Изучение темы: «Простейшие» в 7-м классе
Урок 1 Тема Среда обитания, строение и дыхание простейших.
Задачи:
1. Способствовать изучению
особенностей строения одноклеточных животных
(простейших)
2. Подвести учащихся к пониманию единства
органического мира.
3. Закрепить умение работать с микроскопом.
4. Использовать активные формы и приемы работы с
целью проявления личного уровня зунов.
Понятия: Одноклеточное животное, дыхание, питание, хищник,
Оборудование:
1. На каждую парту микроскоп.
2. Таблицы “Простейшие” “Хламидомонада”.
3. Микропрепараты “Амеба”, “Эвглена”,
“Туфелька”.
Ход урока.
1. Вступительное слово учителя:
Простейшие — это первая группа
животных, которую мы изучаем.
1) строение,
2) среда обитания,
3) как дышат; как размножаются; как передвигаются?
На эти вопросы мы с вами и будем отвечать.
2. Изучение нового материала.
1) История открытия – сообщение учащегося.“300 лет назад жил в г. Дельфте, что в Голландии шлифовальщик стекол Антон Левенгук. Через свои стекла он рассматривал окружающий его мир. Взяв застоявшуюся воду из бочки, он увидел в ней движущиеся организмы. Левенгук очень удивился и назвал их ничтожнейшими зверушками. Позднее ученые дали им название простейшие. Заслуга Левенгука перед наукой велика: во-первых, он открыл не видимых невооружённым глазом животных. И, во-вторых, он сделал микроскоп орудием изучением природы”.
2) Изучение строения простейших.
– Приготовьте таблицу (1) как на доске. Надписи сделайте точно также.
Надписи сделайте точно также.
царство |
животные |
||
Вид |
Амеба обыкновенная |
Эвглена зеленая |
Туфелька обыкновенная |
Строение |
Ложноножка Оболочка Цитоплазма Ядро Пищеварительная вакуоль
|
Жгутик Стигма Сократительная вакуоль Оболочка Хлоропласт Цитоплазма Ядро Запасные питательные вещества |
Реснички Сократительная вакуоль Пищеварительная вакуоль Ядро М. Рот Порошица ЦитоплазмаОболочка |
Среда |
Загрязненная |
С гнилыми листьями |
Загрязненная |
Движение |
Ложноножки |
Жгутики |
Реснички |
Питание |
Гетеротрофное, хищник |
Гетеротрофное, автотрофное |
Гетеротрофное, хищник |
Кислород поступает через поверхность |
Кислород поступает через поверхность | Кислород поступает через поверхность | |
 Ядро
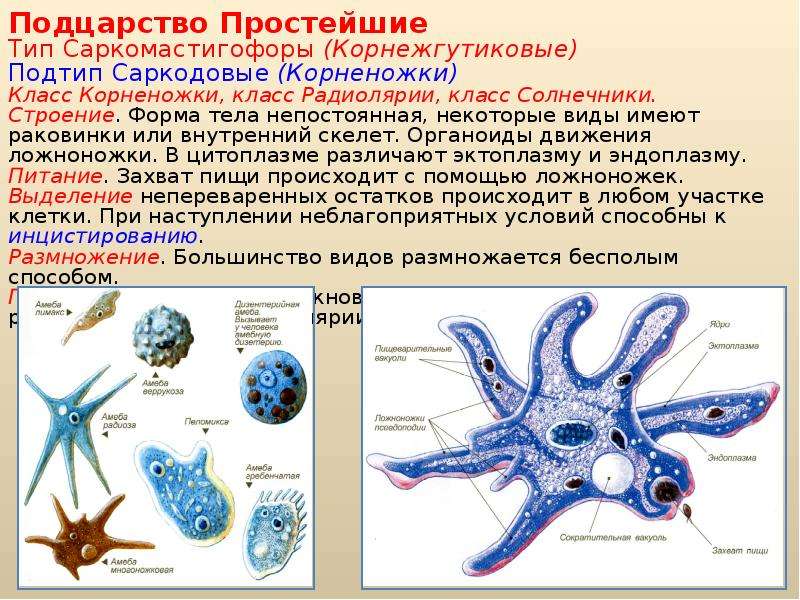
ЯдроПодцарство Простейшие. Характерные черты организации. Классификация. Значение для медицины
Характерные черты организации. Классификация. Значение для медицины
· Царство Животные
· Подцарство Многоклеточные (metazoa)
· Подцартсво Простейшие (одноклеточные; protozoa). Выделяют 7 типов.
В подцарстве одноклеточные с токи зрения медицинской паразитологии интерес вызывают 3 типа: саркомастигофоры (sarcomastigophora), апикомплексы (apicomplexa), инфузории (ciliphora). Простейшие – это гетеротрофные и автотрофные организмы, одноклеточные, которые могут могу быть одноядерными или многоядерными и в ряде случаев образуют колонии.
Простейшие были открыты Левенгуком. В настоящее время известно более 39 тыс. видов простейших. Понятие о простейших как одноклеточных организмах было сформулировано лишь в 19 веке 2 учеными Зибольдом и Келлкиером. Согласно новым принципам классификации организмов жгутиковые и саркодные объединяются в тип саркомастиговоры и насчитывают около 25 тыс. видов. В основу деления простейших на типы положены принципы строения их ядерного аппарата, органелл движения, типов размножения и жизненных циклов.
Деление клеток у многоклеточных приводит к росту организма, а у одноклеточных – к размножению. Жизненный цикл простейших представляет собой циклически повторяющийся отрезок развития вида между двумя одноименными стадиями. Средние размеры простейших измеряются как правило несколькими десятками микрометров. Самые мелкие – 2-4 мкм, самые крупные – до 1 мм.
(2)
Форма тела у простейших очень разнооьразная (непостоянная, постоянная). Также как и тип симметрии (внешняя билатеральная, лучевая). Строение клетки простейших хар-ся всеми основными признаками клеточного строения эукариот. Клетка простейших состоит из цитоплазмы и одного или нескольких ядер. Поверх мембраны могут располагаться оболочки (слизистая,хитиноидная), может образовываться скелет (известковый или кремниевый органический). В цитоплазме различается 2 слоя: наружный прозрачный – эктоплазма, внутренний зернистый – эндоплазма (где располагаются все органеллы). У простейших органеллами движения являются псевдоподии, жгутики (небольшое количесвто на переднем колнце тела), реснички, ундулирующая мембрана.
У человека могут паразитировать представители амеб, споровиков, жгутиконосцев и ресничных инфузорий. Кроме этого простейшие встречаются в различных средах жизни. Чаще всего в морских и пресных водах. Простейшие паразитируют в различных тканях и полостях тела хозяина, проникая в его организм в соответствии с эволюционно сложившимся механизмом передачи. Это может быть алементарный способ, трансмессивный, половой путь и др.
Это может быть алементарный способ, трансмессивный, половой путь и др.
Простейшие у человека паразитируют в ротовой полости (ротовая амеба, entamoeba gingivalis; ротовая трихомонада), в тонкой кишке (лямблия кишечная, кишечная кокцидия, криптоспоридиум), в мочеполовых путях (вагинальная или урогенетальная трихомонада), остальные простейшие локализуются в толстом кишечнике. Все простейшие – паразиты, рамзножаются в организме человека простым делением и только некоторым из них свойственен половой путь размножения. Диагностика протозоозов (заболевания, вызва. Простейшими паразитами) основывается на обнаружении их в крови (плазмодий, трипаносомы), в костном мозге (лейшмания Донована), фекалиях (возбудители амебиаза, балантидиаз), выделениях из мочеполовых путей (урогенетальная трихомонада), иммунологические методы (токсоплазмоз).
В пищеварительном тракте у человека паразитируют более 15 видов простейших, из них амеб – 7 видов, жгутиковых – 6, споровиков – 2, инфузорий – 1.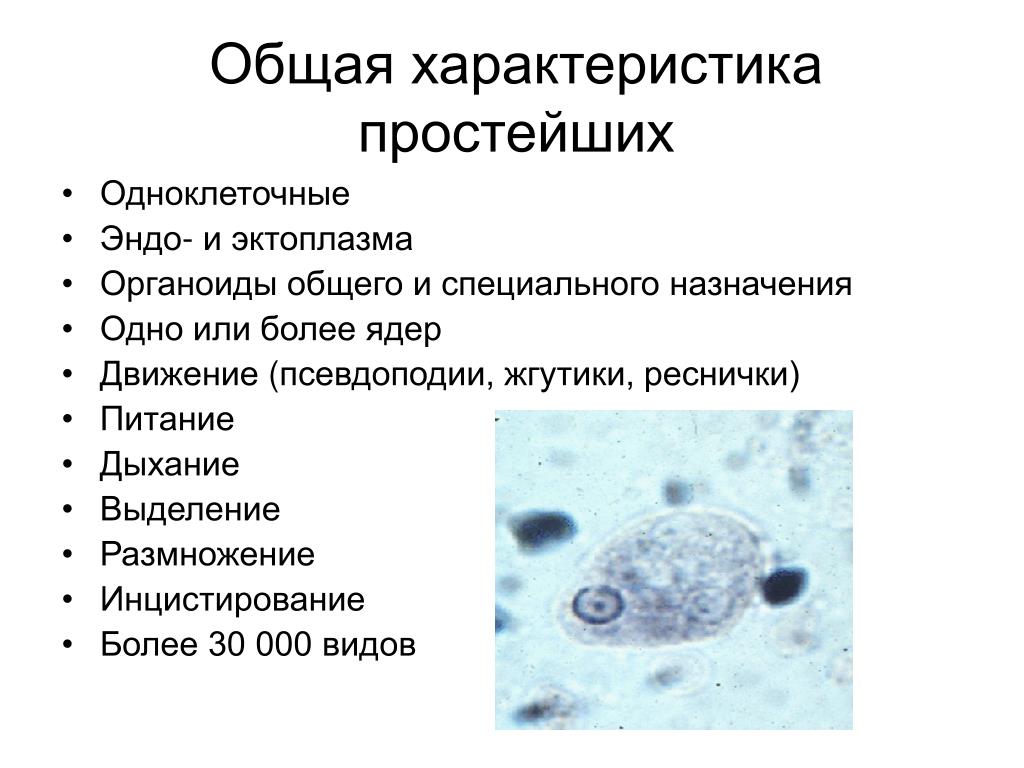 Простейшие существуют в теле человека в вегетативной форме и в форме цисты. Циста – пассивная стадия. Вегетатвиная – тропазоид – активная стадия. Есть простейшие, которые не образую цист (трихомонада кишечная и ротовая, ротовая амеба). Вегетативные формы во внешней среде неустойчивы и поэтому быстро погибают. Цисты образуются из вегетативных форм нижних отделов кишечника. У споровиков в результате полового процесса образуются ооцисты, которые по строению отличаются от цист других простейших (у ктоксоплазмы ооциста включает 2 цисты под 1 оболочкой). 1 организм-паразит, размножившись, способен привести к гибели хозяина, т.к. большинство простейших имеют время генерации от 6 до 24 часов.
Простейшие существуют в теле человека в вегетативной форме и в форме цисты. Циста – пассивная стадия. Вегетатвиная – тропазоид – активная стадия. Есть простейшие, которые не образую цист (трихомонада кишечная и ротовая, ротовая амеба). Вегетативные формы во внешней среде неустойчивы и поэтому быстро погибают. Цисты образуются из вегетативных форм нижних отделов кишечника. У споровиков в результате полового процесса образуются ооцисты, которые по строению отличаются от цист других простейших (у ктоксоплазмы ооциста включает 2 цисты под 1 оболочкой). 1 организм-паразит, размножившись, способен привести к гибели хозяина, т.к. большинство простейших имеют время генерации от 6 до 24 часов.
К типу саркомастигофоры относятся свободноживущие и паразитические простейшие, органиодами движения которых являются псевдоподии и жгутики. В настоящее время объединяются в 1 тип из-за наличия переходных форм, обладающих сразу 2 типами органелл (мастигоамебы), половой процесс у саркомастигофор – это копуляция, но большинство видов разножаются бесполым путем. Тип включает в себя подтип саркодовые, подтип жгутиконосцы, подтип аполины. У саркодовых наибольшее значение имеет класс кореножки (ризопода), отряд амебы. Саркодовые не имеют постоянной формы тела, покрыты толькой мембранной, не имеют уплотненных оболочек, но могут образовывать раковину или внутренний скелет. Большинство саркодовых – свободноживущие виды. Передвигаются с помощью псевдоподий или за счет циркуляции цитоплазмы. Жгутики могут образовываться на кратковренменной стадии развития (гаметы, зооспоры).
Тип включает в себя подтип саркодовые, подтип жгутиконосцы, подтип аполины. У саркодовых наибольшее значение имеет класс кореножки (ризопода), отряд амебы. Саркодовые не имеют постоянной формы тела, покрыты толькой мембранной, не имеют уплотненных оболочек, но могут образовывать раковину или внутренний скелет. Большинство саркодовых – свободноживущие виды. Передвигаются с помощью псевдоподий или за счет циркуляции цитоплазмы. Жгутики могут образовываться на кратковренменной стадии развития (гаметы, зооспоры).
Наука о паразитических простейших — возбудителях болезней человека, носит название медицинская протозоология.
Морфологическая характеристика. Тело простейших имеет микроскопические размеры и представлено одной клеткой. Клетка эукариотическая. Органоиды простейших осуществляют функции целого организма.
Органоиды передвижения — псевдоподии, жгутики или реснички.
Органоиды питания — пищеварительные вакуоли. Непереваренные остатки пищи выбрасываются наружу.
Органоиды выделения и осморегуляции — сократительные вакуоли. У паразитических и морских форм этот тип вакуоли может отсутствовать.
Органоиды защиты — трихоцисты. Существу001ют у инфузорий.
Размножаются простейшие бесполым и половым путем. Бесполое размножение: бинарное и множественное (шизогония) деления. Половой процесс: копуляция и конъюгация.
Жизненный цикл. Стадии развития в цикле часто повторяются с определенной закономерностью: зигота, бесполое поколение, половое поколение и вновь зигота. Имеются активно питающиеся, подвижные стадии — трофозоиты. Простейшие способны к инцистированию. Среда обитания: свободноживущих — в пресной и морской воде, в почве, паразитических форм — в жидких средах и клетках организмов.
Классификация: 1. Класс Саркодовые — Sarcodina; 2. Класс Жгутиковые — Flagellata s. Mastigophora; 3. Класс Инфузории — Infusoria; 4. Класс Споровики — Sporozoa.
Паразитические виды встречаются в каждом классе
ЛЕКЦИЯ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
План
1.История изучения.
2.Компоненты клетки.
3.Формы тела, покровы, симметрия.
4.Органеллы движения, способы движения.
5.Питание простейших. Органеллы питания.
6.Сократительная вакуоль и ее функции.
7.Дыхание простейших.
8.Поведение простейших.
9. Размножение и жизненные циклы.
10. Классификации.
Животные населяют разнообразные среды. Большинство встречается в пресных и соленых водоемах, часть видов обитает в почве, множествоведет паразитический образ жизни. В настоящее время известно свыше 39 тыс. видов
1. Историяизучения
Изучение простейших началось значительно позже, чем изучение большинства других групп животных. Первооткрывателем этих животных стал А. ван Левенгук (1632–1723), который описал и зарисовал различных простейших и назвал их animalcula. В 1718 г. Жабло изучал строение инфузорий. Амебы открыты Р. Розенхофом в 1755 г. К. Линней в своей «Системе природы» (1759 г.) всех ему известных простейших объединил в один род, который назвал Chaos infusorium. О. Ф. Мюллер в труде « Animalcula infusoria» (1770 г.) приводит описание 377 видов микроскопических организмов, главным образом простейших. Воззрения исследователей на простейших в 18 и начале 19 века носили противоречивый характер. Х. Эренберг считал, что простейшие это сложно организованные существа, обладающие различными системами органов, отличаются от других животных размерами. Ф. Дюжарден, напротив, утверждал, что простейшие не обладают никакой внутренней организацией, построены из бесструктурного вещества – саркоды. Название Protozoa введено в науку в 1820 г. Гольдфусом, он относил сюда не только простейших, но и многих других животных. В 1845 г. К. Зибольд и Р. А. Келликер сформулировали представление о простейших как об одноклеточных организмах. Во второй половине 19 века большую роль в изучении простейших сыграл О.
В 1718 г. Жабло изучал строение инфузорий. Амебы открыты Р. Розенхофом в 1755 г. К. Линней в своей «Системе природы» (1759 г.) всех ему известных простейших объединил в один род, который назвал Chaos infusorium. О. Ф. Мюллер в труде « Animalcula infusoria» (1770 г.) приводит описание 377 видов микроскопических организмов, главным образом простейших. Воззрения исследователей на простейших в 18 и начале 19 века носили противоречивый характер. Х. Эренберг считал, что простейшие это сложно организованные существа, обладающие различными системами органов, отличаются от других животных размерами. Ф. Дюжарден, напротив, утверждал, что простейшие не обладают никакой внутренней организацией, построены из бесструктурного вещества – саркоды. Название Protozoa введено в науку в 1820 г. Гольдфусом, он относил сюда не только простейших, но и многих других животных. В 1845 г. К. Зибольд и Р. А. Келликер сформулировали представление о простейших как об одноклеточных организмах. Во второй половине 19 века большую роль в изучении простейших сыграл О. Бючли. Он положил начало исследованию форм размножения. Исследования Мопа связаны с изучением размножения
Бючли. Он положил начало исследованию форм размножения. Исследования Мопа связаны с изучением размножения
Зоология беспозвоночных. Конспект лекций | 21 |
ЛЕКЦИЯ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
1. История изучения
инфузорий. В конце 19 и начале 20 века внимание исследователей привлекает изучение паразитических простейших и циклов их развития (работы Р. Шаудина, П. П. Грасси, Н. Я. Данилевского). В 20 веке изучается строение, физиология, размножение простейших разных групп (исследования М. У. Калкинса, П. Вудруфа, Р. Гертвига, С. И. Метальникова, В. Т. Шевякова, В. А. Догеля, В. М. Марциновского, Ю. А. Филипченко и др.). За последние годы объем знаний об одноклеточных существенно вырос. Выделяются группы организмов с разными планами строения, которых рассматривают внутри царства Protista.
Понятие «простейшие» отражает морфологические особенности: соответствие строения простейших схеме строения клетки многоклеточного животного. В физиологическом отношении простейшие являются целостными организмами, они эквивалентны организму многоклеточных животных.
В физиологическом отношении простейшие являются целостными организмами, они эквивалентны организму многоклеточных животных.
2. Компонентыклетки
Компоненты тела можно подразделить на три группы: общеклеточные структуры, специальные органеллы и включения.
К общеклеточным структурам относятся: цитоплазма, ядро, митохондрии, эндоплазматическая сеть, рибосомы, лизосомы, аппарат Гольджи, центриоль.
Цитоплазма подразделяется на экто- и эндоплазму (у лучевиков на внекапсулярную и внутрикапсулярную цитоплазму). Цитоплазма ограничена снаружи клеточной мембраной (фосфоролипидный биослой). К плазматической мембране снаружи прилегает гликокаликс. Он образован белками и углеводными цепями, отходящими от наружной поверхности мембраны, содержит рецепторные молекулы и связан с информационной системой клетки. С помощью гликокаликса клетки способны накапливать из окружающей среды различные вещества, которые затем включаются в нее путем эндоцитоза. Плазмалемма некоторых инфузорий окружена мембраноподобной структурой (перилеммой).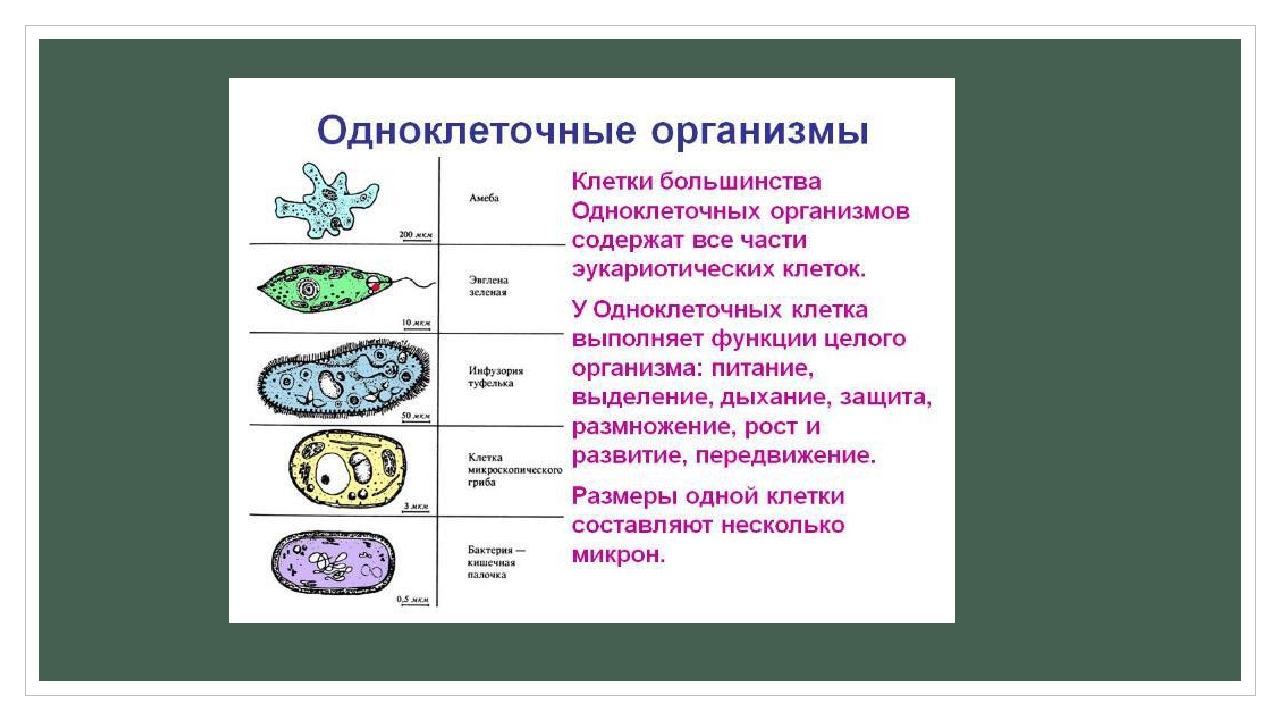 Под мембраной жгутиконосцев располагается перипласт, представленный белковыми или целлюлозными пластинками. В цитоплазме постоянно происходит изменение состояния белков – агрегатного состояния цитоплазмы – состояние золя и состояние геля. В состоянии золя цитоплазма характеризуется текучестью, при переходе в состояние геля – утратой текучести, повышением плотности.
Под мембраной жгутиконосцев располагается перипласт, представленный белковыми или целлюлозными пластинками. В цитоплазме постоянно происходит изменение состояния белков – агрегатного состояния цитоплазмы – состояние золя и состояние геля. В состоянии золя цитоплазма характеризуется текучестью, при переходе в состояние геля – утратой текучести, повышением плотности.
Внеклеточными образованиями являются чешуйки, системы фибрилл, внеклеточные домики, у растительных протистов клеточные стенки.
Имеется либо одно ядро, либо несколько ядер. Ядро имеет двухслойную мембрану с многочисленными порами, кариоплазму, в которой распределены хроматин и ядрышки. В зависимости от числа ядер простейшие подразделяются на моноэнергидных и полиэнергидных. Организмы с многочисленными одинаковыми ядрами называются гомокариотными. Если ядра клетки различаются между собой – организмы гетерокариотные. Явление,
Зоология беспозвоночных. Конспект лекций | 22 |
ЛЕКЦИЯ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
2. Компоненты клетки
при котором ядра выполняют разные функции (мелкое – микронуклеус – генеративное и крупное – макронуклеус – соматическое), называется ядерным дуализмом. Это характерно для инфузорий: каждая клетка содержит от 1 до 20 диплоидных микронуклеусов, количество макронуклеусов также изменчиво. Макронуклеусы имеют разнообразную форму и структурно сложны.
Специальными органеллами клетки являются: сократительные и пищеварительные вакуоли, микрофиламенты, микротрубочки, экструсомы, порошица, стигма, жгутики и реснички. Микрофиламенты – нити сократительного белка актина – участвуют в процессах сокращения, клеточном делении, образуют фибриллы. Микротрубочки – полые цилиндры, стенка которых состоит из полимеров тубулина – выполняют функцию цитоскелета, принимают участие в делении ядра, в формировании ротового аппарата, удерживают
вопределенном положении органеллы, участвуют в динамических процессах
вклетках (определенные типы пузырьков транспортируются вдоль лент из микротрубочек), входят в состав жгутиков, ресничек. Экструсомы встречаются у жгутиконосцев, инфузорий, саркодовых. Это разнообразные по форме пузырьки, в ответ на раздражение животные выделяют их содержимое наружу. Известно 10 типов экструсом.
Экструсомы встречаются у жгутиконосцев, инфузорий, саркодовых. Это разнообразные по форме пузырьки, в ответ на раздражение животные выделяют их содержимое наружу. Известно 10 типов экструсом.
Включениями являются: липидные капельки, белковые кристаллы, гранулы резервных полисахаридов, симбиотические организмы.
3. Форматела, покровы, симметрия
Форма тела разнообразная, имеются животные с непостоянной формой.
Покровы тела образованы мембраной, пелликулой. Пелликула представляет собой уплотненный периферический слой цитоплазмы с опорными белковыми фибриллами (микрофиламентами, микротрубочками). У инфузорий краевая цитоплазма (кортекс) устроена сложно. В состав кортекса инфузорий входят: пелликула, эпиплазма и комплекс кинетосом.
Унекоторых простейших имеется скелет. Скелет может быть внешним
ивнутренним (например, у аксоподии), органическим, неорганическим или смешанным. Чешуйки и домики являются внешними опорными структурами. Чешуйки формируются в диктиосомах и выделяются наружу путем экзоцитоза. Домики имеют органическую основу, в которую встраиваются органические или неорганические части.
Домики имеют органическую основу, в которую встраиваются органические или неорганические части.
Часть одноклеточных животных в ответ на неблагоприятные условия внешней среды образуют цисты. Циста может являться этапом в жизненном цикле животного. Материалом для формирования цист может служить хитин, целлюлоза, кремнезем и др.
Типы симметрии разнообразны. У протист выделяют девять типов симметрии: анаксонная (симметрия отсутствует), сферическая (характерно неопределенно большое число осей и плоскостей симметрии, которые пере-
Зоология беспозвоночных. Конспект лекций | 23 |
ЛЕКЦИЯ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
3. Форма тела, покровы, симметрия
секаются в одной точке), неопределенно полиаксонная (выделяют центр симметрии, большое, но конечное число осей симметрии, положение их не является строго постоянным), правильно полиаксонная (характерно определенное число осей симметрии, расходятся под определенными углами), ставраксонная гомополярная – радиальная (имеется одна ось симметрии, которая пересекается плоскостью симметрии так, что оба полюса ее одинаковые), ставраксонная гетерополярная (в этом случае ось симметрии не пересекается плоскостью), монаксонная гетерополярная (при данном типе центр симметрии отсутствует, полюса различаются, выделяется плоскость симметрии), билатеральная (элементом симметрии является только плоскость симметрии), вращательная (имеется только ось симметрии).
4. Органеллыдвижения, способыдвижения
Выделяют следующие органеллы движения:
•Псевдоподии. Типы псевдоподий: лобоподии, филоподии, ризоподии и аксоподии. Способ движения – амебоидный. У амеб найдены актиновые и миозиновые филаменты, благодаря деятельности которых возникает ток цитоплазамы. Существует две гипотезы, объясняющие амебоидное движение: гипотеза потока цитоплазмы под давлением и гипотеза сокращения фронтальной зоны. В настоящее время поддерживается гипотеза сокращения актинмиозиновых комплексов кортикальной зоны эктоплазмы почти по всей длине клетки (за исключением уроида – заднего отдела – и самого переднего конца).
•Жгутики. Один или несколько. При наличии нескольких жгутиков в зависимости от длины они подразделяются на изоконтные (равные по длине) и гетероконтные. Выделяют жгутик (локомоторная часть, ундулиподия), переходную зону, кинетосому и корешки. В состав жгутика входит аксонема. Аксонема представляет собой совокупность микротрубочек (9х2+2- конфигурация) и структур из микрофиламентов. Микротрубочки представлены девятью периферическими двойными (дуплеты – 9 х 2) и двумя одиночными (синглеты – 2) центральными микротрубочками. Дуплеты связаны друг с другом и центральными трубочками. В кинетосоме центральные трубочки заканчиваются в аксиальном зерне, а дуплеты становятся триплетами (9 х 3 + 0- конфигурация). На концевом участке жгутика центральные трубочки окружены девятью синглетами. Кинетосома прикрепляется к ядру или мембране цитоскелетными корешками. Корешковая система представлена микротрубочками и микрофиламентами. Они расходятся под определенным углом от кинетосомы. У эвгленовых и кинетопластид вдоль аксонемы проходит параксиальный пучок, у динофлагеллят рядом с аксионемой располагается сократимый пучок филаментов. Способы биения жгутиков: в одной плоскости (унипланарный) или по спирали (геликоидальный). У некоторых паразитических жгутиконосцев вдоль тела проходит аксостиль (состоит из лент микротрубочек, начинающихся от кинетосом). Изгибание аксостиля обеспечивает змеевидное
Микротрубочки представлены девятью периферическими двойными (дуплеты – 9 х 2) и двумя одиночными (синглеты – 2) центральными микротрубочками. Дуплеты связаны друг с другом и центральными трубочками. В кинетосоме центральные трубочки заканчиваются в аксиальном зерне, а дуплеты становятся триплетами (9 х 3 + 0- конфигурация). На концевом участке жгутика центральные трубочки окружены девятью синглетами. Кинетосома прикрепляется к ядру или мембране цитоскелетными корешками. Корешковая система представлена микротрубочками и микрофиламентами. Они расходятся под определенным углом от кинетосомы. У эвгленовых и кинетопластид вдоль аксонемы проходит параксиальный пучок, у динофлагеллят рядом с аксионемой располагается сократимый пучок филаментов. Способы биения жгутиков: в одной плоскости (унипланарный) или по спирали (геликоидальный). У некоторых паразитических жгутиконосцев вдоль тела проходит аксостиль (состоит из лент микротрубочек, начинающихся от кинетосом). Изгибание аксостиля обеспечивает змеевидное
Зоология беспозвоночных. | 24 |
 Конспект лекций
Конспект лекцийЛЕКЦИЯ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
4. Органеллы движения, способы движения
движение. У немногих представителей имеется гаптонема (состоит из 6–7 микротрубочек и цистерны эндоплазматической сети – ЭС), она расположена между двумя жгутиками. Движение – свертывание в рулон. Жгутики многих простейших несут «волоски» – мастигонемы. Когда по такому жгутику по направлению к свободному концу проходит волна, за счет мастигонем жгутик тянет за собой клетку.
•Реснички. Строение подобно строению жгутика. Способ движения – гребля. Работа цилиатуры координируется таким образом, что соседние реснички находятся в одинаковой фазе биения, а по сравнению с соседними ресничными рядами их биение сдвинуто по фазе. Благодаря этому по поверхности клетки пробегают сменяющие друг друга волны. Такая волна называется метахрональной.
•Мионемы – волоконца, сокращающие клетку, и ламеллы, вытягивающие клетку до исходного состояния.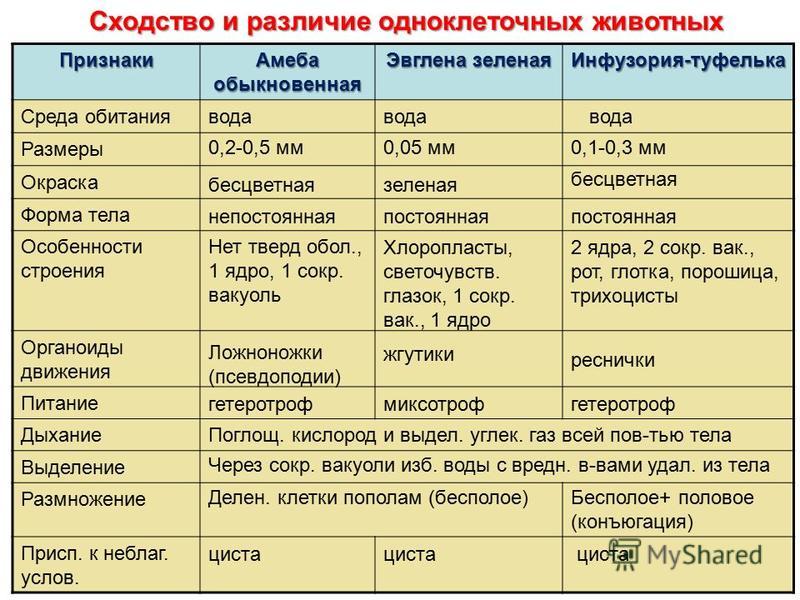 Такая система сокращения – вытягивания распространена у инфузорий. Ламеллы представлены микротрубочками, тянутся от переднего конца клетки к заднему концу. Мионемы образованы микрофиламентами.
Такая система сокращения – вытягивания распространена у инфузорий. Ламеллы представлены микротрубочками, тянутся от переднего конца клетки к заднему концу. Мионемы образованы микрофиламентами.
•Метаболия (эвгленоидное движение) – перистальтические волны де-
формации клетки.
Скольжение у трофических стадий грегарин, спорозоитов кокцидий произходит за счет колебания гребней пелликулы.
Прикрепительные органеллы.
Для протист характерны различные способы прикрепления: внутри- и внеклеточные стебельки, пальцевидные выросты, выделения слизистого материала, присоски, эпимериты. Способ образования стебельков различен. Присоски разных представителей образованы различающимися структурными единицами. В присосках дипломонад структурным компонентом являются кольцевидно расположенные микротрубочки, прикрепительный диск – триходин построен из белковых элементов.
5.Питаниепростейших. Органеллыпитания
Упростейших встречаются следующие типы питания: автотрофное, гетеротрофное и миксотрофное. Способ питания автотрофов голофитный (происходит фотосинтез, зеленый пигмент локализован в хроматофорах, резервные вещества – парамил, крахмал). Гетеротрофные животные питаются готовыми органическими веществами. Способы питания – голозойный (заглатывание оформленной твердой пищи путем фагоцитоза) и сапрофитный (питание растворенными органическими веществами путем пиноцитоза). Миксотрофный тип питания – смешанный тип. Способы захвата пищи различны (использование токсицистов, создание токов воды к клетке или клеточному рту, палочковые аппараты из микротрубочек, щупальца и др.).
Способ питания автотрофов голофитный (происходит фотосинтез, зеленый пигмент локализован в хроматофорах, резервные вещества – парамил, крахмал). Гетеротрофные животные питаются готовыми органическими веществами. Способы питания – голозойный (заглатывание оформленной твердой пищи путем фагоцитоза) и сапрофитный (питание растворенными органическими веществами путем пиноцитоза). Миксотрофный тип питания – смешанный тип. Способы захвата пищи различны (использование токсицистов, создание токов воды к клетке или клеточному рту, палочковые аппараты из микротрубочек, щупальца и др.).
Зоология беспозвоночных. Конспект лекций | 25 |
ЛЕКЦИЯ 2. ОБЩАЯ ХАРАКТЕРИСТИКА ПРОСТЕЙШИХ
5. Питание простейших. Органеллы питания
Органеллы питания: пищеварительные или пиноцитозные вакуоли, у некоторых представителей имеются клеточный рот (цитостом), клеточная глотка (цитофаринкс) и клеточное анальное отверстие – порошица (цитопиг, цитопрокт).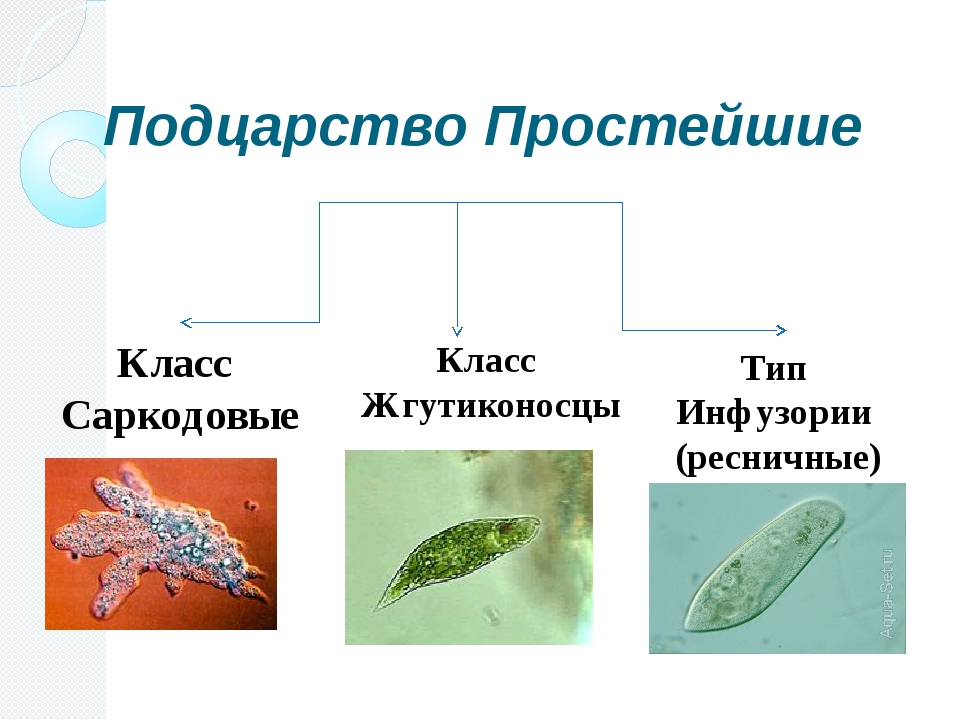 Пиноцитозные вакуоли образуются путем эндоцитоза (впячивания клеточной мембраны, впячивания отшнуровываются от мембраны в виде пузырьков). Пиноцитоз подразделяют на микропиноцитоз (поступает вода, ионы и мелкие молекулы, скорость поступления вещества зависит от его концентрации в окружающей среде) и макропиноцитоз (поглощаются белки и другие макромолекулы, скорость превышает ожидаемую исходя из градиента концентрации). Когда пища оказывается в клетке, лизосомы (содержат кислоты и гидролитические ферменты) сливаются с эндоцитозными пузырьками (пищеварительными вакуолями). Вакуоль перемещается в клетке, наблюдаются изменения в ее размерах (выделяют несколько фаз). Движение вакуоли по кругу (инфузории) – циклоз. Начало пищеварения проходит в щелочной среде, которая сменяется кислой, и заканчивается в щелочной среде. Непереваренные компоненты выводятся во внешнюю среду в результате слияния вакуоли с клеточной мембраной (экзоцитоз).
Пиноцитозные вакуоли образуются путем эндоцитоза (впячивания клеточной мембраны, впячивания отшнуровываются от мембраны в виде пузырьков). Пиноцитоз подразделяют на микропиноцитоз (поступает вода, ионы и мелкие молекулы, скорость поступления вещества зависит от его концентрации в окружающей среде) и макропиноцитоз (поглощаются белки и другие макромолекулы, скорость превышает ожидаемую исходя из градиента концентрации). Когда пища оказывается в клетке, лизосомы (содержат кислоты и гидролитические ферменты) сливаются с эндоцитозными пузырьками (пищеварительными вакуолями). Вакуоль перемещается в клетке, наблюдаются изменения в ее размерах (выделяют несколько фаз). Движение вакуоли по кругу (инфузории) – циклоз. Начало пищеварения проходит в щелочной среде, которая сменяется кислой, и заканчивается в щелочной среде. Непереваренные компоненты выводятся во внешнюю среду в результате слияния вакуоли с клеточной мембраной (экзоцитоз).
6. Сократительнаявакуольиеефункции
Комплекс сократительной вакуоли включает вакуоль и спонгиом, у некоторых (преимущественно инфузорий) сократительную пору. Спонгиом располагается возле сократительной вакуоли. Формы спонгиома: везикулярный (в виде пузырьков) и тубулярный (в виде постоянно присутствующих трубочек). Цикл пульсации вакуоли – чередование систолы (сокращения) и диастолы (наполнения). Число вакуолей варьируется у разных представителей. Сократительные вакуоли отсутствуют у большинства паразитических видов простейших и у обитателей соленых водоемов. У динофлагеллят их функцию выполняют пузулы – трубчатые впячивания мембраны, окруженные вакуолярной системой.
Спонгиом располагается возле сократительной вакуоли. Формы спонгиома: везикулярный (в виде пузырьков) и тубулярный (в виде постоянно присутствующих трубочек). Цикл пульсации вакуоли – чередование систолы (сокращения) и диастолы (наполнения). Число вакуолей варьируется у разных представителей. Сократительные вакуоли отсутствуют у большинства паразитических видов простейших и у обитателей соленых водоемов. У динофлагеллят их функцию выполняют пузулы – трубчатые впячивания мембраны, окруженные вакуолярной системой.
Функции сократительной вакуоли: выведение избытка воды, поддержание постоянной концентрации и определенного соотношения ионов в цитоплазме (ионная регуляция). Избыточная вода поступает в клетку дополнительно с пищей за счет осмоса: когда осмотическое давление в цитоплазме превышает таковое в окружающей среде.
Зоология беспозвоночных. Конспект лекций | 26 |
Простейшие — Большая советская энциклопедия
Просте́йшие
(Protozoa)
тип одноклеточных животных из группы эукариотов (См. Эукариоты). П. отличаются от всех других эукариотов, относимых к многоклеточным (См. Многоклеточные), тем, что их организм состоит из одной клетки, т. е. высший уровень организации у них клеточный. Почти все П. микроскопических размеров, по различны по уровню морфо-физиологической дифференцировки. Так, Амёбы устроены относительно просто (не имеют дифференцированных органоидов захвата пищи, движения, сокращения и т.п.), инфузории же обладают сложной организацией (имеют поверхностные пелликулярные структуры, опорные и сократительные фибриллы, органоиды движения — реснички и их производные, специальные органоиды захвата пищи, защиты и т.п.). Всем П. присущи типичная клеточная ультраструктура и комплекс органоидов общего назначения: митохондрии, эндоплазматическая сеть, элементы аппарата Гольджи, рибосомы, лизосомы. Ядро окружено типичной двухмембранной оболочкой с порами, содержит кариоплазму, хромосомы (в интерфазном ядре они обычно находятся в деспирализованном состоянии) и нуклеоли.
Эукариоты). П. отличаются от всех других эукариотов, относимых к многоклеточным (См. Многоклеточные), тем, что их организм состоит из одной клетки, т. е. высший уровень организации у них клеточный. Почти все П. микроскопических размеров, по различны по уровню морфо-физиологической дифференцировки. Так, Амёбы устроены относительно просто (не имеют дифференцированных органоидов захвата пищи, движения, сокращения и т.п.), инфузории же обладают сложной организацией (имеют поверхностные пелликулярные структуры, опорные и сократительные фибриллы, органоиды движения — реснички и их производные, специальные органоиды захвата пищи, защиты и т.п.). Всем П. присущи типичная клеточная ультраструктура и комплекс органоидов общего назначения: митохондрии, эндоплазматическая сеть, элементы аппарата Гольджи, рибосомы, лизосомы. Ядро окружено типичной двухмембранной оболочкой с порами, содержит кариоплазму, хромосомы (в интерфазном ядре они обычно находятся в деспирализованном состоянии) и нуклеоли.
Известно 25—30 тыс. видов П. Число же существующих в природе видов П., вероятно, в несколько раз больше, т.к. из-за микроскопических размеров и технических трудностей фауна П. недостаточно исследована, ежегодно описываются сотни новых видов. П. делят на 5 классов: Саркодовые, Жгутиковые, Споровики, Инфузории, Книдоспоридии. Имеется несколько прогрессивных филогенетических линий, ведущих к образованию крупных таксонов — фораминифер, радиолярий, инфузорий, у которых морфо-физиологическая дифференцировка наиболее сложная. Пути морфо-физиологического прогресса в пределах П. отличаются от таковых у многоклеточных. Для прогрессивной эволюции П. характерна полимеризация органоидов, высокий уровень полиплоидии, дифференцировка ядер на генеративные и вегетативные (у инфузорий их называют Микронуклеус и Макронуклеус). Для многих П. характерны циклы развития, выражающиеся в закономерном чередовании бесполого и полового размножения. Особой сложности достигают жизненные циклы у паразитических П. из класса споровиков (см. Кокцидии).
из класса споровиков (см. Кокцидии).
П. широко распространены в природе и занимают существенное место в цепях питания (См. Цепи питания) во многих биоценозах и биосфере в целом. Многие П. (жгутиковые, радиолярии, инфузории) входят в состав морского Планктона, где нередко, быстро размножаясь, достигают огромного количества. Они служат важным звеном в питании морского зоопланктона, особенно веслоногих ракообразных. Многие П. (фораминиферы, инфузории) входят и в состав морского Бентоса, обитающего от литорали до самых больших глубин. Описана фауна инфузорий, населяющих поверхностные слои морских песков. Ряд П. входит в состав пресноводного планктона и бентоса. Видовой состав П. пресных вод служит показателем степени их сапробности, т. е.
4.6.2. Подцарство Одноклеточные или Простейшие. Общая характеристика
4.6.2. Подцарство Одноклеточные или Простейшие. Общая характеристика
Общая характеристика
Основные термины и понятия, проверяемые в экзаменационной работе: амебы, балантидий, жгутиковые, инфузории, кокцидии, малярийный плазмодий, пищеварительная вакуоль, половой прогресс, порошица, саркодовые, сократительная вакуоль, споровики, эвглена зеленая.
Тело простейших животных состоит из одной клетки, осуществляющей все функции жизнедеятельности. Представители этого подцарства обладают всеми свойствами самостоятельного организма. Свободноживущие простейшие имеют дополнительные органоиды движения, питания, выделения, защиты и т.д. Некоторые из этих органоидов временные (ложноножки амебы), некоторые постоянные (жгутик эвглены, реснички инфузорий).
Среди простейших встречаются внутриклеточные паразиты, у которых отсутствуют пищеварительные вакуоли и органеллы движения; у них сложный жизненный цикл, иногда со сменой хозяина; они обладают высокой плодовитостью.
Особенности жизнедеятельности простейших: Обитают простейшие в воде, влажной почве, в других организмах. Для большинства из них характерно гетеротрофное питание: некоторые представители (эвгленовые) питаются смешанным способом. Днем они могут фотосинтезировать, ночью поглощают готовые органические вещества. Поглощают пищу эти животные разными способами: путем фагоцитоза и пиноцитоза, а также путем диффузии. Продукты обмена удаляются через сократительные вакуоли или же путем диффузии. Свободноживущие простейшие дышат кислородом, растворенным в воде или атмосферным воздухом. Паразитические простейшие живут в бескислородной среде и извлекают энергию для своей жизнедеятельности путем гликолиза Простейшие обладают раздражимостью. Они реагируют на изменение химического состава окружающей среды, на свет. Размножаются бесполым и половым способами. Бесполое размножение осуществляется путем митотического деления клеточных ядер; половое – путем копуляции или конъюгации. Неблагоприятные условия переживают в виде цисты.
Для большинства из них характерно гетеротрофное питание: некоторые представители (эвгленовые) питаются смешанным способом. Днем они могут фотосинтезировать, ночью поглощают готовые органические вещества. Поглощают пищу эти животные разными способами: путем фагоцитоза и пиноцитоза, а также путем диффузии. Продукты обмена удаляются через сократительные вакуоли или же путем диффузии. Свободноживущие простейшие дышат кислородом, растворенным в воде или атмосферным воздухом. Паразитические простейшие живут в бескислородной среде и извлекают энергию для своей жизнедеятельности путем гликолиза Простейшие обладают раздражимостью. Они реагируют на изменение химического состава окружающей среды, на свет. Размножаются бесполым и половым способами. Бесполое размножение осуществляется путем митотического деления клеточных ядер; половое – путем копуляции или конъюгации. Неблагоприятные условия переживают в виде цисты.
Роль простейших в природе и жизни человека:
– являются непременными участниками круговорота веществ и энергии в экосистемах, выступая в роли микро– консументов и редуцентов;
– образуют геологические залежи известняка, мела;
– являются объектами научного исследования;
– многие являются паразитами человека и животных, а также возбудителями заболеваний.
Многообразие простейших. Класс Саркодовые. Свободноживущие представители: амеба обыкновенная, фора– миниферы, радиолярии. Размер клетки 0,2—0,7 мм. Амеба живет в пресноводных, илистых водоемах. Цитоплазма образует выросты – псевдоподии, или ложноножки, служащие для передвижения и фагоцитоза. Клетка типичного эукариотического строения. Есть пищеварительные и сократительные вакуоли. Выполняет все функции самостоятельного организма. К патогенным простейшим этого типа относится дизентерийная амеба, вызывающая амебную дизентерию у человека. Паразитирует в кишечнике человека.
Класс Жгутиковые. Представители этого класса имеют постоянную форму тела, благодаря наличию уплотненной клеточной оболочки.
Животные передвигаются с помощью одного или нескольких жгутиков. Свободноживущие формы обитают в воде, влажной почве. Среди них есть как фотосинтезирующие, так и нефотосинтезирующие организмы. Паразитические простейшие (лямблия печеночная, трипаносома) и другие обитают в других организмах.
Паразитические простейшие (лямблия печеночная, трипаносома) и другие обитают в других организмах.
Эвглена зеленая имеет веретенообразную форму тела. Размер клетки около 0,05 мм. Передвигается эвглена с помощью жгутика – цитоплазматического выроста, состоящего из тонких фибрилл. На переднем конце находится светочувствительный глазок. В цитоплазме, помимо всех, характерных для животных клеток, органелл, находятся хроматофоры, содержащие хлорофилл. На свету эвглена способна к фотосинтезу. Поэтому ее относят к промежуточным, между растениями и животными, эволюционным формам. Размножается эвглена бесполым путем, делением надвое по продольной оси. Половое размножение осуществляется путем копуляции (слияния клеток).
К колониальным формам жгутиковых относится вольвокс.
Тип Споровики. К этому типу относятся паразитические простейшие, представителем которых является малярийный паразит. Его жизненный цикл протекает со сменой хозяев (комар – человек), которые, заражая друг друга, способствуют распространению паразита. При укусе человека самкой малярийного комара в его кровь проникают возбудители малярии – малярийный плазмодий. Он начинает активно размножаться в клетках печени человека. После размножения паразиты превращаются в другую стадию, поселяющуюся в эритроцитах крови. Бесполое размножение паразита в крови сопровождается массовой гибелью эритроцитов, выходом в кровь новых паразитов и токсичных продуктов их обмена, вызывающих лихорадку. Этот процесс цикличен, поэтому и лихорадка носит характер периодических приступов. Для дальнейшего развития возбудители должны попасть в желудок комара, где происходит их половое размножение и снова в слюнные железы самки.
При укусе человека самкой малярийного комара в его кровь проникают возбудители малярии – малярийный плазмодий. Он начинает активно размножаться в клетках печени человека. После размножения паразиты превращаются в другую стадию, поселяющуюся в эритроцитах крови. Бесполое размножение паразита в крови сопровождается массовой гибелью эритроцитов, выходом в кровь новых паразитов и токсичных продуктов их обмена, вызывающих лихорадку. Этот процесс цикличен, поэтому и лихорадка носит характер периодических приступов. Для дальнейшего развития возбудители должны попасть в желудок комара, где происходит их половое размножение и снова в слюнные железы самки.
Тип Инфузории. Класс ресничные инфузории. Тип насчитывает около 6 тыс. видов.
Представители – инфузория-туфелька, инфузория-трубач.
Инфузория-туфелька – животное размером 0,1—0,3 мм.
Ее клеточная оболочка покрыта ресничками, служащими для передвижения. В клетке два ядра – вегетативное, полиплоидное и генеративное, диплоидное. Ротовое углубление на теле образует ротовую воронку, переходящую в клеточный рот, ведущий в глотку. В глотке формируются пищеварительные вакуоли, переваривающие пищу. Непереваренные остатки пищи удаляются через отверстие – порошицу.
Ротовое углубление на теле образует ротовую воронку, переходящую в клеточный рот, ведущий в глотку. В глотке формируются пищеварительные вакуоли, переваривающие пищу. Непереваренные остатки пищи удаляются через отверстие – порошицу.
У инфузории-туфельки две сократительные вакуоли, расположенные в противоположных концах тела. Через них выводится избыток воды и продукты обмена веществ.
Размножение инфузории происходит как бесполым, так и половым путями. При бесполом размножении происходит продольное деление клетки. При половом процессе между двумя инфузориями образуется цитоплазматический мостик. Полиплоидные (большие) ядра разрушаются, а диплоидные (малые) ядра делятся мейозом с образованием четырех гаплоидных ядер, три из которых погибает, а четвертое делится пополам, но уже митозом. Образуется два ядра. Одно – стационарное и другое – мигрирующее. Затем между инфузориями происходит обмен мигрирующими ядрами. Потом стационарное и мигрировавшее ядра сливаются, особи расходятся и в них снова образуются большое и малое ядра.
К паразитическим инфузориям относится паразит толстого кишечника человека – инфузория-балантидий.
ПРИМЕРЫ ЗАДАНИЙ
Часть С
А1. Таксон, в который объединяются все простейшие, называется
1) царство
2) подцарство
3) тип
4) класс
А2. У простейших нет
1) тканей 3) обмена веществ
2) органоидов 4) полового размножения
А3. При полном окислении 1 молекулы глюкозы у амебы вырабатывается АТФ в количестве
1) 18 г/моль 3) 9 г/моль
2) 2 г/моль 4) 38 г/моль
А4. К паразитическим простейшим относится
1) амеба протей 3) трипаносома
2) эвглена зеленая 4) радиолярия
А5. Через сократительную вакуоль у инфузории происходит
1) удаление твердых продуктов жизнедеятельности
2) выделение жидких продуктов жизнедеятельности
3) выведение половых клеток – гамет
4) газообмен
А6. Заражение человека малярийным паразитом происходит при попадании в его организм
1) крови комара 3) личинок комара
2) слюны комара 5) яиц комара
А7.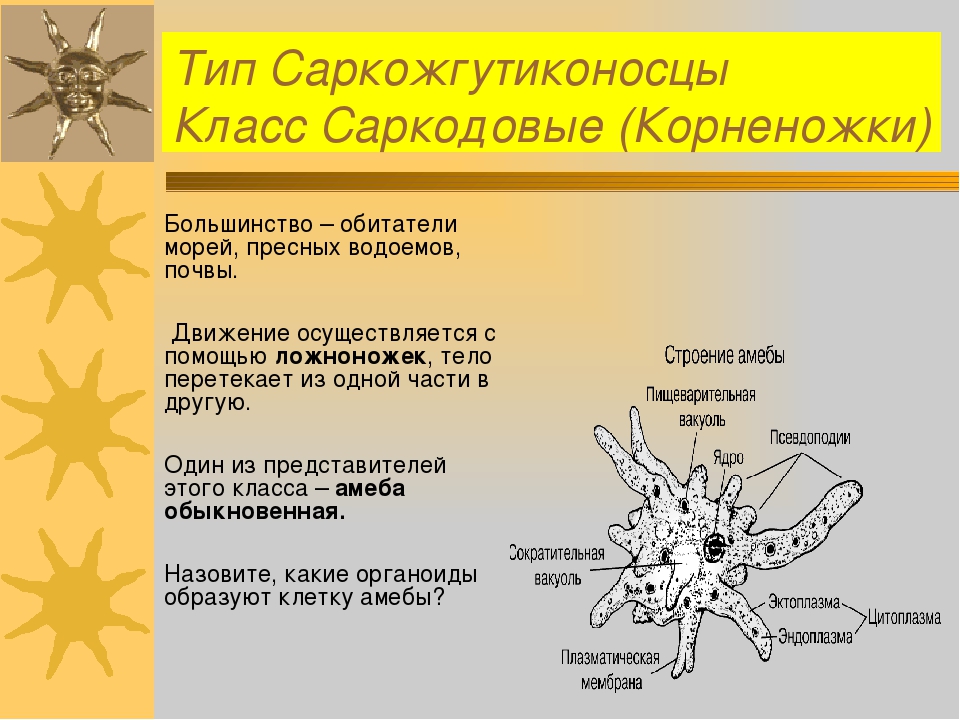 Бесполое размножение малярийного плазмодия происходит в
Бесполое размножение малярийного плазмодия происходит в
1) эритроцитах человека
2) эритроцитах и желудке комара
3) лейкоцитах человека
4) эритроцитах и клетках печени человека
А8. Какой из органоидов отсутствует в клетках инфузорий?
1) ядро 3) митохондрии
2) хлоропласты 4) аппарат Гольджи
А9. Что общего между эвгленой и хлореллой?
1) присутствие в клетках гликогена
2) способность к фотосинтезу
3) анаэробное дыхание
4) наличие жгутиков
А10. Среди инфузорий не встречаются
1) гетеротрофные организмы
2) аэробные организмы
3) автотрофные организмы
4) паразитические формы
А11. Наиболее сложно устроена
амеба обыкновенная 3) малярийный плазмодий
эвглена зеленая 4) инфузория-туфелька
А12. При похолодании, других неблагоприятных условиях свободно живущие простейшие
1) образуют колонии 3) образуют споры
2) активно двигаются 4) образуют цисты
Часть В
В1. Выберите простейших, ведущих свободный образ жизни
Выберите простейших, ведущих свободный образ жизни
1) инфузория стентор 4) лямблия
2) амеба протей 5) стилонихия
3) трипаносома 6) балантидий
В2. Соотнесите представителя простейших с признаком, который у него есть
Часть С
С1. Почему аквариумисты выращивают культуру инфузорий на молоке?
С2. Найдите ошибки в приведенном тексте, исправьте их, укажите номера предложений, в которых они сделаны. 1. Простейшие (одноклеточные) организмы обитают только в пресных водах. 2. Клетка простейших – это самостоятельный организм, со всеми функциями живой системы. 3. В отличие от клеток многоклеточных организмов клетки всех простейших имеют одинаковую форму. 4. Простейшие питаются частицами твердой пищи, бактериями. 5. Непереваренные остатки пищи удаляются через сократительные вакуоли. 6. Некоторые простейшие имеют хроматофоры, содержащие хлорофилл, и способны к фотосинтезу.
Данный текст является ознакомительным фрагментом.
Читать книгу целиком
Поделитесь на страничкеСледующая глава >
Жгутиконосцы, инфузории. Болезни рыб, вызываемые простейшими
- ГДЗ
- 1 Класс
- Окружающий мир
- 2 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Литература
- Окружающий мир
- 3 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Окружающий мир
- 4 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Окружающий мир
- 5 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Биология
- История
- География
- Литература
простейших — Викивиды
| Этот таксон является (или может быть) парафилетическим |
Таксонавигация [править]
Superregnum: Eukaryota
Regnum: Protozoa
Тип: Euglenozoa — Kinetoplastida — Rhizopoda — Microspora — Protozoa incertae sedis
Protozoa Goldfuss, 1817 г.
Народные имена [править]
Alemannisch: Protozoen
azərbaycanca: Protozoylar
беларуская: Прасцейшыя
català: Protozou
čeština: Prvoci
Prvoci
Urthiere (устаревшее написание)
español: Protozoo
français: Protozoaire
hrvatski: Praživotinje
magyar: Protozoa
interlingua: Protozoa
0 Protozoa0 Protozoa0 Limburgs: Protozoa
Plattdüütsch: Protozoa
norsk nynorsk: Protozo
norsk: Protozo
polski: Pierwotniaki
Protozoa0 Protozoa0: 9rio 170009: 9rio 170009: 9 Английский:
Protozoa
slovenčina: Prvoky
svenska: Urdjur
தமிழ்: முன்னுயிரிகள் (புரோட்டோசூவா)
Ссылки [править]
Goldfuss, Georg August: Ueber die Entwicklungsstufen des Thieres , Нюрнберг, Леонард Шраг (1817) [1].
Дополнительные ссылки [править]
- Беккер, Э. Р. и Кюн Л. Б. 1939: Роды простейших . 65 с. (Мимеография), [2].
- Mauch, E., Schmedtje, U., Maetze, A. & Fischer, F. 2003: Taxaliste der Gewässerorganismen Deutschlands zur Kodierung biologischer Befunde , [3]. (Со списком определяющей литературы с комментариями.)
См. Также [править]
Альтернативные классификации [править]
[Примечание: в большинстве классификаций Protozoa в настоящее время считается полифилетическим таксоном.Некоторые авторы (например, Кавалье-Смит) все еще используют его как парафилетический таксон, хотя большинство систематиков предпочитают использовать только монофилетические таксоны. См. Более современную классификацию ранее размещенных здесь групп в Eukaryota.]
Гольдфус (1820) [править]
Из Handbuch der Zoologie .
Класс Простейшие
фон Зибольд (1848) [править]
Lehrbuch der vergleichenden .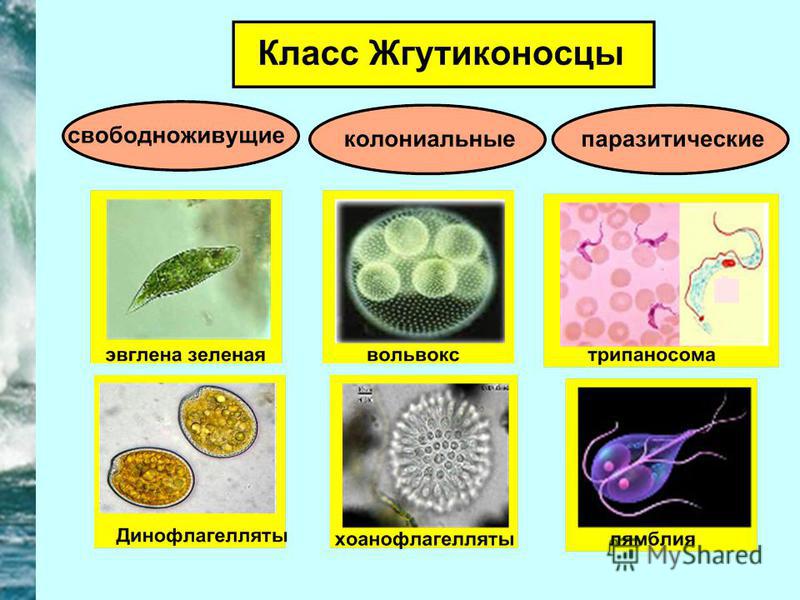 Vol. 1, Wirbellose Thiere, von C. Th. против Зибольда, 1848 г.Vol. 2, Wirbelthiere, von H. Stanius, 1846, [4].
Vol. 1, Wirbellose Thiere, von C. Th. против Зибольда, 1848 г.Vol. 2, Wirbelthiere, von H. Stanius, 1846, [4].
Тип Простейшие
Оуэн (1860) [править]
Из Палеонтология или систематический обзор вымерших животных и их геологических отношений .
Королевство Простейшие
Hogg (1860) [править]
Хогг, Дж. (1860 г.). О различиях растений и животных и о четвертом царстве природы . Эдинбург Нью Фил. J. , Н.С. 12: 216–225 [5].
- Минеральное царство
- Растительное царство
- Царство животных
- Первобытное королевство (Regnum Primigenum)
Haeckel (1874) [править]
Из Геккеля Э (1874 г.). Die Gastrea-Theorie, die phylogenetische Classification des Tierreiches und die Homologie der Keimblatter. Jenaische Zeitschrift fur Naturwissenschaft , 8, 1–55, [6].
См. См. Kluge (2004-2014), [7].
[стр. 32:]
1. Абластерия
Абластерия Простейшие Простейшие Простейшие 2. Дибластерия Gastraeada Zoophyta Metazoa Spongiae 3. Трибластерия Acalephae 4. Тетрабластерия Акоеломи Vermes 5. Пентабластерия Coelomati Mollusca Typozoa Иглокожие Членистоногие Позвоночные
[П.33:]
- Bilateria = Sphenota
- Vermes I + Vermes II [Sammtlische Würmer]
- Mollusca [Molluscken]
- Иглокожие [Echinotermen]
- Членистоногие [Arthropoden]
- Vertebrata [Vertebraten]
[P. 52:]
- Thierreichs
- Простейшие
- Metazoa [= Blastozoa, p.
 19]
19]- Anaemaria
- Haemataria = Pentablasteria
Lankester (1877) [править]
Ланкестер, Э.R. 1877. Заметки по эмбриологии и классификации животного мира: включает пересмотр предположений о происхождении и значении зародышевых листков. Ежеквартальный журнал микроскопических исследований , 68: 399-454, [8].
Сэвилл-Кент (1880-1882) [править]
Из Сэвилл-Кент, W. (1880–1882). Инструкция Инфузории . Лондон: Д. Бог, 3 тома, [9].
Под-царство Protozoa [стр. 36]
Простейшие царства [стр. 211]
Bütschli (1880-1889) [править]
От Bütschli, O.(1880–1889). Классификация и порядок вещей доктора Х. Г. Бронна: wissenschaftlich dargestellt in Wort und Bild: wissenschaftlich dargestellt in Wort und Bild. Кольцо Эрстера: простейшие. Abt. I, Sarkodina und Sporozoa, 1880-82 [10]. Abt. II, Мастигофора, 1883–87 [11]. Abt. III, Infusoria und System der
Radiolaria, 1887-89 [12]. К.Ф. Зима, Лейпциг.
К.Ф. Зима, Лейпциг.
Тип Простейшие
Ланкестер (1885, 1891) [править]
Ланкестер, Э. Р. (1885). Статья «Простейшие».В: Британская энциклопедия (9-е изд.), 19: 830–866,
[13].
Ланкестер, Э. Р. (1891). Статья «Простейшие». В: Зоологические статьи, внесенные в Британскую энциклопедию , [14].
Блохманн (1886) [править]
Die mikroskopische Tierwelt des Süßwassers , 1-е изд., 1886, [15].
Простейшие
Геккель (1889 г.) [править]
Haeckel, E. 1889. Natürliche Schöpfungsgeschichte , 8-е изд, [16].См. Стр. 452, 453, 462, 508.
- Protistenreich
- Protophyta (Protista Vegetalia)
- Класс I. Фитарча
- Фитомонера
- Chromaceae
- Ordnung 2. Chroococceae
- Ordnung 3. Nostochineae
- Класс II. Диатомеи
- Класс III. Космарии (Desmidieae)
- Класс IV. Пальмеллярии (Palmellaceae)
- Класс V.
 Siphoneae (Coeloblastae)
Siphoneae (Coeloblastae)
- Простейшие (Protista animalia)
- Зоарча (Zoocytoda)
- Zoomonera
- Бактерии
- Сфаэробактерии
- Рабдобактерии
- Cytarcha
- Лобоса (Амебина)
- Gregarinae (Sporozoa)
- Инфузории
- Flagellata (Мастигофора)
- Реснички
- Acinetae (Suctoria)
- Rhizopoda
- Mycetozoa (Myxomycetes)
- Heliozoa
- Таламария (Foraminifera)
- Радиолярия
- Pflanzenreich
- Thallota (Thallophyta)
- Водоросли (Phycophyta)
- Zygnemaceae
- Conferveae
- Fucoideae
- Флориды
- Characeae
- Грибы (Inophyta)
- Prothallota (Мезофита)
- Phanerogamae (Anthophyta)
- Gymnospermae
- Cycadeae
- Хвойные
- Gnetaceae
- покрытосемянных
- Thierreich
- кишечнополостные (Coelenterata, Zoophyta, Anaemaria)
- Gastraeades
- Губки
- Книдария
- Платоды
- Целомария (Bilaterata, Bilateria, Haemataria)
- Гельминты
- Моллюска
- Иглокожие
- Сочленений
- Туниката
- Позвоночные
Клаус (1891) [править]
Из Lehrbuch der zoologie .
Простейшие
Геккель (1894) [править]
Из Systematische phylogenie , 3 тома (том 1, Protisten und Pflanzen, 1894; том 2, Invertebrata, 1896; том 3, Vertebrata, 1895). [17].
[Том. 1, стр. 90-91:]
Пфланцен
- Protophyta (плазмодома Protista) [Vol. 1, стр. 96]
- Archephyta (Прогонелла)
- Пробионты
- Хромовые (Phycochromaceae)
- Алгарии (Paulosporata)
- Paulotomeae
- Конъюгаты
- Диатомеи
- Альгетты (Zoosporata)
- Мастигота
- Мелеталия
- Siphoneae
- Metaphyta (гистоны плазмодомии) [Vol.1, стр. 256]
- Thallophyta
- Водоросли [Vol. 1, стр. 302]
- Chlorophyceae
- Charaphyceae
- Phaeophyceae
- Rhodophyceae
- Mycetes [Vol. 1, стр. 316]
- Аскомицеты (Ascodiomycetes)
- Basimycetes (Базидиомицеты)
- Лихены [Vol. 1, стр. 325]
- Diaphyta [Vol. 1, стр. 330]
- Bryophyta [Vol. 1, стр. 336]
- Pteridophyta
- Anthophyta
- Gymnospermae
- Покрытосемянное
Thieren
- Protozoa (Protista plasmophaga) [Vol.1, стр. 138]
- Архезоа (= Зоарчега)
- Бактерии (= Bactromonera)
- Coccillida
- Bacillida
- Спириллида
- Зоомонера (= ул. Монера)
- Грибки (= Sporozoa)
- Ризопода
- Инфузории
- Metazoa (Гистоны плазмофаги)
- кишечник (акеломия)
- Gastraeades
- Губки
- Книдария
- Платоды
- Билатерия (Coelomaria)
- Гельминты
- Моллюска
- Сочленений
- Иглокожие
- Туниката
- Vertebrata
Levander (1894) [править]
Левандер, К.М. (1894). Materialien zur Kenntnis der Wasserfauna in der Umgebung Helsingfors, mit besonderer Berücksichtigung der Meeresfauna. I: простейшие. Acta Societatis pro Fauna et Flora Fennica 12 (2): 3–155, 3 табл., 1 таблица, [18].
Простейшие
Блохманн (1895) [править]
Блохманн, Ф. (1895). Die mikroskopische Tierwelt des Süßwassers. Abt. Я, простейшие. В: Кирхнер О. и Блохманн Г., Die mikroskopische Pflanzen- und Tierwelt des Süßwassers .2. Aufl., Teil II. Гамбург: Gräfe & Sillem, [19].
Простейшие
Delage & Hérouard (1896) [редактировать]
Traité de zoologie concrète , t. 1, [20].
Kalberlah & Schoenichen (1900) [править]
Kalberlah, A. & Schoenichen, W. B. (1900). Эйферта Einfachste Lebensformen des Tier- und Pflanzenreiches: Naturgeschichte der mikroskopischen Süsswasserbewohner . 3-е изд. Брауншвейг: Б. Гериц, [21].
- Pflanzliche Organismen
- Tierische Organismen
Дофлейн (1901) [править]
Дофлейн, Ф.(1901). Die Protozoen als Parasiten und Krankheitserreger , 1-е изд. Дополнительные издания: Lehrbuch der Protozoenkunde .
Дофлейн (1902) [править]
Дофлейн, Ф. (1902). Das System der Protozoen. Archiv für Protistenkunde 1: 169–192, [22].
Lankester et al. (1903, 1909) [править]
Ланкестер, Э. Р. (ред.) Трактат по зоологии , т. 1, фас. 1 (1909), фас. 2 (1903), [23].
Простейшие
Хартог (1906) [править]
Хартог, М.(1906). Простейшие. В: Кембриджская естественная история . Vol. 1, Protozoa, Porifera, Coelenterata и Echinodermata. (ред. С. Ф. Хармера и А. Э. Шипли). Лондон: Macmillan, [24].
Простейшие
Дофлейн (1911) [править]
Из Lehrbuch der Protozoenkunde , 3-е изд., 1911 г. (1-е изд. Под названием Die Protozoen als Parasiten und Krankheitserreger , 1901).
Простейшие
Минчин (1912) [править]
Введение в изучение простейших со специальной ссылкой на паразитические формы , 1-е изд., [25].
Простейшие
Люэ (1913) [редактировать]
Из Люэ, М. 1913: Erstes Urreich der Tiere. В: Lang A, редактор. Handbuch der morphologie der wirbellosen Tiere. Йена: Г. Фишер. 416 с.
Простейшие
Поче (1913) [править]
Poche, F. 1913. Das System der Protozoa. Archiv für Protistenkunde 30: 125–321, [26].
Phylum Protozoa Goldfuss, 1817 г.
Hartmann (1919) [редактировать]
От Хартманна, М.»Allgemeine Morphologie und Physiologie der Protozoen». В: Lehrbuch der Mikrobiologie . Фридбергер, Э; Pfeiffer, R. [ред.] G. Fischer, 1919, [27], [28].
Простейшие
Calkins (1926) [править]
Из Биология простейших .
Тип Простейшие
Веньон (1926) [править]
Веньон, К. М. 1926. Протозоология, руководство для врачей, ветеринаров и зоологов . Байер, Тиндаль и Кокс: Лондон; Нью-Йорк: W.Дерево. Тт. 1 и 2, [29].
Тип простейших
Sandon (1927) [редактировать]
Сандон, Х., 1927. Состав и распространение фауны простейших в почве . Лондон: Оливер и Бойд, [30].
Простейшие
Уэйлс (1937-1943) [править]
Уэйлс, Г. (1937). Тихоокеанская фауна Канады: 1. Простейшие. 1а. Лобоса; lb. Reticulosa; 1c. Heliozoa; 1г. Радиолярии. с. 1-14. Торонто: Univ. Toronto Press, Совет по исследованиям рыболовства Канады.
Wailes, G.H. (1939). Тихоокеанская фауна Канады. 1. Простейшие. 1e. Мастигофора. pp. 1-45, Fig. 138. Toronto: Univ. Toronto Press, Совет по исследованиям рыболовства Канады.
Wailes, G.H. (1943). Тихоокеанская фауна Канады: 1. Простейшие. 1f. Реснички; 1гр. Суктория. с. 1-46. Торонто: Univ. Toronto Press, Совет по исследованиям рыболовства Канады.
Простейшие
Баркли (1949) [править]
Баркли, Фред А. «Un esbozo de clasificación de los organos.» Revista de la Facultad Nacional de Agronomia , Universidad de Antioquia, Медельин. 10: 83–103, [31].
- Mychota (Monera, Anucleobionta)
- Протиста
- Euthallophyta
- Родофита
- Хризофита
- Pyrrophyta
- Euglenophyta
- Gonidiophyta
- Charophyta
- Phaeophyta
- Myxophyta
- Микофита
- Простейшие
- Sarcodina
- Фораминиферы
- Актиноподы
- Грибки
- Реснички
- Suctoria
- Мезозоа
- Паразоа
- Euplanta
- Euanimalia
Jahn & Jahn (1949) [править]
От Яна, Т.Л. и Ф. Ф. Ян, 1949. Как узнать простейшие . Браун, Dubuque, lowa.
Grassé et al. (1952-1994) [править]
Из Грассе, П.-П. (Ред.), 1952–1994. Traité de Zoologie . Vol. 1, пт. I, Protozoaires: Flagellés , 1952. Vol. 1, пт. II, Простейшие: Rhizopodes, Actinopodes, Sporozoaires, Cnidosporidies , 1953. Vol. 2, пт. I, Infusoires ciliés , 1984. Vol. 2, пт. II, Infusoires ciliés , 1994. Masson, Paris.См. Фернандес-Галиано (1964), [32].
Règne animal
Холл (1953) [править]
Из Холла, Р.П., 1953. Протозоология . Прентис-Холл, Нью-Йорк, [33]
Простейшие
Pennak (1953) [редактировать]
Пеннак Р.В. 1953. Пресноводные беспозвоночные США . 1-е изд. Рональд, Нью-Йорк, [34].
Тип простейших
Кудо (1954) [редактировать]
Из Протозоология , 4-е изд.
Тип Protozoa Goldfuss
Эдмондсон (1959) [править]
Эдмондсон, W.T. (ред.). 1959. Пресноводная биология . 2-е изд. Вили, Нью-Йорк, [35].
Протиста
Protozoa Images, Stock Photo & Vectors
В настоящее время вы используете более старую версию браузера, и ваша работа может быть не оптимальной. Пожалуйста, подумайте об обновлении. Учить больше. ImagesImages homeCurated collectionsPhotosVectorsOffset ImagesCategoriesAbstractAnimals / WildlifeThe ArtsBackgrounds / TexturesBeauty / FashionBuildings / LandmarksBusiness / FinanceCelebritiesEditorialEducationFood и DrinkHealthcare / MedicalHolidaysIllustrations / Clip-ArtIndustrialInteriorsMiscellaneousNatureObjectsParks / OutdoorPeopleReligionScienceSigns / SymbolsSports / RecreationTechnologyTransportationVectorsVintageAll categoriesFootageFootage homeCurated collectionsShutterstock SelectShutterstock ElementsCategoriesAnimals / WildlifeBuildings / LandmarksBackgrounds / TexturesBusiness / FinanceEducationFood и DrinkHealth CareHolidaysObjectsIndustrialArtNaturePeopleReligionScienceTechnologySigns / SymbolsSports / RecreationTransportationEditorialAll categoriesEditorialEditorial главнаяРазвлеченияНовостиРоялтиСпортМузыкаМузыка домойПремиумBeatИнструментыShutterstock EditorМобильные приложенияПлагиныИзменение размера изображенияКонвертер файловСоздатель коллажейЦветовые схемыБлогГлавная страница блогаДизайнВидеоКонтроллерНовости
PremiumBeat blogEnterpriseЦена ing Войти
Зарегистрироваться
Меню
ФильтрыОчистить всеВсе изображения- Все изображения
- Фото
- Векторы
- Иллюстрации
- Редакционные
- Музыка
- Видеоматериалы
8
- Поиск по изображению
90azo Самые актуальные
Свежий контент
Тип изображения Все изображения
Фото
Векторы
Иллюстрации
Ориентация Все ориентации
Горизонтально
Определение, типы, болезни, профилактика и многое другое
Патогены — это организмы, которые могут вызывать болезнь.Различные типы патогенов и тяжесть вызываемых ими заболеваний очень разнообразны.
В этой статье мы рассмотрим различные патогены, их влияние на людей и болезни, которые они вызывают. Мы также объясняем, как распространяются патогены и как снизить риск заражения.
Патоген приносит болезнь своему хозяину. Другое название возбудителя — инфекционный агент, поскольку они вызывают инфекции. Как и в случае с любым организмом, патогены отдают приоритет выживанию и воспроизводству.
Иммунная система человеческого организма действует как защита от патогенов.Организм может легко бороться с некоторыми патогенами, но другие потенциально смертельны.
Существует пять основных типов патогенов:
Бактерии
Бактерии — это микроскопические патогены, которые быстро размножаются после попадания в организм. Они могут выделять токсины, которые повреждают ткани и вызывают болезни.
Врачи обычно назначают антибиотики для лечения бактериальных инфекций, но некоторые бактерии становятся устойчивыми к этим лекарствам.
Однако не все бактерии являются патогенными.В организме существует множество типов безвредных бактерий, некоторые из которых могут даже поддерживать основные функции организма.
Вирусы
Вирус, который меньше бактерий, проникает в клетку-хозяина. Затем он реплицируется, производя сотни и тысячи новых вирусов, которые продолжают заражать другие клетки-хозяева. Вирусы могут передаваться от человека к человеку различными путями, в том числе:
- через респираторные капли, которые перемещаются по воздуху
- через контакт с кровью человека с инфекцией
- через контакт с жидкостями организма человека, инфицированного инфекцией
Грибы
Существуют тысячи видов грибов, некоторые из которых вызывают болезни у людей.Распространенные грибковые заболевания кожи включают микоз стопы и стригущий лишай. Эти состояния заразны и могут передаваться от человека к человеку.
Исследование, проведенное в Trends in Microbiology , показало, что у грибковых патогенов развивается способность к памяти. Они могут использовать сигналы в теле, чтобы предвидеть неминуемую угрозу своему выживанию, к которой они затем могут подготовиться.
Протисты
Эти одноклеточные организмы вызывают болезни у своего хозяина. Они заражают другие организмы, чтобы выжить и размножаться.
Простейшие патогены поражают растения и продовольственные культуры. Продукты, содержащие простейшие, могут вызвать дизентерию — инфекцию кишечника, вызывающую диарею.
Протистские патогены также могут быть паразитическими и жить в других организмах, таких как комары. Протисты вызывают малярию через укусы комаров.
Паразитические черви
Паразитические черви, также известные как гельминты, достаточно велики, чтобы их можно было увидеть невооруженным глазом, и они могут жить во многих частях тела.Некоторые черви включают:
- Плоские черви: К ним относятся ленточные черви, которые обитают в кишечнике.
- Колючие черви: Этот вид червей обитает в кишечнике.
- Круглые черви: Эти черви могут выжить в желудочно-кишечном тракте и лимфатической системе.
Патогены могут вызывать множество различных заболеваний, некоторые из которых более серьезны, чем другие. Человеческие тела богаты питательными веществами и могут обеспечить патогену идеальную среду для роста и размножения.
Степень тяжести инфекций, вызываемых патогенами, может быть разной. Некоторые инфекции могут быть легкими, в то время как другие могут быть опасными для жизни. Например, простуда — это легкая вирусная инфекция по сравнению со смертельной болезнью, вызванной вирусом Эбола.
Болезни, вызываемые бактериальными патогенами, включают:
Некоторые ученые считают, что вирусы не являются живыми организмами. Вот некоторые причины:
- у них нет клеток
- они не могут воспроизводиться, не вторгаясь в живую клетку
- они не реагируют активно на изменения в окружающей среде
К вирусным инфекциям относятся:
Около 300 видов грибов патогенны для человека.Как и бактерии и вирусы, они могут оказывать значительное влияние на здоровье человека.
Грибки вызывают множество различных типов заболеваний, в том числе:
- астму
- инфекции кожи и ногтей
- инфекции легких, такие как пневмония
- инфекции кровотока
- менингит
Протозойные заболевания являются причиной большинства протистических заболеваний. Простейшие — это одноклеточные микроорганизмы, которые питаются другими микроорганизмами, органическими тканями и мусором.К протистским заболеваниям относятся:
- дизентерия
- малярия
- Африканский трипаносомоз или сонная болезнь
Паразитические черви вызывают множество болезней, в том числе:
Патогены могут распространяться различными путями. Например, прямой контакт кожа к коже во время секса может привести к инфекциям, передаваемым половым путем (ИППП).
При кашле или чихании болезнетворные микроорганизмы могут распространяться через крошечные капельки в воздухе. Эти капли могут содержать микроорганизмы, которыми дышат другие люди.
Микроорганизмы также могут попадать прямо в кишечник, когда человек употребляет зараженную пищу или воду.
Укусы инфицированных насекомых также могут распространять болезнь. Например, клещи с бактериальной инфекцией могут вызвать болезнь Лайма, если кого-то укусили, а комары с вирусной инфекцией могут вызвать болезнь, вызванную вирусом Зика.
Наряду с поддержанием хорошего общего состояния здоровья человек может предпринять определенные шаги, чтобы снизить риск заражения патогенами. К ним относятся:
- Регулярное мытье и сушка рук: Регулярное мытье рук имеет решающее значение для предотвращения распространения болезней.Люди должны мыть их не менее 20 секунд водой с мылом или дезинфицирующим средством на спиртовой основе. Это особенно важно после занятий на открытом воздухе, прикосновений к животным, посещения туалета, ухода за больными, чихания или кашля.
- Будьте в курсе рекомендованных вакцин: Вакцины против инфекционных заболеваний, таких как грипп, коклюш и корь, доступны для младенцев, детей и взрослых. Вакцины стимулируют иммунную систему распознавать патогены, уничтожать их и предотвращать будущие инфекции.
- Поддержание чистоты поверхностей: Загрязненные поверхности или предметы могут передавать болезни. Исследование, проведенное в аэропорту Финляндии, обнаружило вирусное заражение в нескольких местах на поверхностях, к которым люди часто прикасаются.
- Уборка кухонь и ванных комнат: Важно поддерживать в чистоте столешницы и кухонные поверхности, особенно перед приготовлением еды. В ванной также может быть высокая концентрация инфекционных агентов.
- Оставаться дома в случае болезни: Люди с инфекциями должны избегать посещения школы, работы и тесного контакта с другими людьми, чтобы снизить вероятность распространения патогена.
- Как избежать укусов насекомых: Люди могут сделать это, надев защитные средства от насекомых и подходящую одежду, такую как рубашки с длинными рукавами, брюки и шляпы, в местах, где они могут встретить насекомых.
- Практика безопасного секса: Использование методов безопасного секса, включая использование презервативов и регулярные проверки сексуального здоровья, может снизить вероятность заражения ИППП.
- Получение медицинской консультации: Человек может поговорить с медицинским работником, если есть признаки инфекции или лечение не работает.
Патогены поражают все живые организмы и вызывают заболевания у людей множеством различных способов.
Они могут повредить ткани или клетки, вырабатывая токсины в процессе репликации.
В то время как некоторые патогены вызывают легкие проблемы, другие могут быть опасными для жизни. Медицинские эксперты продолжают изучать болезни, вызываемые патогенами, чтобы больше узнать об их причинах и способах их лечения.
Паразиты — простейшие
Простейшие теперь имеют свой собственный порядок в живом царстве, хотя когда-то их считали крошечными животными, поскольку они были подвижными и не были зелеными, как растения — теперь у них есть отдельное царство в соответствии со схемой классификации для всей жизни на Земле.Многие простейшие паразиты на человеке и вызывают от легких до чрезвычайно опасных заболеваний — некоторые из них паразитируют на человеке и из-за своего крошечного размера образуют простейших из всех внутренних паразитов. Клеточная организация простейших намного сложнее, а размер намного больше, хотя, как и бактерии, все простейшие являются одноклеточными организмами — весь организм состоит только из одной клетки. Тип окружающей среды, в которой обитает большинство простейших, очень космополитичен, и их распространение повсеместно, они встречаются почти в любой водной среде и во влажных местах, включая пруды, реки, озера, аквариумы, канавы и т. Д.
Мазь для ногтей
Лучшее, 100% натуральное средство для ежедневного ухода за ногтями в идеальной форме.
Большинство простейших не причиняют вреда животным или людям и считаются свободноживущими, то есть им не требуется хозяин для жизни и размножения. Однако есть несколько простейших, которые являются паразитами, среди этих паразитических организмов некоторые могут вызвать чрезвычайно серьезные проблемы со здоровьем у людей, которым не повезло заразиться простейшими.Первым патогенным простейшим, идентифицированным еще в 1875 году, было простейшее. Паразитические простейшие были разновидностью амебы под названием Entamoeba histolytica, которая также известна как дизентерия, вызывающая амебу — она очень распространена и поражает людей во всем мире. Антон ван Левенгук, один из первых и, возможно, один из величайших ранних микроскопистов и клеточных биологов, заметил, что в его собственных фекалиях скользят простейшие. Крошечная микроскопическая форма жизни принадлежала к роду Giardia, и с момента открытия Левенгука такие паразиты, как Giardia lamblia и простейшие виды Cryptosporidium, были связаны с причиной болезненных кишечных инфекций и вспышек тяжелой диареи во всем мире.Вода, загрязненная фекалиями человека или животных, обычно содержит этих двух паразитических простейших, и употребление такой загрязненной воды является обычной причиной заболеваний.
Мазь от грибка
Вся сила фармацевтических фунгицидов — но без агрессивных химикатов.
Бабезиоз
Паразитические простейшие, известные как Babesia microti, ответственны за вспышки этого заболевания. Паразит проникает в эритроциты различных животных, которых заражает, и вызывает типичные симптомы, столь характерные для этого состояния.Основным резервуаром болезни являются грызуны, способ передачи болезни — через клещей, возможно, тех же видов, которые участвуют в передаче болезни Лайма — таким образом, клещи являются основными переносчиками болезни и распространяют ее. от грызунов-хозяев к другим животным, включая человека, путем укуса. Распространенное заболевание, называемое техасской лихорадкой, иногда также известное как клещевая лихорадка крупного рогатого скота, которое поражает крупный рогатый скот, вызывается простейшими, связанными с этим видом.
Состояние вызывает симптомы у пораженных людей, все эти симптомы, по-видимому, связаны или похожи на симптомы, наблюдаемые у пациентов, страдающих малярией, которая также вызывается простейшими.Физические симптомы включают стойкую усталость и физическую усталость в течение семи дней после первоначального контакта с паразитом, за этим начальным периодом следует период, когда у пациента появляется внезапный озноб, а также другие состояния, такие как лихорадка, сильная головная боль и болезненность мышц в целом. тело. Некоторые больные часто прогрессируют до анемии и почечной недостаточности, хотя большинство симптомов легкие, и состояние в основном проходит через несколько недель. Однако известны случаи смертельного исхода в некоторых случаях, особенно в отношении пациентов старших возрастных групп.Для этого состояния не существует специальных лекарств, хотя симптоматическое лечение с использованием тех же лекарств, которые используются для лечения малярии, было опробовано с некоторым успехом — профилактика является лучшей политикой в отношении этого заболевания, и случаи заболевания редки.
Болезнь Шагаса
Простейшее заболевание Темы — болезнь Шагаса — серьезная проблема для здоровья в Латинской Америке. За эти состояния отвечает простейший организм, известный в науке как Trypanosoma cruzi, болезнь широко распространена в Центральной и Южной Америке, где она представляет собой проблему для общественного здравоохранения.Переносчиком этой болезни является насекомое, известное как целующиеся клопы, иногда называемое «конусообразным насекомым». Люди, живущие в районах с маргинальной жилой инфраструктурой, зараженной этим насекомым, могут заразиться семейством болезни Шагаса. Насекомые-переносчики относятся к семейству насекомых Reduviidae, подсемейству Triatominae и роду Triatoma — это насекомое снова очень распространено в Южной Америке. Опоссумы и броненосцы, а также инфицированные люди служат резервуарами для простейших, насекомое, которое является кровососом, передает паразита из таких источников неинфицированным животным и людям — инфекции обычно распространяются среди людей в одной семье — насекомое передает паразита от хозяин за хозяином, поскольку он сосет кровь и оставляет отходы на новых хозяевах.
Восстанавливающее средство для кожи
Усовершенствованный, 100% натуральный ревитализатор, который сделает вашу кожу сияющей и молодой.
После того, как клоп поедает инфицированное млекопитающее, он передает простейшие, которые уже прошли по его кишечнику, с отходами, которые он оставляет на коже неинфицированного животного-хозяина или человека. Когда человек непроизвольно царапает эту область, отходы в области укусов насекомых загрязняют рану, и повторяющиеся царапины вызывают ушибы кожи — простейшие также часто передаются через глаза — когда человек трет отходы, простейшие попадают в тело, используя сломанный участок кожи.Тропическое заболевание было идентифицировано в 1912 году бразильским врачом Карлосом Шагасом — отсюда и название болезнь Шагаса.
Появление небольшой красной язвочки на коже или развитие опухших и покрасневших, воспаленных глаз являются наиболее частыми ранними симптомами заболевания у человека. Через несколько недель язва на пораженном участке кожи, кажется, заживает и выделяет твердую корку, которая вскоре исчезает, и остается небольшой темный шрам в этой области — если человеку повезет. В других случаях первоначальные симптомы могут сменяться появлением лихорадки, развитием отека лимфатических узлов и узлов, а также развитием кожной сыпи по всему телу.Тяжелые формы болезни Шагаса могут поражать сердце, и такие симптомы, как боли в груди и внезапная одышка, становятся обычным явлением. Также могут возникнуть судороги, если заболевание поражает нервную систему — в таких случаях в непрерывных конвульсиях может быть вовлечено все тело. В некоторых случаях это состояние может привести к летальному исходу — это остается риском для здоровья.
Лямблиоз (бобровая лихорадка)
Эта лихорадка является паразитарным заболеванием человека, которое является наиболее распространенной формой паразитарного заболевания в некоторых частях Соединенных Штатов и других районах Северной Америки, в частности, в западных регионах Канады.Причина лихорадки — крошечный паразитический простейший организм, который называется Giardia lamblia. Паразитический организм остается на внутренней поверхности кишечника человека и, в частности, в больших количествах обнаруживается в тонком кишечнике пораженных людей. Обычным физическим симптомом, помимо повышения температуры тела, является стойкая инфекционная диарея.
Обычный источник распространения паразита — употребление зараженной пищи и воды. Если не поддерживать строгие и надлежащие санитарные условия, многие детские центры и дома престарелых становятся районами, в которых часто происходят вспышки лямблиоза, поэтому дети становятся одними из первых, кто становится жертвами паразита.Путь передачи паразита в большинстве случаев заражения происходит через попадание паразитарных цист в пищеварительный тракт вместе с зараженной пищей или водой — такие источники обычно содержат фекальные отходы людей и других животных-переносчиков.
Примерно через три недели после первоначального заражения паразитом хроническая диарея начинает поражать людей, страдающих лямблиозом — это обычные временные рамки для появления этого симптома у большинства пациентов.В дополнение к этому симптому у пораженных людей также появляются симптомы тошноты и другие жалобы, такие как стойкий и сильный дискомфорт в животе. Пищеварительная система нарушается во время длительных периодов инфекции, конечным результатом может быть внезапная потеря веса у пораженного человека — это происходит из-за очень плохого всасывания переваренной пищи из области кишечника. Человек может оставаться инфицированным в течение нескольких лет подряд, хотя в некоторых случаях у пораженных людей начинается спонтанное выздоровление от симптомов.
Раньше считалось, что основным резервуаром простейших паразитов в дикой природе являются бобры — так считалось в течение многих лет, пока недавние доказательства не показали обратного. В настоящее время известно, что домашний скот и овцы являются основными резервуарами болезни, согласно данным недавних исследований. Сейчас считается, что бобры, как и люди, вероятно, могут быть вторичным хозяином, а не резервуаром для паразита. Было показано, что такие животные, как бобры, полевки, ондатры и крысы-кенгуру, обладают способностью переносить этого паразита — все они могут действовать как хозяева, похожие на людей.
Сонная болезнь (африканский трипаносомоз)
Мелкая рыба, такая как простейшие паразиты, вызывает опасное заболевание, известное как сонная болезнь — это смертельное заболевание, широко распространенное в некоторых районах Африки. Простейшие — это трипаносома — разновидность нематодных червей, этот червь может плавать в крови своих жертв, ударяя хлыстом, как хвостом, или, точнее, своим жгутиком. Печально известная муха цеце несет ответственность за передачу этого смертельного заболевания. Муха кусает и сосет кровь, как только паразит попадает в организм.Человек медленно теряет сознание, и смерть наступает, если паразиту удается проникнуть в области спинного и головного мозга своих жертв. Шансы умереть от болезни очень высоки, если человек инфицирован, а сонная болезнь, как известно, уничтожает все домашние запасы мяса в районе — поскольку этот паразит также заражает домашний скот, например, крупный рогатый скот. Смертность как среди людей, так и среди животных высока в районах, где болезнь носит эндемический характер, и профилактика является лучшим вариантом против нее.Среда обитания мухи цеце широко распространена в тропической Африке, и некоторые ученые и эксперты в области здравоохранения считают, что с повышением температуры из-за глобального потепления существует возможность значительного расширения этой среды обитания. В таком случае муха цеце, как и паразит, может заразить космополитичную территорию. Это не единственный наихудший сценарий для одного насекомого-переносчика; Обычно считается, что глобальное потепление может расширить ареал обитания других насекомых-переносчиков, а также привести к распространению большего количества тропических паразитов в другие регионы мира.
Лейшманиоз
Паразитические простейшие лейшмании ответственны за это изнурительное и часто смертельное заболевание, которое широко распространено в некоторых тропических и субтропических регионах мира. Крошечная кровососущая песчаная муха ответственна за распространение этого смертоносного паразитического простейшего — эта муха является единственным переносчиком паразита. Состояние, известное как висцеральный лейшманиоз, иначе известное как кала-азар, является наиболее летальной стадией прогрессирования инфекции. В эндемичных районах болезнь способна уничтожить две трети населения некоторых серьезно пострадавших деревень — это особенно верно в отношении сельских районов в тропиках, где медицинская помощь плохая или полностью недоступна.Постоянная лихорадка, темно-серая кожа, наличие тяжелой анемии — одни из первых физических симптомов кала-азар, кроме того, сильно увеличиваются селезенка и печень, что приводит к неприглядному вздутию живота. область. Болезнь может стать смертельной очень быстро, если симптомы остаются без лечения в течение длительного периода времени. Кала-азар обычно лечится с помощью лекарств, содержащих сурьму тяжелых металлов — такие лекарства являются единственным эффективным средством против паразита.Белые кровяные тельца в организме — это область, в которой простейшие лейшмании фактически проживают в организме — это защитные клетки, которые должны были поглотить простейшие, когда началась инфекция. Белые кровяные тельца поглощают простейшее, но не могут его разрушить или переварить. В результате простейшие лейшмании процветают в тех самых белых кровяных тельцах, от которых зависит его собственная защита. Лейкоциты, в которых проживают простейшие, со временем разрушаются, подобно заболеванию, подобному ВИЧ, что, естественно, наносит огромный ущерб иммунной системе больного человека.Оппортунистические и смертельные бактериальные инфекции, такие как пневмония или дизентерия, преодолевают слабую и неэффективную защиту организма, так как лейкоциты в большом количестве умирают из-за простейших. Если это продолжается, человек умирает от симптомов.
Малярия
Настоящее значение слова «малярия» — просто «плохой воздух». Этот плохой воздух должен был вызывать у людей перемежающуюся лихорадку, когда они впервые столкнулись с этой болезнью. Действительно, древнегреческий врач Гиппократ предположил, что это было вызвано появлением миазмов на человеке — это смертельные туманы, которые предположительно нарушили тщательный баланс, существующий в четырех жидкостях тела, — что является древним медицинским верованием в кровь. мокрота, черная желчь и желтая желчь.С другой стороны, согласно древней китайской медицинской системе, дисбаланс между инь и ян в организме должен был вызвать приступообразную лихорадку, а именно малярию, а дисгармония между этими двумя противоположными мужскими и женскими жизненными силами в организме приписывалась быть причиной состояния. Несколько столетий назад великий систематик и биолог Карол Линней выдвинул гипотезу о том, что причиной малярии может быть присутствие крошечных взвешенных частиц глины в питьевой воде, эти взвешенные твердые частицы, как предполагается, привели к закупорке кровеносных сосудов и нарушение кровообращения должно было вызвать болезнь.Все это были теории о причине малярии, выдвинутые до появления микроскопа и в самом начале современной науки, истинный возбудитель малярии был идентифицирован только в 1898 году — британский доктор Росс правильно объяснил бы. Это произошло с укусом инфицированного комара, и он продолжил идентифицировать смертельный патоген как тип простейших.
Только укус самок комаров вида anopheles ответственен за передачу малярии людям и некоторым другим млекопитающим, которые могут быть инфицированы.Самки анофелесов — единственные москиты, которые являются переносчиками малярии человека; самцы комаров не кусаются и не могут распространять болезнь. Когда самка комара Anopheles кусает свою жертву, она высасывает кровь через прокол в коже особи и в то же время вводит тысячи нитевидных личинок простейших, называемых спорозоитами — эти личинки принадлежат к видам простейших плазмодиев. Оказавшись в теле жертвы, спорозоиты отправятся через кровоток прямо в печень, где каждая из личинок начинает инфицировать и жить в собственной клетке печени.В этих клетках печени спорозоиты собираются и начинают быстро делиться, многократно размножаясь, производя до сорока тысяч спор на каждую личинку. По прошествии двух недель производство и размножение спор в клетках печени продолжается, в течение этого периода нет абсолютно никаких признаков или симптомов болезни, и человек здоров. Однако здесь закладываются первые семена полномасштабной малярии, которые начинают вторжение в организм.
Разрыв клетки печени, в которой образовались споры, вызывает первые клинические признаки и является начальной стадией атаки на организм — у человека наблюдается сильное потоотделение и на этой стадии развивается высокая температура.Теперь лопнувшие клетки печени начинают выпускать многочисленные споры прямо в кровоток жертвы. На этом этапе каждая отдельная спора теперь вторгается в эритроцит, где она будет находиться какое-то время, медленно расходуя содержание гемоглобина в клетке и увеличиваясь в размере. Затем спора делится или раскалывается на множество более мелких фрагментов — это бесполое средство образования тысяч других спор — научное название спор на этой стадии — мерозоиты. Наконец, воля красных кровяных телец также лопается, и это высвобождает все присутствующие в них мерозоиты, а затем вторгаются в новые клетки крови по соседству.Этот циклический эффект, а также повторяющееся инфицирование и разрушение красных кровяных телец в конечном итоге приводят к хроническому малярийному заболеванию и могут привести к смерти человека.
Разрыв и вторжение красных кровяных телец — это очень своевременное дело, и существует удивительная синхронность всего процесса заражения в кровотоке жертвы. Основные причины одновременного разрыва миллионов инфицированных эритроцитов с высвобождением их мерозоитов до сих пор полностью не изучены и требуют изучения.Худшие приступы тряски и озноба происходят из-за своевременного и массового разрыва красных кровяных телец в организме.
Окончательный процесс трансформации этих мерозоитов в мужские или женские гаметоциты также начинает происходить после нескольких раундов инфицирования клеток крови и их последующего разрушения — эти гаметоциты являются репродуктивными клетками паразита плазмодия, а на этой стадии инфекции — человеческих клеток. фазу жизненного цикла паразита можно считать завершенной.Если пациенту повезет, он или она выжили бы на этой стадии, однако на этой стадии он заразен, потому что, если самка комара Anopheles высосет эту зараженную малярией кровь, она также поглотит миллионы паразитических клеток гаметоцитов. Гаметоциты простейших будут претерпевать дальнейшее развитие в яйцеклетки и сперматозоиды в теле самки комара, начиная следующую стадию у их нового хозяина, и оплодотворение этих гамет будет происходить в теле насекомого.
Цикл завершается, когда оплодотворенные яйца паразита превращаются в нитевидные спорозоиты в теле комара, а затем они попадают в слюнные железы комара, готовые для передачи новому человеку-хозяину.Именно в это время самка комара становится смертельным переносчиком малярийного паразита, и ее укус переносит новые спорозоиты, чтобы начать новый цикл внутри другого человека-хозяина.
Очень интересный факт заключается в том, что люди с серповидноклеточной анемией и даже те, кто страдает ее более легкой формой, или те, у кого есть черта серповидно-клеточной анемии в красных кровяных тельцах, вероятно, имеют гораздо большую защиту от малярии. предлагает. Серповидно-клеточная анемия деформирует красные кровяные тельца тела, и это изменение формы клеток напрямую препятствует и предотвращает проживание простейших внутри клетки.Это эволюционный ответ на малярию, поскольку люди, страдающие серповидно-клеточной анемией, как правило, в основном сконцентрированы в регионах мира с самым высоким уровнем заражения малярией — серповидно-клеточная форма красных кровяных телец является только общим генетическим дефектом в человеческих популяциях, круглый год подверженных малярийным заболеваниям.
Итак, как именно лечат малярию? И какое оружие применяется против болезни. Лекарства и противомалярийные препараты являются одними из инструментов, с помощью которых врачи и ученые непосредственно лечат малярию. Болезнь также можно контролировать, регулируя популяцию насекомых-переносчиков.Цитата «Лечите пациента, а не комара» часто использовала Роберт Кох, блестящий микробиолог, получивший известность благодаря выявлению туберкулеза. Единственным противомалярийным лекарством, которое было доступно в его дни — а это было в конце девятнадцатого века, было лекарство растительного происхождения, известное как хинин.
Противомалярийный препарат хинин — это медицинский препарат, полученный из коры хинного дерева, он веками использовался в традиционных противомалярийных средствах туземцами Южной Америки.На сегодняшний день он все еще используется как отличное противомалярийное лекарство во многих частях мира. К сожалению, высокие дозировки препарата токсичны и могут вызвать глухоту при использовании в высоких дозах, необходимых для полного удаления паразита плазмодия из организма. До получения менее вредных производных в 1930-х годах его использование было обширным и универсальным — теперь его заменяют менее вредные производные, синтезированные из того же растения. Хлорохин — это основное синтезируемое лекарство, используемое в наши дни для лечения малярии.Однако в качестве препарата выбора при лечении малярии хлорохин в большинстве стран мира был вытеснен другим противомалярийным мефлохином.
Другие полезные травы
Мазь для кожи
100% натуральная формула для всех проблем с кожей. Отлично подходит для диабетиков.
комментариев
Frontiers | Роль инфузорий простейших в рубце
Введение
Впервые описанные в 1843 году, простейшие Rumen с их поразительной внешностью считались важными для благополучия своего хозяина.Однако, несмотря на то, что простейшие могут составлять до 50% биомассы в рубце, роль простейших в микробной экосистеме рубца остается неясной (Williams and Coleman, 1992). Здесь мы оцениваем недавнюю информацию о роли простейших инфузорий в микробной экосистеме рубца.
Разнообразие и филогения простейших инфузорий рубца
С тех пор, как они были впервые обнаружены Gruby и Delafond (1843), исследования простейших в рубце основывались на морфологической идентификации с помощью оптической микроскопии.В настоящее время нет коллекций культур инфузорий рубца, поэтому исследователи должны использовать микрофотографии для идентификации (Ogimoto, Imai, 1981; Williams and Coleman, 1992) или штриховые рисунки (Dogiel, 1927; Kofoid and MacLennan, 1930, 1932, 1933; Latteur , 1968, 1969, 1970; Dehority, 1993). Широко признано, что микроскопическая идентификация и подсчет представляют собой золотой стандарт для анализа структуры сообщества инфузорий в исследованиях рубца (Williams and Coleman, 1992). Однако полиморфная природа этих микробов (Dehority, 1994, 2006) требует от исследователя высокого уровня опыта для идентификации инфузорий рубца на уровне родов или видов (Dehority, 2008).Таким образом, необходимы альтернативные способы идентификации простейших, поскольку микроскопические методы трудоемки и весьма требовательны.
Ранние молекулярные исследования простейших ресничек рубца были сосредоточены на секвенировании генов 18S рРНК для выяснения внутренней филогении класса Litostomatea (Embley et al., 1995; Wright et al., 1997). Embley et al. (1995) были первыми, кто описал монофилетическое положение анаэробных инфузорий рубца Dasytricha ruminantium и Polyplastron multivesiculatum на основе генов 18S рРНК, а Wright et al.(1997), Wright and Lynn (1997a, b) предоставили молекулярные данные для Diplodinium dentatum , Entodinium caudatum , Epidinium caudatum , Eudiplodinium maggii , Isotrichaintechinalis108 Oyinoscopusa , .
С тех пор были выполнены многочисленные качественные и количественные исследования разнообразия инфузорий; в этих исследованиях использовались различные методы, включая полиморфизм длины рестрикционных фрагментов (RFLP; Regensbogenova et al., 2004a, Tymensen et al., 2012b), денатурирующий градиентный гель-электрофорез (DGGE; Regensbogenova et al., 2004b, McEwan et al., 2005; de la Fuente et al., 2009; Belanche et al., 2010a, Kittelmann and Janssen, 2011), ПЦР в реальном времени (qPCR; Sylvester et al., 2004; Skillman et al., 2006; Belanche et al., 2010a, Kittelmann and Janssen, 2011), флуоресцентная гибридизация in situ (FISH; Xia et al. ., 2014) и секвенирование следующего поколения (Kittelmann et al., 2013, 2015; Ishaq, Wright, 2014; Moon-van der Staay et al., 2014).
Микроскопия имеет несколько преимуществ перед молекулярными методами на основе ПЦР для изучения простейших инфузорий. Во-первых, хотя подавляющее большинство кишечных инфузорий охарактеризовано морфологически, для многих наблюдаемых родов и видов отсутствуют эталонные последовательности гена 18S рРНК. Во-вторых, как обсуждается ниже, вариация числа копий генов рибосомной РНК в разных родах или в разных условиях роста может исказить наблюдаемые пропорции этих родов в образце (Medinger et al., 2010). Исследования с использованием обзоров генов 18S рРНК показывают очевидное более высокое разнообразие инфузорий, чем оценка с помощью обычных морфологических методов (Moon-van der Staay et al., 2014).
Как и другие инфузории, простейшие в рубце содержат два типа ядер: микроядро и макронуклеус. Микроядро имеет четко видимые хромосомы, диплоидно и синтезирует только следы РНК. Макронуклеус не содержит различимых хромосом, имеет во много раз больше диплоидного количества ДНК, делится амитотически и обеспечивает практически всю РНК, необходимую для вегетативной жизни клетки (Prescott and Murti, 1974).Чрезвычайно высокое число копий рДНК в макронуклеарном геноме инфузорий, как было обнаружено ранее (Gong et al., 2013), можно понять, учитывая жизненный цикл, большой размер клеток и быстрый рост этих организмов (Cavalier-Smith, 2005; Zhu et al., 2005; Godhe et al., 2008). В инфузориях рубца, в то время как небольшие простейшие, такие как Entodinium , имели тенденцию быть недостаточно представленными, более крупные простейшие, такие как Epidinium или Polyplastron , имели тенденцию быть чрезмерно представлены методом пиросеквенирования по сравнению с микроскопическим подсчетом (Kittelmann et al ., 2015). Аналогично, Epidinium caudatum , который примерно в пять раз больше по объему, чем Entodinium caudatum (Dehority, 1993), также имеет примерно в пять раз больше копий гена 18S рРНК, закодированных в его геноме (Sylvester et al., 2009). Эти результаты согласуются с выводом о том, что методы на основе ПЦР дают более низкие оценки численности мелких Entodinium spp. (Tymensen et al., 2012a) и завышенные оценки численности, например, Polyplastron spp.(Исхак и Райт, 2014). Знание числа копий и вариаций последовательностей рДНК в отдельных эукариотах важно как для интерпретации обзоров разнообразия на основе рДНК, так и при использовании 18s рДНК для количественной оценки биомассы простейших. (Crosby, Criddle, 2003; Thornhill et al., 2007; Herrera et al., 2009; Amaral-Zettler et al., 2011). На сегодняшний день не опубликовано исследований, посвященных количеству копий на клетку и вариациям рДНК у простейших в рубце. Это особенно верно, поскольку исследования штрих-кодирования и микробного разнообразия на основе рДНК с использованием высокопроизводительного секвенирования становятся все более популярными (Amaral-Zettler et al., 2009) и молекулярные инструменты, основанные на исследованиях маркерных генов, в настоящее время широко используются для изучения разнообразия других микробов (Schlötterer, 2004; Case et al., 2007; Langille et al., 2013). Исследования на инфузориях, не относящихся к рубцу, показали, что вариация числа копий рДНК между видами инфузорий и внутри них, подчеркивая сложность использования подхода на основе порядковых номеров рДНК для вывода об относительной численности микробных эукариотических клеток в образцах окружающей среды (Wintzingerode et al., 1997 ; Мединджер и др., 2010). Таким образом, новейшие методы, основанные на генах 18S рРНК, могут быть ненадежными при оценке α-разнообразия или относительной численности различных родов и видов в данной выборке, хотя они могут надежно определять тенденции относительной численности родов и видов между разными образцами (β- разнообразие). Необходимы дополнительные исследования, сравнивающие молекулярные и традиционные методы (de la Fuente et al., 2009; Tymensen et al., 2012b; Kittelmann et al., 2015).
Функциональные гены простейших инфузорий рубца
Хотя существуют многочисленные копии рДНК в макронуклеусах инфузорий, вполне вероятно, что только небольшая часть этих генов транскрипционно активна в инфузориях при любых данных условиях роста, как ранее было показано для др. Эукариот (Reeder, 1999).Исследования на Tetrahymena thermophila , Paramecium tetraurelia и Oxytricha trifallax выявили важную роль некодирующих РНК в событиях перестройки генома (Feng and Guang, 2013). У спиротрихозных инфузорий, таких как Euplotes , Stylonychia и Oxytricha , более 95% микроядерной ДНК удаляется с образованием макронуклеуса во время полового размножения (Swart et al., 2013), что является механизмом, позволяющим клетка адаптироваться во время стресса.Более того, в макронуклеусе оставшийся геном сильно фрагментирован, и эти фрагменты сортируются и переупорядочиваются под руководством транскриптов из родительского макронуклеуса с образованием генов, кодирующих белок (Nowacki et al., 2011). Ни один из этих процессов у простейших рубца до сих пор не изучен из-за трудностей получения полноразмерных геномов инфузорий рубца.
Несмотря на неоднократные попытки, оказалось невозможным поддерживать простейшие в рубце в аксенической культуре. Таким образом, было трудно окончательно установить роль простейших инфузорий в рубце и, в частности, разрушение волокон.Ранние исследования стремились выделить и охарактеризовать ферменты целлюлозы, гемицеллюлазы и ксиланазы из промытых препаратов простейших (Howard et al., 1960; Bailey et al., 1962; Clayet et al., 1992), однако присутствие как внеклеточных, так и внутриклеточных симбиотических бактерии в препаратах простейших затрудняли уверенность в том, что изолированная активность действительно имеет протозойное происхождение (Delfosse-Debusscher et al., 1979; Thines-Sempoux et al., 1980). Мы разработали методы клонирования и экспрессии генов инфузорий в фаге λ (Eschenlauer et al., 1998), используя FISH для подтверждения протозойной идентичности экспрессируемых генов (Newbold et al., 2005) и разрабатывая биоинформатические индексы для подтверждения инфузорийного происхождения генов (McEwan et al., 2000). Используя эти методы, мы и другие специалисты смогли выделить и охарактеризовать гены ряда простейших в рубце (McEwan et al., 1999; Newbold et al., 2005; Belzecki et al., 2007; Boxma et al., 2007). Сюда входит широкий спектр фибролитических ферментов, ряд из которых, как было обнаружено, содержит несколько доменов со связывающими доменами и предполагаемые химерные конструкции, которые наблюдаются, что свидетельствует о высокоразвитой фибролитической способности инфузорий рубца (Devillard et al., 1999, 2003; Takenaka et al., 1999, 2004; Wereszka et al., 2004; Bera-Maillet et al., 2005). Это наблюдение было подтверждено недавним метагеномным скринингом простейших глюкозидаз и эукариотических метатранскриптомов, который подтвердил, что в простейших рубца присутствует широкий спектр разнообразных гликозидгидролаз (Findley et al., 2011; Qi et al., 2011).
На основе крупномасштабного конструирования и анализа филогении более 4000 библиотек экспрессируемых последовательностей из инфузорий рубца Entodinium caudatum , Eudiplodinium maggii , Metadinium medium , Diploplastron affine 1010, ec Polyplastron affine 1010, ec , Isotricha prostoma , Isotricha Кишечник и Dasytricha ruminantium .Ricard et al. (2006) пришли к выводу, что имеются обширные доказательства горизонтального переноса генов (HGT; 148 из 3563 неизбыточных генов) от бактерий и архей в геномах ресничек рубца. Среди кандидатов на ГПГ они сообщили о чрезмерном представительстве (> 75%) генов, участвующих в метаболизме, в частности, в катаболизме сложных углеводов (Ricard et al., 2006), предполагая, что ГПГ мог играть важную роль в обеспечении инфузорий рубца. адаптироваться к новым нишам в рубце и что фибролитические гены были приобретены простейшими из бактериальных источников (Findley et al., 2011).
Роль простейших в рубце
Несмотря на то, что эти простейшие составляют значительную часть биомассы рубца, их роль в ферментации рубца и их вклад в метаболизм и питание хозяина по-прежнему вызывает серьезные споры (Williams and Coleman, 1992). В последнем разделе этой статьи мы рассмотрим различные стратегии управления плотностью простейших в рубце; однако большинство этих диетических вмешательств также приводят к изменениям функции рубца, что затрудняет оценку эффекта простейших как таковых рубца.Простейшие в рубце не необходимы животному для выживания и дефаунации (удаления простейших из рубца с использованием широкого спектра химических и физических методов), а животные, свободные от простейших, использовались для изучения роли инфузорий простейших в функции рубца без пострадали от диетических вмешательств (Williams and Coleman, 1992). Эта статья не ставит своей целью провести полный обзор последствий дефауна, но объединяет наиболее важные публикации со времен превосходного и полного обзора, проведенного Уильямсом и Коулманом (1992).Таким образом, был проведен метаанализ для изучения основных эффектов дефаунации на основании 23 исследований in vivo , включающих 48 сравнений (Kreuzer et al., 1986; Vermorel and Jouany, 1989; Ushida et al., 1990; Frumholtz, 1991 ; Nagaraja et al., 1992; Hegarty et al., 1994, 2008; Faichney et al., 1999; Chandramoni et al., 2001; Machmuller et al., 2003; Eugène et al., 2004; Ozutsumi et al., 2005; Ohene-Adjei et al., 2007; Yáñez-Ruiz et al., 2007a, b; Bird et al., 2008; Morgavi et al., 2008, 2012; Belanche et al., 2011, 2012b, 2015; Mosoni et al., 2011; Чжоу и др., 2011). Большинство исследований проводилось на овцах (87%), остальные — на крупном рогатом скоте (13%). Изоляция новорожденных животных от их матерей (40%), использование моющих средств и других химикатов (35%, с использованием лаурилсульфата натрия, алканов, синперонового NP9, пероксида кальция, сульфата меди и т. Д.) И манипуляции с рубцом (25%, опорожнение и промывание рубца), были использованы для достижения дефаунации животных. Испытания, в которых влияние добавок, кроме дефауна, было значительным, были исключены из метаанализа.Точно так же не были включены исследования с использованием монофаунизированных животных или селективной дефауны. Большинство данных (75%) были получены из испытаний, в которых животных кормили на поддерживающем уровне, а остальные испытания были из производственных испытаний. Животных в основном кормили смешанными рационами (90%), состоящими из кормов с добавлением концентрата, в то время как 10% рационов состояли исключительно из кормов. Данные о ферментации рубца и выбросы метана были представлены в большинстве испытаний (69%), в то время как информация о пищеварении (31%), продуктивности животных (12%) и микробных популяциях рубца (10%) была менее обширной.Производство метана измерялось в камерах (75% в открытых камерах или респираторных калориметрах) или с помощью метода индикатора SF 6 (гексафторид серы) (25% исследований).
Множественные сравнения были включены из отдельной публикации с множественными исследованиями. Для каждого сравнения, включенного в анализ, величина эффекта рассчитывалась как натуральный логарифм отношения ответа (среднее значение в исчерпанном лечении, деленное на среднее значение в контрольном лечении) и дисперсии отношения, рассчитанной на основе сообщенного стандартного отклонения или стандарта. ошибка среднего для каждого сравнения (Viechtbauer, 2010).Все эффекты дефаунации были взвешены в соответствии с количеством наблюдений ( n ) в каждом сравнении. Мета-анализ был рассчитан с использованием модели случайных эффектов с оценкой Дерсимониан-Лэрда (Dersimonian and Laird, 1989) для оценки неоднородности (τ 2 ) в пакете Metafor R для каждой категории отдельно, как показано ниже:
, где θ i = истинная величина эффекта в i-м исследовании, μ = общая истинная величина эффекта и e i = случайное отклонение от общей величины эффекта [ u i ∼ N (0, τ 2 )] (Viechtbauer, 2010).Для оценки коэффициента отклика значения ниже единицы указывают на отрицательный результат, а значения выше единицы указывают на положительный эффект дефаунации на этот конкретный параметр.
Этот метаанализ подтвердил многие из ранее сообщенных результатов (Williams and Coleman, 1992) и помог прояснить ряд областей, в которых результаты все еще были противоречивыми. На рис. 1 показаны основные эффекты дефаунации на функцию рубца, указывающие на то, что физические и химические характеристики среды рубца были изменены дефаунацией; хотя характер наблюдаемых изменений не всегда был последовательным.Дефауна не повлияла на объем рубца и скорость оборота твердых веществ, в то время как скорость оборота жидкости имела тенденцию к снижению (–14%, P = 0,07). Удаление простейших из рубца значительно снижает усвояемость ОВ рубца (–7%, P = 0,008) и особенно NDF (–20%, P = 0,040) и усвояемость ADF (–16%, P = 0,100) , вероятно, в результате потери фибролитической активности простейших. Однако эта активность, по-видимому, различается в разных группах простейших: крупные Ophyroscolecidae , такие как Epidinium , Polyplastron и Eudiplodinium , обладают большей эндоглюканазной и ксиланазной активностью (Williams and Coleman, 1992).С другой стороны, Entodinium spp. имеют только слабую активность (Williams, Coleman, 1992). Точно так же Dasytricha обладает активностью глюкозидазы и целлобиозидазы, но незначительной фибролитической активностью (Takenaka et al., 2004). Низкая перевариваемость в рубце у дефаунированных животных частично компенсируется более выраженным пострубцовым перевариванием, что приводит к менее выраженным различиям в показателях общей перевариваемости тракта для OM (–4%, P = 0,089), CP (–3%, P = 0.034), NDF (–11%, P <0,001) и ADF (–9%, P <0,001; рис. 2). Другим компенсирующим фактором может быть сдвиг в сторону более энергетически эффективных реакций в рубце (меньшее количество выбросов метана) и меньшая метаболическая энергия, необходимая обезвожившимся животным для устранения избытка мочевины в результате более низкого расщепления бактериального белка и уровня аммиака в рубце. Несмотря на этот компенсаторный механизм, снижение перевариваемости корма, вероятно, будет основным недостатком дефауна, поскольку оно может ограничить потребление корма и эффективность использования корма на производственных уровнях потребления; Это в сочетании с отсутствием коммерчески жизнеспособного подхода (см. ниже) к дефаунации означает, что дефаунация не рекомендуется в качестве стратегии снижения выбросов метана в фермерских условиях (Христов и др., 2013).
Рис. 1. Мета-анализ, описывающий влияние дефаунации на функцию рубца. Для каждого параметра предоставляется информация о количестве исследований, наблюдениях (n) и диапазоне значений. На графике показаны средние эффекты и 95% конфиденциальные интервалы. Значения ниже единицы указывают на отрицательный эффект, тогда как значения выше единицы указывают на положительное влияние отключения на этот конкретный параметр.
Рис. 2. Мета-анализ, описывающий эффекты дефаунации животных. Для каждого параметра предоставляется информация о количестве исследований, наблюдениях (n) и диапазоне значений. На графике показаны средние эффекты и 95% конфиденциальные интервалы. Значения ниже единицы указывают на отрицательный эффект, тогда как значения выше единицы указывают на положительное влияние отключения на этот конкретный параметр.
Общая концентрация и производство продуктов ферментации также различаются у фаунированных и лишившихся животных животных. Снижение аммиака в рубце (–26%, P <0.001), вероятно, является наиболее последовательным из наблюдаемых эффектов элиминации простейших и, по-видимому, обусловлен снижением расщепления бактериального белка и разлагаемости кормового белка в отсутствие простейших в рубце (Williams and Coleman, 1992). Более того, было продемонстрировано, что, хотя хищничество бактерий простейшими в рубце зависит от размера простейших, холотрих простейшие имеют гораздо более низкую хищническую активность, чем энтодиниоморфиды (Belanche et al., 2012a) и, в конечном итоге, более низкое влияние на концентрацию аммиака в рубце (Belanche et al. ., 2015) и микробного белкового потока двенадцатиперстной кишки (Ivan et al., 2000a; Иван, 2009). Однако это негативное влияние энтодиниоморфидов на белковый поток двенадцатиперстной кишки можно переоценить из-за отсутствия надежного маркера для измерения протозойного кровотока (Broderick and Merchen, 1992). В предыдущих обзорах сообщалось о более высоких уровнях лактата у обезвоженных животных (Williams and Coleman, 1992), поскольку простейшие потребляют лактат быстрее, чем бактерии (Newbold et al., 1986). Наш метаанализ показал численное увеличение концентрации лактата у дефунтированных животных (+ 11%), но это увеличение было незначительным, возможно потому, что набор данных, использованный в этом метаанализе, в основном состоял из рационов с высоким содержанием кормов, которые менее склонны к продвижению. эти конкретные нарушения питания.
Хотя рН рубца не изменился, более низкая концентрация ЛЖК, наблюдаемая у дефаунированных животных (–5%, P = 0,013), по-видимому, подчеркивает роль простейших в синтезе ЛЖК и деградации корма. Способность простейших поглощать экзогенные жирные кислоты (Karnati et al., 2009) может отвлекать больше углерода на выработку ЛЖК, а не на синтез жирных кислот, и в конечном итоге увеличивать выработку ЛЖК. Что еще интереснее, наши данные показали, что дефаунация существенно снижает бутират (–22%, P <0.001), слегка повышенное содержание ацетата (+ 3%, P <0,001) и не влияло на молярные пропорции пропионата. В предыдущем метаанализе (Noziere et al., 2011) сообщалось, что усвояемость NDF положительно коррелирует с молярной долей ацетата ( r = 0,95), но отрицательно с пропионатом ( r = –0,94) и бутиратом ( r = –0.91), поэтому наши наблюдения могут указывать на то, что изменение молярных пропорций ЛЖК, вызванное дефаунацией, по-видимому, в основном вызвано уменьшением переваривания клетчатки.Исходя из стехиометрии, такой сдвиг в выработке ЛЖК в рубце должен привести к снижению продукции метана, поскольку менее метаболический H 2 будет доступен в качестве субстрата для метаногенеза (Demeyer et al., 1996). Однако влияние остановки на производство метана все еще не ясно; в различных обзорах, объединяющих in vivo, и in vitro, исследований (Hegarty, 1999) или просто in vivo (Morgavi et al., 2010), был сделан вывод, что удаление простейших из рубца приведет к 13 и 10 .Снижение добычи метана на 5%, что полностью согласуется с нашими результатами, основанными на более поздних исследованиях (–11%, P = 0,074). В соответствии с этими наблюдениями, в метаанализе, содержащем 28 экспериментов и 91 обработку, было сообщено о значительной линейной зависимости между выбросами метана и концентрацией простейших ( r = 0,96) с уменьшением выхода метана в среднем на 8,14 г / кг. DMI на каждую логарифмическую единицу снижения концентрации простейших (Guyader et al., 2014), предполагая, что простейшие играют каталитическую роль в метаногенезе рубца.Причины более низких выбросов метана у дефонированных животных до сих пор остаются спорными, поскольку количество простейших в рубце объясняет только одну часть (примерно 47%) изменчивости выбросов метана, что указывает на то, что метаногенез в рубце представляет собой сложный процесс, в котором участвуют несколько микробов (Morgavi и др., 2010). Возможен ряд механизмов, с помощью которых простейшие могут усиливать метаногенез, в зависимости от их способности продуцировать H 2 в своих гидрогеносомах (митохондриально-подобной органелле), их способности принимать эпи- и эндосимбиотические метаногены и защищать их от кислорода. токсичность (Fenchel and Finlay, 2006).Это взаимодействие является типичным примером межвидового переноса H 2 , который благоприятствует как метаногенам, так и простейшим (Ushida et al., 1997). Одна из гипотез состоит в том, что дефауна приводит к снижению количества метаногенов, которые являются единственными продуцентами метана в рубце, но наш мета-анализ показал, что это снижение уровней метаногенов не было значительным (–3%, P = 0,48). Альтернативная гипотеза предполагает, что дефаунация приводит к элиминации метаногенов, связанных с простейшими, что можно рассматривать как одно из наиболее активных сообществ метаногенов в рубце (Finlay et al., 1994), однако эта более поздняя гипотеза, основанная на замещении метаногеновых сообществ, различающихся по своей метаногенной активности, требует дальнейшего изучения. Несколько авторов изучали эндосимбиотическую популяцию архей у простейших (Sharp et al., 1998; Irbis, Ushida, 2004; Regensbogenova et al., 2004b; Tymensen et al., 2012a), и большинство из них согласны с тем, что Methanobrevibacter sp . является преобладающим родом, а вклад Methanomicrobium sp. и Methanomassiliicoccales в эндосимбиотическом сообществе метаногенов варьирует и может указывать на различия между группами простейших.Развитие молекулярных методов за последние десятилетия позволило коллегам дополнительно изучить эти микробные взаимодействия, обнаружив, что метаногены инфузорий эндосимбиотиков отличаются от свободноживущих метаногенов рубца (Tokura et al., 1997; Sharp et al., 1998). В недавней публикации также было продемонстрировано, что у холотриких простейших есть разные эндосимбиотические метаногены, чем у энтодиниоморфидов (Belanche et al., 2014), возможно, потому, что у любого из холотрих простейших есть более активные гидрогеносомы, чем у энтодиниоморфидов (Paul et al., 1990) и / или быстрый синтез гликогена простейшими холотрихами в присутствии избытка углеводов генерирует больше водорода (Hall, 2011; Denton et al., 2015). Эти различия могут объяснять большее влияние голотричных простейших на метаногенез рубца по сравнению с энтодиниоморфидами (Belanche et al., 2015). Это более позднее наблюдение основано на последовательной инокуляции овец, свободных от фауны, различными группами простейших, поэтому его следует интерпретировать с осторожностью из-за возможных смешанных эффектов лечения и периода.
Влияние дефаунации на микробную экосистему рубца не ограничивается только популяцией метаногена, и было продемонстрировано, что у дефаунированных животных было значительно больше популяций бактерий в рубце (+ 9%, P <0,001), чем у контрольных животных. Причина этого, по-видимому, основана на доступности экологической ниши для бактерий, когда простейшие не присутствуют в рубце, в сочетании с устранением простейших хищников (Williams and Coleman, 1992).Недавние исследования с использованием молекулярных методов показали, что дефаунация также изменяет структуру бактериального сообщества, что приводит к упрощению популяционной структуры и снижению бактериального разнообразия (Belanche et al., 2012b, 2015). Наш метаанализ выявил снижение концентрации фибролитических микробов, таких как анаэробные грибы (–92%, P <0,001), Ruminococcus albus (–34%, P <0,001) и Ruminococcus flavefaciens ( –22%, P <0.001) в отсутствие простейших. Это наблюдение предполагает, что переваривание клетчатки в рубце - сложная задача, которая требует симбиотического сотрудничества нескольких фибролитических микробов, включая простейшие рубца, для выполнения начальных стадий колонизации клетчатки и пищеварения; поэтому отсутствие простейших в рубце, по-видимому, оказывает пагубное влияние на этот фибролитический консорциум и, в конечном итоге, на переваривание клетчатки. Однако Hsu et al. (1991) сообщили об увеличении зооспор грибов в рубце у лишившихся жизни животных, возможно, в результате устранения хищничества простейших и конкуренции за питательные вещества (простейшие vs.грибки), очевидно, необходимы дополнительные исследования, чтобы полностью понять взаимодействие простейших с другими микробами рубца при различных режимах питания.
Влияние присутствия простейших в рубце на выживаемость патогенов в рубце и выделение патогенов — еще одна область, представляющая интерес. Как отмечалось выше, простейшие в рубце поглощают и переваривают широкий спектр бактерий (Williams and Coleman, 1992) и могут уменьшить выделение потенциальных патогенов от животного, хотя эффект сильно зависит от состава присутствующей популяции простейших (Stanford et al. ., 2010). Однако также было показано, что простейшие в рубце увеличивают патогенность некоторых патогенов, покидающих рубец (Rasmussen et al., 2005; Carlson et al., 2007), что позволяет предположить, что в этой области требуется дополнительная работа.
Рисунок 2 суммирует влияние простейших на продуктивность животных как косвенный эффект дефаунации на функцию рубца. Дефауна улучшила конверсию корма в результате более низкого потребления СВ (–2%, P <0,001) и большего среднесуточного привеса (+ 9%, P <0.001). Эти положительные эффекты дефауна особенно очевидны при низком качестве диеты, при которой средний дневной прирост был низким, возможно, из-за более низкой доступности ферментируемой энергии и перевариваемого в рубце белка для микробов рубца (Williams and Coleman, 1992). В предыдущем метаанализе эффект дефаунации оказался более выраженным, когда отношение N / NDF было ниже 6, а процент концентрата ниже 40% (Eugène et al., 2004). Так как животные, лишенные жизни, имеют более низкую усвояемость корма, поглощенная энергия ниже, чем у обычных животных.Таким образом, лучшая степень конверсии корма после отключения может быть главным образом связана с более высокой эффективностью использования поглощенных питательных веществ. Действительно, наш мета-анализ показал, что дефауна способствует большей эффективности использования энергии для откорма (+ 11%, P = 0,001), возможно, в результате более низкой выработки тепла (–5%, P = 0,010). Другая гипотеза, объясняющая лучшую продуктивность дефунтированных животных, основана на более эффективном использовании пищевого белка. Существуют четкие доказательства того, что простейшие холотрицы покидают рубец медленнее, чем бактерии (Abe et al., 1981). Однако количество и скорость потока простейших в нижний отдел кишечника является предметом многочисленных споров. Firkins et al. (2007), обнаружили, что поструминальный поток простейших пропорционален биомассе простейших в рубце. Хотя подсчет простейших в рубце и сычуговой жидкости показал, что количество сычужных сычуг составляло только 6–64% от подсчета жидкости в рубце (Punia et al., 1992), количество простейших в свободной жидкости рубца является ненадежным индикатором биомассы и оттока простейших, поскольку большинство простейших в рубце (63–90%) обнаруживаются либо связанными с частицами корма, либо секвестрированными в стенке рубца (Hook et al., 2012). Использование специфических маркеров простейших, таких как 2-аминоэтилфосфоновая кислота, подвергается сомнению, поскольку она присутствует в бактериях и в кормах (Ling and Buttery, 1978). Протозойный отток, измеренный с использованием общих микробных маркеров (например, 15 N и 35 S и пуриновых оснований), микробных N минус специфических бактериальных маркеров (таких как 2,6-диаминопимелиновая кислота, DAPA) давал различные оценки протозойных отток, а иногда и отрицательные значения из-за методологических ограничений используемых маркеров (Broderick and Merchen, 1992).Недавние исследования, основанные на использовании 18S рРНК в качестве нового маркера простейших, показали, что, хотя простейшие могут составлять до 60% микробной биомассы рубца, они редко превышают 20% потока микробных белков в тонкий кишечник (Sylvester et al. , 2004, 2005; Yáñez-Ruiz et al., 2006; Belanche et al., 2011, 2012b). Однако эти новые методы не лишены собственных ограничений: Sylvester et al. (2005) наблюдали, что до 48% простейшего азота на самом деле было азотом от заражающих бактерий, что могло привести к раздуванию пула простейшего азота в рубце, особенно когда животные имеют низкое потребление корма и низкую скорость прохождения твердых частиц (Dijkstra, 1994).Аналогичным образом Belanche et al. (2010b) наблюдали большую деградацию протозойной ДНК по сравнению с бактериальной ДНК во время сычужного переваривания, что могло привести к недооценке оттока простейшего азота из рубца. Несмотря на эти ограничения, эти новые данные, кажется, подтверждают, что простейшие в рубце частично секвестрируются в рубце. Однако эта секвестрация в рубце не одинакова для всех групп простейших; holotrich простейшие связываются с частицами корма после кормления из-за их сильного хемотаксиса по отношению к сахарам (Diaz et al., 2014), но быстро мигрируют в вентральную ретикулоруму, чтобы предотвратить вымывание из рубца (Karnati et al., 2007), напротив, энтодиниоморфиды также связываются с кормом из-за их умеренного хемотаксиса в отношении глюкозы и пептидов, но не проявляют такое же сродство к стенке рубца (Diaz et al., 2014) и, таким образом, вытекает из рубца с фазой твердых частиц (Hook et al., 2012).
Простейшие предшествуют бактериям в качестве своего основного источника белка (Williams and Coleman, 1992), и в результате дефаунация делает рубец более эффективным с точки зрения протеосинтеза, увеличивая дуоденальный поток микробного белка (+ 30%, P <0.001) и общий поток неаммиачного азота (+ 31%, P <0,001). Дефаунация также увеличивала эффективность синтеза микробного белка (+ 27%, P = 0,008) в результате как лучшего микробного протеосинтеза, так и более низкого усвоения ОМ. Время генерации протозойных намного выше, чем у бактерий, поэтому энергетические потребности для поддержания выше, если выразить их как соотношение белков, покидающих рубец (Williams and Coleman, 1992). В результате присутствие простейших отрицательно сказывается на общей энергетической эффективности экосистемы рубца.Кроме того, дефаунация также может изменять состав бактерий рубца (Belanche et al., 2012b) и, в конечном итоге, аминокислотный профиль поступления белка двенадцатиперстной кишки, способствуя увеличению определенных аминокислот, таких как лейцин, треонин и аргинин (Ivan, 2009 ), но не лизин, который считается основной лимитирующей аминокислотой у высокопродуктивных животных (Hristov and Jouany, 2005). После серии экспериментов in vivo , в которых ягнят, свободных от фауны, постепенно инокулировали простейшие виды (т.e., Isotricha Кишечник , Dasytricha ruminantium , Polyplastron multivesiculatum , Epidinium ecaudatum , Eudiplodinium maggi , и Ivan Entodinium caudatum, в разных порядках, b. 2009), Иван пришел к выводу, что холотрих простейшие поглощают лишь очень небольшое количество бактерий рубца и оказывают незначительное влияние на дуоденальный поток NAN и метаболизм белков. Точно так же простейшие голотрих не оказали значительного влияния на переваривание клетчатки.Это вместе с вкладом голотрихов в метаногенез рубца (Belanche et al., 2015), по-видимому, указывает на то, что присутствие голотрихов простейших в рубце не имеет большого значения для производства жвачных животных, если не используются диеты с высоким содержанием углеводов, и в этом случае присутствие голотричных простейших может быть полезным в результате их способности поглощать и накапливать зерна крахмала и растворимые углеводы (Williams and Coleman, 1992). Такое поглощение высоко ферментируемых углеводов предотвращает альтернативную бактериальную ферментацию, которая в противном случае снизила бы pH и увеличила бы возникновение молочнокислого ацидоза (Mackie et al., 1978). Напротив, целлюлолитические простейшие ( Polyplastron , Epidinium и Eudiplodinium ) могут быть полезными для животных, которых кормили волокнистой пищей. Однако эти фибролитические простейшие и Entodinium spp. обладают значительным потенциалом поглощения и разложения бактерий (Belanche et al., 2012a) и могут быть вредными с точки зрения использования белка жвачным животным-хозяином. Следовательно, их присутствие в рубце может быть бесполезным для животных, получающих диету с низким содержанием белка, поскольку их благоприятное фибролитическое действие будет уравновешено чувствительностью фибролитических бактерий и анаэробных грибов к дефициту азота в рубце (Belanche et al., 2012c). Таким образом, похоже, что дефауна уменьшила потери азота с мочой здесь (–10%, P <0,001) из-за комбинации более низкой пищевой деградации CP в рубце и бактериального расщепления нижней части рубца. Однако большая изменчивость, наблюдаемая с точки зрения баланса азота, подчеркивает тот факт, что дефаунация может по-разному влиять на общую эффективность использования азота в зависимости от диеты, потребляемой животным, в частности, уровня потребления и скорости прохождения частиц через тракт (Дейкстра , 1994).
Влияние дефаунации на метаболизм липидов в рубце менее определено. Простейшие вносят вклад в общую липолитическую активность микробов, но их роль в биогидрировании менее изучена (Williams and Coleman, 1992), хотя они действительно вносят значительный вклад в отток ненасыщенных жирных кислот, покидающих рубец (Yáñez-Ruiz et al., 2006). Используя рацион бычков, получавших рационы с различным содержанием хлорофилла, было продемонстрировано, что высокие уровни полиненасыщенных жирных кислот в клетках простейших, по-видимому, связаны с потреблением хлоропластов (Huws et al., 2009). Это поглощение хлоропластов, по-видимому, специфично для энтодиниоморфидов, поскольку не было обнаружено захваченных хлоропластов у голотриких простейших (Huws et al., 2012). Таким образом, простейшие, по-видимому, защищают ненасыщенные жирные кислоты хлоропластов от биогидрирования рубца, увеличивая дуоденальные потоки моно- и полиненасыщенных жирных кислот. Наш метаанализ согласуется с этим наблюдением, и дефаунация способствовала увеличению насыщенных жирных кислот (+ 4%, P = 0,046) и численному снижению моно- и полиненасыщенных жирных кислот (–13 и –10% соответственно), что возможно, не достигли значимости из-за эффектов, зависящих от диеты.
Результаты, представленные в нашем метаанализе, должны быть тщательно интерпретированы, поскольку большинство исследований имело методологические ограничения из-за внутренней сложности процесса дефаунации, ведущего к:
(1) Путаница между эффектом дефаунации и периодом при использовании одних и тех же экспериментальных животных во времени,
(2) Почти все исследования, оценивающие размер пула простейших, не смогли сообщить о дуоденальных потоках протозойной биомассы, а наоборот , более того, поток простейших часто недооценивается с использованием традиционных микробных маркеров (Broderick and Merchen, 1992),
(3) Исследования, оценивающие производство метана, не учитывали другие переменные эффекты, кроме численности простейших (т.е., усвояемость NDF),
(4) Животных часто кормят в количествах, намного ниже производственных уровней потребления,
(5) В некоторых исследованиях период адаптации после дефаунации / восстановления слишком короток для того, чтобы микробная экосистема могла полностью адаптироваться,
(6) Устранение простейших из рубца изменяет экологическую структуру рубца, и в конечном итоге альтернативные группы микробов могут занять экологическую нишу, ранее заполненную простейшими.
Таким образом, необходимы дополнительные исследования с использованием современных технологий для количественной оценки активности простейших (т.е., фибролитические и протеолитические), а также размер пула простейших в рубце относительно оттока простейшего азота для лучшего понимания роли простейших в метаболизме жвачных. Эти исследования должны проводиться на производственных уровнях потребления, чтобы действительно оценить эффект дефаунации на единицу произведенного продукта (молока или мяса).
Манипуляции с простейшими в рубце
Текущие опасения по поводу роли домашнего скота в глобальном предупреждении побудили исследователей искать стратегии манипулирования простейшими в рубце для уменьшения производства метана.Как уже упоминалось ранее, существует линейная зависимость между концентрацией простейших и выбросами метана (Guyader et al., 2014), и было подсчитано, что от 9 до 37% продукции метана в рубце можно отнести к метаногенам, связанным с простейшими в рубец (Finlay et al., 1994; Newbold et al., 1995; Machmuller et al., 2003).
Большинство подходов к дефаунации редко приводят к полному удалению простейших из рубца, их эффективность во многом зависит от состава диеты (Hegarty, 1999).Лечение, обычно используемое для частичной или полной дефауны рубца, включает: химические вещества, токсичные для простейших (сульфат меди, диоктилсульфосукцинат, этоксилат или алканаты спирта, пероксид кальция), ионофоры, липиды и сапонины (Williams and Coleman, 1992; Jouany, 1996; Hook et al., 2010).
Недавний метаанализ Guyader et al. (2014) показали одновременное снижение численности простейших и выбросов метана в 31% из 70 исследований, в которых использовались различные стратегии сокращения выбросов метана.В большинстве исследований липиды использовались в качестве стратегии борьбы с простейшими / снижения уровня метана. Авторы также сообщили, что антипротозойный эффект липидов зависит от состава жирных кислот, причем жирные кислоты со средней длиной цепи более эффективны, чем полиненасыщенные жирные кислоты, в контроле численности простейших. Было показано, что добавки, богатые полиненасыщенными жирными кислотами, такими как линолевая кислота (C18: 2 из сои и подсолнечника) и линоленовая кислота (C18: 3 из льняного семени), оказывают негативное влияние на выработку метана (4.Снижение на 1 и 4,8% на процентную единицу добавленных липидов, соответственно, Martin et al., 2010). Антиметаногенный эффект полиненасыщенных жирных кислот был связан с их токсическим действием на целлюлозолитические бактерии (Nagaraja et al., 1997) и простейшие (Doreau and Ferlay, 1995). Хотя первоначально предполагалось, что биогидрирование полиненасыщенных жирных кислот в рубце может представлять собой альтернативу метаногенезу H 2 (Lennarz, 1966), теперь считается, что значение биогидрирования для общего стока H 2 является small (Nagaraja et al., 1997). Было показано, что жирные кислоты со средней длиной цепи обладают сильным антипротозойным действием. В нескольких исследованиях сообщалось о снижении метаногенеза в рубце при добавлении лауриновой кислоты (C12: 0) и миристиновой кислоты (C14: 0) либо в чистых формах, либо в продуктах, богатых этими жирными кислотами (кокосовое масло), под in vitro (Dohme et al. ., 2001; Soliva et al., 2004) или in vivo в условиях (Machmuller et al., 2003; Jordan et al., 2006). Martin et al. (2010) сообщили, что жирные кислоты со средней длиной цепи, в основном содержащиеся в кокосовом масле, привели к снижению метана на 7.3% на процентную единицу добавленного липида. Однако наблюдаемый эффект подавления метана не всегда был связан со снижением концентрации простейших (Machmuller et al., 2003), что может быть связано с прямым действием жирных кислот со средней длиной цепи на метаногены (Dohme et al., 1999; Panyakaew et al. др., 2013). Недавнее исследование (Faciola and Broderick, 2014) показало, что кокосовое масло и лауриновая кислота снижают количество простейших на 40%, тогда как лауриновая кислота изменяет усвояемость клетчатки, кокосовое масло — нет.Однако сообщалось о сокращении выработки метана и сопутствующем уменьшении потребления сухого вещества, когда кокосовое масло и лауриновая кислота использовались в качестве дезорганизующих агентов (Христов и др., 2013), что потенциально ограничило бы их практическое использование в фермерских хозяйствах. Очевидно, что необходимы дополнительные исследования, чтобы выяснить, как использовать масла и жирные кислоты для борьбы с простейшими в рубце.
В литературе предполагается, что сапонины смягчают метаногенез, главным образом, за счет уменьшения количества простейших, в то время как конденсированные танины действуют как за счет уменьшения количества простейших, так и за счет прямого токсического воздействия на метаногены, тогда как эфирные масла действуют в основном путем прямого токсического действия на метаногены (Cieslak и другие., 2013). В соответствии с этой информацией, метаанализ влияния фитохимических веществ на метаногенез (Patra, 2010) показал, что изменения численности простейших следовали линейной зависимости с изменениями выработки метана сапонинами ( r = 0,69) и танинами ( r = 0,55), но эта связь была слабее ( r = 0,45) по отношению к эфирным маслам. Ингибирование метана сероорганическими соединениями не было связано с изменениями в популяции простейших (Patra, 2010), поскольку такие соединения специфически ингибируют метаногенные археи.Для растений, содержащих танин, антиметаногенная активность приписывается группе конденсированных танинов. Было высказано предположение, что дубильные вещества имеют прямое влияние на метаногены в рубце и косвенное влияние на производство водорода из-за снижения переваривания клетчатки (Tavendale et al., 2005). Что касается их воздействия на простейшие, в некоторых исследованиях не сообщалось об их эффекте, в то время как другие показали снижение количества простейших в присутствии танинов (обзор Patra and Saxena, 2009). Это несоответствие, вероятно, связано с различиями в структуре и дозе конденсированного танина.Сапонины в доступной литературе показывают более стойкое ингибирующее действие на простейшие в рубце. Гоэль и Маккар (2012) предположили, что риск нарушения функции рубца и, как следствие, снижения продуктивности животных выше с дубильными веществами, чем с сапонинами, и когда они используются для уменьшения выработки метана, эффективный диапазон концентраций дубильных веществ уже, чем сапонинов. Сапонины представляют собой гликозилированные тритерпены или стероиды, где сапонин представляет собой агликон, а гликон представляет собой углеводную единицу, состоящую из моносахарида или олигосахарида меньшего размера.Наиболее частыми источниками сапонинов, используемых в питании жвачных животных, являются Yucca schidigera , богатая стериновыми сапонинами (10%), и Quillaja saponaria , содержащая тритерпеновые сапонины. В последнее время изучаются и другие источники сапонинов, такие как сапонины чая (Hu et al., 2005; Guo et al., 2008). Антипротозойный эффект сапонинов связан с их взаимодействием со стероловым фрагментом, который присутствует в мембране простейших (Patra and Saxena, 2009). Было высказано предположение, что сапонины с одним и тем же агликоном могут по-разному влиять на простейшие в зависимости от состава и расположения сахара (Wina et al., 2006). Этот эффект кажется временным из-за разрыва гликозидной связи микробами рубца (Newbold et al., 1997). Подавление простейших в рубце сапонинами или сапонинсодержащими растениями постоянно наблюдается в исследованиях in vitro (Wina et al., 2005). Однако исследования in vivo показали, что антипротозойный эффект сапонинов имеет тенденцию исчезать через 7–14 дней приема (Wina et al., 2005; Patra and Saxena, 2009). Было высказано предположение, что когда сапонины дегликозилируются в сапогенины микробами рубца, они становятся неактивными (Newbold et al., 1997). Таким образом, можно предположить, что комбинация сапонинов с ингибиторами гликозидазы позволит избежать дегликозилирования, поддерживая интактный сапонин и, следовательно, активность в рубце. Предварительные исследования in vitro (Ramos-Morales et al., 2014) с использованием 2,5-дигидроксиметил-3,4-дигидроксипирролидина (DMDP) в качестве ингибитора гликозидазы в сочетании с экстрактом растений, богатым сапонинами, показали потенциал этого подхода. для поддержания активности сапонинов с течением времени. Другой подход, который изучается нашей исследовательской группой, — это синтез химически модифицированного сапонина без естественных гликозидных связей, поэтому ферментативное расщепление будет структурно запрещено.Влияние сапонинов на ферментацию рубца не подтвердилось. Эти расхождения, по-видимому, связаны с химической структурой и дозировкой сапонинов, составом рациона, а также адаптацией микроорганизмов к сапонинам (Wina et al., 2005; Patra and Saxena, 2009).
Австралийские исследователи продемонстрировали потенциал вакцинации против метаногенов как метода уменьшения выбросов метана (Wright et al., 2004). Хотя эта технология все еще развивается, она предоставляет множество возможностей для долгосрочного сокращения выбросов метана.Точно так же простейшие, как поставщики водорода для метаногенов или ацетогены, которые конкурируют за водород с метаногенами, могут быть возможными целями вакцины для сокращения выбросов метана. Иммунологический подход был изучен для дефаунации (Williams et al., 2008). Вакцинация овец энтодинией или смешанными протозойными антигенами уменьшала количество простейших, и антитела IgG, выработанные против простейших в рубце, оставались активными и продолжали связывать клетки-мишени в течение до 8 часов. Однако не наблюдалось никакого действия in vivo на простейшие в рубце.Было высказано предположение, что причины отсутствия эффекта могут быть связаны с недостаточным количеством специфического Ig, доставляемого в слюне, необходимостью адъюванта для оптимизации выработки антител в слюне, нацеливания на другие антигены простейших для создания более сильного иммунного ответа.
Заключение
Со времени знаменательной публикации «Rumen Protozoa» Уильямса и Коулмана (1992) в нашем понимании простейших рубца наблюдается устойчивый, но, возможно, не впечатляющий прогресс.Появление молекулярных методов привело к появлению множества публикаций, касающихся разнообразия простейших в рубце, и, хотя, как обсуждалось выше, используемые методы имеют внутри себя ограничения в их способности точно количественно определять отдельные роды простейших, они предоставили новое понимание разнообразия простейших инфузорий. у разных видов жвачных, в разных географических регионах и при разных диетических условиях. В области дефаунации наблюдается неуклонный прогресс, и хотя на данном этапе на рынке еще не поступило коммерчески доступной техники дефаунации, вполне вероятно, что экстракты растений можно использовать для борьбы с простейшими в рубце; если не устранить их полностью.Работа над последствиями уничтожения простейших в значительной степени сосредоточена на их роли в метаногенезе, что отражает текущие опасения относительно роли жвачных животных в производстве парниковых газов. Однако использование метаанализа существующих данных в сочетании с новыми исследованиями дефаунации помогло прояснить наше понимание роли простейших в рубце, как показано на рисунках 1 и 2. Однако, возможно, это развитие молекулярных методов для клонирования и характеристики простейших. гены, первоначально от одного вида, но в последнее время из метагеномных и транскриптомных источников, которые, по-видимому, предлагают наибольший, но пока еще в значительной степени не реализованный, потенциал, чтобы помочь по-настоящему выяснить роль простейших в рубце и в отсутствие прогресса в развитии аксенических Культура простейших в рубце Необходимо приложить больше усилий для характеристики активности простейших в рубце с помощью молекулярных методов.
Авторские взносы
Все авторы в равной степени внесли вклад в подготовку данной публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
CN и AB выражают признательность за поддержку Исследовательскому совету биотехнологии и биологических наук Великобритании через грант номер BB / J0013 / 1.
Ссылки
Abe, M., Iriki, T., Tobe, N., and Shibui, H. (1981). Секвестрация простейших голотрихов в ретикуло-рубце крупного рогатого скота. Заявл. Environ. Microbiol. 41, 758–765.
PubMed Аннотация | Google Scholar
Амарал-Цеттлер, Л. А., МакКлимент, Э. А., Даклоу, Х. В., и Хьюз, С. М. (2009). Метод изучения разнообразия протистанов с использованием массивно-параллельного секвенирования гипервариабельных участков v9 малых субъединичных генов рибосомной РНК. PLoS ONE 4: e6372. DOI: 10.1371 / journal.pone.0006372
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Амарал-Цеттлер, Л. А., Зеттлер, Э. Р., Теру, С. М., Паласиос, К., Агилера, А., и Амилс, Р. (2011). Структура микробного сообщества на древе жизни в крайнем Рио-Тинто. ISME J. 5, 42–50. DOI: 10.1038 / ismej.2010.101
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Абесия, Л., Holtrop, G., Guada, J. A., Castrillo, C., de la Fuente, G., et al. (2011). Изучение влияния присутствия или отсутствия простейших на ферментацию рубца и вклад микробных белков в химус. J. Anim. Sci. 89, 4163–4174. DOI: 10.2527 / jas.2010-3703
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Бальселлс, Дж., Де Ла Фуэнте, Г., Яньес-Руиз, Д. Р., Фондевила, М., и Каллеха, Л. (2010a). Описание развития экосистемы рубца с помощью ПЦР-анализа у ягнят на молочном, отнятом от груди и в откорме в системе интенсивного откорма. J. Anim. Physiol. Anim. Nutr. 94, 648–658. DOI: 10.1111 / j.1439-0396.2009.00952.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче А., Эрроа И. Р., Балселлс Дж. И Каллеха Л. (2010b). Использование количественной ПЦР в реальном времени для оценки выживания in vitro определенных последовательностей генов ДНК микробов рубца в имитированных сычужных условиях. J. Anim. Physiol. Anim. Nutr. 94, 204–211. DOI: 10.1111 / j.1439-0396.2008.00901.х
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., Мурби, Дж. М., и Ньюболд, К. Дж. (2012a). Расщепление бактериального белка разными простейшими группами рубца. J. Anim. Sci. 90, 4495–4504. DOI: 10.2527 / jas.2012-5118
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., Пинлоче, Э., Ньюболд, К. Дж., И Балселлс, Дж. (2012b). Влияние диеты и отсутствия простейших на микробное сообщество рубца и на репрезентативность бактериальных фракций, используемых при определении синтеза микробного белка. J. Anim. Sci. 90, 3924–3936. DOI: 10.2527 / jas.2011-4802
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Доро, М., Эдвардс, Дж. Э., Мурби, Дж. М., Пинлоче, Э. и Ньюболд, К. Дж. (2012c). Изменения в микробиоте рубца из-за типа углеводов и уровня белка, потребляемого молочным скотом, связаны с изменениями ферментации рубца. J. Nutr. 142, 1684–1692. DOI: 10.3945 / jn.112.159574
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., и Ньюболд, К. Дж. (2014). Изучение метаногеновых сообществ, связанных с различными популяциями простейших в рубце. FEMS Microbiol. Ecol. 90, 663–677. DOI: 10.1111 / 1574-6941.12423
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче А., де ла Фуэнте Г. и Ньюболд К. Дж. (2015). Влияние прогрессивной инокуляции овец, свободных от фауны, холотобоязненными простейшими и всей фауной на ферментацию рубца, микробное разнообразие и выбросы метана. FEMS Microbiol. Ecol. 362, 1–10. DOI: 10.1093 / фемсек / fiu026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Белжецкий, Г., Ньюболд, К. Дж., МакЭван, Н. Р., Макинтош, Ф. М., и Михаловски, Т. (2007). Характеристика амилолитических свойств простейших инфузорий рубца Eudiplodinium maggii . J. Anim. Feed Sci. 16, 590–606.
Google Scholar
Bera-Maillet, C., Devillard, E., Cezette, M., Jouany, J.П., Форано Э. (2005). Ксиланазы и карбоксиметилцеллюлазы простейших в рубце Polyplastron multivesiculatum , Eudiplodinium maggii и Entodinium sp. FEMS Microbiol. Lett. 244, 149–156. DOI: 10.1016 / j.femsle.2005.01.035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берд, С. Х., Хегарти, Р. С., Вудгейт, Р. (2008). Сохранение эффектов дефаунации на пищеварение и выработку метана у овец. Aust. J. Exp. Agric. 48, 152–155. DOI: 10.1071 / EA07298
CrossRef Полный текст | Google Scholar
Boxma, B., Ricard, G., van Hoek, A.H., Severing, E., Moon-van der Staay, S.Y., van der Staay, G.W., et al. (2007). Гидрогеназа [FeFe] Nyctotherus ovalis имеет химерное происхождение. BMC Evol. Биол. 7: 230. DOI: 10.1186 / 1471-2148-7-230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бродерик, Г.А., Мерчен, Н.Р. (1992). Маркеры для количественного определения синтеза микробного белка в рубце. J. Dairy Sci. 75, 2618–2632.
PubMed Аннотация | Google Scholar
Карлсон, С. А., Шарма, В. К., МакКаддин, З. П., Расмуссен, М. А., и Франклин, С. К. (2007). Участие гена Salmonella геномного острова 1 в рубце, опосредованном простейшими, усиление инвазии для множественной устойчивости к антибиотикам Salmonella enterica серовар Typhimurium . Заражение.Иммун. 75, 792–800. DOI: 10.1128 / IAI.00679-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Case, R. J., Boucher, Y., Dahllöf, I., Holmström, C., Doolittle, W.F., and Kjelleberg, S. (2007). Использование генов 16S рРНК и rpoB в качестве молекулярных маркеров для исследований микробной экологии. Заявл. Environ. Microbiol. 73, 278–288. DOI: 10.1128 / AEM.01177-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кавальер-Смит, Т.(2005). Экономика, скорость и размер имеют значение: эволюционные силы, движущие миниатюризацией и расширением ядерного генома. Ann. Бот. 95, 147–175. DOI: 10.1093 / aob / mci010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чандрамони, Джадхао, С. Б., Тивари, К. М., Хак, Н., Мурари Лал и Хан, М. Ю. (2001). Энергетический метаболизм и выработка метана у фаунированных и дефонтированных овец, которых кормили двумя рационами с одинаковым соотношением концентратов и грубых кормов (70: 30), но разного состава. Азиатско-Австралийский. J. Anim. Sci. 14, 1238–1244. DOI: 10.5713 / ajas.2001.1238
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cieslak, A.
0 Limburgs: Protozoa
Plattdüütsch: Protozoa
norsk nynorsk: Protozo
norsk: Protozo
polski: Pierwotniaki
Protozoa0 Protozoa0: 9rio 170009: 9rio 170009: 9 Английский:
Protozoa
slovenčina: Prvoky
svenska: Urdjur
தமிழ்: முன்னுயிரிகள் (புரோட்டோசூவா)
Ссылки [править]
Goldfuss, Georg August: Ueber die Entwicklungsstufen des Thieres , Нюрнберг, Леонард Шраг (1817) [1].
Дополнительные ссылки [править]
- Беккер, Э. Р. и Кюн Л. Б. 1939: Роды простейших . 65 с. (Мимеография), [2].
- Mauch, E., Schmedtje, U., Maetze, A. & Fischer, F. 2003: Taxaliste der Gewässerorganismen Deutschlands zur Kodierung biologischer Befunde , [3]. (Со списком определяющей литературы с комментариями.)
См. Также [править]
Альтернативные классификации [править]
[Примечание: в большинстве классификаций Protozoa в настоящее время считается полифилетическим таксоном.Некоторые авторы (например, Кавалье-Смит) все еще используют его как парафилетический таксон, хотя большинство систематиков предпочитают использовать только монофилетические таксоны. См. Более современную классификацию ранее размещенных здесь групп в Eukaryota.]
Гольдфус (1820) [править]
Из Handbuch der Zoologie .
Класс Простейшие
фон Зибольд (1848) [править]
Lehrbuch der vergleichenden . Vol. 1, Wirbellose Thiere, von C. Th. против Зибольда, 1848 г.Vol. 2, Wirbelthiere, von H. Stanius, 1846, [4].
Тип Простейшие
Оуэн (1860) [править]
Из Палеонтология или систематический обзор вымерших животных и их геологических отношений .
Королевство Простейшие
Hogg (1860) [править]
Хогг, Дж. (1860 г.). О различиях растений и животных и о четвертом царстве природы . Эдинбург Нью Фил. J. , Н.С. 12: 216–225 [5].
- Минеральное царство
- Растительное царство
- Царство животных
- Первобытное королевство (Regnum Primigenum)
Haeckel (1874) [править]
Из Геккеля Э (1874 г.). Die Gastrea-Theorie, die phylogenetische Classification des Tierreiches und die Homologie der Keimblatter. Jenaische Zeitschrift fur Naturwissenschaft , 8, 1–55, [6].
См. См. Kluge (2004-2014), [7].
[стр. 32:]
1. Абластерия Простейшие Простейшие Простейшие 2. Дибластерия Gastraeada Zoophyta Metazoa Spongiae 3. Трибластерия Acalephae 4. Тетрабластерия Акоеломи Vermes 5. Пентабластерия Coelomati Mollusca Typozoa Иглокожие Членистоногие Позвоночные
[П.33:]
- Bilateria = Sphenota
- Vermes I + Vermes II [Sammtlische Würmer]
- Mollusca [Molluscken]
- Иглокожие [Echinotermen]
- Членистоногие [Arthropoden]
- Vertebrata [Vertebraten]
[P. 52:]
- Thierreichs
- Простейшие
- Metazoa [= Blastozoa, p. 19]
- Anaemaria
- Haemataria = Pentablasteria
Lankester (1877) [править]
Ланкестер, Э.R. 1877. Заметки по эмбриологии и классификации животного мира: включает пересмотр предположений о происхождении и значении зародышевых листков. Ежеквартальный журнал микроскопических исследований , 68: 399-454, [8].
Сэвилл-Кент (1880-1882) [править]
Из Сэвилл-Кент, W. (1880–1882). Инструкция Инфузории . Лондон: Д. Бог, 3 тома, [9].
Под-царство Protozoa [стр. 36]
Простейшие царства [стр. 211]
Bütschli (1880-1889) [править]
От Bütschli, O.(1880–1889). Классификация и порядок вещей доктора Х. Г. Бронна: wissenschaftlich dargestellt in Wort und Bild: wissenschaftlich dargestellt in Wort und Bild. Кольцо Эрстера: простейшие. Abt. I, Sarkodina und Sporozoa, 1880-82 [10]. Abt. II, Мастигофора, 1883–87 [11]. Abt. III, Infusoria und System der
Radiolaria, 1887-89 [12]. К.Ф. Зима, Лейпциг.
Тип Простейшие
Ланкестер (1885, 1891) [править]
Ланкестер, Э. Р. (1885). Статья «Простейшие».В: Британская энциклопедия (9-е изд.), 19: 830–866,
[13].
Ланкестер, Э. Р. (1891). Статья «Простейшие». В: Зоологические статьи, внесенные в Британскую энциклопедию , [14].
Блохманн (1886) [править]
Die mikroskopische Tierwelt des Süßwassers , 1-е изд., 1886, [15].
Простейшие
Геккель (1889 г.) [править]
Haeckel, E. 1889. Natürliche Schöpfungsgeschichte , 8-е изд, [16].См. Стр. 452, 453, 462, 508.
- Protistenreich
- Protophyta (Protista Vegetalia)
- Класс I. Фитарча
- Фитомонера
- Chromaceae
- Ordnung 2. Chroococceae
- Ordnung 3. Nostochineae
- Класс II. Диатомеи
- Класс III. Космарии (Desmidieae)
- Класс IV. Пальмеллярии (Palmellaceae)
- Класс V. Siphoneae (Coeloblastae)
- Простейшие (Protista animalia)
- Зоарча (Zoocytoda)
- Zoomonera
- Бактерии
- Сфаэробактерии
- Рабдобактерии
- Cytarcha
- Лобоса (Амебина)
- Gregarinae (Sporozoa)
- Инфузории
- Flagellata (Мастигофора)
- Реснички
- Acinetae (Suctoria)
- Rhizopoda
- Mycetozoa (Myxomycetes)
- Heliozoa
- Таламария (Foraminifera)
- Радиолярия
- Pflanzenreich
- Thallota (Thallophyta)
- Водоросли (Phycophyta)
- Zygnemaceae
- Conferveae
- Fucoideae
- Флориды
- Characeae
- Грибы (Inophyta)
- Prothallota (Мезофита)
- Phanerogamae (Anthophyta)
- Gymnospermae
- Cycadeae
- Хвойные
- Gnetaceae
- покрытосемянных
- Thierreich
- кишечнополостные (Coelenterata, Zoophyta, Anaemaria)
- Gastraeades
- Губки
- Книдария
- Платоды
- Целомария (Bilaterata, Bilateria, Haemataria)
- Гельминты
- Моллюска
- Иглокожие
- Сочленений
- Туниката
- Позвоночные
Клаус (1891) [править]
Из Lehrbuch der zoologie .
Простейшие
Геккель (1894) [править]
Из Systematische phylogenie , 3 тома (том 1, Protisten und Pflanzen, 1894; том 2, Invertebrata, 1896; том 3, Vertebrata, 1895). [17].
[Том. 1, стр. 90-91:]
Пфланцен
- Protophyta (плазмодома Protista) [Vol. 1, стр. 96]
- Archephyta (Прогонелла)
- Пробионты
- Хромовые (Phycochromaceae)
- Алгарии (Paulosporata)
- Paulotomeae
- Конъюгаты
- Диатомеи
- Альгетты (Zoosporata)
- Мастигота
- Мелеталия
- Siphoneae
- Metaphyta (гистоны плазмодомии) [Vol.1, стр. 256]
- Thallophyta
- Водоросли [Vol. 1, стр. 302]
- Chlorophyceae
- Charaphyceae
- Phaeophyceae
- Rhodophyceae
- Mycetes [Vol. 1, стр. 316]
- Аскомицеты (Ascodiomycetes)
- Basimycetes (Базидиомицеты)
- Лихены [Vol. 1, стр. 325]
- Diaphyta [Vol. 1, стр. 330]
- Bryophyta [Vol. 1, стр. 336]
- Pteridophyta
- Anthophyta
- Gymnospermae
- Покрытосемянное
Thieren
- Protozoa (Protista plasmophaga) [Vol.1, стр. 138]
- Архезоа (= Зоарчега)
- Бактерии (= Bactromonera)
- Coccillida
- Bacillida
- Спириллида
- Зоомонера (= ул. Монера)
- Грибки (= Sporozoa)
- Ризопода
- Инфузории
- Metazoa (Гистоны плазмофаги)
- кишечник (акеломия)
- Gastraeades
- Губки
- Книдария
- Платоды
- Билатерия (Coelomaria)
- Гельминты
- Моллюска
- Сочленений
- Иглокожие
- Туниката
- Vertebrata
Levander (1894) [править]
Левандер, К.М. (1894). Materialien zur Kenntnis der Wasserfauna in der Umgebung Helsingfors, mit besonderer Berücksichtigung der Meeresfauna. I: простейшие. Acta Societatis pro Fauna et Flora Fennica 12 (2): 3–155, 3 табл., 1 таблица, [18].
Простейшие
Блохманн (1895) [править]
Блохманн, Ф. (1895). Die mikroskopische Tierwelt des Süßwassers. Abt. Я, простейшие. В: Кирхнер О. и Блохманн Г., Die mikroskopische Pflanzen- und Tierwelt des Süßwassers .2. Aufl., Teil II. Гамбург: Gräfe & Sillem, [19].
Простейшие
Delage & Hérouard (1896) [редактировать]
Traité de zoologie concrète , t. 1, [20].
Kalberlah & Schoenichen (1900) [править]
Kalberlah, A. & Schoenichen, W. B. (1900). Эйферта Einfachste Lebensformen des Tier- und Pflanzenreiches: Naturgeschichte der mikroskopischen Süsswasserbewohner . 3-е изд. Брауншвейг: Б. Гериц, [21].
- Pflanzliche Organismen
- Tierische Organismen
Дофлейн (1901) [править]
Дофлейн, Ф.(1901). Die Protozoen als Parasiten und Krankheitserreger , 1-е изд. Дополнительные издания: Lehrbuch der Protozoenkunde .
Дофлейн (1902) [править]
Дофлейн, Ф. (1902). Das System der Protozoen. Archiv für Protistenkunde 1: 169–192, [22].
Lankester et al. (1903, 1909) [править]
Ланкестер, Э. Р. (ред.) Трактат по зоологии , т. 1, фас. 1 (1909), фас. 2 (1903), [23].
Простейшие
Хартог (1906) [править]
Хартог, М.(1906). Простейшие. В: Кембриджская естественная история . Vol. 1, Protozoa, Porifera, Coelenterata и Echinodermata. (ред. С. Ф. Хармера и А. Э. Шипли). Лондон: Macmillan, [24].
Простейшие
Дофлейн (1911) [править]
Из Lehrbuch der Protozoenkunde , 3-е изд., 1911 г. (1-е изд. Под названием Die Protozoen als Parasiten und Krankheitserreger , 1901).
Простейшие
Минчин (1912) [править]
Введение в изучение простейших со специальной ссылкой на паразитические формы , 1-е изд., [25].
Простейшие
Люэ (1913) [редактировать]
Из Люэ, М. 1913: Erstes Urreich der Tiere. В: Lang A, редактор. Handbuch der morphologie der wirbellosen Tiere. Йена: Г. Фишер. 416 с.
Простейшие
Поче (1913) [править]
Poche, F. 1913. Das System der Protozoa. Archiv für Protistenkunde 30: 125–321, [26].
Phylum Protozoa Goldfuss, 1817 г.
Hartmann (1919) [редактировать]
От Хартманна, М.»Allgemeine Morphologie und Physiologie der Protozoen». В: Lehrbuch der Mikrobiologie . Фридбергер, Э; Pfeiffer, R. [ред.] G. Fischer, 1919, [27], [28].
Простейшие
Calkins (1926) [править]
Из Биология простейших .
Тип Простейшие
Веньон (1926) [править]
Веньон, К. М. 1926. Протозоология, руководство для врачей, ветеринаров и зоологов . Байер, Тиндаль и Кокс: Лондон; Нью-Йорк: W.Дерево. Тт. 1 и 2, [29].
Тип простейших
Sandon (1927) [редактировать]
Сандон, Х., 1927. Состав и распространение фауны простейших в почве . Лондон: Оливер и Бойд, [30].
Простейшие
Уэйлс (1937-1943) [править]
Уэйлс, Г. (1937). Тихоокеанская фауна Канады: 1. Простейшие. 1а. Лобоса; lb. Reticulosa; 1c. Heliozoa; 1г. Радиолярии. с. 1-14. Торонто: Univ. Toronto Press, Совет по исследованиям рыболовства Канады.
Wailes, G.H. (1939). Тихоокеанская фауна Канады. 1. Простейшие. 1e. Мастигофора. pp. 1-45, Fig. 138. Toronto: Univ. Toronto Press, Совет по исследованиям рыболовства Канады.
Wailes, G.H. (1943). Тихоокеанская фауна Канады: 1. Простейшие. 1f. Реснички; 1гр. Суктория. с. 1-46. Торонто: Univ. Toronto Press, Совет по исследованиям рыболовства Канады.
Простейшие
Баркли (1949) [править]
Баркли, Фред А. «Un esbozo de clasificación de los organos.» Revista de la Facultad Nacional de Agronomia , Universidad de Antioquia, Медельин. 10: 83–103, [31].
- Mychota (Monera, Anucleobionta)
- Протиста
- Euthallophyta
- Родофита
- Хризофита
- Pyrrophyta
- Euglenophyta
- Gonidiophyta
- Charophyta
- Phaeophyta
- Myxophyta
- Микофита
- Простейшие
- Sarcodina
- Фораминиферы
- Актиноподы
- Грибки
- Реснички
- Suctoria
- Мезозоа
- Паразоа
- Euplanta
- Euanimalia
Jahn & Jahn (1949) [править]
От Яна, Т.Л. и Ф. Ф. Ян, 1949. Как узнать простейшие . Браун, Dubuque, lowa.
Grassé et al. (1952-1994) [править]
Из Грассе, П.-П. (Ред.), 1952–1994. Traité de Zoologie . Vol. 1, пт. I, Protozoaires: Flagellés , 1952. Vol. 1, пт. II, Простейшие: Rhizopodes, Actinopodes, Sporozoaires, Cnidosporidies , 1953. Vol. 2, пт. I, Infusoires ciliés , 1984. Vol. 2, пт. II, Infusoires ciliés , 1994. Masson, Paris.См. Фернандес-Галиано (1964), [32].
Règne animal
Холл (1953) [править]
Из Холла, Р.П., 1953. Протозоология . Прентис-Холл, Нью-Йорк, [33]
Простейшие
Pennak (1953) [редактировать]
Пеннак Р.В. 1953. Пресноводные беспозвоночные США . 1-е изд. Рональд, Нью-Йорк, [34].
Тип простейших
Кудо (1954) [редактировать]
Из Протозоология , 4-е изд.
Тип Protozoa Goldfuss
Эдмондсон (1959) [править]
Эдмондсон, W.T. (ред.). 1959. Пресноводная биология . 2-е изд. Вили, Нью-Йорк, [35].
Протиста
Protozoa Images, Stock Photo & Vectors
В настоящее время вы используете более старую версию браузера, и ваша работа может быть не оптимальной. Пожалуйста, подумайте об обновлении. Учить больше. ImagesImages homeCurated collectionsPhotosVectorsOffset ImagesCategoriesAbstractAnimals / WildlifeThe ArtsBackgrounds / TexturesBeauty / FashionBuildings / LandmarksBusiness / FinanceCelebritiesEditorialEducationFood и DrinkHealthcare / MedicalHolidaysIllustrations / Clip-ArtIndustrialInteriorsMiscellaneousNatureObjectsParks / OutdoorPeopleReligionScienceSigns / SymbolsSports / RecreationTechnologyTransportationVectorsVintageAll categoriesFootageFootage homeCurated collectionsShutterstock SelectShutterstock ElementsCategoriesAnimals / WildlifeBuildings / LandmarksBackgrounds / TexturesBusiness / FinanceEducationFood и DrinkHealth CareHolidaysObjectsIndustrialArtNaturePeopleReligionScienceTechnologySigns / SymbolsSports / RecreationTransportationEditorialAll categoriesEditorialEditorial главнаяРазвлеченияНовостиРоялтиСпортМузыкаМузыка домойПремиумBeatИнструментыShutterstock EditorМобильные приложенияПлагиныИзменение размера изображенияКонвертер файловСоздатель коллажейЦветовые схемыБлогГлавная страница блогаДизайнВидеоКонтроллерНовости
PremiumBeat blogEnterpriseЦена ing Войти
Зарегистрироваться
Меню
ФильтрыОчистить всеВсе изображения- Все изображения
- Фото
- Векторы
- Иллюстрации
- Редакционные
- Музыка
- Видеоматериалы
8
- Поиск по изображению
90azo Самые актуальные
Свежий контент
Тип изображения Все изображения
Фото
Векторы
Иллюстрации
Ориентация Все ориентации
Горизонтально
Определение, типы, болезни, профилактика и многое другое
Патогены — это организмы, которые могут вызывать болезнь.Различные типы патогенов и тяжесть вызываемых ими заболеваний очень разнообразны.
В этой статье мы рассмотрим различные патогены, их влияние на людей и болезни, которые они вызывают. Мы также объясняем, как распространяются патогены и как снизить риск заражения.
Патоген приносит болезнь своему хозяину. Другое название возбудителя — инфекционный агент, поскольку они вызывают инфекции. Как и в случае с любым организмом, патогены отдают приоритет выживанию и воспроизводству.
Иммунная система человеческого организма действует как защита от патогенов.Организм может легко бороться с некоторыми патогенами, но другие потенциально смертельны.
Существует пять основных типов патогенов:
Бактерии
Бактерии — это микроскопические патогены, которые быстро размножаются после попадания в организм. Они могут выделять токсины, которые повреждают ткани и вызывают болезни.
Врачи обычно назначают антибиотики для лечения бактериальных инфекций, но некоторые бактерии становятся устойчивыми к этим лекарствам.
Однако не все бактерии являются патогенными.В организме существует множество типов безвредных бактерий, некоторые из которых могут даже поддерживать основные функции организма.
Вирусы
Вирус, который меньше бактерий, проникает в клетку-хозяина. Затем он реплицируется, производя сотни и тысячи новых вирусов, которые продолжают заражать другие клетки-хозяева. Вирусы могут передаваться от человека к человеку различными путями, в том числе:
- через респираторные капли, которые перемещаются по воздуху
- через контакт с кровью человека с инфекцией
- через контакт с жидкостями организма человека, инфицированного инфекцией
Грибы
Существуют тысячи видов грибов, некоторые из которых вызывают болезни у людей.Распространенные грибковые заболевания кожи включают микоз стопы и стригущий лишай. Эти состояния заразны и могут передаваться от человека к человеку.
Исследование, проведенное в Trends in Microbiology , показало, что у грибковых патогенов развивается способность к памяти. Они могут использовать сигналы в теле, чтобы предвидеть неминуемую угрозу своему выживанию, к которой они затем могут подготовиться.
Протисты
Эти одноклеточные организмы вызывают болезни у своего хозяина. Они заражают другие организмы, чтобы выжить и размножаться.
Простейшие патогены поражают растения и продовольственные культуры. Продукты, содержащие простейшие, могут вызвать дизентерию — инфекцию кишечника, вызывающую диарею.
Протистские патогены также могут быть паразитическими и жить в других организмах, таких как комары. Протисты вызывают малярию через укусы комаров.
Паразитические черви
Паразитические черви, также известные как гельминты, достаточно велики, чтобы их можно было увидеть невооруженным глазом, и они могут жить во многих частях тела.Некоторые черви включают:
- Плоские черви: К ним относятся ленточные черви, которые обитают в кишечнике.
- Колючие черви: Этот вид червей обитает в кишечнике.
- Круглые черви: Эти черви могут выжить в желудочно-кишечном тракте и лимфатической системе.
Патогены могут вызывать множество различных заболеваний, некоторые из которых более серьезны, чем другие. Человеческие тела богаты питательными веществами и могут обеспечить патогену идеальную среду для роста и размножения.
Степень тяжести инфекций, вызываемых патогенами, может быть разной. Некоторые инфекции могут быть легкими, в то время как другие могут быть опасными для жизни. Например, простуда — это легкая вирусная инфекция по сравнению со смертельной болезнью, вызванной вирусом Эбола.
Болезни, вызываемые бактериальными патогенами, включают:
Некоторые ученые считают, что вирусы не являются живыми организмами. Вот некоторые причины:
- у них нет клеток
- они не могут воспроизводиться, не вторгаясь в живую клетку
- они не реагируют активно на изменения в окружающей среде
К вирусным инфекциям относятся:
Около 300 видов грибов патогенны для человека.Как и бактерии и вирусы, они могут оказывать значительное влияние на здоровье человека.
Грибки вызывают множество различных типов заболеваний, в том числе:
- астму
- инфекции кожи и ногтей
- инфекции легких, такие как пневмония
- инфекции кровотока
- менингит
Протозойные заболевания являются причиной большинства протистических заболеваний. Простейшие — это одноклеточные микроорганизмы, которые питаются другими микроорганизмами, органическими тканями и мусором.К протистским заболеваниям относятся:
- дизентерия
- малярия
- Африканский трипаносомоз или сонная болезнь
Паразитические черви вызывают множество болезней, в том числе:
Патогены могут распространяться различными путями. Например, прямой контакт кожа к коже во время секса может привести к инфекциям, передаваемым половым путем (ИППП).
При кашле или чихании болезнетворные микроорганизмы могут распространяться через крошечные капельки в воздухе. Эти капли могут содержать микроорганизмы, которыми дышат другие люди.
Микроорганизмы также могут попадать прямо в кишечник, когда человек употребляет зараженную пищу или воду.
Укусы инфицированных насекомых также могут распространять болезнь. Например, клещи с бактериальной инфекцией могут вызвать болезнь Лайма, если кого-то укусили, а комары с вирусной инфекцией могут вызвать болезнь, вызванную вирусом Зика.
Наряду с поддержанием хорошего общего состояния здоровья человек может предпринять определенные шаги, чтобы снизить риск заражения патогенами. К ним относятся:
- Регулярное мытье и сушка рук: Регулярное мытье рук имеет решающее значение для предотвращения распространения болезней.Люди должны мыть их не менее 20 секунд водой с мылом или дезинфицирующим средством на спиртовой основе. Это особенно важно после занятий на открытом воздухе, прикосновений к животным, посещения туалета, ухода за больными, чихания или кашля.
- Будьте в курсе рекомендованных вакцин: Вакцины против инфекционных заболеваний, таких как грипп, коклюш и корь, доступны для младенцев, детей и взрослых. Вакцины стимулируют иммунную систему распознавать патогены, уничтожать их и предотвращать будущие инфекции.
- Поддержание чистоты поверхностей: Загрязненные поверхности или предметы могут передавать болезни. Исследование, проведенное в аэропорту Финляндии, обнаружило вирусное заражение в нескольких местах на поверхностях, к которым люди часто прикасаются.
- Уборка кухонь и ванных комнат: Важно поддерживать в чистоте столешницы и кухонные поверхности, особенно перед приготовлением еды. В ванной также может быть высокая концентрация инфекционных агентов.
- Оставаться дома в случае болезни: Люди с инфекциями должны избегать посещения школы, работы и тесного контакта с другими людьми, чтобы снизить вероятность распространения патогена.
- Как избежать укусов насекомых: Люди могут сделать это, надев защитные средства от насекомых и подходящую одежду, такую как рубашки с длинными рукавами, брюки и шляпы, в местах, где они могут встретить насекомых.
- Практика безопасного секса: Использование методов безопасного секса, включая использование презервативов и регулярные проверки сексуального здоровья, может снизить вероятность заражения ИППП.
- Получение медицинской консультации: Человек может поговорить с медицинским работником, если есть признаки инфекции или лечение не работает.
Патогены поражают все живые организмы и вызывают заболевания у людей множеством различных способов.
Они могут повредить ткани или клетки, вырабатывая токсины в процессе репликации.
В то время как некоторые патогены вызывают легкие проблемы, другие могут быть опасными для жизни. Медицинские эксперты продолжают изучать болезни, вызываемые патогенами, чтобы больше узнать об их причинах и способах их лечения.
Паразиты — простейшие
Простейшие теперь имеют свой собственный порядок в живом царстве, хотя когда-то их считали крошечными животными, поскольку они были подвижными и не были зелеными, как растения — теперь у них есть отдельное царство в соответствии со схемой классификации для всей жизни на Земле.Многие простейшие паразиты на человеке и вызывают от легких до чрезвычайно опасных заболеваний — некоторые из них паразитируют на человеке и из-за своего крошечного размера образуют простейших из всех внутренних паразитов. Клеточная организация простейших намного сложнее, а размер намного больше, хотя, как и бактерии, все простейшие являются одноклеточными организмами — весь организм состоит только из одной клетки. Тип окружающей среды, в которой обитает большинство простейших, очень космополитичен, и их распространение повсеместно, они встречаются почти в любой водной среде и во влажных местах, включая пруды, реки, озера, аквариумы, канавы и т. Д.
Мазь для ногтей
Лучшее, 100% натуральное средство для ежедневного ухода за ногтями в идеальной форме.
Большинство простейших не причиняют вреда животным или людям и считаются свободноживущими, то есть им не требуется хозяин для жизни и размножения. Однако есть несколько простейших, которые являются паразитами, среди этих паразитических организмов некоторые могут вызвать чрезвычайно серьезные проблемы со здоровьем у людей, которым не повезло заразиться простейшими.Первым патогенным простейшим, идентифицированным еще в 1875 году, было простейшее. Паразитические простейшие были разновидностью амебы под названием Entamoeba histolytica, которая также известна как дизентерия, вызывающая амебу — она очень распространена и поражает людей во всем мире. Антон ван Левенгук, один из первых и, возможно, один из величайших ранних микроскопистов и клеточных биологов, заметил, что в его собственных фекалиях скользят простейшие. Крошечная микроскопическая форма жизни принадлежала к роду Giardia, и с момента открытия Левенгука такие паразиты, как Giardia lamblia и простейшие виды Cryptosporidium, были связаны с причиной болезненных кишечных инфекций и вспышек тяжелой диареи во всем мире.Вода, загрязненная фекалиями человека или животных, обычно содержит этих двух паразитических простейших, и употребление такой загрязненной воды является обычной причиной заболеваний.
Мазь от грибка
Вся сила фармацевтических фунгицидов — но без агрессивных химикатов.
Бабезиоз
Паразитические простейшие, известные как Babesia microti, ответственны за вспышки этого заболевания. Паразит проникает в эритроциты различных животных, которых заражает, и вызывает типичные симптомы, столь характерные для этого состояния.Основным резервуаром болезни являются грызуны, способ передачи болезни — через клещей, возможно, тех же видов, которые участвуют в передаче болезни Лайма — таким образом, клещи являются основными переносчиками болезни и распространяют ее. от грызунов-хозяев к другим животным, включая человека, путем укуса. Распространенное заболевание, называемое техасской лихорадкой, иногда также известное как клещевая лихорадка крупного рогатого скота, которое поражает крупный рогатый скот, вызывается простейшими, связанными с этим видом.
Состояние вызывает симптомы у пораженных людей, все эти симптомы, по-видимому, связаны или похожи на симптомы, наблюдаемые у пациентов, страдающих малярией, которая также вызывается простейшими.Физические симптомы включают стойкую усталость и физическую усталость в течение семи дней после первоначального контакта с паразитом, за этим начальным периодом следует период, когда у пациента появляется внезапный озноб, а также другие состояния, такие как лихорадка, сильная головная боль и болезненность мышц в целом. тело. Некоторые больные часто прогрессируют до анемии и почечной недостаточности, хотя большинство симптомов легкие, и состояние в основном проходит через несколько недель. Однако известны случаи смертельного исхода в некоторых случаях, особенно в отношении пациентов старших возрастных групп.Для этого состояния не существует специальных лекарств, хотя симптоматическое лечение с использованием тех же лекарств, которые используются для лечения малярии, было опробовано с некоторым успехом — профилактика является лучшей политикой в отношении этого заболевания, и случаи заболевания редки.
Болезнь Шагаса
Простейшее заболевание Темы — болезнь Шагаса — серьезная проблема для здоровья в Латинской Америке. За эти состояния отвечает простейший организм, известный в науке как Trypanosoma cruzi, болезнь широко распространена в Центральной и Южной Америке, где она представляет собой проблему для общественного здравоохранения.Переносчиком этой болезни является насекомое, известное как целующиеся клопы, иногда называемое «конусообразным насекомым». Люди, живущие в районах с маргинальной жилой инфраструктурой, зараженной этим насекомым, могут заразиться семейством болезни Шагаса. Насекомые-переносчики относятся к семейству насекомых Reduviidae, подсемейству Triatominae и роду Triatoma — это насекомое снова очень распространено в Южной Америке. Опоссумы и броненосцы, а также инфицированные люди служат резервуарами для простейших, насекомое, которое является кровососом, передает паразита из таких источников неинфицированным животным и людям — инфекции обычно распространяются среди людей в одной семье — насекомое передает паразита от хозяин за хозяином, поскольку он сосет кровь и оставляет отходы на новых хозяевах.
Восстанавливающее средство для кожи
Усовершенствованный, 100% натуральный ревитализатор, который сделает вашу кожу сияющей и молодой.
После того, как клоп поедает инфицированное млекопитающее, он передает простейшие, которые уже прошли по его кишечнику, с отходами, которые он оставляет на коже неинфицированного животного-хозяина или человека. Когда человек непроизвольно царапает эту область, отходы в области укусов насекомых загрязняют рану, и повторяющиеся царапины вызывают ушибы кожи — простейшие также часто передаются через глаза — когда человек трет отходы, простейшие попадают в тело, используя сломанный участок кожи.Тропическое заболевание было идентифицировано в 1912 году бразильским врачом Карлосом Шагасом — отсюда и название болезнь Шагаса.
Появление небольшой красной язвочки на коже или развитие опухших и покрасневших, воспаленных глаз являются наиболее частыми ранними симптомами заболевания у человека. Через несколько недель язва на пораженном участке кожи, кажется, заживает и выделяет твердую корку, которая вскоре исчезает, и остается небольшой темный шрам в этой области — если человеку повезет. В других случаях первоначальные симптомы могут сменяться появлением лихорадки, развитием отека лимфатических узлов и узлов, а также развитием кожной сыпи по всему телу.Тяжелые формы болезни Шагаса могут поражать сердце, и такие симптомы, как боли в груди и внезапная одышка, становятся обычным явлением. Также могут возникнуть судороги, если заболевание поражает нервную систему — в таких случаях в непрерывных конвульсиях может быть вовлечено все тело. В некоторых случаях это состояние может привести к летальному исходу — это остается риском для здоровья.
Лямблиоз (бобровая лихорадка)
Эта лихорадка является паразитарным заболеванием человека, которое является наиболее распространенной формой паразитарного заболевания в некоторых частях Соединенных Штатов и других районах Северной Америки, в частности, в западных регионах Канады.Причина лихорадки — крошечный паразитический простейший организм, который называется Giardia lamblia. Паразитический организм остается на внутренней поверхности кишечника человека и, в частности, в больших количествах обнаруживается в тонком кишечнике пораженных людей. Обычным физическим симптомом, помимо повышения температуры тела, является стойкая инфекционная диарея.
Обычный источник распространения паразита — употребление зараженной пищи и воды. Если не поддерживать строгие и надлежащие санитарные условия, многие детские центры и дома престарелых становятся районами, в которых часто происходят вспышки лямблиоза, поэтому дети становятся одними из первых, кто становится жертвами паразита.Путь передачи паразита в большинстве случаев заражения происходит через попадание паразитарных цист в пищеварительный тракт вместе с зараженной пищей или водой — такие источники обычно содержат фекальные отходы людей и других животных-переносчиков.
Примерно через три недели после первоначального заражения паразитом хроническая диарея начинает поражать людей, страдающих лямблиозом — это обычные временные рамки для появления этого симптома у большинства пациентов.В дополнение к этому симптому у пораженных людей также появляются симптомы тошноты и другие жалобы, такие как стойкий и сильный дискомфорт в животе. Пищеварительная система нарушается во время длительных периодов инфекции, конечным результатом может быть внезапная потеря веса у пораженного человека — это происходит из-за очень плохого всасывания переваренной пищи из области кишечника. Человек может оставаться инфицированным в течение нескольких лет подряд, хотя в некоторых случаях у пораженных людей начинается спонтанное выздоровление от симптомов.
Раньше считалось, что основным резервуаром простейших паразитов в дикой природе являются бобры — так считалось в течение многих лет, пока недавние доказательства не показали обратного. В настоящее время известно, что домашний скот и овцы являются основными резервуарами болезни, согласно данным недавних исследований. Сейчас считается, что бобры, как и люди, вероятно, могут быть вторичным хозяином, а не резервуаром для паразита. Было показано, что такие животные, как бобры, полевки, ондатры и крысы-кенгуру, обладают способностью переносить этого паразита — все они могут действовать как хозяева, похожие на людей.
Сонная болезнь (африканский трипаносомоз)
Мелкая рыба, такая как простейшие паразиты, вызывает опасное заболевание, известное как сонная болезнь — это смертельное заболевание, широко распространенное в некоторых районах Африки. Простейшие — это трипаносома — разновидность нематодных червей, этот червь может плавать в крови своих жертв, ударяя хлыстом, как хвостом, или, точнее, своим жгутиком. Печально известная муха цеце несет ответственность за передачу этого смертельного заболевания. Муха кусает и сосет кровь, как только паразит попадает в организм.Человек медленно теряет сознание, и смерть наступает, если паразиту удается проникнуть в области спинного и головного мозга своих жертв. Шансы умереть от болезни очень высоки, если человек инфицирован, а сонная болезнь, как известно, уничтожает все домашние запасы мяса в районе — поскольку этот паразит также заражает домашний скот, например, крупный рогатый скот. Смертность как среди людей, так и среди животных высока в районах, где болезнь носит эндемический характер, и профилактика является лучшим вариантом против нее.Среда обитания мухи цеце широко распространена в тропической Африке, и некоторые ученые и эксперты в области здравоохранения считают, что с повышением температуры из-за глобального потепления существует возможность значительного расширения этой среды обитания. В таком случае муха цеце, как и паразит, может заразить космополитичную территорию. Это не единственный наихудший сценарий для одного насекомого-переносчика; Обычно считается, что глобальное потепление может расширить ареал обитания других насекомых-переносчиков, а также привести к распространению большего количества тропических паразитов в другие регионы мира.
Лейшманиоз
Паразитические простейшие лейшмании ответственны за это изнурительное и часто смертельное заболевание, которое широко распространено в некоторых тропических и субтропических регионах мира. Крошечная кровососущая песчаная муха ответственна за распространение этого смертоносного паразитического простейшего — эта муха является единственным переносчиком паразита. Состояние, известное как висцеральный лейшманиоз, иначе известное как кала-азар, является наиболее летальной стадией прогрессирования инфекции. В эндемичных районах болезнь способна уничтожить две трети населения некоторых серьезно пострадавших деревень — это особенно верно в отношении сельских районов в тропиках, где медицинская помощь плохая или полностью недоступна.Постоянная лихорадка, темно-серая кожа, наличие тяжелой анемии — одни из первых физических симптомов кала-азар, кроме того, сильно увеличиваются селезенка и печень, что приводит к неприглядному вздутию живота. область. Болезнь может стать смертельной очень быстро, если симптомы остаются без лечения в течение длительного периода времени. Кала-азар обычно лечится с помощью лекарств, содержащих сурьму тяжелых металлов — такие лекарства являются единственным эффективным средством против паразита.Белые кровяные тельца в организме — это область, в которой простейшие лейшмании фактически проживают в организме — это защитные клетки, которые должны были поглотить простейшие, когда началась инфекция. Белые кровяные тельца поглощают простейшее, но не могут его разрушить или переварить. В результате простейшие лейшмании процветают в тех самых белых кровяных тельцах, от которых зависит его собственная защита. Лейкоциты, в которых проживают простейшие, со временем разрушаются, подобно заболеванию, подобному ВИЧ, что, естественно, наносит огромный ущерб иммунной системе больного человека.Оппортунистические и смертельные бактериальные инфекции, такие как пневмония или дизентерия, преодолевают слабую и неэффективную защиту организма, так как лейкоциты в большом количестве умирают из-за простейших. Если это продолжается, человек умирает от симптомов.
Малярия
Настоящее значение слова «малярия» — просто «плохой воздух». Этот плохой воздух должен был вызывать у людей перемежающуюся лихорадку, когда они впервые столкнулись с этой болезнью. Действительно, древнегреческий врач Гиппократ предположил, что это было вызвано появлением миазмов на человеке — это смертельные туманы, которые предположительно нарушили тщательный баланс, существующий в четырех жидкостях тела, — что является древним медицинским верованием в кровь. мокрота, черная желчь и желтая желчь.С другой стороны, согласно древней китайской медицинской системе, дисбаланс между инь и ян в организме должен был вызвать приступообразную лихорадку, а именно малярию, а дисгармония между этими двумя противоположными мужскими и женскими жизненными силами в организме приписывалась быть причиной состояния. Несколько столетий назад великий систематик и биолог Карол Линней выдвинул гипотезу о том, что причиной малярии может быть присутствие крошечных взвешенных частиц глины в питьевой воде, эти взвешенные твердые частицы, как предполагается, привели к закупорке кровеносных сосудов и нарушение кровообращения должно было вызвать болезнь.Все это были теории о причине малярии, выдвинутые до появления микроскопа и в самом начале современной науки, истинный возбудитель малярии был идентифицирован только в 1898 году — британский доктор Росс правильно объяснил бы. Это произошло с укусом инфицированного комара, и он продолжил идентифицировать смертельный патоген как тип простейших.
Только укус самок комаров вида anopheles ответственен за передачу малярии людям и некоторым другим млекопитающим, которые могут быть инфицированы.Самки анофелесов — единственные москиты, которые являются переносчиками малярии человека; самцы комаров не кусаются и не могут распространять болезнь. Когда самка комара Anopheles кусает свою жертву, она высасывает кровь через прокол в коже особи и в то же время вводит тысячи нитевидных личинок простейших, называемых спорозоитами — эти личинки принадлежат к видам простейших плазмодиев. Оказавшись в теле жертвы, спорозоиты отправятся через кровоток прямо в печень, где каждая из личинок начинает инфицировать и жить в собственной клетке печени.В этих клетках печени спорозоиты собираются и начинают быстро делиться, многократно размножаясь, производя до сорока тысяч спор на каждую личинку. По прошествии двух недель производство и размножение спор в клетках печени продолжается, в течение этого периода нет абсолютно никаких признаков или симптомов болезни, и человек здоров. Однако здесь закладываются первые семена полномасштабной малярии, которые начинают вторжение в организм.
Разрыв клетки печени, в которой образовались споры, вызывает первые клинические признаки и является начальной стадией атаки на организм — у человека наблюдается сильное потоотделение и на этой стадии развивается высокая температура.Теперь лопнувшие клетки печени начинают выпускать многочисленные споры прямо в кровоток жертвы. На этом этапе каждая отдельная спора теперь вторгается в эритроцит, где она будет находиться какое-то время, медленно расходуя содержание гемоглобина в клетке и увеличиваясь в размере. Затем спора делится или раскалывается на множество более мелких фрагментов — это бесполое средство образования тысяч других спор — научное название спор на этой стадии — мерозоиты. Наконец, воля красных кровяных телец также лопается, и это высвобождает все присутствующие в них мерозоиты, а затем вторгаются в новые клетки крови по соседству.Этот циклический эффект, а также повторяющееся инфицирование и разрушение красных кровяных телец в конечном итоге приводят к хроническому малярийному заболеванию и могут привести к смерти человека.
Разрыв и вторжение красных кровяных телец — это очень своевременное дело, и существует удивительная синхронность всего процесса заражения в кровотоке жертвы. Основные причины одновременного разрыва миллионов инфицированных эритроцитов с высвобождением их мерозоитов до сих пор полностью не изучены и требуют изучения.Худшие приступы тряски и озноба происходят из-за своевременного и массового разрыва красных кровяных телец в организме.
Окончательный процесс трансформации этих мерозоитов в мужские или женские гаметоциты также начинает происходить после нескольких раундов инфицирования клеток крови и их последующего разрушения — эти гаметоциты являются репродуктивными клетками паразита плазмодия, а на этой стадии инфекции — человеческих клеток. фазу жизненного цикла паразита можно считать завершенной.Если пациенту повезет, он или она выжили бы на этой стадии, однако на этой стадии он заразен, потому что, если самка комара Anopheles высосет эту зараженную малярией кровь, она также поглотит миллионы паразитических клеток гаметоцитов. Гаметоциты простейших будут претерпевать дальнейшее развитие в яйцеклетки и сперматозоиды в теле самки комара, начиная следующую стадию у их нового хозяина, и оплодотворение этих гамет будет происходить в теле насекомого.
Цикл завершается, когда оплодотворенные яйца паразита превращаются в нитевидные спорозоиты в теле комара, а затем они попадают в слюнные железы комара, готовые для передачи новому человеку-хозяину.Именно в это время самка комара становится смертельным переносчиком малярийного паразита, и ее укус переносит новые спорозоиты, чтобы начать новый цикл внутри другого человека-хозяина.
Очень интересный факт заключается в том, что люди с серповидноклеточной анемией и даже те, кто страдает ее более легкой формой, или те, у кого есть черта серповидно-клеточной анемии в красных кровяных тельцах, вероятно, имеют гораздо большую защиту от малярии. предлагает. Серповидно-клеточная анемия деформирует красные кровяные тельца тела, и это изменение формы клеток напрямую препятствует и предотвращает проживание простейших внутри клетки.Это эволюционный ответ на малярию, поскольку люди, страдающие серповидно-клеточной анемией, как правило, в основном сконцентрированы в регионах мира с самым высоким уровнем заражения малярией — серповидно-клеточная форма красных кровяных телец является только общим генетическим дефектом в человеческих популяциях, круглый год подверженных малярийным заболеваниям.
Итак, как именно лечат малярию? И какое оружие применяется против болезни. Лекарства и противомалярийные препараты являются одними из инструментов, с помощью которых врачи и ученые непосредственно лечат малярию. Болезнь также можно контролировать, регулируя популяцию насекомых-переносчиков.Цитата «Лечите пациента, а не комара» часто использовала Роберт Кох, блестящий микробиолог, получивший известность благодаря выявлению туберкулеза. Единственным противомалярийным лекарством, которое было доступно в его дни — а это было в конце девятнадцатого века, было лекарство растительного происхождения, известное как хинин.
Противомалярийный препарат хинин — это медицинский препарат, полученный из коры хинного дерева, он веками использовался в традиционных противомалярийных средствах туземцами Южной Америки.На сегодняшний день он все еще используется как отличное противомалярийное лекарство во многих частях мира. К сожалению, высокие дозировки препарата токсичны и могут вызвать глухоту при использовании в высоких дозах, необходимых для полного удаления паразита плазмодия из организма. До получения менее вредных производных в 1930-х годах его использование было обширным и универсальным — теперь его заменяют менее вредные производные, синтезированные из того же растения. Хлорохин — это основное синтезируемое лекарство, используемое в наши дни для лечения малярии.Однако в качестве препарата выбора при лечении малярии хлорохин в большинстве стран мира был вытеснен другим противомалярийным мефлохином.
Другие полезные травы
Мазь для кожи
100% натуральная формула для всех проблем с кожей. Отлично подходит для диабетиков.
комментариев
Frontiers | Роль инфузорий простейших в рубце
Введение
Впервые описанные в 1843 году, простейшие Rumen с их поразительной внешностью считались важными для благополучия своего хозяина.Однако, несмотря на то, что простейшие могут составлять до 50% биомассы в рубце, роль простейших в микробной экосистеме рубца остается неясной (Williams and Coleman, 1992). Здесь мы оцениваем недавнюю информацию о роли простейших инфузорий в микробной экосистеме рубца.
Разнообразие и филогения простейших инфузорий рубца
С тех пор, как они были впервые обнаружены Gruby и Delafond (1843), исследования простейших в рубце основывались на морфологической идентификации с помощью оптической микроскопии.В настоящее время нет коллекций культур инфузорий рубца, поэтому исследователи должны использовать микрофотографии для идентификации (Ogimoto, Imai, 1981; Williams and Coleman, 1992) или штриховые рисунки (Dogiel, 1927; Kofoid and MacLennan, 1930, 1932, 1933; Latteur , 1968, 1969, 1970; Dehority, 1993). Широко признано, что микроскопическая идентификация и подсчет представляют собой золотой стандарт для анализа структуры сообщества инфузорий в исследованиях рубца (Williams and Coleman, 1992). Однако полиморфная природа этих микробов (Dehority, 1994, 2006) требует от исследователя высокого уровня опыта для идентификации инфузорий рубца на уровне родов или видов (Dehority, 2008).Таким образом, необходимы альтернативные способы идентификации простейших, поскольку микроскопические методы трудоемки и весьма требовательны.
Ранние молекулярные исследования простейших ресничек рубца были сосредоточены на секвенировании генов 18S рРНК для выяснения внутренней филогении класса Litostomatea (Embley et al., 1995; Wright et al., 1997). Embley et al. (1995) были первыми, кто описал монофилетическое положение анаэробных инфузорий рубца Dasytricha ruminantium и Polyplastron multivesiculatum на основе генов 18S рРНК, а Wright et al.(1997), Wright and Lynn (1997a, b) предоставили молекулярные данные для Diplodinium dentatum , Entodinium caudatum , Epidinium caudatum , Eudiplodinium maggii , Isotrichaintechinalis108 Oyinoscopusa , .
С тех пор были выполнены многочисленные качественные и количественные исследования разнообразия инфузорий; в этих исследованиях использовались различные методы, включая полиморфизм длины рестрикционных фрагментов (RFLP; Regensbogenova et al., 2004a, Tymensen et al., 2012b), денатурирующий градиентный гель-электрофорез (DGGE; Regensbogenova et al., 2004b, McEwan et al., 2005; de la Fuente et al., 2009; Belanche et al., 2010a, Kittelmann and Janssen, 2011), ПЦР в реальном времени (qPCR; Sylvester et al., 2004; Skillman et al., 2006; Belanche et al., 2010a, Kittelmann and Janssen, 2011), флуоресцентная гибридизация in situ (FISH; Xia et al. ., 2014) и секвенирование следующего поколения (Kittelmann et al., 2013, 2015; Ishaq, Wright, 2014; Moon-van der Staay et al., 2014).
Микроскопия имеет несколько преимуществ перед молекулярными методами на основе ПЦР для изучения простейших инфузорий. Во-первых, хотя подавляющее большинство кишечных инфузорий охарактеризовано морфологически, для многих наблюдаемых родов и видов отсутствуют эталонные последовательности гена 18S рРНК. Во-вторых, как обсуждается ниже, вариация числа копий генов рибосомной РНК в разных родах или в разных условиях роста может исказить наблюдаемые пропорции этих родов в образце (Medinger et al., 2010). Исследования с использованием обзоров генов 18S рРНК показывают очевидное более высокое разнообразие инфузорий, чем оценка с помощью обычных морфологических методов (Moon-van der Staay et al., 2014).
Как и другие инфузории, простейшие в рубце содержат два типа ядер: микроядро и макронуклеус. Микроядро имеет четко видимые хромосомы, диплоидно и синтезирует только следы РНК. Макронуклеус не содержит различимых хромосом, имеет во много раз больше диплоидного количества ДНК, делится амитотически и обеспечивает практически всю РНК, необходимую для вегетативной жизни клетки (Prescott and Murti, 1974).Чрезвычайно высокое число копий рДНК в макронуклеарном геноме инфузорий, как было обнаружено ранее (Gong et al., 2013), можно понять, учитывая жизненный цикл, большой размер клеток и быстрый рост этих организмов (Cavalier-Smith, 2005; Zhu et al., 2005; Godhe et al., 2008). В инфузориях рубца, в то время как небольшие простейшие, такие как Entodinium , имели тенденцию быть недостаточно представленными, более крупные простейшие, такие как Epidinium или Polyplastron , имели тенденцию быть чрезмерно представлены методом пиросеквенирования по сравнению с микроскопическим подсчетом (Kittelmann et al ., 2015). Аналогично, Epidinium caudatum , который примерно в пять раз больше по объему, чем Entodinium caudatum (Dehority, 1993), также имеет примерно в пять раз больше копий гена 18S рРНК, закодированных в его геноме (Sylvester et al., 2009). Эти результаты согласуются с выводом о том, что методы на основе ПЦР дают более низкие оценки численности мелких Entodinium spp. (Tymensen et al., 2012a) и завышенные оценки численности, например, Polyplastron spp.(Исхак и Райт, 2014). Знание числа копий и вариаций последовательностей рДНК в отдельных эукариотах важно как для интерпретации обзоров разнообразия на основе рДНК, так и при использовании 18s рДНК для количественной оценки биомассы простейших. (Crosby, Criddle, 2003; Thornhill et al., 2007; Herrera et al., 2009; Amaral-Zettler et al., 2011). На сегодняшний день не опубликовано исследований, посвященных количеству копий на клетку и вариациям рДНК у простейших в рубце. Это особенно верно, поскольку исследования штрих-кодирования и микробного разнообразия на основе рДНК с использованием высокопроизводительного секвенирования становятся все более популярными (Amaral-Zettler et al., 2009) и молекулярные инструменты, основанные на исследованиях маркерных генов, в настоящее время широко используются для изучения разнообразия других микробов (Schlötterer, 2004; Case et al., 2007; Langille et al., 2013). Исследования на инфузориях, не относящихся к рубцу, показали, что вариация числа копий рДНК между видами инфузорий и внутри них, подчеркивая сложность использования подхода на основе порядковых номеров рДНК для вывода об относительной численности микробных эукариотических клеток в образцах окружающей среды (Wintzingerode et al., 1997 ; Мединджер и др., 2010). Таким образом, новейшие методы, основанные на генах 18S рРНК, могут быть ненадежными при оценке α-разнообразия или относительной численности различных родов и видов в данной выборке, хотя они могут надежно определять тенденции относительной численности родов и видов между разными образцами (β- разнообразие). Необходимы дополнительные исследования, сравнивающие молекулярные и традиционные методы (de la Fuente et al., 2009; Tymensen et al., 2012b; Kittelmann et al., 2015).
Функциональные гены простейших инфузорий рубца
Хотя существуют многочисленные копии рДНК в макронуклеусах инфузорий, вполне вероятно, что только небольшая часть этих генов транскрипционно активна в инфузориях при любых данных условиях роста, как ранее было показано для др. Эукариот (Reeder, 1999).Исследования на Tetrahymena thermophila , Paramecium tetraurelia и Oxytricha trifallax выявили важную роль некодирующих РНК в событиях перестройки генома (Feng and Guang, 2013). У спиротрихозных инфузорий, таких как Euplotes , Stylonychia и Oxytricha , более 95% микроядерной ДНК удаляется с образованием макронуклеуса во время полового размножения (Swart et al., 2013), что является механизмом, позволяющим клетка адаптироваться во время стресса.Более того, в макронуклеусе оставшийся геном сильно фрагментирован, и эти фрагменты сортируются и переупорядочиваются под руководством транскриптов из родительского макронуклеуса с образованием генов, кодирующих белок (Nowacki et al., 2011). Ни один из этих процессов у простейших рубца до сих пор не изучен из-за трудностей получения полноразмерных геномов инфузорий рубца.
Несмотря на неоднократные попытки, оказалось невозможным поддерживать простейшие в рубце в аксенической культуре. Таким образом, было трудно окончательно установить роль простейших инфузорий в рубце и, в частности, разрушение волокон.Ранние исследования стремились выделить и охарактеризовать ферменты целлюлозы, гемицеллюлазы и ксиланазы из промытых препаратов простейших (Howard et al., 1960; Bailey et al., 1962; Clayet et al., 1992), однако присутствие как внеклеточных, так и внутриклеточных симбиотических бактерии в препаратах простейших затрудняли уверенность в том, что изолированная активность действительно имеет протозойное происхождение (Delfosse-Debusscher et al., 1979; Thines-Sempoux et al., 1980). Мы разработали методы клонирования и экспрессии генов инфузорий в фаге λ (Eschenlauer et al., 1998), используя FISH для подтверждения протозойной идентичности экспрессируемых генов (Newbold et al., 2005) и разрабатывая биоинформатические индексы для подтверждения инфузорийного происхождения генов (McEwan et al., 2000). Используя эти методы, мы и другие специалисты смогли выделить и охарактеризовать гены ряда простейших в рубце (McEwan et al., 1999; Newbold et al., 2005; Belzecki et al., 2007; Boxma et al., 2007). Сюда входит широкий спектр фибролитических ферментов, ряд из которых, как было обнаружено, содержит несколько доменов со связывающими доменами и предполагаемые химерные конструкции, которые наблюдаются, что свидетельствует о высокоразвитой фибролитической способности инфузорий рубца (Devillard et al., 1999, 2003; Takenaka et al., 1999, 2004; Wereszka et al., 2004; Bera-Maillet et al., 2005). Это наблюдение было подтверждено недавним метагеномным скринингом простейших глюкозидаз и эукариотических метатранскриптомов, который подтвердил, что в простейших рубца присутствует широкий спектр разнообразных гликозидгидролаз (Findley et al., 2011; Qi et al., 2011).
На основе крупномасштабного конструирования и анализа филогении более 4000 библиотек экспрессируемых последовательностей из инфузорий рубца Entodinium caudatum , Eudiplodinium maggii , Metadinium medium , Diploplastron affine 1010, ec Polyplastron affine 1010, ec , Isotricha prostoma , Isotricha Кишечник и Dasytricha ruminantium .Ricard et al. (2006) пришли к выводу, что имеются обширные доказательства горизонтального переноса генов (HGT; 148 из 3563 неизбыточных генов) от бактерий и архей в геномах ресничек рубца. Среди кандидатов на ГПГ они сообщили о чрезмерном представительстве (> 75%) генов, участвующих в метаболизме, в частности, в катаболизме сложных углеводов (Ricard et al., 2006), предполагая, что ГПГ мог играть важную роль в обеспечении инфузорий рубца. адаптироваться к новым нишам в рубце и что фибролитические гены были приобретены простейшими из бактериальных источников (Findley et al., 2011).
Роль простейших в рубце
Несмотря на то, что эти простейшие составляют значительную часть биомассы рубца, их роль в ферментации рубца и их вклад в метаболизм и питание хозяина по-прежнему вызывает серьезные споры (Williams and Coleman, 1992). В последнем разделе этой статьи мы рассмотрим различные стратегии управления плотностью простейших в рубце; однако большинство этих диетических вмешательств также приводят к изменениям функции рубца, что затрудняет оценку эффекта простейших как таковых рубца.Простейшие в рубце не необходимы животному для выживания и дефаунации (удаления простейших из рубца с использованием широкого спектра химических и физических методов), а животные, свободные от простейших, использовались для изучения роли инфузорий простейших в функции рубца без пострадали от диетических вмешательств (Williams and Coleman, 1992). Эта статья не ставит своей целью провести полный обзор последствий дефауна, но объединяет наиболее важные публикации со времен превосходного и полного обзора, проведенного Уильямсом и Коулманом (1992).Таким образом, был проведен метаанализ для изучения основных эффектов дефаунации на основании 23 исследований in vivo , включающих 48 сравнений (Kreuzer et al., 1986; Vermorel and Jouany, 1989; Ushida et al., 1990; Frumholtz, 1991 ; Nagaraja et al., 1992; Hegarty et al., 1994, 2008; Faichney et al., 1999; Chandramoni et al., 2001; Machmuller et al., 2003; Eugène et al., 2004; Ozutsumi et al., 2005; Ohene-Adjei et al., 2007; Yáñez-Ruiz et al., 2007a, b; Bird et al., 2008; Morgavi et al., 2008, 2012; Belanche et al., 2011, 2012b, 2015; Mosoni et al., 2011; Чжоу и др., 2011). Большинство исследований проводилось на овцах (87%), остальные — на крупном рогатом скоте (13%). Изоляция новорожденных животных от их матерей (40%), использование моющих средств и других химикатов (35%, с использованием лаурилсульфата натрия, алканов, синперонового NP9, пероксида кальция, сульфата меди и т. Д.) И манипуляции с рубцом (25%, опорожнение и промывание рубца), были использованы для достижения дефаунации животных. Испытания, в которых влияние добавок, кроме дефауна, было значительным, были исключены из метаанализа.Точно так же не были включены исследования с использованием монофаунизированных животных или селективной дефауны. Большинство данных (75%) были получены из испытаний, в которых животных кормили на поддерживающем уровне, а остальные испытания были из производственных испытаний. Животных в основном кормили смешанными рационами (90%), состоящими из кормов с добавлением концентрата, в то время как 10% рационов состояли исключительно из кормов. Данные о ферментации рубца и выбросы метана были представлены в большинстве испытаний (69%), в то время как информация о пищеварении (31%), продуктивности животных (12%) и микробных популяциях рубца (10%) была менее обширной.Производство метана измерялось в камерах (75% в открытых камерах или респираторных калориметрах) или с помощью метода индикатора SF 6 (гексафторид серы) (25% исследований).
Множественные сравнения были включены из отдельной публикации с множественными исследованиями. Для каждого сравнения, включенного в анализ, величина эффекта рассчитывалась как натуральный логарифм отношения ответа (среднее значение в исчерпанном лечении, деленное на среднее значение в контрольном лечении) и дисперсии отношения, рассчитанной на основе сообщенного стандартного отклонения или стандарта. ошибка среднего для каждого сравнения (Viechtbauer, 2010).Все эффекты дефаунации были взвешены в соответствии с количеством наблюдений ( n ) в каждом сравнении. Мета-анализ был рассчитан с использованием модели случайных эффектов с оценкой Дерсимониан-Лэрда (Dersimonian and Laird, 1989) для оценки неоднородности (τ 2 ) в пакете Metafor R для каждой категории отдельно, как показано ниже:
, где θ i = истинная величина эффекта в i-м исследовании, μ = общая истинная величина эффекта и e i = случайное отклонение от общей величины эффекта [ u i ∼ N (0, τ 2 )] (Viechtbauer, 2010).Для оценки коэффициента отклика значения ниже единицы указывают на отрицательный результат, а значения выше единицы указывают на положительный эффект дефаунации на этот конкретный параметр.
Этот метаанализ подтвердил многие из ранее сообщенных результатов (Williams and Coleman, 1992) и помог прояснить ряд областей, в которых результаты все еще были противоречивыми. На рис. 1 показаны основные эффекты дефаунации на функцию рубца, указывающие на то, что физические и химические характеристики среды рубца были изменены дефаунацией; хотя характер наблюдаемых изменений не всегда был последовательным.Дефауна не повлияла на объем рубца и скорость оборота твердых веществ, в то время как скорость оборота жидкости имела тенденцию к снижению (–14%, P = 0,07). Удаление простейших из рубца значительно снижает усвояемость ОВ рубца (–7%, P = 0,008) и особенно NDF (–20%, P = 0,040) и усвояемость ADF (–16%, P = 0,100) , вероятно, в результате потери фибролитической активности простейших. Однако эта активность, по-видимому, различается в разных группах простейших: крупные Ophyroscolecidae , такие как Epidinium , Polyplastron и Eudiplodinium , обладают большей эндоглюканазной и ксиланазной активностью (Williams and Coleman, 1992).С другой стороны, Entodinium spp. имеют только слабую активность (Williams, Coleman, 1992). Точно так же Dasytricha обладает активностью глюкозидазы и целлобиозидазы, но незначительной фибролитической активностью (Takenaka et al., 2004). Низкая перевариваемость в рубце у дефаунированных животных частично компенсируется более выраженным пострубцовым перевариванием, что приводит к менее выраженным различиям в показателях общей перевариваемости тракта для OM (–4%, P = 0,089), CP (–3%, P = 0.034), NDF (–11%, P <0,001) и ADF (–9%, P <0,001; рис. 2). Другим компенсирующим фактором может быть сдвиг в сторону более энергетически эффективных реакций в рубце (меньшее количество выбросов метана) и меньшая метаболическая энергия, необходимая обезвожившимся животным для устранения избытка мочевины в результате более низкого расщепления бактериального белка и уровня аммиака в рубце. Несмотря на этот компенсаторный механизм, снижение перевариваемости корма, вероятно, будет основным недостатком дефауна, поскольку оно может ограничить потребление корма и эффективность использования корма на производственных уровнях потребления; Это в сочетании с отсутствием коммерчески жизнеспособного подхода (см. ниже) к дефаунации означает, что дефаунация не рекомендуется в качестве стратегии снижения выбросов метана в фермерских условиях (Христов и др., 2013).
Рис. 1. Мета-анализ, описывающий влияние дефаунации на функцию рубца. Для каждого параметра предоставляется информация о количестве исследований, наблюдениях (n) и диапазоне значений. На графике показаны средние эффекты и 95% конфиденциальные интервалы. Значения ниже единицы указывают на отрицательный эффект, тогда как значения выше единицы указывают на положительное влияние отключения на этот конкретный параметр.
Рис. 2. Мета-анализ, описывающий эффекты дефаунации животных. Для каждого параметра предоставляется информация о количестве исследований, наблюдениях (n) и диапазоне значений. На графике показаны средние эффекты и 95% конфиденциальные интервалы. Значения ниже единицы указывают на отрицательный эффект, тогда как значения выше единицы указывают на положительное влияние отключения на этот конкретный параметр.
Общая концентрация и производство продуктов ферментации также различаются у фаунированных и лишившихся животных животных. Снижение аммиака в рубце (–26%, P <0.001), вероятно, является наиболее последовательным из наблюдаемых эффектов элиминации простейших и, по-видимому, обусловлен снижением расщепления бактериального белка и разлагаемости кормового белка в отсутствие простейших в рубце (Williams and Coleman, 1992). Более того, было продемонстрировано, что, хотя хищничество бактерий простейшими в рубце зависит от размера простейших, холотрих простейшие имеют гораздо более низкую хищническую активность, чем энтодиниоморфиды (Belanche et al., 2012a) и, в конечном итоге, более низкое влияние на концентрацию аммиака в рубце (Belanche et al. ., 2015) и микробного белкового потока двенадцатиперстной кишки (Ivan et al., 2000a; Иван, 2009). Однако это негативное влияние энтодиниоморфидов на белковый поток двенадцатиперстной кишки можно переоценить из-за отсутствия надежного маркера для измерения протозойного кровотока (Broderick and Merchen, 1992). В предыдущих обзорах сообщалось о более высоких уровнях лактата у обезвоженных животных (Williams and Coleman, 1992), поскольку простейшие потребляют лактат быстрее, чем бактерии (Newbold et al., 1986). Наш метаанализ показал численное увеличение концентрации лактата у дефунтированных животных (+ 11%), но это увеличение было незначительным, возможно потому, что набор данных, использованный в этом метаанализе, в основном состоял из рационов с высоким содержанием кормов, которые менее склонны к продвижению. эти конкретные нарушения питания.
Хотя рН рубца не изменился, более низкая концентрация ЛЖК, наблюдаемая у дефаунированных животных (–5%, P = 0,013), по-видимому, подчеркивает роль простейших в синтезе ЛЖК и деградации корма. Способность простейших поглощать экзогенные жирные кислоты (Karnati et al., 2009) может отвлекать больше углерода на выработку ЛЖК, а не на синтез жирных кислот, и в конечном итоге увеличивать выработку ЛЖК. Что еще интереснее, наши данные показали, что дефаунация существенно снижает бутират (–22%, P <0.001), слегка повышенное содержание ацетата (+ 3%, P <0,001) и не влияло на молярные пропорции пропионата. В предыдущем метаанализе (Noziere et al., 2011) сообщалось, что усвояемость NDF положительно коррелирует с молярной долей ацетата ( r = 0,95), но отрицательно с пропионатом ( r = –0,94) и бутиратом ( r = –0.91), поэтому наши наблюдения могут указывать на то, что изменение молярных пропорций ЛЖК, вызванное дефаунацией, по-видимому, в основном вызвано уменьшением переваривания клетчатки.Исходя из стехиометрии, такой сдвиг в выработке ЛЖК в рубце должен привести к снижению продукции метана, поскольку менее метаболический H 2 будет доступен в качестве субстрата для метаногенеза (Demeyer et al., 1996). Однако влияние остановки на производство метана все еще не ясно; в различных обзорах, объединяющих in vivo, и in vitro, исследований (Hegarty, 1999) или просто in vivo (Morgavi et al., 2010), был сделан вывод, что удаление простейших из рубца приведет к 13 и 10 .Снижение добычи метана на 5%, что полностью согласуется с нашими результатами, основанными на более поздних исследованиях (–11%, P = 0,074). В соответствии с этими наблюдениями, в метаанализе, содержащем 28 экспериментов и 91 обработку, было сообщено о значительной линейной зависимости между выбросами метана и концентрацией простейших ( r = 0,96) с уменьшением выхода метана в среднем на 8,14 г / кг. DMI на каждую логарифмическую единицу снижения концентрации простейших (Guyader et al., 2014), предполагая, что простейшие играют каталитическую роль в метаногенезе рубца.Причины более низких выбросов метана у дефонированных животных до сих пор остаются спорными, поскольку количество простейших в рубце объясняет только одну часть (примерно 47%) изменчивости выбросов метана, что указывает на то, что метаногенез в рубце представляет собой сложный процесс, в котором участвуют несколько микробов (Morgavi и др., 2010). Возможен ряд механизмов, с помощью которых простейшие могут усиливать метаногенез, в зависимости от их способности продуцировать H 2 в своих гидрогеносомах (митохондриально-подобной органелле), их способности принимать эпи- и эндосимбиотические метаногены и защищать их от кислорода. токсичность (Fenchel and Finlay, 2006).Это взаимодействие является типичным примером межвидового переноса H 2 , который благоприятствует как метаногенам, так и простейшим (Ushida et al., 1997). Одна из гипотез состоит в том, что дефауна приводит к снижению количества метаногенов, которые являются единственными продуцентами метана в рубце, но наш мета-анализ показал, что это снижение уровней метаногенов не было значительным (–3%, P = 0,48). Альтернативная гипотеза предполагает, что дефаунация приводит к элиминации метаногенов, связанных с простейшими, что можно рассматривать как одно из наиболее активных сообществ метаногенов в рубце (Finlay et al., 1994), однако эта более поздняя гипотеза, основанная на замещении метаногеновых сообществ, различающихся по своей метаногенной активности, требует дальнейшего изучения. Несколько авторов изучали эндосимбиотическую популяцию архей у простейших (Sharp et al., 1998; Irbis, Ushida, 2004; Regensbogenova et al., 2004b; Tymensen et al., 2012a), и большинство из них согласны с тем, что Methanobrevibacter sp . является преобладающим родом, а вклад Methanomicrobium sp. и Methanomassiliicoccales в эндосимбиотическом сообществе метаногенов варьирует и может указывать на различия между группами простейших.Развитие молекулярных методов за последние десятилетия позволило коллегам дополнительно изучить эти микробные взаимодействия, обнаружив, что метаногены инфузорий эндосимбиотиков отличаются от свободноживущих метаногенов рубца (Tokura et al., 1997; Sharp et al., 1998). В недавней публикации также было продемонстрировано, что у холотриких простейших есть разные эндосимбиотические метаногены, чем у энтодиниоморфидов (Belanche et al., 2014), возможно, потому, что у любого из холотрих простейших есть более активные гидрогеносомы, чем у энтодиниоморфидов (Paul et al., 1990) и / или быстрый синтез гликогена простейшими холотрихами в присутствии избытка углеводов генерирует больше водорода (Hall, 2011; Denton et al., 2015). Эти различия могут объяснять большее влияние голотричных простейших на метаногенез рубца по сравнению с энтодиниоморфидами (Belanche et al., 2015). Это более позднее наблюдение основано на последовательной инокуляции овец, свободных от фауны, различными группами простейших, поэтому его следует интерпретировать с осторожностью из-за возможных смешанных эффектов лечения и периода.
Влияние дефаунации на микробную экосистему рубца не ограничивается только популяцией метаногена, и было продемонстрировано, что у дефаунированных животных было значительно больше популяций бактерий в рубце (+ 9%, P <0,001), чем у контрольных животных. Причина этого, по-видимому, основана на доступности экологической ниши для бактерий, когда простейшие не присутствуют в рубце, в сочетании с устранением простейших хищников (Williams and Coleman, 1992).Недавние исследования с использованием молекулярных методов показали, что дефаунация также изменяет структуру бактериального сообщества, что приводит к упрощению популяционной структуры и снижению бактериального разнообразия (Belanche et al., 2012b, 2015). Наш метаанализ выявил снижение концентрации фибролитических микробов, таких как анаэробные грибы (–92%, P <0,001), Ruminococcus albus (–34%, P <0,001) и Ruminococcus flavefaciens ( –22%, P <0.001) в отсутствие простейших. Это наблюдение предполагает, что переваривание клетчатки в рубце - сложная задача, которая требует симбиотического сотрудничества нескольких фибролитических микробов, включая простейшие рубца, для выполнения начальных стадий колонизации клетчатки и пищеварения; поэтому отсутствие простейших в рубце, по-видимому, оказывает пагубное влияние на этот фибролитический консорциум и, в конечном итоге, на переваривание клетчатки. Однако Hsu et al. (1991) сообщили об увеличении зооспор грибов в рубце у лишившихся жизни животных, возможно, в результате устранения хищничества простейших и конкуренции за питательные вещества (простейшие vs.грибки), очевидно, необходимы дополнительные исследования, чтобы полностью понять взаимодействие простейших с другими микробами рубца при различных режимах питания.
Влияние присутствия простейших в рубце на выживаемость патогенов в рубце и выделение патогенов — еще одна область, представляющая интерес. Как отмечалось выше, простейшие в рубце поглощают и переваривают широкий спектр бактерий (Williams and Coleman, 1992) и могут уменьшить выделение потенциальных патогенов от животного, хотя эффект сильно зависит от состава присутствующей популяции простейших (Stanford et al. ., 2010). Однако также было показано, что простейшие в рубце увеличивают патогенность некоторых патогенов, покидающих рубец (Rasmussen et al., 2005; Carlson et al., 2007), что позволяет предположить, что в этой области требуется дополнительная работа.
Рисунок 2 суммирует влияние простейших на продуктивность животных как косвенный эффект дефаунации на функцию рубца. Дефауна улучшила конверсию корма в результате более низкого потребления СВ (–2%, P <0,001) и большего среднесуточного привеса (+ 9%, P <0.001). Эти положительные эффекты дефауна особенно очевидны при низком качестве диеты, при которой средний дневной прирост был низким, возможно, из-за более низкой доступности ферментируемой энергии и перевариваемого в рубце белка для микробов рубца (Williams and Coleman, 1992). В предыдущем метаанализе эффект дефаунации оказался более выраженным, когда отношение N / NDF было ниже 6, а процент концентрата ниже 40% (Eugène et al., 2004). Так как животные, лишенные жизни, имеют более низкую усвояемость корма, поглощенная энергия ниже, чем у обычных животных.Таким образом, лучшая степень конверсии корма после отключения может быть главным образом связана с более высокой эффективностью использования поглощенных питательных веществ. Действительно, наш мета-анализ показал, что дефауна способствует большей эффективности использования энергии для откорма (+ 11%, P = 0,001), возможно, в результате более низкой выработки тепла (–5%, P = 0,010). Другая гипотеза, объясняющая лучшую продуктивность дефунтированных животных, основана на более эффективном использовании пищевого белка. Существуют четкие доказательства того, что простейшие холотрицы покидают рубец медленнее, чем бактерии (Abe et al., 1981). Однако количество и скорость потока простейших в нижний отдел кишечника является предметом многочисленных споров. Firkins et al. (2007), обнаружили, что поструминальный поток простейших пропорционален биомассе простейших в рубце. Хотя подсчет простейших в рубце и сычуговой жидкости показал, что количество сычужных сычуг составляло только 6–64% от подсчета жидкости в рубце (Punia et al., 1992), количество простейших в свободной жидкости рубца является ненадежным индикатором биомассы и оттока простейших, поскольку большинство простейших в рубце (63–90%) обнаруживаются либо связанными с частицами корма, либо секвестрированными в стенке рубца (Hook et al., 2012). Использование специфических маркеров простейших, таких как 2-аминоэтилфосфоновая кислота, подвергается сомнению, поскольку она присутствует в бактериях и в кормах (Ling and Buttery, 1978). Протозойный отток, измеренный с использованием общих микробных маркеров (например, 15 N и 35 S и пуриновых оснований), микробных N минус специфических бактериальных маркеров (таких как 2,6-диаминопимелиновая кислота, DAPA) давал различные оценки протозойных отток, а иногда и отрицательные значения из-за методологических ограничений используемых маркеров (Broderick and Merchen, 1992).Недавние исследования, основанные на использовании 18S рРНК в качестве нового маркера простейших, показали, что, хотя простейшие могут составлять до 60% микробной биомассы рубца, они редко превышают 20% потока микробных белков в тонкий кишечник (Sylvester et al. , 2004, 2005; Yáñez-Ruiz et al., 2006; Belanche et al., 2011, 2012b). Однако эти новые методы не лишены собственных ограничений: Sylvester et al. (2005) наблюдали, что до 48% простейшего азота на самом деле было азотом от заражающих бактерий, что могло привести к раздуванию пула простейшего азота в рубце, особенно когда животные имеют низкое потребление корма и низкую скорость прохождения твердых частиц (Dijkstra, 1994).Аналогичным образом Belanche et al. (2010b) наблюдали большую деградацию протозойной ДНК по сравнению с бактериальной ДНК во время сычужного переваривания, что могло привести к недооценке оттока простейшего азота из рубца. Несмотря на эти ограничения, эти новые данные, кажется, подтверждают, что простейшие в рубце частично секвестрируются в рубце. Однако эта секвестрация в рубце не одинакова для всех групп простейших; holotrich простейшие связываются с частицами корма после кормления из-за их сильного хемотаксиса по отношению к сахарам (Diaz et al., 2014), но быстро мигрируют в вентральную ретикулоруму, чтобы предотвратить вымывание из рубца (Karnati et al., 2007), напротив, энтодиниоморфиды также связываются с кормом из-за их умеренного хемотаксиса в отношении глюкозы и пептидов, но не проявляют такое же сродство к стенке рубца (Diaz et al., 2014) и, таким образом, вытекает из рубца с фазой твердых частиц (Hook et al., 2012).
Простейшие предшествуют бактериям в качестве своего основного источника белка (Williams and Coleman, 1992), и в результате дефаунация делает рубец более эффективным с точки зрения протеосинтеза, увеличивая дуоденальный поток микробного белка (+ 30%, P <0.001) и общий поток неаммиачного азота (+ 31%, P <0,001). Дефаунация также увеличивала эффективность синтеза микробного белка (+ 27%, P = 0,008) в результате как лучшего микробного протеосинтеза, так и более низкого усвоения ОМ. Время генерации протозойных намного выше, чем у бактерий, поэтому энергетические потребности для поддержания выше, если выразить их как соотношение белков, покидающих рубец (Williams and Coleman, 1992). В результате присутствие простейших отрицательно сказывается на общей энергетической эффективности экосистемы рубца.Кроме того, дефаунация также может изменять состав бактерий рубца (Belanche et al., 2012b) и, в конечном итоге, аминокислотный профиль поступления белка двенадцатиперстной кишки, способствуя увеличению определенных аминокислот, таких как лейцин, треонин и аргинин (Ivan, 2009 ), но не лизин, который считается основной лимитирующей аминокислотой у высокопродуктивных животных (Hristov and Jouany, 2005). После серии экспериментов in vivo , в которых ягнят, свободных от фауны, постепенно инокулировали простейшие виды (т.e., Isotricha Кишечник , Dasytricha ruminantium , Polyplastron multivesiculatum , Epidinium ecaudatum , Eudiplodinium maggi , и Ivan Entodinium caudatum, в разных порядках, b. 2009), Иван пришел к выводу, что холотрих простейшие поглощают лишь очень небольшое количество бактерий рубца и оказывают незначительное влияние на дуоденальный поток NAN и метаболизм белков. Точно так же простейшие голотрих не оказали значительного влияния на переваривание клетчатки.Это вместе с вкладом голотрихов в метаногенез рубца (Belanche et al., 2015), по-видимому, указывает на то, что присутствие голотрихов простейших в рубце не имеет большого значения для производства жвачных животных, если не используются диеты с высоким содержанием углеводов, и в этом случае присутствие голотричных простейших может быть полезным в результате их способности поглощать и накапливать зерна крахмала и растворимые углеводы (Williams and Coleman, 1992). Такое поглощение высоко ферментируемых углеводов предотвращает альтернативную бактериальную ферментацию, которая в противном случае снизила бы pH и увеличила бы возникновение молочнокислого ацидоза (Mackie et al., 1978). Напротив, целлюлолитические простейшие ( Polyplastron , Epidinium и Eudiplodinium ) могут быть полезными для животных, которых кормили волокнистой пищей. Однако эти фибролитические простейшие и Entodinium spp. обладают значительным потенциалом поглощения и разложения бактерий (Belanche et al., 2012a) и могут быть вредными с точки зрения использования белка жвачным животным-хозяином. Следовательно, их присутствие в рубце может быть бесполезным для животных, получающих диету с низким содержанием белка, поскольку их благоприятное фибролитическое действие будет уравновешено чувствительностью фибролитических бактерий и анаэробных грибов к дефициту азота в рубце (Belanche et al., 2012c). Таким образом, похоже, что дефауна уменьшила потери азота с мочой здесь (–10%, P <0,001) из-за комбинации более низкой пищевой деградации CP в рубце и бактериального расщепления нижней части рубца. Однако большая изменчивость, наблюдаемая с точки зрения баланса азота, подчеркивает тот факт, что дефаунация может по-разному влиять на общую эффективность использования азота в зависимости от диеты, потребляемой животным, в частности, уровня потребления и скорости прохождения частиц через тракт (Дейкстра , 1994).
Влияние дефаунации на метаболизм липидов в рубце менее определено. Простейшие вносят вклад в общую липолитическую активность микробов, но их роль в биогидрировании менее изучена (Williams and Coleman, 1992), хотя они действительно вносят значительный вклад в отток ненасыщенных жирных кислот, покидающих рубец (Yáñez-Ruiz et al., 2006). Используя рацион бычков, получавших рационы с различным содержанием хлорофилла, было продемонстрировано, что высокие уровни полиненасыщенных жирных кислот в клетках простейших, по-видимому, связаны с потреблением хлоропластов (Huws et al., 2009). Это поглощение хлоропластов, по-видимому, специфично для энтодиниоморфидов, поскольку не было обнаружено захваченных хлоропластов у голотриких простейших (Huws et al., 2012). Таким образом, простейшие, по-видимому, защищают ненасыщенные жирные кислоты хлоропластов от биогидрирования рубца, увеличивая дуоденальные потоки моно- и полиненасыщенных жирных кислот. Наш метаанализ согласуется с этим наблюдением, и дефаунация способствовала увеличению насыщенных жирных кислот (+ 4%, P = 0,046) и численному снижению моно- и полиненасыщенных жирных кислот (–13 и –10% соответственно), что возможно, не достигли значимости из-за эффектов, зависящих от диеты.
Результаты, представленные в нашем метаанализе, должны быть тщательно интерпретированы, поскольку большинство исследований имело методологические ограничения из-за внутренней сложности процесса дефаунации, ведущего к:
(1) Путаница между эффектом дефаунации и периодом при использовании одних и тех же экспериментальных животных во времени,
(2) Почти все исследования, оценивающие размер пула простейших, не смогли сообщить о дуоденальных потоках протозойной биомассы, а наоборот , более того, поток простейших часто недооценивается с использованием традиционных микробных маркеров (Broderick and Merchen, 1992),
(3) Исследования, оценивающие производство метана, не учитывали другие переменные эффекты, кроме численности простейших (т.е., усвояемость NDF),
(4) Животных часто кормят в количествах, намного ниже производственных уровней потребления,
(5) В некоторых исследованиях период адаптации после дефаунации / восстановления слишком короток для того, чтобы микробная экосистема могла полностью адаптироваться,
(6) Устранение простейших из рубца изменяет экологическую структуру рубца, и в конечном итоге альтернативные группы микробов могут занять экологическую нишу, ранее заполненную простейшими.
Таким образом, необходимы дополнительные исследования с использованием современных технологий для количественной оценки активности простейших (т.е., фибролитические и протеолитические), а также размер пула простейших в рубце относительно оттока простейшего азота для лучшего понимания роли простейших в метаболизме жвачных. Эти исследования должны проводиться на производственных уровнях потребления, чтобы действительно оценить эффект дефаунации на единицу произведенного продукта (молока или мяса).
Манипуляции с простейшими в рубце
Текущие опасения по поводу роли домашнего скота в глобальном предупреждении побудили исследователей искать стратегии манипулирования простейшими в рубце для уменьшения производства метана.Как уже упоминалось ранее, существует линейная зависимость между концентрацией простейших и выбросами метана (Guyader et al., 2014), и было подсчитано, что от 9 до 37% продукции метана в рубце можно отнести к метаногенам, связанным с простейшими в рубец (Finlay et al., 1994; Newbold et al., 1995; Machmuller et al., 2003).
Большинство подходов к дефаунации редко приводят к полному удалению простейших из рубца, их эффективность во многом зависит от состава диеты (Hegarty, 1999).Лечение, обычно используемое для частичной или полной дефауны рубца, включает: химические вещества, токсичные для простейших (сульфат меди, диоктилсульфосукцинат, этоксилат или алканаты спирта, пероксид кальция), ионофоры, липиды и сапонины (Williams and Coleman, 1992; Jouany, 1996; Hook et al., 2010).
Недавний метаанализ Guyader et al. (2014) показали одновременное снижение численности простейших и выбросов метана в 31% из 70 исследований, в которых использовались различные стратегии сокращения выбросов метана.В большинстве исследований липиды использовались в качестве стратегии борьбы с простейшими / снижения уровня метана. Авторы также сообщили, что антипротозойный эффект липидов зависит от состава жирных кислот, причем жирные кислоты со средней длиной цепи более эффективны, чем полиненасыщенные жирные кислоты, в контроле численности простейших. Было показано, что добавки, богатые полиненасыщенными жирными кислотами, такими как линолевая кислота (C18: 2 из сои и подсолнечника) и линоленовая кислота (C18: 3 из льняного семени), оказывают негативное влияние на выработку метана (4.Снижение на 1 и 4,8% на процентную единицу добавленных липидов, соответственно, Martin et al., 2010). Антиметаногенный эффект полиненасыщенных жирных кислот был связан с их токсическим действием на целлюлозолитические бактерии (Nagaraja et al., 1997) и простейшие (Doreau and Ferlay, 1995). Хотя первоначально предполагалось, что биогидрирование полиненасыщенных жирных кислот в рубце может представлять собой альтернативу метаногенезу H 2 (Lennarz, 1966), теперь считается, что значение биогидрирования для общего стока H 2 является small (Nagaraja et al., 1997). Было показано, что жирные кислоты со средней длиной цепи обладают сильным антипротозойным действием. В нескольких исследованиях сообщалось о снижении метаногенеза в рубце при добавлении лауриновой кислоты (C12: 0) и миристиновой кислоты (C14: 0) либо в чистых формах, либо в продуктах, богатых этими жирными кислотами (кокосовое масло), под in vitro (Dohme et al. ., 2001; Soliva et al., 2004) или in vivo в условиях (Machmuller et al., 2003; Jordan et al., 2006). Martin et al. (2010) сообщили, что жирные кислоты со средней длиной цепи, в основном содержащиеся в кокосовом масле, привели к снижению метана на 7.3% на процентную единицу добавленного липида. Однако наблюдаемый эффект подавления метана не всегда был связан со снижением концентрации простейших (Machmuller et al., 2003), что может быть связано с прямым действием жирных кислот со средней длиной цепи на метаногены (Dohme et al., 1999; Panyakaew et al. др., 2013). Недавнее исследование (Faciola and Broderick, 2014) показало, что кокосовое масло и лауриновая кислота снижают количество простейших на 40%, тогда как лауриновая кислота изменяет усвояемость клетчатки, кокосовое масло — нет.Однако сообщалось о сокращении выработки метана и сопутствующем уменьшении потребления сухого вещества, когда кокосовое масло и лауриновая кислота использовались в качестве дезорганизующих агентов (Христов и др., 2013), что потенциально ограничило бы их практическое использование в фермерских хозяйствах. Очевидно, что необходимы дополнительные исследования, чтобы выяснить, как использовать масла и жирные кислоты для борьбы с простейшими в рубце.
В литературе предполагается, что сапонины смягчают метаногенез, главным образом, за счет уменьшения количества простейших, в то время как конденсированные танины действуют как за счет уменьшения количества простейших, так и за счет прямого токсического воздействия на метаногены, тогда как эфирные масла действуют в основном путем прямого токсического действия на метаногены (Cieslak и другие., 2013). В соответствии с этой информацией, метаанализ влияния фитохимических веществ на метаногенез (Patra, 2010) показал, что изменения численности простейших следовали линейной зависимости с изменениями выработки метана сапонинами ( r = 0,69) и танинами ( r = 0,55), но эта связь была слабее ( r = 0,45) по отношению к эфирным маслам. Ингибирование метана сероорганическими соединениями не было связано с изменениями в популяции простейших (Patra, 2010), поскольку такие соединения специфически ингибируют метаногенные археи.Для растений, содержащих танин, антиметаногенная активность приписывается группе конденсированных танинов. Было высказано предположение, что дубильные вещества имеют прямое влияние на метаногены в рубце и косвенное влияние на производство водорода из-за снижения переваривания клетчатки (Tavendale et al., 2005). Что касается их воздействия на простейшие, в некоторых исследованиях не сообщалось об их эффекте, в то время как другие показали снижение количества простейших в присутствии танинов (обзор Patra and Saxena, 2009). Это несоответствие, вероятно, связано с различиями в структуре и дозе конденсированного танина.Сапонины в доступной литературе показывают более стойкое ингибирующее действие на простейшие в рубце. Гоэль и Маккар (2012) предположили, что риск нарушения функции рубца и, как следствие, снижения продуктивности животных выше с дубильными веществами, чем с сапонинами, и когда они используются для уменьшения выработки метана, эффективный диапазон концентраций дубильных веществ уже, чем сапонинов. Сапонины представляют собой гликозилированные тритерпены или стероиды, где сапонин представляет собой агликон, а гликон представляет собой углеводную единицу, состоящую из моносахарида или олигосахарида меньшего размера.Наиболее частыми источниками сапонинов, используемых в питании жвачных животных, являются Yucca schidigera , богатая стериновыми сапонинами (10%), и Quillaja saponaria , содержащая тритерпеновые сапонины. В последнее время изучаются и другие источники сапонинов, такие как сапонины чая (Hu et al., 2005; Guo et al., 2008). Антипротозойный эффект сапонинов связан с их взаимодействием со стероловым фрагментом, который присутствует в мембране простейших (Patra and Saxena, 2009). Было высказано предположение, что сапонины с одним и тем же агликоном могут по-разному влиять на простейшие в зависимости от состава и расположения сахара (Wina et al., 2006). Этот эффект кажется временным из-за разрыва гликозидной связи микробами рубца (Newbold et al., 1997). Подавление простейших в рубце сапонинами или сапонинсодержащими растениями постоянно наблюдается в исследованиях in vitro (Wina et al., 2005). Однако исследования in vivo показали, что антипротозойный эффект сапонинов имеет тенденцию исчезать через 7–14 дней приема (Wina et al., 2005; Patra and Saxena, 2009). Было высказано предположение, что когда сапонины дегликозилируются в сапогенины микробами рубца, они становятся неактивными (Newbold et al., 1997). Таким образом, можно предположить, что комбинация сапонинов с ингибиторами гликозидазы позволит избежать дегликозилирования, поддерживая интактный сапонин и, следовательно, активность в рубце. Предварительные исследования in vitro (Ramos-Morales et al., 2014) с использованием 2,5-дигидроксиметил-3,4-дигидроксипирролидина (DMDP) в качестве ингибитора гликозидазы в сочетании с экстрактом растений, богатым сапонинами, показали потенциал этого подхода. для поддержания активности сапонинов с течением времени. Другой подход, который изучается нашей исследовательской группой, — это синтез химически модифицированного сапонина без естественных гликозидных связей, поэтому ферментативное расщепление будет структурно запрещено.Влияние сапонинов на ферментацию рубца не подтвердилось. Эти расхождения, по-видимому, связаны с химической структурой и дозировкой сапонинов, составом рациона, а также адаптацией микроорганизмов к сапонинам (Wina et al., 2005; Patra and Saxena, 2009).
Австралийские исследователи продемонстрировали потенциал вакцинации против метаногенов как метода уменьшения выбросов метана (Wright et al., 2004). Хотя эта технология все еще развивается, она предоставляет множество возможностей для долгосрочного сокращения выбросов метана.Точно так же простейшие, как поставщики водорода для метаногенов или ацетогены, которые конкурируют за водород с метаногенами, могут быть возможными целями вакцины для сокращения выбросов метана. Иммунологический подход был изучен для дефаунации (Williams et al., 2008). Вакцинация овец энтодинией или смешанными протозойными антигенами уменьшала количество простейших, и антитела IgG, выработанные против простейших в рубце, оставались активными и продолжали связывать клетки-мишени в течение до 8 часов. Однако не наблюдалось никакого действия in vivo на простейшие в рубце.Было высказано предположение, что причины отсутствия эффекта могут быть связаны с недостаточным количеством специфического Ig, доставляемого в слюне, необходимостью адъюванта для оптимизации выработки антител в слюне, нацеливания на другие антигены простейших для создания более сильного иммунного ответа.
Заключение
Со времени знаменательной публикации «Rumen Protozoa» Уильямса и Коулмана (1992) в нашем понимании простейших рубца наблюдается устойчивый, но, возможно, не впечатляющий прогресс.Появление молекулярных методов привело к появлению множества публикаций, касающихся разнообразия простейших в рубце, и, хотя, как обсуждалось выше, используемые методы имеют внутри себя ограничения в их способности точно количественно определять отдельные роды простейших, они предоставили новое понимание разнообразия простейших инфузорий. у разных видов жвачных, в разных географических регионах и при разных диетических условиях. В области дефаунации наблюдается неуклонный прогресс, и хотя на данном этапе на рынке еще не поступило коммерчески доступной техники дефаунации, вполне вероятно, что экстракты растений можно использовать для борьбы с простейшими в рубце; если не устранить их полностью.Работа над последствиями уничтожения простейших в значительной степени сосредоточена на их роли в метаногенезе, что отражает текущие опасения относительно роли жвачных животных в производстве парниковых газов. Однако использование метаанализа существующих данных в сочетании с новыми исследованиями дефаунации помогло прояснить наше понимание роли простейших в рубце, как показано на рисунках 1 и 2. Однако, возможно, это развитие молекулярных методов для клонирования и характеристики простейших. гены, первоначально от одного вида, но в последнее время из метагеномных и транскриптомных источников, которые, по-видимому, предлагают наибольший, но пока еще в значительной степени не реализованный, потенциал, чтобы помочь по-настоящему выяснить роль простейших в рубце и в отсутствие прогресса в развитии аксенических Культура простейших в рубце Необходимо приложить больше усилий для характеристики активности простейших в рубце с помощью молекулярных методов.
Авторские взносы
Все авторы в равной степени внесли вклад в подготовку данной публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
CN и AB выражают признательность за поддержку Исследовательскому совету биотехнологии и биологических наук Великобритании через грант номер BB / J0013 / 1.
Ссылки
Abe, M., Iriki, T., Tobe, N., and Shibui, H. (1981). Секвестрация простейших голотрихов в ретикуло-рубце крупного рогатого скота. Заявл. Environ. Microbiol. 41, 758–765.
PubMed Аннотация | Google Scholar
Амарал-Цеттлер, Л. А., МакКлимент, Э. А., Даклоу, Х. В., и Хьюз, С. М. (2009). Метод изучения разнообразия протистанов с использованием массивно-параллельного секвенирования гипервариабельных участков v9 малых субъединичных генов рибосомной РНК. PLoS ONE 4: e6372. DOI: 10.1371 / journal.pone.0006372
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Амарал-Цеттлер, Л. А., Зеттлер, Э. Р., Теру, С. М., Паласиос, К., Агилера, А., и Амилс, Р. (2011). Структура микробного сообщества на древе жизни в крайнем Рио-Тинто. ISME J. 5, 42–50. DOI: 10.1038 / ismej.2010.101
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Абесия, Л., Holtrop, G., Guada, J. A., Castrillo, C., de la Fuente, G., et al. (2011). Изучение влияния присутствия или отсутствия простейших на ферментацию рубца и вклад микробных белков в химус. J. Anim. Sci. 89, 4163–4174. DOI: 10.2527 / jas.2010-3703
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Бальселлс, Дж., Де Ла Фуэнте, Г., Яньес-Руиз, Д. Р., Фондевила, М., и Каллеха, Л. (2010a). Описание развития экосистемы рубца с помощью ПЦР-анализа у ягнят на молочном, отнятом от груди и в откорме в системе интенсивного откорма. J. Anim. Physiol. Anim. Nutr. 94, 648–658. DOI: 10.1111 / j.1439-0396.2009.00952.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче А., Эрроа И. Р., Балселлс Дж. И Каллеха Л. (2010b). Использование количественной ПЦР в реальном времени для оценки выживания in vitro определенных последовательностей генов ДНК микробов рубца в имитированных сычужных условиях. J. Anim. Physiol. Anim. Nutr. 94, 204–211. DOI: 10.1111 / j.1439-0396.2008.00901.х
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., Мурби, Дж. М., и Ньюболд, К. Дж. (2012a). Расщепление бактериального белка разными простейшими группами рубца. J. Anim. Sci. 90, 4495–4504. DOI: 10.2527 / jas.2012-5118
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., Пинлоче, Э., Ньюболд, К. Дж., И Балселлс, Дж. (2012b). Влияние диеты и отсутствия простейших на микробное сообщество рубца и на репрезентативность бактериальных фракций, используемых при определении синтеза микробного белка. J. Anim. Sci. 90, 3924–3936. DOI: 10.2527 / jas.2011-4802
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Доро, М., Эдвардс, Дж. Э., Мурби, Дж. М., Пинлоче, Э. и Ньюболд, К. Дж. (2012c). Изменения в микробиоте рубца из-за типа углеводов и уровня белка, потребляемого молочным скотом, связаны с изменениями ферментации рубца. J. Nutr. 142, 1684–1692. DOI: 10.3945 / jn.112.159574
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., и Ньюболд, К. Дж. (2014). Изучение метаногеновых сообществ, связанных с различными популяциями простейших в рубце. FEMS Microbiol. Ecol. 90, 663–677. DOI: 10.1111 / 1574-6941.12423
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче А., де ла Фуэнте Г. и Ньюболд К. Дж. (2015). Влияние прогрессивной инокуляции овец, свободных от фауны, холотобоязненными простейшими и всей фауной на ферментацию рубца, микробное разнообразие и выбросы метана. FEMS Microbiol. Ecol. 362, 1–10. DOI: 10.1093 / фемсек / fiu026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Белжецкий, Г., Ньюболд, К. Дж., МакЭван, Н. Р., Макинтош, Ф. М., и Михаловски, Т. (2007). Характеристика амилолитических свойств простейших инфузорий рубца Eudiplodinium maggii . J. Anim. Feed Sci. 16, 590–606.
Google Scholar
Bera-Maillet, C., Devillard, E., Cezette, M., Jouany, J.П., Форано Э. (2005). Ксиланазы и карбоксиметилцеллюлазы простейших в рубце Polyplastron multivesiculatum , Eudiplodinium maggii и Entodinium sp. FEMS Microbiol. Lett. 244, 149–156. DOI: 10.1016 / j.femsle.2005.01.035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берд, С. Х., Хегарти, Р. С., Вудгейт, Р. (2008). Сохранение эффектов дефаунации на пищеварение и выработку метана у овец. Aust. J. Exp. Agric. 48, 152–155. DOI: 10.1071 / EA07298
CrossRef Полный текст | Google Scholar
Boxma, B., Ricard, G., van Hoek, A.H., Severing, E., Moon-van der Staay, S.Y., van der Staay, G.W., et al. (2007). Гидрогеназа [FeFe] Nyctotherus ovalis имеет химерное происхождение. BMC Evol. Биол. 7: 230. DOI: 10.1186 / 1471-2148-7-230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бродерик, Г.А., Мерчен, Н.Р. (1992). Маркеры для количественного определения синтеза микробного белка в рубце. J. Dairy Sci. 75, 2618–2632.
PubMed Аннотация | Google Scholar
Карлсон, С. А., Шарма, В. К., МакКаддин, З. П., Расмуссен, М. А., и Франклин, С. К. (2007). Участие гена Salmonella геномного острова 1 в рубце, опосредованном простейшими, усиление инвазии для множественной устойчивости к антибиотикам Salmonella enterica серовар Typhimurium . Заражение.Иммун. 75, 792–800. DOI: 10.1128 / IAI.00679-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Case, R. J., Boucher, Y., Dahllöf, I., Holmström, C., Doolittle, W.F., and Kjelleberg, S. (2007). Использование генов 16S рРНК и rpoB в качестве молекулярных маркеров для исследований микробной экологии. Заявл. Environ. Microbiol. 73, 278–288. DOI: 10.1128 / AEM.01177-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кавальер-Смит, Т.(2005). Экономика, скорость и размер имеют значение: эволюционные силы, движущие миниатюризацией и расширением ядерного генома. Ann. Бот. 95, 147–175. DOI: 10.1093 / aob / mci010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чандрамони, Джадхао, С. Б., Тивари, К. М., Хак, Н., Мурари Лал и Хан, М. Ю. (2001). Энергетический метаболизм и выработка метана у фаунированных и дефонтированных овец, которых кормили двумя рационами с одинаковым соотношением концентратов и грубых кормов (70: 30), но разного состава. Азиатско-Австралийский. J. Anim. Sci. 14, 1238–1244. DOI: 10.5713 / ajas.2001.1238
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cieslak, A.
0: 9rio 170009: 9rio 170009: 9 Английский:
Vol. 1, Wirbellose Thiere, von C. Th. против Зибольда, 1848 г.Vol. 2, Wirbelthiere, von H. Stanius, 1846, [4]. Абластерия- Vermes I + Vermes II [Sammtlische Würmer]
- Mollusca [Molluscken]
- Иглокожие [Echinotermen]
- Членистоногие [Arthropoden]
- Vertebrata [Vertebraten]
- Простейшие
- Metazoa [= Blastozoa, p. 19]
- Anaemaria
- Haemataria = Pentablasteria
К.Ф. Зима, Лейпциг.- Protophyta (Protista Vegetalia)
- Класс I. Фитарча
- Фитомонера
- Chromaceae
- Ordnung 2. Chroococceae
- Ordnung 3. Nostochineae
- Класс II. Диатомеи
- Класс III. Космарии (Desmidieae)
- Класс IV. Пальмеллярии (Palmellaceae)
- Класс V. Siphoneae (Coeloblastae)
- Класс I. Фитарча
- Простейшие (Protista animalia)
- Зоарча (Zoocytoda)
- Zoomonera
- Бактерии
- Сфаэробактерии
- Рабдобактерии
- Cytarcha
- Лобоса (Амебина)
- Gregarinae (Sporozoa)
- Инфузории
- Flagellata (Мастигофора)
- Реснички
- Acinetae (Suctoria)
- Rhizopoda
- Mycetozoa (Myxomycetes)
- Heliozoa
- Таламария (Foraminifera)
- Радиолярия
- Зоарча (Zoocytoda)
- Thallota (Thallophyta)
- Водоросли (Phycophyta)
- Zygnemaceae
- Conferveae
- Fucoideae
- Флориды
- Characeae
- Грибы (Inophyta)
- Водоросли (Phycophyta)
- Prothallota (Мезофита)
- Phanerogamae (Anthophyta)
- Gymnospermae
- Cycadeae
- Хвойные
- Gnetaceae
- покрытосемянных
- Gymnospermae
- кишечнополостные (Coelenterata, Zoophyta, Anaemaria)
- Gastraeades
- Губки
- Книдария
- Платоды
- Целомария (Bilaterata, Bilateria, Haemataria)
- Гельминты
- Моллюска
- Иглокожие
- Сочленений
- Туниката
- Позвоночные
- Archephyta (Прогонелла)
- Пробионты
- Хромовые (Phycochromaceae)
- Алгарии (Paulosporata)
- Paulotomeae
- Конъюгаты
- Диатомеи
- Альгетты (Zoosporata)
- Мастигота
- Мелеталия
- Siphoneae
- Thallophyta
- Водоросли [Vol. 1, стр. 302]
- Chlorophyceae
- Charaphyceae
- Phaeophyceae
- Rhodophyceae
- Mycetes [Vol. 1, стр. 316]
- Аскомицеты (Ascodiomycetes)
- Basimycetes (Базидиомицеты)
- Лихены [Vol. 1, стр. 325]
- Водоросли [Vol. 1, стр. 302]
- Diaphyta [Vol. 1, стр. 330]
- Bryophyta [Vol. 1, стр. 336]
- Pteridophyta
- Anthophyta
- Gymnospermae
- Покрытосемянное
- Архезоа (= Зоарчега)
- Бактерии (= Bactromonera)
- Coccillida
- Bacillida
- Спириллида
- Зоомонера (= ул. Монера)
- Бактерии (= Bactromonera)
- Грибки (= Sporozoa)
- Ризопода
- Инфузории
- кишечник (акеломия)
- Gastraeades
- Губки
- Книдария
- Платоды
- Билатерия (Coelomaria)
- Гельминты
- Моллюска
- Сочленений
- Иглокожие
- Туниката
- Vertebrata
- Euthallophyta
- Родофита
- Хризофита
- Pyrrophyta
- Euglenophyta
- Gonidiophyta
- Charophyta
- Phaeophyta
- Myxophyta
- Микофита
- Простейшие
- Sarcodina
- Фораминиферы
- Актиноподы
- Грибки
- Реснички
- Suctoria
- Мезозоа
- Паразоа
8
Самые актуальные
Свежий контент
Тип изображенияВсе изображения
Фото
Векторы
Иллюстрации
ОриентацияВсе ориентации
Горизонтально
Определение, типы, болезни, профилактика и многое другое
Патогены — это организмы, которые могут вызывать болезнь.Различные типы патогенов и тяжесть вызываемых ими заболеваний очень разнообразны.
В этой статье мы рассмотрим различные патогены, их влияние на людей и болезни, которые они вызывают. Мы также объясняем, как распространяются патогены и как снизить риск заражения.
Патоген приносит болезнь своему хозяину. Другое название возбудителя — инфекционный агент, поскольку они вызывают инфекции. Как и в случае с любым организмом, патогены отдают приоритет выживанию и воспроизводству.
Иммунная система человеческого организма действует как защита от патогенов.Организм может легко бороться с некоторыми патогенами, но другие потенциально смертельны.
Существует пять основных типов патогенов:
Бактерии
Бактерии — это микроскопические патогены, которые быстро размножаются после попадания в организм. Они могут выделять токсины, которые повреждают ткани и вызывают болезни.
Врачи обычно назначают антибиотики для лечения бактериальных инфекций, но некоторые бактерии становятся устойчивыми к этим лекарствам.
Однако не все бактерии являются патогенными.В организме существует множество типов безвредных бактерий, некоторые из которых могут даже поддерживать основные функции организма.
Вирусы
Вирус, который меньше бактерий, проникает в клетку-хозяина. Затем он реплицируется, производя сотни и тысячи новых вирусов, которые продолжают заражать другие клетки-хозяева. Вирусы могут передаваться от человека к человеку различными путями, в том числе:
- через респираторные капли, которые перемещаются по воздуху
- через контакт с кровью человека с инфекцией
- через контакт с жидкостями организма человека, инфицированного инфекцией
Грибы
Существуют тысячи видов грибов, некоторые из которых вызывают болезни у людей.Распространенные грибковые заболевания кожи включают микоз стопы и стригущий лишай. Эти состояния заразны и могут передаваться от человека к человеку.
Исследование, проведенное в Trends in Microbiology , показало, что у грибковых патогенов развивается способность к памяти. Они могут использовать сигналы в теле, чтобы предвидеть неминуемую угрозу своему выживанию, к которой они затем могут подготовиться.
Протисты
Эти одноклеточные организмы вызывают болезни у своего хозяина. Они заражают другие организмы, чтобы выжить и размножаться.
Простейшие патогены поражают растения и продовольственные культуры. Продукты, содержащие простейшие, могут вызвать дизентерию — инфекцию кишечника, вызывающую диарею.
Протистские патогены также могут быть паразитическими и жить в других организмах, таких как комары. Протисты вызывают малярию через укусы комаров.
Паразитические черви
Паразитические черви, также известные как гельминты, достаточно велики, чтобы их можно было увидеть невооруженным глазом, и они могут жить во многих частях тела.Некоторые черви включают:
- Плоские черви: К ним относятся ленточные черви, которые обитают в кишечнике.
- Колючие черви: Этот вид червей обитает в кишечнике.
- Круглые черви: Эти черви могут выжить в желудочно-кишечном тракте и лимфатической системе.
Патогены могут вызывать множество различных заболеваний, некоторые из которых более серьезны, чем другие. Человеческие тела богаты питательными веществами и могут обеспечить патогену идеальную среду для роста и размножения.
Степень тяжести инфекций, вызываемых патогенами, может быть разной. Некоторые инфекции могут быть легкими, в то время как другие могут быть опасными для жизни. Например, простуда — это легкая вирусная инфекция по сравнению со смертельной болезнью, вызванной вирусом Эбола.
Болезни, вызываемые бактериальными патогенами, включают:
Некоторые ученые считают, что вирусы не являются живыми организмами. Вот некоторые причины:
- у них нет клеток
- они не могут воспроизводиться, не вторгаясь в живую клетку
- они не реагируют активно на изменения в окружающей среде
К вирусным инфекциям относятся:
Около 300 видов грибов патогенны для человека.Как и бактерии и вирусы, они могут оказывать значительное влияние на здоровье человека.
Грибки вызывают множество различных типов заболеваний, в том числе:
- астму
- инфекции кожи и ногтей
- инфекции легких, такие как пневмония
- инфекции кровотока
- менингит
Протозойные заболевания являются причиной большинства протистических заболеваний. Простейшие — это одноклеточные микроорганизмы, которые питаются другими микроорганизмами, органическими тканями и мусором.К протистским заболеваниям относятся:
- дизентерия
- малярия
- Африканский трипаносомоз или сонная болезнь
Паразитические черви вызывают множество болезней, в том числе:
Патогены могут распространяться различными путями. Например, прямой контакт кожа к коже во время секса может привести к инфекциям, передаваемым половым путем (ИППП).
При кашле или чихании болезнетворные микроорганизмы могут распространяться через крошечные капельки в воздухе. Эти капли могут содержать микроорганизмы, которыми дышат другие люди.
Микроорганизмы также могут попадать прямо в кишечник, когда человек употребляет зараженную пищу или воду.
Укусы инфицированных насекомых также могут распространять болезнь. Например, клещи с бактериальной инфекцией могут вызвать болезнь Лайма, если кого-то укусили, а комары с вирусной инфекцией могут вызвать болезнь, вызванную вирусом Зика.
Наряду с поддержанием хорошего общего состояния здоровья человек может предпринять определенные шаги, чтобы снизить риск заражения патогенами. К ним относятся:
- Регулярное мытье и сушка рук: Регулярное мытье рук имеет решающее значение для предотвращения распространения болезней.Люди должны мыть их не менее 20 секунд водой с мылом или дезинфицирующим средством на спиртовой основе. Это особенно важно после занятий на открытом воздухе, прикосновений к животным, посещения туалета, ухода за больными, чихания или кашля.
- Будьте в курсе рекомендованных вакцин: Вакцины против инфекционных заболеваний, таких как грипп, коклюш и корь, доступны для младенцев, детей и взрослых. Вакцины стимулируют иммунную систему распознавать патогены, уничтожать их и предотвращать будущие инфекции.
- Поддержание чистоты поверхностей: Загрязненные поверхности или предметы могут передавать болезни. Исследование, проведенное в аэропорту Финляндии, обнаружило вирусное заражение в нескольких местах на поверхностях, к которым люди часто прикасаются.
- Уборка кухонь и ванных комнат: Важно поддерживать в чистоте столешницы и кухонные поверхности, особенно перед приготовлением еды. В ванной также может быть высокая концентрация инфекционных агентов.
- Оставаться дома в случае болезни: Люди с инфекциями должны избегать посещения школы, работы и тесного контакта с другими людьми, чтобы снизить вероятность распространения патогена.
- Как избежать укусов насекомых: Люди могут сделать это, надев защитные средства от насекомых и подходящую одежду, такую как рубашки с длинными рукавами, брюки и шляпы, в местах, где они могут встретить насекомых.
- Практика безопасного секса: Использование методов безопасного секса, включая использование презервативов и регулярные проверки сексуального здоровья, может снизить вероятность заражения ИППП.
- Получение медицинской консультации: Человек может поговорить с медицинским работником, если есть признаки инфекции или лечение не работает.
Патогены поражают все живые организмы и вызывают заболевания у людей множеством различных способов.
Они могут повредить ткани или клетки, вырабатывая токсины в процессе репликации.
В то время как некоторые патогены вызывают легкие проблемы, другие могут быть опасными для жизни. Медицинские эксперты продолжают изучать болезни, вызываемые патогенами, чтобы больше узнать об их причинах и способах их лечения.
Паразиты — простейшие
Простейшие теперь имеют свой собственный порядок в живом царстве, хотя когда-то их считали крошечными животными, поскольку они были подвижными и не были зелеными, как растения — теперь у них есть отдельное царство в соответствии со схемой классификации для всей жизни на Земле.Многие простейшие паразиты на человеке и вызывают от легких до чрезвычайно опасных заболеваний — некоторые из них паразитируют на человеке и из-за своего крошечного размера образуют простейших из всех внутренних паразитов. Клеточная организация простейших намного сложнее, а размер намного больше, хотя, как и бактерии, все простейшие являются одноклеточными организмами — весь организм состоит только из одной клетки. Тип окружающей среды, в которой обитает большинство простейших, очень космополитичен, и их распространение повсеместно, они встречаются почти в любой водной среде и во влажных местах, включая пруды, реки, озера, аквариумы, канавы и т. Д.
Мазь для ногтей
Лучшее, 100% натуральное средство для ежедневного ухода за ногтями в идеальной форме.
Большинство простейших не причиняют вреда животным или людям и считаются свободноживущими, то есть им не требуется хозяин для жизни и размножения. Однако есть несколько простейших, которые являются паразитами, среди этих паразитических организмов некоторые могут вызвать чрезвычайно серьезные проблемы со здоровьем у людей, которым не повезло заразиться простейшими.Первым патогенным простейшим, идентифицированным еще в 1875 году, было простейшее. Паразитические простейшие были разновидностью амебы под названием Entamoeba histolytica, которая также известна как дизентерия, вызывающая амебу — она очень распространена и поражает людей во всем мире. Антон ван Левенгук, один из первых и, возможно, один из величайших ранних микроскопистов и клеточных биологов, заметил, что в его собственных фекалиях скользят простейшие. Крошечная микроскопическая форма жизни принадлежала к роду Giardia, и с момента открытия Левенгука такие паразиты, как Giardia lamblia и простейшие виды Cryptosporidium, были связаны с причиной болезненных кишечных инфекций и вспышек тяжелой диареи во всем мире.Вода, загрязненная фекалиями человека или животных, обычно содержит этих двух паразитических простейших, и употребление такой загрязненной воды является обычной причиной заболеваний.
Мазь от грибка
Вся сила фармацевтических фунгицидов — но без агрессивных химикатов.
Бабезиоз
Паразитические простейшие, известные как Babesia microti, ответственны за вспышки этого заболевания. Паразит проникает в эритроциты различных животных, которых заражает, и вызывает типичные симптомы, столь характерные для этого состояния.Основным резервуаром болезни являются грызуны, способ передачи болезни — через клещей, возможно, тех же видов, которые участвуют в передаче болезни Лайма — таким образом, клещи являются основными переносчиками болезни и распространяют ее. от грызунов-хозяев к другим животным, включая человека, путем укуса. Распространенное заболевание, называемое техасской лихорадкой, иногда также известное как клещевая лихорадка крупного рогатого скота, которое поражает крупный рогатый скот, вызывается простейшими, связанными с этим видом.
Состояние вызывает симптомы у пораженных людей, все эти симптомы, по-видимому, связаны или похожи на симптомы, наблюдаемые у пациентов, страдающих малярией, которая также вызывается простейшими.Физические симптомы включают стойкую усталость и физическую усталость в течение семи дней после первоначального контакта с паразитом, за этим начальным периодом следует период, когда у пациента появляется внезапный озноб, а также другие состояния, такие как лихорадка, сильная головная боль и болезненность мышц в целом. тело. Некоторые больные часто прогрессируют до анемии и почечной недостаточности, хотя большинство симптомов легкие, и состояние в основном проходит через несколько недель. Однако известны случаи смертельного исхода в некоторых случаях, особенно в отношении пациентов старших возрастных групп.Для этого состояния не существует специальных лекарств, хотя симптоматическое лечение с использованием тех же лекарств, которые используются для лечения малярии, было опробовано с некоторым успехом — профилактика является лучшей политикой в отношении этого заболевания, и случаи заболевания редки.
Болезнь Шагаса
Простейшее заболевание Темы — болезнь Шагаса — серьезная проблема для здоровья в Латинской Америке. За эти состояния отвечает простейший организм, известный в науке как Trypanosoma cruzi, болезнь широко распространена в Центральной и Южной Америке, где она представляет собой проблему для общественного здравоохранения.Переносчиком этой болезни является насекомое, известное как целующиеся клопы, иногда называемое «конусообразным насекомым». Люди, живущие в районах с маргинальной жилой инфраструктурой, зараженной этим насекомым, могут заразиться семейством болезни Шагаса. Насекомые-переносчики относятся к семейству насекомых Reduviidae, подсемейству Triatominae и роду Triatoma — это насекомое снова очень распространено в Южной Америке. Опоссумы и броненосцы, а также инфицированные люди служат резервуарами для простейших, насекомое, которое является кровососом, передает паразита из таких источников неинфицированным животным и людям — инфекции обычно распространяются среди людей в одной семье — насекомое передает паразита от хозяин за хозяином, поскольку он сосет кровь и оставляет отходы на новых хозяевах.
Восстанавливающее средство для кожи
Усовершенствованный, 100% натуральный ревитализатор, который сделает вашу кожу сияющей и молодой.
После того, как клоп поедает инфицированное млекопитающее, он передает простейшие, которые уже прошли по его кишечнику, с отходами, которые он оставляет на коже неинфицированного животного-хозяина или человека. Когда человек непроизвольно царапает эту область, отходы в области укусов насекомых загрязняют рану, и повторяющиеся царапины вызывают ушибы кожи — простейшие также часто передаются через глаза — когда человек трет отходы, простейшие попадают в тело, используя сломанный участок кожи.Тропическое заболевание было идентифицировано в 1912 году бразильским врачом Карлосом Шагасом — отсюда и название болезнь Шагаса.
Появление небольшой красной язвочки на коже или развитие опухших и покрасневших, воспаленных глаз являются наиболее частыми ранними симптомами заболевания у человека. Через несколько недель язва на пораженном участке кожи, кажется, заживает и выделяет твердую корку, которая вскоре исчезает, и остается небольшой темный шрам в этой области — если человеку повезет. В других случаях первоначальные симптомы могут сменяться появлением лихорадки, развитием отека лимфатических узлов и узлов, а также развитием кожной сыпи по всему телу.Тяжелые формы болезни Шагаса могут поражать сердце, и такие симптомы, как боли в груди и внезапная одышка, становятся обычным явлением. Также могут возникнуть судороги, если заболевание поражает нервную систему — в таких случаях в непрерывных конвульсиях может быть вовлечено все тело. В некоторых случаях это состояние может привести к летальному исходу — это остается риском для здоровья.
Лямблиоз (бобровая лихорадка)
Эта лихорадка является паразитарным заболеванием человека, которое является наиболее распространенной формой паразитарного заболевания в некоторых частях Соединенных Штатов и других районах Северной Америки, в частности, в западных регионах Канады.Причина лихорадки — крошечный паразитический простейший организм, который называется Giardia lamblia. Паразитический организм остается на внутренней поверхности кишечника человека и, в частности, в больших количествах обнаруживается в тонком кишечнике пораженных людей. Обычным физическим симптомом, помимо повышения температуры тела, является стойкая инфекционная диарея.
Обычный источник распространения паразита — употребление зараженной пищи и воды. Если не поддерживать строгие и надлежащие санитарные условия, многие детские центры и дома престарелых становятся районами, в которых часто происходят вспышки лямблиоза, поэтому дети становятся одними из первых, кто становится жертвами паразита.Путь передачи паразита в большинстве случаев заражения происходит через попадание паразитарных цист в пищеварительный тракт вместе с зараженной пищей или водой — такие источники обычно содержат фекальные отходы людей и других животных-переносчиков.
Примерно через три недели после первоначального заражения паразитом хроническая диарея начинает поражать людей, страдающих лямблиозом — это обычные временные рамки для появления этого симптома у большинства пациентов.В дополнение к этому симптому у пораженных людей также появляются симптомы тошноты и другие жалобы, такие как стойкий и сильный дискомфорт в животе. Пищеварительная система нарушается во время длительных периодов инфекции, конечным результатом может быть внезапная потеря веса у пораженного человека — это происходит из-за очень плохого всасывания переваренной пищи из области кишечника. Человек может оставаться инфицированным в течение нескольких лет подряд, хотя в некоторых случаях у пораженных людей начинается спонтанное выздоровление от симптомов.
Раньше считалось, что основным резервуаром простейших паразитов в дикой природе являются бобры — так считалось в течение многих лет, пока недавние доказательства не показали обратного. В настоящее время известно, что домашний скот и овцы являются основными резервуарами болезни, согласно данным недавних исследований. Сейчас считается, что бобры, как и люди, вероятно, могут быть вторичным хозяином, а не резервуаром для паразита. Было показано, что такие животные, как бобры, полевки, ондатры и крысы-кенгуру, обладают способностью переносить этого паразита — все они могут действовать как хозяева, похожие на людей.
Сонная болезнь (африканский трипаносомоз)
Мелкая рыба, такая как простейшие паразиты, вызывает опасное заболевание, известное как сонная болезнь — это смертельное заболевание, широко распространенное в некоторых районах Африки. Простейшие — это трипаносома — разновидность нематодных червей, этот червь может плавать в крови своих жертв, ударяя хлыстом, как хвостом, или, точнее, своим жгутиком. Печально известная муха цеце несет ответственность за передачу этого смертельного заболевания. Муха кусает и сосет кровь, как только паразит попадает в организм.Человек медленно теряет сознание, и смерть наступает, если паразиту удается проникнуть в области спинного и головного мозга своих жертв. Шансы умереть от болезни очень высоки, если человек инфицирован, а сонная болезнь, как известно, уничтожает все домашние запасы мяса в районе — поскольку этот паразит также заражает домашний скот, например, крупный рогатый скот. Смертность как среди людей, так и среди животных высока в районах, где болезнь носит эндемический характер, и профилактика является лучшим вариантом против нее.Среда обитания мухи цеце широко распространена в тропической Африке, и некоторые ученые и эксперты в области здравоохранения считают, что с повышением температуры из-за глобального потепления существует возможность значительного расширения этой среды обитания. В таком случае муха цеце, как и паразит, может заразить космополитичную территорию. Это не единственный наихудший сценарий для одного насекомого-переносчика; Обычно считается, что глобальное потепление может расширить ареал обитания других насекомых-переносчиков, а также привести к распространению большего количества тропических паразитов в другие регионы мира.
Лейшманиоз
Паразитические простейшие лейшмании ответственны за это изнурительное и часто смертельное заболевание, которое широко распространено в некоторых тропических и субтропических регионах мира. Крошечная кровососущая песчаная муха ответственна за распространение этого смертоносного паразитического простейшего — эта муха является единственным переносчиком паразита. Состояние, известное как висцеральный лейшманиоз, иначе известное как кала-азар, является наиболее летальной стадией прогрессирования инфекции. В эндемичных районах болезнь способна уничтожить две трети населения некоторых серьезно пострадавших деревень — это особенно верно в отношении сельских районов в тропиках, где медицинская помощь плохая или полностью недоступна.Постоянная лихорадка, темно-серая кожа, наличие тяжелой анемии — одни из первых физических симптомов кала-азар, кроме того, сильно увеличиваются селезенка и печень, что приводит к неприглядному вздутию живота. область. Болезнь может стать смертельной очень быстро, если симптомы остаются без лечения в течение длительного периода времени. Кала-азар обычно лечится с помощью лекарств, содержащих сурьму тяжелых металлов — такие лекарства являются единственным эффективным средством против паразита.Белые кровяные тельца в организме — это область, в которой простейшие лейшмании фактически проживают в организме — это защитные клетки, которые должны были поглотить простейшие, когда началась инфекция. Белые кровяные тельца поглощают простейшее, но не могут его разрушить или переварить. В результате простейшие лейшмании процветают в тех самых белых кровяных тельцах, от которых зависит его собственная защита. Лейкоциты, в которых проживают простейшие, со временем разрушаются, подобно заболеванию, подобному ВИЧ, что, естественно, наносит огромный ущерб иммунной системе больного человека.Оппортунистические и смертельные бактериальные инфекции, такие как пневмония или дизентерия, преодолевают слабую и неэффективную защиту организма, так как лейкоциты в большом количестве умирают из-за простейших. Если это продолжается, человек умирает от симптомов.
Малярия
Настоящее значение слова «малярия» — просто «плохой воздух». Этот плохой воздух должен был вызывать у людей перемежающуюся лихорадку, когда они впервые столкнулись с этой болезнью. Действительно, древнегреческий врач Гиппократ предположил, что это было вызвано появлением миазмов на человеке — это смертельные туманы, которые предположительно нарушили тщательный баланс, существующий в четырех жидкостях тела, — что является древним медицинским верованием в кровь. мокрота, черная желчь и желтая желчь.С другой стороны, согласно древней китайской медицинской системе, дисбаланс между инь и ян в организме должен был вызвать приступообразную лихорадку, а именно малярию, а дисгармония между этими двумя противоположными мужскими и женскими жизненными силами в организме приписывалась быть причиной состояния. Несколько столетий назад великий систематик и биолог Карол Линней выдвинул гипотезу о том, что причиной малярии может быть присутствие крошечных взвешенных частиц глины в питьевой воде, эти взвешенные твердые частицы, как предполагается, привели к закупорке кровеносных сосудов и нарушение кровообращения должно было вызвать болезнь.Все это были теории о причине малярии, выдвинутые до появления микроскопа и в самом начале современной науки, истинный возбудитель малярии был идентифицирован только в 1898 году — британский доктор Росс правильно объяснил бы. Это произошло с укусом инфицированного комара, и он продолжил идентифицировать смертельный патоген как тип простейших.
Только укус самок комаров вида anopheles ответственен за передачу малярии людям и некоторым другим млекопитающим, которые могут быть инфицированы.Самки анофелесов — единственные москиты, которые являются переносчиками малярии человека; самцы комаров не кусаются и не могут распространять болезнь. Когда самка комара Anopheles кусает свою жертву, она высасывает кровь через прокол в коже особи и в то же время вводит тысячи нитевидных личинок простейших, называемых спорозоитами — эти личинки принадлежат к видам простейших плазмодиев. Оказавшись в теле жертвы, спорозоиты отправятся через кровоток прямо в печень, где каждая из личинок начинает инфицировать и жить в собственной клетке печени.В этих клетках печени спорозоиты собираются и начинают быстро делиться, многократно размножаясь, производя до сорока тысяч спор на каждую личинку. По прошествии двух недель производство и размножение спор в клетках печени продолжается, в течение этого периода нет абсолютно никаких признаков или симптомов болезни, и человек здоров. Однако здесь закладываются первые семена полномасштабной малярии, которые начинают вторжение в организм.
Разрыв клетки печени, в которой образовались споры, вызывает первые клинические признаки и является начальной стадией атаки на организм — у человека наблюдается сильное потоотделение и на этой стадии развивается высокая температура.Теперь лопнувшие клетки печени начинают выпускать многочисленные споры прямо в кровоток жертвы. На этом этапе каждая отдельная спора теперь вторгается в эритроцит, где она будет находиться какое-то время, медленно расходуя содержание гемоглобина в клетке и увеличиваясь в размере. Затем спора делится или раскалывается на множество более мелких фрагментов — это бесполое средство образования тысяч других спор — научное название спор на этой стадии — мерозоиты. Наконец, воля красных кровяных телец также лопается, и это высвобождает все присутствующие в них мерозоиты, а затем вторгаются в новые клетки крови по соседству.Этот циклический эффект, а также повторяющееся инфицирование и разрушение красных кровяных телец в конечном итоге приводят к хроническому малярийному заболеванию и могут привести к смерти человека.
Разрыв и вторжение красных кровяных телец — это очень своевременное дело, и существует удивительная синхронность всего процесса заражения в кровотоке жертвы. Основные причины одновременного разрыва миллионов инфицированных эритроцитов с высвобождением их мерозоитов до сих пор полностью не изучены и требуют изучения.Худшие приступы тряски и озноба происходят из-за своевременного и массового разрыва красных кровяных телец в организме.
Окончательный процесс трансформации этих мерозоитов в мужские или женские гаметоциты также начинает происходить после нескольких раундов инфицирования клеток крови и их последующего разрушения — эти гаметоциты являются репродуктивными клетками паразита плазмодия, а на этой стадии инфекции — человеческих клеток. фазу жизненного цикла паразита можно считать завершенной.Если пациенту повезет, он или она выжили бы на этой стадии, однако на этой стадии он заразен, потому что, если самка комара Anopheles высосет эту зараженную малярией кровь, она также поглотит миллионы паразитических клеток гаметоцитов. Гаметоциты простейших будут претерпевать дальнейшее развитие в яйцеклетки и сперматозоиды в теле самки комара, начиная следующую стадию у их нового хозяина, и оплодотворение этих гамет будет происходить в теле насекомого.
Цикл завершается, когда оплодотворенные яйца паразита превращаются в нитевидные спорозоиты в теле комара, а затем они попадают в слюнные железы комара, готовые для передачи новому человеку-хозяину.Именно в это время самка комара становится смертельным переносчиком малярийного паразита, и ее укус переносит новые спорозоиты, чтобы начать новый цикл внутри другого человека-хозяина.
Очень интересный факт заключается в том, что люди с серповидноклеточной анемией и даже те, кто страдает ее более легкой формой, или те, у кого есть черта серповидно-клеточной анемии в красных кровяных тельцах, вероятно, имеют гораздо большую защиту от малярии. предлагает. Серповидно-клеточная анемия деформирует красные кровяные тельца тела, и это изменение формы клеток напрямую препятствует и предотвращает проживание простейших внутри клетки.Это эволюционный ответ на малярию, поскольку люди, страдающие серповидно-клеточной анемией, как правило, в основном сконцентрированы в регионах мира с самым высоким уровнем заражения малярией — серповидно-клеточная форма красных кровяных телец является только общим генетическим дефектом в человеческих популяциях, круглый год подверженных малярийным заболеваниям.
Итак, как именно лечат малярию? И какое оружие применяется против болезни. Лекарства и противомалярийные препараты являются одними из инструментов, с помощью которых врачи и ученые непосредственно лечат малярию. Болезнь также можно контролировать, регулируя популяцию насекомых-переносчиков.Цитата «Лечите пациента, а не комара» часто использовала Роберт Кох, блестящий микробиолог, получивший известность благодаря выявлению туберкулеза. Единственным противомалярийным лекарством, которое было доступно в его дни — а это было в конце девятнадцатого века, было лекарство растительного происхождения, известное как хинин.
Противомалярийный препарат хинин — это медицинский препарат, полученный из коры хинного дерева, он веками использовался в традиционных противомалярийных средствах туземцами Южной Америки.На сегодняшний день он все еще используется как отличное противомалярийное лекарство во многих частях мира. К сожалению, высокие дозировки препарата токсичны и могут вызвать глухоту при использовании в высоких дозах, необходимых для полного удаления паразита плазмодия из организма. До получения менее вредных производных в 1930-х годах его использование было обширным и универсальным — теперь его заменяют менее вредные производные, синтезированные из того же растения. Хлорохин — это основное синтезируемое лекарство, используемое в наши дни для лечения малярии.Однако в качестве препарата выбора при лечении малярии хлорохин в большинстве стран мира был вытеснен другим противомалярийным мефлохином.
Другие полезные травы
Мазь для кожи
100% натуральная формула для всех проблем с кожей. Отлично подходит для диабетиков.
комментариев
Frontiers | Роль инфузорий простейших в рубце
Введение
Впервые описанные в 1843 году, простейшие Rumen с их поразительной внешностью считались важными для благополучия своего хозяина.Однако, несмотря на то, что простейшие могут составлять до 50% биомассы в рубце, роль простейших в микробной экосистеме рубца остается неясной (Williams and Coleman, 1992). Здесь мы оцениваем недавнюю информацию о роли простейших инфузорий в микробной экосистеме рубца.
Разнообразие и филогения простейших инфузорий рубца
С тех пор, как они были впервые обнаружены Gruby и Delafond (1843), исследования простейших в рубце основывались на морфологической идентификации с помощью оптической микроскопии.В настоящее время нет коллекций культур инфузорий рубца, поэтому исследователи должны использовать микрофотографии для идентификации (Ogimoto, Imai, 1981; Williams and Coleman, 1992) или штриховые рисунки (Dogiel, 1927; Kofoid and MacLennan, 1930, 1932, 1933; Latteur , 1968, 1969, 1970; Dehority, 1993). Широко признано, что микроскопическая идентификация и подсчет представляют собой золотой стандарт для анализа структуры сообщества инфузорий в исследованиях рубца (Williams and Coleman, 1992). Однако полиморфная природа этих микробов (Dehority, 1994, 2006) требует от исследователя высокого уровня опыта для идентификации инфузорий рубца на уровне родов или видов (Dehority, 2008).Таким образом, необходимы альтернативные способы идентификации простейших, поскольку микроскопические методы трудоемки и весьма требовательны.
Ранние молекулярные исследования простейших ресничек рубца были сосредоточены на секвенировании генов 18S рРНК для выяснения внутренней филогении класса Litostomatea (Embley et al., 1995; Wright et al., 1997). Embley et al. (1995) были первыми, кто описал монофилетическое положение анаэробных инфузорий рубца Dasytricha ruminantium и Polyplastron multivesiculatum на основе генов 18S рРНК, а Wright et al.(1997), Wright and Lynn (1997a, b) предоставили молекулярные данные для Diplodinium dentatum , Entodinium caudatum , Epidinium caudatum , Eudiplodinium maggii , Isotrichaintechinalis108 Oyinoscopusa , .
С тех пор были выполнены многочисленные качественные и количественные исследования разнообразия инфузорий; в этих исследованиях использовались различные методы, включая полиморфизм длины рестрикционных фрагментов (RFLP; Regensbogenova et al., 2004a, Tymensen et al., 2012b), денатурирующий градиентный гель-электрофорез (DGGE; Regensbogenova et al., 2004b, McEwan et al., 2005; de la Fuente et al., 2009; Belanche et al., 2010a, Kittelmann and Janssen, 2011), ПЦР в реальном времени (qPCR; Sylvester et al., 2004; Skillman et al., 2006; Belanche et al., 2010a, Kittelmann and Janssen, 2011), флуоресцентная гибридизация in situ (FISH; Xia et al. ., 2014) и секвенирование следующего поколения (Kittelmann et al., 2013, 2015; Ishaq, Wright, 2014; Moon-van der Staay et al., 2014).
Микроскопия имеет несколько преимуществ перед молекулярными методами на основе ПЦР для изучения простейших инфузорий. Во-первых, хотя подавляющее большинство кишечных инфузорий охарактеризовано морфологически, для многих наблюдаемых родов и видов отсутствуют эталонные последовательности гена 18S рРНК. Во-вторых, как обсуждается ниже, вариация числа копий генов рибосомной РНК в разных родах или в разных условиях роста может исказить наблюдаемые пропорции этих родов в образце (Medinger et al., 2010). Исследования с использованием обзоров генов 18S рРНК показывают очевидное более высокое разнообразие инфузорий, чем оценка с помощью обычных морфологических методов (Moon-van der Staay et al., 2014).
Как и другие инфузории, простейшие в рубце содержат два типа ядер: микроядро и макронуклеус. Микроядро имеет четко видимые хромосомы, диплоидно и синтезирует только следы РНК. Макронуклеус не содержит различимых хромосом, имеет во много раз больше диплоидного количества ДНК, делится амитотически и обеспечивает практически всю РНК, необходимую для вегетативной жизни клетки (Prescott and Murti, 1974).Чрезвычайно высокое число копий рДНК в макронуклеарном геноме инфузорий, как было обнаружено ранее (Gong et al., 2013), можно понять, учитывая жизненный цикл, большой размер клеток и быстрый рост этих организмов (Cavalier-Smith, 2005; Zhu et al., 2005; Godhe et al., 2008). В инфузориях рубца, в то время как небольшие простейшие, такие как Entodinium , имели тенденцию быть недостаточно представленными, более крупные простейшие, такие как Epidinium или Polyplastron , имели тенденцию быть чрезмерно представлены методом пиросеквенирования по сравнению с микроскопическим подсчетом (Kittelmann et al ., 2015). Аналогично, Epidinium caudatum , который примерно в пять раз больше по объему, чем Entodinium caudatum (Dehority, 1993), также имеет примерно в пять раз больше копий гена 18S рРНК, закодированных в его геноме (Sylvester et al., 2009). Эти результаты согласуются с выводом о том, что методы на основе ПЦР дают более низкие оценки численности мелких Entodinium spp. (Tymensen et al., 2012a) и завышенные оценки численности, например, Polyplastron spp.(Исхак и Райт, 2014). Знание числа копий и вариаций последовательностей рДНК в отдельных эукариотах важно как для интерпретации обзоров разнообразия на основе рДНК, так и при использовании 18s рДНК для количественной оценки биомассы простейших. (Crosby, Criddle, 2003; Thornhill et al., 2007; Herrera et al., 2009; Amaral-Zettler et al., 2011). На сегодняшний день не опубликовано исследований, посвященных количеству копий на клетку и вариациям рДНК у простейших в рубце. Это особенно верно, поскольку исследования штрих-кодирования и микробного разнообразия на основе рДНК с использованием высокопроизводительного секвенирования становятся все более популярными (Amaral-Zettler et al., 2009) и молекулярные инструменты, основанные на исследованиях маркерных генов, в настоящее время широко используются для изучения разнообразия других микробов (Schlötterer, 2004; Case et al., 2007; Langille et al., 2013). Исследования на инфузориях, не относящихся к рубцу, показали, что вариация числа копий рДНК между видами инфузорий и внутри них, подчеркивая сложность использования подхода на основе порядковых номеров рДНК для вывода об относительной численности микробных эукариотических клеток в образцах окружающей среды (Wintzingerode et al., 1997 ; Мединджер и др., 2010). Таким образом, новейшие методы, основанные на генах 18S рРНК, могут быть ненадежными при оценке α-разнообразия или относительной численности различных родов и видов в данной выборке, хотя они могут надежно определять тенденции относительной численности родов и видов между разными образцами (β- разнообразие). Необходимы дополнительные исследования, сравнивающие молекулярные и традиционные методы (de la Fuente et al., 2009; Tymensen et al., 2012b; Kittelmann et al., 2015).
Функциональные гены простейших инфузорий рубца
Хотя существуют многочисленные копии рДНК в макронуклеусах инфузорий, вполне вероятно, что только небольшая часть этих генов транскрипционно активна в инфузориях при любых данных условиях роста, как ранее было показано для др. Эукариот (Reeder, 1999).Исследования на Tetrahymena thermophila , Paramecium tetraurelia и Oxytricha trifallax выявили важную роль некодирующих РНК в событиях перестройки генома (Feng and Guang, 2013). У спиротрихозных инфузорий, таких как Euplotes , Stylonychia и Oxytricha , более 95% микроядерной ДНК удаляется с образованием макронуклеуса во время полового размножения (Swart et al., 2013), что является механизмом, позволяющим клетка адаптироваться во время стресса.Более того, в макронуклеусе оставшийся геном сильно фрагментирован, и эти фрагменты сортируются и переупорядочиваются под руководством транскриптов из родительского макронуклеуса с образованием генов, кодирующих белок (Nowacki et al., 2011). Ни один из этих процессов у простейших рубца до сих пор не изучен из-за трудностей получения полноразмерных геномов инфузорий рубца.
Несмотря на неоднократные попытки, оказалось невозможным поддерживать простейшие в рубце в аксенической культуре. Таким образом, было трудно окончательно установить роль простейших инфузорий в рубце и, в частности, разрушение волокон.Ранние исследования стремились выделить и охарактеризовать ферменты целлюлозы, гемицеллюлазы и ксиланазы из промытых препаратов простейших (Howard et al., 1960; Bailey et al., 1962; Clayet et al., 1992), однако присутствие как внеклеточных, так и внутриклеточных симбиотических бактерии в препаратах простейших затрудняли уверенность в том, что изолированная активность действительно имеет протозойное происхождение (Delfosse-Debusscher et al., 1979; Thines-Sempoux et al., 1980). Мы разработали методы клонирования и экспрессии генов инфузорий в фаге λ (Eschenlauer et al., 1998), используя FISH для подтверждения протозойной идентичности экспрессируемых генов (Newbold et al., 2005) и разрабатывая биоинформатические индексы для подтверждения инфузорийного происхождения генов (McEwan et al., 2000). Используя эти методы, мы и другие специалисты смогли выделить и охарактеризовать гены ряда простейших в рубце (McEwan et al., 1999; Newbold et al., 2005; Belzecki et al., 2007; Boxma et al., 2007). Сюда входит широкий спектр фибролитических ферментов, ряд из которых, как было обнаружено, содержит несколько доменов со связывающими доменами и предполагаемые химерные конструкции, которые наблюдаются, что свидетельствует о высокоразвитой фибролитической способности инфузорий рубца (Devillard et al., 1999, 2003; Takenaka et al., 1999, 2004; Wereszka et al., 2004; Bera-Maillet et al., 2005). Это наблюдение было подтверждено недавним метагеномным скринингом простейших глюкозидаз и эукариотических метатранскриптомов, который подтвердил, что в простейших рубца присутствует широкий спектр разнообразных гликозидгидролаз (Findley et al., 2011; Qi et al., 2011).
На основе крупномасштабного конструирования и анализа филогении более 4000 библиотек экспрессируемых последовательностей из инфузорий рубца Entodinium caudatum , Eudiplodinium maggii , Metadinium medium , Diploplastron affine 1010, ec Polyplastron affine 1010, ec , Isotricha prostoma , Isotricha Кишечник и Dasytricha ruminantium .Ricard et al. (2006) пришли к выводу, что имеются обширные доказательства горизонтального переноса генов (HGT; 148 из 3563 неизбыточных генов) от бактерий и архей в геномах ресничек рубца. Среди кандидатов на ГПГ они сообщили о чрезмерном представительстве (> 75%) генов, участвующих в метаболизме, в частности, в катаболизме сложных углеводов (Ricard et al., 2006), предполагая, что ГПГ мог играть важную роль в обеспечении инфузорий рубца. адаптироваться к новым нишам в рубце и что фибролитические гены были приобретены простейшими из бактериальных источников (Findley et al., 2011).
Роль простейших в рубце
Несмотря на то, что эти простейшие составляют значительную часть биомассы рубца, их роль в ферментации рубца и их вклад в метаболизм и питание хозяина по-прежнему вызывает серьезные споры (Williams and Coleman, 1992). В последнем разделе этой статьи мы рассмотрим различные стратегии управления плотностью простейших в рубце; однако большинство этих диетических вмешательств также приводят к изменениям функции рубца, что затрудняет оценку эффекта простейших как таковых рубца.Простейшие в рубце не необходимы животному для выживания и дефаунации (удаления простейших из рубца с использованием широкого спектра химических и физических методов), а животные, свободные от простейших, использовались для изучения роли инфузорий простейших в функции рубца без пострадали от диетических вмешательств (Williams and Coleman, 1992). Эта статья не ставит своей целью провести полный обзор последствий дефауна, но объединяет наиболее важные публикации со времен превосходного и полного обзора, проведенного Уильямсом и Коулманом (1992).Таким образом, был проведен метаанализ для изучения основных эффектов дефаунации на основании 23 исследований in vivo , включающих 48 сравнений (Kreuzer et al., 1986; Vermorel and Jouany, 1989; Ushida et al., 1990; Frumholtz, 1991 ; Nagaraja et al., 1992; Hegarty et al., 1994, 2008; Faichney et al., 1999; Chandramoni et al., 2001; Machmuller et al., 2003; Eugène et al., 2004; Ozutsumi et al., 2005; Ohene-Adjei et al., 2007; Yáñez-Ruiz et al., 2007a, b; Bird et al., 2008; Morgavi et al., 2008, 2012; Belanche et al., 2011, 2012b, 2015; Mosoni et al., 2011; Чжоу и др., 2011). Большинство исследований проводилось на овцах (87%), остальные — на крупном рогатом скоте (13%). Изоляция новорожденных животных от их матерей (40%), использование моющих средств и других химикатов (35%, с использованием лаурилсульфата натрия, алканов, синперонового NP9, пероксида кальция, сульфата меди и т. Д.) И манипуляции с рубцом (25%, опорожнение и промывание рубца), были использованы для достижения дефаунации животных. Испытания, в которых влияние добавок, кроме дефауна, было значительным, были исключены из метаанализа.Точно так же не были включены исследования с использованием монофаунизированных животных или селективной дефауны. Большинство данных (75%) были получены из испытаний, в которых животных кормили на поддерживающем уровне, а остальные испытания были из производственных испытаний. Животных в основном кормили смешанными рационами (90%), состоящими из кормов с добавлением концентрата, в то время как 10% рационов состояли исключительно из кормов. Данные о ферментации рубца и выбросы метана были представлены в большинстве испытаний (69%), в то время как информация о пищеварении (31%), продуктивности животных (12%) и микробных популяциях рубца (10%) была менее обширной.Производство метана измерялось в камерах (75% в открытых камерах или респираторных калориметрах) или с помощью метода индикатора SF 6 (гексафторид серы) (25% исследований).
Множественные сравнения были включены из отдельной публикации с множественными исследованиями. Для каждого сравнения, включенного в анализ, величина эффекта рассчитывалась как натуральный логарифм отношения ответа (среднее значение в исчерпанном лечении, деленное на среднее значение в контрольном лечении) и дисперсии отношения, рассчитанной на основе сообщенного стандартного отклонения или стандарта. ошибка среднего для каждого сравнения (Viechtbauer, 2010).Все эффекты дефаунации были взвешены в соответствии с количеством наблюдений ( n ) в каждом сравнении. Мета-анализ был рассчитан с использованием модели случайных эффектов с оценкой Дерсимониан-Лэрда (Dersimonian and Laird, 1989) для оценки неоднородности (τ 2 ) в пакете Metafor R для каждой категории отдельно, как показано ниже:
, где θ i = истинная величина эффекта в i-м исследовании, μ = общая истинная величина эффекта и e i = случайное отклонение от общей величины эффекта [ u i ∼ N (0, τ 2 )] (Viechtbauer, 2010).Для оценки коэффициента отклика значения ниже единицы указывают на отрицательный результат, а значения выше единицы указывают на положительный эффект дефаунации на этот конкретный параметр.
Этот метаанализ подтвердил многие из ранее сообщенных результатов (Williams and Coleman, 1992) и помог прояснить ряд областей, в которых результаты все еще были противоречивыми. На рис. 1 показаны основные эффекты дефаунации на функцию рубца, указывающие на то, что физические и химические характеристики среды рубца были изменены дефаунацией; хотя характер наблюдаемых изменений не всегда был последовательным.Дефауна не повлияла на объем рубца и скорость оборота твердых веществ, в то время как скорость оборота жидкости имела тенденцию к снижению (–14%, P = 0,07). Удаление простейших из рубца значительно снижает усвояемость ОВ рубца (–7%, P = 0,008) и особенно NDF (–20%, P = 0,040) и усвояемость ADF (–16%, P = 0,100) , вероятно, в результате потери фибролитической активности простейших. Однако эта активность, по-видимому, различается в разных группах простейших: крупные Ophyroscolecidae , такие как Epidinium , Polyplastron и Eudiplodinium , обладают большей эндоглюканазной и ксиланазной активностью (Williams and Coleman, 1992).С другой стороны, Entodinium spp. имеют только слабую активность (Williams, Coleman, 1992). Точно так же Dasytricha обладает активностью глюкозидазы и целлобиозидазы, но незначительной фибролитической активностью (Takenaka et al., 2004). Низкая перевариваемость в рубце у дефаунированных животных частично компенсируется более выраженным пострубцовым перевариванием, что приводит к менее выраженным различиям в показателях общей перевариваемости тракта для OM (–4%, P = 0,089), CP (–3%, P = 0.034), NDF (–11%, P <0,001) и ADF (–9%, P <0,001; рис. 2). Другим компенсирующим фактором может быть сдвиг в сторону более энергетически эффективных реакций в рубце (меньшее количество выбросов метана) и меньшая метаболическая энергия, необходимая обезвожившимся животным для устранения избытка мочевины в результате более низкого расщепления бактериального белка и уровня аммиака в рубце. Несмотря на этот компенсаторный механизм, снижение перевариваемости корма, вероятно, будет основным недостатком дефауна, поскольку оно может ограничить потребление корма и эффективность использования корма на производственных уровнях потребления; Это в сочетании с отсутствием коммерчески жизнеспособного подхода (см. ниже) к дефаунации означает, что дефаунация не рекомендуется в качестве стратегии снижения выбросов метана в фермерских условиях (Христов и др., 2013).
Рис. 1. Мета-анализ, описывающий влияние дефаунации на функцию рубца. Для каждого параметра предоставляется информация о количестве исследований, наблюдениях (n) и диапазоне значений. На графике показаны средние эффекты и 95% конфиденциальные интервалы. Значения ниже единицы указывают на отрицательный эффект, тогда как значения выше единицы указывают на положительное влияние отключения на этот конкретный параметр.
Рис. 2. Мета-анализ, описывающий эффекты дефаунации животных. Для каждого параметра предоставляется информация о количестве исследований, наблюдениях (n) и диапазоне значений. На графике показаны средние эффекты и 95% конфиденциальные интервалы. Значения ниже единицы указывают на отрицательный эффект, тогда как значения выше единицы указывают на положительное влияние отключения на этот конкретный параметр.
Общая концентрация и производство продуктов ферментации также различаются у фаунированных и лишившихся животных животных. Снижение аммиака в рубце (–26%, P <0.001), вероятно, является наиболее последовательным из наблюдаемых эффектов элиминации простейших и, по-видимому, обусловлен снижением расщепления бактериального белка и разлагаемости кормового белка в отсутствие простейших в рубце (Williams and Coleman, 1992). Более того, было продемонстрировано, что, хотя хищничество бактерий простейшими в рубце зависит от размера простейших, холотрих простейшие имеют гораздо более низкую хищническую активность, чем энтодиниоморфиды (Belanche et al., 2012a) и, в конечном итоге, более низкое влияние на концентрацию аммиака в рубце (Belanche et al. ., 2015) и микробного белкового потока двенадцатиперстной кишки (Ivan et al., 2000a; Иван, 2009). Однако это негативное влияние энтодиниоморфидов на белковый поток двенадцатиперстной кишки можно переоценить из-за отсутствия надежного маркера для измерения протозойного кровотока (Broderick and Merchen, 1992). В предыдущих обзорах сообщалось о более высоких уровнях лактата у обезвоженных животных (Williams and Coleman, 1992), поскольку простейшие потребляют лактат быстрее, чем бактерии (Newbold et al., 1986). Наш метаанализ показал численное увеличение концентрации лактата у дефунтированных животных (+ 11%), но это увеличение было незначительным, возможно потому, что набор данных, использованный в этом метаанализе, в основном состоял из рационов с высоким содержанием кормов, которые менее склонны к продвижению. эти конкретные нарушения питания.
Хотя рН рубца не изменился, более низкая концентрация ЛЖК, наблюдаемая у дефаунированных животных (–5%, P = 0,013), по-видимому, подчеркивает роль простейших в синтезе ЛЖК и деградации корма. Способность простейших поглощать экзогенные жирные кислоты (Karnati et al., 2009) может отвлекать больше углерода на выработку ЛЖК, а не на синтез жирных кислот, и в конечном итоге увеличивать выработку ЛЖК. Что еще интереснее, наши данные показали, что дефаунация существенно снижает бутират (–22%, P <0.001), слегка повышенное содержание ацетата (+ 3%, P <0,001) и не влияло на молярные пропорции пропионата. В предыдущем метаанализе (Noziere et al., 2011) сообщалось, что усвояемость NDF положительно коррелирует с молярной долей ацетата ( r = 0,95), но отрицательно с пропионатом ( r = –0,94) и бутиратом ( r = –0.91), поэтому наши наблюдения могут указывать на то, что изменение молярных пропорций ЛЖК, вызванное дефаунацией, по-видимому, в основном вызвано уменьшением переваривания клетчатки.Исходя из стехиометрии, такой сдвиг в выработке ЛЖК в рубце должен привести к снижению продукции метана, поскольку менее метаболический H 2 будет доступен в качестве субстрата для метаногенеза (Demeyer et al., 1996). Однако влияние остановки на производство метана все еще не ясно; в различных обзорах, объединяющих in vivo, и in vitro, исследований (Hegarty, 1999) или просто in vivo (Morgavi et al., 2010), был сделан вывод, что удаление простейших из рубца приведет к 13 и 10 .Снижение добычи метана на 5%, что полностью согласуется с нашими результатами, основанными на более поздних исследованиях (–11%, P = 0,074). В соответствии с этими наблюдениями, в метаанализе, содержащем 28 экспериментов и 91 обработку, было сообщено о значительной линейной зависимости между выбросами метана и концентрацией простейших ( r = 0,96) с уменьшением выхода метана в среднем на 8,14 г / кг. DMI на каждую логарифмическую единицу снижения концентрации простейших (Guyader et al., 2014), предполагая, что простейшие играют каталитическую роль в метаногенезе рубца.Причины более низких выбросов метана у дефонированных животных до сих пор остаются спорными, поскольку количество простейших в рубце объясняет только одну часть (примерно 47%) изменчивости выбросов метана, что указывает на то, что метаногенез в рубце представляет собой сложный процесс, в котором участвуют несколько микробов (Morgavi и др., 2010). Возможен ряд механизмов, с помощью которых простейшие могут усиливать метаногенез, в зависимости от их способности продуцировать H 2 в своих гидрогеносомах (митохондриально-подобной органелле), их способности принимать эпи- и эндосимбиотические метаногены и защищать их от кислорода. токсичность (Fenchel and Finlay, 2006).Это взаимодействие является типичным примером межвидового переноса H 2 , который благоприятствует как метаногенам, так и простейшим (Ushida et al., 1997). Одна из гипотез состоит в том, что дефауна приводит к снижению количества метаногенов, которые являются единственными продуцентами метана в рубце, но наш мета-анализ показал, что это снижение уровней метаногенов не было значительным (–3%, P = 0,48). Альтернативная гипотеза предполагает, что дефаунация приводит к элиминации метаногенов, связанных с простейшими, что можно рассматривать как одно из наиболее активных сообществ метаногенов в рубце (Finlay et al., 1994), однако эта более поздняя гипотеза, основанная на замещении метаногеновых сообществ, различающихся по своей метаногенной активности, требует дальнейшего изучения. Несколько авторов изучали эндосимбиотическую популяцию архей у простейших (Sharp et al., 1998; Irbis, Ushida, 2004; Regensbogenova et al., 2004b; Tymensen et al., 2012a), и большинство из них согласны с тем, что Methanobrevibacter sp . является преобладающим родом, а вклад Methanomicrobium sp. и Methanomassiliicoccales в эндосимбиотическом сообществе метаногенов варьирует и может указывать на различия между группами простейших.Развитие молекулярных методов за последние десятилетия позволило коллегам дополнительно изучить эти микробные взаимодействия, обнаружив, что метаногены инфузорий эндосимбиотиков отличаются от свободноживущих метаногенов рубца (Tokura et al., 1997; Sharp et al., 1998). В недавней публикации также было продемонстрировано, что у холотриких простейших есть разные эндосимбиотические метаногены, чем у энтодиниоморфидов (Belanche et al., 2014), возможно, потому, что у любого из холотрих простейших есть более активные гидрогеносомы, чем у энтодиниоморфидов (Paul et al., 1990) и / или быстрый синтез гликогена простейшими холотрихами в присутствии избытка углеводов генерирует больше водорода (Hall, 2011; Denton et al., 2015). Эти различия могут объяснять большее влияние голотричных простейших на метаногенез рубца по сравнению с энтодиниоморфидами (Belanche et al., 2015). Это более позднее наблюдение основано на последовательной инокуляции овец, свободных от фауны, различными группами простейших, поэтому его следует интерпретировать с осторожностью из-за возможных смешанных эффектов лечения и периода.
Влияние дефаунации на микробную экосистему рубца не ограничивается только популяцией метаногена, и было продемонстрировано, что у дефаунированных животных было значительно больше популяций бактерий в рубце (+ 9%, P <0,001), чем у контрольных животных. Причина этого, по-видимому, основана на доступности экологической ниши для бактерий, когда простейшие не присутствуют в рубце, в сочетании с устранением простейших хищников (Williams and Coleman, 1992).Недавние исследования с использованием молекулярных методов показали, что дефаунация также изменяет структуру бактериального сообщества, что приводит к упрощению популяционной структуры и снижению бактериального разнообразия (Belanche et al., 2012b, 2015). Наш метаанализ выявил снижение концентрации фибролитических микробов, таких как анаэробные грибы (–92%, P <0,001), Ruminococcus albus (–34%, P <0,001) и Ruminococcus flavefaciens ( –22%, P <0.001) в отсутствие простейших. Это наблюдение предполагает, что переваривание клетчатки в рубце - сложная задача, которая требует симбиотического сотрудничества нескольких фибролитических микробов, включая простейшие рубца, для выполнения начальных стадий колонизации клетчатки и пищеварения; поэтому отсутствие простейших в рубце, по-видимому, оказывает пагубное влияние на этот фибролитический консорциум и, в конечном итоге, на переваривание клетчатки. Однако Hsu et al. (1991) сообщили об увеличении зооспор грибов в рубце у лишившихся жизни животных, возможно, в результате устранения хищничества простейших и конкуренции за питательные вещества (простейшие vs.грибки), очевидно, необходимы дополнительные исследования, чтобы полностью понять взаимодействие простейших с другими микробами рубца при различных режимах питания.
Влияние присутствия простейших в рубце на выживаемость патогенов в рубце и выделение патогенов — еще одна область, представляющая интерес. Как отмечалось выше, простейшие в рубце поглощают и переваривают широкий спектр бактерий (Williams and Coleman, 1992) и могут уменьшить выделение потенциальных патогенов от животного, хотя эффект сильно зависит от состава присутствующей популяции простейших (Stanford et al. ., 2010). Однако также было показано, что простейшие в рубце увеличивают патогенность некоторых патогенов, покидающих рубец (Rasmussen et al., 2005; Carlson et al., 2007), что позволяет предположить, что в этой области требуется дополнительная работа.
Рисунок 2 суммирует влияние простейших на продуктивность животных как косвенный эффект дефаунации на функцию рубца. Дефауна улучшила конверсию корма в результате более низкого потребления СВ (–2%, P <0,001) и большего среднесуточного привеса (+ 9%, P <0.001). Эти положительные эффекты дефауна особенно очевидны при низком качестве диеты, при которой средний дневной прирост был низким, возможно, из-за более низкой доступности ферментируемой энергии и перевариваемого в рубце белка для микробов рубца (Williams and Coleman, 1992). В предыдущем метаанализе эффект дефаунации оказался более выраженным, когда отношение N / NDF было ниже 6, а процент концентрата ниже 40% (Eugène et al., 2004). Так как животные, лишенные жизни, имеют более низкую усвояемость корма, поглощенная энергия ниже, чем у обычных животных.Таким образом, лучшая степень конверсии корма после отключения может быть главным образом связана с более высокой эффективностью использования поглощенных питательных веществ. Действительно, наш мета-анализ показал, что дефауна способствует большей эффективности использования энергии для откорма (+ 11%, P = 0,001), возможно, в результате более низкой выработки тепла (–5%, P = 0,010). Другая гипотеза, объясняющая лучшую продуктивность дефунтированных животных, основана на более эффективном использовании пищевого белка. Существуют четкие доказательства того, что простейшие холотрицы покидают рубец медленнее, чем бактерии (Abe et al., 1981). Однако количество и скорость потока простейших в нижний отдел кишечника является предметом многочисленных споров. Firkins et al. (2007), обнаружили, что поструминальный поток простейших пропорционален биомассе простейших в рубце. Хотя подсчет простейших в рубце и сычуговой жидкости показал, что количество сычужных сычуг составляло только 6–64% от подсчета жидкости в рубце (Punia et al., 1992), количество простейших в свободной жидкости рубца является ненадежным индикатором биомассы и оттока простейших, поскольку большинство простейших в рубце (63–90%) обнаруживаются либо связанными с частицами корма, либо секвестрированными в стенке рубца (Hook et al., 2012). Использование специфических маркеров простейших, таких как 2-аминоэтилфосфоновая кислота, подвергается сомнению, поскольку она присутствует в бактериях и в кормах (Ling and Buttery, 1978). Протозойный отток, измеренный с использованием общих микробных маркеров (например, 15 N и 35 S и пуриновых оснований), микробных N минус специфических бактериальных маркеров (таких как 2,6-диаминопимелиновая кислота, DAPA) давал различные оценки протозойных отток, а иногда и отрицательные значения из-за методологических ограничений используемых маркеров (Broderick and Merchen, 1992).Недавние исследования, основанные на использовании 18S рРНК в качестве нового маркера простейших, показали, что, хотя простейшие могут составлять до 60% микробной биомассы рубца, они редко превышают 20% потока микробных белков в тонкий кишечник (Sylvester et al. , 2004, 2005; Yáñez-Ruiz et al., 2006; Belanche et al., 2011, 2012b). Однако эти новые методы не лишены собственных ограничений: Sylvester et al. (2005) наблюдали, что до 48% простейшего азота на самом деле было азотом от заражающих бактерий, что могло привести к раздуванию пула простейшего азота в рубце, особенно когда животные имеют низкое потребление корма и низкую скорость прохождения твердых частиц (Dijkstra, 1994).Аналогичным образом Belanche et al. (2010b) наблюдали большую деградацию протозойной ДНК по сравнению с бактериальной ДНК во время сычужного переваривания, что могло привести к недооценке оттока простейшего азота из рубца. Несмотря на эти ограничения, эти новые данные, кажется, подтверждают, что простейшие в рубце частично секвестрируются в рубце. Однако эта секвестрация в рубце не одинакова для всех групп простейших; holotrich простейшие связываются с частицами корма после кормления из-за их сильного хемотаксиса по отношению к сахарам (Diaz et al., 2014), но быстро мигрируют в вентральную ретикулоруму, чтобы предотвратить вымывание из рубца (Karnati et al., 2007), напротив, энтодиниоморфиды также связываются с кормом из-за их умеренного хемотаксиса в отношении глюкозы и пептидов, но не проявляют такое же сродство к стенке рубца (Diaz et al., 2014) и, таким образом, вытекает из рубца с фазой твердых частиц (Hook et al., 2012).
Простейшие предшествуют бактериям в качестве своего основного источника белка (Williams and Coleman, 1992), и в результате дефаунация делает рубец более эффективным с точки зрения протеосинтеза, увеличивая дуоденальный поток микробного белка (+ 30%, P <0.001) и общий поток неаммиачного азота (+ 31%, P <0,001). Дефаунация также увеличивала эффективность синтеза микробного белка (+ 27%, P = 0,008) в результате как лучшего микробного протеосинтеза, так и более низкого усвоения ОМ. Время генерации протозойных намного выше, чем у бактерий, поэтому энергетические потребности для поддержания выше, если выразить их как соотношение белков, покидающих рубец (Williams and Coleman, 1992). В результате присутствие простейших отрицательно сказывается на общей энергетической эффективности экосистемы рубца.Кроме того, дефаунация также может изменять состав бактерий рубца (Belanche et al., 2012b) и, в конечном итоге, аминокислотный профиль поступления белка двенадцатиперстной кишки, способствуя увеличению определенных аминокислот, таких как лейцин, треонин и аргинин (Ivan, 2009 ), но не лизин, который считается основной лимитирующей аминокислотой у высокопродуктивных животных (Hristov and Jouany, 2005). После серии экспериментов in vivo , в которых ягнят, свободных от фауны, постепенно инокулировали простейшие виды (т.e., Isotricha Кишечник , Dasytricha ruminantium , Polyplastron multivesiculatum , Epidinium ecaudatum , Eudiplodinium maggi , и Ivan Entodinium caudatum, в разных порядках, b. 2009), Иван пришел к выводу, что холотрих простейшие поглощают лишь очень небольшое количество бактерий рубца и оказывают незначительное влияние на дуоденальный поток NAN и метаболизм белков. Точно так же простейшие голотрих не оказали значительного влияния на переваривание клетчатки.Это вместе с вкладом голотрихов в метаногенез рубца (Belanche et al., 2015), по-видимому, указывает на то, что присутствие голотрихов простейших в рубце не имеет большого значения для производства жвачных животных, если не используются диеты с высоким содержанием углеводов, и в этом случае присутствие голотричных простейших может быть полезным в результате их способности поглощать и накапливать зерна крахмала и растворимые углеводы (Williams and Coleman, 1992). Такое поглощение высоко ферментируемых углеводов предотвращает альтернативную бактериальную ферментацию, которая в противном случае снизила бы pH и увеличила бы возникновение молочнокислого ацидоза (Mackie et al., 1978). Напротив, целлюлолитические простейшие ( Polyplastron , Epidinium и Eudiplodinium ) могут быть полезными для животных, которых кормили волокнистой пищей. Однако эти фибролитические простейшие и Entodinium spp. обладают значительным потенциалом поглощения и разложения бактерий (Belanche et al., 2012a) и могут быть вредными с точки зрения использования белка жвачным животным-хозяином. Следовательно, их присутствие в рубце может быть бесполезным для животных, получающих диету с низким содержанием белка, поскольку их благоприятное фибролитическое действие будет уравновешено чувствительностью фибролитических бактерий и анаэробных грибов к дефициту азота в рубце (Belanche et al., 2012c). Таким образом, похоже, что дефауна уменьшила потери азота с мочой здесь (–10%, P <0,001) из-за комбинации более низкой пищевой деградации CP в рубце и бактериального расщепления нижней части рубца. Однако большая изменчивость, наблюдаемая с точки зрения баланса азота, подчеркивает тот факт, что дефаунация может по-разному влиять на общую эффективность использования азота в зависимости от диеты, потребляемой животным, в частности, уровня потребления и скорости прохождения частиц через тракт (Дейкстра , 1994).
Влияние дефаунации на метаболизм липидов в рубце менее определено. Простейшие вносят вклад в общую липолитическую активность микробов, но их роль в биогидрировании менее изучена (Williams and Coleman, 1992), хотя они действительно вносят значительный вклад в отток ненасыщенных жирных кислот, покидающих рубец (Yáñez-Ruiz et al., 2006). Используя рацион бычков, получавших рационы с различным содержанием хлорофилла, было продемонстрировано, что высокие уровни полиненасыщенных жирных кислот в клетках простейших, по-видимому, связаны с потреблением хлоропластов (Huws et al., 2009). Это поглощение хлоропластов, по-видимому, специфично для энтодиниоморфидов, поскольку не было обнаружено захваченных хлоропластов у голотриких простейших (Huws et al., 2012). Таким образом, простейшие, по-видимому, защищают ненасыщенные жирные кислоты хлоропластов от биогидрирования рубца, увеличивая дуоденальные потоки моно- и полиненасыщенных жирных кислот. Наш метаанализ согласуется с этим наблюдением, и дефаунация способствовала увеличению насыщенных жирных кислот (+ 4%, P = 0,046) и численному снижению моно- и полиненасыщенных жирных кислот (–13 и –10% соответственно), что возможно, не достигли значимости из-за эффектов, зависящих от диеты.
Результаты, представленные в нашем метаанализе, должны быть тщательно интерпретированы, поскольку большинство исследований имело методологические ограничения из-за внутренней сложности процесса дефаунации, ведущего к:
(1) Путаница между эффектом дефаунации и периодом при использовании одних и тех же экспериментальных животных во времени,
(2) Почти все исследования, оценивающие размер пула простейших, не смогли сообщить о дуоденальных потоках протозойной биомассы, а наоборот , более того, поток простейших часто недооценивается с использованием традиционных микробных маркеров (Broderick and Merchen, 1992),
(3) Исследования, оценивающие производство метана, не учитывали другие переменные эффекты, кроме численности простейших (т.е., усвояемость NDF),
(4) Животных часто кормят в количествах, намного ниже производственных уровней потребления,
(5) В некоторых исследованиях период адаптации после дефаунации / восстановления слишком короток для того, чтобы микробная экосистема могла полностью адаптироваться,
(6) Устранение простейших из рубца изменяет экологическую структуру рубца, и в конечном итоге альтернативные группы микробов могут занять экологическую нишу, ранее заполненную простейшими.
Таким образом, необходимы дополнительные исследования с использованием современных технологий для количественной оценки активности простейших (т.е., фибролитические и протеолитические), а также размер пула простейших в рубце относительно оттока простейшего азота для лучшего понимания роли простейших в метаболизме жвачных. Эти исследования должны проводиться на производственных уровнях потребления, чтобы действительно оценить эффект дефаунации на единицу произведенного продукта (молока или мяса).
Манипуляции с простейшими в рубце
Текущие опасения по поводу роли домашнего скота в глобальном предупреждении побудили исследователей искать стратегии манипулирования простейшими в рубце для уменьшения производства метана.Как уже упоминалось ранее, существует линейная зависимость между концентрацией простейших и выбросами метана (Guyader et al., 2014), и было подсчитано, что от 9 до 37% продукции метана в рубце можно отнести к метаногенам, связанным с простейшими в рубец (Finlay et al., 1994; Newbold et al., 1995; Machmuller et al., 2003).
Большинство подходов к дефаунации редко приводят к полному удалению простейших из рубца, их эффективность во многом зависит от состава диеты (Hegarty, 1999).Лечение, обычно используемое для частичной или полной дефауны рубца, включает: химические вещества, токсичные для простейших (сульфат меди, диоктилсульфосукцинат, этоксилат или алканаты спирта, пероксид кальция), ионофоры, липиды и сапонины (Williams and Coleman, 1992; Jouany, 1996; Hook et al., 2010).
Недавний метаанализ Guyader et al. (2014) показали одновременное снижение численности простейших и выбросов метана в 31% из 70 исследований, в которых использовались различные стратегии сокращения выбросов метана.В большинстве исследований липиды использовались в качестве стратегии борьбы с простейшими / снижения уровня метана. Авторы также сообщили, что антипротозойный эффект липидов зависит от состава жирных кислот, причем жирные кислоты со средней длиной цепи более эффективны, чем полиненасыщенные жирные кислоты, в контроле численности простейших. Было показано, что добавки, богатые полиненасыщенными жирными кислотами, такими как линолевая кислота (C18: 2 из сои и подсолнечника) и линоленовая кислота (C18: 3 из льняного семени), оказывают негативное влияние на выработку метана (4.Снижение на 1 и 4,8% на процентную единицу добавленных липидов, соответственно, Martin et al., 2010). Антиметаногенный эффект полиненасыщенных жирных кислот был связан с их токсическим действием на целлюлозолитические бактерии (Nagaraja et al., 1997) и простейшие (Doreau and Ferlay, 1995). Хотя первоначально предполагалось, что биогидрирование полиненасыщенных жирных кислот в рубце может представлять собой альтернативу метаногенезу H 2 (Lennarz, 1966), теперь считается, что значение биогидрирования для общего стока H 2 является small (Nagaraja et al., 1997). Было показано, что жирные кислоты со средней длиной цепи обладают сильным антипротозойным действием. В нескольких исследованиях сообщалось о снижении метаногенеза в рубце при добавлении лауриновой кислоты (C12: 0) и миристиновой кислоты (C14: 0) либо в чистых формах, либо в продуктах, богатых этими жирными кислотами (кокосовое масло), под in vitro (Dohme et al. ., 2001; Soliva et al., 2004) или in vivo в условиях (Machmuller et al., 2003; Jordan et al., 2006). Martin et al. (2010) сообщили, что жирные кислоты со средней длиной цепи, в основном содержащиеся в кокосовом масле, привели к снижению метана на 7.3% на процентную единицу добавленного липида. Однако наблюдаемый эффект подавления метана не всегда был связан со снижением концентрации простейших (Machmuller et al., 2003), что может быть связано с прямым действием жирных кислот со средней длиной цепи на метаногены (Dohme et al., 1999; Panyakaew et al. др., 2013). Недавнее исследование (Faciola and Broderick, 2014) показало, что кокосовое масло и лауриновая кислота снижают количество простейших на 40%, тогда как лауриновая кислота изменяет усвояемость клетчатки, кокосовое масло — нет.Однако сообщалось о сокращении выработки метана и сопутствующем уменьшении потребления сухого вещества, когда кокосовое масло и лауриновая кислота использовались в качестве дезорганизующих агентов (Христов и др., 2013), что потенциально ограничило бы их практическое использование в фермерских хозяйствах. Очевидно, что необходимы дополнительные исследования, чтобы выяснить, как использовать масла и жирные кислоты для борьбы с простейшими в рубце.
В литературе предполагается, что сапонины смягчают метаногенез, главным образом, за счет уменьшения количества простейших, в то время как конденсированные танины действуют как за счет уменьшения количества простейших, так и за счет прямого токсического воздействия на метаногены, тогда как эфирные масла действуют в основном путем прямого токсического действия на метаногены (Cieslak и другие., 2013). В соответствии с этой информацией, метаанализ влияния фитохимических веществ на метаногенез (Patra, 2010) показал, что изменения численности простейших следовали линейной зависимости с изменениями выработки метана сапонинами ( r = 0,69) и танинами ( r = 0,55), но эта связь была слабее ( r = 0,45) по отношению к эфирным маслам. Ингибирование метана сероорганическими соединениями не было связано с изменениями в популяции простейших (Patra, 2010), поскольку такие соединения специфически ингибируют метаногенные археи.Для растений, содержащих танин, антиметаногенная активность приписывается группе конденсированных танинов. Было высказано предположение, что дубильные вещества имеют прямое влияние на метаногены в рубце и косвенное влияние на производство водорода из-за снижения переваривания клетчатки (Tavendale et al., 2005). Что касается их воздействия на простейшие, в некоторых исследованиях не сообщалось об их эффекте, в то время как другие показали снижение количества простейших в присутствии танинов (обзор Patra and Saxena, 2009). Это несоответствие, вероятно, связано с различиями в структуре и дозе конденсированного танина.Сапонины в доступной литературе показывают более стойкое ингибирующее действие на простейшие в рубце. Гоэль и Маккар (2012) предположили, что риск нарушения функции рубца и, как следствие, снижения продуктивности животных выше с дубильными веществами, чем с сапонинами, и когда они используются для уменьшения выработки метана, эффективный диапазон концентраций дубильных веществ уже, чем сапонинов. Сапонины представляют собой гликозилированные тритерпены или стероиды, где сапонин представляет собой агликон, а гликон представляет собой углеводную единицу, состоящую из моносахарида или олигосахарида меньшего размера.Наиболее частыми источниками сапонинов, используемых в питании жвачных животных, являются Yucca schidigera , богатая стериновыми сапонинами (10%), и Quillaja saponaria , содержащая тритерпеновые сапонины. В последнее время изучаются и другие источники сапонинов, такие как сапонины чая (Hu et al., 2005; Guo et al., 2008). Антипротозойный эффект сапонинов связан с их взаимодействием со стероловым фрагментом, который присутствует в мембране простейших (Patra and Saxena, 2009). Было высказано предположение, что сапонины с одним и тем же агликоном могут по-разному влиять на простейшие в зависимости от состава и расположения сахара (Wina et al., 2006). Этот эффект кажется временным из-за разрыва гликозидной связи микробами рубца (Newbold et al., 1997). Подавление простейших в рубце сапонинами или сапонинсодержащими растениями постоянно наблюдается в исследованиях in vitro (Wina et al., 2005). Однако исследования in vivo показали, что антипротозойный эффект сапонинов имеет тенденцию исчезать через 7–14 дней приема (Wina et al., 2005; Patra and Saxena, 2009). Было высказано предположение, что когда сапонины дегликозилируются в сапогенины микробами рубца, они становятся неактивными (Newbold et al., 1997). Таким образом, можно предположить, что комбинация сапонинов с ингибиторами гликозидазы позволит избежать дегликозилирования, поддерживая интактный сапонин и, следовательно, активность в рубце. Предварительные исследования in vitro (Ramos-Morales et al., 2014) с использованием 2,5-дигидроксиметил-3,4-дигидроксипирролидина (DMDP) в качестве ингибитора гликозидазы в сочетании с экстрактом растений, богатым сапонинами, показали потенциал этого подхода. для поддержания активности сапонинов с течением времени. Другой подход, который изучается нашей исследовательской группой, — это синтез химически модифицированного сапонина без естественных гликозидных связей, поэтому ферментативное расщепление будет структурно запрещено.Влияние сапонинов на ферментацию рубца не подтвердилось. Эти расхождения, по-видимому, связаны с химической структурой и дозировкой сапонинов, составом рациона, а также адаптацией микроорганизмов к сапонинам (Wina et al., 2005; Patra and Saxena, 2009).
Австралийские исследователи продемонстрировали потенциал вакцинации против метаногенов как метода уменьшения выбросов метана (Wright et al., 2004). Хотя эта технология все еще развивается, она предоставляет множество возможностей для долгосрочного сокращения выбросов метана.Точно так же простейшие, как поставщики водорода для метаногенов или ацетогены, которые конкурируют за водород с метаногенами, могут быть возможными целями вакцины для сокращения выбросов метана. Иммунологический подход был изучен для дефаунации (Williams et al., 2008). Вакцинация овец энтодинией или смешанными протозойными антигенами уменьшала количество простейших, и антитела IgG, выработанные против простейших в рубце, оставались активными и продолжали связывать клетки-мишени в течение до 8 часов. Однако не наблюдалось никакого действия in vivo на простейшие в рубце.Было высказано предположение, что причины отсутствия эффекта могут быть связаны с недостаточным количеством специфического Ig, доставляемого в слюне, необходимостью адъюванта для оптимизации выработки антител в слюне, нацеливания на другие антигены простейших для создания более сильного иммунного ответа.
Заключение
Со времени знаменательной публикации «Rumen Protozoa» Уильямса и Коулмана (1992) в нашем понимании простейших рубца наблюдается устойчивый, но, возможно, не впечатляющий прогресс.Появление молекулярных методов привело к появлению множества публикаций, касающихся разнообразия простейших в рубце, и, хотя, как обсуждалось выше, используемые методы имеют внутри себя ограничения в их способности точно количественно определять отдельные роды простейших, они предоставили новое понимание разнообразия простейших инфузорий. у разных видов жвачных, в разных географических регионах и при разных диетических условиях. В области дефаунации наблюдается неуклонный прогресс, и хотя на данном этапе на рынке еще не поступило коммерчески доступной техники дефаунации, вполне вероятно, что экстракты растений можно использовать для борьбы с простейшими в рубце; если не устранить их полностью.Работа над последствиями уничтожения простейших в значительной степени сосредоточена на их роли в метаногенезе, что отражает текущие опасения относительно роли жвачных животных в производстве парниковых газов. Однако использование метаанализа существующих данных в сочетании с новыми исследованиями дефаунации помогло прояснить наше понимание роли простейших в рубце, как показано на рисунках 1 и 2. Однако, возможно, это развитие молекулярных методов для клонирования и характеристики простейших. гены, первоначально от одного вида, но в последнее время из метагеномных и транскриптомных источников, которые, по-видимому, предлагают наибольший, но пока еще в значительной степени не реализованный, потенциал, чтобы помочь по-настоящему выяснить роль простейших в рубце и в отсутствие прогресса в развитии аксенических Культура простейших в рубце Необходимо приложить больше усилий для характеристики активности простейших в рубце с помощью молекулярных методов.
Авторские взносы
Все авторы в равной степени внесли вклад в подготовку данной публикации.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
CN и AB выражают признательность за поддержку Исследовательскому совету биотехнологии и биологических наук Великобритании через грант номер BB / J0013 / 1.
Ссылки
Abe, M., Iriki, T., Tobe, N., and Shibui, H. (1981). Секвестрация простейших голотрихов в ретикуло-рубце крупного рогатого скота. Заявл. Environ. Microbiol. 41, 758–765.
PubMed Аннотация | Google Scholar
Амарал-Цеттлер, Л. А., МакКлимент, Э. А., Даклоу, Х. В., и Хьюз, С. М. (2009). Метод изучения разнообразия протистанов с использованием массивно-параллельного секвенирования гипервариабельных участков v9 малых субъединичных генов рибосомной РНК. PLoS ONE 4: e6372. DOI: 10.1371 / journal.pone.0006372
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Амарал-Цеттлер, Л. А., Зеттлер, Э. Р., Теру, С. М., Паласиос, К., Агилера, А., и Амилс, Р. (2011). Структура микробного сообщества на древе жизни в крайнем Рио-Тинто. ISME J. 5, 42–50. DOI: 10.1038 / ismej.2010.101
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Абесия, Л., Holtrop, G., Guada, J. A., Castrillo, C., de la Fuente, G., et al. (2011). Изучение влияния присутствия или отсутствия простейших на ферментацию рубца и вклад микробных белков в химус. J. Anim. Sci. 89, 4163–4174. DOI: 10.2527 / jas.2010-3703
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Бальселлс, Дж., Де Ла Фуэнте, Г., Яньес-Руиз, Д. Р., Фондевила, М., и Каллеха, Л. (2010a). Описание развития экосистемы рубца с помощью ПЦР-анализа у ягнят на молочном, отнятом от груди и в откорме в системе интенсивного откорма. J. Anim. Physiol. Anim. Nutr. 94, 648–658. DOI: 10.1111 / j.1439-0396.2009.00952.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче А., Эрроа И. Р., Балселлс Дж. И Каллеха Л. (2010b). Использование количественной ПЦР в реальном времени для оценки выживания in vitro определенных последовательностей генов ДНК микробов рубца в имитированных сычужных условиях. J. Anim. Physiol. Anim. Nutr. 94, 204–211. DOI: 10.1111 / j.1439-0396.2008.00901.х
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., Мурби, Дж. М., и Ньюболд, К. Дж. (2012a). Расщепление бактериального белка разными простейшими группами рубца. J. Anim. Sci. 90, 4495–4504. DOI: 10.2527 / jas.2012-5118
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., Пинлоче, Э., Ньюболд, К. Дж., И Балселлс, Дж. (2012b). Влияние диеты и отсутствия простейших на микробное сообщество рубца и на репрезентативность бактериальных фракций, используемых при определении синтеза микробного белка. J. Anim. Sci. 90, 3924–3936. DOI: 10.2527 / jas.2011-4802
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., Доро, М., Эдвардс, Дж. Э., Мурби, Дж. М., Пинлоче, Э. и Ньюболд, К. Дж. (2012c). Изменения в микробиоте рубца из-за типа углеводов и уровня белка, потребляемого молочным скотом, связаны с изменениями ферментации рубца. J. Nutr. 142, 1684–1692. DOI: 10.3945 / jn.112.159574
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче, А., де ла Фуэнте, Г., и Ньюболд, К. Дж. (2014). Изучение метаногеновых сообществ, связанных с различными популяциями простейших в рубце. FEMS Microbiol. Ecol. 90, 663–677. DOI: 10.1111 / 1574-6941.12423
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беланче А., де ла Фуэнте Г. и Ньюболд К. Дж. (2015). Влияние прогрессивной инокуляции овец, свободных от фауны, холотобоязненными простейшими и всей фауной на ферментацию рубца, микробное разнообразие и выбросы метана. FEMS Microbiol. Ecol. 362, 1–10. DOI: 10.1093 / фемсек / fiu026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Белжецкий, Г., Ньюболд, К. Дж., МакЭван, Н. Р., Макинтош, Ф. М., и Михаловски, Т. (2007). Характеристика амилолитических свойств простейших инфузорий рубца Eudiplodinium maggii . J. Anim. Feed Sci. 16, 590–606.
Google Scholar
Bera-Maillet, C., Devillard, E., Cezette, M., Jouany, J.П., Форано Э. (2005). Ксиланазы и карбоксиметилцеллюлазы простейших в рубце Polyplastron multivesiculatum , Eudiplodinium maggii и Entodinium sp. FEMS Microbiol. Lett. 244, 149–156. DOI: 10.1016 / j.femsle.2005.01.035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берд, С. Х., Хегарти, Р. С., Вудгейт, Р. (2008). Сохранение эффектов дефаунации на пищеварение и выработку метана у овец. Aust. J. Exp. Agric. 48, 152–155. DOI: 10.1071 / EA07298
CrossRef Полный текст | Google Scholar
Boxma, B., Ricard, G., van Hoek, A.H., Severing, E., Moon-van der Staay, S.Y., van der Staay, G.W., et al. (2007). Гидрогеназа [FeFe] Nyctotherus ovalis имеет химерное происхождение. BMC Evol. Биол. 7: 230. DOI: 10.1186 / 1471-2148-7-230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бродерик, Г.А., Мерчен, Н.Р. (1992). Маркеры для количественного определения синтеза микробного белка в рубце. J. Dairy Sci. 75, 2618–2632.
PubMed Аннотация | Google Scholar
Карлсон, С. А., Шарма, В. К., МакКаддин, З. П., Расмуссен, М. А., и Франклин, С. К. (2007). Участие гена Salmonella геномного острова 1 в рубце, опосредованном простейшими, усиление инвазии для множественной устойчивости к антибиотикам Salmonella enterica серовар Typhimurium . Заражение.Иммун. 75, 792–800. DOI: 10.1128 / IAI.00679-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Case, R. J., Boucher, Y., Dahllöf, I., Holmström, C., Doolittle, W.F., and Kjelleberg, S. (2007). Использование генов 16S рРНК и rpoB в качестве молекулярных маркеров для исследований микробной экологии. Заявл. Environ. Microbiol. 73, 278–288. DOI: 10.1128 / AEM.01177-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кавальер-Смит, Т.(2005). Экономика, скорость и размер имеют значение: эволюционные силы, движущие миниатюризацией и расширением ядерного генома. Ann. Бот. 95, 147–175. DOI: 10.1093 / aob / mci010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чандрамони, Джадхао, С. Б., Тивари, К. М., Хак, Н., Мурари Лал и Хан, М. Ю. (2001). Энергетический метаболизм и выработка метана у фаунированных и дефонтированных овец, которых кормили двумя рационами с одинаковым соотношением концентратов и грубых кормов (70: 30), но разного состава. Азиатско-Австралийский. J. Anim. Sci. 14, 1238–1244. DOI: 10.5713 / ajas.2001.1238
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cieslak, A.