Паразитические простейшие
Автор статьи Азбукина Н.В.
- Excavata
Мы начнем знакомство с паразитическими простейшими с жгутиконосцев.
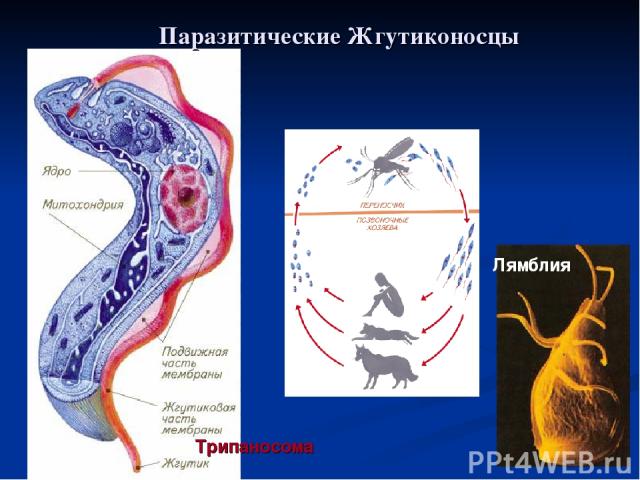
Одним из самых известных представителей является трипаносома (Trypanosoma). Этот организм обладает особой органеллой – кинетопластом, являющимся видоизмененной огромной митохондрией. Кинетопласт содержит митохондриальный геном, представляющий собой множество копий ДНК, организованной в макси- и миникольца. В миникольцах закодирована РНК, редактирующая генетическую информацию максиколец. Кроме того, в зрелой форме трипаносомы имеют ундулирующуя мембрану – пластинку, натягивающуюся между жгутиком и клеточной мембраной паразита, служащую для движения. По мере развития жгутик и кинетопласт смещаются с заднего конца тела на передний. Еще одной особенностью трипаносом является их способность к антигенной вариабельности. Другими словами, паразиты время от времени меняют состав гликопротеинового слоя на мембране, и из-за этого иммунная система тратит больше времени на их распознавание.
Trypanosoma среди клеток крови
Другой представитель жгутиконосцев Лямблия (Giardia) паразитирует в верхнем отделе тонкого кишечника человека – на ворсинках двенадцатиперстной кишки. Они имеют тело грушевидной формы, 8 жгутиков, 2 ядра, присоску на брюшной стороне. Питаются полупереваренным содержимым кишечника путем пиноцитоза.
Жизненный цикл лямблий
Лейшмании (Leishmania) являются внутриклеточными паразитами и переносятся москитами. Основными хозяевами являются люди, собаки, лошади. Эти паразиты фагоцитируются сначала нейтрофилами, потом макрофагами и обитают в их фагосомах, поэтому невидимы для клеток иммунной системы. Вызывают лейшманиозы – кожные язвы.
- Alveolata
Следующие представители простейших относятся к типу Apicomplexa, названному так из-за наличия у этих организмов апикального комплекса, состоящего из кольца, коноида, роптрий и микронем, обеспечивающие проникновение в клетки хозяина. Форма тела этих организмов постоянна за счет белковой пелликулы. Кроме того, эти организмы раньше были фотосинтезирующими, а сейчас у них остался апикопласт, содержащий кольцевую молекулу ДНК и сохранивший функцию синтеза жирных кислот.
 После этого происходит слияние гамет, образуется зигота, первое деление которой является редукционным, в результате которого образуются спороцисты. Цикл замкнулся.
После этого происходит слияние гамет, образуется зигота, первое деление которой является редукционным, в результате которого образуются спороцисты. Цикл замкнулся.Тип Apicomplexa делится на два класса: Conoidasida и Aconoidasida. Приставка A- в латинских корнях обозначает отрицание, то есть у одних представителей есть коноид, а у других нет. К Коноидным относят подкласс Грегарин, паразитов насекомых. Как правило, это внеклеточные паразиты размер которых может достигать 16 мм, обитающие в кишечнике. Их тело состоит из трех отделов : эпимерит, служащий для прикрепления к стенке кишечника, протомерит и дейтеромерит. При вступлении в стадию полового размножение гамонты образуют сизигий, то есть попарно «встают паравозиком», покрываются общей цистой. Внутри нее они делятся, образуют гаметы, которые сливаясь, образуют зиготу. Зигота затем образует 8 спорозоитов. Как можно заметить, то для этих организмов характерно отсутствие мерогонии (стадии бесполого размножения).
Кроме Грегарин к Коноидным относится подкласс Кокцидий, включающих в себя паразитов позвоночных, например кроликов Eimeria magna. Все стадии развития этого организма проходят в кролике. Цисты очень «живучи», их можно убить только крутым кипятком или выжиганием, на пастбищах сохраняются годами. Поэтому этот паразит приносит колоссальный ущерб сельскому хозяйству.
Все стадии развития этого организма проходят в кролике. Цисты очень «живучи», их можно убить только крутым кипятком или выжиганием, на пастбищах сохраняются годами. Поэтому этот паразит приносит колоссальный ущерб сельскому хозяйству.
Еще одним распространенными представителеми этого подкласса являются организмы рода Toxoplasma. Основным хозяином (то есть в котором происходит половое размножение) является кошка, а промежуточным – мыши, свиньи, люди. Особо опасно заражение женщин во время беременности, так как происходит также и заражение плода.
Доказано, что мыши, зараженные токсоплазмой, перестают бояться кошек, их даже превлекает запах кошачьей мочи. Также имеются исследования о влиянии токсоплазмы на поведение людей, а также на развитие шизофрении. По предварительной оценке около 65% населения Земли является носителем токсоплазмы, многие из которых даже не подозревают об этом!
Жизненный цикл Toxoplasma
К второму классу Aconoidasida относятся преставители кровяных споровиков, к которым относится знакомый всем малярийный плазмодий.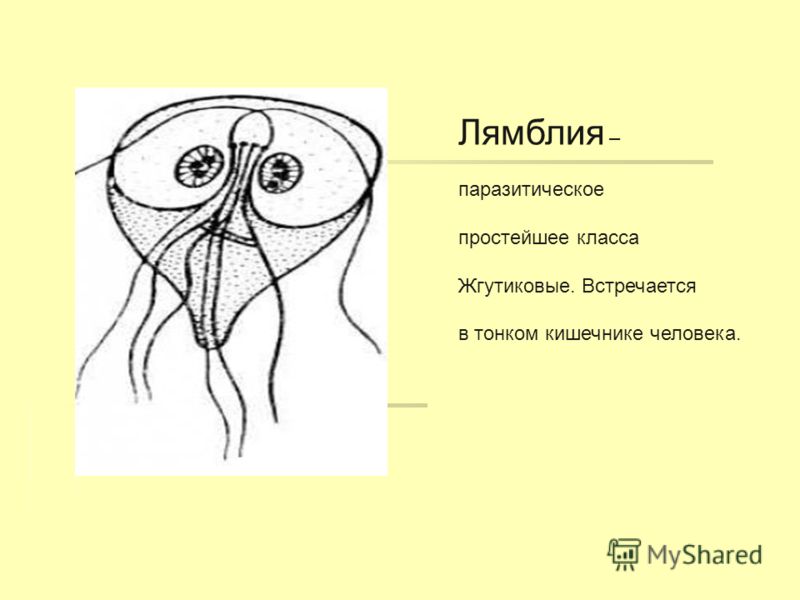 Промежуточным хозяином является человек, а окончательным- комар. У малярийного комара необычная зигота, имеющая псевдоподии и обладающая подвижностью.
Промежуточным хозяином является человек, а окончательным- комар. У малярийного комара необычная зигота, имеющая псевдоподии и обладающая подвижностью.
Жизненный цикл соответствует лейкартовской триаде, стадии мерогонии сопряжены с приступами лихорадки. Если говорить подробнее, то когда плазмоий выхоит в кровоток, имунная система обнаруживает его, начинает бороться, поэтому повышается температура и проч. Потом мерозоиты снова возвращаются в эритроциты, там размножаются и через какое-то время снова синхронно выходят в кровоток. Для разных видов плазмодиев характерна разная продолжительность этого периода, поэтому различают трех- семидневные лихорадки.
Малярия – очень опасное и по сей день распространенное заболевание, по оценке ВОЗ около 200 млн людей в год заражаются малярией, и 700 тысяч людей умирают от этого заболевания. Интересно, что за изучение малярии и лекарства от нее было присуждено 4 Нобелевские премии по физиологии и медицине.
Совсем недавно этих организмов определяли как один из классов споровиков (Apocomplexa), но сейчас выделили в отдельный тип, так как у них нет чередования мерогонии и спорогони, кроме того они имеют особые споры со створками, обеспечивающими плавучесть, и стрекательными капсулами, позволяющими им прикрепляться к стенке кишечника хозяина.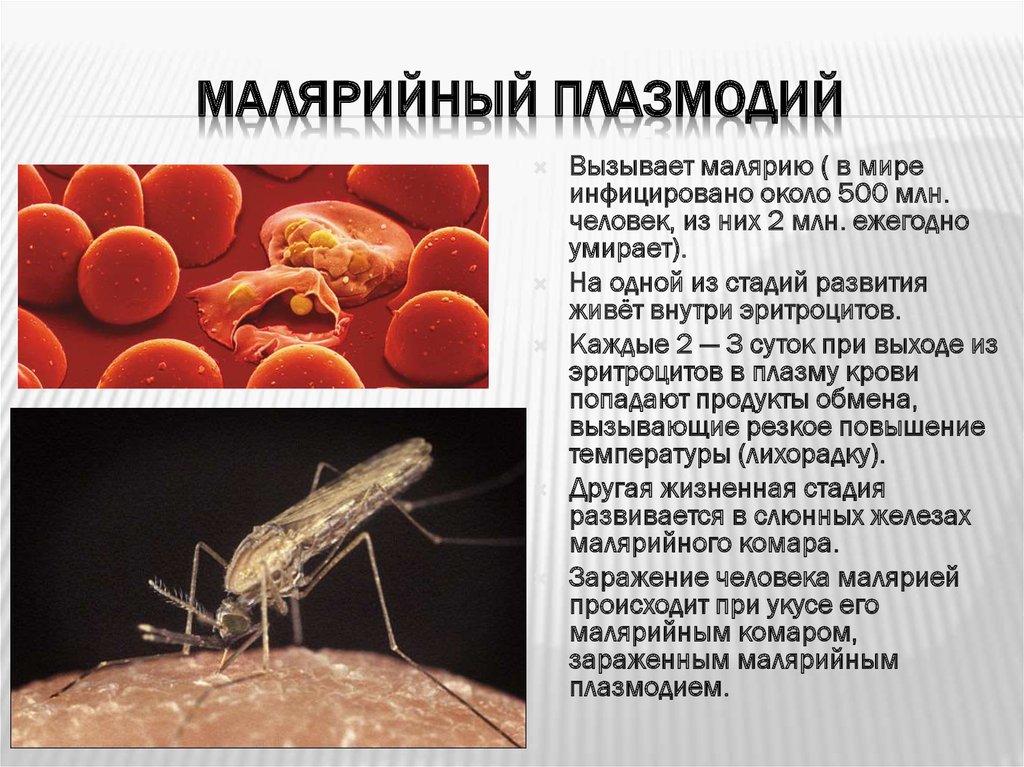
Хозяином этих организмов чаше всего выступают рыбы и амфибии, однако некоторые паразитируют и на насекомых. Взрослый организм диполоиен, представлен многоклеточным плазмодием, образует шишки на внутренних органах хозяина. У взрослого организма есть вегетативные ядра (там, где идет транскрипция белков, необходимых для жизнедеятельности) и генеративные, служащие для размножения. Генеративные ядра делятся несколько раз (причем один раз мейозом, то есть споры – гаплоидны), продуцируя двуядерные споры. Через заднюю кишку они попадают в воду, где заглатываются другими организмами. В новом хозяине оболочки спор разрываются, выходит двуядерная амеба, ядра которой впоследствии сливаются, образуя диплоидный набор хромосом. Эта амеба впоследствии даст начало новому плазмодию. Книдоспоридии наносят массовый ущерб рыбоводческим хозяйствам, вызывая массовую гибель рыб, особеннно молоди.
Тип Микроспоридии.
Это особые внутриклеточные паразиты, раньше относившиеся к споровикам, обладающие очень маленьким геномом (сопоставимым с бактериальным). Кроме того у них редуцированы многие органеллы (митохондрии, лизососмы, аппарат Гольджи). Систематическое положение этих организмов спорно: по молекулярно- генетическим данным они близки к грибам, но пока непонятно, как их определять: как сестринскую к грибам группу или включать в Fungi. Споры этих организмов имеют особый аппарат эуструзии для проникновения внутрь клетки хозяина. Все представители этого типа являются паразитами как беспозвоночных, так и позвоночных животных, и даже человека. Поражают медоносных пчел, гусениц тутового шелкопряда (болезнь – пебрина), промысловые виды рыб, нанося сельскому хозяйству миллионные убытки.
Кроме того у них редуцированы многие органеллы (митохондрии, лизососмы, аппарат Гольджи). Систематическое положение этих организмов спорно: по молекулярно- генетическим данным они близки к грибам, но пока непонятно, как их определять: как сестринскую к грибам группу или включать в Fungi. Споры этих организмов имеют особый аппарат эуструзии для проникновения внутрь клетки хозяина. Все представители этого типа являются паразитами как беспозвоночных, так и позвоночных животных, и даже человека. Поражают медоносных пчел, гусениц тутового шелкопряда (болезнь – пебрина), промысловые виды рыб, нанося сельскому хозяйству миллионные убытки.
# Зоология беспозвоночных
Конспект урока по биологии в 7 классе на тему: Паразитические простейшие. Значение простейших
Обновлено: 22.09.2022
Цель урока: создать условия для усвоения знаний о паразитических простейших и их опасности для человека.
Задачи:
Образовательная: организовать работу для усвоения знаний о простейших, об их разнообразии и их значении в жизни человека.
Развивающая: способствовать формированию умений проводить определение видовой принадлежности пресноводных простейших; продолжить формирование умений сравнивать и формулировать выводы. Развивать умения выполнять практические задания, работать в парах и самостоятельно.
Воспитательная: способствовать формированию материалистического мировоззрения. Экологическое воспитание.
Методы: словестный, наглядный, практический.
Вид урока: комбинированный.
Ход урока
I. Изучение нового материала
Беседа учителя с учащимися о живых организмах:
1) Какие функции выполняет клетка одноклеточного животного?
2) Какие организмы называют паразитами?
Учитель рассказывает о паразитических простейших и о наиболее опасных и распространенных болезнях, вызываемых ими. Учащиеся фиксируют информацию в виде таблицы:
Паразитические простейшие
Название болезни
Особенности протекания
Распространение
Внедряются в оболочку кишечника и питаются кровью, истощая организм
Мухами и с грязными руками
Живут в крови и спинномозговой жидкости, вызывая слабость, озноб, сонливость
Кровососущая муха цеце
Присасывается к клеткам кишечника, вызывая его расстройство
Грязная еда, руки и предметы
Образуют цисты в мозге, глазах, внутренних органах
В крови уничтожаются эритроциты, больной страдает от малокровия
Поражение органов пищеварительной системы, истощение
Затем учитель рассказывает учащимся о значении простейших. Учащиеся фиксируют информацию в виде схемы:
Учащиеся фиксируют информацию в виде схемы:
Далее учащиеся приступают к лабораторной работе.
Лабораторная работа №1
Тема : «Изучение мела под микроскопом».
Цель : изучить мел и определить его происхождение.
Оборудование : микроскопы, мел, презентация.
• Рассмотрите фотографии мела под микроскопом и с помощью рис. 14 из учебника на стр. 28 найдите части раковин радиолярий и фораминифер.
• Зарисуйте увиденное.
• Сделайте вывод о происхождении мела.
Примерный вариант вывода:
Вывод: мел состоит из остатков скелета простейших радиолярий и фораминивер.
I I. Рефлексия
Учащиеся делают вывод урока, отвечая на вопрос: Что вы узнали сегодня на уроке?
Паразитические простейшие
Простейшие — это одноклеточные организмы, имеющие ядра. Другими словами, простейшие — это одноклеточные эукариоты. Представителями простейших являются амебы, эвглены, инфузории, а также множество других организмов. Среди простейших немало паразитов, то есть организмов, питающихся за счет других живых организмов, при этом паразиты вредят своему хозяину, часто доводят его до гибели, но не умерщвляют его непосредственно. Представителями паразитических простейших являются дизентерийная амеба, малярийные плазмодии, представители трипаносом, лейшманий, лямблий, кокцидий, микроспоридий и др. Они являются паразитами животных и человека.
Среди простейших немало паразитов, то есть организмов, питающихся за счет других живых организмов, при этом паразиты вредят своему хозяину, часто доводят его до гибели, но не умерщвляют его непосредственно. Представителями паразитических простейших являются дизентерийная амеба, малярийные плазмодии, представители трипаносом, лейшманий, лямблий, кокцидий, микроспоридий и др. Они являются паразитами животных и человека.
Одним из самых широко распространенных и опасных заболеваний, вызываемых паразитическими простейшими, является малярия. Ее вызывают некоторые виды плазмодиев. Таких плазмодиев называют малярийными плазмодиями. Они паразитируют в клетках многоклеточных организмов, в том числе человека, который заражается ими при укусе определенного вида комара. Вид комара, который является переносчиком малярийного плазмодия, называют малярийным комаром.
Попав в кровь человека, плазмодий достигает печени, где увеличивается в размерах и размножается. Клетки плазмодия возвращаются в кровь и поражают эритроциты, в которых питаются и размножаются. Когда малярийные плазмодии выходят из эритроцитов, то их отходы жизнедеятельности попадают в кровяное русло и отравляют организм человека. У больного в это время поднимается высокая температура.
Когда малярийные плазмодии выходят из эритроцитов, то их отходы жизнедеятельности попадают в кровяное русло и отравляют организм человека. У больного в это время поднимается высокая температура.
Если больного человека снова укусит малярийный комар, то теперь уже плазмодий попадет в тело комара. Здесь паразит размножается половым способом. Дальше комар может заразить другого человека.
Другим распространенным паразитическим простейшим является дизентерийная амеба. В отличие от амебы обыкновенной, у дизентерийной более короткие и толстые ложноножки. Живет и питается она в толстом кишечнике и до поры до времени безвредна. Но когда дизентерийные амебы начинают внедряться в стенку кишечника и разрушать эритроциты крови, то начинается заболевание амебная дизентерия (амебиаз). При этом кишечник воспаляется. С током крови паразиты попадают в печень и усугубляют заболевание. Человек заражается дизентерийной амебой с помощью их цист, которые попадают в желудочно-кишечный тракт с пищей.
Паразитический простейший, относящийся к трипаносомам, вызывает у человека сонную болезнь. Данное заболевание встречается в Африке и переносит его муха цеце. Трипаносома паразитирует в крови и спинномозговой жидкости. У человека поднимается температура, возникают головные боли и др. Если во время не начать лечение, то через некоторое время нарушается сон и умственная активность.
Многие виды лейшманий могут вызывать у человека кожный лейшманиоз. При этом на коже образуется язва. Лейшманиозом человек заражается при укусах москитов, которые являются переносчиками паразитического простейшего.
§36 Паразитические простейшие, значение простейших, ответы на вопросы
ГДЗ Биология 7 класс Пасечник Суматохин Калинова
1. Где обитают простейшие?
2. Какие организмы называют паразитами?
1. Простейшие могут обитать в океане, море, пресных водоемах, почве, в организмах других животных.
2. Паразиты – это организмы, которые живут за счет других организмов, называемых хозяевами.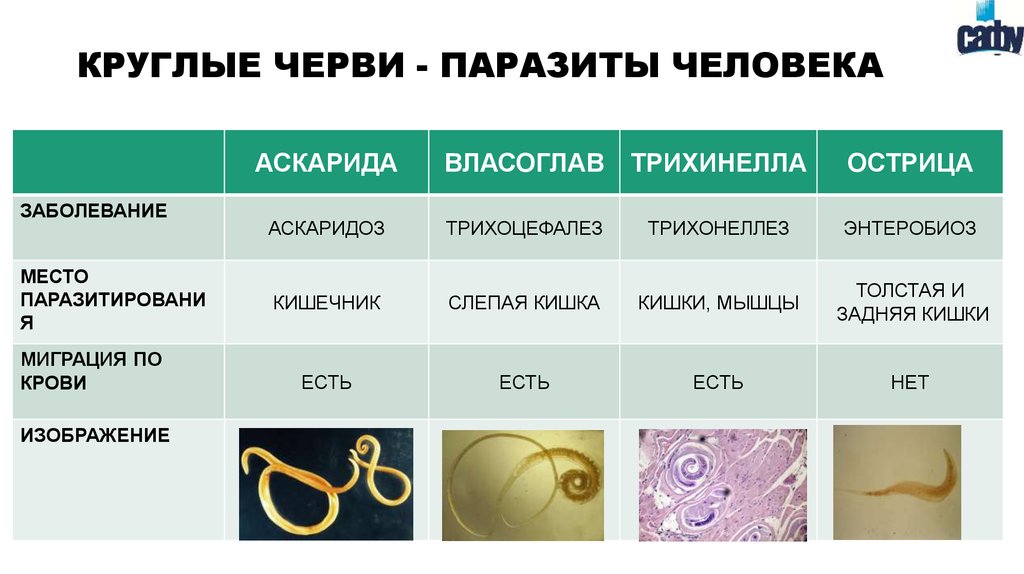
1. Какой вред здоровью человека наносит малярийный плазмодий?
2. Расскажите об образе жизни известных вам простейших-паразитов.
3. Какое значение имеют простейшие в природе?
1. Малярийный плазмодий вызывает малярию, протекающую с приступами лихорадки, изменениями в крови, увеличением печени и селезенки. Различают четыре формы малярии: трехдневную, четырехдневную, тропическую, а также овалемалярию. Источником болезни является больной малярией человек, а переносчиком – самка малярийного комара. Самка комара, заражаясь при сосании крови больного, становится способной передавать плазмодии. Здоровый человек заражается при укусе комара, инфицированного плазмодиями, с чьей слюной возбудители проникают в организм. С током крови плазмодии попадают в печень, где проходят первый (тканевой) цикл развития, переходя затем в кровь и внедряясь в эритроциты. Здесь они совершают второй (эритроцитарный) цикл развития, заканчивающийся распадом эритроцитов и вы ходом в кровь больного возбудителей, что сопровождается приступом лихорадки.
2.
Трипаносома
Трипаносома – это одноклеточный организм, бактерия, принадлежащий к семейству трипосоматид. Ведет исключительно паразитический образ жизни – в природе как самостоятельный микроорганизм не встречается. Переносчиками являются насекомые – муха цеце, триатомовые клопы, самки слепней Табанус и мухи-жигалки рода стомоксис. Трипаносомы разных видов вызывают разные заболевания — сонная болезнь, болезнь Шагаса, случная болезнь непарнокопытных, су-ауру или трипаносомоз парнокопытных.
Трипаносома – опасный паразит: обосновавшись в каком-либо органе тела млекопитающего, она полностью разрушает его, вызывая соответствующую симптоматику.
Лейшмании
Вызывают заболевание лейшманиоз. Заражение происходит через «посредника». Например, москит, укусивший больное животное, всасывает вместе с кровью промастиготы лейшманий. Когда зараженная самка москита кусает человека, ей приходится срыгивать часть содержимого, заполнившего пищевод. Таким образом, промастиготы попадают в кровь жертвы.
Лямблии
Лямблии (Lamblia intestinalis) – паразиты очень мелких размеров (в среднем 18 мкм), обитают в двух жизненных формах – трофозиод (вегетативная стадия) и цисты. Вегетативная форма является патогенной и инвазивной — способной к заражению, именно эта форма жизни вызывает симптомы у человека, т.к в этом цикле возбудитель имеет присасывающий диск на нижней поверхности тела, с помощью него он крепится к стенке тонкого кишечника и начинает свой паразитический образ жизни.
Заболевание, вызываемое лямблиями, называют лямблиоз. Человека считают носителем инфекции, если в нем есть лямблии, но никаких симптомов их присутствия не возникает.
3. Простейшие участвуют в биологической очистке водоёмов. Они служат ценным кормом для мальков рыб, личинок моллюсков и раков. Простейшие, обитающие в желудках коров, оленей и антилоп, помогают этим травоядным животным переваривать пищу. Почвенные простейшие способствуют процессу почвообразования.
Как можно использовать простейших-паразитов в хозяйственной деятельности человека?
Презентация на тему «Паразитические простейшие» 7 класс
Помогите другим пользователям — будьте первым, кто поделится своим мнением об этой презентации.
Аннотация к презентации
Презентация «Паразитические простейшие» содержит общую характеристику простейших и более подробное описание 2-х видов паразитов: дизентерийной амебы и кровяного споровика, которые способны вызывать заболевания у человека. Слайды содержат изображения простейших паразитов.
Краткое содержание
- Общая характеристика;
- Простейшие в теле человека;
- Кровяной споровик;
- Дизентерийная амеба.
Для проведения урока учителем
Презентация сделана учеником для получения оценки
Содержание
Паразитические простейшие
- Паразитические простейшие – это одноклеточные животные, которые живут за счет других организмов, называемых хозяевами. Только в организме человека паразитирует около 30 видов простейших.
- Паразитический образ жизни ведут простейшие, относящиеся к типу споровиков. Они паразитируют в организмах червей, насекомых, зверей и человека. К споровикам относят около 4 тыс.
 простейших.
простейших. - Паразиты разрушают ткани хозяина и отравляют его организм ядовитыми выделениями.
Слайд 3
Простейшие в теле человека
В теле человека встречаются различные простейшие, которые проявляют неодинаковое отношение к своему хозяину. Одни из этих форм являются истинными паразитами, так как питаются частями тела хозяина, производят разрушение его тканей, вызывают интоксикацию своими ядовитыми выделениями и в конце концов дают известный симптомокомплекс специфического заболевания (патогенные простейшие). Примером таких паразитов является дизентерийная амеба, питающаяся при известных условиях эритроцитами человека и разъедающая стенку кишки, малярийный плазмодий и др. Такие простейшие являются болезнетворными, или патогенными.
Слайд 4
Кровяной споровик – малярийный плазмодий
Для человека опасен кровяной споровик – малярийный плазмодий. Он обитает в клетках крови человека и имеет микроскопические размеры. Этот паразит распространяется малярийными комарами и вызывает опасное заболевание людей – малярию.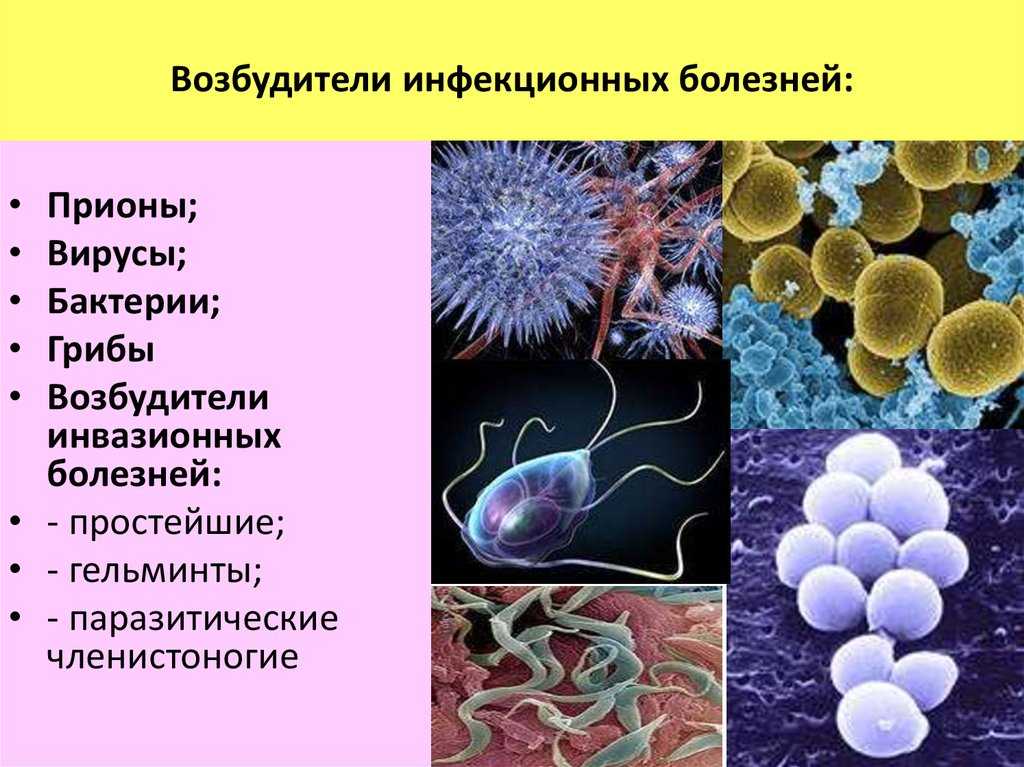 Возбудителя малярии в 1880 г. описал французский врач А. Лаверан.
Возбудителя малярии в 1880 г. описал французский врач А. Лаверан.
Слайд 5
Дизентерийная амеба
Дизентерийная амеба была открыта в 1875 г. русским ученым Ф. А. Лешем, работавшим в Петербургской медико-хирургической академии. Первая форма дизентерийной амебы (тканевая) живет в тканях кишечника человека. В этой форме дизентерийная амеба является болезнетворной (патогенной) и вызывает заболевание – дизентерию, или амебиаз. Вегетативная крупная форма амебы проникает в подслизистую стенку кишечника, питается его тканями, вызывая изъязвления стенки кишечника.
Слайд 6
Строение Дизентерийной амебы
- Сравнительно крупное, расположенное в центре ядра, шаровидное ядрышко.
- Хроматин — это вещество, в состав которого входят ДНК и белки.
Слайд 7
Паразитические простейшие значение простейших 7 класс
Согласно статистическим данным Всемирной организации здравоохранения, в мире более 4,5 млрд. человек заражены различными паразитами.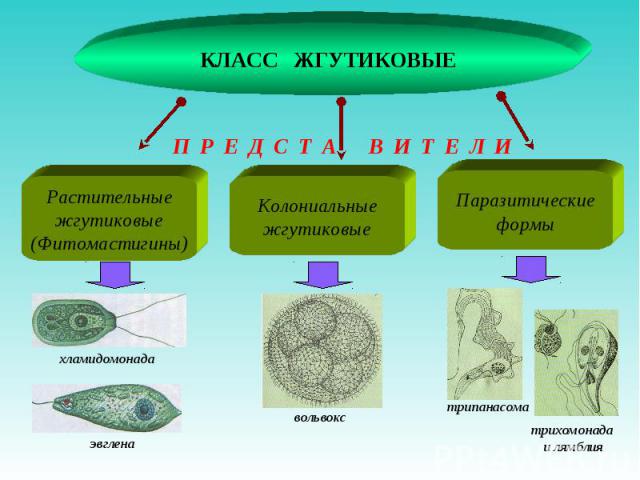 В развивающихся странах, зараженность теми или иными паразитами может достигать 93-97%.
В развивающихся странах, зараженность теми или иными паразитами может достигать 93-97%.
Существует огромное количество разнообразных паразитов, которые могут существовать во внутренних органах человека или населять его кожу и волосы. В том числе паразитарные заболевания могут вызывать и простейшие организмы, которые представляют группу одноклеточных или колониальных организмов. Большинство простейших — микроорганизмы, но некоторые могут достигают размеров в несколько миллиметров и хорошо видны невооружённым глазом. Настоящих многоклеточных форм среди простейших нет.
Способов попадания паразитов в организм человека довольно много, однако, одним из наиболее часто встречающихся путей заражения является заражение через пищевые продукты. Плохо промытые овощи, фрукты или зелень, недостаточно прожаренное или сваренное мясо, рыба, птица, неправильно обработанные продукты питания, могут привести к заражению простейшими.
Давайте же попробуем разобраться, какие простейшие, передаются с пищей и встречаются наиболее часто в нашей стране, а также как свести к минимуму риск заражения ими.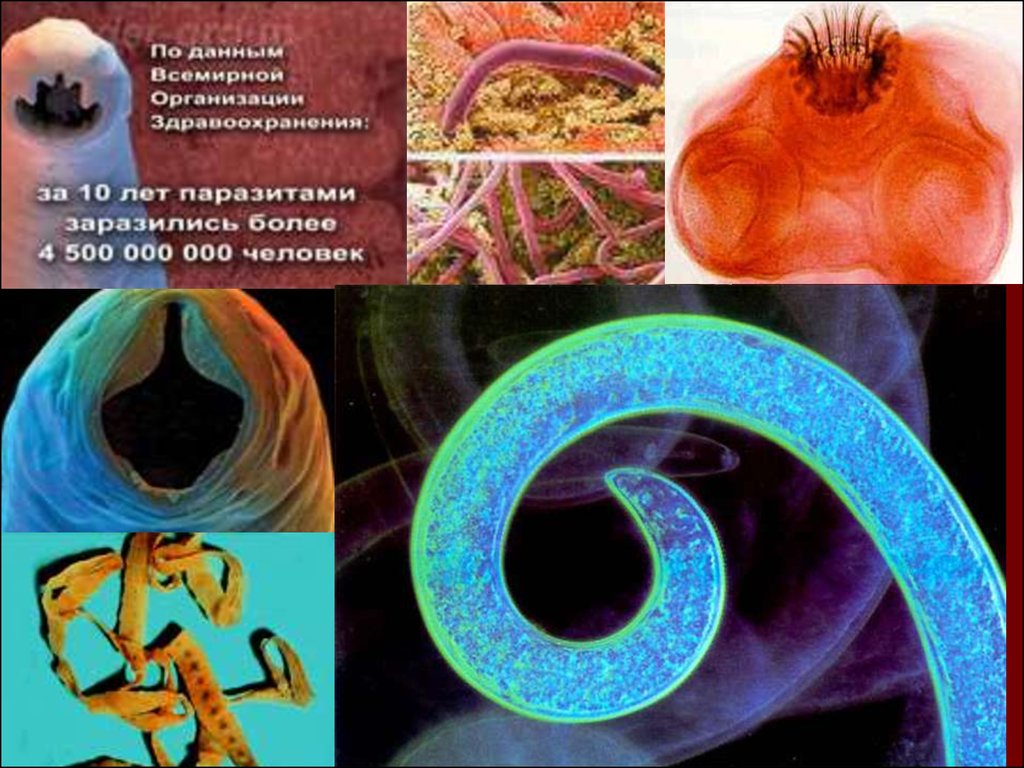
Простейшие:
Заболевание, вызывается лямблиями и поражающей кишечник человека. Лямблиоз может протекать бессимптомно, в этом случае заболевание выявляется случайно, после проведения лабораторных исследований. Клинически заболевание проявляется умеренными болями в животе, вздутием живота, тошнотой, изжогой, расстройством стула. У человека, зараженного лямблиями, могут наблюдаться повышенная утомляемость, нарушение аппетита, снижение массы тела, бледность кожных покровов, мышечные подергивания.
В случае гибели лямблий, продукты их распада всасываются из кишечника и способствуют возникновению различных форм аллергических реакций: кожного зуда, крапивницы, бронхиальной астмы.
Значительную роль в развитии заболевания играет состояние слизистой оболочки тонкой кишки и кислотность желудочного сока. У людей с пониженной кислотностью желудочного сока, нарушением слизистой оболочки кишечника, риск заражения лямблиозом, а также тяжесть клинических проявлений возрастают.
Возбудителем балантидоза является балантидий кишечный — единственный представитель инфузорий, паразитирующий в толстом кишечнике человека. Заболевание характеризуется изъязвлениями стенки толстой кишки с признаками общей интоксикации организма, но чаще протекает с незначительными клиническими проявлениями или вовсе бессимптомно. Источниками заболевания являются свиньи. Человек заболевает, как правило, при употреблении немытых овощей, ягод и зелени, где в качестве удобрений использовались свиные фекалии.
Заболевание характеризуется изъязвлениями стенки толстой кишки с признаками общей интоксикации организма, но чаще протекает с незначительными клиническими проявлениями или вовсе бессимптомно. Источниками заболевания являются свиньи. Человек заболевает, как правило, при употреблении немытых овощей, ягод и зелени, где в качестве удобрений использовались свиные фекалии.
Возникновению признаков заболевания способствует ряд факторов: нарушение питания, снижение активности иммунной системы, переохлаждение организма, воспалительные заболевания кишечника и другие. В местах локализации паразитов возникает воспалительно-язвенный процесс, происходит отмирание клеток кишечника с образованием язв. У больных возникает общая интоксикация, появляются боли в животе, понос, в фекалиях – слизь и кровь. Наиболее опасными осложнениями балантидиаза являются прободение язв с развитием перитонита и кишечные кровотечения.
Возбудителем амебиаза является дизентерийная амеба, обитающая в кишечнике человека. Заболевание характеризуется частым водянистым стулом с примесью крови и слизи, болями в животе, лихорадкой и обезвоживанием организма.
Заболевание характеризуется частым водянистым стулом с примесью крови и слизи, болями в животе, лихорадкой и обезвоживанием организма.
Заболеваемость амебиазом довольно высока — ежегодно регистрируются несколько сотен миллионов больных во всех странах. Для нескольких десятков тысяч больных заболевание заканчивается летальным исходом. В России эндемичными районами являются Закавказье, Калмыкия, Астраханская область и районы, примыкающие к среднеазиатским республикам.
Если место обитания амебы в просвете толстой кишки, то паразитирование не приводит к возникновению клинических признаков заболевания. Если же дизентерийная амеба проникает в ткани слизистой оболочки толстой кишки, то образуются язвы. У больных появляются сильные боли в животе, развивается диарея (до 15 и более раз в сутки). Отличительный признак амебиаза – стул напоминает слизь и окрашен кровью («малиновое желе»). При разрушении сосудов стенки кишечника может возникнуть массивное кровотечение.
Больные амебной дизентерией подлежат обязательной госпитализации.
Возбудителем токсоплазмоза является токсоплазма. Заболевание характеризуется хроническим течением, поражением нервной системы, увеличением печени и селезенки, поражением скелетных мышц и миокарда.
Цикл развития токсоплазмы довольно сложный: происходит не только со сменой хозяев, но и чередованием жизненных стадий паразита.
Окончательные хозяева паразита – представители семейства кошачьих — американская рысь, бенгальский тигр и домашние кошки.
Большинство случаев токсоплазмоза протекает бессимптомно. На характер проявлений заболевания существенное влияние оказывает общее состояние организма, а также активность иммунной системы.
У лиц с нарушениями иммунной системы заболевание приобретает особенно тяжелый характер. Увеличение количества очагов размножения паразита приводит к воспалительным изменениям в органах с последующим отмиранием тканей. Наиболее частое и грозное осложнение токсоплазмоза – энцефалит, летальность при котором достигает до 90%.
Профилактика паразитозов
Чтобы не допустить заражения простейшими, нужно соблюдать несколько правил, а именно:
Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания — обратитесь к врачу.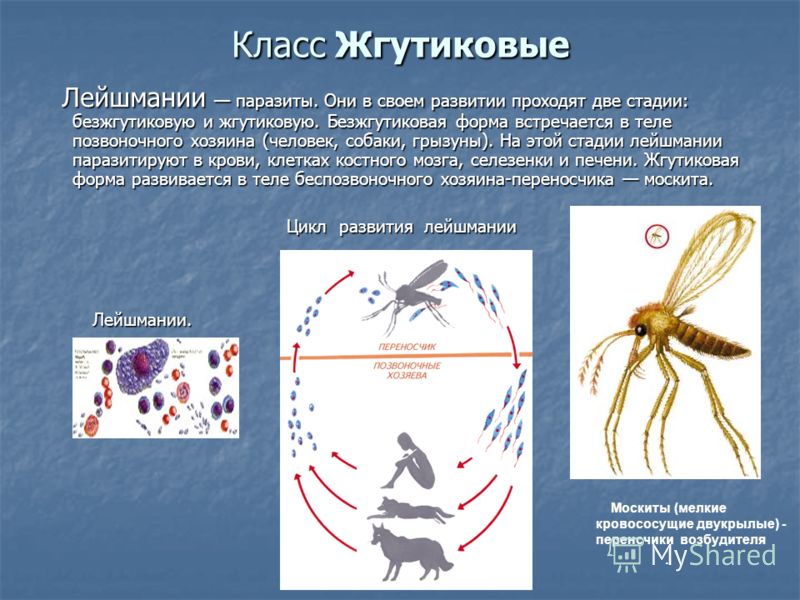
Читайте также:
- Как защитить себя и своих близких от вирусной инфекции. Осторожно Грипп!
- О моем перерождении в паразита
- Удаление бородавок лазером
- Мазь тридерм при грибке
- Вирус папилломы человека и рак головы и шеи: что нужно знать?
Возбудитель сонной болезни прячется от иммунитета благодаря разрывам двойной цепочки ДНК
Наука
Возбудители сонной болезни прячутся от иммунной системы, постоянно меняя поверхностную оболочку. Делают они это весьма активно, разрывая свою ДНК для запуска генной конверсии. По иронии, тот же механизм помогает нашим иммунным клеткам синтезировать миллионы вариантов антител.
«Гонка вооружений» между паразитами или инфекционными агентами с одной стороны и иммунной системой с другой заставляет обе стороны проявлять максимум изобретательности. И человеку вместе с остальными млекопитающими здесь действительно есть чем гордиться: естественные барьеры, секреторные антитела слюны и слезы, узкая специализация иммунокомпетентных клеток и, безусловно, гуморальный и клеточный иммунитет, основанный на сверхточном распознавании поверхностных молекул-мишеней чужеродного организма.
И человеку вместе с остальными млекопитающими здесь действительно есть чем гордиться: естественные барьеры, секреторные антитела слюны и слезы, узкая специализация иммунокомпетентных клеток и, безусловно, гуморальный и клеточный иммунитет, основанный на сверхточном распознавании поверхностных молекул-мишеней чужеродного организма.
Глобальная микроугроза
Естественный отбор, прежде не дававший распространяться страшным заболеваниям, больше не работает. По иронии…
06 ноября 11:27
Микроскопические «враги» не остаются в долгу. Они усложняют свой жизненный цикл, чтобы как можно шире распространиться по организму хозяина, как, например, малярийный плазмодий. Они поражают сами иммунокомпетентные органы и клетки, как ВИЧ. Они используют клетки первой линии защиты, чтобы как можно быстрей добраться до основной мишени, а могут даже прятаться внутри тех, кто казалось бы, должен их попросту переварить, как возбудители туберкулеза микобактерии. А перечислить всех, кто использует для обмана нашей иммунной системы многочисленные системы ложных сигналов, вообще не представляется возможным.
А перечислить всех, кто использует для обмана нашей иммунной системы многочисленные системы ложных сигналов, вообще не представляется возможным.
Но, безусловно, первую строчку в списке ухищрений, на которые идут микробы, занимает изменчивость — способность изменять поверхностные маркеры, по которым происходит адресное узнавание врага, когда он повторно проникает в наш организм. Нина Папавизилиу из Института имени Рокфеллера в Нью-Йорке и ее коллеги сумели объяснить активную перестройку «портрета» Trypanosoma brucei, вызывающей сонную болезнь.
Оказалось, что ее поверхностные гликопротеины меняются благодаря тому же механизму, который обеспечивает многообразие антител в нашем организме, — двухцепочечному разрыву ДНК с последующей конверсией генов.
Безусловно, изменчивость характерна для всех живых организмов и именно ей мы обязаны многообразием форм и яркостью красок, но механизмы этой изменчивости разнообразны и сами по себе. Развитием устойчивости к антибиотикам у синегнойной палочки или появлением нового штамма гриппа мы обязаны «пассивному пути»: разнообразные случайные мутации возникают у всех организмов, особенно при размножении, и если эта мутация оказывается жизнеопределяющей (до неузнаваемости меняет поверхность вируса или вдруг позволяет какому-то ферменту разрушать антибиотик), она закрепляется в будущих поколениях.
Второй вариант изменчивости — «направленный» поиск. Например, перестройка хромосом при образовании половых клеток, которой мы обязаны многочисленными отличиями от родителей.
К последнему методу и прибегают возбудители сонной болезни человека и крупного рогатого скота при проникновении в организм.
Сонная болезнь,
или африканский трипаносомоз – заболевание людей и животных, вызываемое паразитическим простейшим вида Trypanosoma brucei, переносчиком которого является муха цеце. Существуют три морфологически идентичных вида возбудителя: T. brucei…
После укуса мухи-цеце трипаносомы, относящиеся к эукариотам и потому являющиеся паразитами, на некоторое время остаются в небольшом шанкре под кожей. Потом они начинают распространяться по организму, вызывая нарушение функции различных органов и систем, завершающееся проникновением в мозг. А иммунная система не может справиться с паразитами из-за большого разнообразия поверхностных гликопротеинов.
Способность активно менять «портрет» была известна у трипаносом и ранее. Это подтверждает, например, тот факт, что частота изменения поверхностного гликопротеина в 3–4 раза возрастает при проникновении в хозяйский организм. Однако до Папавизилиу и соавторов публикации в Nature никому не удавалось понять механизм этого феномена.
Это подтверждает, например, тот факт, что частота изменения поверхностного гликопротеина в 3–4 раза возрастает при проникновении в хозяйский организм. Однако до Папавизилиу и соавторов публикации в Nature никому не удавалось понять механизм этого феномена.
Авторы-коллеги предположили, что молекулярный механизм такой же, как и у наших B-клеток, почти случайно набирающих из нескольких вариантов генов комплекса гистосовместимости «детали» для создания антител. Чтобы проверить гипотезу, ученые искусственно создали двухцепочечные разрывы ДНК в области гена, кодирующего поверхностный гликопротеин.
Это привело к возрастанию изменчивости в 250 раз.
После восстановления целостности нитей последовательность гена гликопротеина была уже совсем другой.
Иммунитет выдал своих врагов
Создать уникальное биологическое оружие, прицельно действующее на обладателей конкретного набора генов, стало…
14 июля 10:09
Поможет ли это в поиске лекарств, впрочем, не ясно.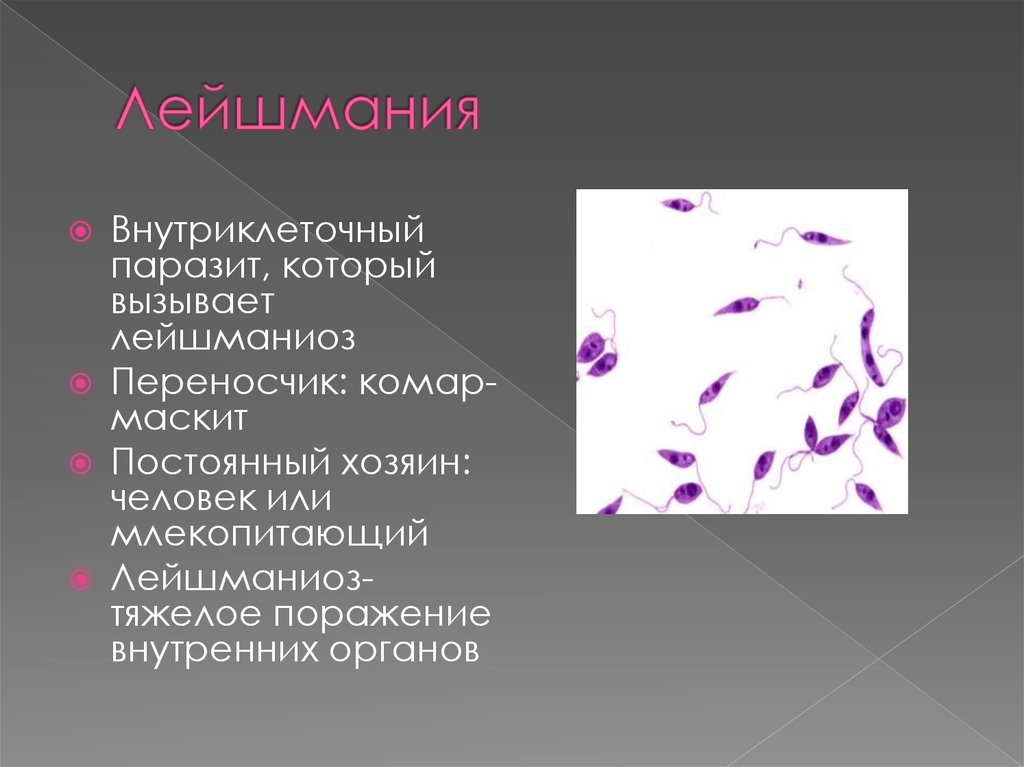 В своем опыте Папавизилиу и ее коллеги для разрыва цепочек ДНК использовали дрожжевой фермент эндонуклеазу, которая может отличаться по своим свойствам от катализаторов этого процесса у самой трипаносомы.
В своем опыте Папавизилиу и ее коллеги для разрыва цепочек ДНК использовали дрожжевой фермент эндонуклеазу, которая может отличаться по своим свойствам от катализаторов этого процесса у самой трипаносомы.
Теперь специалисты активно интересуются деталями естественного процесса. В первую очередь сигналами, вызывающими разрыв ДНК при проникновении в хозяйский организм, и ферментами, сначала разрушающими, а потом восстанавливающими целостность цепочки. Такие знания вполне могут пригодиться в борьбе с болезнями. Тем более что тот же механизм может обеспечивать жизнеспособность грибка Candida albicans, который является важной причиной осложнений при СПИДе и других нарушениях иммунитета, а также возбудителей боррелиоза и гонореи.
Паразитарные заболевания
Паразитарные заболевания человека(от греч.parasitos — «нахлебник»)– это большая группа заболеваний из раздела «инфекционные болезни», причиной их являются одноклеточные и многоклеточные паразиты.
Паразит — это живой организм, который живет или находится в организме хозяина и получает пищу от него или за его счет, то есть ведет паразитический образ жизни. Все они умеют жить в организме человека, некоторые совершенно незаметно, а некоторые могут нанести серьезный урон здоровью.
Все они умеют жить в организме человека, некоторые совершенно незаметно, а некоторые могут нанести серьезный урон здоровью.
Возбудители паразитарных заболеваний
Существует три основных класса паразитов, которые могут вызывать заболевания у людей:
Простейшие — это микроскопические одноклеточные организмы, которые могут быть свободноживущими или паразитическими по природе. Они способны размножаться у людей, что способствует их выживанию, а также позволяет развиваться серьезным инфекциям только из одного организма.
Гельминты (от греч.helmins – «черви») — это крупные многоклеточные организмы, которые обычно видны невооруженным глазом на стадии взросления. Как и простейшие, гельминты могут быть либо свободно живущими, либо паразитическими по природе. В своей взрослой форме гельминты не могут размножаться у людей.
Эктопаразиты.Этот термин обычно используется более узко для обозначения организмов, таких как клещи, блохи и вши, которые прикрепляются или прячутся в кожу и остаются там длительный период времени (от недель до месяцев). Членистоногие могут самостоятельно вызывать заражение, а могут являться переносчиками других заболеваний.
Членистоногие могут самостоятельно вызывать заражение, а могут являться переносчиками других заболеваний.
Список паразитарных инфекций (наиболее значимых на территории Российской Федерации): амебиаз, аскаридоз, гименолепидоз, дифиллоботриоз, клонорхоз, криптоспоридиоз, лямблиоз, малярия (завозные случаи), описторхоз, тениаринхоз, тениоз, токсокароз, токсоплазмоз, трихинеллез, трихоцефалез, цистицеркоз, энтеробиоз, эхинококкоз.
Признаки паразитарных заболеваний.
Проявления их могут быть разнообразны и зависят от вида и локализации паразита, а также от уровня иммунитета человека, который является их хозяином.
Простейшие чаще всего вызывают диарею и другие желудочно-кишечные симптомы. Гельминтозные инфекции могут вызывать боль в животе, диарею (понос), мышечные боли, кашель, поражения кожи, недоедание, потерю веса, нарушения координации движения и речи, судороги и многие другие симптомы в зависимости от конкретного организма и тяжести инфекции.
Диагностика и лечение паразитарных заболеваний
Диагностика паразитарных заболеваний включает:
· Клинический анализ крови
· Анализ крови на специфические антитела и антигены паразитов
· Мазок крови
· Анализ кала на яйца глист и паразитов
· Эндоскопические методы исследований (например, колоноскопия, в случае сложной диагностики)
· Рентгенография,магнитно-резонансная томография (МРТ), компьютерная томография (КТ) при серьезных поражениях внутренних органов паразитами.
План лечения будет зависеть от конкретного диагноза.Как правило, назначаютсяпрепараты в виде таблеток, иногда требуется стационарное лечение, вплоть до хирургического вмешательства.
Также могут быть рекомендованы другие методы лечения, чтобы облегчить ваши симптомы (диета, режим потребления воды).
Профилактика паразитарных болезней
Профилактика всегда проще лечения и есть достаточно простые способы защитить себя
Не употребляйте в пищу:
· недоваренную рыбу, крабов и моллюсков.
· недоваренное мясо;
· сырые водные растения
· сырые овощи, которые могли быть загрязнены фекалиями человека или животных.
Паразиты могут жить в природных источниках воды, поэтомуво время плавания:
· не глотайте воду.
· не разрешайте детям испражняться в воду, берите детей в туалет и проверяйте подгузники каждый час, меняйте подгузники в ванной или в зоне смены подгузников, а не у бассейна, чтобы избежать попадания микробов в бассейн
· не плавайте и не позволяйте детям плавать, если они болеют диареей.
· сделайте свой мини-осмотр (загрязненность)
· принимайте душ не менее 1 минуты, прежде чем погрузиться в воду.
Домашние животные могут переносить паразитов и передавать их людям. Зоонозная болезнь — это болезнь, распространяющаяся между животными и людьми. Иногда люди с зоонозными инфекциями не имеют симптомов. У других людей могут быть такие симптомы, как диарея, мышечные боли и лихорадка.
У других людей могут быть такие симптомы, как диарея, мышечные боли и лихорадка.
Регулярный ветеринарный уход защитит вашего питомца и вашу семью.
Есть простые шаги, которые вы можете предпринять, чтобы защитить себя и свою семью от заболеваний, переносчиками которых могут быть животные.
Убедитесь, что ваш питомец находится под присмотром ветеринара,
Практикуйте четыре правила:
1. Быстро соберите экскременты животных и утилизируйте правильно. Обязательно мойте руки после обращения с домашними отходами.
2. Часто мойте руки, особенно после прикосновения к животным, и избегайте контакта с фекалиями животных.
3. Соблюдайте надлежащие процедуры обращения с пищевыми продуктами, чтобы снизить риск заражения зараженной пищей.
4. Для людей с ослабленной иммунной системой будьте особенно осторожны при контакте с животными, которые могут передавать эти инфекции
Автор: врач экспертной клиники ID-Clinic Михаил Андреевич Савченко
Тип Споровики.
 Паразитические простейшие. Значение простейших
Паразитические простейшие. Значение простейшихУрок 8. Беспозвоночные животные [биология 7 класс]
Видеофрагмент подробно и доступно излагает информацию о паразитических простейших, особенностях строения споровиков в связи с их паразитическим образом жизни. Рассказывает о таком опасном заболевании, как малярия, причинах её возникновения и методах борьбы с ней. Цель урока – узнать больше о разнообразии паразитических простейших, понять какой вред они наносят другим животным и человеку.
Конспект урока «Тип Споровики. Паразитические простейшие. Значение простейших»
Тип
споровики включает около 2 тыс. видов простейших. Все представители данного
типа являются паразитами, что нашло отражение в упрощении строения
клетки. Паразитические простейшие – одноклеточные животные, которые
живут за счёт других организмов, называемых хозяевами. Свободно существовать в
окружающей среде вне хозяина споровики не могут.
Свободно существовать в
окружающей среде вне хозяина споровики не могут.
Как и все паразитические простейшие, споровики не имеют сократительных вакуолей, у них отсутствуют какие-либо органоиды движения. Пищеварительных вакуолей у них тоже нет, поэтому питание происходит всей поверхностью тела. Другой характерной особенностью споровиков является сложный жизненный цикл со сменой хозяев. Хозяева паразитов – различные позвоночные и беспозвоночные животные (черви, насекомые, млекопитающие), человек.
Обитают споровики в органах пищеварения, выделения, размножения и в крови животных и человека.
Они разрушают ткани хозяина, отравляют организм своими ядовитыми выделениями, вызывают различные заболевания, которые зачастую могут приводить к гибели.
Для
человека опасны споровики, которые являются возбудителями малярии. Это малярийный
плазмодий. На определённых стадиях своего развития он паразитирует на эритроцитах
– красных клетках крови человека. В организме человека развиваются плазмодии
трёх видов, которые являются возбудителями самой распространённой трехдневной
малярии, четырехдневной малярии и наиболее опасной тропической малярии.
Все виды широко распространены в странах с тропическим, субтропическим климатом
и экваториальным климатом. Малярийные комары живут в районах, где не бывает
низких температур и выпадает много осадков.
В организме человека развиваются плазмодии
трёх видов, которые являются возбудителями самой распространённой трехдневной
малярии, четырехдневной малярии и наиболее опасной тропической малярии.
Все виды широко распространены в странах с тропическим, субтропическим климатом
и экваториальным климатом. Малярийные комары живут в районах, где не бывает
низких температур и выпадает много осадков.
Малярийный плазмодий имеет микроскопические размеры, распространяется малярийными комарами. Переносчиками паразита являются только самки, которым для развития яиц необходимо насосать крови. И, как говорилось ранее, у человека вызывает опасное заболевание – малярию.
Рассмотрим жизненный цикл малярийного плазмодия.
В организме комара плазмодии размножаются в его кишечнике и скапливаются в слюнных железах.
При
укусе вместе со слюной комара в кровь человека проникают малярийные плазмодии. Попав в кровяное русло, они разносятся по всему телу. В клетках крови они
питаются, растут, размножаются, разрушают клетки крови человека, выделяют
ядовитые вещества.
Попав в кровяное русло, они разносятся по всему телу. В клетках крови они
питаются, растут, размножаются, разрушают клетки крови человека, выделяют
ядовитые вещества.
После разрушения эритроцита, плазмодии оказываются в крови, и у человека начинается приступ лихорадки – повышается температура, появляются озноб и слабость, мышечные и головные боли. В течение нескольких часов температура повышается до 40 °С. Продолжительность приступа невелика, и через 6 -8 часов температура снижается до нормальной. Прекращение симптомов происходит из-за того, что плазмодии не находятся долго в крови, а проникают в новые эритроциты. Приступы повторяются через каждые два-три дня, и это ведёт к истощению организма.
Малярийных паразитов комар всасывает с кровью зараженного человека.
Всемирной организацией здравоохранения разработаны и реализованы меры борьбы с малярией:
Поэтому
теперь от малярии умирает гораздо меньше людей, чем раньше.
К споровикам относится паразит грегарина. Грегарина является внеклеточным паразитом различных беспозвоночных, прежде всего насекомых. Тело этих простейших имеет продолговатую форму или реже округлую форму. На переднем конце тела грегарины имеют структуру с выростами и крючочками, позволяющую паразиту прикрепляться к стенке органа хозяина. Далее следует передний отдел и задний, несущий ядро.
Характерные признаки класса Споровики:
· внутриклеточные паразиты;
· отсутствуют органоиды движения;
· отсутствуют сократительная и пищеварительная вакуоли;
· постоянная форма тела;
· 1 ядро;
· гетеротрофный тип питания;
· сложный цикл развития.
Кроме
споровиков к паразитическим простейшим относятся некоторые представители
саркодовых, жгутиковых и инфузорий. Рассмотрим некоторых представителей данных
групп.
Рассмотрим некоторых представителей данных
групп.
К паразитическим саркодовым относится дизентерийная амёба. Она попадает в организм человека в виде цист, которые он заглатывает при несоблюдении правил гигиены (немытые руки, овощи, фрукты, сырая вода). В пищеварительном тракте человека оболочки цист растворяются. В кишечнике человека амебы питаются, размножаются и повреждают стенки кишечника, тем самым вызывая тяжелое заболевание амёбиаз. В народе амёбиаз называют «болезнью грязных рук».
К паразитическим жгутиковым относятся лямблия, трихомонада, трипаносома.
Лямблии
поражают
кишечник, печень и половые железы человека. Клетка лямблии похожа на
разрезанную пополам грушу, имеет 2 ядра и 8 жгутиков. Она попадает в организм человека
в виде цист. Заражение человека происходит при употреблении в пищу плохо
вымытых фруктов и овощей, при несоблюдении правил личной гигиены. Заражение
лямблиями вызывает заболевание лямблиоз, которое сопровождается
кишечными расстройствами.
Заражение
лямблиями вызывает заболевание лямблиоз, которое сопровождается
кишечными расстройствами.
В организме человека паразитирует инфузория балантидий. Подробно с её строением вы знакомились при изучении темы «инфузории».
Значение простейших в природе и жизни человека
Рассмотрим значение простейших. Несмотря на малые размеры тела, простейшие имеют большое значение в природе и жизни человека.
· Они участвуют в круговороте веществ;
· Поедают бактерий, тем самым обеспечивают биологическую очистку водоёмов;
· служат пищей головастикам, малькам рыб и другим водным животным;
· В морях и океанах раковины фораминифер, оседая на дно, образуют горные породы и залежи известняка;
· Являются возбудителями паразитарных заболеваний человека и животных.
Предыдущий урок 7 Тип Инфузории, или Ресничные
Следующий урок 9
Многоклеточные животные. Тип Губки
Тип Губки
Получите полный комплект видеоуроков, тестов и презентаций Беспозвоночные животные [биология 7 класс]
Чтобы добавить комментарий зарегистрируйтесь или войдите на сайт
Токсоплазма — паразит, манипулирующий человеческой культурой • Константин Попадьин • Новости науки на «Элементах» • Биология, Медицина, Психология
Чтобы завершить свой сложный жизненный цикл и попасть в конечного носителя, паразиты умело манипулируют поведением своих промежуточных хозяев. А что если таким промежуточным хозяином оказался человек? Недавно было показано, что широко распространенный в человеческой популяции паразит Toxoplasma gondii влияет на поведение не только конкретного человека, на котором он паразитирует, но и определяет особенности культуры целой нации, в той или иной степени зараженной этим паразитом.
Toxoplasma gondii — это паразитическое простейшее, жизненный цикл которого проходит через двух хозяев: промежуточного (любое теплокровное позвоночное, например мышь) и конечного (любой представитель семейства кошачьих, например домашняя кошка). Находясь в промежуточном хозяине, паразит способен к бесполому размножению. Чтобы кошка с большей вероятностью съела зараженную мышку, Toxoplasma gondii изменяет поведение мышей — они становятся более активными и смелыми, не боящимися ни кошек, ни их запаха, а также в них пробуждается интерес исследования новых территорий. В результате кошки съедают зараженных смельчаков, и паразит достигает своей цели — попадая в конечного хозяина, он приступает к половому размножению.
В современном мире кошки редко едят людей, однако зараженность токсоплазмозом в человеческой популяции в среднем около 35%. Если токсоплазма в небольшом количестве попадает в здорового человека, она покрывается оболочкой (превращается в цисту) и более для человека не опасна.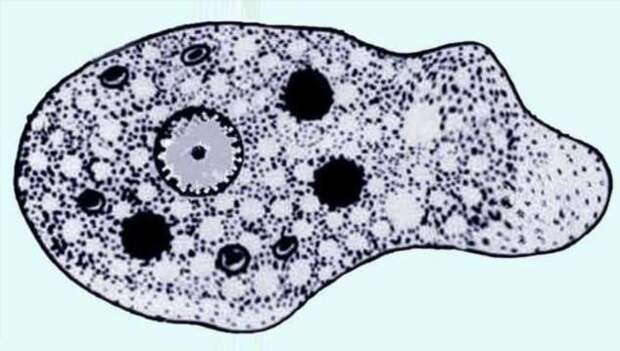 Наибольший вред токсоплазма может принести эмбриону, если мать заразилась в первом или втором триместре беременности. При таком врожденном токсоплазмозе увеличивается вероятность поражения токсоплазмой центральной нервной системы, глаз и других внутренних органов, что может даже привести к внутриутробной гибели плода.
Наибольший вред токсоплазма может принести эмбриону, если мать заразилась в первом или втором триместре беременности. При таком врожденном токсоплазмозе увеличивается вероятность поражения токсоплазмой центральной нервной системы, глаз и других внутренних органов, что может даже привести к внутриутробной гибели плода.
Помимо чисто физического вреда, наносимого токсоплазмой при инцистировании в тканях организма, существует и другой вред, определяемый взаимодействием паразита с иммунной системой организма-хозяина. Например, при инцистировании в головном мозге, токсоплазма вызывает локальный иммунный ответ хозяйского организма, в результате которого увеличивается уровень дофамина — нейромодулятора, влияющего на активность некоторых отделов лимбической системы — отдела головного мозга, отвечающего за эмоциональный настрой. У крыс, например, эта связь четко изучена, и показано, что зараженные токсоплазмой особи имеют повышенный уровень дофамина. Более того, обработка зараженных крыс антагонистом дофамина или специфичными лекарствами против токсоплазмы нормализует их поведение — они снова боятся кошек и у них угасает интерес к исследованию новых территорий. У человека искусственно вызванный избыток дофамина (при принятии стимулирующих веществ, таких как производные амфетамина) приводит к психозам, проявления которых практически не отличаются от симптомов шизофрении, или же обостряет латентную или уже начавшуюся шизофрению. Таким образом, потенциальное влияние токсоплазмы на поведение человека, скорее всего, обусловлено специфическим ответом нашей иммунной системы — а именно, увеличенным синтезом дофамина.
У человека искусственно вызванный избыток дофамина (при принятии стимулирующих веществ, таких как производные амфетамина) приводит к психозам, проявления которых практически не отличаются от симптомов шизофрении, или же обостряет латентную или уже начавшуюся шизофрению. Таким образом, потенциальное влияние токсоплазмы на поведение человека, скорее всего, обусловлено специфическим ответом нашей иммунной системы — а именно, увеличенным синтезом дофамина.
Интересно отметить, что паразит по-разному влияет на женщин и мужчин. Так, зараженные женщины становятся более интеллигентными, уравновешенными, сознательными, теплыми и отзывчивыми, а вот мужчины становятся менее интеллигентными и более грубыми. В чем причина такого различия — не ясно. Может быть, без влияния паразита обнажается истинная сущность мужского и женского начала? Вряд ли. Скорее всего, это просто результат действия токсоплазмы на организмы с разным гормональным фоном, и здесь нет никаких эволюционных механизмов, приводящих к тому, что интеллигентную женщину и неинтеллигентного мужчину проще съесть кошке. Однако оба зараженных пола — и мужчины и женщины — предрасположены к повышенному чувству вины (guilt proneness), неуверенности и беспокойству. Вот именно эти изменения в эмоциональном настрое «жертвы» могут быть эволюционным механизмом выживания токсоплазмы, ведь проблемы с эмоциональной регуляцией понижают способность к четкому и логичному мышлению, мешают бороться со стрессом и принимать правильные решения (убегать от голодной злой кошки, вместо того чтобы гладить ее).
Однако оба зараженных пола — и мужчины и женщины — предрасположены к повышенному чувству вины (guilt proneness), неуверенности и беспокойству. Вот именно эти изменения в эмоциональном настрое «жертвы» могут быть эволюционным механизмом выживания токсоплазмы, ведь проблемы с эмоциональной регуляцией понижают способность к четкому и логичному мышлению, мешают бороться со стрессом и принимать правильные решения (убегать от голодной злой кошки, вместо того чтобы гладить ее).
Поскольку чувство вины и связанные с ним симптомы также коррелируют с невротизмом (neurotiсism) — одной из основных психологических характеристик разных культур человечества, — Кевин Лафферти (Kevin D. Lafferty) из Калифорнийского университета в Санта-Барбаре заметки выдвинул предположение о зависимости между средней зараженностью разных национальностей токсоплазмозом и их невротизмом. Работа Кевина Лафферти опубликована в Трудах Королевского общества.
Описание культурного пространства разных национальностей — очень интересная область науки. В ней изучаются факторы, формирующие особенности культур, и ищутся те характеристики, которые могут наиболее адекватно отображать эти различия между национальностями. Одним из наиболее признанных подходов сейчас является выделение пяти персональных признаков (один из них невротизм), из которых и складывается культурное пространство (более подробно см. Big Five personality traits или McCrae et al. 2005, Personality Profiles of Cultures: Aggregate Personality Traits, Pdf, 910 Кб).
В ней изучаются факторы, формирующие особенности культур, и ищутся те характеристики, которые могут наиболее адекватно отображать эти различия между национальностями. Одним из наиболее признанных подходов сейчас является выделение пяти персональных признаков (один из них невротизм), из которых и складывается культурное пространство (более подробно см. Big Five personality traits или McCrae et al. 2005, Personality Profiles of Cultures: Aggregate Personality Traits, Pdf, 910 Кб).
Невротизм обратен эмоциональной стабильности и высокие его значения наблюдаются у народностей, которые по небольшим поводам долго и глубоко переживают негативные эмоции и стрессы. Например, наиболее высокие значения невротизма наблюдаются в Венгрии, Бразилии, Китае, Франции, Италии и Аргентине, тогда как наиболее низкие (более толерантные к стрессам) значения в Норвегии, Австрии, Швеции, Швейцарии, Германии и США.
Для проверки своей гипотезы Лафферти нашел опубликованные данные о значении невротизма в 39 странах, а также значения средней зараженности токсоплазмозом (она определялась по зараженности молодых беременных женщин).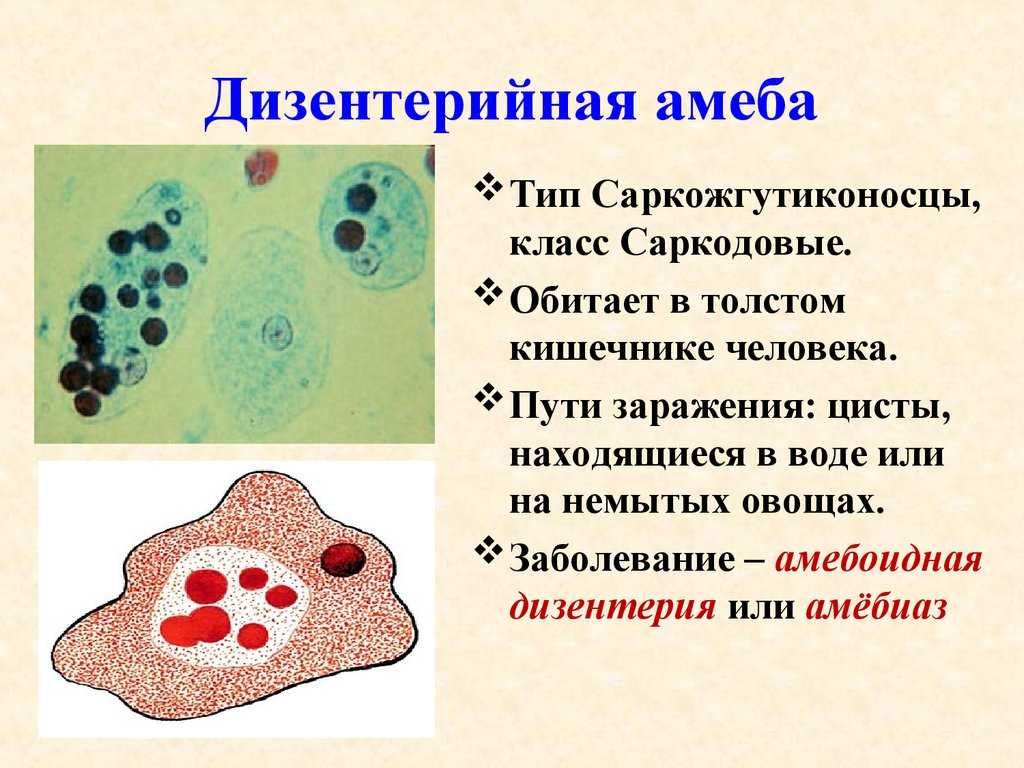 Статистический анализ подтвердил положительную корреляцию между этими двумя величинами: то есть чем больше процент зараженных токсоплазмозом людей, тем выше значения невротизма в данной стране. Таким образом, выше перечисленные страны классифицируются примерно также по степени зараженности токсоплазмой, как и по значениям невротизма. Если поверить, что эта зависимость описывает причинно-следственную связь между присутствием паразита и его влиянием на человека, то мы придем к неутешительному выводу, что одна из основных психологических осей культуры человека определяется паразитом!
Статистический анализ подтвердил положительную корреляцию между этими двумя величинами: то есть чем больше процент зараженных токсоплазмозом людей, тем выше значения невротизма в данной стране. Таким образом, выше перечисленные страны классифицируются примерно также по степени зараженности токсоплазмой, как и по значениям невротизма. Если поверить, что эта зависимость описывает причинно-следственную связь между присутствием паразита и его влиянием на человека, то мы придем к неутешительному выводу, что одна из основных психологических осей культуры человека определяется паразитом!
Много ли смысла в этой положительной корреляции? Возможно, что прямой причинно-следственной связи нет совсем — например, более переживающие народы (с высоким уровнем невротизма) привыкли снимать частый и долгий стресс путем общения с домашними животными, в том числе и с кошками. При этом повышенный уровень зараженности паразитом является просто следствием обилия кошек, а настоящая причина повышенного невротизма некоторых национальностей остается неясной (например, генетически обусловленный повышенный уровень синтеза дофамина в головном мозге).
Но не исключено, что на наше поведение большое влияние действительно оказывают паразиты. Ведь влияют же на нашу культуру окружающие нас растения и животные? Так что нам пора отдавать себе отчет в том, что доброй половиной наших негативных эмоций мы обязаны паразиту Toxoplasma gondii, рассчитывающему на то, что, пока мы будем злиться, притаившийся в ближайшем зоопарке тигр сможет нас съесть.
Источник:
K. D. Lafferty. Can the common brain parasite, Toxoplasma gondii, influence human culture? // Proceedings of the Royal Society B: Biological science (2006). DOI:10.1098/rspb.2006.3641 (FirstCite Early Online Publishing).
Константин Попадьин
См. также:
Cat Parasite May Affect Cultural Traits in Human Populations — пресс-релиз Калифорнийского университета в Санта-Барбаре.
Паразитические простейшие | АТСС
Делиться
ATCC предлагает множество паразитических простейших
Паразитические простейшие — это одноклеточные организмы, обладающие способностью проникать в ткани и клетки человека и других животных, вызывая ряд медицинских последствий, от неопасных до серьезных заболеваний. Эти микроорганизмы встречаются во многих средах обитания и могут передаваться фекально-оральным путем, половым путем или от членистоногих-переносчиков, таких как клещи и комары.
Эти микроорганизмы встречаются во многих средах обитания и могут передаваться фекально-оральным путем, половым путем или от членистоногих-переносчиков, таких как клещи и комары.
Обнаружение и лечение паразитических простейших может быть сложной задачей, что делает потребность в точной диагностике и эффективных противопаразитарных средствах еще более актуальной. Для поддержки исследований этих патогенов АТСС предоставляет более 1000 штаммов паразитических простейших, включая возбудителей малярии, токсоплазмоза, лямблиоза, лейшманиоза, бабезиоза и болезни Шагаса.
Откройте для себя нашу коллекцию паразитических простейших ниже, чтобы помочь вам в ваших исследованиях уже сегодня.
Посмотрите наш вебинар, чтобы узнать о переносимых клещами простейших
Рекомендуемые товары
Лямблии кишечные Улучшите свои кишечные исследования с помощью штамма, выделенного из тонкой кишки человека.
заказать сегодня
Вагинальная трихомонадаИспользуйте этих простейших паразитов, передающихся половым путем, для изучения устойчивости к метронидазолу, антиоксидантной защиты и метаболизма фосфолипидов.
заказать сейчас
Plasmodium falciparumИспользуйте эти лекарственно-устойчивые трансмиссивные простейшие, чтобы помочь вам в разработке тестов и методов лечения малярии. Этот штамм устойчив к хлорохину и чувствителен к пириметамину.
заказать сегодня
Уточнить по
Работает на Биоз Подробнее о Биозе
- Формат продукта:
- Произведено из:
- Диапазон спецификации:
- Концентрация:
- Масса:
- Класс антител:
- Источник:
- Иммуногенные виды:
- Обозначение штамма:
- Депонировано как:
- Тип штамма:
- Том:
- Условия хранения:
- Организм:
- Ткань:
- Болезнь:
- Тип ячейки:
- Тип клона:
- Информация о доставке:
- Организм:
- Информация о доставке:
Сравнивать
Вы не можете сравнить более 5 продуктов.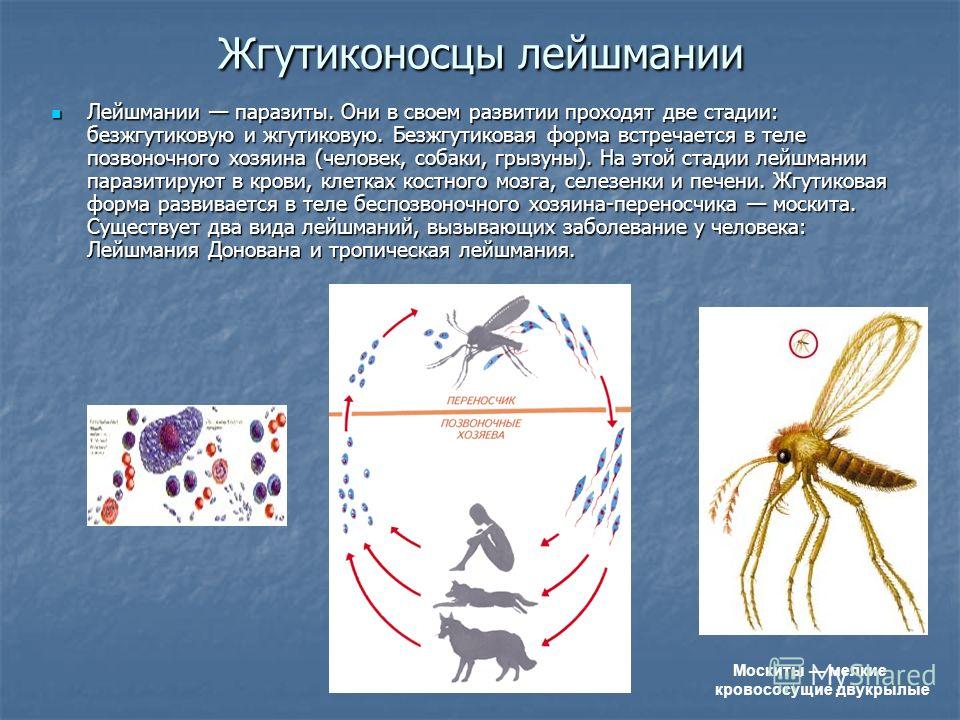
Этот лист продукта недоступен в Интернете. Мы предоставляем этот лист продукта только клиентам, которые приобрели этот продукт уровня биобезопасности 3. Если вы приобрели этот продукт, обратитесь в службу технической поддержки LGC для получения данного описания продукта.
Паспорт безопасности Скачать
Откройте паспорт безопасности для этого продукта, чтобы загрузить его.
Выберите язык Язык
ошибкаОтмена
Этот паспорт безопасности в настоящее время недоступен в Интернете. Нажмите кнопку ниже, чтобы получить дополнительную информацию о паспорте безопасности.
Проверьте информацию паспорта безопасности
сравнить продукты
Сравнить продукты ()
Продукт 1
Продукт 2
Продукт 3
Товар 4
Продукт 5
Анализ паразитических простейших на уровне отдельных клеток с использованием микрофлюидной импедансной цитометрии
Abstract
В настоящее время существует несколько технологий, позволяющих обнаруживать, идентифицировать и анализировать жизнеспособность простейших патогенов, включая Cryptosporidium и/или Giardia в уровень одиночной (оо)кисты.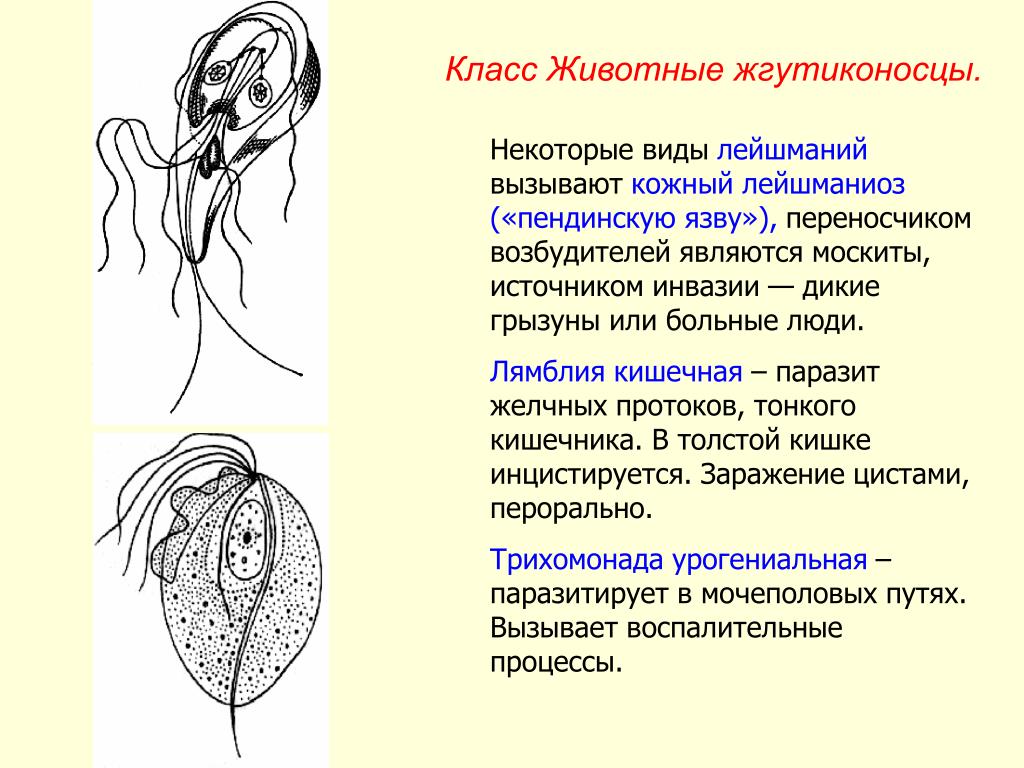 Мы сообщаем об использовании микрофлюидной импедансной цитометрии (MIC) для характеристики электрических (импедансных) свойств переменного тока отдельных паразитов и демонстрации быстрой дискриминации на основе жизнеспособности и вида. В частности, МИК использовали для идентификации живых и неактивных C. parvum ооцист с достоверностью более 90%, а также обнаружение поврежденных и/или эксцистированных ооцист. Кроме того, была достигнута дискриминация Cryptosporidium parvum , Cryptosporidium muris и Giardia lamblia с достоверностью более 92%. Подсчет и идентификация (оо)цист могут быть выполнены за несколько минут, что позволяет сократить время идентификации и трудозатраты по сравнению с существующими методами обнаружения.
Мы сообщаем об использовании микрофлюидной импедансной цитометрии (MIC) для характеристики электрических (импедансных) свойств переменного тока отдельных паразитов и демонстрации быстрой дискриминации на основе жизнеспособности и вида. В частности, МИК использовали для идентификации живых и неактивных C. parvum ооцист с достоверностью более 90%, а также обнаружение поврежденных и/или эксцистированных ооцист. Кроме того, была достигнута дискриминация Cryptosporidium parvum , Cryptosporidium muris и Giardia lamblia с достоверностью более 92%. Подсчет и идентификация (оо)цист могут быть выполнены за несколько минут, что позволяет сократить время идентификации и трудозатраты по сравнению с существующими методами обнаружения.
Введение
Простейшие патогены Cryptosporidium и Giardia ответственны за большое глобальное бремя болезней, которое влияет на здоровье как людей, так и домашнего скота и, в свою очередь, оказывает значительное влияние на экономику 1,2, 3 . В глобальном масштабе бремя заболевания человека Cryptosporidium оценивается в 250–500 миллионов случаев в год 1 . Вспышка криптоспоридиоза, передающегося через воду, в Милуоки в 1993 году заразила 400 000 человек и обошлась в 9 долларов.6 миллионов 3 , 4 , а вспышки произошли совсем недавно в Австралии 5 , Великобритании 6 и Швеции 7 . По оценкам, в Великобритании вспышка Cryptosporidium в Ланкашире, начавшаяся в августе 2015 г., за шесть недель затронула 300 000 домохозяйств, что обошлось прибл. 15 миллионов фунтов стерлингов 6 . При продолжающемся росте населения мира интенсификация сельского хозяйства и растущий спрос на питьевую воду, вероятно, приведут к увеличению заболеваемости людей и животных 1, 4 . Поэтому патогены, передающиеся через пищу и воду, вызывают растущую озабоченность правительств и промышленности во всем мире.
В глобальном масштабе бремя заболевания человека Cryptosporidium оценивается в 250–500 миллионов случаев в год 1 . Вспышка криптоспоридиоза, передающегося через воду, в Милуоки в 1993 году заразила 400 000 человек и обошлась в 9 долларов.6 миллионов 3 , 4 , а вспышки произошли совсем недавно в Австралии 5 , Великобритании 6 и Швеции 7 . По оценкам, в Великобритании вспышка Cryptosporidium в Ланкашире, начавшаяся в августе 2015 г., за шесть недель затронула 300 000 домохозяйств, что обошлось прибл. 15 миллионов фунтов стерлингов 6 . При продолжающемся росте населения мира интенсификация сельского хозяйства и растущий спрос на питьевую воду, вероятно, приведут к увеличению заболеваемости людей и животных 1, 4 . Поэтому патогены, передающиеся через пищу и воду, вызывают растущую озабоченность правительств и промышленности во всем мире.
Ооцисты Cryptosporidium образуются в больших количествах, высокоинфекционны, устойчивы в воде, устойчивы к хлорированию, деформируются и способны ускользать от современных систем фильтрации в небольшом количестве 2, 4, 8,9,10,11,12 . При наличии менее 10 ооцист, способных вызвать серьезную инфекцию человека 13, 14 , поэтому необходим мониторинг наличия этого патогена в воде 15, 16 . Стандартный метод выделения и обнаружения ооцист Cryptosporidium , присутствующих в источниках очищенной воды, EPA 1623.1 17 , обычно включает обработку 50–100 л воды (, например, , на месте) или 1000 л (например, ). ., на очистных сооружениях) до объема 50 мкл, прежде чем высококвалифицированные микроскописты подтвердят обнаружение с помощью специализированных методов 18 . Этот процесс также одновременно извлекает цисты Giardia 9.род 0158 – простейшие паразиты, которые также могут вызывать гастроэнтерит при проглатывании человеком 17 .
При наличии менее 10 ооцист, способных вызвать серьезную инфекцию человека 13, 14 , поэтому необходим мониторинг наличия этого патогена в воде 15, 16 . Стандартный метод выделения и обнаружения ооцист Cryptosporidium , присутствующих в источниках очищенной воды, EPA 1623.1 17 , обычно включает обработку 50–100 л воды (, например, , на месте) или 1000 л (например, ). ., на очистных сооружениях) до объема 50 мкл, прежде чем высококвалифицированные микроскописты подтвердят обнаружение с помощью специализированных методов 18 . Этот процесс также одновременно извлекает цисты Giardia 9.род 0158 – простейшие паразиты, которые также могут вызывать гастроэнтерит при проглатывании человеком 17 .
Возможность быстрого подсчета и различения ооцист Cryptosporidium ( например, ., после восстановления через EPA 1623.1) на основе статуса жизнеспособности и/или видов в автоматизированном процессе сократит время обнаружения, снизит уровень человеческого вмешательства, помогают лучше оценить риск для здоровья человека и способствуют экономии ресурсов 19 .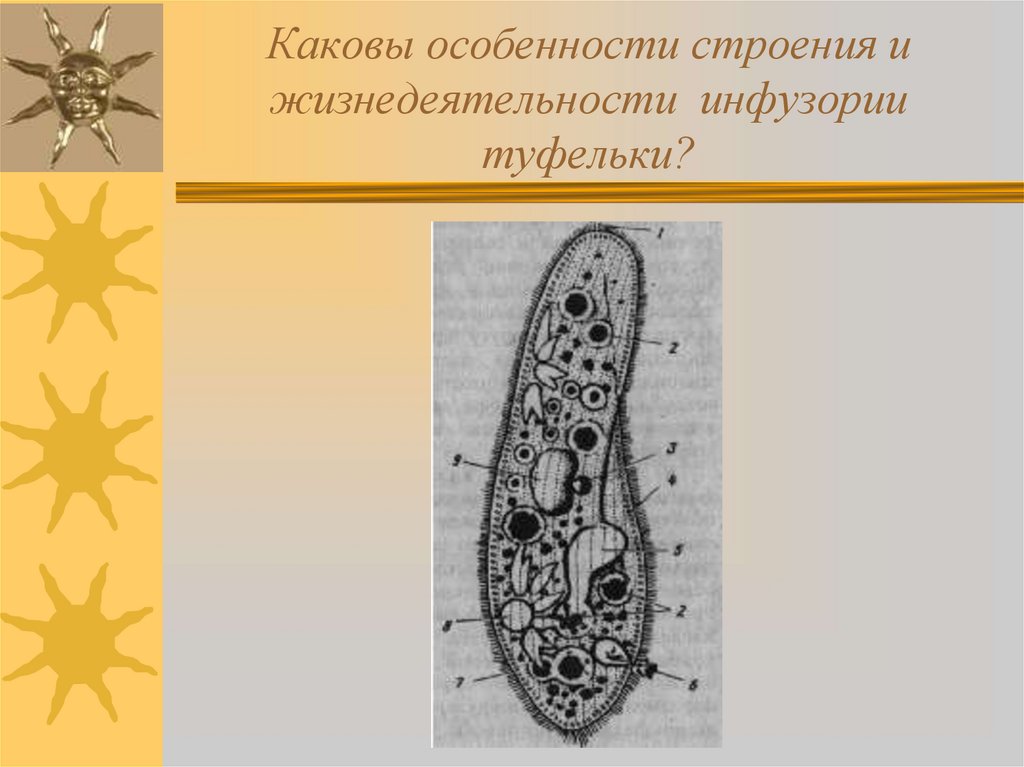 Чтобы различать живые ооцисты и ооцисты, инактивированные температурой, собственные электрические свойства отдельных C. parvum были измерены с использованием электрокинетических методов переменного тока, таких как диэлектрофорез 20, 21 и/или электровращение 22, 23 . Измерения проводились либо в партии 20, 22, 23 , требовалось длительное время обработки 21 , либо требовалась микроскопия для оценки реакции ооцист на прикладное поле 20,21,22,23 . Электрокинетические методы переменного тока использовались для сортировки на уровне рода, напр. Криптоспоридии и Giardia . Унни и др. . 24 продемонстрировал диэлектрофоретическое разделение Cryptosporidium muris и Giardia lamblia на чипе в пакетном режиме, но не указал предел обнаружения. Однако, насколько нам известно, такие подходы не использовались для различения альтернативных видов Cryptosporidium , и было высказано предположение, что разделение C.
Чтобы различать живые ооцисты и ооцисты, инактивированные температурой, собственные электрические свойства отдельных C. parvum были измерены с использованием электрокинетических методов переменного тока, таких как диэлектрофорез 20, 21 и/или электровращение 22, 23 . Измерения проводились либо в партии 20, 22, 23 , требовалось длительное время обработки 21 , либо требовалась микроскопия для оценки реакции ооцист на прикладное поле 20,21,22,23 . Электрокинетические методы переменного тока использовались для сортировки на уровне рода, напр. Криптоспоридии и Giardia . Унни и др. . 24 продемонстрировал диэлектрофоретическое разделение Cryptosporidium muris и Giardia lamblia на чипе в пакетном режиме, но не указал предел обнаружения. Однако, насколько нам известно, такие подходы не использовались для различения альтернативных видов Cryptosporidium , и было высказано предположение, что разделение C. parvum и C. muris может оказаться невозможным, так как оба демонстрируют лишь незначительную разницу в диэлектрических свойствах 24 .
parvum и C. muris может оказаться невозможным, так как оба демонстрируют лишь незначительную разницу в диэлектрических свойствах 24 .
Единственное использование системы анализа электрического импеданса для характеристики электрических свойств жизнеспособных и нежизнеспособных ооцист C. parvum было продемонстрировано Houssin et al . 25 . Результаты показали 15-процентную разницу в величине импеданса с буфером с низкой проводимостью, содержащим либо жизнеспособные, либо нежизнеспособные C. parvum , с заявленным пределом обнаружения <10 ооцист мкл −1 . Измерения проводились в массе, когда аликвоты, содержащие ооцисты, как жизнеспособные, так и нежизнеспособные, загружались в лунки для образцов, при этом измерялся импеданс и сравнивался с контролем (предварительно измеренные значения только суспендирующей жидкости). Однако ооцисты в любом реальном образце, например, ., полученные с помощью EPA 1623.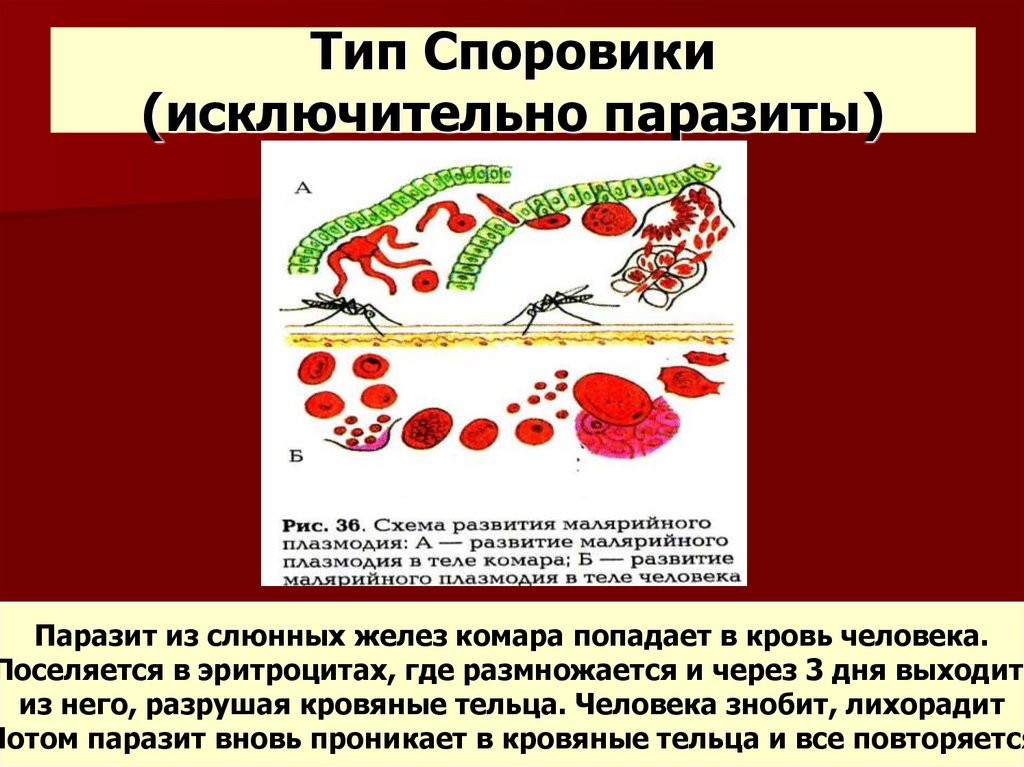 1, могут иметь смешанную жизнеспособность; следовательно, необходимо, чтобы любая импедансная система, используемая для обнаружения, могла определять статус жизнеспособности отдельных ооцист. Насколько нам известно, нет отчетов об анализе импеданса различных видов Cryptosporidium или одновременный импедансный анализ видов Cryptosporidium и Giardia .
1, могут иметь смешанную жизнеспособность; следовательно, необходимо, чтобы любая импедансная система, используемая для обнаружения, могла определять статус жизнеспособности отдельных ооцист. Насколько нам известно, нет отчетов об анализе импеданса различных видов Cryptosporidium или одновременный импедансный анализ видов Cryptosporidium и Giardia .
Микрофлюидная импедансная цитометрия (MIC) представляет собой микрофлюидную систему, в которой используются микроэлектроды для измерения импеданса отдельных частиц 26, 27 . Взвешенные частицы проходят через микроканал с высокой скоростью (можно использовать скорость потока до ~80 мкл мин -1), и когда они проходят через область обнаружения, измеренный ток дает сигнал, пропорциональный импедансу частиц. Этот метод не требует маркировки, требует минимальной обработки образца и измеряет до 1000 частиц в секунду 28 . По сравнению с другими технологиями непрерывного микрожидкостного разделения/дискриминации 29, 30 , MIC обеспечивает быструю обработку проб и высокую производительность. МИК использовали для различения различных типов клеток, включая стволовые клетки 31 , лейкоциты 32 , 33 и опухолевые клетки в цельной крови 34 . Однако применимость MIC для обнаружения паразитических простейших еще не была широко изучена, и предыдущая работа была сосредоточена на анализе инфицированных клеток, а не на прямом анализе простейших 35, 36 .
МИК использовали для различения различных типов клеток, включая стволовые клетки 31 , лейкоциты 32 , 33 и опухолевые клетки в цельной крови 34 . Однако применимость MIC для обнаружения паразитических простейших еще не была широко изучена, и предыдущая работа была сосредоточена на анализе инфицированных клеток, а не на прямом анализе простейших 35, 36 .
В этой статье мы описываем использование MIC для обнаружения отдельных простейших (оо)кист в системе с непрерывным потоком. Мы характеризуем биофизические свойства живых и инактивированных температурой патогенных для человека паразитов и оцениваем состав образцов путем анализа электрических данных. Также обнаруживаются единичные ооцисты C. parvum и C. muris , а также цисты G. lamblia . Обнаружение и анализ жизнеспособности/видов ооцист в непрерывном потоке с использованием MIC предлагает значительный прогресс по сравнению с другими подходами как с точки зрения сбора информации на уровне одной (оо)цисты, так и с точки зрения скорости сбора данных.
Материалы и методы.
Обзор системы. Короче говоря, платиновые электроды (шириной 30 мкм и толщиной 200 нм) были нанесены на стеклянную пластину. Микроканалы с размерами поперечного сечения 200 мкм × 30 мкм, ведущие в зону обнаружения 40 мкм × 30 мкм, были определены в фоторезисте SU8 перед термическим соединением двух пластин. После нарезки впускной и выпускной каналы были протравлены с помощью CO
2 лазер (Mini 18, Epilog Laser). Чипы удерживались в держателе, напечатанном на 3D-принтере, в котором размещались гидравлические насадки и электрические соединения. Держатель был установлен на предметном столике размером x — y — z с оптикой для изображения области обнаружения. Шприцевой насос (Fusion 400, Chemyx) использовали для введения образца из шприца со скоростью потока 40 мкл мин -1 . На рис. 1 показана схема чипа импеданса. К двум верхним электродам прикладывали синусоидальные напряжения на двух одновременных частотах с помощью цифрового анализатора импеданса (h3FLI, Zurich Instruments).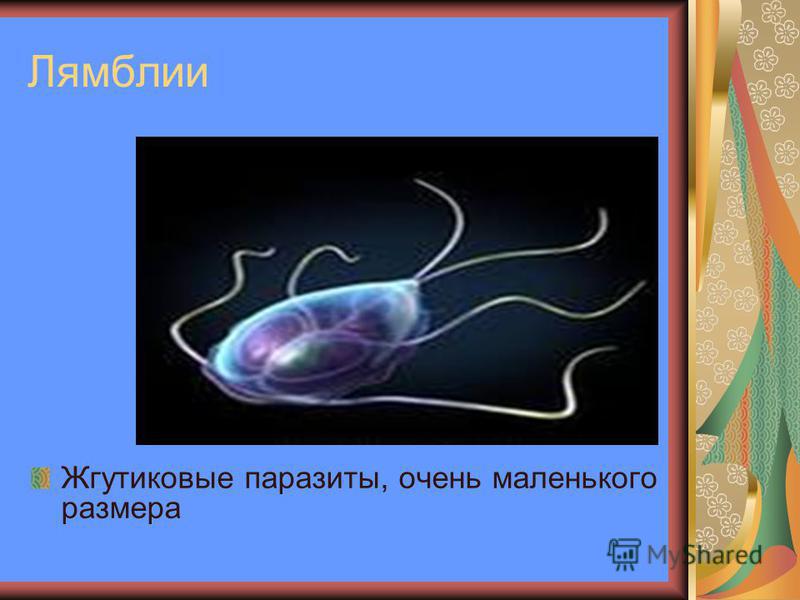 Первая частота, называемая опорной частотой, применялась на уровне 18,3 МГц, а вторая (зондовая) частота варьировалась в диапазоне от 250 кГц до 50 МГц (рис. 1). Напряжение варьировалось от 1,5 В до 10 В в зависимости от проводимости буфера и частоты сигнала. На частотах выше 20 МГц сигналы ослабляются входным и выходным фильтрами синхронного усилителя, достигая -6 дБ на частоте 50 МГц. Поэтому для частот ≥20 МГц приложенное напряжение увеличивалось постепенно, чтобы учесть это затухание. Сначала была рассчитана средняя величина сигнала для трех ближайших зондирующих частот ниже 20 МГц\(\,({\bar{Z}}_{<20MHz})\). Затем коэффициент усиления рассчитывался путем деления этого среднего значения на сигнал средней амплитуды для каждой частоты (например, {Z}}_{50MHZ}\) для 50 MHz). Пример можно найти на дополнительном рисунке S1. Повышение напряжения минимизировало потери сигнала, не влияя на сбор или анализ данных. При любой частоте зондирования данные записывались в течение примерно 30 с, прежде чем частота зондирования была увеличена.
Первая частота, называемая опорной частотой, применялась на уровне 18,3 МГц, а вторая (зондовая) частота варьировалась в диапазоне от 250 кГц до 50 МГц (рис. 1). Напряжение варьировалось от 1,5 В до 10 В в зависимости от проводимости буфера и частоты сигнала. На частотах выше 20 МГц сигналы ослабляются входным и выходным фильтрами синхронного усилителя, достигая -6 дБ на частоте 50 МГц. Поэтому для частот ≥20 МГц приложенное напряжение увеличивалось постепенно, чтобы учесть это затухание. Сначала была рассчитана средняя величина сигнала для трех ближайших зондирующих частот ниже 20 МГц\(\,({\bar{Z}}_{<20MHz})\). Затем коэффициент усиления рассчитывался путем деления этого среднего значения на сигнал средней амплитуды для каждой частоты (например, {Z}}_{50MHZ}\) для 50 MHz). Пример можно найти на дополнительном рисунке S1. Повышение напряжения минимизировало потери сигнала, не влияя на сбор или анализ данных. При любой частоте зондирования данные записывались в течение примерно 30 с, прежде чем частота зондирования была увеличена. Ток, протекающий через оба нижних электрода, преобразовывался в напряжение с помощью трансимпедансных усилителей. Дифференциальный усилитель использовался для получения одиночного выходного сигнала. Синхронное усиление (HF2LI, Zurich Instruments) использовалось для разделения амплитуды и фазы на каждой частоте, на основе которых были построены графики рассеяния импеданса.
Ток, протекающий через оба нижних электрода, преобразовывался в напряжение с помощью трансимпедансных усилителей. Дифференциальный усилитель использовался для получения одиночного выходного сигнала. Синхронное усиление (HF2LI, Zurich Instruments) использовалось для разделения амплитуды и фазы на каждой частоте, на основе которых были построены графики рассеяния импеданса.
Схематическое изображение микрофлюидного импедансного цитометра. (a) Область обнаружения состоит из пар параллельно расположенных электродов шириной 30 мкм, разделенных расстоянием 50 мкм. Электроды изготавливаются внутри микрожидкостного канала шириной 40 мкм и высотой 30 мкм. Патогены, взвешенные в PBS (показан C. parvum ), перемещаются по каналу потоком под давлением. Синусоидальные напряжения на двух дискретных частотах (ƒ 1 , ƒ 2 ) подаются на верхние электроды, а разница в токе, протекающем через нижние электроды, измеряется специальной схемой обнаружения.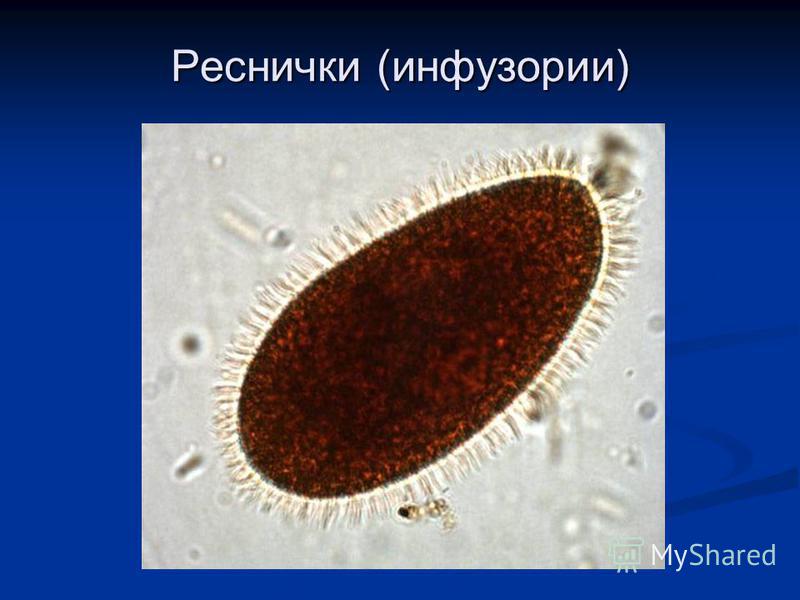 Схема состоит из трансимпедансных усилителей, преобразующих ток (I) в напряжение (V), и дифференциального усилителя. Отклик на каждой применяемой частоте демодулируется с использованием синхронного усиления (HF2LI, Zurich Instruments). ( b ) Идеализированный дифференциальный сигнал, когда ооциста проходит через центр области обнаружения; шаги последовательности аннотируются (t 0 , т 1 , т 2 , т 3 и т 4 ).
Схема состоит из трансимпедансных усилителей, преобразующих ток (I) в напряжение (V), и дифференциального усилителя. Отклик на каждой применяемой частоте демодулируется с использованием синхронного усиления (HF2LI, Zurich Instruments). ( b ) Идеализированный дифференциальный сигнал, когда ооциста проходит через центр области обнаружения; шаги последовательности аннотируются (t 0 , т 1 , т 2 , т 3 и т 4 ).
Изображение в полный размер
Между образцами чипы промывали 1 мл этанола (70%) и 1 мл деионизированной воды с помощью шприца. Поверхности электродов ежедневно очищали от мусора путем введения в систему 1 М раствора гидроксида натрия в количестве 50 мкл·мин 90 177 -1 90 178 в течение ~ 4 минут перед промывкой 1 мл деионизированной воды.
Подготовка проб
В нашем анализе необработанных и термоинактивированных 9 образцов использовались четыре различных исходных образца.0157 С. парвум .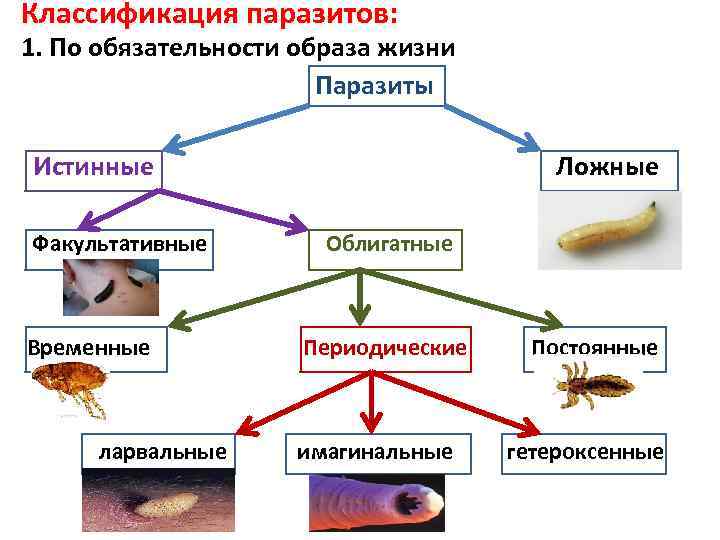 Эти образцы, все полученные в суспензиях, содержащих PBS + 2% антибиотиков + 0,01% Tween 20, были следующими:
Эти образцы, все полученные в суспензиях, содержащих PBS + 2% антибиотиков + 0,01% Tween 20, были следующими:
Партию C. parvum (WC1) от телят, полученную от Waterborne Inc. (штамм Айова; США), использовали в течение одного месяца после размножения.
Вторая партия C. parvum (WC2) от телят и партия 9 от мышей0157 C. parvum (WM) от Waterborne Inc. (штамм Айова) использовали в течение 1–2 месяцев после размножения ооцист.
C. parvum (MC), полученный от телят, полученный для Moredun Scientific (Moredun Strain; Великобритания), использовали в возрасте примерно 2–3 месяцев.
Из каждой партии были независимо приготовлены образцы, содержащие необработанные или инактивированные нагреванием C. parvum , и импеданс был измерен отдельно. В каждом образце ооцисты разводили до конечной концентрации ~100 ооцист мкл −1 ПБС. Влияние различной проводимости буфера исследовали с использованием различных концентраций PBS в диапазоне от 0,76 до 7,10 См м -1 . Каждый буфер также содержал 0,1% Tween 20. Калибровочные полистироловые шарики диаметром 7 мкм (Microparticles GmbH) добавляли к каждому образцу до конечной концентрации 100 шариков мкл -1 . Шарики из полистирола имеют известный размер и электрические свойства 34 , а также имеют постоянный импеданс во всем диапазоне частот, используемом в этих экспериментах. Измеренный сигнал импеданса каждой оо(цисты) нормализовали по отношению к шарику путем деления на средний импеданс полистироловых шариков для каждой частоты, таким образом удаляя любые нелинейности.
В каждом образце ооцисты разводили до конечной концентрации ~100 ооцист мкл −1 ПБС. Влияние различной проводимости буфера исследовали с использованием различных концентраций PBS в диапазоне от 0,76 до 7,10 См м -1 . Каждый буфер также содержал 0,1% Tween 20. Калибровочные полистироловые шарики диаметром 7 мкм (Microparticles GmbH) добавляли к каждому образцу до конечной концентрации 100 шариков мкл -1 . Шарики из полистирола имеют известный размер и электрические свойства 34 , а также имеют постоянный импеданс во всем диапазоне частот, используемом в этих экспериментах. Измеренный сигнал импеданса каждой оо(цисты) нормализовали по отношению к шарику путем деления на средний импеданс полистироловых шариков для каждой частоты, таким образом удаляя любые нелинейности.
Для анализа импеданса живых Cryptosporidium spp . и Г . lamblia , отдельные образцы, содержащие C . парвум , С .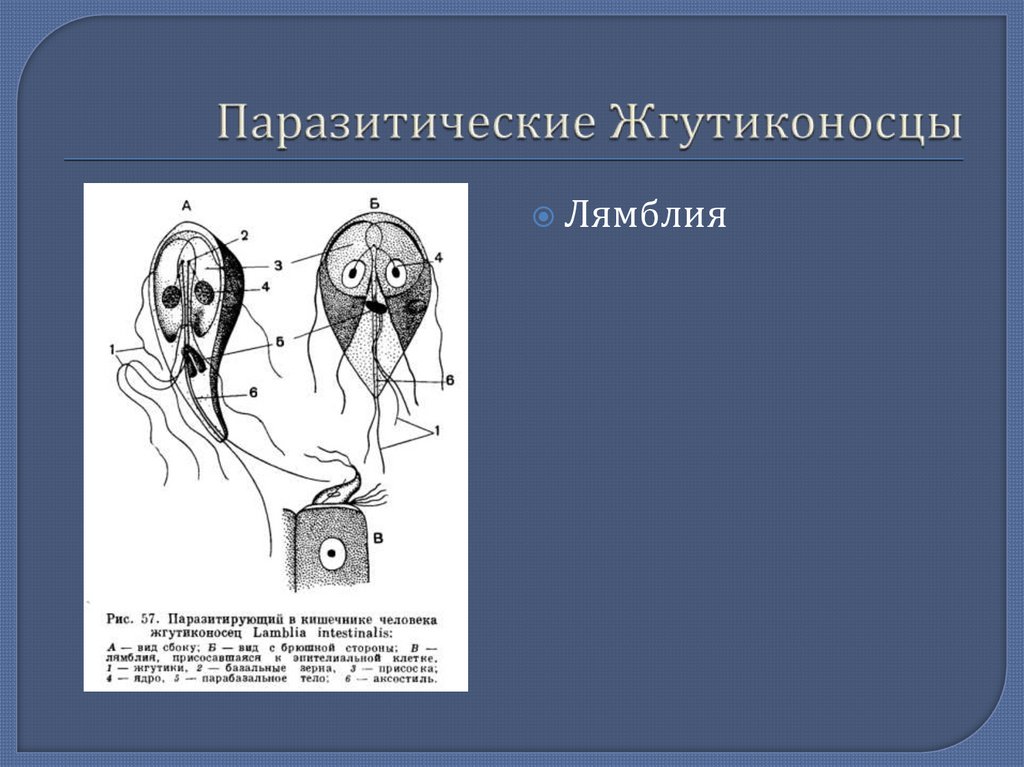 muris или G. lamblia (полученные от Waterborne Inc.) готовили в PBS + 0,1% Tween 20 (σ m = 1,6 S m -1 ). Смешанные образцы разбавляли, чтобы они содержали в общей сложности ~100 (оо) цист и 100 гранул мкл -1 образца; были подготовлены смешанные образцы, включающие равное количество каждого патогена. Все образцы вводили в систему и измеряли, как описано ранее.
muris или G. lamblia (полученные от Waterborne Inc.) готовили в PBS + 0,1% Tween 20 (σ m = 1,6 S m -1 ). Смешанные образцы разбавляли, чтобы они содержали в общей сложности ~100 (оо) цист и 100 гранул мкл -1 образца; были подготовлены смешанные образцы, включающие равное количество каждого патогена. Все образцы вводили в систему и измеряли, как описано ранее.
Инактивация ооцист
Для инактивации ооцист образцы, приготовленные, как описано для необработанного C. parvum в предыдущем разделе, подвергали термообработке при 70 °C в течение 5 минут в нагревательном блоке (Stuart Scientific, Великобритания). перед измерением. Это лечение описано в литературе как минимальное лечение, необходимое для индукции потери инфекционности у мышиной модели 20, 38 .
Анализ данных
На языке Matlab (R2016a) был написан специальный скрипт для обработки данных и статистического анализа. Для того чтобы сделать надежный статистический вывод относительно каждой популяции, образцы, содержащие каждую популяцию, сначала измеряли отдельно.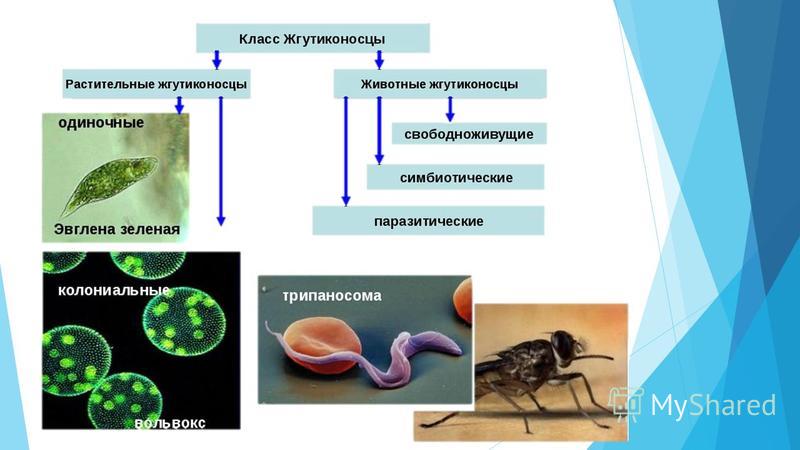 Используя сравнение необработанных и термоинактивированных C. parvum , например, сигнал импеданса сначала нормализовали относительно частотно-независимой характеристики импеданса полистироловых шариков. Затем популяции ооцист C. parvum были отобраны из более мелких остатков и более крупных эталонных шариков с использованием нормализованных данных импеданса, собранных на эталонной частоте (18,3 МГц). После стробирования наносили на график и анализировали нормализованную реакцию импеданса гейтированных патогенов на частоте зонда.
Используя сравнение необработанных и термоинактивированных C. parvum , например, сигнал импеданса сначала нормализовали относительно частотно-независимой характеристики импеданса полистироловых шариков. Затем популяции ооцист C. parvum были отобраны из более мелких остатков и более крупных эталонных шариков с использованием нормализованных данных импеданса, собранных на эталонной частоте (18,3 МГц). После стробирования наносили на график и анализировали нормализованную реакцию импеданса гейтированных патогенов на частоте зонда.
Принимая во внимание разброс и ориентацию данных и допуская нормальное распределение данных, можно было оценить уровень дискриминации, достигнутый по обоим измерениям импеданса (действительному и мнимому или непрозрачности и размеру) на заданной частоте между любыми двумя населения. Во-первых, была рассчитана ковариационная матрица, чтобы определить направления, в которых двумерные данные одной совокупности варьировались больше всего — наибольший и наименьший собственные векторы этой матрицы указывают направление разброса данных, а величина этого разброса определялась собственные значения матрицы.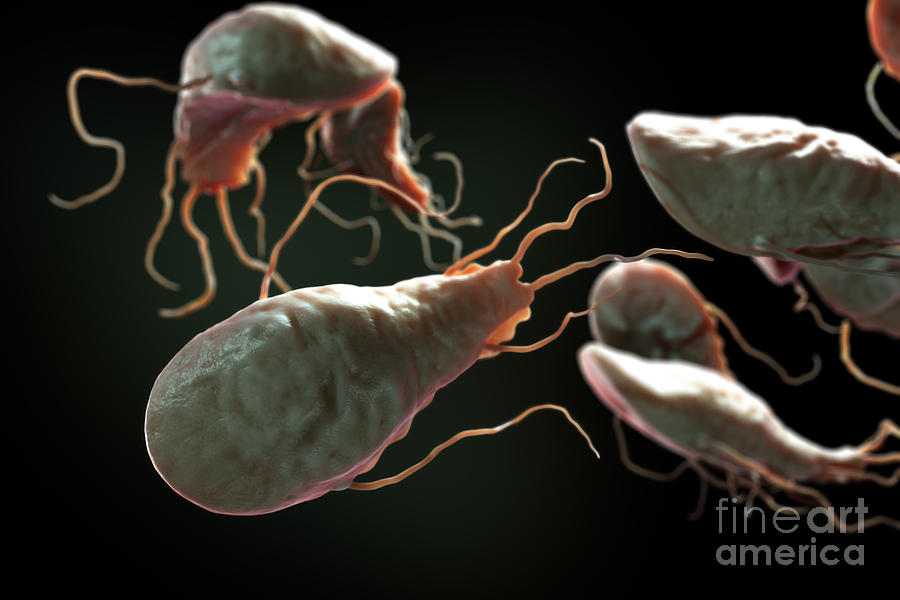 Во-вторых, в отношении распространения и ориентации каждой популяции были построены доверительные эллипсы, содержащие все события в пределах 1–3 стандартных отклонений от среднего, чтобы определить позиции равновероятностного отклонения между двумя кластерами, т. е. пересечение эквивалентных эллипсов достоверности определяет границу, на которой точка данных имеет равную вероятность принадлежать к любой совокупности. Затем к точкам пересечения подгоняли линию равновероятностного отклонения, а затем указывали достоверность различения – , т.е. ., путем подсчета количества ооцист из каждой популяции, нанесенной по обе стороны от линии равновероятностного отклонения.
Во-вторых, в отношении распространения и ориентации каждой популяции были построены доверительные эллипсы, содержащие все события в пределах 1–3 стандартных отклонений от среднего, чтобы определить позиции равновероятностного отклонения между двумя кластерами, т. е. пересечение эквивалентных эллипсов достоверности определяет границу, на которой точка данных имеет равную вероятность принадлежать к любой совокупности. Затем к точкам пересечения подгоняли линию равновероятностного отклонения, а затем указывали достоверность различения – , т.е. ., путем подсчета количества ооцист из каждой популяции, нанесенной по обе стороны от линии равновероятностного отклонения.
Анализ эксцистации
Показатели жизнеспособности живых 9Образцы 0157 C. parvum были оценены с использованием анализа эксцистации in vitro , разработанного в Исследовательском институте Моредуна (Великобритания) 39 , для оценки качества исходного образца C. parvum и предоставления эталона для сравнения необработанных и инактивированных нагреванием C. парвум . По завершении анализа ооцисты визуализировали с использованием дифференциально-интерференционно-контрастного (ДИК) микроскопа (BX50, Olympus) и подсчитывали количество интактных ооцист и пустых оболочек, пока не было зарегистрировано 250 событий. Для каждого образца проводили три подсчета. Для расчета процента эксцистирования использовали следующую формулу: количество пустых оболочек/(количество пустых оболочек + количество неповрежденных ооцист). Средние показатели жизнеспособности выборки для исходных 9Образцы запасов 0157 C. parvum : 95% для WC1, 86% для WC2; 87% для ВМ; и 88% для МС. См. Таблицу S1 для получения более подробных результатов анализа эксцистации.
парвум . По завершении анализа ооцисты визуализировали с использованием дифференциально-интерференционно-контрастного (ДИК) микроскопа (BX50, Olympus) и подсчитывали количество интактных ооцист и пустых оболочек, пока не было зарегистрировано 250 событий. Для каждого образца проводили три подсчета. Для расчета процента эксцистирования использовали следующую формулу: количество пустых оболочек/(количество пустых оболочек + количество неповрежденных ооцист). Средние показатели жизнеспособности выборки для исходных 9Образцы запасов 0157 C. parvum : 95% для WC1, 86% для WC2; 87% для ВМ; и 88% для МС. См. Таблицу S1 для получения более подробных результатов анализа эксцистации.
Оптическая проточная цитометрия
Обычный проточный цитометрический анализ суспензий паразитов проводили при 35 мкл мин -1 с использованием проточного цитометра BD Accuri C6 (BD Biosciences) с двумя лазерами (488 нм и 640 нм). Данные были экспортированы в виде стандартных файлов FCS, а сигналы прямого и бокового рассеяния света (FSC и SSC) были проанализированы с использованием Matlab.
Доступность данных
Все данные, подтверждающие это исследование, находятся в открытом доступе в репозитории Саутгемптонского университета по адресу http://doi.org/10.5258/SOTON/D0047.
Результаты и обсуждение
Диэлектрические свойства
Простейшей электрической моделью, описывающей свойства ячейки, является так называемая модель с одной оболочкой, как показано на рис. 2(а). Как правило, биологические клетки являются изолирующими на низких частотах (кГц) из-за наличия липидной клеточной мембраны, но становятся все более проводящими на более высоких частотах (МГц) из-за емкостной связи через мембрану 25, 34, 40 . Таким образом, для жизнеспособной клетки импеданс на низких частотах (<1 МГц) измеряет объем клетки, а на средних частотах (0,5–10 МГц) измеряет емкость мембраны, которая отражает инвагинацию и свертка мембраны. На более высоких частотах электрическое поле емкостно связано с мембраной, а сигнал импеданса отражает свойства цитоплазмы и любого ядра, которое может присутствовать. На рисунке 2(b) показан конкретный случай ооцисты, содержащей четыре спорозоита, содержащих ядра, и различные органеллы. По аналогии с типичной клеткой на низких частотах можно измерить только объем интактных ооцист. В режиме МГц импеданс ооцисты сначала становится функцией емкости внешней стенки (обычно на частотах 1–10 МГц), прежде чем проводимость внутренней части сильно повлияет на отклик на высоких частотах (> 10 МГц) 27 . Следовательно, в промежуточном диапазоне частот жизнеспособные ооцисты эквивалентного размера могут демонстрировать различия в своем импедансе из-за различий в составе и/или структуре внешней и внутренней стенки. По мере дальнейшего увеличения частоты сигнала, т.е. ., >30 МГц, на импеданс дополнительно влияют спорозоиты и ядра 27 .
На рисунке 2(b) показан конкретный случай ооцисты, содержащей четыре спорозоита, содержащих ядра, и различные органеллы. По аналогии с типичной клеткой на низких частотах можно измерить только объем интактных ооцист. В режиме МГц импеданс ооцисты сначала становится функцией емкости внешней стенки (обычно на частотах 1–10 МГц), прежде чем проводимость внутренней части сильно повлияет на отклик на высоких частотах (> 10 МГц) 27 . Следовательно, в промежуточном диапазоне частот жизнеспособные ооцисты эквивалентного размера могут демонстрировать различия в своем импедансе из-за различий в составе и/или структуре внешней и внутренней стенки. По мере дальнейшего увеличения частоты сигнала, т.е. ., >30 МГц, на импеданс дополнительно влияют спорозоиты и ядра 27 .
Иллюстрация частотно-зависимого диэлектрического отклика однослойной частицы или C. parvum ооцист. ( a ) Упрощенная модель с одной оболочкой часто используется для аппроксимации диэлектрических свойств клеток/частиц. Клетка/частица, окруженная оболочкой (мембраной), испытывает однократную диэлектрическую дисперсию в диапазоне частот 1–10 МГц (называемую β-релаксацией). ( b ) Для более сложных биологических структур, таких как ооциста C. parvum , упрощенная модель с одной оболочкой не может точно рассчитать диэлектрические свойства компонентов ооцисты. Например, 9Ооцисты 0157 C. parvum , вероятно, испытывают множественные релаксации из-за поляризации последовательных оболочек по мере увеличения частоты. Отметим, что частотное окно для этих релаксаций зависит от проводимости взвешенной среды.
Клетка/частица, окруженная оболочкой (мембраной), испытывает однократную диэлектрическую дисперсию в диапазоне частот 1–10 МГц (называемую β-релаксацией). ( b ) Для более сложных биологических структур, таких как ооциста C. parvum , упрощенная модель с одной оболочкой не может точно рассчитать диэлектрические свойства компонентов ооцисты. Например, 9Ооцисты 0157 C. parvum , вероятно, испытывают множественные релаксации из-за поляризации последовательных оболочек по мере увеличения частоты. Отметим, что частотное окно для этих релаксаций зависит от проводимости взвешенной среды.
Изображение с полным размером
Разрушение избирательно проницаемой мембраны биологической клетки обычно измеряют с помощью красителя жизнеспособности, такого как йодистый пропидий (PI). При попадании в клетки с нарушенными мембранами PI связывается с ДНК, интеркалируя между основаниями, что значительно увеличивает флуоресценцию. Если скомпрометированная клеточная мембрана проницаема для PI, ионы смогут свободно проходить через нее (9). 0157 т. е. ., ячейка электрически негерметична, рис. 3), что проявляется в изменении импеданса на низких частотах 34 . Следовательно, нежизнеспособные клетки могут казаться меньше в электрическом, но не в оптическом объеме. Нежизнеспособные клетки становятся неспособными регулировать перенос ионов и поддерживать осмотическое давление так же эффективно, как жизнеспособные клетки 34, 41 , как, например, в случае ооцисты C. parvum , что схематично показано на рис. 3a. Способствующим фактором способности жизнеспособных ооцист сохраняться в окружающей среде в течение длительного времени является их строгая регуляция переноса ионов. После инактивации ооцисты больше не регулируют перенос ионов так же эффективно, как жизнеспособные ооцисты. Таким образом, внутренний состав инактивированной ооцисты может быть нарушен. Например, микроскопические изображения необработанного (рис. 3b) и термоинактивированного (рис. 3c) C. parvum показывают, что внутренние компоненты кажутся менее гранулированными в инактивированных нагреванием ооцистах, что может указывать на изменение их внутреннего состава.
0157 т. е. ., ячейка электрически негерметична, рис. 3), что проявляется в изменении импеданса на низких частотах 34 . Следовательно, нежизнеспособные клетки могут казаться меньше в электрическом, но не в оптическом объеме. Нежизнеспособные клетки становятся неспособными регулировать перенос ионов и поддерживать осмотическое давление так же эффективно, как жизнеспособные клетки 34, 41 , как, например, в случае ооцисты C. parvum , что схематично показано на рис. 3a. Способствующим фактором способности жизнеспособных ооцист сохраняться в окружающей среде в течение длительного времени является их строгая регуляция переноса ионов. После инактивации ооцисты больше не регулируют перенос ионов так же эффективно, как жизнеспособные ооцисты. Таким образом, внутренний состав инактивированной ооцисты может быть нарушен. Например, микроскопические изображения необработанного (рис. 3b) и термоинактивированного (рис. 3c) C. parvum показывают, что внутренние компоненты кажутся менее гранулированными в инактивированных нагреванием ооцистах, что может указывать на изменение их внутреннего состава. Следовательно, суспендируя ооцисты в неизоосмотических условиях, можно вызвать изменение ионного состава ооцисты и, следовательно, изменить ее промежуточный и/или высокочастотный импеданс.
Следовательно, суспендируя ооцисты в неизоосмотических условиях, можно вызвать изменение ионного состава ооцисты и, следовательно, изменить ее промежуточный и/или высокочастотный импеданс.
Диаграмма, показывающая структуру как необработанных (живых), так и термоинактивированных (нежизнеспособных) C. parvum ооцист. ( a ) Схема, показывающая гипотетический эффект тепловой инактивации на ооцисты, взвешенные в PBS. Ооциста C. parvum обычно состоит из трехслойной внешней стенки, которая содержит четыре голых спорозоита и окруженное мембраной остаточное тело. При инактивации на направление переноса ионов влияет проводимость/осмоляльность буфера. ( b ) Микроскопические изображения необработанных и ( c ) термоинактивированных ооцист (в PBS), полученные с помощью масляной иммерсии (100x) ДИК-микроскопии.
Изображение полного размера
Импедансный анализ паразитов
На рис. 4 показаны экспериментальные данные импеданса для четырех различных необработанных популяций C. parvum , взвешенных в 0,5 × PBS (σ м =7 10 м 7 1 0,76 9 S 90,76 опорная частота 18,3 МГц. Была выбрана высокая эталонная частота, чтобы можно было исследовать внутренние свойства ооцист 31, 34 . Величина импеданса (|Z | ) отложена на оси x , а фаза импеданса (ΦZ) на y — ось, где каждая точка данных представляет одну ооцисту, а ее цвет отражает плотность данных. Рис. 4 Каждая точка данных представляет собой одну ооцисту, а цвет представляет плотность данных. Первые 3000 обнаруженных событий нанесены на график в каждом подмножестве рисунка, а популяции ооцист и раковин аннотированы в соответствии с анализом, описанным в разделе моделирования. ( a ) Ооцисты телят (штамм Айова), измеренные в течение 1 месяца после размножения ооцист. ( b ) Ооцисты, размножаемые мышами (штамм Айова), используются в течение 1–2 месяцев.
4 показаны экспериментальные данные импеданса для четырех различных необработанных популяций C. parvum , взвешенных в 0,5 × PBS (σ м =7 10 м 7 1 0,76 9 S 90,76 опорная частота 18,3 МГц. Была выбрана высокая эталонная частота, чтобы можно было исследовать внутренние свойства ооцист 31, 34 . Величина импеданса (|Z | ) отложена на оси x , а фаза импеданса (ΦZ) на y — ось, где каждая точка данных представляет одну ооцисту, а ее цвет отражает плотность данных. Рис. 4 Каждая точка данных представляет собой одну ооцисту, а цвет представляет плотность данных. Первые 3000 обнаруженных событий нанесены на график в каждом подмножестве рисунка, а популяции ооцист и раковин аннотированы в соответствии с анализом, описанным в разделе моделирования. ( a ) Ооцисты телят (штамм Айова), измеренные в течение 1 месяца после размножения ооцист. ( b ) Ооцисты, размножаемые мышами (штамм Айова), используются в течение 1–2 месяцев. (c) Различные образцы ооцист телят (штамм Айова), использованные в течение 1–2 месяцев. ( d ) C. parvum (штамм Моредун) от телят, измерено в течение 2–3 месяцев.
(c) Различные образцы ооцист телят (штамм Айова), использованные в течение 1–2 месяцев. ( d ) C. parvum (штамм Моредун) от телят, измерено в течение 2–3 месяцев.
Полноразмерное изображение
При старении в окружающей среде, например, ., воздействии естественного стресса, такого как ультрафиолетовое излучение и перепады температуры, или, альтернативно, при транспортировке и обработке образцов при использовании в лабораторных условиях некоторые ооцисты могут разрушаться, деформироваться или даже эксцистировать 42 . В этих случаях инфекционные спорозоиты могут быть потеряны, и, следовательно, ооцисты становятся «пустыми оболочками» или «призраками». Пустые скорлупы обычно обнаруживаются в пробах питьевой воды из окружающей среды 43 , и данные импеданса на рис. 4 показывают наличие таких поврежденных/эксцистированных ооцист. Как показано в следующем разделе, оболочки появляются в виде отдельного кластера ниже основной популяции ооцист и имеют меньшую величину импеданса (поскольку они электрически негерметичны) и фазу в среднем. Подграфики на рис. 4a–d расположены в порядке возрастания возраста. Интересно, что младшему (а) и старшему (г) 9Образцы 0157 C. parvum имеют наименьшее и наибольшее количество раковин соответственно (% раковин в a = 6, b = 6, c = 8 и d = 41), что означает, что количество раковин увеличивается с возрастом окружающей среды.
Подграфики на рис. 4a–d расположены в порядке возрастания возраста. Интересно, что младшему (а) и старшему (г) 9Образцы 0157 C. parvum имеют наименьшее и наибольшее количество раковин соответственно (% раковин в a = 6, b = 6, c = 8 и d = 41), что означает, что количество раковин увеличивается с возрастом окружающей среды.
Моделирование импеданса
Обычно диэлектрическое поведение биологических клеток в суспензии определяется теорией смесей Максвелла (ММТ) 44 , которая описывает связь между комплексным импедансом суспензии, свойствами клеток, суспензионной средой и объемная доля 45 . Оболочечные модели, основанные на ММТ, использовались для моделирования диэлектрических свойств ячеек 45,46,47 . Модели с одинарной оболочкой описывают клетки только с мембраной и без внутренней части (см. рис. 2), а модели с двойной оболочкой применяются для более сложных клеток с внутренними органеллами. Зависимое от частоты поведение этих моделей можно представить в терминах релаксации Дебая 48 . Одиночные оболочки имеют одну релаксацию, а двойные оболочки — две (первая связана с клеточной мембраной, а вторая — с внутренними органеллами, рис. 2б) 27, 45 . Таким образом, целостность и сложность клетки можно вывести, подгоняя данные к этим моделям.
Одиночные оболочки имеют одну релаксацию, а двойные оболочки — две (первая связана с клеточной мембраной, а вторая — с внутренними органеллами, рис. 2б) 27, 45 . Таким образом, целостность и сложность клетки можно вывести, подгоняя данные к этим моделям.
Частотно-зависимый импеданс для средней популяции необработанных и инактивированных нагреванием ооцист C. parvum был измерен в диапазоне от 250 кГц до 50 МГц с проводимостью буфера σ Рис. 5. Каждая точка данных представляет собой среднее значение 1000 ооцист на частоту зондирования.
Рисунок 5 Спектр импеданса для необработанного (UTP) и термоинактивированного C. parvum (HIP) в диапазоне частот от 250 кГц до 50 МГц, для буферной проводимости σ м = 0,76 См м −1 . Среднее значение действительной (\(\Re \)(Z)) и мнимой (\(\Im \)(Z)) частей импеданса для оболочек (( a ) и ( b )) и ооцист (( c ) и ( d )), соответственно, нанесены (звездочки). Затем два набора данных были смоделированы с использованием подгонки одинарной оболочки (SSF) для оболочек (( a ) и (b )) и подгонки двойной оболочки (DSF) для ооцист (9).0333 (c) и (d) ), и нанесены на данные (сплошные линии).
Затем два набора данных были смоделированы с использованием подгонки одинарной оболочки (SSF) для оболочек (( a ) и (b )) и подгонки двойной оболочки (DSF) для ооцист (9).0333 (c) и (d) ), и нанесены на данные (сплошные линии).
Изображение полного размера
Данные импеданса для пустых оболочек в необработанных и термоинактивированных суспензиях C. parvum были смоделированы путем подгонки данных к одной дебаевской релаксации, представляющей одиночную подгонку оболочки (SSF) — рис. 5a, б. Как и ожидалось, простая структура оболочки ооцисты, в которой отсутствуют спорозоиты или другие органеллы, хорошо подходит для однократной релаксации. Кроме того, термоинактивация суспензии приводит к физическому разрушению мембраны, что приводит к явному снижению величины низкочастотного импеданса (рис. 5а). Основная популяция ооцист не может быть приспособлена к одной модели оболочки (см. Дополнительный рисунок S2), что означает, что эта модель не полностью отражает тонкости структуры ооцисты. Однако модель с двойной оболочкой обеспечивает хорошее соответствие, как показано на рис. 5c, d, 9.0157, т. е. ., он правильно представляет релаксацию мембраны ооцисты (~ 1–10 МГц), за которой следует расслабление внутреннего содержимого (> 10 МГц). Кроме того, как видно для оболочек, тепловая инактивация и последующее разрушение внешней стенки вызывают небольшое падение импеданса на низких частотах (рис. 5c). Таким образом, простая модель подтверждает правильность идентификации двух популяций (рис. 4) и возможность исключения раковин C. parvum из будущего анализа ооцист.
Однако модель с двойной оболочкой обеспечивает хорошее соответствие, как показано на рис. 5c, d, 9.0157, т. е. ., он правильно представляет релаксацию мембраны ооцисты (~ 1–10 МГц), за которой следует расслабление внутреннего содержимого (> 10 МГц). Кроме того, как видно для оболочек, тепловая инактивация и последующее разрушение внешней стенки вызывают небольшое падение импеданса на низких частотах (рис. 5c). Таким образом, простая модель подтверждает правильность идентификации двух популяций (рис. 4) и возможность исключения раковин C. parvum из будущего анализа ооцист.
Оптимальная проводимость суспензии
Средний импеданс всех необработанных и инактивированных нагреванием популяций C. parvum , взвешенных в буфере проводимости (σ м = 1,51 См м -1 ) показан на рис. 6a,b. С увеличением проводимости буфера частота релаксации внешней стенки, как и ожидалось, смещается в сторону более высоких частот. На рисунке 6 также показано, что диэлектрические свойства необработанного и термоинактивированного C.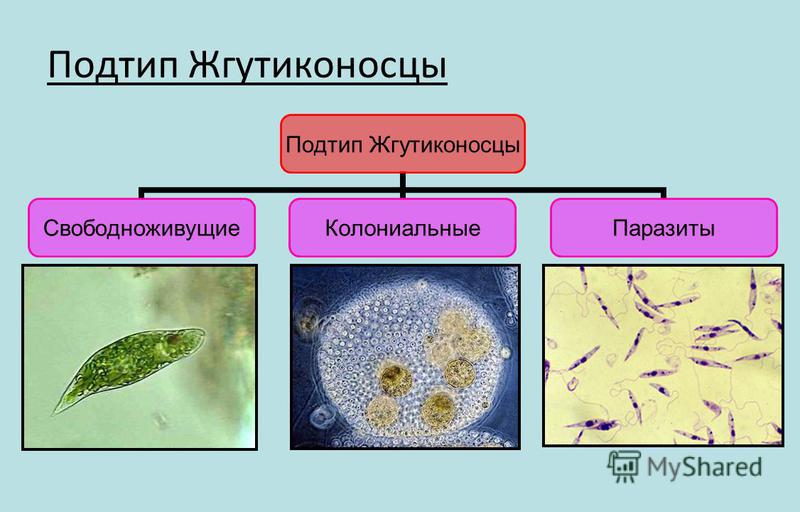 parvum различаются, особенно на более высоких частотах (см. рис. 6c, e), где на сигнал влияют свойства внутренней части ооцисты. Как правило, C. parvum Внутренняя часть ооцисты состоит из четырех (содержащих ядра) спорозоитов – инфекционных агентов ооцисты – и окруженного мембраной остаточного тела. Остаточное тело содержит большое липидное тело, многочисленные гранулы амилопектина, кристаллическое белковое включение, рибосомы и цитомембраны 4 . Для термоинактивированного C. parvum целостность и/или состав внутренней жидкости и внутренних органелл предположительно были нарушены, а разница осмоляльности между внутренней частью ооцисты и суспендирующей средой была больше в 1xPBS, чем в 0,5xPBS-буфере (σ м = 0,76 См м −1 ) – см. рис. 5в,г. Таким образом, разница в высокочастотном импедансе популяций паразитов была больше у повторно суспендированных в PBS (рис. 6). Эти данные свидетельствуют о том, что паразиты подвергались воздействию гиперосмотических условий при взвешивании в PBS.
parvum различаются, особенно на более высоких частотах (см. рис. 6c, e), где на сигнал влияют свойства внутренней части ооцисты. Как правило, C. parvum Внутренняя часть ооцисты состоит из четырех (содержащих ядра) спорозоитов – инфекционных агентов ооцисты – и окруженного мембраной остаточного тела. Остаточное тело содержит большое липидное тело, многочисленные гранулы амилопектина, кристаллическое белковое включение, рибосомы и цитомембраны 4 . Для термоинактивированного C. parvum целостность и/или состав внутренней жидкости и внутренних органелл предположительно были нарушены, а разница осмоляльности между внутренней частью ооцисты и суспендирующей средой была больше в 1xPBS, чем в 0,5xPBS-буфере (σ м = 0,76 См м −1 ) – см. рис. 5в,г. Таким образом, разница в высокочастотном импедансе популяций паразитов была больше у повторно суспендированных в PBS (рис. 6). Эти данные свидетельствуют о том, что паразиты подвергались воздействию гиперосмотических условий при взвешивании в PBS. Данные на рис. 6a,b также указывают на отсутствие различий ни в величине, ни в фазе, независимо от штамма или источника ооцист C. parvum .
Данные на рис. 6a,b также указывают на отсутствие различий ни в величине, ни в фазе, независимо от штамма или источника ооцист C. parvum .
Полное сопротивление 9 Ом0157 C. parvum в PBS. ( a ) Величина импеданса и ( b ) фаза необработанного (красный) и термоинактивированного (синий) C. parvum в зависимости от частоты; данные представляют собой среднее значение ± ± SD для всех четырех образцов 90 157 C. parvum 90 158. Низкочастотный, промежуточный и высокочастотный импеданс необработанного и термоинактивированного C. parvum нанесены на график вместе при ( c ) 250 кГц, ( d ) 2,5 МГц и (e) 25 МГц. Аннотированные эллипсы достоверности содержат 50% каждой совокупности, а цвет каждой точки данных представляет нормализованную близость каждого события к соответствующему среднему значению совокупности. Необработанные и термоинактивированные суспензии, содержащие ооцисты из всех четырех исходных образцов, измеряли отдельно.
Увеличенное изображение
Суспензия популяций ооцист в гиперосмотическом буфере с высокой электропроводностью (5x PBS; инактивированный C. parvum . На рисунке 7 показан низкочастотный, промежуточный и высокочастотный импеданс C. parvum в 5x PBS. Высокочастотный импеданс позволяет четко различать популяции паразитов, и эта разница усиливается за счет увеличения проводимости взвешенной среды. Зеленая линия указывает линию равновероятностного отклонения между популяциями ( , т. е. ., где обнаруженное событие имеет равные шансы принадлежать к любой популяции) и является оценкой достоверности различия между необработанными и инактивированными нагреванием C. parvum . Например, на частоте 50 МГц, где было достигнуто наиболее четкое различение (рис. 7с), 91% событий справа от границы равной вероятности были от необработанной популяции C. parvum и 92% событий от слева были из термоинактивированного C. parvum населения. Таким образом, МИК позволила идентифицировать необработанные или инактивированные нагреванием C. parvum с достоверностью более 90% в этих экспериментальных условиях. Данные, содержащиеся в таблице S2, показывают достоверность идентификации, достигнутую для каждого отдельного образца, и демонстрируют, что возраст образца и/или жизнеспособность образца отрицательно влияют на уровень дискриминации.
parvum с достоверностью более 90% в этих экспериментальных условиях. Данные, содержащиеся в таблице S2, показывают достоверность идентификации, достигнутую для каждого отдельного образца, и демонстрируют, что возраст образца и/или жизнеспособность образца отрицательно влияют на уровень дискриминации.
График рассеяния импеданса для C. parvum в 5x PBS. Данные импеданса на низких, средних и высоких частотах для необработанных и термоинактивированных C. parvum нанесены вместе на ( a ) 500 кГц, ( b ) 5 МГц и ( c ) 50 МГц. Аннотированные эллипсы достоверности содержат ~ 50% необработанных или инактивированных нагреванием популяций (1000 событий для каждой популяции). Зеленая линия указывает на границу равной вероятности между двумя популяциями. Цвет каждой точки данных представляет нормализованную близость события к соответствующему среднему значению генеральной совокупности.
Полноразмерное изображение
Анализ видов
Сопротивление патогенов, которые обычно выделяют с помощью EPA 1623.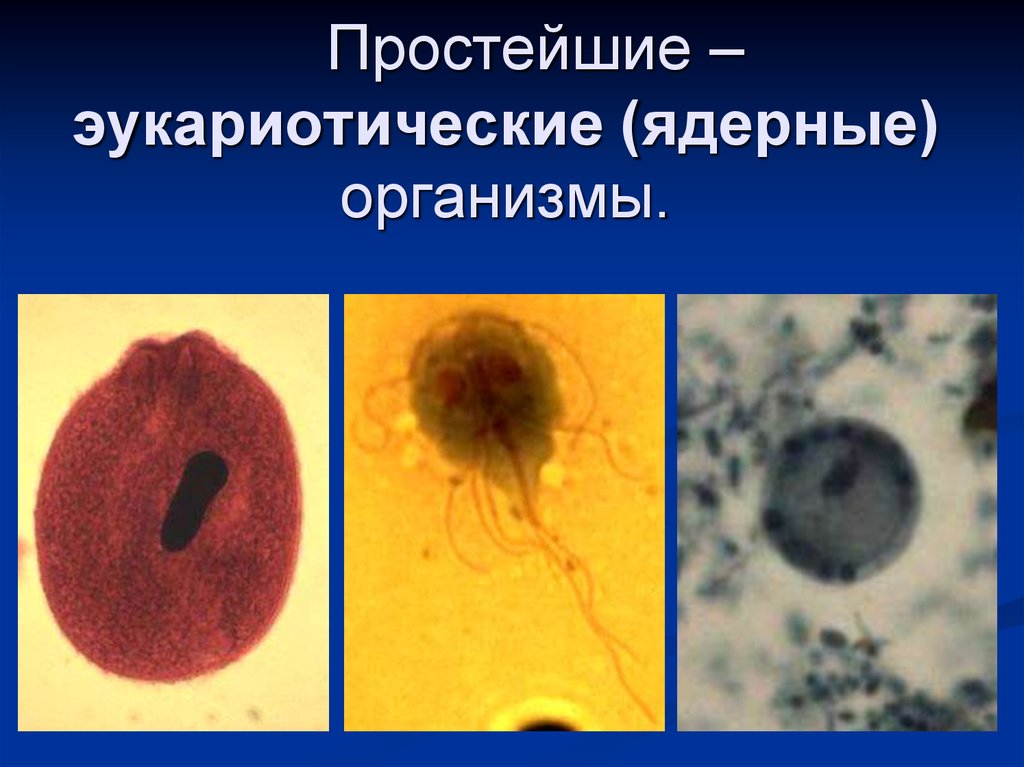 1, измеряли для оценки пригодности MIC для различения передающихся через воду паразитов на уровне видов и родов. В частности, были проанализированы образцы, содержащие C. parvum (CP), C. muris (CM) и G. lamblia (GL), суспендированные в PBS (σ m = 1,61 S m -1 ). Чтобы правильно ввести и охарактеризовать импеданс каждой популяции, образцы сначала измерялись независимо — дополнительный рисунок S3. Кубический корень из величины низкочастотного импеданса пропорционален диаметру частицы 28, 34 . Используя эти данные, средний диаметр паразитов был определен как CP = 4,3 ± 0,3 мкм, CM = 5,9 ± 0,4 мкм и GL = 9,3 ± 0,4 мкм, что хорошо коррелирует с размерами, полученными при оптической визуализации 49, 80,9017
1, измеряли для оценки пригодности MIC для различения передающихся через воду паразитов на уровне видов и родов. В частности, были проанализированы образцы, содержащие C. parvum (CP), C. muris (CM) и G. lamblia (GL), суспендированные в PBS (σ m = 1,61 S m -1 ). Чтобы правильно ввести и охарактеризовать импеданс каждой популяции, образцы сначала измерялись независимо — дополнительный рисунок S3. Кубический корень из величины низкочастотного импеданса пропорционален диаметру частицы 28, 34 . Используя эти данные, средний диаметр паразитов был определен как CP = 4,3 ± 0,3 мкм, CM = 5,9 ± 0,4 мкм и GL = 9,3 ± 0,4 мкм, что хорошо коррелирует с размерами, полученными при оптической визуализации 49, 80,9017
Разницу в импедансе между популяциями можно визуализировать несколькими способами, но наиболее четкое различие получается из диаграммы рассеяния высокочастотной фазы (на 18,3 МГц) по сравнению с величиной низкочастотного импеданса (на 250 кГц), как показано на рис. 8. Эти данные эффективно отображают изменения внутренней структуры/состава 9Размер 0157 против . На рисунке показаны данные для 1000 событий, полученные путем измерения выборки, содержащей все различные популяции. Используя те же ворота, что и на рисунках 5 и 6, можно было идентифицировать CP с достоверностью 98 %, CM с достоверностью 93 % и GL с достоверностью более 99 %. Эти результаты показывают, что анализ импеданса позволяет различать эти патогены с высокой степенью достоверности, особенно по сравнению с обычной проточной цитометрией (рис. 8b).
8. Эти данные эффективно отображают изменения внутренней структуры/состава 9Размер 0157 против . На рисунке показаны данные для 1000 событий, полученные путем измерения выборки, содержащей все различные популяции. Используя те же ворота, что и на рисунках 5 и 6, можно было идентифицировать CP с достоверностью 98 %, CM с достоверностью 93 % и GL с достоверностью более 99 %. Эти результаты показывают, что анализ импеданса позволяет различать эти патогены с высокой степенью достоверности, особенно по сравнению с обычной проточной цитометрией (рис. 8b).
Импеданс против оптического обнаружения паразитов. ( a ) Диаграмма рассеяния фазы на частоте 18,3 МГц в зависимости от величины импеданса на частоте 250 кГц для всех видов паразитов, измеренных вместе в смешанной выборке. Аннотированные эллипсы достоверности содержат ~ 50% каждой совокупности. Зеленые линии указывают границы, определяющие равновероятностное отклонение между соседними популяциями. Цвет каждой точки данных представляет нормализованную близость события к соответствующему среднему значению генеральной совокупности. ( b ) Данные обычной проточной цитометрии (SSC против FSC) для всех видов паразитов в PBS. Индивидуальные данные оптического рассеяния для каждой популяции нанесены вместе. В общей сложности 1000 событий нанесены как на ( a ), так и на ( b ).
Цвет каждой точки данных представляет нормализованную близость события к соответствующему среднему значению генеральной совокупности. ( b ) Данные обычной проточной цитометрии (SSC против FSC) для всех видов паразитов в PBS. Индивидуальные данные оптического рассеяния для каждой популяции нанесены вместе. В общей сложности 1000 событий нанесены как на ( a ), так и на ( b ).
Изображение в полный размер
По оценкам, два вида C. parvum и C. hominis ответственны за более чем 90% случаев криптоспоридиоза у людей 51 . Однако другие виды ( C. meleagridis , C. canis, C. cuniculus и C. felis ) также связаны с инфицированием человека и считаются «основными патогенными для человека видами» 49, 52 . С точки зрения размеров ооцист все эти «основные» виды демонстрируют сходство и обычно описываются как эллиптические ооцисты в диапазоне размеров 4,0–5,0 × 5,0–5,5 мкм 49 .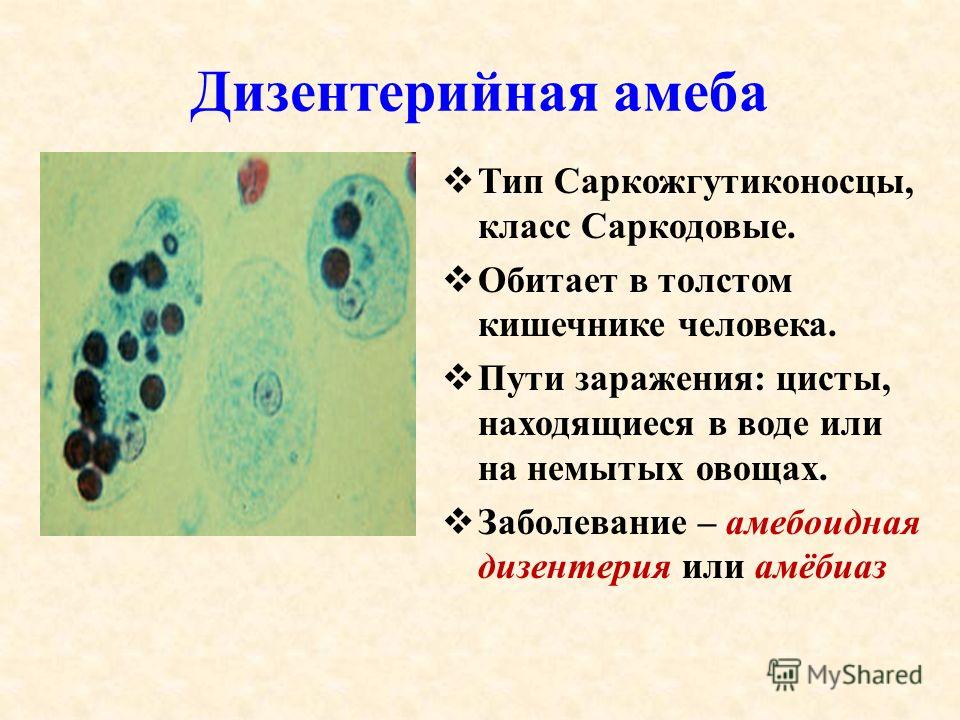 Наши результаты показывают, что MIC может с высокой степенью достоверности отличить ооцисты такого размера от других видов Cryptosporidium , размеры которых не входят в этот диапазон и которые не представляют или практически не представляют опасности для здоровья человека (как мы продемонстрировали с C. parvum и C. muris ). Такие виды включают (но не ограничиваются ими) более крупные C. andersoni , C. baileyi и C. muris , а также более мелкие C. xiaoi и C. ryanae . 49 , которые были идентифицированы среди видов с низким уровнем риска, загрязняющих питьевую воду в Великобритании в течение одного года 53 . Однако необходим анализ отдельных видов, имеющих сходные размеры, чтобы оценить, могут ли межвидовые вариации стенки ооцисты и внутренней структуры/состава способствовать различению таких ооцист на основе импеданса.
Наши результаты показывают, что MIC может с высокой степенью достоверности отличить ооцисты такого размера от других видов Cryptosporidium , размеры которых не входят в этот диапазон и которые не представляют или практически не представляют опасности для здоровья человека (как мы продемонстрировали с C. parvum и C. muris ). Такие виды включают (но не ограничиваются ими) более крупные C. andersoni , C. baileyi и C. muris , а также более мелкие C. xiaoi и C. ryanae . 49 , которые были идентифицированы среди видов с низким уровнем риска, загрязняющих питьевую воду в Великобритании в течение одного года 53 . Однако необходим анализ отдельных видов, имеющих сходные размеры, чтобы оценить, могут ли межвидовые вариации стенки ооцисты и внутренней структуры/состава способствовать различению таких ооцист на основе импеданса.
Заключение и перспективы
Использование MIC впервые дает возможность оценить вид и статус жизнеспособности на уровне одной (оо)цисты, с дополнительными преимуществами быстрой автоматизированной обработки и неразрушающего метода без маркировки.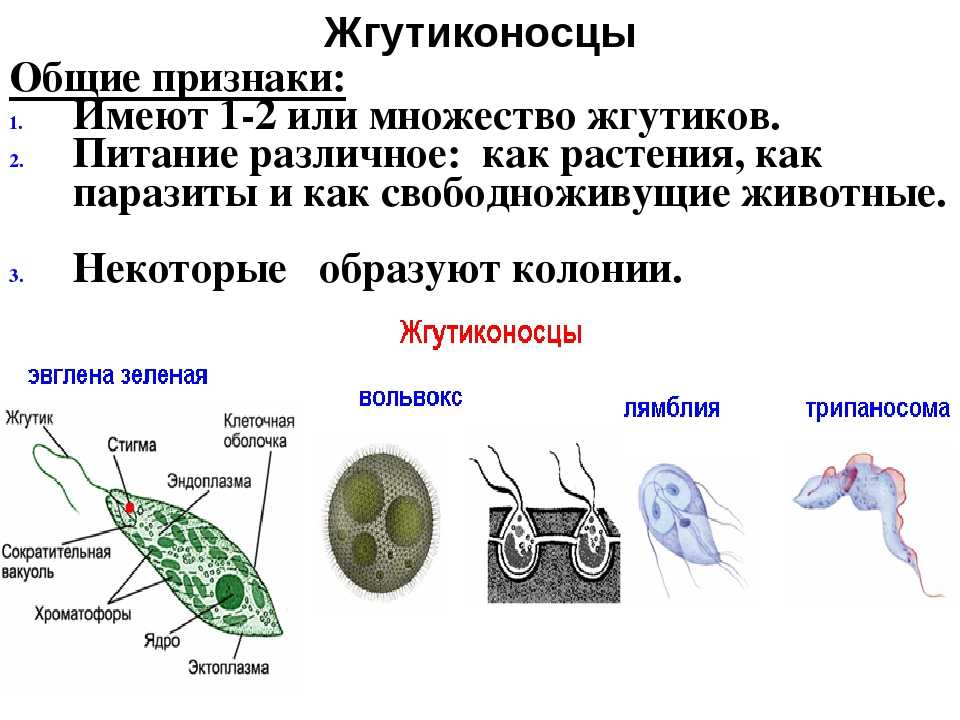 метод. Импедансные свойства C. parvum ооцист различного происхождения, штамма и возраста измеряли в буферах различной проводимости. Данные об импедансе популяций ооцист и раковин в каждом образце были приспособлены либо к модели с одинарной, либо к двойной оболочке. Результаты показали, что можно было определить наличие поврежденных/эксцистированных ооцист (называемых раковинами) и что количество раковин увеличивалось с возрастом окружающей среды.
метод. Импедансные свойства C. parvum ооцист различного происхождения, штамма и возраста измеряли в буферах различной проводимости. Данные об импедансе популяций ооцист и раковин в каждом образце были приспособлены либо к модели с одинарной, либо к двойной оболочке. Результаты показали, что можно было определить наличие поврежденных/эксцистированных ооцист (называемых раковинами) и что количество раковин увеличивалось с возрастом окружающей среды.
Способность MIC исследовать различные слои структуры ооцисты означает, что анализ импеданса обеспечивает более четкое различение, чем оптическая проточная цитометрия — эквивалентный метод без меток (см. Дополнительный рисунок S4). Данные показывают, что необработанные и термоинактивированные C. parvum может быть идентифицирован с достоверностью более 90% на высоких частотах при суспендировании в 5x PBS. Предполагается, что контраст лучше всего проявляется в этой проводимости, потому что нежизнеспособный паразит теряет способность регулировать перенос ионов, что приводит к направленному внутрь потоку ионов, который увеличивает электрический «контраст» между необработанными и инактивированными нагреванием C. parvum . . Кроме того, были идентифицированы одиночные C. parvum , C. muris и G. lamblia (oo) цисты с более чем 92% уверенности по высокочастотной фазе по сравнению с низкочастотными данными магнитуды (при приостановке в PBS).
parvum . . Кроме того, были идентифицированы одиночные C. parvum , C. muris и G. lamblia (oo) цисты с более чем 92% уверенности по высокочастотной фазе по сравнению с низкочастотными данными магнитуды (при приостановке в PBS).
Эта технология потенциально может быть полезна для предприятий водоснабжения, например, ., системы предупреждения, которая быстрее идентифицирует отдельные Cryptosporidium и Giardia (oo)цисты после восстановления через EPA 1623.1. Как было продемонстрировано, система также может одновременно обеспечивать индикацию жизнеспособности и видов извлеченных (оо)цист, что может снизить потребность в квалифицированных микроскопистах и связанную с этим потребность в рабочей силе. Эту систему также можно использовать для мониторинга простейших пищевого происхождения или для анализа ветеринарных или клинических образцов.
Для широкого использования этого метода в, например, . , мониторинге воды, это исследование должно быть расширено, чтобы продемонстрировать, что ооцисты Cryptosporidium , которые были инактивированы другими способами, кроме тепловой инактивации, также могут быть отделены от жизнеспособных ооцист. Будущая работа должна также оценить пригодность МИК для различения других видов Cryptosporidium , которые представляют/не представляют значительного риска для здоровья человека, а также охарактеризовать эффективность работы с (оо)цистами из различных матриц, включая воду, пищу и образцы фекалий. Метод также следует сравнить на предмет замены «золотого стандарта» протоколов инфекционности животных.
, мониторинге воды, это исследование должно быть расширено, чтобы продемонстрировать, что ооцисты Cryptosporidium , которые были инактивированы другими способами, кроме тепловой инактивации, также могут быть отделены от жизнеспособных ооцист. Будущая работа должна также оценить пригодность МИК для различения других видов Cryptosporidium , которые представляют/не представляют значительного риска для здоровья человека, а также охарактеризовать эффективность работы с (оо)цистами из различных матриц, включая воду, пищу и образцы фекалий. Метод также следует сравнить на предмет замены «золотого стандарта» протоколов инфекционности животных.
При условии дальнейшей валидации с использованием образцов из окружающей среды эту технологию можно внедрить в существующую нормативную базу для мониторинга переносимых через воду простейших, чтобы обеспечить быструю оценку жизнеспособности ооцист и, таким образом, обеспечить более точную оценку риска для здоровья человека. Например, существующий метод обнаружения использует иммуномагнитную сепарацию (IMS) для обогащения паразитов, что дает объем ~ 50 мкл с извлеченными ооцистами 16 . Увеличив проводимость образца этого продукта, можно будет отличить необработанный и термоинактивированный C. parvum с использованием MIC. Система могла бы работать с повышенным расходом 28 , поэтому можно было бы обработать образец гораздо меньше, чем за час. Наконец, также можно просматривать результаты в режиме реального времени, что позволит предприятиям водоснабжения быстро выявлять потенциально опасную питьевую воду.
Увеличив проводимость образца этого продукта, можно будет отличить необработанный и термоинактивированный C. parvum с использованием MIC. Система могла бы работать с повышенным расходом 28 , поэтому можно было бы обработать образец гораздо меньше, чем за час. Наконец, также можно просматривать результаты в режиме реального времени, что позволит предприятиям водоснабжения быстро выявлять потенциально опасную питьевую воду.
Ссылки
Всемирная организация здравоохранения. Руководство по обеспечению качества питьевой воды – четвертое издание . (Всемирная организация здравоохранения, 2011 г.).
Дэвис А. П. и Чалмерс Р. М. Криптоспоридиоз. Британский медицинский журнал 339 , doi:10.1136/bmj.b4168 (2009).
Джекс, А. Р., Смит, Х. В., Монис, П. Т., Кэмпбелл, Б.
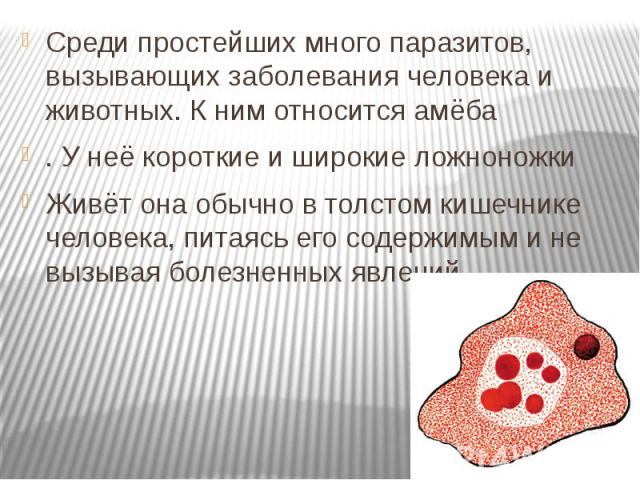 Э. и Гассер, Р. Б. Cryptosporidium — Биотехнологические достижения в обнаружении, диагностике и анализе генетической изменчивости. Достижения биотехнологии 26 , 304–317 (2008).
Э. и Гассер, Р. Б. Cryptosporidium — Биотехнологические достижения в обнаружении, диагностике и анализе генетической изменчивости. Достижения биотехнологии 26 , 304–317 (2008).КАС Статья пабмед Google ученый
Fayer, R. & Xiao, L. Cryptosporidium and Cryptosporidiosis, второе издание. (Тейлор и Фрэнсис, 2012).
Дейл, К., Кирк, М., Синклер, М., Холл, Р. и Ледер, К. Зарегистрированные вспышки желудочно-кишечных заболеваний, передаваемых через воду, в Австралии в основном связаны с рекреационным воздействием. Австралийский и новозеландский журнал общественного здравоохранения 34 , 527–530 (2010).
Артикул пабмед Google ученый
Эллиот, А. Дж. и др. . Потенциальное влияние сообщений СМИ на синдромальный эпиднадзор: пример возможного заражения Cryptosporidium в Северо-Западной Англии, август-сентябрь 2015 г.
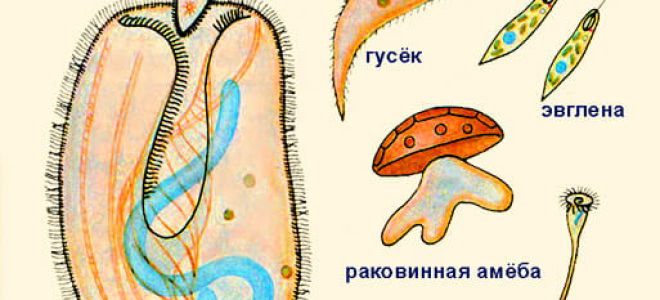 Евронадзор 21 (2016).
Евронадзор 21 (2016).Видерстрём, М. и др. . Крупная вспышка инфекции Cryptosporidium hominis, передающейся через системы водоснабжения, Швеция. Новые инфекционные болезни 20 , 581–589 (2014).
Артикул пабмед ПабМед Центральный Google ученый
Šlapeta, J. Криптоспоридиоз и виды Cryptosporidium у животных и человека: тридцатицветная радуга? Международный журнал паразитологии 43 , 957–970 (2013).
Артикул пабмед Google ученый
Smith, H.V. & Nichols, R.A.B. Cryptosporidium: Обнаружение в воде и пищевых продуктах. Экспериментальная паразитология 124 , 61–79 (2010).
Артикул пабмед Google ученый
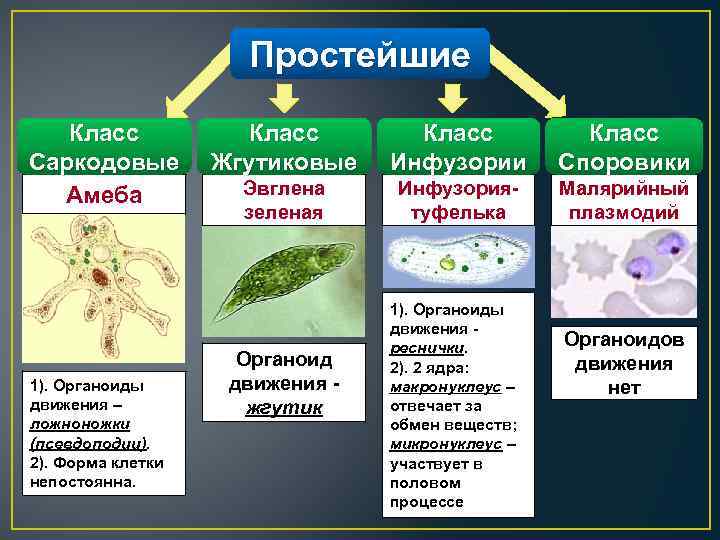 «>
«>Чен Ф., Хуанг К., Цинь С., Чжао Ю. и Пан К. Сравнение жизнеспособности и инфекционности 9Ооцисты 0157 Cryptosporidium parvum , хранящиеся в растворе дихромата калия и хлорированной водопроводной воде. Ветеринарная паразитология 150 , 13–17 (2007).
КАС Статья пабмед Google ученый
Берд, Т. и Уолц, Дж. Профили силы взаимодействия между ооцистами Cryptosporidium parvum и поверхностями кремнезема. Экологические науки и технологии 39 , 9574–9582 (2005).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
МакГрат, Дж. С. и др. . Оценка деформируемости переносимых водой простейших с использованием зонда для силовой микроскопии с микрофлюидной поддержкой. PLoS Один 11 , e0150438 (2016).

Артикул пабмед ПабМед Центральный Google ученый
Okhuysen, P.C., Chappell, C.L., Crabb, JH, Sterling, C.R. & DuPont, HL. Вирулентность трех различных изолятов Cryptosporidium parvum для здоровых взрослых. Журнал инфекционных болезней 180 , 1275–1281 (1999).
КАС Статья пабмед Google ученый
Кинг, Б. Дж. и Монис, П. Т. Критические процессы, влияющие на выживание ооцист Cryptosporidium в окружающей среде. Паразитология 134 , 309–323 (2007).
КАС Статья пабмед Google ученый
Шотландская вода. Годовой отчет о качестве воды за 2013 г. . (Шотландская вода, 2013 г.).
Служба охраны окружающей среды и наследия.
 Руководство по мониторингу Cryptosporidium в очищенной воде в Северной Ирландии . (Служба окружающей среды и наследия, 2003 г.).
Руководство по мониторингу Cryptosporidium в очищенной воде в Северной Ирландии . (Служба окружающей среды и наследия, 2003 г.).АООС США. Метод 1623.1: Cryptosporidium и Giardia в воде путем фильтрации/IMS/FA. (Агентство по охране окружающей среды США, 2012 г.).
Уздечка, Х. и др. . Обнаружение Cryptosporidium в миниатюрных жидкостных устройствах. Исследование воды 46 , 1641–1661 (2012).
КАС Статья пабмед Google ученый
Мост, JW и др. . Взаимодействие с водным сектором на благо общественного здравоохранения: передающиеся через воду патогены и болезни в развитых странах. Бюллетень Всемирной организации здравоохранения 88 , 873–875 (2010).
Артикул пабмед ПабМед Центральный Google ученый
 «>
«>Вс, Ю.-Х. и др. . Количественное диэлектрофоретическое отслеживание для характеристики и разделения устойчивых субпопуляций Cryptosporidium parvum. Аналитик 139 , 66–73 (2014).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
Куинн, К., Арчер, Г., Беттс, В. и О’Нил, Дж. Дозозависимый диэлектрофоретический ответ ооцист Cryptosporidium, обработанных озоном. Письма по прикладной микробиологии 22 , 224–228 (1996).
КАС Статья пабмед Google ученый
Гоутер, А. Д., Берт, Дж. П. и Петик, Р. Комбинированное устройство для диэлектрофореза бегущей волны и электровращения: применение для определения концентрации и жизнеспособности Cryptosporidium. Journal of Physics D-Прикладная физика 30, L65 (1997).
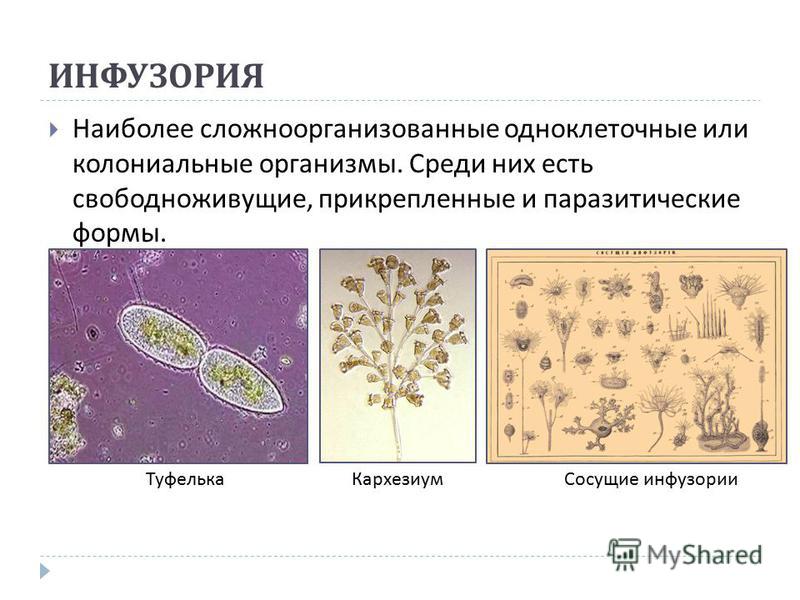
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Далтон К., Гоутер А., Берт Дж. и Смит Х. Анализ паразитов методом электровращения. Журнал прикладной микробиологии 96 , 24–32 (2004).
КАС Статья пабмед Google ученый
Унни, Х. Н. и др. . Характеристика и разделение клеток Cryptosporidium и Giardia с использованием встроенного диэлектрофореза. Биомикрофлюидика 6 , 012805 (2012).
Артикул Google ученый
Хуссен, Т., Фолле, Дж., Фолле, А., Деи-Кас, Э. и Сенез, В. Анализ паразитов, загрязняющих воду, без использования меток с помощью электрохимической импедансной спектроскопии. Биосенсоры и биоэлектроника 25 , 1122–1129 (2010).

КАС Статья пабмед Google ученый
Морган, Х., Сан, Т., Холмс, Д., Гавад, С. и Грин, Н. Г. Диэлектрическая спектроскопия одиночных клеток. Journal of Physics D: Прикладная физика 40 , 61 (2007).
ОБЪЯВЛЕНИЕ КАС Статья Google ученый
Сан, Т. и Морган, Х. Одноклеточная микрожидкостная импедансная цитометрия: обзор. Микрофлюидика и нанофлюидика 8 , 423–443 (2010).
КАС Статья Google ученый
Спенсер, Д., Эллиот, Г. и Морган, Х. Безоболочный комбинированный оптический и импедансный микроцитометр. Лаборатория на чипе 14 , 3064–3073 (2014).
КАС Статья пабмед Google ученый
 «>
«>МакГрат, Дж. С., Хименес, М. и Бридл, Х. Детерминированное боковое смещение для разделения частиц: обзор. Лаборатория на чипе 14 , 4139–4158 (2014).
КАС Статья пабмед Google ученый
Бхагат, А.А.С. и др. . Микрофлюидика для разделения клеток. Медицинская и биологическая инженерия и вычислительная техника 48 , 999–1014 (2010).
Артикул Google ученый
Песня, Х. и др. . Микрожидкостный импедансный проточный цитометр для определения состояния дифференцировки стволовых клеток. Лаборатория на чипе 13 , 2300–2310 (2013).
КАС Статья пабмед Google ученый
Холмс, Д.
 и др. . Анализ и дифференцировка лейкоцитов с использованием высокоскоростной микрофлюидной импедансной цитометрии одиночных клеток. Лаборатория на чипе 9 , 2881–2889 (2009).
и др. . Анализ и дифференцировка лейкоцитов с использованием высокоскоростной микрофлюидной импедансной цитометрии одиночных клеток. Лаборатория на чипе 9 , 2881–2889 (2009).КАС Статья пабмед Google ученый
Хан, Х., ван Беркель, К., Гвайер, Дж., Капретто, Л. и Морган, Х. Микрофлюидный лизис крови человека для анализа лейкоцитов с использованием одноклеточной импедансной цитометрии. Аналитическая химия 84 , 1070–1075 (2011).
Артикул пабмед Google ученый
Спенсер Д., Холлис В. и Морган Х. Микрофлюидная импедансная цитометрия опухолевых клеток в крови. Биомикрофлюидика 8 , 064124 (2014).
Артикул пабмед ПабМед Центральный Google ученый
 «>
«>Кюттель, К. и др. . Безэтикеточное обнаружение инфицированных Babesia bovis эритроцитов с помощью импедансной спектроскопии на проточном цитометре, изготовленном из микрофибры. Акта тропика 102 , 63–68 (2007).
Артикул пабмед Google ученый
Ду, Э. и др. . Электроимпедансная микропроточная цитометрия для характеристики болезненных состояний клеток. Лаборатория на чипе 13 , 3903–3909 (2013).
КАС Статья пабмед ПабМед Центральный Google ученый
Холмс, Д., Ше, Дж. К., Роуч, П. Л. и Морган, Х. Иммуноанализ на основе шариков с использованием проточного цитометра с микрочипом. Лаборатория на чипе 7 , 1048–1056 (2007).
КАС Статья пабмед Google ученый
 «>
«>Fayer, R. Влияние высокой температуры на инфекционность ооцист Cryptosporidium parvum в воде. Прикладная и экологическая микробиология 60 , 2732–2735 (1994).
КАС пабмед ПабМед Центральный Google ученый
Blewett, D. In Proceedings of the First International Workshop on Cryptosporidiosis. Эдинбург: Ассоциация исследования болезней животных . 85–95.
Schwan, H. In Engineering in Medicine and Biology Society, 1994. Инженерные достижения: новые возможности для инженеров-биомедиков. Материалы 16-й ежегодной международной конференции IEEE . А70–А71 том. 71 (IEEE).
Шафи, Х., Сано, М.Б., Хенсли, Э.А., Колдуэлл, Дж.Л. и Давалос, Р.В. Селективное выделение живых/мертвых клеток с помощью бесконтактного диэлектрофореза (cDEP). Лаборатория на чипе 10 , 438–445 (2010).
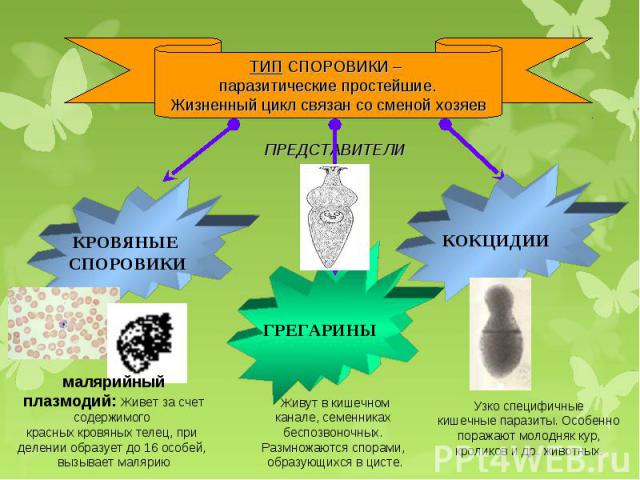
КАС Статья пабмед Google ученый
Агентство по охране окружающей среды. Методы исследования вод и связанных с ними материалов . (Агентство по охране окружающей среды, 2010 г.).
Костржинская М. и др. . Три протокола подготовки проб для обнаружения Cryptosporidium parvum на основе полимеразной цепной реакции в пробах окружающей среды. Журнал микробиологических методов 35 , 65–71 (1999).
КАС Статья пабмед Google ученый
Максвелл, Дж. К. Трактат об электричестве и магнетизме . Том. 1 (Кларендон пресс, 1881 г.).
Morgan, H. & Green, N.G. AC Электрокинетика: коллоиды и наночастицы . (Издательство Исследовательских Исследований, 2003).
 «>
«>Миллер, Р. Д. и Джонс, Т. Б. Электроориентация эллипсоидальных эритроцитов. Теория и эксперимент. Биофизический журнал 64 , 1588 (1993).
ОБЪЯВЛЕНИЕ КАС Статья пабмед Google ученый
Сан, Т., Грин, Н. Г., Гавад, С. и Морган, Х. Аналитический анализ электрического поля и чувствительности для двух конструкций микрофлюидных импедансных цитометров. ИЭТ Нанобиотехнологии 1 , 69–79 (2007).
КАС Статья пабмед Google ученый
Дебай, П. и Фалькенхаген, Х. Дисперсия проводимости и диэлектрической проницаемости сильных электролитов, Phys . Том. 29 (З, 1928).
Чалмерс, Р. М. и Катцер, Ф. В поисках Cryptosporidium: применение достижений в обнаружении и диагностике. Тенденции паразитологии 29 , 237–251 (2013).

Артикул пабмед Google ученый
LeChevallier, M.W., Norton, W.D. & Lee, R.G. Наличие Giardia и Cryptosporidium spp. в поверхностном водоснабжении. Прикладная и экологическая микробиология 57 , 2610–2616 (1991).
КАС пабмед ПабМед Центральный Google ученый
Бузид М., Хантер П. Р., Чалмерс Р. М. и Тайлер К. М. Патогенность и вирулентность криптоспоридий. Обзоры клинической микробиологии 26 , 115–134 (2013).
КАС Статья пабмед ПабМед Центральный Google ученый
Fratamico, P., Liu, Y. & Kathariou, S. Геномы пищевых и водных патогенов . (Американское общество микробиологии, 2011 г.).
 «>
«>Николс Р., Коннелли Л., Салливан К. и Смит Х. Идентификация видов и генотипов Cryptosporidium в шотландских сырых и питьевых водах в течение одногодичного периода мониторинга. Прикладная и экологическая микробиология 76 , 5977–5986 (2010).
КАС Статья пабмед ПабМед Центральный Google ученый
Скачать ссылки
Благодарности
Авторы признательны Совету по научно-техническим средствам, EPSRC и Европейской комиссии в рамках проекта ITN по сортировке частиц без этикеток (LAPASO) в рамках программы «Люди» (Действия Марии Кюри) Седьмой рамочной программы Европейского Союза FP7/2007. -2013 г. по договору гранта РЭА № 607350 ИНН на финансирование. Мы также хотели бы поблагодарить Scottish Water за их постоянную поддержку.
Информация об авторе
Примечания об авторе
Дж. С. МакГрат и К.
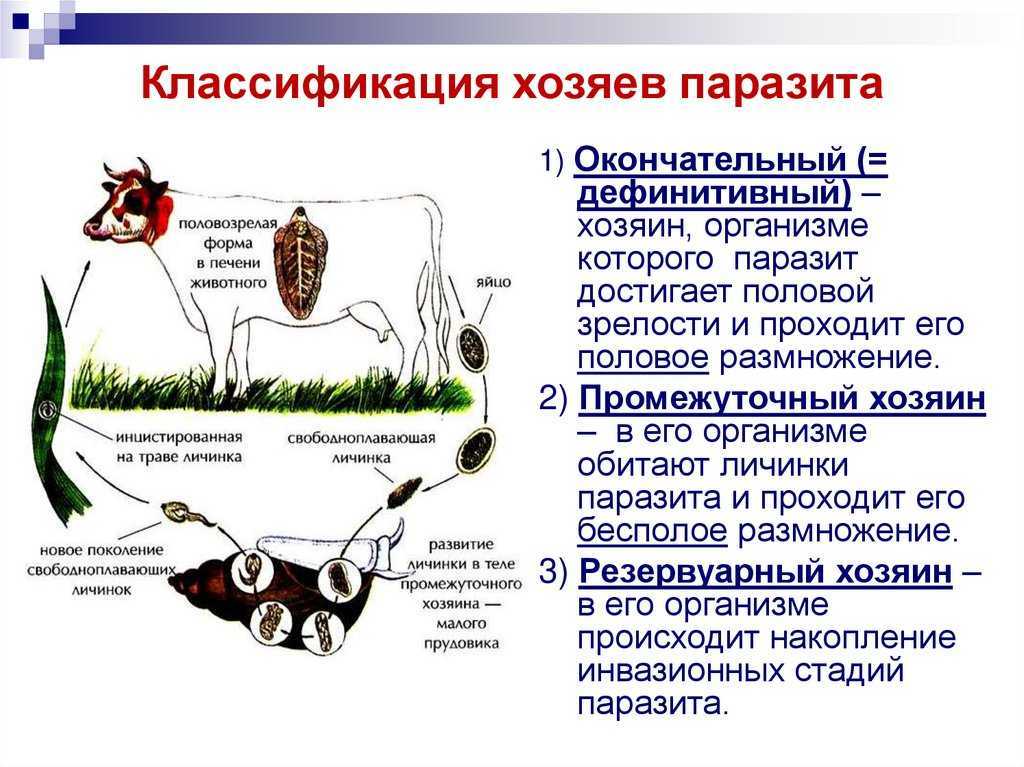 Онрадо внесли равный вклад в эту работу.
Онрадо внесли равный вклад в эту работу.
Авторы и принадлежности
Институт биологической химии, биофизики и биоинженерии, Школа инженерных и физических наук, Университет Хериот-Ватт, Эдинбург, EH24 4AS, Великобритания
. S. S. S. McGrath & H. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. L. Факультет физических и инженерных наук и Институт наук о жизни, Саутгемптонский университет, Саутгемптон, SO17 1BJ, Соединенное Королевство
C. Honrado, D. Spencer & H. Morgan
Moredun Scientific, Pentlands Science Park, Bush Loan, Penicuik, Midlothian, EH36 0PZ, Соединенное Королевство
B. Horton
авторы
- . McGrath
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- C.
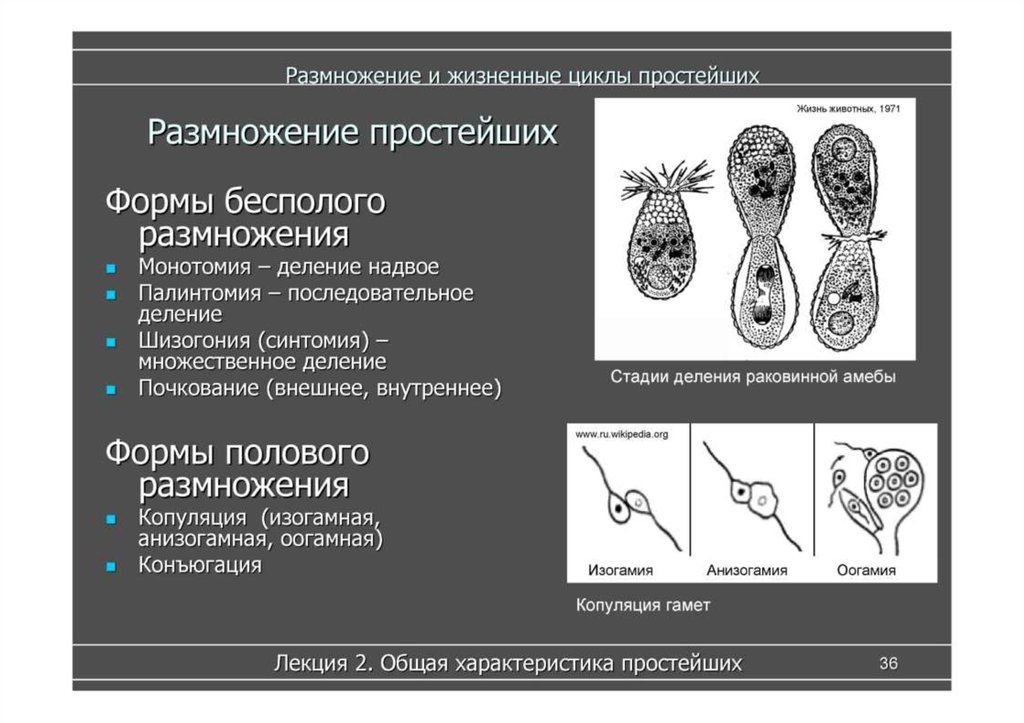 Honrado
HonradoПросмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- D. Spencer
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- B. Horton
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- H. L. Bridle
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Х. Морган
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
D.C. and H.M. спроектировал и изготовил систему обнаружения. Дж. М., К. Х., Д. К., Х. Б. и Х.М. придумали и разработали эксперименты. Дж.М., К.Х. и Б.Х. выполнил экспериментальную работу. Х.Б. и Х.М. предоставленные реагенты/материалы/инструменты для анализа. Дж.М., К.Х. и DC создали рукопись и представили рисунки. округ Колумбия и Х.М. проанализировал данные. Дж. М., округ Колумбия, Х. Б. и Х.М. пересмотрел рукопись.
Дж.М., К.Х. и DC создали рукопись и представили рисунки. округ Колумбия и Х.М. проанализировал данные. Дж. М., округ Колумбия, Х. Б. и Х.М. пересмотрел рукопись.
Автор, ответственный за переписку
Переписка с Х. Морган.
Заявление об этике
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих интересов.
Дополнительная информация
Все данные, подтверждающие это исследование, находятся в открытом доступе в репозитории Университета Саутгемптона по адресу http://doi.org/10.5258/SOTON/D0047.
Примечание издателя: Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Электронный дополнительный материал
41598_2017_2715_MOESM1_ESM.doc
Анализ паразитических простейших на уровне отдельных клеток с использованием микрофлюидной импедансной цитометрии — Дополнительная информация
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4. 0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или превышает разрешенное использование, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или превышает разрешенное использование, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Эту статью цитирует
Электрический импеданс как показатель здоровья клеток микроводорослей
- Цзянье Суй
- Фатима Фофлонкер
- Мехди Джаванмар
Научные отчеты (2020)
Автоматизация сортировки отдельных клеток на основе изображений в капельной микрофлюидике
- Мухсинкан Сесен
- Грэм Уайт
Научные отчеты (2020)
Микрожидкостное разделительное устройство на основе двойного неодимового магнита
- Хён Ги Ке
- Пак Пён Сон
- Бон Гын Чунг
Научные отчеты (2019)
Одноразовые микрожидкостные микромиксеры для эффективного улавливания ооцист Cryptosporidium parvum из проб воды
- Л.
 Диегес
Диегес - М. Зима
- Б. Тьерри
Журнал биологической инженерии (2018)
- Л.
Интеграция мониторинга скорости роста на основе импеданса в микрожидкостную платформу клеточных культур для микроскопии живых клеток
- Кетки Чавла
- Себастьян К.
 Бюргель
Бюргель - Андреас Хирлеманн
Микросистемы и наноинженерия (2018)
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Паразитарные инфекции | Определение и обучение пациентов
Что такое паразитарная инфекция?
Паразиты — это организмы, которые живут за счет других организмов или хозяев, чтобы выжить. Некоторые паразиты не оказывают заметного влияния на своих хозяев. Другие растут, размножаются или вторгаются в системы органов, вызывая заболевание их хозяев, что приводит к паразитарной инфекции.
Паразитарные инфекции представляют большую проблему в тропических и субтропических регионах мира. Малярия — одно из самых смертоносных паразитарных заболеваний. Паразитарные инфекции также могут возникать в Соединенных Штатах. Общие паразитарные инфекции, обнаруженные в Соединенных Штатах, включают:
- трихомониаз
- лямблиоз
- криптоспоридиоз
- токсоплазмоз
Симптомы паразитарных инфекций варьируются в зависимости от организма. Например:
- Трихомониаз — это инфекция, передающаяся половым путем, вызываемая паразитом, который часто не вызывает никаких симптомов. В некоторых случаях это может вызвать зуд, покраснение, раздражение и необычные выделения в области половых органов.
- Лямблиоз может вызывать диарею, газы, расстройство желудка, жирный стул и обезвоживание.
- Криптоспоридиоз может вызывать желудочные спазмы, боль в животе, тошноту, рвоту, обезвоживание, потерю веса и лихорадку.

- Токсоплазмоз может вызывать гриппоподобные симптомы, включая увеличение лимфатических узлов и мышечные боли или боли, которые могут длиться более месяца.
Паразитарные инфекции могут быть вызваны тремя типами организмов:
- простейшими
- гельминтами
- эктопаразитами
Простейшие — это одноклеточные организмы, которые могут жить и размножаться внутри вашего тела. Некоторые инфекции, вызванные простейшими, включают лямблиоз. Это серьезная инфекция, которой можно заразиться через питьевую воду, зараженную простейшими Giardia .
Гельминты — это многоклеточные организмы, которые могут жить в вашем теле или вне его. Они более известны как черви. К ним относятся плоские черви, ленточные черви, колючие черви, и аскариды.
Эктопаразиты — это многоклеточные организмы, которые живут на вашей коже или питаются ею. К ним относятся некоторые насекомые и паукообразные, такие как комары, блохи, клещи и клещи.
Паразитарные инфекции могут распространяться разными путями. Например, простейшие и гельминты могут распространяться через зараженную воду, продукты питания, отходы, почву и кровь. Некоторые из них могут передаваться половым путем. Некоторые паразиты распространяются насекомыми, которые действуют как переносчики или переносчики болезни. Например, малярию вызывают паразитические простейшие, переносимые комарами, когда они питаются людьми.
Любой может заразиться паразитарной инфекцией. Но некоторые люди подвергаются большему риску, чем другие. У вас больше шансов заразиться паразитарной инфекцией, если вы:
- имеете ослабленную иммунную систему или уже болеете другим заболеванием
- живете или путешествуете в тропических или субтропических регионах мира
- не имеете доступа к чистой питьевой воде
- плавать в озерах, реках или прудах, где распространены Giardia или другие паразиты
- работать в сфере ухода за детьми, регулярно работать с почвой или работать в других условиях, где вы постоянно контактируете с фекалиями
Уличные кошки могут контактировать с инфицированными грызунами и птицами. Это делает их владельцев более склонными к заражению токсоплазмозом, типом простейших. Токсоплазмоз может быть очень опасным для беременных женщин и их развивающихся детей. Инфекция распространяется через кошачьи фекалии. Если вы беременны, важно, чтобы кто-то другой ежедневно чистил лоток.
Это делает их владельцев более склонными к заражению токсоплазмозом, типом простейших. Токсоплазмоз может быть очень опасным для беременных женщин и их развивающихся детей. Инфекция распространяется через кошачьи фекалии. Если вы беременны, важно, чтобы кто-то другой ежедневно чистил лоток.
Паразитарные инфекции можно диагностировать несколькими способами. Например, ваш врач может выполнить или заказать:
- Анализ крови
- Анализ кала: при таком исследовании у вас берут образец кала и проверяют на наличие паразитов и их яиц.
- Эндоскопия или колоноскопия: эти анализы могут быть назначены, если результаты исследования кала неубедительны. Пока вы находитесь под действием седативных средств, врач введет тонкую гибкую трубку через рот или прямую кишку в пищеварительную систему, чтобы осмотреть желудочно-кишечный тракт.
- Рентген, магнитно-резонансная томография (МРТ) или компьютерная аксиальная томография (КТ): эти сканирования используются для выявления признаков поражений или повреждений органов, вызванных паразитами.
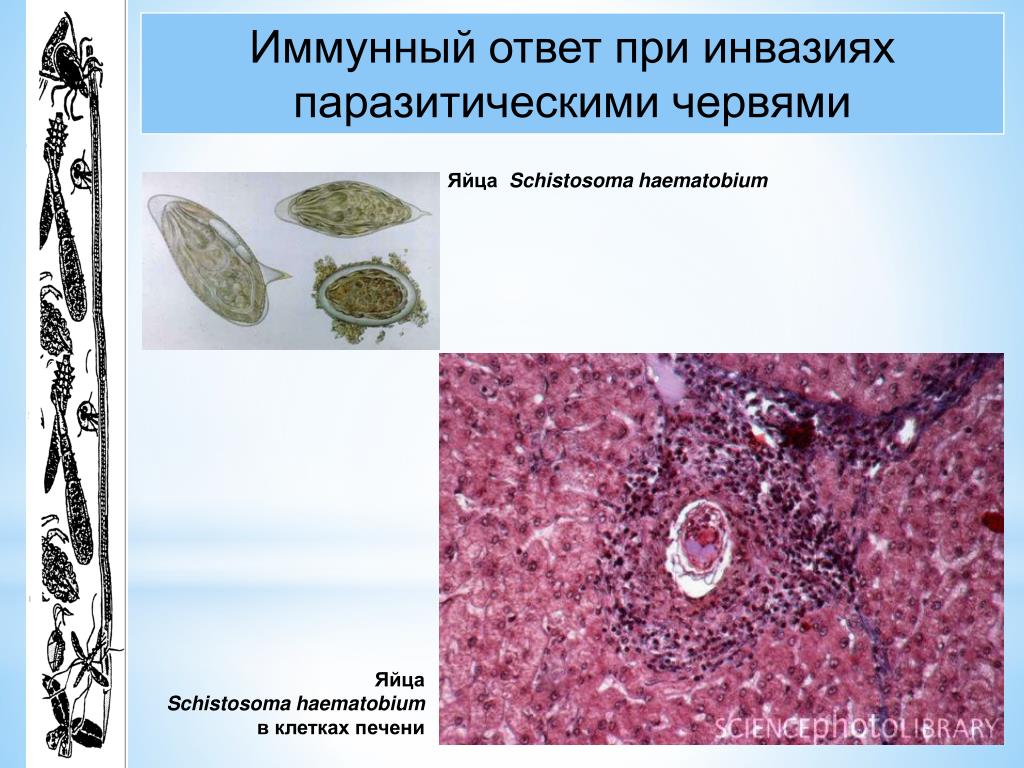
Ваш врач может также назначить анализы для проверки на наличие бактерий или других вещей, которые могут вызывать инфекции.
Ваш план лечения будет зависеть от вашего конкретного диагноза. Как правило, врач прописывает лекарства. Например, они могут прописать лекарства для лечения трихомониаза, лямблиоза или криптоспоридиоза. Они, вероятно, не будут назначать лекарства от токсоплазмоза, если вы не беременны и в остальном здоровы, если только у вас нет тяжелой и продолжительной инфекции.
Ваш врач может также порекомендовать другие методы лечения для облегчения ваших симптомов. Например, многие паразитарные инфекции могут вызывать диарею, которая часто приводит к обезвоживанию организма. Ваш врач, скорее всего, порекомендует вам пить много жидкости, чтобы восполнить потери.
Есть несколько шагов, которые вы можете предпринять, чтобы снизить риск заражения паразитарной инфекцией:
- Практикуйте безопасный секс с использованием презерватива.

- Регулярно мойте руки, особенно после контакта с сырой пищей или фекалиями.
- Приготовьте пищу до рекомендуемой внутренней температуры.
- Во время путешествия пейте чистую воду, в том числе бутилированную.
- Не глотайте воду из озер, ручьев или прудов.
- Избегайте кошачьего туалета и фекалий во время беременности.
Если вы подозреваете у себя паразитарную инфекцию, запишитесь на прием к врачу. Они могут помочь диагностировать причину ваших симптомов и порекомендовать план лечения. Своевременное лечение поможет остановить распространение инфекции среди других людей.
ПАРАЗИТЫ-ПРОСТЕЙШИЕ – Лабораторные занятия по микробиологии
Перейти к содержимому
Определить репрезентативные простейшие патогены
Описать жизненные циклы и уникальные характеристики репрезентативных простейших патогенов
Определите структурные характеристики основных групп микроорганизмов
Сравните и сопоставьте прокариотические и эукариотические клетки
Сравните и сопоставьте физиологию и биохимию различных групп микроорганизмов
Описывать симптомы, сопутствующий возбудитель, передачу, течение, лечение и профилактику распространенных инфекционных заболеваний
Применять различные лабораторные методы для идентификации типов микроорганизмов
Демонстрация под микроскопом:
Entamoeba histolytica трофозоитов и цист – общее увеличение 1000X
Trichomonas vaginalis трофозоитов – 1000X общее увеличение 9001mblia Giardia 9
0020 trophozoites and cysts– 1000X total magnificationBalantidium coli trophozoites and cysts – 1000X total magnification
Plasmodium blood smear – 1000X total magnification
Cryptosporidium parvum oocysts – 1000X total magnification
Trypanosoma blood smear – 1000-кратное общее увеличение
Toxoplasma gondii трофозоиты – 1000-кратное общее увеличение
https://goo. gl/photos/uUb8o3mReoEN5ZMJ9
gl/photos/uUb8o3mReoEN5ZMJ9
Все простейшие находятся на стадии жизни, называемой трофозоитами. Стадия «трофа» активно питается, в основном подвижна и отвечает за симптомы у хозяина. Некоторые простейшие могут образовывать стадию кисты — покоящуюся, неактивную стадию, которая защищает организм из-за образующейся толстой стенки. Эти кисты позволяют простейшим выживать в суровых условиях окружающей среды за пределами хозяина и обеспечивают передачу организма другим хозяевам. Проглатывание кисты с зараженной пищей или водой является распространенным способом заражения многими протозойными инфекциями.
Классификация микроорганизмов динамична и часто меняется по мере поступления новой информации. Простейшие не является официальным термином, используемым в классификации микроорганизмов. Скорее, это термин, используемый для группировки эукариотических организмов, которые являются одноклеточными и не имеют клеточной стенки. Эти организмы часто объединяются в подгруппы в зависимости от их средств передвижения. Некоторые ученые называют эти двигательные группы типами; другие называют их классами. Мы просто сгруппируем эти организмы в соответствии с их способом передвижения, чтобы у нас были общие средства общения, когда мы обсуждаем этих паразитов.
Некоторые ученые называют эти двигательные группы типами; другие называют их классами. Мы просто сгруппируем эти организмы в соответствии с их способом передвижения, чтобы у нас были общие средства общения, когда мы обсуждаем этих паразитов.
Entamoeba histolytica передвигаются, выдавливая цитоплазматические отростки, называемые псевдоножками или «ложными ножками». Во всем мире ежегодно происходит около 50 миллионов случаев инвазивной болезни E. histolytica , что приводит к 100 000 смертей. Это представляет собой верхушку айсберга, потому что только у 10-20% инфицированных людей появляются симптомы. В США это чаще всего наблюдается у путешественников или иммигрантов из тропических стран с плохими санитарными условиями. Во время путешествия пейте воду в бутылках или кипятите воду в течение 1 минуты, чтобы уничтожить паразита. Метронидазол (Флагил) может использоваться для лечения инфекции.
Метронидазол (Флагил) может использоваться для лечения инфекции.
Entamoeba вызывает заболевание, амебную дизентерию или амебиаз. Дизентерия характеризуется кровавой диареей, которая может привести к сильному обезвоживанию организма. Люди заражаются Entamoeba при проглатывании цист с зараженной пищей или водой. Трофы лежат в просвете толстой кишки. Они вторгаются в крипты слизистой оболочки, питаются эритроцитами (эритроциты часто видны внутри окрашенных Entamoeba образцов). Симптомы обычно легкие. В редких случаях организм может образовывать язвы кишечника. Изъязвление стенки кишечника вызывает симптомы дизентерии (кровавый стул, содержащий гной и слизь). Entamoeba мигрирует с кровью в печень, где может вызвать тяжелые абсцессы в течение 2-4 недель.
Трофозоиты можно отличить от цист. Трофозоиты неправильной формы, видны ложноножки. Трофическая форма имеет одно ядро, тогда как циста содержит максимум четыре ядра. Каждое ядро может содержать темную окрашивающуюся точку, называемую кариосомой. Сигарообразные хроматиновые тела также можно увидеть либо на стадии кисты, либо на стадии трофии. Стадия кисты отличается почти идеальной сферической формой.
Сигарообразные хроматиновые тела также можно увидеть либо на стадии кисты, либо на стадии трофии. Стадия кисты отличается почти идеальной сферической формой.
Naegleria fowleri встречается на песчаном дне водоемов с теплой водой, озер и горячих источников. Он возбуждается, когда люди прыгают или ныряют в мелкие водоемы. В соленой воде не встречается. Чаще всего это наблюдается во время засухи, когда уровень воды низок, а деятельность на воде с большей вероятностью поднимает песок и грязь со дна пруда или озера. Считается, что носовые пробки помогают предотвратить инфекцию. Микроорганизм очень инвазивен, и если он попадает высоко в носовые ходы, он может пройти через обонятельный нерв в мозг, где он вызывает смертельную инфекцию, называемую первичным амебным менингоэнцефалитом (ПАМ). Это разрушение мозговой ткани вызывает такие симптомы, как бактериальный менингит, такие как головная боль, лихорадка, тошнота, рвота и ригидность затылочных мышц, а также судороги и галлюцинации. Смерть обычно наступает в течение трех-семи дней после появления симптомов, несмотря на лечение. 9Инфекции 0019 Naegleria fowleri встречаются редко. По данным CDC, с 2011 по 2020 год в США было зарегистрировано 33 случая заражения.
Смерть обычно наступает в течение трех-семи дней после появления симптомов, несмотря на лечение. 9Инфекции 0019 Naegleria fowleri встречаются редко. По данным CDC, с 2011 по 2020 год в США было зарегистрировано 33 случая заражения.
Acanthamoeba — свободноживущая пресноводная амеба, вызывающая глазные инфекции. Восемьдесят пять процентов инфекций в Соединенных Штатах связаны с неправильной чисткой контактных линз. Инфекция может вызвать стойкое нарушение зрения или слепоту. Он также может вызывать инвазивный энцефалит у лиц с ослабленным иммунитетом. Несмотря на то, что все еще можно заразиться, если человек не носит контактные линзы, большинство случаев можно предотвратить путем надлежащего хранения и обращения с контактными линзами – с использованием подходящих чистящих растворов для очистки линз, мытья рук перед контактными линзами, отказа от плавания, использования горячей воды. ношение линз в ванне или душе, ношение и замена линз в соответствии с предписанным графиком, использование чистого раствора для мытья линз, замена футляров для хранения не реже одного раза в три месяца и регулярное обследование глаз. Большинство людей будут подвергаться воздействию Acanthamoeba в течение жизни, но большинство из них не заразятся.
Большинство людей будут подвергаться воздействию Acanthamoeba в течение жизни, но большинство из них не заразятся.
Trichomonas vaginalis не имеет стадии кисты, поэтому она не может долго выживать вне хозяина. Передается незащищенным сексом. Это причина трихомониаза, обычно называемого «пинг-понговым вагинитом», потому что он может передаваться от одного полового партнера к другому. В Соединенных Штатах около 2 миллионов человек инфицированы, но только у 30% проявляются какие-либо симптомы трихомониаза. Хотя большинство инфекций протекают бессимптомно, у мужчин они могут вызывать инфекции предстательной железы и придатка яичка. Женщины жалуются на частое мочеиспускание, зуд, жжение и выделения из влагалища. Диагноз часто ставится, когда люди обращаются за медицинской помощью по поводу того, что, по их мнению, является инфекцией мочевыводящих путей.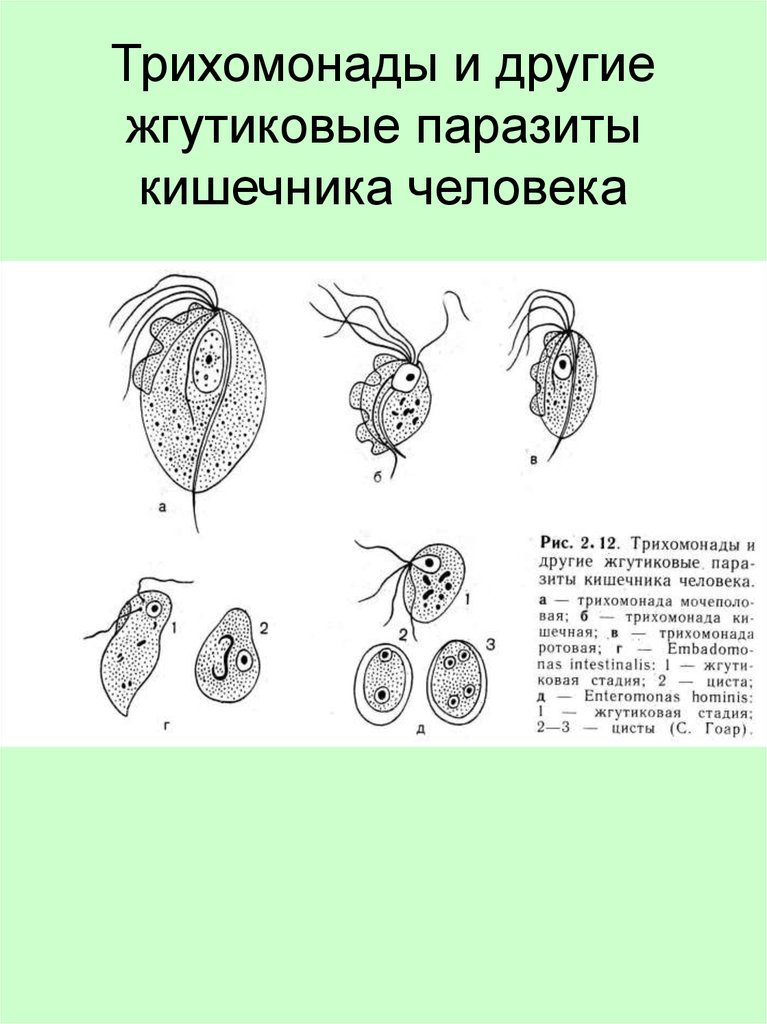 Флагил используется для лечения инфекций. Диагноз обычно ставится при обнаружении трофозоитов каплевидной формы во влажном препарате выделений из влагалища или уретры. На переднем конце клетки могут быть видны от трех до пяти жгутиков в виде пучка. T. vaginalis будет демонстрировать быструю, отрывистую, стремительную подвижность, перемещаясь по полю зрения.
Флагил используется для лечения инфекций. Диагноз обычно ставится при обнаружении трофозоитов каплевидной формы во влажном препарате выделений из влагалища или уретры. На переднем конце клетки могут быть видны от трех до пяти жгутиков в виде пучка. T. vaginalis будет демонстрировать быструю, отрывистую, стремительную подвижность, перемещаясь по полю зрения.
Giardia lamblia уникальны по внешнему виду на стадии трофозоита или цисты. Троф представляет собой грушевидный жгутик с широким закругленным передним и заостренным задним концами. Овальный вогнутый сосательный диск занимает около трех четвертей плоской вентральной поверхности, которая прикрепляется к слизистой оболочке пищеварительного тракта хозяина. Два ядра, каждое с большой центральной кариосомой, напоминающей «глаза на лице», помогают отличить этого паразита от других простейших. Может присутствовать от четырех до пяти пар жгутиков. Стадия кисты имеет овальную форму, и обычно можно наблюдать от двух до четырех ядер.
Может присутствовать от четырех до пяти пар жгутиков. Стадия кисты имеет овальную форму, и обычно можно наблюдать от двух до четырех ядер.
Общее название для Giardia — «паразит туриста», в результате случаев, приобретенных теми, кто пьет воду, не очищая ее. Такие животные, как бобр, лось и олень, могут переносить паразита и откладывать цисты в ручьях при дефекации. Паразит обычно встречается у кошек, собак, овец, оленей и коров. При попадании в организм нового хозяина кисты без повреждений проходят через желудочный сок и эксцистируются в двенадцатиперстной кишке. Таким образом, трофы Giardia обнаруживаются в верхнем отделе тонкой кишки. Инфекция Giardia является наиболее распространенным кишечным паразитарным заболеванием в Соединенных Штатах, которым ежегодно заражаются более 1 миллиона человек. Симптомы часто появляются в течение 1-3 недель после заражения. Кишечные симптомы включают дискомфорт в животе, сильную диарею, метеоризм, мальабсорбцию и потерю веса.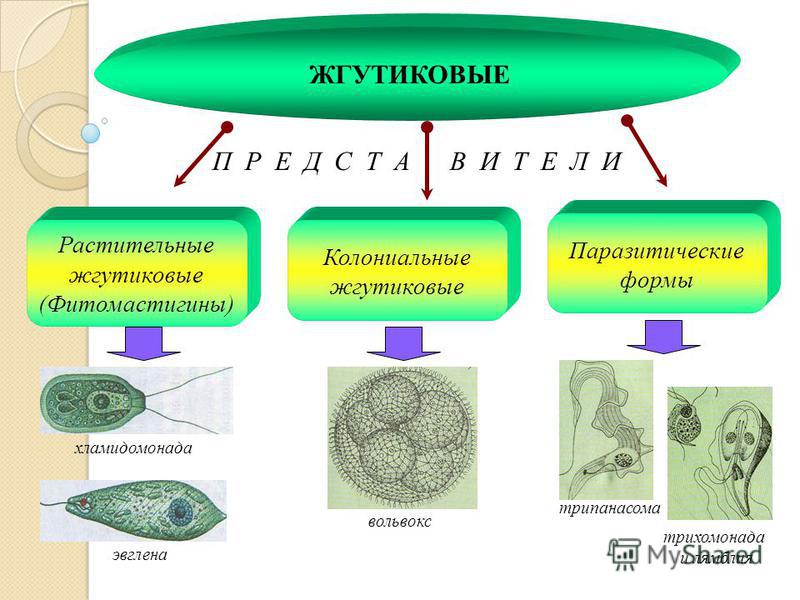 У большинства людей инфекция длится 2-6 недель, Giardia редко кого-то убивает. Кипячение воды в течение пяти-десяти минут или фильтрация воды могут предотвратить заражение. Известно, что Giardia способна выжить после приема таблеток хлора и йода. Есть несколько рецептов, доступных для лечения, в том числе Flagyl.
У большинства людей инфекция длится 2-6 недель, Giardia редко кого-то убивает. Кипячение воды в течение пяти-десяти минут или фильтрация воды могут предотвратить заражение. Известно, что Giardia способна выжить после приема таблеток хлора и йода. Есть несколько рецептов, доступных для лечения, в том числе Flagyl.
Трофозоиты Trypanosoma под микроскопом выглядят идентичными. Трипаносомы перемещаются волнообразным спиральным движением, производимым сократительным жгутиком и ундулирующей мембраной. Они видны между кровяными тельцами в мазках крови инфицированных больных. Трипаносомы не имеют стадии кисты. Поскольку эти паразиты находятся в кровотоке, их называют гемофлагеллятами. Трипаносомы способны к чрезвычайной антигенной изменчивости, которая подвергает стрессу иммунную систему хозяина. Похоже, что Trypanosoma может продуцировать не менее тысячи различных антигенных оболочек.
Trypanosoma трофозоиты Trypanosoma brucei (trip-an’o-so’muh/bru’see) Trypanosoma brucei — простейшее, вызывающее африканскую сонную болезнь.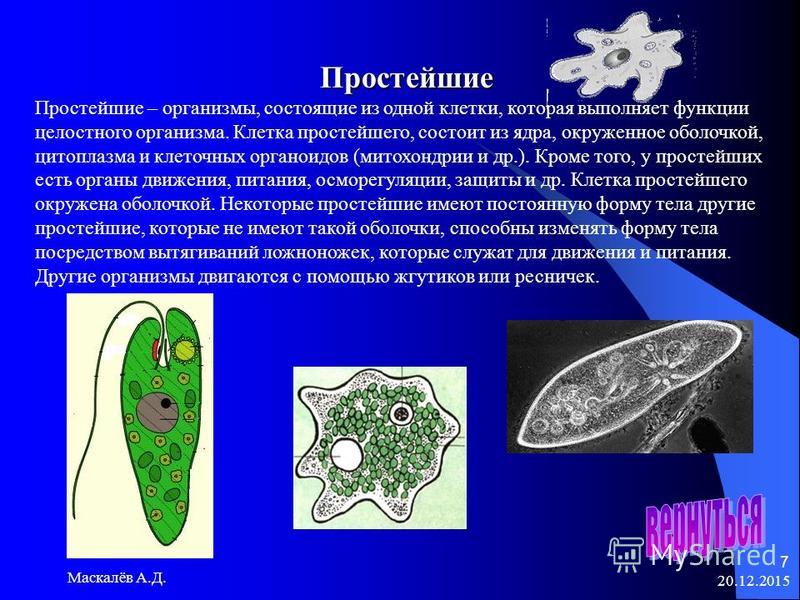 Существует два подвида патогенных Trypanosoma – Trypanosoma brucei gambiense в Западной Африке и Trypanosoma brucei rhodesiense в Восточной Африке. Оба переносятся мухой цеце (вид Glossina ), которая встречается только в сельских районах Африки. Хотя инфекция не встречается в Соединенных Штатах, исторически она была серьезной проблемой общественного здравоохранения в некоторых регионах Африки к югу от Сахары. С 2015 года Всемирная организация здравоохранения ежегодно регистрирует менее 100 случаев заболевания. Сонную болезнь можно вылечить с помощью пентамидина на ранних стадиях, но если ее не лечить, она может привести к летальному исходу.
Существует два подвида патогенных Trypanosoma – Trypanosoma brucei gambiense в Западной Африке и Trypanosoma brucei rhodesiense в Восточной Африке. Оба переносятся мухой цеце (вид Glossina ), которая встречается только в сельских районах Африки. Хотя инфекция не встречается в Соединенных Штатах, исторически она была серьезной проблемой общественного здравоохранения в некоторых регионах Африки к югу от Сахары. С 2015 года Всемирная организация здравоохранения ежегодно регистрирует менее 100 случаев заболевания. Сонную болезнь можно вылечить с помощью пентамидина на ранних стадиях, но если ее не лечить, она может привести к летальному исходу.
T. brucei вводятся хозяину через укус инфицированной мухи цеце. Паразиты размножаются в месте заражения. Затем они проникают в лимфатическую ткань, вызывая симптомы лихорадки, головной боли, тошноты, рвоты и поражения центральной нервной системы, что приводит к коме и смерти. Муха цеце поражает не только людей, но и крупный рогатый скот, лошадей, коз и свиней. Более четырех миллионов квадратных миль Африки непригодны для выращивания домашнего скота из-за присутствия мух цеце.
Более четырех миллионов квадратных миль Африки непригодны для выращивания домашнего скота из-за присутствия мух цеце.
Trypanosoma cruzi — простейшее, вызывающее болезнь Шагаса. T. cruzi ограничен западным полушарием, включая Калифорнию, юг Соединенных Штатов, Центральную и Южную Америку. Чаще всего встречается в сельских районах Латинской Америки, где широко распространена бедность. Вектором T. cruzi является редувиидный жук (целующийся жук). Люди заражаются болезнью Шагаса (южноамериканский трипаносомоз), когда зараженный жук кусает кожу человека и впоследствии испражняется в рану. Инфекция может быть легкой или бессимптомной. Вокруг укуса клопа может быть лихорадка или опухоль. Паразиты также могут быть обнаружены в циркулирующей крови. Болезнь Шагаса возникает сразу после заражения и может длиться несколько недель или месяцев. У 20-30% инфицированных людей острая инфекция может привести к тяжелому воспалению сердечной мышцы или головного мозга и оболочек вокруг головного мозга.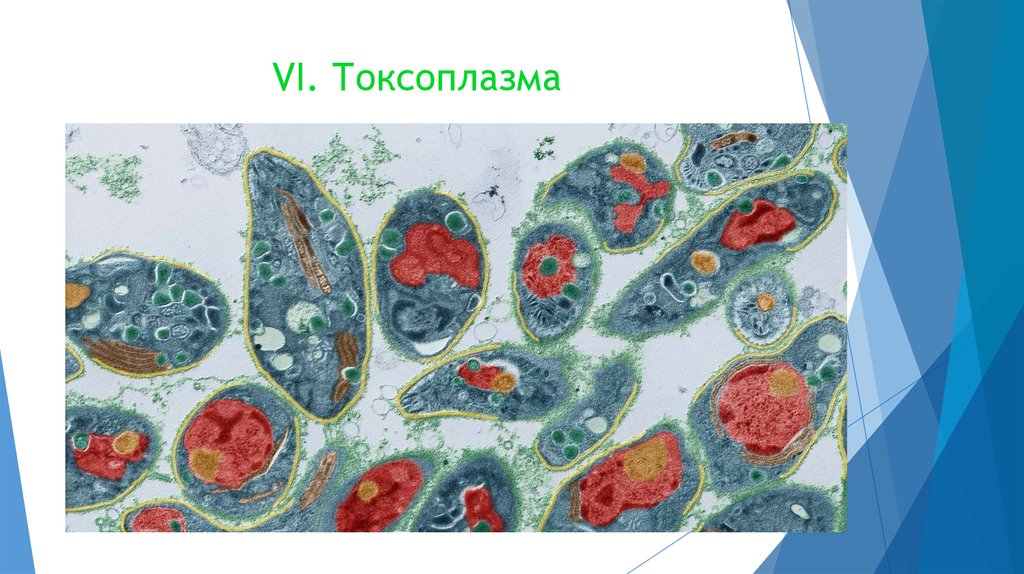
Balantidium coli — единственный известный реснитчатый паразит, инфицирующий человека. Паразит встречается во всем мире, но редко встречается в Соединенных Штатах. Он наиболее распространен в тропических и субтропических регионах, в районах, где разводят свиней, поскольку они являются обычным животным резервуаром. Паразит передается фекально-оральным путем. Наиболее частыми клиническими проявлениями инфекции являются хронические рецидивирующие поносы, чередующиеся с запорами. Balantidium coli обитают в просвете толстой кишки и могут проникать на слизистую кишечника, вызывая изъязвление. Balantidium легко узнать по большому размеру. Трофозит имеет размеры от 50 до 130 мкм в длину и 20-70 мкм в ширину. Это крупнейшее кишечное простейшее человека.
Balantidium coli киста Balantidium coli трофозоит Простейшие без подвижности Плазмодий , Криптоспоридий и Toxoplasma — все роды, сгруппированные как апикомплексы. Организмы этой группы не имеют средств передвижения. Все они облигатные паразиты; свободноживущих форм нет. Их сложные жизненные циклы характеризуются чередованием бесполых и половых циклов у разных хозяев. Хозяева, содержащие форму паразита, размножающегося половым путем, называются окончательными хозяевами. Хозяева, у которых происходит бесполое размножение, называются промежуточными хозяевами.
Организмы этой группы не имеют средств передвижения. Все они облигатные паразиты; свободноживущих форм нет. Их сложные жизненные циклы характеризуются чередованием бесполых и половых циклов у разных хозяев. Хозяева, содержащие форму паразита, размножающегося половым путем, называются окончательными хозяевами. Хозяева, у которых происходит бесполое размножение, называются промежуточными хозяевами.
Plasmodium вызывает малярию, одну из распространенных причин гемолитической анемии во всем мире. Существует четыре вида Plasmodium – P. vivax (наиболее распространенный), P . ovale , P. malariae и P. falciparum (наиболее летальные). Типичными симптомами являются лихорадка, озноб, головная боль, мышечная боль и потливость. По данным ВОЗ, во всем мире регистрируется около 229 миллионов случаев малярии в год, в основном (94%) в Африканском регионе. Случаи малярии, диагностированные в Соединенных Штатах, в основном наблюдались у путешественников и иммигрантов, возвращающихся в США из стран, эндемичных по малярии.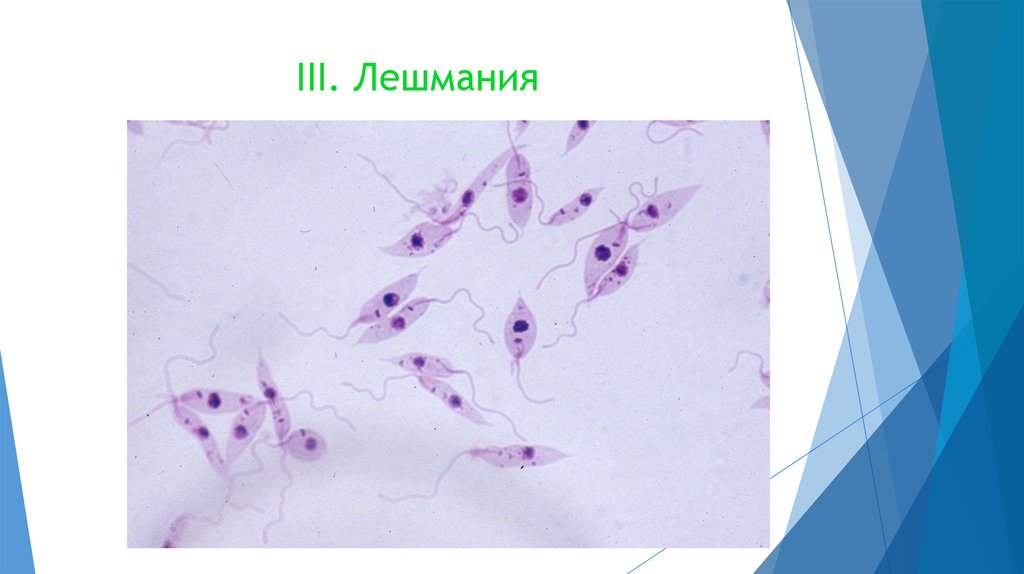
Малярия является основной причиной смерти от паразитов в мире. Для продолжения существования он должен чередовать половые и бесполые циклы. Прерывание любого жизненного цикла будет контролировать болезнь. Меры, принимаемые для прерывания жизненного цикла, включают попытки уничтожить комара Anopheles , защитить хозяина от укусов с помощью химических репеллентов и противомоскитных сеток, профилактическое лечение путешественников в районах повышенного риска, лечение активных случаев различными противопаразитарными препаратами. В последние годы во всем мире резко возросло число устойчивых к лекарственным препаратам штаммов. Контакт с CDC поможет определить наилучший профилактический препарат и препарат выбора для лечения в зависимости от состояния здоровья пациента и региона, в котором он заразился малярией.
Plasmodium – Бесполая стадия у человека
Человек заражается Plasmodium через укус инфицированной самки комара Anopheles . Комар передает спорозоитов Plasmodium через свою слюну, когда он вставляет свой хоботок в кожу человека, чтобы получить кровяную пищу. Кровь обеспечивает питание для яиц, которые она отложит. Введенные в кровоток спорозоиты в течение сорока минут покидают кровеносную систему и проникают в паренхиматозные клетки печени. В клетках печени спорозоиты размножаются бесполым путем. Затем они высвобождаются и проникают в эритроциты, инициируя фазу кровотока инфекции.
Комар передает спорозоитов Plasmodium через свою слюну, когда он вставляет свой хоботок в кожу человека, чтобы получить кровяную пищу. Кровь обеспечивает питание для яиц, которые она отложит. Введенные в кровоток спорозоиты в течение сорока минут покидают кровеносную систему и проникают в паренхиматозные клетки печени. В клетках печени спорозоиты размножаются бесполым путем. Затем они высвобождаются и проникают в эритроциты, инициируя фазу кровотока инфекции.
В эритроцитах инфицированного хозяина происходит бесполый цикл, известный как шизогония. Этот процесс приводит к образованию от четырех до тридцати шести новых паразитов в каждой эритроците. Развиваются незрелые трофозоиты, называемые «кольцевыми» формами, которые затем увеличиваются, превращаясь в зрелые трофозоиты, заполняя большую часть паразитированных эритроцитов. Бесполое размножение происходит при расщеплении ядерного материала и цитоплазмы трофозоитов. В конце шизогонического цикла инфицированные клетки крови разрываются, высвобождая мерозоиты, которые, в свою очередь, заражают новые эритроциты. Лизис эритроцитов высвобождает продукты метаболизма паразитов и эритроцитов. Эти токсичные материалы вызывают симптомы малярии – озноб, лихорадку, тошноту, рвоту и головную боль. Всплески лихорадки происходят с различными интервалами в 24, 48 или 72 часа в зависимости от вида Плазмодий присутствует. Лихорадочный период может длиться несколько часов, заканчиваясь стадией обильного потоотделения. Этот цикл повторяется много раз.
Лизис эритроцитов высвобождает продукты метаболизма паразитов и эритроцитов. Эти токсичные материалы вызывают симптомы малярии – озноб, лихорадку, тошноту, рвоту и головную боль. Всплески лихорадки происходят с различными интервалами в 24, 48 или 72 часа в зависимости от вида Плазмодий присутствует. Лихорадочный период может длиться несколько часов, заканчиваясь стадией обильного потоотделения. Этот цикл повторяется много раз.
Plasmodium – Половая стадия комара Anopheles
В эритроцитах также могут появляться мужские и женские гаметоциты половой стадии. Когда комар кусает зараженного человека, он набирает в желудок кровь, которая может содержать мужские и женские гаметоциты. В кишечнике комара мужские гаметоциты образуют сперматозоиды, а женские – яйцеклетку. Происходит оплодотворение. Образовавшаяся зигота может внедряться в стенку кишечника и продуцировать многочисленные спорозоиты. Спорозоиты мигрируют через ткани комара в слюнные железы, где они будут введены следующему хозяину-человеку, когда комар снова примет кровь.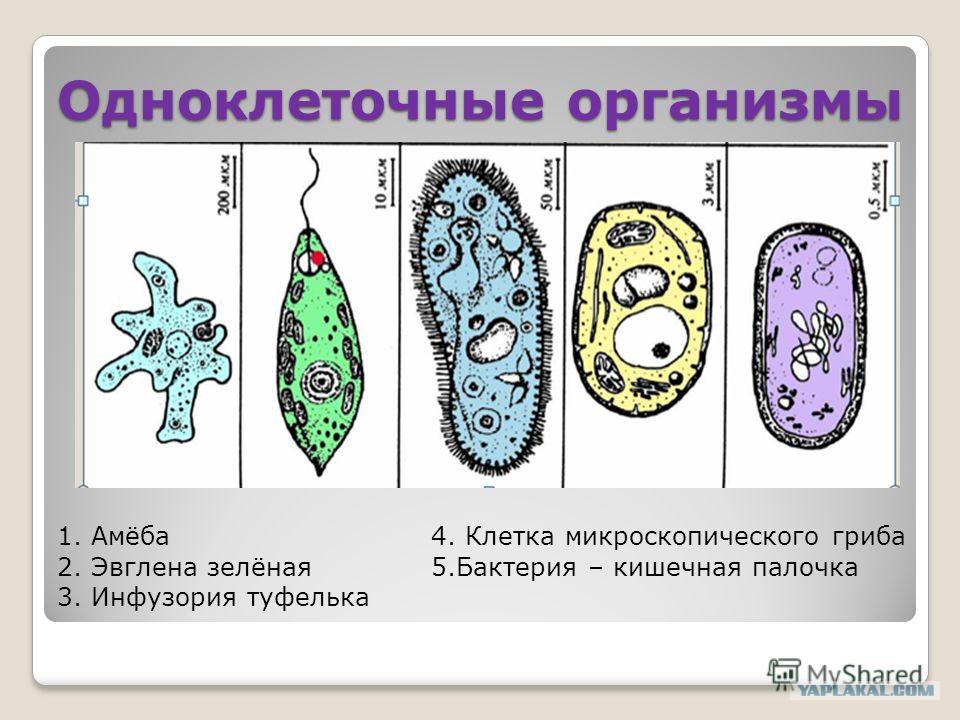 Затем бесполый цикл продолжается у нового хозяина.
Затем бесполый цикл продолжается у нового хозяина.
Toxoplasma gondii встречается во всем мире, и более 40 миллионов человек в США инфицированы, хотя у очень немногих из них есть симптомы. Иммунная система здорового человека может удерживать паразита от заражения. В группу риска развития тяжелых инфекций входят беременные женщины, которые могут передать инфекцию своему будущему ребенку, и лица с ослабленным иммунитетом. В то время как женщина обычно протекает бессимптомно, у плода могут развиться тяжелые врожденные дефекты, такие как ретинохориоидит, умственная отсталость, часто из-за гидроцефалии или «воды в мозгу», судороги, слепота и смерть. T. gondii был признан известным оппортунистом, который может вызывать смерть у больных СПИДом, поскольку они страдают от последствий ослабленной иммунной системы. Хотя большинству людей лечение не требуется, симптоматические инфекции у людей, возникающие не у беременных женщин, можно лечить пириметамином и сульфадиазином.
Наиболее распространенным путем передачи Toxoplasma является проглатывание тканевой формы паразита с недоваренным, зараженным мясом (особенно свинина, баранина и оленина) или случайное проглатывание через посуду или обращение с сырым мясом. Мытье рук и достаточное приготовление мяса перед употреблением являются лучшей профилактикой.
Другая форма передачи может быть через инфицированные кошачьи фекалии. Кошки могут заразиться Toxoplasma , поедая инфицированную мышь или крысу. Кошка является окончательным хозяином и обычно не проявляет симптомов. Трофозоиты Toxoplasma размножаются половым путем в кишечном тракте кошки. Ооцисты, содержащие восемь спорозоитов, выделяются с фекалиями кошек и могут загрязнять пищу или воду, чем могут быть проглочены другими животными. Если ооцисты проглатываются человеком (или любым млекопитающим или птицей), спорозоиты будут размножаться бесполым путем в тканях мозга, глаз или скелетных мышц нового хозяина.
Беременным женщинам рекомендуется избегать всех ненужных контактов с кошками, особенно с их лотками. Ооцистам требуется не менее 24 часов после прохождения для образования спор. Таким образом, один из потенциальных методов минимизации воздействия T . gondii спорозоитов – ежедневная смена кошачьего туалета. Как только ооцисты образуют споры и становятся инфекционными, они могут оставаться инфекционными в почве до 18 месяцев. Ящики для мусора можно дезинфицировать кипятком. Детские песочницы должны иметь крышку, чтобы кошки не испражнялись в них.
Диагностика токсоплазмоза часто основывается на серологическом измерении продукции антител. Повышение титра антител будет свидетельствовать о текущей инфекции. При наличии ткани или жидкости, предположительно содержащих паразитов, их можно привить лабораторным мышам, которые очень восприимчивы к инфекции. На положительный результат указывает фактическое выделение токсоплазмы из мышей или выработка у них антител к токсоплазме . Toxoplasma встречается в жидкостях организма в виде серповидных организмов.
Toxoplasma встречается в жидкостях организма в виде серповидных организмов.
Cryptosporidium , впервые обнаруженный у людей в 1976 году, был признан одной из наиболее распространенных причин заболеваний, передающихся через воду (вода для отдыха и питьевая вода) у людей в Соединенных Штатах. Паразит встречается во всех регионах США и во всем мире. Паразит может заразить каждое семейство позвоночных и может легко преодолевать барьеры видов-хозяев. Передача от человека к человеку Cryptosporidium , вероятно, встречается часто. Сообщалось о вспышках криптоспоридиоза среди детей в детских садах, больницах и после случаев заражения через воду. Вспышки произошли после воздействия в бассейнах и аквапарках. Недавно общественные бассейны и аквапарки в районе Феникса были закрыты из-за заражения воды Cryptosporidium .
Исследование, проведенное в начале 1990-х годов, показало, что ооцист Cryptosporidium parvum присутствуют в 65-95% воды в США. Имели место крупные вспышки из-за загрязненного общественного водоснабжения. Вспышка в штате Мэн, связанная с яблочным сидром, была вызвана зараженным теленком, пасущимся в районе, где яблоки собирали с земли. Один инфицированный теленок может выделять до 10 миллиардов ооцист за один день! Ооцисты могут оставаться активными до 18 месяцев при хранении в куче навоза.
Имели место крупные вспышки из-за загрязненного общественного водоснабжения. Вспышка в штате Мэн, связанная с яблочным сидром, была вызвана зараженным теленком, пасущимся в районе, где яблоки собирали с земли. Один инфицированный теленок может выделять до 10 миллиардов ооцист за один день! Ооцисты могут оставаться активными до 18 месяцев при хранении в куче навоза.
Cryptosporidium вторгается и размножается в эпителиальных клетках, выстилающих пищеварительный и дыхательный тракты человека. Округлые ооцисты подвергаются спорогенезу внутри клетки-хозяина и сразу же становятся инфекционными при попадании в фекалии. Каждая зрелая ооциста содержит четыре спорозоита. Инфекция распространяется фекально-оральным путем – ооцисты передаются с фекалиями одного зараженного животного и попадают в организм другого.
Болезнь, вызванная Cryptosporidium , называется криптоспоридиозом. Наиболее распространенным симптомом является водянистая диарея, но другие симптомы включают спазмы желудка, обезвоживание, тошноту и рвоту.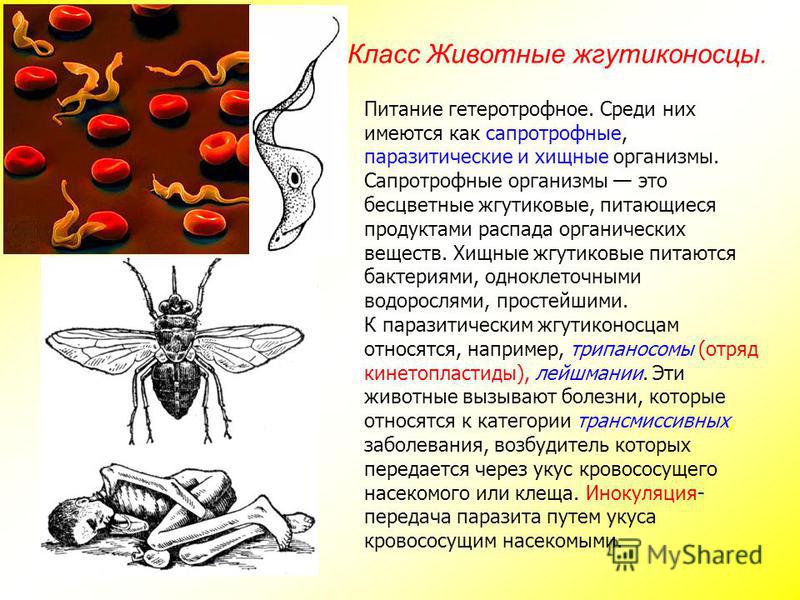 Считается, что это серьезная причина заболеваемости и смертности, особенно у детей младшего возраста, живущих в развивающихся странах. У людей со здоровой иммунной системой диарея длится от трех до двенадцати дней, однако кисты могут выделяться в течение 2 недель после исчезновения симптомов. У лиц с иммунодефицитом, таких как больные СПИДом, могут возникнуть опасные для жизни холероподобные заболевания. Криптоспоридиоз является одной из наиболее важных оппортунистических инфекций больных СПИДом.
Считается, что это серьезная причина заболеваемости и смертности, особенно у детей младшего возраста, живущих в развивающихся странах. У людей со здоровой иммунной системой диарея длится от трех до двенадцати дней, однако кисты могут выделяться в течение 2 недель после исчезновения симптомов. У лиц с иммунодефицитом, таких как больные СПИДом, могут возникнуть опасные для жизни холероподобные заболевания. Криптоспоридиоз является одной из наиболее важных оппортунистических инфекций больных СПИДом.
Толстая стенка ооцисты делает Cryptosporidium чрезвычайно устойчивым к большинству распространенных дезинфицирующих средств. Они могут выжить в неразбавленном отбеливателе в течение нескольких часов и выдержат стандартное хлорирование питьевой воды. Наиболее эффективным способом предотвращения заражения является микрофильтрация воды с использованием фильтров с размером пор не более 0,2 мкм.
Диагноз устанавливается при обнаружении ооцист в образцах стула инфицированных пациентов.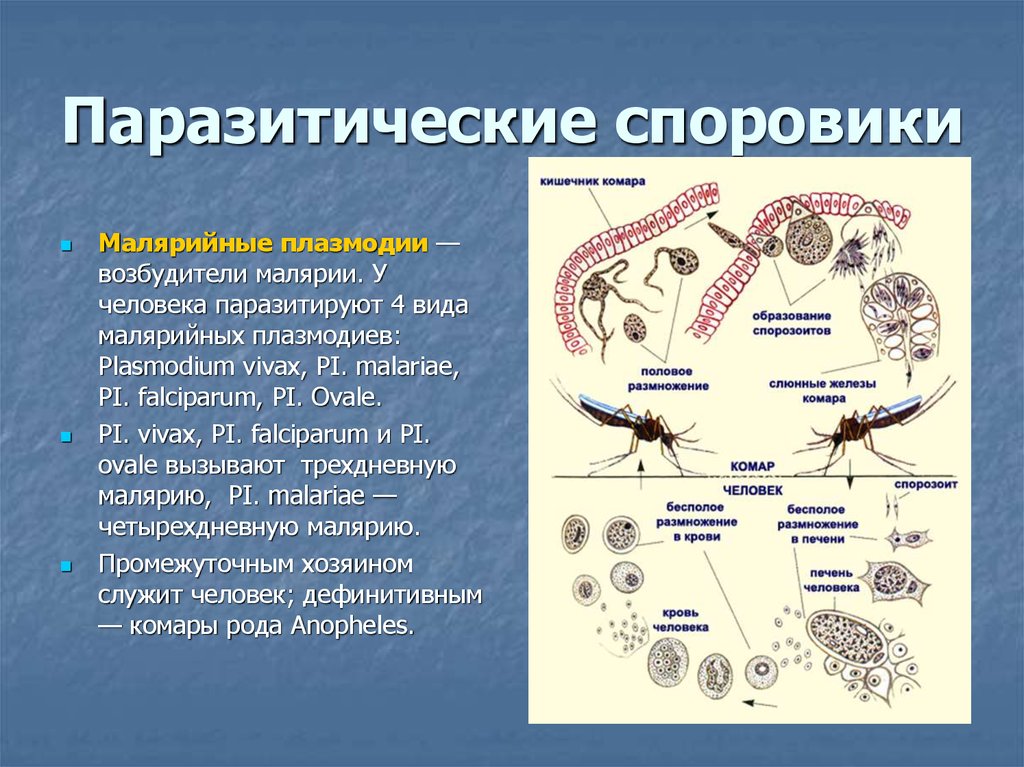 Ооцисты трудно увидеть при обычном окрашивании. Можно использовать модифицированный кислотостойкий краситель, при котором окрашенные ооцисты выглядят ярко-красными на сине-зеленом фоне. Иногда в стенке ооцисты можно увидеть четыре спорозоита. Хотя большинство здоровых людей выздоравливают без лечения, нитазоксанид является препаратом выбора для лечения криптоспоридиоза.
Ооцисты трудно увидеть при обычном окрашивании. Можно использовать модифицированный кислотостойкий краситель, при котором окрашенные ооцисты выглядят ярко-красными на сине-зеленом фоне. Иногда в стенке ооцисты можно увидеть четыре спорозоита. Хотя большинство здоровых людей выздоравливают без лечения, нитазоксанид является препаратом выбора для лечения криптоспоридиоза.
Нарисуйте на рабочем листе демонстрацию работы микроскопа.
ДР. БРЮС ЭЙМС
Вы предпочитаете покупать продукты, не тестированные на животных? Тест Эймса был разработан американским биохимиком доктором Брюсом Эймсом в 1970-х годах для определения того, является ли химическое вещество мутагеном. В тесте Эймса используется мутантный штамм Salmonella , который не может продуцировать аминокислоту гистидин. Если химическое вещество обращает мутацию вспять, это мутаген Сальмонелла . Химические вещества, которые являются мутагенными по тесту Эймса, могут быть дополнительно протестированы на животных для оценки их способности вызывать мутации/рак у людей.
Лицензия
Лабораторные упражнения по микробиологии Энн Мейсон М.С. и Джилл Рэймонд, доктор философии. находится под лицензией Creative Commons Attribution-NonCommercial 4.0 International License, если не указано иное.
Поделиться этой книгой
Поделиться в Твиттере
Интегративные подходы к пониманию мастерства в манипулировании цитокиновыми сетями хозяина простейшими паразитами с акцентом на виды Plasmodium и Leishmania
Введение
Паразитические простейшие являются причиной некоторых основных заболеваний человека, от которых ежегодно страдают несколько миллионов человек, что приводит к значительной заболеваемости и смертности, а также к снижению экономической активности. Были достигнуты определенные успехи в снижении заболеваемости этими заболеваниями благодаря более совершенным стратегиям вмешательства, но в отсутствие эффективных вакцин такие болезни, как малярия, лейшманиоз, трипаносомоз, по-прежнему представляют собой серьезную проблему для общественного здравоохранения. Эти простейшие обычно заражаются хроническими инфекциями, подтверждая их успех в уклонении и манипулировании защитой хозяина и метаболическими процессами для их выживания, размножения и передачи. Многие из этих патогенных простейших адаптировались к внутриклеточной среде обитания, как это видно при инфекциях, вызванных Plasmodium spp., Leishmania spp. и другие. Внутриклеточная ниша делает их уязвимыми для лизосомальных ферментов, активных промежуточных продуктов кислорода и обнаружения цитозольными сенсорами инфекции, но также обеспечивает некоторую защиту от адаптивного иммунитета (1). Это динамическое взаимодействие хозяина и патогена приводит к активации ряда внутриклеточных и межклеточных биохимических сигнальных процессов, ведущих к синтезу диффундирующих эффекторных молекул, которые включают цитокины и активные формы кислорода. «Самые ранние стадии инфекции — это первая возможность паразита закрепиться внутри своего хозяина, и, наоборот, это также шанс хозяина быстро и эффективно отреагировать на излечение или, по крайней мере, контроль над инфекцией» (2).
Эти простейшие обычно заражаются хроническими инфекциями, подтверждая их успех в уклонении и манипулировании защитой хозяина и метаболическими процессами для их выживания, размножения и передачи. Многие из этих патогенных простейших адаптировались к внутриклеточной среде обитания, как это видно при инфекциях, вызванных Plasmodium spp., Leishmania spp. и другие. Внутриклеточная ниша делает их уязвимыми для лизосомальных ферментов, активных промежуточных продуктов кислорода и обнаружения цитозольными сенсорами инфекции, но также обеспечивает некоторую защиту от адаптивного иммунитета (1). Это динамическое взаимодействие хозяина и патогена приводит к активации ряда внутриклеточных и межклеточных биохимических сигнальных процессов, ведущих к синтезу диффундирующих эффекторных молекул, которые включают цитокины и активные формы кислорода. «Самые ранние стадии инфекции — это первая возможность паразита закрепиться внутри своего хозяина, и, наоборот, это также шанс хозяина быстро и эффективно отреагировать на излечение или, по крайней мере, контроль над инфекцией» (2). Недавние исследования показывают, что патогены, в том числе простейшие, модулируют среду клетки-хозяина, манипулируя транскриптомом хозяина посредством эпигенетических модификаций, помимо нацеливания на основные сигнальные узлы метаболических, иммунных процессов и процессов клеточного цикла, чтобы способствовать их росту, размножению и выживанию (3–9).). Многие простейшие секретируют эффекторные молекулы, которые активно модулируют иммунный транскриптом хозяина, изменяя уровни цитокинов и сигнализируя либо о том, чтобы избежать иммунных процессов, как на стадиях печени P. falciparum , либо стимулировать их рост, как это наблюдается на стадиях крови этого патогена.
Недавние исследования показывают, что патогены, в том числе простейшие, модулируют среду клетки-хозяина, манипулируя транскриптомом хозяина посредством эпигенетических модификаций, помимо нацеливания на основные сигнальные узлы метаболических, иммунных процессов и процессов клеточного цикла, чтобы способствовать их росту, размножению и выживанию (3–9).). Многие простейшие секретируют эффекторные молекулы, которые активно модулируют иммунный транскриптом хозяина, изменяя уровни цитокинов и сигнализируя либо о том, чтобы избежать иммунных процессов, как на стадиях печени P. falciparum , либо стимулировать их рост, как это наблюдается на стадиях крови этого патогена.
Учитывая разнообразие патогенеза простейших, в этом обзоре основное внимание будет уделено манипулированию и захвату сетей цитокинов штаммами Leishmania и Plasmodium spp. для их выживания в организме человека. Мы выделим несколько недавно опубликованных репрезентативных исследований, основанных на омике и системной биологии, на паразитов Leishmania и Plasmodium , чтобы понять модуляцию сетей цитокинов и хемокинов в хозяине паразитом для достижения их персистенции в хозяине посредством иммунного ускользания.
Цитокины и регуляция цитокинов
Цитокины представляют собой небольшие молекулы иммунной системы, синтезируемые различными типами клеток, которые благодаря связыванию со своими рецепторами, присутствующими на множестве клеток, опосредуют активацию, дифференцировку и взаимодействие иммунных клеток для поддержания иммунного гомеостаза (10, 11). Синтез и регуляция экспрессии цитокинов зависит от типа стимула, типа клетки и состояния ее активации (12–14). Экспрессия генов цитокинов также регулируется эпигенетическими модификациями, которые включают метилирование ДНК, модификации гистонов и взаимодействия хроматина более высокого порядка (15, 16), а также посттранскрипционную регуляцию с помощью механизмов, опосредованных микроРНК (16–19).). Дифференцировка иммунных клеток как в субпопуляциях Т-клеток, так и в фенотипах макрофагов определяется и регулируется цитокиновым окружением (4, 16, 20, 21) и эпигенетическими модификациями в локусах генов цитокинов (22, 23). Отмечено, что перекрестное взаимодействие цитокинов между IFNα/β и TNF-α происходит на уровне хроматина, где IFN в дополнение к регуляции генов, передающих сигналы интерферона, также потенцируют гены TNF (4). Точно так же новые данные свидетельствуют об обширных перекрестных взаимодействиях между белками семейства NLR воспалительного комплекса для секреции IL-1β и IL-18 и другими интегрированными сигналосомами цитокинов, облегчающими интеграцию различных путей для оптимального иммунного ответа (24). Сообщается, что h4K27, энхансер метилтрансферазы гомолога 1 zeste, способствует продукции воспалительных цитокинов, запускаемой TLR, путем подавления TLR-негативного регулятора toll-взаимодействующего белка, тем самым способствуя полной активации врожденного иммунного ответа против вторгающихся патогенов (25).
Точно так же новые данные свидетельствуют об обширных перекрестных взаимодействиях между белками семейства NLR воспалительного комплекса для секреции IL-1β и IL-18 и другими интегрированными сигналосомами цитокинов, облегчающими интеграцию различных путей для оптимального иммунного ответа (24). Сообщается, что h4K27, энхансер метилтрансферазы гомолога 1 zeste, способствует продукции воспалительных цитокинов, запускаемой TLR, путем подавления TLR-негативного регулятора toll-взаимодействующего белка, тем самым способствуя полной активации врожденного иммунного ответа против вторгающихся патогенов (25).
Управление передачей сигналов цитокинов простейшими патогенами
Внутриклеточные простейшие модулируют экспрессию генов цитокинов и передачу сигналов с помощью некоторых общих тем, которые включают нацеливание на факторы транскрипции (15, 23), статус фосфорилирования сигнальных молекул, таких как STAT, молекул иммунных контрольных точек, таких как CTLA-4 и PD-1 управляет регуляторными путями (26), а также киназами (5, 6, 27).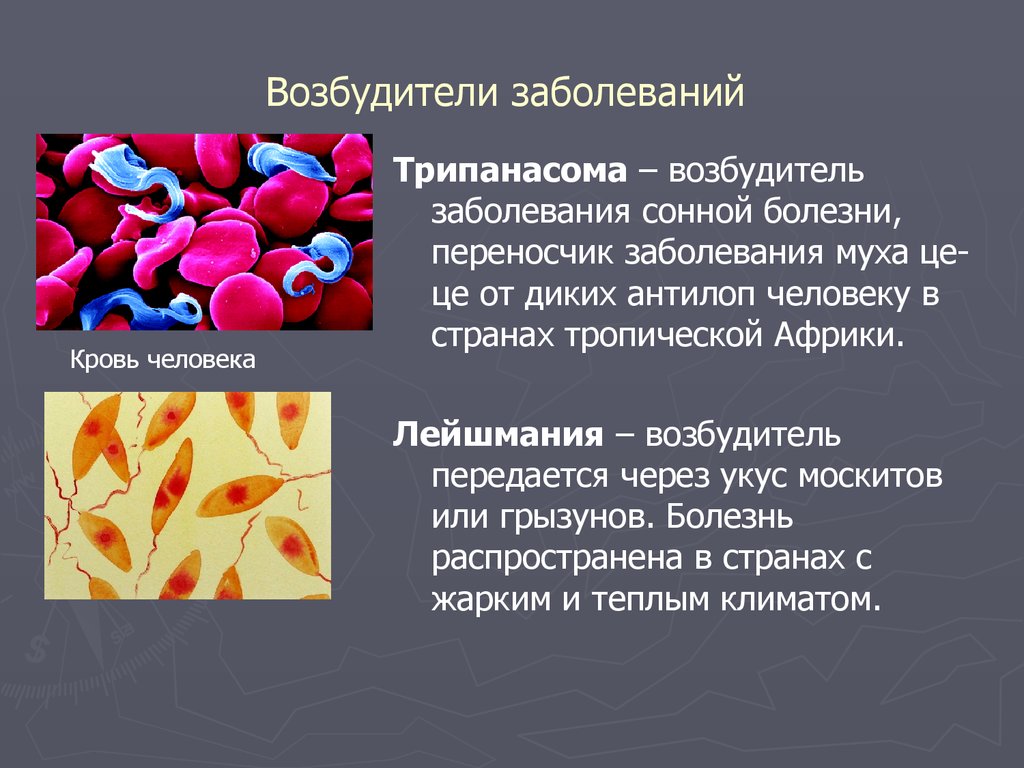 Пути, на которые обычно нацелены патогены, включают NF-κB, клеточный цикл, интерфероны, MAP-киназу JAK-STAT и пути, опосредованные рецепторами TLR и NLR из-за их широкого спектра функций и основной связи с геномом хозяина (28–30).
Пути, на которые обычно нацелены патогены, включают NF-κB, клеточный цикл, интерфероны, MAP-киназу JAK-STAT и пути, опосредованные рецепторами TLR и NLR из-за их широкого спектра функций и основной связи с геномом хозяина (28–30).
Toxoplasma spp. секретируют плотный гранулярный белок (GRA) и белки Rhoptry, которые активируют киназы хозяина и обладают киназной активностью, соответственно, в клетку-хозяина, которые путем фосфорилирования STAT3 и STAT6, ядерной транслокации NF-κB или состояния активации путей MAPK модулируют уровни IL- 4, ИЛ-6, ИЛ-12 и ИФН-g (31–35). «Ингибитор транскрипции STAT1 T. gondii представляет собой еще один секреторный белок, который задействует комплекс ремоделирования нуклеосом и деацецитлазы хозяина для блокирования транскрипции генов, опосредованной STAT1» (36). Trypanasoma cruzi модулирует путь NF-κB с помощью TLR и опосредованной NLR передачи сигналов для благоприятного окружения цитокинов (37–39). , 41) и аргиназы для его выживания и репликации.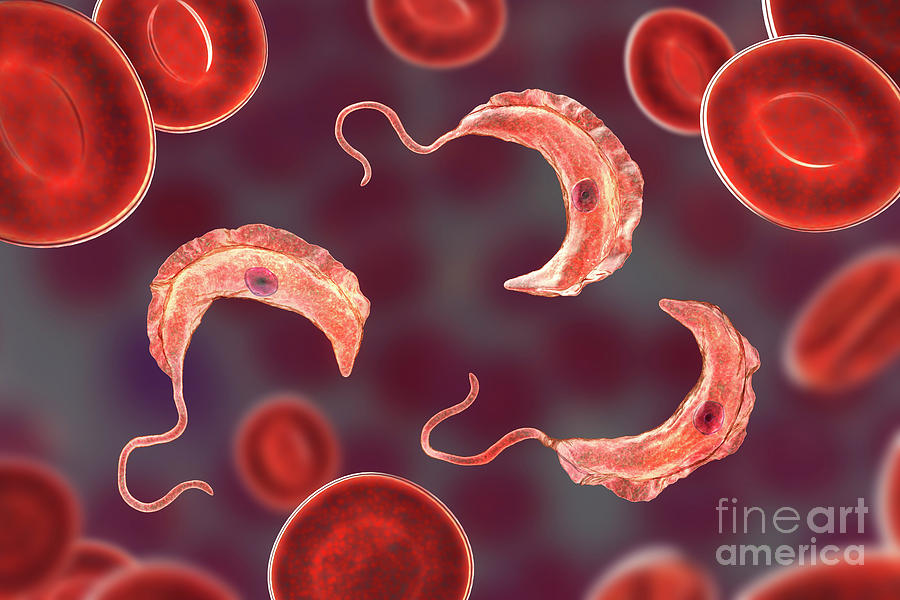
Малярия, вызванная Plasmodium spp. Apicomplexa phylum, была сильнейшей эволюционной селективной силой в новейшей истории человечества, сформировала геном человека (42) и является одной из основных причин смертности детей в возрасте до 5 лет, особенно в Африканском регионе ВОЗ, унося жизнь ребенка каждые 2 мин (43). Жизненный цикл паразита сложен и проходит в несколько стадий у человека и у комара (самка 9).0157 Anopheles spp.) со специфическими для стадии генами и белковыми сигнатурами (44). Вкратце, спорозоиты, привитые человеку-хозяину укусом инфицированного комара, попадают в печень, где созревают в мерозоиты, которые заражают эритроциты, чтобы продолжить бесполый цикл, а также развиваются в гаметоциты, которые после оплодотворения в кишечнике комара развиваются и созревают в спорозоиты.
На печеночных стадиях паразита иммунный ответ хозяина имеет тенденцию быть толерогенным, а циркумспорозоитный белок, как было замечено, ингибирует НАДФН-оксидазу и ИЛ-12 и подавляет секрецию ИЛ-6 и ФНО-α с одновременным повышением уровней ИЛ-10, позволяя паразиту избежать обнаружения иммунной системой (45, 46).
Воспаление признано основным признаком иммунного ответа на кровяные стадии инфекции Plasmodium (47). Примечательно, что клинические проявления заболевания связаны с эритроцитарной стадией инфекции. Ранний и точно сбалансированный воспалительный ответ с повышением уровней провоспалительных ИЛ-12, ИФН-γ, ФНО-α, ИЛ-1β и ИЛ-6, а также противовоспалительных ИЛ-10 и ТФР-β необходим для разрешение паразитемии и болезни (48–52). Однако патологическая активация повышенных уровней тех же самых провоспалительных цитокинов (цитокиновый шторм) в сочетании с более низкими уровнями регуляторных механизмов связывают с синдромами тяжелой и церебральной малярии (14, 53–57). В недавнем исследовании изучались уровни различных биомаркеров иммунного ответа и были обнаружены высокие концентрации sCDI63 и фракталкина, которые участвуют в подавлении иммунного ответа и модуляции противовоспалительных реакций при бессимптомной малярии (58). Эти авторы также сообщили о высоких уровнях неоптерина, что связано с усилением клеточно-опосредованного иммунного ответа и активацией макрофагов у пациентов с тяжелой и церебральной малярией, что указывает на общее устойчивое состояние воспаления, подтверждающее гипотезу об интенсивной и продолжительной воспалительной реакции при тяжелой и церебральной малярии. больных малярией.
больных малярией.
Тогда возникает вопрос: почему и как паразит может вызвать интенсивную воспалительную реакцию, которая потенциально может привести к летальному исходу, что может ограничить передачу паразита и, следовательно, не в интересах патогена? Ответ, по-видимому, заключается в (а) усиленной экспрессии молекул адгезии на эндотелиальных клетках провоспалительными цитокинами (IFNγ и TNFα) (59) и (b) потребностью в эндотелиальной адгезии, опосредованной мембранным белком 1 P. falciparum ( PfEMP1) с CD36 и рецептором эндотелиального белка С (EPCR) (60, 61). С точки зрения паразитов секвестрация эндотелия необходима, чтобы избежать клиренса в селезенке и облегчить falciparum созревание мерозоитов. Очень разнообразные белки PfEMP1, кодируемые генами паразита var , содержат домены, подобные Даффи-связыванию, и междоменные области, богатые цистеином (CIDR). Большинство доменов CIDRα1 связываются с EPCR, а CIDRα2–6 связываются с CD36 (60, 61). Примечательно, что взаимодействие EPCR с его лигандом, активированным протеином C (APC), играет роль в противовоспалительных функциях, коагуляционном гомеостазе и функциях защиты эндотелиального барьера (62), а его блокада этих функций взаимодействием PfEMP1-EPCR, что постулируется, способствует к церебральной малярийной патологии (59, 61). Интересно, что Смит и др. (61) обнаружили повышенную связь тяжелой формы малярии с изолятами, содержащими домен CIDRα1, связывающий EPCR, что подтверждает данное утверждение. Сообщается также, что взаимодействия с CD36 ингибируют синтез IL-12 и подавляют созревание дендритных клеток (ДК) и активацию Т-клеток.
Интересно, что Смит и др. (61) обнаружили повышенную связь тяжелой формы малярии с изолятами, содержащими домен CIDRα1, связывающий EPCR, что подтверждает данное утверждение. Сообщается также, что взаимодействия с CD36 ингибируют синтез IL-12 и подавляют созревание дендритных клеток (ДК) и активацию Т-клеток.
Таким образом, не исключено, что паразит манипулирует NF-κB и интерфероновым путем 1-го типа, чтобы вызвать воспаление. PAMP, полученные из Plasmodium , которые включают якоря GPI, мотивы CpG, мотивы, богатые AT, и гемазоин, обнаруживаются PRR хозяина, которые включают TLR, NLR и AIM2 на клетках линии моноцитов/макрофагов и на DC (61, 63–63). 65). Эти взаимодействия лиганд-рецептор инициируют опосредованную MyD88 и STING-IRF3 передачу сигналов вниз по течению, что приводит к активации путей NF-κB и IRF3 и синтезу провоспалительных цитокинов и интерферона α/β (55, 65–68). Именно преувеличенная активация этих путей, «опосредованная провоспалительным праймингом IFN-γ с экстремальными уровнями провоспалительных медиаторов» с сопутствующей потерей регуляторных цитокинов, управляет патогенезом малярии (46, 57, 68). Также было высказано предположение, что, помимо стимулирования воспаления, P. falciparum путем подавления экспрессии GATA3 подавляет IL-10 и SOCS3, которые необходимы для контроля воспаления, возможно, за счет использования пути IFNα/β, как показано на рисунке 1.
Также было высказано предположение, что, помимо стимулирования воспаления, P. falciparum путем подавления экспрессии GATA3 подавляет IL-10 и SOCS3, которые необходимы для контроля воспаления, возможно, за счет использования пути IFNα/β, как показано на рисунке 1.
Рисунок 1 . Гипотетическая модель, обобщающая вероятные механизмы тяжелого воспаления при малярии. Молекулы паразитов, такие как гемазоин, ДНК, богатая Pf AT, рекрутируются TLR и TLR независимыми (STING) путями (63, 69). Высокая нагрузка ДНК, богатой Pf AT, может привести к повышенным уровням TRAF3 и IFN-α. И IFN-α, в свою очередь, подавляет экспрессию GATA3 в клетках Th3, что приводит к низким уровням IL-10 и, следовательно, к подавлению SOCS3 (68). Кроме того, низкие уровни IL-2 и T-bet не могут опосредовать переключение с IFN-γ+/IL-10- на IFN-γ+/IL-10+ Th2-клеток, что требует уровней T-bet и IL-2. также объясняют низкие уровни IL-10. Наконец, пониженная регуляция SOCS3, который, как известно, опосредует противовоспалительные функции IL-10, не в состоянии регулировать усиленный провоспалительный ответ. Еще одним фактором, способствующим тяжелому воспалению при малярии, является высокая распространенность IL-8-251T/A, который увеличивает экспрессию IL-8 для усиленного рекрутирования и активации нейтрофилов воспалительных клеток, что приводит к повышенной активации NF-κB через IL-1β-опосредованный путь.
Еще одним фактором, способствующим тяжелому воспалению при малярии, является высокая распространенность IL-8-251T/A, который увеличивает экспрессию IL-8 для усиленного рекрутирования и активации нейтрофилов воспалительных клеток, что приводит к повышенной активации NF-κB через IL-1β-опосредованный путь.
Лейшманиоз, вызываемый Leishmania spp. является проблемой общественного здравоохранения: во всем мире зарегистрировано 1,3 миллиона случаев лейшманиоза, что усугубляется наличием нескольких эффективных лекарств (70) и вакцин (71, 72). Будучи внутриклеточным паразитом, он должен преодолевать механизмы сопротивления хозяина и использовать среду хозяина для выживания. В контексте паразитов метаболизм Leishmania обладает уникальной метаболической организацией, которая может перенаправлять метаболиты, поглощение которых ограничено в различных средах хозяина в сторону синтеза специфических метаболитов биомассы; тем самым обеспечивая новые механизмы метаболической адаптации (73, 74). С точки зрения хозяина, вклад специфических факторов вирулентности в подавление иммунитета или неспособность хозяина генерировать достаточный иммунный ответ против паразита, что способствует заражению. Стратегия выживания Leishmania предназначен для модулирования сигнальных путей макрофагов после проникновения в фаголизосому. В зависимости от типа инфекции и паразитарной нагрузки генерируются либо заживляющие Th-1, либо незаживающие Th-2 иммунные ответы, но подробный механизм плохо изучен. Это можно в значительной степени понять в отношении взаимодействия молекул паразита с сигнальными путями хозяина для подавления иммунитета хозяина против инфекции (71).
С точки зрения хозяина, вклад специфических факторов вирулентности в подавление иммунитета или неспособность хозяина генерировать достаточный иммунный ответ против паразита, что способствует заражению. Стратегия выживания Leishmania предназначен для модулирования сигнальных путей макрофагов после проникновения в фаголизосому. В зависимости от типа инфекции и паразитарной нагрузки генерируются либо заживляющие Th-1, либо незаживающие Th-2 иммунные ответы, но подробный механизм плохо изучен. Это можно в значительной степени понять в отношении взаимодействия молекул паразита с сигнальными путями хозяина для подавления иммунитета хозяина против инфекции (71).
Во время инвазии поверхностные молекулы Leishmania взаимодействуют с белками толл-подобных рецепторов, присутствующими на мембране макрофагов (75). Активация TLR запускает нижестоящие сигнальные пути, такие как путь MAPK, опосредованный RAS-RAF, канонический и неканонический путь NF-κB, путь JAK-STAT, путь PI3K-PLC Gamma и путь JNK (76). Впоследствии несколько факторов транскрипции, таких как ERK1/2, NF-kB, NFAT, AP1, STAT3, активируются, что инициирует синтез и секрецию нескольких цитокинов, факторов роста, хемокинов и антимикробных молекул, ответственных за иммунный ответ хозяина во время инфекции (77).
Впоследствии несколько факторов транскрипции, таких как ERK1/2, NF-kB, NFAT, AP1, STAT3, активируются, что инициирует синтез и секрецию нескольких цитокинов, факторов роста, хемокинов и антимикробных молекул, ответственных за иммунный ответ хозяина во время инфекции (77).
Однако при хронической инфекции (рис. 2) антигенные молекулы паразита Leishmania активируют белки фосфатазы в макрофагах, например, SHP-1 и PTP1B, что приводит к дефосфорилированию и дезактивации отдельных сигнальных путей (78). ). Это приводит к подавлению экспрессии iNOS и оксида азота в инфицированных макрофагах, тем самым нарушая микробицидные функции клетки и создавая состояние подавления иммунитета, благоприятное для продолжения выживания патогена внутри АПК. Одновременно резко снижается продукция цитокинов, таких как IL-12 и TNF-α. Такие изменения в картине экспрессии цитокинов антигенпрезентирующих клеток приводят к изменению фенотипических ответов Т-клеток, которые теперь начинают проявлять смещение в сторону незаживающего иммунного ответа Th-2, который характеризуется повышенной продукцией Цитокины IL-4, IL-10, IL-13 и TGF-β (79) и подавление IFN-γ, который регулирует реакцию заживления Th-1 (71).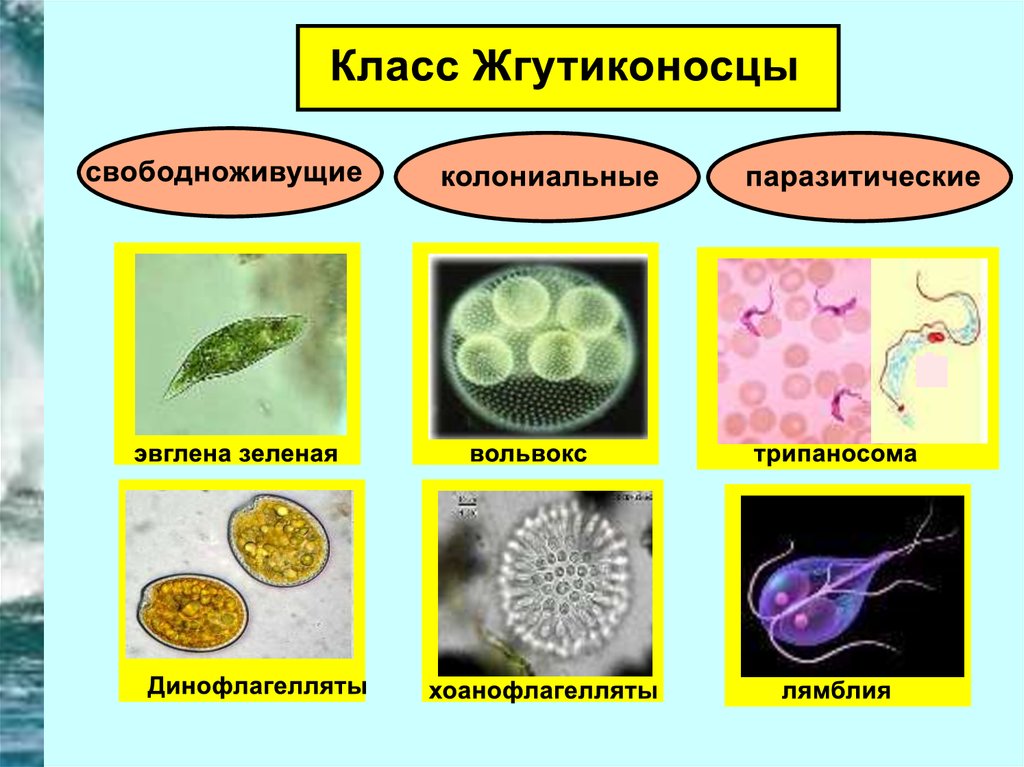 Факторы транскрипции T-bet и GATA3 играют ключевую роль в регуляции соотношения Th-1/Th-2 во время инфекции (80). Leishmania также ингибирует способность клетки-хозяина к презентации антигена другим иммунным клеткам путем подавления экспрессии генов MHC класса II (81) и путем модулирования взаимодействия костимулирующих молекул B7-1/CD28 (82) и CD40/CD40L (83).
Факторы транскрипции T-bet и GATA3 играют ключевую роль в регуляции соотношения Th-1/Th-2 во время инфекции (80). Leishmania также ингибирует способность клетки-хозяина к презентации антигена другим иммунным клеткам путем подавления экспрессии генов MHC класса II (81) и путем модулирования взаимодействия костимулирующих молекул B7-1/CD28 (82) и CD40/CD40L (83).
Рисунок 2 . Иммуномодуляция паразитом Leishmania : антигены Leishmania препятствуют сигнальному каскаду макрофага и способствуют невосстановительной реакции Th-2, которая способствует выживанию паразита внутри хозяина.
Различия в антигенной провокации хозяина приводят к различиям в экспрессии белков макрофагов, как это наблюдается при висцеральных и кожных инфекциях (84). Разница в профиле экспрессии белков макрофагов, о чем свидетельствует повышенная продукция COX2 и продукции PGE2 в случае 9Инфекция 0157 L. donovani (в отличие от L. major ) (85) указывает на то, что разные виды Leishmania избирательно активируют или ингибируют различные пути хозяина из-за различий в антигенной нагрузке. Кроме того, в исследовании было отмечено, что L. donovani , известный как возбудитель висцерального лейшманиоза, в редких случаях может вызывать кожный лейшманиоз (86). Такое поведение инфекции L. donovani может быть связано с устойчивостью хозяина к заболеванию, которое ограничивает распространение инфекции на внутренние органы и удерживает ее локализованной в кожных областях (86).
Кроме того, в исследовании было отмечено, что L. donovani , известный как возбудитель висцерального лейшманиоза, в редких случаях может вызывать кожный лейшманиоз (86). Такое поведение инфекции L. donovani может быть связано с устойчивостью хозяина к заболеванию, которое ограничивает распространение инфекции на внутренние органы и удерживает ее локализованной в кожных областях (86).
Регуляторные Т-клетки CD4+ CD25+ также играют важную роль в регуляции персистенции паразита L. major внутри хозяина. Ингибирование стимулирующих T-reg цитокинов, таких как IL-10, приводит к элиминации патогена у хозяина (87). Однако при лейшманиозе низкая продукция цитокинов IFN-γ и IL-12 приводит к повышенной пролиферации клеток T-reg, что приводит к повторной активации паразитов Leishmania внутри хозяина (87).
Интегративные подходы, основанные на системной биологии, для понимания взаимодействия хозяина и паразита и закономерностей коэволюции при протозойных заболеваниях
Во время взаимодействия хозяев и простейших паразитов оба оказывают взаимное селективное давление друг на друга, что может способствовать быстрой взаимной адаптации.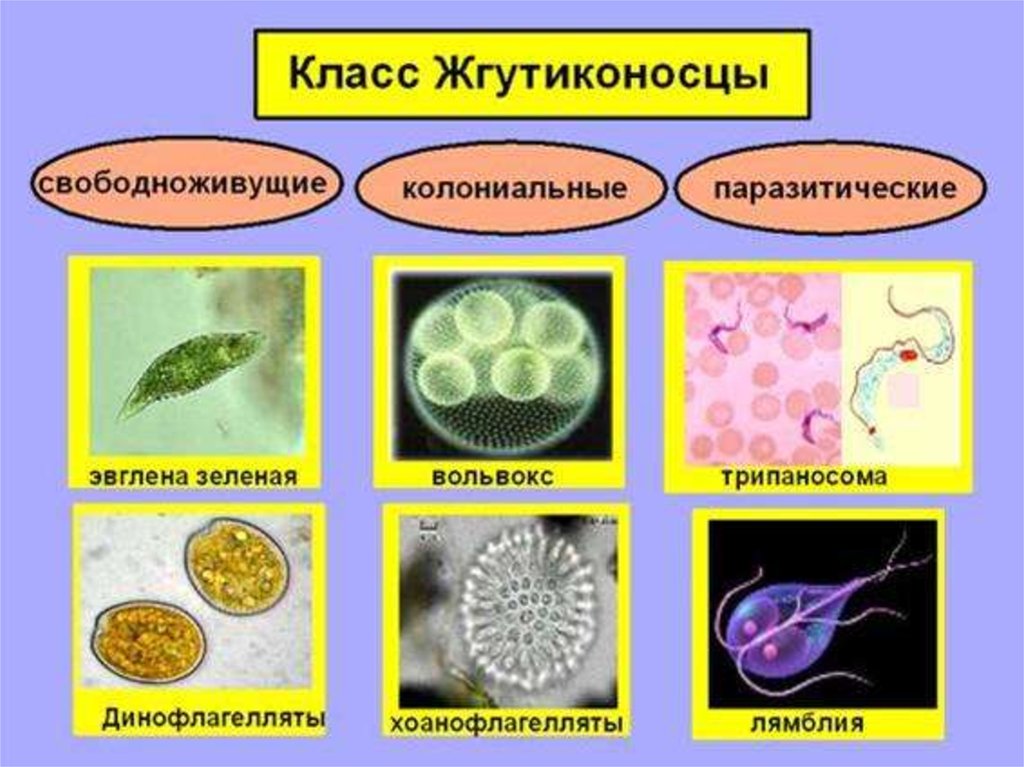 Различные стадии жизненного цикла паразита представляют собой еще один уровень сложности (88). Значительное количество молекулярных, омических, клинических, эпидемиологических, а также экологических данных было получено в 9 лет.0157 in vitro и in vivo уровней с использованием различных патогенов и соответствующих заболеваний. Интегративный анализ таких дискретно сгенерированных и локализованных данных о хозяине и вариантах простейших паразитов в лабораторных, а также естественных популяциях является наиболее важной необходимостью для выявления сложных механизмов эволюции простейших посредством иммунного ускользания во время коэволюции хозяин-паразит. Публичные ресурсы, такие как EuPathDB (89), Pathogen-Host Interactions (90), ProtozoaDB (91) вместе с базами данных по видам простейших чрезвычайно полезны для сбора полезной информации для инициирования комплексного анализа на основе систем. Ключевые этапы такого интегративного подхода включают генерацию/сбор данных, организацию данных, интеграцию данных, интегративное построение сети, сетевой анализ и, наконец, компьютерное математическое моделирование и прогнозное моделирование (92).
Различные стадии жизненного цикла паразита представляют собой еще один уровень сложности (88). Значительное количество молекулярных, омических, клинических, эпидемиологических, а также экологических данных было получено в 9 лет.0157 in vitro и in vivo уровней с использованием различных патогенов и соответствующих заболеваний. Интегративный анализ таких дискретно сгенерированных и локализованных данных о хозяине и вариантах простейших паразитов в лабораторных, а также естественных популяциях является наиболее важной необходимостью для выявления сложных механизмов эволюции простейших посредством иммунного ускользания во время коэволюции хозяин-паразит. Публичные ресурсы, такие как EuPathDB (89), Pathogen-Host Interactions (90), ProtozoaDB (91) вместе с базами данных по видам простейших чрезвычайно полезны для сбора полезной информации для инициирования комплексного анализа на основе систем. Ключевые этапы такого интегративного подхода включают генерацию/сбор данных, организацию данных, интеграцию данных, интегративное построение сети, сетевой анализ и, наконец, компьютерное математическое моделирование и прогнозное моделирование (92). Например, с использованием реконструированной модели метаболизма генома Leishmania infantum 9.0158 адаптаций (73) определили устойчивость метаболической сети паразита к случайным ошибкам и продемонстрировали широкий выбор паразитов для достижения оптимального выживания (73).
Например, с использованием реконструированной модели метаболизма генома Leishmania infantum 9.0158 адаптаций (73) определили устойчивость метаболической сети паразита к случайным ошибкам и продемонстрировали широкий выбор паразитов для достижения оптимального выживания (73).
Недавний прогресс в методах, основанных на RNA-Seq, облегчил одновременное секвенирование транскриптомов как хозяина, так и паразита (включая немодельных паразитов) (93). В первом в своем роде эксперименте с секвенированием РНК в контрольных нейтрофилах человека во время праймирования провоспалительными цитокинами (TNF-α и GM-CSF) Wright et al. показали быструю экспрессию обычного набора транскриптов цитокинов, хемокинов и рецепторов клеточной поверхности (CXCL1, CXCL2, IL1A, IL1B, IL1RA, ICAM1) (9).4). Они продемонстрировали полезность этого подхода для определения функциональных изменений в нейтрофилах после воздействия цитокинов. Во время крупномасштабного анализа 116 пациентов с малярией, заразивших 90 157 паразитов P. falciparum 90 158, Yamagishi et al. выявили переменное поведение полевых малярийных паразитов, которое было гораздо более сложным, чем поведение, наблюдаемое в лабораторных условиях (95). Питтман и др. создали крупномасштабный интерактом T. gondii -хозяин, используя двойное профилирование транскрипции мышей и паразита во время острой и хронической инфекции (96) продемонстрировать влияние развития паразита на транскрипцию генов хозяина, а также эпигенетическое влияние среды хозяина на транскрипцию генов паразита. В различных общесистемных исследованиях малярийных паразитов сообщалось о посттранскрипционном (97) и трансляционном (98) контроле на различных этапах жизненного цикла паразита. Одним из таких контролирующих механизмов является задержка трансляции, при которой экспрессия белка в паразите активно приостанавливается для экспрессируемых транскриптов мРНК. Это было показано в P. falciparum , что, подавляя более 30% своих генов, паразит быстро адаптируется к новым условиям внутри хозяина, оставаясь незамеченным для иммунной системы хозяина, и подвергается переключению в процессе развития, чтобы выжить (99).
falciparum 90 158, Yamagishi et al. выявили переменное поведение полевых малярийных паразитов, которое было гораздо более сложным, чем поведение, наблюдаемое в лабораторных условиях (95). Питтман и др. создали крупномасштабный интерактом T. gondii -хозяин, используя двойное профилирование транскрипции мышей и паразита во время острой и хронической инфекции (96) продемонстрировать влияние развития паразита на транскрипцию генов хозяина, а также эпигенетическое влияние среды хозяина на транскрипцию генов паразита. В различных общесистемных исследованиях малярийных паразитов сообщалось о посттранскрипционном (97) и трансляционном (98) контроле на различных этапах жизненного цикла паразита. Одним из таких контролирующих механизмов является задержка трансляции, при которой экспрессия белка в паразите активно приостанавливается для экспрессируемых транскриптов мРНК. Это было показано в P. falciparum , что, подавляя более 30% своих генов, паразит быстро адаптируется к новым условиям внутри хозяина, оставаясь незамеченным для иммунной системы хозяина, и подвергается переключению в процессе развития, чтобы выжить (99).
Заключение и перспективы на будущее
Стратегии нападения, используемые простейшим патогеном при инфекциях человека, явно различаются. В отличие от этого, по-видимому, существует широкий консенсус в отношении основных сигнальных узлов, которыми манипулируют патогены. Было бы целесообразно проанализировать взаимодействия хозяина и патогена на клеточном, молекулярном и системном уровне, чтобы отличить вирулентные инфекции с потенциалом летального исхода от бессимптомных или неосложненных инфекций с ограниченной заболеваемостью. Можно предположить, что иммунорегуляторные механизмы, обеспечивающие толерантность к заболеваниям, отличаются от иммунных и метаболических реакций на тяжелые заболевания и требуют определения в крупных глобальных исследованиях с использованием различных систем простейших патогенов. Однако, несмотря на наличие огромного количества многомерных данных о взаимодействии хозяина и простейших, функциональная характеристика и аннотация геномов паразитов сильно ограничены из-за отсутствия как генетических инструментов, так и ресурсов у простейших. Учитывая размер, неоднородность и сложность данных о взаимодействии хозяина и паразита, разработка новых вычислительных инструментов и удобных для пользователя методов интеграции разнородных «больших данных» облегчит восполнение недостающих звеньев. Это будет полезно для лучшего понимания эволюционной гонки вооружений между хозяином и паразитом и, наконец, для эффективного лечения и контроля протозойных заболеваний у людей.
Учитывая размер, неоднородность и сложность данных о взаимодействии хозяина и паразита, разработка новых вычислительных инструментов и удобных для пользователя методов интеграции разнородных «больших данных» облегчит восполнение недостающих звеньев. Это будет полезно для лучшего понимания эволюционной гонки вооружений между хозяином и паразитом и, наконец, для эффективного лечения и контроля протозойных заболеваний у людей.
Авторские вклады
SB: дизайн рукописи и участие во введении и разделах о цитокинах, малярии и токсоплазме и перспективах на будущее, PB: участие в разделе, посвященном интегративным подходам, основанным на системной биологии, для понимания взаимодействия паразит-хозяин и перспектив на будущее, RRS: внес вклад в разделы по лейшманиозу и Trypanasoma , AM: внес вклад в раздел о малярии, NS: внес вклад в раздел о цитокинах и регуляции цитокинов, SP: внес вклад в раздел о Toxoplasma , MB: внес вклад в раздел Toxoplasma и PG: внес вклад в разделы лейшманиоз и трипанасома . SP и MB: участвовали в разделе, посвященном манипулированию сигналами цитокинов простейшими паразитами.
SP и MB: участвовали в разделе, посвященном манипулированию сигналами цитокинов простейшими паразитами.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Рецензент DN и главный редактор заявили о своей совместной принадлежности.
Благодарности
Мы хотели бы поблагодарить всех участников исследования и клинических сотрудников за грант и инфраструктурную поддержку Комиссии по университетским грантам. Это исследование финансировалось Департаментом биотехнологии (№ грантов: BT/22/NE/2011, BT/CP/11/NE/TBP/2010 и BT/PR14958/BID/7/537/2015).
Финансирование
Эта работа была поддержана Институциональным биотехнологическим центром Департамента биотехнологии (№ гранта: BT/22/NE/2011) и Департаментом биотехнологии (№ гранта: BT/CP/11/NE/TBP/2010) . Эта работа поддержана грантом, предоставленным RRS Департаментом биотехнологии, проект правительства Индии [BT/PR14958/BID/7/537/2015].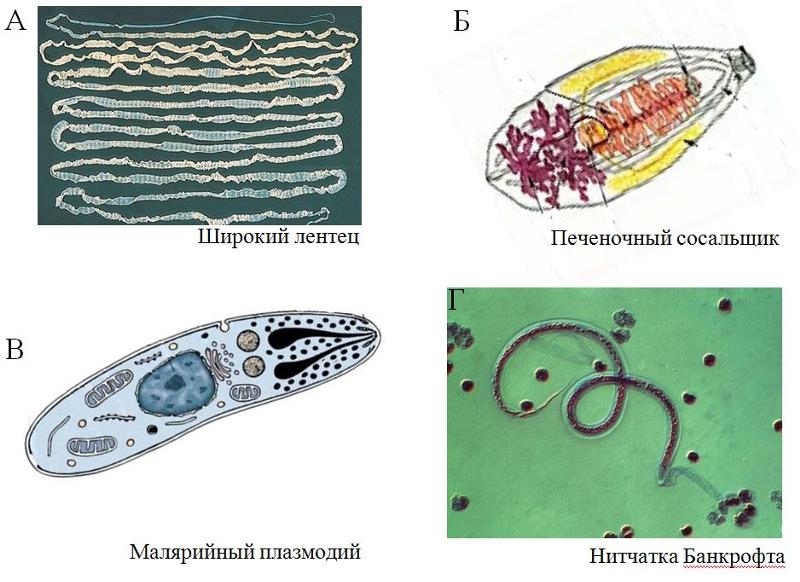 Авторы также отмечают поддержку инфраструктуры со стороны Комиссии по университетским грантам. PG признателен за получение стипендии для младших исследователей от CSIR, Индия». и SB выражает благодарность за клиническую поддержку профессору Какати, AMC&H, Дибругарх.
Авторы также отмечают поддержку инфраструктуры со стороны Комиссии по университетским грантам. PG признателен за получение стипендии для младших исследователей от CSIR, Индия». и SB выражает благодарность за клиническую поддержку профессору Какати, AMC&H, Дибругарх.
Ссылки
1. Сакс Д., Шер А. Уклонение от врожденного иммунитета паразитическими простейшими. Nat Immunol (2002) 3:1041–7. doi:10.1038/ni1102-1041
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
2. Макговерн К.Э., Уилсон Э.Х. Темная сторона освещена: изображение Toxoplasma gondii на протяжении десятилетий. Векторы-паразиты (2013) 6:334. doi:10.1186/1756-3305-6-334
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
3. де Монерри NCS, Ким К. Патогены захватывают эпигеном Новый взгляд на взаимодействие хозяина и патогена. Am J Pathol (2014) 184:897–911. doi:10.1016/j.ajpath.2013.12.022
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
4. Park SH, Kang K, Giannopoulou E, Qiao Y, Kang K, Kim G, et al. Интерфероны типа I и цитокин TNF совместно перепрограммируют эпигеном макрофагов, способствуя активации воспаления. Nat Immunol (2017) 18:1104. doi:10.1038/ni.3818
Park SH, Kang K, Giannopoulou E, Qiao Y, Kang K, Kim G, et al. Интерфероны типа I и цитокин TNF совместно перепрограммируют эпигеном макрофагов, способствуя активации воспаления. Nat Immunol (2017) 18:1104. doi:10.1038/ni.3818
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
5. Бугдур А., Тардье И., Хакими М.А. Toxoplasma экспортирует белки плотных гранул за пределы вакуоли в ядро клетки-хозяина и перестраивает экспрессию генома хозяина. Cell Microbiol (2014) 16:334–43. doi:10.1111/cmi.12255
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
6. Шнайдер А.Г., Абдаллах Д.С.А., Бутчер Б.А., Денкерс Э.Ю. toxoplasma gondii запускает фосфорилирование и ядерную транслокацию STAT1 дендритных клеток, одновременно блокируя транскрипционную активность STAT1, индуцированную гамма-интерфероном. PLoS One (2013) 8. doi:10.1371/journal.pone.0060215
CrossRef Полный текст | Google Scholar
7. Эсколл П., Мондино С., Роландо М., Букризер С. Нацеливание патогенных бактерий на органеллы хозяина: сложная стратегия подрывной деятельности. Nat Rev Microbiol (2016) 14:5–19. doi:10.1038/nrmicro.2015.1
Эсколл П., Мондино С., Роландо М., Букризер С. Нацеливание патогенных бактерий на органеллы хозяина: сложная стратегия подрывной деятельности. Nat Rev Microbiol (2016) 14:5–19. doi:10.1038/nrmicro.2015.1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
8. Hanein D. Toxoplasma gondii нацеливается на актиновый цитоскелет хозяина во время инвазии, рисунок GO. Biophys J (2013) 104:140a. doi:10.1016/j.bpj.2012.11.796
Полный текст CrossRef | Google Scholar
9. Lee SH, Lee BY, Min DY, Kim JM, Ahn MH. Роль цитоскелета в инвазии клеток-хозяев внутриклеточными простейшими Токсоплазма гондии . J Microbiol Biotechnol (2002) 12:628–34.
Google Scholar
10. Джонс М.Дж., Фейес А.П., Кобор М.С. Метилирование ДНК, генотип и экспрессия генов: кто за рулем, а кто за рулем? Геном Биол (2013) 14:126. doi:10.1186/gb-2013-14-7-126
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
11.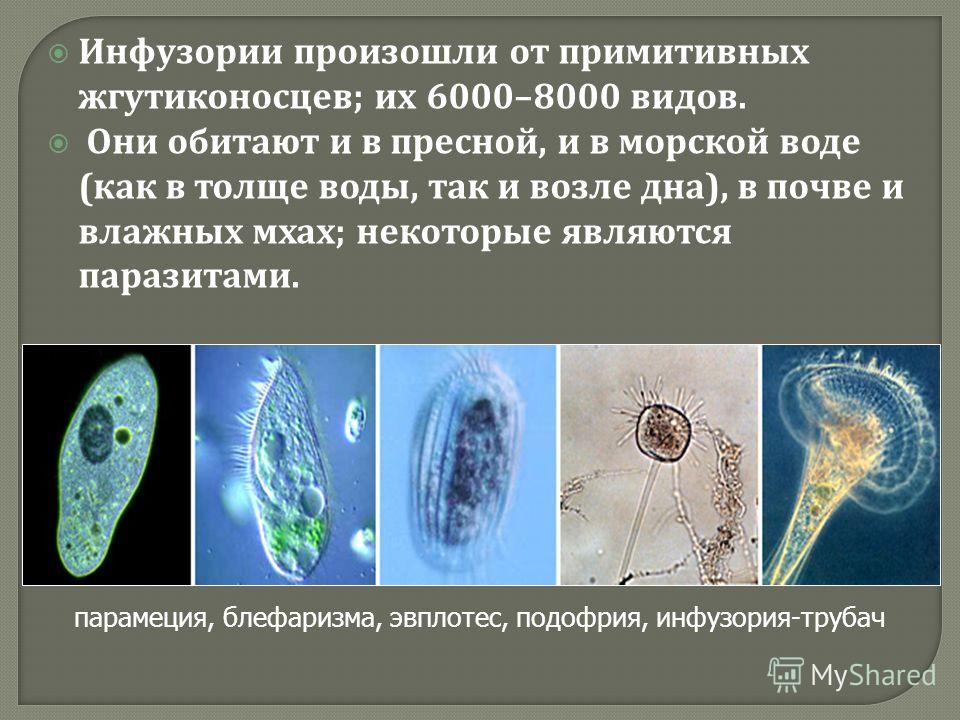 Ивасаки А., Меджитов Р. Регуляция адаптивного иммунитета врожденной иммунной системой. Наука (2010) 327:291–5. doi:10.1126/science.1183021
Ивасаки А., Меджитов Р. Регуляция адаптивного иммунитета врожденной иммунной системой. Наука (2010) 327:291–5. doi:10.1126/science.1183021
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
12. Майер-Барбер К.Д., Ян Б. Столкновение титанов цитокинов: контррегуляция воспалительных реакций, опосредованных интерлейкином-1 и интерфероном I типа. Cell Mol Immunol (2017) 14:22–35. doi:10.1038/cmi.2016.25
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
13. Кубо М., Мотомура Ю. Транскрипционная регуляция противовоспалительного цитокина IL-10 в приобретенных иммунных клетках. Фронт Иммунол (2012) 3:275. doi:10.3389/fimmu.2012.00275
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
14. Freitas do Rosario AP, Lamb T, Spence P, Stephens R, Lang A, Roers A, et al. IL-27 способствует продукции IL-10 эффекторными Th2 CD4+ T-клетками: критический механизм защиты от тяжелой иммунопатологии во время малярийной инфекции.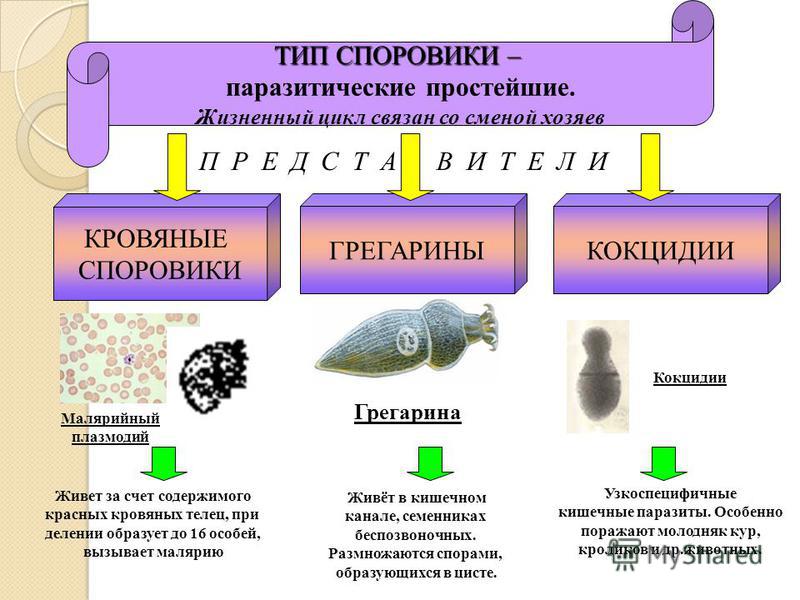 J Immunol (2012) 188:1178–90. doi:10.4049/jimmunol.1102755
J Immunol (2012) 188:1178–90. doi:10.4049/jimmunol.1102755
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
15. Фальво Ю.В., Ясенский Л.Д., Крюденьер Л., Гольдфельд А.Е. Эпигенетический контроль экспрессии генов цитокинов: регуляция локуса TNF/LT и дифференцировка Т-хелперов. Adv Immunol (2013) 118:37–128. doi:10.1016/B978-0-12-407708-9.00002-9
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
16. Mia S, Warnecke A, Zhang XM, Malmstrom V, Harris RA. Оптимизированный протокол для макрофагов M2 человека с использованием M-CSF и IL-4/IL-10/TGF-бета дает доминирующий иммуносупрессивный фенотип. Scand J Immunol (2014) 79:305–14. doi:10.1111/sji.12162
CrossRef Full Text | Google Scholar
17. Xiao C, Rajewsky K. Контроль микроРНК в иммунной системе: основные принципы. Cell (2009) 136:26–36. doi:10.1016/j.cell.2008.12.027
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
18. Балтимор Д., Болдин М.П., О’Коннелл Р.М., Рао Д.С., Таганов К.Д. МикроРНК: новые регуляторы развития и функционирования иммунных клеток. Нат Иммунол (2008) 9:839–45. doi:10.1038/ni.f.209
Балтимор Д., Болдин М.П., О’Коннелл Р.М., Рао Д.С., Таганов К.Д. МикроРНК: новые регуляторы развития и функционирования иммунных клеток. Нат Иммунол (2008) 9:839–45. doi:10.1038/ni.f.209
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
19. Асирватам А.Дж., Магнер В.Дж., Томази Т.Б. микроРНК регуляция генов цитокинов. Цитокин (2009) 45:58–69. doi:10.1016/j.cyto.2008.11.010
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
20. Макговерн К.Э., Уилсон Э.Х. Роль хемокинов и трафик иммунных клеток при паразитарных инфекциях. Curr Immunol Rev (2013) 9:157–68. дои: 10.2174/1573395509666131217000000
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
21. Diaz-Gandarilla JA, Osorio-Trujillo C, Hernandez-Ramirez VI, Talamas-Rohana P. Активация PPAR вызывает поляризацию макрофагов M1 посредством ингибирования cPLA(2)-COX-2, активируя выработку ROS против Leishmania mexicana. . Biomed Res Int (2013) 2013:215283. doi:10.1155/2013/215283
. Biomed Res Int (2013) 2013:215283. doi:10.1155/2013/215283
CrossRef Полный текст | Google Scholar
22. Lee CG, Sahoo A, Im SH. Эпигенетическая регуляция экспрессии генов цитокинов в Т-лимфоцитах. Yonsei Med J (2009) 50:322–30. doi:10.3349/ymj.2009.50.3.322
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
23. Zhu JF, Jankovic D, Oler AJ, Wei G, Sharma S, Hu GQ, et al. Фактор транскрипции T-bet индуцируется несколькими путями и предотвращает эндогенную программу клеток Th3 во время ответов клеток Th2. Иммунитет (2012) 37:660–73. doi:10.1016/j.immuni.2012.09.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
24. Баркер Б.Р., Taxman DJ, Ting JP. Перекрестная регуляция между IL-1beta/IL-18, процессирующим инфламмасому, и другими воспалительными цитокинами. Curr Opin Immunol (2011) 23:591–7. doi:10.1016/j.coi.2011.07.005
Полный текст CrossRef | Google Scholar
25. Liu Y, Zhang Q, Ding Y, Li X, Zhao D, Zhao K, et al. Гистон-лизин-метилтрансфераза Ezh2 способствует продукции воспалительных цитокинов, запускаемой TLR, путем подавления Tollip. J Immunol (2015) 194:2838–46. doi:10.4049/jimmunol.1402087
Liu Y, Zhang Q, Ding Y, Li X, Zhao D, Zhao K, et al. Гистон-лизин-метилтрансфераза Ezh2 способствует продукции воспалительных цитокинов, запускаемой TLR, путем подавления Tollip. J Immunol (2015) 194:2838–46. doi:10.4049/jimmunol.1402087
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
26. Вайкс М.Н., Левин С.Р. Блокада иммунных контрольных точек при инфекционных заболеваниях. Nat Rev Immunol (2018) 18:91–104. doi:10.1038/nri.2017.112
CrossRef Полный текст | Google Scholar
27. Cheeseman K, Weitzman JB. Взаимодействия хозяин-паразит: интимные эпигенетические отношения. Cell Microbiol (2015) 17:1121–32. doi:10.1111/cmi.12471
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
28. Гош Д., Штумхофер Дж.С. Вы видите то, что вижу я: распознавание простейших паразитов толл-подобными рецепторами. Карр Иммунол Ред. (2013) 9:129–40. doi:10.2174/1573395509666131203225929
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
29.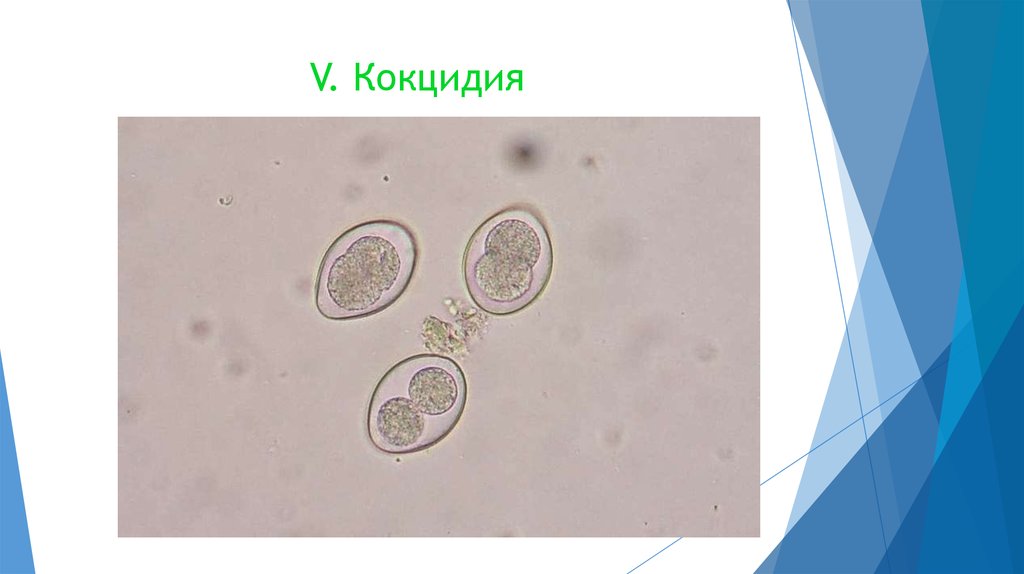 Сибли Л.Д. Инвазия и внутриклеточное выживание простейших паразитов. Immunol Rev (2011) 240:72–91. doi:10.1111/j.1600-065X.2010.00990.x
Сибли Л.Д. Инвазия и внутриклеточное выживание простейших паразитов. Immunol Rev (2011) 240:72–91. doi:10.1111/j.1600-065X.2010.00990.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
30. Kim BH, Shenoy AR, Kumar P, Bradfield CJ, MacMicking JD. IFN-индуцируемые ГТФазы в защите клетки-хозяина. Микроб-хозяин клетки (2012) 12:432–44. doi:10.1016/j.chom.2012.09.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
31. Du J, An R, Chen L, Shen Y, Chen Y, Cheng L, et al. Фактор вирулентности Toxoplasma gondii ROP18 ингибирует путь NF-kappaB хозяина, способствуя деградации p65. J Biol Chem (2014) 289:12578–92. doi:10.1074/jbc.M113.544718
CrossRef Полный текст | Google Scholar
32. Saeij JPJ, Coller S, Boyle JP, Jerome ME, White MW, Boothroyd JC. Toxoplasma кооптирует экспрессию гена хозяина путем инъекции гомолога полиморфной киназы. Природа (2007) 445:324–7. doi:10.1038/nature05395
doi:10.1038/nature05395
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
33. Rosowski EE, Lu D, Julien L, Rodda L, Gaiser RA, Jensen KD, et al. Штамм-специфическая активация пути NF-kappaB с помощью GRA15, нового белка плотных гранул Toxoplasma gondii . J Exp Med (2011) 208:195–212. doi:10.1084/jem.20100717
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
34. Франко М., Шастри А.Дж., Бутройд Дж.К. Инфекция Toxoplasma gondii специфически индуцирует c-Myc хозяина и гены, которые регулирует этот ключевой транскрипционный фактор. Эукариотическая клетка (2014) 13:483–93. doi:10.1128/EC.00316-13
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
35. Peixoto L, Chen F, Harb OS, Davis PH, Beiting DP, Brownback CS, et al. Интегративные геномные подходы выделяют семейство специфичных для паразитов киназ, которые регулируют ответы хозяина. Микроб-хозяин клетки (2010) 8:208–18. doi:10.1016/j.chom.2010.07.004
doi:10.1016/j.chom.2010.07.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
36. Bougdour A, Durandau E, Brenier-Pinchart MP, Ortet P, Barakat M, Kieffer S, et al. Подрывная деятельность клетки-хозяина с помощью Toxoplasma GRA16, экспортируемого белка плотных гранул, который нацелен на ядро клетки-хозяина и изменяет экспрессию генов. Микроб-хозяин клетки (2013) 13:489–500. doi:10.1016/j.chom.2013.03.002
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
37. Cardoso MS, Reis-Cunha JL, Bartholomeu DC. Уклонение от иммунного ответа Trypanosoma cruzi во время острой инфекции. Фронт Иммунол (2015) 6:659. doi:10.3389/fimmu.2015.00659
CrossRef Full Text | Google Scholar
38. Родригес М.М., Оливейра А.С., Беллио М. Иммунный ответ на Trypanosoma cruzi : роль толл-подобных рецепторов и перспективы разработки вакцин. J Parasitol Res (2012) 2012:507874.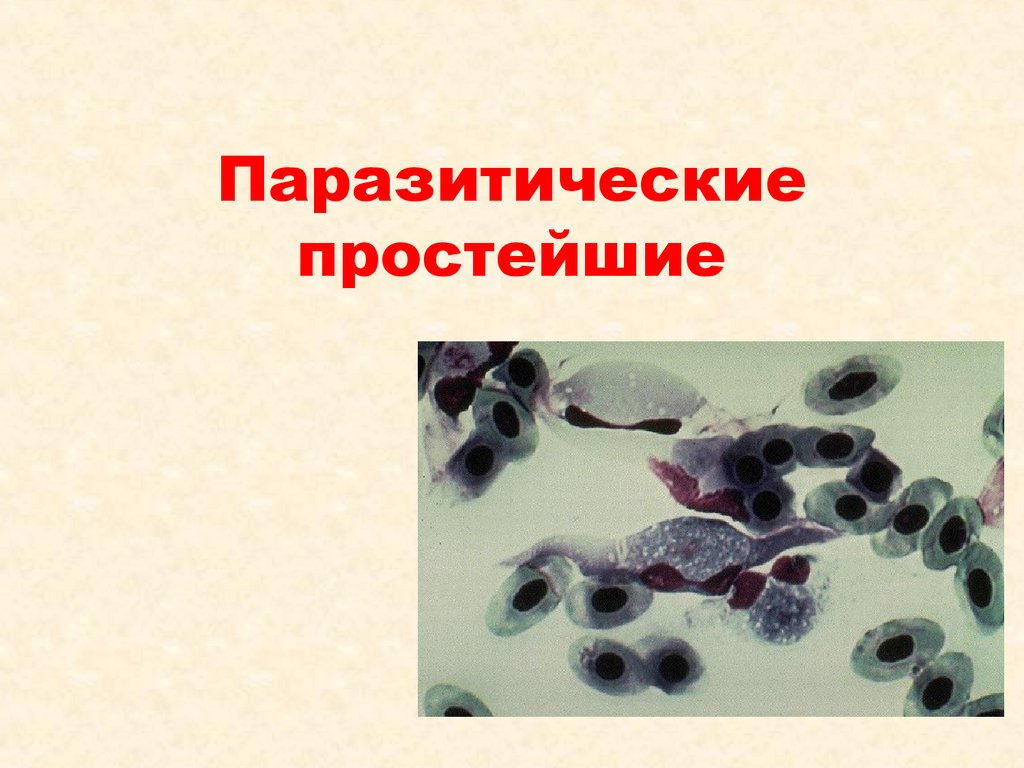 дои: 10.1155/2012/507874
дои: 10.1155/2012/507874
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
39. Silva GK, Gutierrez FR, Guedes PM, Horta CV, Cunha LD, Mineo TW, et al. Передний край: ответы, зависящие от домена 1 олигомеризации, связывающего нуклеотиды, объясняют устойчивость мышей к инфекции Trypanosoma cruzi . J Immunol (2010) 184:1148–52. doi:10.4049/jimmunol.04
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
40. ДосРейс Г.А. Уклонение от иммунных ответов на Trypanosoma cruzi , этиологический агент болезни Шагаса. Braz J Med Biol Res (2011) 44:84–90. doi:10.1590/S0100-879X2011007500005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
41. Cribb P, Perdomo V, Alonso VL, Manarin R, Barrios-Payan J, Marquina-Castillo B, et al. Trypanosoma cruzi высокой подвижности группы B (TcHMGB) может действовать как медиатор воспаления на клетки млекопитающих. Plos Negl Trop Dis (2017) 11:e0005350. doi:10.1371/journal.pntd.0005350
doi:10.1371/journal.pntd.0005350
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
42. Хедрик П.В. Популяционная генетика малярийной резистентности человека. Наследственность (2011) 107:283–304. doi:10.1038/hdy.2011.16
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
43. Всемирная организация здравоохранения. Информационный бюллетень о малярии № 94 . Доступно на сайте www.who.int/mediacentre/factsheets/fs375/en/
Google Scholar
44. Joice R, Narasimhan V, Montgomery J, Sidhu AB, Oh K, Meyer E, et al. Вывод состава стадии развития по экспрессии генов при малярии человека. PLoS Comput Biol (2013) 9:e1003392. doi:10.1371/journal.pcbi.1003392
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
45. Zheng H, Tan Z, Xu W. Стратегии уклонения от иммунитета преэритроцитарных малярийных паразитов. Медиаторы Inflamm (2014) 2014:362605. doi:10. 1155/2014/362605
1155/2014/362605
Полный текст CrossRef | Google Scholar
46. Бертолино П., Боуэн Д.Г. Малярия и печень: иммунологические прятки или подрыв иммунитета изнутри? Front Microbiol (2015) 6:41. doi:10.3389/fmicb.2015.00041
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
47. Кларк И.А., Бадд А.С., Аллева Л.М., Кауден В.Б. Малярийное заболевание человека: следствие выброса воспалительных цитокинов. Малар Дж. (2006) 5:85. doi:10.1186/1475-2875-5-85
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
48. Rovira-Vallbona E, Moncunill G, Bassat Q, Aguilar R, Machevo S, Puyol L, et al. Низкие антитела против Plasmodium falciparum и несбалансированные провоспалительные цитокины связаны с тяжелой малярией у мозамбикских детей: исследование случай-контроль. Малар Дж. (2012) 11:181. doi:10.1186/1475-2875-11-181
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
49. Anyona SB, Kempaiah P, Raballah E, Ouma C, Were T, Davenport GC, et al. Функциональные промоторные гаплотипы интерлейкина-18 обусловливают предрасположенность к тяжелой малярийной анемии и детской смертности. Infect Immun (2011) 79:4923–32. doi:10.1128/IAI.05601-11
Anyona SB, Kempaiah P, Raballah E, Ouma C, Were T, Davenport GC, et al. Функциональные промоторные гаплотипы интерлейкина-18 обусловливают предрасположенность к тяжелой малярийной анемии и детской смертности. Infect Immun (2011) 79:4923–32. doi:10.1128/IAI.05601-11
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
50. Ayimba E, Hegewald J, Segbena AY, Gantin RG, Lechner CJ, Agosssou A, et al. Провоспалительные и регуляторные цитокины и хемокины у младенцев с неосложненной и тяжелой малярией Plasmodium falciparum . Clin Exp Immunol (2011) 166:218–26. doi:10.1111/j.1365-2249.2011.04474.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
51. Boeuf PS, Loizon S, Awandare GA, Tetteh JKA, Addae MM, Adjei GO, et al. Взгляд на нарушение регуляции производства ФНО и ИЛ-10 при малярии: последствия для понимания тяжелой малярийной анемии. Малар Дж. (2012) 11:253. doi:10.1186/1475-2875-11-253
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
52.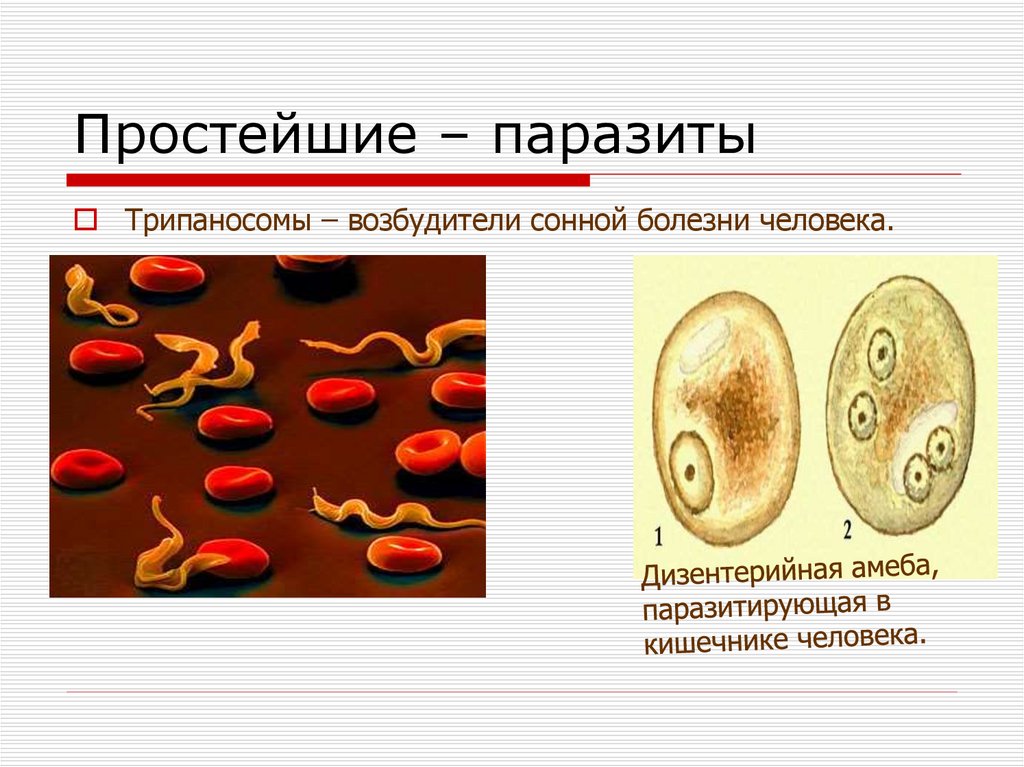 Mendonca VRR, Queiroz ATL, Lopes FM, Andrade BB, Barral-Netto M. Создание сети иммунного ответа хозяина при малярии Plasmodium vivax . Малар Дж. (2013) 12:69. doi:10.1186/1475-2875-12-69
Mendonca VRR, Queiroz ATL, Lopes FM, Andrade BB, Barral-Netto M. Создание сети иммунного ответа хозяина при малярии Plasmodium vivax . Малар Дж. (2013) 12:69. doi:10.1186/1475-2875-12-69
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
53. Ockenhouse CF, Hu WC, Kester KE, Cummings JF, Stewart A, Heppner DG, et al. Общие и различные пути передачи сигналов иммунного ответа, обнаруженные в паттернах экспрессии генов мононуклеарных клеток периферической крови при досимптомной и клинически очевидной малярии. Infect Immun (2006) 74:5561–73. doi:10.1128/IAI.00408-06
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
54. Sharma S, DeOliveira RB, Kalantari P, Parroche P, Goutagny N, Jiang Z, et al. Врожденное иммунное распознавание богатого AT мотива ДНК «стебель-петля» в геноме Plasmodium falciparum. Иммунитет (2011) 35:194–207. doi:10.1016/j.immuni.2011.05.016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
55. Маханта А., Кар С.К., Какати С., Баруа С. Усиленное воспаление при тяжелой малярии связано со снижением уровня экспрессии ИЛ-10 и нейтрофилов. Innate Immun (2015) 21:546–52. doi:10.1177/1753425
Маханта А., Кар С.К., Какати С., Баруа С. Усиленное воспаление при тяжелой малярии связано со снижением уровня экспрессии ИЛ-10 и нейтрофилов. Innate Immun (2015) 21:546–52. doi:10.1177/1753425
1277
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
56. Сторм Дж., Крейг А.Г. Патогенез церебральной малярии-воспаление и цитоадгезия. Front Cell Infect Microbiol (2014) 4:100. doi:10.3389/fcimb.2014.00100
Полный текст CrossRef | Google Scholar
57. Lourembam SD, Sawian CE, Baruah S. Нарушение регуляции экспрессии цитокинов при осложненной малярии falciparum с повышением TGF-бета и IFN-гамма и снижением IL-2 и IL-12. Цитокин (2013) 64:503–8. doi:10.1016/j.cyto.2013.08.007
Полный текст CrossRef | Google Scholar
58. Tahar R, Albergaria C, Zeghidour N, Ngane VF, Basco LK, Roussilhon C. Плазменные уровни восьми различных медиаторов и их потенциал в качестве биомаркеров различных клинических состояний малярии у африканских детей. Малар Дж. (2016) 15:337. doi:10.1186/s12936-016-1378-3
Малар Дж. (2016) 15:337. doi:10.1186/s12936-016-1378-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
59. Engwerda CR, Mynott TL, Sawhney S, De Souza JB, Bickle QD, Kaye PM. Локально активированный лимфотоксин альфа, а не системный фактор некроза опухоли альфа, является основным медиатором мышиной церебральной малярии. J Exp Med (2002) 195:1371–7. doi:10.1084/jem.20020128
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
60. Се Ф.Л., Тернер Л., Болла Дж.Р., Робинсон К.В., Лавстсен Т., Хиггинс М.К. Структурная основа связывания CD36 малярийным паразитом. Nat Commun (2016) 7:12837. doi:10.1038/ncomms12837
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
61. Бернабеу М., Данцигер С.А., Аврил М., Ваз М., Бабар П.Х., Брейзер А.Дж. и др. Тяжелая форма малярии у взрослых связана со специфическими типами адгезии PfEMP1 и высокой биомассой паразита. Proc Natl Acad Sci U S A (2016) 113:E3270–9. doi:10.1073/pnas.1524294113
doi:10.1073/pnas.1524294113
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
62. Саранджи П.П., Ли Х.В., Ким М. Действие активированного протеина С при воспалении. Br J Haematol (2010) 148:817–33. doi:10.1111/j.1365-2141.2009.08020.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
63. Gazzinelli RT, Kalantari P, Fitzgerald KA, Golenbock DT. Врожденное ощущение малярийных паразитов. Nat Rev Immunol (2014) 14:744–57. doi:10.1038/nri3742
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
64. Franchi L, Munoz-Planillo R, Nunez G. Восприятие микробов и реакция на них через инфламмасомы. Nat Immunol (2012) 13:325–32. doi:10.1038/ni.2231
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
65. Kalantari P, DeOliveira RB, Chan J, Corbett Y, Rathinam V, Stutz A, et al. Двойное вовлечение инфламмасом NLRP3 и AIM2 к Plasmodium — производный гемозоина и ДНК во время малярии. Cell Rep (2014) 6:196–210. doi:10.1016/j.celrep.2013.12.014
Cell Rep (2014) 6:196–210. doi:10.1016/j.celrep.2013.12.014
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
66. Дурай П., Говиндарадж Р.Г., Чой С. Структура и динамическое поведение подсемейства толл-подобных рецепторов 2, запускаемых малярийными гликозилфосфатидилинозитолами Plasmodium falciparum . FEBS J (2013) 280:6196–212. doi:10.1111/febs.12541
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
67. Пичьянгкул С., Йонгванитчит К., Кум-Арб У., Хемми Х., Акира С., Криг А.М. и соавт. Паразиты кровяной стадии малярии активируют плазмоцитоидные дендритные клетки человека и мышиные дендритные клетки посредством зависимого от рецептора 9 пути, подобного толл-подобному. J Immunol (2004) 172:4926–33. doi:10.4049/jimmunol.172.8.4926
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
68. Маханта А., Баруа С. Низкая экспрессия GATA3 и T-bet коррелирует со сниженной регуляцией IL-10 при тяжелой форме falciparum малярия. Clin Transl Immunology (2015) 4:e49. doi:10.1038/cti.2015.30
Clin Transl Immunology (2015) 4:e49. doi:10.1038/cti.2015.30
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
69. Шахам Ш.Х. Регулирование IL-10 при осложненной малярии . [диссертация/магистерская диссертация], Университет Тезпур.
Реферат PubMed | Google Scholar
70. Крофт С.Л., Зайферт К., Ярдли В. Текущий сценарий разработки лекарств от лейшманиоза. Индиан J Med Res (2006) 123:399–410.
Реферат PubMed | Google Scholar
71. Гангули П., Чоудхури С., Саркар Р.Р. Идентификация регуляторного переключения Th2/Th3 для стимулирования реакции заживления при лейшманиозе: вычислительный подход. EURASIP J Bioinform Syst Biol (2015) 2015(1):13. doi:10.1186/s13637-015-0032-7
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
72. Кумар Р., Энгверда К. Вакцины для профилактики лейшманиоза. Clin Transl Immunology (2014) 3:e13. doi:10.1038/cti.2014.4
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
73. Субраманиан А., Саркар Р.Р. Раскрытие тайны метаболических адаптаций с использованием масштабной модели генома Leishmania infantum . Научный представитель (2017) 7:10262. doi:10.1038/s41598-017-10743-x
Субраманиан А., Саркар Р.Р. Раскрытие тайны метаболических адаптаций с использованием масштабной модели генома Leishmania infantum . Научный представитель (2017) 7:10262. doi:10.1038/s41598-017-10743-x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
74. Saunders EC, Ng WW, Kloehn J, Chambers JM, Ng M, McConville MJ. Индукция строгого метаболического ответа на внутриклеточных стадиях Leishmania mexicana приводит к повышенной зависимости от митохондриального метаболизма. PLoS Pathog (2014) 10:e1003888. doi:10.1371/journal.ppat.1003888
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
75. Туон Ф.Ф., Амато В.С., Бача Х.А., Альмусави Т., Дуарте М.И., Амато Нето В. Толл-подобные рецепторы и лейшманиоз. Infect Immun (2008) 76:866–72. doi:10.1128/IAI.01090-07
Полный текст CrossRef | Google Scholar
76. Фариа М.С., Рейс Ф.К., Лима А.П. Толл-подобные рецепторы в Инфекции Leishmania : опекуны или промоутеры? J Parasitol Res (2012) 2012:
7. doi:10.1155/2012/
doi:10.1155/2012/
7
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
77. Бхардвадж С., Сривастава Н., Судан Р., Саха Б. Лейшмани а мешает передаче сигналов клетки-хозяина для разработки стратегии выживания. J Biomed Biotechnol (2010) 2010:109189. doi:10.1155/2010/109189
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
78. Shio MT, Hassani K, Isnard A, Ralph B, Contreras I, Gomez MA, et al. Передача сигналов клетками-хозяевами и механизмы уклонения от лейшманиоза. J Trop Med (2012) 2012:819512. doi:10.1155/2012/819512
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
79. Каммингс Х.Э., Туладхар Р., Сатоскар А.Р. Цитокины и их STAT при кожном и висцеральном лейшманиозе. J Biomed Biotechnol (2010) 2010:294389. doi:10.1155/2010/294389
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
80. Диас Ю.Р., Рохас Р. , Вальдеррама Л., Саравиа Н.Г. Экспрессия T-bet, GATA-3 и Foxp3 и продукция цитокинов Th2/Th3 в клинических исходах инфекции человека видами Leishmania (Viannia). J Infect Dis (2010) 202:406–15. doi:10.1086/653829
, Вальдеррама Л., Саравиа Н.Г. Экспрессия T-bet, GATA-3 и Foxp3 и продукция цитокинов Th2/Th3 в клинических исходах инфекции человека видами Leishmania (Viannia). J Infect Dis (2010) 202:406–15. doi:10.1086/653829
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
81. Olivier M, Gregory DJ, Forget G. Подрывные механизмы, с помощью которых паразиты Leishmania могут избежать иммунного ответа хозяина: сигнальная точка зрения. Clin Microbiol Rev (2005) 18:293. doi:10.1128/CMR.18.2.293-305.2005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
82. Kaye PM, Rogers NJ, Curry AJ, Scott JC. Недостаточная экспрессия костимулирующих молекул на инфицированных Leishmania макрофагах. Eur J Immunol (1994) 24:2850–4. doi:10.1002/eji.1830241140
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
83. Soong L, Xu JC, Grewal IS, Kima P, Sun J, Longley BJ Jr. , et al. Нарушение взаимодействия лигандов CD40-CD40 приводит к повышенной чувствительности к Инфекция Leishmania amazonensis . Иммунитет (1996) 4:263–73. doi:10.1016/S1074-7613(00)80434-3
, et al. Нарушение взаимодействия лигандов CD40-CD40 приводит к повышенной чувствительности к Инфекция Leishmania amazonensis . Иммунитет (1996) 4:263–73. doi:10.1016/S1074-7613(00)80434-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
84. McMahon-Pratt D, Alexander J. Справедлива ли парадигма патогенеза и защиты Leishmania major для кожного лейшманиоза Нового Света или висцерального заболевания? Immunol Rev (2004) 201:206–24. doi:10.1111/j.0105-2896.2004.00190.x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
85. Gregory DJ, Sladek R, Olivier M, Matlashewski G. Сравнение эффектов инфекции Leishmania major или Leishmania donovani на экспрессию генов макрофагов. Infect Immun (2008) 76:1186–92. doi:10.1128/IAI.01320-07
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
86. МакКолл Л.И., Чжан В.В., Матлашевски Г. Детерминанты развития висцерального лейшманиоза.![]() PLoS Pathog (2013) 9:e1003053. doi:10.1371/journal.ppat.1003053
PLoS Pathog (2013) 9:e1003053. doi:10.1371/journal.ppat.1003053
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
87. Mendez S, Reckling SK, Piccirillo CA, Sacks D, Belkaid Y. Роль CD4(+) CD25(+) регуляторных Т-клеток в реактивации персистирующего лейшманиоза и контроле сопутствующего иммунитета. J Exp Med (2004) 200:201–10. doi:10.1084/jem.20040298
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
88. Хилл А.В., Джепсон А., Плебански М., Гилберт С.К. Генетический анализ коэволюции хозяина и паразита при малярии человека. Philos Trans R Soc Lond B Biol Sci (1997) 352:1317–25. doi:10.1098/rstb.1997.0116
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
89. Aurrecoechea C, Barreto A, Basenko EY, Brestelli J, Brunk BP, Cade S, et al. EuPathDB: ресурс базы данных геномики эукариотических патогенов. Nucleic Acids Res (2017) 45:D581–91. doi:10.1093/nar/gkw1105
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
90. Урбан М., Пант Р., Рагунатх А., Ирвин А.Г., Педро Х., Хаммонд-Косак К.Е. База данных взаимодействий патоген-хозяин (база PHI): дополнения и будущие разработки. Nucleic Acids Res (2015) 43:D645–55. doi:10.1093/nar/gku1165
Урбан М., Пант Р., Рагунатх А., Ирвин А.Г., Педро Х., Хаммонд-Косак К.Е. База данных взаимодействий патоген-хозяин (база PHI): дополнения и будущие разработки. Nucleic Acids Res (2015) 43:D645–55. doi:10.1093/nar/gku1165
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
91. Давила А.М., Мендес П.Н., Вагнер Г., Чоке Д.А., Куадрат Р.Р., Либерман Ф. и другие. ProtozoaDB: динамическая визуализация и исследование геномов простейших. Nucleic Acids Res (2008) 36:D547–52. doi:10.1093/nar/gkm820
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
92. Суонн Дж., Джамшиди Н., Льюис Н.Е., Винзелер Э.А. Системный анализ взаимодействия паразит-хозяин. Wiley interdiscip Rev Syst Biol Med (2015) 7:381–400. doi:10.1002/wsbm.1311
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
93. Вестерманн А.Дж., Горски С.А., Фогель Дж. Двойная последовательность РНК патогена и хозяина. Nat Rev Microbiol (2012) 10:618–30. doi:10.1038/nrmicro2852
Nat Rev Microbiol (2012) 10:618–30. doi:10.1038/nrmicro2852
CrossRef Полный текст | Google Scholar
94. Райт Х.Л., Томас Х.Б., Мутс Р.Дж., Эдвардс С.В. RNA-Seq выявляет активацию как общих, так и специфичных для цитокинов путей после праймирования нейтрофилов. PLoS One (2013) 8:e58598. doi:10.1371/journal.pone.0058598
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
95. Yamagishi J, Natori A, Tolba MEM, Mongan AE, Sugimoto C, Katayama T, et al. Интерактивный транскриптомный анализ больных малярией и инфицированных Plasmodium falciparum . Genome Res (2014) 24:1433–44. doi:10.1101/gr.158980.113
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
96. Питтман К.Дж., Алиота М.Т., Нолл Л.Дж. Двойное транскрипционное профилирование мышей и Toxoplasma gondii при острой и хронической инфекции. BMC Genomics (2014) 15:806. doi:10.1186/1471-2164-15-806
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
97. Дериг С., Райнер Дж. К., Шерф А., Тобин А. Б. Посттрансляционные модификации белков у малярийных паразитов. Nat Rev Microbiol (2015) 13:160–72. doi:10.1038/nrmicro3402
Дериг С., Райнер Дж. К., Шерф А., Тобин А. Б. Посттрансляционные модификации белков у малярийных паразитов. Nat Rev Microbiol (2015) 13:160–72. doi:10.1038/nrmicro3402
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
98. Цуй Л.В., Линднер С., Мяо Дж. Трансляционная регуляция во время стадийных переходов у малярийных паразитов. Ann NY Acad Sci (2015) 1342:1–9. doi:10.1111/nyas.12573
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
99. Bunnik EM, Chung DWD, Hamilton M, Ponts N, Saraf A, Prudhomme J, et al. Полисомное профилирование выявило трансляционный контроль экспрессии генов у человеческого малярийного паразита Plasmodium falciparum . Genome Biol (2013) 14:R128. doi:10.1186/gb-2013-14-11-r128
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Parasitic Protozoa — 2nd Edition
Select country/regionUnited States of AmericaUnited KingdomAfghanistanÅland IslandsAlbaniaAlgeriaAmerican SamoaAndorraAngolaAnguillaAntigua and BarbudaArgentinaArmeniaArubaAustraliaAustriaAzerbaijanBahamasBahrainBangladeshBarbadosBelgiumBelizeBeninBermudaBhutanBoliviaBonaire, Sint Eustatius and SabaBosnia and HerzegovinaBotswanaBrazilBritish Indian Ocean TerritoryBritish Virgin IslandsBruneiBulgariaBurkina FasoBurundiCambodiaCameroonCanadaCanary IslandsCape VerdeCayman IslandsCentral African RepublicChadChileChinaChristmas IslandCocos (Keeling) IslandsColombiaComorosCongoCook IslandsCosta RicaCroatiaCubaCuraçaoCyprusCzech RepublicDemocratic Republic of the КонгоДанияДжибутиДоминикаДоминиканская РеспубликаЭквадорЕгипетСальвадорЭкваториальная ГвинеяЭритреяЭстонияЭфиопияФолклендские (Мальвинские) островаФарерские островаФедеративные Штаты МикронезииФиджиФинляндияФранцияФранцузская ГвианаФранцузская ПолинезияГабонГамбияГрузияГерманияГанаГибралтарГрецияГринлан dGrenadaGuadeloupeGuamGuatemalaGuernseyGuineaGuinea-BissauGuyanaHaitiHondurasHong KongHungaryIcelandIndiaIndonesiaIranIraqIrelandIsle of ManIsraelItalyJamaicaJapanJerseyJordanKazakhstanKenyaKiribatiKuwaitKyrgyzstanLaoLatviaLesothoLiberiaLibyaLiechtensteinLuxembourgMacaoMacedoniaMadagascarMalawiMalaysiaMaldivesMaliMaltaMarshall IslandsMartiniqueMauritaniaMauritiusMayotteMexicoMoldovaMonacoMongoliaMontenegroMontserratMoroccoMozambiqueMyanmarNamibiaNepalNetherlandsNew CaledoniaNew ZealandNicaraguaNigerNiueNorfolk IslandNorth KoreaNorthern Mariana IslandsNorwayOmanPakistanPalauPanamaPapua New GuineaParaguayPeruPhilippinesPitcairnPolandPortugalPuerto RicoQatarRéunionRomaniaRwandaSaint BarthélemySaint HelenaSaint Kitts and NevisSaint LuciaSaint Martin (French part)Saint Pierre and MiquelonSaint Vincent and the GrenadinesSamoaSan MarinoSao Tome and PrincipeSaudi ArabiaSenegalSerbiaSeychellesSierra LeoneSingaporeSint Maarten (Dutch part)SlovakiaSloveniaSolomon IslandsSomaliaSouth AfricaSouth G eorgia and the South Sandwich IslandsSouth KoreaSouth SudanSpainSri LankaSudanSurinameSvalbard and Jan MayenSwazilandSwedenSwitzerlandSyriaTaiwanTajikistanTanzaniaThailandTimor LesteTogoTokelauTongaTrinidad and TobagoTunisiaTurkeyTurkmenistanTurks and Caicos IslandsTuvaluUgandaUkraineUnited Arab EmiratesUruguayUS Virgin IslandsUzbekistanVanuatuVatican CityVenezuelaVietnamWallis and FutunaWestern SaharaYemenZambiaZimbabwe
Варианты покупкиЭлектронная книга $72,95
Налог с продаж рассчитывается при оформлении заказа
Бесплатная доставка по всему миру
Нет минимального заказа
Описание
Обновленное и значительно расширенное второе издание Parasitic Protozoa врачам, ветеринарам и ученым-исследователям, занимающимся болезнями, вызываемыми простейшими у человека, а также у домашних и диких животных, включая рыб, моллюсков и насекомых, а также у наиболее часто считающихся позвоночными животными.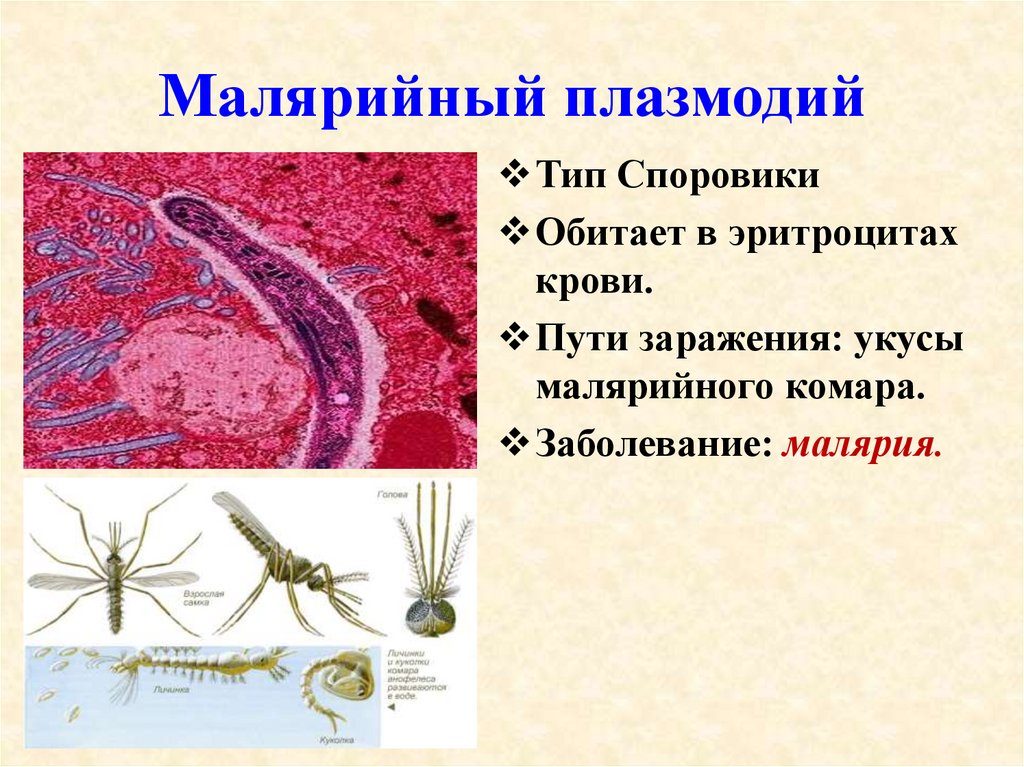 Каждый раздел содержит информацию о возбудителях болезней, лечении, диагностике и эпидемиологии болезней, вызываемых различными простейшими. Книга не ограничивается этими медицинскими предметами, но рассматривает таксономию, морфологию и метаболизм организмов таким образом, чтобы представлять интерес для ученых и аспирантов, работающих в области протозоологии. Все издание, опубликованное в десяти томах, организовано таким образом, что темы, представляющие общий интерес, занимают отдельные тома.
Каждый раздел содержит информацию о возбудителях болезней, лечении, диагностике и эпидемиологии болезней, вызываемых различными простейшими. Книга не ограничивается этими медицинскими предметами, но рассматривает таксономию, морфологию и метаболизм организмов таким образом, чтобы представлять интерес для ученых и аспирантов, работающих в области протозоологии. Все издание, опубликованное в десяти томах, организовано таким образом, что темы, представляющие общий интерес, занимают отдельные тома.
Содержание
-
Соавторы
Предисловие ко второму изданию
Предисловие к первому изданию
Глава I Птичий кокцидиоз
I. Введение: Значение кокцидий птиц
II· Таксономия и номенклатура III
III. Основные жизненные циклы 92 913 A. Стадии жизненного цикла 92 913 B. Краткий обзор жизненного цикла типичной эймерии: гомоксенный жизненный цикл 92 913 C. Уникальные аспекты жизненного цикла Crytosporidium: гомоксенный паразит, способный к аутоинфекции своего хозяина
г.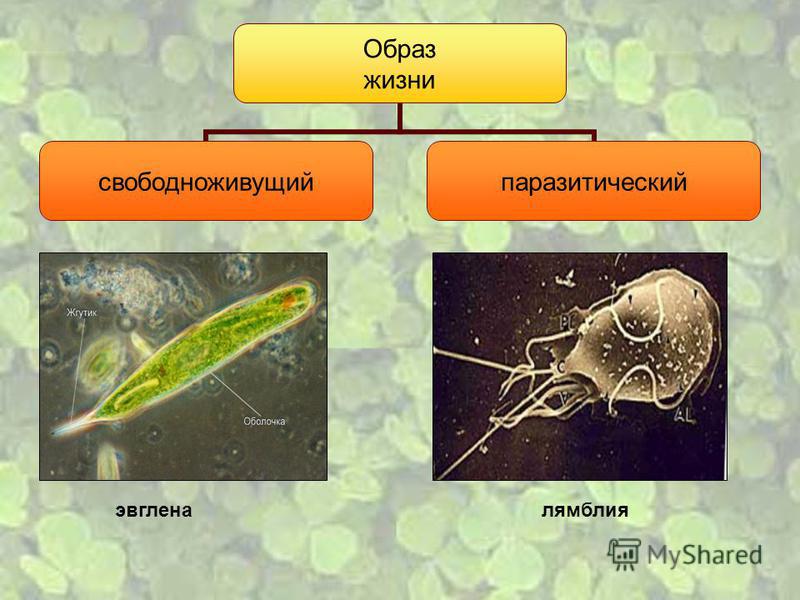 IV. Кокцидии птичьих хозяев
IV. Кокцидии птичьих хозяев
A. Классические кокцидии
B. Птичий криптоспоридиоз
C. Птичий токсоплазмоз
V. Генетика кокцидий
A. Половая дифференциация и клонирование
B. Штаммовая изменчивость кокцидиальной вирулентности
C. Перекрестное оплодотворение
D. Раннее развитие
VI. Хозяйственная специфичность кокцидий
VII. Сайтоспецифичность кокцидий
VIII. Жизнеспособность кокцидиальных ооцист
IX. Метаболизм кокцидий
X. Патофизиологические изменения, вызванные заражением кокцидиями
XI. Влияние дозы и частоты введения ооцист на кокцидиоз
XII. Влияние возраста хозяина на кокцидиоз
XIII. Влияние кишечных бактерий и пищевого статуса хозяина на кокцидиоз
XIV. Взаимодействие с вирусами
XV. Взаимодействия между видами Eimeria
XVI. Иммунитет против инфекции Eimeria 92 913 A. Эндогенные стадии, инициирующие иммунный ответ 92 913 B. Стадии, затронутые иммунным ответом 92 913 C. Продолжительность иммунного ответа 929-13 D. Иммунизация с использованием различных путей инокуляции 929-13 E. Механизмы с использованием различных путей инокуляции 929-13 F. Практические аспекты иммунитета к кокцидиям 929-13 XVII. Иммунизация против кокцидиоза
Иммунизация с использованием различных путей инокуляции 929-13 E. Механизмы с использованием различных путей инокуляции 929-13 F. Практические аспекты иммунитета к кокцидиям 929-13 XVII. Иммунизация против кокцидиоза
A. Аттенуация Eimeria
B. Иммунизация цыплят индукцией инфекции и введением антигенов паразитов
XVIII. Борьба с помощью химиотерапии
A. История лечения кокцидиозом
B. Типы используемых препаратов
C. Перспективы химиотерапии и других форм борьбы с кокцидиозом в будущем
D. Экономические аспекты химиотерапии
Ссылки
Глава 2 Кокцидии млекопитающих
I. Введение
II. Жизненные циклы
A. Спорогония
B. Эксцистация
C. Эндогенное развитие
D. Специфика хозяина
III. Кокцидиоз человека и домашних млекопитающих
A. Человек
B. Нечеловеческие приматы
C. Лошади
D. Крупный рогатый скот
E. Овцы
F. Козы
G. Свиньи
H. Собаки
I. Кошки
J. Лабораторные мыши
K Лабораторные крысы
L. Лабораторные кролики
Лабораторные кролики
M. Другие лабораторные животные
IV. Ультраструктура и клеточное проникновение 929-13 A. Ультраструктура подвижных стадий 929-13 B. Ультраструктура мерогонических стадий 929-13 C. Ультраструктура гамогонических стадий 929-13 D. Проникновение клеток-хозяев 929-13 V. Развитие in Vitro 929-13 A. Клеточные культуры 929-13 B. Эмбрионы птиц 929-13 VI. Иммунитет 929–13 A. Антигены 929–13 B. Поликлональные и моноклональные антитела 929–13 C. Клеточный иммунитет 929–13 VII. Caryospora: необычные кокцидии 92 913 A. Жизненный цикл Caryospora
B. Заболевание у первичных хозяев
C. Заболевание у вторичных хозяев
D. Естественно встречающиеся инфекции Caryospora у собак
VIII. Резюме и выводы
Список литературы
Глава 3 Грегарины
I. Введение
II. Жизненный цикл эвгрегарина и различные типы трофозоитов 929-13 А. Жизненный цикл эвгрегарина 929-13 Б. Различные типы трофозоитов 929-13 III. Клеточная и молекулярная организация разных стадий 929–13 А. Молекулярная организация клеточной коры трофозоитов
Молекулярная организация клеточной коры трофозоитов
B. Цитоскелет и клеточная подвижность трофозоитов
C. Связь разнообразия прикрепления грегарин апекса трофозоитов с питанием
D. Цитоплазматические органеллы трофозоитов
E. Сизигия и узнавание клеток между гомантами
F. Гамогонии
G. Гаметы и Фекондация: вариации 9 + 0, 6 + 0 или 3 + 0 аксонемных паттернов в жгутиках мужских гамет
H. Спорогоны
I. Использование характеристик «спороцист» и спороцист в качестве критериев для таксономии грегарин
J. Переход от спорозоита к трофозоиту у эвгрегарин
IV. Взаимодействия хозяин-паразит: изобилие грегарин 929-13 A. Поведение грегарин 929-13 B. Электрофизиологические свойства кортикальных мембран трофозоитов 929-13 C. Эндосимбионты и гиперпаразитемия 929-13 D. Экспериментальный контроль жизненных циклов грегарин 929-13 E. Корреляции между жизненными циклами хозяина и паразита и хронология жизненных циклов грегарин
V. Шизогония и таксономия грегарин
A. Статус Selenidiidae и определение архигрегарин
Статус Selenidiidae и определение архигрегарин
B. Шизогония и неогрегарины
VI. Заключение
Литература
Глава 4 Haemogregarinidae и Lankesterellidae
I. Введение
II. Haemogregarinidae
A. Таксономия
B. Морфология
C. Жизненные циклы
D. Патогенез
III. Lankesterellidae
A. Таксономия
B. Морфология
C. Жизненные циклы
IV. Заключительные замечания
Литература
Глава 5 Роды Leucocytozoon, Haemoproteus и Hepatocystis
I. Введение
г. II. Таксономия Haemosporina/Haemospororida
III. Leucocytozoon, Sambon, 1908
A. Морфология и жизненные циклы
B. Патология
V. Haemoproteus, Kruse, 1890
A. Морфология и жизненные циклы
B. Патология
и гемопротеус
VI. Hepatocystis, Levaditi and Schoen, 1932, emend, Garnham, 1951
A. Морфология и жизненные циклы
B. Переносчики и экология Hepatocystis Kochi
C. Патология
VII. Сводка 304
Список литературы
ИНДЕКС
Подробная информация о продукте
- № Страницы: 323
- Язык: Английский
- Copyright: © Academic Press 1993
- .
