и биологическая эволюция. Какие пройдены ею этапы в древнейшие эры Земли.
Какие пройдены ею этапы в древнейшие эры Земли.
Принимаем, что Земля сформировалась 4,5 миллиарда лет назад и была окружена первичной атмосферой: аммиак, метан, пары воды, азот, окись углерода, углекислый газ, водород, азот.
Эры (Начало и конец в миллиардах лет назад) Биохимические и биологические объекты Катархей 4,5—3 Аминокислоты, пептиды, пурины, пиримидины, нуклеиды, порфирины. Затем микросферы, предклетки. Архей 3–1,9 Первые живые клетки (доядерные, анаэробные). Анаэробные хемотрофные бактерии. Нитчатые и округлые водоросли (сине-зеленые?). Протерозой: Нижний 1,9–1,6 Строматолиты сине-зеленых водорослей и онколиты бактерий. Фотосинтезирующие бактерии. Верхний (или рифей) 1,6–0,57 Организмы с клеточными ядрами. Многоклеточные животные, близкие к медузам, червям, морским перьям, членистоногим.Цифры приведены приблизительные, поскольку у разных авторов неодинаковые датировки эпох (как и их названия). Часто и возраст Земли не всеми указывается одинаковый. Многое еще в этих вопросах остается спорным.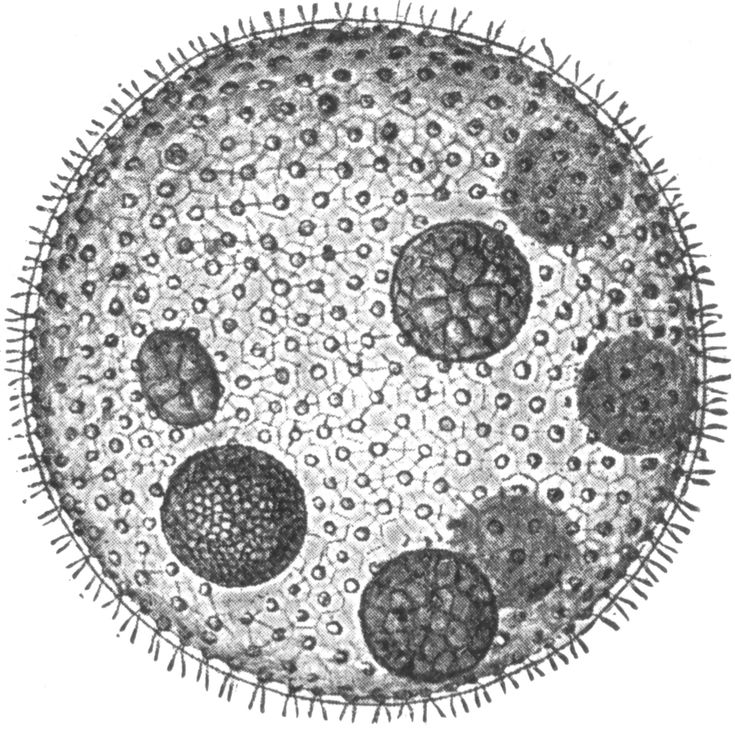
Каждое живое существо в самом себе несёт следы своих древних предков
Каждое живое существо в самом себе несёт следы своих древних предков Возьмём для примера человека. Человек благодаря труду чрезвычайно высоко поднялся над миром животных. Но в строении тела человека, в его сердце, лёгких, желудке, кишках, мускулах, скелете много сходного
ПЕРВОЕ ЗНАКОМСТВО
ПЕРВОЕ ЗНАКОМСТВО Вы найдете у них нечто вроде республики, а также память и предусмотрительность… при встречах они узнают друг друга. Какое тут бывает оживление! Как суетливо они останавливаются и расспрашивают друг друга! Мы видим, как стирается камень под их стопами,
Первое знакомство
Первое знакомство
На разных континентах
Астрономы давно ведут учет звезд. Конечно, нельзя думать, что все небесные светила уже сосчитаны. Однако в самом подробном из астрономических каталогов значится более полутораста тысяч звезд, до девятой величины включительно.
Конечно, нельзя думать, что все небесные светила уже сосчитаны. Однако в самом подробном из астрономических каталогов значится более полутораста тысяч звезд, до девятой величины включительно.
Глава 10 Третий полевой сезон в Хадаре: „первое семейство”
Глава 10 Третий полевой сезон в Хадаре: „первое семейство” Когда Дон показал мне первый коленный сустав, я отправил его назад, велев вернуться с целым скелетом. И ему пришлось найти Люси. Тогда я вновь велел ему вернуться и привезти останки нескольких особей. И на
Первое наземное
Первое наземное Вместе с этими растениями произрастали странные бактериальные колонии, достигавшие 70 см в поперечнике, и мхи. Во влажных зарослях сложилось силурийское наземное сообщество. Среди безлистных стебельков уже бегали многоножки, тригонотарбы (греч.
ПЕРВОЕ ВПЕЧАТЛЕНИЕ ОТ ЗЕМЛИ
ПЕРВОЕ ВПЕЧАТЛЕНИЕ ОТ ЗЕМЛИ Когда путешественник проходит по плодоносным равнинам, где тихие воды в своем постоянном течении питают богатую растительность, где покой земли с многочисленным населением, с цветущими селениями, богатыми городами, величественными зданиями,
5 «Первое, вольное вдохновенье»
5
«Первое, вольное вдохновенье»
Кого мы выбираем?
Где-то в ожиданье одиноком
Томится одинокая душа,
Но ждет она, надеется, — и к сроку
С другой душою встретится, спеша.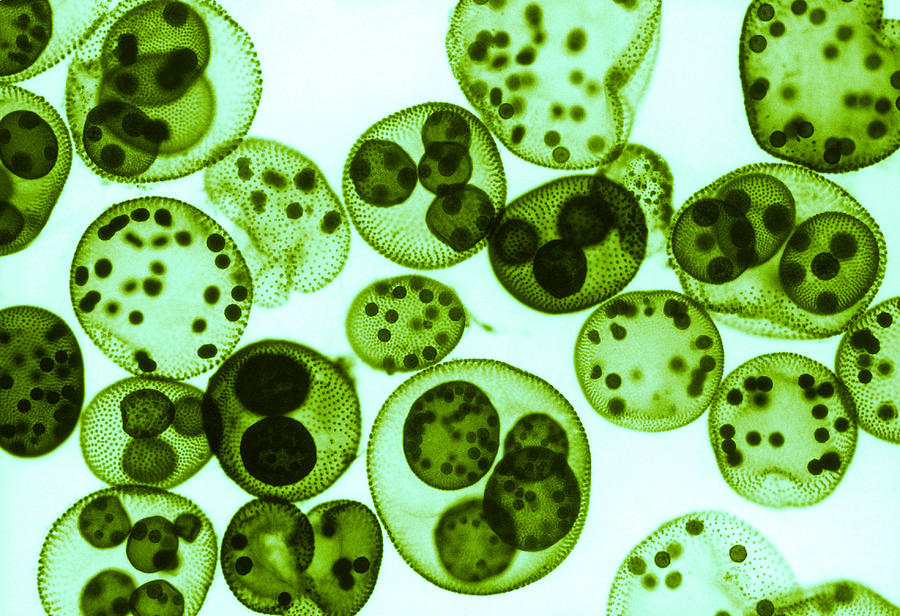 И воедино, словно два потока,
Сольются, тихой нежностью дыша,
И вечность распахнет свои врата,
И тьма уж не
И воедино, словно два потока,
Сольются, тихой нежностью дыша,
И вечность распахнет свои врата,
И тьма уж не
Каким было первое млекопитающее?
Каким было первое млекопитающее? В 1999 году в китайской провинции Ляонинь найден прекрасно сохранившийся скелет древнейшего млекопитающего, которому примерно 120 миллионов лет. Это настоящая химера, промежуточное звено между рептилиями и млекопитающими: передняя часть
ПЕРВОЕ ЗНАКОМСТВО
ПЕРВОЕ ЗНАКОМСТВО На разных континентах Астрономы давно ведут учет звезд. Конечно, нельзя думать, что все небесные светила уже сосчитаны. Однако в самом подробном из астрономических каталогов значится более полутораста тысяч звезд, до девятой величины включительно.
Социальное существо
Социальное существо
Когда щенку исполняется 21 день, он буквально внезапно становится общественным существом, полностью осознающим индивидуальность не только своих братьев и сестер, но и матери, и своего хозяина.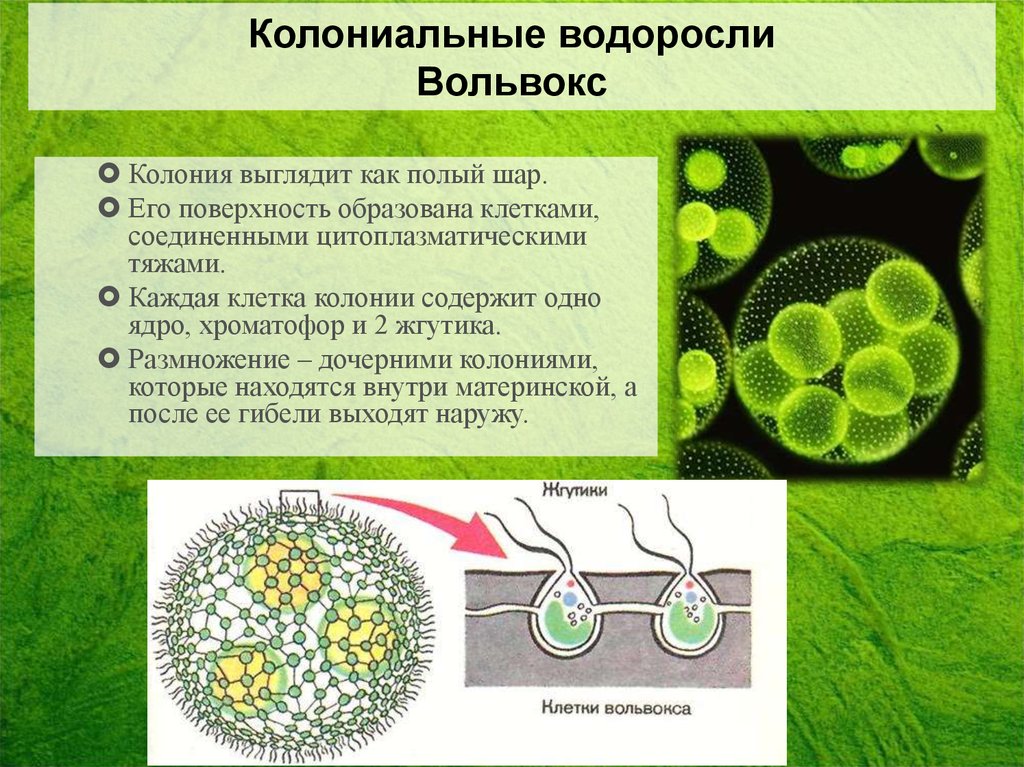 Он смотрит на других людей, на собак и других животных и
Он смотрит на других людей, на собак и других животных и
Социальное существо
Социальное существо Когда щенку исполняется 21 день, он буквально внезапно становится общественным существом, полностью осознающим индивидуальность не только своих братьев и сестер, но и матери, и своего хозяина. Он смотрит на других людей, на собак и других животных и
Первое уточнение
Первое уточнение Кое-кто из скорых на руку службистов, прослышав краем уха, что муравьи могут быть полезны в деле защиты леса от вредителей и что некоторые виды лесных муравьёв надо поэтому всячески охранять, а можно даже и расселять, тут же принялся с воодушевлением
Кагуан – существо непонятное
Кагуан – существо непонятное
Одни знатоки уверяют, что кагуан, или шерстокрыл (ростом он с кошку), насекомоядный зверь, нечто вроде летающей землеройки.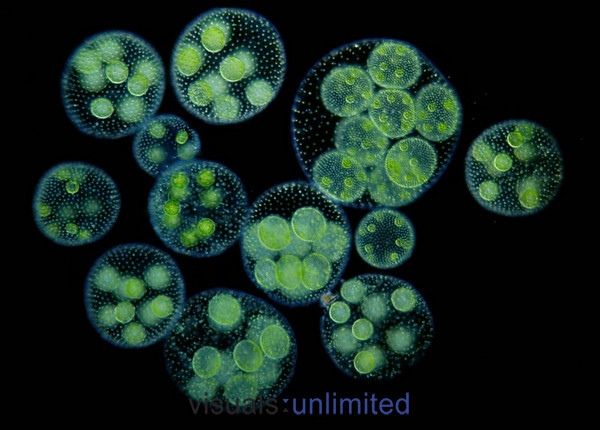 Другие не согласны: он лемур (летающий, конечно). Наконец, третьи доказывают: кагуан ни то и ни другое, а особое, в
Другие не согласны: он лемур (летающий, конечно). Наконец, третьи доказывают: кагуан ни то и ни другое, а особое, в
Фрукторианство — первое знакомство…
Фрукторианство — первое знакомство… Фрукторианство — это уникальный образ питания, подразумевающий употребление исключительно растительной пищи. Рацион фрукторианца состоит из всевозможных фруктов и овощей, а основное отличие этого типа питания от других диет
Глава 7 TIGR, мое первое детище
Глава 7 TIGR, мое первое детище Секрет моих успехов кроется исключительно в моем упорстве. Луи Пастер Луи Пастер – один из гигантов науки. Он добился огромных успехов в микробиологии и иммунологии. Открыл принцип стерилизации, который был назван в его честь
Volvox: красивая микроскопическая форма жизни
Volvox: красивая микроскопическая форма жизни
| VOLVOX Морис Smith & Ken Jones 1997 Кен Джонс и Журнал Micscape | Одна из самых красивых живых форм, плавающих в наших прудах — Volvox: вращающаяся колония
ярко-зеленые клетки.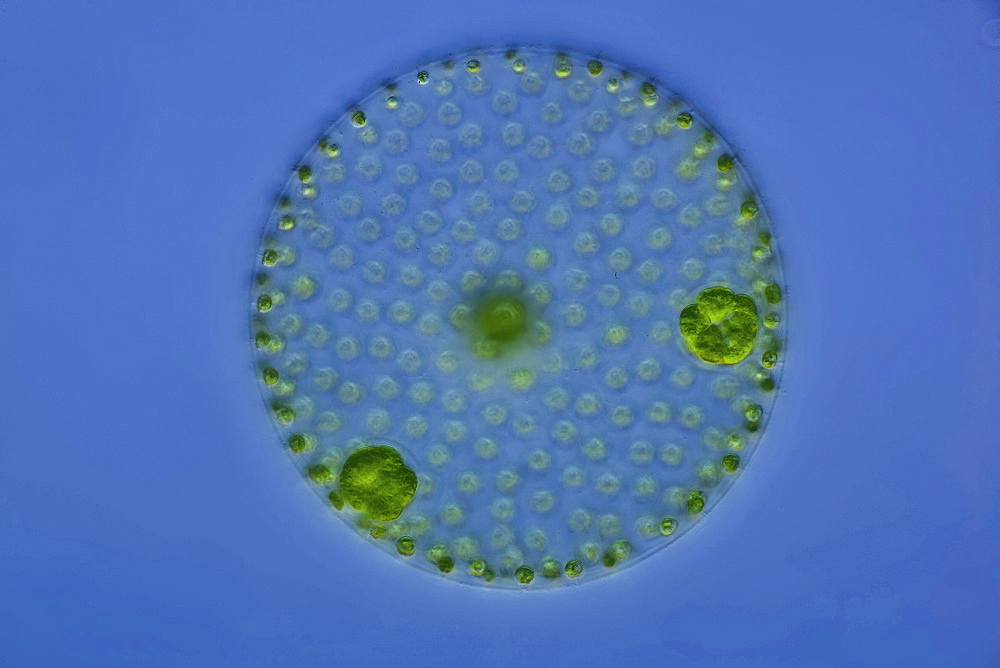 Эта статья из журнала Micscape Magazine знакомит вас с их потрясающим присутствием. Все изображения
и видеофрагменты ведущего микроскописта-любителя Кена Джонса, чьи видеоработы известны любителям
по всей Великобритании. Эта статья из журнала Micscape Magazine знакомит вас с их потрясающим присутствием. Все изображения
и видеофрагменты ведущего микроскописта-любителя Кена Джонса, чьи видеоработы известны любителям
по всей Великобритании. |
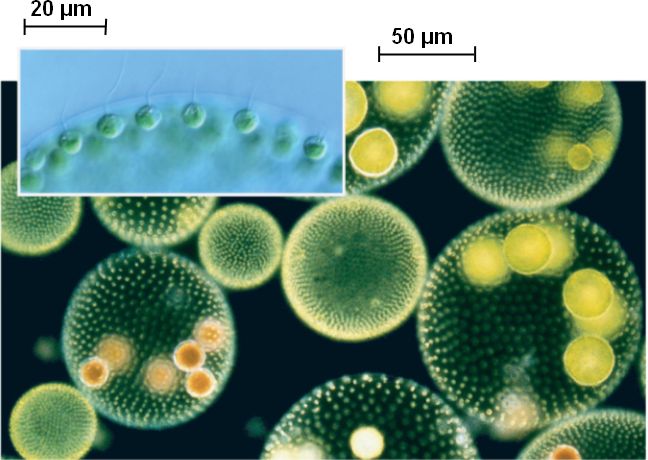
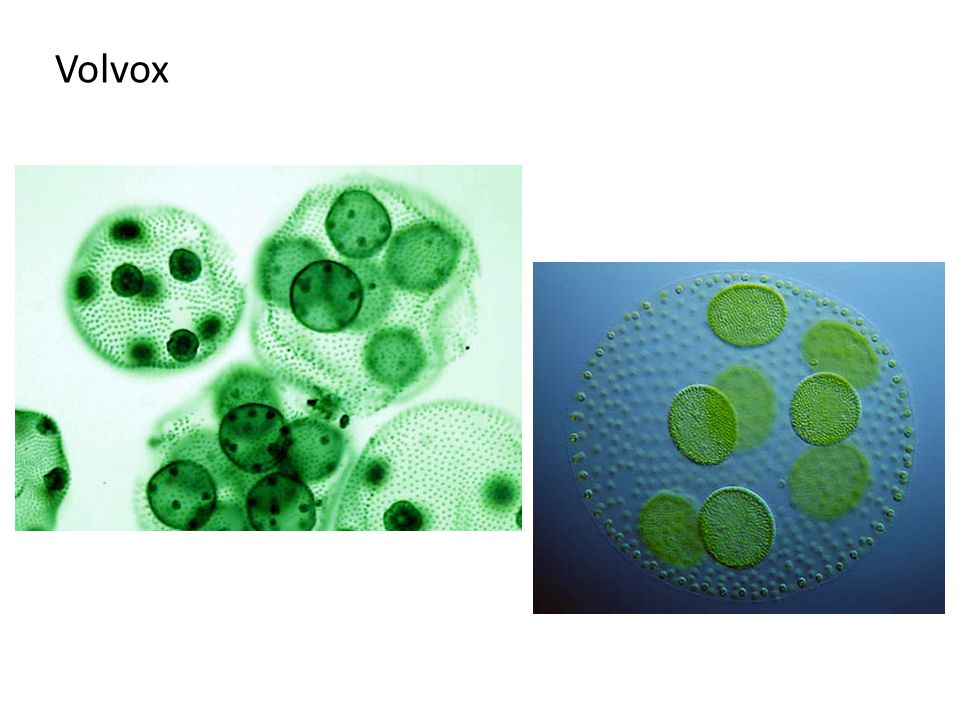
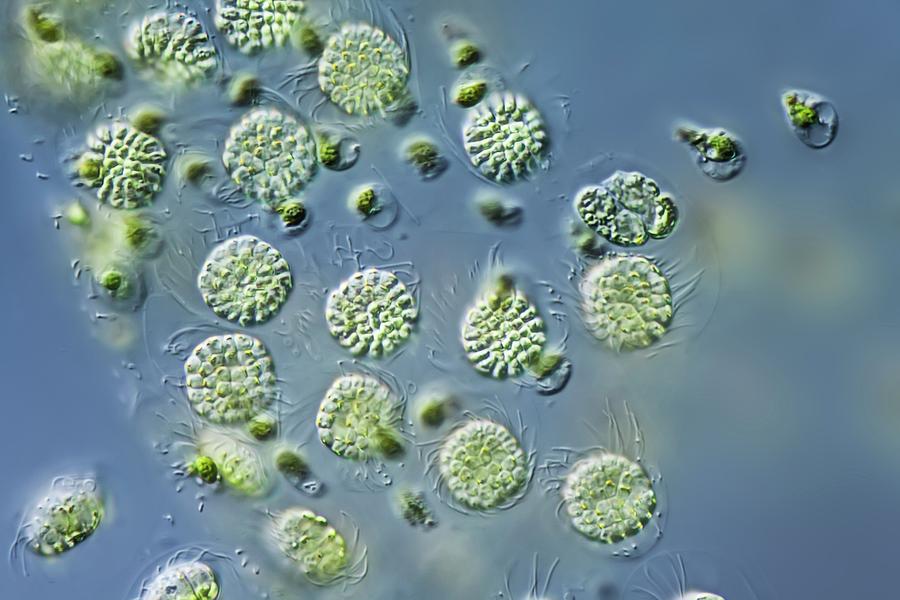
| Водоросли — это простые растительные организмы, встречающиеся во всех влажных средах. Их размеры варьируются от микроскопических форм до простых макроскопических форм «рдестых водорослей» и крупных морских водорослей. Всем интересно учиться но микроскописты в целом согласны с тем, что одним из самых красивых является Volvox. Это сферические колонии зеленых клеток, цепляющихся за полупрозрачный полый шарик слизи. А одна колония может состоять из более чем 500 клеток, каждая из которых имеет крошечную пару хлыстообразных хвостов (жгутиков) — и все клетки синхронно взмахивают жгутиками, толкая колонию по воде. |
Очень большие колонии могут превышать 1 мм в диаметре и легко видны невооруженным глазом.
 Многие будут
содержат дочерние клетки, а иногда даже внучатых клеток на различных стадиях развития внутри полой внутренней части земного шара.
Многие будут
содержат дочерние клетки, а иногда даже внучатых клеток на различных стадиях развития внутри полой внутренней части земного шара. | Кен Джонс, ведущий микроскопист-любитель, специализируется на видеосъемке прудовой жизни. Один из в его видеороликах запечатлены загадочные и грациозные движения Volvox, и он любезно разрешил нам показать некоторые фрагменты из его фильма на этой странице. | Одинокая колония вольвоксов кружится в темных водах пруда. Дочерние клетки только видно внутри. |
 Посмотрите, как часть сферы уже отсутствует. Его поглотило крошечное существо.
Посмотрите, как часть сферы уже отсутствует. Его поглотило крошечное существо. | Здесь Volvox показан в значительном увеличении. Загляните внутрь большего слева. Вы видите ярко-зеленые дочерние клетки внутри? Обратите внимание на вольвокс рядом с ним и чуть ниже не содержит такое потомство. |
Micscape Примечание редактора. Эта страница только что проверила ваш браузер и обнаружила что не все изображения отображаются. Вам необходимо обновить браузер до версии 3.0 или 4.0 для корректного отображения этой страницы!
Нравится миры внутри миров — дочерние клетки
можно рассматривать как развивающиеся колониальные сферы внутри
полой внутренней части более крупной.
Во всей жизни, независимо от масштаба, есть постоянная борьба и конкуренция
для выживания.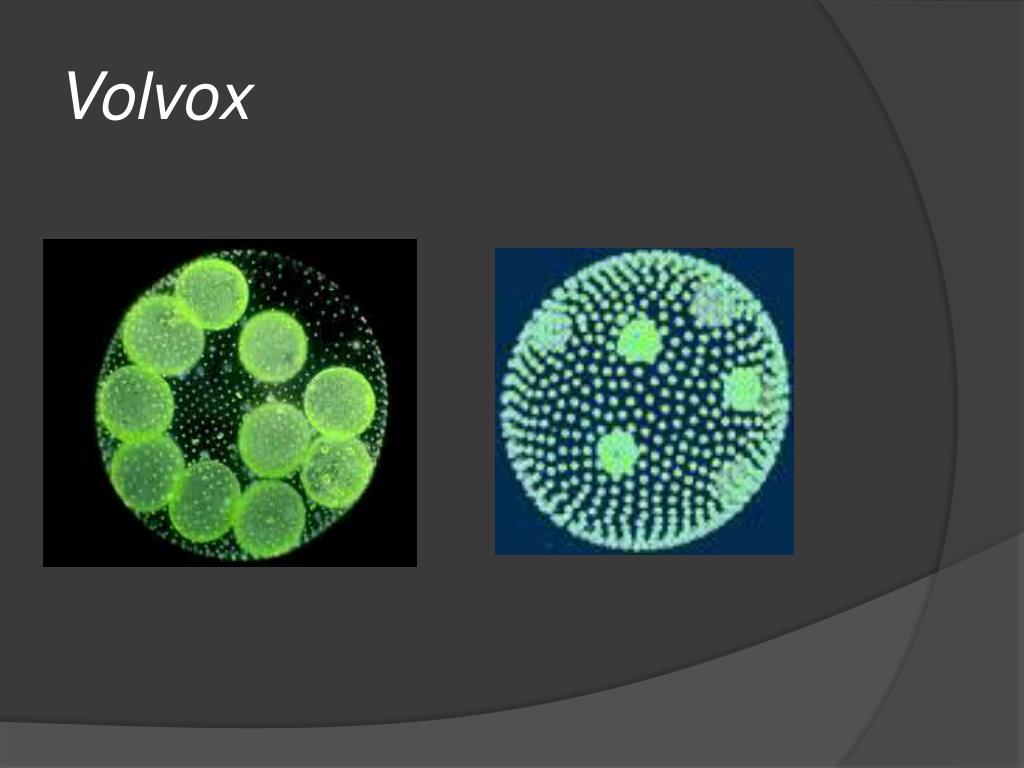 Пруд – это космос в миниатюре. Разнообразные формы жизни, как животные, так и растения, конкурируют за
энергоносители, продукты питания, пространство и другие ресурсы. Пруд – это космос в миниатюре. Разнообразные формы жизни, как животные, так и растения, конкурируют за
энергоносители, продукты питания, пространство и другие ресурсы.Потрясающе красивый Volvox может быть атакован и съеден крошечными микроскопическими животными. как коловратки — существа, которые хотя и почти невидимы невооруженным глазом, но способны выискивать свою добычу. |
Здесь снова можно увидеть, как коловратка атакует вольвокса. это размыто по скорости его движения в воде при падении — передний конец с разинутой пастью обращен вниз ближе к нижней части экрана.
Единственный верный способ насладиться всем этим действием, красотой и чудесами — выйти на улицу.
в пруд, принеси немного воды из пруда и сам посмотри под микроскопом. Это еще куча
весело, чем пытаться увидеть их здесь, на наших страницах через Интернет. Когда вы начнете, вы сможете делать то, что
Кен Джонс сделал и делает свои собственные потрясающие видео для себя.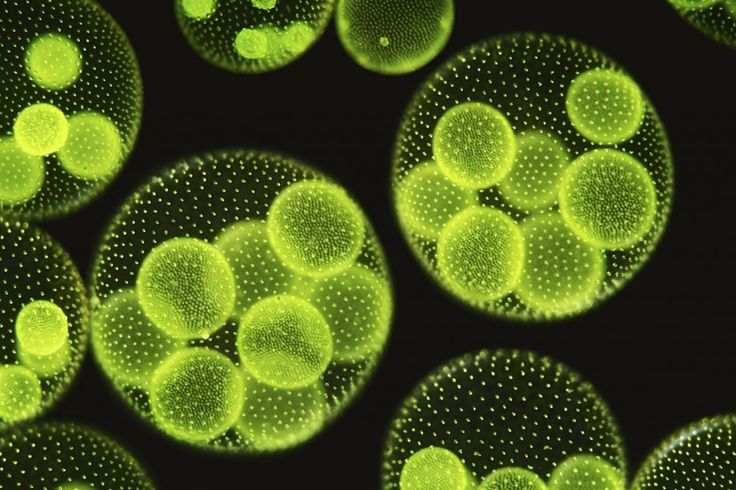
Если вы новичок или микроскопист-любитель, вам может быть трудно визуализировать
как клетки Volvox и других сферических колониальных водорослей взаимодействуют друг с другом. Здесь справа колония водорослей
очень похожи на вышеперечисленные. Его зовут Уроглена Волксокс. Хотя вы не можете видеть жгутики в каждой клетке,
вы можете видеть сами отдельные клетки, когда колония вращается.
Эти колонии обычно имеют диаметр до 500 (0,5 мм) с каждой клеткой.
примерно 10 (0,10 мм) в длину. В отличие от зеленого Volovox, клетки Uroglena обычно имеют золотисто-коричневый цвет.
Видео здесь показывает клетки более зеленого цвета, чем они обычно выглядят под микроскопом; Это
в связи с улучшениями в видеоряде, чтобы облегчить просмотр предмета на нашей странице.
Пожалуйста
примечание: это бесплатный ресурс , предоставленный
Микроскопия-Великобритания. Работаем 7 лет
без оплаты, чтобы создать один из самых богатых контентом сайтов в Интернете. Наши расходы растут.
Наши расходы растут.
Если вы считаете, что этот ресурс стоит держать в свободном доступе для всех, возможно, вы могли бы
хотите рассмотреть вопрос о пожертвовании лишь небольшой суммы, чтобы помочь?
Пожалуйста, нажмите здесь, если вы хотите рассмотреть
небольшое пожертвование.
Это действительно помогло бы!
Вернуться на главную страницу Microscopy-UK Композиция текста и страницы и звуки Мориса Смита 1997 |
Примечание редактора: Некоторые версии Netscape 3. 0 не запускаются
avi файлы фильмов правильно. Если основные изображения на этой странице не двигаются постоянно, как это должно быть, попробуйте нажать
на них дважды быстро с помощью мыши! Эта страница предназначена для просмотра с установленным как минимум Netscape 4.0 или
Браузеры Explorer 4.0. 0 не запускаются
avi файлы фильмов правильно. Если основные изображения на этой странице не двигаются постоянно, как это должно быть, попробуйте нажать
на них дважды быстро с помощью мыши! Эта страница предназначена для просмотра с установленным как минимум Netscape 4.0 или
Браузеры Explorer 4.0. |
Onview.net Ltd, Microscopy-UK, и все участники, начиная с 1995 г. Все права сдержанный. Основной сайт находится по адресу www.microscopy-uk.org.uk, а полное зеркало — по адресу www.microscopy-uk.net.
Биология 104 Эмбриология 24 января 2005 г. Альберт ХаррисКак (и как?) одноклеточные организмы стали многоклеточнымиVolvox и Dictyostelium/ становятся многоклеточными совершенно по-разному . Volvox — род многоклеточных растений. Каждый отдельный вольвокс представляет собой полую сферу из клеток. Обратите внимание на сходство между Volvox и родом одноклеточных водорослей под названием Chlamydomonas . Другими словами, вид X Volvox имеет последовательности генов, которые больше похожи на вид Q Chlamydomonas, а иногда и на вид B Eudorina. Принимая во внимание, что вид Y Volvox имеет последовательности генов, которые больше похожи на вид R Chlamydomonas и вид C Eudorina. Это является убедительным доказательством того, что морфология, форма, внешний вид и образ жизни вольвоксов эволюционировали снова и снова от Chlamydomonas (конечно, в далеком прошлом), и что виды, классифицированные как Eudorina и Pandorina, являются живыми примерами промежуточных стадий в этом эволюционном достижении многоклеточности (такое использование слова «достижение» может отражать многоклеточный шовинизм). Различные виды Volvox развивались отдельно, три или более раз, всегда из одноклеточных водорослей, которые можно было бы классифицировать как Chlamydomonas. Никто не видел, как это происходило, как и не снимали покадровую видеозапись эрозии Гранд-Каньона, но доказательства из генетических базовых последовательностей очевидны. Высшие растения произошли от зеленых водорослей (а не от бурых водорослей, таких как ламинария, или от красных водорослей).
Chlamydomonas и Volvox классифицируются как зеленые водоросли (но они не являются предками высших растений) Совсем другой способ стать многоклеточным встречается у группы «животных» (?), называемых « Клеточные слизевики ». Являются ли они грибоподобными амебоидными животными? Или это животноподобные амебоидные грибы?
Наиболее изученным видом в этой группе является Dictyostelium discoideum, , который был обнаружен выпускником Университета Северной Каролины по имени Кеннет Рапер в 1930-х годах. Сотни различных исследовательских лабораторий по всему миру сосредоточились на изучении этого вида. |
 (« Конвергентная Эволюция «)
(« Конвергентная Эволюция «)
 Однако…..
Однако….. Эти слизни удлиняются и ползают в течение нескольких часов или меньше, пока не найдут подходящее место, а затем часть клеток дифференцируется в клетки стебля, а большинство дифференцируется в споровые клетки.
В результате этого процесса «плодоношения» образуется высокий стебель с примерно сферической массой спор на его верхнем конце. «Грекс» на латыни означает стадо (стадное).
Эти слизни удлиняются и ползают в течение нескольких часов или меньше, пока не найдут подходящее место, а затем часть клеток дифференцируется в клетки стебля, а большинство дифференцируется в споровые клетки.
В результате этого процесса «плодоношения» образуется высокий стебель с примерно сферической массой спор на его верхнем конце. «Грекс» на латыни означает стадо (стадное). 8.19A: Архепластиды — Биология LibreTexts
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 9849
- Безграничный
- Безграничный
Цели обучения
- Описать взаимосвязь между красными водорослями, зелеными водорослями и наземными растениями
Красные водоросли и зеленые водоросли входят в надгруппу Archaeplastida. Хорошо задокументировано, что наземные растения произошли от общего предка этих простейших; их ближайшие родственники находятся в этой группе. Молекулярные данные подтверждают, что все Archaeplastida являются потомками эндосимбиотических отношений между гетеротрофным протистом и цианобактерией. Красные и зеленые водоросли включают одноклеточные, многоклеточные и колониальные формы
Хорошо задокументировано, что наземные растения произошли от общего предка этих простейших; их ближайшие родственники находятся в этой группе. Молекулярные данные подтверждают, что все Archaeplastida являются потомками эндосимбиотических отношений между гетеротрофным протистом и цианобактерией. Красные и зеленые водоросли включают одноклеточные, многоклеточные и колониальные формы
Красные водоросли
Красные водоросли, или родофиты, в основном многоклеточные, лишены жгутиков и варьируются по размеру от микроскопических одноклеточных протистов до крупных многоклеточных форм, сгруппированных в неофициальную категорию морских водорослей. Жизненный цикл красных водорослей представляет собой чередование поколений. Некоторые виды красных водорослей содержат фикоэритрин, дополнительные фотосинтетические пигменты красного цвета, которые превосходят зеленый оттенок хлорофилла, благодаря чему эти виды имеют различные оттенки красного. Другие протисты, классифицируемые как красные водоросли, лишены фикоэритрина и являются паразитами.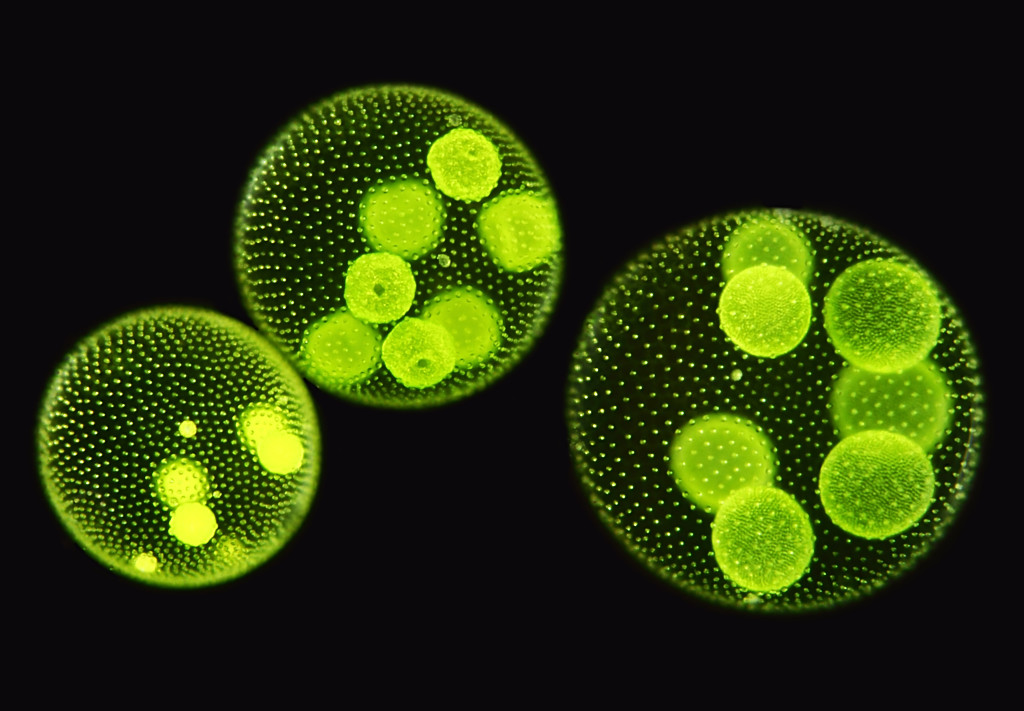 Красные водоросли распространены в тропических водах, где они были обнаружены на глубине 260 метров. Другие красные водоросли существуют в наземной или пресноводной среде.
Красные водоросли распространены в тропических водах, где они были обнаружены на глубине 260 метров. Другие красные водоросли существуют в наземной или пресноводной среде.
Зеленые водоросли: Chlorophytes и Charophytes
Наиболее многочисленной группой водорослей являются зеленые водоросли. Зеленые водоросли имеют сходные черты с наземными растениями, особенно в отношении строения хлоропластов. Хорошо подтверждено, что эта группа простейших имеет относительно недавних общих предков с наземными растениями. Зеленые водоросли подразделяются на хлорофиты и харофиты. Харофиты являются ближайшими живыми родственниками наземных растений, напоминая их по морфологии и стратегиям размножения. Харофиты распространены во влажных местах обитания, где их присутствие часто сигнализирует о здоровой экосистеме.
Хлорофиты отличаются большим разнообразием форм и функций. Хлорофиты населяют преимущественно пресноводные и влажные почвы; они являются обычным компонентом планктона. Chlamydomonas представляет собой простой одноклеточный хлорофит с грушевидной морфологией и двумя противоположными передними жгутиками, которые направляют этого простейшего к свету, воспринимаемому его глазным пятном. Более сложные виды хлорофитов демонстрируют гаплоидные гаметы и споры, которые напоминают Chlamydomonas .
Более сложные виды хлорофитов демонстрируют гаплоидные гаметы и споры, которые напоминают Chlamydomonas .
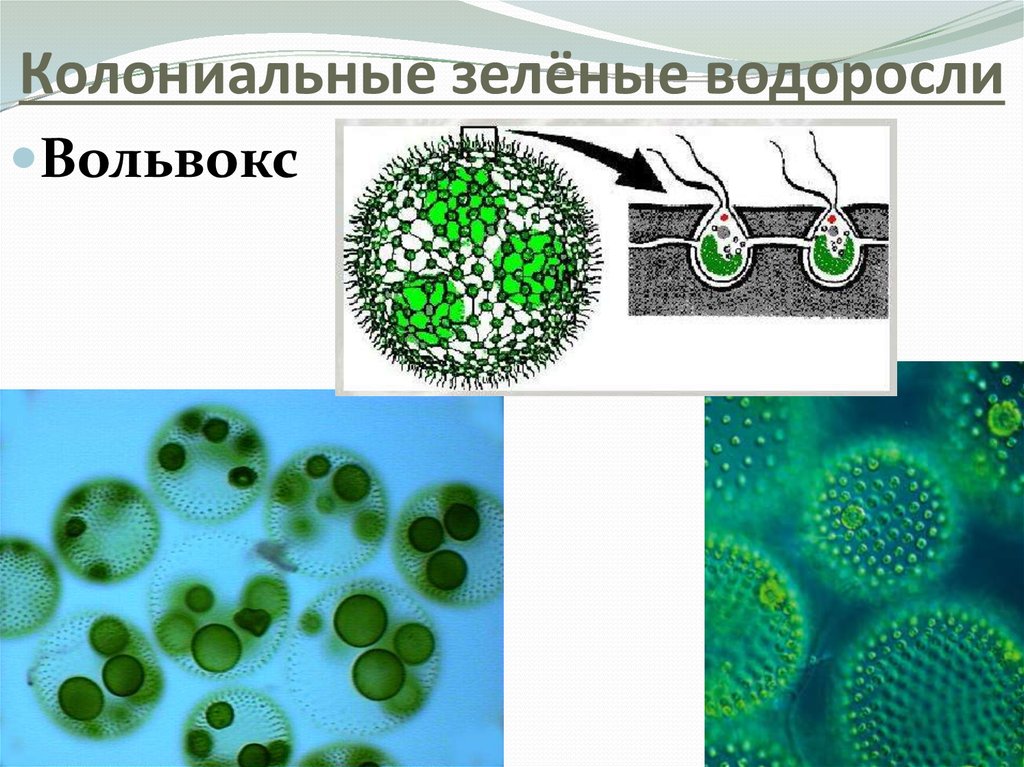
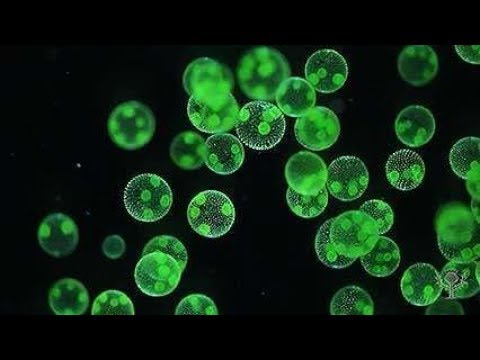
Хлорофит Volvox — один из немногих примеров колониального организма, который в некотором роде ведет себя как совокупность отдельных клеток, а в другом — как специализированные клетки многоклеточного организма. Колонии Volvox содержат от 500 до 60 000 клеток, каждая с двумя жгутиками, заключенными в полую сферическую матрицу, состоящую из студенистого секрета гликопротеина. Отдельные клетки Volvox перемещаются скоординированно и связаны между собой цитоплазматическими мостиками. Только несколько клеток размножаются, образуя дочерние колонии, что является примером основной специализации клеток в этом организме.
Рисунок: Volvox aureus: Volvox aureus — зеленая водоросль надгруппы Archaeplastida. Этот вид существует в виде колонии, состоящей из клеток, погруженных в гелеобразный матрикс и переплетенных друг с другом с помощью волосовидных цитоплазматических отростков.
Среди хлорофитов представлены настоящие многоклеточные организмы, такие как морской салат, Ulva . Кроме того, некоторые хлорофиты существуют в виде крупных многоядерных одиночных клеток. Виды рода Caulerpa имеют уплощенную, похожую на папоротник листву и могут достигать 3 метров в длину. Виды Caulerpa подвергаются ядерному делению, но их клетки не завершают цитокинез, оставаясь вместо этого массивными и сложными одиночными клетками.
Рисунок: Caulerpa Taxifolia : Caulerpa Taxifolia представляет собой хлорофит, состоящий из одной клетки, потенциально содержащей тысячи ядер.Ключевые моменты
- Архепластиды обычно связаны с их родством с наземными растениями; кроме того, молекулярные данные показывают, что Archaeplastida произошли от эндосимбиотических отношений между гетеротрофным протистом и цианобактерией.
- Красные водоросли (родофиты) классифицируются как Archaeplastida и чаще всего характеризуются наличием красного пигмента фикоэритрина; однако есть красные водоросли, в которых отсутствуют фикоэритрины, и их можно отнести к паразитам.
- Красные водоросли обычно существуют в виде многоклеточных простейших, у которых отсутствуют жгутики; однако они также могут существовать как одноклеточные организмы.
- Зеленые водоросли являются наиболее многочисленной группой водорослей и могут быть дополнительно классифицированы как хлорофиты и харофиты.
- Харофиты — это зеленые водоросли, которые напоминают наземные растения и являются их ближайшими живыми родственниками.
- Хлорофиты — это зеленые водоросли, имеющие широкий спектр форм; они могут быть одноклеточными, многоклеточными или колониальными.
Ключевые термины
- эндосимбиотик : живущий в теле или клетках другого организма
- планктон : общий термин для всех организмов, плавающих в море
Эта страница под названием 8.19A: Archaeplastida распространяется под лицензией CC BY-SA 4.0 и была создана, изменена и/или курирована Boundless.