Строение водорослей, подготовка к ЕГЭ по биологии
Водоросли относятся к низшим растениям, наиболее примитивным: у них отсутствует разделение организма на стебель, корень и листья. Спешу заметить, что термин «низшие растения» — отжившее понятие, использовавшееся в ботанике до второй половины XX века.
Современная биология не считает дифференциацию тканей определяющим различием, сейчас существенным считают фундаментальные различия в строение клеток, обмене веществ. Тем не менее, во многих устаревших пособиях этот термин используется, и я обязан предупредить вас о нем.
Наука о водорослях называется альгология (от лат. alga — морская трава, водоросль и греч. λόγος — учение).
Среди водорослей есть одноклеточные и многоклеточные, некоторые водоросли достигают в длину 100-200 метров. Способ питания водорослей автотрофный: они синтезируют органические вещества в процессе фотосинтеза.
Клетки водорослей характеризуются наличием клеточной стенки (из целлюлозы и гликопротеинов — от греч. glykys сладкий (углеводы) + греч. prōtos — первый, важнейший (белок)) Органоиды располагаются в цитоплазме (син. — внеядерной протоплазме), где также располагается(-ются) один или несколько хроматофоров. Размножение происходит бесполым, вегетативным или половым путем.
Тело водорослей представлено слоевищем (син. — талломом) — недифференцированным скоплением клеток. С помощью ризоидов (от др.-греч. ῥίζα — корень и εἶδος — вид)
водоросли прикрепляются к субстрату (камням, коралловым полипам), функцию всасывания ризоиды не выполняют. У водорослей отсутствуют настоящие ткани, механических
тканей нет, так как таллом водоросли поддерживается (парит) в толще воды.
Хроматофор (от греч. chroma — цвет и phoros — несущий) — органелла в клетке водоросли, аналогичная хлоропласту и осуществляющая фотосинтез. Отличается от хлоропласта упрощенным строением, меньшим размером и иным составом хлорофилла. Внешне отличаются между собой по форме, хроматофор может быть: чашевидный, спиралевидный, в виде незамкнутых колец, цилиндрические, лентовидные, дисковидные. В хроматофорах находятся пигменты, которые придают окраску растению.
Система вакуолей в клетках водорослей развита отлично, в подвижных клетках водорослей можно обнаружить пульсирующие (сократительные) вакуоли. Их основная функция — поддержание
постоянного осмотического давления внутри клетки. Вообразите: в глубине океана находится клетка водоросли, в которую постоянно поступает много воды.
Также у многих подвижных водорослей в клетках присутствует светочувствительный глазок (стигма), что обуславливает их чувствительность к свету — фототаксис. Подвижные водоросли стремятся занять как можно более освещенное место, чтобы активно шел процесс фотосинтеза.
Жизненный цикл водорослей
Жизненные циклы водорослей разнообразны, обусловлены рядом экологических факторов. Мы разберем жизненный цикл на примере зеленой водоросли ульвы (морского салата).
Для начала отметим, что в целом жизненный цикл водорослей представляет собой чередование двух фаз: гаплоидной (гаметофита) и диплоидной (спорофита). Гаплоидной фазой называется
фаза, при которой клеточные ядра содержат непарный (половинный) набор хромосом. К гаплоидной фазе всегда принадлежат гаметы: сперматозоиды, спермии (отличающиеся от сперматозоидов отсутствием
жгутика), яйцеклетки.
При слиянии двух гамет: яйцеклетки (n) и спермия (n) образуется зигота (2n) из которой развивается спорофит (2n), таким образом, в спорофите восстанавливается диплоидный набор хромосом. В зооспорангии на спорофите в результате мейоза образуются зооспоры (n), которые делятся митозом, порастают и образуют мужские и женские гаметофиты (n). Клетки гаметофитов делятся митозом, образуются гаметы (n), которые сливаются в зиготу (2n), цикл замыкается.
Типы половых процессов
У водорослей выделяют несколько типов полового процесса:
- Изогамия — копулирующие элементы (гаметы) не отличаются друг от друга, подвижны
- Анизогамия — от греч. anisos неравный и gamos брак (гетерогамия) — при таком типе копулирующие элементы различаются по размерам, форме, величине, поведению
- Оогамия — от др. греч. ᾠόν яйцо и γάμος брак — копулирующие элементы резко отличаются друг от друга: крупная женская гамета без жгутиков обычно с мужской мелкой подвижной гаметой.
 Допустимо считать оогамию в некотором смысле подтипом анизогамии.
Допустимо считать оогамию в некотором смысле подтипом анизогамии.
Особо стоит выделить тип полового процесса — конъюгацию. Конъюгация отличается тем, что сливаются не гаметы, а обычные вегетативные клетки, лишенные жгутиков. Клетки соединяются друг с другом с помощью боковых выростов, формируется копуляционный (конъюгационный) канал, по которому содержимое из одной клетки перетекает в другую — образуется зигоспора. В дальнейшем из зигоспоры развивается новая водоросль.
Отметим, что зооспора представляет собой подвижную клетку, которая способна двигаться в воде с помощью жгутиков. Образуется она в зооспорангии. Зооспора участвует в бесполом размножении
у многих водорослей и простейших грибов. У некоторых водорослей имеются апланоспоры (гр. aplanes неподвижный + spora семя) — неподвижные безжгутиковые споры. Зооспоры и апланоспоры
выходят в окружающую среду, разрывая стенки спорангия, в котором они находятся.
Значение водорослей
В Мировом океане водоросли составляют основную часть биомассы. Именно они являются главными продуцентами (производителями) органического вещества, преобразуя в ходе фотосинтеза энергию солнечного света в энергию химических связей. Значение водорослей для человека трудно переоценить: содержащиеся в них вещества необходимы для нормального роста и развития животных и человека (к примеру, морская капуста (ламинария) отличается большим содержанием йода.)
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Порядок вольвоксовые (Volvocales) — это… Что такое Порядок вольвоксовые (Volvocales)?
Порядок вольвоксовых объединяет наиболее высокоорганизованных представителей класса. Сюда относятся исключительно ценобиальные и колониальные формы. Но и здесь отдельные клетки, слагающие ценобии и колонии, построены по типу хламидомонады. Однако, в отличие от одноклеточных форм, при размножении клетки здесь не расходятся, а срастаются своими оболочками или остаются соединенными общей слизью. Форма общей слизистой оболочки, количество клеток и способ их размещения в ней, а также способ срастания клеток в комплексы являются постоянными признаками и широко используются в систематике этого порядка. Обычно порядок вольвоксовых подразделяют на три семейства. Один из наиболее типичных представителей семейства спондиломоровых (Spondylonioraceae)— пироботрис стройный (Pyrobotrys gracilis, рис. 207, 1).
Он характеризуется способом срастания клеток, расположением их ярусами и отсутствием общей слизистой оболочки вокруг гроздевидного ценобия.
 Они попарно копулируют и дают зиготу, при помощи которой вид переживает неблагоприятные условия до следующего года.
Они попарно копулируют и дают зиготу, при помощи которой вид переживает неблагоприятные условия до следующего года.В чистых водах рек и озер можно встретить четырехжгутиковую форму, относящуюся к этому семейству, например спондиломорум четверной (Spondylomorum quaternarium, рис. 207, 2), сходный с предыдущим видом. Весьма своеобразный по строению вид — хлоркорона богемская (Chlorcorona bohemica, рис. 207, 3), в ценобиях которой клетки соединены слизистыми тяжами в два 4-клеточных ромбических яруса.
Второе семейство — 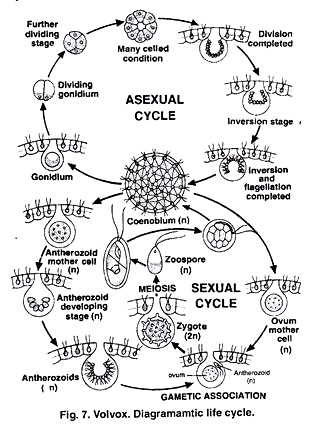 В пробе воды из такого растаявшего снега при малом увеличении мы увидим шаровидные или эллипсоидные ценобии, внутри которых обычно расположено 8 клеток более или менее веретеновидпой формы. Клетки покрыты лишь тонким и нежным перипластом, позволяющим им вырабатывать разветвленные протоплазматические выросты, которыми они упираются в стенку цепобия. Внутри ценобия клетки расположены поясом по экватору, а их жгутики выходят наружу. В хлоропласте каждой клетки содержится по 2—5 пиреноидов и округло-треугольный глазок, расположенный на наружной его стороне. Пульсирующих вакуолей несколько, и они разбросаны по всему протопласту. В клетках некоторых ценобиев можно увидеть дробление протопласта на 8, реже 4 или 16 участков. Это начало образования зооспор, которые, сформировавшись, не выходят из оболочки материнской клетки, а размещаются в ней по ее экватору и слагают новый ценобий. Кроме зооспор, в клетках ценобия может образоваться от 4 до 32 веретеновидных изогамет, которые, попарно копулируя внутри ценобия, дают одну зиготу.
В пробе воды из такого растаявшего снега при малом увеличении мы увидим шаровидные или эллипсоидные ценобии, внутри которых обычно расположено 8 клеток более или менее веретеновидпой формы. Клетки покрыты лишь тонким и нежным перипластом, позволяющим им вырабатывать разветвленные протоплазматические выросты, которыми они упираются в стенку цепобия. Внутри ценобия клетки расположены поясом по экватору, а их жгутики выходят наружу. В хлоропласте каждой клетки содержится по 2—5 пиреноидов и округло-треугольный глазок, расположенный на наружной его стороне. Пульсирующих вакуолей несколько, и они разбросаны по всему протопласту. В клетках некоторых ценобиев можно увидеть дробление протопласта на 8, реже 4 или 16 участков. Это начало образования зооспор, которые, сформировавшись, не выходят из оболочки материнской клетки, а размещаются в ней по ее экватору и слагают новый ценобий. Кроме зооспор, в клетках ценобия может образоваться от 4 до 32 веретеновидных изогамет, которые, попарно копулируя внутри ценобия, дают одну зиготу.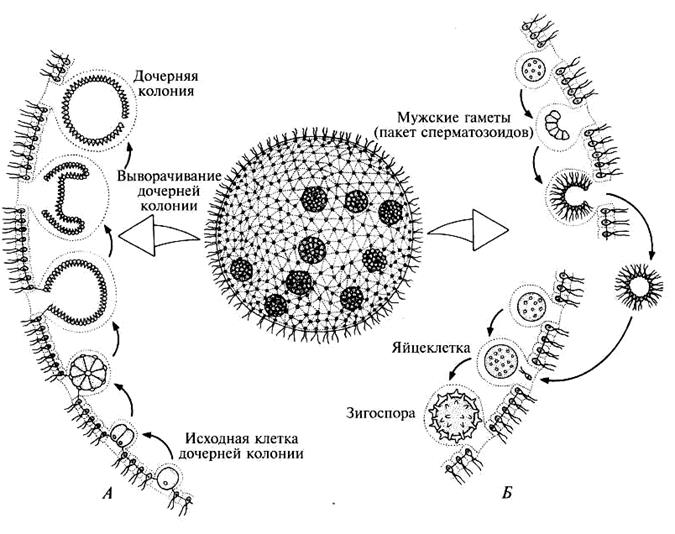 Последняя накапливает гематохром, приобретает красную окраску и переходит в состояние покоя. Через некоторое время она дает новый ценобий.
Последняя накапливает гематохром, приобретает красную окраску и переходит в состояние покоя. Через некоторое время она дает новый ценобий.
Последнее семейство — собственно вольвоксовые (Volvocaceae)—представлено ценобиальными и колониальными формами. Из ценобиалышх форм наиболее обычны представители родов гониум (Gonium) и эвдорина (Eudorina). У первого из них клетки ценобия расположены в один слой, пакетиком, окруженным общей слизистой обверткой.
Яркое впечатление производит под микроскопом гониум пекторальный (Gonium pectorale, рис. 208, 1) во время движения, когда его 16-клеточный ценобий парит в толще воды, напоминая сказочный ковер-самолет: клетки ценобия, дружно двигая жгутиками, свисающими вниз бахромой, несут над собой общую слизистую обвертку.
У широко распространенных видов эвдорины ценобий обычно эллипсоидные, состоят из 32 клеток, расположенных не в одной плоскости, а по периферии в поверхностном слое общей слизи в 5—8 рядов. Благодаря согласованному движению жгутиков всех 32 клеток ценобий довольно быстро передвигается в толще воды. Обычно в ценобии эвдорины все клетки одинаковые (Eudorina elegans, рис. 208, 2) и все способны, как и у гониума, к образованию бесполым путем новых дочерних ценобиев.
Благодаря согласованному движению жгутиков всех 32 клеток ценобий довольно быстро передвигается в толще воды. Обычно в ценобии эвдорины все клетки одинаковые (Eudorina elegans, рис. 208, 2) и все способны, как и у гониума, к образованию бесполым путем новых дочерних ценобиев.
У пандорины (Pandorina morum, рис. 208, 3) в ценобиях обычно 16 клеток, и расположены они не рыхло, как у эвдорины, а компактно, соприкасаясь боковыми сторонами, в связи с чем обычно приобретают многогранную форму.
Указанные ценобиальные формы вольвоксовых встречаются довольно часто, обычно в небольших стоячих водоемах, где летом нередко вызывают зеленое «цветение» воды. Размножаются они в основном бесполым способом, путем образования в каждой клетке молодых ценобиев. К концу вегетации у них осуществляется и половой процесс — в виде изогамии у гониумов, гетерогамии у пандорин и оогамии у эвдорин. Во всех случаях образовавшаяся зигота переходит в состояпие покоя.
Колониальные формы вольвоксовых представлены лишь видами рода вольвокс (Volvox). Это наиболее высокоорганизованные представители класса. Их колонии (рис. 209) имеют вид слизистых, диаметром до 2 мм шаров, в периферическом слое которых расположено множество, до 20, а у некоторых и до 50 тыс. хламидомонадо-подобных клеток, сросшихся своими боковыми ослизненными стенками друг с другом и соединенных одна с другой протоплазменными тяжами (плазмодесмами). Здесь уже наблюдается дифференциация или специализация клеток в пределах колонии. Громадное большинство составляют вегетативные клетки, слагающие периферическую часть колонии и не принимающие участие в размножении. Между вегетативными клетками разбросаны более крупные репродуктивные клетки. Около десятка из них — клетки бесполого размножения, так называемые партеногонидии, которые в результате многократных делений дают начало молодым, дочерним колониям внутри материнской. Половое размножение представлено типичной оогамией. Оогонии и антеридии возникают также из репродуктивных клеток. В грушевидных, с темно-зеленым содержимым оогониях возникает одна яйцеклетка.
Это наиболее высокоорганизованные представители класса. Их колонии (рис. 209) имеют вид слизистых, диаметром до 2 мм шаров, в периферическом слое которых расположено множество, до 20, а у некоторых и до 50 тыс. хламидомонадо-подобных клеток, сросшихся своими боковыми ослизненными стенками друг с другом и соединенных одна с другой протоплазменными тяжами (плазмодесмами). Здесь уже наблюдается дифференциация или специализация клеток в пределах колонии. Громадное большинство составляют вегетативные клетки, слагающие периферическую часть колонии и не принимающие участие в размножении. Между вегетативными клетками разбросаны более крупные репродуктивные клетки. Около десятка из них — клетки бесполого размножения, так называемые партеногонидии, которые в результате многократных делений дают начало молодым, дочерним колониям внутри материнской. Половое размножение представлено типичной оогамией. Оогонии и антеридии возникают также из репродуктивных клеток. В грушевидных, с темно-зеленым содержимым оогониях возникает одна яйцеклетка. В антеридиях образуется по 64 желтоватой окраски антерозоида, сложенных в виде пакетика. После оплодотворения яйцеклетка превращается в ооспору с многослойной, шиповатой снаружи оболочкой и богатым запасными веществами внутренним содержимым. После периода покоя ооспора дает начало новой колонии вольвокса.
В антеридиях образуется по 64 желтоватой окраски антерозоида, сложенных в виде пакетика. После оплодотворения яйцеклетка превращается в ооспору с многослойной, шиповатой снаружи оболочкой и богатым запасными веществами внутренним содержимым. После периода покоя ооспора дает начало новой колонии вольвокса.
Наиболее широко распространенными являются два вида этого рода — вольвокс шаровидный (Volvox globator) с однодомными колониями и вольвокс золотистый (Volvox aureus), колонии которого двудомны. В центральных районах европейской части СССР оба эти вида часто встречаются в старицах рек и прудах, нередко вызывая даже «цветение» воды (рис. 209).
Из вышесказанного видно, что класс вольвоксовых (Volvocophyceae)—это сравнительно однородная группа отдела зеленых водорослей (Chlorophyta). Они характеризуются только одной монадной структурой тела. Таллом их преимущественно одноклеточный, у некоторых типично ценобиальный, у немногих колониальный. Строение клетки в общем однородное, почти у всех видов хламидомонадоподобное.
Строение клетки в общем однородное, почти у всех видов хламидомонадоподобное.
Вольвоксовые не являются тупиковой ветвью в эволюции зеленых водорослей. Потеряв подвижность, представители вольвоксовых на каком-то этапе исторического развития передали эстафету протококковым водорослям.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
Многоклеточные зеленые водоросли. Спирогира
Многоклеточные водоросли, в отличие от одноклеточных, мы можем разглядеть и потрогать. Иногда они нас радуют своими декоративными свойствами, например, в аквариумах, иногда раздражают — скажем, когда при сходе с берега в воду мы оскальзываемся на камнях, покрытых зеленым одеялом, или когда не можем привольно поплавать в море потому, что прибоем нанесло много противных, дурно пахнущих водорослей. Внешний данные водорослей очень различаются — от едва заметных сфер и тонких нитей до больших пластин. Для зеленых водорослей характерны богатые оттенки зеленого цвета, — эту особенность определяет преобладание пигмента хлорофилла. Ниже мы рассмотрим особенности нескольких видов многоклеточных зеленых водорослей: спирогиры, кладофоры, улотрикса, ульвы и вольвокса.
Для зеленых водорослей характерны богатые оттенки зеленого цвета, — эту особенность определяет преобладание пигмента хлорофилла. Ниже мы рассмотрим особенности нескольких видов многоклеточных зеленых водорослей: спирогиры, кладофоры, улотрикса, ульвы и вольвокса.
Спирогира. Строение и размножение
Спирогира — нитчатая водоросль, принадлежащая к семейству зигнемовых и классу конъюгатов. Обитает в прудах, в стоячей воде, где ее густая масса образует тину.
Строение
1. Тело спирогиры представлено ниточкой, в которой в один ряд выстроились цилиндрические клетки. Снаружи каждая нить покрыта слизистым чехлом.
2. Хроматофор спиральный, имеет вид закрученной ленты.
3. Крупное ядро с ядрышком.
4. Большая вакуоль.
Размножение
Спирогира способна размножаться как бесполым способом, так и половым. При бесполом размножении идет процесс фрагментации, нить водоросли рвется на отдельные участки, из которых формируются новые новая спирогиры.
При бесполом размножении идет процесс фрагментации, нить водоросли рвется на отдельные участки, из которых формируются новые новая спирогиры.
Половое размножение спирогиры идет путем конъюгации.
1. Между клетками разных спирогир при их сближении образуются выросты, как будто перекидываются мостики.
2. Живое содержимое по этим «трубочкам» перетекает из одной клетки в другую, сливаясь.
3. Образуется зигота.
4. Зигота делится мейозом, давая в результате четыре клетки. Три из них отмирают, а одна образует новую спирогиру.
5. Гаметы не образуются, так как у конъюгатов нет подвижных стадий размножения: зооспор и гамет.
Volvox — Volvox — qaz.wiki
Volvox является полифилетической род из chlorophyte зеленых водорослей в семье вольвоксовой . Он образует сферические колонии до 50 000 клеток. Они живут в различных пресноводных средах обитания, и впервые о них сообщил Антони ван Левенгук в 1700 году. Вольвоксы произошли от одноклеточных предков примерно 200 миллионов лет назад .
Вольвоксы произошли от одноклеточных предков примерно 200 миллионов лет назад .
Описание
Колония Volvox : 1) Хламидомоноподобная клетка, 2) Дочерняя колония, 3) Цитоплазматические мостики, 4) Межклеточный гель, 5) Репродуктивная клетка, 6) Соматическая клетка.Вольвокс — полифилетический род в кладе вольвоциновых зеленых водорослей. Каждая зрелая колония вольвокса состоит из тысяч клеток двух дифференцированных типов клеток: многочисленных жгутиковых соматических клеток и меньшего числа половых клеток, лишенных сомы, которые встроены в поверхность полой сферы или ценобия, содержащего внеклеточный матрикс, состоящий из гликопротеины .
Взрослые соматические клетки состоят из одного слоя с обращенными наружу жгутиками. Клетки плавают скоординированно, с четкими передним и задним полюсами. У клеток есть передние глазные пятна, которые позволяют колонии плыть навстречу свету. Клетки колоний в более базальной кладе Euvolvox связаны между собой тонкими нитями цитоплазмы , называемыми протоплазматами.![]() Номер ячейки указывается при разработке и зависит от количества раундов деления.
Номер ячейки указывается при разработке и зависит от количества раундов деления.
Размножение
Вольвокс является факультативно половым и может размножаться как половым, так и бесполым путем. В лаборатории чаще всего наблюдается бесполое размножение; относительная частота полового и бесполого размножения в дикой природе неизвестна. Переход от бесполого размножения к половому может быть вызван условиями окружающей среды и производством феромона, вызывающего пол. Устойчивые к высыханию диплоидные зиготы получаются после успешного оплодотворения.
Бесполая колония включает как соматическую (вегетативную) клетку, которые не размножаются, и большой, не подвижен gonidia в интерьере, которые производят новые колонии через повторное деление. При половом размножении образуются два типа гамет . Виды Volvox могут быть однодомными и раздельнополыми . Мужские колонии выпускают многочисленные пакеты спермы, в то время как в женских колониях отдельные клетки увеличиваются и становятся оогаметами, или яйцеклетками.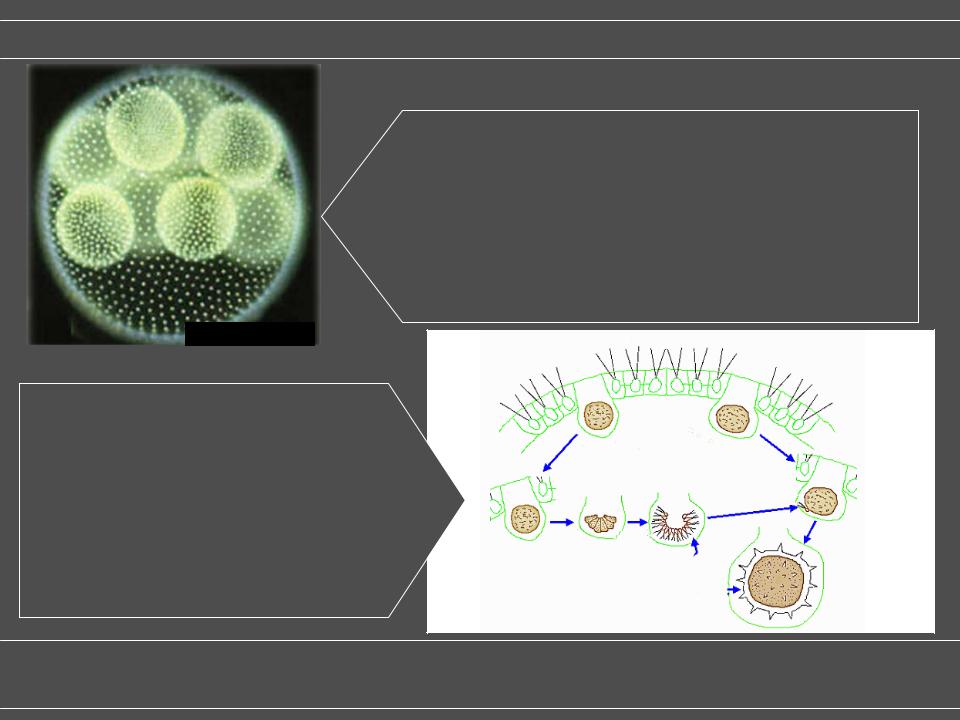
Кирк и Кирк показали, что производство феромонов, вызывающих пол, может быть вызвано в соматических клетках коротким тепловым шоком, которым подвергаются бесполые растущие организмы. Возбуждение секса при тепловом шоке опосредовано окислительным стрессом, который, вероятно, также вызывает окислительное повреждение ДНК. Было высказано предположение, что переключение на сексуальный путь является ключом к выживанию в условиях стресса окружающей среды, включая жару и засуху . В соответствии с этой идеей, индукция секса включает в себя путь передачи сигнала, который также индуцируется у Volvox при ранении.
Инверсия колонии
Инверсия колоний — это особая характеристика во время развития в отряде Volvocaceae, которая приводит к появлению новых колоний, жгутики которых обращены наружу. Во время этого процесса бесполые репродуктивные клетки (гонидии) сначала подвергаются последовательным клеточным делениям с образованием эмбриона вогнутой формы или чашеобразной формы, состоящего из одного слоя клеток. Сразу после этого клеточный слой оказывается наизнанку по сравнению с конфигурацией взрослого человека — апикальные концы протопластов зародыша, из которых образуются жгутики, ориентированы внутрь плакеи. Затем эмбрион претерпевает инверсию, во время которой клеточный слой переворачивается с образованием сфероидальной дочерней колонии с апикальными концами и жгутиками дочерних протопластов, расположенными снаружи. Этот процесс обеспечивает соответствующее передвижение сфероидальных колоний Volvocaceae. Механизм инверсии широко исследовался на клеточном и молекулярном уровнях с использованием модельного вида Volvox carteri .
Сразу после этого клеточный слой оказывается наизнанку по сравнению с конфигурацией взрослого человека — апикальные концы протопластов зародыша, из которых образуются жгутики, ориентированы внутрь плакеи. Затем эмбрион претерпевает инверсию, во время которой клеточный слой переворачивается с образованием сфероидальной дочерней колонии с апикальными концами и жгутиками дочерних протопластов, расположенными снаружи. Этот процесс обеспечивает соответствующее передвижение сфероидальных колоний Volvocaceae. Механизм инверсии широко исследовался на клеточном и молекулярном уровнях с использованием модельного вида Volvox carteri .
 (e) Схематическое изображение клеток в области, отмеченной (d). ПК: лопастные ячейки, два разных вида иллюстрируют анизотропную форму; SC: веретеновидные клетки; красная линия: положение цитоплазматических мостиков (ЦБ). (е) 3D-изображения одного эмбриона V. globator на трех последовательных стадиях инверсии. (g) Оптические среднесагиттальные сечения эмбриона на рисунке (f). (h) Прослеженные контуры клеточного листа, наложенные на секции в (g), с кривизной, обозначенной цветом κ. (i) Поверхности вращения, вычисленные по усредненным контурам.
(e) Схематическое изображение клеток в области, отмеченной (d). ПК: лопастные ячейки, два разных вида иллюстрируют анизотропную форму; SC: веретеновидные клетки; красная линия: положение цитоплазматических мостиков (ЦБ). (е) 3D-изображения одного эмбриона V. globator на трех последовательных стадиях инверсии. (g) Оптические среднесагиттальные сечения эмбриона на рисунке (f). (h) Прослеженные контуры клеточного листа, наложенные на секции в (g), с кривизной, обозначенной цветом κ. (i) Поверхности вращения, вычисленные по усредненным контурам.Среды обитания
Вольвокс — это род пресноводных водорослей, обитающих в прудах и канавах, даже в неглубоких лужах. По словам Чарльза Джозефа Чемберлена ,
«Лучше всего искать его в более глубоких прудах, лагунах и канавах, которые получают много дождевой воды. Говорят, что там, где вы найдете Лемну , вы, скорее всего, найдете Вольвокса ; и это правда, что такие вода благоприятна, но затенение неблагоприятное.
Посмотрите, где вы найдете Sphagnum , Vaucheria , Alisma , Equisetum fluviatile , Utricularia , Typha и Chara . Доктор Ньювланд сообщает, что Pandorina , Eudorina и Gonium обычно входят в состав зеленой накипи на валяется на полях, где содержатся свиньи. Жгутиконосица эвглена часто ассоциируется с этими формами ».
История
Антони ван Левенгук впервые сообщил о наблюдениях Вольвокса в 1700 году.
После нескольких рисунков Генри Бейкера (1753 г.) Линней (1758 г.) описал род Volvox с двумя видами: V. globator и V. chaos . Volvox chaos — это амеба, известная теперь как Chaos (genus) sp.
Эволюция
Предки Volvox перешли от одиночных клеток к многоклеточным колониям по крайней мере 200 миллионов лет назад , в триасовый период . Оценка с использованием последовательностей ДНК примерно 45 различных видов вольвоциновых зеленых водорослей, включая Volvox , предполагает, что переход от одиночных клеток к недифференцированным многоклеточным колониям занял около 35 миллионов лет.
Ссылки
внешние ссылки
| Викискладе есть медиафайлы по теме Volvox . |
Wikizero — Вольвокс
Из Википедии, бесплатной энциклопедии
| Вольвокс | |
|---|---|
| Колония Volvox | |
промежуточные ранги | |
| Volvox L., 1758 | |
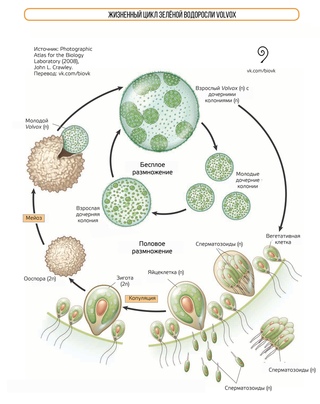
Во́львокс (лат. Volvox) — род подвижных колониальных организмов, относящийся к отделу зелёных водорослей. Обитают в стоячих пресных водоёмах. При массовом размножении вызывают цветение воды, окрашивая её в зелёный цвет.
Антони ван Левенгук был первым, кто в 1700 году обратил внимание и описал вольвокс. Вероятнее всего это была первая идентификация в истории микроводорослей. Через свой микроскоп он наблюдал зрелищную зелёную сферу, которая неторопливо вращаясь, перемещалась вперёд. За такое поведение он и получил своё латинское название — подвижные шаровые водоросли (Volvox)[1].
Вероятнее всего это была первая идентификация в истории микроводорослей. Через свой микроскоп он наблюдал зрелищную зелёную сферу, которая неторопливо вращаясь, перемещалась вперёд. За такое поведение он и получил своё латинское название — подвижные шаровые водоросли (Volvox)[1].
Размер одной колонии — до 3 мм. Колония шарообразная, включает от 200 до 10 тысяч клеток[2]. Клетки соединены протоплазматическими нитями, в центре колонии имеется полость, содержащая жидкую слизь. Клетки внешнего слоя схожи по строению с хламидомонадой. Они имеют по два жгутика, обращённых наружу.
Размножение[править | править код]
В пределах колонии вольвокса наблюдается специализация клеток. Большинство клеток — вегетативные. Между ними разбросаны генеративные клетки, принимающие участие в процессе размножения. Половой процесс — оогамия. Генеративные клетки крупные и не имеют жгутиков. В оогониях развиваются яйцеклетки, имеющие зелёную окраску из-за запасных веществ. В антеридиях развиваются сперматозоиды. После проникновения сперматозоида в оогоний происходит слияние гамет и образование зиготы (2n), которая также имеет название ооспоры. Ооспора прорастает обычно весной. Происходит сперва мейоз, а затем множество митотических делений. Новый вольвокс вновь гаплоиден. В жизненном цикле лишь зигота имеет двойной набор хромосом.
В антеридиях развиваются сперматозоиды. После проникновения сперматозоида в оогоний происходит слияние гамет и образование зиготы (2n), которая также имеет название ооспоры. Ооспора прорастает обычно весной. Происходит сперва мейоз, а затем множество митотических делений. Новый вольвокс вновь гаплоиден. В жизненном цикле лишь зигота имеет двойной набор хромосом.
Бесполое размножение осуществляется посредством особых клеток — партеногонидий. Происходит перпендикулярное поверхности шара митотическое деление этих клеток. В результате формируется пластинка, которая выворачивается, смыкается краями и образует дочерний шар. Дочерние клетки высвобождаются, разрастаясь и разрывая материнский организм[3].
Объект интересен как яркий пример колониального организма среди водорослей. Исследование особенностей строения вольвокса даёт возможность сделать предположение, что развитие организмов от одноклеточных к многоклеточным происходило через колониальные формы.
Согласно базе данных AlgaeBase род охватывает следующие виды[4]:
- Vovox africanus G.
 S.West, 1910
S.West, 1910 - Volvox amboensis M.F. Rich et Pocock, 1933
- Vovox aureus Ehrenb., 1832 — Вольвокс золотистый
- Volvox barberi W.R. Shaw, 1922
- Volvox capensis M.F. Rich et Pocock, 1933
- Volvox carteri F. Stein, 1873
- Volvox chaos L., 1758
- Volvox dissipatrix (W.R. Shaw) Printz, 1927
- Volvox fertilis Nayal
- Volvox gigas Pocock, 1933
- Volvox globator L., 1758 — Вольвокс шаровидный
- Volvox merrillii W.R. Shaw
- Volvox migulae (W.R. Shaw) Printz, 1927
- Volvox obversus (W.R. Shaw) Printz, 1927
- Volvox ovolis Pocock ex Nozaki et A.W.Coleman, 2011
- Volvox pocockiae R.C. Starr, 1970
- Volvox polychlamys Korshikov, 1938 — Вольвокс многооболочковый
- Vovox rousseletii G.S.West, 1910
- Volvox perglobator J.H.Powers
- Volvox pilula O.F.Müll.
- Volvox powersii (W.R. Shaw) Printz, 1927
- Volvox spermatosphaera Power, 1908
- Volvox tertius Art.
 Meyer, 1896 — Вольвокс третий
Meyer, 1896 — Вольвокс третий
Читать книгу Альгология и микология. Практикум Н. А. Лемезы : онлайн чтение
1.13. Отдел Зеленые водоросли (chlorophyta)
Отдел включает одноклеточные, колониальные, ценобиальные и многоклеточные организмы зеленого цвета разной морфологической структуры таллома, кроме амебоидной, размером от 1 мкм до нескольких десятков сантиметров. Хроматофоры содержат хлорофилл а и Ь, а– и β– каротины, ксантофиллы: неоксантин, виолаксантин, зеаксантин, антераксантин и др. Клетки у большинства видов одеты целлюлозной, целлюлозно-пектиновой или пектиновой оболочкой, одно– или многоядерные. Хроматофоры могут быть различной формы, как правило, с пиреноидами. Запасное вещество – крахмал, редко масло. У подвижных зеленых водорослей есть глазок (стигма), 2 – 4 и более жгутиков, обычно одинаковой длины.
Размножение осуществляется путем деления клетки, фрагментации нити или колонии на части, а также при помощи зоо– или апланоспор. Половой процесс – изо-, гетеро-, оогамия, хологамия и конъюгация. Для некоторых водорослей характерно чередование поколений – споро– и гаметофита. Они обитают в пресных водоемах, некоторые в морях и влажных местах.
Для некоторых водорослей характерно чередование поколений – споро– и гаметофита. Они обитают в пресных водоемах, некоторые в морях и влажных местах.
Классификация зеленых водорослей окончательно не разработана, однако чаще всего их группируют в 5 классов (вольвокофициевые, протококкофициевые, улотрихофициевые, сифонофициевые и конъюгатофициевые) по основным ступеням морфологической дифференцировки таллома, начиная от примитивных подвижных одноклеточных организмов и кончая более совершенными прикрепленными многоклеточными формами.
Класс Вольвокофициевые (Volvocophyceae), или Равножгутиковые (Isocontae)К классу Вольвокофициевые относятся наиболее примитивные одноклеточные, ценобиальные и колониальные представители отдела Зеленые водоросли, имеющие монадную структуру тела. У немногих форм клетки голые, у остальных покрыты оболочкой, снабжены двумя или четырьмя жгутиками равной длины (отсюда название равножгутиковые). Размножаются бесполым и половым путем. Половой процесс может быть изо-, гетеро– и оогамным.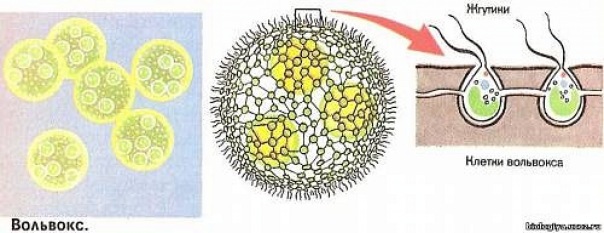 Водоросли способны переходить в пальмеллевидное состояние. Рост ценобия происходит за счет увеличения размеров клеток, а колонии – за счет их деления.
Водоросли способны переходить в пальмеллевидное состояние. Рост ценобия происходит за счет увеличения размеров клеток, а колонии – за счет их деления.
Среди этой группы водорослей встречаются фото-, гетеро– и миксотрофы. В цикле их развития доминирует гаплоидная фаза, диплоидны лишь зиготы и покоящиеся споры. К ним относят свыше 250 видов, встречающихся в мелких пресноводных водоемах. Деление класса на порядки во многом определяется особенностями морфологической дифференцировки таллома.
Порядок Хламидомонадальные (Chlamydomonadales). Сюда относятся одноклеточные водоросли с плотной оболочкой, с двумя или четырьмя равными жгутиками. У ряда представителей оболочки инкрустированы солями кальция, кремнезема или железа. Они могут быть цельными и двустворчатыми, с различными выростами и без них. В зависимости от строения оболочки выделяют 3 семейства, основное из них – хламидомонадовые.
Род хламидомонада ( Сhlamydomonas) включает многочисленные виды, обитающие в лужах, канавах, мелких водоемах, почве, на наземных субстратах (как аэрофиты), на снегу.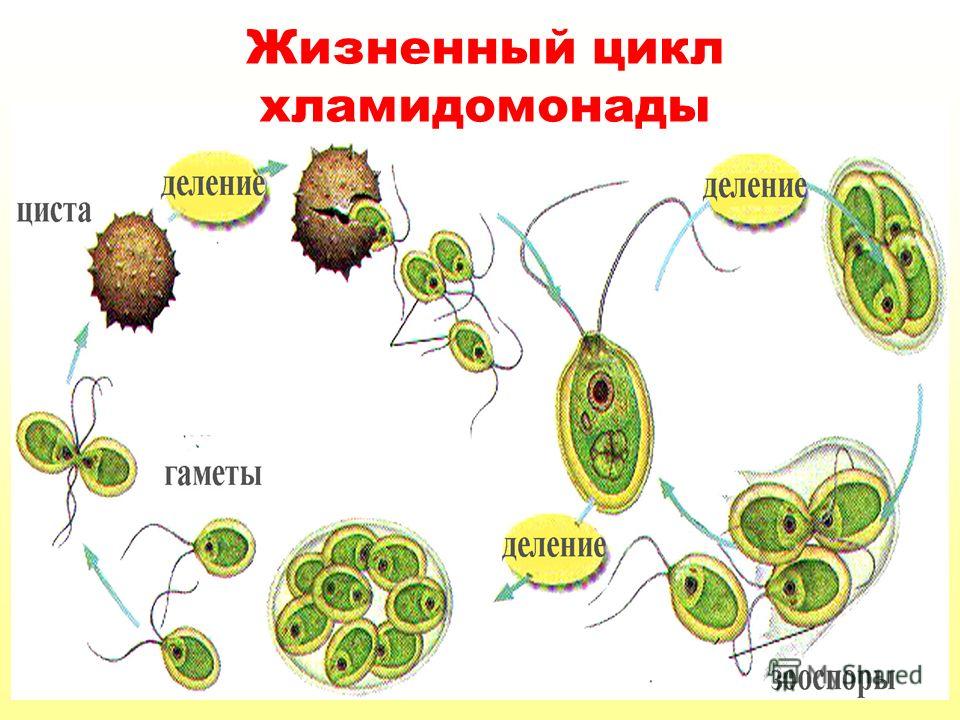 При их массовом развитии вода часто принимает зеленую окраску.
При их массовом развитии вода часто принимает зеленую окраску.
Сферическая или эллипсовидная клетка одета оболочкой, нередко отстающей от протопласта на его заднем конце. На переднем конце хламидомонада несет 2 жгутика, с помощью которых клетка активно передвигается в воде. Протопласт содержит 1 ядро, обычно чашевидный хроматофор (в который погружены 1 – 2 пиреноида и глазок) и пульсирующие вакуоли, находящиеся в передней части клетки (рис. 1.37).
Рис. 137. Chlamydomonas: a – вегетативная особь; б – пальмеллевидная стадия; в — деление клетки; г – мододые особи внутри материнской клетки
При благоприятных условиях хламидомонады размножаются бесполым и половым путем. При бесполом размножении клетка останавливается, теряет жгутики и протопласт последовательно делится на 2, 4 или 8 частей, которые одеваются собственной оболочкой, вырабатывают жгутики и превращаются в зооспоры, покидающие материнскую клетку. От последней они отличаются только размерами. После непродолжительного роста молодые особи приступают к размножению. Половой процесс у большинства видов хламидомонады изогамный, реже гетеро– или даже оогамный. Зигота прорастает после периода покоя, как правило, четырьмя гаплоидными зооспорами. Гаметы образуются внутри материнской клетки также, как и зооспоры, но в большем количестве (32 – 64) и соответственно меньших размеров.
После непродолжительного роста молодые особи приступают к размножению. Половой процесс у большинства видов хламидомонады изогамный, реже гетеро– или даже оогамный. Зигота прорастает после периода покоя, как правило, четырьмя гаплоидными зооспорами. Гаметы образуются внутри материнской клетки также, как и зооспоры, но в большем количестве (32 – 64) и соответственно меньших размеров.
При недостатке кислорода и подсыхании водоемов хламидомонады переходят в пальмеллевидное состояние: у них исчезают жгутики, выделяется обильная слизь и клетка начинает делиться.
Известно 500 – 600 видов хламидомонад, из них 18 в Беларуси. Широко распространены X. Акимовой (Chl. akimovii), X. сфагновая (Chl. sphagnicola), X. Дебари (Chl. debaryana), X. неподвижная (Chl. immobills) и др. Они являются хорошими санитарами и обеспечивают биологическое самоочищение воды.
Порядок Вольвокальные (Volvocales). Включает наиболее высокоорганизованных представителей класса. Сюда относятся только ценобиальные и колониальные формы. Клетки их построены по типу хламидомонад, но они срастаются краями оболочек или соединяются общей слизью.
Сюда относятся только ценобиальные и колониальные формы. Клетки их построены по типу хламидомонад, но они срастаются краями оболочек или соединяются общей слизью.
Род гониум (Gonium) объединяет виды, ценобии которых имеют форму пластинок, состоящих из 4 – 32 двухжгутиковых клеток, расположенных в один слой и соединенных слизью (рис. 1.38).
У наиболее распространенного вида G. pectorale клетки соединяются друг с другом углами, образуя в общей слизистой оболочке однослойный 16-, реже 8 – 4-клеточный пакетик. Их жгутики направлены в одну сторону. Размножение бесполое – делением протопласта на 4 – 16 частей. После формирования оболочки, жгутиков и слизи зооспоры образуют новый ценобий внутри материнской клетки, откуда выходят в воду. Половой процесс изогамный.
Известно 7 видов, в Беларуси 2 (Г. пекторальный (G. pectorale) и Г. общественный (G. sociale)). Встречаются в реках, озерах и мелких водоемах.
Родэвдорина (Eudorina) (рис. 1.39) образует слизистые эллипсовидные ценобии, по периферии которых под плотным слоем слизи расположены 16 – 64 клетки, а центр занят менее плотной слизью. Клетки ориентированы таким образом, что их передние концы со жгутиками направлены к периферии ценобия. Это имеет важное значение для движения эвдорины. Размножение бесполое – делением протопласта на зооспоры, половой процесс оогамный. У двудомных организмов антеридии и оогонии формируются в разных ценобиях. При неблагоприятных условиях переходят в пальмеллевидное состояние.
1.39) образует слизистые эллипсовидные ценобии, по периферии которых под плотным слоем слизи расположены 16 – 64 клетки, а центр занят менее плотной слизью. Клетки ориентированы таким образом, что их передние концы со жгутиками направлены к периферии ценобия. Это имеет важное значение для движения эвдорины. Размножение бесполое – делением протопласта на зооспоры, половой процесс оогамный. У двудомных организмов антеридии и оогонии формируются в разных ценобиях. При неблагоприятных условиях переходят в пальмеллевидное состояние.
Рис. 1.38. Gonium. Ценобий
Рис. 1.39. Eudorina elegans. Ценобий
Известно 9 видов эвдорин, распространенных в небольших стоячих водоемах, где летом часто вызывают «цветение» воды. В Беларуси и других странах Европы широко распространенным видом является Э. изящная (Е. elegans).
У рода пандорина (Pandorina) (рис. 1.40), как и у эвдорины, ценобии микроскопически мелкие, образованы клетками хламидомонадного типа, эллипсоидальные или шаровидные, состоящие из 16 – 32 двухжгутиковых клеток, расположенных по периферии. Клетки лежат в слизи очень тесно, давят друг на друга, вследствие чего имеют яйце-, конусовидную или многогранную формы. Общая полость очень мала. Размножение бесполое (путем последовательного деления протопласта всех клеток ценобия на 16 – 32 дочерних). Половой процесс гетерогамный или изогамный. Ценобии двудомные.
Рис. 1.40. Pandorina. Ценобий
Известно 2 наиболее распространенных вида: П. ежевиковая (P. morum) встречается в различных водоемах и часто вызывает «цветение» воды, П. харьковская (P. charkoviensis) отмечена в почве.
Род вольвокс (Volvox) (рис. 1.41) включает наиболее высокоорганизованных представителей класса. Их ценобии имеют вид слизистых, подвижных, диаметром 1 – 2 мм шаров, в периферическом слое которых расположено 500 – 60 000 хламидомонадоподобных клеток, сросшихся своими боковыми стенками друг с другом и соединенных одна с другой плазмодесмами.
В отличие от колониальных вольвокофициевых, где все клетки одинаковы и в равной мере способны к размножению, у вольвокса наблюдается дифференцировка, или специализация клеток на вегетативные, не способные к размножению, и репродуктивные. Последних немного, и они расположены в задней (по движению) части ценобия. Около десятка из них – клетки бесполого размножения, так называемые партеногонидии, которые в результате многократных делений образуют дочерний ценобий внутри материнского. Половой процесс у вольвокса оогамный. Оогонии и антеридии формируются из репродуктивных клеток. В грушевидных, с темно-зеленым содержимым оогониях возникает 1 яйцеклетка. В ангеридиях образуется 32 – 64 вытянутых двухжгутиковых сперматозоида, сложенных в виде пакетика (пластинки). После оплодотворения формируется зигота (ооспора) с многослойной шиповатой снаружи оболочкой. После периода покоя ооспора прорастает в молодую особь. Вольвоксы чаще бывают одно-, реже двудомными.
Рис. 1.41 Volvox: а – общий вид ценобия; б – часть пенобия с яйцеклетками (1) и сперматозоидами (2)
Известно 17 видов, из них 3 встречаются в Беларуси. В. шаровидный (Vglobator) и В. золотистый (Vaureus) распространены в прудах, старицах, мелководных озерах. Массовое развитие вызывает «цветение» воды.
Задания
1. Рассмотреть и зарисовать строение клетки хламидомонады. Отметить пектиновую оболочку, две пульсирующие вакуоли вблизи переднего конца тела, красный глазок (стигму) рядом с вакуолью, чашевидный хроматофор, пиреноид, ядро, два равных жгутика. Чтобы рассмотреть последние, надо окрасить препарат 2%-м раствором метиленовой сини или раствором Люголя. Для этого раствор по каплям наносят на предметное стекло с одного края покровного стекла, а с противоположного края оттягивают воду полоской фильтровальной бумаги.
2. Рассмотреть и зарисовать ценобии вольвокса, гониума, эвдорины и пандорины. Отметить у вольвокса вегетативные клетки, составляющие колонию, плазмодесмы, репродукnивные клетки (располагаются в задней части ценобия, более крупные, чем вегетативные клетки), оогонии и антеридии, дочерние особи. Обратить внимание на количество клеток и их расположение в каждом ценобии.
Вопросы и задания для самоконтроля
1. Какие типы структуры таллома характерны для отдела Зеленые водоросли?
2. Назовите пигменты и продукты ассимиляции зеленых водорослей.
3. Охарактеризуйте строение клетки вольвокофициевых, роль всех ее структурных компонентов.
4. Каково строение и размножение ценобиальных форм вольвокофициевых?
5. Опишите строение и жизненный цикл вольвокса.
6. Каково происхождение вольвокофициевых и каковы направления их эволюции?
Класс Протококкофициевые (Protococcophyceae), или Хлорококкофициевые (Chlorococcophyceae)Объединяет коккоидные формы, т.е. одноклеточные, ценобиальные, колониальные, изредка многоклеточные организмы, неподвижные в вегетативном состоянии. Они имеют микроскопические размеры, и лишь водяная сеточка достигает 1 м.
У протококкофициевых преобладает бесполое размножение при помощи зоо-, гемизоо– и автоспор. Гемизооспоры, или амебоиды, – самые примитивные специализированные клетки бесполого размножения. Они не имеют жгутиков и оболочек, способны передвигаться амебоидно. Половой процесс наблюдается редко. Обычно это изогамия, реже гетеро– и оогамия. Покоящимися стадиями служат акинеты и цисты.
В теоретическом отношении протококкофициевые интересны тем, что в процессе эволюции зеленых водорослей на этом этапе впервые возникла и утвердилась коккоидная, типично растительная структура тела с целлюлозными оболочками, от которых развились новые ступени морфологической дифференциации таллома: сифональная, сифонокладальная, нитчатая, разнонитчатая и пластинчатая.
Протококкофициевые встречаются во всех водоемах, в планктоне и бентосе, в наземных и почвенных биоценозах. По количеству видов они уступают лишь диатомовым.
В классе выделяют 3 порядка: Вакуольные, Хлорококкальные и Прототрихальные. Наиболее типичным представителем данного класса является порядок Хлорококкальные. Вакуольные (Vacuolates) являются переходным звеном между вольвокальными и протококальными, а прототрихальные (Prototrichales) — между протококальными и улотрихальными.
Порядок Хлорококкальные (Chlorococcales). Объединяет водоросли с наиболее выраженной коккоидной структурой, т.е. имеются плотные оболочки, а у вегетативных клеток отсутствуют пульсирующие вакуоли. Нет у них ни глазка, ни псевдоцилий, не наблюдается и тенденции к образованию многоклеточного таллома. Преимущественно одноклеточные формы, хотя есть колониальные и ценобиальные. Размножение хлорококковых осуществляется зоо– и автоспорами. Половой процесс изо-, гетеро– и оогамный. В порядке более 20 семейств.
Род хлорелла (Chlorella) широко распространен в пресных водоемах, на сырой земле, коре деревьев, входит в состав лишайников. Одиночные шаро– или эллипсовидные клетки одеты гладкой оболочкой, содержат пристенный, цельный, рассеченный или лопастный чашевидный хроматофор с пиреноидом и ядро (рис. 1.42).
Размножается хлорелла автоспорами, которые образуются по 2 – 4, 16 – 64 в клетке. Известны покоящиеся стадии – акинеты. Половое размножение не отмечено.
Практическое значение хлореллы чрезвычайно велико. Она используется для очистки сточных вод на сахарных и других заводах, для регенерации воздуха в замкнутых биологических системах (например, в космических кораблях). Белки хлореллы содержат все незаменимые аминокислоты. В Беларуси отмечено 5 видов. Широко распространены X. обыкновенная (Ch. vulgaris), X. почвенная (Ch. terricola), X. эллипсовидная (Ch. ellipsoidea).
Рис. 1.42. Chlorella: а – вегетативная клетка; б – образование автоспор; в – выход автоспор из материнской клетки
Рис. 1.43. Chlorococcum: а – взрослая клетка; б – образование зооспор; в – выход зооспор из материнской клетки; г – зооспора; д — молодые особи
Род хлорококк (Chlorococcum) (рис. 1.43) включает представителей с шар о– или эллипсовидными клетками, с гладкой, у отдельных видов слоистой, оболочкой. Отличительным признаком их является наличие глубокочашевидного, толстого, постенного, с одним крупным пиреноидом хроматофора. Ядро одно, в старых клетках их несколько. Внешне виды данного рода сходны с хлореллой, но первые крупнее и размножаются двухжгутиковыми зооспорами, изредка апланоспорами. Распространены в тех же биотопах, что и хлорелла.
Известно более 40 видов, в том числе 4 в Беларуси. Наиболее часто встречаются X. наземный (Ch. humicola), X. инфузионный (Ch. infusionum).
Род гидродикцион, или водяная сеточка (llydrodictyon). интересен тем, что его ценобий имеет вид сетчатого мешка, размеры которого могут достигать в длину до 1 м (чаще 15 – 20 см), а в ширину – 10 – 15 см. В состав ценобия входит несколько тысяч клеток. Они крупные, длиной до 1 – 1,5 см, цилиндрической формы, одеты целлюлозными оболочками. Клетки соединяются между собой (по 3 – 4) концами таким образом, что образуют 4 – 6-угольные пустотные ячейки и внешне напоминают рыболовную сеть (рис. 1.44).
Рис. 1.44. Hydrodictyon: а – молодая сеточка внутри материнской клетки; б — часть сеточки
Протопласт их содержит сетчатый хроматофор с многочисленными пиреноидами и до 20 000 – 30 000 ядер. Центральную часть клетки занимает крупная вакуоль.
При бесполом размножении в любой из клеток ценобия образуется до 20 000 зооспор, внутри которой они теряют жгутики и складываются в новую сеточку. Освобождение дочернего ценобия происходит после ослизнення оболочки материнской клетки. В дальнейшем ценобий растет за счет увеличения размеров клеток.
Половой процесс изогамный. В клетках развиваются мелкие двухжгутиковые гаметы (до 30 000), которые выходят в воду и, попарно копулируя, образуют зиготу. После периода покоя зигота прорастает 2 – 4-гаплоидными зооспорами. Каждая зооспора превращается в многоугольную звездообразную пластинку – полиэдр. Он увеличивается в размерах, становится многоядерным и распадается на двухжгутиковые зооспоры, которые внутри полиэдра слагают водяную сеточку.
Обитает гидродикцион в реках, прудах, ямах, богатых азотистыми соединениями.
Известно 5 видов. В нашей стране встречается 1 вид – Г. сеточный (H. reticulatum).
У водорослей рода педиаструм (Pediastrum) ценобии плоские, пластинчатые, округлой или звездчатой формы, состоят из 4 – 128 клеток, которые плотно прилегают друг к другу или образуют межклетники в виде отверстий в ценобии (рис. 1.45). Клетки одноядерные (старые многоядерные), периферийные, в отличие от внутренних сильно выемчатые, с 1 – 4 лопастями и выростами. Оболочки клеток гладкие, бородавчатые, морщинистые, складчатые. Хроматофор пристенный с одним пиреноидом. Размножение бесполое – зооспорами. Они выходят из клеток в слизистом пузыре и, еще находясь в нем, складываются в дочерний ценобий. Половой процесс и развитие зиготы сходны с половым процессом водяной сеточки и включают в себя стадию полиэдра. Известны стадии покоя – акинеты.
Рис. 1.45, Pediastrum: 1 – вегетативная клетка; 2 – клетка с зооспорами;3 – пустые клетки после выхода зооспор
В Беларуси установлены 13 видов и множество разновидностей.
Наиболее известны П. угловатый (P. angulosum), П. бориана (P. boryanum), П. двухлучевой (P. biradiatum), П. простой (P. simplex), П. удвоенный (P. duplex) Встречаются в планктоне озер, водохранилищ, рек, на влажных скалах.
У видов рода сценедесмус (Scenedesmus) (рис. 1.46) ценобии имеют вид плоских, прямых или изогнутых пластинок из 4 – 8 (реже 2 – 32) продолговатых или округлых клеток, соединенных боками в простые или двойные ряды. Оболочки гладкие или с бородавками, ребрами, шипами или сосочками, часто на концах клеток с длинными шипами или рогами. Хлоропласт пластинчатый, с боковым пиреноидом и центральным ядром. Размножается автоспорами, которые внутри материнской клетки слагаются в дочерний ценобий.
Рис. 1.46. Scenedesmus. Ценобий
Сценедесмус широко распространен в планктоне, часто встречается в прибрежной зоне среди нитчаток, мхов и др.
Известно более 200 видов, из них 26 отмечены в нашей республике. Часто встречаются С. четырехрогий (S. quadricauda), С. остроконечный (S. acuminatus), С. мелкозубчатый (S. denticulatus) и др.
Задания
1. Приготовить препараты хлореллы и хлорококка и рассмотреть при малом и большом увеличении микроскопа. Зарисовать отдельную клетку. Отметить оболочку, ядро, чашевидный хроматофор с пиреноидом, цитоплазму.
2. Рассмотреть и зарисовать форму клеток гидродикциона, характер их сочленения при образовании ячеек сеточки. Отметить оболочку клетки, сетчатый хроматофор с большим количеством пиреноидов, многоядерность, постенное расположение цитоплазмы. Найти и зарисовать часть молодой (дочерней) сеточки.
3. Рассмотреть и зарисовать общий вид ценобия педиаструма и сценедесмуса. Обратить внимание на форму клеток, характер сочленения их друг с другом, отсутствие в некоторых клетках педиаструма внутреннего содержимого (пустые клетки) после выхода из них зооспор.
Вопросы для самоконтроля
1. Каковы особенности строения клетки протококкофициевых?
2. Какие типы размножения характерны для протококкофициевых?
3. Каково различие между зоо-, аплано-, гемизоо– и автоспорами? Какие из них являются более совершенными и почему?
4. Каковы общие черты в образовании колоний у протококкофициевых и равножгутиковых?
5. Каковы особенности цикла развития водяной сеточки?
6. Каково теоретическое и практическое значение протококкофициевых водорослей?
Класс Улотрихофициевые (Ulothrichophyceae)Класс объединяет нитчатые и пластинчатые (паренхимитозные) одноядерные водоросли. Среди нитчатых форм есть свободноживущие и прикрепленные, ветвящиеся и неветвящиеся, разнонитчатые, живущие одиночно или скоплениями. Пластинчатые формы бывают одно-, двух– или местами многослойными, распростертыми по субстрату и прикрепленными только в одном месте.
Улотрихофициевые являются продолжением эволюционной линии зеленых водорослей с более высокой ступенью организации. Это выражается в многоклеточности их представителей и способности всех клеток делиться и наращивать таллом.
В классе 7 порядков, из которых большой интерес представляют Улотрихальные, Ульвальные, Хетофоральные и Эдогониальные.
Порядок Улотрихальные (Ulothrichales). Слоевище улотрихальных построено по типу однорядной неразветвленной нити. Клетки таллома похожи друг на друга по строению и функции, за исключением базальной, которая образует прикрепляющийся к субстрату ризоид. Все клетки, кроме базальной, способны делиться и участвовать в росте водоросли, а также формировать репродуктивные клетки (споры и гаметы). Порядок включает более 16 родов.
Виды родаулотрикс (Ulothrix) (рис. 1.47) встречаются преимущественно в пресных, реже морских и солоноватых водоемах, а также в почве. Водоросли прикрепляются к подводным предметам, формируя ярко-зеленые кустики размером до 10 см и более.
Неразветвленные нити улотрикса, состоящие из одного ряда цилиндрических или бочонковидных клеток с толстыми целлюлозными оболочками, прикрепляются к субстрату конической базальной клеткой или ризоидом. Характерной особенностью представителей данного рода является строение хроматофора. У улотрикса опоясанного он имеет вид постенной пластинки, края которой не смыкаются, а образуют незамкнутый поясок. У других видов хроматофор представляет собой замкнутое кольцо (цилиндр). Хорошо заметны многочисленные пиреноиды. Нити улотрикса нарастают в длину за счет деления слагающих их клеток. Вегетативное размножение осуществляется фрагментацией нити на короткие сегменты или акинетами. Для собственно бесполого размножения служат четырехжгутиковые зооспоры, реже – апланоспоры. Зооспоры образуются по 2 – 16 (32) во всех клетках нити, кроме базальной. Они выходят из материнской клетки в слизистой обертке, которая вскоре расплывается. После периода движения зооспора останавливается, сбрасывая один за другим жгутики, прикрепляется боком к субстрату и прорастает в нить.
Рис. 1.47. Ulothrix: а, б — общий вид и фрагмент нити с зооспорами и гаметами; в – зооспора; г, д — гаметы и их копуляция
Половой процесс изогамный. После слияния двухжгутиковых гамет зигота вначале двигается, затем оседает на дно водоема, вырабатывает плотную оболочку и слизистую ножку, прикрепляющую ее к субстрату. Это покоящийся спорофит (диплонт). После периода покоя он прорастает 4-16 (большей частью 8) четырехжгутиковыми зооспорами с редукцией числа хромосом. Таким образом, у улотрикса опоясанного происходит гетероморфная смена половой и бесполой форм развития: нитчатый многоклеточный гаметофит сменяется одноклеточным спорофитом. Для некоторых морских видов улотрикса характерен изоморфный цикл развития: зигота прорастает непосредственно в диплоидную нить, а редукция происходит при формировании зооспор.
Известно около 30 видов улотрикса, из них 6 встречаются в Беларуси. Чаще других распространены У. опоясанный (U. zonata), У утонченный (U. tenuissima), У изменчивый (U. varia bills) и др.
Порядок Ульвальные (Ulvales). Включает водоросли со слоевищами паренхиматозного строения пластинчатой, трубчатой или мешковидной формы и относительно крупных размеров. Слоевища ульвальных слабо дифференцированы и состоят почти из однотипных клеток. Лишь у основания они более крупные и снабжены ризоидными отростками, с помощью которых осуществляется прикрепление водорослей. Строение клеток ульвальных сходно со строением клеток улотрихальных. Размножаются ульвальные вегетативным, собственно бесполым (четырехжгутиковыми зооспорами, реже апланоспорами) и половым (изо– и гетерогамия) путями. Специальных органов размножения у них нет; зооспоры и гаметы образуются в вегетативных клетках.
В цикле развития большинства ульвальных осуществляется изоморфная смена генераций.
Рис. 1.48. Ulva. Внешний вид таллома
Обитают ульвальные в основном в морях, реже в пресных водоемах. В порядке 4 семейства, центральным и наиболее обширным из которых является семейство ульвовых.
Водоросли рода ульва (Ulva) (рис. 1.48) представляют собой двухслойное слоевище в виде широкой (до 15 см) пластинки длиной до 25 см ярко-зеленой окраски. На начальных стадиях развития ульвы образуется однорядная нить, которая переходит в трубчатую стадию, затем стенки трубки смыкаются и в дальнейшем она растет как двухслойная пластина. Для ульвы характерна изоморфная смена поколений. Спорофит размножается одно– или четырехжгутиковыми зооспорами, а гаметофит – половым путем – копуляцией изогамет. Зигота прорастает в диплоидный спорофит, у которого любая клетка может функционировать как зооспорангий. Перед образованием зооспор происходит редукционное деление ядра. Из гаплоидных зооспор развиваются гаплоидные гаметофиты, которые по внешнему виду не отличаются от спорофита.
Род представлен небольшим количеством видов, из которых широко распространены У. жесткая (U rigidш) и У. латук (U. lactyca). В ряде стран некоторые виды ульвы употребляют в пищу («морской салат»).
Род энтероморфа (Enteromorpha) характеризуется трубчатым строением таллома. Стенка трубки однослойная, таллом полый, простой или ветвящийся, вначале прикрепленный, позднее часто свободно плавающий (рис. 1.49).
Рис. 1.49. Enteromorpha: а – внешний вид таллома; б – поперечный разрез стенки таллома
На ранних стадиях таллом имеет вид двухслойной пластинки, как и у ульвы, но в дальнейшем слои расходятся, сохраняя связь по краям, в результате чего между ними образуется полость.
Род характеризуется изоморфной сменой форм развития. При прорастании зооспор и зигот энтероморфа, как и все ульвовые, проходит стадию однорядной нити, которая позднее преобразуется в трубчатое слоевище.
Встречается преимущественно в морских водоемах, реже – в пресноводных; растет на твердых грунтах, каменистых склонах в слабозагрязненных местах, где занимает до 60-70% поверхности дна.
Порядок Хетофоральные (Chaetophorales). Включает разнонитчатые ветвящиеся формы. Подавляющее большинство водорослей этого порядка имеет на слоевище волоски или щетинки. Бесполое размножение происходит при помощи четырехжгутиковых зоо– и апланоспор. Половой процесс изо-, гетеро– и оогамный. Гаметы двух– или четырехжгутиковые.
Многие хетофоральные обнаруживают черты высокой специализации в строении вегетативной и репродуктивной систем.
Распространены в пресных водоемах, реже встречаются в морях и на почве. В порядке 5 семейств. Наибольший интерес представляют 2 из них: хетофоровые и трентеполиевые. Из семейства хетофоровых (Chaetophoraceae) наиболее типичны роды стигеоклониум, драпарнальдия и плеврококк.
Водоросли рода стигеоклониум (Stigeoclonium) встречаются в водоемах в виде небольших нежных кустиков, прикрепленных к субстрату ризоидами или подошвой, представленной в виде стелющихся по субстрату нитей. От них берут начало восходящие нити, супротивно или дихотомически ветвящиеся, конечные клетки которых заканчиваются длинным бесцветным волоском (рис. 1.50). Строение клетки и размножение такое же, как и у улотрикса.
Обитают стигеоклониумы (род включает более 30 видов, из них 4 в Беларуси) в пресных и солоноватых водах в виде обрастаний разных субстратов. Широко распространены С. щетинконосный (St. setigerum), С. тонкий (St. tenue), С. наполненный (St. farctum).
Виды рода драпарнальдия (Draparnaldia) представляют собой слизистые светло-зеленые кустики, состоящие из многократно ветвящихся однорядных нитей с четкой дифференциацией на ствол и боковые ветви (ассимиляторы). Клетки ствола крупные, прозрачные, иногда слегка вздутые. Хроматофор в них продырявленный, с большим числом пиреноидов. Боковые ветви короткие, расположены мутовками и состоят из сильно ветвящихся тонких нитей, оканчивающихся длинными волосками. Прикрепляется драпарнальдия ризоидом (рис. 1.51). Половой процесс изогамный или гетерогамный, бесполое размножение осуществляется четырехжгутиковыми зооспорами.
Рис. 1.50. Stigeoclonium. Общий вид таллома
Рис. 1.51. Drapamaldia. Общий вид таллома
Известно 19 видов драпарнальдии, распространенных в прибрежной полосе озер, проточных холодных вод и растущих на разных субстратах. В Беларуси 2 вида. Широко распространены Д. скрученная (D. glomerata), Д. перистая (D. plumosa), Д. простая (D. simplex) и др.
Водоросли рода плеврококк (Pleurococcus) встречаются в виде зеленого порошистого налета на коре деревьев, на стенах, камнях и старых заборах. Клетки плеврококка округлые, одиночные или сросшиеся вместе по 3 – 4 и более, часто образуют характерные пакетики. Они одеты довольно толстой целлюлозной оболочкой с одним хроматофором, как правило, без пиреноида (рис. 1.52).
Размножение только вегетативное. Материнские оболочки после деления срастаются с дочерними. Часто в результате деления в одном направлении образуются нити, поэтому многие систематики считают плеврококк многоклеточной водорослью, имеющей редуцированный гетеротрихальный таллом.
Рис. 1.52. Pleurococcus. Общий вид таллома
Известно 5 видов. В Беларуси чаще других встречается П. обыкновенный (Р. vulgaris).
Род трентеполия (Trentepohlia) одноименного семейства включает аэрофильные водоросли, обитающие на коре деревьев, камнях, деревянных зданиях, образуя на них налет кирпично-красного или желтого цвета (за счет гематохрома, растворенного в каплях масла). Гетеротрихальный таллом состоит из стелющихся по субстрату и вертикальных нитей. Первые – короткие, разветвленные, без ризоидов, нередко распадающиеся на отдельные клетки, которые разносятся токами воздуха или воды, обеспечивая тем самым вегетативное размножение и расселение. Клетки их овальные, одеты толстыми, часто слоистыми оболочками. Хроматофоры в виде дисков, без пиреноидов. На стелющихся нитях образуются шаровидные или эллипсовидные гаметангии (рис. 1.53).
Восходящие нити образованы более вытянутыми цилиндрическими клетками. На концах этих нитей образуются овальные или шаровидные зооспорангии. Они легко отделяются и переносятся ветром или водой. Размножение бесполое, двух-, четырехжгутиковыми зооспорами, половое – двухжгутиковыми изогаметами, которые развиваются в сидячих гаметангиях. Гаметы копулируют не всегда. Они могут прорастать в нити, как зооспоры, либо превращаться в апланоспоры. Вегетативное размножение осуществляется участками нитей.
Рис. 1.53. Trentepoliliat; a – общий вид нити и клетки, отделившиеся от нее;б — нить с пустыми гаметангиями; в – нить с зооспорангиями на конце
Известно около 60 видов трентеполии, в Беларуси их 4. Широко распространены Т. затененная (Т. umbrina), Т. золотистая ( Т. aurea), Т. смолистая ( Т. piceana) и др.
Порядок Эдогониальные (Oedogoniales). Он объединяет нитчатые, одноядерные, преимущественно неветвящиеся прикрепленные формы, с особыми «колпачками», которые образуются на оболочках клеток при их делении. Зооспоры и сперматозоиды многожгутиковые, жгутики располагаются венцом. Половой процесс оогамный. Порядок включает одно семейство эдогониевых (Oedogoniaceae), представленное тремя родами.
Процесс жизненного цикла продукта — Блок-схема | График жизненного цикла продукта | График жизненного цикла продукта — шаблон
Распространение инноваций
Нормальное распределение
Five Force модель
Схема герметичного ковша
Диаграмма воронки
Лестница лояльности клиентов
Модель для шести рынков
График жизненного цикла продукта
График продленного жизненного цикла продукта
Континуальная диаграмма потребительских товаров
Блок ступенчатой диаграммы
Блок схемы ступеней
Жизненный цикл дефекта / ошибки при тестировании программного обеспечения
- Главная
Тестирование
- Назад
- Agile-тестирование
- BugZilla
- Cucumber
- Тестирование базы данных
- ETL-тестирование
- Jmeter
- JIRA
- Назад
- JUnit
- LoadRunner
- Ручное тестирование
- Мобильное тестирование
- Mantis
- Почтальон
- QTP
- Назад
- Центр качества (ALM)
- RPA
- SAP Testing
- Selenium
- SoapU
- Управление тестированием
- TestLink
SAP
- Назад
- ABAP
- APO
- Начинающий
- Basis
- BODS
- BI
- BPC
- CO
- Назад
- CRM
- Crystal Reports
- FICO
- HANA
- HR
- MM
- QM 900 Заработная плата
- Назад
- PI / PO
- PP
- SD
- SAPUI5
- Безопасность
- Менеджер решений
- Successfactors
- SAP Tutorials
Интернет
- Назад
- Apache
- AngularJS
- ASP.Net
- C
- C #
- C ++
- CodeIgniter
- СУБД
- JavaScript
- Назад
- Java
- JSP
- Kotlin
- Linux
- MariaDB
- MS Access
- MYSQL
- Node. js
- Perl
- Назад
- PHP
- PL / SQL
- PostgreSQL
- Python
- ReactJS
- Ruby & Rails
- Scala
- SQL
- SQLite
- Назад
- SQL Server
- UML
- VB.Net
- VBScript
- Веб-службы
- WPF
Обязательно изучите!
- Назад
- Бухгалтерский учет
- Алгоритмы
- Android
- Блокчейн
- Бизнес-аналитик
- Создание веб-сайта
- Облачные вычисления
- COBOL
- Дизайн компилятора
- Назад
- Встроенные системы
- E
IELTS AC Writing Task 1: диаграмма, жизненный цикл лягушки, с подробными объяснениями и лучшим модельным ответом
Это еще одна статья IELTS AC Writing Task 1 , в которой рассказывается, как написать диаграмму цикла на жизненном цикле лягушки и как использовать разные формы глаголов в ответе на задачу 1.Взгляните на заголовок диаграммы, а затем попробуйте написать ответ самостоятельно. Затем найдите здесь лучший модельный ответ и сравните с ним свое письмо. Между ответами должно быть какое-то несходство. Однако обратите внимание, есть ли большая разница в использовании времени, особенно глагольных структур. Этот простой, но простой модельный ответ, представленный здесь, должен дать минимум 7,5–8,0 баллов в письменном задании 1. Некоторые важные словари из типового ответа также объясняются в конце этого поста.
Теперь давайте посмотрим на название и изображение.
На схеме показан жизненный цикл лягушки.
А теперь рассмотрите следующие моменты, прежде чем писать свой ответ для этой диаграммы.
Посмотрите еще раз на заголовок и картинку и попробуйте определить время, которое вы будете использовать в своем ответе.
Пункт 1: Какую структуру времени вам следует использовать?
Прошлое и будущее время не используются.Итак, ответ, который вы напишете, не должен содержать прошедшее или будущее время, вам нужно использовать настоящее время в каждом написанном вами абзаце. Если вы напишете в этом ответе какое-либо предложение в прошлой или будущей форме, вы обязательно будете наказаны.
Попробуйте написать свой ответ в настоящем неопределенном времени. Вот несколько структур с примерами предложений:
Есть + есть / есть + число + существительное + расширение
На схеме 6 ступеней …… … .
Субъект + am / is / are + прилагательное + существительное
Это непрерывный процесс.
Или,
На диаграмме видно, что это непрерывный процесс.
Предмет (3-е лицо множественного числа) + основной глагол без s / es + расширение
Головастики цепляются за водные растения.
Пункт 2: Сколько абзацев нужно написать?
Если внимательно изучить схему, можно увидеть кое-что интересное. Посмотрите на различия в цвете и информацию с правой стороны. Схема состоит из двух частей; уровень воды и уровень земли.Это означает, что жизненный цикл лягушки делится на две части, а именно на период грунта и период воды. Итак, вы можете описать схему в двух основных частях. В теле 1 / параграфе 1 вы можете описать водную фазу, а в теле 2 / параграфе 2 вы можете обсудить наземную фазу.
Пункт 3: Как описать цикл?
Цикловые диаграммы в основном отображают природные процессы. В большинстве случаев вам дадут такую диаграмму по экологическим вопросам, таким как круговорот воды, загрязнение, движение газов, жизненные циклы животных и т. Д.Ярлыки на схемах предоставят вам необходимую информацию. Вы должны использовать информацию в своем ответе. Вам следует попытаться использовать глаголы из информации и изменить форму глаголов или использовать синонимы для этих глаголов в своем ответе на письменное задание 1. Вы не должны использовать один и тот же глагол снова и снова; лучше найди им подходящие синонимы. Есть разные этапы, обозначенные стрелкой. Это составляет последовательность для цикла. Вы не должны пропустить последовательность, когда пишете свой ответ.На схеме может быть указана дополнительная информация. Вы должны попробовать и их использовать.
Пункт 4: Количество слов?
Количество слов имеет решающее значение для обоих письменных заданий IELTS. Итак, вы должны иметь представление о том, сколько нужно написать для разных письменных задач. Некоторые диаграммы могут заставить вас написать более 200 слов, потому что нужно охватить очень много информации. Однако написание слишком большого количества слов не принесет вам более высоких баллов. Более 150 слов означают 151 слово или больше.Так как вы не получите дополнительных оценок за лишние слова, не пишите слишком много слов. Попробуйте написать несколько сжатых предложений, чтобы вам не приходилось писать больше слов, чем необходимо.
Пункт 5: Личное мнение? Никогда!
Это будет опасная работа и наверняка будет стоить вам баллов или очков, если вы выскажете какое-либо мнение, причины, воздействия или последствия и решения, если они не показаны на диаграмме. Никогда их вообще не пиши. Избегайте использования любых слов, ссылок или фраз, которые указывают на мнение, причины, воздействия или эффекты и решения.
Учтите вышеупомянутые моменты и получите отличный балл.
Теперь давайте посмотрим на модельный ответ.
Модель ответа:
Диаграмма описывает разные стадии жизненного цикла лягушки-амфибии. В целом жизненный цикл состоит из 8 стадий, начиная со спаривания и заканчивая развитием во взрослых лягушек.
Цикл начинается со спаривания взрослых самцов и самок лягушек на уровне земли. Впоследствии самки лягушки откладывают на воду большое количество яиц, внутри каждой из которых находится эмбрион.На следующем этапе из яйца вылупляются детеныши лягушек, и из них появляются маленькие головастики, которые цепляются за водные растения. Постепенно эти головастики увеличиваются в размерах и начинают формировать различные конечности.
Следующий этап — превращение головастиков в молодых лягушек. Их хвосты становятся длиннее, чем раньше, а конечности, такие как жабры, значительно увеличиваются. На следующем этапе головастики начинают меняться, так как их фигура увеличивается в размерах, появляются задние лапы. Затем появляются их передние лапы, и на этом этапе также начинается легочное дыхание.Таким образом, их путешествие по воде заканчивается и начинается наземная жизнь. После этого их хвосты начинают укорачиваться, которые исчезают на следующей стадии. Наконец, молодые лягушки превращаются во взрослых лягушек, которые теперь снова готовы к спариванию.
Количество слов: 198
Вот еще несколько ответов на Задачу 1:
Технологическая схема производства яблочного сока
Технологическая карта производства свитера
Технологическая схема производства оливкового масла
Схема написания рассказа
Две карты, показывающие изменения в промышленной деревне
Две карты, показывающие изменения в городе
Диаграмма жизненного цикла тунца
Диаграмма жизненного цикла тутового шелкопряда
Схема цикла движения воды
3.5 2 голоса
Рейтинг статьи
Что такое SDLC (жизненный цикл разработки программного обеспечения) Методологии этапов
Что такое жизненный цикл разработки программного обеспечения (SDLC)? Изучите этапы, методологии, процессы и модели SDLC
Жизненный цикл разработки программного обеспечения (SDLC) — это структура, которая определяет этапы разработки программного обеспечения на каждом этапе. Он охватывает подробный план создания, развертывания и обслуживания программного обеспечения.
SDLC определяет полный цикл разработки, то есть все задачи, связанные с планированием, созданием, тестированием и развертыванием программного продукта.
Жизненный цикл разработки программного обеспечения
SDLC — это процесс, который определяет различные этапы разработки программного обеспечения для предоставления высококачественного продукта. Этапы SDLC охватывают полный жизненный цикл программного обеспечения, то есть от начала до вывода продукта из эксплуатации.
Соблюдение процесса SDLC ведет к систематической и дисциплинированной разработке программного обеспечения.
Цель:
Целью SDLC является предоставление высококачественного продукта в соответствии с требованиями заказчика.
SDLC определил свои фазы как сбор требований, проектирование, кодирование, тестирование и сопровождение. Для систематического предоставления Продукта важно придерживаться этапов.
Например, Программное обеспечение должно быть разработано, и команда разделена для работы над функцией продукта, и ей разрешено работать так, как они хотят.Один из разработчиков решает сначала разработать дизайн, тогда как другой решает сначала написать код, а другой — часть документации.
Это приведет к сбою проекта, из-за чего необходимо иметь хорошие знания и понимание среди членов команды, чтобы предоставить ожидаемый продукт.
Цикл SDLC
Цикл SDLC представляет собой процесс разработки программного обеспечения.
Ниже приведено схематическое представление цикла SDLC:
Фазы SDLC
Ниже представлены различные фазы:
- Сбор и анализ требований
- Дизайн
- Реализация или кодирование
- Тестирование
- Развертывание
- Техническое обслуживание
# 1) Сбор и анализ требований
На этом этапе от клиента собирается вся необходимая информация для разработки продукта в соответствии с его ожиданиями.Любые неясности должны быть разрешены только на этом этапе.
Бизнес-аналитик и менеджер проекта назначают встречу с заказчиком, чтобы собрать всю информацию, например, что заказчик хочет построить, кто будет конечным пользователем, какова цель продукта. Перед созданием продукта очень важно понимание или знание продукта.
Например, Клиент хочет иметь приложение, которое включает денежные транзакции. В этом случае требование должно быть четким, например, какие транзакции будут выполняться, как они будут выполняться, в какой валюте они будут проводиться и т. Д.
После того, как сбор требований завершен, выполняется анализ, чтобы проверить возможность разработки продукта. В случае возникновения неясностей, устанавливается звонок для дальнейшего обсуждения.
После того, как требование ясно понято, создается документ SRS (Спецификация требований к программному обеспечению). Этот документ должен быть полностью понят разработчиками, а также должен быть рассмотрен заказчиком для использования в будущем.
# 2) Дизайн
На этом этапе требование, содержащееся в документе SRS, используется в качестве входных данных, а архитектура программного обеспечения, используемая для реализации разработки системы, определяется.
# 3) Реализация или кодирование
Реализация / кодирование начинается, как только разработчик получает проектный документ. Дизайн программного обеспечения переведен в исходный код. На этом этапе реализуются все компоненты программного обеспечения.
# 4) Тестирование
Тестирование начинается после завершения кодирования и выпуска модулей для тестирования. На этом этапе разработанное программное обеспечение тщательно тестируется, и все обнаруженные дефекты передаются разработчикам для их исправления.
Повторное тестирование, регрессионное тестирование проводится до тех пор, пока программное обеспечение не будет соответствовать ожиданиям клиента.Тестировщики обращаются к документу SRS, чтобы убедиться, что программное обеспечение соответствует стандарту заказчика.
# 5) Развертывание
После тестирования продукта он развертывается в производственной среде или выполняется первое UAT (пользовательское приемочное тестирование) в зависимости от ожиданий клиента.
В случае UAT создается копия производственной среды, и заказчик вместе с разработчиками выполняет тестирование. Если клиент найдет приложение, как ожидалось, то клиент предоставит согласие на запуск.
# 6) Техническое обслуживание
После развертывания продукта в производственной среде разработчики позаботятся о техническом обслуживании продукта, т.
Модели жизненного цикла разработки программного обеспечения
Модель жизненного цикла программного обеспечения — это описательное представление цикла разработки программного обеспечения. Модели SDLC могут иметь другой подход, но основные фазы и действия остаются одинаковыми для всех моделей.
# 1) Модель водопада
Модель водопада — самая первая модель, которая используется в SDLC. Она также известна как линейная последовательная модель.
В этой модели результат одного этапа является входом для следующего этапа. Разработка следующего этапа начинается только после завершения предыдущего этапа.
- Во-первых, выполняется сбор и анализ требований. После того, как требование заморожено, можно начинать только проектирование системы. Здесь созданный документ SRS является выходом для этапа требований и действует как вход для проектирования системы.
- В архитектуре и дизайне программного обеспечения системного проектирования создаются документы, которые служат исходными данными для следующего этапа, то есть реализации и кодирования.
- На этапе внедрения выполняется кодирование, и разработанное программное обеспечение является исходными данными для следующего этапа, то есть тестирования.
- На этапе тестирования разработанный код тщательно тестируется для выявления дефектов в программном обеспечении. Дефекты регистрируются в средстве отслеживания дефектов и повторно проверяются после исправления. Ведение журнала ошибок, повторное тестирование и регрессионное тестирование продолжаются до тех пор, пока программное обеспечение не будет запущено.
- На этапе развертывания разработанный код перемещается в производство после утверждения заказчиком.
- Любые проблемы в производственной среде решаются разработчиками, которые находятся на обслуживании.
Жизненный цикл системы | Редакция World
На этом этапе участвуют:
Настройка системы в соответствии с проектной спецификацией
Тестирование, проводимое с использованием плана, чтобы убедиться, что все части системы работают правильно с нормальными, экстремальными и ошибочными данными
- Обычные тестовые данные используются для проверки того, что система может обрабатывать данные, которые можно ожидать при повседневном использовании.
- Данные экстремальных испытаний используются для проверки того, что система может справиться с данными, лежащими в границах допустимого.
- Ошибочные (или исключительные) тестовые данные используются для проверки того, что система может идентифицировать неправильные данные и отклонять их
Тестирование с использованием нормальных, экстремальных и ошибочных данных
Установка новой системы
Может включать:
- Установка любого нового оборудования и программного обеспечения;
- Перенос данных из существующей системы в новую;
- Обучение пользователей работе с новой системой
Изготовление документации
Техническая документация
- техническое задание на проектирование системы;
- схемы систем;
- диаграммы потоков данных;
- описание различных частей системы и того, что каждая из них делает;
- макеты экранов и дизайн пользовательского интерфейса;
- план тестирования.
Пользовательская документация
- описание того, для чего предназначена система;
- минимальные аппаратные и программные требования системы;
- инструкция по загрузке и запуску системы;
- подробные инструкции по эксплуатации каждой части системы;
- Сообщения об ошибках, их значение и способы их устранения.
- Где получить дополнительную помощь, например по телефону поддержки и интерактивным учебным материалам.
Обзор после внедрения
Выполняется после того, как новая система проработает несколько недель или месяцев для выявления любых модификаций, которые могут потребоваться.
Что такое цикл разработки программного обеспечения?
Веб-разработчики и разработчики программного обеспечения во всем мире используют жизненный цикл разработки программного обеспечения (SDLC) для успешной реализации технических проектов. Отрасль программного обеспечения следует SDLC для проектирования, разработки и тестирования проектов перед развертыванием.При эффективном выполнении SDLC создает высококачественное программное обеспечение, которое соответствует ожиданиям клиентов и завершается в срок и в установленные сроки. Поскольку он определяет, какие задачи должны выполняться на каждом этапе процесса разработки программного обеспечения, SDLC — это метод контроля качества и способ гарантировать, что группы технических разработчиков остаются на одной странице.
SDLC возник в 1960-х годах как способ разработки крупномасштабных функциональных бизнес-систем. Согласно книге Global Business Information Technology , в то время деятельность информационных систем «вращалась вокруг тяжелой обработки данных и обработки чисел».В целом, этапы процесса с тех пор не сильно изменились. Он по-прежнему состоит из подробного плана, описывающего разработку, обслуживание и усовершенствования конкретного программного обеспечения. Согласно Techopedia, в организациях по разработке программного обеспечения или в командах разработчиков нетехнологических компаний жизненный цикл «определяет методологию повышения качества программного обеспечения и всего процесса разработки». Системные инженеры и разработчики используют SDLC для планирования, проектирования, построения, тестирования и доставки информационных систем.Чтобы добиться успеха, они должны выполнять это структурированно и методично, потому что каждый этап зависит от успешных результатов предыдущего.
Шесть этапов, один процесс
Типичный SDLC состоит из следующих этапов:
1. Планирование
Этот этап, который также включает анализ требований, является наиболее фундаментальной частью процесса SDLC. Это выполняется в первую очередь командой разработчиков, а также предложениями клиентов, заинтересованных сторон, исследованиями рынка, продажами и другими.Этот этап является основой анализа требований. Иногда команды изучают своих конкурентов, чтобы определить, как выделиться. После того, как все уяснят спецификации, команда может использовать информацию, полученную из внешних источников, для планирования основного подхода к проекту и выявления потенциальных рисков. Еще один аспект этапа планирования — это анализ затрат и выгод. Это включает определение того, как проект способствует достижению бизнес-целей организации.
2. Анализ
Этап анализа процесса SDLC включает определение целей проекта как функций. Затем группа разработчиков определяет, какие операции должно выполнять предполагаемое приложение. В общем, анализ требует сбора и интерпретации фактов, а также диагностики проблем с текущей системой и рекомендаций по улучшениям. Частично этого можно достичь путем изучения информационных потребностей конечных пользователей и устранения несоответствий. Думайте об этом этапе как о шаге решения проблемы.Команда разработчиков определяет, где находятся препятствия, и определяет способы их устранения с помощью нового проекта. Цель состоит в том, чтобы устранить препятствия, мешающие успешному внедрению нового продукта. На этом этапе создается документ спецификации требований к программному обеспечению (SRS), в котором определяется объем проекта. После получения одобрения руководства проект можно начинать.
3. Дизайн
На этом этапе команда использует SRS для разработки оптимальной архитектуры продукта.Требования SRS диктуют подходы к проектированию, которые включены в спецификацию проектной документации (DDS). Заинтересованные стороны просматривают этот документ, и на основе их отзывов выбирается подход к разработке. Необходимо учитывать оценку рисков, исследование рынка, модульность дизайна, бюджет и временные ограничения. Это определит архитектурные модули вместе с коммуникацией и представлением потока данных, включая внешние и сторонние модули. Внутренний дизайн архитектуры должен быть определен полностью и подробно.Компоненты, которые следует включить, — это макеты экранов, бизнес-правила, диаграммы процессов и другая документация. Разработчики и инженеры-программисты должны иметь возможность разработать и поставить систему на основе этой информации с минимальными дополнениями.
4. Реализация
Этот этап — начало собственно разработки. Во время реализации команда создает продукт и создает программный код, соответствующий DDS. На этом этапе разработчикам важно следовать руководствам по кодированию, определенным их организацией.Разработчики используют различные инструменты программирования, такие как интерпретаторы, отладчики и компиляторы для генерации кода, а также языки программирования высокого уровня, такие как C, C ++, Java и другие. Разработчики определяют язык для использования в зависимости от типа программного обеспечения и требований заказчика. В конце этого этапа программное обеспечение запускается в производство.
5. Тестирование и интеграция
Этот этап SDLC включает объединение отдельных частей проекта в специальную среду тестирования для проверки на наличие ошибок, ошибок и других проблем.На этапе тестирования продукт проверяется, чтобы гарантировать, что дефекты будут зарегистрированы, отслежены, исправлены и снова протестированы до тех пор, пока продукт не будет соответствовать стандартам качества. Некоторые компании не рассматривают тестирование как официальный этап процесса. Они рассматривают это как часть обслуживания. В этих случаях он может называться обзором после внедрения. При тестировании задаются такие важные вопросы, как:
- Отвечает ли новая система бизнес-требованиям и целям?
- Насколько это надежно?
- Остались ли ошибки?
- Функционирует ли он в соответствии с утвержденными функциональными требованиями?
На этом этапе также рекомендуется оценить сам процесс разработки, чтобы определить, есть ли аспекты, которые не пошли по плану.Команда разработчиков также должна изучить любые аспекты готового продукта, которые не оправдали ожиданий клиентов. Это позволяет команде исправить ошибки и несоответствия для следующего проекта.
6. Техническое обслуживание
Когда тестирование завершено и продукт готов к развертыванию, пора выпустить его на рынок. Иногда это происходит поэтапно или сразу, в зависимости от бизнес-стратегии организации. Например, продукт может быть выпущен только для текущих клиентов.По отзывам, изменения могут быть внесены до того, как произойдет полное развертывание. После выпуска продукта заказчику проводится техническое обслуживание. При необходимости команда вносит улучшения в программное обеспечение или вносит запросы на изменение. Конечная цель этапа технического обслуживания — обеспечить актуальность и высокое качество продукта. Он включает в себя постоянную оценку производительности системы.
Преимущества SDLC оправдывают его широкое использование в организациях любого типа. Для тех, кто заинтересован в карьере разработчика программного обеспечения, важно понимание этой фундаментальной структуры.Различные этапы SDLC гарантируют, что заинтересованные стороны и технические команды имеют четкое представление об ожидаемой производительности, а также о плане, обеспечивающем достижение желаемых результатов. «До тех пор, пока соблюдаются изложенные планы SDLC, удобство использования и успех программного обеспечения гарантированы», — говорит GeekInterview.com.
Разработка программного обеспечения в Husson
Требования к образованию для разработчиков программного обеспечения продолжают расти. Получение степени бакалавра может помочь людям добиться успеха в карьере в этой быстрорастущей области.По данным Бюро статистики труда, с 2014 по 2024 год занятость разработчиков программного обеспечения вырастет на 17 процентов — намного быстрее, чем в среднем по всем профессиям. Основная причина быстрого роста — большой рост спроса на компьютерное программное обеспечение.
Husson University предлагает онлайн-программу бакалавриата по разработке программного обеспечения, которая предназначена для обучения студентов навыкам, необходимым для разработки, создания и модификации корпоративного программного обеспечения или специализированных служебных программ.Практическая программа университета учит студентов анализировать потребности клиентов и разрабатывать программные решения с помощью таких процессов, как SDLC.